
Plains bison and wood bison: COSEWIC assessment and status report 2013

Bison bison bison - THREATENED
Bison bison athabascae - SPECIAL CONCERN
2013
Table of Contents
- Document Information
- COSEWIC Assessment Summary
- COSEWIC Executive Summary
- Technical Summary (Plains Bison)
- Technical Summary (Wood Bison)
- Preface
- Wildlife Species Description and Significance
- Distribution
- Population Units to be Assessed by COSEWIC
- Habitat
- Biology
- Population Sizes and Trends
- Threats and Limiting Factors
- Protection, Status, and Ranks
- Acknowledgements and Authorities Contacted
- Information Sources
- Biographical Summary of Report Writer(s)
- Collections Examined
List of Figures
- Figure 1. Key morphological differences between mature Wood Bison (top) and Plains Bison bulls. Line drawing courtesy of Wes Olson.
- Figure 2. Photographs of mature Wood Bison (top) and Plains Bison (bottom) bulls during summer at Elk Island National Park. Note the morphological and pelage differences as per Figure 1. Photographs courtesy of Wes Olson.
- Figure 3. Photographs of adult female Wood Bison (top) and Plains Bison (bottom) during summer at Elk Island National Park. Photographs courtesy of Wes Olson.
- Figure 4. Historical (pre-settlement) distribution of Wood Bison and Plains Bison in North America. Modified from Gates et al. (2010). Polygons courtesy of Keith Aune, Wildlife Conservation Society.
- Figure 5. Decade interval timeline of Plains Bison captures and translocations for lineages used to populate Canadian Parks spanning 1860 to 1930. Sources: Coder 1975, Dary 1974, Ogilvie 1979, and Lothian 1981.
- Figure 6. Approximate original (pre-settlement) range of Plains Bison in Canada (dashed red line) and current distribution of five subpopulations considered in this assessment. Striped polygon is the original distribution of Wood Bison. Note that McCusker River range (3) occurs at the periphery of Wood Bison, close to the edge of the original range of Plains Bison.
- Figure 7. Approximate original (pre-settlement) range of Wood Bison in Canada (dashed red line) and current distribution of 9 subpopulations considered in this assessment. Striped polygon approximates the original distribution of Plains Bison.
List of Tables
- Table 1. Translocations and establishment of Plains and Wood Bison wild subpopulations considered in this assessment since the 1960s. Elk Island National Park (EINP) was the source of stock for many herds.
- Table 2. Status and trends of Plains Bison subpopulations included in the designatable unit. See Population Sizes and Trends for details on survey methods for each subpopulation (e.g., for some subpopulation total numbers represent minimum counts) and specific information on age structure where available.
- Table 3. Status and trends of subpopulations included in the Wood Bison designatable unit. Population trends were based on comparing current estimates (total individuals) with those from the earliest year with survey data within three generations (ca. 1989). See Population Sizes and Trends for details on survey methods for each subpopulation (e.g., some subpopulation total numbers represent minimum counts) and specific information on age structure where available.
Document Information

Committee on the Status of Endangered Wildlife in Canada (COSEWIC) status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2013. COSEWIC assessment and status report on the Plains Bison Bison bison bison and the Wood Bison Bison bison athabascae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xv + 109 pp. (Species at Risk Public Registry website).
Previous report(s):
COSEWIC. 2004. COSEWIC assessment and status report on the plains bison Bison bison bison in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 71 pp.
COSEWIC. 2000. COSEWIC assessment and update status report on the wood bison Bison bison athabascae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 29 pp.
Ruckstuhl, K.E. 2000. Update COSEWIC status report on the wood bison Bison bison athabascae in COSEWIC assessment and status report on wood bison Bison bison athabascae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-29 pp.
Wood Bison Recovery Team. 1988. Update COSEWIC status report on the wood bison Bison bison athabascae in Canada. Committee on Status of Endangered Wildlife in Canada. Ottawa. 103 pp.
Novakowski, N.S. 1978. COSEWIC status report on the wood bison Bison bison athabascae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 30 pp.
Production note:
COSEWIC would like to acknowledge Cormack Gates and Thomas Jung for writing the status report on the Plains Bison (Bison bison bison) and Wood Bison (Bison bison athabascae) in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Justina Ray, Co-chair of the COSEWIC Terrestrial Mammals Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-953-3215
Fax: 819-994-3684
COSEWIC E-mail
COSEWIC web site
Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur le Bison des prairies (Bison bison bison) et le Bison des bois (Bison bison athabascae) au Canada.
Cover illustration/photo:
Wood Bison (left) and Plains Bison (right) -- Photographs courtesy of Wes Olson.
©Her Majesty the Queen in Right of Canada, 2014.
Catalogue No. CW69-14/379-2014E-PDF
ISBN 978-1-100-23570-7

COSEWIC Assessment Summary
Assessment Summary - November 2013
- Common name
- Plains Bison
- Scientific name
- Bison bison bison
- Status
- Threatened
- Reason for designation
- This bison occurs in only five isolated wild subpopulations in Canada. There are approximately 1,200 to 1,500 mature individuals, of which about half occur in one subpopulation located outside of the historical range. The total number of individuals has increased by 36% since the last assessment in 2004, but the total remains a tiny fraction of their numbers of 200 years ago. Currently they occupy less than 0.5% of their original range in Canada. This animal continues to face a number of threats to its persistence. Further increases in population size or the addition of new subpopulations is curtailed by fragmented or unsuitable habitat that is often managed to exclude bison. An overall decline is projected for wild subpopulations because they are managed to control or reduce population size and are subject to unpredictable but potentially catastrophic future events, mainly disease outbreaks and extreme weather.
- Occurrence
- British Columbia, Alberta, Saskatchewan
- Status history
- Designated Threatened in May 2004. Status re-examined and confirmed in November 2013.
Assessment Summary - November 2013
- Common name
- Wood Bison
- Scientific name
- Bison bison athabascae
- Status
- Special Concern
- Reason for designation
- This bison only occurs in the wild in Canada. There are currently 5,136 to 7,172 mature individuals in nine isolated wild subpopulations. The population has increased since 1987, mostly due to the establishment of new wild subpopulations within the original range. About 60% of the overall population is included in Wood Buffalo National Park and surrounding areas, and is affected by two cattle diseases, bovine brucellosis and tuberculosis. Two wild subpopulations have recently experienced significant mortality events demonstrating the inherent vulnerability of small isolated populations. The Mackenzie herd decreased by 53% due to an outbreak of anthrax and the Hay-Zama decreased by 20% due to starvation during a severe winter. Further increases to the population size or the addition of new wild subpopulations is not likely, as recovery is constrained by fragmented or unsuitable habitat, road mortality, disease management associated with livestock and commercial bison operations, and disease outbreaks.
- Occurrence
- Yukon, Northwest Territories, British Columbia, Alberta, Manitoba
- Status history
- Designated Endangered in April 1978. Status re-examined and designated Threatened in April 1988 and May 2000. Status re-examined and designated Special Concern in November 2013.
COSEWIC Executive Summary
Plains Bison
Bison bison bison
and the
Wood Bison
Bison bison athabascae
Wildlife Species Description and Significance
The American bison is a member of the wild cattle family and is the largest land mammal in North America. The two recognized subspecies--Plains Bison (Bison bison bison) and Wood Bison (B. b. athabascae)--have distinct morphology, body shape, size, and pelage patterns. Phylogenetic divisions between them remain despite a massive translocation of Plains Bison into the remnant Wood Bison population during the 1920s, which has had a substantial impact on their genetic and distributional integrity.
Bison once served as both an ecological and cultural keystone species, having a disproportionate influence on ecological processes and biodiversity in socio-ecological systems it occupied. This animal has been important to the material and spiritual cultures of many Aboriginal peoples. Since the 1970s, Bison have also increased in economic and commercial importance. This report provides information necessary to assess the wild component of the species, in keeping with COSEWIC guidelines.
Distribution
The late Holocene, pre-Columbian range of the American Bison extended from the desert grasslands of northern Mexico to the meadow systems in interior Alaska and from the woodlands of Manitoba to the Rocky Mountains. The continental divide between Alberta and British Columbia marked the approximate western extent of Plains Bison. The transition between Parklands and the Boreal Forest marked the northern extent of Plains Bison and southern limit of Wood Bison.
Both Wood Bison and Plains Bison populations declined sharply during the 1800s, largely as a result of unsustainable hunting. By the end of the 19th century the Plains Bison had been extirpated from the wild in Canada, but a small number of Wood Bison remained in what is now Wood Buffalo National Park. In 2013, wild Plains Bison occurred in five isolated subpopulations: three in Saskatchewan, one in Alberta and one in British Columbia--about 2% of their original range. There are 9 wild Wood Bison subpopulations ranging in Alberta, British Columbia, Manitoba, Yukon and Northwest Territories, occupying about 5% of their original range.
Habitat
The most important habitats for Wood and Plains Bison are those producing winter forage, consisting primarily of grasses, sedges, and rushes. Plains Bison habitat included prairie grasslands and adjacent mixed woodlands in Manitoba, central Saskatchewan, and southwestern Alberta. Conversion of native prairies to crop and livestock agriculture occurred rapidly after bison were eliminated. Loss of native rangelands is still taking place, albeit at a reduced rate. The potential for conflicts with crop agriculture and livestock grazing, including programs to control the spread of bovine tuberculosis and brucellosis from wild bison, all limit population and range expansion for wild Plains and Wood Bison in much of their range.
Biology
Female Bison typically produce their first, single calf (rarely twins) at three years of age and reproductive senescence occurs after 13 to 15 years of age. Fecundity varies between individuals and among populations depending on nutrition and heredity. Generation time for Bison is estimated at eight years. Males as young as 1.5 years can reproduce in well-nourished, captive populations, but full morphological and behavioural maturity (adulthood) is not achieved until six or seven years of age. Sub-adult males rarely have an opportunity to breed in the presence of adult males. Competition for mating opportunities among adult males is an important aspect of the evolutionary ecology of bison. Wolves, Grizzly Bears, and Coyotes are the primary predators.
Population Sizes and Trends
All wild Plains Bison subpopulations in Canada today are the descendants of approximately 81 ancestors captured in three locations in the 1870s and 1880s, and persist as a tiny fraction of their original numbers (tens of millions in North America). The 5 wild subpopulations are in Elk Island National Park and four others originating from that source. One new subpopulation was established in Grasslands National Park since the last COSEWIC status assessment in 2004. There are an estimated 2,335-2,573 Plains Bison, 1,204-1,490 of which are mature individuals. This represents a 36% increase since 2004, although one subpopulation is currently in decline. Overall, there is an unquantified but increasing trend over the past 3 generations.
The ca. 250 Wood Bison that persisted in what is now Wood Buffalo National Park into the late 1800s grew to 1,500-2,000 individuals when the Wood Buffalo National Park was established in 1922. Political exigencies resulted in the translocation of more than 6,000 Plains Bison to the Park in the late 1920s where Wood and Plains Bison subsequently interbred. All Wood Bison existing today are descendants of this mixed ancestry although have remained morphologically and genetically distinct from Plains Bison and are separately managed. Two translocations from Wood Buffalo National Park occurred during the 1960s, including one to Elk Island National Park to establish a disease-free population to support recovery. This subpopulation has directly or indirectly been the source of stock to establish 7 other subpopulations, one since the last assessment in 2001. There are between 7,642-10,458 Wood Bison in 9 wild subpopulations, of which 5,213-7,191 are mature individuals. This represents a substantial increase over the past ca. 3 generations (1987: 1,827) through significant recovery efforts, and a 47% increase since 2000. Although 8 of the 9 wild subpopulations have increased in number since the last COSEWIC assessment, 2 have experienced significant mortality since 2012 due to disease (anthrax) and starvation following a severe winter. All but 2 subpopulations number fewer than 500 individuals. The Greater Wood Buffalo National Park meta-population represents about 60% of the Canadian population of wild Wood Bison today, and they are diseased.
Threats and Limiting Factors
The overall calculated threat impact based on the World Conservation Union-Conservation Measures Partnership ‘unified threats classification system’ is Very High for Plains Bison and High for Wood Bison. The highest impact threat facing both is hunting and population control. Social intolerance due to perceived competition with other ungulates, disease transmission, property damage, and human safety is a significant factor determining policies that reduce, control, and limit the number of wild Plains and Wood Bison in large landscapes. Unregulated hunting of some subpopulations constrains effective population size below a threshold where small population effects may negatively impact viability.
Disease (livestock-borne and native, e.g., anthrax) and severe weather are other threats that have caused significant mortality events, both recently and historically. The continued existence of reportable cattle diseases in the Greater Wood Buffalo National Park Wood Bison meta-population is the largest threat in terms of geographic scale and potential to impact neighbouring subpopulations. Plains Bison habitat loss from conversion of native range to croplands is ongoing with the sale of public rangelands being a significant threat. Wild Plains Bison are primarily limited by land tenure and use patterns, and by land use, grazing, and animal management policies. Road mortality (Wood Bison) and genetic introgression from cattle and private bison holdings serve as localized threats for both Wood and Plains Bison. Both are also limited by founder effects and small population sizes (< 500).
Protection, Status, and Ranks
Plains Bison in Canada have no status under the federal Species at Risk Act. They are classified as wildlife in the provincial wildlife acts of British Columbia and Saskatchewan, but are not wildlife under provincial wildlife acts in Alberta or Manitoba. In Alberta and Manitoba, all bison considered livestock. Plains Bison are not listed under the U.S. Endangered Species Act, despite successive petitions to do so.
Wood Bison are listed as Threatened under Schedule I of the federal Species at Risk Act upon proclamation in June 2003. Wood Bison are classed as wildlife in the wildlife acts of Manitoba, Alberta, British Columbia, and the Northwest Territories. In Yukon, they are classified as a Transplanted Species in the Yukon Wildlife Act. Wood Bison is listed on Appendix II of CITES, and under the U.S. Endangered Species Act as Threatened. Globally, the IUCN Red List ranks American Bison (both subspecies) as Near Threatened. NatureServe has assigned a global rank of G4 to American Bison, with national ranks of N4 for U.S. and N3N4 for Canada. The global (and national) rank for Wood Bison is G4T2Q (N2N3) and Plains Bison G4TU (N3N4). Canada's General Status program considers American Bison (both subspecies) as At Risk and At Risk in Yukon, Northwest Territories, Alberta and Manitoba, May be at Risk in British Columbia, and Sensitive in Saskatchewan.
Technical Summary (Plains Bison)
Bison bison bison
Plains Bison
Bison des prairies
- Range of occurrence in Canada (province/territory/ocean):
- Manitoba (historical), Saskatchewan, Alberta, British Columbia
Demographic Information
Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines (2008) is being used)
ca. 8 yrs
Is there an [observed, inferred, or projected] continuing decline in number of mature individuals?
No
Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations]
None
[Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations].
Increase of ca. 36% in total number of individuals in 13 yrs.
[Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations].
Uncertain (increases will be constrained by human conflict)
[Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future.
Overall increasing trend
Are the causes of the decline clearly reversible and understood and ceased?
N/A
Are there extreme fluctuations in number of mature individuals?
No
Extent and Occupancy Information
Estimated extent of occurrence
403,592 km2
Index of area of occupancy (IAO)
(Always report 2x2 grid value).
14,764 km2
Is the total population severely fragmented?
Possibly. Close to half the population persists in small and isolated habitat patches relative to the original distribution. Population expansion is ultimately constrained by socio-political context.
Number of locationsFootnote∗
5
Is there an [observed, inferred, or projected] continuing decline in extent of occurrence?
No
Is there an [observed, inferred, or projected] continuing decline in index of area of occupancy?
No
Is there an [observed, inferred, or projected] continuing decline in number of populations?
No
Is there an [observed, inferred, or projected] continuing decline in number of locationsFootnote∗?
No
Is there an [observed, inferred, or projected] continuing decline in [area, extent and/or quality] of habitat?
Yes. Some ongoing degradation and loss. Livestock grazing and human conflict limit availability of potential habitat. Increasing industrial access threatens the habitat of at least one subpopulation.
Are there extreme fluctuations in number of populations?
No
Are there extreme fluctuations in number of locationsFootnote∗?
No
Are there extreme fluctuations in extent of occurrence?
No
Are there extreme fluctuations in index of area of occupancy?
No
| Subpopulation | N Mature Individuals (estimated) |
|---|---|
| Pink Mountain (BC) | 629-699 |
| Elk Island National Park (AB) | 204-300 |
| Grasslands National Park (AB) | 189 |
| McCusker River (SK) | 51-113 |
| Sturgeon River (SK) | 130-188 |
| Total Plains Bison population (see Table 2) | 1,204-1,490 |
Quantitative Analysis
Probability of extinction (PoE) in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years].
The PoE has not been quantified
Threats (actual or imminent, to populations or habitats)
The highest impact threat is hunting (both controlled and unregulated), used to manage growth, prevent range expansion, and reduce bison-human conflicts, including discouraging bison use of agricultural areas and roadsides. Another high-medium impact threat comes from a variety of cattle-borne and native (anthrax) pathogens. Agricultural settlement and land conversion in the grasslands and parkland areas is ongoing, and the sale of public rangelands is an increasing concern. Land tenure and use patterns, and land use, grazing, and animal management policies constrain recovery prospects. A lower but locally significant threat includes genetic introgression from cattle and privately owned bison.
Rescue Effect (immigration from outside Canada)
Status of outside population(s)? The United Stated Forest Service classifies Plains Bison as “Not Sensitive in Region 2 and Not of Concern” by its Species Conservation Program assessment (USDA Forest Service 2009). The rationale for this classification is that populations and habitats are currently stable or increasing.
Is immigration known or possible?
No immigration from wild populations
Would immigrants be adapted to survive in Canada?
Yes
Is there sufficient habitat for immigrants in Canada?
No
Is rescue from outside populations likely?
No rescue from wild populations
Status History
- COSEWIC:
- Designated Threatened in May 2004. Status re-examined and confirmed in November 2013.
Author of Technical Summary: Cormack Gates and Justina Ray
Status and Reasons for Designation
- Status:
- Threatened
- Alpha-numeric code:
- C2a(i)
- Reason for Designation:
- This bison occurs in only five isolated wild subpopulations in Canada. There are approximately 1,200 to 1,500 mature individuals, of which about half occur in one subpopulation located outside of the historical range. The total number of individuals has increased by 36% since the last assessment in 2004, but the total remains a tiny fraction of their numbers of 200 years ago. Currently they occupy less than 0.5% of their original range in Canada. This animal continues to face a number of threats to its persistence. Further increases in population size or the addition of new subpopulations is curtailed by fragmented or unsuitable habitat that is often managed to exclude bison. An overall decline is projected for wild subpopulations because they are managed to control or reduce population size and are subject to unpredictable but potentially catastrophic future events, mainly disease outbreaks and extreme weather.
- Criterion A:
- (Decline in Total Number of Mature Individuals): Not applicable. The overall trend over the past 3 generations is increasing.
- Criterion B:
- (Small Distribution Range and Decline or Fluctuation): Not applicable. Both the EO and IAO exceed thresholds for this criterion.
- Criterion C:
- (Small and Declining Number of Mature Individuals): Meets Threatened C2a(i) since the total number of mature individuals is fewer than 10,000, and there is a projected continuing decline in number of mature individuals because: 1) management practices are in place to prevent the expansion of most subpopulations beyond their current size; 2) the largest subpopulation is being managed to reduce numbers; 3) there is a strong potential for reductions caused by catastrophic future events, and 4) no subpopulation is estimated to contain more than 1,000 mature individuals.
- Criterion D:
- (Very Small or Restricted Population): Not applicable. Almost meets Threatened D1 with a population estimated between 1,200-1,500 mature individuals. Although restricted to five locations, does not meet D2 because the locations are each isolated by considerable geography, and no one threatening event would impact all or even several subpopulations in a very short period of time enough to render the population endangered or extinct in a short period of time.
- Criterion E:
- (Quantitative Analysis): Not applicable.
Technical Summary (Wood Bison)
Bison bison athabascae
Wood Bison
Bison des bois
- Range of occurrence in Canada (province/territory/ocean):
- Manitoba, Saskatchewan (historical), Alberta, British Columbia, Yukon, Northwest Territories
Demographic Information
Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines(2008) is being used)
ca. 8 yrs
Is there an [observed, inferred, or projected] continuing decline in number of mature individuals?
No. Total population has increased by almost 400% in the last 3 generations, following several reintroductions.
Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations]
Total population is increasing
[Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations].
Total population has increased in the last 3 generations
[Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations].
Uncertain.
[Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future.
Increased rapidly in past 10 years. Uncertain in future 3 generations.
Are the causes of the decline clearly reversible and understood and ceased?
N/A
Are there extreme fluctuations in number of mature individuals?
Doesn’t meet the definition, although severe mortality events have removed up to half of some subpopulations, and the largest has fluctuated 2-fold over two decades.
Extent and Occupancy Information
Estimated extent of occurrence
1,187,546 km2
Index of area of occupancy (IAO)
(Always report 2x2 grid value).
101,492 km2
Is the total population severely fragmented?
No. Although some subpopulations persist in isolated fragments, 60% of population is in Greater Wood Buffalo National Park ecosystem.
Number of locationsFootnote∗.1
9
Is there an [observed, inferred, or projected] continuing decline in extent of occurrence?
No
Is there an [observed, inferred, or projected] continuing decline in index of area of occupancy?
No
Is there an [observed, inferred, or projected] continuing decline in number of populations?
No
Is there an [observed, inferred, or projected] continuing decline in number of locationsFootnote∗.1?
No
Is there an [observed, inferred, or projected] continuing decline in [area, extent and/or quality] of habitat?
Possibly
Are there extreme fluctuations in number of populations?
No
Are there extreme fluctuations in number of locationsFootnote∗.1?
No
Are there extreme fluctuations in extent of occurrence?
No
Are there extreme fluctuations in index of area of occupancy?
No
| Subpopulation | N Mature Individuals (estimated) |
|---|---|
| Greater Wood Buffalo meta-population (NT & AB) | 3,164-4,226 |
| Elk Island National Park (AB) | 207 |
| Mackenzie (NT) | 344-705 |
| Nahanni (NT & BC) | 150-444 |
| Aishihik (YT) | 763-956 |
| Nordquist (BC) | 85-95 |
| Etthithun (BC & AB) | 116-134 |
| Hay-Zama (AB) | 283 |
| Chitek Lake (MB) | 155-190 |
| Total Wood Bison population (see Table 3) | 5,213-7,191 |
Quantitative Analysis
Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years].
PoE has not been quantified
Threats (actual or imminent, to populations or habitats)
The highest impact threat is hunting (both controlled and unregulated), used to manage growth, prevent range expansion, and reduce bison-human conflicts, including discouraging bison use of agricultural areas and roadsides. An additional high-medium impact threat comes from a variety of cattle-borne and native (anthrax) pathogens and severe weather events, which have generated high mortality events within various subpopulations both historically and recently. Lower but locally significant threats include road mortality and genetic introgression from cattle and privately owned bison.
Rescue Effect (immigration from outside Canada)
Status of outside population(s)? 100% of the wild population is in Canada.
Is immigration known or possible?
No
Would immigrants be adapted to survive in Canada?
Yes
Is there sufficient habitat for immigrants in Canada?
No
Is rescue from outside populations likely?
No
Status History
- COSEWIC:
- Designated Endangered in April 1978. Status re-examined and designated Threatened in April 1988 and May 2000. Status re-examined and designated Special Concern in November 2013.
Author of Technical Summary: Cormack Gates and Justina Ray
Status and Reasons for Designation
- Status:
- Special Concern
- Alpha-numeric code:
- Not applicable
- Reason for Designation:
- This bison only occurs in the wild in Canada. There are currently 5,136 to 7,172 mature individuals in nine isolated wild subpopulations. The population has increased since 1987, mostly due to the establishment of new wild subpopulations within the original range. About 60% of the overall population is included in Wood Buffalo National Park and surrounding areas, and is affected by two cattle diseases, bovine brucellosis and tuberculosis. Two wild subpopulations have recently experienced significant mortality events demonstrating the inherent vulnerability of small isolated populations. The Mackenzie herd decreased by 53% due to an outbreak of anthrax and the Hay-Zama decreased by 20% due to starvation during a severe winter. Further increases to the population size or the addition of new wild subpopulations is not likely, as recovery is constrained by fragmented or unsuitable habitat, road mortality, disease management associated with livestock and commercial bison operations, and disease outbreaks.
- Criterion A:
- (Decline in Total Number of Mature Individuals): Not applicable. The overall trend over the past 3 generations is increasing.
- Criterion B:
- (Small Distribution Range and Decline or Fluctuation): Not applicable. Both the EO and IAO exceed thresholds for this criterion.
- Criterion C:
- (Small and Declining Number of Mature Individuals): Not applicable, the number of mature individuals is fewer than 10,000 but there is no observed, projected or inferred continuing decline.
- Criterion D:
- (Very Small or Restricted Total Population): Not applicable. The population is estimated at greater than 1,000 mature individuals (D1) and the number of locations (9) exceeds the threshold (D2).
- Criterion E:
- (Quantitative Analysis): Not applicable.
Preface
This updated report is based on evidence collected since the 2000 COSEWIC status update for the Wood Bison, Bison bison athabascae,in Canada (Ruckstuhl 2000), the 2001 Recovery Plan for Wood Bison (Gates et al. 2001a), and the 2004 Status Report for Plains Bison, Bison bison bison, in Canada (COSEWIC 2004). This is the first COSEWIC status assessment to consider both Plains Bison and Wood Bison in one report.
Assessed as Endangered, Wood Bison were one of 6 species evaluated by COSEWIC at the committee’s first species assessment meeting in 1978. In 1988, Wood Bison were reassessed as Threatened due to increases in both distribution and abundance. A subsequent review by COSEWIC in 2000 maintained a Threatened status. Wood Bison were listed as Threatened under Schedule 1 of SARA when the Act came into force in 2003.
COSEWIC assessed Plains Bison for the first time in 2004, giving it Threatened status. Plains Bison are not, however, currently included on the legal list under Schedule 1 of the Act. In 2005, the Governor in Council decided not to list Plains Bison because of “potential economic implications for the Canadian bison industry”, pledging instead to work “with provincial governments, the bison industry and other stakeholders to develop an approach for the recovery of wild plains bison” (Order Amending Schedules 1 to 3 to the Species at Risk Act, 2005: 1769).
This report provides information necessary to assess the wild component of the species, with “wild” defined in keeping with COSEWIC's Guidelines for Manipulated Populations (COSEWIC 2010a). Considerable new information has been generated for Plains Bison and Wood Bison since 2004 and 2000, respectively. Available Aboriginal Traditional Knowledge (ATK) was gathered and presented in a COSEWIC ATK Source Report (COSEWIC 2011a) and COSEWIC ATK Assessment Report (COSEWIC 2012). New subpopulations have been established, including a new Plains Bison subpopulation in Saskatchewan and a new Wood Bison subpopulation in British Columbia. New inventories have been conducted for most populations since the last status reports.

COSEWIC History
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC Mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC Membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2013)
Wildlife Species
A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
- Special Concern (SC)Footnote*
- A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
- Not at Risk (NAR)Footnote**
- A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
- Data Deficient (DD)Footnote***
- A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife Species Description and Significance
Name and Classification
Bison belong to the order Artiodactyla, suborder Ruminantia, family Bovidae, subfamily Bovinae, tribe Bovini (wild cattle), and the genus Bison (Brands 1989-2005). The genus Bison is represented as two extant species, European Bison (B. bonasus) and American Bison (B. bison). American Bison are further segregated into two subspecies, Plains Bison (B. b. bison) and Wood Bison (B. b. athabascae) (Wilson and Reeder 2005).
Linneaus (1758) first classified bison in the genus Bos with other cattle-like mammals. Hamilton Smith (in Griffith et al. 1827) classified bison to a sub-genus (Bison) on the basis of their morphological distinctiveness, which Knight (1849) then elevated to the level of genus (Skinner and Kaisen 1947). Linnaeus (1758) recognized two species of bison, the European form (B. bonasus) and American Bison (B. bison). Rhoads (1898) considered Wood Bison (B. b. athabascae) to be a subspecies of American Bison distinct from Plains Bison (B. b. bison). The IUCN American Bison Specialist Group reviewed the taxonomy of American Bison (Gates et al. 2010), upholding the conventions described above, but noting controversies concerning both genus and subspecies designations (Boyd et al. 2010), which continue (Cronin et al. 2013).
More recently, mitochondrial DNA analyses suggest that American Bison (Bison bison) may be a junior synonym of Steppe Bison (Bison priscus; B. Shapiro, University of California Santa Cruz, pers. comm.).
Morphological Description
American Bison are the largest land mammal in North America. The body is tall and narrow, its height accentuated by vertical (spinous) processes of the thoracic vertebrae anchoring muscles and ligaments forming the hump. In males, the head is massive and is likely used in intraspecific combat. The head is highly protected by thick hair, a thick dermal shield, and a lattice of bony structures isolating the cranium from the crown of the skull. Body size is sexually dimorphic, with males outweighing females (Reynolds et al. 2003). In a similar environment at Elk Island National Park, the asymptotic weight of Wood Bison males was 880 kg vs. 739 kg for Plains Bison males. The mature weight of females was also larger in Wood Bison than Plains Bison: 540 kg vs. 440 kg (Reynolds et al. 2003).
Plains and Wood Bison differ in cranial and skeletal morphology (McDonald 1981, van Zyll de Jong 1986). Geographic variation in Plains Bison was continuous (clinal) along a north–south axis, with smaller animals in the eastern and southern portion of the range and larger ones in the north, including the Canadian prairies (Reynolds et al. 2003). In contrast, van Zyll de Jong (1986) found a discontinuity in skeletal morphology between Plains Bison and Wood Bison. Plains Bison and Wood Bison can also be distinguished by external morphology (van Zyll de Jong et al. 1995), including the anterior slope of the hump, location of the highest point on the hump, angle of the hump, cape variegation and demarcation, upper front leg hair, frontal display hair, ventral neck mane, and beard (Figures 1-3). Geist (1991) suggested these differences are environmentally induced. However, a study by van Zyll de Jong et al. (1995) showed the traits are not affected by geographic location, indicating they are genetically controlled.
Figure 1. Key morphological differences between mature Wood Bison (top) and Plains Bison bulls. Line drawing courtesy of Wes Olson.
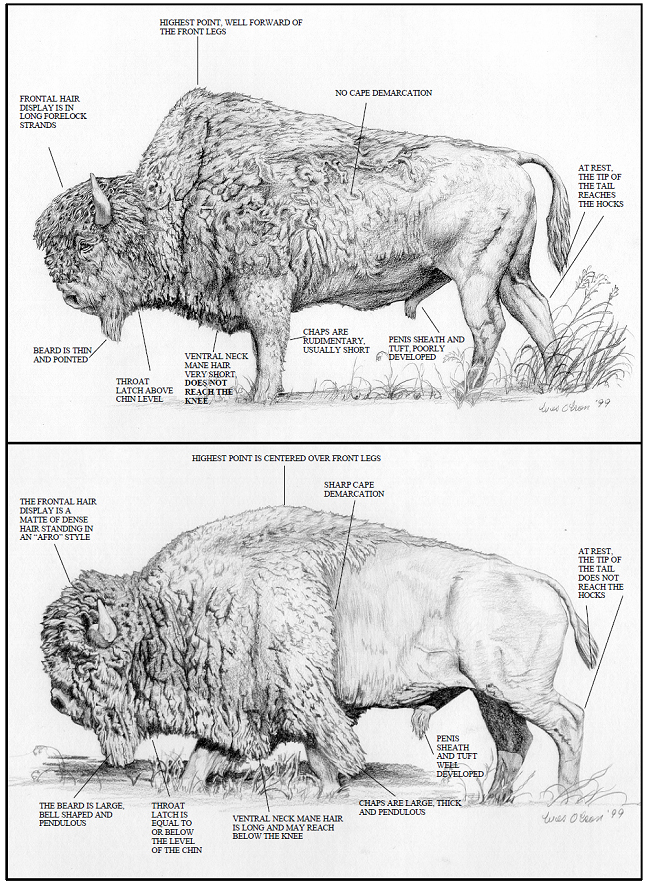
Long description for Figure 1
Annotated illustrations showing adult male Wood Bison (top image) and Plains Bison (bottom image). The annotations point out key morphological differences between the two bison. These include frontal hair display (Wood Bison: long forelock strands; Plains Bison: matte of dense hair); cape demarcation (Wood Bison: no demarcation; Plains Bison: sharp demarcation); beard (Wood Bison: pointed; Plains Bison: bell-shaped and pendulous); highest point (Wood Bison: forward of front legs; Plains Bison: centred over front legs); tail length (Wood Bison: reaches hocks; Plains Bison: does not reach hocks); throat latch (Wood Bison: above chin; Plains Bision: equal to or below level of chin); ventral neck mane hair (Wood Bison: short, above knee; Plains Bison: long and may reach below knee); penis sheath and tuft (Wood Bison: poorly developed; Plains Bison: well developed); chaps (Wood Bison: rudimentary; Plains Bison: large, thick and pendulous). Further details can be found in the preceding/next paragraph(s).
Figure 2. Photographs of mature Wood Bison (top) and Plains Bison (bottom) bulls during summer at Elk Island National Park. Note the morphological and pelage differences as per Figure 1. Photographs courtesy of Wes Olson.

Figure 3. Photographs of adult female Wood Bison (top) and Plains Bison (bottom) during summer at Elk Island National Park. Photographs courtesy of Wes Olson.

Population Spatial Structure and Variability
In North America, fossil evidence indicates there was likely a single species, the Steppe Bison, Bison priscus, with variable species/sub-species designations throughout Beringia (Guthrie 1990). Bison moved south to occupy central North America when the ice sheets advanced (McDonald 1981). During the Late Pleistocene, bison underwent a gradual reduction in body size (Guthrie 1980; van Zyll de Jong 1993). During the Wisconsonian Glaciation (110,000-12,000 years B.P), Beringian and central populations became separated as the Laurentide continental ice sheet extended into western Canada during 20,000-13,000 years B.P. (Burns 1996, Wilson 1996). These two genetic lineages were recognizable in the fossil record, constituting a northern clade and a southern clade (Shapiro et al. 2004). At various times, depending on the extent of ice cover, the two clades oscillated between being isolated and intermingling. The extent of isolation of the clades is unknown (B. Shapiro, University of California Santa Cruz, pers. comm.). American Bison are currently believed to belong to the southern clade, distinct from apparently extinct Beringian bison, with a most recent common ancestor between 22,000 and 15,000 years B.P. (Shapiro et al. 2004). Ancient DNA patterns analyzed by Shapiro et al. (2004) and current patterns analyzed by Wilson et al. (2008) supported the hypothesis that extant American Bison descended solely from the southern clade, which existed south of the ice sheet before the Last Glacial Maximum. However, recent populations from southwestern Yukon and Alaska may have been from the northern clade (G. Zazula, Government of Yukon, pers. comm.). Until recently, it was believed that there was no evidence of genetic contribution by Beringian bison to the genome of modern American Bison; however, the emerging evidence is complex and suggests that perhaps both the northern and southern clades are descendant, and genetically indistinguishable, from Steppe Bison (B. Shapiro, University of California Santa Cruz, pers. comm.; see Name and Classification).
