
COSEWIC Assessment and Status Report on the Proud Globelet Patera pennsylvanica in Canada - 2015

Photo: Robert Forsyth © Environment Canada
Long description for Figure
Three photos providing different views of the fresh shells of recently dead Proud Globelets (Patera pennsylvanica). A general characteristic of this genus is the imperforate umbilicus. Unlike other species of Patera, Proud Globelet lacks a parietal tooth-like protuberance on the aperture wall, has a higher spire and has a last whorl that is more markedly descending at the aperture. Shells are thin, regularly striate, yellowish olive with 5.75 to 6 whorls in adults. The lip of the aperture is white and narrowly reflected with a low prominence on the inner rim of the baso-columellar margin.
- Document Information
- Assessment Summary
- Executive Summary
- Technical Summary
- Preface
- Wildlife Species Description and Significance
- Distribution
- Habitat
- Biology
- Population Sizes and Trends
- Threats and Limiting Factors
- Protection, Status and Ranks
- Acknowledgements and Authorities Contacted
- Information Sources
- Biographical Summary of Report Writers
- Collections Examined
- Figure 1. A) Best conserved shells of Proud Globelet (Patera pennsylvanica) found in the south part of the Black Oak Heritage Forest in the City of Windsor, Essex County, during the survey of summer 2013. B) Shell found in the soil with partial periostracum.
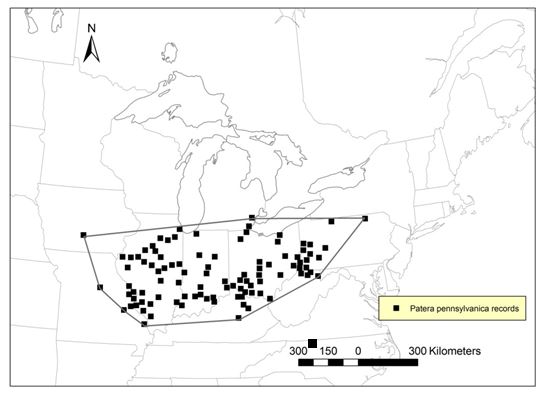
- Figure 2. Global range of Proud Globelet, Patera pennsylvanica, based on Hubricht (1985) and updated with georeferenced data from Canadian museums (Appendix 1) as well as from the Global Biodiversity Information Facility (GBIF 2013) collected from different museums worldwide (Appendix 3). Data include records without date and with dates from 1882 to 2013. The global range was estimated using the minimum convex polygon method (534,453 km2) by the report writers. All non-georeferenced data from GBIF, Canadian museums and Coppolino (2009) were verified to be within the range boundaries defined by the convex polygon.
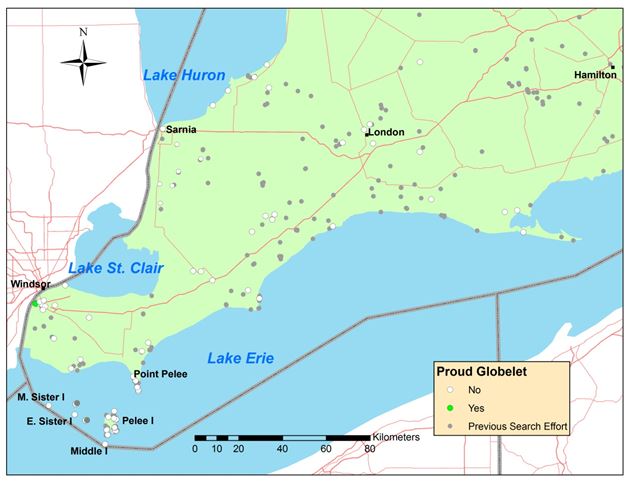
- Figure 3. Map of southwestern Ontario showing the search effort for Proud Globelet (Patera pennsylvanica). Grey dots: sites surveyed between 1992 and 2012 by M.J. Oldham and A. Nicolai for terrestrial gastropods and white dots: sites surveyed in 2013 specifically targeting P. pennsylvanica unsuccessfully (no Proud Globelets found). Green dot: successfully surveyed sites in 2013 targeting P. pennsylvanica.
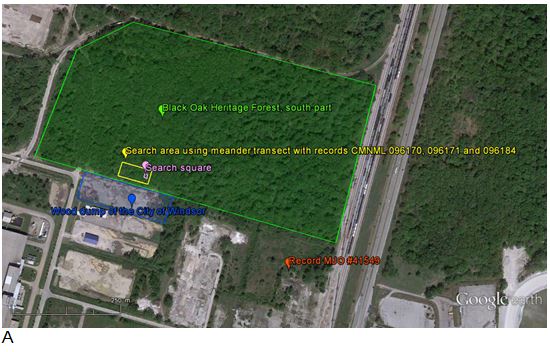
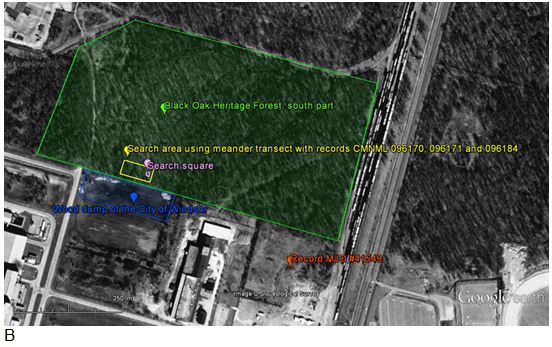
- Figure 4. Search area (yellow) and search square (pink) for live individuals in the south part of the Black Oak Heritage Forest in Windsor, Essex County (green), represented on aerial photographs from 2011 (A) and 1999 (B). The intensive search area corresponds to the zone where shells were found in 1992 (CMNML 096171) and 1996 (CMNML 096170) by M.J. Oldham, and on May 3, July 28 and August 27 (CMNML 096184), 2013 by A. Nicolai (see Appendix 1 and 2). The intensive search area is located close to the bordering wood dump (blue) managed by the City of Windsor. One shell was found on September 5, 2013 by M.J. Oldham (# 41549) south of Black Oak Heritage Forest on a former light industrial area (orange). All records are from habitats that have not substantially changed since 1999. Aerial imagery containing point locations has been removed. Contact the COSEWIC Secretariat for access.
- Figure 5. Wood dump operated by the City of Windsor, Essex County, bordering the south part of the Black Oak Heritage Forest. View of the south entrance of the park with the wood dump on the right side. (Photo credit: Annegret Nicolai.)
- Figure 6. Square (10 m x 10 m, flagging tape can be seen on two edges) for the intensive search for live individuals of the Proud Globelet (Patera pennsylvanica) in the zone of shell occurrence in the Black Oak Heritage Forest in the City of Windsor. (Photo credit: Annegret Nicolai.)
- Figure 7. The new bridge crossing the Detroit River and connecting Highway 75 in Detroit with Highway 401 in Windsor. The construction site in Windsor (blue with dashed line within the red rectangle) is located just north of the Black Oak Heritage Forest (red transparent square within the red rectangle) (modified from Samuel 2009).
