
Atlantic walrus (Odobenus rosmarus rosmarus): COSEWIC assessment and status report 2017
High Arctic population
Central-Low Arctic population
Nova Scotia-Newfoundland-Gulf of St. Lawrence population
Atlantic Walrus

High Arctic population - Special concern
Central-Low Arctic population - Special concern
Nova Scotia-Newfoundland-Gulf of St. Lawrence population - Extinct
2017
Table of contents
- Table of contents
- COSEWIC assessment summary - Atlantic Walrus - High Arctic population
- COSEWIC assessment summary - Atlantic Walrus - Central/low Arctic population
- COSEWIC assessment summary - Atlantic Walrus - Nova Scotia-Newfoundland-Gulf of St Lawrence population
- COSEWIC Executive summary
- Technical summary - High Arctic population
- Technical summary - Central-low Arctic population
- Technical summary - Nova Scotia-Newfoundland-Gulf of St. Lawrence population
- Preface
- Wildlife species description and significance
- Distribution
- Habitat
- Biology
- Population sizes and trends
- Threats and limiting factors
- Protection, status and ranks
- Acknowledgements
- Information sources
- Biographical summary of report writers
- Personal communications/authorities contacted
List of figures
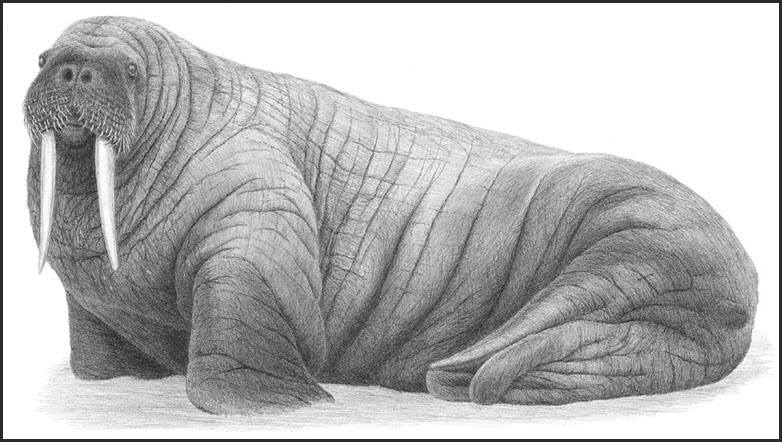
- Figure 1. Drawing of an adult male Atlantic Walrus (Odobenus rosmarus rosmarus).
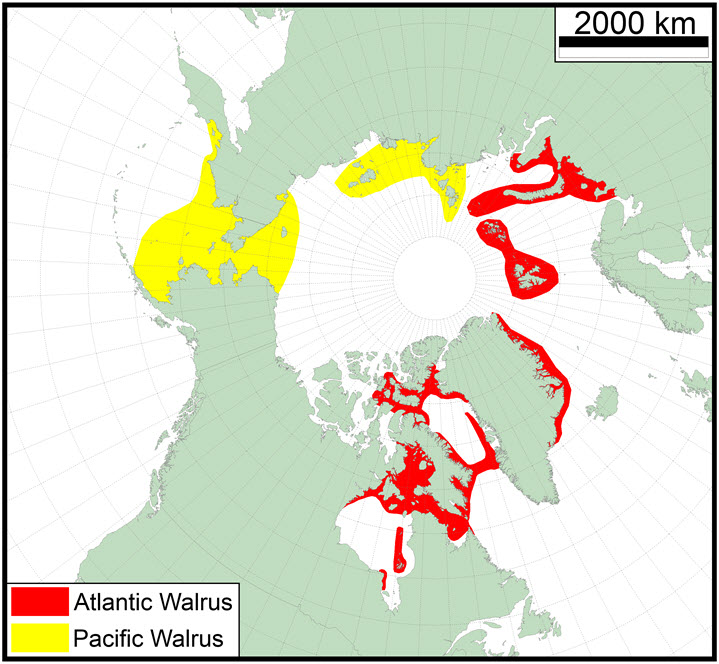
- Figure 2. Approximate present distribution of Atlantic (red polygons) and Pacific (yellow polygons) Walrus (based on Gjertz and Wiig 1994; Born et al. 1995; Witting and Born 2005; COSEWIC 2006; IUCN 2008; Stewart 2008; Boltunov et al. 2010; Garlich-Milleret al. 2011; LGL Limited and North/South Consultants Inc. 2011; Lydersen et al. 2012; Elliot et al. 2013; Heide-Jørgensenet al. 2013, 2014; Andersen et al. 2014; Dietz et al. 2014; R.E.A. Stewart et al. 2014c; Hammill et al. 2016a, b).
- Figure 3. Eastern Arctic place names used in text.
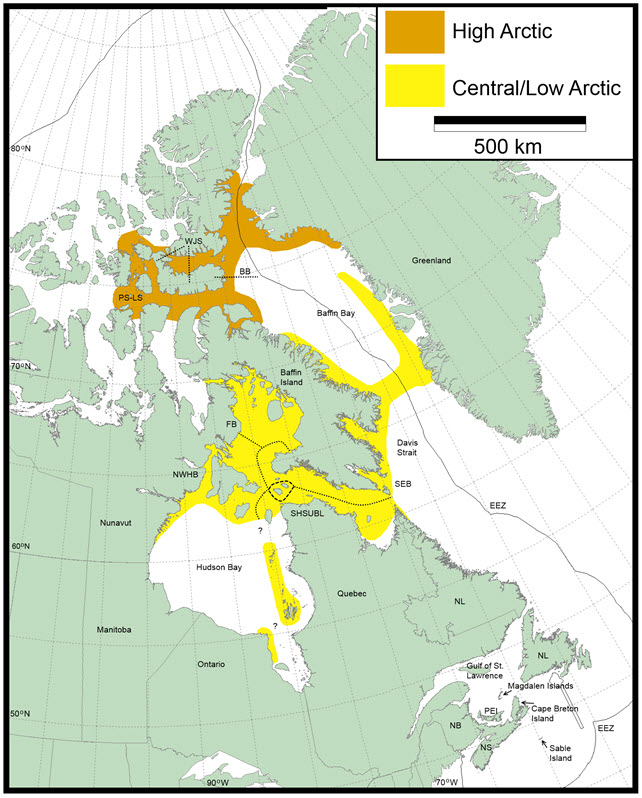
- Figure 4. Approximate distributions of the two extant Atlantic Walrus DUs in Canada. Walrus management stocks within these populations include: BB = Baffin Bay, FB = Foxe Basin, NWHB = North and West Hudson Bay, PS-LS = Penny Strait-Lancaster Sound, SEB = South and East Baffin, SHSUBL = South Hudson Strait-Ungava Bay-Labrador, and WJS = western Jones Sound. Question marks (?) indicate uncertainty with respect to distributions and/or movements. Nottingham and Salisbury islands are included with the SEB stock for population survey estimates but Walrus have historically been hunted at these islands by Inuit from both Baffin Island and Nunavik.
- Figure 5. Walrus hauled out at a rocky uglit in northern Foxe Basin on 28 August 2010 (Photo credit: R.E.A. Stewart, DFO). This was one of several islets in the area that were fully occupied by Walrus.
- Figure 6. Annual landed catches of Walrus from populations in the eastern Canadian Arctic by subsistence and sport hunters, 1980 to 2015 (see Table 2).
List of tables
- Table 1. Walrus population size estimates by population and management stock. Only recent estimates are provided, except for the NWHB management stock, where no recent surveys are available. Some estimates are considered negatively biased (see text), and most do not cover the entire range of the population/stock. Sources of past survey data are listed.
- Table 2. Annual landed catches reported from Atlantic Walrus populations in Canada, 1980-2015. Sources: Subsistence: D.B. Stewart et al. 2014a and references therein, DFO Iqaluit unpubl. data; Sport: DFO Iqaluit unpubl. data.
- Participants in the DFO (2012) Pre-COSEWIC Walrus meeting
List of appendices
- Appendix 1. Threats assessment worksheet for high Arctic population, Atlantic Walrus.
- Appendix 2. Threats assessment worksheet for central-low Arctic population, Atlantic Walrus.
Document information
COSEWIC Assessment and Status Report on the Atlantic Walrus Odobenus rosmarus rosmarus in Canada 2017
High Arctic population
Central-Low Arctic population
Nova Scotia-Newfoundland-Gulf of St. Lawrence population

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2017. COSEWIC assessment and status report on the Atlantic Walrus Odobenus rosmarus rosmarus, High Arctic population, Central-Low Arctic population and Nova Scotia-Newfoundland-Gulf of St. Lawrence populationin Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xxi + 89 pp. (Species at Risk Public Registry website).
Previous report(s):
COSEWIC. 2000. COSEWIC assessment and status report on the Atlantic walrus Odobenus rosmarus rosmarus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 24 pp.
Richard, P. 1987. COSEWIC status report on the Atlantic walrus Odobenus rosmarus rosmarus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-24 pp.
Production note:
COSEWIC acknowledges Arctic Biological Consultants (D. Bruce Stewart and Jeff W. Higdon) for writing the status report on Atlantic Walrus (High Arctic population, Central-Low Arctic population, and Nova Scotia-Newfoundland-Gulf of St. Lawrence population), Odobenus rosmarus rosmarus, in Canada, prepared with the financial support of Environment and Climate Change Canada. This report was overseen and edited by David Lee and Hal Whitehead, Co-Chairs of the COSEWIC Marine Mammals Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment and Climate Change Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
Website: COSEWIC
E-mail: COSEWIC
Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur le Morse de l'Atlantique (Odobenus rosmarus rosmarus), population du Haut-Arctique, population du centre de l’Arctique et du Bas-Arctique et population de la Nouvelle-Écosse, de Terre-Neuve et du golfe du Saint-Laurent, au Canada.
Cover illustration/photo:
Atlantic Walrus - Photo credit: J.W. Higdon.
COSEWIC - Assessment summary - Atlantic Walrus - High Arctic population
Assessment summary - April 2017
- Common name
- Atlantic Walrus - High Arctic population
- Scientific name
- Odobenus rosmarus rosmarus
- Status
- Special Concern
- Reason for designation
- This population numbers in the low thousands. It is not clear whether the aggregated hunting pressure in Canada and Greenland is sustainable under current management regimes. The population may become threatened if commercial shipping related to industrial development in the Arctic increases because this species is sensitive to human disturbance.
- Occurrence
- Nunavut, Arctic Ocean
- Status history
- The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate populations: Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Northwest Atlantic population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern. Split into three populations in April 2017. The High Arctic population was designated Special Concern in April 2017.
COSEWIC - Assessment summary - Atlantic Walrus - Central/low Arctic population
Assessment summary - April 2017
- Common name
- Atlantic Walrus - Central/Low Arctic population
- Scientific name
- Odobenus rosmarus rosmarus
- Status
- Special Concern
- Reason for designation
- This population appears fairly stable in its core areas of northern Hudson Bay and Foxe Basin, following historical declines. There is evidence for substantial population declines in southern and eastern Hudson Bay. These changes are likely a result of unsustainable hunting. While reported catches in Canada have been declining, there is concern that harvest may be underestimated. The population may become threatened if commercial shipping related to industrial development in the Arctic increases, particularly because the species is sensitive to human disturbance, and proposed routes run through core habitat (e.g. Foxe Basin).
- Occurrence
- Nunavut, Manitoba, Quebec, Newfoundland and Labrador, Arctic Ocean, Atlantic Ocean.
- Status history
- The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate populations: Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Northwest Atlantic population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern. Split into three populations in April 2017. The Central / Low Arctic population was designated Special Concern in April 2017.
COSEWIC - Assessment summary - Atlantic Walrus - Nova Scotia-Newfoundland-Gulf of St Lawrence population
Assessment summary - April 2017
- Common name
- Atlantic Walrus - Nova Scotia-Newfoundland-Gulf of St Lawrence population
- Scientific name
- Odobenus rosmarus rosmarus
- Status
- Extinct
- Reason for designation
- This population was hunted to extinction by the mid 1800’s. Sporadic recent sightings of individuals and small groups in the Gulf of St Lawrence and off Nova Scotia are not considered evidence of re-establishment, and there is no evidence of breeding in the region.
- Occurrence
- Quebec, New Brunswick, Prince Edward Island, Nova Scotia, Newfoundland and Labrador, Atlantic Ocean
- Status history
- The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate populations: Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Nova Scotia - Newfoundland - Gulf of St Lawrence population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern. Split into three populations in April 2017. The Nova Scotia - Newfoundland - Gulf of St Lawrence population was designated Extinct in April 2017.
COSEWIC - Executive summary
Atlantic Walrus
Odobenus rosmarus rosmarus
High Arctic population
Central-Low Arctic population
Nova Scotia-Newfoundland-Gulf of St. Lawrence population
Wildlife species description and significance
Walrus are large gregarious pinnipeds with upper canine teeth that grow into long tusks and a moustache of quill-like vibrissae. Newborns are ~120 cm long and 55 kg; males can grow to ~315 cm (~1100 kg) and females to ~277 cm (~800 kg). The Atlantic Walrus, Odobenus rosmarus rosmarus (Linnaeus, 1758) is one of two extant subspecies, the other being the Pacific Walrus (O. r. divergens). Atlantic Walrus traditionally provided important staples in the subsistence economy of the eastern Canadian Arctic and Greenland. The hunts are still of great social and cultural significance, and the economic value of the meat and ivory is substantial. The Walrus is important as the only living species in its family and a key link in the Arctic food web between bivalve molluscs and humans.
Distribution
The Atlantic Walrus ranged historically from the central Canadian Arctic east to the Kara Sea, north to Svalbard and south to Nova Scotia. There are three Walrus designatable units (DUs) in Canada: High Arctic, Central-Low Arctic and Nova Scotia-Newfoundland-Gulf of St. Lawrence (extinct). The High Arctic and Central-Low Arctic DUs are shared with Greenland. There is some evidence that the Central Arctic and Low Arctic animals form separate populations, but there is currently not enough information to support distinct DUs. The High Arctic and Central-Low Arctic DUs are distinguished by the degree of genetic interchange, geographical distribution, and movements. Walrus distribution appears to have contracted to areas that are less accessible to people and possibly in response to previous loss of ice cover.
Habitat
Atlantic Walrus occupy a large range but have a quite narrow ecological niche. They require large areas of shallow water (80 m or less) with bottom substrates that support a productive bivalve community, the reliable presence of open water over feeding areas, and suitable ice or land nearby upon which to haul out. Polynyas and other open-water areas are important during the winter.
Biology
Walrus haul out on ice and land, sometimes in large herds. They can travel long distances by swimming or by riding ice floes, but their seasonal movements are poorly understood. Walrus are polygynous and males compete intensely for females from February through April. Implantation of the embryo is delayed until late June or early July, and gestation is active for about 11 months. Most calves are born in late May and early June; they can suckle for up to 27 months and once weaned eat primarily bivalve molluscs. Females ovulate for the first time at 4 to 11 years and give birth to a single calf about every three years until reproductive senescence. The resulting birth rate is ~0.30 calves per fecund female per year and an annual gross production rate of ~10%. Generation time may be about 21 years and longevity over 35 years. Rates of mortality from human hunting and Polar Bear (Ursus maritimus) predation are unknown. Little is known of diseases of Walrus or of their response to pathogens.
Population sizes and trends
High Arctic DU1: occupies Penny Strait-Lancaster Sound (PS-LS), western Jones Sound (WJS), and Baffin Bay (BB). Aerial surveys in August 2009 yielded best estimates of about 727 Walrus in PS-LS, 503 in WJS, and 1,251 in BB (east coast of Ellesmere Island), for a total of 2,481. The PS-LS and WJS stocks seem stable over three decades. The main distributional change is in the Avanersuaq (Thule) area of west Greenland, where BB Walrus were once abundant in summer but are now absent.
Central-Low Arctic DU2: consists of Central Arctic subpopulations occupying Foxe Basin (FB), north and west Hudson Bay (NWHB), south and east Baffin (SEB), and southern Hudson Strait-Ungava Bay-Labrador (SHSUBL), and Low Arctic (LA) subpopulations occupying south and east Hudson Bay and northern James Bay. A September 2011 survey of FB haulouts (uglit in Inuktitut; singular ugli) estimated 10,379 Walrus. These estimates were much greater than previous estimates for this stock but there is no evidence for a trend. In September 2014 an aerial survey of uglit in NWHB yielded a corrected estimate of 5,500 Walrus. In summer 2005-2008, aerial surveys of uglit estimated 2,100-2,500 Walrus in the Hoare Bay area of SEB. There is no recent survey estimate from SHSUBL. An April 2012 survey estimated the population wintering in Hudson Strait at 6,020 Walrus, probably from various stocks (NWHB, SHSUBL, SEB). These Central Arctic stocks are likely reduced from their historical levels but no trend can be established and survey coverage is incomplete. Inuit have observed changes in Walrus distribution and seasonal availability. In September 2014 an aerial survey of the Low Arctic habitat yielded an estimate of 200 Walrus. Some past opportunistic counts in the Low Arctic were higher but existing data are insufficient to assess whether the population has recently declined. These suggest a minimum population size of 18,900 Walrus (including immature animals) in DU2.
Nova Scotia-Newfoundland-Gulf of St. Lawrence DU3: Extinct ca. 1850 by hunting, this population was once abundant in the southwestern Gulf of St. Lawrence and Scotian Shelf areas. Occasional recent sightings are not considered a sign of re-establishment, and there is no indication of breeding in the area.
In Canada, the total abundance of Atlantic Walrus is estimated at ca. 21,400. This estimate is negatively biased due to incomplete survey coverage and methodological issues that preclude trend assessment. The distribution of age classes is unknown.
Threats and limiting factors
Atlantic Walrus populations may be limited or threatened by hunting activities, noise disturbance, and industrial development activities. Their narrow ecological niche and restricted seasonal distribution make Walrus relatively easy for hunters to locate and vulnerable to environmental changes. Hunting is the cause of most known mortality and probably the most consistent limiting factor. Stock size and structure, survival rate, sustainable catch rate and levels, and rate of removal for all Canadian stocks are unknown. However, Fisheries and Oceans Canada (DFO) has used the best available information to estimate total allowable removal (TAR) for some stocks. The reported landed catch has been declining despite an increasing Inuit population. In the current decade (2010-2015) Canadian hunters may have removed about 10 Walrus annually from the High Arctic DU, and 381 from the Central-Low Arctic DU (over half from FB). It is not known whether the decreasing landings are due to less hunt effort, lower hunter success, changes in rates of reporting or some combination of these factors. Rates of struck and lost are uncertain. Walrus that summer in the Canadian High Arctic and SEB waters may be hunted in Greenland waters. There is evidence that recent Greenland catches of about 86 Walrus annually from the High Arctic have not been sustainable, and Walrus have largely disappeared from their west Greenland habitat in summer. More information is needed on movement patterns between Canada and Greenland and total hunting mortality from both countries to inform management of these shared stocks.
Human disturbances may cause Walrus to stampede into the water, interfere with feeding, increase energy expenditures, mask communications, impair thermoregulation and increase stress levels. Prolonged or repeated disturbances may cause Walrus to abandon their uglit. Their ability to recolonize areas and to adapt to non-threatening disturbances is unknown. Threats posed to Walrus in Canada by industrial activities and tourism are currently low but expected to increase rapidly within the decade. Climatic changes may expose Walrus to greater hunting pressure and alter trophic dynamics. The effects of chemical contaminants are unknown but tissue levels are typically low, except for cadmium and lead from natural sources and organochlorines in animals that eat seals.
Protection, status, and ranks
Atlantic Walrus in Canada are afforded protection by regulations that manage hunting and trade in Walrus products (Fisheries Act, Marine Mammal Regulations SOR/93-56, 1993). Hunts in Nunavut and Nunavik are co-managed by the Nunavut Wildlife Management Board (NWMB) and Nunavik Marine Region Wildlife Board (NMRWB), under the applicable sections of their respective land claims agreements, with scientific advice from DFO, which manages Walrus in other jurisdictions in cooperation with other agencies. Community knowledge and Aboriginal Traditional Knowledge (ATK) are also considered in the management of Walrus. Four Nunavut communities have catch quotas. Elsewhere, “an Indian or Inuk other than a beneficiary” can kill up to four Walrus per year without a licence; other hunters require a licence. Trade in edible parts is prohibited, except among First Nations and Inuit. A DFO permit is required to transport Walrus parts within Canada, except for First Nations or Inuit hunters who are returning home after the hunt. A Scientific Research Licence from DFO is required to do research in Walrus habitat and applicants must demonstrate community support. Live capture is permitted only under licence.
Existing National Parks, Wildlife Areas, and other federal lands afford little habitat protection for Walrus. The Nunavut Land Use Plan that is in preparation may protect Walrus haulouts in Nunavut by setting vessel approach setbacks of 5 km and prohibiting certain land uses, such as mineral extraction.
The Atlantic Walrus is listed on Appendix III of the Convention on International Trade in Endangered Species (CITES), which requires a CITES export permit to be issued before Walrus products can be exported from Canada. There is no formal cooperation between Canada and Greenland in the management of shared Atlantic Walrus populations.
Technical summary - High Arctic population
- Scientific name:
- Odobenus rosmarus rosmarus
- English name:
- Atlantic Walrus, High Arctic population / Population du Haut-Arctique
- Alternate name:
- Atlantic Walrus (English), Morse de l’Atlantique (French), Aivik (Inuktitut)
- Range of occurrence in Canada:
- Nunavut, Arctic Ocean
Demographic information
| Summary items | Information |
|---|---|
| Generation time [(age at first reproduction + age at last reproduction) / 2] | 21 years [i.e., (7 y + ~35 y) / 2] |
| Is there an observed continuing decline in number of mature individuals? | No. |
| Estimated percent of continuing decline in total number of mature individuals within 2 generations? | The Penny Strait-Lancaster Sound and western Jones Sound stocks appear stable. Unknown for the Baffin Bay stock. |
| Observed percent reduction or increase in total number of mature individuals over the last 3 generations? | Unknown |
| Projected percent reduction or increase in total number of mature individuals over the next 3 generations | Unknown |
| Observed percent reduction or increase in total number of mature individuals over any 3 generations period, over a time period including both the past and the future. | Unknown |
| Are the causes of the decline clearly reversible and understood and ceased? | Not applicable |
| Are there extreme fluctuations in number of mature individuals? | No |
Extent and occupancy information
| Summary items | Information |
|---|---|
| Estimated extent of occurrence (calculated as the area of a minimum convex polygon around the range polygon for the species distribution in Canada, with land area removed, using a Canada Albers Equal-Area projection (ArcView 3.3), and excluding the range in Greenland) | 415 457 km2 |
| Index of area of occupancy (IAO) (calculated as the number of 2 km x 2 km grid cells using the same projection and software, and including the range in Greenland) | 245,720 km2 |
| Is the population severely fragmented? | No |
| Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term. |
Not applicable |
| Is there an observed, inferred, or projected continuing decline in extent of occurrence? | Walrus were once abundant in summer in the Avanersuaq (Thule) area of Greenland, but are now absent in summer. Changes may occur in response to shipping and climate change but the direction of change is unknown and may vary seasonally and geographically. |
| Is there an observed, inferred, or projected continuing decline in index of area of occupancy? | See above. |
| Is there an observed, inferred, or projected continuing decline in Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term. |
No |
| Is there an observed, inferred, or projected continuing decline in Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term. |
Unknown |
| Is there an observed, inferred, or projected continuing decline in area, extent and/or quality of habitat? | Yes. Declines in seasonal ice habitat have occurred over the past decade. Projected increases in open-water shipping may reduce habitat quality in the Pond Inlet-Milne Inlet area over the next decade. |
| Are there extreme fluctuations in number of populations? | No |
| Are there extreme fluctuations in number of locations? | No |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | No |
Number of mature individuals
| Summary items | Information |
|---|---|
| Population | ~2,500; known to be negatively biased (minimum estimate); made up of ~727 in Penny Strait-Lancaster Sound, ~503 in western Jones Sound, and ~1,251 in Baffin Bay, along the east coast of Ellesmere Island |
| N Mature Individuals | No complete comprehensive population survey has ever been undertaken, and survivorship is also unknown so the number of mature animals is unknown. |
| Total | ~2,500 |
Quantitative analysis
| Summary items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. | Not available, suitable data for quantitative analysis lacking. |
Threats (direct, from highest impact to least, as per IUCN threats calculator)
| Summary items | Information |
|---|---|
| Was a threats calculator completed for this species? YES, 24 January 2017 (Appendix 1) |
Hunting : The population appears able to sustain current Canadian removal rates but is also hunted in Greenland waters, where hunts may not be sustainable. Harvests may not be evenly partitioned among the stocks, and stock discreteness is uncertain. Disturbance : Walrus are sensitive to noise and habitat disturbances. Human activities on or near occupied uglit (haulout sites) can stampede herds causing mortality. Repeated disturbances can cause habitat abandonment. The effects of long-term exposure to tourism and open-water shipping are unknown but may be serious. Climate change : Walrus haul out on sea ice for much of the year and this habitat has been changing rapidly. The effects of climate change on Walrus populations are difficult to predict. Climate amelioration that increases interactions with humans may impact Walrus populations more than other environmental factors. |
Rescue effect
| Summary items | Information |
|---|---|
| Status of outside population(s)? | Animals summering in West Greenland have nearly disappeared |
| Is immigration known or possible? | Immigration is possible |
| Would immigrants be adapted to survive in Canada? | Yes. |
| Is there sufficient habitat for immigrants in Canada? | Yes. But the reason Walrus disappeared from an area may determine whether it is reoccupied. |
| Is rescue from outside populations likely? | Unlikely as there are few animals left in West Greenland |
Data sensitive species
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
Status history
| Summary items | Information |
|---|---|
| COSEWIC: The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate populations: | Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Northwest Atlantic population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern. Split into three populations in April 2017. The High Arctic population was designated Special Concern in April 2017. |
High Arctic Population - Status and reasons for designation:
| Summary items | Information |
|---|---|
| Status: | Special Concern |
| Alpha-numeric code | Not applicable |
| Reasons for designation | This population numbers in the low thousands. It is not clear whether the aggregated hunting pressure in Canada and Greenland is sustainable under current management regimes. The population may become threatened if commercial shipping related to industrial development in the Arctic increases because this species is sensitive to human disturbance. |
Applicability of criteria
| Summary items | Information |
|---|---|
| Criterion A (Decline in Total Number of Mature Individuals) | Not applicable. No evidence for decline. |
| Criterion B (Small Distribution Range and Decline or Fluctuation) | Not applicable. EOO and IAO exceed thresholds. |
| Criterion C (Small and Declining Number of Mature Individuals) | Not applicable, although near to qualifying for Threatened under C2a(i) because: the population size is less than 10,000 mature individuals; hunting pressure may cause a continuing decline; and the largest of the three subpopulations is slightly higher than 1,000. |
| Criterion D (Very Small or Restricted Population) | Not applicable. Population exceeds thresholds. |
| Criterion E (Quantitative Analysis) | No applicable analysis. |
Technical summary - Central-low Arctic population
- Scientific name:
- Odobenus rosmarus rosmarus
- English name:
- Atlantic Walrus, Central-Low Arctic population / Population du centre de l’Arctique et du Bas-Arctique
- Alternate name:
- Atlantic Walrus (English), Morse de l’Atlantique (French), Aivik (Inuktitut)
- Range of occurrence in Canada:
- Newfoundland and Labrador, Manitoba, Nunavut, and Quebec; Arctic Ocean and Atlantic Ocean (Labrador Sea)
Demographic information
| Summary items | Information |
|---|---|
| Generation time [(age at first reproduction + age at last reproduction) / 2] | 21 years [i.e., (7 y + ~35 y) / 2] |
| Is there an observed continuing decline in number of mature individuals? | No. |
| Estimated percent of continuing decline in total number of mature individuals within 2 generations? | No population trends are known but the Foxe Basin stock is substantially larger than was previously observed or estimated. |
| Observed percent reduction or increase in total number of mature individuals over the last 3 generations? | Unknown |
| Projected percent reduction or increase in total number of mature individuals over the next 3 generations | Unknown |
| Observed percent reduction or increase in total number of mature individuals over any 3 generations period, over a time period including both the past and the future. | Unknown |
| Are the causes of the decline clearly reversible and understood and ceased? | No |
| Are there extreme fluctuations in number of mature individuals? | No |
Extent and occupancy information
| Summary items | Information |
|---|---|
| Estimated extent of occurrence (calculated as the area of a minimum convex polygon around the range polygon for the species’ distribution in Canada, with land area removed, using a Canada Albers Equal-Area projection (ArcView 3.3), and excluding the range in Greenland) | 1 759 137 km2 |
| Index of area of occupancy (IAO) (calculated as the number of 2 km x 2 km grid cells using the same projection and software, and including the range in Greenland) | 778,288 km2 |
| Is the population severely fragmented? | No |
| Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term. |
Not applicable |
| Is there an observed, inferred, or projected continuing decline in extent of occurrence? | Walrus are no longer reported from areas of eastern James Bay and western Baffin Bay, as well as summer habitat off west Greenland that they used in the historical past. No trends in extent of occurrence can be established over the past 2 generations from the data available. Changes may occur in response to shipping and climate change but the direction of change is unknown and may vary seasonally and geographically. |
| Is there an observed, inferred, or projected continuing decline in index of area of occupancy? | See above. |
| Is there an observed, inferred, or projected continuing decline in number of populations? | No |
| Is there an observed, inferred, or projected continuing decline in Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term. |
Unknown |
| Is there an observed, inferred, or projected continuing decline in area, extent and/or quality of habitat? | Yes. Declines in seasonal ice habitat have occurred over the past decade. Projected increases in year-round shipping may reduce habitat quality in Hudson Strait and Foxe Basin over the next decade. |
| Are there extreme fluctuations in number of populations? | No |
| Are there extreme fluctuations in Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term.? |
No |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | No |
Number of mature individuals (in each population)
| Summary items | Information |
|---|---|
| Population | Surveys suggest a minimum of ~18,900 in this DU (including immature animals); made up of ~10,400 in Foxe Basin, ~6,000 in Hudson Strait in winter, ~2,300 in the Hoare Bay area, and ~200 in the Low Arctic. |
| N Mature Individuals | No complete comprehensive population survey has ever been undertaken, and survivorship is also unknown so the number of mature animals is unknown. |
| Total | ~18,900 |
Quantitative analysis
| Summary items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. | Not available, suitable data for quantitative analysis lacking. |
Threats (direct, from highest impact to least, as per IUCN threats calculator)
| Summary items | Information |
|---|---|
| Was a threats calculator completed for this species? YES, 24 January 2017 (Appendix 2) |
Hunting : The ability to sustain current hunting removal rates is uncertain. Some Walrus that summer in Canada winter in Greenlandic waters and are hunted in both jurisdictions. The partitioning of harvests among stocks is unknown. Hunting in Greenland may be unsustainable. Disturbance : Walrus are sensitive to noise and habitat disturbances. Human activities on or near occupied uglit (haulout sites) can stampede herds causing mortality. Repeated disturbances can cause habitat abandonment. The effects of long-term exposure to year-round shipping and other projected commercial developments in their habitat are unknown but may be serious. Climate change : Walrus haul out on sea ice for much of the year and this habitat has been changing rapidly. The effects of climate change on Walrus populations are difficult to predict. Climate amelioration that increases interactions with humans may impact Walrus populations more than other environmental factors. |
Rescue effect
| Summary items | Information |
|---|---|
| Status of outside population(s)? | Animals summering in West Greenland have nearly disappeared |
| Is immigration known or possible? | Immigration is possible |
| Would immigrants be adapted to survive in Canada? | Yes. |
| Is there sufficient habitat for immigrants in Canada? | Yes. But the reason Walrus disappeared from an area may determine whether it is reoccupied. |
| Is rescue from outside populations likely? | Unlikely as there are few animals left in West Greenland |
Data sensitive species
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
Status history
| Summary items | Information |
|---|---|
| COSEWIC: The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate population | Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Northwest Atlantic population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern. Split into three populations in April 2017. The Central-Low Arctic population was designated Special Concern in April 2017. |
Central/Low Arctic Population - Status and reasons for designation
| Summary items | Information |
|---|---|
| Status | Special Concern |
| Alpha-numeric code | Not applicable |
| Reasons for designation: | This population appears fairly stable in its core areas of northern Hudson Bay and Foxe Basin, following historical declines. There is evidence for substantial population declines in southern and eastern Hudson Bay. These changes are likely a result of unsustainable hunting. While reported catches in Canada have been declining, there is concern that harvest may be underestimated. The population may become threatened if commercial shipping related to industrial development in the Arctic increases, particularly because the species is sensitive to human disturbance, and proposed routes run through core habitat (e.g. Foxe Basin). |
Applicability of criteria
| Summary items | Information |
|---|---|
| Criterion A (Decline in Total Number of Mature Individuals) | Not applicable. No decline data. |
| Criterion B (Small Distribution Range and Decline or Fluctuation) | Not applicable. EOO and IAO exceed thresholds. |
| Criterion C (Small and Declining Number of Mature Individuals) | Not applicable. No evidence of decline and population exceeds thresholds. |
| Criterion D (Very Small or Restricted Population) | Not applicable. Population exceeds thresholds. |
| Criterion E (Quantitative Analysis) | No applicable analysis. |
Technical summary - Nova Scotia-Newfoundland-Gulf of St. Lawrence population
- Scientific name:
- Odobenus rosmarus rosmarus
- English name:
- Atlantic Walrus, Nova Scotia-Newfoundland-Gulf of St. Lawrence Population / Population de la Nouvelle-Écosse, de Terre-Neuve et du golfe du Saint-Laurent
- Alternate name:
- Atlantic Walrus (English), Morse de l’Atlantique (French), Bastugobajijik (Mi'kmaw)
- Range of occurrence in Canada:
- former range New Brunswick, Newfoundland and Labrador, Nova Scotia, Prince Edward Island, and Quebec (southwestern Gulf of St. Lawrence and Scotian Shelf), Atlantic Ocean
Demographic information
| Summary items | Information |
|---|---|
| Generation time [(age at first reproduction + age at last reproduction) / 2] | 21 years [i.e., (7 y + ~35 y) / 2] |
| Is there an observed continuing decline in number of mature individuals? | No. This population has been extinct since the mid-1800s, so many of the questions below are not applicable |
| Estimated percent of continuing decline in total number of mature individuals within 2 generations? | Not applicable |
| Observed percent reduction or increase in total number of mature individuals over the last 3 generations? | Not applicable |
| Projected percent reduction or increase in total number of mature individuals over the next 3 generations | Not applicable |
| Observed percent reduction or increase in total number of mature individuals over any 3 generations period, over a time period including both the past and the future. | Unknown. |
| Are the causes of the decline clearly reversible and understood and ceased? | Commercial hunting, which was the primary cause of this Walrus population’s extinction, is no longer permitted. Since extinction the human population and associated activities have increased and would pose new threats to any Walrus in the region. |
| Are there extreme fluctuations in number of mature individuals? | Not applicable |
Extent and occupancy information
| Summary items | Information |
|---|---|
| Estimated extent of occurrence | Not applicable |
| Index of area of occupancy (IAO) | Not applicable |
| Is the population severely fragmented? | Not applicable |
| Number of “locations” See Definitions and Abbreviations on site Web du COSEPAC and IUCN (Feb 2014) for more information on this term. |
Not applicable |
| Is there an observed, inferred, or projected continuing decline in extent of occurrence? | Not applicable |
| Is there an observed, inferred, or projected continuing decline in index of area of occupancy? | Not applicable |
| Is there an observed, inferred, or projected continuing decline in number of populations? | Not applicable |
| Is there an observed, inferred, or projected continuing decline in number of locations? | Not applicable |
| Is there an observed, inferred, or projected continuing decline in area, extent and/or quality of habitat? | Yes. Human populations and activities in the area formerly occupied by this population are likely to continue increasing in the future, affecting the extent of suitable Walrus habitat. |
| Are there extreme fluctuations in number of populations? | Not applicable |
| Are there extreme fluctuations in number of locations? | Not applicable |
| Are there extreme fluctuations in extent of occurrence? | Not applicable |
| Are there extreme fluctuations in index of area of occupancy? | Not applicable |
Number of mature individuals (in each population)
| Summary items | Information |
|---|---|
| Population | Not applicable |
| N Mature Individuals | Not applicable |
| Total | blank |
Quantitative analysis
| Summary items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. | Not applicable. Already extinct. |
Threats
| Summary items | Information |
|---|---|
| This population is extinct so there are only rare vagrants in the area formerly frequented by this DU. Human activities will hinder the re-establishment of Walrus in the area formerly occupied by the Nova Scotia-Newfoundland-Gulf of St. Lawrence Population (DU3). | n/a |
Rescue effect
| Summary items | Information |
|---|---|
| Status of outside population(s)? | No members of DU alive anywhere |
| Is immigration known or possible? | No |
| Would immigrants be adapted to survive in Canada? | Not applicable. |
| Is there sufficient habitat for immigrants in Canada? | Unknown. The relationship between the species’ life history requirements and human populations and activities has not been studied. |
| Is rescue from outside populations likely? | No. |
Data sensitive species
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
Status history
| Summary items | Information |
|---|---|
| COSEWIC: The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate populations | Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Nova Scotia-Newfoundland-Gulf of St. Lawrence population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern. Split into three populations in April 2017. The Nova Scotia-Newfoundland-Gulf of St. Lawrence population was designated Extinct in April 2017. |
Nova Scotia-Newfoundland-Gulf of St. Lawrence population - Status and reasons for resignation:
| Summary items | Information |
|---|---|
| Status | Extinct |
| Alpha-numeric code | Not applicable |
| Reasons for designation | This population was hunted to extinction by the mid-1800s. Sporadic recent sightings of individuals and small groups in the Gulf of St. Lawrence and off Nova Scotia are not considered evidence of re-establishment, and there is no evidence of breeding in the region. |
Applicability of criteria
| Summary items | Information |
|---|---|
| Criterion A (Decline in Total Number of Mature Individuals) | Not applicable. |
| Criterion B (Small Distribution Range and Decline or Fluctuation) | Not applicable. |
| Criterion C (Small and Declining Number of Mature Individuals) | Not applicable. |
| Criterion D (Very Small or Restricted Population) | Not applicable. |
| Criterion E (Quantitative Analysis) | Not applicable. |
Preface
Since the last COSEWIC assessment in 2006, genetic (Andersen et al. 2009, 2014; Shafer et al. 2014) and satellite-telemetry (Stewart 2008; Dietz et al. 2014) data have been gathered that support differentiation of the High Arctic and Central-Low Arctic populations, and confirm movement of individuals from West Greenland to southeastern Baffin Island and from northwest Greenland well into the Canadian Arctic archipelago (NAMMCO 2015). Genetic markers also revealed a sex bias in favour of male dispersal and female philopatry (Andersen et al. 2014). A recent study by McLeod et al. (2014) suggests that Walrus from the population that was extirpated from the Canadian Maritimes were a morphologically and genetically distinctive group that was on a different evolutionary path from other Walrus found in the North Atlantic.
