
Evening grosbeak (Coccothraustes vespertinus): COSEWIC assessment and status report 2016
Evening Grosbeak

Special concern
2016
Table of contents
- Table of contents
- COSEWIC Assessment summary
- COSEWIC Executive summary
- Technical summary
- Wildlife species description and significance
- Distribution
- Habitat
- Biology
- Population sizes and trends
- Sampling effort and methods
- Christmas bird count
- Project FeederWatch
- North american breeding bird survey
- Breeding bird atlases
- Forest bird monitoring program
- Checklist program in Quebec (Étude des populations d’oiseaux du Québec, ÉPOQ)
- Migration count at Observatoire d’oiseaux de Tadoussac (OOT)
- Fort Liard landbird monitoring program
- Abundance
- Fluctuations and trends
- Christmas bird count
- Project FeederWatch
- North American breeding bird survey
- Alberta breeding bird atlas
- Ontario breeding bird atlas
- Quebec breeding bird atlas
- Checklist program in Quebec (Étude des populations d’oiseaux du Québec, ÉPOQ)
- Forest bird monitoring program
- Migration count at the Observatoire d’oiseaux de Tadoussac (OOT) QC
- Fort Liard landbird monitoring program
- Population trend summary
- Rescue effect
- Sampling effort and methods
- Threats and limiting factors
- Threats
- Residential and commercial development (housing and urban areas – low threat)
- Transportation and service corridors (roads and railroads – low threat)
- Biological resource use (logging and wood harvesting – low threat)
- Natural system modification (unknown threat)
- Pollution (industrial and military effluents – unknown threat)
- Agriculture and aquaculture (annual and perennial non-timber/livestock farming and ranching – negligible threat)
- Climate change and severe weather (negligible threat)
- Limiting factors
- Diseases and parasites
- Number of locations
- Threats
- Protection, status, and ranks
- Acknowlegements and authorities contacted
- Information sources
- Biographical summary of report writer
- Appendix 1. Threats assessment for Evening Grosbeak
List of figures
- Figure 1. Adult male Evening Grosbeak (photo Carl Savignac).
- Figure 2. North American range of Evening Grosbeak. The species is found year-round in the green area, but can winter irregularly up to the dotted lines except for the subspecies C.v. montana, which is resident in Arizona and Mexico.
- Figure 3. Canadian range and extent of occurrence of Evening Grosbeak.
- Figure 4. Relative quality of Evening Grosbeak habitat in the boreal and hemiboreal zone of North America based on climate and vegetation cover.
- Figure 5. Relative abundance (average number of birds/route/year) of Evening Grosbeak calculated for each square of latitude and longitude between 1987 and 2006 during the breeding period according to the North American Breeding Bird Survey (BBS). Grey areas = not sampled by BBS; white areas = sampled, but no Evening Grosbeak detected.
- Figure 6. Expected counts (with 95% confidence interval) from an average CBC circle, in Canada from 1970 through 2012, after accounting for variations in effort within circles, and sampling bias among circles, years, and regions.
- Figure 7. Ten-year rolling trend for Evening Grosbeak between 1980 and 2012 in Canada based on Christmas Bird Count data.
- Figure 8. Trends in the percentage of feeders visited and the average number of birds seen at feeders in four regions of North America from Project FeederWatch, 1989-2014. Far North = Northwest Territories; Northwest Region = British Columbia and Alberta; Central Region = Saskatchewan and Manitoba; Northeast Region = Ontario, Quebec, and Atlantic provinces.
- Figure 9. Annual abundance index in Canada between 1970 and 2012 (with 95% confidence interval) according to a hierarchical Bayesian model of BBS data (Environment Canada 2014).
- Figure 10. Ten-year rolling trend for Evening Grosbeak between 1980 and 2014 in Canada, based on Breeding Bird Survey data.
- Figure 11. Distribution of Evening Grosbeaks in Ontario during the period 001-2005. Squares with black dots correspond to plots in which Evening Grosbeak was found in the first atlas period (1980-1985), but not in the second atlas period (2001-2005), while squares with yellow dots indicate presence in the second but not the first atlas period.
- Figure 12a. Distribution of Evening Grosbeaks in Quebec during the first atlas period (1984-1989).
- Figure 12b. Distribution of Evening Grosbeaks in Quebec during the second atlas period 2010-2014.
- Figure 13. Distribution of Evening Grosbeaks in the Maritimes provinces during the period 2006-2010, with dots showing comparison with the first atlas period of 1986-1990.
- Figure 14. Annual abundance index of Evening Grosbeak in Quebec between 1970 and 2014 according to the Checklist program in Quebec.
- Figure 15. Trend in abundance index (total number/time effort) of migrating Evening Grosbeaks at the Observatoire d’oiseaux de Tadoussac from 1996-2015. Only birds that were detected moving generally south were included in the analysis.
- Figure 16. Map of BBS trends for Evening Grosbeak in the United States and Canada for the period 1966 to 2012.
- Figure 17. Area of forest defoliated by Spruce Budworm in Ontario, Quebec and in Canada between 1966 and 2009, by province (Venier, unpubl. data). Only provinces with defoliated area values ≥ 1 million ha during any year are shown.
- Figure 18. Association between the total area of forest defoliated by Spruce Budworm and the BBS annual abundance index for Evening Grosbeak between 1974 and 2012 in Canada. Regression adjusted using generalized least squares with errors following a first-order autoregressive process.
List of tables
- Table 1. Evening Grosbeak densities obtained in various Canadian provinces using the Canadian Breeding Bird Census mapping method (Kennedy et al. 1999). Data were grouped according to a period prior to Spruce Budworm (1965-1970) and during the outbreak (1970-1985).
- Table 2. Estimated population and relative abundance of Evening Grosbeak in the Canadian provinces according to BBS data (Partners in Flight Science Committee 2013).
- Table 3. Annual long-term (1970-2012) and short-term (2002-2012) trends from CBC for Evening Grosbeak in Canada (Smith, unpubl. data) with 95% lower (LCI) and upper (UCI) credible intervals. Results in bold are statistically significant declines, i.e., 95% credible intervals do not overlap zero.
- Table 4. Annual long-term (1970-2014, unless indicated otherwise) and short-term (2004-2014) trends from BBS for Evening Grosbeak in Canada (Smith, pers. comm.) with 95% lower (LCI) and upper (UCI) credible intervals. Results in bold are statistically significant declines, i.e., 95% credible intervals do not overlap zero.
- Table 5. NatureServe (2015) and General Status of Species in Canada (GSSC) (CESCC 2011) ranks assigned to Evening Grosbeak (all subspecies)
Document information
COSEWIC Assessment and Status Report on the Evening Grosbeak Coccothraustes vespertinus in Canada - 2016

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
Committee on the Status of Endangered Wildlife in Canada (COSEWIC) status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2016. COSEWIC assessment and status report on the Evening Grosbeak Coccothraustes vespertinus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xi + 64 pp. (Species at Risk Public Registry website).
Production note:
COSEWIC acknowledges Carl Savignac for writing the status report on the Evening Grosbeak, Coccothraustes vespertinus, in Canada, prepared with the financial support of Environment and Climate Change Canada. This report was overseen and edited by Jon McCracken, Co-chair of the COSEWIC Birds Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
E-mail: COSEWIC E-mail
Website: COSEWIC
Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur le Gros-bec errant (Coccothraustes vespertinus) au Canada.
Cover illustration/photo:
Evening Grosbeak - Photo credit: Carl Savignac.
COSEWIC
Assessment summary
Assessment summary - November 2016
- Common name
- Evening Grosbeak
- Scientific name
- Coccothraustes vespertinus
- Status
- Special Concern
- Reason for designation
- This large finch is widely distributed across Canada’s forests, but has exhibited significant long-term declines (77-90%) over most of its range, since 1970. Over the past decades, some data suggest a further decline of nearly 40%, while other data indicate stabilization at a lower level. Threats to the species include reduced availability of mature and old-growth mixed wood and conifer forests, collisions with windows, and mortality associated with feeding on grit and salt along roads in winter.
- Occurrence
- Yukon, Northwest Territories, British Columbia, Alberta, Saskatchewan, Manitoba, Ontario, Quebec, New Brunswick, Prince Edward Island, Nova Scotia, Newfoundland and Labrador
- Status history
- Designated Special Concern in November 2016.
COSEWIC
Executive summary
Evening Grosbeak
Coccothraustes vespertinus
Wildlife species description and significance
Evening Grosbeak is a stocky, boldly coloured songbird, with a massive greenish-yellow bill. Adult males have a dark brown head with a brilliant yellow supercilium; the brown of the head transitions to yellow upperparts and belly, contrasting with a black tail and black wings, with a distinct patch of all-white secondaries. Adult females and juveniles are generally greyish-brown with some yellow on the nape and flanks and black and white wings and tail. In summer, this species can be a major predator of the Spruce Budworm and helps in the natural control of this insect pest. In winter it is a familiar visitor to bird feeders.
Distribution
Evening Grosbeak breeds in Canada, the United States, and Mexico. In Canada, its distribution includes all Canadian provinces and territories except Nunavut. In the United States, the species breeds primarily in northern New England and some western states. In winter, it is nomadic and can range widely, depending on the quantity of seeds produced in the boreal forest. Historically, this species was restricted to western North America, but expanded eastward in the late 19th and early 20th centuries.
Habitat
Optimal Evening Grosbeak breeding habitat generally includes open, mature mixedwood forests, where fir species and/or White Spruce are dominant, and Spruce Budworm is abundant. Outside the breeding season, the species seems to depend largely on seed crops from various trees such as firs and spruces in the boreal forest, but is also attracted to ornamental trees that produce seeds or fruit, and bird feeders stocked with sunflower seeds.
Biology
Evening Grosbeak is socially monogamous and is not territorial during the breeding period. Pairs typically arrive on their breeding grounds from mid- to late May, and the nesting season can extend until early September. The nest is an open cup made of twigs or rootlets located in the canopy of trees, with conifers preferred over deciduous trees. There is one clutch per year with an average size of 3 to 4 eggs; re-nesting may occur if the initial clutch fails. Incubation typically lasts 12 to 14 days, and fledglings leave the nest at 13 to 14 days old. The age at first breeding is one year.
Population sizes and trends
The Canadian Evening Grosbeak population is estimated to be approximately 2,200,000 mature individuals. Trends are difficult to evaluate for nomadic species, but data from the Christmas Bird Count (CBC) and Breeding Bird Survey (BBS) show similar long-term declines. The CBC, which samples sites throughout the entire wintering range, indicates a significant overall decline of 3.4% per year from 1970 to 2012, corresponding to a cumulative decline of 76.6%, although from 2002 to 2012 there was a non-significant increase of 3.1% per year. The BBS primarily monitors the southern portion of the Evening Grosbeak’s breeding range, and indicates a significant annual decline of 5.2% between 1970 and 2014, for a population decline of 90% over 44 years. BBS data for the most recently available ten-year period (2004 to 2014) show an ongoing significant decrease of 5.0% per year in Canada, for a cumulative decline of 42%. Short-term (2004-2014) BBS trends are also negative in all provinces, but the trend is significant only in Manitoba, Ontario, and Quebec. Provincial breeding bird atlases, the Étude des populations d’oiseaux du Québec (ÉPOQ), and Project FeederWatch also generally show declining trends. Observatoire d’oiseaux de Tadoussac (QC) data suggest that Evening Grosbeak numbers were low from 1998 to 2011, but have increased considerably from 2012 to 2015. The Fort Liard Songbird Monitoring Project in the southern Northwest Territories showed a stable trend for 1998 to 2011. Overall, long-term trends are strongly negative across many sources of data; there is more variability among short-term trends, with some indicating ongoing declines, and others reflecting stability or increasing numbers in certain regions.
Threats and limiting factors
Fluctuations of Spruce Budworm populations, which naturally occur every 25-40 years in eastern Canada and every 26 years in western Canada, are likely a key factor in fluctuations of the Evening Grosbeak population since 1970. Known threats to Evening Grosbeak include mortality caused by window strikes while birds are visiting feeders in winter, reduction of mature and old-growth mixedwood forests due to commercial forest management, and mortality due to road collisions when individuals feed on grit and road salt. Mortality related to ingestion of sodium chloride along roadsides may also be a threat. Over the long term, there may be a contraction of breeding habitat due to climate change.
Protection, status and ranks
In Canada, Evening Grosbeak and its nests and eggs are protected under the Migratory Birds Convention Act, 1994. NatureServe considers Evening Grosbeak secure in Canada, imperilled in Prince Edward Island, and vulnerable in Yukon, Manitoba, and New Brunswick; in other provinces and territories, the species is considered either secure or probably secure.
Technical summary
- Scientific name:
- Coccothraustes vespertinus
- English name:
- Evening Grosbeak
- French name:
- Gros-bec errant
- Range of occurrence in Canada:
- Yukon, Northwest Territories, British Columbia, Alberta, Saskatchewan, Manitoba, Ontario, Quebec, New Brunswick, Prince Edward Island, Nova Scotia, Newfoundland and Labrador
Demographic information
| Summary items | Information |
|---|---|
| Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines (2011) is being used) | approximately 3-4 yrs |
| Is there an [observed, inferred, or projected] continuing decline in number of mature individuals? | Yes |
| Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations] | n/a |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations]. | 42% Based on Canadian Breeding Bird Survey data for 2004-2014 showing a mean annual decline of 5.0% |
| [Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations]. | Unknown, but possibly increasing in parts of eastern Canada if Spruce Budworm outbreaks persist |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future. | Unknown, but possibly increasing in parts of eastern Canada if Spruce Budworm outbreaks persist |
| Are the causes of the decline a. clearly reversible and b. understood and c. ceased? | The causes of the decline are not clearly understood or reversible, and probably not ceased. Natural Spruce Budworm outbreaks may reverse the decline temporarily in some regions. |
| Are there extreme fluctuations in number of mature individuals? | No overall, although the population does fluctuate in association with the cycle of Spruce Budworm, and fluctuations can be extreme in some areas. |
Extent and occupancy information
| Summary items | Information |
|---|---|
| Estimated extent of occurrence - Based on a minimum convex polygon of the species’ range map (see Figure 3) |
6,633,526 km2 |
| Index of area of occupancy (IAO) - Based on a 2 km x 2 km grid, with estimated average density of 5 pairs / km2 and a population of 1.1 million pairs | ~220,000 km2 |
| Is the population “severely fragmented” i.e. is >50% of its total area of occupancy in habitat patches that are (a) smaller than would be required to support a viable population, and (b) separated from other habitat patches by a distance larger than the species can be expected to disperse? | No |
| Number of “locations” (use plausible range to reflect uncertainty if appropriate) | Unknown, but >10 |
| Is there an [observed, inferred, or projected] decline in extent of occurrence? | Potentially, but likely long-term, projected based on decreasing Balsam Fir forests in southern Canada |
| Is there an [observed, inferred, or projected] decline in index of area of occupancy? | Yes, observed based on breeding bird atlas data |
| Is there an [observed, inferred, or projected] decline in number of subpopulations? | The number of subpopulations is unknown |
| Is there an [observed, inferred, or projected] decline in number of “locations”? | Unknown |
| Is there an [observed, inferred, or projected] decline in [area, extent and/or quality] of habitat? | Possibly, based on reduced diversity in forest structure, but may be limited in occurrence |
| Are there extreme fluctuations in number of subpopulations? | No |
| Are there extreme fluctuations in number of “locations”? | Unlikely |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | Unlikely |
Number of mature individuals (in each subpopulation)
| Subpopulations (give plausible ranges) | N Mature individual |
|---|---|
| Total (Based on BBS data corrected with point counts from the Ontario Breeding Bird Atlas (Partners in Flight Science Committee 2013) |
2,200,000 |
Quantitative analysis
| Summary items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. | Not conducted |
Threats (actual or imminent, to populations or habitats, from highest impact to least)
| Summary items | Information |
|---|---|
| Was a threats calculator completed for this species and if so, by whom? | Yes, March 2015 (Dave Fraser, Marcel Gahbauer, Carl Savignac, Bruno Drolet, Julie Perrault, Steve Van Wilgenburg, Mary Sabine) |
| Overall threats are low, but include: |
|
Rescue effect (immigration from outside Canada)
| Summary items | Information |
|---|---|
| Status of outside population(s) most likely to provide immigrants to Canada. | Overall decline in the United States, but with some states near the border showing an increasing trend according to BBS data |
| Is immigration known or possible? | Yes, known |
| Would immigrants be adapted to survive in Canada? | Yes |
| Is there sufficient habitat for immigrants in Canada? | Yes |
| Are conditions deteriorating in Canada? | Yes, but to a minor extent in the short term |
| Are conditions for the source population deteriorating? | Yes, decline of Balsam Fir forests in the U.S. part of the Evening Grosbeak’s range is predicted due to climate change (Williams and Liebhold 1997; Matthews et al. 2004; Siegel et al. 2014) |
| Is the Canadian population considered to be a sink? | Unknown |
| Is rescue from outside populations likely? | Unknown |
Data sensitive species
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
Status history
| Summary items | Information |
|---|---|
| Designated Special Concern in November 2016 | Designated Special Concern in November 2016 |
Status and reasons for designation:
| Summary items | Information |
|---|---|
| Recommended Status | Special Concern |
| Alpha-numeric codes | Not applicable |
| Reasons for designation | This large finch is widely distributed across Canada’s forests, but has exhibited significant long-term declines (77-90%) over most of its range, since 1970. Over the past decades, some data suggest a further decline of nearly 40%, while other data indicate stabilization at a lower level. Threats to the species include reduced availability of mature and old-growth mixed wood and conifer forests, collisions with windows, and mortality associated with feeding on grit and salt along roads in winter. |
Applicability of criteria
| Summary items | Information |
|---|---|
| Criterion A (Decline in Total Number of Mature Individuals) | Not applicable. Potentially qualifies as Threatened under A2b, given evidence of a 42% decline in the Canadian breeding population over the most recent 10-year period of Breeding Bird Survey results, but other trend data including Christmas Bird Count results suggest a more stable population over the same period. |
| Criterion B (Small Distribution Range and Decline or Fluctuation) | Not applicable, as EOO and IAO greatly exceed thresholds. |
| Criterion C (Small and Declining Number of Mature Individuals) | Not applicable, as the population greatly exceeds thresholds. |
| Criterion D (Very Small or Restricted Population) | Not applicable, as the population greatly exceeds thresholds. |
| Criterion E (Quantitative Analysis) | Not undertaken. |
COSEWIC history
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2016)
- Wildlife species
- A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
-
Special concern (SC)
(Note: Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.) - A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
-
Not at risk (NAR)
(Note: Formerly described as “Not in any category”, or “No designation required.”) - A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
-
Data deficient (DD)
(Note: Formerly described as “Indeterminate” from 1994 to 1999 or “ISIBD” [insufficient scientific information on which to base a designation] prior to 1994. Definition of the [DD] category revised in 2006.) - A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment and Climate Change Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife species description and significance
Name and classification
The common name of Coccothraustes vespertinus (Cooper 1825) is Evening Grosbeak. The French name is Gros-bec errant (American Ornithologists’ Union 1998). The taxonomy is:
Class: Birds
Order: Passeriformes
Family: Fringillidae
Species: Coccothraustes vespertinus
Morphological description
Evening Grosbeak (Figure 1) is a stocky passerine with colourful plumage and a heavy greenish-yellow bill, slightly smaller than an American Robin (Turdus migratorius) (16.5–18.0 cm; 53–74 g; Gillihan and Byers 2001). Adult males have a dark brown head with a bold yellow supercilium; the brown of the head blends to a bright yellow belly and scapulars, contrasting with a short black tail and black wings with a large white patch on the secondaries. Adult females and juveniles are generally greyish-brown with a yellowish nape and flanks; the primaries are white at the base, while the secondaries are grey and white.
In Canada, Evening Grosbeak is unlikely to be confused with other passerine species due to its distinctive colouration and large bill.
Population spatial structure and variability
There have been no molecular or genetic studies conducted on the Canadian Evening Grosbeak population (Gillihan and Byers 2001).
Long description for Figure 1
Photo of a male Evening Grosbeak, showing the colourful plumage and a heavy greenish-yellow bill. Adult males have a dark brown head with a bold yellow supercilium; the brown of the head blends to a bright yellow belly and scapulars, contrasting with a short black tail and black wings with a large white patch on the secondaries.
Designatable units
Three subspecies of Evening Grosbeak are recognized in North America, two of which breed in Canada, C.v. vespertinus and C.v. brooksi (Clements 2007). C.v. vespertinus breeds from Alberta to the Maritimes and overwinters from the breeding range down to the southeastern United States; C.v. brooksi breeds from British Columbia and southwestern Alberta in the Rocky Mountains (Godfrey 1986) to the southwestern United States and overwinters from the breeding range to as far south as Texas (Clements 2007). Limits of range boundaries between C.v. vespertinus and C.v. brooksi are poorly known, but based on differences in flight calls between the two subspecies, Sewall et al. (2004) suggest the range of C.v. brooksi in Canada possibly includes most of British Columbia, southern Alberta and southwestern Saskatchewan; in contrast, the range of C.v. vespertinus is north and east of these latter limits, extending from Yukon to the Atlantic Coast.