- Figure 8. Dusky Arion (Arion subfuscus) foraging on a fungus in Black Oak Heritage Forest, Windsor, Essex County. (Photo credit: Annegret Nicolai.)
- Appendix 1. Records of Proud Globelet (Patera pennsylvanica) in Canadian museums and private collections. CMN – Canadian Museum of Nature. CMNML – CMN accession number. ROM – Royal Ontario Museum. ROMCN – ROM accession number. Geographic coordinates have been removed. Contact the COSEWIC Secretariat for access.
- Appendix 2. Terrestrial gastropods found during the search for Proud Globelet (Patera pennsylvanica) in and near the Black Oak Heritage Forest, Windsor, Essex County, in 2013. Observers were Annegret Nicolai (AN), Michael Oldham (MJO), Jane Bowles (JMB) and Mykola Merkulov (MM).* denotes introduced exotic species.
- Appendix 3. Museum records of Proud Globelet, Patera pennsylvanica, in the Global Biodiversity Information Facility (GBIF) database. FLMNH - Florida Museum of Natural History, MCZ - Museum of Comparative Zoology at Harvard University, LI - Biologiezentrum Linz Oberösterreich, NMNH - National Museum of Natural History, Smithsonian Institution, ANSP - Academy of Natural Sciences Philadelphia, NMR - Natural History Museum Rotterdam, FMNH - Field Museum of Natural History, NTSRV - NatureServe Central Databases, MACN - Museo Argentino de Ciencias Naturales. Geographic coordinates have been removed. Contact the COSEWIC Secretariat for access.
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Cananda
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2015. COSEWIC assessment and status report on the Proud Globelet Patera pennsylvanica in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xi + 41 pp.
COSEWIC would like to acknowledge Annegret Nicolai, the University of Western Ontario, and Michael J. Oldham for writing the status report on the Proud Globelet, Patera pennsylvanica, in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Dwayne Lepitzki, Co-chair of the COSEWIC Molluscs Specialist Subcommittee.
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
E-mail: COSEWIC E-mail
Website: COSEWIC
Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur la Patère de Pennsylvanie (Patera pennsylvanica) au Canada.
Proud Globelet -- Robert Forsyth (Black Oak Heritage Forest, April 19 1996, collector: Michael J. Oldham, CMNML 096170).
Proud Globelet, Patera pennsylvanica is a terrestrial snail in the family Polygyridae. The yellowish, round shell (15-20 mm diameter) lacks a tooth-like protuberance at the shell opening compared to other species of the genus. The sole known Canadian population occurred in and near the Black Oak Heritage Forest owned by the City of Windsor. Although the ecological significance of Proud Globelet is unknown, gastropods, in general, play important roles in forest ecosystem functioning via nutrient cycling and soil building processes.
Proud Globelet is found from southwestern Ontario south to Iowa and Missouri and east to Pennsylvania. The species’ entire range, nearly all of which is in the U.S., is about 534,453 km2. Canada has less than 0.001% of the global range. Empty, fresh shells were found in 1992 and 1996 in Windsor. Empty, weathered shells were found in 2013 in the same place and nearby. No live individual was ever recorded in Canada. This species was not found elsewhere in southwestern Ontario in gastropod surveys from 1916 to 2013.
Proud Globelet generally occurs on wooded hillsides or in ravines. In Canada, the species has been reported in a sandy oak forest and a disturbed light industrial site. Food requirements for Proud Globelet might be fungi, leaf litter and fresh plant material, but some Polygyridae are carnivorous. In Ontario, trends observed in the habitat of Proud Globelet include a general reduction in oak forests and reduction in biodiversity, the latter potentially affecting the snail communities.
Very little is known about the biology of Proud Globelet. From other species in the polygyrid family, it has been inferred that mating occurs in fall or early spring and oviposition in spring to late summer. Clutch size ranges between 20 and 80 eggs that hatch about 20 to 60 days after oviposition. Growth rate and, consequently, adult size (reached after 1-2 years) are highly variable. Growth periods correspond to activity periods from spring to fall. Sexual maturity is reached after 2 to 3 years, and lifespan has been estimated to range between 3 and 5 years. Snails are ectotherms and prone to freezing in winter or dehydration in summer. Different strategies have evolved to enable the species to survive extreme temperatures and drought besides going into dormancy. Hibernation in the Polygyridae extends from early October until mid-April in temperate regions. Aestivation occurs occasionally during prolonged heat and drought periods. Snails rely on humidity-buffered refugia and snow cover for dormancy survival. Most Polygyridae are active at dusk or during the night with dispersal for colonization of new habitat being slow, around 35 m in 3 years. Predation and parasites can be a source of mortality for land snails.
Seventeen person-hours of search effort in different light and weather conditions were spent trying to find live individuals in a 200 m x 100 m square plot in the Black Oak Heritage Forest and in a nearby former light industrial area in 2013. The plot encompasses the area where the snail had been previously found. Fourteen empty shells (estimated to be 5-15 years old) were found under leaf litter in the upper 5 cm of the soil in addition to other snail and slug species in the forest plot; Proud Globelet was not found elsewhere in the forest but one shell also was found south of the forest in the nearby former light industrial area. The complete absence of live individuals and the age of the shells found in 2013 suggest that the population has substantially declined since 1996; there is a strong likelihood that Proud Globelet has disappeared from this area and from Canada as it has only ever been found in this area in southwestern Ontario. Rescue from the U.S. population is unlikely because the Detroit and St. Clair rivers and Lake Erie are dispersal barriers.
Human impacts such as pollution, garbage accumulation, intensive recreational use and changes to soil composition and hydrology can affect the snail population. Introduced species, such as plants, earthworms and other gastropods can affect native snail populations through alteration of the soil nutrient cycle, reduction of leaf litter and inter-specific competition. The introduced slug Dusky Arion, Arion subfuscus, was abundant at the site where Proud Globelet shells were found and was observed feeding on fungi. Climate change can have a large impact on snail survival. In temperate regions, climate change will involve increases in both average temperature and the frequency of extreme weather events such as heat waves, drought, and high precipitation, as well as an absence of insulating snow cover. Snails are limited by their low dispersal or escape capacity, relatively long generation time, low physiological resistance to fluctuating environmental factors, susceptibility to bioaccumulation of toxic agents, and limited genetic flow.
Global Rank: G4 (Apparently Secure), National Rank (Canada): N1 (Critically Imperilled), National Rank (US): N4 (Apparently Secure). Sub-national ranks are “critically imperilled” (S1) in Ontario and West Virginia, “critically imperilled” to “imperilled” (S1S2) in Pennsylvania. Michigan listed Proud Globelet as a species of special concern.
Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines (2011) is being used).
- 3-5 years (in Polygyridae)
Is there an [observed, inferred, or projected] continuing decline in number of mature individuals?
- N/A (no live individuals)
Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations] (52-60 years)
- N/A (no live individuals)
[Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations] (78-90 years)
Unknown but could be close to 100%
Fresh dead shells observed in 1992 and 1996 but only old weathered shells (5-15 years old) observed in 2013.
[Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations] (78-90 years).
- N/A (no live individuals)
[Observed, estimated, inferred or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future.