Recent Walrus population data are available from surveys of uglit (haulout sites; singular ugli) in Jones Sound and Lancaster Sound-Penny Strait (R.E.A. Stewart et al. 2014a), northern Foxe Basin (Stewart et al. 2013; Hammill et al. 2016a), southeastern Baffin Island (R.E.A. Stewart et al. 2014c), Hudson Bay-Davis Strait and south and east Hudson Bay (Hammill et al. 2016b); surveys of coastal waters and ice edges in western Nares Strait (R.E.A. Stewart et al. 2014b); and transect surveys conducted in West Greenland waters (Heide-Jørgensen et al. 2013, 2014). Satellite tagging studies coupled with these surveys provide new data on movements and haulout behaviour that can be used to adjust the survey estimates.
Walrus are more common and widely distributed in central and southern Foxe Basin than was previously known (LGL Limited and North/South Consultants Inc. 2011). In at least some summers they may be distributed almost continuously from northern Foxe Basin to Hudson Strait.
A catch history for Atlantic Walrus in the eastern Canadian Arctic has been compiled by hunt type, biological population and management stock (D.B. Stewart et al. 2014a). This compilation provides a starting point for modelling historical populations to inform recovery and management. Aboriginal Traditional Knowledge (ATK) (or Inuit Qaujimajatuqangit, traditional ecological knowledge, or local ecological knowledge in Walrus range jurisdictions) of Atlantic Walrus has been compiled and assessed for COSEWIC (Goulet unpubl.), and gathered to support climate change research on Walrus (Martinez-Levasseur et al. 2016).
Modelling studies have been conducted to gain insight into haulout behaviour (Doniol-Valcroze et al. 2016), population dynamics (Hammill et al. 2016c), abundance estimates and total allowable removals (DFO 2016; Hammill et al. 2016a, b). An integrated fisheries management plan has been completed for Walrus in the Nunavut Settlement Area and will be implemented in 2017 (A. McPhee, DFO Winnipeg, pers. comm. 2017), and DFO Science is developing a 5-year research plan for Walrus (Stewart et al. in press). Threats and effects pathways related to non-renewable resource developments and shipping on Atlantic Walrus have been reviewed (Kasser and Weidmer 2012; Stewart et al. 2012; D.B. Stewart et al. 2014b), as has international trade and management (Shadbolt et al. 2014; Wiig et al. 2014).
COSEWIC history
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2016)
- Wildlife species
- A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
-
Special concern (SC)
(Note: Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.) - A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
-
Not at risk (NAR)
(Note: Formerly described as “Not in any category”, or “No designation required.”) - A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
-
Data deficient (DD)
(Note: Formerly described as “Indeterminate” from 1994 to 1999 or “ISIBD” [insufficient scientific information on which to base a designation] prior to 1994. Definition of the [DD] category revised in 2006.) - A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment and Climate Change Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife species description and significance
Name and classification
The Atlantic Walrus, Odobenus rosmarus rosmarus (Linnaeus, 1758), is one of two living subspecies, the other being the Pacific Walrus (O. r. divergens) (Lindqvist et al. 2009) (Figure 1 and Figure 2). These two subspecies may represent the product of range fragmentation and differentiation during a glacial phase of extensive Arctic sea ice (Harrington 2008). Common names for the species include Walrus (English, Dutch), morse (French), aivik/aiviq (Inuktitut), bastugobajijik (Mi’kmaw), hvalros (Danish), mursu (Finnish), rostungur (Icelandic), hvalross (Norwegian), morzh (Russian), morsa (Spanish, Portuguese), and valross (Swedish).
Long description for Figure 1
Illustration of an adult male Atlantic Walrus. Adults of both sexes have tusks (long upper canines), but those of the male are longer and broader. The neck of the adult male is large and muscular and covered by thick, cornified skin.
Long description for Figure 2
Map outlining the approximate present distribution of the Atlantic Walrus and the Pacific Walrus. Atlantic Walrus range from the central Canadian Arctic in the west to the Kara Sea in the east and south to Nova Scotia. Pacific Walrus are found in the Bering, Chukchi, Laptev, and East Siberian seas, with occasional vagrants in western Canadian Arctic waters.
Morphological description
The Walrus is a large marine mammal with front and hind limbs that have developed into flippers (Figure 1). The front flippers can support the animal in an upright position while the back flippers are structured and function like the hind flippers of otariid seals. Adults of both sexes have tusks, which are long upper canines, and a moustache of quill-like whiskers. Adult males are larger than females and have longer, broader tusks (Mansfield 1966; Garlich-Miller and Stewart 1998). Newborn Walrus have a coat of silver grey hairs that is soon lost and replaced by a short, sparse coat of brown hair.
The sparsely haired skin is cinnamon brown but can appear pink on a warm day, or almost white after a long, cold dive. Adult males have a large muscular neck that is covered by thick, cornified skin. At birth Atlantic Walrus are about 120 cm long and about 55 kg (Mansfield 1958); males grow to about 315 cm (~1100 kg) and females to about 277 cm (~800 kg) (Garlich-Miller and Stewart 1998).
Detailed biological reviews are available for the Walrus (e.g., Fay 1985; Kasser and Weidmer 2012) and for its Atlantic (Reeves 1978; Born et al. 1995; Stewart 2002) and Pacific subspecies (e.g., Fay 1982; Garlich-Miller et al. 2011), as is a bibliography current to January 1993 (Stewart 1993).
Population spatial structure and variability
Three designatable units of Atlantic Walrus are recognized in Canada on the basis of genetics and other indicators of separation. The two extant DUs are composed of different management stocks that have been identified to facilitate hunt management and are used to organize the information that follows. Born et al. (1995) provide a detailed discussion of the seasonal distribution of Atlantic Walrus in Canada.
High Arctic population (DU1)
This population is shared with Greenland (Figure 4), and was formerly referred to as the North Water (Baffin Bay-Eastern Canadian Arctic) population (Born et al. 1995). There are significant genetic differences between this population and other Walrus populations in Canada (DNA microsatellites; Shafer et al. 2014) and between it and the population in central West Greenland (DNA microsatellites and mtDNA; Andersen and Born 2000; Andersen et al. 2014). Some male-mediated genetic exchange may occur with the Walrus in central West Greenland but female-mediated gene flow has apparently been restricted. Observations of Walrus at Prince Patrick and Melville islands and near Taloyoak (Spence Bay) have been interpreted to represent extra-limital occurrences of Walrus from this population (Harrington 1966).
Geographical isolation of the High Arctic population is also supported by information on Walrus distribution and movements (Born et al. 1995). Walrus are sometimes found in the Gulf of Boothia south to Pelly (Brice-Bennett 1976) and Committee Bays and to Crown Prince Frederik Island (70°02'N, 86°50'W) (Loughrey 1959; Anders 1966; Nunavut Department of Environment 2010). These animals are believed to come from the north, either as strays (Loughrey 1959) or when ice does not break up in Barrow Strait (Riewe 1976). Scientists and Inuit consider regular movement of Walrus through Fury and Hecla Strait to be unlikely (Loughrey 1959; Mansfield 1959; Davis et al. 1980; Garlich-Miller cited in Stewart 2002).
There may be a gap in Walrus distribution along the east coast of Baffin Island between Clyde River and Pond Inlet (Born et al. 1995). In the 1970s, hunters from Clyde Inlet travelled north to Scott Inlet to hunt Walrus (Kemp 1976) and hunters from Pond Inlet travelled south to the Cape Macculloch area (Lands Directorate 1981). The intervening stretch of the east Baffin coast is remote from both communities, seldom visited in summer, and has not recently been surveyed.
Three management stocks have been tentatively identified within the High Arctic population on the basis of satellite-linked radio tagging studies, observed seasonal distribution, and lead isotope ratios (206Pb/207Pb and 208Pb/207Pb) in the teeth (Stewart 2008; NAMMCO 2011; see also Outridge et al. 2003). These stocks are located in western Jones Sound (WJS), Penny Strait-Lancaster Sound (PS-LS), and Baffin Bay (BB). The results of genetic studies show no clear differentiation between the WJS animals and the PS-LS animals, but a small difference was detected between the WJS and BB animals using microsatellites (FST = 0.02, p=0.03, Shafer et al. 2014; see also de March et al. 2002). Differences in lead isotope ratios are useful for stock discrimination as an indicator that Walrus harvested by a particular community are exploiting substantially different geological/geographical habitats (Outridge et al. 2003).
This proposed stock structure requires further investigation. In June 2015, the Greenland Institute of Natural Resources (GINR) attached satellite tags to 21 Walrus at Wolstenholme Fiord (NAMMCO 2015). These animals moved west across the North Water to the east coast of Ellesmere Island. Some then moved north along the east coast of Ellesmere Island, some went far west into Jones Sound and 3 Walrus went south of Devon Island into Lancaster Sound where they headed west to Cornwallis Island. These new tracking data confirm that the Baffin Bay subpopulation of Walrus extends farther west into the Canadian High Arctic than was previously known. This raises uncertainty with respect to stock structure in the High Arctic population and to hunt management. Further research is needed to establish the degree of genetic exchange and validity of current stock delineations.
Central-low Arctic population (DU2)
This population is shared with Greenland (Figure 4) (Richard and Campbell 1988; Born et al. 1995; Stewart 2002, 2008; Shafer et al. 2014). It is separated from the High Arctic population on the basis of apparent geographical distributions (Born et al. 1995) and genetic differences (Buchanan et al. 1998; de March et al. 2002; Andersen et al. 2014; Shafer et al. 2014). Walrus in West Greenland and at southeastern Baffin Island could not be distinguished from one another genetically (Andersen et al. 2014). Born et al. (1995) considered animals at Digges and Mansel islands to be part of the Central Arctic population.
The homogeneity of the Walrus population in the central Arctic remains unresolved. Geographical distributions, lead isotope ratios and growth patterns suggest that Walrus in northern Foxe Basin, central Foxe Basin, and northern Hudson Bay-Davis Strait constitute separate management stocks (Stewart 2008). However, genetic analyses have been unable to differentiate between the animals in these areas (de March et al. 2002; Shafer et al. 2014). Differences in the dental lead isotope ratios of Walrus landed by Akulivik and Coral Harbour suggest these Walrus inhabit different chemical environments than those taken from northern Foxe Basin (Outridge and Stewart 1999; Outridge et al. 2003). However, these lead isotope signatures suggest that some of the Hall Beach Walrus visit northeastern Hudson Bay (Outridge et al. 2003; Stewart et al. 2003). Animals sampled from Foxe Basin in the 1980s and 1990s were significantly larger than those sampled from northern Hudson Bay in the 1950s (Garlich-Miller and Stewart 1998). Inuit have observed differences in body size and tusk length between Nottingham Island (hunted by Akulivik) and Coats Island (hunted by Coral Harbour), and between the Chesterfield Inlet and Repulse Bay areas (Fleming and Newton 2003).
Walrus are widely distributed in the relatively shallow waters of Foxe Basin, where they live year-round (Figure 4) (Mansfield 1959; Loughrey 1959; Crowe 1969; Beaubier 1970; Brody 1976; Orr et al. 1986; Nunavut Department of Economic Development and Transportation 2008). Distance may not completely separate them from Walrus to the north and south but it must limit interchange (see above). Inuit elders recognize two groups of Walrus in Foxe Basin on the basis of differences in the animals’ size, colour, flavour, and distribution (DFO 2002). Differences in dental lead isotope ratios of Walrus landed by Igloolik and Hall Beach suggest that these communities take animals from different local stocks (Outridge et al. 2003; Stewart et al. 2003).
There is some north-south movement of Walrus in Foxe Basin but no evidence of concerted movement to or from Hudson Strait (Anderson and Garlich-Miller 1994). Walrus winter in both areas, so they presumably do not move en masse to seek wintering habitat. The species’ seasonal distribution in southeastern Foxe Basin is poorly known, although between ca. 1915 and 1940 many Walrus were landed in the Cape Dorchester area of Baffin Island (Reeves and Mitchell 1986; D.B. Stewart et al. 2014a).
Walrus move between central West Greenland and southeastern Baffin Island (Dietz et al. 2014), but the amount of genetic exchange is unknown. Differences in the patterns and levels of organochlorine contaminants in their blubber indicate that Walrus sampled in West Greenland and southeastern Baffin Island (Loks Land) feed in different areas and/or on different prey (Muir et al. 2000). There are significant genetic differences between the animals in central West Greenland and those in the Baffin Bay stock of the High Arctic population (DNA microsatellites and mtDNA; Andersen and Born 2000; Andersen et al. 2014). Some male-mediated genetic exchange may occur from central West Greenland to Baffin Bay but female-mediated gene flow has apparently been restricted.
For the purpose of later discussions on hunting, the central Arctic portion of DU2 will be discussed in terms of four management stocks that are useful for framing discussions in geographical and jurisdictional terms but may bear little relation to any genetic distances. These management stocks are: Foxe Basin (FB), North and West Hudson Bay (NWHB), South and East Baffin (SEB), and Southern Hudson Strait-Ungava Bay-Labrador (SHSUBL).
The Low Arctic portion of DU2 was formerly known as the South and East Hudson Bay population, and extends from the Ottawa Islands south to the Ekwan Point area of western James Bay (Figure 4). Lack of genetic, population abundance, and movement data from these animals is an important gap in knowledge of Canadian Walrus. If these animals are from a genetically distinct population it will be important to retain the genetically adaptive potential they represent as the most southerly remaining population in a period of climatic change, and they should be considered as a potential separate DU.
Walrus from the Low Arctic can be distinguished from those in Foxe Basin by differences in their organochlorine signatures (Muir et al. 1995), tissue metal concentrations (Wagemann and Stewart 1994), and dental lead isotope ratios (Outridge and Stewart 1999). Walrus in the Low Arctic population can also be distinguished from those in Hudson Strait by differences in their dental lead isotope ratios (Outridge and Stewart 1999; Outridge et al. 2003). On average, animals landed by the communities of Akulivik and Inukjuak inhabit geochemically different habitats over most of their lifetimes. People from Akulivik traditionally hunt Walrus at Nottingham Island in Hudson Strait whereas the people from Inukjuak hunt mostly at the Ottawa, King George, or Sleeper archipelagos (Olpinski 1993; Portnoff 1994; Reeves 1995; Brooke 1997). However, over the past decade (i.e., since these isotope studies were conducted) hunters from Inukjuak have been hunting at Nottingham and Salisbury islands as Walrus at Ottawa Islands are more likely to be infected with Trichinella (S. Oplinski, Makivik, pers. comm. 2014) so any future comparisons of samples must be done with care.
During traditional knowledge interviews in 2013-2014, several Inukjuak Elders reported that Walrus had been seen around the community more often in the previous 10 years than in the past (Trent University and Makivik Corporation 2015). They also believe that Walrus in the Sleeper Islands belong to a different stock than those at Nottingham and Salisbury islands in Hudson Strait.
The apparent historical decline in abundance of Walrus in the Low Arctic has not been accompanied by a similar decline in the Coats Island area, which suggests that immigration from Hudson Strait or northern Hudson Bay is limited (Born et al. 1995).
Within the Low Arctic population, the relationship between animals in the Sleeper and Belcher archipelagos and those to the south near Cape Henrietta Maria and inside James Bay is unknown.
Nova Scotia-Newfoundland-Gulf of St. Lawrence population (DU3)
This Walrus population was historically abundant in the southwestern Gulf of St. Lawrence and the Scotian Shelf areas that offered extensive shallow water habitat with flat sandy bottoms
and an abundance of shellfish (Allen 1880; Reeves 1978; Born et al. 1995). Their original distribution in the St. Lawrence extended as far up-river as Rivière-Ouelle. Many Walrus hauled out on the Seven Islands (Sept-Îles) and Magdalen Islands in Quebec (Shuldham 1775), Miscou Island in New Brunswick (Perley 1850), on the North Cape of Prince Edward Island (Stewart 1806), Cape Breton Island and Sable Island in Nova Scotia (Allen 1880), and on the Ramea Islands off the southern coast of Newfoundland. They were also present on Anticosti Island (Allen 1880). Cow Head in western Newfoundland is named after Walrus, and fossil remains (700-12,000 years old) have been found along the coast of Gros Morne National Park (D. Whitaker, Parks Canada, pers. comm. 2017).
Walrus were considered plentiful in the Gulf of St. Lawrence, at Cape Breton Island, and at Sable Island (Allen 1880). Shuldham (1775) estimated that 7,000 to 8,000 Walrus hauled out in spring at the Magdalen Islands and that on occasion 1500 or 1600 were taken in one hunt (see also Patterson 1891; Warburton 1903). Heavy exploitation throughout the 1600s and 1700s resulted in the extirpation of Walrus from the Sable Island area by the end of the 18th century (Gilpin 1869:126, Allen 1880). A substantial Walrus fishery existed at Sable Island at least until the mid-1600s (Allen 1930) and one ship’s crew reportedly took 1,500 there in 1591 (Allen 1880).
In northeastern New Brunswick, French settlers employed by the “Royal Company of Miscou” were killing up to 300 or 400 Walrus at a time in the 1600s in the Miscou Island area, for their skins, oil and tusks (Perley 1850; Hogan 1986). Walrus were reportedly common there during fall and winter until the late 1700s (Ganong 1904) but were extirpated prior to 1850 (Perley 1850). Walrus were killed in “considerable numbers” near the North Cape of Prince Edward Island between 1770 and 1775 but by the early 1800s they were scarce and seldom seen ashore (Stewart 1806), and by the late 1820s appear to have been extirpated from the island (Sobey 2007; see also Hogan 1986). Allen (1930) reported the only known catch from New England waters, an immature Walrus (with 5-6 inch tusks) taken in December 1734 at Plymouth, MA. However, there is evidence that Walrus were once present along the east coast of North America south to South Carolina prior to its discovery by Europeans (Allen 1880).
Walrus from this extinct population appear to represent a morphologically and genetically distinctive group that was on a different evolutionary path from other Walrus found in the North Atlantic (McLeod et al. 2014). The extinct Walrus appear to have been larger animals, with larger and more robust tusks, skulls and mandibles. Their mtDNA control region haplotypes were unique to the region and a greater average number of nucleotide differences were found between the regions than within either group (i.e., DUs1-2 vs DU3). The extinction of this population following extensive commercial hunting represents a reduction in the adaptive potential of Atlantic Walrus (McLeod et al. 2014).
Designatable units (DU)
In 2006, COSEWIC treated Walrus in the eastern Canadian Arctic and in the vicinity of Prince Edward Island-Nova Scotia-Newfoundland-Gulf of St. Lawrence as a single designatable unit (DU) (COSEWIC 2006). New information on the genetics, movements, and distributions of these Walrus supports the existence of at least two extant DUs in Canada, and one extinct DU.
As detailed in the previous section, analyses of 10 microsatellite loci clearly identified two groups of Walrus that correspond to the High Arctic (DU1) and Central Arctic (DU2) groups (Shafer et al. 2014). The degree of differentiation between the groups was relatively low (FST = 0.07) (see also Andersen and Born 2000). Andersen et al. (2009) examined 11 microsatellite loci in Walrus from throughout the North Atlantic, including East Greenland and Svalbard. Their results suggest that Walrus in the North Atlantic have a recent common ancestry, making FST values more representative of time since the split than of contemporary migration rates. Estimates of contemporary migration rates were also very low, suggesting minimal contemporary gene flow. Whether this is an artifact of fragmentation caused by human activities or a “natural” phenomenon is unknown. Satellite tagging of Walrus showed extensive movement within each DU but no movement between them (Dietz et al. 2014; Stewart 2008). There also appear to be natural disjunctions in the species’ distribution, east and west of Baffin Island, that isolate these populations from one another although there are no known barriers to movement.
The High Arctic and Central-Low Arctic populations meet two criteria for “discreteness” as they have evidence of genetic distinctiveness and there is a disjunction in their range. Their genetic differentiation is not deep but they meet several other criteria for “significance”. In particular, they persist in different ecological settings likely to give rise to local adaptations (Lancaster Sound marine region for DU1 and four other marine regions for DU2; Parks Canada Agency 2012). The two DUs inhabit marine regions that have different environmental conditions, particularly related to the relative availability of shallow coastal feeding habitats and the quality, extent, and duration of sea ice cover, which may have led to differences in the population’s adaptation (e.g., breeding, haulout, and foraging behaviours) and their responses to climate change.
Some data suggest that the Central-Low Arctic DU could be split further, primarily by separating a Low Arctic DU. However, genetic affiliations and seasonal movements of the Low Arctic population are unknown. Also, southeast Baffin Island Walrus might be divided from those in Hudson Strait as suggested by the genetic work of Andersen et al. (2009, 2014), but other studies found no differences (Shafer et al. 2014).
Special significance
Walrus are the only living representatives of the Family Odobenidae, which originated ca. 18 mya, and have anatomical and life history characteristics, such as long tusks and aquatic nursing, that are unique among the pinnipeds. They are an important link in the Arctic food web between benthic invertebrates and humans. The species has played an important role in Canadian culture (D.B. Stewart et al. 2014a). It has been hunted since prehistoric times by Indigenous cultures for subsistence. In the past, Inuit used Walrus ivory to construct harpoon foreshafts and heads, toggles and handles, and parts for dog harnesses, to shoe sledges, and to make protective edges on kayak paddles (see D.B. Stewart et al. 2014a and references therein). The thick skin was used to make tents, boats, oil containers, rope and for other purposes. From the earliest European settlement until ca. 1928, Walrus were also hunted, and sometimes killed in large numbers, by non-Indigenous Canadians. The products of these hunts were important for subsistence and in some areas supported thriving commercial enterprises (e.g., Stewart 1806; Allen 1880; Born et al. 1995; Sobey 2007; D.B. Stewart et al. 2014a; Shadbolt et al. 2014). Skins, oil, and ivory were exported in large quantities.
Walrus continue to be important to Inuit in both cultural and economic terms (e.g., Goulet unpubl.). Some families spend the summer at traditional hunting camps, thus helping to maintain aspects of an ancient culture. Such cultural value is difficult to measure in economic terms (Gustavson et al. 2008). Anderson and Garlich-Miller (1994) estimated the net economic value of products (i.e., meat, ivory, baculum (penis bone)) from the 1992 summer Walrus hunt to Igloolik and Hall Beach at $160,000 to $659,000. The lower figure did not consider the effects on Inuit health of substituting foods imported from the south for nutritious Walrus meat (see also Loring 1996). These studies did not assess the monetary returns accruing from the limited sport hunts that have taken place since 1995.
Inuit, and occasionally First Nations, now hunt Walrus mainly for their meat, which is eaten or fed to dogs, and for their ivory tusks, which are either sold intact or carved for sale (Freeman 1964; Schwartz 1976; Anderson and Garlich-Miller 1994; Born et al. 1995; Bennett and Rowley 2004; DFO 2013; D.B. Stewart et al. 2014a; see also Goulet unpubl.; Shadbolt et al. 2014). There has been less need for Walrus meat for dog food in the past half century with the increased use of snowmobiles. Walrus are killed and eaten on a seasonal basis depending upon availability, which varies among communities (Fleming and Newton 2003). The ivory tusks and the baculum become the property of the hunter who shot the Walrus but the meat is typically shared in the community. It may be boiled and eaten fresh, frozen for winter consumption, or aerobically fermented to make igunaq (e.g., Orr et al. 1986; Anderson and Garlich-Miller 1994; Paniaq 1998; Qamaniq 1999). Walrus products are occasionally sold at the country food market in Iqaluit, with elders having preferred access (Gatehouse 2012), and there is some inter-settlement trade in igunaq (Priest and Usher 2004:155; Aarluk Consulting Inc. 2005). Inuit consider molluscs in Walrus stomachs to be a delicacy (J.W. Higdon, pers. obs.). First Nations along the coasts of Hudson Bay and James Bay occasionally hunted Walrus in the past to feed dog teams and made rope from the tough hide but only ate Walrus when there was no other food (Fleming and Newton 2003).
Distribution
Global range
Walrus have a discontinuous circumpolar Arctic and sub-Arctic distribution (Reeves 1978; Brenton 1979; Fay 1981, 1985; Cronin et al. 1994; Lindqvist et al. 2009). The Pacific Walrus are found in the Bering, Chukchi, Laptev and East Siberian Seas, with occasional vagrants being seen in western Canadian Arctic waters. Atlantic Walrus range from the central Canadian Arctic in the west to the Kara Sea in the east and south to Nova Scotia (historically common, now very rare) (Figure 2). There are two well-separated populations within this range, one to the east of Greenland and the other to the west as there is no evidence that would suggest Walrus move around the southern tip of Greenland or across the northern coast of Greenland.
Canadian range
In Canada, Atlantic Walrus range from Bathurst and Prince of Wales islands eastward to Davis Strait and from James Bay north to Kane Basin (Figure 3 and Figure 4). There are occasional records of Walrus in the Canadian Arctic west of this area (Harrington 1966; Stewart and Burt 1994). Those north and east of Victoria Island have tentatively been considered Atlantic Walrus on the basis of limited taxonomic information; those to the south and west as Pacific Walrus. Walrus appear to be rare along the Manitoba coast. Walrus are rare nowadays south of the Hebron–Okak Bay (57°28'N, 62°20' W) area of the Labrador coast (Mercer 1967; Born et al. 1995). Lone animals, usually young males, have been observed every 2 or 3 years on the pack ice or landfast ice edge south to Nain, Labrador (Communities of Labrador et al. 2005:2; B. Sjare, pers. comm. 2013). Several recent sightings have occurred along the coast of Torngat Mountains National Park, in Saglek Bay and in Eclipse Channel (D. Whitaker, Parks Canada, pers. comm. 2017). Since 1993, there have also been 3 or 4 sightings of lone animals, usually young males, along the east and south coasts of Newfoundland (B. Sjare, pers. comm. 2013; see also CBC 2014), and a few in Nova Scotia (Kingsley 1998; Camus 2003; Richer 2003). A lone Walrus that appeared to be in poor health was present in Witless Bay, on the Avalon Peninsula of Newfoundland, for several days in summer 2015 (D. Whitaker, Parks Canada, pers. comm. 2017).