In terms of discreteness, plumage differences between C.v. vespertinus and C.v. brooksi are weak and clinal (Gillihan and Byers 2001): C.v. vespertinus can be distinguished from C.v. brooksi by its broader yellow supercilium and slightly shorter bill (Gillihan and Byers 2001), and females and juvenilesare darker brown (Godfrey 1985). However, Haiman (2011) compared a series of morphological measurements on birds from the two Canadian subspecies and found no significant differences, but noted differences in their flight calls. In general, C.v. brooksi has a high, clear, untrilled flight call compared with C.v. vespertinus, which has a more ringing or trilled call (Sibley 2000; Sewall et al. 2004; Haiman 2011). Flight calls usually associated with C.v. vespertinus have never been reported west of the Rockies in British Columbia, suggesting a geographical segregation between C.v. vespertinus and C.v. brooksi (Sewall et al. 2004; Haiman 2011). The two subspecies also differ by their occupation of different eco-geographical regions: C.v. brooksi being found mainly in the Southern Mountain and in the Pacific regions and C.v. vespertinus mainly in the Boreal and in the Atlantic regions, although there seems to be considerable overlap in the breeding range of the two subspecies along the Continental Divide in the southern Rocky Mountains (Federation of Alberta Naturalists 2007). Finally, discreteness between the two subspecies may also be shown by their distinctive use of wintering grounds: C.v. brooksi in Canadashows little movement and largely remains in British Columbia orwithin adjacent western provinces and states (Brewer et al. 2000), while C.v. vespertinus winters in eastern Canada and from the Midwest and east to the Atlantic Coast (Brewer et al. 2000). Further research is needed to determine genetic differences.
While there appear to be some differences between C.v. brooksi and C.v. vespertinus, they are generally poorly defined. In the absence of other information to support their distinction, a clearly defined separation in range, or evidence of discreteness or significance, it is appropriate to treat Evening Grosbeak as a single designatable unit.
Special significance
Evening Grosbeak is an important predator of the Spruce Budworm (Choristoneura fumiferana) in North America and is known to affect the life cycle of the Spruce Budworm during outbreaks of the insect (Blais and Parks 1964; Crawford et al. 1983; Takekawa and Garton 1984). A total of eight species and 11 subspecies of Spruce Budworm exist in North America (Freeman 1967), with C. feumiferana found from Alberta east to the Maritimes, and Jack Pine Budworm (C. pinus pinus) and a complex of C. occidentalis, C. biennis and C. lambertiana found in southern Alberta and British Columbia (Lumley and Sperling 2010). Evening Grosbeak contributes to the natural control of Spruce Budworm (Blais and Parks 1964; Venier et al. 2009; Venier and Holmes 2010), which is highly detrimental to economic forestry activities involving harvest of fir (Abies spp.) and spruce (Picea spp.) (Takekawa and Garton 1984). The economic value of Evening Grosbeak predation on Spruce Budworm in Washington State was estimated at between $790 and $1,270/km2 in 1979, more than half of the per-km2 cost of spraying with the insecticide Sevin-4-oil ($1,820/km2; Takekawa and Garton 1984).
Across Canada in winter, Evening Grosbeak was formerly a familiar species present at bird feeders, and among the most frequently banded (and subsequently encountered) songbirds in Canada (Brewer et al. 2000), although declines in recent decades have greatly reduced frequency of observation and banding.
No Aboriginal Traditional Knowledge associated with Evening Grosbeak was found for Canada.
Distribution
Global range
The current breeding range of Evening Grosbeak corresponds to boreal and other conifer-dominated forests in each of the provinces and territories, except Nunavut, as well as in the western (Montana, Idaho, Washington, California, Arizona, Nevada, Utah, Colorado, New Mexico, Wyoming and South Dakota, Figure 2) and northeastern United States (Maine, Massachusetts, Vermont, New York, Minnesota, Wisconsin and Michigan in United States (Figure 2). A resident population of Evening Grosbeak is also found in northern and central Mexico (Figure 2). The winter range is largely the same, although in some years individuals irrupt south of the breeding range, occasionally as far as the southern United States.
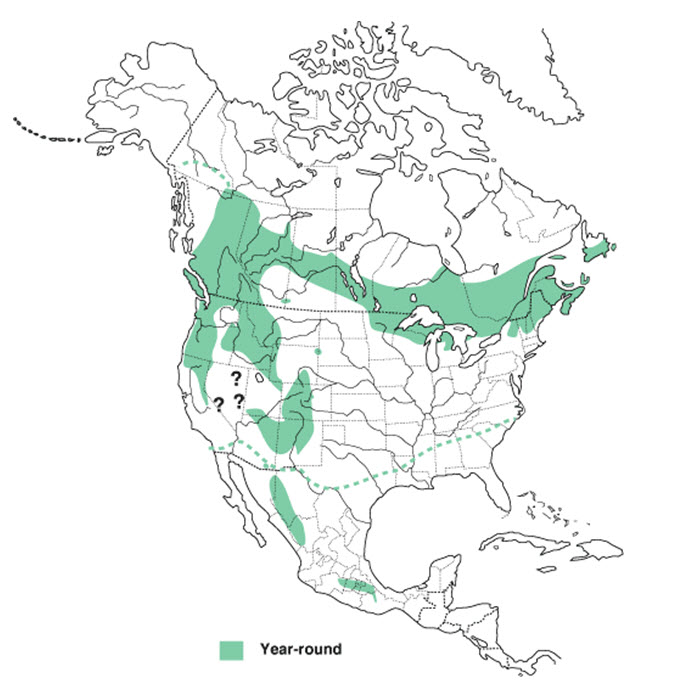
Long description for Figure 2
Map showing the year-round range (shaded areas) of the Evening Grosbeak in North America. A dotted line indicates the northern limit of the winter range. The current breeding range of Evening Grosbeak corresponds to boreal and other conifer-dominated forests in all of Canada’s provinces and territories, except Nunavut, as well as in the western and northeastern United States. A resident population of Evening Grosbeak is also found in northern and central Mexico.
Canadian range
The historical range of Evening Grosbeak has changed considerably since the mid-1800s, when the species was primarily found breeding in northwestern North America and was considered rare to uncommon east of the Rockies (Gillihan and Byers 2001; Sabine 2010). Evening Grosbeak was infrequently found in eastern North America until the late 1800s, when they began a range expansion from the west. Pulses of expansion occurred during 1883-1890, 1901-1917, and between the early 1940s and the mid-1950s (Speirs 1968; Brunton 1994). First documented breeding records in eastern Canada vary from 1920 in Ontario (Godfrey 1985) to around 1940 in the Maritimes (Sabine 2010). The most accepted explanation for the Evening Grosbeak range expansion is the increase in intensity and size of Spruce Budworm outbreaks in the eastern Canada during the first decades of the 1900s (Ouellet 1974; Bolgiano 2004). The range expansion has also been linked to extensive planting of Manitoba Maple (Acer negundo) as windbreaks in the Prairies and as an ornamental tree in eastern cities in the early 1900s (Taverner 1921; Erskine 1992), the increase of Pin Cherry (Prunus pensylvanica)due to large areas of fires and forest clearing in the early 1900s (Brunton 1994) and the increased number of bird feeders installed in inhabited regions throughout North America (Root and Weckstein1994).
The Canadian range of Evening Grosbeak currently extends from Yukon to Newfoundland and Labrador, and includes parts of all provinces and territories except Nunavut (Figure 3). More specifically, the northern limit of the Canadian breeding range of Evening Grosbeak is bounded by the La Biche and Beaver rivers in southern Yukon (Sinclair et al. 2003), the Skeena River area in British Columbia (Martell 2015), Fort Liard in the Northwest Territories (Gillihan and Byers 2001), La Crète in Alberta (Federation of Alberta Naturalists 2007), Cree Lake in Saskatchewan, Opasquia Provincial Park and Moosonee in Ontario (Hoar 2007), southern Hudson Bay and the Mingan Archipelago National Park of Canada Reserve in Quebec (Atlas of the breeding birds of Quebec [ABBQ] 2014), most of the island of Newfoundland, and the Maritime provinces (BSC 2015). The southern limit of its range corresponds to southern British Columbia, east of the Coast Mountains, central Alberta (Red Deer), central Saskatchewan (Saskatoon), southern Manitoba, Lake Simcoe in southern Ontario, southern Quebec (Outaouais and Estrie) and southern New Brunswick and Nova Scotia (Sinclair et al. 2003; Hoar 2007; Federation of Alberta Naturalists 2007 and BSC 2014, 2015; Martell 2015; Figure 3).
The extent of occurrence (EOO) of Evening Grosbeak in Canada covers roughly 6,633,526 km2, while the index of area of occupancy (IAO) is approximately 220,000 km2. The area of occupancy estimate is based on an average of 1 pair/0.2 km2 obtained in 33 plots of grid maps for all of Canada (Table 1; Kennedy et al. 1999) multiplied by an estimated population of approximately 1,100,000 breeding pairs for all of Canada (Partners in Flight Science Committee 2013).
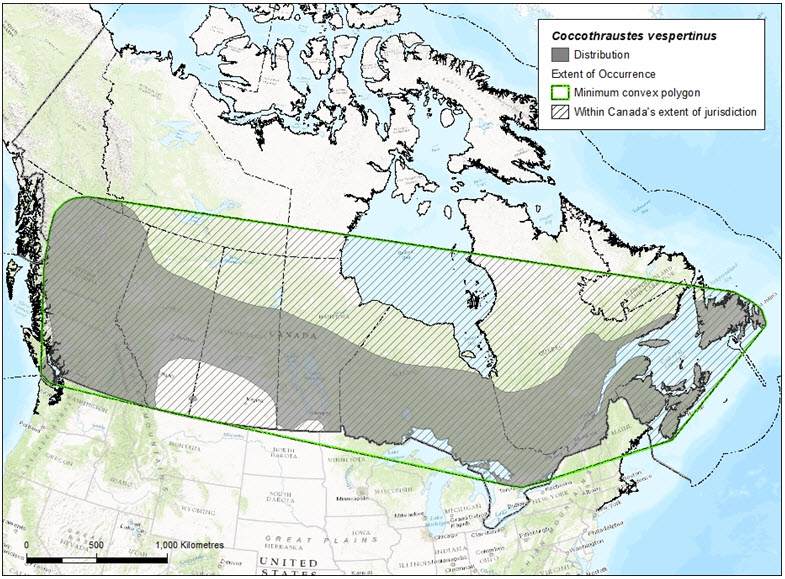
Long description for Figure 3
Map indicating the range (shading) and extent of occurrence (hatching) of the Evening Grosbeak in Canada.
| Provinces | Period | Number of plots | Density ( territories/ km2 ± standard error |
|---|---|---|---|
| British Columbia | 1965-1970 | 2 | 19.0 ± 6.0 |
| British Columbia | 1970-1985 | 2 | 8.3 ± 2.7 |
| Alberta | 1970-1985 | 3 | 3.5 ± 2.5 |
| Manitoba | 1965-1970 | 1 | 18.0 ± 0.0 |
| Ontario | 1965-1970 | 14 | 6.6 ± 4.0 |
| Ontario | 1970-1985 | 4 | 34.8 ± 21.8 |
| Quebec | 1970-1985 | 5 | 0.1 ± 0.0 |
| New Brunswick | 1965-1970 | 2 | 10 |
Search effort
Data on the distribution of Evening Grosbeak in Canada come primarily from the breeding bird atlas work carried out since the 1980s in Ontario (Cadman et al., 2007), Quebec (Gauthier and Aubry 1995; ABBQ 2014), Alberta (Federation of Alberta Naturalists 2007), British Columbia (Martell 2015), Manitoba (BSC 2014), and in the Maritimes (BSC 2015). Since around 2010, checklists filled by birders on eBird (2015) also provide valuable data on Evening Grosbeak distribution in Canada. A model of the distribution of Evening Grosbeak is also provided by the Boreal Avian Modeling Project (2014).
Habitat
Habitat requirements
The distribution of Evening Grosbeak in Canada closely matches the limits of the boreal shield, boreal plain and mountain cordillera ecozones, which total more than 200 million ha of forest (Government of Canada 2013). These regions are generally dominated by fir, spruce, larch (Larix spp.), pine (Pinus spp.) and aspen (Populus spp.).
Nesting habitat of C.v. vespertinus generally features large mature and old mixedwood forest stands with a high proportion of fir, White Spruce (Picea glauca) or Trembling Aspen (Populus tremuloides), with a diversified structure and a relatively open canopy (Langelier 1983; Peck and James 1987; Schieck et al. 1995; Anthony et al. 1996; Hobson et al. 2000; Hobson and Bayne 2000a,b; Cumming and Diamond 2002; Sinclair et al. 2003; Schieck and Song 2006; Venier et al. 2009). In Manitoba and Alberta, Evening Grosbeak has also been reported nesting in urban mixedwood areas (Speirs 1968), but this appears to be generally uncommon.
In coastal British Columbia, C.v. brooksi is found in open forests dominated by Douglas-fir (Pseudotsuga menziesii), Bigleaf Maple (Acer macrophyllum), Paper Birch (Betula papyrifera), Arbutus (Arbutus menziesii), Red-osier Dogwood (Cornus stolonifera), Vine Maple (Acer circinatum) and other species where it forages on berries and seeds (Campbell et al. 2001). In interior British Columbia, it favours stands of Douglas-fir, Ponderosa Pine (Pinus ponderosa), spruce, and Trembling Aspen, with understorey vegetation dominated by Douglas Maple (Acer glabrum), Chokecherry (Prunus virginiana), Pin Cherry, Saskatoon (Amelanchier alnifolia) or Red-osier Dogwood (Merkins and Booth 1998; Campbell et al. 2001). Overall, Evening Grosbeak is found in British Columbia from sea level to high mountainous forest (i.e., up to 1950 m), mainly where the density of berries and seeds is high (Martell 2015). In the northwestern United States, Langelier (1983) found that Evening Grosbeak numbers varied greatly among similarly structured forests, with breeding abundance closely linked to densities of Western Spruce Budworm (C. occidentalis).
The Boreal Avian Modeling Project (BAM 2014) provides a habitat model for Evening Grosbeak using data from hundreds of thousands of point counts along with climate and vegetation cover across Canada. It shows that areas of high-quality habitat based on abundance data are located mainly in northwestern Ontario (BCR 12), central Quebec (BCRs 8, 12, and 14), New Brunswick, Nova Scotia (BCR 14) and eastern and central Newfoundland (Figure 4). However, spatio-temporal coverage in the BAM data is uneven, and the estimated distribution of high versus low quality habitat may be influenced by the spatio-temporal distribution of budworm outbreaks, which has not yet been incorporated in BAM modelling (Van Wilgenburg, pers. comm. 2015).
Winter habitat selection has been poorly studied but seems to be associated with food patches containing berries and seeds (Campbell et al. 2001). In urban and suburban areas, Evening Grosbeaks are attracted to trees that produce large, winged seeds, especially Manitoba Maple, as well as a wide variety of berry-producing ornamental plants (Campbell et al. 2001). It is also a frequent visitor to bird feeders, especially those containing sunflower seeds (Gillihan and Byers 2001).
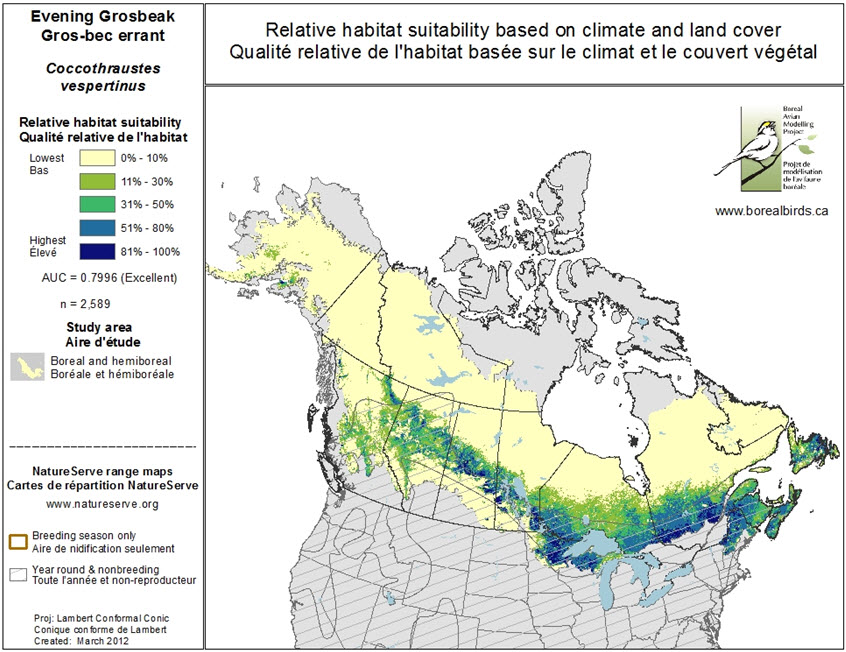
Long description for Figure 4
Map illustrating the relative quality of Evening Grosbeak habitat in the boreal and hemiboreal zone of North America based on climate and vegetation cover. (Habitat quality indicated by colour.)
Habitat trends
Habitat available for C.v. vespertinus has likely increased since 1850 as the interval between forest fires in Canada lengthened, resulting in higher Balsam Fir (Abies balsamifera) abundance across the boreal forest (Bergeron and Leduc 1998). However, since the 1950s there has likely been some decline due to the expansion of large-scale forestry activities, and predicted effects of climate change may cause some further reductions (Williams and Liebhold 1997; Matthews et al. 2004; Bonter and Harvey 2008; Siegel et al. 2014). For example, >20 million ha of boreal forest (mainly mature and old stands) were harvested in Canada between 1975 and 2010, with an additional 6 million ha predicted to be harvested by 2020 (Wells 2011). Clearcut logging can include a short rotation time (i.e., 40-70 years) between harvests, which limits forests from reaching maturity (Schieck and Song 2006) and achieving a diverse forest structure (Bonter and Harvey 2008; Wells 2011), both of which are important for Evening Grosbeak. Moreover, the practice of harvesting Balsam Fir and replanting with faster-growing species such as Black Spruce (Picea mariana) or Jack Pine (Pinus banksiana) in eastern Canada may reduce availability of suitable habitat for Evening Grosbeak through changing forest composition in favour of tree species known to be less susceptible to Spruce Budworm outbreaks (Morin et al. 2008). In northwestern Canada, mature mixedwood forests are also managed using techniques that reduce tree diversity in favour of Trembling Aspen stands (Kirk et al. 1996; Hobson and Bayne 2000). However, increased use of natural regeneration and attempting to replant to maintain similar tree species composition post-harvest may reduce the impacts of forest harvesting in western Canada if forest rotation ages are sufficiently long for favourable Evening Grosbeak habitat to develop (Van Wilgenburg, pers. comm. 2015).
A number of studies suggest that climate change will reduce the total area of Balsam Fir forests in eastern Canada by the end of the century, which could result in a decline in optimal breeding habitat for Evening Grosbeak (Williams and Liebhold 1997; Matthews et al. 2004; Siegel et al. 2014). Moreover, with climate change, fire frequency is expected to increase, especially in central and western Canada (Bergeron and Flannigan 1995; Krawchuk and Cumming 2011), which could result in a decrease in the quantity of suitable habitat for Evening Grosbeak. In portions of the Atlantic provinces (especially Newfoundland and Cape Breton), overbrowsing by Moose (Alces americanus) and deer may limit regeneration of suitable habitat (Gosse et al. 2011; Rae et al. 2014). Under these scenarios, availability of optimal habitat for Evening Grosbeak is likely to decrease in Canada.
Further studies on potential effects of forest management and climate change on Evening Grosbeak habitat are required to better estimate future habitat trends for this species.
Biology
Few reports contain detailed information on the breeding biology of Evening Grosbeak in Canada, but key sources include Campbell et al. (2001), Peck and James (1987), Kennedy et al. (1999) and Gauthier and Aubry (1995). Most studies conducted on Evening Grosbeak during the breeding period are from the western United States (Fee and Bekoff 1986; Bekoff et al. 1987). Dispersal and migration are particularly well studied through banding and recovery of banded birds (Brewer et al. 2000; Hoffman 2009), due to the ease of capturing this species at bird feeders in winter. For a review of species ecology in North America see Gillihan and Byers (2001).
Life cycle and reproduction
Evening Grosbeak is socially monogamous, although polygyny has been observed during major Spruce Budworm outbreaks (Fee and Bekoff 1986). In April, several weeks prior to the breeding period, migrating flocks of Evening Grosbeaks break up and the birds form pairs (Bekoff et al. 1987). In British Columbia and in the western United States, most breeding pairs begin nesting by early June (Bekoff and Scott 1989; Campbell et al. 2001). In the eastern part of the species’ range, courtship and mating occur from early April to May (Shaub 1954; Downs 1956). Nest building, carried out solely by the female (Scott and Bekoff 1991), usually occurs between mid-May and mid-June (Gillihan and Byers 2001). The nest is a loose cup made of coniferous and deciduous twigs or rootlets, with conifers often preferred over deciduous trees (Peck and James 1987). The nest building/egg laying stage lasts an average of six days (Scott and Bekoff 1991).