The two modern North American forms (Plains Bison and Wood Bison) diverged as geographic variants about 5,000 years ago (van Zyll de Jong 1986), with the Wood Bison being the most recent wild form (Stephenson et al. 2001). The validity of subspecies designations for American Bison has been controversial (Geist 1991, Cronin et al. 2013) and arguments remain inconclusive, largely as a matter of the recent divergence of these two forms, the lack of reproductive isolation, and the complicating influence of the significant population bottlenecks and founder events in the past 150 years (see below).
The genomes of extant Wood and Plains Bison became entwined due to the translocation of Plains Bison into the range of the remnant Wood Bison population in the 1920s (see History of Decline and Early Restoration of American Bison in Canada). Despite mixing, Wood Bison in Wood Buffalo National Park and descendant populations are morphologically distinct from Plains Bison (van Zyll de Jong et al. 1995), and ATK suggests the Ronald Lake animals to be pure Wood Bison (Athabasca Chipewyn First Nation 2010). Furthermore, distinctive morphological traits for Plains and Wood Bison are independent of local environmental conditions (van Zyll de Jong et al. 1995). Therefore they are not ecotypic variants as suggested by Geist (1991). A more detailed review of the results of morphological studies is provided by Wilson and Zittlau (2004).
Polziehn et al. (1996) sampled maternally inherited, mitochondrial DNA (mtDNA) from nine bison populations, including Wood Buffalo National Park (WBNP) and Elk Island National Park (EINP) Wood and Plains Bison, and six other Plains Bison populations. Of 11 haplotypes identified, four were unique to Wood Bison and four others were shared between Wood and Plains Bison, with the WBNP population exhibiting the most variability. The presence of unique haplotypes in Wood Bison potentially supports the inference that Wood Bison and Plains Bison were diverging owing to geographic isolation prior to the release of Plains Bison into WBNP (Wilson and Zittlau 2004). In a recent study, Douglas et al. (2011) sequenced the entire mtDNA genome of 43 American Bison, including two Wood Bison from EINP. Two unique haplotypes were found in the Wood Bison but they did not group together among the 16 bison mtDNA haplotypes identified. The authors suggested two explanations: either Wood Bison were never a genetically distinct form, or the unique mtDNA sequences were derived from the introduction of Plains Bison into Wood Bison subpopulations in the 1920s. An alternative hypothesis could also be that the two unique haplotypes were present in Wood Bison prior to introduction of Plains Bison.
Bork et al. (1991) compared restriction fragment length polymorphisms (RFLPs) for four nuclear DNA genes in samples from Elk Island National Park Wood and Plains Bison. Two of 28 fragments were polymorphic and their frequencies were distinctly different between Wood and Plains Bison. The results were consistent with recent geographic isolation of the two forms as suggested by van Zyll de Jong (1986), who came to the same conclusion based on spatial discontinuity in skeletal morphology between the two forms. In another study of neutral nuclear DNA, Wilson and Strobeck (1999) examined relationships between 11 bison populations using microsatellite markers. Although all populations were unique, the authors found genetic distances between Wood and Plains Bison populations were larger than between populations within subspecies. Samples from Wood Buffalo National Park, the descendant Mackenzie Bison and Elk Island National Park Wood Bison subpopulations were genetically similar, leading the authors to conclude that Wood Bison and Plains Bison are functioning as distinct entities. Furthermore, of the 370 individual bison examined, 98.6% were assigned to their a priori defined respective subspecies, further suggesting that extant Wood and Plains Bison nuclear genomes are distinctive. In subsequent research, using the same methods, only 4 of 258 (1.55%) bison sampled from the Slave River Lowlands (a subunit of the Greater Wood Buffalo National Park meta-population) were assigned to Plains Bison (Wilson 2001). Single Bison have recently been studied with single nucleotide polymorphism (SNP) microarrays designed for the bovine genome (e.g., Decker et al. 2009, Douglas et al. 2011). Using the Illumina Bovine SNP50 chip array, Pertoldi et al. (2010) found significant differentiation between Plains and Wood Bison.
Designatable Units
Wood and Plains Bison satisfy the criteria for discrete and evolutionarily significant designatable units (DUs; COSEWIC 2011b), whether or not they are classified as subspecies or geographic variants. Specifically:
- There is evidence of genetic distinctiveness between populations of Wood and Plains Bison including inherited traits (skeletal and external morphology), and genetic markers (see Population Spatial Structure and Variability).
- A natural disjunction existed between the original ranges of Wood and Plains Bison, such that movement between separated regions was limited. Currently, there is potential for contact between extant free-ranging Wood Bison and Plains Bison populations in northeastern British Columbia where free-ranging subpopulations of both occupy separate ranges, and the escape of captive commercial Plains Bison could lead to mixing with range of free-ranging Wood Bison, but this occurs outside the original range for Plains Bison. Management policies limit the potential for movement between free-ranging Plains and Wood Bison subpopulations in northeastern British Columbia (Harper et al. 2000), and provincial agricultural regulations may mitigate the risk of escapes from captive commercial herds.
- Wood and Plains Bison populations originally occupied different eco-geographic regions. Different environmental conditions in the range of the Plains Bison (prairies and parklands) compared with the Boreal and Northern Mountain range of the Wood Bison (meadows in a matrix of boreal forest) likely gave rise to different local adaptations (heritable morphology, and behaviour) that continue to distinguish the two forms despite the translocation of a large number of Plains Bison into Wood Buffalo National Park during the 1920s.
Despite the evident recent divergence and close phylogenic relationship of Wood Bison and Plains Bison determined through mtDNA and nuclear DNA analysis, there is agreement among many authors that sufficient molecular distinctions exist between them to warrant separate conservation emphasis (e.g. Wilson and Strobeck 1999, Douglas et al. 2011; see Population Spatial Structure and Variability). This phylogenetic division has also been corroborated at a phenotypical level, by ATK holders, notably in the Athabasca Chipewyn traditional area (Athabasca Chipewyn First Nation 2010).
In spite of the continuing debate about whether or not these entities are subspecies (e.g., Cronin et al. 2013), morphological and ecological distinctions between the two variants, both historically (van Zyll de Jong 1986) and following mixing of Wood and Plains Bison in Wood Buffalo National Park (van Zyll de Jong et al. 1995), also support the conclusion of extant geographically discrete units.
Special Significance
Ecological significance
American Bison are the largest extant herbivore in North America. The original range of this species extended from the arid grasslands of northern Mexico (List et al. 2007) to the boreal meadow systems of interior Alaska (Stephenson et al. 2001; Figure 4). Bison have been described as a foundation (Freese et al. 2007), keystone species (Knopf, 1996), or landscape transformer (Centre for Indigenous Environmental Resources Inc., 2008). Bison is an interactive species, meaning its ‘‘virtual or effective absence leads to significant changes in some features of its ecosystem(s)’’ (Soule et al., 2003: 1239). In the Canadian grasslands, bison were the dominant herbivore prior to European settlement (Kay et al. 1999, Bush and Rowell 2000, White et al. 2001, Epp and Dyck 2002, Langemann 2004).
Figure 4. Historical (pre-settlement) distribution of Wood Bison and Plains Bison in North America. Modified from Gates et al. (2010). Polygons courtesy of Keith Aune, Wildlife Conservation Society.
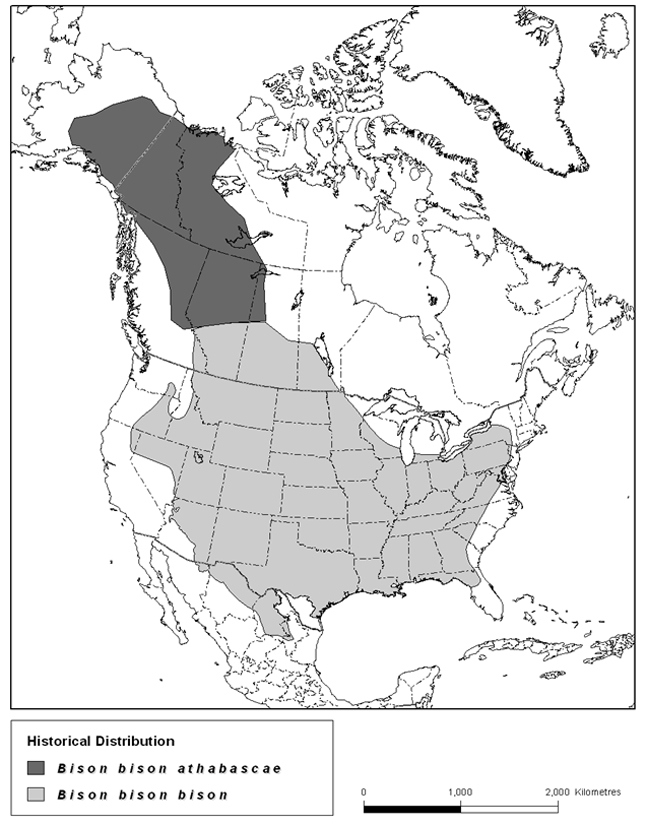
Long description for Figure 4
Map of North America showing the pre-settlement distribution (polygons) of the Wood Bison and Plains Bison, which extended from northern Mexico to interior Alaska. Further details can be found in the preceding/next paragraph(s).
Bison grazing in combination with fire creates heterogeneous vegetation patches (Knapp et al. 1999, Coppedge and Shaw 1998, Fuhlendorf et al. 2009) that vary in forage quality and maturity (Coppock et al. 1983a,b, Krueger 1986). Bison grazing modifies plant species composition (Fahnestock and Knapp 1993, Damhoureyeh and Hartnett 1997) and horizontal and vertical structure (Coppock and Detling 1986, Virchow and Hyngstrom 2002) through alteration of light penetration and soil moisture, and nutrient cycling (Day and Detling 1990, Frank 1998, 2000, Frank and McNaughton 1993, Frank and Evans 1997). Wallows are small depressions in the ground created when Bison root in the ground and roll in the exposed soil (Polley and Collins 1984). Vegetation composition and structure, soil texture, soil moisture, phosphorus availability and pH in wallows differ from the surrounding undisturbed sites, contributing to the heterogeneity of grasslands (Polley and Collins 1984, Hartnett et al. 1997), favouring establishment of colonizing plant species. Wallowing and other disturbance of sand dunes by Bison is thought to have contributed to a geomorphological cascade benefiting other species associated with active sand dunes, including several species that are currently at risk (Fox et al. 2012).
Bison facilitate the dispersal of seeds when they become temporarily attached to the hair coat (Berthoud 1892; Rosas et al. 2008) or pass intact through the digestive tract (Gokbulak 2002). Bison grazing and soil disturbance influence patterns of abundance and distribution of a number of other species (Bragg 1940, England and deVoss 1969, Knapp et al. 1999, Rosas et al. 2005, Jonas and Joern 2007). Bison are an important prey species for Wolves (Canis lupus) when other more susceptible species are relatively rare (Carbyn and Trottier 1987, 1988, Larter et al. 1994, Joly and Messier 2004a,b, Smith et al. 2000, Gates et al. 2005), and wolf-killed bison may be used by a host of scavengers (see Interspecific Interactions).
Cultural and economic significance
The American Bison has a history rich in paleontology, paleoecology, and archaeology; perhaps no other species has so profoundly shaped early cultures and emerging politics of a continent. Humans and bison have a shared history dating back to the first human occupants of Beringia. Literature on human-bison interactions is encyclopedic in scope (see reviews by Frison [1991], Isenberg [2000], Stephenson et al. [2001] and Potter et al. [2010]).
Bison were an important faunal element in the subsistence economy of the first western hemisphere colonists (Potter et al. 2010). By 10,000 B.P., human hunters were killing bison with projectiles launched with atlatls and later bows. They improved hunting efficiency by driving small groups of bison into natural and artificial traps such as arroyos, mud or sand, and “pounds” of timber and brush to corner a dozen or more bison (Reeves 1978, Frison 1991, Barsh and Marlor 2003).
Throughout the Holocene, until about 150 years ago, bison were a highly profitable and preferred food source for many Native American groups. This was especially so on the Great Plains where they provided a staple food, clothing, shelter, and material for tools (Geist 1996, Roe 1970). The bison was a central aspect of oral tradition, rituals, dances, and ceremonies of native peoples of the Plains (Wissler 1927), and it remains symbolically important in the cultural traditions of many First Nations and Metis people. In a brief, well-documented span of about 40 years Aboriginal communal Plains Bison hunting came to an end in Canada in 1879 once bison became scarce (Foster 1992, Flores 1996, Isenberg 2000, TM-SPHA 2008).
Contemporary Aboriginal conceptions about bison are now largely historical, rather than acquired and tested through direct experience (Barsh and Marlor 2003, COSEWIC 2012). Although there has been a renewal of interest in propagating bison by some Aboriginal groups for sustaining or reinforcing spiritual and cultural connections, the commercial potential of bison as a production commodity is most important for others (Potter et al. 2010). The pragmatic use of bison for food, the relationship between local control over food production and land, food security, tribal sovereignty, and decreasing reliance on outside sources for food and commodities are emerging as other important reasons for propagating bison (Potter et al. 2010, COSEWIC 2012).
Conservation and recovery of wild bison is a concern to several Aboriginal communities in Canada (COSEWIC 2012). Governments of the Yukon, British Columbia, and the Northwest Territories have addressed complaints from some communities about bison entering settlements, vehicle collisions, disturbance to traps and trap lines, trampling of berry patches, competition between bison and valued food species and with horses grazed in bison habitat, and fears about encountering bison while on the land. For example, a First Nations government in British Columbia is demanding the removal of a one subpopulation that ranges within their traditional territory for many of these reasons (COSEWIC 2012). On the other hand, in southwest Yukon, initial concerns described in the 1998 Wood Bison management plan (Yukon Department of Renewable Resources 1998) may have decreased somewhat with the initiation of bison hunting. Bison have become an increasingly valued resource and tolerance for them has increased somewhat over time.
Bison provide a number of community benefits. They are hunted as wildlife in Saskatchewan, Alberta, British Columbia, NWT and the Yukon. Subsistence use of bison in the NWT helps reduce dependence on store-bought foods and contributes to a healthy diet (NWT Environment and Natural Resources 2010). Private businesses provide guiding and outfitting services for hunters in Alberta, British Columbia, the Northwest Territories and the Yukon. Nature tours to see bison are offered by businesses in NWT and Yukon. Local handicrafts featuring, or using material from, bison have developed in some communities.
In the early 1980s, Canada’s Elk Island National Park began regular sales of surplus bison to agricultural producers. Surplus animals were also sold by some parks in the United States beginning at about the same time. Private ownership and production of bison as a market commodity have developed dramatically since 1980 (Renecker et al. 1989, Freese et al. 2007).
Distribution
Original Distribution
The original range of American Bison during the late Holocene extended from the desert grasslands of northern Mexico (List et al. 2007) to interior Alaska (Stephenson et al. 2001), and from New England to Florida in the east, then westward to Nevada and parts of the Great Basin (Figure 4). Sanderson et al. (2008) estimated the area of original North American range of American Bison (Figure 4) as 8.96 million km2, a larger area than any other indigenous ungulate in North America. Even with the inclusion of commercial herds, American Bison currently occupy <1% of their historical range (ca. 1500; Sanderson et al. 2008).
Wood Bison
Physical remains and Aboriginal knowledge from the late Holocene until about 200 years ago indicate that Wood Bison originally occupied a small area of western Saskatchewan, much of northern Alberta, northeastern British Columbia, the western Northwest Territories, Yukon, and much of Alaska (van Zyll de Jong 1986, Stephenson et al. 2001, Farnell et al. 2004, Heffner 2008, Figure 4). The area originally occupied by Wood Bison calculated from mapped data presented by Sanderson et al. (2008) was 2,524,000 km2, with approximately 79% of the area in Canada (1,993,960 km2) and 21% in Alaska.
Plains Bison
The geographic division between the original range of Wood Bison and Plains Bison aligns approximately with the interface between the Boreal Forest and Parklands in Alberta (Figure 4). This demarcation is derived from the seminal work of van Zyll de Jong (1986) who based his conclusions on craniometric (skull measurements) of recent and sub-fossil skeletal material, identifying a discontinuity in size and morphology of bison specimens found north and south of this line. Plains Bison moved into the Parkland during the winter months (Campbell et al. 1994); hence, there was the potential for Wood Bison and Plains Bison to be sympatric in this area. However, Plains Bison moved south in spring before the rut, thereby reducing the chance of interbreeding (van Zyll de Jong et al.1995).
The original range of the Plains Bison extended from the transition between Parklands and Boreal Forest in the Prairie Provinces south to the arid shrub steppe and grasslands of northern Mexico (Figure 4). They ranged from the New England states to Florida in the east, then to the continental divide in the west. Plains Bison were most abundant in the grasslands of the Great Plains (Isenberg 2000). In Canada, the Plains Bison was abundant throughout the grasslands and parklands of the Prairie Provinces, where most undertook seasonal movements from summer ranges in the grasslands to fall and winter ranges in the parklands and intermountain valleys in Alberta (Campbell et al. 1994, White et al. 2001), while smaller numbers may have been year-round residents (Epp 1988). The global area originally occupied by Plains Bison calculated from mapped data presented by Sanderson et al. (2008) was 6,965,000 km2. About 86% of their original range was in the United States, with the remainder in Canada (11%; 752,363 km2) and Mexico (3%).
History of Decline and Early Restoration of American Bison in Canada
Plains Bison
Historical and archaeological records show that Plains Bison thrived in their greatest abundance in the grasslands of the Great Plains (Malainey and Sherriff 1996, Shaw and Lee 1997). Explorers, settlers, and Euroamerican hunters described enormous herds of Plains Bison, with population estimates ranging from 15 to 100 million (Dary 1974; Shaw 1995). In the 1890s, naturalist Ernest Thompson Seton offered a widely accepted pre-human settlement estimate for American Bison as 60 million (Dary 1974; McHugh 1972; Roe 1970). Shaw (1995) revised the estimate down to 30 million. Wild Plains Bison were rapidly depleted between 1840 and 1880 in Canada and the United States, owing primarily to market hunting to serve demands for meat and hides (Hornaday, 1889; Isenberg, 2000). In Canada, extirpation followed a parallel course driven by commercial demand for hides and subsistence needs of First Nations and Metis peoples (Roe 1970). Extirpation was abetted by ineffective conservation legislation and enforcement (Hewitt 1919) and occurred swiftly in Canada. Hornaday (1889) estimated eight Plains Bison remained in Canada in 1888. (Coder 1975) and Roe (1970) suggested they were extirpated from the country within the next two years.
The history of salvage and translocation of a few surviving Plains Bison is important for understanding potential bottleneck and founder effects on the genetic diversity of Plains Bison and the foundation of wild Plains Bison populations in Canada (Figure 5). With the exception of one wild Plains Bison population that survived in Yellowstone National Park (23 individuals; Meagher 1973), the foundation stock for all other subpopulations in existence today originated from approximately 116 bison captured by a handful of private citizens. Five herds played a key role in the establishment of all current North American Plains Bison subpopulations (Coder 1975). The 81 founding ancestors of all Plains Bison in the public domain in Canada were captured in three locations (Dary 1974, Coder 1975, Ogilvie 1979, Neufeld 1992). The history of these animals in relation to the establishment of Canadian subpopulations is discussed in Gates (in press).
Figure 5. Decade interval timeline of Plains Bison captures and translocations for lineages used to populate Canadian Parks spanning 1860 to 1930. Sources: Coder 1975, Dary 1974, Ogilvie 1979, and Lothian 1981.
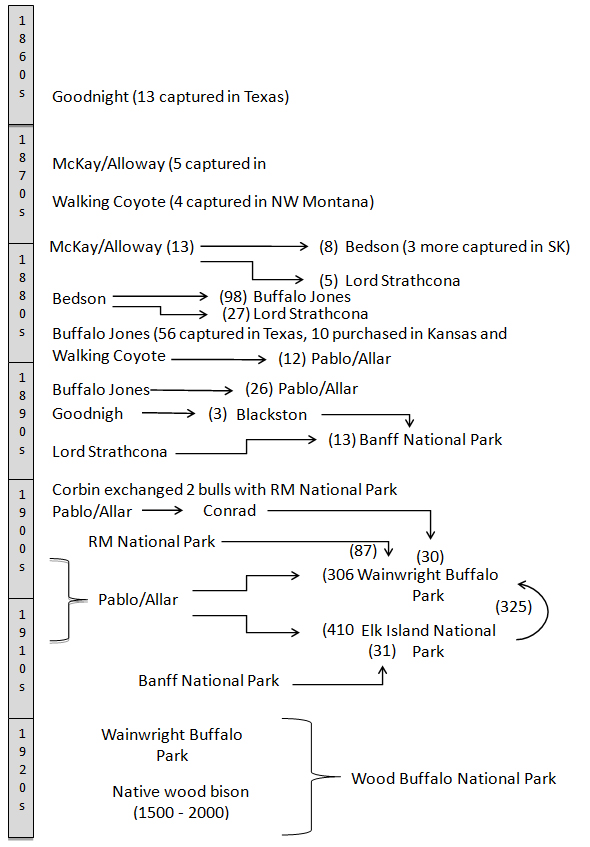
Long description for Figure 5
Diagram showing decade-interval timeline of Plains Bison captures and translocations for lineages used to populate Canadian parks from 1860 to 1930. Further details can be found in the preceding/next paragraph(s).
Wood Bison
Wood Bison were far less abundant with a more fragmented distribution than Plains Bison owing to the limited availability and configuration of their preferred grazing habitats in the vast boreal forest matrix (Gates et al. 1992, Larter et al. 2000, Gates et al. 2001a. Soper (1941) offered a highly speculative estimate of the number of Wood Bison population in 1800 as 168,000, based on extrapolating the number and distribution of bison in Wood Buffalo National Park during the 1930s. The estimate did not account for regional variability in habitat availability and was applied to a more limited range than documented later by Stephenson et al. (2001). Therefore, Soper (1941) may have underestimated the original population of Wood Bison.
Wood Bison were widely distributed in Alaska and adjacent Yukon until late in the Holocene. Information from oral narratives indicated that Wood Bison persisted in small numbers in Alaska and the Yukon into the early years of the 20th century (Lotenberg 1996, Stephenson et al. 2001).
Heavy exploitation following the advent of the fur trade played a major role in the decline of Wood Bison elsewhere in Canada (Gates et al. 1992); habitat changes (loss of boreal grasslands), and severe winters were also contributing factors (Soper 1941, Stephenson et al. 2001). Wood Bison were nearly eliminated from their range during the late 1800s, coinciding with the rapid decline of Plains Bison (Raup 1933). In 1888, the total population was estimated at between 500 and 600 animals (Schultz 1888). By 1891, only 300 Wood Bison remained in the wilderness between Great Slave Lake and the Peace-Athabasca Delta (Ogilvie 1893). The population reached an estimated low of approximately 250 during 1896-1900 (Soper 1941). After 1900, Wood Bison were occasionally observed across their range in Canada, but numbers were insignificant except in the Slave River lowlands and the Peace-Athabasca region.
One of the last reports of bison in northeastern British Columbia was in 1906 (MacGregor 1952, cited by Gates et al. 1992). The last recorded bison in the range of Wood Bison in British Columbia was shot at Lower Post in 1939 (Clarke 1945, cited by Lotenberg 1996). A small herd of bison was seen at Grainger Creek near Nahanni Butte, NWT in 1919 (Gates et al. 1992). This was the last recorded sighting of bison in the NWT outside the Greater Wood Buffalo National Park area.
With enhanced protection, the Wood Bison population increased slowly to approximately 500 by 1914 (Banfield and Novakowski 1960). Wood Buffalo National Park was established in 1922 in an attempt to save the Wood Bison from extinction and to protect its habitat (Soper 1941; Lothian 1981). The total number of Wood Bison at the time was estimated at 1,500 and 2,000 (Siebert 1925; Soper 1941).
Between 1925 and 1928, 6,673 young Plains Bison--‘surplus’ animals from the Wainwright Buffalo Park Plains Bison herd in west central Alberta--were transported to Wood Buffalo National Park, where resident Wood Bison numbered approximately 1,500 to 2,000 individuals in 1922 (Lothian 1981). Although an unknown number of individuals survived the transfer (Bradley and Wilmhurst 2005), this introduction of Plains Bison into Wood Bison habitat resulted in interbreeding between these subspecies (see Population Spatial Structure and Variability) and the resulting admixed population was the foundation stock for all current day Wood Bison. Some ATK holders believe that the Ronald Lake and the Firebag River animals are pure Wood Bison and not part of the mixture (Athabasca Chipewyn First Nation 2010).
The introduction of Plains Bison into Wood Bison range was heavily criticized by the American Society of Mammalogists (Howell 1925) and by individual biologists, who believed hybridization with indigenous Wood Bison would result in the loss of Wood Bison and that the population would become infected with tuberculosis known to be present in the Wainwright herd (Harper 1925; Saunders 1925). Following the introduction of Plains Bison the number of bison in Wood Buffalo National Park increased to an estimated 12,000 by 1934 (Soper 1941). Raup (1933) speculated that the wood buffalo “as a race” was rapidly disappearing, but suggested an intact northern herd still existed. In 1959, five specimens were collected from a herd of about 200 animals near the Nyarling River and were determined by Banfield and Novakowski (1960) to be morphologically representative of Wood Bison. The taxonomic affiliation of bison in other parts of the region was not studied until the 1990s (van Zyll de Jong et al. 1995, Wilson and Strobeck 1999).
In 1963 and 1965, bison were captured at Needle Lake/Nyarling River area in northwestern Wood Buffalo National Park to establish a captive-breeding herd from a genetic source thought to be free of Plains Bison introgression. This assumption was later proven false based on genetic evidence (Wilson and Strobeck 1999). Bison obtained from these two captures are ancestral to all Wood Bison subpopulations outside the Greater Wood Buffalo National Park area.
Current Distribution
In North America today, < 20,000 (~4%) of approximately 500,000 Plains Bison are in subpopulation units managed principally for conservation purposes (Boyd 2003). The number of individuals in conservation has changed little since 1930 in spite of an increase in the number of subpopulations over time (Freese et al. 2007). Gates and Ellison (2010) enumerated 62 “conservation herds” on the continent, 87% of which are located within original Plains Bison range, including some display herds. There are 49 such herds in 19 U.S. states, and one in the state of Chihuahua, Mexico. Most are small (< 400 animals), with the largest in the U.S. residing in the Greater Yellowstone Area. Most extant Wood Bison occur in Canada (see below), with only one captive herd in Alaska and two captive herds in the Sakha Republic (Yakutia), Russia.
This assessment considers five and nine Canadian subpopulations that are “wild by nature” (see Population Units to Be Assessed by COSEWIC) within the Plains Bison and Wood Bison designatable units, respectively (see Designatable Units).
Plains Bison
In Canada, there are five subpopulations of Plains Bison in Canada that are considered “wild by nature” (Figure 6). Details regarding the establishment and origin of founder stock are presented in Table 1.
Figure 6. Approximate original (pre-settlement) range of Plains Bison in Canada (dashed red line) and current distribution of five subpopulations considered in this assessment. Striped polygon is the original distribution of Wood Bison. Note that McCusker River range (3) occurs at the periphery of Wood Bison, close to the edge of the original range of Plains Bison.
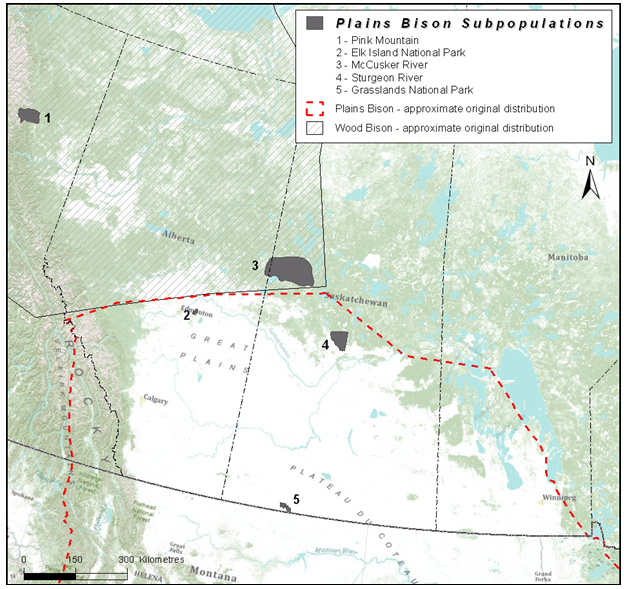
Long description for Figure 6
Map showing approximate original (pre-settlement) range of Plains Bison in Canada (outlined) and current distribution of five subpopulations considered in this assessment (shaded polygons; Pink Mountain, Elk Island National Park, McCusker River, Sturgeon River, and Grasslands National Park) . The map also shows the original distribution of Wood Bison (hatched polygon). Further details can be found in the preceding/next paragraph(s).
| DU | Subpopulation | Source | Year | Number |
|---|---|---|---|---|
| Plains | Sturgeon River, SK | EINP, then Big River, SK | 1969 | 10-22 |
| Plains | McCusker River, SK | EINP then Big River, SK | 1969 | 17 |
| Plains | Pink Mountain, BC | EINP | 1971 | 48 |
| Plains | Grasslands Nat. Park, SK | EINP | 2005 | 71 |
| Wood | Mackenzie, NWT | Wood Buffalo National Park (WBNP) | 1963 | 18 |
| Wood | Elk Island National Park (EINP), AB | WBNP | 1965 | 11 |
| Wood | Elk Island National Park (EINP), AB | WBNP | 1968 | 2 |
| Wood | Nahanni, NWT | EINP | 1981 | 28 |
| Wood | Nahanni, NWT | Moose Jaw Wild Animal Park, SK (originated from EINP) | 1989 | 12 |
| Wood | Nahanni, NWT | EINP | 1998 | 59 |
| Wood | Hay-Zama, AB | EINP | 1984 | 29 |
| Wood | Chitek Lake, MB | Waterhen Wood Ranch (originated from EINP) | 1991 | 13 |
| Wood | Chitek Lake, MB | Waterhen Wood Ranch | 1993 | 9 |
| Wood | Aishihik Lake, YT | EINP | 1986 | 34 |
| Wood | Aishihik Lake, YT | Moose Jaw Wild Animal Park | 1989 | 10 |
| Wood | Aishihik Lake, YT | EINP | 1990 | 50 |
| Wood | Aishihik Lake, YT | EINP & Metro Toronto Zoo | 1992 | 48 |
| Wood | Nordquist, BC | EINP | 1995 | 49 |
| Wood | Nordquist, BC | EINP | 1999 | 19 |
| Wood | Etthithun Lake, BC | EINP | 2000 | 24 |
The EO of Plains Bison is approximately 397,338 km2, encompassing extensive areas of unsuitable and unoccupied habitat. The IAO, or the area within the EO that is occupied by Plains Bison, is about 14,764 km2, based on a 2x2 grid, representing about 2% of the original Canadian range. The EO and IAO have increased since the last assessment because of the addition of a new fenced subpopulation in southern Saskatchewan managed by Parks Canada (Grassland National Park subpopulation - 2005). Three of the five Plains Bison subpopulations qualifying as components of the DU (see Designatable Units) are found in COSEWIC’s Boreal ecological area, while two are in the Prairie ecological area.
Wood Bison
There are nine subpopulations of Wood Bison in Canada that are considered “wild by nature” (Figure 7). With all extant Wood Bison herds outside Canada (in Alaska and Russia) being captive, 100% of the global wild Wood Bison population resides in Canada. Details regarding the establishment and origin of founder stock are presented in Table 1.
Figure 7. Approximate original (pre-settlement) range of Wood Bison in Canada (dashed red line) and current distribution of 9 subpopulations considered in this assessment. Striped polygon approximates the original distribution of Plains Bison.
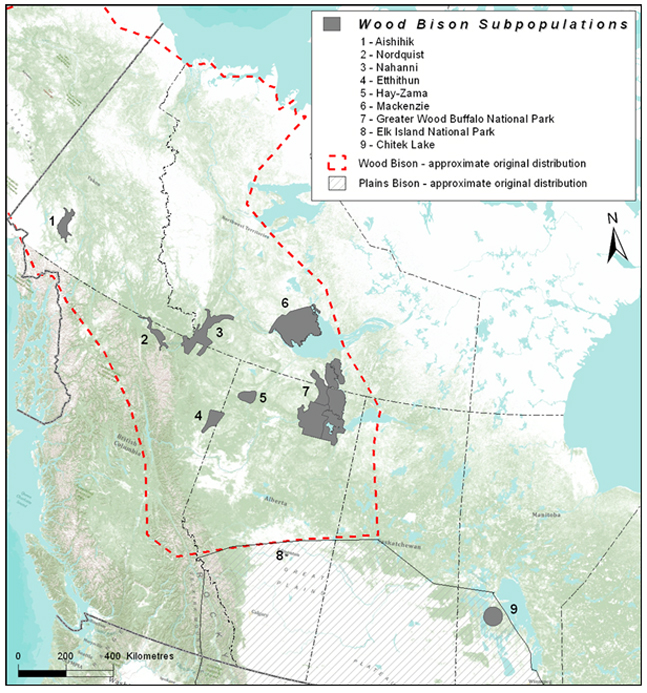
Long description for Figure 7
Map showing approximate original (pre-settlement) range of Wood Bison in Canada (outlined) and current distribution of nine subpopulations considered in this assessment (shaded polygons; Aishihik, Nordquist, Nahanni, Etthithun, Hay-Zama, Mackenzie, Greater Wood Buffalo National Park, Elk Island National Park, Chitek Lake). The map also shows the original distribution of Plains Bison (hatched polygon). Further details can be found in the preceding/next paragraph(s).
Bison from Wood Buffalo National Park are ancestral to all extant subpopulations (Table 1). Wood Bison were historically distributed across COSEWIC’s Boreal and Northern Mountains ecological regions, and were probably more numerous in the former (Soper 1941). Through concerted conservation effort, representative Wood Bison subpopulations are extant in both of the historical COSEWIC ecological areas where they formerly occurred, with most subpopulations in the Boreal ecological area. Bison control areas aimed at reducing the spread of brucellosis and tuberculosis (Harper et al. 2000, Gates et al. 2001a, Nishi 2010, Government of Alberta 2011) from the Greater Wood Buffalo National Park meta-population, range occupation by extra-limital Plains Bison (Pink Mountain subpopulation; Harper et al. 2000), and low “wildlife stakeholder acceptance capacity” (Decker and Purdy 1988; Carpenter et al. 2000) severely limit potential range expansion by existing subpopulations (e.g., Aishihik subpopulation [Government of Yukon 2012], or the establishment of new subpopulations within the original range [Gates et al. 2001a]). All subpopulations are or have been subject to management actions to discourage range expansion into unwanted areas. In effect, little of the remaining 94.7% of the original range is currently available for Wood Bison restoration given these limitations.
The EO of Wood Bison is about 1,144,329 km2, encompassing a large area outside of the original range and currently unoccupied and unsuitable habitat. The IAO (occupied habitat) is considerably smaller at about 120,528 km2, based on a 2x2 grid (J. Wu, Environment Canada, pers. comm.), representing about 6% of the original range. The EO increased since the last assessment because of the addition of a new free-ranging subpopulation in northeastern British Columbia (Etthithun subpopulation) in 2002, established as part of the recovery effort for Wood Bison (Thiessen 2010).