Long description for Figure 3
Map of the eastern Arctic showing place names used in the text of this status report.
Long description for Figure 4
Map outlining the approximate distributions of the two extant Atlantic Walrus designatable units in Canada: Central-Low Arctic and High Arctic. Also indicated are the Walrus management stocks within these populations: Baffin Bay, Foxe Basin, North and West Hudson Bay, Penny Strait-Lancaster Sound, South and East Baffin, South Hudson Strait-Ungava Bay-Labrador, and western Jones Sound.
Extent of occurrence and area of occupancy
Atlantic Walrus have an extent of occurrence (EOO) of 1,997,081 km2 in Canada and index of area of occupancy (IAO) of 256,002 grid cells, each 2 km by 2 km. These numbers have increased since the last status update (COSEWIC 2006), largely due to new survey coverage of southern and central Foxe Basin. The changes do not signify any increase in population size or distribution but rather are the result of improved documentation. The High Arctic and Central-Low Arctic DUs have EOO of ca. 415 457 km2 and ca. 1 759 137 km2, respectively; and IAO of 245,720 and 778,288 km2, respectively.
The EOO values were calculated as the area of a minimum convex polygon around the range polygon for the species as a whole, with land area removed, using a Canada Albers Equal-Area projection in ArcView 3.3 (ESRI Inc., Redlands, CA), and excluding the range in Greenland. The IAO was calculated as the number of 2 km x 2 km grid cells using the same projection and software, and including the range in Greenland.
Search effort
The distribution maps for Walrus in Canada were developed based on data from the references cited in the “Information sources” section. Many uncertainties remain regarding the animals’ offshore movements in Baffin Bay and Davis Strait and their seasonal presence in Foxe Basin south of South Spicer Island and in James Bay and much of Hudson Bay. Information on search effort to locate populations and estimate abundance is presented below under the headings “Dispersal and migration” and “Population sizes and trends.”
Habitat
Habitat requirements
Atlantic Walrus require large areas of shallow water (80 m or less) with productive bivalve communities, open water over these feeding areas, and suitable ice or land nearby upon which to haul out (Davis et al. 1980). For a marine mammal species, this is a relatively narrow ecological niche (Born et al. 1995).
Walrus often gather in large herds. They are associated with moving pack ice for much of the year. When ice is lacking in summer and fall, they tend to congregate and haul out on land in a few predictable sites (Mansfield 1973). Uglit are often situated on low, rocky shores with steep or shelving subtidal zones where animals have easy access to the water for feeding and quick escape from predators (Figure 5) (Mansfield 1959; Salter 1979a, b; Miller and Boness 1983; R.E.A. Stewart et al. 2013, 2014a, c; Trent University and Makivik Corporation 2015). The animals generally move to more sheltered areas when there are strong onshore winds and heavy seas (Mansfield 1959).

Long description for Figure 5
Aerial photo of Atlantic Walruses at a rocky haul-out in northern Foxe Basin
Habitat trends
Suitable Walrus habitat is decreasing as human activities expand. Hunting and noise disturbance caused by motorized transportation have caused herds to abandon uglit near communities in favour of less accessible islands and shores (Kopaq 1987; Born et al. 1995; Immaroitok 1996; Kupaaq 1996; Paniaq 2005). Whether these animals would eventually habituate to disturbance and reoccupy abandoned uglit if hunting ceased is unknown. Shipping-related disturbances of Walrus habitat are expected to increase substantially within the next decade (see Threats).
Post-glacial rebound is slowly raising existing uglit relative to sea level over much of the eastern Canadian Arctic. The rate of coastal emergence is likely to slow and in some areas may reverse in response to water level rise caused by climate warming (e.g., Tsuji et al. 2009). In southern Hudson Bay, near Cape Henrietta Maria, the rate of rebound is about 1.2 m per century (Webber et al. 1970). Some uglit that were once islands in the Winisk area have become part of the mainland, reducing their use by Walrus (Fleming and Newton 2003). Declining use of habitat in the Attawapiskat area has also been attributed to coastal changes, which could, however, be offset by the emergence of new shoals with consequent shifts in Walrus distributions.
Little is known of the seasonal use by Walrus of habitats in southern Foxe Basin, along the northeastern coast of Baffin Island, and in southeastern Hudson Bay-James Bay. These are important knowledge gaps that make it difficult to assess how changes in these habitats may affect Walrus populations. Historically the Cape Dorchester area of southern Foxe Basin supported many Walrus (D.B. Stewart et al. 2014a). Walrus are still present in southern Foxe Basin but their seasonal habitat use of the area, which is likely to become a major year-round shipping route, is poorly known (LGL Limited and North/South Consultants Inc. 2011; D.B. Stewart et al. 2014b). There is believed to be a gap in the species’ distribution along the northeastern coast of Baffin Island that contributes to separation of the High Arctic and Central Arctic populations. Changes in use of this habitat could have important ramifications for these populations. Habitat use by Walrus is poorly known in southeastern Hudson Bay and James Bay.
Biology
Life cycle and reproduction
Walrus are gregarious and polygynous, and mature males compete intensely for females (Le Boeuf 1986; Sjare and Stirling 1996; Fay 1981). Rather little is known about reproductive behaviour since mating occurs in the water in remote areas from February through April (Born 1990, 2003; Sjare and Stirling 1996). Detailed observations of breeding behaviour were made at a High-Arctic polynya surrounded by fast ice (Dundas Island - 76°05’N, 94°58’W; Sjare and Stirling 1996), where males competed for, and defended access to, females for up to five days. During the breeding season, males vocalize (sing) underwater to communicate dominance and attract females. A female was likely to mate with the male that was attending the herd when she came into estrus.
Sea ice stability may be an important determinant of breeding behaviour (Sjare and Stirling 1996). The mating system in fast-ice habitat apparently differs from what has been described for Pacific Walrus breeding in pack ice. In pack ice Pacific Walrus exhibit behaviour suggestive of a lek or lek-like mating system in which several mature males vocalize from small, defended territories (Fay et al. 1984). It is not known whether the behaviour of Atlantic Walrus breeding in pack ice is like that observed in Pacific Walrus.
Female Atlantic Walrus ovulate for the first time between 4 and 11 years of age (Garlich-Miller and Stewart 1999; Born 2001). Of 79 females from northern Foxe Basin, all aged 7 years or older had ovulated, but not all had become pregnant (Garlich-Miller and Stewart 1999). Age at first pregnancy ranged from 5 to 12 years. Some females in the North Water became pregnant at age 4 years, while others did not ovulate until they were 11 (Born 2001). The average age at first ovulation was about 6 (Born 2001). The average age of first reproduction by female Walrus cannot be calculated based on the data available but has been estimated at 7 years for the purpose of estimating generation time. Males mature between 7 and 13 years of age (Born 2003).
Walrus age is estimated from the growth layers in the cementum of the lower canines (Mansfield 1958; Garlich-Miller et al. 1993; Stewart et al. 1993). These layers correspond closely to known ages of captives (0-15 years; Fay 1982), but age validation studies have not been conducted on wild Atlantic Walrus. Over 35 cemental growth layers have been counted in wild Atlantic Walrus, suggesting that layering occurs throughout the life of an animal (Mansfield 1958). Loss of fetal dentine suggests that ages may be underestimated, so Walrus may live longer than 35 years and generation time may be underestimated (Stewart and Stewart 2005). Growth layers in the mandible are not reliable indicators of age in mature Walrus and underestimate the ages of males that are more than 19 years old and females over 9 years, probably due to resorption and slower bone growth (Garlich-Miller et al. 1993).
Newborns are ~ 120 cm long and ~ 55 kg, and both sexes grow quickly, reaching ~ 200 cm and ~ 340 kg by age two (Mansfield 1958, 1967). Male Walrus grow larger than females and geographical differences are apparent (Knutsen and Born 1994; Garlich-Miller and Stewart 1998). Adults males average ~ 305 cm in length and weigh ~ 900 kg, and adult females average ~ 260 cm in length and weight ~ 570 kg (Mansfield 1967). The largest male Walrus measured from Foxe Basin weighed about 1100 kg and the largest female about 800 kg (Garlich-Miller 1994).
Females give birth on average once every three years (Mansfield 1958; Garlich-Miller and Stewart 1999; Born 2001). Most mature females ovulate every second year (Born 1990; Garlich-Miller and Stewart 1999). Reported pregnancy rates among fecund females were 0.29 in Foxe Basin, 0.35 in northern Hudson Bay (Mansfield 1958), and 0.346 in the North Water (Born 2001). Garlich-Miller and Stewart (1999) found a pregnancy rate of 0.33 and a birth rate of 0.30 in northern Foxe Basin.
Female Walrus in NW Greenland are in estrous from at least mid-January (collection started 19 January, no December samples) until late June (Born 2001). Breeding in Canada is thought to occur about the same time (January-April; Stirling et al. 1983). Following mating and fertilization, implantation of the embryo is delayed until late June or early July (Born 1990, 2001; Garlich-Miller and Stewart 1999). Active gestation lasts about 11 months resulting in a mean birth date of 20 June or earlier in the North Water (Born 2001). However, Born (2001) noted that newborn calves were observed by scientists and hunters between 4 February and 11 November, and cited several other studies that showed an extended birth period. Most pregnancies involve a single foetus but twins have been reported (Inukshuk 1996; see also Fay et al. 1991). Mansfield (1973) estimated the gross annual production rate, or proportion of newborns in the population, at 11%. Recent counts at High Arctic uglit in August, not corrected for sex and age segregation among uglit, suggest a calf production rate of about 10% (Stewart 2002).
The generation time of Atlantic Walrus, or average age of parenthood, is uncertain due to gaps in knowledge of population demographics, survival rates, and length of the reproductive period. The relative numbers of adult females of a given age are unknown, as is the age of the oldest reproducing female. Lacking empirical data, COSEWIC (2006) followed the approach Pianka (1988) suggested to obtain a rough estimate, namely:
Generation time = (age of first reproduction + age at last reproduction) / 2
This yields a precautionary generation time of 21 y [i.e., (7 y + 35 y)/2]. This approach is similar to the third calculation method recommended by IUCN (2013), where:
Generation time = age of first reproduction + z (length of the reproductive period)
When z = 0.5 is used in the absence of empirical data on survivorship and the relative fecundity of young vs. old individuals in the population the results are the same. Lowry et al. (2008) argue that COSEWIC (2006) overestimated generation time as young are more common than old animals in the population and that senescence may occur. They suggested a generation time of 15 y, based on the average age of female Walrus landed by subsistence hunters in Alaska (Garlich-Miller et al. 2006). This approach may underestimate generation time. Older females may be underrepresented, as hunters tend to select for younger animals with more tender meat and older males with large tusks. The reproductive period may be longer, as ages are negatively biased (Garlich-Miller et al. 1993) and age at senescence has not been documented. Mature female Pacific Walrus also calve at 2-year intervals (Fay 1982) on average while Atlantic Walrus in Foxe Basin calve at 3-year intervals (Mansfield 1958; Garlich-Miller and Stewart 1999; Born 2001). Consequently the estimate for Pacific Walrus may not be transferrable to Atlantic Walrus and the value of 21 y has been retained until the data needed to calculate a more accurate generation time are available.
Young Walrus can be suckled for up to 25 or even 27 months (Fisher and Stewart 1997). Females take their calves to sea while they forage for food (Kovacs and Lavigne 1992); the young can be nursed in the water (Loughrey 1959; Miller and Boness 1983). Calves moult in their first summer and each summer thereafter (Mansfield 1958). Mothers and calves produce stereotyped vocalizations that enable individual identification, and mothers can recognize their calf’s vocalizations (Charrier et al. 2009). Females, and the herd as a whole, are intensely protective of the young, so calf survival is high relative to that of most other pinnipeds. Adoption may be widespread and important to Pacific Walrus (Fay 1982) but has not been reported in Atlantic Walrus.
Hunting by humans is the greatest known cause of mortality in most areas within Canada, as well as Greenland. Survival of the young is probably high, owing to the intense maternal care they receive, although they are vulnerable to trampling when a herd is stampeded. Mortality from predation is probably low, given the animals’ large size, aggressive defence and dangerous tusks. But competitive fighting during the breeding season may increase the natural mortality of males (Fay 1985). The mortality rates from pathogens and parasites are unknown.
Physiology
Walrus are well adapted to cold and ice. They reduce heat loss during extreme cold by constricting blood flow to their peripheral vascular system and vice versa in extreme warm weather (Ray and Fay 1968). Their thick skin (2–4 cm) and blubber (1-15 cm; Fay 1985) enable them to sleep on the ice at –31°C with a strong wind blowing (Bruemmer 1977). They huddle together and reduce their exposed skin surface by curling into a “foetal position” when it is cold. Walrus in the Quataq area show evidence of skin damage from solar radiation but local Inuit hunters and Elders had not observed a decrease in the condition of the skin of the Atlantic Walrus over the long term that could be linked to ozone loss or climate change (Martinez-Levasseur et al. 2016).
Behaviour
Walrus hauled out on the land spend most of their time resting, often lying in contact with one another (Figure 5) (Salter 1979a; Miller and Boness 1983). This inactivity enables them to maintain high, stable temperatures in their skin and appendages, which may be crucial during the moult, and possibly for the healing of wounds and the survival of young calves (Fay and Ray 1968; Ray and Fay 1968). When hauling out on sea ice in Nares Strait in July and August they show diurnal variation in their haulout pattern and are more apt to be hauled out during afternoon and evening (R.E.A. Stewart et al. 2014b). In August they spend about 33% of each day hauled out of the water, and about 18% of their time at the surface. The number of animals hauled out on land at a particular site can vary widely from day to day and year to year (Mansfield and St. Aubin 1991; Gaston and Ouellet 1997). While comparatively slow and awkward on land, Walrus are good swimmers. Their cruising speed seldom exceeds 6 or 8 km/h but they can accelerate to about 30 km/h for a short time when chased (Bruemmer 1977). Foraging trips can last 72 h between haulouts (Born et al. 2003).
Conflicts are common at uglit, where animals must gain and defend space to avoid being crowded (Miller 1975, 1976, 1982; Salter 1979a,b; Miller and Boness 1983), but less so in the water. Large body size and long tusks characterize dominant animals. Tusks are used in threat displays by both sexes and are important in fighting. Females with calves favour the central and seaward areas of the ugli, where the calves are better protected from Polar Bears (Ursus maritimus) (Miller 1982; Miller and Boness 1983). Males in mixed herds tend to occupy the inland sites. In the water, males tend to be separated from females with offspring, possibly owing to differences in food requirements and to time and energy budgets related to nursing (Miller and Boness 1983).
Conflicts observed on the haulout beaches in summer and fall are minor compared with the serious battles that take place in the breeding season (Sjare and Stirling 1996; B. Sjare, DFO, pers. comm. 2005). Most of the fights in the breeding season occur in the water and go undetected (there have been few studies of breeding behaviour). The proportion of males injured in the breeding season might not be large, but those actively breeding or struggling to establish themselves can sustain serious puncture wounds, gashes, loss of eyes and tusk breakage. In addition, breeding males lose a lot of weight through vocalizing almost continuously and not feeding during February, March and April.
Walrus use a wide variety of vocalizations both in and out of the water to communicate threats, submission, and distress and to maintain contact between females and calves (Miller and Boness 1983; Miller 1985; Stirling et al. 1987; Sjare and Stirling 1996; Sjare et al. 2003; Stirling and Thomas 2003; Charrier et al. 2009).
Sensitivity to disturbance at uglit
The level of response by Walrus on land to aircraft overflights depends upon the distance and altitude of approach (Salter 1979a). Walrus on Bathurst Island at Brooman Point (75°31’N, 97°24’W) raised their heads to locate the source of the disturbance when a Bell 206 helicopter was up to 8 km away, oriented toward the sea when it was within 1.3 km and sometimes escaped into the water immediately thereafter. Pacific Walrus at Round Island, Alaska, dispersed when a jet aircraft passed overhead at an altitude of about 9,000 m (Okonek et al. 2009), and when a plane flew within 800 m (Okonek et al. 2010).
The response to disturbance may affect population dynamics by causing stampedes, interfering with feeding and increasing energy expenditures-particularly on the part of calves-and by masking communications, impairing thermoregulation and increasing stress levels (Stewart et al. (ed.) 1993). At some ‘rocky’ summer haulout sites tusk breakage may be a problem if animals startle and stampede into the water (B. Sjare, DFO, pers. comm. 2005). Prolonged or repeated disturbances may cause Walrus to abandon uglit (Salter 1979a).
Young Walrus and those in poor condition are vulnerable to trampling if herds are stampeded onshore or offshore. These stampedes typically result in few deaths. However, in one incident at St. Lawrence Island in the Bering Sea, where at least 537 Pacific Walrus died, trampling may have been one cause of the mortality (Fay and Kelly 1980). Over the past decade, stampedes have contributed to numerous large-scale mortality events among Pacific Walrus (USFWS 2014). Similar events have not been reported for Atlantic Walrus, although stampedes do cause some mortality (Loughrey 1959).
Diving behaviour
Gjertz et al. (2001) used dive recorders to study the diving behaviour of 9 male Atlantic Walrus at Svalbard. On average the animals spent 56 h in the water followed by 20 h hauled out on land. They dove to a maximum depth of 67 m and stayed submerged up to 24 min. Foraging dives to a mean depth of 22.5 m lasted an average of 6 min. Satellite-linked data loggers indicated that adult male Walrus can dive to at least 250 m in both summer and winter (Born et al. 2005).
Ice entrapment
Information on the entrapment of Walrus in ice is scant. Walrus can break ice with their tusks to keep holes open and can climb out onto the ice using their tusks. Large male Pacific Walrus can break through ice up to 20 cm thick by ramming it from below with their heavy, dense skull (Bruemmer 1977; see also Riewe and Amsden 1979). Atlantic Walrus can travel over the ice for at least 6 km when they are frozen out (Calvert and Stirling 1990; see also Piugattuk 1986; Ijjangiaq 1990)-typically in a straight line regardless of obstacles (Freuchen 1921). They also can become stranded inland and die (Siakuluk 1996).
Dispersal and migration
Walrus can swim or ride ice floes long distances but their seasonal movements in the Canadian Arctic are poorly known. Tagged animals crossed Davis Strait over the shallowest and narrowest part (Dietz et al. 2014), although Walrus that cross from east Greenland to Svalbard travel at least 700 km and cross water over 2500 m deep (Born and Gjertz 1993). Annual seasonal site fidelity in both summer and winter seems to be strong, at least in northeast Greenland (Born et al. 2005) and Svalbard (Freitas et al. 2009). Walrus in these areas follow the same seasonal migration pattern regardless of annual variations in ice and temperature regimes. In Svalbard, males actively travel through areas of dense ice cover in winter, regardless of sea ice advances and retreats (Freitas et al. 2009). Walrus feeding on shallow banks do not seem to be easily displaced by moving pack ice (Jay et al. 2010; Dietz et al. 2014). Summer habitat use appears to be driven by feeding requirements and the availability of uglit or sea ice (Freitas et al. 2009).
High Arctic DU1
The substantial migrations of Walrus that Freuchen (1921) and Vibe (1950) described, northward in the spring along the west coast of Greenland and southward in the fall along the east coast of Baffin Island, no longer seem to occur (Born et al. 1995). However, some Walrus cross from Greenland to Ellesmere Island in the spring and presumably return in the fall. Recent tagging has followed the movement of some Walrus from northwest Greenland westward in the Canadian Arctic Archipelago to Cornwallis Island (NAMMCO 2015). In May 2009 Walrus were distributed in a belt across the southern part of the North Water from Greenland to Ellesmere Island at about 76°30’N, over both shallow and deep (>500 m) water (Heide-Jørgensen et al. 2013). During the open-water period Walrus are concentrated along the east coast of Ellesmere Island and are rare in the waters off northwest Greenland (R.E.A. Stewart et al. 2014b).
Walrus move west via Lancaster Sound into the Canadian Arctic archipelago as the ice edge recedes in spring (Greendale and Brousseau-Greendale 1976; Priest and Usher 2004; Stewart 2008). The main migration occurs from mid-June to mid-July, mostly along the north side of Lancaster Sound, although some animals move deep into Pond, Milne and Admiralty inlets (Schwartz 1982). Some enter bays and inlets along the south coast of Devon Island; others continue west into Barrow Strait, north into Wellington Channel, or south into Prince Regent Inlet (Read and Stephansson 1976; Riewe 1976; Davis et al. 1978). They move ashore as ice dissipates. Hunters from Resolute suggest that there is a concerted and brief eastward migration out of the region via Lancaster Sound in the fall (Stewart 2002). Some Penny Strait and Lancaster Sound Walrus share habitat. A Walrus tagged in August 1993 at Bathurst Island was killed in early June 1994 in Milne Inlet of Baffin Island, about 750 km by sea to the east (Stewart 2002). Another animal at the same Bathurst Island ugli in 1993 had wintered ~120 km to the northeast near Dundas Island (B. Sjare cited in Stewart 2002). There is also a westward movement of Walrus from Baffin Bay into Jones Sound in early August (Davis et al. 1978), and catch patterns suggest a return eastward movement past Grise Fiord in the fall (Stewart 2008).
Some Walrus from this population winter at the ice edges of eastern Jones Sound or Lancaster Sound, or in the North Water (Born et al. 1995). Others appear to winter within the archipelago in the Cardigan Strait–Fram Sound and Penny Strait–Queens Channel areas, in the Hell Gate and Dundas Island polynyas, and in other areas with small polynyas or rotten ice (Riewe 1976; Davis et al. 1978; Killian and Stirling 1978; Stirling et al. 1983; Sjare and Stirling 1996).
Central-low Arctic DU2
The seasonal movements of Walrus in Foxe Basin are apparently in response to changing ice conditions (Mansfield 1958; Loughrey 1959). Movements have been observed between summering areas around the islands in northern Foxe Basin-particularly the Spicers, and wintering areas along the floe edge that forms along the north side of Rowley Island and extends southward, parallel to the Melville Peninsula, to about 67°30’N (Loughrey 1959; Orr et al. 1986). There is also some north-south movement by Walrus in northern Foxe Basin (Anderson and Garlich-Miller 1994). Observations along a north-south transect through central Foxe Basin in April to October 2008 suggest that Walrus are more common in southcentral Foxe Basin in June and August than was previously known, and uncommon there earlier and later in the year (LGL Limited and North/South Consultants Inc. 2011: 134-138).
Analyses of lead isotope signatures in the teeth of male Walrus landed by Hall Beach residents (Stewart et al. 2003) support assertions by Degerbøl and Freuchen (1935) that some animals from this population move to Southampton Island and by Loughrey (1959) that some go to Hudson Strait; but do not indicate that these are seasonal movements. Indeed these animals may travel farther south to the Sleeper Islands where hunters from Inukjuak often kill Walrus. These movements do not necessarily indicate regular gene flow. Significant seasonal movements of Walrus through Fury and Hecla Strait are thought unlikely (Loughrey 1959; Mansfield 1959; Davis et al. 1980; Guinn and Stewart 1988; Garlich-Miller cited in Stewart 2002).
Where environmental conditions permit, some animals remain in northern Hudson Bay, Hudson Strait and Ungava Bay year-round, apparently moving inshore and offshore in response to changes in the ice. Others appear to undertake significant seasonal migrations. Evidence for the extent of these movements is circumstantial, as it is based on local observations. Whether the wintering and migratory animals represent different subpopulations is unknown (Stewart 2002).
Walrus occupy the north side of Chesterfield Inlet in the spring, are absent near the community in summer, and are present in the Chesterfield Inlet–Roes Welcome Sound area in winter (Brice-Bennett 1976; Fleming and Newton 2003). They occur in Wager Bay when ice is minimal, and Inuit indicate that they prefer areas with strong currents. Walrus are common in the Repulse Bay area but less so when the summer ice concentration remains high. Their presence also depends on the strength of the current, which varies each summer. When the current is stronger they sometimes approach within 60 km of Repulse Bay in the fall; they are sometimes seen at the floe edge in winter.
Walrus are present year-round in northern Hudson Bay and western Hudson Strait (Orr and Rebizant 1987; Elliot et al. 2013). Tagging studies in the mid-1950s at Bencas, Coats, and Southampton islands, using harpoon-head tags (147 tagged, 4 recaptured), revealed only local movements (Mansfield 1958; Loughrey 1959). However, hunters report seasonal movements in response to changing ice conditions (Orr and Rebizant 1987). Walrus occur off the floe edge along the south and east coasts of Southampton Island and west and southwest coasts of Foxe Peninsula in winter, favour the floating pack ice of Evans Strait and Hudson Strait in late spring and summer, and move ashore to uglit as pack ice dissipates. During a late winter survey of Hudson Strait (10 March to 2 April 2012), Walrus were seen more frequently along the coasts in relatively shallow water with areas of lighter ice composed of nilas and small ice pans (Elliot et al. 2013). Abundance peaked in water depths of 100 m, and in areas of open water with adjacent ice cover (<50% ice cover). In the fall Walrus are concentrated at or near uglit on Bencas, Walrus, Coats, Mill, Nottingham, and Salisbury islands and on western Foxe Peninsula (Orr and Rebizant 1987; Hammill et al. 2016b).
Inuit from Akulivik and Ivujivik have seen Walrus moving northward from Hudson Bay into Hudson Strait in the fall (Figure 4; Reeves 1995; Fleming and Newton 2003). Walrus remain in the Ivujivik area year-round but are seldom seen near Akulivik in summer (Fleming and Newton 2003). Salisbury Island (Akulliq) and “Pilik” Island, which does not appear on maps, are important sites for these animals. In the early 1990s, Ivujivik hunters would go to Salisbury Island when they did not see Walrus elsewhere in winter.
There is a general westward movement of Walrus through Ungava Bay and Hudson Strait in summer to Nottingham (Tutjaat) and Salisbury islands, with a return movement in the fall (Degerbøl and Freuchen 1935; Loughrey 1959; Trent University and Makivik Corporation 2015). Currie (1963) described an influx of Walrus to the southeast coast of Akpatok Island in Ungava Bay as soon as ice conditions permitted in June or early July, and their subsequent dispersal in late July or August northwest past Cape Hopes Advance into Hudson Strait, with a return migration following the same general route but further offshore the cape in September and October. Smith et al. (1979) observed a large influx of Walrus, apparently from Hudson Strait, into the Hall Peninsula area in mid-September. Some Walrus are present year-round near Nottingham and Salisbury Islands, where strong currents maintain polynyas through the winter (Kemp 1976; Orr and Rebizant 1987). Inuit hunters and elders in Nunavik report that Walrus in Hudson Strait are now migrating earlier (Trent University and Makivik Corporation 2015). In the 1990s Walrus were seen at Paul Island, near Nain, Labrador (Communities of Labrador et al. 2005:26). Several recent sightings have occurred along the coast of Torngat Mountains National Park (D. Whitaker, Parks Canada, pers. comm. 2017). “Lost” or dead and drifting Walrus are seen occasionally farther south in the Makkovik area-most recently ca. 2012 (M. Bishop, pers. comm. 2013).
Some Walrus that summer at southeastern Baffin Island winter off West Greenland (Dietz et al. 2014). Walrus are also present far offshore in the pack ice of Davis Strait (Vibe 1967; Born et al. 1995). The Walrus that haul out on pack ice off West Greenland in the winter do not use land haulouts in Greenland when the ice disappears, as they did historically. In 2005 to 2008, tagged Walrus (n=6) left Store Hellefiske Banke off West Greenland during the period 7 April to 25 May and took 7 days on average to swim an average distance of 338 km across Davis Strait to southeastern Baffin Island (Dietz et al. 2014).
There is no evidence for a concerted movement of Walrus into or out of southeastern Hudson Bay. Instead, there are local seasonal movements between the rocky sites where animals haul out during the ice-free period and their wintering areas (Freeman 1964). In both the Belcher and Sleeper archipelagos, Walrus are present at the floe edge in winter and move into the islands and onshore as the pack dissipates in summer (Fleming and Newton 2003; Hammill et al. 2016b; P. Kattuk, Mayor of Sanikiluaq and Z. Novalinga, Sanikiluaq Environmental Committee, pers. comm. 1993). The winter whereabouts of animals that summer along the Ontario coast are unknown, and the question of whether they move between this area and the Belchers is unanswered.
Lead isotope signatures in the teeth suggest that some males have moved between Foxe Basin and eastern Hudson Bay (Stewart et al. 2003), but these isotopic records spanned several years and do not necessarily indicate seasonal movements.
Nova Scotia-Newfoundland-Gulf of St. Lawrence DU3
No information has been found describing the historical patterns of movement of this population.
Interspecific interactions
Prey
Atlantic Walrus feed primarily on benthic invertebrates, especially bivalve molluscs (Degerbøl and Freuchen 1935; Vibe 1950; Mansfield 1958; Fisher 1989; Fisher and Stewart 1997). Foraging Walrus use their sensitive whiskers to identify suitable prey and expose them using a flipper or a jet of water from the mouth (Loughrey 1959; Fay 1981; Kastelein and van Gaalen 1988; Kastelein et al. 1990; Levermann et al. 2003). They forage mostly at depths of 10 to 80 m (Vibe 1950; Mansfield 1958; Born et al. 2003) and dives can last 24 min (Gjertz et al. 2001). Seasonal feeding patterns are not well known, although Walrus may feed more intensively in the fall. Males and females have similar diets but the females have a more efficient digestion (Fisher 1989; Fisher et al. 1992).
Walrus stomachs often contain only the feet or siphons of bivalves (Vibe 1950; Mansfield 1958; Fisher 1989; Welch and Martin-Bergmann 1990; Fisher and Stewart 1997), but direct observations in the wild show that they remove most of the soft parts of their bivalve prey (Vibe 1950; Born et al. 2003). These soft parts, and prey such as polychaetes, echiurids and sipunculids, are digested quickly so their dietary importance may be underestimated (Sheffield et al. 2001).
The estimated daily gross energy intake by an instrumented 1200 kg male Walrus off east Greenland (Born et al. 2003) was considerably higher than that reported from studies of captive Walrus (Fisher et al. 1992; Kastelein et al. 2000), suggesting that food requirements may be higher than that assumed by some fishery models applied to Walrus (Acquarone et al. 2006). Feeding rates in captivity-and probably also in the wild-vary with age, sex, reproductive status, and season (Kastelein et al. 2000).
Besides benthic invertebrates, Atlantic Walrus eat Ringed Seals (Pusa hispida), Bearded Seals (Erignathus barbatus), fishes and squids (Vibe 1950; Mansfield 1958; Hantzsch 1977:388). Inuit note that predation on seals is most prevalent in areas where deep water makes it harder for Walrus to reach the bottom (Gunn et al. 1988:24; Piugaattuk 1990; Kappianaq 1992, 1997). Observations on Pacific Walrus suggest that most seal-eating is predation rather than scavenging (Lowry and Fay 1984). Atlantic Walrus also prey on seabirds such as Thick-billed Murres (Uria lomvia) (Gjertz 1990; Donaldson et al. 1995; Mallory et al. 2004), and scavenge dead whales (Degerbøl and Freuchen 1935; Mansfield 1958). Some large bulls will eat young Walrus (Degerbøl and Freuchen 1935).
Competition
Bearded Seals and Pacific Walrus compete for clams in some areas (Lowry and Frost 1981); the same is likely true for Atlantic Walrus. The presence of Walrus tends to drive away Ringed Seals (Reeves 1995).
Predation
Polar Bears prey on Atlantic Walrus; with some of the bears dying in the attempt (Freuchen 1921; Loughrey 1959; Killian and Stirling 1978). Walrus are most vulnerable to predation when they are frozen out of their breathing holes or when they must rely on a very limited area of open water for breathing and haulout, particularly where rough ice provides bears cover for stalking (Calvert and Stirling 1990). Sub-adult Walrus are more vulnerable than adults, which are aggressive and possess large tusks for defence. Polar Bears in Foxe Basin derived more of their ingested biomass from Walrus (7% ± 1%) than did bears elsewhere in the Canadian Arctic (Thiemann et al. 2008). Walrus consumption was greatest among large adult male bears and increased with age for both sexes.