Evening Grosbeaks usually produce one clutch per year (Gillihan and Byers 2001), and a second clutch is seldom initiated following nest failure (Scott and Bekoff 1991). Mean clutch size of Evening Grosbeak in Ontario is 3 to 4 eggs (n=5 nests; Peck and James 1987). In Ontario, incubation appears to begin between mid-June and mid-July (James et al. 1976; Peck and James 1987). In British Columbia, nests with eggs were found from late April to late July (Campbell et al. 2001). The duration of incubation in Colorado is 12 to 14 days (n=64 nests; Scott and Bekoff 1991).
Generally, the brooding/fledging period occurs from the third week of June to the last week of July (Gillihan and Byers 2001). The young leave the nest in 13 to 14 days (Gillihan and Byers 2001). In Colorado, most nestlings fledged by late July (Scott and Bekoff 1991).
According to a set of models developed for studying the nesting phenology of Evening Grosbeak in Canada using Project NestWatch data (BSC 2013), the general nesting period (from first egg-laying until the young have naturally left the vicinity of the nest) may start somewhere from mid-May to early June and may extend until mid-August to early September, with greater likelihood from end of May to end of August; before and after these dates the probability of an active nest is lower (Rousseu and Drolet, in prep.).
No data are available on the nesting success of Evening Grosbeak in Canada. In Colorado, 35 of 64 nests (54.7%) followed during breeding were successful (fledged at least one young) (Bekoff et al. 1989). Successful nests in Colorado produced on average 2.90 ± 0.98 (SD) young per nest (Gillihan and Byers 2001). In the same study, four nests failed during building, 16 during incubation and nine during brooding (Bekoff et al. 1989). Nest failure was due primarily to predation, severe weather, and nest abandonment during nest building.
There are no data published on response of either clutch size or nesting/fledging success in response to Spruce Budworm density/outbreak conditions, but other boreal forest songbird species such as Tennessee Warbler (Oreothlypis peregrina) are known to show positive numerical responses in clutch size (Venier et al. 2009).
There are no specific data on the age at which Evening Grosbeak reaches sexual maturity (Gillihan and Byers, 2001), but it likely breeds in the first spring following hatching, similarly to other boreal songbirds of its size.
Survival
Hoffman (2009) used banding recovery data on Evening Grosbeak from across the species’ range (breeding and wintering grounds) in North America from 1955-2008 (n=14,224 birds) to model an average annual survival rate (i.e., maximum likelihood estimates of survival and reporting parameters) of 72.4% (95% CI: 35.8 to 85.0%). However, it fluctuated considerably among years, with survival rates high in years with low irruptions and vice versa. This pattern remained consistent until the early 1980s, but after that became less predictable (Hoffman, 2009).
Hoffman (2009) presented three hypotheses to explain the relationship between survival and irruptions. Survival in wintering areas is density-dependent, such that an irruption with a large number of birds exceeds the wintering region’s carrying capacities or results in more birds in “sink” regions, thereby decreasing survival rates. Secondly, there might be a cost to irrupting such that an increased number of birds irrupting results in more mortalities. Finally, periods of high survival increase breeding opportunities, which results in a birth pulse, increasing the number of young birds in the population.
The longevity record for Evening Grosbeak in the wild is 15 years and 3 months (Klimkiewicz and Futcher 1987). The maximum recorded age in Canada is 14 years and 9 months (Brewer et al. 2000). However, survival rates estimated by Hoffman (2009), suggest a mean generation time of 3-4 years.
Dispersal and migration
In Canada, and more specifically east of the Rocky Mountains, Evening Grosbeak is a nomadic species with irruptive migratory movements (Bock and Lepthien 1976; Brewer et al. 2000; Campbell et al. 2001), defined as a massive immigration to a particular region in which the proportion of individuals that participate and the distance they travel varies greatly from year to year (Newton 2008). Banding data from across North America indicate that Evening Grosbeaks show little fidelity to wintering sites (Aubry and Laporte 1990; Brewer et al. 2000). For example, while many other species establish winter territories, data from banded Evening Grosbeaks at nine feeding stations in the city of Québec region between 1983 and 1989 indicate a recapture rate within a season of less than 10% (Aubry and Laporte 1990), while Gillihan and Byers (2001) reported a 2.2% recapture rate within winters in New York State.
Evening Grosbeaks can migrate in large flocks of up to several hundred birds (Gillihan and Byers 2001). Fall migratory movements in Evening Grosbeak have been associated with variability in the food supply in the boreal forest, notably coniferous cone production (Bock and Lepthien 1976; Koenig and Knops 2001; Venier et al. 2009). Evening Grosbeak migratory movements can reach up to 3,400 km (Brewer et al. 2000). C.v. brooksi appears to have relatively short movements, with most recaptured birds from British Columbia being in adjacent provinces or states (i.e., Idaho, Alberta, and Oregon; Brewer et al. 2000). In contrast, C.v. vespertinus banded in Ontario spent other winters from Michigan east to the Atlantic Coast, overlapping broadly with wintering birds captured from the Prairie provinces (Brewer et al. 2000). Meanwhile, birds wintering in Quebec and the Maritime provinces spent other winters in the Appalachian states and southern New England states to Virginia and west to Texas; and birds wintering in the Prairie provinces spent other winters in Minnesota, Wisconsin and Michigan (Prescott 1992; Brewer et al. 2000).
The timing of the onset of spring migration of Evening Grosbeak is variable and can be difficult to ascertain because of the mixing of local residents and winter visitors. In British Columbia, spring movements begin in late March or early April, and reach a peak in the last week of April, then decline through May (Campbell et al. 2001). In Alberta, spring movements are observed primarily from mid-March to mid-May (Pinel et al. 1993). In Quebec, spring migration of Evening Grosbeak occurs in time for breeding to align with the larval and pupal stages of Spruce Budworm, which occur from mid-June to early July (Blais and Parks 1964). Campbell et al. (2001) report that, following the breeding period, small flocks of adults with fledglings become evident in most regions of British Columbia by late July. In British Columbia’s Okanagan Valley, Evening Grosbeaks return to the valleys in early August, with increasing flock sizes building into September (Cannings et al. 1987). Fall movements are poorly documented in British Columbia, but probably occur in September and October (Campbell et al. 2001).
In central and eastern Canada, males usually winter farther north than females (Prescott 1991). This difference may be due to social dominance behaviour or to the ability of males to better tolerate harsher winter conditions (Prescott 1991). No differences were observed in winter distribution between first-year birds and adults (Prescott 1991).
Diet and feeding behaviour
The diet of Evening Grosbeak during the breeding season is composed mainly of invertebrates, especially Spruce Budworm larvae and pupae (>80%; Mitchell 1952; Blais and Park 1964).
Spruce Budworm ecology has been studied intensively in Canada, as it is an important forest pest for the forestry industry. The periodicity of outbreaks varies geographically and outbreaks have a well-documented 25-40 year cycle in the eastern boreal forest (Royama 1984; Price et al. 2013) and every 26 years in British Columbia (Burleigh et al. 2002). Factors that trigger outbreaks include generally consecutive dry summers or spring and autumn droughts (Ives 1974). Gray (2008) found that spatial variability in outbreak pattern was best explained by climate (i.e., average summer minimum and maximum temperature), forest composition, and spatial location. In Ontario, the spatial distribution of historical defoliation by Spruce Budworm was found to be related to winter maximum and minimum temperatures, forest content of Balsam Fir and White Spruce, and spring and summer minimum temperatures (Candeau and Fleming 2011). In eastern Canada, Spruce Budworm outbreaks are linked to the presence of mature and old Balsam Fir stands (Morin et al. 2008). In the Montane Boreal White and Black Spruce biogeoclimatic subzone of British Columbia, budworm development is limited by cool and wet summers (Burleigh et al. 2002).
During the twentieth century, Spruce Budworm outbreaks in eastern Canada occurred in 1910-1920, 1945-1955, and 1968-1985 and increased in intensity over time, defoliating approximately 10, 25, and 55 million hectares, respectively (Morin et al. 2008). Morin et al. (2008) also argue that large-scale clear-cutting, replanting with White Spruce (a tree more susceptible to budworm), and fire suppression may have contributed to larger expanses of susceptible forest, thus leading to bigger budworm outbreaks. Outbreaks occur somewhat synchronously over extensive areas (Candau et al. 1998; Gray et al. 1999; Williams and Liebhold 2000), but duration varies regionally (Candau et al. 1998; Gray et al. 1999).
Blais and Park (1964) proposed that during spring migration and probably during the beginning of the breeding season, Evening Grosbeaks are attracted to outbreak areas in large numbers. The year following collapse of an outbreak, very few Evening Grosbeaks are usually seen in the area, suggesting they left for other forests that have higher budworm densities (Blais and Park 1964). Banding data from across North America indicate that during periods of low budworm density, the maximum likelihood estimates of survival and reporting parameters of Evening Grosbeaks are very low in comparison to levels during Spruce Budworm outbreaks (Hoffman 2009).
To a lesser extent, Evening Grosbeaks also feed on other defoliating insects that occur across the boreal forest during the breeding season such as Forest Tent Caterpillar (Malacosoma disstria), Jack Pine Budworm, Larch Sawfly (Pristiphora erichsonii), and Large Aspen Tartrix (C. conflictana)(Sutton and Tardif 2008; Government of Canada 2011). For example, Forest Tent Caterpillar has a long history of periodic outbreaks in Canada (especially in mature and old Trembling Aspen forest), which usually last 3-6 years in any one locality. Between 1969 and 1980, outbreaks were reported in every province except Newfoundland and Labrador. Provinces with highest outbreaks areas were Manitoba, Alberta, and Ontario (Price et al. 2013). More studies are needed to assess the importance of these insects in the diet of Evening Grosbeak.
The stomach contents of 88 winter specimens from across the species’ range contained 40% fruit seeds, 38% winged seeds, 15% conifer seeds and 7% miscellaneous seeds and other plant material (Gabrielson 1924). The importance of tree seed-crops in winter, particularly of the genera Acer, Pinus, Cornus, Prunus, and Juniperus have been also reported by other studies (Martin et al. 1951; Gillihan and Byers 2001). Koenig and Knops (2001) found that Evening Grosbeaks irrupt out of their normal winter range when the seed-crop productivity of coniferous trees (mainly from genera Abies, Picea, and Pinus) is low, which happens every 2-3 years. A prevalent hypothesis is that widespread masting in the boreal forest at high latitudes is driven primarily by favourable climate during the two to three consecutive years required to initiate and mature seed crops in most conifers (Koenig and Knops 2001). Seed production is usually much reduced in the years following masting, driving Evening Grosbeaks to search elsewhere for food and overwintering habitat.
In winter, flocks of Evening Grosbeaks are often observed along roadsides ingesting salt particles and grit (Campbell et al. 2001). It is during winter that Evening Grosbeak has the greatest need to ingest grit to aid in the digestion of seeds (Mineau and Brownlee 2005).
Interspecific interactions
During the breeding period, agonistic interactions have been reported with several species of forest birds, including Hairy Woodpecker (Picoides villosus), Eastern Phoebe (Sayornis phoebe), American Robin and Brown-headed Cowbird (Molothrus ater; Downs 1956). At feeding stations, it is reported that Common Redpoll (Carduelis flammea) and Pine Siskin (Spinus pinus), which are unable to husk sunflower seeds, look for the presence of Evening Grosbeaks and feed on particles of sunflower seeds falling from their bills (Balph and Balph 1979).
There are a number of known predators of adult Evening Grosbeaks, including domestic cats (Bekoff 1995; Blancher 2013) and several diurnal and nocturnal raptors (Gillihan and Byers 2001). Nest predators include Common Raven (Corvus corax; Bekoff et al. 1989).
Home range and territory
Evening Grosbeak is not territorial during the breeding period (Scott 1990) and could nest in small groups or loose colonies (Speirs 1968). Recapture data (n=10 birds) from across eastern Canada indicate that 80% of birds had dispersed up to 950 km from the previous year’s breeding sites, suggesting that this species shows little breeding site fidelity (Brewer et al. 2000).
Home range size varies considerably and is likely associated with budworm density (Venier et al. 2009). In Ontario for example, the number of Evening Grosbeak territories counted using territory-mapping techniques in a mixedwood forest between 1979 and 1983 increased from 0 territories/km2 during low Spruce Budworm levels to 22.2 territories/km2 during epidemic levels (Venier et al. 2009). Data using the same survey method across Canada show a lower density in Ontario from 1965-1970 than from 1970-1985, which corresponded with a major Spruce Budworm outbreak (Kennedy et al. 1999; Table 1). In British Columbia, densities were high from 1965-1970 and lower during the budworm outbreak occurring in Eastern Canada (Table 1). Table 1 shows that territory densities in Quebec were low during the peak of the outbreak, which is difficult to explain. Although these density data provide information on breeding density for several provinces, results should be interpreted with caution, as different plots were surveyed in the two time periods and sample size is generally low.
Behaviour and adaptability
Evening Grosbeak is well known for its gregarious behaviour in winter where wintering groups of more than 250 birds can be seen (eBird 2015), especially at bird feeders.
Since the 1900s, Evening Grosbeak is also known to have adapted to the large scale planting of ornamental Manitoba Maples as farm windbreaks as well as in parks, along roadsides and in cities in eastern Canada (Taverner 1921; Forbush 1929; Speirs 1968; Gillihan and Byers 2001). Evening Grosbeaks appear to show a clear preference for the seeds of Manitoba Maple, which stay on the tree during fall and winter (Taverner 1921).
Population sizes and trends
Sampling effort and methods
Christmas bird count
The Christmas Bird Count (CBC), which began in the early 1900s, estimates Evening Grosbeak population trends in North America in winter (Cornell University 2014) and is considered one of the most important sampling efforts to assess Evening Grosbeak trends across the species’ range. At each CBC, observers record all species observed within a 24-km diameter circle on a single day between December 14 and January 5 (Sauer et al. 1996). The main advantage of this method lies in the fact that it surveys Evening Grosbeak throughout the species’ wintering range (Sauer et al. 1996). The CBC can be particularly valuable for species such as Evening Grosbeak, which have part of their breeding range in relatively inaccessible regions (Cornell University 2014). However, given the irruptive nature of Evening Grosbeak movements, there can be high interannual variation in results, and therefore long-term trends are more meaningful than those for shorter periods. Bolgiano (2004) provides a trend of the percent of CBCs reporting Evening Grosbeak between 1940 and 2001, and Environment Canada (2014) has conducted a trend analysis of the abundance index (effort-adjusted) for the period 1970-2012.
Project FeederWatch
Project FeederWatch (PFW) is a joint program of Bird Studies Canada and the Cornell Lab of Ornithology (Lepage and Francis 2002). PFW is a long-term North American winter survey of birds (since 1976) that visits feeders at backyards and elsewhere. Today, approximately 15,000 citizens participate in this project. The participants must periodically report the largest number of individuals of each species they see at their feeders from November to April. The purpose of PFW is to monitor movements of winter bird populations across North America and to identify long-term trends in species abundance and distribution. It is thus possible to study the variation in the percentage of feeders visited by Evening Grosbeaks and the average number of individuals per site (Bonter and Harvey 2008). The advantage of PFW is that it follows a more detailed protocol than the CBC, given that it is conducted only at feeders (Lepage and Francis 2002). The trends described by PFW for Evening Grosbeak are generally correlated with CBC trends (Lepage and Francis 2002). The PFW database is also used for associating bird occurrence and abundance observations with gridded climate data because of its continuous winter sampling protocol, annual time series (1989-present), and large sample size (>10,000 participants per year; Strong et al. 2015).
North american breeding bird survey
The North American Breeding Bird Survey (BBS) is a survey of breeding bird populations in North America (Sauer et al. 2014). Data on the abundance of breeding birds including Evening Grosbeak are collected by volunteers on fixed 39.2-km routes consisting of 50 stops with a 400-m radius spaced 0.8 km apart (Environment Canada 2014). In Canada, the surveys are generally conducted in June, during the breeding period of most birds. BBS routes are started at 0.5 h before sunrise and take approximately five hours to complete.
For species such as Evening Grosbeak with fluctuating annual numbers, a “rolling trend” graph can provide a more effective overview of trends than the standard 10-year trend based just on end points. The rolling trend graph plots a series of 10-year trend values ending in each year over a specific period, thereby showing how the trends change over time. It highlights how much the interpretation of a short-term trend depends on the particular year of assessment, and whether the trend changes over time (Smith, pers. comm. 2016).
Although BBS is conducted across Canada, there are some caveats with respect to monitoring Evening Grosbeak: 1) incomplete sampling of the Canadian population, because BBS coverage is sparse in the northern parts of the breeding range (Machtans et al. 2014); 2) relatively poor detectability at 3-min point counts due to the low frequency of songs and calls; and 3) a possible detection bias due to the variability in the peak of the species’ breeding period between years and/or regions, which could be as late as early July to early August (Drolet, pers. comm. 2016). All the same, because these limitations remain consistent over time and the BBS has a standardized design and relatively stable extent of coverage, the BBS is believed to be valuable for assessing Evening Grosbeak trends.
Breeding bird atlases
Comparison of breeding bird atlases conducted in the 1980s-1990s and again with similar methods since 2000 in Alberta (Federation of Alberta Naturalists 2007), Ontario (Cadman et al. 2007), Quebec (ABBQ 2014), and the Maritimes (BSC 2015) provide valuable data at the provincial/regional scale. Data were gathered by volunteers in 10x10 km squares during the breeding season (Cadman et al. 2007). For some provinces, the percent change in the distribution of Evening Grosbeak over a period of 20 years was calculated by comparing the percentage of squares with breeding evidence in the first atlas period to that in the second atlas period, adjusting for observation effort (e.g., squares having a minimum of 20 hours of atlassing; Cadman et al. 2007). In the second round of atlassing, relative abundance was also estimated by point counts in all provinces except Alberta.
The main constraint of this method, which is based on breeding evidence, lies in the fact that comparisons are made on the basis of squares/plots with or without bird occurrences during the breeding period rather than directly on the basis of species abundance. Nonetheless, the comparison of the probability of observation between the two survey periods is considered adequate for estimating Evening Grosbeak trends because of the large number of samples gathered during the two periods and the standardized methodology used (Cadman et al. 2007). In addition, this program generally covers the entire breeding range of the species in a given region (Cadman et al. 2007).
Forest bird monitoring program
The Forest Bird Monitoring Program (FBMP) began in Ontario in 1987 to provide information on population trends and habitat associations of birds that breed in the forest interior (Government of Canada 2008). Each year, between 50 and 150 sites are surveyed by volunteers, who make two 10-minute visits to five point-count stations per site. Although the FBMP primarily targets 52 species (not including Evening Grosbeak), it yields data on occurrence and relative abundance for more than 100 species at those sites. The program was designed to investigate spatial and temporal patterns in mature forest-related birds, with monitoring sites selected in off-road sites in core areas of large, mature forests that are protected from active forest management. These surveys are intended to assess populations within intact forest ecosystems, and can be used for comparison to general population trends from randomly selected sites across the broad landscape (Francis et al. 2009). Evening Grosbeak population trends are currently provided for the period 1987-2007 (Government of Canada 2008).
Checklist program in Quebec (Étude des populations d’oiseaux du Québec, ÉPOQ)
The Checklist program in Quebec began in 1969. It manages thousands of checklists produced by volunteers and is the basic reference for determining Evening Grosbeak population trends in Quebec (Cyr and Larivée 1995). Similar to the CBC, ÉPOQ winter data cover the species’ entire wintering area in Quebec (Cyr and Larivée 1995). ÉPOQ also has the advantage of covering areas of the boreal forest during the breeding season that are not covered by BBS. Unlike the BBS however, it does not systematically cover the same sectors from year to year, and is therefore subject to a larger detection bias. ÉPOQ trends are nonetheless correlated with BBS trends (Dunn et al. 1996).
Migration count at Observatoire d’oiseaux de Tadoussac (OOT)
Since 1996, the OOT has conducted annual monitoring of migratory Evening Grosbeaks in the fall using a standardized protocol of visual counting to study fluctuations in the abundance and productivity of several boreal species (Explos-Nature 2016). Each morning during the fall migratory period (August 24 -November 25), birds are visually counted for 5 hours when they pass over an observer located on the coast of the St. Lawrence River (Explos-Nature 2016). For Evening Grosbeak, an average of 350 hours/year was used to correct the abundance index by search effort (Explos-Nature 2016).
Fort Liard landbird monitoring program
The Canadian Wildlife Service undertook a 14-year monitoring project between 1998 and 2011 in the southwestern Northwest Territories to compare breeding songbird trends obtained from point counts and BBS surveys in a relatively undisturbed boreal landscape with a more disturbed region (i.e., northern Alberta) and at the national level (Machtans et al. 2014).
Abundance
BBS data from Canada suggest that abundance is greatest in southern British Columbia and the Maritime provinces (up to 15 birds/route), intermediate in Ontario and Quebec, and lowest in the Prairie provinces (<2.8 birds/route) (Environment Canada 2014; Figure 5). The extent of BBS coverage in North America and relative abundance data over the period 1987-2006 is shown in Figure 5.