Unknown but could be close to 100%
Fresh dead shells observed in 1992 and 1996 but only old weathered shells (5-15 years old) observed in 2013.
Are the causes of the decline clearly reversible and understood and ceased?
- No
- No
- No
Are there extreme fluctuations in number of mature individuals?
- N/A (no live individuals)
Estimated extent of occurrence
- 4 km²
Index of area of occupancy (IAO, 2 x 2 km2grid values)
- 4 km²
Is the population severely fragmented?
- No
- No
Number of locations
- 1-2
Is there an observed continuing decline in extent of occurrence?
- N/A (no live individuals)
Is there an observed continuing decline in index of area of occupancy? (Current Area of Occupancy is 65% of historical; see Figure 4)
- N/A (no live individuals)
Is there an [observed, inferred, or projected] continuing decline in number of populations?
- N/A (no live individuals)
Is there an [observed, inferred, or projected] continuing decline in number of locations?Footnote1
- N/A (no live individuals)
Is there an [observed, inferred, or projected] continuing decline in quality of habitat?
- Yes, inferred, quality
Are there extreme fluctuations in number of populations?
- N/A (no live individuals)
Are there extreme fluctuations in number of locations?Footnote1
- N/A (no live individuals)
Are there extreme fluctuations in extent of occurrence?
- N/A (no live individuals)
Are there extreme fluctuations in index of area of occupancy?
- N/A (no live individuals)
Black Oak Heritage Forest, City of Windsor
0 Mature Individuals Alive
(15 shells of adults and juveniles)
Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years].
- N/A (no live individuals)
Was a threats calculator completed for this species: No (no live individuals)
- Human intrusions and disturbances
- Natural system modifications
- Pollution
- Climate change and extreme weather
What additional limiting factors are relevant?
Low dispersal or escape capacity, relatively long generation time, low physiological resistance to fluctuating environmental factors, susceptibility to bioaccumulation of toxic agents, and limited genetic flow.
Status of outside population(s) most likely to provide immigrants to Canada.
- Michigan: SNR (by NatureServe but Special Concern in Michigan publication)
- Ohio: SNR
- Pennsylvania: S1S2
- New York: not present
Is immigration known or possible?
- Yes
Would immigrants be adapted to survive in the SLE?
- Unknown
Is there sufficient habitat for immigrants in the SLE?
- Unknown
Are conditions deteriorating in Canada?Footnote2
- Yes
Are conditions for the source population deteriorating?Footnote2
- Unknown
Is the Canadian population considered to be a sink?Footnote2
- N/A
Is rescue from outside populations likely?
- No
Is this a data-sensitive species?
- No
COSEWIC: Designated Endangered in May 2015.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Kingdom: Animalia
Phylum: Mollusca
Class: Gastropoda
Order: Pulmonata
Suborder: Stylommatophora
Family: Polygyridae
Genus: Patera
Species: Patera pennsylvanica(Green 1827) – Proud Globelet
Proud Globelet has been recognized as a species since 1827. Although placed in various genera, including Mesodon andPolygyra (see Pilsbry 1940), the species is currently assigned to the genus Patera(Emberton 1991; Turgeon et al.1998). Emberton’s systematic revision assigned Proud Globelet to the subgenus Ragdaleorbis, and it is the sole species in this subgenus within Patera.
There are 13 species in the genus Patera, two of which inhabit the eastern United States and Canada (Grimm et al. 2010). Besides Proud Globelet, one apparently introduced population of Flat Bladetooth, Patera appressa, has been recorded in Ontario (Grimm et al. 2010).
A general characteristic of this genus is the imperforate umbilicus (hole at the central part of the underside of the shell) (Pilsbry 1940). Morphologically, Proud Globelet is unlike any other species of Patera, which accounts for its placement in the subgenus Ragsdaleorbis. Proud Globelet (Figure 1) lacks a parietal tooth-like protuberance on the aperture wall, has a higher spire (conical point at top of the shell) and the last whorl is more markedly descending at the aperture (Grimm et al. 2010). Shells are thin, regularly striate, yellowish olive with 5¾ to 6 whorls in adults. The lip of the aperture is white and narrowly reflected with a low prominence on the inner rim of the baso-columellar margin. Adult shells measure between 10 and 15 mm high and between 15 and 20 mm in diameter.

Photo: Annegret Nicolai © Environment Canada, 2015
Long description for Figure 1
Two photos of Proud Globelet shells found in the south part of the Black Oak Heritage Forest in the City of Windsor, Essex County, Ontario, in the summer of 2013. Photo A shows 11 of the best conserved shells. Photo B shows a shell with a partial periostracum.
Canada’s only known population of Proud Globelet is/was located in and near the Black Oak Heritage Forest, City of Windsor, Essex County, Ontario (Table 1; Appendix 1 and 2; Grimm 1996; NatureServe 2013). No genetic or population structure data are available. No other populations are known to exist in Canada and dispersal between potential sites has been greatly reduced with anthropogenic changes to the landscape. Less than 5% of the Essex Region is forested (ERCA 2002). Most patches of remaining forest are less than 10 ha. Habitat is highly fragmented with many possible barriers to forest snail dispersal, such as roads, agricultural fields and urban areas. When migration is limited between low-density populations there might be a high degree of genetic structuring due to reduced gene flow as observed in other Polygyridae (Anderson 2007).
The sole Canadian population is/was within the COSEWIC Great Lakes Plains National Ecological Area (COSEWIC 2011). Genetic data and evidence of local adaptations are unavailable. Therefore, the Canadian occurrence is a single designatable unit.