Killer Whales (Orcinus orca) prey upon Pacific Walrus (Lowry et al. 1987; Melnikov and Zagrebin 2005; Kryukovaa et al. 2012), but Inuit observations suggest that they do not prey on Atlantic Walrus in Canada or Greenland (Ferguson et al. 2012; Westdal et al. 2013).
Diseases and parasites
The susceptibility of Walrus to mortality from disease is not well understood. Serological testing of 210 Atlantic Walrus from sites in the eastern Canadian Arctic did not find antibodies to influenza A virus, which can cause high mortality of seals and has been found in Ringed Seals and Beluga Whales (Delphinapterus leucas) (Nielsen et al. 2001a). If Walrus are susceptible to this virus, the absence of seropositive animals could mean that the animals sampled had not been exposed to the virus or that all of the infected animals had died. There is serological evidence for sporadic infection of Walrus in the Igloolik area with Brucella sp., a bacterium that can cause reproductive failure (Nielsen et al. 1996, 2001b, 2004). Morbillivirus antibodies are common in Walrus from the eastern Canadian Arctic (Nielsen et al. 2000, 2004; Phillipa et al. 2004), indicating exposure to the phocine distemper virus or a related virus. Serological testing of Foxe Basin Walrus for pathogens found antibodies for canine adenovirus, canine distemper virus, dolphin morbillivirus, phocine distemper virus, and dolphin rhabdovirus (Phillipa et al. 2004). The pathology of all these viruses in Walrus is unknown.
Walrus are commonly infected by the helminth nematode Trichinella nativa Britov and Boev 1972, which causes trichinellosis (or trichinosis) in humans (Campbell 1988; Pozio et al. 1992; Serhir et al. 2001). This parasite was identified in earlier literature as T. spiralis (e.g., Brown et al. 1948, 1950; Fay 1960; Born et al. 1982; see also Manning 1961). Outbreaks of trichinosis due to eating uncooked (i.e., fresh, frozen, igunaq) or undercooked (i.e., not “well done”) Walrus meat are recurrent among Inuit in the eastern Canadian Arctic (Viallet et al. 1986; MacLean et al. 1992; Heinzig 1996; Serhir et al. 2001; Hill 2003; George 2008; Ndao 2011; CBC 2012; Larrat et al. 2012; Rogers 2013, 2015; S. Olpinski, Makivik pers. comm. 2014; Nunatsiaq News 2016). The pathology of this parasite in Walrus is unknown but its presence may have contributed to a decline in Walrus catches by affected communities in the 1980s and early 1990s (Olpinski 1991; Brooke 1992; Larrat et al. 2012). The prevalence of Trichinella in Walrus is an ongoing concern for hunters in Nunavut and Nunavik. The Pathology Laboratory at Makivik Corporation’s Nunavik Research Centre receives and tests Walrus tongue samples from communities for Trichinella.
Population sizes and trends
In the late 1800s, declining Bowhead Whale (Balaena mysticetus) abundance prompted commercial whalers and traders in the eastern Canadian Arctic to diversify and increase their hunting of Walrus and other species (D.B. Stewart et al. 2014a). Many Walrus were killed before commercial and sport hunting of the species was banned in Canada in 1928 (Canada 1928: P.C. 1036). Since then the species has been reserved for Indigenous use although a limited sport hunt was reopened in 1995. Walrus populations in the eastern Canadian Arctic (Richard and Campbell 1988; Born et al. 1995; D.B. Stewart et al. 2014a) and West Greenland (Witting and Born 2005, 2014) may still be much lower than they were prior to the large commercial hunts.
There has been a general shift in Walrus distribution away from human communities to areas that are relatively inaccessible (Kopaq 1987; Born et al. 1995; Kuppaq 1996; Immaroitok 1996; Paniaq 2005). This is not a new phenomenon and is related to changes in technology (Brody 1976). It began with the introduction of whaleboats in the 1920s, which extended hunting ranges and enabled open-water hunting; accelerated with the introduction of motorized technology ca. 1940-60; and continues as the range and speed of boats increase (see also Crowe 1969; Beaubier 1970; Orr et al. 1986). The extent to which distributional changes reflect declines as opposed to shifts in the Walrus populations is not always clear but in the absence of evidence of the latter it is prudent to assume numbers have been reduced (DFO 2002). An exception to the pattern of Walrus distributions shifting away from communities has been observed in the vicinity of Kangiqsujuaq, where Walrus are increasing in number and remain localized despite subsistence hunting (S. Olpinski, Makivik, pers. comm. 2014).
Inuit living around Hudson Bay have variously attributed the disappearance of Walrus from traditional hunting areas to natural shifts in the species’ distribution, poor and wasteful hunting techniques, and industrial development (Fleming and Newton 2003; DFO 2013). In the past, unregulated hunting from motorboats disturbed animals at uglit in the Belcher and Sleeper islands, and along the west coast of Hudson Bay. Walrus remains were sometimes discarded at the uglit and, together with sinking losses, tainted both the uglit and feeding grounds causing herds to leave the area. Inuit recognize the sensitivity of Walrus to habitat disturbance and to the mortality of other Walrus in their traditional knowledge.
“When I was growing up, I remember, my father and the others used to say never try to kill a Walrus where you think it will sink right into the feeding areas, or never cut up the Walrus where they usually bask or rest. The elders used to say never to leave the guts near the islands where they bask. If you do that the Walrus will move away from there.” (Zach Novalinga, Sanikiluaq in Fleming and Newton 2003:16).
Recent surveys have greatly improved knowledge of Walrus abundance in Canada but large gaps remain. Data on the number of Walrus present in an area often consist simply of counts of animals hauled out at known areas of concentration, or opportunistic sightings, and cannot be used to estimate the total Canadian Walrus population (Richard and Campbell 1988). Only the most recent abundance estimates correct for animals that are submerged beyond view or for haulout dynamics, which determine the proportion of a herd that is on land or ice or in the water at the time of the survey (R.E.A. Stewart et al. 2013, 2014a-c; Hammill et al. 2016a, b). Ugli occupancy varies widely within and among years, making even large changes in population size difficult to detect. Changes in population abundance, ice patterns, and human activity may all lead to the use of new uglit. Population estimates in general could be improved by obtaining broader survey coverage in a short period to reduce double-counting uncertainty. More detailed information is also required on the movement of Walrus between Greenland and Canada and the summer dispersal of these animals within Canada.
Available population estimates for extant Atlantic Walrus populations and stocks in Canada are summarized in Table 1 and detailed in Appendix 1 The total estimated abundance of Walrus in Canada from the studies in Table 1 is ca. 21,400 animals, with ca. 2,500 in the High Arctic population (DU1), ca. 18,900 in the Central-Low Arctic population (DU2), and zero in the Nova Scotia-Newfoundland-Gulf of St. Lawrence population (DU3). These estimates (except that for DU3) are negatively biased due to incomplete survey coverage and methodological differences and unknown stock composition on wintering grounds. The age-class distributions of the stocks are unknown.
| DU | Stock | Population estimate | Year(s) | Method | Source | Trend | Sources for past surveys |
|---|---|---|---|---|---|---|---|
| High Arctic DU | PS-LS | 727 (CV 0.07, 95% CI 623 - 831) |
Aug. 2009 | Haulout counts (minimum counted population (MCP)), adjusted for animals at sea. | Stewart et al. 2014a | Stable since late 1970s | Davis et al. 1978 |
| High Arctic DU | WJS | 503 (CV 0.07, 95% CI 473 - 534) |
Aug. 2008 | Haulout counts, MCP adjusted for animals at sea. | Stewart et al. 2014a | Stable since late 1970s | Davis et al. 1978 |
| High Arctic DU | BB | 1,251 (CV 1.00, 95% CI 1226) | Aug. 2009 | Haulout counts, MCP adjusted for activity using satellite-tagged animals. | Stewart et al. 2014b | Unknown | Finley and Renaud 1980; Richard et al. 1998; Witting and Born 2005 |
| High Arctic DU | blank | 1,759 (CV 0.29) | May 2010 | Aerial line-transect survey, corrected for perception and availability biases, includes Greenland waters. | Heide-Jørgensen et al. 2013 | Unknown | blank |
| Central-Low Arctic DU | NFB/CFB | 10,379 (CV 0.42)a | Sept. 2011 | Haulout counts, adjusted for activity using satellite-tagged animals; MCP = 6043, corrected estimates ranged from 8,152 (CV 0.02) to 13,452 (CV 0.43) (also see re-analyses by Hammill et al. 2016a). | Stewart et al. 2013 | Unknown, no evidence for decline | Orr et al. 1986; Richard 1993; Hammill et al. 2016b |
| Central-Low Arctic DU | NWHB | 5,500 (95% CI = 2,000-15,900) (rounded to nearest 100)a | Sept. 2014 | Haulout counts adjusted using several different methods, data reported for entire area surveyed – “Hudson Bay Davis Strait stock”: Simple Count (SC) = 2,144; MCP = 3,418; Bounded Count (BC) = 5,969. Portion reported for NWHB based on animals counted (78% of simple count) in that area. | Hammill et al. 2016b | Unknown | Loughrey 1959; Mansfield 1962; Mansfield and St. Aubin 1991; Richard 1993 |
| Central-Low Arctic DU | SEB | 1,500 (95% CI = 500-4,400) (rounded to nearest 100)a | Sept. 2014 | See NWHB above - for SEB area SC = 467, MCP = 582, ca. 22% of total SC for “Hudson Bay Davis Strait stock”. | Hammill et al. 2016b | Unknown | MacLaren Atlantic Limited 1978; MacLaren Marex Inc. 1979, 1980a+b; Smith et al. 1979; Richard 1993 |
| Central-Low Arctic DU | blank | 2,100 - 2,500 | Aug.-Sept. 2007 | Haulout counts in Hoare Bay area, MCP adjusted by proportion of tags ‘dry’ on survey morning (2,102, 95% CI = MCP - 4,482) and by percent time tagged animals spent hauled out on survey day (2,502, 95% CI 1,660 – 3,345). | R.E.A. Stewart et al. 2014c | Unknown | MacLaren Atlantic Limited 1978. |
| Central-Low Arctic DU | blank | 1,408 (CV 0.22, 95% CI 922 - 2,150) |
Mar.-Apr. 2012 | Winter aerial line-transect surveys in West Greenland waters, corrected for availability and perception biases. | Heide-Jørgensen et al. 2014 | Unknown | blank |
| Central-Low Arctic DU | SHSUBL | ???? | Sept. 2014 | Haulout surveys conducted in 2014, only two animals seen (on Charles Island), excluding Walrus counted in area of overlap with SEB (Salisbury and Nottingham islands). Historical catches and observations suggest reduction in abundance and distribution. | Hammill et al. 2016b (also see text) | Unknown | blank |
| Central-Low Arctic DU | Mix: NWHB, SHSUBL, SEB? |
4,675 (CV 0.45, 95% CI 1,845 - 11,842) 6,020 (CV 0.40, 95% CI 2,485 - 14,585) |
Mar.- Apr. 2012 | Aerial strip-transect survey of Hudson Strait, corrected for availability and perception biases. Two replicates, second incomplete due to weather (higher estimate). Stock composition of Walrus wintering in this area is unknown, most animals seen in central and western Hudson Strait, along both coasts (Baffin Island and Nunavik). | Elliot et al. 2013 | Unknown | blank |
| Central-Low Arctic DU | Low Arctic | 200 (95% CL = 70–570) (rounded to the nearest 10 animals) | Sept. 2014 | Haulout counts adjusted using several different methods, SC = 58; MCP = 101; BC = 196 | Hammill et al. 2016b | Unknown | Opportunistic counts by OMNR staff at shoals near Cape Henrietta Maria (see text). |
| All Canadian Walrus populations | blank | ca. 21,400b | blank | Limited data for many stocks or populations; negatively biased. | blank | Unknown | blank |
a Hammill et al. (2016c) report model-based estimates of population size for these stocks in 2014 (see text).
b Total calculated using: 727 (PS-LS), 503 (WJS), and 1,251 (BB) for High Arctic; 10,379 (FB), 2,300 (SEB – midpoint of estimate range for Hoare Bay), and 6,020 (mixed: NWHB, SEB, SHSUBL) for Central Arctic; 200 for Low Arctic; population totals rounded to nearest 50 for discussion. Replacing the March-April 2012 Hudson Strait counts with the 2014 “Hudson Bay Davis Strait stock” estimate provides a similar total count.
Rescue effect
The ability of Atlantic Walrus to re-colonize areas where populations have been depleted or extirpated is unknown. The rarity of animals along the Atlantic coast of Canada since the Nova Scotia-Newfoundland-Gulf of St. Lawrence DU became extinct in the 1850s suggests that re-colonization is exceedingly slow at best. Walrus off west Greenland have become very scarce and Atlantic Walrus from the populations in east Greenland and Svalbard are unlikely to offer any immediate prospect of rescue for populations in the eastern Canadian Arctic. Whether this would change if the animals had a compelling reason to relocate, such as overabundance or disturbance, is unknown.
Threats and limiting factors
Walrus are gregarious and valuable, with a narrow trophic niche and restricted seasonal distribution that makes them relatively easy for hunters to locate and vulnerable to environmental changes (Born et al. 1995). Hunting continues to be an important factor for Atlantic Walrus populations in Canada but industrial development and climate change may become increasingly important. Shipping on a massive scale may soon disrupt Walrus habitats in Hudson Strait and Foxe Basin year-round. At recent workshops and hearings Inuit and scientists have expressed concern about the potential impacts from non-renewable resource exploration and development as well as disturbance by tourism (e.g., Qikiqtani Inuit Association 2011, 2012, 2013, 2014; DFO 2013; A. McPhee, DFO pers. comm. 2014). Other issues have also been raised related to the conservation and management of Walrus, including the need to identify important Walrus habitat, improve harvest and harvest loss reporting, collect and use local knowledge, develop appropriate local by-laws and best management practices, and to develop a formal management plan for the species (A. McPhee, DFO pers. comm. 2014; Wiig et al. 2014).
A Threats assessment for the High Arctic DU produced an overall threat impact score of “high” (Appendix 1). Medium-low impact threats included transportation and service corridors such as shipping lanes and flight paths and human intrusions and disturbance (recreational activities, work and other activities). Medium impact threats included biological resource use (i.e., subsistence harvesting), and low impact threats included natural system modifications and pollution. Unknown impact threats included invasive and other problematic species and climate change (e.g., habitat shifting and alteration, temperature extremes). A Threats assessment for the Central-Low Arctic DU produced an overall threat impact score of “high” to “very high” (Appendix 2). High to medium impact threats included transportation and service corridors such as shipping lanes and flight paths. Medium impact threats were biological resource use (i.e., harvesting) and natural system modifications (e.g., from water management activities), and medium to low-impact threats were human intrusions and disturbance such as recreation activities. Pollution was considered a low impact threat, and unknown impact threats included energy production and mining, invasive and other problematic species, and climate change impacts.
Energy production and mining
Oil and gas drilling activity is uncertain given the moratorium on new oil and gas leasing announced in 2016 by the Government of Canada, which is in effect for 5 years at minimum (CBC 2016). It is uncertain as to whether or not the moratorium includes seismic exploration. The effects, if any, of pulsed noise from seismic exploration on Walrus are unknown, as is the ability of Walrus to habituate to noise.
Transportation and service corridors
Shipping-related disturbance of Walrus habitat in Hudson Strait and Foxe Basin will increase substantially during the next decade and even more by mid-century (D.B. Stewart et al. 2014b). These changes are being driven primarily by demand for mineral resources and by climate change. Portions of these shipping routes and others in the eastern Arctic are already travelled by grain carriers sailing to and from Churchill, MB, merchant vessels loaded with cargo or fuel (e.g., sealift) for communities and other developments, tourist cruise ships, fishing vessels, and government vessels (scientific, Coast Guard, Navy) (Stewart and Howland 2009; Chan et al. 2012; D.B. Stewart et al. 2014b). There is little information on Walrus response to vessels. McFarland and Aerts (2015) studied vessel disturbance of Pacific Walrus in water and hauled out on sea ice, and the most energetic behaviour responses (diving and changing course/speed) predominantly occurred when the vessel was within 500 m. Salter (1979a) recorded responses of Atlantic Walrus at a High Arctic uglit. Walrus responded to some flights by helicopters and fixed-wing aircraft, but to none of six approaches by small boats (Zodiacs) at distances of 1.8 to 7.7 km from the haulout. Underwater noise might disrupt the transmission of important sounds made by Walrus, such as songs during the breeding season and mother-calf communications (Moore et al. 2012; Stewart et al. 2012).
The Mary River Iron Mine project poses the threat of a very significant increase in ship traffic. The main project includes a port in Steensby Inlet, Foxe Basin, and shipping along the “southern” route through Hudson Strait. While this project has been postponed for the present (BIMC 2013), it has government approval to export 18 Mt of iron ore annually over a period of at least 21 years (BIMC 2012; NIRB 2012). This will require about 102 round trips annually by ice-breaking vessels each capable of carrying about 180,000 t of ore. The Mary River Iron Mine could also increase annual iron ore exports by 3.35 Mt without triggering further environmental review (i.e., 121 c.f. 102 round trips annually; NIRB 2012), and it has numerous other iron deposits that may be mined either in sequence or in parallel. The ore would be loaded at Steensby Inlet in northern Foxe Basin and delivered via Hudson Strait to Rotterdam in the Netherlands. In late winter 2012, the highest densities of Walrus in Hudson Strait were observed within 3 km of the planned year-round shipping route; densities decreased steadily with distance from the route (Elliot et al. 2013). Scientists and Inuit have expressed concerns regarding the potential effects of shipping activities including underwater shipping noise, ice habitat alteration, uglit disturbance, sediment mobilization, introduction of non-indigenous species via ballast water, accidental oil spills, and ship strikes on Walrus distribution and abundance (e.g., Megannety 2011; Stewart et al. 2012; Qikiqtani Inuit Association 2011, 2012, 2013, 2014; Trent University and Makivik Corporation 2015).
Ship traffic along sections of the same routes could increase much more within the next decade if other metal mining projects near southern Ungava Bay (Nunatsiaq News 2012a), Deception Bay (Xstrata Nickel 2011; www.canadianroyalties.com/en/), western Foxe Basin (Advanced Explorations Inc. 2012; Saul et al. 2012), the Belcher Islands (Nunatsiaq News 2012b), and in the Kivalliq (Areva 2011; Agnico Eagle 2014) proceed to development or expand (see also Gavrilchuk and Lesage 2014). Boat and barge traffic along the Hudson Bay and James Bay coasts of Ontario is expected to increase with development of the Victor diamond mine and clean up of existing DEW Line sites (J. Fitzsimmons, C. Risley, and C. Chenier, OMNR, pers. comm. 2014). Farther north, the Mary River Iron Mine project’s “early revenue phase” has government approval to export 3.5 Mt of ore annually (as of 2016) during the summer via Milne Inlet on northern Baffin Island to Rotterdam (BIMC 2013; NIRB 2014). This will require about 55 return trips annually by ore carriers via Pond Inlet and Baffin Bay, potentially affecting Walrus in these areas. The project is now proposing an expansion of activities through the northern shipping route (BIMC 2016).
Biological resource use
Subsistence hunting
Data on the historical catches of Walrus in Canada are incomplete and vary widely in quality. The 1980-2015 subsistence catch data presented are from the recent catch history (D.B. Stewart et al. 2014a), which elaborates on their sources and quality, and from DFO catch statistics (DFO Iqaluit and DFO Quebec unpubl. data) (Table 2; Figure 6). This period followed enactment of the Walrus Protection Regulations under the Fisheries Act (Canada 1980: P.C. 1980 -1216), which reduced the number of Walrus “an Indian or Inuk” could hunt and kill in one year from seven to four (Section 3), except where new annual community quotas were scheduled instead for Coral Harbour: 60, Sanikiluaq: 10, Arctic Bay: 10, and Clyde River: 20. Details of the methodology behind these numbers, and uncertainties are given in Appendices . Sources of uncertainty in reported numbers include rates of reporting harvests, and rates at which animals were injured or killed but lost. These rates likely vary between areas of the Arctic, as well as with time (). These uncertainties, together with those in population estimates and life history measures, mean that, while reported catches have declined (Table 2; Figure 6), it is uncertain whether current levels of hunting in Canada and Greenland are sustainable.
| Yearc | High Arcticd | Central-Low Arctic Subsistence |
Central-Low Arctic Sport |
Central-Low Arctic Total |
Canadian Catches |
|---|---|---|---|---|---|
| 1980 | 15 | 500 | -- e | 500 | 515 |
| 1981 | 20 | 618 | -- e | 618 | 638 |
| 1982 | 36 | 656 | -- e | 656 | 692 |
| 1983 | 18 | 619 | -- e | 619 | 637 |
| 1984 | 19 | 447 | -- e | 447 | 466 |
| 1985 | 15 | 419 | -- e | 419 | 434 |
| 1986 | 15 | 418 | -- e | 418 | 433 |
| 1987 | 22 | 365 | -- e | 365 | 387 |
| 1988 | 15 | 440 | -- e | 440 | 455 |
| 1989 | 15 | 311 | -- e | 311 | 326 |
| 1990 | 25 | 391 | -- e | 391 | 416 |
| 1991 | 19 | 420 | -- e | 420 | 439 |
| 1992 | 30 | 440 | -- e | 440 | 470 |
| 1993 | 20 | 453 | -- e | 453 | 473 |
| 1994 | 40 | 398 | -- e | 398 | 438 |
| 1995 | 16 | 246 | 1 | 247 | 263 |
| 1996 | 11 | 347 | 3 | 350 | 361 |
| 1997 | 14 | 434 | 9 | 443 | 457 |
| 1998 | 24 | 396 | 8 | 404 | 428 |
| 1999 | 13 | 457 | 10 | 467 | 480 |
| 2000 | 12 | 400 | 7 | 407 | 419 |
| 2001 | 8 | 174 | 15 | 189 | 197 |
| 2002 | 4 | 191 | 16 | 207 | 211 |
| 2003 | 14 | 275 | 15 | 290 | 304 |
| 2004 | 10 | 122 | 10 | 132 | 142 |
| 2005 | 4 | 272 | 17 | 289 | 293 |
| 2006 | 5 | 438 | 9 | 447 | 452 |
| 2007 | 6 | 197 | 0 | 197 | 203 |
| 2008 | n/dc | 181 | 4 | 185 | 185 |
| 2009 | 9 | 221 | 9 | 230 | 239 |
| 2010 | 9 | 290 | 8 | 298 | 307 |
| 2011 | 6 | 207 | 12 | 219 | 225 |
| 2012 | 2 | 334 | 8 | 342 | 344 |
| 2013 | 0 | 57 | 17 | 74 | 74 |
| 2014 | 17 | 166 | 9 | 175 | 192 |
| 2015 | 2 | 146 | 18 | 164 | 166 |
| Total | 510 | 12147 | 205 | 12352 | 13161 |
c Years are DFO reporting year (e.g., 1995 = 1 April 1995 to 31 March 1996).
d n/d = no data.
e indicates no sport hunts were conducted.
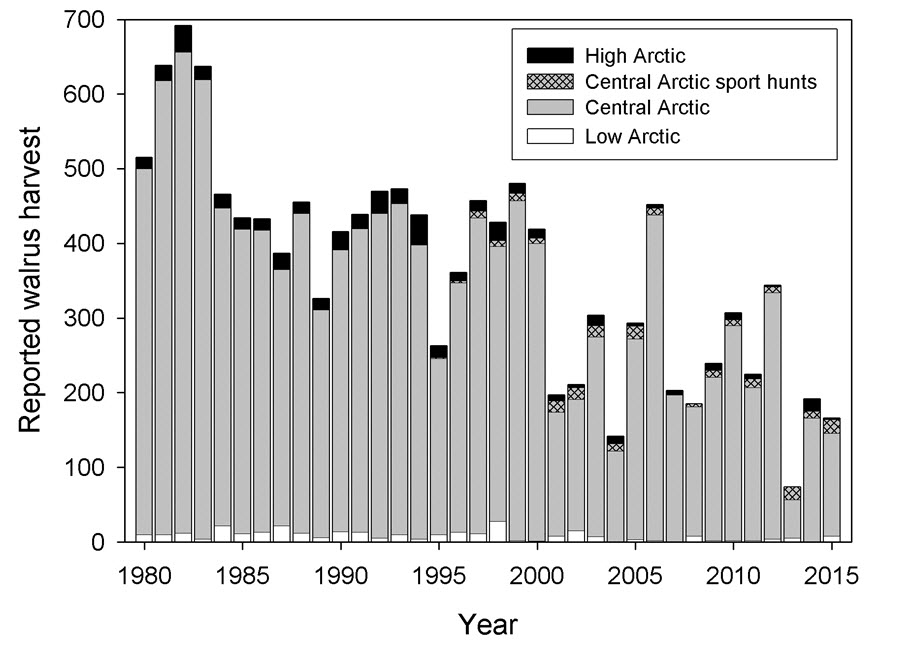
Long description for Figure 6
Chart illustrating trends in annual landed catches of Walrus from populations in the eastern Canadian Arctic by subsistence and sport hunters, from 1980 to 2015.
Sport hunting
From 1928 through 1994, only Inuit and First Nations could hunt Atlantic Walrus in Canada. In 1995, a limited hunt was opened for non-resident hunters in order to benefit communities with nearby Walrus populations. Since then applications for sport hunts have been approved annually by the Nunavut Wildlife Management Board (NWMB) and DFO has then issued licences under the Marine Mammal Regulations (s. 4) (A. Currie DFO Iqaluit, pers. comm. 2012; A. McPhee, DFO Winnipeg, pers. comm. 2014) (Table 2; Figure 6). Most animals are taken from northern Foxe Basin and some from northern Hudson Bay. Sport hunters landed a total of 53 Walrus in 1995-2001, 71 in 2002-2008, and 81 in 2009-2015. A recent decline is related to Igloolik suspending sport hunts and other tourism activities in 2008 through 2010 over concerns that this activity was disturbing Walrus, driving them farther from the community and making the subsistence hunt more difficult (CBC 2008; D.B. Stewart et al. 2014a). During this closure, sport landings from the Coral Harbour area increased.
Non-resident hunters must travel with local guides approved by the Hunters and Trappers Organization (HTO) (A. McPhee, DFO pers. comm. 2014). Sport Hunt Reporting Cards have recently been developed by DFO and co-management organizations. These cards require the sport hunter to provide DFO with hunt-related information upon completion of each hunt (e.g., number of strikes, gender, struck and lost information, location, number harvested, etc.). In 2011/12, sampling for a number of biological parameters was included as a licence condition. This requires the sport hunter to provide DFO with liver, kidney, skin and muscle samples, tusk lengths and girth measurements. The hunter can take the tusks and cape but must leave the meat in the village.
Commercial fisheries
Commercial fisheries that overlap the Walrus’ range may compete directly for food, damage feeding habitats, and cause disturbance. Scallop dragging has been tried in Cumberland Sound, Hudson Strait, and Ungava Bay, and along the Nunavik coast of Hudson Bay, but these fisheries were uneconomical and are no longer operating (Stewart et al. 1993; Lambert and Prefontaine 1995; Stewart and Howland 2009). Scallops occur mostly on gravelly substrates whereas Walrus’ forage primarily on bivalves buried in soft bottoms. Open-water trawl or drag fisheries for shrimp, turbot, cod, or other species would not compete directly with Walrus for food but could disturb Walrus and their feeding habitat. Ship noise could displace Walrus from their uglit and interfere with their communication (Salter 1979a; Born et al. 1995; Stewart 2002).
Human intrusions and disturbance
Machine noise, particularly from aircraft, disturbs Walrus, which may stampede into the water causing significant calf mortality and spontaneous abortion of fetuses (Salter 1979a; Born et al. 1995; Okonek et al. 2009, 2010). The response depends upon many factors related to the characteristics of the aircraft and its flight, environmental conditions, and the demography and activity state of the affected Walrus. There is concern that frequent aerial tourist traffic may disturb Walrus hauled out along the Ontario coast of Hudson Bay (C. Chenier, DNR Cochrane, pers. comm. 2003).
The reactions of Walrus to vessel noise vary depending upon their previous experiences (Born et al. 1995). Animals from hunted populations tend to be skittish when approached by boats but when asleep can sometimes be approached within 10-20 m. Ice breaking activities cause Pacific Walrus to enter the water: females and calves when the ship is within 500–1000 m and males when it is within 100–300 m. They move 20–25 km away from the disturbance if it continues but return after it stops.
Tourist activities can cause Walrus to stampede (Cody 2003). There is concern among Inuit and scientists not only that disturbance from tourism may cause stampedes but also that it could drive herds further into the pack ice or away from their traditional uglit (Stewart 2002; Dueck 2003; C. Chenier, DNR Cochrane, pers. comm. 2003). This concern prompted the Igloolik HTO to ban all forms of tourism related to Walrus in northern Foxe Basin from ca. May 2008 through May 2011 (CBC News 2008; Gagnon 2011). International tours have since resumed bringing visitors to view Walrus at uglit in Foxe Basin in July and August (https://www.windowsonthewild.com/canada/northwest-territories-nunavut/bowhead-whales-walrus-of-foxe-basin/). Cruise tourism in the eastern Canadian Arctic has increased over the past quarter century (Stewart et al. 2007, 2010). Inuit in Pond Inlet and Clyde River have expressed concern about the added stress to marine mammals caused by cruise ships (J. Alooloo and M. Kotierk, pers. comm. 2013). Studies are underway at Svalbard to assess the effects of disturbance by tourism on haulout dynamics (Lydersen and Kovacs 2014).
Natural system modifications
The impacts on Walrus of inland hydroelectric developments that alter the seasonality of freshwater runoff into Hudson and James bays are unknown (Stewart and Lockhart 2005). Inuit in the Belcher Islands have expressed concern that these developments are reducing winter currents, and contributing to heavier ice conditions that harm overwintering marine birds and mammals (Panel Report 2006:346; Stewart and Hamilton 2007). Fresh water released to meet winter power demands may be diluting the offshore surface waters, enabling ice to form more rapidly and entrap wildlife (J. Heath cited in George 2013).
Invasive and other problematic species
Predation by Polar Bears and Killer Whales may increase in response to ongoing climate change as Walrus are forced to make greater use of terrestrial sites and spend more time in open water (Garlich-Miller et al. 2011). Young Walrus will be most at risk from predation by these species. Walrus are consumed most often by older male Polar Bears, and there is spatial variation in the importance of Walrus as a prey item, with Foxe Basin bears feeding on a larger proportion of Walrus than those in other areas of the Canadian Arctic (Thiemann et al. 2007, 2008). Killer Whale occurrence is increasing in the eastern Canadian Arctic (Higdon and Ferguson 2009; Higdon et al. 2014). The species does prey on Pacific Walrus (Kryukova et al. 2012) but Inuit indicate that Killer Whales in eastern Canadian Arctic waters rarely if ever feed on Walrus (Ferguson et al. 2012). While Killer Whales do not appear to regularly hunt Atlantic Walrus, they might acquire this behaviour and learn to hunt them successfully in the future if hunting opportunities increase. However, given the abundance of other prey species that are easier and safer to hunt it seems unlikely that Killer Whales will become a major predator of Atlantic Walrus in the next decade. Disease transmission could increase in response to increased ugli use (Burek et al. 2008). Walrus might also be exposed to novel pathogens and parasites as vector species expand their distributions northward.