Long description for Figure 5
Map illustrating the relative abundance (average number of birds per route per year) of the Evening Grosbeak calculated for each square of latitude and longitude between 1987 and 2006 during the breeding period according to the North American Breeding Bird Survey. (Relative abundance indicated by shading.)
According to the Partners in Flight Population Estimates database (Partners in Flight Science Committee 2013), which is based on a combination of BBS count data from 1998-2007 and Ontario Breeding Bird Atlas point counts from 2001-2005 (Blancher et al. 2013), the North American Evening Grosbeak population is an estimated 3.9 million breeding individuals. Canada accounts for 56.9% of the North American population, or roughly 2.2 million breeding birds (Table 2). Across Canada, the largest population of Evening Grosbeak is in British Columbia with an estimated 600,000 breeding adults, corresponding largely to C.v. brooksi (Table 2). The highest numbers of C.v. vespertinus are in Ontario and Quebec with 500,000 breeding individuals each, though density is greatest in Nova Scotia (Table 2). These abundance estimates likely correspond to normal population levels between peaks of Spruce Budworm outbreaks because data are mostly available for years with low to intermediate Spruce Budworm abundances. Therefore during outbreaks, numbers are expected to be considerably greater.
| Province / State / Territory |
Population size (adult birds) |
% of global population | Relative abundance from BBS (birds/route) |
Standard deviation of relative abundance | Number of BBS routes | Number of routes detecting Evening Grosbeak |
|---|---|---|---|---|---|---|
| YK | 0 | 0 | n/a | n/a | n/a | n/a |
| NWT/NU | 0 | 0 | n/a | n/a | n/a | n/a |
| BC | 600,000 | 14.5 | 0.69 | 0.19 | 111 | 49 |
| AB | 60,000 | 1.4 | 0.10 | 0.04 | 137 | 15 |
| SK | 40,000 | 1 | 0.07 | 0.05 | 55 | 7 |
| MB | 200,000 | 5 | 0.37 | 0.18 | 66 | 16 |
| ON | 500,000 | 12.8 | 0.59 | 0.13 | 221 | 55 |
| QC | 500,000 | 13.4 | 0.47 | 0.08 | 99 | 70 |
| NB | 110,000 | 2.6 | 1.62 | 0.32 | 31 | 24 |
| NS | 200,000 | 5.4 | 4.42 | 0.70 | 29 | 29 |
| PEI | 1,300 | 0 | 0.25 | 0.20 | 4 | 2 |
| NL | 30,000 | 0.8 | 0.09 | 0.06 | 23 | 8 |
| Total | 2,211,300 | 56.9 | blank | blank | blank | blank |
According to the Ontario Breeding Bird Atlas, Evening Grosbeak is most abundant (1.21-14.33 birds / 25 point-count stations) in Bird Conservation Region 12 (Boreal Hardwood Transition) (Hoar 2007), a region characterized by coniferous and northern hardwood forests, nutrient-poor soils, and numerous forested wetlands (NABCI International 2014).
In British Columbia, the highest probability of observation occurs in the Southern Interior Ecoprovince, spilling into neighbouring portions of the Coast and Mountains, Southern Interior Mountains and Central Interior ecoprovinces (Martell 2015). Atlas point counts indicate highest abundance between 1,000 and 1,250 m, corresponding with the mid-elevation plateaus in this area, and in the Interior Douglas-fir and Montane Spruce biogeoclimatic zones that dominate those plateaus (Martell 2015).
Fluctuations and trends
Until the late 1800s, Evening Grosbeak bred mainly west of the Rocky Mountains and was considered a rare visitor to the eastern provinces (Gillihan and Byers 2001; Bolgiano 2004). Since the early 1900s (Taverner 1921), it has expanded its range to eastern Canada (Bolgiano 2004). The increase in the Evening Grosbeak population in eastern Canada has been variably attributed to the planting of Manitoba Maples, colonization of large areas by Pin Cherry, and attraction to Spruce Budworm outbreaks in eastern Canada in the 20th century (Morris et al. 1958; Blais and Parks 1964; Speirs, 1968; Ouellet 1974; Crawford et al. 1983; Bolgiano 2004), arising from the reduced interval between forest fires and allowing for an increase of Balsam Fir abundance which in turn generated particularly severe Spruce Budworm outbreaks from 1910 onward (Bergeron and Leduc 1998).
Christmas bird count
Analysis of CBC data for North America, which reflect primarily the Canadian breeding population, indicate a significant decline of -3.4% per year from 1970-2012 (95% CI: -6.5 to -0.9; Smith, unpubl. data), representing a cumulative decline of 76.6% over 42 years. From 2002-2012, the North American trend was a non-significant increase of 3.1% per year (95% CI: -1.5 to 8.8; A. Smith, unpubl. data). This latter trend appears to have started around 1997, following the long declining trend documented from 1970 (Figure 6).
Within Canada, CBC data show a non-significant decline of -1.1% per year between 1970 and 2012 (95% CI: -4.4 to 2.3; Table 3, A. Smith, unpubl. data), and a short-term (2002-2012) non-significant increase of 4.4% per year (95% CI: -1.6 to 12.3; Table 3, Smith unpubl. data). The annual abundance index fluctuated greatly during an overall decline from 1971 to 1998, but largely stabilized between 1999 and 2012 (Figure 6). The 10-year trend ending in any particular year has fluctuated over time, with declines steepest in the early 1980s and from 1996 to 2002, contrasting with periods of stability or modest increases from 1985 to 1991 and 2008 to 2012 (Figure 7; Smith, unpubl. data). However, these rolling trends have wide credible intervals, in all but two years crossing zero. At the provincial scale, CBC data show the greatest long-term (1970-2012) declines in Ontario and New Brunswick, at -7.1% and -11.9% per year respectively, while there are have been large short-term increases in Alberta, Saskatchewan, and Quebec (Table 3).
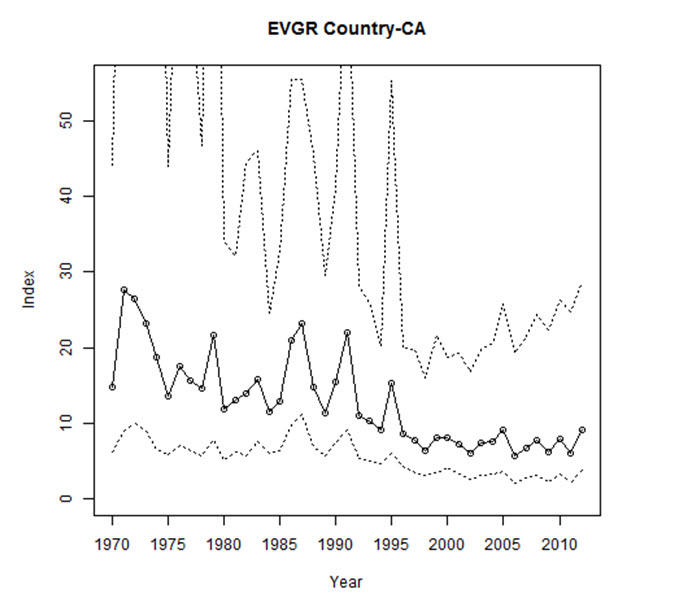
Long description for Figure 6
Chart containing expected counts (with 95-percent confidence intervals) of Evening Grosbeaks from an average Christmas Bird Count circle in Canada, from 1970 through 2012, after accounting for variations in effort within circles, and sampling bias among circles, years, and regions.
| Province | Period | Annual trend | LCI | UCI | Number of CBCs |
|---|---|---|---|---|---|
| Canada | 1970-2012 | -1.12 | -4.37 | 2.29 | 318 |
| Canada | 2002-2012 | 4.36 | -1.64 | 12.3 | 271 |
| BC | 1970-2012 | 0.84 | -4.24 | 6.14 | 96 |
| BC | 2002-2012 | 1.53 | -5.67 | 11.0 | 87 |
| AB | 1970-2012 | -3.00 | -7.67 | 1.93 | 52 |
| AB | 2002-2012 | 12.90 | -2.05 | 36.4 | 45 |
| SK | 1970-2012 | -2.57 | -11.5 | 7.36 | 14 |
| SK | 2002-2012 | 35.0 | -17.9 | 155.0 | 13 |
| MB | 1970-2012 | 0.77 | -4.16 | 6.3 | 27 |
| MB | 2002-2012 | -0.92 | -16.5 | 21.0 | 20 |
| ON | 1970-2012 | -11.9 | -22.3 | -0.68 | 52 |
| ON | 2002-2012 | -12.5 | -48.5 | 44.7 | 46 |
| QC | 1970-2012 | -0.78 | -5.76 | 4.41 | 19 |
| QC | 2002-2012 | 23.8 | 2.89 | 48.4 | 13 |
| NB | 1970-2012 | -7.06 | -11.4 | -1.74 | 20 |
| NB | 2002-2012 | 2.96 | -10.1 | 20.7 | 16 |
| NS | 1970-2012 | -4.17 | -12.1 | 5.17 | 36 |
| NS | 2002-2012 | -14.10 | -38.2 | 13.6 | 28 |
| NT | 1970-2012 | -1.63 | -5.38 | 2.23 | 3 |
| NT | 2002-2012 | -1.2 | -13.5 | 13.0 | 3 |
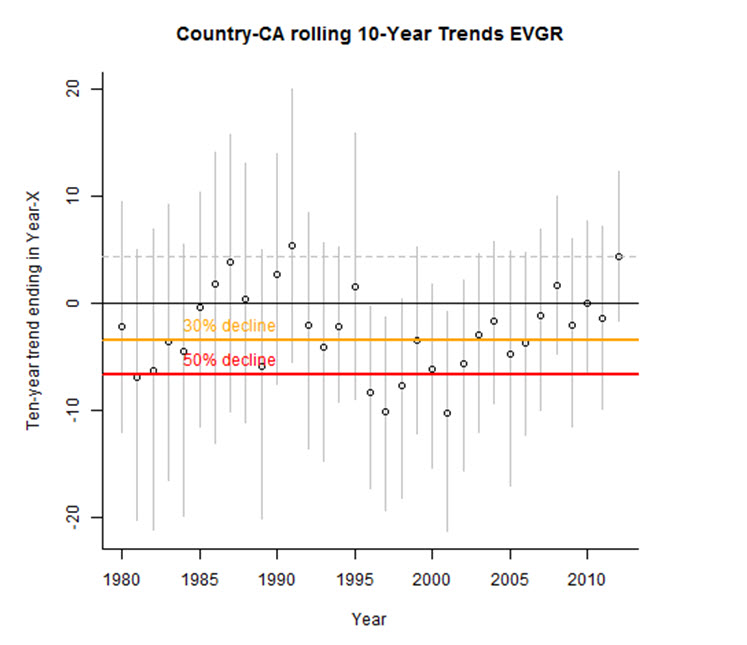
Long description for Figure 7
Chart illustrating 10-year rolling trend in Evening Grosbeak abundance between 1980 and 2012 in Canada based on Christmas Bird Count data.
Project FeederWatch
Data analysis for the period from 1989 to 2006 show a contraction of the species’ winter range, primarily in the Rocky Mountain, Great Lakes, Atlantic Canada and Appalachian regions (Bonter and Harvey 2008). Between 1989 and 1994, the mean count at feeders was highest in Quebec, Maritimes, Saskatchewan, Manitoba, Northwest Territories, British Columbia, and Alberta (mean count >16 birds). Between 1995 and 2000, there was a substantial range contraction with a decrease in the mean count of Evening Grosbeaks everywhere except in the Northwest Territories and Newfoundland, where counts remained relatively high (Bonter and Harvey 2008). Between 2001 and 2006, Evening Grosbeaks continued to decline significantly at feeders across Canada except in Newfoundland, Saskatchewan, and Manitoba. The mean flock size of Evening Grosbeaks at feeding stations showed a significant decline of 27%, from 11.8 birds for the period 1989-1994 to 8.6 birds for the period 2001-2006 (Bonter and Harvey 2008). In addition, the proportion of sites reporting Evening Grosbeaks also decreased from 0.194 in 1989-1994 to 0.097 in 2001-2006, a 50% decline (Bonter and Harvey 2008).
Recent data analysis from this project suggest that the average flock size of Evening Grosbeak at feeders has fluctuated somewhat between 1990 and 2014, but in most regions showing only a small overall decline; the percentage of feeders visited by Evening Grosbeaks was greatest in western provinces, with particularly high values between 1989 and 1993 (Figure 8; Cornell University 2015).
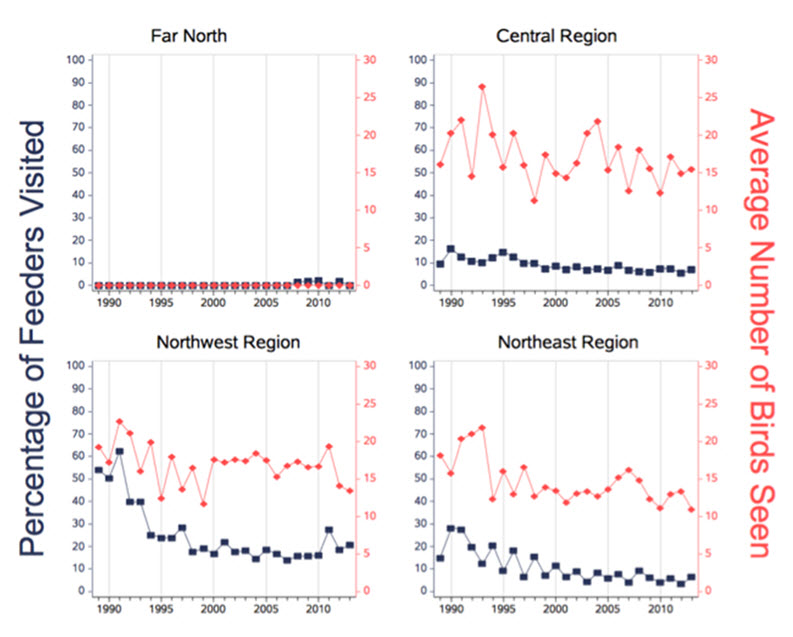
Long description for Figure 8
Four chart panels illustrating trends in the percentage of feeders visited by Evening Grosbeaks and the average number of these birds seen at feeders in four regions of North America from Project FeederWatch, between 1989 and 2014. The regions are: Northwest Territories; British Columbia and Alberta; Saskatchewan and Manitoba; and Ontario, Quebec, and the Atlantic provinces.
North American breeding bird survey
For the period 1970-2014, Evening Grosbeak has a trend of -5.2% per year across Canada (n = 448 routes, overall reliability level = high, 95% credible interval [CI]: -6.5, -4.0), which represents a 90% population decline over 44 years; for 2004-2014, there was a significant decline of 5.0% per year (n = 379 routes, overall reliability level = medium, 95% CI: -7.9, -2.0), or a 42% cumulative decline (Figure 9, Table 4; Smith, pers. comm. 2016). The 10-year trend ending in any particular year fluctuates greatly, near a 50% cumulative decline from 1980 to the early 1990s, then slowing down for approximately a decade before experiencing the steepest rate of decline between 2003 and 2007, and then again abating somewhat since then, although remaining distinctly negative (Figure 10; Smith, unpubl. data). Although the rolling trends have wide credible intervals, they have been entirely negative since 1999, and annual estimates have been at or below the threshold of a 30% decline over ten years throughout this period.
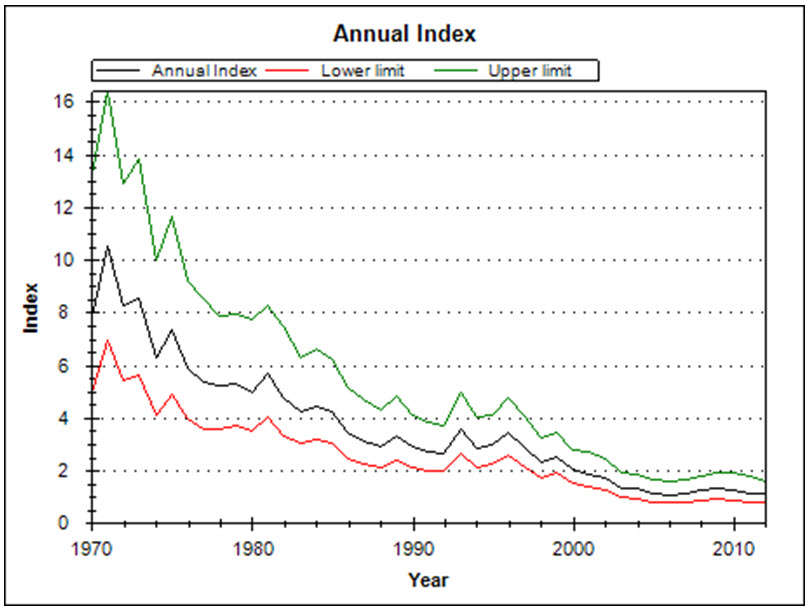
Long description for Figure 9
Chart showing the annual abundance index for Evening Grosbeaks in Canada between 1970 and 2012 (with 95-percent confidence intervals) according to a hierarchical Bayesian model of Breeding Bird Survey data.
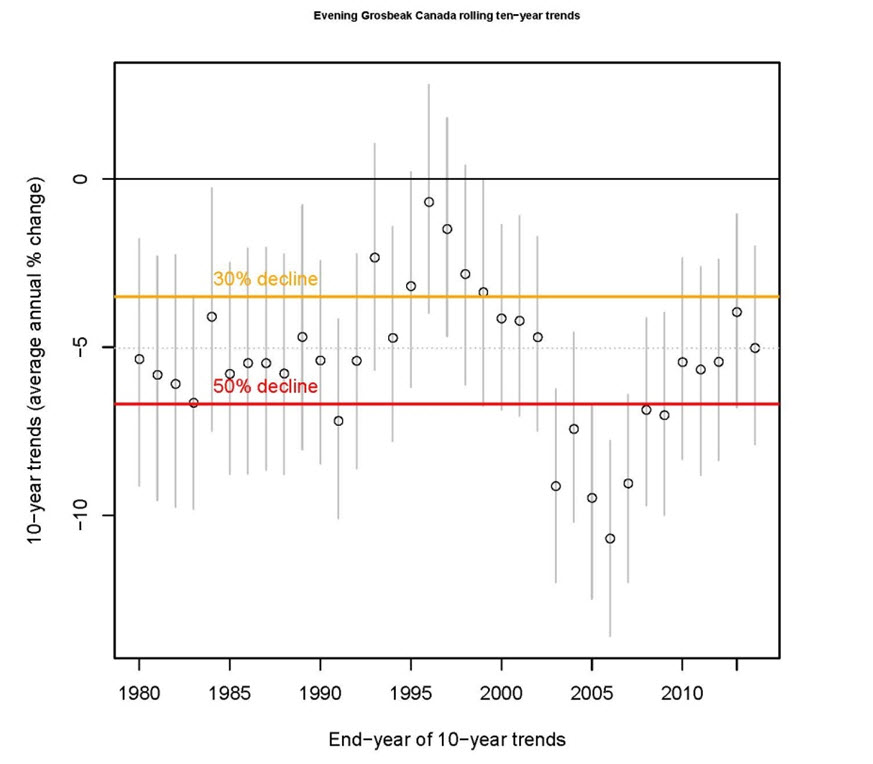
Long description for Figure 10
Chart illustrating the 10-year rolling trend in Evening Grosbeak abundance between 1980 and 2014 in Canada based on Breeding Bird Survey data.
| Province | Period | Annual trend | LCI | UCI | Number of routes |
|---|---|---|---|---|---|
| Canada | 1970-2014 | -5.19 | -6.51 | -4.04 | 448 |
| Canada | 2004-2014 | -5.03 | -7.90 | -2.00 | 379 |
| British Columbia | 1970-2014 | -4.07 | -7.18 | -1.42 | 85 |
| British Columbia | 2004-2014 | -1.08 | -9.61 | 8.47 | 66 |
| Alberta | 1970-2014 | -1.03 | -7.32 | 3.37 | 33 |
| Alberta | 2004-2014 | -0.56 | -15.6 | 12.0 | 30 |
| Saskatchewan | 1972-2014 | -2.51 | -8.30 | 2.22 | 13 |
| Saskatchewan | 2004-2014 | -1.73 | -13.5 | 15.5 | 6 |
| Manitoba | 1970-2014 | -4.02 | -10.3 | 1.91 | 7 |
| Manitoba | 2004-2014 | -6.41 | -12.7 | -0.08 | 21 |
| Ontario | 1970-2014 | -5.81 | -7.97 | -3.65 | 102 |
| Ontario | 2004-2014 | -5.86 | -11.1 | -1.05 | 87 |
| Quebec | 1970-2014 | -5.14 | -7.00 | -3.21 | 137 |
| Quebec | 2004-2014 | -5.82 | -10.4 | -1.52 | 102 |
| Newfoundland and Labrador | 1979-2014 | -1.44 | -6.81 | 4.2 | 7 |
| Newfoundland and Labrador | 2004-2014 | -2.09 | -15.2 | 5.82 | 7 |
| New Brunswick | 1970-2014 | -8.95 | -11.5 | -6.45 | 35 |
| New Brunswick | 2004-2014 | -11.2 | -22.0 | 0.75 | 27 |
| Nova Scotia and Prince Edward Island | 1970-2014 | 1.3 | -2.16 | 4.84 | 36 |
| Nova Scotia and Prince Edward Island | 2004-2014 | -6.09 | -17.2 | 5.64 | 33 |
Significant long-term declines of 4.1 to 9.0% per year have also been estimated for British Columbia, Ontario, Quebec and New Brunswick (Table 4); Ontario and Quebec also have significant short-term declines, as does Manitoba (Table 4; Smith, pers. comm. 2016). The only positive long-term trend is in Nova Scotia and Prince Edward Island (+1.3%) but it is not significant (n = 36 routes, 95% CI: -2.2, 4.8).