Search Effort
American Bison rarely go unnoticed on the landscape, and hence their distribution is well defined. Their large body size, propensity to form large groups, particularly during calving and the rut, and preference for open habitats makes them conspicuous. Periodic population surveys (see Sampling Effort and Methods), combined with use of the land by Aboriginal People, other residents and travellers, and aircraft traffic, offer ample opportunity to incidentally locate any unknown subpopulations. The likelihood of unknown subpopulations of free-ranging American Bison in Canada is negligible.
Population Units to be Assessed by COSEWIC
All extant Wood Bison and Plains Bison occur in population units subject to various past or present manipulations by humans. Hence, some discussion is required to define which units (subpopulations) are components of each DU in the quantitative assessment to determine status (COSEWIC 2010a). Both SARA and COSEWIC define “a wildlife species” as being “wild by nature” (COSEWIC 2010b). This determination is based on whether populations included in the assessment 1) function both ecologically and evolutionarily in a manner that maintains the wild nature of bison in the long term and 2) are genetically or geographically distinct from populations managed for purposes other than conservation (COSEWIC 2010b).
1) “Wild by Nature” Bison
Manipulations contemplated in the COSEWIC Guidelines on Manipulated Populations (COSEWIC 2010a) include introduced/re-introduced, hybrid, supplemented and captive populations. The only native Wood Bison subpopulation that survived in the wild through the early 1900s (Slave River Lowlands within Wood Buffalo National Park) was subjected to a massive translocation of Plains Bison. Unlike Wood Bison, Plains Bison were extirpated from the wild in Canada, but the Elk Island National Park subpopulation has served as source of stock for establishing or augmenting numerous other subpopulations.
The meaning of the concept “wild by nature” is germane to assessing bison subpopulations to be included in Plains and Wood Bison DUs. To be useful for biodiversity conservation the definition ought to serve the dual objectives of maintaining patterns of adaptive variation and formational processes existing in nature. Moritz (2002) cast the problem of conservation in terms of protecting both ‘pattern’ and ‘process’. Pattern can be equated to local adaptations and geographic variations, and its conservation can be accomplished by the identification and protection of groups of populations, at least over the short term. In contrast, protecting evolutionary processes requires maintaining what Moritz terms the “context” for natural selection to operate.
Species, subspecies, varieties or geographically or genetically distinct subpopulations of American Bison that are “wild by nature” reflect patterns of local adaptation and geographic variation arising from species-formational processes, and occur in locations where conditions support natural selection. The central questions for determining if a bison subpopulation is functionally “wild by nature” involves the degree to which natural selection has been and continues to be the primary formational process, and by corollary, the extent to which humans actively manage breeding, survival, disease, behaviour, and movements. Some indicators have been used to evaluate the presence of natural selection in bison subpopulations (Sanderson et al., 2008; Gates and Ellison 2010; Gross et al. 2010). These include the presence of effective predators (predation), mating competition among mature males, population size and structure, the absence of supplemental feeding and occurrence of resource limitation, differential fecundity and survival, and the ability of bison to move in response to spatio-temporal resource gradients. Exposure to pathogens is also a major factor influencing animal fitness, diversity, and evolution (Altizer et al. 2003).
Privately owned commercial bison herds
Plains Bison occur in the greatest abundance and widest distribution in private captive herds managed primarily for commercial propagation (Freese et al. 2007, Gates and Ellison 2010). Private interest in captive rearing of bison occurred at a low level in Canada until the 1970s when alternative livestock production was promoted (Renecker et al. 1989). Beginning in the 1970s in the United States, hundreds of surplus park animals were sold annually to ranchers, and the number of bison in commercial holdings soon outnumbered those in conservation herds (Freese et al.2007). By 1985 there were about 5,000 bison on farms in Canada. The farmed bison population peaked in 2006 at 195,728, and then declined to 124,848 in 2011 from 1,211 farms (last census), with most in Alberta (46%), Saskatchewan (32%), and Manitoba (11%) (Gates in press). In 2008 in the United States and Canada, there were about 400,000 bison on ~4000 farms (Freese et al. 2007; Gates and Ellison 2010). People involved in the commercial bison industry own approximately 97.4% of all bison in Canada.
COSEWIC generally does not consider as part of the “wildlife species” being assessed “any manipulated populations established for purposes other than species conservation (for example, those established for commercial purposes), provided the population is geographically or genetically distinct from the wildlife species under assessment, and there is no intention that the population contribute to the wild population” (COSEWIC 2010a). Some Aboriginal groups do run bison herds for species conservation and they partly fund and operate it through a commercial enterprise (COSEWIC 2012). Similar to the process of domestication (Trut et al. 2009), management procedures for commercial populations of bison create substantially different selective pressures compared to what they would experience in the wild. Therefore, in keeping with both the concept of “wild by nature” (as defined for bison in this report) and the COSEWIC guideline excluding any manipulated populations established for purposes other than species conservation, privately owned commercial herds were not included in this assessment as components of Wood Bison or Plains Bison DUs.
Subpopulations managed specifically for conservation
A Plains or Wood Bison subpopulation that is wild by nature is one where the management policy or plan fosters natural selection as an evolutionary process, in addition to conserving the ecological roles of bison. Consideration was given to which were functioning as wild subpopulations and to factors that may limit that function, including:
- Population structure. Competition for mating opportunities is an important selective force contributing to fitness in contrast to control of breeding (artificial selection) under domestication. Maintaining a ratio of adult males to reproductively mature females typical of large non-manipulated populations, and hence allowing mating competition to occur as a natural selection process, is an important criterion for assessing the wild nature of a bison subpopulation. If culling occurs, it is with the intent to emulate natural mortality (highest in juveniles), and old animals are allowed to die in situ.
- Supplemental feeding and disease treatment: Supplemental feeding or treatment of disease can increase fertility and survival and thus moderate the influence of natural selection on a population and on individual fitness. Intentional or inadvertent selection for tameness and reduced flight responses leads to lack of predator recognition or reduced ability to flee (Menzel and Beck 2000). Wild subpopulations are not supplementally fed to enhance productivity or survival. With pathogens having the potential to act as significant drivers of fitness, population dynamics, and genetic diversity, management practices may reduce or eliminate the natural pathogen fauna while at the same time causing exposure to introduced pathogens (from cattle and other domestic livestock) (Altizer et al. 2011). Wild subpopulations are those where any management interventions that occur as a result of introduced disease do not affect ecological or evolutionary processes.
- Movement and habitat heterogeneity: A wild subpopulation is free-ranging (i.e., not confined to small areas < 200 km2 [Sanderson et al. 2008] by a perimeter fence or other habitat barriers), such that densities are suitable for social interactions, ecological functions are maintained, and seasonal movements within and between ranges is unimpeded (Sanderson et al. 2008). Wild bison should be exposed to seasonal and spatial patterns of resource availability and be able to move in response to resource gradients (Gross et al. 2010).
- Predation: Natural predation (primarily by wolves in the case of bison) serves as a key selective force in the evolution of species (Dawkins and Krebs, 1979). The presence of predators is well-documented to influence behaviour and has implications for fitness, as well as lethal and demographic effects (Preisser and Bolnick, 2008; Creel and Christianson, 2008).
Plains Bison
The 2010 IUCN status report for American Bison (Gates and Ellison, 2010) identified 10 Plains Bison subpopulations in Canada of 62 in North America that are managed by public agencies or private organizations for conservation purposes (including education). Not all can be considered wild by nature, rather only those where bison can continue to evolve under conditions that support natural selection and continued evolution of the species. Four of the 10 “conservation herds” in Canada are educational display herds. These are small (ca. 10- 50 animals), intensively managed herds confined in fenced paddocks. They are maintained for historical and educational purposes and no attempt is made to manage them as wild populations. They therefore meet none of the criteria described above. These include: Riding Mountain National Park (Manitoba), Waterton Lakes National Park (Alberta), Buffalo Pound Provincial Park (Saskatchewan), and Canadian Forces Base-Wainwright (Alberta).
In 1996, the Nature Conservancy of Canada (NCC) acquired 53 km2 of land for the Old Man on His Back (OMB) Nature Prairie and Heritage Conservation Area, located in southwestern Saskatchewan in the mixed grasslands. NCC translocated 50 Plains Bison from Elk Island National Park in 2003. At present, OMB are highly managed within a population objective of 60 to 130 bison. The OMB population is not functioning as a wild population owing to significant limitations: there are no adult males and hence no mating competition, predators are absent, the herd is small (60 to 130) with no opportunity for growth, available range is small and fenced, water is provided in winter, supplemental feeding is provided depending on severity of winter conditions, and herd size is managed by removing juveniles and mature bulls (R. Grilz, Nature Conservancy of Canada, pers. comm.). It was therefore not included in the Plains Bison DU for assessment by COSEWIC.
Each of the remaining five Canadian “conservation herds” (sensu Gates and Ellison, 2010) of Plains Bison have most or all of the four wild attributes, and were deemed to be functioning as wild populations for the purposes of this assessment. From west to east:
Pink Mountain (British Columbia):
This introduced free-ranging subpopulation in northeastern British Columbia is extra-limital for Plains Bison and is located in the original range of Wood Bison. The Pink Mountain subpopulation was established in 1971 with 48 Plains Bison purchased from Elk Island National Park by a private rancher (COSEWIC 2004). Within a year of their translocation to the ranch the bison escaped captivity to become a free-ranging population. Population size and distribution are managed through regulated hunting. The herd ranges in a 3,200 km2 area in the upper Sikanni and Halfway River valleys; Rowe 2006).
In accordance with COSEWIC’s Guidelines for Manipulated Populations (COSEWIC 2010a), population units resulting from “benign extra-limital introductions” are to be included as part of the wildlife species being assessed only if there is no suitable habitat remaining within the natural range of the wildlife species in Canada. Options for restoration of Plains Bison within their original range are severely limited, both because of extensive habitat conversion and fragmentation and because socio-political conflicts mean that most lands are managed to exclude bison (see Habitat Trends). Moreover, as the most demographically robust, disease-free unit within the Plains Bison DU, the Pink Mountain subpopulation has a net positive impact on the conservation of Plains Bison and is therefore included in the assessment.
The Pink Mountain Plains Bison subpopulation is subject to a full range of natural ecological processes, including disease and wolf predation, and its movements are unrestricted by fences (D. Fraser, B.C. Ministry of Environment, pers. comm.). Salt blocks are used in winter to encourage bison to stay off the highway (C. Thiessen, B.C. Ministry of Natural Resource Operations, pers. comm.), but there is no supplemental feeding. Pink Mountain bison are functioning as a wild population, without limitations.
Elk Island National Park (Alberta):
In 1906, the federal government was petitioned by a local group of five businessmen to establish an elk sanctuary west of Edmonton. Elk Island National Park was granted federal park status in 1913. The park has played a central historical and contemporary role in the conservation of Plains Bison in North America, being the source of multiple established subpopulations.
The Elk Island National Park Plains Bison subpopulation is confined to the northern portion of the park in a 136 km2 fenced area of parkland habitat. Since the 1970s the herd has ranged from 400-1000 until around 2009, when the park started managing the adult population at about 400 through translocations and sales (Parks Canada Agency 2009). It does not receive supplemental feeding except for baiting during passive captures. Population size is mainly regulated by non-selective annual or semi-annual removal of animals, especially calves and yearlings. Although removal of individuals to control population increase might limit natural selection acting on this cohort, random selection of those removed should result in unbiased fitness or adaptive outcomes. Older animals are allowed to die naturally in the park. Occasionally a bison bull is removed if it is considered dangerous for park visitors. Wolves have recently been observed in the south (Wood Bison area) portion of the park and the adjacent Blackfoot Grazing Reserve and have since reproduced (Martha Allen, Parks Canada, pers. comm.). Habitat management is accomplished with prescribed burning and herbicide application to manage invasive plants. There are no other manipulations. Elk Island Plains Bison are functioning as a wild population, with a limitation that they are confined to an area of < 200 km2 by fencing.
McCusker River (Saskatchewan):
This subpopulation originated from 17 Elk Island National Park bison that settled in the Big River Community Pasture west of Prince Albert National Park in 1969, and were captured and moved in June 1969 to Vermette Lake north of Cold Lake Air Weapons Range in western Saskatchewan. The subpopulation is one of two free-ranging populations within or on the periphery of the original Plains Bison range. The subpopulation ranges in a ~2,500 km2 area that is unfenced. Wolves are present (Saskatchewan Environment and Resource Management, 2000). Sex and age structure of the bison population are not manipulated. McCusker River Bison are functioning as a wild population.
Sturgeon River (Saskatchewan):
In 1969, approximately 50 Plains Bison (36 females, 14 males) were translocated from Elk Island National Park to the Thunder Hills north of Montreal Lake and Prince Albert National Park in Saskatchewan. Following release, some bison moved elsewhere. Taking into account calves that may have been born in 1969, the number of founders of the Sturgeon River subpopulation is estimated at between 10 and 22 individuals (SRPB Management Planning Coordinating Committee 2013). This herd is the only free-ranging (unfenced) Plains Bison subpopulation within the core of the original range of Plains Bison in Canada.
The Sturgeon River subpopulation ranges in a 1,053 km2 area in and around southeastern Prince Albert National Park (S. Cherry, Parks Canada, pers. comm.). On average, First Nations people take ca. 20 Plains Bison from the subpopulation outside the national park annually; this take has been skewed towards females (SRPB Management Planning Coordinating Committee 2013). Lone bulls considered a threat by landowners may be removed (S. Cherry, Parks Canada, pers. comm.; R. Tether, Saskatchewan Ministry of Environment, pers. comm.). No other manipulations or management interventions occur. This subpopulation is subject to all natural ecological and evolutionary processes. Wolf predation is an important limiting factor (Fortin et al. 2009). An outbreak of anthrax occurred in 2008; 28 carcasses were found and none were treated (Shury et al., 2009). Sturgeon River Bison are functioning as a wild population.
Grasslands National Park (Saskatchewan):
Grassland National Park is situated near the southwestern border of Saskatchewan in the mixed grasslands natural sub-region. In December 2005 the park received 71 Plains Bison from Elk Island National Park. They were released into a 181 km2 fenced area (west - Larson Block) and the subpopulation grew to about 350 by the fall of 2012. The target bison population is 300-350, which may be increased pending the outcome of research on the impacts of grazing at this stocking level. Recent purchases of privately owned freehold land and provincial Crown leased land adjacent to the park also offer opportunities for enlargement of the enclosed area (W. Olson, Parks Canada Agency, pers. comm.).
There have been no removals for population management to date. However a draft management plan calls for non-selective removal of juveniles to control the population at the target level (Wes Olson, Parks Canada, pers. comm.). Hence the stable population structure should be similar to wild subpopulations. There are no effective predators of bison remaining in this system, although some species (e.g., Wolves, Cougars, and Grizzly Bears [Ursus arctos]) are experiencing signs of recovery in the region (e.g., Montana) and the functional role of Coyotes (Canis latrans) within the system. The newly established Grasslands National Park Bison are already largely functioning as a wild population, albeit with more limitations than the other four subpopulations.
Wood Bison
Nine “conservation herds” identified by Gates et al. (2010) meet most or all of the above criteria (three without any limitations), and were deemed to be functioning as wild populations for the purposes of this assessment:
Aishihik (Yukon):
In March 1986, 34 Wood Bison from Elk Island National Park were released in a 5 km2 fenced holding area near Carmacks, Yukon. Additional shipments were made in 1989, (10), 1990 (50), and 1992 (48), with the total bison imported as founders being 142. Twenty-one Wood Bison were released in March 1988, followed by additional releases each year until 1992, bringing the total released to 170. The bison successfully established in the Aishihik Lake area where the subpopulation currently ranges in an area of about 11,000 km2. The subpopulation is subject to regulated hunting by sport hunters at a rate of about 10% annually (Government of the Yukon 2012) to limit population size. In addition, a few animals (none in the last 3 years) have been culled when they posed a threat of collisions on the Alaska Highway or they encroached on agricultural lands (Government of the Yukon 2012). The current population objective for the herd is to maintain it at or near 1,000 animals after hunting season (Government of the Yukon 2012). The new management plan also identifies management zones into which bison will not be allowed to expand. Predation by wolves occurs (Jung 2011). Aishihik bison are functioning as a wild population without limitations.
Nordquist, British Columbia/Yukon:
Forty-nine Wood Bison from Elk Island National Park were reintroduced into the Aline Lake area east of Liard Hotsprings in northern British Columbia in 1995 (Harper and Gates 2000). Soon after they were released they dispersed westward, taking up residence in the Alaska Highway corridor. They found abundant grazing along the cleared land adjacent to the highway that was vegetated with agronomic plant species. In recent years they extended their range along the highway corridor. The Nordquist herd ranges in an 11,000 km2 area with areas of concentration along the Alaska Highway, from Muncho Lake, British Columbia to as far west as Watson Lake, Yukon (Rowe 2007, Leverkus 2012). The Nordquist subpopulation is subject to predation by wolves, and all other natural limiting factors and ecological processes are present (D. Fraser, BC Ministry of Environment, pers. comm.). No hunting or culling occurs. Predation by wolves occurs (Harper et al. 2000). The Nordquist subpopulation is functioning as a wild population.
Nahanni (Northwest Territories /British Columbia/Yukon):
The Nahanni subpopulation was established in 1980 with the reintroduction of 28 Wood Bison translocated from Elk Island National Park to near Nahanni Butte in southwestern NWT. By 1981, Wood Bison numbers in the Nahanni area had declined to 14 individuals through accidental deaths and dispersal (Gates et al. 2001a). It was augmented in 1989 with 12 Wood Bison, followed in 1998 by 59 Wood Bison released north of Fort Liard (Gates et al. 2001a; Larter and Allaire 2007). The 7,590 km2 winter range (Larter and Allaire 2013) occurs in three jurisdictions, the NT (4,308 km2), British Columbia (2,797 km2), and Yukon (485 km2). Based upon recent GPS collar data the annual range has increased to an estimated 11,700 km2, albeit not as a result of an increase in population size (N. Larter, NWT Department of Environment and Natural Resources, pers. comm.). Seasonal movements are not restricted by human infrastructure or management interventions. Regulated hunting was initiated in 1998. A maximum of seven male-only tags are available annually by a quota system; the annual quota has never been taken (N. Larter, pers. comm.). Nuisance animals are occasionally culled if they pose a risk to human safety in or near communities. There are no other manipulations. The Nahanni subpopulation is exposed to a full range of natural ecological and evolutionary processes and is functioning as a wild population.
Etthithun (British Columbia/Alberta):
An initial attempt to establish a wild subpopulation of Wood Bison in 1996 on the Etthithun River in northeastern British Columbia failed when the released animals moved 100 km south into an agricultural area and mixed with a commercial Plains Bison herd (Harper and Gates 2000). A second attempt was made in 1999 when 19 Wood Bison were translocated from Elk Island National Park to a fenced site near Etthithun Lake. In 2002, 43 bison were released from the enclosure (Rowe and Backmeyer 2006). Range expansion into Alberta was first observed in 2008 (D. Moyles, Alberta Sustainable Resource Development, pers. comm.). The subpopulation is subject to wolf predation and seasonal movements within an area of ~5,000 km2 are unimpeded by fences. Nuisance animals in agricultural areas are culled (6 in the past 10 years; D. Fraser and C. Thiessen, BC Ministry of Environment, pers. comm.). There are no other manipulations. As such, the Etthithun subpopulation is exposed to a full range of natural ecological and evolutionary processes, and is functioning as a wild population.
Hay-Zama (Alberta):
In 1984, 29 Wood Bison were translocated from Elk Island National Park to a fenced area near Hay Lakes in the northwestern Alberta. A free-ranging subpopulation was established in 1993 when 48 Wood Bison escaped from the enclosure (Mitchell and Gates 2001). The subpopulation has since grown at a moderate rate and animals are taken by sport hunters and First Nations to limit range expansion and the herd size (Government of Alberta 2011). Individual animals presenting a risk to human safety are also removed (L. Fullerton, Alberta Sustainable Resource Development, pers. comm.). There are no other manipulations. The Hay-Zama subpopulation occupies an area of ~9,000 km2 and is exposed to a full range of natural ecological and evolutionary processes. Licensed hunting begun in 2008. Hay-Zama bison are functioning as a wild population without limitations.
Mackenzie (Northwest Territories):
In the winter of 1962-1963, 77 bison were captured near the Nyarling River in northwestern Wood Buffalo National Park to establish a captive-breeding herd of Wood Bison (Wood Bison Recovery Team 1987). In August 1963, 16 bison were moved to the Mackenzie Bison Sanctuary near Fort Providence, NWT (Gates and Larter 1990). The herd ranges over a vast area of 21,000 km2 west of Great Slave Lake in NWT, from the Mackenzie River north almost to Yellowknife. Limited regulated hunting is permitted and bison that occasionally enter towns or villages are hazed away or shot to prevent injury to people or property damage (T. Armstrong, NWT Department of Environment and Natural Resources, pers. comm.). MacKenzie bison are functioning as a wild population without limitations.
Greater Wood Buffalo National Park (Alberta/Northwest Territories):
Bison in the Wood Buffalo National Park and surrounding subpopulations represent the largest meta-population of wild Wood Bison. The history of its establishment was discussed earlier (see History of Decline and Early Restoration of American Bison in Canada). The Greater Wood Buffalo National Park meta-population ranges in a vast area (~58,000 km2) in Alberta and NWT. It is composed of the following interacting units (the extent of interactions presumably depends on proximity): Slave River Lowlands; Wentzel Lake; Wabasca and Mikkwa River areas west of the park in Alberta, and the Ronald Lake herd south of the park, also in Alberta. Bison hunting by non-aboriginal residents and non-residents is prohibited in the park and in the Slave River Lowlands. Unregulated hunting of bison occurs in other subpopulations surrounding the park. Bison in the Wood Buffalo National Park meta-population are functioning as a wild population without limitations.
Elk Island National Park (Alberta):
In 1965, 40 bison were captured in the Nyarling River area of Wood Buffalo National Park and transported to an enclosure at Fort Smith, NWT (Novakowski and Stevens 1965). Concern about anthrax in wild bison in the Slave River Lowlands led to a decision to establish a captive breeding herd well outside the region, and 23 were successfully moved to the southern portion of Elk Island National Park in 1965. Two additional calves were translocated in 1968. Brucellosis was detected in the new herd. All animals originating from Wood Buffalo National Park were subsequently culled and the 11 remaining juveniles were vaccinated. The herd was declared free of brucellosis in 1972 and has remained so through annual testing since then (see Nishi et al. 2002a, Halbert et al. 2005).
The Elk Island National Park Wood Bison subpopulation ranges in a 58 km2 area in the southern portion of the park surrounded by a high game fence, and is separated from the Plains Bison herd by a highway and another game fence. The herd is maintained at 350 to 450 to remove surplus individuals with removals intended to emulate natural mortality by focusing non-selectively on calves and yearlings. There is no supplemental feeding, although baiting is used to capture animals. Older animals are allowed to succumb to natural factors in the park; one pack of wolves recently moved into the area. Habitat management is accomplished through prescribed burning. There are no other manipulations. Wolves have recently been observed in the south part of the park (in the Wood Bison area) and in the adjacent Blackfoot Grazing Reserve and have since reproduced (Martha Allen, Parks Canada, pers. comm.). Elk Island Wood Bison are largely functioning as a wild population with limitations of being confined to a small area (< 200 km2).
Chitek Lake (Manitoba):
In 1981 the Waterhen First Nation (now Skownan First Nation) proposed the development of a commercial Wood Bison ranch on its land that would also produce surplus Wood Bison for release to the wild (Payne 1987). The first shipment of 34, consisting of surplus stock from several zoos in western Canada, arrived in February 1984. Animals were released into a 2.5 km2 holding area near the community of Waterhen; the next year the animals were transferred into a 23 km2 fenced pasture.
By 2009, the subpopulation had increased to about 300 (Joynt 2010), and occupied an area of approximately 3,800 km2. It is extra-limital with respect to the original range of Wood Bison, although in an ecological area that was a likely zone of overlap between Wood and Plains Bison during winter months. All ecological and evolutionary processes are present, including wolf predation (B. Joynt, Manitoba Conservation, pers. comm.). With the exception of a small number of bison that threatened public safety or that became a concern for agriculturalists, animals in this subpopulation are not hunted or culled. Mineral blocks are provided as bait in winter to help keep animals distributed within the core range (B. Joynt, Manitoba Conservation, pers. comm.), but there is no supplemental feeding. There are no other manipulations. Chitek Lake bison are functioning as a wild population.
Summary
In summary, five Plains Bison and nine Wood Bison subpopulations in Canada are functioning as wild populations, occurring in locations where prevailing conditions support natural selection and continued evolution of the species, with minimal human interventions. All but three of these 14 subpopulations (two Plains Bison and one Wood Bison) are functioning as such without limitations; the most frequent limitation for these three subpopulations is being confined to a small area or not being subject to natural predation.
2) Geographic or Genetic Distinctiveness
COSEWIC guidelines for manipulated populations (COSEWIC 2010a) also stipulate that “wild by nature” populations included in the assessment must be geographically or genetically distinct from other manipulated populations that are not included. At the population scale, the nine wild Wood Bison subpopulations in this assessment, each of which is largely isolated from the others, can all be considered to be geographically separated from commercial bison herds, which tend to be fenced. Although there are over 1,000 commercial bison herds in Canada, the five wild Plains Bison herds are all largely geographically separated from them. Exceptions include a privately owned Plains Bison ranch located on the eastern boundary of Prince Albert National Park (Shury et al. 2009), and commercial herds in the vicinity of the Pink Mountain subpopulation.
The legacy of hybridization between Wood and Plains Bison, and between bison and cattle, combined with the extreme population bottleneck in the 1880s as a result of the near-extinction of the species, complicate the genetic picture. Although evidence has been provided that several Plains Bison herds are genetically distinct (Wilson and Strobeck 1999, Halbert and Derr 2008), there have been no analyses examining whether the collective set of wild subpopulations being assessed here are genetically distinguishable from all other Plains or Wood Bison. Given both isolation and the common practice of purposeful selection over several generations for traits favourable for human needs, it is likely that privately owned commercial and display bison have accumulated genetic differences characteristic of adaptation to the captive environment. However, this picture may be complicated by the common practice of Elk Island National Park to sell surplus Plains Bison to agricultural producers, which began in the early 1980s.
In conclusion, subpopulations of bison in Canada that have been established for purposes other than conservation are confined, which largely precludes the possibility for individuals to interbreed with the 14 wild Plains and Wood Bison subpopulations under consideration in this assessment. They are therefore geographically distinct from one another, but may not yet be genetically distinct.
Habitat
Habitat Requirements
The original range of American Bison encompassed 22 major habitat types (classification adapted by Sanderson et al. 2008 from eco-region classes mapped by Ricketts et al. 1999). Eighteen of these habitats occur in the original continental range of Plains Bison and seven in that of Wood Bison (Gates and Ellison 2010). Despite the variety of habitats used by bison, members of both DUs are predominantly grazers; grasses and sedges generally dominate diet composition (Reynolds et al. 2003, Gogan et al. 2010), regardless of availability. Bison tend to show strong selection for open habitats, particularly grasslands and sedge meadows. Forested areas are used primarily for thermal and escape cover.
Diet breadth varies among ecoregional habitats and between seasons (see comprehensive review by Gogan et al. 2010). A few examples relevant to bison ranges in Canada are as follows: In the Mackenzie Bison Range, NT (Subarctic Boreal Forest; glaciated plain, meadows in forest matrix), Wood Bison foraged during the winter almost exclusively on grasses and sedges in meadow communities (Larter and Gates 1990). The summer diet was a diverse mix of sedges, grasses, and shrubs (predominantly willow). Lichens, available in forests adjacent to meadows, became an important element of the fall diet. In the Northern Mixed Grasslands grasses and sedges dominated diets in all seasons (88-96%: Gogan et al. 2010, COSEWIC 2012). At the Parkland/Boreal Forest interface in central Saskatchewan, sedges, grasses and rushes comprised 81-99% of seasonal diets (Fortin et al. 2002). One frequently cited exception to typical diet composition of bison is the Plains Bison introduced to the Farewell Lake area (Arctic Lowland Taiga) in interior Alaska (Waggoner and Hinkes 1986). Sedge and grass communities were limited there to small patches scattered on alluvial gravel bars, the beds of dry glacial lakes, margins of shallow lakes, and to an area of forest and shrublands burned 9 years earlier. The summer diet, based on fecal samples collected in June, was composed of 94% willow (Salix spp.). The fall diet (early October) was entirely shrubs: 60% willow, and 40% silverberry (Elaeagnus spp.) and buffaloberry (Shepherdia spp.). Summer range was not limiting for this subpopulation; where grasses and sedges are available they were used heavily, and sedges were the main source of winter forage (Campbell and Hinkes 1983). Rather than an exception, the example of the Farewell herd supports the inference, consistent with the other examples, that bison require grazing habitat that provides sedges and grasses, particularly during the most limiting season (winter) and where they have the flexibility to select the most available nutrient-rich foliage in other seasons.
Other examples support the flexibility of foraging habitat selection, yet are consistent with what is understood about the ecological niche of bison. The Nahanni subpopulation is strongly associated with the Liard River and riverine islands (Larter and Allaire 2007) that provide abundant stands of Salix and Equisitum used in summer as forage. The Aishihik subpopulation seasonally uses alpine meadows (T. Jung, Yukon Department of Environment, unpublished data). The Nordquist subpopulation almost exclusively uses the grassy roadside verge along the Alaska Highway (Thiessen 2010).
Bison habitat selection varies across the range of landscapes they occupy during seasonal movements. Plains Bison in Canada commonly moved over long distances from open grasslands to mixed woodlands in the fall and winter (Roe 1970, Louis Riel Institute 2011). In an analysis of grassland communities associated with the historically observed movements of the herds in the Prairie Provinces, Morgan (1980) concluded that the availability of superior forage appeared to be the primary stimulus of major bison movement patterns each year. Metis ATK holders historically recognized two ‘grand divisions’ of ‘buffalo’, the Grand Coteau/Red River and the Saskatchewan herds (Louis Riel Institute 2011). Based on differences in radio-carbon isotope concentrations in warm season C4 grasses typical of the prairies and cool season grasses common in the Parklands and Foothills, Chisholm et al. (1986) demonstrated that bison in the Canadian plains used both parkland vegetation and mixed grass prairie.
Research in the Mackenzie Bison Range and Wood Buffalo National Park revealed that even while small in extent (~10% or less of landscapes) and patchily distributed, graminoid (sedge and grass) meadows are the most used habitats by Wood Bison (Larter and Gates 1991a,b, Larter et al. 2000). Such habitats are associated mainly with watercourses and the fringes of water bodies. Several geographically disparate studies have demonstrated that, given a choice, both Plains and Wood Bison preferentially forage in high biomass lowland meadows dominated by sedges and grasses in the winter (Mackenzie: Larter and Gates 1991a,b; Slave River Lowlands: Reynolds et al. 1978; Elk Island National Park: Telfer and Cairns, 1986; Yellowstone National Park: Barmore 2003, DelGiudice et al. 2001). In forested landscapes these habitat patches are connected by well-used trails that bison follow to seek forage and avoid wolves (Gates et al. 2005, Fortin et al. 2009). In mountainous habitats, bison have been observed to forage in lowland meadows in winter then move to higher elevations to feed on phenologically less mature plants that green up later in the spring and summer. Seasonal habitat shifts following elevation gradients may reduce grazing intensity in valley bottoms in summer, sparing forage there for winter use (Frank and McNaughton 1992). In Yukon, Wood Bison primarily foraged in lowland meadows and open forest areas in valley bottoms in winter when the populations were at low densities (Fischer and Gates 2005). During a period of recent population growth, bison have expanded the range of habitats they occupy to include alpine tundra (T. Jung, Yukon Department of Environment, unpublished data).
Forage availability and quality, hence habitat suitability and carrying capacity, varies among plant communities in the boreal forest. In addition to a primary focus on grasses and sedges, Wood Bison use the new growth of willow during spring and early summer, some forb species during summer, and fruticose lichens in autumn (Larter and Gates 1991a,b). Habitat selection is narrow during winter when bison forage almost exclusively in meadows and graminoid-dominated shrublands; selection is broader in other seasons. Foraging efficiency and habitat selection, and site and diet selection are constrained by forage biomass density and quality (Hudson and Frank, 1987; Fortin et al. 2003). Bison can be expected to avoid or sample habitats where forage biomass is below a biomass density threshold (Hudson and Frank, 1987; Fortin et al. 2003), and where preferred food is not available. In an area west of Wood Buffalo National Park, Strong and Gates (2009) found sustainable stocking rates for Wood Bison varied among eight broadly defined vegetation types (Treed Uplands, Treed Lowlands, Mixed Tall Shrub/Sedge, Closed-canopied Willow, and Open-canopied Willow, Meadow, Wetland Grass, Wetland Sedge). Meadows and wetlands produced higher foraging efficiency thresholds than treed communities. The authors concluded that treed upland habitats in northern Alberta are marginal summer foraging habitats and predicted they would not be used in winter by Wood Bison due to the low availability and quality of senesced forage under snow cover. They added that the regional carrying capacity for bison in the Lower Peace River district depends on numerous small wetland and non-forested patches of vegetation distributed in the forest matrix. In combination, they represent only 1.8% of the landscape north of 57.5°N (160,700 km2) in Alberta, which limits the capacity of the region to support Wood Bison restoration.
Habitat Trends
Plains Bison habitat
The original range of the Plains Bison included mixed grasslands, parklands, the eastern slopes of the Rocky Mountains, and tall grass prairie and mixed woodlands in Manitoba. Today the habitat mosaic present in prairie landscapes largely reflects the early agricultural history of the Prairie Provinces. Cattle ranchers began to move into the region beginning in about 1870. By 1930, most of the land cultivated today had been converted for crop production.
The Canadian prairies have been dramatically altered by the cumulative effects of cultivation, irrigation, access infrastructure, petroleum and natural gas development, mining, urban growth, electrical transmission lines and other developments (Forrest et al. 2004). As development progressed following settlement and the human population grew, roads associated with industrial and rural development increased. By the mid-1990s more than 90,000 km of highways, roads, and wellsite access trails existed in the Grassland Natural Region of Alberta (Alberta Environmental Protection 1997). In the prairie region of Canada, 24% of native mixedgrass prairie remains intact and less than 1% of tallgrass prairie (Gauthier and Wiken 2003; ref). The Aspen Parkland has been reduced to about 25% remaining natural cover; Moist Mixed Grassland to 30% and Mixed Grassland to 45% (Riley et al. 2007). The remaining parcels of native habitat lie scattered in a sea of agriculture, with the largest intact areas in southeast Alberta and southwest Saskatchewan, and along major rivers. Pressure on remaining native landscapes is incessant: between 1971 and 2001, native range in Saskatchewan declined by 20% (Saskatchewan Environment 2005). Already intensively used for oil and gas extraction, new technologies such as hydrological fracturing of geological formations and wind energy development are intensifying the human footprint on the prairies.
Today, habitat occupied by wild Plains Bison exists in two national parks and at the northern periphery of agriculturally developed (arable) lands. Within Plains Bison habitat in Canada, the predominant land use is seasonal cattle grazing. In these areas significant institutional, historical and cultural barriers exist for shifting land use to include restoration of Plains Bison as a wild species.
Wood Bison habitat
Unlike the grassland and mixed-woodland habitat of Plains Bison, agricultural development and settlement have negatively affected relatively small areas within the original range of Wood Bison in boreal eco-regions. Crop and livestock production are most developed in the Peace River corridor extending north to Fort Vermillion west of Wood Buffalo National Park (Hamley 1992, Bowen 2002) and from Peace River Alberta west to Fort St. John in British Columbia.