Pollution
Cadmium (Cd) concentrations in the soft tissue of Atlantic Walrus from Foxe Basin and northeastern Hudson Bay are high relative to other mammals (0.03-130.9 µg·g–1 wet wt; Outridge et al. 1994; Wagemann and Stewart 1994), as are their lead (Pb) concentrations (0.02–0.58 µg·g–1 wet wt; Wagemann and Stewart 1994). Mercury (Hg) levels in the muscle ranged from 0.02 to 1.34 µg·g–1 wet wt (Wagemann and Stewart 1994; Wagemann et al. 1995). They were higher on average (0.11 µg·g–1 wet wt, SD 0.13) than those sampled from the Thule area of Greenland in the late 1970s (0.06 µg·g–1 wet wt, SD 0.03; Born et al. 1981). The sources of cadmium, lead, and mercury in Atlantic Walrus in northern Foxe Basin appear to be natural (Outridge et al. 1997, 2002). Metal concentrations in the tissue of these Walrus paralleled that in the tissue of local clams, with the exception of cadmium (Wagemann and Stewart 1994). No temporal trend was found in liver mercury concentration (range 1-5 µg·g–1 wet wt) of Walrus in Foxe Basin over the period 1982 through 2008 (Gaden and Stern 2010). The effects of these and other chemical contaminants on Walrus are unknown (Wagemann and Stewart 1994; de March et al. 1998; Fisk et al. 2003).
Levels of organochlorines in Walrus tissues are generally low because they primarily feed low in the food web. Walrus typically have 4–10 times lower concentrations of organochlorine contaminants than Beluga Whales from the same area but a similar pattern of residues (Norstrom and Muir 2000). The highest levels are found in individuals that are thought to eat seals, which accumulate these contaminants in their fat (Muir et al. 1995).
The direct and indirect effects of petroleum on Walrus have not been studied. Born et al. (1995) believed that several aspects of the species’ ecology may make it vulnerable to oil pollution, in particular, its gregariousness, which may spread oil from animal to animal, its preference for coastal areas and loose pack ice where oil may be more likely to accumulate, and its reliance on benthic molluscs which may accumulate petroleum hydrocarbons or succumb to the oil. Walrus populations may be most vulnerable to harm from oil spills during the calving period, and calves may be the most vulnerable component of the population.
Climate change
The eastern Canadian Arctic has been experiencing major changes in ice conditions (Parkinson and Cavalieri 2008; Sahanatien and Derocher 2012; Parkinson 2014) that are bound to affect Walrus ecology (D.B. Stewart et al. 2014b). The direct effects of climate change (warming or cooling) on Atlantic Walrus are likely limited and not necessarily negative but the potential for indirect effects is worrisome. Sea ice cover does not appear to be a critical determinant of their populations, since various areas of their habitat are seasonally ice-free and their pristine distribution extended far south of its present limits (Laidre et al. 2008). A decrease in the extent and duration of Arctic sea ice in response to warming might increase food availability for Walrus by improving access to feeding areas in shallow inshore waters that are currently covered in winter by landfast ice (Born et al. 2003; Born and Wiig 2005; Laidre et al. 2008). Atlantic Walrus are clearly capable of thriving on boreal bivalves if there is a northward movement of these species in response to warming.
Warming and loss of ice cover might cause a trophic shift from ‘ice algae – benthos’ to a ‘phytoplankton–zooplankton’ dominance (Piepenburg 2005). This shift toward a more pelagic food web could reduce benthic production and thereby food for Walrus (Bluhm and Gradinger 2008). In the Bering Sea, between ca. 1995 and 2005, an increase in air and ocean temperatures coincided with a reduction in sea ice, a reduction in benthic prey populations, an increase in pelagic fish populations, and geographical displacement of marine mammal distributions (Grebmeier et al. 2006).
Ocean acidification related to increased atmospheric CO2 may also alter trophic dynamics by reducing the availability of calcium to marine invertebrates, including bivalves, and by altering host-pathogen relationships in favour of pathogens, changing the abundance and composition of Walrus prey (Azetsu-Scott et. al. 2010; Garlich-Miller et al. 2011; Kroeker et al. 2013; Asplund et al. 2014). The likelihood of such trophic changes, their time horizon, and possible effects on Walrus are unknown.
Atlantic Walrus may be less vulnerable to ice loss than Pacific Walrus, as Atlantic Walrus’ feeding areas tend to be closer to terrestrial uglit and fewer animals occupy these uglit, except perhaps in south-central Foxe Basin. Fischbach et al. (2009) have postulated that the mortality of 131 young Pacific Walrus along the Chukchi Sea coast near Icy Cape, Alaska, in mid-September 2009 was related to loss of sea ice cover over the continental shelf but this cannot be confirmed. A large mortality event involving Pacific Walrus at Wrangel Island in 2007 was also coincident with light ice conditions and Polar Bear predation (Ovsyanikov et al. 2007). In the event of ice loss Atlantic Walrus would not likely experience the same need to increase energy expenditure to gain access to distant foraging areas or face the same degree of increased intraspecific competition for food resources near the uglit. Females with dependent young are the animals most likely to be affected by changes in energy expenditure or competition (Garlich-Miller et al. 2011).
If sea ice is not available Walrus must haul out on land. The fossil record suggests that between 9000 and 1000 y BP, when Walrus occupied areas along the east coast of Canada, the summer surface water temperatures there may have ranged between 12 and 15°C (Miller 1997). Whether these animals summered in these relatively warm waters or moved north into cooler waters is unknown. Within the last few hundred years they clearly thrived in the Gulf of St. Lawrence and on the Scotian Shelf (Shuldham 1775; Perley 1850; Gilpin 1869; Stewart 1806; Allen 1880). Behavioural and physiological responses to changes in air temperature suggest that Pacific Walrus calves can maintain their body temperature at an air temperature of 18°C in still air and shade or under equivalent conditions (Fay and Ray 1968; Ray and Fay 1968). Above this temperature they withdraw into the water to avoid overheating. Air temperatures at or above this level for an extended period might disrupt normal feeding, moulting, and calving schedules.
The indirect effects of climate change may pose a greater threat to Walrus than the change itself. Winter hunting pressure on Walrus may decrease as ice conditions deteriorate and become less predictable (Laidler 2009). The duration of open-water access to Walrus may increase in response to ice loss, and Walrus may also become more concentrated at terrestrial haulouts (Born and Wiig 2005; NAMMCO 2006), but whether hunting pressure will increase in response is unknown. Careful regulation of hunting may be required to prevent Walrus disappearing from uglit as they did in west and northwest Greenland during the 20th century (Born and Wiig 2005). Earlier loss of sea ice could prompt Arctic marine fisheries to expand into areas that have not hitherto been fished commercially (Christiansen et al. 2014), thereby increasing the potential for interactions between Walrus and fisheries.
Climatic warming could also have unanticipated impacts. An increase in the occurrence of thunderstorms, for example, might disturb Walrus and result in mortality as observed at Round Island, Alaska (Okonek and Snively 2005). If other prey species are limited, Walrus might eat more seals and thereby increase the incidence of Trichinella infection (Garlich-Miller et al. 2011) although the life history of T. nativa is not well known. A recent study in Nunavik used a combination of histological analyses of skin sections and traditional ecological knowledge (TEK) to study the effects of solar ultraviolet radiation (UV) on Walrus (Trent University and Makivik Corporation 2015; Martinez-Levasseur et al. 2016). Histological analyses provided data on the prevalence of UV-induced cellular lesions and interviews with experienced Inuit Walrus hunters and Elders provided information on incidences and temporal changes of UV-induced gross lesions. A range of skin abnormalities consistent with UV damage were detected at the microscopic scale, but these types of UV effects were not widely observed at the whole-animal level (e.g., absence of skin blistering and eye cataracts) by the hunters interviewed, indicating no relation between increased sun radiation secondary to ozone loss and Walrus health (Trent University and Makivik Corporation 2015; Martinez-Levasseur et al. 2016).
Protection, status and ranks
The Nova Scotia-Newfoundland-Gulf of St. Lawrence population of Atlantic Walrus in Canada, formerly the Northwest Atlantic population, is currently listed on Schedule 1 of the Species at Risk Act as Extirpated (see also Reeves 1978; Richard and Campbell 1988). Fisheries and Oceans Canada does not consider recovery of this population to be feasible at this time (DFO 2008). COSEWIC reassessed this species (under the new name) as Extinct in 2017. The Atlantic Walrus in Canada was originally treated by COSEWIC as two separate populations: Eastern Arctic population (Not at Risk in April 1987 and May 2000) and Nova Scotia-Newfoundland-Gulf of St. Lawrence population (Extirpated in April 1987 and May 2000). In April 2006, COSEWIC included both populations in a single designatable unit for Atlantic Walrus in Canada, and the species was designated Special Concern (COSEWIC 2006). Atlantic Walrus was split into three populations in April 2017, and each population received a separate risk status: High Arctic population, Special Concern; Central-Low Arctic population, Special Concern; and Nova Scotia-Newfoundland-Gulf of St. Lawrence population, Extinct. In 2008, the Nunavut Wildlife Management Board and Government of Canada signed a memorandum of understanding to harmonize the designation of rare, threatened, and endangered species under the Nunavut Agreement and the listing of wildlife species at risk under the Species at Risk Act (AANDC 2011) but no similar agreement is in place for Nunavik (M. Hammill, DFO Mont-Joli, pers. comm. 2013). The Province of Quebec has included the Atlantic Walrus on its “List of wildlife species likely to be designated threatened or vulnerable” (Liste des espèces susceptibles d’être désignées menacées ou vulnérables) under the Act Respecting Threatened or Vulnerable Species (CQLR, c E-12.01) (Loi sur les espèces menacées ou vulnérables) (RLRQ, c E-12.01) (Gouvernement du Québec - Liste des mammiferes, In French only). A socioeconomic analysis of the benefits and costs of listing the Atlantic Walrus under the Species at Risk Act, and undertaking recovery or management actions, was completed in 2008 (Gustavson et al. 2008).
Legal protection and status
Regulations on the hunting of Walrus in Canada, and on the international trade in Walrus parts, afford limited protection for Atlantic Walrus populations in Canada (Shadbolt et al. 2014; Wiig et al. 2014).
Walrus hunting in Nunavut is co-managed by the NWMB, which under the Nunavut Agreement (NLCA) is the primary instrument of wildlife management and regulator of access to wildlife in the Nunavut Settlement Area (NSA), and also has recommending authority in adjacent areas. The NWMB board consists of four Inuit-appointed and four government-appointed members, plus a chairperson. Fisheries and Oceans Canada (DFO) advises the NWMB and hunting communities on sustainable hunting levels and they in turn use this information to manage hunting. Regional Wildlife Organizations (RWOs) and the community-level Hunters and Trappers Organizations (HTOs) also play a role in the management of Walrus hunting, in part by providing local and traditional knowledge to inform the NWMB recommendations. In 2012, the NWMB established that the basic needs level for Walrus under the NLCA (s. 5.6.25) should be equal to the total allowable harvest, thereby reserving all Walrus harvests in the NSA for Inuit (NWMB staff, pers. comm. 2014).
In Nunavik, Walrus co-management is led by the Nunavik Marine Region Wildlife Board (NMRWB), established in 2009, subsequent to ratification of the Nunavik Inuit Land Claims Agreement (NILCA). The NMRWB consists of three Makivik-appointed and three government-appointed members and a chairperson and is the main instrument of wildlife management for the Nunavik Marine Region (NMR). At the local level, each Nunavik community has a wildlife organization (LNUK, local “nunavimmi umajutvijiit katajuaqatigninga”), which consults with a regional RNUK composed of four elected executive members from amongst presidents of the community LNUKs (analogous to the HTO and RWO system in Nunavut).
While the NWMB and NMRWB are the main instruments of wildlife management and the main regulators of access to wildlife in the NSA and NMR, the Government of Canada retains ultimate responsibility for wildlife management, and thereby conservation. Hunting is regulated under the Fisheries Act and the Marine Mammal Protection Regulations by DFO, which manages Walrus in other jurisdictions in cooperation with other agencies. Two Walrus co-management working groups (Foxe Basin and Baffin Bay-High Arctic) worked together to complete an Integrated Fisheries Management Plan (IFMP) for Walrus in Nunavut (DFO 2013:13; A. McPhee, DFO Winnipeg pers. comm. 2017). The IFMP is being implemented and used to provide direction for the sustainable management of Walrus in the Nunavut Settlement Area. In the event that an IFMP is inconsistent or in conflict with obligations under land claims agreements, the provisions of the land claims agreements will prevail to the extent of the inconsistency or conflict (NWMB staff, pers. comm. 2014).
Under Marine Mammal Regulations (SOR/93-56) of the Canadian Fisheries Act (R.S.C., 1985, c. F-14) “an Indian or Inuk other than a beneficiary” can hunt and kill four Walrus in one year without a licence, except where community quotas limit annual catches. “Beneficiary” refers to a beneficiary of the Western Arctic (Inuvialuit) Claims Settlement Act or James Bay and Northern Québec Native Claims Settlement Act. Since 1980, Coral Harbour has had an annual catch quota of 60 Walrus, Sanikiluaq 10, Arctic Bay 10, and Clyde River 20 (Canada 1980: P.C. 1980 -1216). In the other communities it is the number of Inuit rather than the number of Walrus that limits the allowable catch (Stewart 2002). Under the Nunavut and Nunavik land claims agreements Inuit harvesting rights can only be limited by their respective board (NWMB or NMRMB) or a minister to the extent necessary to effect a valid conservation purpose (NLCA S. 5.3.3; NILCA S.5.5.3). Therefore, until the current system creates a conservation concern, there is no justification for implementing stricter measures (M. O’Connor, NMRWB, pers. comm. 2014).
Non-Indigenous harvesters require a licence under the Marine Mammal Regulations or Aboriginal Communal Fishing Licence Regulation to hunt Walrus (DFO 2002; Hall 2003). Sport hunts are managed by limiting the number of licences approved annually.
Within Canada buying, selling, trading or bartering edible Walrus parts is prohibited except by “an Indian or Inuk other than a beneficiary”, within the Northwest Territories, Nunavut, the Yukon Territory, Quebec or Newfoundland (Marine Mammal Regulations SOR/93-56). Beneficiaries of the Western Arctic (Inuvialuit) Claims Act and the James Bay and Northern Québec Native Land Claims Settlement Act must conduct any trade in Walrus parts in accordance with the agreement in which the beneficiary is enrolled. The regulations prohibit disturbing Walrus, attempting to kill them except in a manner that is designed to kill quickly, hunting without equipment to retrieve, waste of edible parts, and abandoning a killed Walrus without making a reasonable effort to retrieve it.
DFO has proposed amendments to the Marine Mammal Regulations (Canada Gazette Vol. 146, No. 12. March 24, 2012) that would prohibit vehicles (except aircraft in flight) from approaching within 100 m of Walrus (year-round), and within 200 m when Walrus are hauled out on ice or 300 m when they are hauled out on shore (1 June-31 October). Flight maneuvers, including landings, takeoffs, and changes in course or altitude intended to bring aircraft closer to Walrus would be prohibited when aircraft are flying at an altitude of less than 304.8 m (1000 ft) within a 0.5 nautical mile radius of the animals. This prohibition would not apply to commercial aircraft operating on a scheduled flight plan. Other exemptions apply to people authorized to hunt Walrus and to employees of various government agencies.
Live capture and tagging are permitted only with a licence, which must be approved by the NWMB in Nunavut and the NMRWB in Nunavik. In 2003, Makivik sent a calf from Nunavik to the Quebec Aquarium, where it survives today; two calves sent there in 2004 did not survive (S. Olpinski, pers. comm. 2014).
A Marine Mammal Transportation Licence from DFO is required to transport Walrus parts within Canada, with the exception of Indigenous harvesters who land the Walrus in one jurisdiction and are returning to their home in another jurisdiction (Marine Mammal Regulations SOR/93-56).
The Atlantic Walrus is listed in Appendix III of the Convention on International Trade in Endangered Species (CITES) (Richard and Campbell 1988; Hall 2003; CITES 2014; Shadbolt et al. 2014). Anyone wishing to export Walrus parts or derivatives from Canada must obtain an export permit from the Canadian CITES administration. A total of 181 CITES export permits were issued from 1992 to 2001 (Hall 2003), and 438 in 2002 to 2011 (A. Tilochand, DFO Tignish, pers. comm. 2012). The apparent increase is not definitive of a change in hunting pressure. A single permit can cover one or many pieces, and the products from a single animal may be included in one or many permits. Awareness of permitting requirements has likely increased over the past decade with the increase in Arctic tourism, and reporting mechanisms also may have changed.
There is no formal agreement between Canada and Greenland for the management of shared stocks of Atlantic Walrus.
Habitat protection and ownership
Existing National Parks, National Wildlife Areas, Migratory Bird Sanctuaries, Indian Reservations, and other lands owned and managed by the Government of Canada afford little protection to Walrus habitat. Walrus haul out in the Polar Bear Pass National Wildlife Area (Davis et al. 1978) but this area only protects a few of their uglit along the east coast of Bathurst Island. They also haul out on Coburg Island in the Nirjutiqarvik (Nirjutiqavvik) National Wildlife Area, and possibly on western Bylot Island in Sirmilik National Park. Walrus may occasionally haul out in the Ninginganiq National Wildlife Area south of Clyde River (R.E.A. Stewart et al. 2014c), at the East Bay Bird Sanctuary on Southampton Island, and the Bowman Bay Wildlife Sanctuary on Baffin Island. They may also haul out along the northeastern coast of Bathurst Island on lands reserved for a proposed North Bathurst Island National Park. Their use of other National Parks and Bird Sanctuaries is very limited. Overall, federal lands offer temporary protection to a few Walrus. Inuit have the right to hunt in National Parks and other conservation areas within Nunavut and Nunatsiavut. In itself, this level of habitat protection is certainly insufficient to ensure the long-term survival of the species. The Lancaster Sound National Marine Conservation Area, which is in the feasibility planning stage, may offer additional protection.
The North Baffin Regional Land Use Plan (Sec. 3.3.8) and Keewatin Regional Land Use Plan (Sec. 2.7) specifically mention protection of Walrus uglit (haulouts), although the level of protection is vague (Nunavut Planning Commission 2000a, b). A Nunavut-wide land use plan is being prepared, and the most recent (June 2016) draft assigned haulout sites a Protected Area Land Use Designation that prohibits incompatible uses and includes setback requirements of up to 5 km (NPC 2016). The Walrus haulouts identified and mapped by NPC (2016) are incomplete, however, and are limited to Foxe Basin only (using data from Stewart et al. 2013). Walrus Island was identified by Coral Harbour as a Community Area of Interest. These areas are also assigned a Protected Area Land Use Designation that will protect some important Walrus haulout sites in the Kivalliq region. Many other Walrus haulout sites have been reported in the literature, and WWF-Canada recently compiled these data and provided the database to the NPC (B. Laforest, WWF-Canada, pers. comm., 2016) for future updates to the plan.
Acknowledgements
This work benefited from the presentations and discussions at the Pre-COSEWIC peer review meeting for the Atlantic Walrus. The meeting was organized by Fisheries and Oceans Canada (DFO) and Chaired by Don Bowen in Iqaluit on February 28 to March 1, 2012. Participants represented many communities, organizations, and jurisdictions. They are listed under Personal Communications and the report writers thank them all for their contributions. At other times, information and advice was gratefully received from Patt Hall, Allison McPhee, and Rob Stewart of DFO in Winnipeg, and many others. Caroline Girard and Anne Marie Cabana (DFO QC) and Amanda Currie, Richard Moore, and Jeremiah Young (DFO NU) contributed unpublished catch data; Chris Chenier and Martyn Obbard of the Ontario Ministry of Natural Resources data on Walrus in Ontario; Mike Bishop (Makkovik, Labrador) and Becky Sjare (DFO NL) information on Walrus in Labrador, Newfoundland, and the Maritimes; and Adrian Tilochand (DFO Tignish, PEI) data on the CITES permits. Dale McGowan (DFO Winnipeg, retired) furnished the fine Walrus drawing by Gerald Kuehl. Jenny Wu (COSEWIC) advised us on calculating the extent of occurrence and index of area of occupancy. Karen Timm (COSEWIC) assisted with organization and communications. Randall Reeves (past Co-chair), Jane Watson and other current or former members of the COSEWIC Marine Mammals Specialist Subcommittee, and Cecile Stewart of Arctic Biological Consultants provided constructive reviews of the draft report. Constructive reviews of the draft provisional and/or interim report were provided by Tim Frasier, Roger Gallant, Mike Hammill, Michael Kingsley, Kim Parsons, Randall Reeves, Rob Stewart, and Hal Whitehead (Co-Chair) (COSEWIC); Nunavut Wildlife Management Board Staff; Isabelle Gauthier (Government of Quebec); Gregor Gilbert and Stas Olpinski (Makivik Corporation); Maha Ghazal and Devon Imrie (Government of Nunavut); Mark O’Connor (Nunavik Marine Region Wildlife Board); Rosemary Curley (Government of Prince Edward Island); Christine Abraham (DFO Ottawa), Paul Blanchfield and Holly Cleator (DFO Science C&A), Melissa Landry (DFO EFM NCR), Allison McPhee (DFO Resource Mgmt. and Aboriginal Affairs, C&A), Jennifer Shaw (DFO Science NCR), Sam Stephenson (DFO SARA Program C&A), Katrina Sullivan (DFO SARA Program NL), and Robert Young (DFO Science C&A); Jay Fitzsimmons and Chris Risley (OMNR Species at Risk Branch), Chris Chenier (Northeast Region OMNR), and Scott Reid (OMNRF); Marie-France Noel and Lisa Pirie (Canadian Wildlife Service); Patrick Nantel and D. Whitaker (Parks Canada); and the Hunting, Fishing and Trapping Coordinating Committee (HFTCC). This work also benefited from the knowledge of participants in the Threats Workshop. The report writers are grateful for your contributions to this work.
Information sources
AANDC (Aboriginal Affairs and Northern Development Canada). 2011. 2008-2010 Nunavut Implementation Panel: Annual Report. Published under the authority of the Minister of Indian Affairs and Northern Development and Federal Interlocutor for Métis and Non-Status Indians. Ottawa, ON.
Aarluk Consulting Inc. 2005. Review of intersettlement trade opportunities for Arctic food products in Nunavut, final report. Prepared for the Department of Environment, Government of Nunavut, Iqaluit, NU. ii + 99 pp.
Acquarone, M., E.W. Born, and J.R. Speakman. 2006. Field metabolic rates of Walrus (Odobenus rosmarus) measured by the doubly labeled water method. Aquat. Mamm. 32(3): 363-369.
Advanced Explorations Inc. 2012. News release: Advanced Explorations Inc. announces filing of feasibility report for Roche Bay Iron Project. Toronto, Ontario. 4 pp. (Marketwire – Sept, 24, 2012)
Agnico Eagle. 2014. Final environmental impact statement (FEIS) – Meliadine Gold Project, Nunavut. April 2014. 11 vols.
Allen, G.M. 1930. The Walrus in New England. J. Mammal. 11: 139-145.
Allen, J.A. 1880. History of North American pinnipeds: a monograph of the Walrus, sea-lions, sea-bears and seals of North America. U.S. Geological and Geographical Survey of the Territories, Misc. Publ. 12: 785 pp.
Anders, G. 1966. Northern Foxe Basin, an area economic survey, 1965. Canada, Department of Northern Affairs and National Resources, Area Economic Survey Report 65–2: iv + 139 pp.
Andersen, L.W. and E.W. Born. 2000. Indications of two genetically different subpopulations of Atlantic Walrus (Odobenus rosmarus rosmarus) in west and northwest Greenland. Can. J. Zool. 78: 1999–2009.
Andersen, L.W., E.W. Born, D.W. Doidge, I. Gjertz, Ø. Wiig, and R.S. Waples. 2009. Genetic signals of historic and recent migration between sub-populations of Atlantic Walrus Odobenus rosmarus rosmarus west and east of Greenland. Endag. Species Res. 9: 197-211.
Andersen, L.W., E.W. Born, R.E.A. Stewart, R. Dietz, D.W. Doidge, and C. Lanthier. 2014. A genetic comparison of West Greenland and Baffin Island (Canada) Walrus: management implications. NAMMCO Sci. Publ. 9: 33-52.
Anderson, L.E. and J. Garlich-Miller. 1994. Economic analysis of the 1992 and 1993 summer Walrus hunts in northern Foxe Basin, Northwest Territories. Can. Tech. Rep. Fish. Aquat. Sci. 2011: iv + 20 pp.
AREVA (AREVA Resources Canada Inc.). 2011. Kiggavik Project environmental impact statement, Tier 1, Vol. 1, Main Document. 20 December 2011. 216 pp.
Asplund, M.E., S.P. Baden, S. Russ, R.P. Ellis, N. Gong, and B.E. Hernroth. 2014. Ocean acidification and host–pathogen interactions: blue mussels, Mytilus edulis, encountering Vibrio tubiashii. Environ. Microbiol. 16: 1029–1039. doi: 10.1111/1462-2920.12307 .
Azetsu-Scott, K., A. Clarke, K. Falkner, J. Hamilton, E.P. Jones, C. Lee, B. Petrie, S. Prinsenberg, M. Starr, and P. Yeats. 2010. Calcium carbonate saturation states in the waters of the Canadian Arctic Archipelago and the Labrador Sea. J. Geophys. Res. 115: C11021, doi:10.1029/2009JC005917.
Beaubier, P.H. 1970. The hunting pattern of the Igluligmiut: with emphasis on the marine environment. Montreal, Québec. M.A. Thesis. Department of Geography, McGill University. ix + 250 pp.
Bennett, J. and S. Rowley. 2004. Uqalurait: an oral history of Nunavut. McGill-Queen’s University Press, Montreal. 473 pp.
BIMC (Baffinland Iron Mines Corporation). 2012. Mary River Project final environmental impact statement. NIRB File No.: 08MN053, February 14, 2012. Toronto, ON.
BIMC (Baffinland Iron Mines Corporation). 2013. Mary River Project, Addendum to Final Environmental Impact Statement, June 2013. 10 volumes.
BIMC (Baffinland Iron Mines Corporation). 2016. Mary River Project. Project update. Final Report. November 30, 2016. 28 pp.
Bluhm, B.A. and R. Gradinger. 2008. Regional variability in food availability for Arctic marine mammals. Ecol. Appl. 18(2) Suppl.: S77-S96.
Boltunov, A.N., S.E. Belkov, Yu.A. Gorbunov, D.T. Menis, and V.S. Semenova. 2010. The Atlantic Walrus of the southeastern Barents Sea and adjacent regions: review of present-day status. WWF-Russia, Marine Mammal Council, Moscow, Russia. 17 pp.
Born, E.W. 1990. Distribution and abundance of Atlantic Walrus (Odobenus rosmarus rosmarus) in Greenland. Prepared by Greenland Home Rule, Department for Wildlife Management, Sjreleboderne 2, 1016 Copenhagen, Denmark for The International Workshop on Population Ecology and Management of Walrus. 26-30 March 1990, Seattle, Washington, USA. 65 pp.
Born, E.W. 2001. Reproduction in female Atlantic Walrus (Odobenus rosmarus rosmarus) from northwestern Greenland. J. Zool. (London) 255: 165-174.
Born, E.W. 2003. Reproduction in male Atlantic Walrus (Odobenus rosmarus rosmarus) from the North Water (N. Baffin Bay). Mar. Mamm. Sci. 19(4): 819-831.
Born, E.W. and I. Gjertz. 1993. A link between Walrus (Odobenus rosmarus) in northeast Greenland and Svalbard. Polar Record 29: 329.
Born, E.W., and Ø. Wiig. 2005. Potential effects on Atlantic Walrus of warming in the Arctic. Working paper to the NAMMCO workshop on Walrus in the North Atlantic, Copenhagen, 11-14 January 2005. 13 pp.
Born, E.W., I. Kraul, and T. Kristensen. 1981. Mercury, DDT and PCB in the Atlantic Walrus (Odobenus rosmarus rosmarus) from the Thule District, North Greenland. Arctic 34: 255–260.
Born, E.W., B. Clausen, and Sv.Aa. Henricksen. 1982. Trichinella spiralis in Walrus from the Thule district, North Greenland, and possible routes of transmission. Z. Saeugetierkd. 47: 246–251.
Born, E.W., I. Gjertz, and R.R. Reeves. 1995. Population assessment of Atlantic Walrus. Norsk Polarinst. Medd. 138: 100 pp.
Born, E.W., S. Rysgaard, G. Ehlmé, M. Sejr, M. Acquarone, and N. Levermann. 2003. Underwater observations of foraging free-Iiving Atlantic Walrus (Odobenus rosmarus rosmarus) and estimates of their food consumption. Polar Biol. 26: 348–357.
Born, E.W., M. Aquarone, L.Ø. Knutsen, and L. Toudal. 2005. Homing behavior in an Atlantic Walrus (Odobenus rosmarus rosmarus). Aquat. Mamm. 31(1): 11-21.
Brenton, C. 1979. Walrus. pp. 55–57. in FAO Fisheries Series No. 5: Mammals in the seas, Volume II. Pinniped species summaries and report on Sirenians. Food and Agriculture Organization of the United Nations, Rome.
Brice-Bennett, C. 1976. Inuit land use in the east central Canadian arctic. pp. 63–81, in M.M.R. Freeman (ed.) Inuit land use and occupancy study, Vol. 1. Canada Department of Indian and Northern Affairs, Ottawa.
Brody, H. 1976. Inuit land use in northern Baffin Island and northern Foxe Basin. pp. 153–172, in M.M.R. Freeman (ed.) Inuit land use and occupancy study, Vol. 1. Canada Department of Indian and Northern Affairs, Ottawa.
Brooke, L.F. 1992. A report on the 1991 beluga whale and Walrus subsistence harvest levels by the Inuit of Nunavik. Canada. Dept. of Fisheries and Oceans, [Aboriginal Fisheries Strategy (Canada)]. i + 16 pp.
Brooke, L.F. 1997. A report on the 1996 Nunavik beluga and Walrus subsistence harvest study. Canada. Dept. of Fisheries and Oceans, [Aboriginal Fisheries Strategy (Canada)]. 45 pp.
Brown, M., J.E. Green, T.J. Boag, and E. Kuitunen-Ekbaum. 1950. Parasitic infections in the eskimos at Igloolik, N.W.T. Can. J. Public Health 41: 508–512.
Brown, M., R.G. Sinclair, L.B. Cronk, G.C. Clark, and E. Kuitunen-Ekbaum. 1948. Intestinal parasites of eskimos on Southampton Island, Northwest Territories. Can. J. Public Health 39: 451–454.
Bruemmer, F. 1977. The gregarious but contentious Walrus. Nat. Hist. 86: 52–61.
Buchanan, F.C., L.D. Maiers, T.D. Thue, B.G.E. deMarch, and R.E.A. Stewart. 1998. Microsatellites from the Atlantic Walrus Odobenus rosmarus rosmarus. Mol. ecol. 7: 1083–1090.
Burek, K.A., F.M.D. Gulland, and T.M. O’Hara. 2008. Effects of climate change on Arctic marine mammal health. Ecol. Appl. 18(2) Suppl.: S126-S134.
Calvert, W. and I. Stirling. 1990. Interactions between polar bears and overwintering Walrus in the central Canadian High Arctic. Int. Conf. Bear Res. and Manage. 8: 351–356.
Campbell, W.C. 1988. Trichinosis revisited - another look at modes of transmission. Parasitology Today 4: 83–86.
Camus, T. 2003. The sunbathing Walrus. Halifax Herald. Thursday 12 June.
Canada, Privy Council.1928. Order in Council P.C. 1036 of June 20, 1928. Canada Gazette Vol. 61, No. 53, 30 June 1928, p. 4227.
Canada, Privy Council. 1980. Order in Council P.C. 1980 -1216 of May 8, 1980. SOR/80-338. Canada Gazette Part II, Vol. 114, No. 10, 28 May 1980, pp. 1860-1862.
CBC News North. 2008. Ban on Walrus tourism divides Igloolik. May 7, 2008, 3:45 pm CT. [Accessed April 2013]
CBC News North. 2012. Hunters urged to get Walrus meat tested for parasite. Posted August 17, 2012, 2:21 PM CT. [Accessed August 2013].
CBC News. 2014. Walrus spotted sunning near Portugal Cove-St. Philip’s. Posted July 19, 2014, 12:20 PM NT. [Accessed September 2014]
CBC News. 2016. U.S., Canada ban offshore drilling in Arctic waters. Posted: Dec 20, 2016 1:41 PM ET. [Accessed February 2017]
Chan, F.T., Bronnenhuber, J.E., Bradie, J.N., Howland, K., Simard, N., and Bailey, S.A. 2012. Risk assessment for ship-mediated introductions of aquatic nonindigenous species to the Canadian Arctic. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/105. vi + 93 p.