In the United States, there was a significant decline of -2.7% per year from 1970 to 2014, equating to a 70% population decline over 44 years (Smith pers. comm. 2016).
Alberta breeding bird atlas
Evening Grosbeak distribution declined in the Boreal Forest and Parkland Natural Regions between the first (1987-1991) and second (2000-2005) survey periods, but remained stable in the Foothills Natural Region, where frequency of occurrence was highest (Federation of Alberta Naturalists 2007).
Ontario breeding bird atlas
The comparison of Evening Grosbeak distribution between the first (1981-1985) and second (2001-2005) survey periods shows a significant decline of 30% in the probability of observation for the province as a whole (Figure 11; Hoar 2007). A significant decline of between 10 and 82% was observed in each of the four atlas regions, and was most pronounced in the Hudson Bay Lowlands.
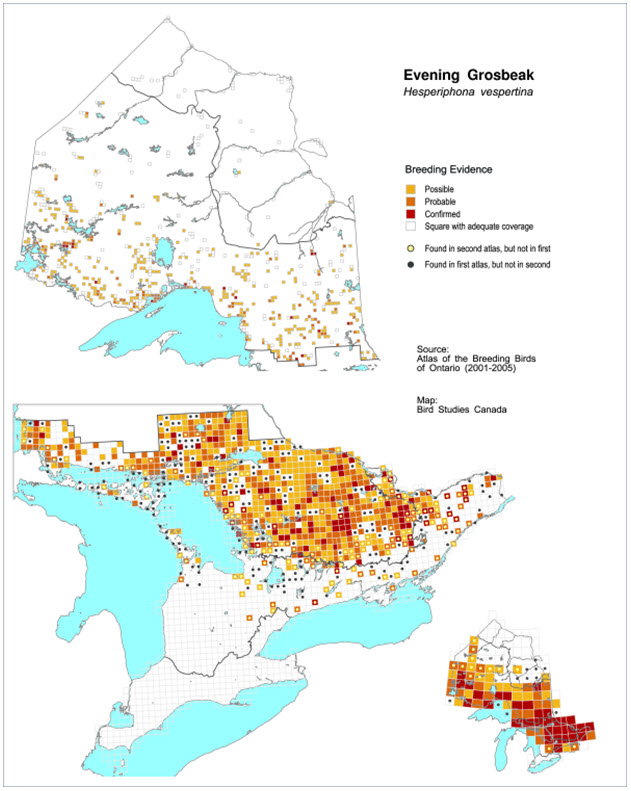
Long description for Figure 11
Three maps. The top map showing the distribution (based on grid cells) of Evening Grosbeaks in Ontario from 2001 to 2005 based on Ontario Breeding Bird Atlas data. The other two maps (of southeastern Ontario and Ontario as a whole) indicate cells in which Evening Grosbeak was found only in the first atlas period (1980 to 1985), and cells in which the birds were found only in the second atlas period (2001 to 2005).
Quebec breeding bird atlas
The visual comparison of regions where Evening Grosbeak was assessed as probable and confirmed breeding between the two atlas periods suggests an increase in squares in the Abitibi-Témiscamingue, North Shore, and Bas-St-Laurent/Gaspésie regions and a decrease in the Outaouais, Laurentian and Mauricie regions during the 2010-2014 period (Figure 12; ABBQ 2014). In all other regions, the number of squares with observations of probable and/or confirmed breeders were similar (Figure 12; ABBQ 2014), but statistical analyses are needed to provide greater accuracy around these estimates.
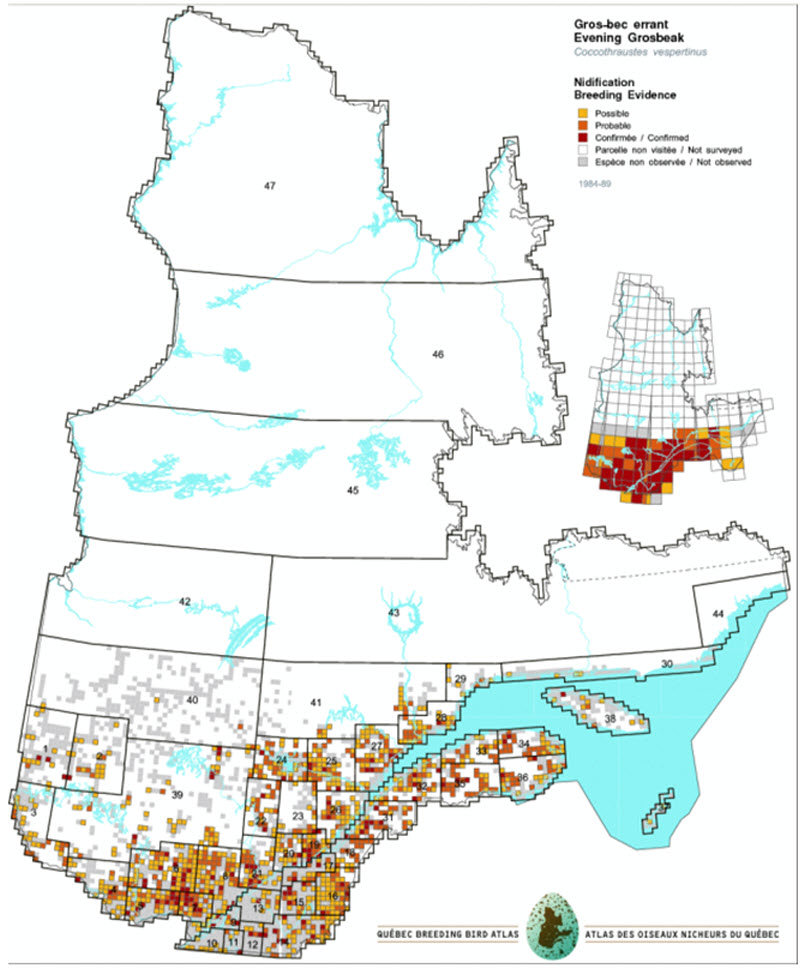
Long description for Figure 12a
Map panel illustrating the distribution (based on grid cells) of Evening Grosbeaks in Quebec during the first Quebec Breeding Bird Atlas period (1984 to 1989).
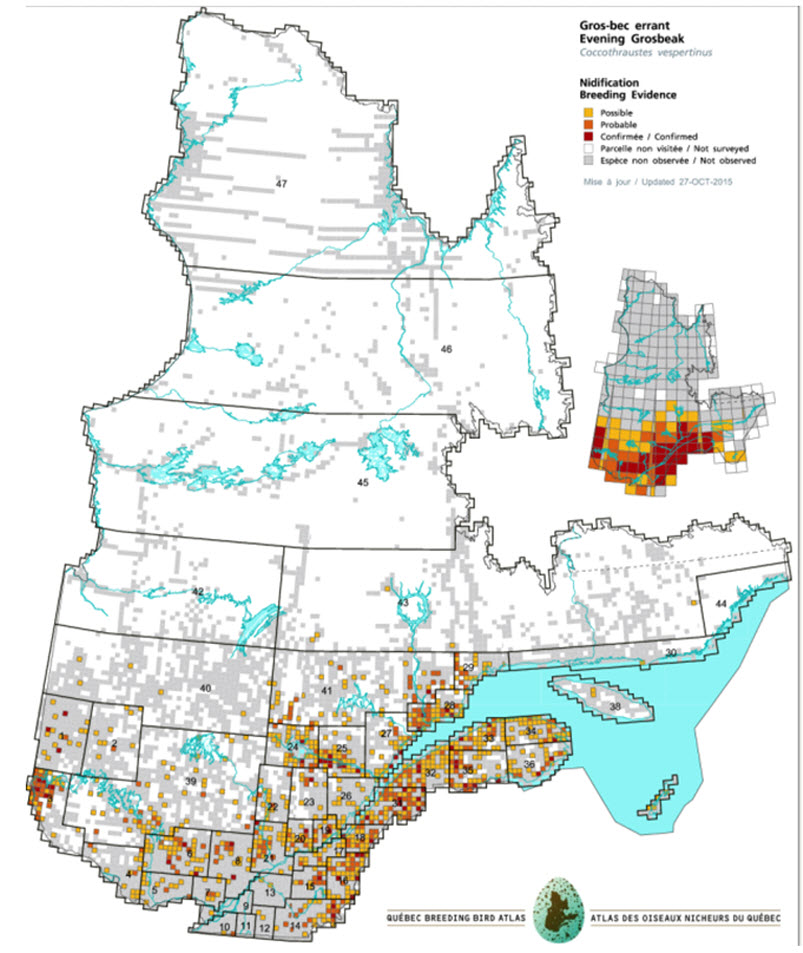
Long description for Figure 12b
Map panel illustrating the distribution (based on grid cells) of Evening Grosbeaks in Quebec during the second atlas period (2010 to 2014) (right panel).
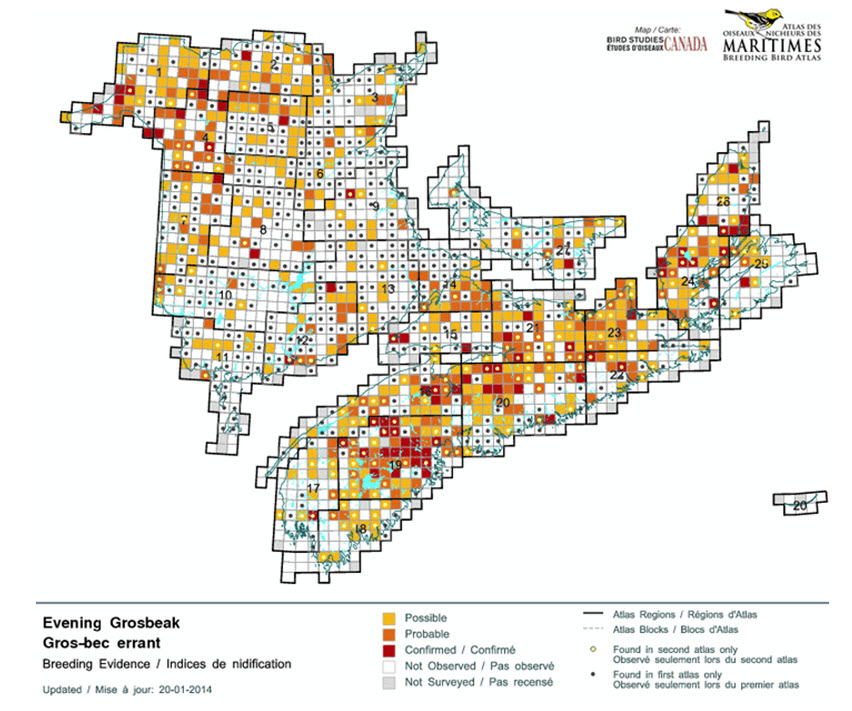
Long description for Figure 13
Map illustrating the distribution (based on grid cells) of Evening Grosbeaks in the Maritimes provinces based on Maritimes Breeding Bird Atlas data. Symbols indicate cells in which Evening Grosbeak was found only in the first atlas period (1986 to 1990), and cells in which the birds were present only in the second atlas period (2006 to 2010).
Checklist program in Quebec (Étude des populations d’oiseaux du Québec, ÉPOQ)
The ÉPOQ database indicates that the abundance index for Evening Grosbeak follows a non-linear relationship for the period 1970 to 2014 (Figure 14; Larivée 2014). If the period is broken at the inflexion point (i.e., 1982), the species abundance index is stable from 1970 to 1982 (R2 = 0.00072, p> 0.05), but is declining sharply by -19% per year (R2 = 0.70, p ≤ 0.001) from 1983 to 2014.
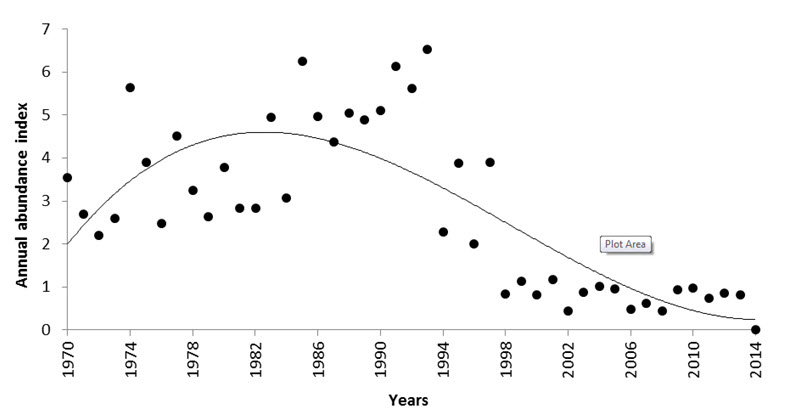
Long description for Figure 14
Chart illustrating change in the annual abundance index of Evening Grosbeaks in Quebec between 1970 and 2014 according to the Checklist program in Quebec.
Forest bird monitoring program
Results from this program show a negative but not significant decline in Ontario from 1987 to 2007 (-1.4% per year, 95% CI: -13.7, 12.8; Government of Canada 2008).
Migration count at the Observatoire d’oiseaux de Tadoussac (OOT) QC
Visual counts of migratory birds from the OOT indicate a peak in the abundance index in 1997 (1422 birds), followed by several smaller peaks of fewer than 500 birds every 2-4 years until 2012, when Evening Grosbeak numbers started to increase noticeably, reaching the second highest peak since the beginning of the survey in 1996 (1120 individuals; Explos-Nature 2016, Figure 15). This increase in number of Evening Grosbeaks is mainly explained by the presence of a major Spruce Budworm outbreak occurring since 2006 in Côte-Nord and Saguenay-Lac-Saint-Jean regions located just north and west from the OOT (Explos-Nature 2016). Aside from the rebound in numbers at Tadoussac since 2012, the trend generally correlates with EPOQ data, which also show numbers fluctuating around a steady low level after a peak in 1997.
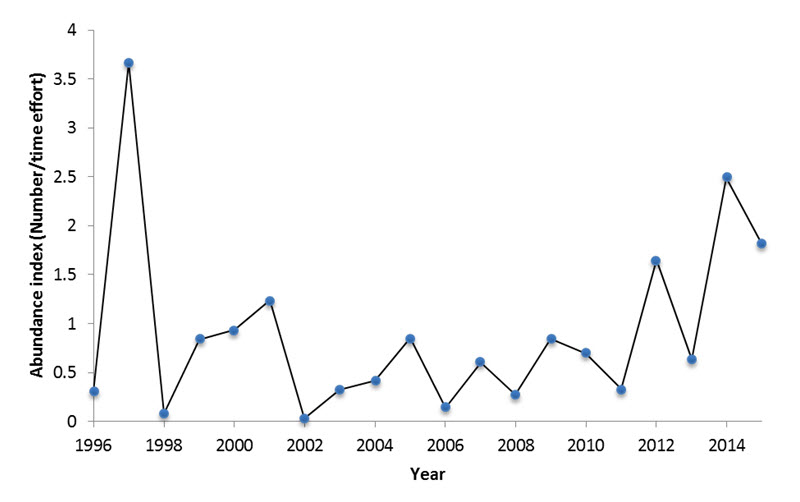
Long description for Figure 15
Chart illustrating the trend in the abundance index of migrating Evening Grosbeaks at the Observatoire d’oiseaux de Tadoussac, Quebec, from 1996 to 2015.
Fort Liard landbird monitoring program
In the southwestern Northwest Territories, the Evening Grosbeak breeding population showed a non-significant declining trend (-2.8% per year ± 8.2, p=0.719) between 1998 and 2011, a pattern that follows a quadratic trend (Machtans et al. 2014).
Population trend summary
A substantial long-term decline of Evening Grosbeaks in Canada is evident from both the CBC (cumulative population loss of 77%) and BBS (cumulative population loss of 86%) databases, and is reflected also in declines recorded by Project FeederWatch, the ÉPOQ database in Quebec, and comparisons between first and second generation breeding bird atlases in Ontario and the Maritimes. Short-term trends are less consistent, with BBS indicating a continuing significant decline of 42% over the most recent decade, but other data sources suggesting a stabilization of the population or even a slight increase in some regions, most notably in relation to the growing Spruce Budworm outbreak in central Quebec.
Rescue effect
In the event of the extirpation of the Canadian Evening Grosbeak population, immigration of individuals from the United States is likely, considering that several bordering northwestern and northeastern states have shown a long-term increase ≥1.5 % per year since 1966 (Figure 16; Sauer et al. 2014).

Long description for Figure 16
Map illustrating Breeding Bird Survey trends (percent change per year, indicated by coloured areas) for the Evening Grosbeak in the United States and Canada between 1966 and 2012.
Threats and limiting factors
Threats
The overall assessment of threats to Evening Grosbeak is of low magnitude, as outlined in Appendix 1: Threats Assessment for Evening Grosbeak. Three individual threats are identified as low, three as negligible, and another two as unknown.
Residential and commercial development (housing and urban areas – low threat)
The primary concern under this category is collision with buildings, given that Evening Grosbeak is among the 10 species in North America most frequently killed by window strikes near feeding stations (Klem 1989; Dunn 1993). Data collected from Project FeederWatch during winter 1989-90 at 5,500 homes show that Evening Grosbeak mortality at windows represented 3.7% of all fatalities detected (n= 945 kills; Dunn 1993). However, this is comparable to the percentage of Evening Grosbeaks observed at feeders where window kills were reported (3.3%) (Dunn 1993), suggesting the species is not disproportionately susceptible to collisions. It is unknown what percentage of Evening Grosbeaks visit feeders. The impact of this threat is expected to be low overall, given that it is ongoing, but likely affects a minority of the population, and of those colliding with windows, some may survive. Bird feeders also have beneficial aspects for the species by providing supplementary food during harsh winter conditions.
Transportation and service corridors (roads and railroads – low threat)
During winter, spring and probably fall, when Evening Grosbeaks are attracted to roads to feed on grit and road salt, collisions with vehicles have been known to be important at least locally (Wilson 1981; Campbell et al. 2001). Examples include 2,000 Evening Grosbeaks found dead along a 16 km stretch of highway in southern British Columbia in the early 1980s (Wilson 1981) and another 500 birds along the same stretch of highway in 1974 (Campbell et al. 2001). These two reports coincide with a Spruce Budworm infestation in this region (Campbell et al. 2001), which suggest that this threat could be more important when the species is at high densities. Given that many road mortalities are unreported, this may be a larger factor than has been documented (Mineau and Brownlee 2005). However, the impact is assessed as low, given that severity and timing values are slight and high respectively.
Biological resource use (logging and wood harvesting – low threat)
The impact of forest harvesting on the Evening Grosbeak population in Canada is difficult to assess in part because of differences in management practices and disturbance history between eastern and western Canada and the lack of studies addressing the response of this species to forestry practices. In the western United States and in Saskatchewan, some studies suggest that Evening Grosbeak is generally absent from second-growth forests in which clearcutting of mature and old-growth stands has occurred (Hutto et al. 1993; Cumming and Diamond 2002; Hayes et al. 2003; Bonter and Harvey 2008). This might be linked to current management practices in many parts of the species’ range favouring short rotation time between cuts (e.g., 40-70 years), which prevents forest structure from diversifying and producing suitable Evening Grosbeak habitat (Hagar et al. 1996; Hayes et al. 2003). In the south-central boreal forest of Central Canada, habitat degradation is also associated with forest management practices that reduce the natural heterogeneity of mature and old mixedwood forests, such as planting one or two fast-growing tree species such as Trembling Aspen and Jack Pine (Hobson and Bayne 2000a). On the other hand, some forestry practices conducted in western North America may favour Evening Grosbeak habitat, such as pre-commercial thinning in homogeneous Douglas-fir stands (Hagar et al. 1996).
Overall impact calculated for this threat is low, as scope for this threat over the next ten years is considered small, although it is ongoing and on a local scale, severity can be serious to moderate.
Natural system modification (unknown threat)
Chemical insecticides, such as DDT (now banned in North America) and fenitrothion were used extensively to control Spruce Budworm from the early 1950s to the 1980s (Blais and Parks 1964; Holmes 1998). In the case of fenitrothion, although its effects on Evening Grosbeak are unknown, a decline in abundance and probable mortality in several passerine species feeding in the crowns of trees following the application of this insecticide have been reported (Buckner 1975; Pearce et al. 1979). In Ontario, Millikin and Smith (1990) and Millikin (1990) have noted that fenitrothion did affect forest arthropods density and forest bird behaviour but had no long-term effect on populations. With organophosphate pesticides, such as trichlorfon (Dylox) and carbaryl (Sevin-4-oil), Zinkl et al. (1977) reported that the enzyme cholinesterase is suppressed in Evening Grosbeaks in treated forests.