Strong and Gates (2009) suggested that agricultural and other land use may have reduced the area of high-quality habitat originally available to bison. Much of the Peace River Parkland was converted to agricultural uses by the mid-1930s and is not available for wild Wood Bison. In addition to agricultural development, post-settlement fire control may have contributed to a reduction in foraging habitat through forest expansion (Lewis 1982: p. 21). A rise in regional temperature (Jacoby and D’Arrigo 1989; Davi et al. 2003) and increased precipitation (Bradley et al. 1987) may have favoured greater forest canopy density and light attenuation causing understory graminoid suppression, and woody plant encroachment into previously open sites (see Raup 1947, Jeffrey 1961, Schwarz and Wein 1997). An increase in forest abundance during the latter half of the 20th century is clearly represented in the palynological record in northern Alberta (Larter and MacDonald 1998). Upon reviewing historical evidence, Strong et al. (2009) suggested the increase in aspen forest in northern Alberta during the mid-twentieth century was a regional phenomenon. They concluded vegetation change in the Lower Peace River District was a consequence of climate change and upland vegetation changes (more trees) were promoted by increased summer water availability. In lowland meadows, seasonal inundation and periodic flooding play an important role in the dynamics of shrub establishment vs. sedge and grass dominance (Timoney et al. 1997; Timoney 2008a, 2009; Strong et al. 2009). Intentional burning adds to the complexity of these dynamics.
Fire is the dominant stand-renewing agent for much of the boreal forest, greatly influencing plant community structure and function and the quality and extent of wildlife habitat (Stocks et al. 2002). Depending on pre-burn site conditions and burn severity, early seral understory can provide forage for bison at biomass densities and of types that support winter foraging in addition to generating abundant leafy material produced by shrubs and flowering plants used by bison as summer forage (Campbell and Hinkes 1983). Aboriginal communities in northern Alberta used fire to influence the local distribution and relative abundance of plant and animal resources in northern Alberta until shortly after World War II (Lewis 1977, 1980). Fire has also been used elsewhere by Aboriginal communities to varying success (Quilan 1999, Kessel 2002). Today, wild fire potentially has less of an impact on habitat patch dynamics than in earlier times due to fire suppression, although prescribed burning for the purposes of meadow and grassland habitat improvement projects does occur (e.g., NWT [Chowns et al. 1998, Quinland et al. 2003], Banff National Park [Sachro et al. 2005, White et al. 2012], and northeastern Alberta [C. Gates, pers. obs.]).
Forestry and oil and gas development are the prominent industrial land uses in Wood Bison range in northern Alberta and British Columbia. Nearly all of the Boreal Forest in Alberta has been allocated under forest management agreements with companies. In combination with the energy sector, access development has expanded rapidly. Access impacts habitat quality for large herbivores both directly and indirectly. Plantings of agronomic species along roadsides provide attractive forage for bison, but potentially creates sink habitats for bison in the ranges of the Mackenzie, Nordquist, Nahanni and Aishihik subpopulations. Increased road access creates opportunities for public access, which is a concern to some communities in areas where Wood Bison are subject to unregulated hunting (Wentzel, Wabasca/Mikkwa, and Ronald Lake). ATK suggests that resource development leads to habitat access and therefore increased hunting pressures (Schramm et al. 2002).
Forestry in particular, owing to large cut areas, and other industrial activities influence stand age patterns and succession of plant communities in the boreal forest with the potential to affect forage abundance and distribution for Wood Bison (Gates et al. 2001a). Redburn et al. (2008) tested this hypothesis in an aspen clearcut in the mixed-wood forest in the Lower Peace River district west of Wood Buffalo National Park, concluding that enhanced forage production in clearcuts could contribute to increasing summer carrying capacity for bison, but sedge and grass meadows in lowlands areas are necessary to support bison during the winter. The reliance of Wood Bison on sedges and grasses in the winter and use of habitats that produce them makes winter habitat availability an important limiting factor for Wood Bison conservation and restoration.
In addition to the influence of habitat structure and composition on carrying capacity, socio-political factors have a strong influence representing acceptance and tolerance limits for wild bison (Nishi et al. 2006, Bidwell 2009). For example, exclusion of bison from disease control areas in Alberta and the NWT (see Threats) serves as a form of functional habitat loss. Bison control areas exclude Wood Bison from approximately 165,000 km2 of their original range in Canada, an area that is about 50% greater than that which they currently occupy. Moreover, the loss of this habitat is within the core area of their current distribution, where, presumably, habitat quality and quantity are greatest (Strong and Gates 2009) and hold the most potential for Wood Bison restoration at larger spatial scales.
Biology
The ecology, life cycle and reproductive biology of bison has recently been reviewed in detail by Reynolds et al. (2003) and Gogan et al. (2010).
Life Cycle and Reproduction
The typical age of first reproduction in cows is 3 to 4 years old, and the youngest is two (McHugh 1958, Fuller 1961, Shaw and Carter 1989, Green and Rothstein 1991, Wilson et al. 2002). Fuller (1966) reported declining fecundity in females after 13 years of age. The proportion of yearlings conceiving and mean age of reproduction vary among subpopulations with factors such as nutrition, size, prior success, density, and environmental factors (Wilson et al. 2002, Reynolds et al. 2003, Gogan et al. 2010).
The variance in reproductive success has been shown to be significantly higher in males than females in both Plains (Berger and Cunningham 1994) and Wood (Wilson et al. 2002) Bison. Males can be fertile at 16 months in nutritionally supplemented populations (Helbig et al. 2007). Two- and three-year-old males are physiologically competent to breed and are sexually active (Halloran 1968, Maher and Byers 1987). However, male bison generally achieve breeding maturity (adulthood) beginning at five to seven years of age (Meagher 1973, Maher and Byers 1987, Rothstein and Griswold 1991, Komers et al. 1992); Wilson et al (2002) recorded reproductive success in 40% of breeding Wood Bison males; most were in the 7- to 14-years age classes and none were under 5. Reproductive effort is influenced by maturity and experience. Adulthood is reflected in mature body size, risky aggressive behaviours, and seasonal segregation, which allow adult males to compete for mating opportunities. However, they are rarely successful in breeding in the presence of older, larger, more experienced bulls (Shult 1972; Mahan 1978; Shull 1985, Komers et al. 1994a,b) and do not tend to participate in the rut until 5 or 6 years, when large enough to achieve high status (Maher and Byers 1987, Lott 2002). In wild populations not subject to selective hunting pressure, the adult bull: cow ratio can exceed 50:100 (Gates et al. 1995, Gogan et al. 2010).
Females are seasonally polyestrous with a cycle of about 19-21 days during the breeding season (Fuller 1966, Kirkpatrick et al. 1993, Wolff 1998). Cows occasionally come into estrus and breed outside the rutting season, resulting in some calves being born outside the typical (April-June) spring calving period (Soper 1941, McHugh 1958, Banfield 1974). Bison are monoparous; twins are rare (Reynolds et al. 2003). Weaning typically occurs between 8 and 12 months (McHugh 1958, Mahan et al. 1978) but yearlings have been observed to suckle (Hornaday 1889, McHugh 1958, Egerton 1962, Green et al. 1993), and more rarely so have two-year-old bison (Green 1996). Nursing duration is longer in older cows and they are less likely to terminate nursing bouts than young cows (Green 1996).
The bison is a polygynous species; mature males compete for multiple mating opportunities during the breeding season, which occurs between June and September with the peak of activity during July and August (Garretson 1927, Soper 1941, Fuller, 1966; Halloran 1968; Lott 1972; Meagher 1973; Banfield 1974; Haugen 1974). The rutting season varies somewhat in timing and duration among locations (Reynolds et al. 2003). Large bulls form separate, smaller groups from cows, calves, and immature males throughout most of the year (Berger and Cunningham 1994; Komers et al. 1993; Meagher 1973). During breeding when males join the mixed groups is when the largest aggregations occur. Body mass and prior reproductive success were predictors of reproductive success at Elk Island National Park (Wilson et al. 2002). The willingness of bulls to risk injury in fighting other males increases with age, suggesting an increase in reproductive effort with age (Komers et al. 1994b).
Bison have been reported to live longer than 20 years in non-supplemented populations (Halloran 1968, Meagher 1973, Berger and Peacock 1988). In Wood Buffalo National Park, Fuller (1966) considered bison older than 15 to be aged. However, they can live longer than 40 years in captivity (Dary 1974). The generation time of American Bison is estimated as 8 years (Hedrick 2009).
Interspecific Interactions
Given the keystone role of bison on the Great Plains, the number and type of documented interspecific interactions are substantial (reviewed by Knapp et al. 1999; Truett et al. 2001, Sanderson et al. 2008; see Special Significance). Undoubtedly, many other species that interact with bison have not yet been documented, particularly parasitic arthropods (e.g., Tessaro 1989). Moreover, an untold number of plant and animal species benefit from, or depend upon, bison for habitat creation or maintenance, including several species of birds (e.g. Soper 1941, Griebel et al. 1998, Coppedge 2009), small mammals (Matlack et al. 2001, Jung et al. 2010) and amphibians (e.g., Gerlanc and Kaufmann 2003), some of which are at risk (Fox et al. 2012). Among the multitude of known and potential interactions between bison and other species, perhaps those that have the most bearing on status are the processes of predation and competition.
As the largest land mammal in North America, preying or scavenging on bison is seemingly profitable. However, their physical size, social organization, and temperament, make bison formidable prey (Fuller 1953; Smith et al. 2000). Gray Wolves, Coyotes, and Grizzly Bears are the only known predators of bison, and they most often focus on calves and yearlings (Carbyn and Trottier 1987, 1988; Larter et al. 1994; Smith et al. 2000; Sheldon et al. 2009); however, adults are occasionally killed by both wolves (Smith et al. 2000, Jung 2011) and bears (Wyman 2002). In some areas, local wolf packs are relatively adept at killing bison (Carbyn and Trottier 1987; 1988; Larter et al. 1994). Wolves have been extirpated from much of Plains Bison range for over a century. In Yukon, it took over 20 years after reintroduction for the first reports of wolf predation to be documented (Jung 2011). Many recently reintroduced Wood Bison are exposed to, but not yet subject to much natural predation. If kill rates increase, predators (especially wolves) may limit bison population growth rates (sensu Messier 1994). In the NWT, areas with bison resulted in increased wolf abundance (Joly and Messier 2000), which led to increased predation of sympatric moose (Alces americanus) and bison because wolves were subsidized by the availability of alternative prey (Larter et al. 1994).
Competition between Wood Bison and other ungulates (primarily moose and caribou) has been a cause for concern by local communities (Fisher and Gates 2005, Jung and Czetwertynski 2013). For example, ATK in Dene Tha traditional area suggests that bison are responsible for moose population declines (Spyce 2009). Focused research has failed to find substantial diet, habitat, or spatial niche overlap between wood bison and moose and caribou (Fisher and Gates 2005, Jung and Czetwertynski 2013). Results are similar for bison and moose and Elk (Cervus canadensis) in Yellowstone National Park (Singer and Norland 1994). Although Bison, Moose, and Caribou are large herbivores that share the landscape, they employ significantly different positions along the grazer-intermediate-browser gradient (Hofmann 1989), and as such have different dietary requirements, which translate to the use of different habitats and spatial distribution (Jung and Czetwertnyski 2013). Mountain sheep (Ovis spp.), however, are also grazers and there is relatively high overlap in the diet of Wood Bison and mountain sheep, where they are sympatric (Singer and Norland 1994; Jung and Czetwertynski 2013). Spatial segregation and differential habitat selection function to minimize the potential for competition between bison and mountain sheep (Jung and Czetwertynski 2013). For Plains Bison, diet and habitat overlap with cattle and domestic sheep is high (Schwartz and Ellis 1981), while that with Pronghorn (Antilocapra americana) is low (Schwartz and Ellis 1981; Krueger 1986). Potential competition between bison and cattle or sheep may be a cause for concern by livestock producers near bison range.
Physiology and Adaptability
The original distributional range of bison in North America (the greatest of any native ungulate), and the large number of eco-regional habitats they occupied (Figure 4; see Sanderson et al. 2008) reflects the breadth of adaptations of this species to various climate regimes and range conditions. Bison populations are able to thrive under desert-like conditions, endure cold winter conditions in the sub-arctic, and to forage through deep-snow-covered mountain valley grasslands and meadows.
Bison exhibit numerous adaptations permitting them to exist under such an extreme range of ecological conditions. Large body size and wooly pelage provide thermal inertia and insulation (Christopherson et al. 1978). The winter hair coat of the American Bison has a greater weight and follicular density per unit area and finer hair than Hereford, Angus, or Shorthorn cattle (Peters and Slen 1964). Like other northern ungulates, bison exhibited reduced metabolism in winter (Rutley and Hudson 2000). Hawley et al. (1981a,b) reported lower dry matter intake of sedge in bison than cattle during winter. Bison are bulk feeders able to process large amounts of low-quality fibrous forage (Hudson and Frank 1987). Unlike other northern ungulates, bison use their massive heads to clear snow to access forage (Reynolds et al. 2003). This foraging behaviour, in combination with a wide muzzle, allows bison to efficiently consume large quantities of coarse forage during a single feeding bout, despite snow cover. Typical of bulk feeders, bison also possess a large rumen in relation to body size and have a relatively slow rate of rumen turnover (Schaefer et al. 1978). Hence they have a long retention time, enabling them to more thoroughly digest a diet high in cellulose. Bison retain low-quality graminoid forage in their reticulo-rumen longer than cattle and digested low-quality forage more efficiently than cattle (Hawley 1978). Yet the digestive efficiency of bison and cattle on high-quality forage (feeds like alfalfa or alfalfa brome hay) was similar (Hawley et al. 1981a).
Forage limitation can affect age of first reproduction of females, pregnancy rates and birth weights (Kirkpatrick et al. 1993, 1996, Reynolds et al. 2003). Calves and adult bulls are the most vulnerable cohorts to nutrition-related mortality under severe winter conditions (DelGiudice et al. 1994).
Dispersal and Migration
Epp (1988) presented arguments that migratory and non-migratory population segments existed simultaneously. He proposed bison of the Great Plains engaged in dual dispersion behaviour, now referred to as partial migration (e.g. Hebblewhite and Merrill 2006), whereby large herds of Plains Bison engaged in long-distance migration while smaller groups resided locally in mixed woodland areas. Avoiding predation at large scales is hypothesized to be one of the key benefits of long-distance migration in ungulates (Fryxell et al. 1988). Hence, ungulates in migratory populations are more abundant than resident population units. Epp (1988) proposed that migratory segments of the Canadian Plains Bison population were more abundant than non-migratory segments for this reason.
Seasonal migration has not been reported in Wood Bison resident in the Boreal Forest, likely owing to the patchy distribution of suitable foraging habitat in a dominantly forest matrix. For example, in winter, bison foraging habitat in the Mackenzie range occurs in discrete, widely spaced meadow patches representing <6% of the landscape within a boreal forest matrix (Larter et al. 1991a,b, Matthews 1992). A pattern of pulsed, density-driven range expansion following the reintroduction of Wood Bison into the area in 1965 was described by Gates and Larter (1990) and Larter et al. (2000). In that case, population growth rate declined as density approached a threshold of approximately 0.55 bison /km2. Then bison spilled over into the next available habitat patch temporarily reducing density, increasing per capita resources, with renewal of the growth rate of the population (Larter et al. 2000); the cycle was repeated twice. Similarly, in Yellowstone National Park once culling was eliminated as a management practice in the late 1960s, the Plains Bison population exhibited density-driven range expansion (Gates et al. 2005). Range expansion briefly compensated for declining per capita food resources (density-related forage depletion), thus maintaining a relatively stable instantaneous density. However, compensation was not exact; population growth rate declined with density because high quality foraging patches were limited in overall area, were patchily distributed, and depleted first, forcing bison to shift to poorer quality patches as density increased. Likely demographic responses are decreased fecundity and increased juvenile mortality. The pattern of increase to high density followed by expansion into new range is similar to that described for exotic ungulates in New Zealand (Caughley 1970a,b).
Population Sizes and Trends
Sampling Effort and Methods
Inventory efforts vary among bison subpopulations. Subpopulations that are fenced (e.g., Elk Island National Park and Grasslands National Park) tend to be monitored most frequently owing to the requirement to manage surplus animals. Survey effort varies widely for free-ranging populations. Some subpopulations are counted periodically (e.g. Aishihik, Hay-Zama, Mackenzie, Wood Buffalo National Park, Sturgeon River), whereas several others are counted infrequently (e.g., Pink Mountain). The McCusker River Plains Bison herd has never been formally inventoried.
Bison subpopulations are estimated in a number of ways. Most are enumerated using total counts (Wolfe and Kimball 1989) made during surveys in winter from low-flying aircraft. In a total count, no attempt is made to correct for the number of animals not observed. These surveys assume that most, if not all, bison are seen during aerial surveys, as they congregate and use open habitats (e.g., Bradley and Wilmshurst 2005, Hegel et al. 2012) during this time of year, and do not run away when approached by aircraft (Fancy 1982). Such surveys are generally undertaken systematically by following standardized line transects. Total counts represent minimum counts and lack confidence limits.
Population numbers for a few Plains and Wood Bison populations have been estimated using methods that result in a measure of sampling variance. For example, strip line transects have been used for several Wood Bison subpopulations (e.g., Bradley and Wilmhurst 2005; Larter et al. 2007; Armstrong et al. 2011; Larter and Allaire 2013; Armstrong 2013), and the Pink Mountain Plains Bison subpopulation (C. Thiessen, BC Ministry of Environment, pers. comm.). A mark-resight method was used to inventory the Aishihik subpopulation (Jung and Egli 2012), and the Sturgeon River subpopulation (Merkle and Fortin in press).
Age and sex classes of bison are often not provided with census data. When individuals are classified into age classes during field counts, it is most common to report either calves or calves plus yearlings and adults ≥ 2 years (see Abundance and Trends). Yet, reproductive maturity is reached at a later age for most (females: 3 years and males 6 years; see Life Cycle and Reproduction). Although multiple age classes can be discerned, this requires viewing from close range (Carbyn 1998). The age of first reproduction of an individual bison is sensitive to a number of physiological and contextual factors (e.g., nutritional condition, whether the population is expanding, habitat quality, group social structure, etc. [Gogan et al. 2010]); (see Life Cycle and Reproduction).
Abundance and Trends
Inventories and trend data for the five Plains Bison and nine Wood Bison subpopulations considered in this assessment are discussed for each and summarized in Tables 2 and 3, respectively. Where available, information on age structure and population trends over the past three generations (ca. 24 years) are presented for each subpopulation, from west to east:
Plains Bison
Pink Mountain, BC:
This subpopulation has grown substantially since establishment in the early 1970s. An inventory conducted in 2003 yielded a presumed total count of 877 animals with no sightability correction. The age structure was modelled at 22 calves, 16 yearlings, and 43 bulls per 100 cows (Rowe 2006), The most recent aerial survey was conducted in 2006, using a stratified random block design Observed individuals were classified into calves, yearlings, cows (>2 years), bulls (2-6 yrs) and mature bulls (≥ 7 years). The 2006 subpopulation was estimated at 1,302 (90% CI= 1233 - 1371) individuals. About 26% were calves or yearlings, and 51% were females ≥ 2 years and males ≥ 7 years (Rowe 2006), yielding a rough estimate of 629-699 mature individuals.
A population modelling effort by Rowe (2006) suggested an average finite rate of growth of approximately 14% per year in the previous 3 years. For the purposes of setting hunting rates, managers have assumed a continued annual growth of 5%, but hunting limits have also increased each year (C. Thiessen, pers. comm.). Rowe’s (2006) modelled estimate of 546 animals in 1989 suggests a population growth of 138% in three generations, assuming the subpopulation has remained stable since 2006, which is unknown given the increasing hunting rate over the same period.
The Pink Mountain subpopulation is managed to control population growth and range expansion, given its proximity to Wood Bison subpopulations in northeastern British Columbia. Since the last survey in 2006, Pink Mountain bison have been assumed to be increasing in number, and the hunting rate was adjusted from ~10% to 16% in an attempt to slow the population increase. In 2007 the population estimate was calculated using the 14% growth rate that was observed between 2003 (873) and 2006 (1,302). From 2009 to present the population estimate has been modelled assuming a 5% growth rate per year from the 2006 population count of 1,302. Although the annual allowable harvest has increased from 2009-2013, the actual number of authorizations issued has remained fairly steady from 2009-2013 with 411 issued for the 2013 hunting season (D. Lirette, B.C. Ministry of Forests, Lands and Natural Resource Operations, pers. comm.).
Elk Island National Park, AB:
The subpopulation is counted annually and managed on a semi-annual basis during handling of bison in facilities to remove surplus animals from the hyper-abundant population. Sex and age structure information is collected on a subset of animals during penning. Productivity is also assessed every summer to determine the ratio of cows to calves. Winter ungulate aerial surveys are flown every year, yielding minimum counts of bison of all ages. Following a 2009 Ungulate Options Analysis to address management of the high densities of bison, elk, moose, and deer in the 194 km2 park, the Plains Bison population goal was lowered to a recommended adult population of 250-275 (PCA 2009). The 2013 Plains Bison estimate was 680; at least 200-300 animals are expected to be removed from the herd in the coming year to translocate for conservation or research projects and sold to auction (M. Allen, Parks Canada, pers. comm). There are an estimated 204-300 mature individuals, using minimum (0.51) and maximum (0.75) proportions from Plains Bison subpopulations with field data (Pink, Sturgeon River, Grasslands).
McCusker River, SK:
This subpopulation has not been formally censused at any time since established in 1969. Referencing local biologists, Wilson and Zittlau (2004) reported that the subpopulation was about 70-100 animals in 2003. A 2011 population estimate of 100-150 (R. Tether, Saskatchewan Ministry of Environment, pers. comm.) was based on anecdotal information provided by observers flying over the air weapons range and ground sightings outside the range in Saskatchewan. Range expansion onto adjacent provincial forest lands supports the notion that the population may have increased modestly since 2003 (R. Tether, Saskatchewan Ministry of Environment, pers. comm.), although this cannot be confirmed. Number of mature individuals is unknown, but estimated at 51-113, using minimum (0.51) and maximum (0.75) proportions from Plains Bison subpopulations with field data (Pink, Sturgeon River, Grasslands).
Sturgeon River, SK:
Annual systematic aerial transects of the core range of the Sturgeon River subpopulation have been conducted since 1996 (SRPB Management Planning Coordinating Committee 2013). No sightability correction factor has been applied in these surveys, and some animals are likely missed when in dense conifer forest. Regardless, the data represent minimum counts and likely provide a reliable indication of trends because of their systematic nature (S. Cherry, Parks Canada, pers. comm.). Merkle et al. (in prep.) developed a count-based model integrating these aerial survey minimum counts since 1996 and habitat selection from GPS radio collar data, deriving estimates with confidence intervals. Although the estimates were generally higher than the minimum counts, they mirrored the trends. Further, Merkle et al. (in prep.) conducted annual field-based population estimates in 2011-2013. Estimates were derived from capture-mark recapture models using identified photographs of individual adult bison, and calf and juvenile to cow ratios observed in the field (Merkle and Fortin in press). In all three years, estimates from aerial transects were close to the capture-mark recapture estimates.
In 1996, the first aerial survey reported a minimum count of 111 individuals (SRPB Management Planning Coordinating Committee 2013). Wilson and Zittlau (2004) reported the subpopulation in 2002 as 320 bison; it continued to grow until peaking at 400-500 bison between 2006-2008 (Parks Canada Agency 2012). The subpopulation has since declined and is currently estimated at 200-250 bison, based on aerial surveys and the maximum number estimated by local First Nations, ranchers, and outfitters (Parks Canada Agency 2012) and mark-recapture methods and count-based models (Merkle et al. in prep). Reasons for the decline include an anthrax outbreak in 2008, increased wolf predation, and hunting (SRPB Management Planning Coordinating Committee 2013). Merkle and Fortin (in press) reported 75% of the 2011 population as adults (> 2 years old), while this proportion was 65% in 2012 (Merkle et al. in prep.). This means there are approximately 130-188 adults, with an unknown proportion of these being mature individuals.
Grasslands National Park, SK:
Since the establishment of this subpopulation in 2005, it has been increasing steadily. Animals are censused annually through ground counts of calves, yearlings and adults ≥ 2 years (Olson and Sissons 2011). The most recent count from May 2013 yielded 317 individuals, including 17 calves (L. James, Parks Canada, pers. comm.). Based on past birth rates for this herd, the 2013 calf crop is expected to be 100 individuals, bringing the total bison population to approximately 400, with 189 (47%) aged three years and older (Wes Olson, pers. comm.).
Summary Plains Bison:
The best current estimate for the total number of individual wild Plains Bison in Canada is 2,333 - 2,571 (Table 2). This represents an overall increase of ca. 543-751 animals (~36%) since the last COSEWIC assessment in 2004 (1,790-1,820; COSEWIC 2004). This increase is a result of the addition of one subpopulation (Grasslands National Park) and a 47% increase in the Pink Mountain subpopulation. During the same time period, however, the Sturgeon River subpopulation declined by ca. 30%. About half of the total Plains Bison population in Canada is composed of the extra-limital Pink Mountain subpopulation.
| Population | Jurisdiction | Est. range (km2) | Est. total population in (year) | Est. Mature individualsFootnotea | Earliest pop. est. (year) | SourcesFootnotec |
|---|---|---|---|---|---|---|
| Elk Island National Park | AB | 136 | 400 (2013)Footnoteb | 204-300 | - | 1 |
| Sturgeon River | SK | 750 | 200-250 (2012) | 130-188 | 111 (1996) | 2 |
| McCusker River | SK | 2,500 | 100-150 (2012) | 51-113 | - | 3 |
| Grasslands National Park | SK | 181 | 402 (2012) | 189 | 71 (2005) | 4 |
| Pink Mountain | BC | 3,200 | 1,233-1,371 (2006) | 629-699 | 877 (2003) | 5 |
| Totals | - | 6,767 | 2,335 - 2,573 | 1,204-1,490 | - | - |
The number of mature individuals (females ≥ 3 years and males ≥6-7 years) in the Plains Bison population in Canada is estimated at 1,204-1,490, based on the best available information (see Table 2 for age structure information). Wild Plains Bison exist today at a tiny fraction of their original numbers (prior to 1850; Gates and Ellison 2010), and only one of the four subpopulations (Pink Mountain) numbers more than 500 individuals. There has, however, been an overall increasing trend in mature individuals over the past three generations.
Wood Bison
Aishihik, YT:
Since its establishment in the early 1990s, the Aishihik subpopulation was censused regularly by total count until 2006. The herd grew from 350 individuals in 1998 to more than 500 in 2000 (Gates et al. 2001a). From 2007 to 2011 a bi-annual mark-re-sight protocol was implemented, using animals paint-balled and later re-sighted from a helicopter (Hegel et al. 2012). In 2007, the population was estimated at 1,089 (90% CI = 970–1309) and in 2009 at 1,151 (90% CI = 998–1335) (Government of Yukon 2012). The most recent population census was conducted in 2011, yielding a population estimate of 1,230 (90% confidence intervals were 1,106-1,385; Jung and Egli 2012). The population has grown at about 10% per year since 1998, despite a hunting rate of about 11.5% per year during the same period (Jung et al. 2012). Growth in recent years (2007-2011) has slowed to about 2.9% per annum, largely due to high hunting levels, but wolf predation and wounding losses may also play a role. The hunt has been liberalized in a concerted effort to reduce the size of the herd (Jung et al. 2012). In 2011, calves were differentiated from the rest of the population; the average percentage of calves observed during the surveys was 19.9%, but number of adults or mature individuals was not reported (Jung and Egli 2012).
Nordquist, BC:
Estimates for the Nordquist subpopulation are based on total counts from aerial surveys and roadside transects (Thiessen 2010). Following its establishment in 2002, the subpopulation was estimated at 97 in 2007 (Rowe 2007) and 117 in 2010 (Thiessen 2010), indicating an approximate doubling of the subpopulation since 2002 (50). The proportion of adults (≥ 2 yrs) ranged from 72-81% during the latter survey (Thiessen 2010). This means there are approximately 84-95 adults, with an unknown proportion of these being mature animals. The subpopulation appears to be relatively stable in recent years, and is primarily limited by traffic accidents (D. Fraser, British Columbia Ministry of Environment, pers. comm.). Mortality from industrial traffic averages about 10 a year (~10%), and has been as high as 33% in one year (C. Thiessen, British Columbia Ministry of Natural Resource Operations, pers. comm.).
Nahanni, NT, BC:
The first aerial surveys of the Nahanni subpopulation were conducted in 1995-1997; in 1998 it was estimated to number ca. 160 individuals (Gates et al. 2001a). The subpopulation was censused in March 2004 and again in March 2011 using strip line transects (Larter et al. 2007; Larter and Allaire 2013). No correction was made for sightability for the 2004 survey, but a correction factor was applied in 2011. Although the coefficient of variation was consequently reduced in 2011, both censuses had similar results of 403 (2004) and 431 (2011; 95% CI= 218-644) bison. Calf vs. non-calf (94%) observations were tallied separately in the latter survey (Larter and Allaire 2013). Number of adult or mature individuals was not reported. Annual sex and age classification surveys are conducted during summer for the subpopulation. Over the past 10 years the number of calves per 100 adult females and the estimated overwinter survival of calves have been relatively stable (Larter and Allaire 2007; N. Larter pers. comm.).
It appears that after several augmentations and slow growth since establishment in 1980 (reviewed in Larter and Allaire 2007), the subpopulation has been stable for at least the last seven years. Since 1998 (ca. two generations), the average exponential rate of increase was 0.074. Limiting factors appear to be occasional drowning, traffic accidents, and hunting, and possibly adult mortality related to excessive tooth wear caused by a diet high in silica (i.e. Equisetum, Larter and Allaire 2007).
Etthithun Lake, BC/AB:
This subpopulation has been censused using total counts from both aerial and road-based surveys. Bison were first noticed east of the Alberta-BC boundary in 2008, after which Alberta undertook separate surveys in that part of the range. The first count following the initial release of 43 animals from the holding paddock in 2002 came from an aerial survey in 2006, where 124 bison were observed; it is believed that 100% of the range was covered (Rowe and Backmeyer 2006). An aerial survey in BC in March 2009 resulted in a count of 156 bison (Thiessen 2010). The most recent count was in March 2010 and resulted in 181 bison (C. Thiessen, British Columbia Ministry of Environment, pers. comm.). During surveys, animals were classified to calf, yearling, 2+ year bulls, and 2+ year cows according to horn morphology and body size (Rowe and Backmeyer 2006, Thiessen 2010). The proportion of adults (≥ 2 yrs) observed were 71% in the 2006 survey (Rowe and Backmeyer 2006) and ranged from 64-74% during road-based surveys conducted in 2009-10 (Thiessen 2010). This means there are approximately 116-134 adults, with an unknown proportion of these being mature animals.
A minimum population size estimate from a survey conducted in 2013 in the Alberta portion of the range yielded 121 bison (including 19 calves). First Nations were allotted 14 permits in 2010 to take bison in the southern edge of their range, in an attempt to discourage range expansion and conflicts with industry and agriculture.
Hay-Zama, AB:
The Hay-Zama subpopulation is counted annually using a total count method. Aerial censuses follow standardized transects (Hermanutz and Fullerton 2011) and result in fairly thorough coverage of the range; however, some animals are inevitably missed when they are in dense cover and no sightability correction factor has been applied. Calves are distinguished from adults. Following establishment of the free-ranging herd of 43 in 1993, it increased to about 130 animals in 2000 (Gates et al. 2001a). Since this time, the subpopulation has increased at an average exponential rate of 0.122 since 2000, peaking at a minimum of 652 animals (including 59 calves) in 2008. Annual licensed hunting began in 2008 and the herd decreased to 561 in 2011. A minimum count survey completed in March 2013 yielded 529 individuals, at which time it became clear that a number of animals had starved to death following unusually severe winter conditions. A total of 75-100 bison died during the spring; the final population estimate for 2013 was 410 (Government of Alberta 2013a). Number of mature individuals is unknown. In prior years hunting had stabilized population size, but the 2013/14 Hay-Zama bison hunting season was suspended (Government of Alberta 2013a).
Mackenzie, NT:
This subpopulation was censused on an approximately biannual basis from 1964 to 1998. Prior to 1989, total count aerial surveys were conducted; a sightability correction factor was used from 1989 for animals in forested habitats in a stratified sampling design (Larter et al. 2000). The subpopulation peaked at about 2,400 bison in 1989 (Larter et al. 2000) and declined from then until 2008 by 35% and from 1998 and 2008 by 18.5%. The latter survey estimated 1,555 bison (95% confidence intervals = 1240-1870), based on aerial quadrat counts of stratified high and low population density blocks (T. Armstrong, Northwest Territories Department of Environment and Natural Resources, pers. comm.). A 2012 aerial survey derived an estimate of 1,531 bison (90% C.I.=1,160 – 2,020; Armstrong 2013). Larter et al. (2000) and Larter and Allaire (2007) reported the results of summer sex and age classification surveys from 1984-1998 and 1999-2006, respectively, as the number of calves and yearlings per 100 adult females ≥2 yrs. Classification surveys were conducted separately from population surveys. In 1993, about 21% were calves or yearlings, and 69% were females ≥ 2 years and males ≥ 7 years (Gates et al. 1995).
Wolf predation, flooding, and forage availability were likely responsible for much of the decline observed between the 1980s and late 1990s (Larter et al. 2000). However, in the summer of 2012, Mackenzie bison experienced a major outbreak of anthrax where 440 bison were confirmed dead over an 8 week period (Government of NWT 2013). Unlike many of the past outbreaks of this disease in bison populations (Wood Buffalo National Park, Slave River Lowlands, Mackenzie) where mature males were over-represented in the mortalities, preliminary findings point to a broader representation of all age/sex classes (B. Elkin, pers. comm.). The population estimate from the survey conducted in 2013 after the outbreak was 714 bison (90% C.I.= 499-1,022) (Armstrong 2013). This represents an estimated 53% decline in one year, and an overall decline in total number of individuals of 70% since 1989 (almost three generations). Applying estimates from Gates et al. (1995), there are approximately 344-705 mature individuals in this subpopulation.
Greater Wood Buffalo, AB, NT:
The Greater Wood Buffalo National Park meta-population is assessed as a single population unit. Differences in the extent, timing and methods used to census bison in the different subunits reduce certainty in population estimates and trends. Aerial surveys of bison in Wood Buffalo National Park began in 1947 using strip line transects (Fuller 1950). From the 1930s to the 1970s the estimated number of bison in Wood Buffalo National Park was 10,000-12,000, stabilized by management interventions such as wolf poisoning and trapping, field slaughters and roundups for vaccinations (Fuller 2002; Bradley and Wilmshurst 2005). Beginning in the 1970s intensive management of predators and disease was discontinued and the population in the park began a long-term decline, reaching an estimated low of about 2,200 animals in 1999. The role of disease in the decline is a matter of debate (Joly and Messier 2004b, Bradley and Wilmshurst 2005).