| Site | Site Name | Effort (person-hours) | Observers | Date(s) | Patera pennsylvanica? | Weather |
|---|---|---|---|---|---|---|
| 1 | Black Oak Heritage Forest, south part, Windsor | 14 | AN, JMB, MJO | May 3, July 28, Aug 27-28, Sep 5 | Shells | 20°C, sunny 20-23°C, humid or rainy |
| 2 | Former light industrial area south of Black Oak Heritage Forest, Windsor | 3 | MJO | Sep 5 | Shell | 22°C, sunny |
| 3 | Black Oak Heritage Forest, north part, Windsor | 4 | AN, MJO, | Apr 29 | No | 18°C, overcast |
| 4 | Devonwood Conservation Area, Windsor | 2 | AN, MJO | April 29 | No | 18°C, overcast |
| 5 | Springgarden Road Park, Windsor | 2 | AN, MJO | April 29 | No | 18°C, overcast |
| 6 | Ojibway Park, Windsor | 5 | AN, MJO, JMB | Apr 29, May 3 | No | 18°C, overcast; 20°C, sunny |
| 7 | Malden Park, Windsor | 2 | AN, JMB | May 3 | No | 20°C, sunny |
| 8 | Oakwood, Windsor | 2 | AN, MM | Aug 27 | No | 22°C, sunny, humid |
| 9 | Brunet Park, La Salle | 1 | AN | Aug 28 | No | 22°C, sunny, humid |
| 10 | South Cameron Woodlot, Windsor | 1 | AN, MM | Aug 28 | No | 22°C, sunny, humid |
| 11 | Peche Island, Windsor | 2 | AN, HU | May 19 | No | 22°C, sunny |
| 12 | Middle Island, Point Pelee National Park, Lake Erie | 6; 12 | RFF, AN, MJO; AN, TD, MM | May 1; Aug 29 | No | 18°C, sunny; 23°C, sunny, humid |
| 13 | East Sister Island Provincial Park, Lake Erie | 4.5 | TD, RFF, AGH, AN, MJO | Apr 30 | No | 18°C, sunny |
| 14 | Middle Sister Island, Lake Erie | 3.5 | TD, RFF, AGH, AN, MJO | Apr 30 | No | 18°C, sunny |
| 15 | Lighthouse Point Provincial Nature Reserve, Pelee Island | 1.5; 2 | RFF, AN, MJO; AN, MM | May 1; Aug 25 | No | 20°C, sunny, dry, windy; 25°C, sunny, humid |
| 16 | Erie Sand and Gravel NCC parcel, Pelee Island | 3.5 | AN, MJO, AGH | May 2 | No | 24°C, sunny, dry |
| 17 | Middle Point Woods – north part, NCC, Pelee Island | 2; 1 | AGH, RFF, MJO, AN; AN | May 2; Aug 25 | No | 24°C, sunny, dry; 24°C, sunny, humid |
| 18 | Gibwood Property, NCC, Pelee Island | 2 | AN, MJO | May 2 | No | 22°C, sunny, dry |
| 19 | Richard and Beryl Ivey Nature Reserve, NCC, Pelee Island | 1.5 | RFF, AGH, AN | May 1 | No | 24°C, sunny |
| 20 | Winery property, Pelee Island | 4 | RFF, AGH, AN, MJO | May 2 | No | 24°C, sunny |
| 21 | Porchuk Property, NCC, Pelee Island | 2 | AN, MJO | May 2 | No | 22°C, sunny |
| 22 | Fish Point Provincial Nature Reserve, Pelee Island | 5 | RFF, AGH, AN | May 1 | No | 18°C, sunny |
| 23 | Fleck Property, Pelee Island | 1 | RFF | May 2 | No | 22°C, sunny |
| 24 | Essex Conservation Authority lands at Stone Road Alvar, Pelee Island | 1 | AGH | May 2 | No | 22°C, sunny |
| 25 | Ontario Nature Stone Road Alvar, Pelee Island | 1; 2 | AGH; AN, MM | May 2; Aug 27 | No | 22°C, sunny; 25°C, sunny, |
| 26 | Cohen Shaughnessy Property, NCC, Pelee Island | 1; 2 | AGH; AN,MM | May 2; Aug 27 | No | 23°C, sunny; 25°C, sunny, humid |
| 27 | Krestel Parcel, NCC, Pelee Island | 1 | AGH | May 1 | No | 20°C, sunny |
| 28 | Middle Point Woods – south part, NCC, Pelee Island | 1.5; 1 | RFF, AGH, AN; AN, MM | May 1, 2; Aug 26 | No | 22°C, sunny; 25°C, sunny, humid |
| 29 | Florian Diamante Nature Reserve, NCC, Pelee Island | 4.5 | AGH, RFF, AN | May 2 | No | 22°C, sunny |
| 30 | Point Pelee National Park, Tip | 3 | AGH, AN, MJO | Apr 28 | No | 15°C, rain, stopped later |
| 31 | Point Pelee National Park, tip to Visitor Centre | 2 | AGH, AN, MJO | Apr 28 | No | 15°C, rain, stopped later |
| 32 | Point Pelee National Park, West Beach Trail | 3 | AGH, AN, MJO | Apr 28 | No | 15°C, rain, stopped later |
| 33 | Point Pelee National Park, Dunes Picnic area | 1 | RFF | April 29 | No | 18°C, overcast |
| 34 | Point Pelee Nataional Park, east of Dunes Picnic area | 1 | AGH | April 29 | No | 18°C, overcast |
| 35 | Point Pelee National Park, Sanctuary Picnic area | 1.5 | AGH, MJO, AN | Apr 28 | No | 17°C, overcast |
| 36 | Oxley Swamp, NCC | 2 | AN, HU | May 20 | No | 26°C, sunny, humid |
| 37 | Cedar Creek CA | 3 | RFF, AGH | April 29 | No | 12°C, overcast |
| 38 | Kopegaron Woods CA | 4 | RFF, AGH, AN, MJO | Apr 29, 30 | No | 14°C, overcast |
| 39 | Two Creeks CA | 2 | MJO | May 18 | No | - |
| 40 | Canard River CA | 2 | AN, MJO | April 29 | No | 18°C, overcast |
| 41 | Maidstone CA | 2 | RFF, AGH | April 29 | No | 14°C, overcast |
| 42 | Rondeau Provincial Park | 3; 3.5 | MJO, JMB; AGH | May 17; Sep 4 | No | 22°C, sunny |
| 43 | Sinclair’s Bush | 2 | MJO, JMB | May 17 | No | - |
| 44 | Thames Grove CA | 1 | AN, JMB | May 3 | No | 20°C, sunny |
| 45 | Moraviantown First Nation | 6 | AN, JMB | June 7 | No | 15°C, overcast, humid |
| 46 | John E. Pearce Provincial Park | 2 | MJO | May 15 | No | - |
| 47 | Newport Forest, TTLT | 1; 2 | AN; AN, HU | April 21; Sep 01 | No | 18°C, sunny, humid; 23°C after rain |
| 48 | Wardsville Woods TTLT | 1 | JMB | May 17 | No | - |
| 49 | Backus Woods, NCC | 1; 3 | MJO; AGH | May 15; Sep 2 | No | 18°C, sunny |
| 50 | St. Williams Conservation Reserve | 2 | MJO | May 15 | No | - |
| 51 | Calton Swamp | 1 | MJO | May 15 | No | - |
| 52 | Lake Whittaker CA | 2 | AN,HU | June 8 | No | 18°C, overcast |
| 53 | Westminster Ponds, London | 1 | AN | April 7 | No | 15°C, overcast |
| 54 | Komoka Provincial Park | 1 | AN, HU | Jan 13 | No | 12°C, humid, no snow |
| 55 | Western University, London | 0.5 | AN | April 15 | No | 18°C, sunny |
| 56 | Canatara Park, Sarnia | 3 | JMB, MJO; AGH; AN, LC | May 16, August 3; Sep 22 | No | 22°C, sunny (Aug 3) |
| 57 | Killaly Meadows, London | 1 | AN | May 4 | No | 20°C, sunny |
| 58 | Lambton United Church Camp | 2 | AGH | August 3 | No | 25°C, sunny |
| 59 | Highland Glen CA | 1 | AGH | August 3 | No | 25°C, sunny |
| 60 | Joany's Woods TTLT | 1 | AN, JMB | April 1 | No | 12°C, overcast |
| 61 | Port Franks | 2 | AGH | August 4 | No | 24°C, sunny |
| 62 | Pinery Provincial Park | 1;1 | AN | May 5; July 07 | No | 20°C, sunny; 25°C, sunny |
| 63 | C.M. Wilson CA | 2 | MJO, JMB | May 16 | No | - |
| 64 | Paxton Wood, Chatham | 2 | MJO, JMB | May 16 | No | - |
| 65 | Skunk’s Misery | 2 | MJO, JMB | May 16 | No | - |
| 66 | Avon trail near St. Mary’s | 1 | AN | Jul 27 | No | 20°C, rain |
| 67 | Long Point Provincial Park | 2 | AGH | Sep 2 | No | 24°C, sunny |
| 68 | Bickford Oak CA | 2 | AN, LC | Sep 22 | No | 18°C, cloudy, wet |
| 69 | Brigden Crown Game Reserve | 2 | AN, LC | Sep 22 | No | 18°C, cloudy, wet |
| 70 | Moore Wildlife Refuge CA | 2 | AN, LC | Sep 22 | No | 18°C, cloudy, wet |
| 71 | Perch Creek CA | 2 | AN, LC | Sep 21 | No | 20°C, cloudy, wet |
| 72 | Floodway CA | 2 | AN, LC | Sep 21 | No | 20°C, cloudy, wet |
| 73 | Petrolia CA | 1 | AN, LC | Sep 22 | No | 18°C, cloudy, wet |
| 74 | Rouge Park, Scarborough | 4 | AN | Sep 14, 15 | No | 22°C, sunny, humid |
| 75 | High Park, Grenadier Pond, Toronto | 1 | MM | Sep 22 | No | 18°C, cloudy, wet |
The only known Canadian population of Proud Globelet occurs/occurred in the Carolinian Forest Region near the northern limit of the species’ global range (Figure 2). This population may be genetically isolated from other populations in the United States and therefore has significance for conservation.