Since the early 1980s, Bacillus thuringiensis (BT), a biological pesticide, and tebufenozide have been the main products used to control Spruce Budworm in Canada(van Frankenhuyzen 1993; Holmes 1998). BT has a very low toxicity to vertebrates, including birds, and the toxin is specific to Lepidoptera larvae. However, the latter are an important food source for insectivorous forest birds (Holmes and Shultz 1988) and their control can have adverse effects on chick development and adult behaviour and energetics (Powell 1984). The effects on Evening Grosbeak may therefore be negligible, but should be studied in more detail.
Intensity of Spruce Budworm outbreaks is likely to be reduced in the near future in the commercial forests due to new approaches that are currently being assessed to control Spruce Budworm, such as remote sensing techniques to detect forest stands that are vulnerable to budworm defoliation (i.e., old and mature Balsam Fir stands). These techniques will be used to perform preventive logging well before outbreaks occur in such a way that fir forest density will be reduced and forest composition will be changed in order to produce unsuitable habitat for budworms (MFFPQ 2013). The impacts of these techniques of budworm prevention on Evening Grosbeak habitat are currently unknown but will likely reduce the amount and quality of habitat available for this species. Impact calculated for this potential threat is ranked as unknown as the scope and the severity are considered unknown.
Pollution (industrial and military effluents – unknown threat)
Among the different types of road salt used in Canada, sodium chloride and calcium chloride are the most widely used on paved roads and are known to be toxic to birds when taken in excess, because the kidney of terrestrial birds is not efficient at removing excess sodium (Bollinger et al. 2005; Mineau and Brownlee 2005). Evening Grosbeak has a particularly strong attraction to salt (Fraser 1985), frequently consuming sodium chloride or calcium chloride along roadsides (Wilson 1981; Fraser 1985; Taylor 1997; Campbell et al. 2001). Detailed data on acute toxicity of salt on Evening Grosbeaks and other songbird species is currently lacking, but exposure modelling studies on other birds (i.e., House Sparrow, Passer domesticus) indicate that overconsumption of sodium chloride could cause a series of symptoms, including reduced vigilance and motor function (Mineau and Brownlee 2005), which might be linked to the likelihood of vehicle strikes when flocks of Evening Grosbeaks feed on roadside (Taylor 1997; Mineau and Brownlee 2005). When taken in excess, sodium chloride could also cause direct mortality due to dehydration if water is not available near feeding sites (Mineau and Brownlee 2005). Moreover, when Evening Grosbeaks eat snow and water containing high concentrations of salt on roadside for hydration, sodium chloride toxicity could cause direct mortality (Mineau and Brownlee 2005). Although the amount of salt used in Canada was higher prior to 1990 compared to modern techniques (Environment Canada 2013), the impact of this threat to Evening Grosbeaks needs further investigation as road salt is still widely and intensively used across parts of the species’ winter range.
The impact of this threat is unknown, as the scope is small and the severity is unknown.
Agriculture and aquaculture (annual and perennial non-timber/livestock farming and ranching – negligible threat)
Evening Grosbeak is sensitive to forest fragmentation caused by agricultural activities carried out at the southern limit of the boreal forest where the species is more abundant in contiguous forest than in forest fragments surrounded by agriculture (Hobson and Bayne 2000b). Ontario Forest Bird Monitoring data suggest that the species is sensitive to habitat availability at the landscape (3.2 km) and regional (24 km) scales (Desrochers et al. 2010). Given that Evening Grosbeak numbers are affected by Spruce Budworm outbreaks over large areas of forest, habitat selection may occur primarily at these larger spatial scales (Desrochers et al. 2010).
Overall impact calculated for this threat is negligible, as habitat changes could be extreme in severity, but are expected to affect a negligible part of the population.
Climate change and severe weather (negligible threat)
The climate in the boreal forest and in a large portion of the Evening Grosbeak’s Canadian range will likely be warmer and drier in the 21st century (Intergovernmental Panel on Climate Change 2007; Xu et al. 2013). A number of studies suggest that climate change will reduce the total area of Evening Grosbeak optimal habitat by the end of the century by causing a contraction of the growing zone of the Balsam Fir (Williams and Liebhold 1997; Matthews et al. 2004; Siegel et al. 2014). These studies suggest that Balsam Fir-dominated forest and Spruce Budworms will be greatly reduced in the northeastern United States and possibly in the southern portion of the breeding range of Evening Grosbeak in eastern Canada due to a northward shift of Balsam Firs and budworms as a result of increasing temperatures. In the northern portion of the species’ range in eastern Canada, a contraction is also predicted to occur as fire will be more frequent and more intense due to a warmer and drier climate which in turn will negatively affect the capacity of fir to survive (Messaoud et al. 2007; Bergeron et al. 2010; Terrier et al. 2013).
Models using climate, land use and topography variables on the projected density of Evening Grosbeak in the boreal and hemiboreal forest in North America, on the other hand, predict that the species will increase by 93% in density by 2100 in the northernmost areas of Canada (Stralberg et al. 2015a). Other models using climate data predict that climatically suitable core habitat range for Evening Grosbeak will increase by 45% for the 2071-2100 period (Stralberg et al. 2015b). During this period, grosbeak densities are predicted to increase particularly in northwestern British Columbia and Alaska. These latter results have to be viewed with caution as they do not take into account changes in the distribution of forests associated with climate change.
Climate change may also directly impact food sources of Evening Grosbeaks such as Spruce Budworm. Several studies generally agree that climate change may negatively affect the distribution and abundance of Spruce Budworm in Canada, notably in eastern provinces. Climate change is predicted to increase the annual temperature average in eastern boreal forest (Johnson et al. 1999), fire ignition, area burned and length of the wildfire season (16% longer; Bergeron and Flannigan 1995; Flannigan et al. 2005; Krawchuk and Cumming 2011) and the average rate of precipitation (by 5%; Plummer et al. 2006). A more humid climate in eastern Canada could decrease the abundance of Spruce Budworm, which usually requires extended periods of drought to reach epidemic levels (Morin et al. 2008). Moreover, climate change could negatively impact Spruce Budworm as they depend on timely availability of new foliage (Nealis and Régnière 2004; Volney and Fleming 2007). In addition to temperature, the timing of insect life cycles and bud flushing is also affected by photoperiod. Thus, it is conceivable that, with rapid change in climate, the synchrony between bud burst and the emergence of feeding stages could become decoupled because of subtle timing differences in responses to cues determining the seasonality of the insect species concerned, causing a negative effect on insect populations (Thomson and Benton 2007).
Impact calculated for this threat is ranked negligible, as scope and severity of climate change and severe weather are expected to be negligible within the next ten years.
Limiting factors
The 25-40 year natural cycle of Spruce Budworm (especially in eastern Canada) is likely to have a greater influence on Evening Grosbeak numbers than any threats, and largely correlates with declines in Evening Grosbeak populations since the 1970s. The total area of forest defoliated by Spruce Budworm in Canada (mainly in Ontario and Quebec) reached a peak of 52 million ha defoliated in 1975, followed by a sharp decline until 1996 when defoliated areas dropped to fewer than 2 million ha annually until 2005 (Figure 17). In Quebec, where the largest total area of defoliated forest was found (> 40 million ha), a first peak was reached in 1975 with over 28 million ha defoliated and a second peak in 1983 with 12.5 million ha defoliated. In this province, defoliated area decreased sharply from 1983-1986 and stayed at a low level until 2009. In contrast, peak defoliation in Ontario was reached five years later (1980) with a total of 20 million ha, but two other less pronounced defoliation peaks occurred in 1987 and 1992, then decreased sharply from 1993 to 1996 and remained low until 2009.
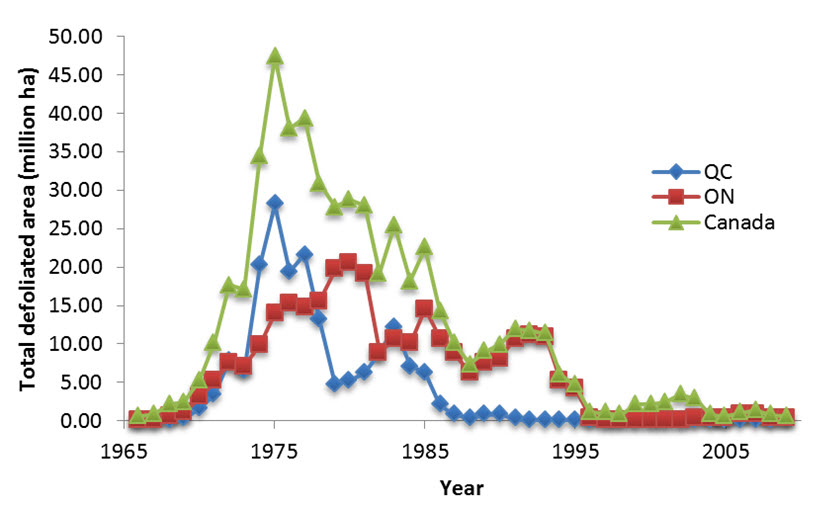
Long description for Figure 17
Chart illustrating trends in area of forest defoliated by Spruce Budworm in Ontario, in Quebec, and Canada as a whole between 1966 and 2009.
Several studies have associated the fluctuation in regional, provincial and even North American Evening Grosbeak populations with fluctuations in Spruce Budworm density or defoliated areas (Bolgiano 2004; Hoffman 2009; Venier et al. 2009). For example, both the abundance index of Evening Grosbeak from CBC data for North America and the total defoliated area in Canada start to increase sharply in mid-1960s and both reached a peak in 1972 and 1975 respectively, then values of both decreased gradually until approximately 1994 (Figure 6 and Figure 17). After 1994, numbers remained low for both until at least 2005.
The percentage of feeders visited from Project FeederWatch and the average number of birds seen at feeders for the northeastern provinces also reflected the peak in defoliated areas around 1989-1990, followed by a decline (Figure 8). This parameter was high in 1989 and 1990, corresponding to the peak in defoliated areas, then decreased in 1991 and remained fairly stable until 2009. Furthermore, a recent analysis indicates that the BBS annual abundance index for Evening Grosbeak in Canada was positively correlated with the total area of forest defoliated by the Spruce Budworm over the 1974-2012 period (generalized least squares regression with errors following a first-order temporal autoregressive process, β = 0.00057, SE = 0.00013, P≤ 0.0001; Figure 18; M. Bélisle unpubl. data). Hoffman (2009) suggests that the trend in Evening Grosbeak population is more likely due to a decrease in the reproduction rate than in the survival rate. Because there are no reliable abundance data from periods prior to Spruce Budworm outbreaks (i.e., before 1970), it is not possible to determine whether the current population is depressed or at levels which are normal for non-outbreak periods (Hoffman 2009). However, because no declining trend in survival was detected using banding data from North America, Hoffman (2009) infers that population levels observed since 1995 are representative of a normal predator-prey cycle.
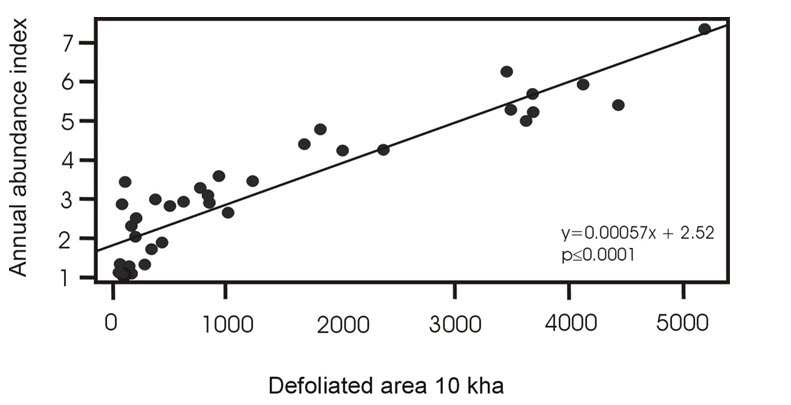
Long description for Figure 18
Chart illustrating the association between the total area of forest defoliated by Spruce Budworm and the Breeding Bird Survey annual abundance index for Evening Grosbeaks between 1974 and 2012 in Canada.
Several lines of evidence suggest that a new major Spruce Budworm outbreak in eastern Canada will occur early in the 21st century (Morin et al. 2008). Since 2006, the total area defoliated by Spruce Budworm in Quebec has increased from less than 100 000 ha in 2005 to 6.3 million ha in 2015 (MFFPQ 2015). The outbreak is now occurring in five regions of this province but is more pronounced in the Côte-Nord (3.7 million ha) and Saguenay-Lac-St-Jean (1 million ha) (MFFPQ 2015) and is now expanding into eastern Ontario and New Brunswick. Evening Grosbeaks appear to have begun to increase in response to this new outbreak. Visual counts of migratory birds in the Côte-Nord region of northern Quebec indicate that the number of Evening Grosbeaks migrating in fall has increased since 2012 (Explos-Nature 2014), corresponding to an increase in defoliation from 1.7 million ha in 2012 to 3.8 million ha in 2015 (MFFPQ 2014, 2015). If this new outbreak reaches the same magnitude as the last outbreak, Evening Grosbeak in eastern Canada may rebound to levels observed in the 1970s and 1980s.
Diseases and parasites
Various types of diseases have been reported as causing mortality in Evening Grosbeak in Canada. They include conjunctivitis caused by Mycoplasma gallisepticum (Mikaelian et al. 2001), salmonellosis (Salmonella enterica Typhimurium) (Locke et al. 1973; Daoust et al. 2000) and infections by a parasitic mite, Knemidokoptes jamaicensis, which causes lesions to the feet and bill (Carothers 1974). At the local scale, the West Nile virus has also caused mortality in Evening Grosbeak in the eastern United States (Komar 2003). The impacts of these diseases on Evening Grosbeak remain unknown, but could be significant given that a number of them are contracted in winter when the species adopts a gregarious behaviour and feeds with other potentially contaminated granivorous species at feeding stations (Mikaelian et al. 2001).
Number of locations
Number of locations for Evening Grosbeak is currently unknown but considering its wide distribution in Canada, it is probably high, and almost certainly >10.
Protection, status, and ranks
Legal protection and status
Evening Grosbeak is protected in Canada under the Migratory Birds Convention Act, 1994 (MBCA). The MBCA prohibits the possession or sale of migratory birds and their nests, as well as the disturbance or destruction of migratory birds, and their eggs and nests, except in cases authorized under the Migratory Birds Regulations. This species is also protected in the United States and Mexico under similar legislation.
In all provinces and territories, Evening Grosbeak is protected under broad provincial regulations such as the Loi sur la conservation et la mise en valeur de la faune (RLRQ, c. C- 61.1) (LCMVF) (Act respecting the conservation and development of wildlife) (CQLR, c. C-61.1) in Quebec or the Fish and Wildlife Conservation Act 1997 in Ontario (Government of Ontario 2012). These regulations generally prohibit the hunt, capture, holding in captivity or selling of wildlife and to destroy or damage nests and eggs.
Non-legal status and ranks
NatureServe (2015) has assigned Evening Grosbeak a global rank of G5 (secure) (Table 5). According to the IUCN Red List, it is considered a species of least concern (IUCN, 2015). In the United States, it has a rank of N5 (secure) (NatureServe, 2015), but it is considered critically imperilled in two states (Virginia and the District of Columbia), imperilled in five states (Wisconsin, Texas, Oklahoma, Maryland and Massachusetts) and vulnerable in four other states (NatureServe, 2015).
Evening Grosbeak is considered secure (N5) in Canada (NatureServe, 2015; Table 5). It is considered imperilled (S2) in Prince Edward Island and vulnerable (S3) in Yukon, Manitoba and New Brunswick (NatureServe, 2015; Table 5). In other provinces and the Northwest Territories, it is considered secure (S5) or apparently secure (S4; Table 5).
| Region | NatureServe Rank* | GSSC Rank |
|---|---|---|
| Global | G5 | - |
| United States | N5 | - |
| Canada | N5 | Secure |
| Yukon | S3B | Sensitive |
| Northwest Territories | S4 | Secure |
| British Columbia | S5 | Secure |
| Alberta | S4 | Secure |
| Saskatchewan | S4B, S4N | Secure |
| Manitoba | S3 | Sensitive |
| Ontario | S4B | Sensitive |
| Quebec | S4 | Secure |
| New Brunswick | S3S4B, S4S5N | Sensitive |
| Nova Scotia | S4B, S5N | Secure |
| Prince Edward Island | S2B, S4N | Sensitive |
| Newfoundland and Labrador | S4B, S5N, SNA | Secure |
* G = is a global status rank; S = rank assigned to a province or state; N= is a national status rank. S1 indicates that a species is critically imperilled because of extreme rarity (often 5 or fewer occurrences) or because of other factor(s) such as a steep decline, making it especially vulnerable to extirpation; S2 indicates that a species is imperilled because of rarity or other factors making it very vulnerable to extirpation, usually with 6 to 20 occurrences or few individuals remaining (i.e., 1,000 to 3,000; S3 indicates that a species is vulnerable at the subnational level because it is rare or uncommon, or found only in a restricted range, or because of other factors making it vulnerable to extirpation; S4 indicates that the species is apparently secure because it is uncommon but not rare, and that there is some cause for long-term concern due to declines or other factors; S5 indicates that a species is secure, because it is common, widespread and abundant.
Habitat protection and ownership
In Canada, suitable breeding habitat for Evening Grosbeak is found primarily on public land in boreal forests (Boreal Avian Modeling Project 2015). It is estimated that 9% of the range of Evening Grosbeak is currently protected by national and provincial parks, migratory bird sanctuaries, national wildlife areas and protected natural areas (Wells 2011). According to Parks Canada’s Biotics Database, Evening Grosbeak is present (during the breeding season, migration or year round) in 14 protected areas managed by the Parks Canada Agency (Parks Canada 2013).
Acknowlegements and authorities contacted
The report writer would like to thank Marco Beaulieu, Marc Bélisle, Yves Bergeron, Pascal Coté, Bruno Drolet, Jonathan Frechette, Louis Imbeau, Adam Smith, Diana Stralberg, Lisa Venier, Peter Taylor and Isabelle Gauthier, Steven L. Van Wilgenburg, Samuel Hache and Isabelle Duclos who were kind enough to share their knowledge on the ecology of Evening Grosbeak and Spruce Budworm. The report writer would also like to thank Marc Bélisle for the unpublished data on the relationship between the BBS abundance index and the total area of forests defoliated by the Spruce Budworm in Canada. Thanks also to the official sponsors of the Ontario Breeding Bird Atlas (Bird Studies Canada, the Canadian Wildlife Service, Ontario Nature, Ontario Field Ornithologists and the Ontario Ministry of Natural Resources) for providing atlas data, and to the thousands of volunteer participants who collected data. Thanks also to the Regroupement QuébecOiseaux, Canadian Wildlife Service and Bird Studies Canada for providing breeding bird atlas data in Quebec and to thousands of participants who collect data for the project. The report writer would like to acknowledge all of the volunteers who participated in the collection of the CBC and BBS data. Funding for this report was provided by the Canadian Wildlife Service of Environment and Climate Change Canada.
Information sources
Anthony, R.G., G.A. Green, E.D. Forsman, and S.K. Nelson. 1996. Avian abundance in riparian zones of three forest types in the Cascade Mountains, Oregon. Wilson Bulletin 108:288-291.
American Ornithologist’s Union. 1998. Checklist of North American birds, 52th suppl. American Ornithologist’s Union, Washington, D.C.
Atlas of the breeding birds of Quebec (ABBQ). 2014. Data obtained in response to a request made to the Atlas office. Regroupement QuébecOiseaux, Environment Canada’s Canadian Wildlife Service and Bird Studies Canada. Québec, Quebec, Canada
Aubry, Y. and P. Laporte. 1990. Les gros-becs aux mangeoires « jamais » les mêmes! Québec Oiseaux1:8-11.
Baicich, P.J. and C.J.O. Harrison. 1997. A guide to the nests, eggs, and nestlings of North American birds. Second Edition. Academic Press, San Diego, California.
Balph, D.F. and M.H. Balph. 1979. Behavioral flexibility of Pine Siskins in mixed species foraging groups. The Condor 81:211-212.
Balshi, M.S., A.D. McGuire, P. Duffy, M.D. Flannigan, J. Walsh, and J. Melillo. 2009. Assessing the response of area burned to changing climate in western boreal North America using a multivariate adaptive regression splines (MARS) approach. Global Change Biology 15:578-600.
Bekoff, M. 1995. Vigilance, flock size, and flock geometry: information gathering by western Evening Grosbeaks (Aves, Fringillidae). Ethology 99:150-161.
Bekoff, M. and A.C. Scott. 1989. Aggression, dominance, and social organization in Evening Grosbeaks. Ethology 83:177-194.
Bekoff, M., A.C. Scott, and D.A. Conner. 1987. Nonrandom nest-site selection in Evening Grosbeaks. The Condor 89:819-829.
Bekoff, M., A.C. Scott, and D.A. Conner. 1989. Ecological analyses of nesting success in Evening Grosbeaks. Oecologia 81:67-74.