The history of aerial census methods and results from 1971-2003 within Wood Buffalo National Park was reviewed by Bradley and Wilmshurt (2005). The most recent census was conducted in 2009 when the number of bison was estimated at 4,958 (90% CI=4,189 - 5,727; Vassal and Kindopp 2010). The estimated population size has fluctuated dramatically over a period of time just over three generations (~1984-2011) but current population estimates are similar to those in the mid-1980s (~5,000 bison; Joly and Messier 2004b). The 2010 estimate includes 2009 census counts from the Slave River Lowlands outside the park in NWT (Armstrong and Cox 2011). Number of mature individuals was not reported (Vassal and Kindopp 2010).
Small groups of bison also range adjacent to the western and southern borders of Wood Buffalo National Park and are considered part of the Greater Wood Buffalo National Park meta-population. A census of the Ronald Lake herd was conducted in a 640 km2 area in February 2010 using mark-resight methodology yielding an estimate of 101 (90% CI=74 -159) (Powell and Morgan 2010). A total of 186 individuals were sighted in a 2013 survey (Government of Alberta 2013b). The number of bison in the Wentzel Lake area was last counted at 200 individuals (Government of Alberta 2011), and there are an additional 11 animals currently residing in the Wabasca River area west of Wood Buffalo National Park (Government of Alberta 2013a). The proportion of adult or mature animals was not reported in the latter surveys.
In aggregate, the approximate current estimate of Greater Wood Buffalo National Park meta-population, including subunits that were surveyed independently between 2011-2013, is 4,586 - 6,124 bison (Table 3). While the meta-population has fluctuated over the past ~ 27 years, the current estimate is close to what it was three generations ago.
| Subpopulation (source)Footnotea.1 | Jurisdictions | Est. range (km2) | Most recent total pop. est. (year) | Earliest pop. est. (year) | Est. mature ind. | Trend (%) (# years)Footnotef |
|---|---|---|---|---|---|---|
| Wood Buffalo N.P.Footnoteb.1 | AB & NT | - | 4,189 - 5,727 (2009) | - | - | - |
| Wentzel/Wabasca | AB | - | 211 (2013) | - | - | - |
| Ronald Lake | AB | - | 186 (2013) | - | - | - |
| Greater Wood Buffalo (1) | AB & NT | 58,000 | 4,586 - 6,124 | ~5,000 (1984) | 3,164-4,226Footnotec.1 | ~0 |
| Elk Island N.P. (2) | AB | 58 | 300 (2013) | - | 207Footnotec.1 | |
| Mackenzie (3) | NT | 21,000 | 499-1,022 (2013) | 2,400 (1989) | 344-705Footnoted | -70 (24) |
| Nahanni (4) | NT & BC | 11,700 | 218-644 (2011) | 160 (1998) | 150-444Footnotec.1 | +169 (13) |
| Aishihik (5) | YT | 11,000 | 1,106-1,385 (2011) | 350 (1998) | 763-956Footnotec.1 | +257 (13) |
| Nordquist (6) | BC | 1,400 | 117 (2010) | 50 (2002) | 84-95Footnotee | +134 (8) |
| Etthithun (7) | BC & AB | 5,000 | 181 (2010) | 43 (2002) | 116-134Footnotee | +321 (8) |
| Hay-Zama (8) | AB | 9,000 | 410-420 (2013) | 130 (2000) | 283Footnotec.1 | +219 (13) |
| Chitek Lake (9) | MB | 3,800 | 225-275 (2009) | 35 (1995) | 155-190Footnotec.1 | +600 (14) |
| Total | - | 121,480 | 7,642 - 10,458 | - | 5,213 - 7,191 | - |
Elk Island National Park, AB:
Elk Island National Park, AB: Similar to Elk Island Plains Bison, Wood Bison are counted on a semi-annual basis during handling in facilities to remove surplus animals from the population. Sex and age structure information is collected on a subset of animals during penning; sex and age structure was assessed while animals were penned in March 2013 (M. Allen, Parks Canada, pers. comm.). Winter ungulate aerial surveys are flown in every year, yielding minimum counts of bison. Following a 2009 Ungulate Options Analysis to address management of the high densities of bison, elk, moose, and deer in the 194 km2 park, the Wood Bison population goal was lowered to a recommended adult population of 260-300 (PCA 2009). The 2013 Wood Bison estimate was 300 (M. Allen, Parks Canada, pers. comm.).
Chitek Lake, MB:
The Chitek Lake herd increased from 35 animals in 1995 to 50 by late 1996. In 2000, the subpopulation was estimated at 70 (Gates et al. 2001a). The most recent aerial survey was in 2009, when minimum count of 208 bison were observed across a portion of the range (B. Joynt, Manitoba Conservation, pers. comm.). Population size was estimated at 225-275, indicating an average annual increase of 12.7% per year since its establishment. Number of adults or mature individuals are unknown. In recent years, however, deep snows and extensive flooding are believed to have reduced the size of the herd (B. Joynt, Manitoba Conservation, pers. comm.). It is believed that the Chitek subpopulation has reached the carrying capacity of its range and range expansion into agricultural areas to the south is considered undesirable (B. Joynt, Manitoba Conservation, pers. comm.).
Summary: Wood Bison:
The total number of Wood Bison in the Canadian subpopulations included in the DU increased from ca.1,827 in 1987 (Ruckstahl 2000) to 6,150 in 2000 (Gates et al. 2001a) to 7,642-10,458 in 2013 (Table 3). Taking the mid-point of the latter estimate (9,050), the Wood Bison population has increased about 395% over the last three generations (24 years) and 47% since just after the last COSEWIC assessment 13 years ago. Much of the increase in numbers has been a result of adding animals to supplement one subpopulation and the reintroduction of animals to establish five new subpopulations. One of the newly established subpopulations (Etthithun -2002) was reintroduced after the last COSEWIC status report. However, the current Wood Bison population is still at <5% of its presumed original level and only three subpopulations currently number >500 individuals. Two subpopulations experienced significant mortality events in 2013. Following a severe winter ca. 20% of the Hay-Zama subpopulation died from starvation and the Mackenzie population decreased an estimated 53% in one year following an outbreak of anthrax. About 60% percent of the DU occurs in the Greater Wood Buffalo meta-population, which is infected with bovine tuberculosis and brucellosis and for which regional control measures are in place to mitigate the risk of these diseases spreading to other subpopulations. Information on age structure of most subpopulations is unknown; however, applying an overall estimated proportion of 69% from Gates et al. (1995) to those subpopulations where field data are unavailable yields an approximate overall number of mature individuals of 5,213 - 7,191 for the Wood Bison population (Table 3).
Rescue Effect
Although several wild Plains Bison subpopulations occur in the United States, they provide no opportunity for natural reestablishment of Canadian subpopulations. The closest free-ranging Plains Bison subpopulation is Yellowstone National Park, about 450 km south of the Canadian border. Although Plains Bison may have moved this distance in historical times, no modern movements of this magnitude have occurred. Moreover, bison would not be allowed to traverse the current landscape at any great distance, through cattle ranching country. Plains Bison in Yellowstone National Park are infected with brucellosis, and animals migrating outside of this protected area are subject to containment measures (Fuller et al. 2007, Bidwell 2009), such that diseased Plains Bison moving naturally into southern Alberta would likely be removed. The nearest disease-free subpopulation of Plains Bison outside of Canada is near Delta Junction, Alaska, about 230 km from the Yukon border and about 2,000 km from the northern extent of the original range of Plains Bison. These bison would likely be removed because of the threat they would pose to Wood Bison conservation in the Yukon.
The original range of Wood Bison once extended into Alaska, but all extant wild Wood Bison currently reside in Canada. Therefore, there is no possibility of rescue effect. Moreover, movements from the largest subpopulation (Greater Wood Buffalo National Park metapopulation) to the nearest-neighbour Wood Bison ranges (Mackenzie or Hay-Zama) are actively controlled against (see Threats).
Threats and Limiting Factors
Limiting Factors
Small Population Size, Genetic Diversity, and Inbreeding Depression
American Bison underwent an extreme bottleneck, from tens of millions to hundreds of animals, in the 1800s. Between 1873 and 1904, fewer than 500 bison from six captive herds and two remnant wild herds (Yellowstone and Wood Buffalo) served as the foundation stock for all Wood and Plains Bison that exist today (Halbert et al. 2005). The current population likely harbours only a small remnant of the species’ original genetic diversity (Wilson and Zittlau 2004).
All but the Greater Wood Buffalo National Park (GWBNP) meta-population of Wood Bison were founded with small numbers of animals. The founder effect and/or genetic drift have affected the levels of genetic diversity in all Wood Bison subpopulations except GWBNP (Wilson and Strobeck 1999, Wilson et al. 2005, McFarlane et al. 2006). Olech (1987) showed how the reproductive success of European bison decreased as inbreeding increased. In Texas, Halbert et al. (2004) chronicled the observed and predicted effect of low levels of genetic diversity on an isolated subpopulation of Plains Bison. Low genetic diversity can limit a population’s ability to adapt to environmental change and can lead to inbreeding depression (Wilson and Strobeck 1999; Halbert et al. 2004).
Hedrick (2009) and Gross et al. (2010) indicated little concern for small population effects on genetic drift and diversity reduction in bison subpopulations greater than 1,000. Subpopulations numbering fewer individuals, however, may experience genetic problems given the polygynous nature of bison and concomitant reductions in effective population sizes. Only two subpopulations of Wood Bison (Aishihik and GWBNP meta-population) and one of Plains Bison (Pink Mountain) are currently >1,000 animals, the minimum safeguard against small population effects on genetic drift and diversity suggested by Hedrick (2009) and Gross et al. (2010). Two of the subpopulations of >1000 animals (Aishihik and Pink Mountain) are currently managed to reduce herd size, while the third (GWBNP) is being managed for geographic expansion due to disease concerns.
No measurable effects of the loss of genetic diversity or inbreeding depression have been observed in Plains or Wood Bison populations in Canada (Wilson and Zittlau 2004). However, the only Canadian Plains Bison subpopulations assessed for genetic diversity were from Elk Island and Wood Buffalo National Parks (Wilson and Strobeck 1999), which were surprisingly high. The level of genetic diversity in the Sturgeon River and McCusker River subpopulations is unknown, but these subpopulations originated from a small number of founders and they have remained reproductively isolated. Low number of founders, small population size, and reproductive isolation, is a cause for concern for Plains Bison. The level of threat from loss of genetic diversity through genetic drift is likely low to moderate for both Plains Bison and Wood Bison in the short-term (i.e. next 3 generations).
Threats
The threat classification below is based on the IUCN-CMP (World Conservation Union-Conservation Measures Partnership) unified threats classification system (Master et al. 2009). Threats were assessed separately for Plains and Wood Bison. Results on the impact, scope, severity and timing of threats are presented in tabular form in Appendix 1 for Plains Bison and Appendix 2 for Wood Bison. The overall calculated and assigned threat impact is Very High for Plains Bison and High for Wood Bison. Narrative descriptions of the threats are provided below in the general order of highest to lowest overall impact threats for both designatable units, although each threat does not have the same impact on Plains and Wood Bison.
High-Very High Impact
Hunting and Population Control (5.1)
Historically, overhunting was a major threat to wild bison, nearly resulting in their extinction by 1900 (Isenberg 2000). Currently, hunting is allowed for three of five Plains Bison subpopulations (Pink Mountain, McCusker River, and Sturgeon River) and 6-7 of nine Wood Bison subpopulations (Aishihik, Etthithun, Hay-Zama, Nahanni, Chitek, and portions of the Wood Buffalo meta-population [i.e. Slave River Lowlands]). A hunting ban of the Mackenzie subpopulation was instated in 2012-2013, following a major anthrax mortality event. Hunting is used as a management tool to limit population growth and range expansion, and reduce bison-human conflicts. Aboriginal groups also take these animals for food and cultural purposes. Non-aboriginal hunting is regulated for all subpopulations, except in Alberta outside of some bison management or control zones and the Slave River Lowlands where hunting by aboriginal people is unrestricted.
Today, direct take of bison through hunting or culling often occurs as a means of controlling the size and extent of individual subpopulations, which is generally dictated by social carrying capacity, or local acceptance and tolerance limits for wild bison. This threat has a Very High impact for Plains Bison, and High Impact for Wood Bison, with a high possibility of leading to reductions in some subpopulations over the next 10 years (Appendix 1; 2). For example, risk intolerance for the potential of bovine tuberculosis and brucellosis spreading from Wood Buffalo National Park to uninfected wild bison subpopulations and to livestock has generated policies and programs to reduce these risks. Colonization of unoccupied habitats near diseased herds is strongly discouraged or prohibited through bison control areas, as is connectivity among adjacent subpopulations. A large bison control area is actively surveyed in the NWT, and unregulated hunting of bison in that area and adjacent to Wood Buffalo National Park in Alberta limits movements and growth of bison populations in these areas. One actively debated option for eradicating disease from the GWBNP meta-population has been to depopulate and replace it with known healthy stock from Elk Island National Park (Shury et al. 2006; Nishi 2010). Increases in number and distribution of the Hay-Zama subpopulation has been actively discouraged in order to protect animals from exposure to disease. The Government of Alberta (2011:3) stated in reference to disease infection: “If this happened, there is a strong probability that the whole herd would have to be culled.”
The recently released management plan for Wood Bison in the Yukon (Government of the Yukon 2012) calls for limits to both population size and distribution. It identifies exclusion zones in areas where bison conflict with community values or are a road hazard. The 5-year management goal for the Aishihik subpopulation is to reduce numbers by approximately 19% (Government of Yukon 2012). Beginning in 2008, the Hay-Zama subpopulation was reduced from about 700 to about 400 animals, and the management goal is to use hunting to keep the subpopulation between 400-600 animals. Altogether, 1,426 and >500 Wood Bison have been taken from the Aishihik and Hay-Zama subpopulations, since hunting began in 1998 and 2008, respectively (Government of Alberta 2011; Government of Yukon 2012).
Among Plains Bison, the McCusker River herd is exposed to unregulated hunting. Hunting pressure will likely increase with public access as the area adjacent to the Cold Lake Air Weapons Range is subject to increasing industrial development (forestry, and oil and gas). Neither population size nor the number taken are monitored (R. Tether, Saskatchewan Ministry of Environment, pers. comm.). The Sturgeon River subpopulation is subject to removals if the number of animals moving onto agricultural lands near the park exceeds acceptance by the landowner community. Overhunting (especially females) has been cited as a key reason behind the current declining trend of this subpopulation (S. Cherry, Parks Canada, pers. comm.). The size and distribution of the Pink Mountain herd is managed by regulated hunting to confine it within a management area and target population range. Elk Island National Park limits the size of its Plains and Wood Bison subpopulations through biennial removals (primarily sales). In addition, bison posing a risk to public safety are occasionally culled.
High-Medium Impact
Invasive and Other Problematic Species (Disease) (8)
Bison are affected by a wide array of pathogens for which the pathobiology and epidemiology are very similar to cattle (Tessaro 1989). The presence of reportable diseases may result in culls of infected herds and herds considered at risk of becoming infected (Nishi et al. 2002a, Gates and Ellison 2010, Nishi 2010). If reportable diseases were to spread to uninfected populations, this will reduce local acceptance of Plains or Wood Bison.
Comprehensive reviews of bison diseases and disease management have been published by Reynolds et al. (2003) and Aune and Gates (2010). The diseases (considered separately in Appendix 1 and 2 as “problematic native species” [8.2] and “invasive non-native species” [8.1]) that have caused the most significant mortality events (due to the disease itself or the management responses) are discussed below.
Anthrax is an infectious bacterial disease caused by the endospore-forming bacterium Bacillus anthracis (Dragon and Rennie 1995, Gates et al. 2001b). It can remain dormant in the soil for long periods of time and causes sporadic outbreaks and mortality. A recent study of phylogeographic patterns of anthrax supports the hypothesis that anthrax in northern Canadian bison originated in Asia and was transported by early human migrants across the Bering Land Bridge into Beringia (Kenefic et al. 2009). The form occurring in outbreaks in bison in Canada has been considered an indigenous species (Ferguson and Laviolette 1992, Gates et al. 2001a,b, Reynolds et al. 2003, Aune and Gates 2010). Humans have, however, played an important role in its proliferation and dispersal (Dragon et al. 1999, Aune and Gates 2010).
After inhalation or ingestion by a susceptible host, endospores germinate and the vegetative form of the bacterium replicates in the bloodstream, releasing toxins that cause septicaemia and death (Dragon and Rennie 1995). Upon release from a carcass, the highly resistant endospores can remain viable in the soil for decades before infecting a new host (Dragon and Rennie 1995, Dragon et al. 2005). Climatic factors, such as season of year, ambient temperature, and drought have a role in promoting anthrax epizootics (Gates et al. 2001b). Outbreaks during summer months occur after flooding, followed by high ambient temperatures, and drought (Gates et al. 2001b). The roles of environmental factors such as soil types and soil disturbances via excavation are poorly defined despite attempts to evaluate these potential factors (Dragon et al. 2005).
Outbreaks in wild bison in Canada have been periodically reported in the Slave River Lowlands and Wood Buffalo National Park (Gates et al. 2001b, Nishi et al. 2002b, Nishi et al. 2007), the Mackenzie Bison Range (Gates et al. 1995), and Prince Albert National Park (Shury et al. 2009). Between 1962 and 1971, anthrax and the associated depopulation and vaccination programs employed to control the disease, were responsible for 2,800 wood bison mortalities (Dragon and Elkin 2001), with several outbreaks occurring in the same areas in subsequent years (Gates and Aune 2010).The highest percent mortality reported in wild bison occurred in the Mackenzie subpopulation in 2012: 440 carcasses were found in an 8-week period, but this was likely an underestimate of the true number of mortalities (B. Elkin, NWT Department of Environment and Natural Resources, pers. comm.). A population count conducted after the outbreak indicated that the subpopulation had been reduced by 53% (see Population Sizes and Trends).
Although it can be devastating, the disease is only likely to emerge in certain environmental conditions, and in accordance with its historical distribution where there are concentrations of endospores. The ecological niche has been modelled for the U.S. (Blackburn et al. 2007), but not for Canada. In addition to the two Wood and one Plains Bison subpopulations that experience anthrax outbreaks, Grasslands and McCusker River subpopulations in Saskatchewan also may be vulnerable.
Bovine brucellosis and tuberculosis. The translocation of Plains Bison into Wood Buffalo National Park in the 1920s (see History of Decline and Early Restoration of American Bison in Canada) was associated with the introduction of bovine brucellosis (Brucella abortus) and tuberculosis (Mycobacterium bovis; Tessaro 1988, Joly and Messier 2004a) to Wood Bison of the GWBNP meta-population.
Bovine brucellosis is an introduced exotic cattle disease that occurs in the GWBNP Wood Bison meta-population. The disease agent is the bacterium Brucella abortus, which primarily causes pathology of the reproductive tract, including abortion. It can also cause bursitis and epididymitis (Tessaro 1989). Transmission is primarily through direct contact with infectious uterine fluids, aborted fetuses, or food, water or soil contaminated by those fluids (Thorne et al. 2001). In an experimental herd Brucella infection results in > 90% abortion rate for first pregnancies (Davis et al. 1990, 1991). Arthritis may also be caused by the disease, resulting in increased susceptibility to predation (Tessaro 1988).
Bovine tuberculosis, caused by the bacterium Mycobacterium tuberculosis, is another exotic cattle disease of bison occurring in bison in the GWBNP meta-population. It is primarily a respiratory disease and is mainly transmitted among herd members by aerosol. It can infect any organ and is evident as abscesses or very small lesions. Tuberculosis also adversely impacts fecundity and survival (Joly and Messier 2004b, 2005). The disease may reduce fertility, weaken infected animals and predispose them to predation, and in advanced cases result in death (Tessaro 1989).
Joly and Messier (2004a) found prevalence rates of tuberculosis and brucellosis to be 49% and 31% respectively in the GWBNP meta-population. Bovine tuberculosis and brucellosis may adversely affect population growth, particularly when combined with wolf predation. Tessaro (1988) estimated that advanced tuberculosis may result in 4-6% mortality in bison; likely as a result of increased susceptibility to predation by wolves. Joly and Messier (2005) found that bison in Wood Buffalo National Park that were positive for both diseases were less likely to be pregnant or to survive the winter than bison positive for one or neither disease. However, there are conflicting interpretations about the impact of the two diseases and synergism with predation with regard to the population dynamics of the GWBNP meta-population (Carbyn et al. 1993, Carbyn 1998, Joly and Messier 2004b, Bradley and Wilmshurst 2005).
Management interventions to contain the geographic extent of these diseases and to prevent them from spreading to adjacent unaffected wild bison herds severely limits recovery potential for Wood Bison (Joly and Messier 2004b, Joly and Messier 2005). Colonization of large tracts of suitable, but unoccupied, habitat is actively discouraged as a means of containing diseased bison of the GWBNP meta-population (Gates et al. 2001a). In addition, population augmentation or the introduction of new animals to establish gene flow to isolated populations is also severely constrained by the potential for introducing these diseases to otherwise unaffected subpopulations. Neither disease has been detected in other wild bison subpopulations in Canada.
Malignant catarrhal fever (MCF) is a typically fatal disease in bison. Sheep-associated MCF (Ovine 2799 herpes virus type 2) does not cause disease in its natural host, but it causes highly lethal infections in bison (Heuschele and Reid 2001), with mortality rates of up to 100% (Schultheiss et al. 2000). In the U.S. it is common in domestic goats (61%) and sheep (53%) (Li et al. 1996). MCF is expressed in two forms, acute and chronic, but regardless, infections proceed rapidly to clinical disease and death ensues in most cases. Some animals recover and remain persistently infected (Schultheiss et al. 1998). Direct contact between bison and domestic sheep is the most likely source of infection. Three of the five wild Plains Bison subpopulations occur in agricultural areas where livestock production is prevalent (Elk Island National Park, Sturgeon River, Grasslands National Park). Grazing of sheep and goats within 5 km of these herds poses a risk of infection with MCF (Schultheiss et al. 2000).
Other cattle-borne pathogens are potential major threats to American Bison. For example, Johne’s disease is an emerging disease of concern for bison conservation. A chronic infection of the gastrointestinal (GI) tract that is fatal, this disease results in severe inflammation of the GI tract, compromising its ability to absorb nutrients. The causative agent of Johne’s disease is Mycobacterium avium subspecies paratuberculosis (MAP; Forde et al. 2013), which is shed from infected animals through their milk or feces. There is often a long sub-clinical phase of up to 10 years where animals infected with MAP show no clinical signs of Johne’s disease. MAP is usually introduced to a population through the arrival of infected individuals, and can spread quickly, particularly in high-density populations. MAP is a hardy organism and difficult to eradicate once established in a host population. There is no known cure or treatment for bovine Johne’s disease. More widely known to affect cattle and sheep, MAP was recently reported by Ellingson et al. (2005) and Sibley et al. (2007) in Plains Bison and Wood Bison, respectively. Subsequently, Forde et al. (2013) sampled all nine subpopulations of Wood Bison considered here for assessment and found MAP present in all of them. Prevalence in Plains Bison subpopulations in Canada is unknown, but may also be 100%. However, it is not known if MAP in bison leads to Johne’s disease, as in cattle and sheep, and no animals have been observed in the wild with clinical signs.
Another significant emerging infectious disease for bison is Mycoplasma bovis, a bacterial pathogen that causes respiratory disease (pnuemonia) and arthritis. In multiple recent outbreaks among commercial bison in the U.S. and Canada, significant mortality has ensued (Woodbury and Windeyer, 2012); 53 of 194 animals, including 45.5% of adult cows were lost to this disease from one herd (Janardhan et al. 2010). Other diseases of conservation concern for American Bison are reviewed by Aune and Gates (2010).
Climate Change and Severe Weather (11)
There have been numerous instances where unusual weather conditions have led to large bison mortality events, particularly for Wood Bison. Historically, thousands of bison were drowned in floods that resulted from the spring melting of large snow packs, and winters characterized by deep snow and ice crusting warming periods were blamed for major die-offs (Dary 1989, cited in Gogan et al. 2010). In contemporary times, occasional mass drownings when groups of bison attempt to cross thin ice in the spring or following spring floods are periodically recorded (Larter et al. 2003).
Incidences of drowning may affect all bison subpopulations at all times of the year; these events are stochastic in nature. During high water flows in the spring and early summer (May to July) bison are sometimes swept away and drowned (Larter and Allaire 2007). Bison become easily trapped once they break through lake or river ice (Carbyn et al. 1993). Reynolds et al. (2003) noted that some bison likely drown each year, as a result of spring flooding, or falling through thin ice. During both fall freeze-up and spring breakup periods, animals attempting to cross rivers or lakes can break through this ice and drown (Larter et al. 2003). Drowning may also be the result of bison being swamped in the wake of large boats on some rivers (i.e. Liard River). Massive mortality events due to drowning have occurred in several Wood Bison subpopulations. For example, a 1974 flood in the Peace-Athabasca Delta of Wood Buffalo National Park killed at least 3,000 bison. Water levels were already high and individuals were weak due to much of their winter range being frozen over (Haynes 1988). In another year, 177 MacKenzie animals drowned after breaking through the spring ice of Falaise Lake (Gates et al. 1991). The Nahanni Bison use both sides of the Liard River Valley; hence swimming across the river is an important component of the ecology of this subpopulation (Larter et al. 2003, Larter and Aillaire 2007). Flooding has reduced habitat in southern Wood Buffalo National Park and the Mackenzie range causing shifts in bison distribution and impacts on population dynamics.
Relatively high foot loading and low chest heights render bison less morphologically adapted to deep snow than other temperate ungulates, such as moose, elk or deer (Telfer and Kelsall 1979, 1984). Moreover, whereas bison rely little on browse during deep snow years (unlike cervids; Telfer and Kelsall 1979), increasing snow depth can decrease their ability to obtain adequate forage, potentially leading to death from starvation. Mature bison have the highest foot loadings, perhaps making them the most vulnerable segment of the population to the deleterious impact of deep snow (Telfer and Kelsall 1979). Deep snow may affect the nutritional status of bison, as well as impede travel and make them more susceptible to predation by wolves than ungulates more adapted to deep snow (Telfer and Kelsall 1984). In years with deep snow conditions, bison mortality increases and calf production and/or survival declines. For example, death from starvation following severe winter conditions was responsible for 75-100 deaths of Hay-Zama Wood Bison in 2013, representing one-fifth of the subpopulation (see Population Sizes and Trends). Calf composition of the Aishihik subpopulation was reduced by about 25% after a deep snow year in winter 2008/2009, when cows were observed to be in poor condition, and there was a recent die-back following a hard winter in the Chitek subpopulation (see Population Sizes and Trends).
Woodhouse et al. (2002) and Isenberg (2000) have suggested severe regional droughts in the mid-19th century contributed to bison declines. Drought may be an issue for Plains Bison in Grasslands National Park where water courses and wetlands are a very small component of this mixed-grasslands landscape.
Climate change has the potential to increase the size and frequency of forest fires (Flannigan et al. 2009, McCoy and Burn 2005), but fire suppression may mitigate these effects. Hence the potential for habitat creation for bison in early seral stages following fire is uncertain. Impacts of fire on forests may benefit the three Plains Bison subpopulations (Pink Mountain, McCusker River, and Sturgeon River) occurring in forested regions.
Agriculture (2)
The potential for recovery of Plains Bison has been substantially reduced by agricultural settlement and land conversion in the grasslands and parkland areas of the Prairie Provinces (see Habitat Trends). The increase in the size and distribution of privately owned captive commercial bison herds in the western provinces also reduces the availability of otherwise suitable rangelands for wild Plains Bison and Wood Bison restoration. Habitat loss from conversion of native range to croplands is still occurring and the most significant threat is from the sale of public rangelands (e.g. Prairie Farm Rehabilitation Administration lands) for farming, particularly in Saskatchewan (Arbuthnott and Schmutz 2013). Wild bison conflict with crop production where they overlap. All Wood Bison subpopulations are affected by expanding agricultural developments, especially the Chitek Lake, Hay-Zama, and Etthithun Lake subpopulations, and the Greater Wood Buffalo National Park meta-population.
The perception of competition with other agricultural interests adjacent to, or within, prospective prairie landscapes that are suitable habitat for bison adds an important dimension to this threat or impediment to recovery (Aune and Wallen 2010). Grazing of livestock and privately owned bison on public lands is a threat to wild Wood Bison for several reasons including exclusion of wild bison from grazing dispositions, intolerance of wild bison, and cattle gene or modified domestic/captive genotype introgression from commercial bison if they intermingle with wild bison (see below). Grazing livestock and commercial bison operations preclude wild bison restoration on some suitable public land. This affects all Plains Bison subpopulations and wild Wood Bison in the Aishihik, Hay-Zama and Chitek Lake subpopulations.
Medium-Low Impact
Introduced Genetic Material (8.3)
Hybridization among Bos species around the world is well-known. Bison and cattle (Bos taurus) are not known to produce hybrids naturally, but were deliberately crossed during the late 1800s for the purposes of creating a heartier beef animal. This experiment was considered a failure and abandoned after 20 years (Coder 1975), and it is uncommon to deliberately cross the two species today, particularly in Canada (Singer 2005). Nevertheless, because the practice was most common at the time when the North American bison population was at its nadir, the genetic legacy is evident today in Plains Bison.
Early published tests of wild Canadian Wood or Plains Bison populations did not detect mtDNA or microsatellite markers attributable to introgression by cattle genes (Polzhein et al. 1995, Ward et al. 1999, Wilson and Strobeck 1999), but the ability of earlier tests to detect this were somewhat limited. The four public Canadian subpopulations included in Halbert et al. (2005)--Elk Island National Park Plains and Wood, MacKenzie, and Wood Buffalo National Park--had no evidence of either mitochondrial or nuclear domestic cattle introgression. Most wild Canadian bison subpopulations were established from Elk Island National Park bison, one of the few sources of American bison germplasm free of domestic cattle introgression (Halbert et al. 2005, Hamilton et al. 2009). In the U.S, genetic introgression from hybridization with cattle has been identified in many modern Plains Bison subpopulations and is nearly ubiquitous among commercial herds (Halbert and Derr, 2007). All 63 individuals tested from the Old Man on His Back Plains Bison herd in Saskatchewan (founded from Elk Island National Park animals) were found to be free of detectable domestic cattle introgression, in contrast to seven privately owned U.S. herds from the same study (Hamilton et al. 2009). Many privately owned bison operations get their stock from Elk Island National Park surplus animals, but similar to U.S. ranches, animals come from elsewhere as well (Singer 2005). Results of genetic testing of private commercial herds in Canada are not publicly available.
Derr et al. (2012) found that bison with cattle mtDNA were consistently smaller (height and weight) across all populations sampled, even after accounting for different environmental conditions. Male bison with cattle mtDNA showed a larger relative reduction in body size than female bison with cattle mtDNA, with the possible fitness consequence of reduced lifetime reproduction (Hedrick 2010). Hence, genomic integrity is important for conservation of wild-type bison. Moreover, there is potential for genetic introgression into wild populations from bison that escape captive commercial herds carrying cattle genes.
The introduction of up to 6,673 Plains Bison from Wainwright, Alberta, into Wood Bison range in Wood Buffalo National Park during the 1920s resulted in hybridization between the two (Fuller 2002; Bradley and Wilmshurst 2005). Hybridization between Wood Bison, and wild Plains Bison or Plains Bison that escape from farms is a threat in some areas. The Pink Mountain subpopulation of Plains Bison is within the original range of Wood Bison and poses a low threat of hybridization with Wood Bison from the Etthithun and Nordquist subpopulations. The management plan for Wood Bison in British Columbia establishes a Bison Control Area to reduce the threat of hybridization by the Pink Mountain subpopulation of Plains Bison (Harper et al. 2000). The effectiveness of this control zone, however, is unknown. Moreover, there are hundreds of bison farms in the Peace River basin in Alberta and adjacent British Columbia. Escapes from these ranches are common and not all escapees are captured. For example, there are three to five bison farms within 80 km of the Hay-Zama subpopulations; one or two escapes occur each year in northwestern Alberta, with a few bison escaping at each event (D. Moyles, Alberta Sustainable Resource Development, pers. comm.). Commercial bison, most of which are Plains Bison, have unknown and variable levels of cattle gene introgression and some may pose a threat to the genetic integrity of wild bison (Singer 2005).
Low Impact
Roads and Railroads (4.1)
Bison are attracted to linear features. Road verges are often treated with agronomic seed mixes that provided high-quality forage, luring and holding bison to road corridors, risking collisions. Bison struck by vehicles are injured or killed, property damage is considerable and human injuries can be serious. Bison use the roadsides as foraging habitat and the roads themselves as travel corridors, particularly in winters with deep snows that impede travel adjacent to the road. Bison are dark-bodied and tend to be particularly difficult for motorists to see during the night (Thiessen 2010). Collisions with motor vehicles may be a source of significant mortality for those bison subpopulations that incorporate linear road corridors into their seasonal or annual home ranges (COSEWIC 2012). The safety hazard posed by bison on highways is also a limiting factor for local bison restoration because it decreases the willingness of local communities to allow the growth of reintroduced subpopulations (e.g., Government of Yukon 2012). Increased public access combined with unregulated hunting means that roads present a very high impact potential for Wood Bison.
Road mortality is of particular concern for the Nordquist Wood Bison subpopulation (COSEWIC 2012). The range of this subpopulation is closely tied to the Alaska Highway and an average of about 15 wood bison are killed per year, mostly by commercial trucks traveling at night (D. Fraser, British Columbia Ministry of Environment, pers. comm.) In heavy snow conditions during a severe winter in 2005/2006, Wood Bison walked on the cleared roadway, resulting in 32 deaths in 2005, and 17 in 2006 (Rowe 2007). It is likely that highway mortality is the most significant threat to this small subpopulation.
Highway 3 in the Northwest Territories runs through the range of the Mackenzie subpopulation and has also become a significant source of mortality for this subpopulation (Nishi 2004). Between 1989 and 2009, 175 wood bison were struck and killed by motor vehicles on Highway 3 (NWT Environment and Natural Resources 2010). Collisions on Highway 3 are increasing, likely due to increased traffic volume and speeds. In recent years, the Hay-Zama subpopulation averaged about 10 wood bison killed per year in collisions with vehicles on the Zama Road (D. Moyles, Alberta Sustainable Resource Development, pers. comm.). Control actions to mitigate risk involve limiting population abundance and distribution. In Yukon, for example, lethal removal (hunting) of animals distributed near the roadside has greatly reduced the occurrence of bison along the Alaska Highway, despite roadsides providing abundant forage for bison. Roads pose a threat for seven of the nine Wood Bison subpopulations considered in this assessment, specifically: Aishihik, Nordquist, Etthithun, Hay-Zama, Mackenzie, Nahanni, and GWBNP. It is considered overall as a moderate impact threat for Wood Bison.
By contrast, wild Plains Bison subpopulations are not located in high traffic areas and this is not expected to change much in the next 10 years, amounting to a likely negligible impact for this DU. The one exception is the McCusker River subpopulations where road access is expected to increase with industrial development (forestry and oil and gas), which is expected to increase in this range.
Low-Negligible Impact
Housing and Urban Areas (1)
For Wood Bison, housing development is present and increasing slowly near urban centres in the ranges of the Aishihik, Nahanni, Mackenzie, Nordquist, and Hay-Zama subpopulations. Bison are often attracted to residential areas; however, they are not tolerated there, and usually lethally removed. The only Plains Bison subpopulations where residential development may have an impact is the Sturgeon River subpopulation. Agricultural land adjacent to Prince Albert National Park is being sold at an increasing rate for rural residential development. The threat of impact on habitat in locations where bison move out of the park is increasing but negligible at the present time.