Charrier, I., T. Aubin, and N. Mathevon. 2009. Mother-calf vocal communication in Atlantic Walrus: a first field experimental study. Anim. Cogn. doi:10.1007/s10071-009-0298-9.
Christiansen, J.S., C.W. Mecklenburg, and O.V. Karamushko. 2014. Arctic marine fishes and their fisheries in light of global change. Global Change Biol. 20: 352–359, doi: 10.1111/gcb.12395 .
CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora). 2014. CITES Appendices I, II and III [accessed August 25, 2014].
Cody, M. 2003. Round Island field report, May 3 - August 10, 2003. Marine Mammals Management, U. S. Fish and Wildlife Service, Anchorage, Alaska. 67 pp.
Communities of Labrador, C. Furgal, M. Denniston, F. Murphy, D. Martin, S. Owens, S. Nickels, and P. Moss-Davies. 2005. Unikkaaqatigiit – putting the human face on climate change: perspectives from Labrador. Ottawa: Joint publication of Inuit Tapiriit Kanatimi, Nasivvik Centre for Inuit Health and Changing Environments at Université Laval and Ajunnginiq Centre at the National Aboriginal Health Organization. iv + 54 pp.
COSEWIC. 2006. COSEWIC assessment and update status report on the Atlantic Walrus Odobenus rosmarus rosmarus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa, ix + 65 pp.
Cronin, M.A., S. Hills, E.W. Born, and J.C. Patton. 1994. Mitochondrial DNA variation in Atlantic and Pacific Walrus. Can. J. Zool. 72: 1035–1043.
Crowe, K.J. 1969. A cultural geography of northern Foxe Basin. Northern Science Research Group: Department of Indian Affairs and Northern Development (NSRG) 69–2: xii + 130 p.
Currie, R.D. 1963. Western Ungava area economic survey. Canada Department of Northern Affairs and National Resources, Area Economic Survey Report 62–2: 103 pp.
Davis, R.A., Koski, W.R., and K.J. Finley. 1978. Numbers and distribution of Walrus in the central Canadian High Arctic. LGL Limited, Environmental Research Associates, 44 Eglinton Avenue West, Toronto, ON. vii + 50 pp.
Davis, R.A., Finley, K.J., and W.J. Richardson. 1980. The present status and future management of Arctic marine mammals in Canada. LGL Limited Environmental Research Associates, Toronto for Science Advisory Board of the Northwest Territories, Yellowknife, NWT. 93 pp.
de March, B.G.E., de Wit, C.A., and D.C.G. Muir (eds.). 1998. Persistent organic pollutants. Chapter 6, pp. 183–371 in AMAP Assessment Report: Arctic pollution issues. Arctic Monitoring and Assessment Programme (AMAP), Oslo, Norway.
de March, B.G.E., L.D. Maiers, and R.E.A. Stewart. 2002. Genetic relationships among Atlantic Walrus (Odobenus rosmarus rosmarus) in the Foxe Basin and Resolute Bay–Bathurst Inlet area. DFO Can. Sci. Advis. Sec. Res. Doc. 2002/92: 20 pp.
Degerbøl, M. and P. Freuchen. 1935. Mammals. Report of the Fifth Thule Expedition 1921–24. Gyldendalske Boghandel, Nordisk Forlag. 2(4–5): 1–278, Copenhagen.
DFO (Fisheries and Oceans Canada). 2002. Atlantic Walrus. DFO Can. Sci. Advis. Sec. Stock Status Rep. E5–17, 18, 19, 20: 19 pp.
DFO (Fisheries and Oceans Canada). 2008. Recovery Strategy for the Atlantic Walrus (Odobenus rosmarus rosmarus), Northwest Atlantic population, in Canada [Proposed]. Species at Risk Act Recovery Strategy Series. Department of Fisheries and Oceans. Ottawa. x + 11 pp.
DFO (Fisheries and Oceans Canada). 2013. Proceedings of the Pre-COSEWIC Peer Review Meeting for Atlantic Walrus (Odobenus rosmarus rosmarus); February 28 to March1, 2012. DFO Can. Sci. Advis. Sec. Proceed. Ser. 2012/041: iv + 29 p.
DFO (Fisheries and Oceans Canada). 2016. Estimates of abundance and total allowable removals for Atlantic Walrus (Odobenus rosmarus rosmarus) in Foxe Basin. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2016/007: 10 pp.
Dietz, R., E.W. Born, R.E.A. Stewart, M.P. Heide-Jørgensen, H. Stern, F. Rigét, L. Toudal, C. Lanthier, M.V. Jensen, and J. Teilmann, 2014. Movements of Walrus (Odobenus rosmarus) between central West Greenland and southeast Baffin Island, 2005-2008. NAMMCO Sci. Publ. 9: 53-74.
Donaldson, G.M., G. Chapdelaine, and J.D. Andrews. 1995. Predation of thick-billed murres, Uria lomvia, at two breeding colonies by polar bears, Ursus maritimus, and Walrus, Odobenus rosmarus. Can. Field-Nat. 109: 112–114.
Doniol-Valcroze, T., Mosnier, A., Hammill, M.O. 2016. Testing estimators of Walrus abundance: insights from simulations of haul-out behaviour. DFO Can. Sci. Advis. Sec. Res. Doc. 2016/040: v + 18 p.
Dueck, L. 2003. Proceedings of the RAP Meeting on Atlantic Walrus, 29–30 January, 2000, Navigator Inn, Iqaluit, NU. DFO Can. Sci. Advis. Sec. Proceed. Ser. 2002/024: 20 pp.
ESWG (Ecological Stratification Working Group). 1995. A national ecological framework for Canada. Agriculture and Argi-food Canada, Research Branch, Centre for Land and Biological Resources Research and Environment Canada, State of the Environment Directorate, Ecozone Analysis Branch, Ottawa/Hull. [Report and National Map at 1:7,500,000 scale.]
Fay, F.H. 1960. Carnivorous Walrus and some Arctic zoonoses. Arctic 13: 111–122
Fay, F.H. 1981. Walrus Odobenus rosmarus (Linnaeus, 1758). pp. 1-23, in S.H. Ridgway and R.J. Harrison (eds.) Handbook of Marine Mammals, Volume 1. The Walrus, sea lions, fur seals, and sea otter. Academic Press, London.
Fay, F.H. 1982. Ecology and biology of the Pacific Walrus, Odobenus rosmarus divergens Illiger. U.S. Department of the Interior, Fish and Wildlife Service, North American Fauna No. 74: vi + 279 pp.
Fay, F.H. 1985. Odobenus rosmarus. Mamm. Species 238: 1–7.
Fay, F.H. and B.P. Kelly. 1980. Mass natural mortality of Walrus (Odobenus rosmarus) at St. Lawrence Island, Bering Sea, Autumn 1978. Arctic 33: 226–245.
Fay, F.H. and C. Ray. 1968. Influence of climate on the distribution of Walrus, Odobenus rosmarus (Linnaeus). I. Evidence from thermoregulatory behavior. Zoologica 53: 1–18.
Fay, F.H., G.C. Ray, and A.A. Kibal’chich. 1984. Time and location of mating and associated behavior of the Pacific Walrus Odobenus rosmarus divergens Illiger. NOAA Tech. Rep. NMFS 12: 89-99.
Fay, F.H., J.J. Burns, A.A. Kibal'chich, and S. Hills. 1991. Incidence of twin fetuses in Walrus, Odobenus rosmarus L. Northwestern Naturalist 72: 110–113.
Ferguson, S.H, J.W. Higdon, and K.H. Westal. 2012. Prey items and predation behavior of killer whales (Orcinus orca) in Nunavut, Canada based on Inuit hunter interviews. Aquat. Biosys. 8(3): 16 pp. doi:10.1186/2046-9063-8-3.
Finley, K.J. and W.E. Renaud. 1980. Marine mammals inhabiting the Baffin Bay north water in winter. Arctic 33: 724–738.
Fischbach, A.S., D.H. Monson, and C.V. Jay. 2009. Enumeration of Pacific Walrus carcasses on beaches of the Chukchi Sea in Alaska following a mortality event, September 2009. U.S. Geological Survey Open-File Report 2009-1291: iv + 10 pp.
Fisher, K.I. 1989. Food habits and digestive efficiency in Walrus, Odobenus rosmarus. M.Sc. Thesis, University of Manitoba, Winnipeg, MB. x + 88 pp.
Fisher, K.I. and R.E.A. Stewart. 1997. Summer foods of Atlantic Walrus, Odobenus rosmarus rosmarus, in northern Foxe Basin, Northwest Territories. Can. J. Zool. 75: 1166–1175.
Fisher, K.I., R.E.A. Stewart, R.A. Kastelein, and L.D. Campbell. 1992. Apparent digestive efficiency in Walrus (Odobenus rosmarus) fed herring (Clupea harengus) and clams (Spisula sp.). Can. J. Zool. 70: 30–36.
Fisk, A.T., Hobbs, K., and D.C.G. Muir (eds.). 2003. Contaminant levels, trends and effects in the biological environment. Canadian Arctic Contaminants Assessment Report II. Canada Department of Indian Affairs and Northern Development, Ottawa, ON. xix + 175 pp.
Fleming, M. and S. Newton. 2003. Hudson Bay TEKMS Report on select Hudson Bay features. Prepared for consideration by Bruce Stewart for incorporation into Hudson Bay Ecosystem Overview, 24 pp.
Freeman, M.M.R. 1964. Observations on the Kayak-Complex, Belcher Islands, N.W.T. Natl Mus. Can. Bull. 194: 56–91.
Freitas, C., K.M. Kovacs, R.A. Ims, M.A. Fedak, C. Lydersen. 2009. Deep into the ice: over-wintering and habitat selection in male Atlantic Walrus. Mar. Ecol. Progr. Ser. 375: 247–261. doi: 10.3354/meps07725
Freuchen, P. 1921. Om hvalrossens forekomst og vandringer ved Grønlands vestkyst (Occurrence and migrations of the Walrus near the west coast of Greenland) . Dansk naturhistorisk forening i København. Videnskabelige meddelelser 72: 237-249. [In Danish. Translated to English in 1973: Can. Transl. Fish. Aquat. Sci. 2383. 14 pp.].
Gaden, A., and G.A. Stern. 2010. Temporal trends in beluga, narwhal and Walrus mercury levels: links to climate change. pp.197-216, in S. Ferguson, M. Mallory, and L. Loseto (ed.) A little less Arctic: top predators in the world’s largest northern inland sea, Hudson Bay. Springer, Dordrecht.
Gagnon, J. 2011. Walrus tourism back on. Nunavut News/North, Monday, May 16, 2011, p. 4. [Accessed April 2013].
Ganong, W.F. 1904. The Walrus in New Brunswick. Bull. Nat. Hist. Soc. New Brunswick II 5: 240-241.
Garlich-Miller, J.L. 1994. Growth and reproduction of Atlantic Walrus (Odobenus rosmarus rosmarus) in Foxe Basin, Northwest Territories, Canada. M.Sc. Thesis, Department of Zoology, University of Manitoba, Winnipeg, MB. 116 pp.
Garlich-Miller, J.L. and R.E.A. Stewart. 1998. Growth and sexual dimorphism of Atlantic Walrus (Odobenus rosmarus rosmarus) in Foxe Basin, Northwest Territories, Canada. Mar. Mamm. Sci. 14: 803–818.
Garlich-Miller, J.L. and R.E.A. Stewart. 1999. Female reproductive patterns and fetal growth of Atlantic Walrus (Odobenus rosmarus rosmarus) in Foxe Basin, Northwest Territories, Canada. Mar. Mamm. Sci. 15: 179–191.
Garlich-Miller, J.L., R.E.A. Stewart, B.E. Stewart, and E.A. Hiltz. 1993. Comparison of mandibular with cemental growth-layer counts for ageing Atlantic Walrus (Odobenus rosmarus rosmarus). Can. J. Zool. 71: 163–167.
Garlich-Miller, J.L., L.T. Quakenbush, and J.F. Bromaghin. 2006. Trends in age structure and productivity of Pacific Walrus harvested in the Bering Strait region of Alaska, 1952-2002. Mar. Mamm. Sci. 22:880-896.
Garlich-Miller, J., J.G. MacCracken, J. Snyder, R. Meehan, M.Myers, J.M. Wilder, E. Lance, and A. Matz. 2011. Status review of the Pacific Walrus (Odobenus rosmarus divergens). U.S. Fish and Wildlife Service, Anchorage, AK. vi + 155 pp.
Gaston, A.J., and H. Ouellet. 1997. Birds and mammals of Coats Island, N.W.T. Arctic 50(2): 101-118.
Gatehouse, J. 2012. Muskox on the menu as Nunavut encourages return to traditional foods. MacLean’s Magazine, Wednesday, August 15, 2012. [Accessed July 2013].
Gavrilchuk, K. and V. Lesage. 2014. Large-scale marine development projects (mineral, oil and gas, infrastructure) proposed for Canada’s North. Can. Tech. Rep. Fish. Aquat. Sci. 3069: viii + 84 pp.
George, J. 2008. Arctic Bay man dies from eating infected Walrus meat. Nunatsiaq News, Nunavut, November 28, 2008.
George, J. 2013. Hydro power plays into Hudson Bay wildlife entrapments: researcher. Nunatsiaq News April 8, 2013. [Accessed April 2013].
Gjertz, I. 1990. Walrus predation on seabirds. Polar Rec. 26: 317.
Gjertz, I. and Ø. Wiig. 1994. Past and present distribution of Walrus in Svalbard. Arctic 47: 34-42.
Gjertz, I., D. Griffiths, B.A. Krafft, C. Lydersen, and Ø. Wiig. 2001. Diving and haul-out patterns of Walrus Odobenus rosmarus on Svalbards. Polar Biol. 24: 314–319.
Goulet, G. Unpublished. Aboriginal Traditional Knowledge Assessment Report, prepared for the ATK Subcommittee of COSEWIC (Committee on the Status of Endangered Wildlife in Canada) on Atlantic Walrus Odobenus rosmarus rosmarus in Canada.
Grebmeier, J.M., J.E. Overland, S.E. Moore, E.V. Farley, E.C. Carmack, L.W. Cooper, K.E. Frey, J.H. Helle, F.A. McLaughlin, and S.L. McNutt. 2006. A major ecosystem shift in the northern Bering Sea. Science 311(10 March 2006): 1461-1464.
Green, D.M. 2005. Designatable units for status assessment of endangered species. Conserv. Biol. 19(6): 1813-1820.
Greendale, R.G. and C. Brousseau-Greendale. 1976. Observations of marine mammal migrations at Cape Hay, Bylot Island, during the summer of 1976. Can. Fish. Mar. Serv. Tech. Rep. 680: 25 pp.
Guinn, B. and D.B. Stewart. 1988. Marine mammals of central Baffin Island, Northwest Territories. Lands Directorate of Environment Canada and Northern Environment Directorate of Indian and Northern Affairs, Northern Land Use Information Series, Background Report No. 6: ii + 65 pp. + map.
Gunn, A., G. Arlootoo, and D. Kaomayak. 1988. The contribution of ecological knowledge of Inuit to wildlife management in the Northwest Territories. pp. 22-30, in M.M.R Freeman and L. N. Carbyn (eds.), Traditional knowledge and renewable resource management in northern regions. University of Alberta, Boreal Institute for Northern Studies, Occasional Publication 23.
Gustavson, K., P. Pearce, J. Knopp, and D. de Kerckhove. 2008. Socioeconomic analysis for Atlantic Walrus. Final report. Prepared by Jacques Whitford Limited Dartmouth NS and Westworth Associates Environmental Ltd., Edmonton AB for Fisheries and Oceans Canada, Sarnia, ON. ii + 45 pp. [Project No. 1021493]
Hall, P. 2003. CITES and the conservation of the Atlantic Walrus (Odobenus rosmarus rosmarus). CITES World 11: 5–6.
Hammill, M.O., P. Blanchfield, J.W. Higdon, D.B. Stewart, S.H Ferguson. 2016a. Estimating abundance and total allowable removals for Walrus in Foxe Basin. DFO Can. Sci. Advis. Sec. Res. Doc. 2016/014. iv + 20 p.
Hammill, M.O., A. Mosnier, J-F Gosselin, J.W. Higdon, D.B. Stewart, T. Doniol-Valcroze, S.H Ferguson and J.B. Dunn. 2016b. Estimating abundance and total allowable removals for Walrus in the Hudson Bay-Davis Strait and south and east Hudson Bay stocks during September 2014. DFO Can. Sci. Advis. Sec. Res. Doc. 2016/036. v + 37 p.
Hammill, M.O., Doniol-Valcroze, T., Mosnier, A. and Gosselin, J.-F. 2016c. Modelling Walrus population dynamics: A direction for future assessments. DFO Can. Sci. Advis. Sec. Res. Doc. 2016/050. v + 47 p.
Hantzsch, B.A. 1977. My life among the Eskimos: Baffinland journeys in the years 1909 to 1911. University of Saskatchewan, Mawdsley Memoir 3: xxii + 395 p. + map. [Translated and edited by L.H. Neatby].
Harrington, C.R. 1966. Extralimital occurrences of Walrus in the Canadian arctic. J. Mammal. 47: 506–513.
Harrington, C.R. 2008. The evolution of Arctic marine mammals. Ecol. Appl. 18(2) Suppl.: S23-S40.
Heinzig, L. 1996. Case reviews – botulism and trichinosis. EpiNorth 8(4): 7.
Heide-Jørgensen, M.P., M.L. Burt, R.G. Hansen, N.H. Nielsen, M. Rasmussen, S. Fossette, and H. Stern. 2013. The significance of the North Water polynya to Arctic top predators. Ambio 42: 596-610.
Heide-Jørgensen, M.P., K.L. Laidre, S. Fossette, M. Rasmussen, N.H. Nielsen, and R.G. Hansen. 2014. Abundance of Walrus in eastern Baffin Bay and Davis Strait. NAMMCO Sci. Publ. 9: 159-171.
Higdon, J.W., and S.H. Ferguson. 2009. Loss of Arctic sea ice causing punctuated change in sightings of killer whales (Orcinus orca) over the past century. Ecol. Appl. 19: 1365-1375.
Higdon, J.W., K.H. Westdal, and S.H. Ferguson. 2014. Distribution and abundance of killer whales (Orcinus orca) in Nunavut, Canada – an Inuit knowledge survey. J. Mar. Biol. Assoc. UK 94(6): 1293-1304.
Hill, M. 2003. GN tests for trichinosis infection in Walrus tongues. Nunatsiaq News January 31, 2003.
Hogan, G. 1986. 'White gold and train oil': the Walrus on P.E.I. The Island Magazine 20 (Fall/Winter): 19-22.
Ijjangiaq, M. 1990. Interviews IE-081 and IE-132. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Immaroitok, E. 1996. Interview IE-377. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Inukshuk, S. 1996. Interview IE-391. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
IUCN (International Union for Conservation of Nature). 2008. Odobenus rosmarus. In: IUCN 2012. IUCN Red List of Threatened Species. Version 2012.2
IUCN (International Union for Conservation of Nature). 2013. Guidelines for Using the IUCN Red List Categories and Criteria (Version PDF; 1.1 Mo). Version 10. Prepared by the Standards and Petitions Subcommittee. [Accessed August 2013].
Jay, C.V., M.S. Udevitz, R. Kwok, A.S. Fischbach, and D.C. Douglas. 2010. Divergent movements of Walrus and sea ice in the northern Bering Sea. Mar. Ecol. Progr. Ser. 407: 293-302.
Kappianaq, G. 1992. Interview IE-234. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Kappianaq, G. 1997. Interview IE-409. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Kasser, J.J., and J.E. Weidmer. 2012. The circumpolar Walrus population in a changing world. Prepared by University of Applied Sciences Van Hall Larenstein, Leeuwarden, The Netherlands for WWF-Netherlands. 68 pp. + appendices 39 pp.
Kastelein, R.A. and M.A. van Gaalen. 1988. The tactile sensitivity of the mystacial vibrissae of a Pacific Walrus (Odobenus rosmarus divergens), part 1. Aquat. Mamm. 14.3: 123–133.
Kastelein, R.A., S. Stevens, and P. Mosterd. 1990. The tactile sensitivity of the mystacial vibrissae of a Pacific Walrus (Odobenus rosmarus divergens), part 2: masking. Aquat. Mamm. 16.2: 78–87.
Kastelein, R.A., N.M. Schooneman, and P.R. Wiepkema. 2000. Food consumption and body weight of captive Pacific Walrus (Odobenus rosmarus divergens). Aquat. Mamm. 26: 175–190.
Kemp, W.B. 1976. Inuit land use in south and east Baffin Island. pp. 125–151. in M.M.R. Freeman (ed.) Inuit land use and occupancy project. Volume 1: Land use and occupancy. Supply and Services Canada, Ottawa, ON.
Killian, H.P.L. and I. Stirling. 1978. Observations on overwintering Walrus in the eastern Canadian High Arctic. J. Mammal. 59: 197–200.
Kingsley, M.C.S. 1998. Walrus, Odobenus rosmarus, in the Gulf and estuary of the St. Lawrence, 1992–1996. Can. Field-Nat. 112: 90–93.
Knutsen, L.Ø. and E.W. Born. 1994. Body growth in Atlantic Walrus (Odobenus rosmarus rosmarus) from Greenland. J. Zool. (Lond.) 234: 371–385.
Kopaq, M. 1987. Interview IE-017. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Kovacs, K.M. and D.M. Lavigne. 1992. Maternal investment in otariid seals and Walrus. Can. J. Zool. 70: 1953–1964.
Kroeker, K.J., R.L. Kordas, R.N. Crim, I.E. Hendriks, L. Ramajo, G.G. Singh C.M. Duarte, and J.P. Gattuso. 2013. Impacts of ocean acidification on marine organisms: quantifying sensitivities and interactions with warming. Global Change Biol. DOI: 10.1111/gcb.12179.
Kryukovaa, N.V., E.P. Kruchenkovab, and D.I. Ivanovc. 2012. Killer Whales (Orcinus orca) hunting for Walrus (Odobenus rosmarus divergens) near Retkyn Spit, Chukotka. Biology Bulletin 39 (9): 768–778.
Kupaaq, M. 1996. Interview IE-360. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Laidler, G.J., J.D. Ford, W.A. Gough, T. Ikummaq, A.S. Gagnon, S. Kowal, K. Qrunnut, and C. Irngaut. 2009. Travelling and hunting in a changing Arctic: assessing Inuit vulnerability to sea ice change in Igloolik, Nunavut. Climatic Change 94: 363–397. DOI 10.1007/s10584-008-9512-z.
Laidre, K.L., Stirling, I., Lowry, L.F., Wiig, O., Heide-Jørgensen, M.P., and Ferguson, S.H. 2008. Quantifying the sensitivity of Arctic marine mammals to climate-induced habitat change. Ecol. Appl. 18(2 Suppl): S97-S125.
Lambert, J. and Préfontaine, G. 1995. The Iceland scallop (Chlamys islandica) in Nunavik. Can. Tech. Rep. Fish. Aquat. Sci. 2071: vii + 40 p.
Lands Directorate. 1981. Nova Zembla Island, District of Franklin, Northwest Territories. Environment Canada and Indian and Northern Affairs Canada, Land Use Information Series Map 38A (1:250,000 scale).
Larrat, S., Simard, M., Lair, S., Bélanger, D., and Proulx, J.F. 2012. From science to action and from action to science: the Nunavik Trichinellosis Prevention Program. Int. J. Circumpolar Health. 71: 18595. doi: 10.3402/ijch.v71i0.18595.
Le Boeuf, B.J. 1986. Sexual strategies of seals and Walrus. New Sci. 1491: 36–39.
Levermann, N., A. Galatius, G. Ehlme, S. Rysgaard, and E.W. Born. 2003. Feeding behaviour of free-ranging Walrus with notes on apparent dextrality of flipper use. BMC Ecology 3: 9. doi:10.1186/1472-6785-3-9
LGL Limited and North/South Consultants Inc. 2011. Appendix 8A-2. Marine mammal baseline. In Baffinland Iron Mines Corporation. Mary River Project Final Environmental Impact Statement, February 2012, Vol. 8. Marine environment.
Lindqvist, C., L. Bachmann, L.W. Andersen, E.W., Born, U. Arnason, K.M. Kovacs, C. Lydersen, A.V. Abramov, and Ø. Wiig. 2009. The Laptev Sea Walrus Odobenus rosmarus laptevi: an enigma revisited. Zool. Scripta 38(2): 113-224.
Loring, E. 1996. The cost-benefit relations of modern Inuit hunting: the Kapuivimiut of Foxe Basin, N.W.T. Canada. M.Sc. Thesis, McGill University, Montreal, Québec. vii + 98 pp.
Loughrey, A.G. 1959. Preliminary investigation of the Atlantic Walrus Odobenus rosmarus rosmarus (Linnaeus). Can. Wildl. Serv. Bull. (Ott.) (Series 1) 14: 123 pp.
Lowry, L.F. and F.H. Fay. 1984. Seal eating by Walrus in the Bering and Chukchi Seas. Polar Biol. 3: 11–18.
Lowry, L.F. and K.J. Frost. 1981. Feeding and trophic relationships of phocid seals and Walrus in the eastern Bering Sea. pp. 813–824, in D.W. Hood and J.A. Calder (ed.) The eastern Bering Sea shelf: oceanography and resources, Vol. 2. University of Washington Press, Seattle, WA.
Lowry, L.F., R.R. Nelson, and K.J. Frost. 1987. Observations of killer whales, Orcinus orca, in western Alaska: sightings, strandings, and predation on other marine mammals. Can. Field-Nat. 101: 6–12.
Lowry, L., K. Kovacs, V. Burkanov (IUCN SSC Pinniped Specialist Group). 2008. Odobenus rosmarus. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.1. Web site: www.iucnredlist.org [Accessed August 2013].
Lydersen, C. and K. Kovacs. 2014. Walrus Odobenus rosmarus research in Svalbard, Norway, 2000-2010. NAMMCO Sci. Publ. 9: 175-190.
Lydersen, C., V.I. Chernook, D.M. Glazov, I.S. Trukhanova, and K.M. Kovacs. 2012. Aerial survey of Atlantic Walrus (Odobenus rosmarus rosmarus) in the Pechora Sea, August 2011. Polar Biol. 35: 1555-1562.
MacLaren Atlantic Limited. 1978. Report on aerial surveys 77–2, 77–3, 77–4 studies of seabirds and marine mammals in Davis Strait, Hudson Strait and Ungava Bay. Unpublished report prepared by MacLaren Atlantic Limited, Dartmouth, NS, for Imperial Oil Limited, Aquitaine Company of Canada, Ltd., and Canada Cities Service Ltd., Calgary, AB. variously paginated. (APOA Projects 134 & 138).
MacLaren Marex Inc. 1979. Report on aerial surveys of birds and marine mammals in the southern Davis Strait between April and December, 1978. Volume 3. Marine mammals. Unpublished report prepared by MacLaren Marex Inc., St. Johns, Newfoundland, for Esso Resources Canada Ltd., Aquitaine Company of Canada, Ltd., and Canada Cities Service Ltd., Calgary, AB. variously paginated. (APOA Project 146).
MacLaren Marex Inc. 1980a. Aerial monitoring of marine birds and mammals: the 1979 offshore drilling programme near southeast Baffin Island. Unpublished report prepared for ESSO Resources Canada Ltd. and Aquitaine Company of Canada Ltd. 88 pp.
MacLaren Marex Inc. 1980b. Surveys for marine mammals along the outer coastline of southeast Baffin Island (August to October 1979). Unpublished report prepared for ESSO Resources Canada Ltd. and Aquitaine Company of Canada Ltd.
MacLean, J.D., L. Poirier, T.W. Gyorkos, J.-F. Proulx, J. Bourgeault, A. Corriveau, S. Illisituk, and M. Staudt. 1992. Epidemiologic and serologic definition of primary and secondary trichinosis in the Arctic. J. Infect. Dis. 165: 908–912.
Mallory, M.L., K. Woo, A.J. Gaston, W.E. Davies, and P. Mineau. 2004. Walrus (Odobenus rosmarus) predation on adult thick-billed murres (Uria lomvia) at Coats Island, Nunavut, Canada. Polar Res. 23(1): 111-114.
Manning, T.H. 1961. Comments on "Carnivorous Walrus and some Arctic zoonoses". Arctic 14: 76–77.
Mansfield, A.W. 1959. The Walrus in the Canadian arctic. Fish. Res. Board Can. Arctic Unit Circ. 2: 1–13.
Mansfield, A.W. 1962. Present status of the Walrus population at Southampton and Coats islands. Fish. Res. Board Can., Arctic Unit, Annual Report and Investigators Summaries April 1, 1961 to March 31, 1962: 41–48.
Mansfield, A.W. 1967. Seals of Arctic and eastern Canada. Fish. Res. Board Can. Bull. 137: vii + 35 p. (Second edition, Revised).
Mansfield, A.W. 1966. The Walrus in Canada's arctic. Can. Geogr. J. 72: 88–95.
Mansfield, A.W. 1973. The Atlantic Walrus Odobenus rosmarus in Canada and Greenland. IUCN (Intl Union Conserv. Nat. Nat. Resour.) Publ. New Ser. Suppl. Pap. 39: 69–79.
Mansfield, A.W. and D.J. St. Aubin. 1991. Distribution and abundance of the Atlantic Walrus, Odobenus rosmarus rosmarus, in the Southampton Island-Coats Island region of northern Hudson Bay. Can. Field-Nat. 105: 95–100.
Martinez-Levasseur, L.M., C.M. Furgal, M.O. Hammill, and G. Burness. 2016. Towards a better understanding of the effects of UV on Atlantic Walrus, Odobenus rosmarus rosmarus: A study combining histological data with local ecological knowledge. PLoS ONE 11(4): e0152122. doi:10.1371/journal.pone.0152122
McFarland, S.E., and L.A.M. Aerts. 2015. Assessing disturbance responses of Pacific Walrus (Odobenus rosmarus divergens) to vessel presence in the Chukchi Sea (Abstract) (PDF version; 68.2 KB)). Chukchi Sea Environmental Studies Program (CSESP), Olgoonik-Fairweather, Fairweather Science, Anchorage, AK. [Accessed February 2017]
McLeod, B.A., T.R. Frasier, and Z. Lucas. 2014. Assessment of the extirpated Maritimes Walrus using morphological and ancient DNA analysis. PLoS ONE 9(6): e99569. doi:10.1371/journal.pone.0099569 .
Megannety, M. 2011. A review of the planned shipping activity for the Baffinland Mary River Project: assessing the hazards to marine mammals and migratory birds, and identifying gaps in proposed mitigation measures. Master of Marine Management thesis, Dalhousie University, Halifax, Nova Scotia. xviii + 137 pp.
Melnikov, V.V. and I.A. Zagrebin. 2005. Killer whale predation in coastal waters of the Chukotka Peninsula. Mar. Mamm. Sci. 21(3): 550-556.
Mercer, M.C. 1967. Records of the Atlantic Walrus, Odobenus rosmarus rosmarus, from Newfoundland. J. Fish. Res. Board Can. 24: 2631–2635.
Miller, E.H. 1975. Walrus ethology. I. The social role of tusks and applications of multidimensional scaling. Can. J. Zool. 53: 590–613.
Miller, E.H. 1976. Walrus ethology. II. Herd structure and activity budgets of summering males. Can. J. Zool. 54: 704–715.
Miller, E.H. 1982. Herd organization and female threat behavior in Atlantic Walrus Odobenus rosmarus rosmarus (L.). Mammalia 46: 29–34.
Miller, E.H. 1985. Airborne acoustic communication in the Walrus Odobenus rosmarus. Natl Geogr. Res. 1: 124–145.
Miller, E.H. and D.J. Boness. 1983. Summer behavior of Atlantic Walrus Odobenus rosmarus rosmarus (L.) at Coats Island, N.W.T. Z. Saeugetierkd. 48: 298–313.
Miller, R.F. 1997. New records and AMS radiocarbon dates on Quaternary Walrus (Odobenus rosmarus) from New Brunswick. Géogr. phys. Quat. 51: 1–5.
Moore, S.E., R.R. Reeves, B.L. Southall, T.J. Ragen, R.S. Suydam, and C.W. Clark. 2012. A new framework for assessing the effects of anthropogenic sound on marine mammals in a rapidly changing Arctic. BioScience 62(3): 289-295.