Bélisle, M. Unpubl. data. 2014. Email correspondence to C. Savignac. November 2014. Associate Professor, Canada Research Chair in Spatial and Landscape Ecology, Department of Biology, Université de Sherbrooke, Québec.
Bergeron, Y, and M.D. Flannigan. 1995. Predicting the effects of climate change on fire frequency in the south-eastern Canadian boreal forest. Water, Air, and Soil Pollution82:437-444.
Bergeron, Y. and A. Leduc. 1998. Relationships between change in fire frequency and mortality due to Spruce Budworm outbreak in the southeastern Canadian boreal forest. Journal of Vegetation Science 9:492-500
Bergeron, Y., D. Cyr, M.P. Girardin, and C. Carcaillet. 2010. Will climate change drive 21st century burn rates in Canadian boreal forest outside of its natural variability: collating global climate model experiments with sedimentary charcoal data. International Journal of Wildland Fire 19: 1127-1139.
Bird Studies Canada (BSC). 2013. Project NestWatch. Bird Studies Canada / Études d'Oiseaux Canada [accessed February 2013]
Bird Studies Canada. 2014. Manitoba Breeding Bird Atlas. [accessed October 2015].
Bird Studies Canada. 2015. Maritimes Breeding Bird Atlas. Maritimes Breeding Bird Atlas. [accessed October 2015].
Blais, J.R. and G.H. Parks. 1964. Interaction of Evening Grosbeak (Hesperiphona vespertina) and Spruce Budworm (Choristoneura fumiferana (Clem.)) in a localized budworm outbreak treated with DDT in Québec. Canadian Journal of Zoology42:1017-1024.
Blancher, P. 2013. Estimated number of birds killed by house cats (Felis catus) in Canada. Avian Conservation and Ecology 8: 3. http://dx.doi.org/10.5751/ACE-00557-080203
Blancher, P.J., K.V. Rosenberg, A.O. Panjabi, B. Altman, A.R. Couturier, W.E. Thogmartin, and the Partners in Flight Science Committee. 2013. Handbook to the Partners in Flight Population Estimates Database, Version 2.0. PIF Technical Series No. 6.
Bock, C.E. and L.W. Lepthien. 1976. Synchronous eruptions of boreal seed-eating birds. American Naturalist 110:559-571.
Bolgiano, N.C. 2004. Cause and effect: changes in boreal bird irruptions in eastern North America relative to the 1970s Spruce Budworm infestation. American Birds58:26-33.
Bollinger, T.K., P. Mineau, and M.L. Wickstrom. 2005. Toxicity of sodium chloride to House Sparrows (Passer domesticus). Journal of Wildlife Disease 41:363-370.
Bonter, D. and M.G. Harvey. 2008. Winter survey data reveal range-wide decline in Evening Grosbeak populations. The Condor 110:376-381.
Boreal Avian Modeling Project. 2015. Evening Grosbeak Coccothraustes vespertinus. [accessed October 2015]
Brewer, D., A. Diamond, E.J. Woodsworth, B.T. Collins and E.H. Dunn. 2000. Canadian Atlas of Bird Banding, Volume 1: Doves, Cuckoos, and Hummingbirds through Passerines, 1921-1995, Occasional Paper, Canadian Wildlife Service, Environment Canada.
Brunton, D.F. 1994. The Evening Grosbeak in Ontario. Pp. 307-314 in M.K. McNicholl and J.L. Cranmer-Byng (eds.). Ornithology in Ontario. Special Publication No. 1. Ontario Field Ornithologists.
Buckner, C.H. 1975. Cited in Fenitrothion: the effects of its use on environmental quality and its chemistry. Associate Committee on Scientific Criteria for Environmental Quality, National Research Council of Canada. NRCC No. 14104. 56 pp.
Burleigh, J.S., R.I., Alfaro, J.H. Borden, and S. Taylor. 2002. Historical and spatial characteristics of Spruce Budworm Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae) outbreaks in northeastern British Columbia. Forest Ecology and Management 168: 301–309.
Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage, and A.R. Couturier (eds.). 2007. Atlas of the breeding birds of Ontario, 2001-2005. Bird Studies Canada, Environment Canada, Ontario Field Ornithologists, Federation of Ontario Naturalists, Ontario Ministry of Natural Resources, and Ontario Nature, Toronto, Ontario. xxii + 706 pp.
Campbell, R.W., N.K. Dawe, I. McTaggart-Cowan, J.M. Cooper, G.W. Kaiser, A.C. Stewart, and M.C.E. McNall. 2001. The Birds of British Columbia. Vol. IV. Passerines: Wood-Warblers through Old World Sparrows. UBC Press. Vancouver, British Columbia.
Canadian Endangered Species Conservation Council (CESCC). 2011. Wild Species 2010: The General Status of Species in Canada. National General Status Working Group.
Cannings, R.A., R.J. Cannings, and S.G. Cannings. 1987. Birds of the Okanagan Valley, British Columbia. Royal British Columbia Museum. Victoria, 420 pp.
Candau, J.-N., R.A. Fleming, and A. Hopkin. 1998. Spatiotemporal patterns of large-scale defoliation caused by the Spruce Budworm in Ontario since 1941. Canadian Journal of Forestry Research 28:1733-1741.
Candau, J.N. and R.A. Fleming. 2011. Forecasting the response of Spruce Budworm defoliation to climate change in Ontario. Canadian Journal of Forest Research 41:1948-1960.
Carothers, S.W., N.J. Sharber, and G.F. Foster. 1974. Scaly-leg (knemidokoptiasis) in a population of Evening Grosbeaks. Wilson Bulletin 86:121-124.
Clements, J.F. 2007. The Clements Checklist of Birds of the World: Sixth edition. Comstock Publishing Associates. Ithaca, New York. 843 pp.
Cornell University. 2014. Birdsource: birding with a purpose. Population trend of the Evening Grosbeak. Christmas Bird Count data show changes in Evening Grosbeak numbers between 1960 and 1998. [accessed October 2014].
Cornell University. 2015. Project FeederWatch [accessed October 2015].
Crawford, H.S., R.W. Titterington, and D.T. Jennings. 1983. Bird predation and Spruce Budworm populations. Journal of Forestry81:433-435.
Cumming, E.E., and A.W. Diamond. 2002. Songbird community composition versus forest rotation age in Saskatchewan boreal mixedwood forest. Canadian Field Naturalist 116:69-75.
Cyr, A. and J. Larivée. 1995. Atlas saisonnier des oiseaux du Québec. Les Presses de l’Université de Sherbrooke et la Société de Loisir ornithologique de l’Estrie, Sherbrooke, Québec.
Daoust, P.-Y. D.G. Busby, L. Ferns, J. Goltz, S. McBurney, C. Poppe, and H. Whitney. 2000. Salmonellosis in songbirds in the Canadian Atlantic provinces during winter-summer 1997-98. Canadian Veterinary Journal 41:54-59.
Desrochers, A., C. Renaud, W.M. Hochachka, and M. Cadman. 2010. Area-sensitivity by forest songbirds: theoretical and practical implications of scale-dependency. Ecography 33:921-931.
Downs, E.H. 1956. Evening Grosbeaks at South Londonderry, Vermont, 1954 and 1955. Bird-Banding 27:166-170.
Drolet, B., pers. comm. 2016. Email correspondence to C. Savignac. May 2016. Head of Population Assessment Unit, Canadian Wildlife Service, Quebec Region.
Dunn, E.H. 1993. Bird mortality from striking residential windows. Journal of Field Ornithology 64:302-309.
Dunn, E.H., J. Larivée, and A. Cyr. 1996. Can checklist programs be used to monitor populations of birds recorded during the migration season? Wilson Bulletin 108:540-549.
eBird. 2015. eBird: An online database of bird distribution and abundance. Cornell Lab of Ornithology, Ithaca, New York. [accessed October 2015].
Environment Canada. 2013. Five-year Review of Progress: Code of Practice for the Environmental Management of Road Salts. [accessed October 2015]
Environment Canada. 2014. North American Breeding Bird Survey – Canadian Trends Website, Data-version 2012. Environment Canada, Gatineau, Québec.
Erskine, A.J. 1992. Atlas of Breeding Birds of the Maritime Provinces. Nimbus Publishing, Halifax, Nova Scotia.
Explos-Nature. 2016. Observatoire d’oiseaux de Tadoussac. Blog des migrations. [accessed August 2016].
Federation of Alberta Naturalists. 2007. The Atlas of Breeding Birds of Alberta: A Second Look. Federation of Alberta Naturalists, Edmonton, Alberta. vii + 626 pp.
Fee, B.A. and M. Bekoff. 1986. Polygyny in the Evening Grosbeak. Wilson Bulletin 98:308.
Flannigan, M.D., K.A. Logan, B.D. Amiro, W.R. Skinner, and B.J. Stocks. 2005. Future area burned in Canada. Climatic Change 72:1-16.
Fleming, R.A. and J.N. Candau. 1998. Influences of climatic change on some ecological processes of an insect outbreak system in Canada's boreal forests and the implications for biodiversity. Environmental Monitoring and Assessment 49:235-249.
Francis, C.M., P.J. Blancher, and R.D. Phoenix. 2009. Bird monitoring programs in Ontario: What have we got and what do we need? The Forestry Chronicle 85:202-217.
Fraser, D. 1985. Mammals, birds and butterflies at sodium sources in Northern Ontario forests. Canadian Field-Naturalist 9:365-367.
Freeman, T.N. 1967. On coniferophagous species of Choristoneura (Lepidoptera: Tortricidae) in North America. I. Some new forms of Choristoneura allied to C. fumiferana. Canadian Entomologist 99:449-455.
Gabrielson, I.N. 1924. In Gillihan, S.W. and B. Byers. 2001. Evening Grosbeak (Coccothraustes vespertinus), The Birds of North America Online (A. Poole, ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online.
Gauthier, J. and Y. Aubry (eds.). 1995. Les oiseaux nicheurs du Québec: Atlas des oiseaux nicheurs du Québec méridional. Association québécoise des groupes d’ornithologues, Société québécoise de protection des oiseaux, Canadian Wildlife Service, Environment Canada, Montreal, Québec, xviii + 1295 pp.
Gillihan, S.W. and B. Byers. 2001. Evening Grosbeak (Coccothraustes vespertinus). The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online.
Godfrey, W.E. 1986. The Birds of Canada, édition révisée, Musées nationaux du Canada, Ottawa (Ontario), 595 p.
Girardin, M.P. M.D. Flannigan, J.C. Tardif and Y. Bergeron. 2008. Climatologie, meteorology et feux de forêt. Sous la direction de S. Gauthier et coll. Dans Aménagement écosystémique en forêt boréale. Presses de l’Université du Québec, Québec, 568 pp.
Gosse, J., L. Hermanutz, B. McLaren, P. Deering, and T. Knight. 2011. Degradation of boreal forests by nonnative herbivores in Newfoundland's national parks: Recommendations for ecosystem restoration. Natural Areas Journal 31:331-339.
Government of Canada. 2008. Ontario Forest Bird Monitoring Program Newsletter. Volume 18, Issue 1, Spring 2008.
Government of Canada. 2011. Natural Resource Canada. Large aspen tortrix (accessed January 2015)
Government of Canada. 2013. Map 1: Terrestrial ecozones and marine ecoregions of Canada [accessed October 2015].
Government of Canada. 2014. National Forestry Database [accessed October 2014].
Government of Ontario. 2012. Fish and Wildlife Conservation Act, 1997
S.O. 1997, CHAPTER 41. [accessed January 2015].
Gray, D.R. 2008. The relationship between climate and outbreak characteristics of the Spruce Budworm in eastern Canada. Climate Change 89:447-449.
Gray, D.R., J. Régnière, and B. Boulet. 1999. Analysis and use of historical patterns of Spruce Budworm defoliation to forecast outbreak patterns in Quebec. Forest Ecology and Management 127:217-231
Hagar, J.C., W.C. McComb, and W.H. Emmingham. 1996. Bird communities in commercially thinned and unthinned Douglas-fir stands in western Oregon. Wildlife Society Bulletin 24:353-366.
Haiman, A.N.K. 2011. Levels of Variation in Evening Grosbeak (Coccothraustes vespertinus) Calls and Morphology. M.Sc. thesis, University of California Davis, California. 53 pp.
Hayes, J.P., J.M. Weikel, and M.M.P. Huso. 2003. Response of birds to thinning young Douglas-fir forests. Ecological Applications 13:1222-1232.
Hoar, T. 2007. Evening Grosbeak. Pp. 626-627 inM.D. Cadman, D.A. Sutherland, G.G. Beck, D. Lepage, and A.R. Couturier (eds.). Atlas of the Breeding Birds of Ontario, 2001–2005. Bird Studies Canada, Environment Canada, Ontario Field Ornithologists, Ontario Ministry of Natural Resources, and Ontario Nature, Toronto, Ontario.
Hobson, K.A. and E. Bayne. 2000a. Breeding Bird Communities in Boreal Forest of Western Canada: Consequences of "Unmixing" the Mixedwoods. The Condor 102:759-769.
Hobson, K.A. and E. Bayne. 2000b. Effects of forest fragmentation by agriculture on avian communities in the southern boreal mixedwoods of western Canada. Wilson Bulletin112:373-387.
Hobson, K.A., D.A. Kirk and A.R. Smith. 2000. A multivariate analysis of breeding bird species of western and central Canadian boreal forests: Stand and spatial effects. Écoscience 7:267-280.
Hoffman, W. 2009. Analysing and interpreting the decline of the Evening Grosbeak (Coccothraustes vespertinus) using annual survival rates estimated from band-recovery data. M.Sc. Thesis, School of Life Sciences, Wildlife Biology and Conservation, Edinburgh Napier University, Scotland. 35 pp.
Holmes, S.B. 1998. Reproduction and nest behaviour of Tennessee warblers, Vermivora peregrina, in forests treated with Lepidoptera-specific insecticides. Journal of Applied Ecology 35:185-194.
Holmes, S.B., C.J. Sanders, D. Fillman, and D.A. Welsh. 2009. Changes in a forest bird community during an outbreak cycle of the Spruce Budworm in northwestern Ontario. Bird Populations 9:13-28.
Holmes, R.T. and J.C. Schultz. 1988. Food availability for forest songbird: Effect of prey distribution and abundance on bird foraging. Canadian Journal of Zoology 66:720-728.
Hutto, R.L., S.J. Hejl, C.R. Preston, and D.M. Finch. 1993. The effects of silvicultural treatment on forest birds in the Rocky Mountains: implications and management recommendations. Pp. 386-391 in D.M. Finch and P.W. Stangel (eds.). Status and Management of Neotropical Migratory Birds. General Technical Report RM-GTR-229. U.S. Forest Service. Fort Collins, Colorado.
IPCC (Intergovernmental Panel on Climate Change). 2007. IPCC Fourth Assessment Report: Climate Change 2007 (AR4). IPCC, Geneva, Switzerland.
IUCN. 2015. The IUCN Red List of Threatened Species. Version 2015-3. [accessed October 2015]
James, R.D., P.L. McLaren, and J.C. Barlow. 1976. Annotated Checklist of the Birds of Ontario. Royal Ontario Museum. Life Sci. Misc. Publ., Toronto.
Ives, W.G.H. 1984. Malacosoma disstria Hübner, forest tent caterpillar (Lepidoptera: Lasiocampidae). Pages 311-319 in J.S. Kelleher and M.A. Hulme (eds.). Biological control programmes against insects and weeds in Canada, 1969-1980. Commonwealth Agricultural Bureau, Slough, England.
Johnson, E.A., K. Miyanishi, and N. O’Brien. 1999. Long-term reconstruction of the fire season in the mixedwood boreal forest of Western Canada. Canadian Journal of Botany 77:1185-1188.
Kennedy, J.A., P. Dilworth-Christie, and A.J. Erskine. 1999. The Canadian Breeding Bird (Mapping) Census Database. Technical Report Series No. 342. Canadian Wildlife Service, Ottawa, Ontario.
Kendeigh, S.C. 1947. Bird population studies in the coniferous forest biome during a Spruce Budworm outbreak. Ontario Department of Lands and Forests, Research Division. Biological Bulletin 1.
Kirk, D.A., A.W. Diamond, K.A. Hobson, and A.R. Smith. 1996. Breeding bird communities of western Canadian boreal forest: relationship to forest type. Canadian Journal of Zoology 74:1749-1770.
Koenig, W.D., and J.M. Knops. 2001. Seed-crop size and eruptions of North American boreal seed-eating birds. Journal of Animal Ecology 70:609-620.
Komar, N., S. Langevin, S. Hinten, N. Nemeth, E. Edwards, D.L. Hettler, B.S. Davis, R.A. Bowen, and M.L. Bunning. 2003. Experimental Infection of North American Birds with the New York 1999 Strain of West Nile Virus. Emerging Infectious Diseases 9:311-322
Klem, Jr., D. 1989. Bird-window collisions. Wilson Bulletin 101:606-620.
Klimkiewicz, M.K. and A.G. Futcher. 1987. Longevity records of North American birds: Coerebinae through Estrildidae. Journal of Field Ornithology 58:318-333.
Krawchuk, M.A. and S.G. Cumming. 2011. Effects of biotic feedback and harvest management on boreal forest fire activity under climate change. Ecological Applications 21:122-136.
Langelier, A. 1983. Habitat selection of common breeding bird species in western Spruce Budworm outbreak areas. M.Sc. thesis, University of Idaho, Moscow, Idaho.
Larivée, J. 2014. Étude des populations d'oiseaux du Québec (Version 2014-10-27) [databases]. Rimouski, Québec. Regroupement QuébecOiseaux.
Lepage, D. and C.M. Francis. 2002. Do feeder counts reliably indicate bird population changes? 21 years of winter bird counts in Ontario, Canada. The Condor 104:255-270.
Locke, L.N., R.B. Shillinger, and T. Jareed. 1973. Salmonellosis in passerine birds in Maryland and West Virginia (PDF Version; 198 KB). Journal of Wildlife Disease 9:144-145.
Lumley, L.M. and F.A.H. Sperling. 2010. Integrating morphology and mitochondrial DNA for species delimitation within the Spruce Budworm (Choristoneura fumiferana) cryptic species complex (Lepidoptera: Tortricidae).Systematic Entomology35:416-428
Machtans, C.S., K.J. Kardynal, and P.A. Smith. 2014. How well do regional or national Breeding Bird Survey data predict songbird population trends at an intact boreal site? Avian Conservation and Ecology 9:5. http://dx.doi.org/10.5751/ACE-00649-090105.
Martell, A. 2015. Evening Grosbeak in Davidson, P.J.A., R.J. Cannings, A.R. Couturier, D. Lepage, and C.M. Di Corrado (eds.). The Atlas of the Breeding Birds of British Columbia, 2008-2012. Bird Studies Canada. Delta, B.C. [accessed November 2015]
Martin, A.C., H.S. Zim, A.L. Nelson. 1951. American Wildlife and Plants: A Guide to Wildlife Food Habits. Dover, New York.
Matthews, S.N., R.J. O’Connor, L.R. Iverson, and A.M. Prasad. 2004. Atlas of climate change effects in 150 bird species of the eastern United States. USDA Forest Service General Technical Report NE-318.
McCorquodale, D. 2015. Evening Grosbeak, pp. 522-523 in Stewart, R.L.M., K.A. Bredin, A.R. Couturier, A.G. Horn, D. Lepage, S. Makepeace, P.D. Taylor, M.-A. Villard, and R.M. Whittam (eds). Second Atlas of Breeding Birds of the Maritime Provinces. Bird Studies Canada, Environment Canada, Natural History Society of Prince Edward Island, Nature New Brunswick, New Brunswick Department of Natural Resources, Nova Scotia Bird Society, Nova Scotia Department of Natural Resources, and Prince Edward Island Department of Agriculture and Forestry, Sackville, New Brunswick.
Merkins, M. and B.P. Booth. 1998. Monitoring changes in wildlife diversity during operational hardwood harvesting: aspen clearcutting in the Dawson Creek Forest District (1992-1997). Unpublished report, British Columbia Ministry of Environment, Lands and Parks, Wildlife Branch, Victoria, British Columbia.
Messaoud, Y., Y. Bergeron, and A. Leduc. 2007. Ecological factors explaining the location of the boundary between the mixedwood and coniferous bioclimatic zones in the boreal biome of eastern North America.Global Ecology and Biogeography 16:90-102
Mikaelian, I., D.H. Ley, R. Claveau, M. Lemieux, and J-P. Bérubé. 2001. Mycoplasmosis in Evening and Pine Grosbeaks with Conjunctivitis in Québec. Journal of Wildlife Diseases 37:826-830.
Millikin, R.L., and J.N.M. Smith. 1990. Sublethal effects of fenitrothion on forest passerines. Journal of Applied Ecology 27:983-1000.
Millikin, R.L. 1990. Effects of fenitrothion on the arthropod food of tree-foraging forest songbirds. Canadian Journal of Zoology 68:2235-2242.
Mineau, P. and L.J. Brownlee. 2005. Road salts and birds: an assessment of the risk with particular emphasis on winter finch mortality. Wildlife Society Bulletin 33:835-841.
Ministère des Forêts, de la Faune et des Parcs du Québec (MFFPQ). 2013. Gros plan sur les forêts. La tordeuse des bourgeons de l’épinette. [accessed December 2014].