Figure 2. Global range of Proud Globelet, Patera pennsylvanica, based on Hubricht (1985) and updated with georeferenced data from Canadian museums (Appendix 1) as well as from the Global Biodiversity Information Facility (GBIF 2013) collected from different museums worldwide (Appendix 3). Data include records without date and with dates from 1882 to 2013. The global range was estimated using the minimum convex polygon method (534,453 km2) by the report writers. All non-georeferenced data from GBIF, Canadian museums and Coppolino (2009) were verified to be within the range boundaries defined by the convex polygon.
© Environment Canada, 2015
Long description for Figure 2
Map outlining the global range of the Proud Globelet, where it occurs or has occurred from southwestern Ontario south to Iowa and Missouri and east to Pennsylvania. The global range covers 534,453 square kilometres based on a minimum convex polygon.
The ecological significance of Proud Globelet is unknown. However, snails and slugs generally play important roles in forest ecosystem functioning, specifically by (i) aiding in decomposition, nutrient cycling and soil building processes (Mason 1970a,b; Jennings and Barkham 1979); (ii) providing food and essential nutrients to wildlife (South 1980; Churchfield 1984; Frest and Johannes 1995; Martin 2000; Nyffeler and Symondson 2001); and (iii) serving as hosts for parasitic worms (e.g., Rowley et al. 1987).
Proud Globelet is unknown to most Canadians. The Ojibway Nature Centre at Windsor published a species at risk list for the Ojibway Prairie Complex in Windsor and included the Proud Globelet (Pratt 2012).
It has no commercial value and is not an agricultural or garden pest.
Aboriginal traditional knowledge was not available for this species.
Proud Globelet is/was distributed from southwestern Ontario south to Iowa and Missouri and east to Pennsylvania (Figure 2).
The global range covers 534,453 km2 as measured by minimum convex polygon by the report writers (Figure 2).
In Canada, the Proud Globelet is/was restricted to the Carolinian Forest region of Ontario, specifically to an area of 200 m x 100 m in the south part of the Black Oak Heritage Forest and to a less than approximately 100 m2 site in a former light industrial area south of the Black Oak Heritage Forest in the City of Windsor (Figures 3 and 4, Grimm 1996). Empty, fresh shells of this species were first found in the Black Oak Heritage Forest in 1992 by Michael J. Oldham. Empty, fresh shells were found again in the same place in 1996 by M.J. Oldham, while only old, weathered shells were found in 2013 by A. Nicolai in the same place and by M.J. Oldham south of the Black Oak Heritage Forest on a former light industrial area (Table 1, Appendix 1 and 2). Fresh empty shells indicate recently dead individuals in 1992 and 1996 suggesting an extant population at that time. No living individuals of Proud Globelet have ever been documented in Canada.
Canada contains less than 0.001% of the global range of the species.
The index of area of occupancy is 4 km2 (i.e., species occurs/occurred in one 2 km x 2 km square).
The extent of occurrence of Proud Globelet in Canada is 4 km2 (extent of occurrence equals index of area of occupancy).
Forsyth’s (2013) references to Canadian occurrences of Proud Globelet were searched. Notable surveys include those conducted on the Lake Erie islands by Clapp (1916) and Goodrich (1916) and the efforts of John Oughton between about 1930 and 1940 (Oughton 1948). Grimm (1996) collected extensively in southern and eastern Ontario between 1970 and the mid-1990s, but his collection is only partially available. Collections at the Royal Ontario Museum, Canadian Museum of Nature and Bishops Mills Natural History Centre have been searched. None of these early general surveys was in the Black Oak Heritage Forest or Windsor area. Other worldwide collections were accessed via the Global Biodiversity Information Facility (GBIF 2013, Appendix 3).
Surveys between 1992 and 2012 also were general land snail searches by M.J. Oldham and A. Nicolai rather than targeted searches for Proud Globelet (Figure 3). From 1992-1996, 92 samples of about 25 species were recorded by M.J. Oldham, each sample containing several specimens of one species. He found fresh shells of recently dead Proud Globelet twice in the Black Oak Heritage Forest: in 1992 and 1996. In 2012 about 360 specimens of about 45 species were collected by A. Nicolai and sent to Robert Forsyth for curation but no Proud Globelet was found.
In 2013, 74 sites in southwestern Ontario were surveyed with a total effort of 233 person-hours to provide information for the preparation of status reports for Proud Globelet (Figure 3, Table 1) and Allogona profunda, Broad-banded Forestsnail (COSEWIC in press). Sites surveyed were mainly small remnants of forest in parks and conservation areas. Living snails and empty shells were searched for visually using a meandering transect on the surface of the ground and under logs, rocks and similar debris. Because Proud Globelet is a relatively large species, it is less likely to have been overlooked than smaller snail species during these surveys; it was found only in the Black Oak Heritage Forest and south of it on a former light industrial area. The snail searches in 2013 resulted in approximatively 210 stored-in-alcohol samples of about 60 species being deposited at the Biodiversity Institute of Ontario (BIO) at Guelph and included in the Barcode of Life Database (BOLD); 200 shell samples of about 40 species also were collected and are currently being curated by Robert Forsyth.
Figure 3. Map of southwestern Ontario showing the search effort for Proud Globelet (Patera pennsylvanica). Grey dots: sites surveyed between 1992 and 2012 by M.J. Oldham and A. Nicolai for terrestrial gastropods and white dots: sites surveyed in 2013 specifically targeting P. pennsylvanicaunsuccessfully (no Proud Globelets found). Green dot: successfully surveyed sites in 2013 targeting P. pennsylvanica.