Oil and Gas Drilling (3.1)
Oil and gas development is considered of potential concern for the Hay-Zama, Etthithun Lake, and Nahanni Wood Bison subpopulations, and the McCusker River subpopulation of Plains Bison. Conventional oil and gas development typically occupies small areas, although untapped areas (e.g., shale deposits) do overlap with bison range (e.g., Nahanni). Human activities associated with exploration and operation of oil and gas fields has the potential to disturb bison, particularly in hunted populations (COSEWIC 2012). Bison in non-hunted or lightly hunted populations readily habituate to human developments, often causing bison nuisance issues. For instance, bison from the Etthithun Lake and Nahanni subpopulations have been lethally removed from oil and gas development areas where they posed a risk to infrastructure. Increased road access associated with resource development can increase the impact of unregulated hunting. Oil and gas development is extensive in the range of the Wood Bison in northern Alberta and British Columbia. The Ronald Lake subpopulation of the GWBNP meta-population is particularly under threat from increased access and habitat loss from proposed oil sands mine developments. Overall, however, the level of impact on Wood Bison from resource development is considered negligible when considered in terms of the proportion of the overall population.
Natural System Modifications (7)
Two dams exist on the Peace River and a third (Site C) is in the environmental assessment process. The potential for changing water regimes in the basin exists and concern has been expressed about potential effects on flood-driven rejuvenation of meadow and marsh habitat. However, published evidence does not support this concern (Timoney 2002, 2006, 2008b). Rather climate change and periodic oscillations in precipitation and hydrology influence vegetation dynamics. The impact of dams and water management is considered a negligible to low impact threat for Wood Bison habitat because the Peace-Athabasca Delta is dynamic and changes as a result of existing dams on the Peace River have not drastically affected vegetation and landscape cover in the delta at large spatial and temporal scales (Timoney 2002, 2006, 2008b).
Recreational Activities (6.1)
Nature-related public recreation is encouraged in national parks where three Plains Bison subpopulationss occur (Sturgeon River (Prince Albert National Park), Elk Island National Park, and Grasslands National Park). This activity appears to cause minimal disturbance and is likely a negligible threat impact. Snowmobile and off-highway vehicles (OHVs) are a threat to some subpopulations. Bison groups with calves respond the strongest to snowmobile disturbance (Fortin and Andruskiw 2003). In a study of the Sturgeon River subpopulation, GPS-collared bison that fled from snowmobiles increased their daily movement rate by 27-30%. The Pink Mountain and Sturgeon River subpopulations’ range receives use by OHVs and snowmobiles. These are hunted subpopulations and may be more sensitive to disturbance by OHVs than if they were not hunted. Recreational OHV and snowmobile use in the McCusker River subpopulation’s range is prohibited in the Cold Lake Air Weapons Range and is low in the adjacent area in Saskatchewan. However, road access is increasing into the McCusker River subpopulation’s range, which in conjunction with unregulated hunting will increase the impact of hunting. Although bison may be disturbed by OHVs and snowmobiles the threat impact level at the population level is negligible.
All Wood Bison subpopulations are subjected to disturbance by snowmobiling. Responses by bison vary with habituation and exposure to hunting. Disturbance by snowmobiles is a high level concern in hunted populations where bison may be displaced from preferred foraging habitats by hunters using snowmobiles. In northwest Alberta, snowmobiles and OHVs are used by crews engaged in industrial activities rather than recreation.
Negligible-No Impact
With respect to Military Exercise (6.2), the McCusker River range includes a large area in the CLAWR bombing range. There is probably some displacement impact associated with low flights and bombing but this is considered negligible. The benefit from excluding public access to the Cold Lake Air Weapons Range is reducing unregulated hunting pressure. For Wood Bison, there is a high frequency of overflights from resource development traffic and research/monitoring activities affecting several subpopulations (e.g., Nahanni and Wood Buffalo National Park), but there is no evidence of any significant response. Fancy (1982) found only 2 of 59 aerial overflights resulted in a reaction by extra-limital Plains Bison in Alaska, and suggested that they readily habituated to aircraft flying as low as 100-150 m above ground level.
Locations
The five Plains and nine Wood Bison subpopulations considered in this assessment are geographically isolated from one another. Each is managed as a distinct unit by a unique set of agencies and stakeholders. The conservation and recovery of each is greatly influenced (and often constrained) by management decisions directed at the subpopulation (e.g., hunting limits, measures to keep the populations separated from each other, cattle and disease management, and culling). Most of the 14 subpopulations (4/5 Plains and 6/9 Wood) are small (< 500 animals) and limited to relatively small areas where range expansion is controlled.
Recent experience demonstrates that the most severe threats facing Plains and Wood Bison subpopulations tend to occur as events that can rapidly affect all individuals of a given population unit (see Threats), thereby meeting the IUCN definition of “location”. For example, since 2008 alone, outbreaks of anthrax and severe weather events have affected four separate subpopulations, acting swiftly and causing non-discriminate mortality in a short period of time (see Threats and Population Sizes and Trends). Disease or weather events are well-known historically (see Threats) and have the potential to strike any subpopulation today and affect all individuals, albeit with unpredictable levels of mortality.
For Wood Bison, there are nine locations, with the most serious plausible threats being: 1) disease (anthrax in Wood Buffalo and Mackenzie), 2) the management response to brucellosis and tuberculosis spread from its current distribution (Wood Buffalo, Mackenzie, Hay-Zama), or 3) a severe winter or unusual spring weather event leading to mass starvation or drowning (all Wood Bison subpopulations, although with varying likelihood). For Plains Bison, there are five locations, with the most serious plausible threats being: 1) disease (anthrax in Sturgeon River and possibly Grasslands and McCusker River, livestock-borne diseases in all subpopulations other than Pink Mountain), or 2) severe winter or unusual spring weather event or drought in all subpopulations, striking with varying likelihood.
Protection, Status, and Ranks
Legal Protection and Status
Legal protection for bison in Canada began in 1877 with the passing of the Buffalo Protection Act. The act was in response to the precipitous decline in Plains Bison, but it was not well-enforced and free-ranging Plains Bison became extirpated from Canada by the 1880s (Hewitt 1921). However, a small population of Wood Bison persisted and in 1894 the Unorganized Territories Game Preservation Act was passed, partly as a response of the decline in Wood Bison (Lothian 1981). In 1922, Wood Buffalo National Park was established as a measure to prevent the extinction of Wood Bison (Soper 1941, Gates et al. 2001a). The past and present legal status of bison in North America is thoroughly reviewed by Aune and Wallen (2010). COSEWIC’s ATK Assessment Report has concluded that “Multiple Aboriginal communities are either actively engaged or are pursuing Plains Bison and Wood Bison relocation and subpopulation co-management” (COSEWIC 2012).
Plains Bison
Plains Bison currently receive no legal protection under SARA, and they are not listed as a species at risk in any province or territory. Plains Bison are legally classified as wildlife in the provincial wildlife acts of British Columbia and Saskatchewan. They are not wildlife under provincial wildlife acts in Alberta or Manitoba where there they are considered livestock. Plains Bison are not listed under any provincial or territorial species at risk legislation. Legal hunting of wild Plains Bison is permitted in British Columbia, through a limited entry hunt (D. Fraser, British Columbia Ministry of Environment, pers. comm.). There is no hunting season for Plains Bison in Saskatchewan. However, there is a small hunt of the Sturgeon River subpopulation by the Big River First Nations and unregulated hunting of the McCusker River subpopulation is a concern (R. Tether, Saskatchewan Ministry of Environment, pers. comm.).
Plains Bison are not listed under the U.S. Endangered Species Act, despite a succession of petitions to do so. Most recently, in February 2011, the U.S. Fish and Wildlife Service announced in the Federal Register that its 90-day finding on a 2009 petition was that listing was not warranted, based on stable to increasing populations (mostly on commercial bison ranches). In contrast to Wood Bison (see below), there has not been a proposal to list Plains Bison on a CITES appendix.
Wood Bison
In Canada, Wood Bison are listed as Threatened under Schedule 1 of SARA. A national recovery plan for Wood Bison in Canada was developed by the National Wood Bison Recovery Team and finalized in 2001 (Gates et al. 2001a). The recovery plan pre-dated the federal Species at Risk Act and was not compliant with the requirements of the act. A SARA-compliant national recovery strategy for Wood Bison in Canada is anticipated.
In 1977, Wood Bison were added to Appendix I of the Convention on the International Trade in Endangered Species (CITES), prohibiting any and all international trade in Wood Bison or their parts. Based on limited population increases, and analyses that indicated that international trade was not a threat, CITES moved Wood Bison from Appendix I to Appendix II in 1987 (Gates et al. 2001a). In accordance with listing on Appendix II of CITES, international trade in Wood Bison or their parts, is allowed, providing it is not detrimental to the population and shipments are well-controlled and tracked through permitting procedures.
Wood Bison are classified as wildlife in the wildlife acts of Manitoba, Alberta, British Columbia, and the Northwest Territories. In Yukon, they are legally classified as a Transplanted Species in the Yukon Wildlife Act. Wood Bison are legally hunted in Yukon, Northwest Territories, and Alberta, through permits issued by the provincial or territorial wildlife management agency. Hunting is regulated in these jurisdictions; Aboriginal people require a permit to take Wood Bison in these jurisdictions, with the exception of bison that wander out of Wood Buffalo National Park, or reside in the Slave River Lowlands. In Alberta, American Bison are classified as wildlife in a special management area that encompasses Hay Zama Lakes then south to the Chichaga River. Where Wood Bison occur outside this area, for example, adjacent to Wood Buffalo National Park they are not afforded any legal protections. Hunting Wood Bison is generally not allowed in Manitoba; however, the Skownon First Nation has occasionally received a permit to remove a problem animal (B. Joynt, Manitoba Conservation, pers. comm.). Hunting is not permitted in British Columbia (D. Fraser, British Columbia Ministry of Environment, pers. comm.). Wood Bison are not protected under species at risk legislation in Manitoba or the Northwest Territories.
Since 1970, Wood Bison were listed as Endangered in Canada in the 1969 U.S. Endangered Species Conservation Act (precursor to the 1973 U.S. Endangered Species Act). A final rule was implemented by the U.S. Fish and Wildlife Service in June 2012 reclassifying the Wood Bison to Threatened under the Endangered Species Act. The decision was based on “compelling evidence that recovery actions have been successful in reducing the risk of extinction associated with the threats identified” (Federal Register/Vol. 77, No. 86/Thursday, May 3, 2012/Rules and Regulations). In January 2013, the U.S. Fish and Wildlife Service proposed to designate a nonessential experimental population of Wood Bison in Alaska under section 10(j) of the Endangered Species Act (ESA) and an associated special rule that would provide a wide range of management options, including assurances that the establishment of the wild subpopulation(s) will not have any unintended consequences for the State, private landowners, industry, or Alaska Natives. If adopted, the proposed rule would give the State of Alaska primary management responsibility for leading and implementing Wood Bison restoration in Alaska (R. Stephenson, Alaska Department of Fish and Game, pers. comm.).
Non-Legal Status and Ranks
Globally, the IUCN Red List ranks American Bison (both Wood and Plains together) as Near Threatened (Gates and Aune 2008). Gates and Aune (2008) provided the following rationale for the current designation: “There has been a modest increase in the number of conservation herds and individuals in populations managed for species conservation and ecological restoration, however, all mature individuals occur within active management programs which if ceased would result in the species qualifying for a threatened status. About 97% of the continental population is managed for private captive commercial propagation; very few of these subpopulations are managed primarily for species conservation and none is managed in the public interest for conservation. Subpopulations managed for conservation purposes in the public interest are typically small (< 400), and populations are widely dispersed with few geographic situations that provide conditions for natural movements between subpopulations.”
Nationally, the General Status program ranks bison as At Risk. This conservation status ranking system does not consider designatable units below the species level. Within the provinces and territories, American Bison is regarded as At Risk in Yukon, Northwest Territories, Alberta and Manitoba, May Be at Risk in British Columbia, and Sensitive in Saskatchewan. The global NatureServe rank for American Bison is G4, while the Canadian and U.S. national ranks are N3N4 and N4, respectively. The Plains Bison is ranked G4TU (unrankable due to a lack of information or substantially conflicting information about status or trends), while the Wood Bison is ranked G4T2Q (imperiled but with questionable taxonomy). The subpopulation ranks (S ranks) for Plains Bison are: SX (presumed extirpated) in British Columbia; S3 (vulnerable) in Saskatchewan; and SNR (not ranked) in Alberta and Manitoba (NatureServe 2011). The British Columbia S rank refers to Plains Bison being extirpated from the central Rocky Mountains, and that the extra-limital Pink Mountain subpopulation was not considered in the assignment of the rank (L. Ramsay, British Columbia Conservation Data Centre, pers. comm.). The S ranks for Wood Bison are: Saskatchewan, SX (presumably extirpated); Alberta, S1 (critically imperiled); British Columbia and Yukon, S2 (imperiled); and Manitoba, SNA (not applicable), where they are considered exotic.
Habitat Protection and Ownership
Wild Plains Bison populations and habitat are protected in Elk Island, Prince Albert and Grasslands National Parks, and in the Cold Lake Air Weapons Range. Wood Bison and habitat are protected in Wood Buffalo National Park and Elk Island National Park.
Other wild bison populations occur outside of protected areas. American Bison are a vagile species and need large, connected landscapes to conserve processes such as migration, and daily and seasonal movements in relation to resource gradients and fleeing from predators (Meagher 1989, Carbyn et al. 1993, Berger 2004). For example, the Sturgeon River population occurs primarily in Prince Albert National Park, Saskatchewan, but now seasonally ranges outside the park where their presence conflicts with agricultural interests (R. Tether, Saskatchewan Ministry of Environment, pers. comm.). Similarly, the McCusker River subpopulation ranges well beyond the boundary of the Cold Lake Air Weapons range in Saskatchewan and is exposed to unregulated take.
Acknowledgements and Authorities Contacted
Preparation of this report was funded by Environment Canada. The writers thank Jenny Wu for administrative and GIS support, Justina Ray, Donna Hulbert, and Dean Tremblay of the COSEWIC Terrestrial Mammals Subcommittee and ATK Subcommittees for advice and support. Members of the Terrestrial Mammal Subcommittee were invaluable in getting this to the finish line, particularly the bison subcommittee (Jeff Bowman, Chris Johnson, Christopher Kyle, and Susan Kutz). Ruben Boles provided key insight into the development of the “wild by nature” concept. Bonnie Fournier (NWT) provided key mapping support. We especially thank the various bison biologists and managers that took the time to complete our online questionnaire and patiently answer our subsequent questions, namely: Terry Armstrong, Seth Cherry, Lyle Fullerton, David Fraser, Barb Johnston, Bryan Joynt, Rhona Kindopp, Gerry Kuzyk, Wes Olson, Delinda Ryerson, Helen Schwantje, Angela Spooner, Rob Tether, Conrad Thiessen, and Greg Wilson. David Fraser generously facilitated the teleconferences on the threats calculator, providing invaluable insight and perspectives.
The following experts generously provided information, advice, information, insight, and opinion toward the writing of the current update status report:
Ackermann, Thomas. Chairman. Bison Producers of Alberta, Lacombe, Alberta.
Allen, Martha, Ecologist Team Leader, Elk Island National Park, Parks Canada Agency, Fort Saskatchewan, Alberta.
Aune, Keith. Wildlife Conservation Society. Bozeman, Montana.
Armstrong, Terry. Bison Ecologist. Northwest Territories Department of Environment and Natural Resources, Government of the Northwest Territories. Fort Smith, Northwest Territories.
Carrière, Suzanne. Ecosystem Management Biologist. Northwest Territories Department of Environment and Natural Resources, Government of the Northwest Territories, Yellowknife, Northwest Territories.
Cherry, Seth. Prince Albert National Park, Parks Canada. Waskesiu Lake, Saskatchewan.
Cool, Norm. Elk Island National Park, Parks Canada. Fort Saskatchewan, Alberta.
Cooper, Alan. Director. Australian Centre for Ancient DNA. University of Adelaide, Australia.
Court, Gord. Provincial Status Wildlife Biologist. Alberta Sustainable Resource Development. Government of Alberta. Edmonton, Alberta.
Derr, James. Professor, Veterinary Pathobiology. Texas A&M University College of Veterinary Medicine, College Station, Texas.
Duncan, Dave. Prairie and Northern Region, Environment Canada, Government of Canada. Edmonton, Alberta.
Duncan, James. A/Director, Manitoba Conservation, Government of Manitoba. Winnipeg, Manitoba.
Elkin, Brett. Disease/Contaminants Specialist. Wildlife Division, Environment and Natural Resources, Government of the Northwest Territories, Yellowknife, NT.
Fraser, David. Scientific Assessment Authority. British Columbia Ministry of Environment, Government of British Columbia, Victoria, British Columbia.
Fullerton, Lyle. Special Project Biologist. Alberta Sustainable Resource Development, Government of Alberta. Peace River, Alberta.
Hamilton, George. Alberta Sustainable Resource Development, Government of Alberta. Edmonton, Alberta.
Handel, Archie. Elk Island National Park, Parks Canada. Fort Saskatchewan, Alberta.
Hulburt, Donna. Co-Chair, ATK Subcommittee, COSEWIC.
Hurd, Tom. Banff National Park, Parks Canada. Banff, Alberta.
Johnston, Barb. Ecosystem Scientist. Waterton Lakes National Park, Parks Canada.
Joynt, Brian. Manitoba Conservation, Government of Manitoba. Winnipeg, Manitoba.
Kassi, Norma. Member. ATK Subcommitee, COSEWIC, Old Crow, Yukon.
Kindopp, Rhona. Wood Buffalo National Park, Parks Canada. Fort Smith, Northwest Territories.
Kremeniuk, Terry. Executive Director, Canadian Bison Association, Regina, Saskatchewan.
Kuzyk, Gerry. Ungulate Specialist. British Columbia Ministry of Environment, Government of British Columbia, Victoria, British Columbia.
Larter, Nic. Manager, Wildlife Research and Monitoring, Dehcho Region, Government of Northwest Territories.
Lirette, Daniel. Wildlife Biologist, Peace Region. Ministry of Forests, Lands and Natural Resource Operations, Peace Region, British Columbia.
McMillan, Stuart. Wood Buffalo National Park, Parks Canada. Fort Smith, Northwest Territories.
Merkle, Jerod. Ph.D. Candidate, Département de Biologie et Centre d’Étude de la Forêt, Université Laval, Québec, Québec.
Moyles, Dave. Senior Wildlife Biologist. Alberta Sustainable Resource Development, Government of Alberta. Peace River, Alberta.
Nantel, Patrick. Conservation Biologist. Ecological Integrity Branch, Parks Canada, Gatineau, Quebec.
Nishi, J.S. Wildlife Biologist, EcoBorealis Consulting Inc. and ALCES Group, Millarville, Alberta.
Olsen, Wes. Grasslands National Park, Parks Canada. Val Marie, Saskatchewan.
Pepper, Jeanette. Zoologist. Saskatchewan Ministry of Environment, Government of Saskatchewan. Regina, Saskatchewan.
Powell, Todd. Manager. Yukon Department of Environment, Government of Yukon. Whitehorse, Yukon.
Ramsay, Leah. Zoologist. British Columbia Conservation Data Centre. Victoria, British Columbia.
Reid, D.G., Conservation Scientist, Wildlife Conservation Society Canada, Whitehorse, YT.
Ryerson, Delinda. Elk Island National Park, Parks Canada. Fort Saskatchewan, Alberta.
Schwantje, Helen. Wildlife Veterinarian. Ministry of Environment, Government of British Columbia, Victoria, British Columbia.
Shapiro, Beth. Associate Professor. University of California, Santa Cruz, California.
Silzer, Mark. President. Canadian Bison Association, Regina, Saskatchewan.
Spooner, Angela. Riding Mountain National Park, Parks Canada. Wasagaming, Manitoba.
Stephenson, Robert. Wood Bison Project Biologist. Alaska Department of Fish and Game, Alaska State Government. Fairbanks, Alaska.
Tether, Rob. Saskatchewan Ministry of Environment, Government of Saskatchewan. Regina, Saskatchewan.
Thiessen, Conrad. Ministry of Environment, Government of British Columbia, Victoria, British Columbia.
Tremblay, Dean. Co-Chair. ATK Subcommittee, COSEWIC. Salmon Arm, British Columbia.
Watkins, William. Zoologist. Manitoba Conservation, Government of Manitoba. Winnipeg, Manitoba.
Wilson, Greg. Wildlife Biologist. Environmental Stewardship Branch, Environment Canada, Government of Canada. Edmonton, Alberta.
Zazula, Grant. Paleontologist. Yukon Plaeontology Program, Department of Tourism and Culture, Government of Yukon. Whitehorse, Yukon.
Information Sources
Alberta Environmental Protection. 1997. The Grasslands Natural Region of Alberta. One of a series of reports prepared for the Special Places 2000 Provincial Coordinating Committee. Alberta Environmental Protection Natural Resources Service Recreation & Protected Areas Division Natural Heritage Protection and Education Branch. Edmonton, Alberta.
Altizer, S., D. Harvell, and E. Friedle. 2003. Rapid evolutionary dynamics and disease threats to biodiversity. Trends in Ecology and Evolution, 18:589-596.
Altizer S., R. Bartel, and B.A. Han. 2011. Animal migration and infectious disease risk. Science, 331:296-302.
Arbuthnott, K.A. and J.K. Schmutz. 2013. PFRA Community Pastures: History and Drama of a Prairie Commons. Canadian Centre for Policy Alternatives - Saskatchewan Office. April 2013: 1-8.
Armstrong, T. 2013. Mackenzie wood bison population estimate. Unpublished Report.
Armstrong. T. and K. Cox. 2011. Project: 2009 Slave River Lowlands Wood Bison Population Estimate. Unpublished Report.
Athabasca Chipewyn First Nation (AFCN) 2010. Supplementary Submission of the Athabasca Chipewyn First Nation to the Federal Statutory Review of the Species at Risk Act [PDF, 90 KB], April, 2010.
Aune, K., and C. Gates. 2010. Reportable or notifiable diseases. Pages 27-37 in C.C. Gates, C.H. Freese, P.J.P. Gogan and M. Kotzman (eds.), American Bison: Status Survey and Conservation Guidelines 2010. IUCN, Gland, Switzerland.
Aune, K. and R. Wallen. 2010. Legal status, policy issues and listings. Pages 63-84 in C.C. Gates, C.H. Freese, P.J.P. Gogan and M. Kotzman (eds.), American Bison: Status Survey and Conservation Guidelines 2010. IUCN, Gland, Switzerland.
Banfield, A. W. F. 1974. The mammals of Canada. University of Toronto Press, Toronto, Ontario, Canada.
Banfield, A.W.F. and N.S. Novakowski. 1960. The survival of the Wood Bison (Bison bison athabascae Rhoads) in the Northwest Territories. Natural History Papers No. 8. National Museum of Man, Ottawa, August 30, 1960. 6pp.
Barmore, W. J. 2003. Ecology of ungulates and their winter range in northern Yellowstone National Park: research and synthesis, 1962-1970. Yellowstone Center for Resources, National Park Service, Mammoth Hot Springs, Wyoming.
Barsh, L. and C. Marlor. 2003. Driving bison and Blackfoot Science. Human Ecology 31:571-593.
Berger, J. 2004. The last mile: how to sustain long-distance migration in mammals. Conservation Biology 18:320-331.
Berger, J. and C. Cunningham. 1994. Bison: Mating and Conservation in Small Populations. Columbia University Press, New York.
Berger, J. and Peacock, M. 1988. Variability in size-weight relationships of Bison bison. Journal of Mammalogy 69:618-624.
Berthoud, E.L. 1892. A peculiar case of plant dissemination. Botanical Gazette 17:321-326.
Bidwell, D. 2009. Bison, boundaries and brucellosis: risk perception and political ecology at Yellowstone. Society & Natural Resources 23:14-30.
Blackburn, J.K., K. M. McNyset, A. Curtis, and M.E. Hugh-Jones. 2007. Modeling the geographic distribution of Bacillus anthracis, the causative agent of Anthrax disease, for the contiguous United States using predictive ecologic niche modeling. American Journal of Tropical Medicine and Hygene, 77: 1103–1110.
Bork, A.M., Strobeck, C.M., Yeh, F.C., Hudson, R.J. and Salmon, R.K. 1991. Genetic relationship of wood and Plains Bison based on restriction fragment length polymorphisms. Canadian Journal of Zoology 69:43-48.
Bowen, D. 2002. Agricultural expansion in northern Alberta. Geographical Review 92: 503-525.
Boyd, D. 2003. Conservation of North American Bison: Status and Recommendations. Master’s Thesis, University of Calgary, Faculty of Environmental Design. 235 pp.
Boyd, D.P., G.A. Wilson and C.C. Gates. 2010. Taxonomy and nomenclature. Pages 13-19 in C.C. Gates C.H. Freese, J.P. Gogan and M. Kotzman (eds) American Bison Status Survey and Conservation Guidelines 2010. IUCN, Gland Switzerland.
Bradley, M., and J. Wilmshurst. 2005. The fall and rise of bison populations in Wood Buffalo National Park: 1971 to 2003. Canadian Journal of Zoology 83:1195-1205.
Bradley, R.S., Diaz, H.F., Eischeid, J.K., Jones, P.D., Kelly, P.M. and Goodess, C.M. 1987: Precipitation fluctuations over northern hemisphere land areas since the mid-19th century. Science 237, 171–75
Bragg, A.N. 1940. Observations on the ecology and natural history of Anura: I Habits, habitat, and breeding of Bufo cognatus Say. American Naturalist 74:322-349, 424-438.
Brands, S.J. (comp.) 1989-2005. Systema Naturae 2000. Amsterdam, The Netherlands.
Burns, J.A. 1996. Vertebrate paleontology and the alleged ice-free corridor: the meat of the matter. Quaternary International 32:107-112.
Bush D., and R. Rowell. 2000. OPTI Canada Long Lake Project, Traditional Land Use Study, AXYS Environmental Consulting Ltd., Calgary, Alberta.
Campbell, B. H., and M. Hinkes. 1983. Winter diets and habitat use of Alaska bison after wildfire. Wildlife Society Bulletin 11:16-21.
Campbell, C., I. Campbell, C. Blythe and J. McAndrews. 1994. Bison extirpation may have caused aspen expansion in western Canada. Ecography 17:360-362.
Carbyn, L. N. 1998. Some aspects regarding wolf predation on bison in Wood Buffalo National Park; Int. Symp. Bison Ecol. Manage. N. Am. 1997: 92-95. Missoula, MT.
Carbyn, L.N., S.M. Oosenbrug, and D.W. Anions. 1993. Wolves, bison and the dynamics related to the Peace-Athabasca Delta in Canada’s Wood Buffalo National Park. Circumpolar Research Series No. 4. Edmonton: Canadian Circumpolar Institute, University of Alberta. 270 p.
Carbyn, L.N. and T. Trottier. 1987. Responses of bison on their calving grounds to predation by wolves in Wood Buffalo National Park. Canadian Journal of Zoology 65:2072-2078.
Carbyn, L.N. and T. Trottier. 1988. Description of wolf attacks on bison calves in Wood Buffalo National Park. Arctic 41:297-302.
Carpenter, L.H., D.J. Decker and J.F. Lipscomb. 2000. Stakeholder acceptance capacity in wildlife management. Human Dimensions of Wildlife 5:5-19.
Caughley, G. 1970a. Eruption of ungulate populations, with emphasis on Himalayan thar in New Zealand. Ecology 51:53–72.
Caughley, G. 1970b. Liberation, dispersal and distribution of Himalayan thar (Hemitragus jemlahicus) in New Zealand. New Zealand Journal of Science 13:220– 239.
Centre for Indigenous Environmental Resources Inc., 2008. Manitoba and Saskatchewan First Nations Species at Risk Lesson Plan Booklet.
Chisholm, B.J. J. Driver, S. Dube and H.P. Schwarcz. 1986. Assessment of prehistoric bison foraging and movement patterns via stable-carbon isotopic analysis. Plains Anthropologist 31:193-205.
Chowns, T., C. Gates, and F. Lepine. 1998. Large scale free burning to im- prove Wood Bison habitat in northern Canada. Pages 205–210 in L. Irby and J. Knight, editors. International Symposium on Bison Ecol- ogy and Management in North America, Montana State University, Bozeman, 4–7 June 1997. Montana State University Press, Bozeman.
Christopherson, R.J., R.J. Hudson, and R.J. Richmond. 1978. Comparative winter bioenergetics of American Bison, yak, Scottish Highland, and Hereford calves. Acta Theriologica 23:49-54.
Clarke, C.H.D. 1945. Biological reconnaissance of the lands adjacent to the Alaska Highway in northern British Columbia and the Yukon Territory. Unpublished report in the National Archives of Canada, RG 85 collection, Vol. 1191, file 400-2, Parts 1-2.
Coder, G.D. 1975. The national movement to preserve the American buffalo in the United States and Canada between 1880 and 1920. PhD dissertation. Ohio State University. 348 pp.
Coppedge, B.R. 2009. Patterns of bison hair use in nests of tallgrass prairie birds. Prairie Naturalist 41: 110-115.
Coppedge, B.R. and J.H. Shaw. 1998. Bison grazing patterns on seasonally burned tallgrass prairie. Journal of Range Management 51:258-264.
Coppock, D.L. and J.K. Detling. 1986. Alteration of bison and black-tailed prairie dog grazing interaction by prescribed burning. Journal of Wildlife Management 50:452-455.
Coppock, D.L., J.K. Detling, J.E. Ellis and M.I. Dyer. 1983a. Plant- herbivore interactions in a North American mixed-grass prairie: I. effects of black-tailed prairie dogs on intraseasonal aboveground plant biomass and nutrient dynamics and plant species diversity. Oecologia 56:1-9.
Coppock, D.L., J.E. Ellis, J.K. Detling and M.I. Dyer. 1983b. Plant-herbivore interactions in a North American mixed-grass prairie. II. responses of bison to modification of vegetation by prairie dogs. Oecologia 56:10-15.
COSEWIC 2004. COSEWIC assessment and status report on the plains bison Bison bison bison in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 71 pp.
COSEWIC 2010a. Appendix E7 – Guidelines on Manipulated Populations. Approved by COSEWIC in April 2010.
COSEWIC 2010b. Instructions for the Preparation of COSEWIC Status Reports. Approved by COSEWIC in April 2010, updated July 2010.
COSEWIC. 2011a. Guidelines for Recognizing Designatable Units. Approved by COSEWIC in November 2011.
COSEWIC. 2011b. Aboriginal Traditional Knowledge Source Report on Plains Bison and Wood Bison Bison bison and Bison bison athabascae. Ottawa, ON.
COSEWIC. 2012. COSEWIC Aboriginal Traditional Knowledge Assessment Report on Plains Bison and Wood Bison, Bison bison and Bison bison athabascae. Ottawa,ON.
Creel, S. and D. Christianson. 2008. Relationships between direct predation and risk effects. Trends in Ecology & Evolution 23:194 –201.
Cronin, M.A., M. D. MacNeil, N. Vu, V. Leesburg, H. Blackburn, and J.N. Derr. 2013. Genetic variation and differentiation of bison (Bison bison) subspecies and cattle (Bos taurus) breeds and subspecies. Journal of Heredity, 104: 500-509.
Damhoureyeh, S.A. and D.C. Hartnett. 1997. Effects of bison and cattle on growth, reproduction, and abundance of five tallgrass prairie forbs. American Journal of Botany 84:1719-1728.
Dary, D. 1974. The Buffalo Book. Saga of an American Symbol. Avon Books. New York, New York.
Dary, D. A. 1974. The buffalo book: The full saga of the American animal. Swallow Press/Ohio University Press, Chicago.
Davi, N.K., Jacoby, G.C. and Wiles, G.C. 2003. Boreal temperature variability inferred from maximum latewood density and tree-ring width data, Wrangell Mountain region, Alaska. Quaternary Research 60, 252–62.
Davis, D.S., Templeton, J.W., Ficht, T.A., Williams, J.D., Kopec, J.D. and Adams, L.G. 1990. Brucella abortus in captive bison: I. serology, bacteriology, pathogenesis and transmission to cattle. Journal of Wildlife Diseases 26:360-371.
Davis, D.S., J.W. Templeton, T.A. Ficht, J.D. Huber, R.D. Angus, and L.G. Adams. 1991. Brucella abortus in bison: II. evaluation of strain 19 vaccination of pregnant cows. Journal of Wildlife Diseases 27:258-264.
Dawkins, R. and J.R. Krebs. 1979. Arms races between and within species. Proceedings of the Royal Society of London Series B Biological Sciences 205: 489–511.
Day, T.A. and J.K. Detling. 1990. Grassland patch dynamics and herbivore grazing preference following urine deposition. Ecology 71:180-188.
Decker, D.J. and K.G. Purdy. 1988. Toward a concept of wildlife acceptance capacity in wildlife management. Wildlife Society Bulletin 16:53-57.
Decker, J.E., Pires, J.C., Conant, G.C., McKay, S.D., Heaton, M.P., Chen, K., Cooper, A., Vilkki, J., Seabury, C.M., Caetano, A.R., Johnson, G.S., Brenneman, R.A., Hanotte, O., Eggert, L.S., Wiener, P., Kim, J., Kim, K.S., Sonstegard, T.S., Van Tassell, C.P., Neibergs, H.L., McEwan, J.C., Brauning, R., Coutinho, L.L., Babar, M.E., Wilson, G.A., McClure, M.C., Rolf, M.M., Kim, J., Schnabel, R.D. & Taylor, J.F. (2009). Resolving the evolution of extant and extinct ruminants with high-throughput genomics. Proceedings of the National Academy of Sciences 106: 18644-18649.
DelGiudice, G. D., F. J. Singer, U. S. Seal, and G. Bowser. 1994. Physiological responses of Yellowstone bison to winter nutritional deprivation. Journal of Wildlife Management 58:24–34.
DelGiudice, G., R. Moen, F. Singer and M. Riggs. 2001. Winter nutritional restriction and simulated body condition of Yellowstone elk and bison before and after the fires of 1988. Wildlife Monographs 147, 60pp.
Derr, J.N., P.W Hedrick, N.D Halbert, L. Plough, L.K Dobson, J. King, C. Duncan, D. Hunter, N.D. Cohen, and D. Hedgecock. 2012. Phenotypic effects of cattle mitochondrial DNA in American Bison. Conservation Biology 26:1130-1136.
Douglas, K.C., N.D. Halbert, C. Kolenda, C. Childers, D.L. Hunter and J.N. Derr. 2011. Complete mitochondrial DNA sequence analysis of Bison bison and bison-cattle hybrids: Function and phylogeny. Mitochondrion 11:166-175.
Dragon, D.C., B.T. Elkin, J.S. Nishi, and T.R. Ellsworth. 1999. A review of anthrax in Canada and implications for research on the disease in northern bison. Journal of Applied Microbiology, 87:208-213.
Dragon, D.C. and B.T. Elkin. 2001. An overview of early anthrax outbreaks in northern Canada: field reports of the Health of Animals Branch, Agriculture Canada, 1962-1971. Arctic, 54:32-40.
Dragon D.C, D.E Bader, J. Mitchell and N. Woollen. 2005. Natural dissemination of Bacillus anthracis spores in northern Canada. Applied Environmental Microbiology 71: 1610–1615.