Muir, D.C.G., M.D. Segstro, K.A. Hobson, C.A. Ford, R.E.A. Stewart, and S. Olpinski. 1995. Can seal eating explain elevated levels of PCBs and organochlorine pesticides in Walrus blubber from eastern Hudson Bay (Canada). Environ. Pollut. 90: 355–348.
Muir, D., E.W. Born, K. Koczansky, and G.A. Stern. 2000. Temporal and spatial trends of persistent organochlorines in Greenland Walrus (Odobenus rosmarus rosmarus). Sci. Total Environ. 2000: 73–86.
NAMMCO. 2006. NAMMCO (North Atlantic Marine Mammal Commission) Scientific Committee Working Group on the stock status of Walrus in the North Atlantic and adjacent seas, Final Report. Copenhagen, 11-14 January 2005. 27 pp.
NAMMCO. 2011. Annual report for 2010. North Atlantic Marine Mammal Commission (NAMMCO), Tromsø, Norway. 501 pp.
NAMMCO. 2015. Report of the 22nd Scientific Committee meeting held 9-12 November 2015 in Tórshavn, Faroe Islands. Final Report. North Atlantic Marine Mammal Commission (NAMMCO), Tromsø, Norway. 196 pp. [Accessed September 2016]
NPC (Nunavut Planning Commission). 2016. Nunavut Land Use Plan. 2016 Draft (June). Nunavut Planning Commission, Cambridge Bay, NU. [Accessed September 2016]
Ndao, M. 2011. Trichinellosis: biology, epidemiology and clinical course. National Reference Center for Parasitology, McGill University Hospital Center, Department of Medicine – Division of Infectious Diseases, Montreal, Québec. Unpublished PowerPoint presentation 45 pp. [Accessed August 2013].
Nielsen, O., K. Nielsen, and R.E.A. Stewart. 1996. Serologic evidence of Brucella spp exposure in Atlantic Walrus (Odobenus rosmarus rosmarus) and ringed seals (Phoca hispida) of Arctic Canada. Arctic 49: 383–386.
Nielsen, O., R.E.A. Stewart, L. Measures, P. Duignan, and C. House. 2000. A morbillivirus antibody survey of Atlantic Walrus, narwhal and beluga in Canada. J. Wildl. Dis. 36: 508–517.
Nielsen, O., A. Clavijo, and J.A. Boughen. 2001a. Serologic evidence of influenza A infection in marine mammals of Arctic Canada. J. Wildl. Dis. 37: 820–825.
Nielsen, O., R.E.A. Stewart, K. Nielsen, L. Measures, and P. Duignan. 2001b. Serologic survey of Brucella spp. antibodies in some marine mammals of North America. J. Wildl. Dis. 37: 89–100.
Nielsen, O., Cobb, D., Stewart, R.E.A., Ryan, A., Dunn, B., Raverty, S., Nielsen, K. and Harwood, L. 2004. Results of a community based disease monitoring program of marine mammals in Arctic Canada. Proceedings of the Oceans 2004 Conference, Kobe, Japan: 1-7.
NIRB (Nunavut Impact Review Board). 2012. NIRB Project Certificate [No.: 005]. NIRB File No. 08MN053, December 28, 2012.
NIRB (Nunavut Impact Review Board). 2014. NIRB Project Certificate [NO.: 005, Amendment 1], (Amendment of Project Certificate to reflect modifications to the Project associated with the Early Revenue Phase), May 28, 2014, Nunavut Impact Review Board, Cambridge Bay, NU. 82 pp.
Norstrom, R.J. and D.C.G. Muir. 2000. Organochlorine contamination in the Arctic marine ecosystem: implications for marine mammals. pp. 38–43, in The Atlantic Coast Contaminants Workshop 2000 "Endocrine disruptors in the marine environment: impacts on marine wildlife and human health". The Marine Environmental Research Institute (MERI), University of Connecticut Department of Pathobiology, and Jackson Laboratory, Bar Harbor, Maine.
Nunatsiaq News. 2012a. Big Nunavik iron mine project heads into federal review. Posted online October 15, 2012, 10:15 am. [Accessed August 2013].
Nunatsiaq News. 2012b. Haig Inlet site could hold 230 million tonnes of iron ore. Posted February 7, 2012, 3:47 pm. [Accessed August 2013].
Nunatsiaq News. 2016. Nunavut-harvested Walrus meat tests positive for trichinella. Nunavut August 29, 2016. Web site: [Accessed September 2016]
Nunavut Department of Economic Development and Transportation. 2008. Nunavut coastal resource inventory – Iglulik Pilot Project. Government of Nunavut, Department of Economic Development and Transportation (now Department of Environment), Fisheries and Sealing Division, Iqaluit, NU. 197 pp.
Nunavut Department of Environment. 2010. Nunavut coastal resource inventory - Arctic Bay. Government of Nunavut, Department of Environment, Fisheries and Sealing Division, Iqaluit, NU. 168 pp.
Nunavut Planning Commission. 2000a. Keewatin Regional Land Use Plan (PDF version; 11.3 MB). Nunavut Planning Commission, Cambridge Bay, NU. 104 pp. [Accessed August 2013].
Nunavut Planning Commission. 2000b. North Baffin Regional Land Use Plan (PDF version; 4.5 MB). Nunavut Planning Commission, Cambridge Bay, NU. 124 pp. [Accessed August 2013]
Okonek, D.C. and M. Snively. 2005. Walrus Islands State Game Sanctuary annual report 2005. Alaska Department of Fish and Game, Division of Wildlife Conservation, Anchorage, Alaska. 46 pp.
Okonek, D.C., Okonek, B., and M. Snively. 2009. Walrus Islands State Game Sanctuary annual report 2008. Alaska Department of Fish and Game, Division of Wildlife Conservation, Anchorage, Alaska. 60 pp.
Okonek, D.C., S.K. Sell, and E.K. Weiss. 2010. Walrus Islands State Game Sanctuary annual management report 2009. Alaska Department of Fish and Game, Division of Wildlife Conservation, Special Areas Management Report. iii + 70 pp.
Olpinski, S. 1991. The 1990 Nunavik beluga whale and Walrus subsistence harvest study. Report submitted to: The Department of Fisheries and Oceans under subcontract to: Kativik Regional Government. v + 22 pp.
Olpinski, S. 1993. The 1992 Nunavik beluga whale and Walrus subsistence harvest study. Report submitted to: The Department of Fisheries and Oceans under subcontract to: The Municipal Corporation of Kuujjuaq, Kuujjuaq, Québec. vi + 36 pp.
Orr, J.R. and T. Rebizant. 1987. A summary of information on the seasonal distribution and abundance of Walrus (Odobenus rosmarus) in the area of northern Hudson Bay and western Hudson Strait, NWT, as collected from local hunters. Can. Data Rep. Fish. Aquat. Sci. 624: iv + 16.
Orr, J.R., B. Renooy, and L. Dahlke. 1986. Information from hunts and surveys of Walrus (Odobenus rosmarus) in northern Foxe Basin, Northwest Territories, 1982-1984. Can. Manuscr. Rep. Fish. Aquat. Sci. 1899: iv + 29 pp.
Outridge, P.M. and R.E.A. Stewart. 1999. Stock discrimination of Atlantic Walrus (Odobenus rosmarus rosmarus) in the eastern Canadian Arctic using lead isotope and element signatures in teeth. Can. J. Fish. Aquat. Sci. 56: 105–112.
Outridge, P.M., D.D. MacDonald, E. Porter, and I.A. Cuthbert. 1994. An evaluation of the ecological hazards associated with cadmium in the Canadian environment. Environ. Rev. 2: 91–107.
Outridge, P.M., R.D. Evans, R. Wagemann, and R.E.A. Stewart. 1997. Historical trends of heavy metals and stable lead isotopes in beluga (Delphinapterus leucas) and Walrus (Odobenus rosmarus) in the Canadian Arctic. Sci. Total Environ. 203: 209–219.
Outridge, P.M., K.A. Hobson, R. McNeely, and A. Dyke. 2002. A comparison of modern and preindustrial levels of mercury in the teeth of beluga in the Mackenzie Delta, Northwest Territories, and Walrus at Igloolik, Nunavut, Canada. Arctic 85: 123–132.
Outridge, P.M., W.J. Davis, R.E.A. Stewart, and E.W. Born. 2003. Investigation of the stock structure of Atlantic Walrus (Odobenus rosmarus rosmarus) in Canada and Greenland using dental Pb isotopes derived from local geochemical environments. Arctic 56: 82–90.
Ovsyanikov, N.G., I.E. Menyushina, and A.V. Bezrukov. 2007. Unusual Pacific Walrus mortality at Wrangel Island in 2007. Marine Mammals of the Holarctic, Odessa. pp. 414-416.
Panel Report (Federal Review Panel for the Eastmain-1-A Diversion Project). 2006. Environmental assessment of the Eastmain-1-A and Rupert Diversion Project. xxiv + iv + 486 pp.
Paniaq, H. 1998. Interview IE-431. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Paniaq, H. 2005. Interview IE-523. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Parkinson, C. L. 2014. Spatially mapped reductions in the length of the Arctic sea ice season. Geophys. Res. Lett., 41, doi:10.1002/2014GL060434.
Parkinson, C.L. and D.J. Cavalieri. 2008. Arctic sea ice variability and trends, 1979–2006. J. Geophys. Res.113(C7) doi: http://dx.doi.org/10.1029/2007JC004558
Parks Canada Agency. 2012. Corporate plan 2012-2013/2016-2017. Parks Canada, Gatineau, QC. v + 36 pp. [Catalogue No.: R61-12/2017E-PDF].
Patterson, G. 1891. The Magdalene Islands. Proc. Trans. Nova Scotian Inst. Nat. Sci. 8(1): 31-57. [Available online at: http://hdl.handle.net/10222/12380]
Perley, M.H. 1850. Reports on the sea and river fisheries of New Brunswick. J. Simpson, Fredericton, NB. ix + 137 pp.
Phillipa, J.D.W., F.A. Leighton, P.Y. Daoust, O. Nielsen, M. Pagliarulo, H. Schwantje, T. Shury, R. van Herwijnen, B.E.E. Martina, T. Kuiken, M.W.G. Van de Bildt, and A.D.M.E. Osterhaus. 2004. Antibodies to selected pathogens in free-ranging terrestrial carnivores and marine mammals in Canada. Veterinary Record 155: 135-140. doi:10.1136/vr.155.5.135.
Piepenburg, D. 2005. Recent research on Arctic benthos: common notions need to be revised, Polar Biol. 28(10): 733–755. doi: 10.1007/s00300-005-0013-5
Pianka, E.R. 1988. Evolutionary ecology. 4th edn. Harper and Row, NY. ix + 468 pp.
Piugattuk, N. 1986. Interview IE-010. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Piugattuk, N. 1990. Interview IE-136. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Portnoff, M. 1994. The 1993 Nunavik beluga whale and Walrus subsistence harvest study. An unpublished report prepared for the Aboriginal Fisheries Strategy of the Department of Fisheries and Oceans, Institut Maurice-Lamontagne, Mont-Joli, QC. iv + 61 p.
Pozio, E., G. LaRosa, K.D. Murrell, and J.R. Lichtenfels. 1992. Taxonomic revision of the genus Trichinella. J. Parasitol. 78: 654–659.
Priest, H., and P.J. Usher. 2004. The Nunavut wildlife harvest study, August 2004, final report. Nunavut, Canada. 822 pp.
Qamaniq, N. 1999. Interview IE-420. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Qikiqtani Inuit Association. 2011. Conformity Technical Review – October 5, 2011, QIA submission of Technical Review Comments titled “Submission of Technical Review Comments”. Appendix D: Fresh water aquatics and marine, 96 pp.
Qikiqtani Inuit Association. 2012. Qikiqtani Inuit Association’s final written submission for Baffinland Iron Mines Corporation, Mary River Project, Final Environmental Impact Statement. Submitted May 30th, 2012 to the Nunavut Impact Review Board (NIRB). 40 pp + appendices.
Qikiqtani Inuit Association. 2013. Qikiqtani Inuit Association’s Technical Review Submission for Baffinland Iron Mines Corporation, Mary River Project, Addendum to the Final Environmental Impact Statement. Submitted October 18, 2013 to the Nunavut Impact Review Board. 176 pp. (variously paginated).
Qikiqtani Inuit Association. 2014. Qikiqtani Inuit Association’s final written submission for Baffinland Iron Mines Corporation, Mary River Project, Addendum to the Final Environmental Impact Statement. Submitted January 13, 2014 to the Nunavut Impact Review Board. 110 pp.
Ray, C. and F.H. Fay. 1968. Influence of climate on the distribution of Walrus, Odobenus rosmarus (Linnaeus). II. evidence from physiological characteristics. Zoologica 53: 19–32.
Read, C.J. and S.E. Stephansson. 1976. Distribution and migration routes of marine mammals in the central Arctic region. Can. Fish. Mar. Serv. Tech. Rep. 667: v + 13 pp.
Reeves, R.R. 1978. Atlantic Walrus (Odobenus rosmarus rosmarus): a literature survey and status report. U.S. Department of the Interior, Fish and Wildlife Service, Wildlife Research Report (Washington, D.C.) 10: ii + 41 pp.
Reeves, R.R. 1995. Walrus of Nunavik. Prepared for Canada Department of Fisheries and Oceans under the Québec Federal Fisheries Development Program (QFFDP), Québec. x + 48 pp.
Reeves, R.R. and E. Mitchell. 1986. Early abundance and distribution of Walrus in eastern Hudson Bay, Hudson Strait, and Ungava Bay, based on historical records: preliminary report. Report to Fisheries and Oceans Canada in fulfillment of contract 12SD.FP 715-5-2664. 64 pp.
Richard, P.R. 1993. Summer distribution and abundance of Walrus in northern Hudson Bay, western Hudson Strait, and Foxe Basin: 1988–1990. Background document prepared for the Arctic Fisheries Science Advisory Meeting, 17–18 February 1993 by Canada Department of Fisheries and Oceans, Central and Arctic Region, Winnipeg, MB. 11 pp. + figures.
Richard, P.R. and R.R. Campbell. 1988. Status of the Atlantic Walrus, Odobenus rosmarus rosmarus, in Canada. Can. Field Nat. 102: 337-350.
Richard, P., J.R. Orr, R. Dietz, and L. Dueck. 1998. Sightings of belugas and other marine mammals in the North Water, late March 1993. Arctic 51: 1–4.
Richer, S. 2003. Nova Scotians awake to find a rare Walrus in their midst. Globe and Mail Thursday June 12th: A3 (illustrated).
Riewe, R.R. 1976. Inuit land use in the High Arctic. pp. 173–184, in M.M.R. Freeman (ed.) Inuit land use and occupancy study, Vol. 1. Canada Department of Indian and Northern Affairs, Ottawa.
Riewe, R.R. and C.W. Amsden. 1979. Harvesting and utilization of pinnipeds by Inuit hunters in Canada's eastern High Arctic. pp. 324–348, in A.P. McCartney (ed) Thule Eskimo culture: an anthropological retrospective. Nat. Mus. Man. Archaeol. Surv. Can. Pap. 88 (Mercury Series).
Rogers, S. 2013. Nunavik public health: Trichinellosis hits 15 people in Inukjuak . Nunatsiaq News Online, Nunavik October 23, 2013. [Accessed September 2016]
Rogers, S. 2015. Nunavut Walrus tests positive for Trichinella, GN warns. Nunatsiaq News online, Nunavut July 30, 2015. [Accessed September 2016]
Roy, C. 1971. La chasse des mammifères marins chez les Ivujivimmiut (The Ivujivimmiut hunt for marine mammals). Cahiers de Géographie du Québec 15: 509–521.
Sahanatien, V. and A.E. Derocher. 2012. Monitoring sea ice habitat fragmentation for polar bear conservation. Anim. Conserv.15(4): 397-407. doi:.
Salter, R.E. 1979a. Site utilization, activity budgets, and disturbance responses of Atlantic Walrus during terrestrial haul-out. Can. J. Zool. 57: 1169–1180.
Salter, R.E. 1979b. Observations on social behaviour of Atlantic Walrus (Odobenus rosmarus (L.)) during terrestrial haul-out. Can. J. Zool. 58: 461–463.
Saul, B., C. Notpol, D. Foster, and 16 others. 2012. Feasibility study on the Roche Bay Iron Ore Project. Report prepared by Tetra Tech Wardrop, Toronto for Advanced Explorations Inc. variously paginated [Document No. 1191360400-REP-R0019-03, 509 pp.]
Schwartz, F.H. 1976. Inuit land use in Hudson Bay and James Bay. pp. 115–120, in M.M.R. Freeman (ed.) Inuit Land Use and Occupancy Project, Volume 1. Milton Freeman Research Limited for the Department of Indian and Northern Affairs, Ottawa, ON.
Schwartz, F.H. 1982. Native land use in the Lancaster Sound area. Can. Dep. Ind. N. Aff. Environ. Stud. 27: 47 pp. + map.
Serhir, B., J.D. MacLean, S. Healey, B. Segal, and L. Forbes. 2001. Outbreak of trichinellosis associated with Arctic Walrus in northern Canada, 1999. Health Canada, Population and Public Health Branch, Canada Communicable Disease Report 27–04: 6 pp.
Shadbolt, T., T. Arnbom, and E.W.T. Cooper. 2014. Hauling Out: International Trade and Management of Walrus. TRAFFIC and WWF-Canada. Vancouver, B.C. viii + 166 pp.
Shafer, A.B.A., C.S. Davis, D.W. Coltman, and R.E.A. Stewart. 2014. Microsatellite assessment of Walrus (Odobenus rosmarus rosmarus) stocks in Canada. NAMMCO Sci. Publ. 9: 15-31.
Sheffield, G., F.H. Fay, H. Feder, and B.P. Kelly. 2001. Laboratory digestion of prey and interpretation of Walrus stomach contents. Mar. Mamm. Sci. 17(2): 310-330.
Shuldham, M. 1775. Account of the sea-cow and the use made of it. Philos. Trans. Roy. Soc. Lon. 65: 249-251.
Siakuluk, N. 1996. Interview IE-385. Archives of the Inullariit Society, Igloolik Research Centre, Igloolik, Nunavut.
Sjare, B. and I. Stirling. 1996. The breeding behavior of Atlantic Walrus, Odobenus rosmarus rosmarus, in the Canadian High Arctic. Can. J. Zool. 74: 897–911.
Sjare, B., I. Stirling, and C. Spencer. 2003. Structural variation in the songs of Atlantic Walrus breeding in the Canadian High Arctic. Aquat. Mamm. 29.2: 297–318.
Smith, T.G., M.H. Hammill, D.W. Doidge, T. Cartier, and G.A. Sleno. 1979. Marine mammal studies in southeastern Baffin Island. Final report to the eastern Arctic Marine Environmental Studies (EAMES) project. Can. Manuscr. Rep. Fish. Aquat. Sci. 1552: 70 pp.
Sobey, D. G. 2007. An analysis of the historical records for the native mammalian fauna of Prince Edward Island. Can. Field-Nat. 121(4): 384-396.
Stewart, B.E. (ed). 1993. The Walrus International Technical and Scientific Committee's bibliography on Walrus, Odobenus rosmarus (L.), to January, 1993. Can. Tech. Rep. Fish. Aquat. Sci. 1923: iv + 191 pp.
Stewart, B.E. and P.M. Burt. 1994. Extralimital occurrences of beluga, Delphinapterus leucas, and Walrus, Odobenus rosmarus, in Bathurst Inlet, Northwest Territories. Can. Field-Nat. 108: 488–490.
Stewart, D.B. and A.L. Hamilton. 2007. Outcomes of the Community Environmental Monitoring Systems (CEMS) Workshop at Sanikiluaq, Nunavut, 22-26 January 2007. Prepared by Arctic Biological Consultants, Winnipeg for the Municipality of Sanikiluaq, Nunavut. iv + 26 p.
Stewart, D.B. and K.L. Howland. 2009. An ecological and oceanographical assessment of the alternate ballast water exchange zone in the Hudson Strait Region. DFO Can. Sci. Advis. Sec. Res. Doc. 2009/008: vi +92 p.
Stewart, D.B. and W.L. Lockhart. 2005. An overview of the Hudson Bay marine ecosystem. Can. Tech Rep. Fish. Aquat. Sci. 2586: vi + 487 p.
Stewart, D.B., R.A. Ratynski, L.M.J. Bernier, and D.J. Ramsey. 1993. A fishery development strategy for the Canadian Beaufort Sea–Amundsen Gulf area. Can. Tech. Rep. Fish. Aquat. Sci. 1910: vi + 127 p.
Stewart, D.B., J.W. Higdon, R.R. Reeves, and R.E.A. Stewart. 2014a. Catch history for Atlantic Walrus (Odobenus rosmarus rosmarus) in the eastern Canadian Arctic. NAMMCO Sci. Publ. 9: 219-313.
Stewart, D.B., J.W. Higdon, and R.E.A. Stewart. 2014b. Development threats and effects pathways of shipping related to non-renewable resource developments on Atlantic Walrus (Odobenus rosmarus rosmarus) in Hudson Strait and Foxe Basin. Prepared by Arctic Biological Consultants Winnipeg and Higdon Wildlife Consulting, Winnipeg for Fisheries and Oceans Central and Arctic Region. ix + 56 pp.
Stewart, E.J., S.E.L. Howell, D. Draper, J. Yackel, and A. Tivy. 2007. Sea ice in Canada’s Arctic: implications for cruise tourism. Arctic 60(4): 370-380.
Stewart, E.J., A. Tivy, S.E.L. Howell, J. Dawson, and D. Draper. 2010. Cruise tourism and sea ice in Canada’s Hudson Bay region. Arctic 63(1): 57-66.
Stewart, J. 1806. An account of Prince Edward Island, in the Gulph of St. Lawrence, North America. W. Winchester and Son, Strand, London. 304 pp.
Stewart, R.E.A. 2002. Review of Atlantic Walrus (Odobenus rosmarus rosmarus) in Canada. Canadian Science Advisory Secretariat Research Document 2002/091: 20 pp.
Stewart, R.E.A. 2008. Refining Walrus stocks in Canada. Arctic 61(3): 292-308.
Stewart, R.E.A., and J.W. Hamilton. 2013. Estimating total allowable removals for Walrus (Odobenus rosmarus rosmarus) in Nunavut using the potential biological removal approach. DFO Can. Sci. Advis. Sec. Res. Doc. 2013/031: iv + 13 pp.
Stewart, R.E.A., and B.E. Stewart. 2005. Comparison of between-tooth age estimates of Atlantic Walrus (Odobenus rosmarus rosmarus) Mar. Mamm. Sci. 21(2): 346–354.
Stewart, R.E.A., P.R. Richard, and B.E. Stewart (ed). 1993. Report of the 2nd Walrus International Technical and Scientific (WITS) Workshop, 11–15 January 1993, Winnipeg, Manitoba, Canada. Can. Tech. Rep. Fish. Aquat. Sci. 1940: viii + 91 pp.
Stewart, R.E.A., P.M. Outridge, and R.A. Stern. 2003. Walrus life-history movements reconstructed from lead isotopes in annual layers of teeth. Mar. Mamm. Sci. 19: 806–818.
Stewart, R.E.A., V. Lesage, J.W. Lawson, H. Cleator and K.A. Martin. 2012. Science Technical Review of the draft Environmental Impact Statement (EIS) for Baffinland’s Mary River Project. DFO Can. Sci. Advis. Sec. Res. Doc. 2011/086. vi + 62 p.
Stewart, R.E.A., J.W. Hamilton, and J.B. Dunn. 2013. Results of Foxe Basin Walrus (Odobenus rosmarus rosmarus) surveys: 2010-2011. DFO Can. Sci. Advis. Sec. Res. Doc. 2013/017. iv + 12 p. (Erratum: February 2014)
Stewart, R.E.A., E.W. Born, J.B. Dunn, W.R. Koski, and A.K. Ryan. 2014a. Use of multiple methods to estimate Walrus (Odobenus rosmarus rosmarus) abundance in the Penny Strait-Lancaster Sound and west Jones Sound stocks, Canada. NAMMCO Sci. Publ. 9: 95-122.
Stewart, R.E.A., E.W. Born, R. Dietz, M.P. Heide-Jørgensen, F.F. Rigét, K. Laidre, M. Villum Jensen, L.Ø. Fossette, and J.B. Dunn. 2014b. Abundance of Atlantic Walrus in in western Nares Strait, Baffin Bay stock, during summer. NAMMCO Sci. Publ. 9: 123-140.
Stewart, R.E.A., E.W. Born, R. Dietz, and A.K. Ryan. 2014c. Estimates of minimum population size for Walrus near southeast Baffin Island, Nunavut. NAMMCO Sci. Publ. 9: 141-157.
Stewart, R.E.A., D.B. Stewart, and J.W. Higdon. in press. Report of the 2016 workshop to inform the DFO 5-year research plan for Atlantic Walrus in Canada. Can.Tech. Rep. Fish. Aquat. Sci. 3200: 45 pp.
Stirling, I. and J.A. Thomas. 2003. Relationships between underwater vocalizations and mating systems in phocid seals. Aquat. Mamm. 29.2: 227–246.
Stirling, I., W. Calvert, and H. Cleator. 1983. Underwater vocalizations as a tool for studying the distribution and relative abundance of wintering pinnipeds in the High Arctic. Arctic 36: 262–274.
Stirling, I., W. Calvert, and C. Spencer. 1987. Evidence of stereotyped underwater vocalizations of male Atlantic Walrus (Odobenus rosmarus rosmarus). Can. J. Zool. 65: 2311–2321.
Thiemann, G.W., S.M. Budge, S.J. Iverson, and I. Stirling. 2007. Unusual fatty acid biomarkers reveal age- and sex-specific foraging in polar bears (Ursus maritimus). Can. J. Zool. 85: 505-517.
Thiemann, G.W., S.J. Iverson, and I. Stirling. 2008. Polar bear diets and Arctic marine food webs: insights from fatty acid analysis. Ecological Monographs 78(4): 591-613.
Trent University and Makivik Corporation. 2015. Using Nunavimmiut Knowledge and Science to study Walrus in Nunavik, Final Report (2013-2015) Inukjuak. Report prepared by L. Martinez-Levasseur, C. Furgal, G. Burness, B. Doidge, M. Simard, and the communities of Inukjuak, Ivujivik, Quaqtaq, Kangiqsualujjuaq, Nunavik, QC. 11 pp.
Tsuji, L.J.S., N. Gomez, J.X. Mitrovica, and R. Kendall. 2009. Post-glacial isostatic adjustment and global warming in Subarctic Canada: implications for islands of the James Bay Region. Arctic 62 (4): 458–467.
USFWS (U.S. Fish and Wildlife Service). 2014. Stock Assessment Report - Pacific Walrus (Odobenus rosmarus divergens): Alaska Stock. Revised: April 2014. U.S. Fish and Wildlife Service, Anchorage, AK. 30 pp. [FWS-R7-ES-2012-0019-0003]
Viallet, J., J.D. MacLean, C.A. Goresky, M. Staudt, G. Routhier, and C. Law. 1986. Arctic trichinosis presenting as prolonged diarrhea. Gastroenterology 91: 938-946.
Vibe, C. 1950. The marine mammals and the marine fauna in the Thule District (northwest Greenland) with observations on ice conditions in 1939–41. Medd. Grønl. 150: 154 pp.
Vibe, C. 1967. Arctic animals in relation to climatic fluctuations. Medd. Grønl. 170: 227 pp.
Wagemann, R. and R.E.A. Stewart. 1994. Concentrations of heavy metals and selenium in tissues and some foods of Walrus (Odobenus rosmarus rosmarus) from the eastern Canadian Arctic and sub-Arctic, and associations between metals, age, and gender. Can. J. Fish. Aquat. Sci. 51: 426–436.
Wagemann, R., W.L. Lockhart, H. Welch, and S. Innes. 1995. Arctic marine mammals as integrators and indicators of mercury in the arctic. Water Air Soil Pollut. 80: 683–693.
Warburton, A.B. 1903. The sea-cow fishery. Acadiensis 3: 116-119.
Webber, P.J., J.W. Richardson, and J.T. Andrews. 1970. Post-glacial uplift and substrate age at Cape Henrietta Maria, southeastern Hudson Bay, Canada. Can. J. Earth Sci. 7: 317–325.
Welch, H.E. and K. Martin-Bergmann. 1990. Does the clam Mya truncata regenerate its siphon after predation by Walrus? Arctic 43: 157–158.
Westdal, K.H., J.W. Higdon, and S.H. Ferguson. 2013. Attitudes of Nunavut Inuit toward Killer Whales (Orcinus orca). Arctic 66(3): 279 – 290.
Wiig, Ø., E.W. Born, and R.E.A. Stewart. 2014. Management of Atlantic Walrus (Odobenus rosmarus rosmarus) in the Arctic Atlantic. NAMMCO Sci. Publ. 9: 315-342.
Witting, L. and E. Born. 2005. An assessment of Greenland Walrus populations. ICES J. Mar. Sci. 62: 266–285.
Witting, L. and E. Born. 2014. Population dynamics of Walrus in Greenland. NAMMCO Sci. Publ. 9: 191-218.
Xstrata Nickel. 2011. Xstrata Nickel Raglan Mine invests in the development of new mining projects in Nunavik. News Release, Rouyn-Noranda, August 2, 2011. 2 pp. [Accessed August 2013].
Biographical summary of report writers
D. Bruce Stewart (MSc), Head of Arctic Biological Consultants, has studied aquatic ecosystems throughout northern Canada since 1976. He has conducted many aquatic field studies and has worked with Inuit on the land studying Arctic Char and in the communities compiling traditional knowledge of marine mammals. Bruce has also provided expert advice on aquatic resources, species at risk, parks initiatives, and resource developments to government, industry, Inuit, and First Nations. In 1991-1993 he was appointed to the Canada-Manitoba Joint Review Panel for the Conawapa Hydroelectric Project. More recently, he has advised the Mackenzie Valley Environmental Impact Review Board on potential aquatic impacts of the Taltson Hydroelectric Expansion Project, and the Qikiqtani Inuit Association on the Mary River Iron Mine Project. Examples of his review work include the COSEWIC status updates for Narwhal and Bearded Seal and An Overview of the Hudson Bay Marine Ecosystem. Bruce has written over 120 publications, reports and articles. Recent works include a catch history for Atlantic Walrus in Canada, a recovery strategy for Lake Sturgeon in Alberta, and an assessment of alternate ballast water exchange zones in the eastern Canadian Arctic.
Jeff W. Higdon is a consulting marine mammal scientist, with a PhD on the biogeography of world pinnipeds and the influence of evolutionary adaptations to sea ice on the distribution patterns of polar species. Since 2005, he has conducted extensive field research on Arctic whales, seals and Walrus, including photo-identification, biopsy sampling, live-capture and tagging, hunt sampling, and hunt monitoring. Other research projects have involved collecting and interpreting Aboriginal traditional knowledge, historical research on marine mammal hunting, spatial analysis of animal movements and habitat selection, and assessments of potential environmental impacts to marine biota from proposed development projects and Arctic shipping. His recent work has focused on species at risk and Arctic monitoring programs, including development of protocols for protected areas monitoring and community-based monitoring. Jeff has worked for government, Inuit, and conservation organizations and written over 40 peer-reviewed scientific papers, book chapters and technical reports.
Personal communications/authorities contacted
Alooloo, J. 2013. Pond Inlet, NU, X0A 0S0.
Bishop, M. 2013. Makkovik, NL, A0P 1J0.
Cabana, Anne Marie. 2016. Aboriginal Programs Agent, Fisheries and Oceans Canada, 104 Dalhousie St., Quebec, QC G1K 7Y7.
Chenier, C. 2003 and 2014. Area Biologist - Cochrane District, Ontario Ministry of Natural Resources and Forestry, P.O. Box 730, Cochrane, ON, P0L 1C0.
Currie, A. 2012. Fisheries and Oceans Canada, Eastern Arctic Area Office, Box 358, Iqaluit, NU, X0A 0H0.
Desrosiers, J. 2003. Biologist. 470, Dolbeau, Québec, QC, G1S 2R5.