© Environment Canada, 2015
Long description for Figure 3
Map of southwestern Ontario showing the sites of surveys targeting Proud Globelet carried out in 2013. Also shown are the sites of surveys by M.J. Oldham and A. Nicolai for terrestrial gastropods between 1992 and 2012.
In the U.S., Proud Globelet generally occurs on wooded hillsides or in ravines, under leaf litter and stones, but also on grassy roadsides (Hubricht 1985). The habitat of the Canadian population is sandy oak forest and a disturbed former light industrial site with building rubble.
The minimal viable habitat patch size for a Proud Globelet population is unknown. In Canada, only a single population of Proud Globelet is known and it is/was confined to a small area in a landscape where natural habitat is highly fragmented. It is unclear what factors caused the apparent disappearance of the population.
Food requirements for Proud Globelet are unknown. Fungi associated with decomposing logs on the forest floor are apparently an important source of food in other Polygyridae (Blinn 1963; Asami 1993). Occasionally some species of Polygyridae appear to be carnivorous (Simpson 1901; Crabb 1928). To counteract cholesterol shortages that occur during reproduction and aestivation, snails can also feed on carrion (Nicolai et al. 2011, 2012). Patera appressa requires fresh plant material in addition to leaf litter to increase growth rate and fitness (Martin and Bergey 2013). According to van Cleave and Foster (1937) as well as Steensma et al. (2009), stinging nettles (Urtica spp.) seem to be an important nutrient source for rapid early juvenile growth and higher fitness in other polygyrids.
Although habitat requirements seem to be similar within the Polygyridae, differences in for example, foraging habits, daily activity patterns, thermal physiology adaptation/plasticity, among species of the same genus can result in different habitat preferences (Walsh and Coles 2006).
Pre-European settlement land cover in southwestern Ontario was dominated by deciduous forest (72%) of which less than 10% were oak forests (Butt et al. 2005). Forest cover is now reduced to 16% in southwestern Ontario, and many oak forests have disappeared or are reduced in size (Butt et al. 2005). Only about 5% of the original forest cover remains in Essex County (ERCA 2002). Many forest remnants might be smaller than the minimal viable habitat patch size for some gastropods. The reduction and fragmentation of habitat could also have led to a decrease in population size accompanied by a loss of genetic diversity affecting the viability of populations.
The biodiversity of oak forests is unique, and Proud Globelet appear to be sensitive to the anthropogenic disturbances that drive biodiversity changes. In the study by Douglas et al. (2013) in Kentucky, the absence of natural Patera appressapopulations was a good indicator of anthropogenic disturbance through its affinity for old growth, undisturbed forests. In the area of the Black Oak Heritage Forest in Windsor where the shells of Proud Globelet were found, old, weathered shells of the native Whitelip (Neohelix albolabris) were also abundant (Appendix 2). However, no live individuals of this species were recorded. The shells were about the same age as the Proud Globelet, and both species might have disappeared at the same time for the same unknown reason. Changes in vegetation affect snails as they use it as habitat, food and winter refugia. The presence of invasive species and the decline of native species change the ecological functioning of the habitat. The absence of live Proud Globelet and Whitelip might indicate a high degree of anthropogenic land use disturbance.
The Black Oak Heritage Forest is a forest remnant with several oak species (Pratt 2012). At the end of the 20th century, recreational use within this forest increased, as evidenced by the appearance of a trail system and the accumulation of garbage (Pratt pers. comm. 2013). At its periphery, urbanization and industrialization have fragmented the habitat (e.g., the railway) and changed environmental factors through soil and air pollution and changes in soil composition, hydrology and light conditions. For example, about 20 years ago a wood dump site (Figures 4 and 5) was installed on the south border of the forest by the City of Windsor (Pratt pers. comm. 2013), which changed habitat conditions and community composition through the introduction of new species. Even though the installation of the wood dump coincides with the first record of Proud Globelet, it is unlikely that snails migrated from the wood dump to the park. The trees in the wood dump came from city-owned land, including parks, road verges and woods, and were the result of urban forestry practices such as thinning, pruning, and removal of hazardous trees. Many of the trees transported to the wood dump are still well preserved, and do not host any woodland gastropod species.
Climate change is also expected to have a large impact on forest ecosystems. The following responses are possible if climatic conditions change beyond the tolerances of species: shifts in timing of life-cycle events; shifts in range boundaries; changes in morphology, reproduction, or genetics; or extinction (Rosenzweig et al. 2007). The consequence of climate change for snail habitat is a general disturbance of ecosystem processes.
Almost no information is available about the biology of Proud Globelet. General aspects of terrestrial snail biology are provided by the review of Barker (2001). Some information on the biology of other Polygyridae should only be considered as likely to be similar in Proud Globelet.
Proud Globelet is an air-breathing (pulmonate), simultaneous hermaphrodite, egg-laying snail (Pilsbry 1940). In general, both members of a mating pair exchange sperm and produce eggs. Mating processes are very species-specific in snails, sperm being exchanged externally in the polygyrid Mesodon genus (Webb 1954). In most snail species, larger individuals lay more eggs than smaller ones (Heller 2001). Mating in Polygyridae occurs in fall or early spring and oviposition in spring to late summer (van Cleave and Foster 1937; Blinn 1963; Steensma et al. 2009). Egg clutches are deposited in shallow holes excavated in moist soil. Clutch size in Polygyridae ranges between 20 and 80 eggs that hatch about 20 to 60 days after oviposition, depending on temperature and moisture.
Growth occurs only during periods of activity (spring to fall), and snails reach their adult shell size after 1 to 2 years. Growth rate is extremely variable within a population, resulting in highly divergent adult sizes. The average annual growth rate ranges from 0.6 to 5.2 mm/month (as measured by shell breadth). Growth rate is highest in summer and fall, but depends also on population density. Sexual maturity is reached after 2 to 3 years and lifespan has been estimated to range between 3 and 5 years (based on studies by Stiven and Foster 1996 and Steensma et al. 2009 of related species of Polygyridae), which is the estimated generation time for Proud Globelet.
Hibernation in Polygyridae extends from early October until mid-April in the temperate region (Blinn 1963). Typical hibernation refugia are shallow depressions in the forest floor covered with leaf litter or at soil depths of 5 to 10 cm (Pearce and Örstan 2006). During hibernation the shell aperture is oriented upwards and sealed with a calcareous epiphragm in other Polygyridae (Blinn 1963); Nicolai (pers. obs.) also photographed an A. profunda in Canada with an epiphragm. Aestivation occurs occasionally during periods of prolonged heat and drought. During aestivation, snails usually stay inactive in moist microhabitats, such as in soil, under leaf litter, and under logs.
Polygyridae can be active both day and night, but often retire to shelter under leaf litter from mid-morning until late afternoon (Blinn 1963). However, most of them are crepuscular or nocturnal, and sympatric species often have different activity patterns (Asami 1993).