Dragon D.C. and Rennie, R.P. 1995. The ecology of anthrax spores: tough but not invincible. Canadian Veterinary Journal. 36:295-301.
Egerton, P.J.M. 1962. The cow–calf relationship and rutting behavior in the American Bison. M.S. Thesis, University of Alberta, Edmonton, Canada.
Ellingson, J.L.E., J.R. Stabel, R.P. Radcliff, R.H. Whitlock, and J.M. Miller. 2005. Detection of Mycobacterium avium subspecies paratuberculosis in free-ranging bison (Bison bison) by PCR. Molecular and Cellular Probes, 19: 219-225.
England, R.E. and A. DeVos. 1969. Influence of animals on pristine conditions on the Canadian grasslands. Journal of Range Management 22:87-94.
Epp, H.T. 1988. Way of the migrant herds. Dual dispersion strategy among bison. Plains Anthrpologist 33:309-330.
Epp. H.T. and I. Dyck. 2002. Early human-bison population interactions in the Plains Ecosystem. Great Plains Research 12:323-327.
Fahnestock, J.T. and A.K. Knapp. 1993. Water relations and growth of tallgrass prairie forbs in response to selective grass herbivory by bison. International Journal of Plant Science 154:432-440.
Fancy, S. 1982. Reaction of bison to aerial surveys in interior Alaska. Canadian Field-Naturalist 96:91.
Farnell, R., P.G. Hare, E. Blake, V. Bowyer, C. Schweger, S.Greer,and R.Gotthardt. 2004. Multidisciplinary Investigations of Alpine Ice Patches in Southwest Yukon, Canada: Paleoenvironmental and Paleobiological Investigations. Arctic 57:247-59.
Ferguson, T. A. and F. Laviolette. 1992. A note on historical mortality in a northern bison population. Arctic 45: 47-50.
Fischer, L.A. and C.C. Gates. 2005. Competition potential between sympatric woodland caribou and Wood Bison in southwestern Yukon, Canada. Canadian Journal of Zoology 83:1162-1173.
Flannigan, M.D., B.J. Stocks, M.R Turetsky, and B.M. Wotton. 2009. Impact of climate change on fire activity and fire management in the circumboreal forest. Global Change Biology, 15: 549-560.Flores, D. 1996. The great contraction. In: C.E. Rankin (ed.), Legacy: new perspectives on the battle of the little Bighorn. Proceedings of the Little Bighorn Legacy Symposium, held in Billings, Montana, August 3-6, 1994, pp.3-22. Montana Historical Society Press, Helena, Montana.
Forde, T., J. De Buck, B. Elkin, S. Kutz, F. van der Meer, and K. Orsel. 2013. Mycobacterium avium subspecies paratuberculosis in wood bison: contrasting results of culture-dependent and molecular analyses. Applied Environmental Microbiology, 79: in press.
Forrest, S.C., Strand, H., Haskins, W.H., Freese, C., Proctor, J. and Dinerstein, E. 2004. Ocean of Grass: a Conservation Assessment for the Northern Great Plains. Northern Plains Conservation Network and World Wildlife Fund Northern Great Plains Program. World Wildlife Fund, Bozeman, Montana.
Fortin, D, Fortin, M-E., Beyer, H.L., Duchesne, T., Courant, S. & Dancose, K. 2009.Group-size-mediated habitat selection and group fusion–fission dynamics of bison under predation risk. Ecology 90: 2480-2490.
Fortin, D., Fryxell, J.M. and Pilote, R. 2002. The temporal scale of foraging decisions in bison. Ecology 83:970-982.
Fortin, D., J.M. Fryxell, L. O'Brodovich and D. Frandsen. 2003. Foraging ecology of bison at the landscape and plant community levels: the applicability of energy maximization principles. Oecologica 134: 219-227.
Fortin, D. and M.C. Andruskiw. 2003. Behavioral response of free-ranging bison to human disturbance. Wildlife Society Bulletin, 31: 804-813.
Foster, J.E. 1992. The Metis and the end of the plains buffalo in Alberta. Alberta: Studies in the Arts and Sciences 3: 61-77. University of Alberta Press.
Fox, T.A., C.H. Hugenholtz., D. Bender, and C.C. Gates. 2012. Can bison play a role in conserving habitat for endangered sandhills species in Canada? Biodiversity and Conservation 21:1441-1455.
Frank, D. 1998. Ungulate regulation of ecosystem processes in Yellowstone National Park: direct and feedback effects. Wildlife Society Bulletin 26:410-418.
Frank, D. 2000. Ungulate stimulation of nitrogen cycling and retention in Yellowstone National Park. Oecologia 123:116-121.
Frank, D. and R.D. Evans. 1997. Effects of native grazers on grassland N cycling in Yellowstone National Park. Ecology 78:2238-2248.
Frank, D.A. and McNaughton, S.J. 1992. The ecology of plants, large mammalian herbivores, and drought in Yellowstone National Park. Ecology 73:2043-2058.
Frank, D.A. and S.J. McNaughton. 1993. Evidence for the promotion of aboveground grassland production by native large herbivores in Yellowstone National Park. Oecologia 96:157-161.
Freese, C., K. Aune, D. Boyd, J. Derr, S. Forrest, C. Gates, P. Gogan, S. Grassel, N. Halbert, K. Kunkel and K. Redford. 2007. Second chance for the Plains Bison. Biological Conservation 136:175-184.
Frison, G.C. 1991. Prehistoric Hunters of the High Plains, Second Edition. Academic Press, Inc., San Diego, California.
Fryxell, J.M., J. Greever and A.R.E. Sinclair. 1988. Why are migratory ungulates so abundant? The American Naturalist 131:781-798.
Fuhlendorf, S., D. Engle, J.D. Kirby and R.G. Hamilton. 2009. Pyric herbivory: rewilding landscapes through the recoupling of fire and grazing. Conservation Biology 23:588-598.
Fuller, W.A. 1950. Aerial census of northern bison in Wood Buffalo National Park and vicinity. Journal of Wildlife Management 14:445-451.
Fuller, W.A. 1953. Behaviour and social organization of the wild bison of Wood Buffalo National Park, Canada. Arctic 13: 2-19.
Fuller, W. A. 1961. The ecology and management of the American Bison. Terre et la Vie 2:286–304.
Fuller, W.A. 1966. The biology and management of the bison of Wood Buffalo National Park. Canadian Wildlife Service Wildlife Management Bulletin Series 1:1-52.
Fuller, W.A. 2002. Canada and the “buffalo”, Bison bison: a tale of two herds. Canadian Field-Naturalist 116:141-159.
Fuller, J.A., R.A. Garrott, P.J. White, K.E. Aune, T.J. Roffe, and J.C. Rhyan. 2007. Reproduction and survival of Yellowstone bison. Journal of Wildlife Management 71:2365-2372.
Garretson, M.S. 1927. A short history of the American Bison. American Bison Society, New York.
Gates C. C. In press. What is a wild bison? A case study of plains bison conservation in Canada. In: Melletti, M. & Burton J. Ecology, Evolution and Behaviour of Wild Cattle: Implications for Conservation. Cambridge University Press.
Gates, C.C. & Aune, K. 2008. Bison bison. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2. <www.iucnredlist.org>. Downloaded on 12 February 2012.
Gates, C.C., T. Chowns and H. Reynolds. 1992. Wood Buffalo at the crossroads. Aberta: Studies in the Arts and Sciences 3: 139-165. University of Alberta Press, Edmonton, AB.
Gates, C.C., Elkin B. and Dragon, D. 1995. Investigation, control and epizootiology of anthrax in an isolated, free-roaming bison population in northern Canada. Canadian Journal of Veterinary Research 59:256-264.
Gates, C.C., B. Elkin and D. Dragon. 2001b. Anthrax. Pages 396 - 412 in E. Williams and I. Barker (eds.) Infectious diseases of wild mammals. Iowa State University Press, Ames.
Gates, C.C., and K. Ellison. 2010. Numerical and geographic status. Pages 55-62 in C.C. Gates, C.H. Freese, P.J.P. Gogan and M. Kotzman (eds.), American Bison: Status Survey and Conservation Guidelines 2010. IUCN, Gland, Switzerland.
Gates, C.C. and N.C. Larter. 1990. Growth and dispersal of an erupting large herbivore population in northern Canada: the Mackenzie Wood Bison (Bison bison athabascae). Arctic 43:231-238.
Gates, C.C., N.C. Larter, and P.K. Komers. 1991. Size and composition of the Mackenzie Bison population in 1989. Renewable Resources File Report No. 93. Yellowknife, Northwest Territories.
Gates, C.C., B. Stelfox, T. Muhly, T. Chowns and R.J. Hudson. 2005. The ecology of bison movements and distribution in and beyond Yellowstone National Park: A critical review with implications for winter use and transboundary population management. Report commissioned by the United States National Park Service, Yellowstone Center for Resources. Faculty of Environmental Design, University of Calgary, Alberta.
Gates, C.C., R.O. Stephenson, H.W. Reynolds, C.G. van Zyll de Jong, H. Schwantje, M. Hoefs, J. Nishi, N. Cool, J. Chisholm and B. Koonz. 2001a. National recovery plan for the Wood Bison (Bison bison athabascae). National Recovery Plan No. 21. Ottawa: Recovery of Nationally Endangered Wildlife (RENEW).
Gates, C.C., R. O. Stephenson, J.P. Gogan, C. H. Freese, and K. Kunkel. 2010. Guidelines for ecological restoration. Pages 103-112 in C.C. Gates, C.H. Freese, P.J.P. Gogan and M. Kotzman (eds.), American Bison: Status Survey and Conservation Guidelines 2010. IUCN, Gland, Switzerland.
Gauthier, D.A and E. Wiken. 2003. Monitoring the conservation of grassland habitats, prairie ecozone, Canada. Environmental Monitoring and Assessment 88:343-364.
Geist, V. 1991. Phantom subspecies:the Wood Bison Bison bison “athabascae” Rhoads 1897 is not a valid taxon, but an ecotype. Arctic 44:283-300.
Geist, V. 1996. Buffalo Nation: History and Legend of the North American Bison. Fifth House Ltd., Saskatoon, Saskatchewan.
Gogan, J.P., N.C. Larter, J.H. Shaw, J.E. Gross, C.C. Gates and J. Truett . 2010. General biology, ecology and demographics. Pages 39-54 in Gates, C. C., C.F. Freese, P. Gogan and M. Kotzman (eds). American Bison: Status survey and conservation guidelines. International Union for the Conservation of Nature, Gland, Switzerland. ISBN 978-2-8317-1149-2.
Gokbulak, F. 2002. Effect of American Bison (Bison bison L.) on the recovery and germinability of seeds of range forage species. Grass and Forage Science 57:395-400.
Government of Alberta. 2011. Managing disease risk in Alberta's wood bison with special focus on bison to the west of Wood Buffalo National Park Edmonton, AB., Government of Alberta. 10 pp.
Government of Alberta. 2013a. Managing disease risk in northern Alberta wood bison outside of Wood Buffalo National Park.
Government of Alberta. 2013b. Ronald Lake Bison (Bison bison) Winter 2012-2013 Activities Progress Report (Final). Unpublished Report.
Government of NWT. 2013. NWT Wood Bison (Bison bison athabascae).
Government of Yukon. 2012. Management Plan for the Aishihik Wood Bison (Bison bison athabascae) Herd in southwestern Yukon. Environment Yukon, Whitehorse, Yukon. 28 pp.
Green, W. C. H. 1996. Age-related differences in nursing behavior among American Bison cows (Bison bison). Journal of Mammalogy 67:739-741.
Green, W. C. H., and A. Rothstein. 1991. Trade-offs between growth and reproduction in female bison. Oecologia 86:521-527.
Green, W. C. H., A. Rothstein, and J. G. Griswold. 1993. Weaning and parent–offspring conflict: variation relative to interbirth interval in bison. Ethology 95:105-125.
Griebel, R.L., S.L. Winter, and A.A. Steuter. 1998. Grassland birds and habitat structure in sandhills prairie managed using cattel or bison plus fire. Great Plains Research 8: 255-268.
Griffith, E. et al. (1827). The Animal Kingdom Arranged in Conformity with Its Organization. London: G. B. Whittaker. pp. 87–89.
Gross, J.E., Halbert, N.D., Derr, J.N., Aune, K., Berger, J., Elkin, B.T., Gates, C.C., Gogan, J.P., Hunter, D., Joly, D.O., Lammers, D.J., Larter, N.C., Licht, D., List, R., Paulson, R.L., Powers, J., Stephenson, R.O., Truett, J., Wallen, R. & Wild, M. 2010. Conservation guidelines for population, genetoc, and disease management. In American Bison status survey and conservation guidelines 2010, pp. 85-101. Eds. Gates, C.C., Freese, C.H., Gogan, J.P. & Kotzman, M. Gland Switzerland: IUCN.
Guthrie, R.D. 1980. Bison and man in North America. Canadian Journal of Anthropology 1:55-73.
Guthrie, R.D. 1990. Frozen Fauna of the Mammoth Steppe. University of Chicago Press, Chicago, Illinois.
Halbert, N.D., and J.N. Derr. 2008. Patterns of genetic variation in US federal bison herds. Molecular Ecology 17:4963-4977.
Halbert, N.D., T. Raudsepp, B.P. Chowdhary, and J.N. Derr. 2004. Conservation genetic analysis of the Texas state bison herd. Journal of Mammalogy 85:924-931.
Halbert, N.D., T.J. Ward, R.D. Schnabel, J.F. Taylor, and J.N. Derr. 2005. Conservation genomics: disequilibrium mapping of domestic cattle chromosomal segments in North American bison populations. Molecular Ecology, 14: 2343–2362.Halbert, N.D., and J.N. Derr. 2007. A comprehensive evaluation of cattle introgression into US federal bison herds. Journal of Heredity 98:1-12.
Halloran, A.F. 1968. Bison (Bovidae) productivity on the Wichita Mountains Wildlife Refuge, Oklahoma. Southwestern Naturalist 13:23-26.
Hamilton, R.G., N.D. Halbert, and J.N Derr. 2009. Bison Herd Genetic Architecture and Management –Working Toward a Species-Wide Conservation Approach. Unpublished report for Rodney Johnson/Katharine Ordway Stewardship Endowment.
Hamley, W. 1992. The farming frontier in northern Alberta. Geographical Journal 158: 286-294.
Harper, F. 1925. Letter to the editor of the Canadian Field-Naturalist. Canadian Field-Naturalist 39:45.
Harper, W.L., J.P. Elliott, I. Hatter and H. Schwantje. 2000. Management Plan for Wood Bison in British Columbia. B.C. Ministry of Environment, Lands and Parks, Victoria, BC. 43 pp.
Harper, W.L. and C.C. Gates. 2000. Recovery of Wood Bison in British Columbia. Pages 915-923 in L. M. Darling, editor. 2000. Proceedings of a Conference on the Biology and Management of Species and Habitats at Risk, Kamloops, B.C., 15 - 19 Feb.,1999. Volume Two. B.C. Ministry of Environment, Lands and Parks, Victoria, B.C. and University College of the Cariboo, Kamloops, B.C. 520pp.
Hartnett, D.C., A.A. Steuter and K.R. Hickman. 1997. Comparative ecology of native and introduced ungulates. Pages 72-101 in Knopf, F. L. and F. B. Samson, eds. Ecology and conservation of Great Plains vertebrates. Springer-Verlag, New York, NY.
Haugen, A.O. 1974. Reproduction in the Plains Bison. Iowa State Journal of Research 49:1-8.
Hawley, A.W.L. 1978. Comparison of forage utilization and blood composition of bison and Hereford cattle. Ph.D. Dissertation, University of Saskatchewan, Saskatoon, Saskatchewan, Canada.
Hawley, A.W.L., D.G. Peden, and W.R. Stricklin. 1981a. Bison and Hereford steer digestion of sedge hay. Canadian Journal of Animal Science 61:165-174.
Hawley, A.W.L., D.G. Peden, H.W. Reynolds, and W.R. Stricklin. 1981b. Bison and cattle digestion of forages from the Slave River Lowlands, Northwest Territories, Canada. Journal of Range Management 34:126-130.
Haynes, G. 1988. Mass deaths and serial predation: comparative taphonomic studies of modern large mammal death sites. Journal of Archaeological Science, 15:219-235.
Hebblewhite, M. and E. Merrill. 2006. Multiscale wolf predation risk for elk: does migration reduce risk. Oecologica 152:377-387.
Hedrick, P.W. 2009. Conservation genetics and North American Bison (Bison bison). Journal of Heredity 100:411-420.
Hedrick, P.W. 2010. Cattle ancestry in bison: explanations for higher mtDNA than autosomal ancestry.
Heffner, T. 2008. The role of glacial lakes in the pre-contact human history of southwest Yukon Territory: A late drainage hypothesis. The Northern Review 29: 85–104.
Hegel, T.M., K. Russell, and T.S. Jung. 2012. Using temporary dye marks to estimate ungulate population abundance in southwestern Yukon, Canada. Rangifer (Special Issue) 20:219-226.
Helbig, L., M.R. Woodbury, J.C. Haigh, and A.D. Barth. 2007. The onset of puberty in North American Bison (Bison bison) bulls. Animal Reproduction Science 97:12-24.
Hermanutz, R. and L. Fullerton. 2011. Bison surveys in the Hay-Zama Lowlands. Unpublished report. Alberta Sustainable Resource Development, Edmonton, Alberta. 10 pages.
Heuschele, W.P. and H.W. Reid. 2001. Malignant catarrhal fever. In: E.S. Williams and I.K. Barker (eds.), Infectious Diseases of Wild Mammals, Third edition, pp.157-163. Iowa State Press. Ames, Iowa.
Hewitt, C.G. 1919. The coming back of the bison. Natural History 19(6):553-565.
Hewitt, C.G. 1921. The buffalo or bison: Its present, past, and future. In: C.G. Hewitt, (ed.), The Conservation of the Wildlife of Canada, pp.113-42. Charles Scribner’s Sons, New York.
Hofmann, R. R. 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 79:443–457.
Hornaday, W.T. 1889. The extermination of the American Bison, with a sketch of its discovery and life history. Pages 367–548 in Report of the United States National Museum, 1886–87, Part 2 [1889].
Howell, A.B. 1925. Letter to the editor of the Canadian Field-Naturalist from the Corresponding Secretary of the American Society of Mammalogists, 13 April 1925. The Canadian Field-Naturalist 39:118
Hudson, R.J., and S. Frank. 1987. Foraging ecology of bison in aspen boreal habitats. Journal of Range Management 40:71-75.
Isenberg, A.C. 2000. The destruction of the bison: An environmental history, 1750-1920. Cambridge: Cambridge University Press. 218 pp.
Jacoby, G.C., Jr and D’Arrigo, R. 1989: Reconstructed northern hemisphere annual temperature since 1671 based on high-latitude tree-ring data from North America. Climate Change 14, 39–59.
Janardhan, K.S., M. Hays, N. Dyer, R.D. Oberst, and B.M. Debey. 2010. Mycoplasma bovis outbreak in a herd of North American bison (Bison bison). Journal Of Veterinary Diagnostic Investigation, 22: 797-801.
Jeffrey, W.W., 1961. A prairie to forest succession in Wood Buffalo National Park, Alberta. Ecology 42, 442e444.
Joly, D.O., and F. Messier. 2000. A numerical response of wolves to bison abundance in Wood Buffalo National Park, Canada. Canadian Journal of Zoology 78:1101–1104.
Joly, D.O. and F. Messier. 2004a. Factors affecting apparent evidence of tuberculosis and brucellosis in Wood Bison. Journal of Animal Ecology 73:623-631.
Joly, D.O. and F. Messier. 2004b. Testing hypotheses of bison population decline, 1970–1999, in Wood Buffalo National Park synergism between exotic disease and predation. Canadian Journal of Zoology 82:1165-1176.
Joly, D.O. and Messier, F. 2005. The effect of bovine tuberculosis and brucellosis on reproduction and survival of Wood Bison in Wood Buffalo National Park. Journal of Animal Ecology 74:543-551.
Jonas, J.L. and Joern, A. 2007. Grasshopper (Orthoptera: Acrididae) communities respond to fire, bison grazing and weather in North American tallgrass prairie: a long term study. Oecologia 153:699-711.
Joynt, B. 2010. Manitoba's wild Wood Bison. Nature Manitoba News 2(6): pages 1 and 10. Official publication of Nature Manitoba, Winnipeg.
Jung, T.S. 2011. Gray wolf (Canis lupus) predation and scavenging on reintroduced American Bison (Bison bison) in southwestern Yukon. Northwestern Naturalist 92:126–130.
Jung, T.S., and S.M. Czetwertynski. 2013. Niche overlap and potential competition between reintroduced bison and other ungulates in southwestern Yukon, Canada. Yukon Department of Environment, Whitehorse, Yukon. TR-13-15. 144 pp.
Jung, T.S., and K. Egli. 2012. Population status of the Aishihik Wood Bison (Bison bison athabascae) population in southwestern Yukon. Technical Report TR-12-19. Yukon Department of Environment, Whitehorse, YT.
Jung, T.S., P.M. Kukka, and A. Milani. 2010. Bison (Bison bison) fur used as drey material by red squirrels (Tamiasciurus hudsonicus): An indication of ecological restoration. Northwestern Naturalist 91:220–222.
Jung, T.S., K. Kuba and K. Egli. 2012. Spatial and temporal patterns in the harvest of Wood Bison (Bison bison athabascae) from southwestern Yukon. Unpublished report. Yukon Department of Environment, Whitehorse, Yukon.
Kay, C.E., C.A. White, I.R. Pengelly and B. Patton. 1999. Long-term ecosystem states and processes in Banff National Park and the central Canadian Rockies. Parks Canada Occasional Report 9. Ottawa, ON.
Kenefic LJ, T. Pearson, R.T. Okinaka, J.M. Schupp, D.M. Wagner, J. Ravel, A.R. Hffmaster, C.P. Trim, W-K Chung, J.A. Beaudry, J.F. Foster. J.I. Mead and P. Keim. 2009. Pre-Columbian origins for North American anthrax. PLoS ONE 4(3): e4813. doi:10.1371/journal.pone.0004813.
Kessel, J.C. van. 2002. Taking care of bison : community perceptions of the Hook Lake Wood Bison Recovery Project in Fort Resolution, NT, Canada. Master of Science Thesis, University of Alberta.
Kirkpatrick, J.F., D.F. Gudermuth, R.L. Flagan, J.C. McCarthy, and B.L. Lasley. 1993. Remote monitoring of ovulation and pregnancy of Yellowstone bison. Journal of Wildlife Management 57:407-412.
Kirkpatrick, J. F., J. C. McCarthy, D. F. Gudermuth, S. E. Shideler, and B. L. Lasley. 1996. An assessment of the reproductive biology of Yellowstone bison (Bison bison) subpopulations using noncapture methods. Canadian Journal of Zoology 74:8–14.
Knapp, A.K., J.M. Blair, J.M. Briggs, S.L. Collins, D.C. Hartnett, L.C. Johnson and E.G. Towne. 1999. The keystone role of bison in North American tallgrass prairie. Bioscience 49:39-50.
Knight, C. 1849. Sketches in natural history: History of the mammalia. London: C. Cox.
Knopf, F.L. 1996. Prairie legacies-Birds. In F.B. Samson and F.L. Knopf (Eds.), Prairie Conservation: Preserving North America’s Most Endangered Ecosystem, pp. 135-148. Covelo, CA: Island Press.
Komers, P.E., F. Messier and C.C. Gates. 1992. Search or relax: the case of bachelor Wood Bison. Behavioral Ecology and Sociobiology 31:195-203.
Komers, , P.E., F. Messier and C.C. Gates. 1993. Group structure in Wood Bison: nutritional and reproductive determinants. Canadian Journal of Zoology 71: 1367-1371.
Komers, P.E., F. Messier and C.C. Gates. 1994a. Plasticity of reproductive behaviour in Wood Bison bulls: when subadults are given a chance. Ethology Ecology & Evolution 6:313-330.
Komers, P.E., F. Messier and C.C. Gates. 1994b. Plasticity of reproductive behaviour in Wood Bison bulls: on risks and opportunities. Ethology Ecology & Evolution 6: 481-495.
Krueger, K. 1986. Feeding relationships among bison, pronghorn, and prairie dogs: an experimental analysis. Ecological Applications 67:760-770.
Langemann, E.G. 2004. Zooarchaeological research in support of a reintroduction of bison to Banff National Park, Canada. Proceedings of the Conference of the International Conference of Archaeozoology 9:79-89.
Larter, N. and D. Allaire. 2007. History and current status of the Nahanni Wood Bison population. Department of Environment and Natural Resources, Government of the Northwest Territories. File Report no. 136. Yellowknife, NWT. 37pp.
Larter, N.C., D.G. Allaire, and T.S. Jung. 2007. Population survey of the Nahanni Wood Bison population, March 2004. NWT Department of Environment and Natural Resources File Report No. 176. Yellowknife, Northwest Territories. 21 pp.
Larter, N.C. and D.G. Allaire. 2013. Population Survey of the Nahanni Wood Bison Population, March 2011. Environment and Natural Resources, Government of the Northwest Territories. Manuscript Report No. 229.
Larter, N.C. and Gates, C.C. 1990. Home ranges of Wood Bison in an expanding population. Journal of Mammalogy 71:604-607.
Larter, N.C., and C.C. Gates. 1991a. Diet and habitat selection of Wood Bison in relation to seasonal changes in forage quantity and quality. Canadian Journal of Zoology 69:2677–2685.
Larter, N.C., and C.C. Gates. 1991b. Seasonal selection of resources by Wood Bison in the Mackenzie Bison Sanctuary, Northwest Territories, Canada. in Global trends in wildlife management: 131–135. B. Bobek, K. Perzanowski and, W. Regelin, eds. Krakow-Warszawa: Swiat Press.
Larter, C.P.S. and MacDonald, G.M., 1998. An 840-year record of fire and vegetation in a boreal white spruce forest. Ecology 79, 106e118.
Larter, N.C., J.S. Nishi, T. Ellsworth, D. Johnson, G. More, and D.G. Allaire. 2003. Observations of Wood Bison swimming across the Liard River, Northwest Territories, Canada. Arctic 56:408-412.
Larter, N.L., A.R.E. Sinclair, and C.C. Gates. 1994 The response of predators to an erupting bison, Bison bison athabascae, population. Canadian Field-Naturalist 108: 318-327.
Larter, N.C., A.R.E. Sinclair, T. Ellsworth, J. Nishi, and C.C. Gates. 2000. Dynamics of reintroduction in an indigenous large ungulate: the Wood Bison of northern Canada. Animal Conservation 3:299-309.
Larter, N.C., A.R.E. Sinclair and C.C. Gates. 1994. The response of predators to an erupting Bison (Bison bison athabascae) population. Canadian Field-Naturalist 108:318-327.
Leverkus, S. 2012. Seasonal range use by Wood Bison in British Columbia. Prepared for: C.D. Thiessen, Ministry of Natural Resource Operations, British Columbia. Contract NRGS12FSJ-008
Lewis, H.T. 1977. "Maskuta: The Ecology of Indian Fires in Northern Alberta." Western Canadian Journal of Anthropology 7:15-52.
Lewis, H.T. 1980. Indian fires in spring: Hunters and gatherers of the Canadian forest shaped their habitat with fire. Natural History 89: 76-78, 82-83.
Lewis, H.T. 1982: A time for burning. Boreal Institute of North Studies. Occasional Publication 17.
Li, H., D.T. Shen, D.A. Jessup, D.P. Knowles, J.R. Gorham, T. Thorne, D. O’Toole, and T.B. Crawford. 1996. Prevalence of antibody to malignant catarrhal fever virus in wild and domesticated ruminants by competitive-inhibition ELISA. Journal of Wildlife Diseases 32:437-43.
Lieffers, V.J., Stadt, K.J., 1994. Growth of understory Picea glauca, Calama- grostis canadensis, and Epilobium angustifolium in relation to overstory light transmission. Can. J. For. Res. 24, 1193–1198.
Linnaeus, C. 1758. Tomus I. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata. Holmiae. (Laurentii Salvii).: [1-4], 1-824.
List, R., G. Ceballos, C. Curtin, P. Gogan, J. Pacheco and J. Truett. 2007. Historic distribution and challenges to bison recovery in the Northern Chihuahuan Desert. Conservation Biology 21:1487-1494.
Lotenberg, G. 1996. History of Wood Bison in the Yukon - a Re- Evaluation Based on Traditional Knowledge and Written Records. Report for Yukon Department of Renewable Resources by Boreal Research Associates, Whitehorse Yukon Territory.
Lothian, W.F. 1981. A History of Canada’s National Parks, Volume IV. Parks Canada. Minster of Supply and Services, Canada, Ottawa, Ontario.
Lott, D.F. 1972. Bison would rather breed than fight. Natural History 81:40-45.
Lott, D.F. 2002. American Bison. A Natural History. University of California Press, Berkeley and Los Angeles.
Louis Riel Institute. 2011. Buffalo Hunt.
MacGregor, J.G. 1952. The land of twelve foot Davis. A history of the Peace River Country. The Institute of Applied Art Ltd. Edmonton, Alberta. 394 pp.
Mahan, B.R. 1978. Aspects of American Bison (Bison bison) social behaviour at fort Niobara National Wildlife Refuge, Valentine Nebraska, with special reference to calves. M.Sc. Thesis, University of Nebraska, Lincoln, Nebraska, USA.
Mahan, B.R., M.P. Munger and H.L. Gunderson. 1978. Analysis of the flehmen display in American Bison (Bison bison). Prairie Naturalist 10:33-42.
Maher, C.R. and Byers, J.A. 1987. Age-related changes in reproductive effort of male bison. Behavioral Ecology and Sociobiology 21:91-96.
Malainey, M.E. and Sherriff, B.L. 1996. Adjusting our perceptions: historical and archaeological evidence of winter on the plains of western Canada. Plains Anthropologist 41(158):333-357.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay, and A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk. NatureServe, Arlington, VA. [Accessed Aug. 23, 2010]
Matlack, R.S., D.W. Kaufman, and G.A. Kaufman. 2001. Influence of grazing by bison and cattle on deer mice in burned tallgrass prairie. American Midland Naturalist 146: 361-368.
Matthews, S.B. 1992. An assessment of bison habitat in the Mills/Mink Lakes area, Northwest Territories, using landsat thematic mapper data. Arctic 44:75–80.
McCoy, V.M., and C.R. Burn. 2005. Potential alteration by climate change of the forest-fire regime in the boreal forest of central Yukon Territory. Arctic, 58:276–285.
McDonald, J.N. 1981. North American Bison, Their Classification and Evolution. University of California Press, Berkeley, California. 316 pp.
McFarlane, K., G.A. Wilson and J. Nishi. 2006. Management strategies for conservation of genetic diversity in Wood Bison (Bison bison athabascae). File report 135. Department of Environment & Natural Resources, Government of the Northwest Territories, Fort Smith, NT.
McHugh, T. 1958. Social behavior of the American buffalo (Bison bison bison). Zoologica 43:1– 40.
McHugh, T. 1972. The time of the buffalo. Alfred A. Knopf, New York.
Meagher, M.M. 1973. The bison of Yellowstone National Park. National Park Service Scientific Monograph Series 1:1-161.
Meagher, M.M. 1989. Range expansion by bison of Yellowstone National Park. Journal of Mammalogy 70:670–685.
Menzel, C.R. and B.B. Beck. 2000. Homing and detour behavior in golden lion tamarin social groups. In: On the Move (Ed. by S. Boinski & P.A. Garber), pp. 299–326. Chicago University Press, Chicago.
Merkle, J.A. and D. Fortin. In press. Likelihood-based photograph identification: application with photographs of free-ranging bison. Wildlife Society Bulletin.
Messier, F. 1994. Ungulate population models with predation: A case study with the North American Moose. Ecology 75:478–488.
Mitchell, J.A. and C.C. Gates. 2001. Status of the Wood Bison (Bison bison athabascae) in Alberta. Alberta Wildlife Status Report No. 38. Fish & Wildlife Division, Resource Status and Assessment Branch, Edmonton, Alberta.
Morgan, R.G. 1980. Bison movement patterns on the Canadian plains: an ecological analysis. Plains Anthropologist. 25:143-160.
Moritz, C. 2002. Strategies to protect biological diversity and the evolutionary processes that sustain it. Systematic Biology 5:238-254.
NatureServe. 2011. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. (Accessed: 15 February 2012 ).
Neufeld, P.L. 1992. Bison conservation: The Canadian story. Manitoba History 24: pages revised. www.mhs.mb.ca/docs/mb_history/24/bisonconservation.shtml
Nishi, J. 2004. Bison-traffic collisions on Highway 3 (1989 – 2003). Unpublished Report. Government of NWT. 27 pp.S. 2002. Surveillance activities under the Northwest Territories Bison Control Area Program (1987 – 2000). Northwest Territories Department of Resources, Wildlife and Economic Development Manuscript Report No. 145. Yellowknife, NT. 32 pp.
Nishi, J.S. 2010. A review of best practices and principles for bison disease issues: Greater Yellowstone and Wood Buffalo Areas. Wildlife Conservation Society and American Bison Society. American Bison Society Working Paper No. 3. 101 pp.
Nishi, J. S., D. C. Dragon, B. T. Elkin, J. Mitchell, T. R. Ellsworth and M. E. Hugh-Jones. 2002b. Emergency response planning for anthrax outbreaks in bison herds of northern Canada: a balance between policy and science. Annals of the New York Academy of Sciences 969: 245-250.
Nishi, J. S., T. R. Ellsworth, N. Lee, D. Dewar, B. T. Elkin and D. C. Dragon. 2007. Northwest Territories. An outbreak of anthrax (Bacillus anthracis) in free-roaming bison in the Northwest Territories, June-July 2006. Canadian Veterinary Journal 48: 37-38.
Nishi, J.S., T. Shury, and B.T. Elkin. 2006. Wildlife reservoirs for bovine tuberculosis (Mycobacterium bovis) in Canada: strategies for management and research. Veterinary Microbiology 112: 325-338.
Nishi, J.S., C. Stephen, and B.T. Elkin. 2002a. Implications of agricultural and wildlife policy on management and eradication of bovine tuberculosis and brucellosis in free-ranging wood bison of northern Canada. Annals of the New York Academy of Sciences 969: 236 – 244.
Northwest Territories Environment and Natural Resources. 2010. Wood Bison management strategy for the Northwest Territories. Environment and Natural Resources, Yellowknife.
Novakowski, N. and W.E. Stevens. 1965. Survival of the Wood Bison (Bison bison athabascae Rhoads) in Canada. Paper presented to the 45th meeting of the American Society of Mammalogists, Winnipeg, 21 June 1965.
Ogilvie, W. 1893. Report on the Peace River and tributaries in 1891. Annual report Department of Interior Canada for 1892. Part 7, 144 pp.
Ogilvie, S.C. 1979. The park buffalo. Calgary - Banff Chapter National and Provincial Parks Association of Cnada, The Universityof Calgary, Calgary, AB.
Olech, W. 1987. Analysis of inbreeding in European bison. Acta Theriologica 32: 373–387.
Olson, W. and R. Sissons. 2011. Plains Bison Monitoring Resource Conservation Annual Report: Grasslands National Park of Canada. Unpublished Parks Canada Report.
Parks Canada Agency. 2009. Ungulate Options Analysis - Final Report Elk Island National Park of Canada.