Dunn, B. 2014. Marine Mammal Research Technician, Fisheries and Oceans Canada, 501 University Crescent, Winnipeg, MB, R3T 2N6
Fitzsimmons, J. 2014. Species at Risk Biologist, Ontario Ministry of Natural Resources and Forestry, 300 Water Street, Peterborough, ON. K9J 8M5.
Galipeau, J. 2004. Conservation/Management Biologist, Nunavut Wildlife Management Board, P.O. Box 1379, Iqaluit, NU, X0A 0H0.
Gilbert, G. 2014. Resource Management Coordinator, Makivik Corporation, 1111 Dr. Frederik-Philips Blvd., 3rd Floor, Ville St Laurent, QC, H4M 2X6.
Girard, C. 2012. Treaty program Officer, Resource and Aboriginal Fisheries Management, Fisheries and Oceans Canada, 104 Dalhousie St., Québec, QC, G1K 7Y7.
Gosselin, J-F. 2014. Biologist, Maurice Lamontagne Institute, Fisheries and Oceans Canada, P.O. Box 1000, 850 route de la Mer, Mont-Joli, QC, G5H 3Z4.
Hamilton, J. 2014. General Delivery, Matlock, MB, R0C 2B0.
Hammill, M. 2013. Research Scientist, Maurice Lamontage Institute, Fisheries and Oceans Canada, P.O. Box 1000, 850 route de la Mer, Mont-Joli, QC, G5H 3Z4.
Irngaut, D. 2010. Igloolik, NU, X0A 0L0.
Kattuk, P. 1993. Mayor of Sanikiluaq, Sanikiluaq, NU, X0A 0W0.
Kotierk, M. 2013. Igloolik, NU. X0A 0L0.
Laforest, B. 2016. WWF-Canada, P.O. Box 1750, Iqaluit, NU XOA OHO.
McPhee, A. 2013, 2016, 2017. Regional Senior Officer-Resource Management and Aboriginal Affairs, Fisheries and Oceans Canada, 501 University Crescent, Winnipeg, MB, R3T 2N6.
Mills, K. 2007. Wyoming Game and Fish Department, PO Box 1353, 432 E. Mill Street, Pinedale, WY, USA 82941.
Novalinga, Z. 1993. Environmental Committee, Sanikiluaq, NU, X0A 0W0.
O’Connor, M. 2014. Nunavik Marine Region Wildlife Board, P.O. Box 433, Inukjuak, QC, J0M 1M0.
Obbard, M. 2014. Research Scientist, Wildlife Research and Monitoring Section, Ontario Ministry of Natural Resources and Forestry, and Adjunct Professor, Environmental and Life Sciences, DNA Building, Trent University, 2140 East Bank Drive, Peterborough, ON K9J 7B8.
Olpinski, S. 2014. Science and Policy Advisor, Makivik Corporation, 1111 Dr. Frederik-Philips Blvd., 3rd Floor, Ville St Laurent, QC, H4M 2X6.
Pattimore, J. 1986. formerly Harvest Coordinator, Baffin Region Inuit Association, Iqaluit, NU, X0A 0H0.
Petersen, S.D. Assiniboine Park Zoo, 2595 Roblin Blvd., Winnipeg, MB R3R 0B8.
Qanatsiaq, S. 2010. Igloolik, NU, X0A 0L0.
Risley, C. 2014. Species at Risk Avian and Marine Specialist, Ontario Ministry of Natural Resources and Forestry, 300 Water Street, Peterborough, ON. K9J 8M5.
Sjare, B. 2005, 2013. Research Scientist, Marine Mammals Section, Fisheries and Oceans Canada, Northwest Atlantic Fisheries Centre, White Hills, P.O. Box 5667, St. John’s, NL, A1C 5X1.
Stewart, R.E.A. 2016. Sila Consultants, 1218 Marchand Road, Howden, MB, R5A 1J6
Tilochand, A. 2012. Certification Officer, Catch Certification Program: Operations Centre, Fisheries and Oceans Canada, 100 School Street, P.O. Box 99, Tiginish, PE, C0B 2B0.
Turgeon, S. 2014. Biologist, Maurice Lamontagne Institute, Fisheries and Oceans Canada, P.O. Box 1000, 850 route de la Mer, Mont-Joli, QC, G5H 3Z4.
Ugarte, F. 2016. Greenland Institute of Natural Resources, P.O. Box 570, DK-3900 Nuuk, Greenland.
Whitaker, D. 2017. Parks Canada, PO Box 130, 3 D.O.T. Dr., Rocky Harbour, Newfoundland and Labrador A0K 4N0.
Young, J. 2016. Fisheries Management Technician, Fisheries and Oceans Canada, P.O. Box 358, Iqaluit, NU X0A 0H0.
| Participant | Affiliation / Community |
|---|---|
| Akkuardjuk, Michel | Arviq Hunters and Trappers Organization (HTO) (Repulse Bay) |
| Arlooktoo, Kiponik | Mayukalik HTO (Kimmirut) |
| Arreak, Lazarus (Interpreter) | Innirvik Support Services Ltd., |
| Bowen, Don (Chair) | DFO (Science, Maritimes region) |
| Cleator, Holly | DFO (Science, Central and Arctic region) |
| Currie, Amanda | DFO (Fisheries Management, Central and Arctic region) |
| Curtis, Martyn | DFO (SARA, Central and Arctic region) |
| Delisle-Alaku, Adamie | Makivik Corporation |
| Hamilton, Jason | DFO (Science, Central and Arctic region) |
| Hidgon, Jeff | COSEWIC status report co-author |
| Idlout, Simon | Resolute HTO |
| Ikkidluak, Elisapee (Interpreter) | Innirvik Support Services Ltd |
| Irngaut, David | Igloolik HTO |
| Irngaut, Paul | Nunavut Tunngavik Inc. |
| Kango, Joshua | Amaruq HTO (Iqaluit) |
| Kaunak, Levi | Hall Beach HTO |
| Kilabuk, Patrick | Pangnirtung HTO |
| Kimmaliardjuk, Eli | Aqigiq HTO (Chesterfield Inlet) |
| Kruger, Lia | DFO (Science, Central and Arctic region) |
| Magera, Anna | Nunavut Wildlife Management Board |
| Martin, Kathleen | DFO (Science, Central and Arctic region) |
| Natanine, Jerry | Nangmautaq HTO (Clyde River) |
| Newkingnak, Toomasie | Nattivak HTO (Qikiqtarjuaq) |
| Ningiuk, Joanassie | DFO (Fishery Officer, Québec region, Inukjuak) |
| Nirlungayuk, Gabriel | Nunavut Tunngavik Inc. |
| Noah, Charlie | Iviq HTO (Grise Fiord) |
| Oovaut , Johnny | Nunavik Marine Region Wildlife Board |
| Oyukuluk, Qaumayuq | Ikajutit HTO (Arctic Bay) |
| Qaunaq, Matthias | Mittimatalik HTO (Pond Inlet) |
| Sala, Harry | Sanikiluaq HTO |
| Schneidmiller, Adam | Nunavut Wildlife Management Board |
| Stephenson, Sam | DFO (SARA, Central and Arctic region) |
| Stewart, Bruce | COSEWIC status report co-author |
| Stewart, Rob | DFO (Science, Central and Arctic region) |
| Tapaungai, Kovianatuliaq | Aiviq HTO (Cape Dorset) |
| Tarqriasuk, Quitsaq | Ivujivik Hunting, Fishing and Trapping Committee |
| Tatty, John | Kivalliq Wildlife Board |
| Trites, Andrew | COSEWIC marine mammal subcommittee |
| Trumbley, Dean | COSEWIC ATK subcommittee co-chair |
| Ugarte, Fernando | Greenland Institute of Natural Resources |
| Whalen, Julie | Torngat Secretariat |
Appendix 1. Threats assessment worksheet for high Arctic population, Atlantic Walrus.
- Species or ecosystem scientific name:
- Odobenus rosmarus rosmarus - High Arctic population
- Date:
- 24/01/2017
- Assessor(s):
- Jeff W. Higdon; D. Bruce Stewart, Hal Whitehead, Dwayne Lepitzki, David Lee, Karen Timm, Jessica Humber, Denis Etiendem, Donna Hurlburt, Mieke Hagesteijn, Mark O'Connor, Kaitlyn Breton-Honeyman, Rob Stewart, Paul Irgnaut, Maha Ghazal, Paul Blanchfield, Gregor Gilbert, Allison MacPhee, Mark Basterfield
- References:
- draft calculator (31 Dec 2016) + 6-month report; telecon 24 Jan 2017
| Threat impact | Threat impact (descriptions) | Level 1 Threat impact counts: high range |
Level 1 Threat impact counts: low range |
|---|---|---|---|
| A | Very high | 0 | 0 |
| B | High | 0 | 0 |
| C | Medium | 3 | 1 |
| D | Low | 2 | 4 |
| - | Calculated overall threat impact: | High | High |
- Assigned overall threat impact:
- B = High
- Overall threat comments
- Generation time = 21 yr so 3 generations = 63 yr (for severity & timing); Subpopulation estimates, aerial surveys, uglit: High Arctic DU1 = 2481: PS-LS (29%), WGS (20%), BB (50%)
| Threat | Threat description | Impact (calculated) | Impact (calculated) description | Scope (next 10 Yrs) | Severity (10 Yrs or 3 Gen.) | Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | blank | Not a Threat | Negligible (<1%) | Neutral or Potential Benefit | Moderate (Possibly in the short term, < 10 yrs) | |
| 1.1 | Housing & urban areas | blank | blank | blank | blank | blank | blank |
| 1.2 | Commercial & industrial areas | blank | blank | blank | blank | blank | blank |
| 1.3 | Tourism & recreation areas | blank | Not a Threat | Negligible (<1%) | Neutral or Potential Benefit | Moderate (Possibly in the short term, < 10 yrs) | Defined as "Tourism and recreation sites with a substantial footprint" and thus excludes disturbance from cruise ships, etc. New development may happen in future but no detailed information. Parks Canada considering a Marine Conservation Area in Lancaster Sound. Also, Inuit Circumpolar Council interested in establishing Northwater Polynya as some sort of marine protected area. Likely any visitor centres could be situated (or footprint from physical development) to have little to no impact to Walrus haulout sites on land. |
| 2 | Agriculture & aquaculture | blank | blank | blank | blank | blank | |
| 2.1 | Annual & perennial non-timber crops | blank | blank | blank | blank | blank | blank |
| 2.2 | Wood & pulp plantations | blank | blank | blank | blank | blank | |
| 2.3 | Livestock farming & ranching | blank | blank | blank | blank | blank | blank |
| 2.4 | Marine & freshwater aquaculture | blank | blank | blank | blank | blank | blank |
| 3 | Energy production & mining | blank | Unknown | Unknown | Unknown | Moderate - Low | Land-based mining and quarrying are not considered but related transportation is considered under section 4.3 (Shipping lanes) and 7.3. (Other ecosystem modifications). Hydropower is considered in 7.2 Dams & Water Management/Use rather than here as "Renewable Energy". |
| 3.1 | Oil & gas drilling | blank | Unknown | Unknown | Unknown | Moderate - Low | Oil and gas drilling is uncertain given the moratorium announced in 2016 by the federal government, which is in effect for 5 years at minimum. Also uncertain is whether or not the moratorium includes seismic exploration. |
| 3.2 | Mining & quarrying | blank | blank | blank | blank | blank | blank |
| 3.3 | Renewable energy | blank | blank | blank | blank | blank | There may be possibility of tidal projects, but considered unlikely. |
| 4 | Transportation & service corridors | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | blank |
| 4.1 | Roads & railroads | blank | blank | blank | blank | blank | |
| 4.2 | Utility & service lines | blank | blank | blank | blank | blank | Proposal for Quintillion Expressnet telecommunications pipeline across Arctic to connect Europe to Nunavut might eventually go through DU habitat. Marine footprint impacts involve paying cable out from slow moving ship, and may not affect Walrus when cable on sea floor. Potential impacts where lines are near haulout sites. |
| 4.3 | Shipping lanes | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | Shipping includes SeaLift and fuel deliveries to Arctic communities in addition to mining companies' shipping for construction, resupply, and product delivery (e.g., ore) out. Shipping lanes are also used by other large vessels like cruise ships, Coast Guard, etc. All types of shipping are likely to increase over the short and long term in response to changing access caused by reduced ice cover, particularly shipping related to resource development and world trade. Displacement, increased stress levels due to disturbance are likely, but there is no direct causal evidence. Severity could increase in future, or may be mitigated. Frequency certainly expected to increase. Baffinland is applying to increase ore production and shipping through Milne Inlet and Eclipse Sound. |
| 4.4 | Flight paths | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Some flight paths could be close to terrestrial haulout sites or potentially disturb animals hauled out on ice. Many research flights occurring but uncertain of extent of disturbance. Ongoing and sporadic in nature. Helicopter and Twin Otter flights may increase in future related to oil and gas interests. May be a bigger effect for Some flight paths could be close to terrestrial haulout sites or potentially disturb animals hauled out on ice. Many research flights are occurring but there is uncertainty about extent of disturbance. Helicopter and Twin Otter flights may increase in future related to oil and gas interests. Severity may be closer to the lower end of the range indicated here. |
| 5 | Biological resource use | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | blank |
| 5.1 | Hunting & collecting terrestrial animals | blank | blank | blank | blank | blank | |
| 5.2 | Gathering terrestrial plants | blank | blank | blank | blank | blank | blank |
| 5.3 | Logging & wood harvesting | blank | blank | blank | blank | blank | blank |
| 5.4 | Fishing & harvesting aquatic resources | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | Subsistence hunting primarily, also limited sport hunt. Harvests appear to be declining based on available reported data and Inuit reports. The reasons for this decline are uncertain. The accuracy of harvest data is uncertain due to inconsistent reporting and lack of accompanying struck and lost data. Periodic verification of reporting accuracy is needed. Canadian animals are hunted in Greenland (where rate thought to be unsustainable). Net entanglements considered possible. |
| 6 | Human intrusions & disturbance | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | blank |
| 6.1 | Recreational activities | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | Arctic tourism activities occur throughout the species' range. Some overlap with 4.3 (Shipping lanes) due to cruise ship tourism, and with 6.3 (Work and other activities) due to recreational hunting and travel. Recreational hunting was discussed under 5.4. Targeted visiting of haulout sites discussed here. Cruise ships and/or excursion tenders may closely approach haulout sites and don't stick to shipping lanes as commercial ships do. Such visits may impact haulouts sequentially (if tourism activities cause abandonment of one haulout, operators will search out another, and so on). In fiords animals may not be able to escape from cruise ships except to swim directly past ship. Severity might increase in future, or may be mitigated. |
| 6.2 | War, civil unrest & military exercises | blank | Negligible | Restricted (11-30%) | Negligible (<1%) | High (Continuing) | Arctic military exercises and Northern Ranger patrols happen on an annual basis, but effects likely to be negligible. |
| 6.3 | Work & other activities | D | Low | Pervasive (71-100%) | Slight (1-10%) | High (Continuing) | Disturbances caused by community activities (e.g., travel, construction, hunting of other species) unrelated to shipping, Walrus hunting, etc. or by research activities may cause Walrus to avoid or abandon suitable habitats. Effects of community harvest of other species was discussed. Severity is towards the lower part of the range. |
| 7 | Natural system modifications | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | blank |
| 7.1 | Fire & fire suppression | blank | blank | blank | blank | blank | blank |
| 7.2 | Dams & water management/use | blank | blank | blank | blank | blank | blank |
| 7.3 | Other ecosystem modifications | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Baffinland plans to seek approval to ship ore during July through December from Milne Inlet via Eclipse Sound, Pond Inlet, and Baffin Bay to ports in Europe but "will make every effort to ship ore during the open water season." (BIMC 2016:19). Physical effects of ballast water discharges in this area affecting Walrus are less likely than along the southern shipping route for DU2 as Walrus are not all that common in Pond and Milne inlets, the water is much deeper, and most vessel traffic is planned for the open water season, at least for now. Impact may be increased depending on type of invasive species introduced with ballast water (Green Crab, Carcinus maenas, interactions for example). |
| 8 | Invasive & other problematic species & genes | blank | Unknown | Unknown | Unknown | High (Continuing) | blank |
| 8.1 | Invasive non-native/alien species | blank | Unknown | Unknown | Unknown | High (Continuing) | Potential for AIS introductions from ballast water discharge or hull-fouling and from ongoing range extensions (e.g., Green Crab). The potential effects on Walrus from trophic changes and novel diseases, parasites, and/or predators are unknown. |
| 8.2 | Problematic native species | blank | Unknown | Unknown | Unknown | High (Continuing) | Polar Bear predation at terrestrial haulouts may increase in response to the longer open water season. With declining sea ice, and a decline in Ringed Seal abundance, there may be a greater reliance by Polar Bears on Walrus haulout sites at certain times of the year. Incidentally, there may be some mortality through trampling of calves which are then consumed by polar bears, but this effect is likely small in scope. Changing predation patterns by both Killer Whales and Polar Bears were discussed. |
| 8.3 | Introduced genetic material | blank | blank | blank | blank | blank | |
| 9 | Pollution | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | blank |
| 9.1 | Household sewage & urban waste water | blank | blank | blank | blank | blank | |
| 9.2 | Industrial & military effluents | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Some industrial activity (e.g., mines), some military (e.g., old DEW Line sites), but effects likely negligible. Most sites not adjacent to important Walrus habitats. SeaLifts and other vessels are transiting through here. If a haulout site is fouled by oil, and no other suitable sites, then there could be population level reductions to consider. Animals would likely become oiled if on shore at event. Overall impact of a single spill could be devastating (especially in winter). May be of higher impact in Central-Low DU. |
| 9.3 | Agricultural & forestry effluents | blank | blank | blank | blank | blank | |
| 9.4 | Garbage & solid waste | blank | blank | blank | blank | blank | Garbage from cruise ships was discussed but entanglement impacts unlikely. Dispersal over long distances of household garbage is common but impacts to Walrus unknown. Microplastics discussed and are globally pervasive but not enough data to determine impacts. |
| 9.5 | Air-borne pollutants | blank | blank | blank | blank | blank | Impacts to food chain were discussed but unknown. |
| 9.6 | Excess energy | blank | blank | blank | blank | blank | Noise from sonar on ships and boats was discussed but impacts unknown. This would include military sonar. |
| 10 | Geological events | blank | blank | blank | blank | blank | blank |
| 10.1 | Volcanoes | blank | blank | blank | blank | blank | blank |
| 10.2 | Earthquakes/tsunamis | blank | blank | blank | blank | blank | blank |
| 10.3 | Avalanches/landslides | blank | blank | blank | blank | blank | blank |
| 11 | Climate change & severe weather | blank | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | blank |
| 11.1 | Habitat shifting & alteration | blank | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | Climate change and changing sea ice dynamics have the potential to affect Walrus throughout their range, but magnitude, severity and time frames for impacts are uncertain. Ocean acidification was discussed; is not at a tipping point yet but impacts to prey sources could be significant. |
| 11.2 | Droughts | blank | blank | blank | blank | blank | |
| 11.3 | Temperature extremes | blank | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | Unfamiliar disturbances from thunder and lightning storms could increase the frequency of stampedes at haulouts. |
| 11.4 | Storms & flooding | blank | Unknown | Unknown | Unknown | Unknown | Storms could affect terrestrial haulout sites that are used by Walrus, depending on the type of habitat (e.g., sand or pebble beaches). |
Appendix 2. Threats assessment worksheet for central-low Arctic population, Atlantic Walrus.
- Species or ecosystem scientific name:
- Odobenus rosmarus rosmarus - Atlantic Walrus – Central-Low Arctic population
- Date:
- 24/01/2017
- Assessor(s):
- Jeff W. Higdon; D. Bruce Stewart, Hal Whitehead, Dwayne Lepitzki, David Lee, Karen Timm, Jessica Humber, Denis Etiendem, Donna Hurlburt, Mieke Hagesteijn, Mark O'Connor, Kaitlyn Breton-Honeyman, Rob Stewart, Paul Irgnaut, Maha Ghazal, Gregor Gilbert, Allison MacPhee, Mark Basterfield
- References:
- draft calculator (31 Dec 2016) + 6-month report; telecon 24 Jan 2017
| Threat impact | Threat impact (descriptions) | Level 1 Threat impact counts: high range |
Level 1 Threat impact counts: low range |
|---|---|---|---|
| A | Very high | 0 | 0 |
| B | High | 1 | 0 |
| C | Medium | 3 | 3 |
| D | Low | 1 | 2 |
| - | Calculated overall threat impact: | Very High | High |
- Overall threat comments
-
It was noted that due to geographical overlap, the overall threat could be modified.
Generation time = 21 yr so 3 generations = 63 yr (for severity & timing); Subpopulation estimates, aerial surveys, uglit: Central-Low Arctic DU2 = 18,579: FB (56%), NWHB (30%), SEB (13%), SHSUBL (? %), LA (1.1%)
| Threat | Threat description | Impact (calculated) | Impact (calculated) description | Scope (next 10 Yrs) | Severity (10 Yrs or 3 Gen.) | Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | blank | blank | blank | blank | blank | |
| 1.1 | Housing & urban areas | blank | blank | blank | blank | blank | blank |
| 1.2 | Commercial & industrial areas | blank | blank | blank | blank | blank | blank |
| 1.3 | Tourism & recreation areas | blank | blank | blank | blank | blank | Defined as "Tourism and recreation sites with a substantial footprint" and thus excludes disturbance from cruise ships, etc. New development may happen in future but no detailed information. Parks Canada has interest in western Hudson Bay and other sites, and DFO has interest in establishing marine protected areas in some other sites, but any designation is unlikely to negatively affect Walrus habitat. Likely any visitor centres, or footprint from physical development, could be situated to have little to no impact to Walrus haulout sites on land. |
| 2 | Agriculture & aquaculture | blank | blank | blank | blank | blank | |
| 2.1 | Annual & perennial non-timber crops | blank | blank | blank | blank | blank | blank |
| 2.2 | Wood & pulp plantations | blank | blank | blank | blank | blank | |
| 2.3 | Livestock farming & ranching | blank | blank | blank | blank | blank | blank |
| 2.4 | Marine & freshwater aquaculture | blank | blank | blank | blank | blank | blank |
| 3 | Energy production & mining | blank | Unknown | Unknown | Unknown | Moderate - Low | Land-based mining and quarrying are not considered but related transportation is considered under section 4.3 (Shipping lanes) and 7.3. (Other ecosystem modifications). Hydropower is considered in 7.2 Dams & Water Management/Use rather than here as "Renewable Energy". |
| 3.1 | Oil & gas drilling | blank | Unknown | Unknown | Unknown | Moderate - Low | Oil and gas drilling is uncertain given the moratorium announced in 2016 by the federal government, which is in effect for 5 years at minimum. Also uncertain is whether or not the moratorium includes seismic exploration. |
| 3.2 | Mining & quarrying | blank | blank | blank | blank | blank | blank |
| 3.3 | Renewable energy | blank | blank | blank | blank | blank | There may be possibility of tidal projects, but considered unlikely. |
| 4 | Transportation & service corridors | BC | High - Medium | Pervasive (71-100%) | Serious - Moderate (11-70%) | High (Continuing) | blank |
| 4.1 | Roads & railroads | blank | blank | blank | blank | blank | |
| 4.2 | Utility & service lines | blank | blank | blank | blank | blank | Proposal for Quintillion Expressnet telecommunications pipeline across Arctic to connect Europe to Nunavut. Proposed line goes along mainland Canada. Secondary line later when funded, which would go up E Baffin Coast to connect northern communities. Marine footprint impacts involve paying cable out from slow moving ship, and may not affect Walrus when cable on sea floor. Potential impacts where lines are near haulout sites. |
| 4.3 | Shipping lanes | BC | High - Medium | Pervasive (71-100%) | Serious - Moderate (11-70%) | High (Continuing) | Shipping includes SeaLift and fuel deliveries to Arctic communities in addition to mining companies' shipping for construction, resupply, and product delivery (e.g., ore) out. Shipping lanes also used by other large vessels like cruise ships, Coast Guard, etc. All types of shipping are likely to increase over the short and long term in response to changing access due to reduction in ice cover, particularly shipping related to resource development and world trade. There may be displacement or increased stress levels due to disturbance in Walrus, but no causal evidence to date. Severity could increase in future, or may be mitigated. Some icebreaker impacts possible in the future. Icebreaking would impact sea ice habitat, with the risk of animal injury or mortality, and the potential for freeze-out, disturbance, and disruption of breeding or other important behaviours. The Mary River Iron Mine on Baffin Island has an approved project that allows year-round shipping through Foxe Basin and Hudson Strait. There is approval in place for 100+ round-trip voyages on the southern route, with year-round shipping by Cape-class ore carriers. This component of the project is on hold for the moment, and shipping is focused on the northern route. |
| 4.4 | Flight paths | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Some flight paths could be close to terrestrial haulout sites or potentially disturb animals hauled out on ice. Many research flights are occurring but the extent of disturbance is uncertain. Helicopter and Twin Otter flights may increase in future related to oil and gas interests. Potential for increased flights due to Baffinland Iron Mine activity. Project-specific Terms and Conditions for Baffinland on minimum flight levels are specific to birds, caribou and other terrestrial species. There is a condition for Baffinland to establish a monitoring program that focuses on Walrus use of Steensby Inlet and their reaction to disturbance from construction activities, aircraft, and vessels. Baffinland has established the following mitigation measure: Small aircraft maintain altitude of 450 m over marine waters when possible. Prohibiting aircraft from flying low over animals for passengers to ‘get a better look’ or for photography. |
| 5 | Biological resource use | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | blank |
| 5.1 | Hunting & collecting terrestrial animals | blank | blank | blank | blank | blank | |
| 5.2 | Gathering terrestrial plants | blank | blank | blank | blank | blank | blank |
| 5.3 | Logging & wood harvesting | blank | blank | blank | blank | blank | blank |
| 5.4 | Fishing & harvesting aquatic resources | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | Subsistence hunting primarily, limited sport hunt. Harvests appear to be declining, based on available reporting and Inuit reports. The reasons for this decline are uncertain. The accuracy of harvest data is uncertain due to inconsistent reporting and lack of accompanying struck and lost data. Periodic verification of reporting accuracy is needed. The severity may be Serious in southeastern Hudson Bay but only Moderate to Slight elsewhere. A portion of the population that summers in Canada is hunted in Greenland. Net entanglements considered. Harvest levels are of more concern in the Low Arctic portion of this DU than in the Central portion. |
| 6 | Human intrusions & disturbance | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | blank |
| 6.1 | Recreational activities | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | Cruise ships less frequent in Foxe Basin than some other parts of the Arctic; however, smaller boat tourism has had past negative impact. Arctic tourism activities happen throughout the species' range. Some overlap with 4.3 (Shipping lanes) due to cruise ship tourism, and with 6.3 (Work and other activities) due to recreational hunting and travel. Recreational hunting was discussed under 5.4. Targeted visiting of haulout sites discussed here. Severity could increase in future, or may be mitigated. |
| 6.2 | War, civil unrest & military exercises | blank | Negligible | Restricted (11-30%) | Negligible (<1%) | High (Continuing) | Arctic military exercises and Northern Ranger patrols happen on an annual basis (DUring winter), but effects likely to be negligible. |
| 6.3 | Work & other activities | D | Low | Pervasive (71-100%) | Slight (1-10%) | High (Continuing) | Disturbances caused by community activities (e.g., travel, construction, hunting of other species) unrelated to shipping, Walrus hunting, etc. or by research activities may cause Walrus to avoid or abandon suitable habitats. Effects of community harvest of other species was discussed. Severity is towards the lower part of the range. |
| 7 | Natural system modifications | C | Medium | Large (31-70%) | Moderate (11-30%) | High - Moderate | blank |
| 7.1 | Fire & fire suppression | blank | blank | blank | blank | blank | |
| 7.2 | Dams & water management/use | D | Low | Small (1-10%) | Serious - Moderate (11-70%) | High (Continuing) | Flow management by hydroelectric developments in southern Hudson Bay and James Bay may have a moderate effect on most Walrus in southeastern Hudson Bay; slight effects on Walrus downstream in northeastern Hudson Bay and Hudson Strait; and no effect on those elsewhere. Changes in under-ice salinity may be an issue as well. Additional plans for future hydroelectric installations may further affect this DU. |
| 7.3 | Other ecosystem modifications | C | Medium | Large (31-70%) | Moderate (11-30%) | High - Moderate | Large-scale ice-breaking activities have been approved to support mining development on Baffin Island via Hudson Strait and Foxe Basin. Ballast water discharge near ports may impact habitat (much more ballast water potential for this DU). Ice-breakers may be of high impact for this DU. Impact may be increased depending on type of invasive species introduced with ballast water (Green Crab interactions for example). |
| 8 | Invasive & other problematic species & genes | blank | Unknown | Unknown | Unknown | High (Continuing) | blank |
| 8.1 | Invasive non-native/alien species | blank | Unknown | Unknown | Unknown | High (Continuing) | Potential for AIS introductions from ballast water discharge or hull-fouling and from ongoing range extensions (e.g., Green Crab). The potential effects on Walrus from trophic changes and novel diseases, parasites, and/or predators are unknown. Trichinosis may be an issue. |
| 8.2 | Problematic native species | blank | Unknown | Unknown | Unknown | High (Continuing) | Polar Bear predation at terrestrial haulouts may increase in response to the longer open water season. With declining sea ice, and a decline in Ringed Seal abundance, there may be a greater reliance by Polar Bears on Walrus haulout sites at certain times of the year. Incidentally, there may be some mortality through trampling of calves which are then consumed by polar bears, but this effect is likely small in scope. Changing predation patterns by both Killer Whales and Polar Bears were discussed. |
| 8.3 | Introduced genetic material | blank | blank | blank | blank | blank | |
| 9 | Pollution | D | Low | Small (1-10%) | Moderate - Slight (1-30%) | High (Continuing) | blank |
| 9.1 | Household sewage & urban waste water | blank | blank | blank | blank | blank | |
| 9.2 | Industrial & military effluents | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | Some industrial activity (e.g., mines), some military (e.g., old DEW Line sites), but effects likely negligible. Most sites not adjacent to important Walrus habitats. SeaLifts and other vessels are transiting through here. If a haulout site is fouled by oil, and no other sites are available, then there could be population level reductions to consider. Animals would likely become oiled if on shore at event. Overall impact of a single spill could be devastating (especially in winter)(and frequency of ships sure to increase). Industrial shipping overlaps areas of high Walrus occupancy. |
| 9.3 | Agricultural & forestry effluents | blank | blank | blank | blank | blank | |
| 9.4 | Garbage & solid waste | blank | blank | blank | blank | blank | Garbage from cruise ships was discussed but entanglement impacts unlikely. Dispersal over long distances of household garbage is common but impacts to Walrus unknown. Microplastics discussed and are globally pervasive but not enough data to determine impacts. |
| 9.5 | Air-borne pollutants | blank | blank | blank | blank | blank | Impacts to food chain were discussed but impacts unknown. |
| 9.6 | Excess energy | blank | Unknown | Pervasive (71-100%) | Unknown | High - Moderate | Noise from sonar on ships and boats was discussed but impacts unknown. This would include military sonar. |
| 10 | Geological events | blank | blank | blank | blank | blank | blank |
| 10.1 | Volcanoes | blank | blank | blank | blank | blank | blank |
| 10.2 | Earthquakes/tsunamis | blank | blank | blank | blank | blank | blank |
| 10.3 | Avalanches/landslides | blank | blank | blank | blank | blank | blank |
| 11 | Climate change & severe weather | blank | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | Ocean acidification? |
| 11.1 | Habitat shifting & alteration | blank | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | Climate change and changing sea ice dynamics have the potential to affect Walrus throughout their range, but magnitude, severity and time frames for impacts are uncertain. Ocean acidification was discussed; is not at a tipping point yet but impacts to prey sources could be significant. |
| 11.2 | Droughts | blank | blank | blank | blank | blank | |
| 11.3 | Temperature extremes | blank | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | Unfamiliar disturbances from thunder and lightening storms could increase the frequency of stampedes at haulouts. |
| 11.4 | Storms & flooding | blank | Unknown | Unknown | Unknown | Unknown | Storms could affect terrestrial haulout sites that are used by Walrus, depending on the type of habitat (e.g., sand or pebble beaches). |