Physiological responses to environmental factors and their plasticity and adaptability have not been investigated in Proud Globelet, nor in Polygyridae. In general, snails require calcium for shell formation. Soil and bedrock calcium availability influence the snail species richness of an area (Nekola 2005) and physiological processes, such as heat resistance in eggs (Nicolai et al. 2013). Heavy metals and pesticides in the soil are accumulated in tissues and may disturb physiological processes (Barker 2001).
Snails in regions with prolonged periods of drought and heat generally aestivate in buffered refuges and seal their shell aperture to avoid evaporation (Barker 2001; Pearce and Örstan 2006). In temperate regions, many species aestivate only in extreme summer conditions for a short period and have developed biochemical stress reactions that protect cells and maintain survival mechanisms, such as membrane fluidity, osmoregulation and enzyme activity (Nicolai et al. 2011). Unusually long heat and drought periods increase mortality.
Snails are ectotherms and prone to freezing in winter. Different strategies that are more or less plastic have evolved to enable survival at sub-zero temperatures and were reviewed in snails by Ansart and Vernon (2003). Because snails hibernate in the soil or the leaf litter, they rely on snow cover for optimal temperature buffering in temperate regions (Nicolai et al. 2011). Mortality during hibernation is around 40% in some populations and drives population dynamics (Peake 1978; Cain 1983).
Many terrestrial gastropods can be reared in captivity with relative ease (see Ansart et al.2014 for a broad study involving short-term rearing of over 30 different species). The long-term success of rearing depends on the knowledge of species’ specific requirements and has not been tested yet in Proud Globelet.
Active dispersal distances are unknown for Proud Globelet, but Polygyridae of similar size move between 120 and 220 cm per day within a home range of 80 to 800 m2, measured with the spooling technique (Pearce 1990). In contrast, mark-recapture methods used for short-term observations underestimate the capacity for movement in snails because many species are homing, e.g., Blinn (1963) found marked snails at distances of 1 to 4 m from hibernation sites. Edworthy et al.(2012) observed a maximum dispersal of 32.2 m over a 3-year study in the polygyrid Allogona townsendiana (Oregon Forestsnail).
The fragmented state of oak forests in the Windsor area due to urban and industrial development as well as roads makes the potential for dispersal among suitable habitat patches unlikely should additional subpopulations exist. Eggs and immature stages are not known to be dispersed by wind, water, or other vectors.
Some terrestrial gastropods can be easily transported by human activity (e.g., cars, horticultural or agricultural products) and therefore be introduced to new habitats (Robinson 1999; Robinson and Slapcinsky 2005). There is no evidence that Proud Globelet is synanthropic or has been transported by humans.
Polygyridae have been noted to be one of the intermediate hosts of the meningeal worm Parelaphostrongylus tenuis (Rowley et al. 1987). Parasitic mites are also common in snails, in general. The infection rate within a population ranges between 45-75%. Depending on the mite species, infections can cause high mortality, reproduction perturbations, and reduced cold hardiness (Baur and Baur 2005). Parasites could therefore be a potential threat, especially in combination with other environmental factors, such as climate change or pollution.
Predation can be a source of mortality for land snails and potential predators have been reviewed by Jordan and Black (2012): “Gastropods are an important food source to a vast number of species, including salamanders, frogs, toads, turtles, snakes, lizards, birds, shrews, voles, moles, rats, mice, chipmunks, and squirrels. Invertebrate predators of terrestrial mollusks include sciomyzid fly larvae, firefly larvae, parasitic wasp larvae, carabid and staphylinid beetles, ants, spiders, and harvestmen.” Introduced predator species or an increase in abundance of native predators due to ecological disturbance can therefore be a potential threat.
Competition with other terrestrial gastropods including exotic species is not documented in the literature but is a potential threat (Whitson 2005; Grimm et al.2010).
In addition to the sampling effort of 1992 and 1996 where fresh, empty shells of recently dead Proud Globelets were found in the south part of the Black Oak Heritage Forest, a total of 17 person-hours of search effort were spent in 2013 in and near this forest (Table 1). The whole forest (north and south part) was searched with an effort of 6 person-hours (Table 1 and Appendix 2). Shells were found only on a small plot at the south border of the forest; one shell also was found further south of the forest in a former light industrial area. In order to increase the probability of encountering live individuals of Proud Globelet, intensive sampling was conducted in the 200 m x 100 m plot in the forest where the shells were initially found with an effort of 14 person-hours (Figure 4, Appendix 2) in different light and weather conditions: (i) warm-humid at dawn, (ii) warm-humid during the morning, (iii) warm-rainy at midday, and (iv) warm-rainy at night. The forest plot was searched using a meandering transect by lifting logs and debris and occasionally digging 5 cm deep in the soil on a surface of 50 cm x 50 cm. Additionally, all observed snails and slugs were collected in the leaf litter and in the first few centimetres of the soil on one square (10 m x 10 m) (Figures 4 and 6) under condition (i) and on one transect under condition (iii). The wood dump of the City of Windsor was also searched. The former light industrial area (Figure 4) was searched as part of a 3 person-hour survey following a transect from the south part of the Black Oak Heritage Forest across the wood dump. The single shell of Proud Globelet was found with other live and dead shells of other snail species in an area estimated to be less than 100 sq. metres.
In total 15 empty, old, weathered shells from juveniles and adults were recorded in this area in 2013. Voucher specimens (shells) were submitted to Robert Forsyth to confirm identification and for curation as well as to Jean-Marc Gagnon of the CMN (Appendix 1). All shells have lost their periostracum (outer protein layer) and the mineral structure was altered (Figure 1). They were estimated to be between 5 and 15 years old following Pearce (2008); shells older than 15 years generally fall into pieces and disintegrate (Nicolai pers. obs.).
Because no live individual has ever been found in Canada, no estimation of abundance of mature individuals can be made. Despite intensive survey efforts only 15 shells were found in 2013, presumably indicating a low level of abundance in the last 15 years. No abundance data are available for other populations of this snail in the U.S.
Figure 4. Search area (yellow) and search square (pink) for live individuals in the south part of the Black Oak Heritage Forest in Windsor, Essex County (green), represented on aerial photographs from 2011 (A) and 1999 (B). The intensive search area corresponds to the zone where shells were found in 1992 (CMNML 096171) and 1996 (CMNML 096170) by M.J. Oldham, and on May 3, July 28 and August 27 (CMNML 096184), 2013 by A. Nicolai (see Appendix 1 and 2). The intensive search area is located close to the bordering wood dump (blue) managed by the City of Windsor. One shell was found on September 5, 2013 by M.J. Oldham (# 41549) south of Black Oak Heritage Forest on a former light industrial area (orange). All records are from habitats that have not substantially changed since 1999. Aerial imagery containing point locations has been removed. Contact the COSEWIC Secretariat for access.
© Environment Canada, 2015
Long description for Figure 4 A and B
Two aerial photos of the south part of the Black Oak Heritage Forest and surrounding area in Windsor, taken in 2011 (Photo A) and 1999 (Photo B). The photos are annotated to show the area of intensive sampling for live individuals of the Proud Globelet (200 by 100 metres), the smaller square area where all observed snails and slugs were collected (10 by 10 metres), the location of a shell found south of the forest in a former light industrial area, and the location of the City of Windsor wood dump.