Ministère des Forêts, de la Faune et des Parcs du Québec (MFFPQ). 2015. Aires infestées par la tordeuse des bourgeons de l’épinette au Québec en 2014 Version 1.1. Direction de la protection des forêts, Service de la gestion des ravageurs forestiers. 17 pp.
Mitchell, R.T. 1952. Consumption of Spruce Budworms by birds in a Maine spruce-fir forest. Journal of Forestry 50:387-389.
Morin, H., D. Laprise, A.-A. Simard, and S. Amouch. 2008. Spruce Budworm outbreak regimes in eastern North America. Pp 155-182 inS. Gauthier, M.-A. Vaillancourt, A. Leduc, L. De Grandpré, D. Kneeshaw, H. Morin, P. Drapeau and Y. Bergeron (eds.). Ecosystem Management in the Boreal Forest. Presse de l’Université du Québec, Québec City, Québec.
Morris, R.F., W.F. Cheshire, C.A. Miller, and D.G. Mott. 1958. The numerical response of avian and mammalian predators during a gradation of the Spruce Budworm. Ecology39:487-494.
NABCI International. 2014. Bird Conservation Regions [accessed November 2014].
NatureServe. 2015. NatureServe Explorer: An online encyclopedia of life [Web application]. Version 7.1. NatureServe, Arlington, Virginia. Web site: http://explorer.natureserve.org [accessed November 2015].
Nealis, V.G. and J. Régnière. 2004. Insect-host relationships influencing distur- bance by the Spruce Budworm in a boreal mixedwood forest. Canadian Journal of Forest Research 34:1870-1882.
Newton, I. 2008. The migration ecology of birds. Academic Press, New York, New York. 976 pp.
Ouellet, H. 1974. The birds of the Monteregian Hills and the Montreal region of Québec, Canada. National Museum of Natural Sciences, Ottawa, Ontario.
Parks Canada. 2013. Biotics Web Explorer. Evening Grosbeak. [accessed November 2014].
Partners in Flight Science Committee. 2013. Population Estimates Database, version 2013. [accessed November 2014].
Pearce, P.A., D.E. Peakall, and A.J. Erskine. 1979. Impact on forest birds of the 1976 Spruce Budworm spray operation in New Brunswick. Canadian Wildlife Service Program Notes No. 97.
Peck, G.K. and R.D. James. 1987. Breeding birds of Ontario: nidiology and distribution. Vol. 2. Royal Ontario Museum, Toronto, Ontario.
Pinel, H.W., W.W. Smith, and C.R. Wershler. 1993. Alberta Birds, 1971-1980. Vol. 2: Passerines. Provincial Museum of Alberta Natural History Occasional Paper 20. Edmonton, Alberta.
Plummer, D.A., D. Caya, A. Frigon, H. Côté, M. Giguère, D. Paquin, S. Biner, R. Harvey, and R. de Elia, 2006: Climate and Climate Change over North America as Simulated by the Canadian RCM. Journal of Climate 19:3112-3132.
Powell, G.V.N. 1984. Reproduction by an altricial songbird, the Red-winged Blackbird, in fields treated with the organophosphate insecticide fenthion. Journal of Applied Ecology 21:83-95.
Prescott, D.R.C. 1991. Winter distribution of age and sex classes in an irruptive migrant, the Evening Grosbeak. The Condor 93:694-700.
Prescott, D.R.C. 1992. Differential migration in the Evening Grosbeak (Coccothraustes vespertinus): a test of hypotheses. PhD dissertation, University of Calgary, Calgary, Alberta.
Price, D.T., R.I. Alfaro, K.J. Brown, M.D. Flannigan, R.A. Fleming, E.H. Hogg, M.P. Girardin, T. Lakusta, M. Johnston, D.W. McKenney, J.H. Pedlar, T. Stratton, R.N. Sturrock, I.D. Thompson, J.A. Trofymow, and L.A. Venier. 2013. Anticipating the consequences of climate change for Canada’s boreal forest ecosystems. Environmental Reviews 21:322-365
Rae, L.F., D.M. Whitaker, and I.G.Warkentin. 2014. Multiscale impacts of forest degradation through browsing by hyperabundant moose (Alces alces) on songbird assemblages. Diversity and Distributions 20:382-395.
Royama, T. 1984. Population dynamics of the Spruce Budworm. Ecological Monograph 54:429-462
Rousseu, F. and B. Drolet. In preparation. The Nesting Phenology of Birds in Canada. Technical Report Series. Canadian Wildlife Service, Québec Region, Environment Canada, Québec, Québec.
Root, T.L. and J.D. Weckstein. 1994. Changes in distribution patterns of select wintering North American birds from 1901 to 1989. A Century of Avifaunal Change in Western North America (J.R. Jehl, Jr., and N.K. Johnson, eds.). Studies in Avian Biology 15:191-201.
Sabine, D.L. 2010. Birds of the Atlantic Maritime Ecozone. Pages 633-691 in D.F. McAlpine and I.M. Smith (eds.).Assessment of Species Diversity in the Atlantic Maritime Ecozone NRC Research Press, Ottawa, Canada.
Sauer, J.R., S. Schwartz, and B. Hoover. 1996.The Christmas Bird Count Home Page. Version 95.1. Patuxent Wildlife Research Center, Laurel, Maryland.
Sauer, J.R., J.E. Hines, J.E. Fallon, K.L. Pardieck, D.J. Ziolkowski, Jr., and W.A. Link. 2014. The North American Breeding Bird Survey, Results and Analysis 1966 - 2012. Version 02.19.2014 USGS Patuxent Wildlife Research Center, Laurel, Maryland.
Schieck, J.M. Nietfeid, and J.B. Stelfox. 1995. Differences in bird species richness and abundance among three successional stages of aspen-dominated boreal forests. Canadian Journal of Zoology 73:1417-1431.
Schieck, J. and S.J. Song. 2006. Changes in bird communities throughout succession following fire and harvest in boreal forests of western North America: literature review and meta-analyses.Canadian Journal of Forest Research 36:1299-1318.
Scott, A.C. 1990. Breeding biology of Evening Grosbeaks. M.Sc. Thesis, University of Colorado, Boulder, Colorado.
Scott, A.C. and M. Bekoff. 1991. Breeding behavior of Evening Grosbeaks. The Condor 93:71-81.
Sewall, K.B., R. Kelsey, and T.P. Hahn. 2004. Discrete variants of Evening Grosbeak flight calls. The Condor 106:161-165.
Shaub, M.S. 1954. Summer appearances of adult and juvenal Evening Grosbeaks. Bird-Banding 25:87-94.
Sibley, D.A. 2000. The Sibley guide to birds. Alfred A. Knopf, New York, New York.
Siegel, R.B., P. Pyle, J.H. Thorne, A.J. Holguin, C.A. Howell, S. Stock, and M.W. Tingley. 2014. Vulnerability of birds to climate change in California's Sierra Nevada. Avian Conservation and Ecology 9:7. http://dx.doi.org/10.5751/ACE-00658-090107.
Sinclair, P.H., W.A. Nixon, C.D. Eckert, and N.L. Hughes. 2003. Birds of the Yukon Territory. UBC Press, Vancouver, British Columbia. 595 pp.
Smith, A., pers. comm. 2016. Email correspondence to C. Savignac. August 2016. Senior Biostatistician, Environmental Stewardship Branch, Canadian Wildlife Service, Ottawa, Ontario.
Speirs, D.H. 1968. Hesperiphona vespertina vespertina (Cooper): Eastern Evening Grosbeak. Pp. 206-237 in Austin, O.L. (ed.). Life histories of North American cardinals, grosbeaks, buntings, towhees, finches, sparrows, and their allies. U.S. National Museum Bulletin 237, Part 1.
Stralberg, D, S.M. Matsuoka, A. Hamann, E.M. Bayne, P. Solymos, F.K.A. Schmiegelow, X. Wang, S.G. Cumming, and S.J. Song. 2015a. Projecting boreal bird responses to climate change: the signal exceeds the noise. Ecological Applications 25:52-69.
Stralberg, D., E.M. Bayne, S.G. Cumming, P. Solymos, S.J. SongandF.K.A. Schmiegelow. 2015b. Conservation of future boreal forest bird communities considering lags in vegetation response to climate change: a modified refugia approach. Diversity and Distributions 21:1112-1128.
Strong, C., B. Zuckerberg, J.L. Betancourt, and W.D. Koenig. 2015. Climatic dipoles drive two principal modes of North American boreal bird irruption Proceedings of the National Academy of Sciences 112:E2795–E2802.
Sutton, A. and J.C. Tardif. 2008. Dynamique des épidémies de la livrée des forets du Manitoba au Nouveau-Brunswick. Pp 193-213 inS. Gauthier, M.-A. Vaillancourt, A. Leduc, L. De Grandpré, D. Kneeshaw, H. Morin, P. Drapeau and Y. Bergeron (eds.). Ecosystem Management in the Boreal Forest. Presse de l’Université du Québec, Québec City, Québec.
Takekawa, J.Y. and E.O. Garton. 1984. How much is an Evening Grosbeak Worth? Journal of Forestry 82:135-140.
Taverner, P.A. 1921. The Evening Grosbeak in Canada. Canadian Field-Naturalist 35:41-45.
Taylor, P. 1997. Evening Grosbeaks attracted to vehicles. Blue Jay 52:138-139.
Terrier, A., M.P. Girardin, C. Périé, P. Legendre, and Y. Bergeron. 2013. Potential changes in forest composition could reduce impacts of climate change on boreal wildfires. Ecological Applications 23:21-30.
The Boreal Avian Modeling Project. 2014. Evening Grosbeak, Coccothraustes vespertinus. [accessed November 2014].
Thomson, A.J. and Benton, R.A. 2007. A 90-year sea warming trend explains outbreak patterns of western Spruce Budworm on Vancouver Island. Forest Chronical 83:867-869.
Van Frankenhuyzen, K. 1993. The challenge of Bacillus thuringiensis. Pp. 1-35 in P.F. Entwistle, J.S. Cory, M.J. Bailey, and S. Higgs (eds.). Bacillus thuringiensis, An Environmental Biopesticide: Theory and Practice. John Wiley and Sons, Toronto, Ontario.
Van Wilgenburg, S.L., pers. comm. 2015. Email correspondence to C. Savignac. September 2015. Wildlife technician, Ecology and Conservation of Migratory Birds National Hydrology Research Center, Saskatoon..
Venier, L.A., J.L. Pearce, D.R. Fillman, D.K. McNicol, and D.A. Welsh. 2009. Effects of Spruce Budworm (Choristoneura fumiferana (Clem.)) outbreaks on boreal mixed-wood bird communities. Avian Conservation and Ecology 4:3.
Venier, L.A and S.B. Holmes. 2010. A review of the interaction between forest birds and eastern Spruce Budworm. Environmental Reviews 18:191-207.
Venier, L.A. Unpubl. data. 2014. Email correspondence to C. Savignac. November 2014. Forest Biologist, Great Lakes Forestry Centre, Sault Ste. Marie, Ontario.
Volney, W.J.A. and R.A. Fleming. 2007. Spruce Budworm (Choristoneura spp.) biotype reactions to forest and climate characteristics. Global Change Biology 13:1630-1643.
Wallace, G.J., W.P. Nickell, and R.F. Bernard. 1961. Bird mortality in the Dutch elm disease program in Michigan.Cranbrook Institute of Science41: 11-44.
Wells, J.V. 2011. Boreal forest threats and conservation status. Pp. 1-6 in J.V. Wells (ed.). Boreal birds of North America: a hemispheric view of their conservation links and significance. Studies in Avian Biology (No. 41). University of California Press, Berkeley, California.
Williams, D.W. and A.M. Liebhold. 1997. Latitudinal shifts in Spruce Budworm (Lepidoptera: Tortricidae) outbreaks and spruce-fir forest distributions with climate change. Acta Phytopathologica et Entomologica Hungarica32:205-215.
Wilson, D.J. 1981. Traffic takes heavy grosbeak toll. British Columbia Naturalist19:18-19.
Williams, D.W. and A.M. Liebhold. 2000. Spatial synchrony of Spruce Budworm outbreaks in eastern North America. Ecology 81:2753-2766
Xu, L., R.B. Myneni, F.S. Chapin III, T.V. Callaghan, J.E. Pinzon, C.J. Tucker, Z. Zhu, J. Bi, P. Ciais, H. Tømmervik, E.S. Euskirchen, B.C. Forbes, S.L. Piao, B.T. Anderson, S. Ganguly, R.R. Nemani, S.J. Goetz, P.S.A. Beck, A.G. Bunn, C. Cao, and J.C. Stroeve. 2013. Temperature and vegetation seasonality diminishment over northern lands. Nature Climate Change 3:581-586.
Zinkl, J.G., C.J. Henny, and L.R. DeWeese. 1977. Brain cholinesterase activities of birds from forests sprayed with trichlorfon (Dylox) and carbaryl (Sevin-4-oil). Bulletin of Environmental Contamination and Toxicology 17:379-386.
Biographical summary of report writer
Carl Savignac is director of Dendroica Environnement et Faune, a consulting firm specializing in avian ecology, conservation of species at risk studies, wetlands conservation and impact assessment of industrial development projects on birds and species at risk. Carl has been studying birds for over 25 years and has conducted numerous field studies in several Canadian provinces and territories. He has led a number of stewardship, research and restoration projects in southern Quebec on the Red-headed Woodpecker, Golden-winged Warbler, Cerulean Warbler, Canada Warbler, Peregrine Falcon and American Ginseng, among others. He has written over 75 scientific reports and publications, including a dozen bird species status reports in Canada and Quebec.
Appendix 1. Threats assessment for Evening Grosbeak
Threats assessment worksheet
- Species or ecosystem scientific name:
- Evening Grosbeak
- Date:
- 18/03/2015
- Assessor(s):
- Dave Fraser, Marcel Gahbauer, Carl Savignac, Bruno Drolet, Julie Perrault, Steve Van Wilgenburg, Mary Sabine
| Threat impact | Threat impact (descriptions) | Level 1 Threat impact counts: high range |
Level 1 Threat impact counts: low range |
|---|---|---|---|
| A | Very high | 0 | 0 |
| B | High | 0 | 0 |
| C | Medium | 0 | 0 |
| D | Low | 3 | 0 |
| - | Calculated overall threat impact: | Low | High |
- Assigned overall threat impact:
- D = Low
- Impact adjustment reasons:
- n/a
- Overall threat comments
- Habitat loss due to forestry operations and mortality from roads and building collisions are recognized as threats for Evening Grosbeak, but all are likely to either be of small scope or slight severity over the next decade.
| Threat ID | Threat | Impact level (calculated) | Impact (calculated) | Scope (next 10 Yrs) | Severity (10 Yrs or 3 Gen.) | Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | D | Low | Restricted - Small (1-30%) | Slight (1-10%) | High (Continuing) | blank |
| 1.1 | Housing & urban areas | D | Low | Restricted - Small (1-30%) | Slight (1-10%) | High (Continuing) | Mortality from hitting windows in buildings is a concern, although at most a minority of the population is likely to be exposed to it. However, Evening Grosbeak was reported among the ten species most frequently killed by window strikes in North America (Klem 1989, Dunn 1993), and Dunn (1993) reported that Evening Grosbeak represented 3.7% of mortalities reported in the winter of 1989-1990 (n=945). |
| 1.2 | Commercial & industrial areas | blank | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Similar issues as above, but more limited in scope, and less likely to pose a hazard given that feeders near houses increase the risk. |
| 1.3 | Tourism & recreation areas | blank | blank | blank | blank | blank | blank |
| 2 | Agriculture & aquaculture | blank | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | blank |
| 2.1 | Annual & perennial non-timber crops | blank | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | Habitat is not suitable for this species. |
| 2.2 | Wood & pulp plantations | blank | blank | blank | blank | blank | blank |
| 2.3 | Livestock farming & ranching | blank | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | blank |
| 2.4 | Marine & freshwater aquaculture | blank | blank | blank | blank | blank | blank |
| 3 | Energy production & mining | blank | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | blank |
| 3.1 | Oil & gas drilling | blank | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | blank |
| 3.2 | Mining & quarrying | blank | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | blank |
| 3.3 | Renewable energy | blank | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | There are no data on whether EVGR is affected by wind turbines. However, they are a diurnal migrant and therefore not likely at much risk in most cases. |
| 4 | Transportation & service corridors | D | Low | Restricted (11-30%) | Slight (1-10%) | High (Continuing) | blank |
| 4.1 | Roads & railroads | D | Low | Restricted (11-30%) | Slight (1-10%) | High (Continuing) | These birds are attracted to road salts, and at risk of vehicle collisions as well as salt poisoning. A minority of the population is expected to be exposed to this risk, and of those, probably only a small fraction would suffer mortality, especially as some of the roads are likely to be lightly traveled. |
| 4.2 | Utility & service lines | blank | blank | blank | blank | blank | blank |
| 4.3 | Shipping lanes | blank | blank | blank | blank | blank | blank |
| 4.4 | Flight paths | blank | blank | blank | blank | blank | blank |
| 5 | Biological resource use | D | Low | Small (1-10%) | Serious - Moderate (11-70%) | High (Continuing) | blank |
| 5.1 | Hunting & collecting terrestrial animals | blank | blank | blank | blank | blank | blank |
| 5.2 | Gathering terrestrial plants | blank | blank | blank | blank | blank | blank |
| 5.3 | Logging & wood harvesting | D | Low | Small (1-10%) | Serious - Moderate (11-70%) | High (Continuing) | Harvesting results in habitat loss for this species, but over the next decade only a small portion of the population is likely to be affected. |
| 5.4 | Fishing & harvesting aquatic resources | blank | blank | blank | blank | blank | blank |
| 6 | Human intrusions & disturbance | blank | blank | blank | blank | blank | blank |
| 6.1 | Recreational activities | blank | blank | blank | blank | blank | blank |
| 6.2 | War, civil unrest & military exercises | blank | blank | blank | blank | blank | blank |
| 6.3 | Work & other activities | blank | blank | blank | blank | blank | blank |
| 7 | Natural system modifications | blank | Unknown | Unknown | Unknown | High (Continuing) | blank |
| 7.1 | Fire & fire suppression | blank | blank | blank | blank | blank | Fire suppression could actually affect the Spruce Budworm which would actually have a positive affect on the habitat for this species. |
| 7.2 | Dams & water management/use | blank | blank | blank | blank | blank | blank |
| 7.3 | Other ecosystem modifications | blank | Unknown | Unknown | Unknown | High (Continuing) | Prevention measures are being taken in QC and NB for Spruce Budworm (spraying, selective logging). Impact of spraying is negligible on Spruce Budworm. Could be a big issue for parts of the country but the impacts are unknown. Food supply could be reduced from spraying. |
| 8 | Invasive & other problematic species & genes | blank | blank | blank | blank | blank | blank |
| 8.1 | Invasive non-native/alien species | blank | blank | blank | blank | blank | blank |
| 8.2 | Problematic native species | blank | blank | blank | blank | blank | blank |
| 8.3 | Introduced genetic material | blank | blank | blank | blank | blank | blank |
| 9 | Pollution | blank | Unknown | Small (1-10%) | Unknown | High (Continuing) | blank |
| 9.1 | Household sewage & urban waste water | blank | blank | blank | blank | blank | blank |
| 9.2 | Industrial & military effluents | blank | Unknown | Small (1-10%) | Unknown | High (Continuing) | Road salt is included here. It may have a toxic effect on EVGR but more research is needed. |
| 9.3 | Agricultural & forestry effluents | blank | blank | blank | blank | blank | Where food supply may be reduced from insect spraying/eradication is dealt with in 7.3 (bt). Other insecticides (Mimic) is being used as well that may affect EVGR but data are lacking |
| 9.4 | Garbage & solid waste | blank | blank | blank | blank | blank | blank |
| 9.5 | Air-borne pollutants | blank | blank | blank | blank | blank | blank |
| 9.6 | Excess energy | blank | blank | blank | blank | blank | blank |
| 10 | Geological events | blank | blank | blank | blank | blank | blank |
| 10.1 | Volcanoes | blank | blank | blank | blank | blank | blank |
| 10.2 | Earthquakes/tsunamis | blank | blank | blank | blank | blank | blank |
| 10.3 | Avalanches/landslides | blank | blank | blank | blank | blank | blank |
| 11 | Climate change & severe weather | blank | Negligible | Negligible (<1%) | Negligible (<1%) | Unknown | blank |
| 11.1 | Habitat shifting & alteration | blank | Negligible | Negligible (<1%) | Negligible (<1%) | Unknown | In the longer term, this could be a more significant threat in terms of habitat shifting of Balsam Fir and Spruce Budworm forest in the east and the United States (climate models available show this). Data for shorter term effects only show changes in phenology but otherwise impacts unknown. |
| 11.2 | Droughts | blank | blank | blank | blank | blank | blank |
| 11.3 | Temperature extremes | blank | blank | blank | blank | blank | blank |
| 11.4 | Storms & flooding | blank | blank | blank | blank | blank | blank |
Classification of Threats adopted from IUCN-CMP, Salafsky et al. (2008).