Parks Canada Agency. 2012. Free-ranging plains bison census 1996-2012. Prince Albert National Park. Waskesiu Lake, Saskatchewan. Canada. Unpublished files.
Payne, C.H. 1987. The Waterhen project: The introduction of the endangered Wood Bison to the Interlake Region of Manitoba and its role in the economic development of the Waterhen Band of Saulteaux Indians. PhD dissertation, University of Manitoba.
Pertoldi, C, M. Tokarska, J.M. Wójcik, et al. 2010. Phylogenetic relationships among the European and American Bison and seven cattle breeds reconstructed using the BovineSNP50 Illumina Genotyping BeadChip. Acta Theriologica 55:97-108.
Peters, H.F. and Slen, S.B. 1964. Hair coat characteristics of bison, domestic cattle x bison hybrids, cattalo, and certain domestic breeds of beef cattle. Canadian Journal of Animal Science 44:48- 57.
Polley, H.W. and S.L. Collins. 1984. Relationship of vegetation and environment in buffalo wallows. American Midland Naturalist 112:178-186.
Polziehn, R.O., R. Beech, J. Sheraton and C. Strobeck. 1996. Genetic relationships among North American Bison populations. Canadian Journal of Zoology 74:738-49.
Polziehn, R.O., C. Strobeck, J. Sheraton, and R. Beech. 1995. Bovine mtDNA discovered in North American Bison populations. Conservation Biology 9:1638-1643.
Potter, B.A., S. Craig Gerlach, and C. Cormack Gates, Delaney P. Boyd, Gerald A. Oetelaar, and James H. Shaw. 2010. History of bison in North America. Pages 5-12 in Gates, C. C., C.F. Freese, P. Gogan and M. Kotzman (eds). American Bison: Status survey and conservation guidelines. International Union for the Conservation of Nature, Gland, Switzerland.
Powell, T. and T. Morgan 2010. Ronald Lake bison (Bison bison) survey, February 2010. Unpublished report. Alberta Sustainable Resource Development Wildlife Division Fort McMurray, Alberta November 2010
Preisser, E.L. and D.I. Bolnick. 2008. When predators don't eat their prey: nonconsumptive predator effects on prey dynamics. Ecology 9:2414-2415
Quinlan, A., M.A. Dale and C.C. Gates. 2003. Effects of prescribed burning on herbaceous and woody vegetation in northern lowland meadows. Restoration Ecology 11:343-350.
Raup, H. M. 1933. Range conditions in the Wood Buffalo Park of western Canada with notes on the history of the Wood Bison (Special Publication 1[2]). American Committee for International Wildlife Protection.
Raup, H.M., 1947. The Botany of Southwestern Mackenzie, Arnold Arboretum Contribution VI. Harvard University, Jamaica Plain.
Redburn, M.J., W.L. Strong and C.C. Gates. 2008. Suitability of boreal mixedwood clearcuts as Wood Bison (Bison bison athabasca) foraging habitat in north-central Alberta, Canada. Forest Ecology and Management 225:2225-2235.
Reeves, B.0.K. 1978. Head-Smashed-In: 5500 years of bison jumping in the Alberta plains. Plains Anthropologist 23:151-174.
Renecker, L.A., Blyth, C.B. and Gates, C.C. 1989. Game production in western Canada. In: R.J. Hudson, K.R. Drew and L.M. Baskin (eds.), Wildlife Production Systems: Economic Utilization of Wild Ungulates, pp.248-267. Cambridge University Press, New York.
Reynolds, H.W., C.C. Gates and R.D. Glaholt. 2003. Bison (Bison bison). In: G. A. Feldhamer, B.C. Thompson, and J. A. Chapman (Eds.), Wild Mammals of North America: Biology, Management, and Conservation Second Edition, pp.1009-1059. Baltimore: The Johns Hopkins University Press.
Reynolds, H. W., R. M. Hansen, and D. G. Peden. 1978. Diets of the Slave River Lowland bison herd, Northwest Territories, Canada. Journal of Wildlife Management 42:581–90.
Rhoads, S.N. 1897. Notes on living and extinct species of North American Bovidae. Proceedings of the Academy of Natural Sciences of Philadelphia, 49:483-502.
Ricketts, T. H., E. Dinerstein, D.M. Olson, C.J. Loucks, W. Eichbaum, D> DellaSalla, K. Kavanagh, P. Hedao, P. Hurley, K. Carney, R. Abell, and S. Walters. 1999. Terrestrial ecoregions of North America: a conservation assessment. Island Press, Washington, D.C.
Riley, J.L., S.E. Green and K.E. Brodribb. 2007. A conservation blueprint for Canada's prairies and parklands. Nature Conservancy of Canada, Toronto, Ontario. 226 pp.
Roe, F.G. 1970. The North American Buffalo. A Critical Study of the Species in its Wild State. Second edition. University of Toronto Press, Toronto, Ontario, Canada.
Rosas, C.A., D.M. Engle and J.H. Shaw. 2005. Potential ecological impact of diet selectivity and bison herd composition. Great Plains Research 15:3-13.
Rosas, C.A., D.M. Engle, J.H. Shaw and M.W. Palmer. 2008. Seed dispersal by Bison bison in tallgrass prairie. Journal of Vegetation Science 19: 769-778.
Rothstein, A., and J.G. Griswold, J.G. 1991. Age and sex preferences for social partners by juvenile bison bulls, Bison bison. Animal Behaviour 41:227-237.
Rowe, M. 2006. 2006 Halfway-Sikanni Plains Bison inventory. BC Ministry of Environment, Fort St. John, BC. 14 pp.
Rowe, M. 2007. 2007 Nordquist Wood Bison inventory. Peace Region Technical Report. Ministry of Environment 400 – 10003 110th Avenue Fort St. John, British Columbia.
Rowe, M. and R. Backmeyer. 2006. Etthithun Wood Bison inventory: March 2006. BC Ministry of Environment, Fort St. John, BC. 6 pp.
Ruckstuhl, K.E. 2000 (In Press). Update COSEWIC status report on the wood bison Bison bison athabascae in COSEWIC assessment and status report on wood bison Bison bison athabascae in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-28 pp.
Rutley, B. and Hudson R.J. 2000. Seasonal energetic parameters of free-grazing bison (Bison bison). Canadian Journal of Animal Science 80(4):663-671.
Sachro, L.L., W.L. Strong and C.C. Gates. 2005. Prescribed burning effects on summer elk forage availability in the subalpine zone, Banff National Park, Canada. Journal of Environmental Manageemnt 77:183-193.
SRPB (Sturgeon River Plains Bison Stewards) Management Planning Coordinating Committee. 2013. Sturgeon River Plains Bison Management Plan. March 2013.
Sanderson, E.W., K. Redford, B. Weber, K. Aune, D. Baldes, J. Berger, D. Carter, C. Curtin, J. Dear, S. Dobrott, E. Fearn, C. Fleener, S. Forrest, C. Gewach, C.C. Gates, J.E. Gross, S. Grassel, J.A. Hilty, M. Jensen, K. Kunkel, D. Lammers, U. List, K. inkowski, T. Olson, C. Pague, P.B. Robertson and B. Stevenson. 2008. The ecological future of the North American Bison: conceiving long-term, large scale conservation of wildlife. Conservation Biology, 22:252-266.
Saskatchewan Environment. 2005. Saskatchewan State of the Environment Report 2005.
Saskatchewan Environment and Resource Management. 2000. Primrose Lake and McCusker River Representative Areas Concept Management Plan. Saskatchewan Environment and Resource Management December 2000.
Saunders, W.E. 1925. Letter to the editor of the Canadian Field-Naturalist. The Canadian Field Naturalist 39:118.
Schaefer, A. L., B. A. Young, and A. M. Chimwano. 1978. Ration digestion and retention times of digesta in domestic cattle (Bos taurus), American Bison (Bison bison), and Tibetan yak (Bos grunniens). Canadian Journal of Zoology 56:2355–58.
Schramm T., in cooperation with; Krogman N., Hudson R.J., Freeman M.M.R. 2002. Caribou Mountains critical ungulate habitat and Traditional Ecological Knowledge study: a GIS analysis, 2002, Sustainable Forest Management Network, University of Alberta, Edmonton, Alberta, Canada.
Schultz, J. 1888. The Great Mackenzie Basin. Report of the Select Committee of the Senate appointed to inquire into the resources of the Great Mackenzie Basin. Session 1888 Ottawa Ontario, Taken from the Senate Debates Ottawa 27th March 1888. Pages 218-244. (cited by Raup 1933).
Schultheiss, P.C., J.K. Collins, L.E. Austgen, and J.C. DeMartini. 1998. Malignant catarrhal fever in bison, acute and chronic cases. Journal of Veterinary Diagnostic Investigation, 10:255-262.
Schultheiss, P.C., J.K. Collins, T.R. Spraker, and J.C. DeMartini. 2000. Epizootic malignant catarrhal fever in three bison herds: differences from cattle and association with ovine herpesvirus-2.. Journal of Veterinary Diagnostic Investigation, 12:497-502.
Schwartz, C.C., and J.E. Ellis 1981. Feeding ecology and niche seperation in some native and domestic ungulates on the shortgrass prairie. Journal of Applied Ecology 18: 343-353.
Schwartz, A.G., and R.W. Wein. 1997. Threatened dry grasslands in the continental boreal forest of Wood Buffalo National Park. Canadian Journal of Botany 75:1363-1370.
Shapiro, B., A.J. Drummond, A. Rambaut, M.C. Wilson, P.E. Matheus, A.V. Sher, O.G. Pybus, M.T. Gilbert, I. Barnes, J. Binladen, E. Willerslev, A.J. Hansen, G.F. Baryshnikov, J.A. Burns, S. Davydov, J.C. Driver, D.G. Froese, C.R. Harington, G. Keddie, P. Kosintsev, M.L. Kunz, L.D. Martin, R.O. Stephenson, J. Storer, R. Telford, S. Zimov and A. Cooper. 2004. Rise and fall of the steppe bison. Science 306:1561-1565.
Shaw, J.H. 1995. How many bison originally populated western rangelands? Rangelands 17(5):148-150.
Shaw, J. H., and T. S. Carter. 1989. Calving patterns among American Bison. Journal of Wildlife Management 53:896-898.
Shaw, J.H. and Lee, M. 1997. Relative abundance of bison, elk, and pronghorn on the southern plains 1806-1857. Plains Anthropologist 42:163-172.
Sheldon, J.W., G. Reed, A.C. Burnett, K. Li, and R.L. Crabtree. 2009. Coyote, Canis latrans, predation on a bison, Bison bison, calf in Yellowstone National Park. Canadian Field-Naturalist 123: 260-261.
Shull, A.M. 1985. Age-specific reproductive behaviour of male American Bison. M. Sc. Thesis, Oklahoma State University, Stillwater, Oklahoma, USA.
Shult, M.J. 1972. American Bison behavior patterns at Wind Cave National Park. Ph.D. Thesis, Iowa State University, Ames, Iowa, USA.
Shury, T.K., D. Fransden and L. O'Brodovich. 2009. Anthrax in free-ranging bison in the Prince Alberta NationalPark area of Saskatchewan in 2008. The Canadian Veterinary Journal 50:152-154.
Shury, T.K., Woodley, S.J. and Reynolds, H.W. 2006. Proceedings of the Bison Diseases Technical Workshop, October 28,29, 2005. Parks Canada, Gatineau, Quebec.
Sibley, J.A., M.R. Woodbury, G.D. Appleyard, and B. Elkin. 2007. Mycobacterium avium subspecies paratuberculosis in bison (Bison bison) from northern Canada. Journal of Wildlife Diseases 43: 775-779.
Siebert, V.F. 1925. Some notes on Canada's so-called wood buffalo. The Canadian Field-Naturalist 39: 204-206.
Singer, B. 2005. Muddying the Gene Pool. Canadian Geographic.
Singer, F. J., and J. E. Norland. 1994. Niche relationships within a guild of ungulate species in Yellowstone National Park, Wyoming, following release from artificial controls. Canadian Journal of Zoology 72:1383–1394.
Skinner, M.F. and O.C. Kaisen. 1947. The fossil bison of Alaska and preliminary revision of the genus. Bulletin of the American Museum of Natural History 89:123-256.
Smith, D., D. Mech, M. Meagher, W. Clary, R. Jaffe, M. Phillips and J. Mack. 2000. Wolf-bison interactions in Yellowstone National Park. Journal of Mammalogy 81:1128-1135.
Soper, J.D. 1941. History, range and home life of the northern bison. Ecological Monographs 11:347-412.
Soule, M.E., Estes, J.A., Berger, J., Martinez del Rio, C., 2003. Ecological effectiveness: conservation goals for interactive species. Conservation Biology 17: 1238–1250.
Stephenson, R.O., S.C. Gerlach., R.D. Guthrie., C.R. Harington., R.O. Mills. and G. Hare, 2001. Wood Bison in late Holocene Alaska and adjacent Canada: Paleontological, archaeological and historical records. In: S.C. Gerlach and M.S. Murray (Eds.), People and Wildlife in Northern North America: Essays in Honor of R. Dale Guthrie, pp.124158. BAR International Series 944. Oxford: British Archaeological Reports.
Stocks BJ, J.A. Mason, J.B. Todd, E.M. Bosch, B.M. Wotton, B.D. Amiro, M.D. Flannigan, K.G. Hirsch, K.A. Logan, D.L. Martell and W.R. Skinner. 2002. Large forest fires in Canada, 1959–1997. Journal of Geophysical Research 107: 8149, doi:10.1029/2001JD000484.
Strong, W.L., and C.C. Gates. 2009. Wood Bison population recovery and forage availability in northwestern Canada. Journal of Environmental Management 90:434-440.
Strong, W.L., M.J. Redburn and C.C. Gates. 2009. Climate and vegetation change during the twentieth century in the Lower Peace River district, northern Alberta, Canada. The Holocene:119:199-207.
Telfer, E. S., and A. L. Cairns. 1986. Resource use by moose versus sympatric deer, wapiti and bison. Alces 22:113–38.
Telfer, E.S. and J.P. Kelsall. 1979. Studies of morphological parameters affecting ungulate locomotion in snow. Canadian Journal of Zoology 57:2153-59.
Telfer, B.S. and J.P. Kelsall. 1984. Adaptation of some large North American mammals for survival in snow. Ecology 65:1828-34.
Spyce, T. 2009. Disruption in Place Attachment: Insights of young Aboriginal adults on the social and cultural impacts of industrial development in Northern Alberta. Master Sc. Thesis, University of Alberta, Alberta, Canada. 226 pp.
Tessaro, S.V. 1988. A descriptive and epizootiologic study of brucellosis and tuberculosis in bison in northern Canada. Ph.D. thesis. University of Saskatchewan, Saskatoon, SK. 320 pages.
Tessaro, S.V. 1989. Review of the diseases, parasites, and miscellaneous pathological conditions of North American Bison. Canadian Veterinary Journal, 31, 174–180.
Thiessen, C. 2010. Peace Wood Bison project: annual report 2009/2010. Peace Region Technical Report. BC Ministry of Environment. Fort St. John, British Columbia. 22 pages.
Thorne, E. T. 2001. Brucellosis. In Infectious diseases of wild mammals, E. S. Williams and I. K. 1. Barker (eds.). Iowa State Press, Ames, Iowa, pp. 372–395.
Timoney, K. 2002. A dying delta? A case study of a wetland paradigm. Wetlands, 22: 282-300.
Timoney, K. 2006. Landscape cover change in the Peace-Athabascae Delta, 1927-2001. Wetlands, 26: 765-778.
Timoney, K. 2008a. Rates of vegetation change in the Peace-Athabasca Delta. Wetlands 28:513-520.
Timoney, K. 2008b. Factors influencing wetland plant communities during a flood-drawdown cycle in the Peace-Athabascae Delta, northern Alberta, Canada. Wetlands 28: 450-463.
Timoney, K. 2009. Three centuries of change in the Peace-Athabasca Delta, Canada. Climatic Change 93:485-515.
Timoney, K., G. Peterson, P. Fargey, M. Peterson, S. McCanny, and R. Wein. 1997. Spring ice-jam flooding of the Peace- Athabasca Delta: evidence of a climatic oscillation. Climatic Change 35:463–83.
Truett, J.C., M. Phillips, K.Kunkel, and R. Miller. 2001. Managing bison to restore biodiversity. Great Plains Research 11: 123-144.
Trut, L., I. Oskina, and A. Kharlamova. 2009. Animal evolution during domestication: the domesticated fox as a model. BioEssays 31:349–360.
Turtle Mountain – Souris Plains Heritage Association (TM-SPHA). 2008. Metis Bison Hunts.
USDA Forest Service. 2009. Species Conservation Program: American Bison Assessment and Rationale.
van Zyll de Jong, C.G. 1986. A Systematic Study of Recent Bison, with Particular Consideration of the Wood Bison (Bison bison athabascae Rhodes 1989). Publications in Natural Sciences No. 6. National Museums of Canada, Ottawa, Ontario.
van Zyll de Jong, C.G. 1993. Origin and geographic variation of recent North American Bison. Alberta: Studies in the Arts and Sciences 3(2):21-35.
van Zyll de Jong, C.G., C. Gates, H. Reynolds and W. Olson. 1995. Phenotypic variation in remnant populations of North American Bison. Journal of Mammalogy 76:391-405.
Vassal, M. and R. Kindopp. 2010. Wood Buffalo National Park Bison Survey, February 2009. Unpublished Parks Canada Report.
Virchow, D.R. and Hygnstrom, S.E. 2002. Estimation of presettlement populations of the black-tailed prairie dog: a reply. Great Plains Research 12:255-260.
Waggoner, V., and M. Hinkes. 1986. Summer and fall browse utilization by an Alaskan bison herd. Journal of Wildlife Management 50: 322-324.
Ward, T.J., J.P. Bielawski, S.K. Davis, J.W. Templeton, and J.N. Derr. 1999. Identification of domestic cattle hybrids in wild cattle and bison species: a general approach using mtDNA markers and the parametric bootstrap. Animal Conservation 2:51-57.
White, C.A., E.G. Langemann, C.C. Gates, C.E. Kay, T. Shury and T.E. Hurd. 2001. Plains Bison restoration in the Canadian Rocky Mountains? Ecological and management considerations. Proceedings George Wright Society Annual Conference on Research and Resource Management in Parks and on Public Lands 11:152-160.
White, C.A., D.D. Perrakis, V.G. Kafka and T. Ennis. 2012. Burning at the edge: Integrating biophysical and eco-cultural fire processes in Canada's parks and protected areas. Fire Ecology 7:74-106.
Wilson, G.A. 2001. Population genetic studies of wood and Plains Bison populations. Unpublished Ph.D. thesis. University of Alberta, Edmonton, Alberta. 156 pp.
Wilson, M.C. 1996. Late Quaternary vertebrates and the opening of the ice-free corridor, with special reference to the genus Bison. Quaternary International 32:97-105.
Wilson, M.C., L. Hills and B. Shapiro. 2008. Late Pleistocene northward-dispersing Bison antiquus from the Bighill Creek Formation, Gallelli Gravel Pit, Alberta, Canada, and the fate of Bison occidentalis. Canadian Journal of Earth Sciences 45:827-859.
Wilson, G., J. Nishi, B. Elkin, and C. Strobeck. 2005. Effects of a recent founding event and intrinsic populations dynamics on genetic diversity in an ungulate population. Conservation Genetics 6: 905-916.
Wilson, G.A., W. Olson, and C. Strobeck. 2002. Reproductive success in Wood Bison (Bison bison athabascae) established using molecular techniques. Canadian Journal of Zoology 80:1537-1548.
Wilson, D.E. and D.M. Reeder (eds.), 2005. Mammal Species of the World. A Taxonomic and Geographic Reference, Third edition. Johns Hopkins University Press, Baltimore, Maryland.
Wilson, G.A. and C. Strobeck. 1999. Genetic variation within and relatedness among wood and Plains Bison populations. Genome 42:483-496.
Wilson, G.A. and K. Zittlau. 2004. COSEWIC status report on Plains Bison, Bison bison bison. 60 pages + appendices.
Wissler, C. 1927. North American Indians of the Plains. Third Edition. American Museum of Natural History, New York.
Wolff, J.O. 1998. Breeding strategies, mate choice, and reproductive success in American Bison. Oikos 83:529-544.
Wolfe, M.L., and J.F. Kimball. 1989. Comparison of bison population estimates with a todal count. Journal of Wildlife Management 53:593-596.
Wood Bison Recovery Team. 1987. Status report on Wood Bison (Bison bison athabascae) in Canada, 1987. Prepared for the Recovery of Nationally Endangered Wildlife Program, Ottawa.
Woodbury M, and C. Windeyer. 2012. Mycoplasma infection in bison. Bison Producers of Alberta. .
Woodhouse, C.A., J.J. Lukas, and P.M. Brown. 2002. Drought in the western Great Plains, 1845–56. Bulletin of the American Meteorological Society, 83:1485-1493.
Wyman, T. 2002. Grizzly bear predation on a bull bison in Yellowstone National Park. Ursus 13: 375-377.
Yukon Department of Renewable Resources. 1998. Yukon bison management plan. Yukon Department of Environment; Whitehorse, Yukon.
Biographical Summary of Report Writer(s)
Cormack Gates is a professor of environmental science and planning in the Faculty of Environmental Design at the University of Calgary. He holds a PhD (1980) in Animal Science (Wildlife Productivity and Management) from the University of Alberta. He is engaged in research and private practice in rural land use planning, impact assessment, conservation design and multi-stakeholder engagement in planning. Prior to taking a position at the University of Calgary in 1998, he spent 18 years working with communities in northern Canada on wildlife and habitat management. He has facilitated numerous collaborative research, multi-stakeholder and management planning initiatives in northern and western Canada and the United States and has actively participated in bison research and conservation since 1983. He is the principal author of Canada’s 2001 National Recovery Plan for Wood Bison, co-author of the Alberta species at risk status report for this animal, and senior author of a report on bison movements and ecology in and beyond Yellowstone National Park prepared for the United States National Park Service. He is the lead editor and author of several chapters of the 2010 IUCN publication ‘American bison status survey and conservation guidelines’. Dr. Gates chaired, and then co-chaired, Canada’s National Wood Bison Recovery Team between 1986 and 2008. He chaired the IUCN Species Survival Commission - Bison Specialist Group for North America between 2001 and 2011 and currently serves as the IUCN Red List authority for the American bison.
Thomas Jung earned an M.Sc. in Wildlife Biology from McGill University. He has worked in varied locations across Canada conducting field-based research on a variety of vertebrates. He has been the Senior Wildlife Biologist for the Yukon government for the past 12 years, where he primarily works on species at risk and biodiversity issues. Tom has been a member of COSEWIC since 2001, and was a member of RENEW from 2001-2008. Since 2003, Tom has been the National Coordinator for the general status assessment of the mammals of Canada. He has been the Co-Chair of national recovery teams for Wood Bison and northern mountain caribou, and also the Southern Lakes Wildlife Coordinating Committee. He is currently an Associate Editor for the Canadian Field-Naturalist and the Northwestern Naturalist, an Adjunct at the University of Alaska-Fairbanks, and a Research Associate at the University of New Mexico. He and his family live near Whitehorse, Yukon.
Collections Examined
None examined.
Appendix 1. Threats calculator results for Plains Bison
Threats Assessment Worksheet
- Species or Ecosystem
- Plains Bison
- Assessor(s):
- D. Fraser and C. Gates with input from jurisdictions in conf call
| Threat Impact | Threat Impact (descriptions) | Level 1 Threat Impact Counts high range |
low range |
|---|---|---|---|
| A | Very High | 2 | 2 |
| B | High | 0 | 0 |
| C | Medium | 1 | 1 |
| D | Low | 1 | 1 |
| - | Calculated Overall Threat Impact: | Very High | Very High |
Overall Threat Comments
5 populations of "wild by nature" plains bison being considered for this assessment: Pink Mountain (1000), Sturgeon River (200-250), McCusker (150), Elk Island NP (400), Grassland NP (320). Total 2070-2120
| Number | Threat | Impact (calculated) Criterion |
Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | - | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | - |
| 1.1 | Housing & urban areas | - | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | More land being bought up in the agricultural area close to Prince Albert, threat of turning into subdivisions cabins in the heart of where the Sturgeon River bison come out of the park. |
| 1.2 | Commercial & industrial areas | - | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | - |
| 2 | Agriculture & aquaculture | D | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | - |
| 2.1 | Annual & perennial non-timber crops | D | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | This is about habitat alienation as a result of croplands. habitat loss is probably near its maximum, not expecting incursion; not expecting more habitat loss from agriculture in the next 10 years. Mostly an issue in SK. |
| 2.3 | Livestock farming & ranching | D | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | Commercial grazing on public lands in Alberta and Manitoba an issue. These land uses preclude recovery on Crown land in these areas where bison being excluded from areas that are grazed by livestock |
| 3 | Energy production & mining | - | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | - |
| 3.1 | Oil & gas drilling | - | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | McCusker herd potential for oil and gas being a threat is increasing; in Grasslands this is limited compared to ranching activity. |
| 3.2 | Mining & quarrying | - | Negligible | Negligible (<1%) | Slight (1-10%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 4 | Transportation & service corridors | - | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | - |
| 4.1 | Roads & railroads | - | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | Herds are not located in high traffic areas and this is not expected to change much in the next 10 years |
| 5 | Biological resource use | A | Very High | Pervasive (71-100%) |
Extreme (71-100%) |
Moderate (Possibly in the short term, < 10 yrs) | - |
| 5.1 | Hunting & collecting terrestrial animals | A | Very High | Pervasive (71-100%) |
Extreme (71-100%) |
Moderate (Possibly in the short term, < 10 yrs) | McCusker herd has no protection, Prince Albert herd is kept low to avoid conflict with agriculture, Pink Mountain herd is hunted to keep population low, Elk Island (currently) and Grasslands National Parks (within next ten years) are controlled owing to small size of the parks. This is mostly about removal for management purposes and all Plains Bison are subject to this threat. Forestry operations and oil and gas activities are increasing in McCusker herd range with increasing access and likely increase in unregulated hunting. |
| 6 | Human intrusions & disturbance | - | Negligible | Small (1-10%) | Negligible (<1%) | High (Continuing) | - |
| 6.1 | Recreational activities | - | Negligible | Small (1-10%) | Negligible (<1%) | High (Continuing) | Recreation is encouraged in the two national park herds (hikers), Pink Mountain receives use by off-road vehicles and snowmobilers. Not as big a deal as for wood bison; bullets are more important for Plains Bison than the snowmobiles themselves. |
| 6.2 | War, civil unrest & military exercises | - | Not a Threat | Small (1-10%) | Neutral or Potential Benefit | High (Continuing) | McCusker: bombing range. There is probably some sort of displacement effect. The beneficial effect of excluding the public from the area should offset any negatives aspects. |
| 7 | Natural system modifications | - | Unknown | Pervasive (71-100%) |
Unknown | Unknown | - |
| 7.3 | Other ecosystem modifications | - | Unknown | Pervasive – Large (31-100%) | Unknown | Unknown | - |
| 8 | Invasive & other problematic species & genes | C | Medium | Large (31-70%) | Moderate (11-30%) | High (Continuing) | - |
| 8.1 | Invasive non-native/alien species | CD | Medium - Low | Restricted (11-30%) | Serious - Moderate (11-70%) | High (Continuing) | Invasive alien species, sheep nearby and cattle ranches. Diseases from interaction with domestics (sheep and cattle). Sheep farm right next to grasslands park, MCF is 100% fatal to bison. At Pink Mountain the threat is very low, but all others are next to agriculture so in a different situation. And there are other livestock diseases that bison are susceptible to. Severity (Helen Schwantze and/or Susan Kutz for details). |
| 8.2 | Problematic native species | D | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | Anthrax (assumed to be native for this exercise). Anthrax has been documented at Prince Albert National Park (100 animals killed in 2007). This threat also includes wolf predation, an increasing threat of wolf predation for both SK populations, and this may be rising. |
| 8.3 | Introduced genetic material | C | Medium | Large (31-70%) | Moderate (11-30%) | High (Continuing) | Mixing with feral or escaped commercial Bison a potential issue in Saskatchewan herds. Mixing with Wood Bison is a potential risk in NE BC that is being managed by limiting the distribution of adjacent Wood and Plains Bison. |
| 11 | Climate change & severe weather | A | Very High | Pervasive (71-100%) |
Extreme (71-100%) |
Moderate (Possibly in the short term, < 10 yrs) | - |
| 11.1 | Habitat shifting & alteration | - | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Fires and forestry create bison habitat. |
| 11.2 | Droughts | CD | Medium - Low | Large (31-70%) | Moderate - Slight (1-30%) |
Moderate (Possibly in the short term, < 10 yrs) | Maybe an issue in Grasslands National Park, particularly in the longer term. |
| 11.3 | Temperature extremes | AB | Very High - High | Pervasive (71-100%) |
Extreme - Serious (31-100%) |
Moderate (Possibly in the short term, < 10 yrs) | - |
| 11.4 | Storms & flooding | CD | Medium - Low | Pervasive - Restricted (11-100%) | Moderate - Slight (1-30%) | Moderate (Possibly in the short term, < 10 yrs) | - |
Appendix 2. Threats calculator results for Wood Bison
Threats Assessment Worksheet
- Species or Ecosystem
- Wood Bison
- Assessor(s):
- Bison Recovery Team, updated by D. Fraser and C. Gates
| Threat Impact | Threat Impact (descriptions) | Level 1 Threat Impact Counts high range |
low range |
|---|---|---|---|
| A | Very High | 0 | 0 |
| B | High | 1 | 1 |
| C | Medium | 2 | 2 |
| D | Low | 3 | 3 |
| - | Calculated Overall Threat Impact: | Very High | Very High |
| Number | Threat | Impact (calculated) Criterion |
Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | - | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | - |
| 1.1 | Housing & urban areas | - | - | Small (1-10%) | Serious (31-70%) | High (Continuing) | Yukon. Nahanni, Mackenzie, Aishihik, Nordquist, Hay-Zama. Habitat alienation from subdivisions |
| 1.2 | Commercial & industrial areas | - | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | Nahanni, Beaver Camp |
| 2 | Agriculture & aquaculture | D | Low | Small (1-10%) | Serious (31-70%) | High (Continuing) | - |
| 2.1 | Annual & perennial non-timber crops | - | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | Aishihik, North Alberta herds |
| 2.3 | Livestock farming & ranching | D | Low | Small (1-10%) | Extreme (71-100%) | High (Continuing) | Hayes-Zama, Chitek Lake (this relates to habitat alienation and not management response, i.e., removal of animals as a result of conflict with livestock/farming) |
| 3 | Energy production & mining | - | Negligible | Negligible (<1%) | Slight (1-10%) | High - Moderate | - |
| 3.1 | Oil & gas drilling | - | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | Facility related, needs more work to document and understand threat. Studies in oil field developed areas that have bison would be useful. |
| 3.2 | Mining & quarrying | - | Negligible | Negligible (<1%) | Slight (1-10%) | Moderate (Possibly in the short term, < 10 yrs) | S. of Wood Buffalo, oil sands mining. |
| 4 | Transportation & service corridors | D | Low | Large (31-70%) | Slight (1-10%) | High (Continuing) | - |
| 4.1 | Roads & railroads | D | Low | Large (31-70%) | Slight (1-10%) | High (Continuing) | All herds are in an area where road and railway impacts are going to be an issue |
| 4.2 | Utility & service lines | - | Unknown | Large - Restricted (11-70%) | Unknown | Unknown | - |
| 4.3 | Shipping lanes | - | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Barge traffic on Liard River |
| 4.4 | Flight paths | - | Negligible | Restricted (11-30%) | Negligible (<1%) | High (Continuing) | Nahanni and Wood BNP especially but most Wood Bison subjected to overflights but 11-30, most of the time there’s no response. |
| 5 | Biological resource use | B | High | Pervasive (71-100%) | Serious (31-70%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 5.1 | Hunting & collecting terrestrial animals | B | High | Pervasive (71-100%) | Serious (31-70%) | Moderate (Possibly in the short term, < 10 yrs) | All herds: management of Wood Bison for social, disease, genetic management involves setting targets, removing “surplus”, and excluding areas using hunting as management tool. Removing animals from highways, farming areas, roads, communities (NWT ,NW Alberta) |
| 5.3 | Logging & wood harvesting | - | - | - | - | - | may be beneficial |
| 6 | Human intrusions & disturbance | D | Low | Large (31-70%) | Slight (1-10%) | High (Continuing) | - |
| 6.1 | Recreational activities | D | Low | Large (31-70%) | Slight (1-10%) | High (Continuing) | Snowmobiling and all-terrain vehicles. All herds are subjected to snowmobiling in particular, although reactions to snowmobiling can be variable. High concern in Yukon in particular where all hunting is from snowmobiles. In NW Alberta it is more from industrial activity than recreation. Increasing concern (esp. in Yukon) that there are impacts on condition and eventually population-level as a result of stress. The severity is a concern but unknown. |
| 6.3 | Work & other activities | - | Negligible | Restricted (11-30%) | Negligible (<1%) | High (Continuing) | Working snowmobiles |
| 7 | Natural system modifications | - | Negligible | Restricted (11-30%) | Negligible (<1%) | High (Continuing) | - |
| 7.1 | Fire & fire suppression | - | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Fire is beneficial, suppression can be a threat |
| 7.2 | Dams & water management/use | - | Negligible | Small (1-10%) | Negligible (<1%) | High (Continuing) | - |
| 7.3 | Other ecosystem modifications | CD | Medium - Low | Restricted (11-30%) | Moderate - Slight(1-30%) | High (Continuing) | Ecosystem modifications as a result of Peace River damming affects Wood Buffalo bison herd |
| 8 | Invasive & other problematic species & genes | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | - |
| 8.1 | Invasive non-native/alien species | C | Medium | Large (31-70%) | Moderate (11-30%) | High (Continuing) | This deals mainly with TB and brucellosis. Extremely low genetic diversity may well limit the subspecies response to novel stressors and environmental change. This does not include roadside plantings of agronomic grasses and legumes that can attract bison to roads and increase vehicle strikes. While a possible threat, it is disease issues in this category are much more important. |
| 8.2 | Problematic native species | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | Mackenzie, WBNP, SRL. Anthrax (assumed to be native for this exercise). Anthrax outbreaks occur at 10-20 year intervals, may kill up to 1/3 of a population, and the experience in NWT in 2012 has demonstrated it could be even more. Wolves as natural predators are important here too; the impact can increase in severity for bison populations weakened by disease (TB and brucellosis). |
| 8.3 | Introduced genetic material | - | Negligible | Pervasive (71-100%) | Negligible (<1%) | High (Continuing) | Hybridization with Cattle and Plains Bison |
| 11 | Climate change & severe weather | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 11.1 | Habitat shifting & alteration | - | Unknown | Large (31-70%) | Unknown | Unknown | Loss of boreal grasslands with climate change, but impacts may be offset by increasing fire |
| 11.2 | Droughts | - | Negligible | Pervasive (71-100%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 11.3 | Temperature extremes | AC | Very High - Medium | Pervasive (71-100%) | Extreme - Moderate (11-100%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 11.4 | Storms & flooding | C | Medium | Large (31-70%) | Moderate (11-30%) | Moderate (Possibly in the short term, < 10 yrs) | Flooding events have impacted > 25% of MacKenzie animals |