Figure 5. Wood dump operated by the City of Windsor, Essex County, bordering the south part of the Black Oak Heritage Forest. View of the south entrance of the park with the wood dump on the right side. (Photo credit: Annegret Nicolai.)

Photo: Annegret Nicolai. © Environment Canada, 2015
Long description for Figure 5
Photo of the wood dump operated by the City of Windsor, bordering the south part of the Black Oak Heritage Forest.
Figure 6. Square (10 m x 10 m, flagging tape can be seen on two edges) for the intensive search for live individuals of the Proud Globelet (Patera pennsylvanica) in the zone of shell occurrence in the Black Oak Heritage Forest in the City of Windsor. (Photo credit: Annegret Nicolai.)

Photo: Annegret Nicolai © Environment Canada, 2015
Long description for Figure 6
Photo of part of the area intensively searched for live individuals of the Proud Globelet in the zone of shell occurrence in the Black Oak Heritage Forest in Windsor. Sections of the flagging tape marking a 10 by 10 metre square can be seen.
Figure 7. The new bridge crossing the Detroit River and connecting Highway 75 in Detroit with Highway 401 in Windsor. The construction site in Windsor (blue with dashed line within the red rectangle) is located just north of the Black Oak Heritage Forest (red transparent square within the red rectangle) (modified from Samuel 2009).
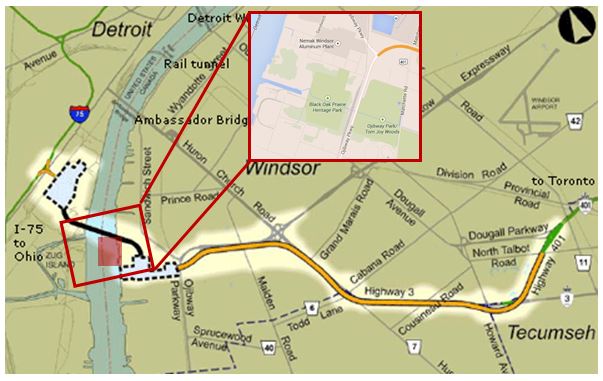
© Environment Canada, 2015
Long description for Figure 7
Map showing the location of the new bridge crossing the Detroit River and connecting Highway 75 in Detroi t with Highway 401 in Windsor. The construction site in Windsor is located just north of the Black Oak Heritage Forest.
Figure 8. Dusky Arion (Arion subfuscus) foraging on a fungus in Black Oak Heritage Forest, Windsor, Essex County. (Photo credit: Annegret Nicolai.)

Photo: Annegret Nicolai. © Environment Canada, 2015
Long description for Figure 8
Photo of Dusky Arion (Arion subfuscus) individuals foraging on a fungus in the Black Oak Heritage Forest, Windsor.
Because no live individuals nor fresh shells of recently dead individuals, as in 1992 and 1996 (see photo on cover page), were found in 2013, population size could not be estimated. The complete absence of live individuals and the age of the shells found in 2013 suggest that the population has substantially declined since 1996, and there is a strong likelihood that Proud Globelet has disappeared from this forest and from Canada as it has only ever been found in and near this forest in southwestern Ontario (see Search Effort, Table 1, Figure 3).
Rescue is unlikely. Natural dispersal from populations in Michigan across the Detroit River is probably rare or non-existent. NatureServe (2013) considers permanent water bodies greater than 30 m wide a dispersal barrier for terrestrial gastropods, and the Detroit and St. Clair rivers and Lake Erie would be formidable barriers to dispersal from adjacent U.S. states.
Because no live individuals or fresh shells of recently dead individuals were found in 2013, the threats calculator (IUCN and CMP 2006; Master et al. 2009) would be difficult to apply as the “scope” and “severity” of the various threats would most likely be “unknown”. Scope is the proportion of the species’ total population that would be expected to be affected by a specific threat within the next 10 years. Severity is the anticipated population reduction in the next 10 years or 3 generations, whichever is longer, of that portion of the population that is exposed to the specific threat. Threats from the past or ongoing threats that probably contributed to the apparent disappearance of the Canadian population and that were observed in the Black Oak Heritage Forest are as follows, using the IUCN and CMP nomenclature and numbering.
A railway crosses the Black Oak Heritage Forest. In the past, the construction of this railroad might have contributed to the decline of the species by the local increase in mortality during construction, maintenance, and operations. Representing a barrier for snails (Baur and Baur 1990) the railway also fragmented the habitat, potentially reducing gene flow, if the population had occupied the entire forest.
A new bridge crossing the Detroit River is expected to be built connecting Highway 75 in Detroit with Highway 401 in Windsor (Figure 7). The New International Trade Crossing or Detroit River International Crossing project is mainly for transportation of goods between Canada and the USA. The connecting Highway 3 between the new bridge and the existing Highway 401 in Windsor is just north of the Black Oak Heritage Forest and the Ojibway Prairie Complex (Figure 7). Although the road, customs inspection plaza and the bridge construction will not directly affect or fragment Proud Globelet habitat, air- and water-borne pollution (e.g., heavy metals and road salt) could significantly increase in the forest due to the expected increase in traffic volume in the area.
The Black Oak Heritage Forest has a high trail density and is intensively visited for recreation (Pratt pers. comm. 2013), but no data on visitor numbers and activities are available. No management plan has been implemented. Trampling by pedestrians is a known threat for some snail species (Baur and Baur 1990; Charrier et al. 2012) and large trails, roads, or railways represent barriers.
The wood dump at the south border of the Black Oak Heritage Forest has changed habitat conditions and might be a source of introduced plant and animal species that have and could further change habitat conditions, and could potentially increase predation or competition.
Urbanization and construction at the periphery of the Black Oak Heritage Forest has destroyed and fragmented the habitat. Construction and then demolition of the light industrial facilities are also likely to alter soil composition, structure and hydrology and have a large impact on snail physiology or behaviour and general ecosystem functioning (Charrier et al. 2012). For instance, changes in soil pH can interfere with calcium absorption, changes in soil structure can alter dormancy refuges or nest conditions for reproduction, and drier soil reduces activity periods and therefore foraging and reproduction. All habitat alterations could have direct consequences on fitness and population dynamics.
Invasive plant species occur throughout southwestern Ontario. Some invasive species are thought to have contributed to declines in larger species of land snails, possibly by altering soil nutrient cycles (Stoll et al.2012).
Among non-native animals, earthworms have one of the greatest impacts on forest ecosystems throughout Canada by altering nutrient distributions, soil pH, soil fauna, and understorey vegetation (Addison 2008). Earthworms reduce leaf litter accumulation and could alter terrestrial snail communities that use leaf litter as habitat (Norden 2010). In the Black Oak Heritage Forest, there were patches of bare earth and just coarse, woody debris, which could be a result of invasive earthworms.
Competition with exotic terrestrial gastropods is a potential threat (Whitson 2005; Grimm et al.2010). The abundance of introduced slug species of the genus Arion in both Proud Globelet occurrence sites was high. The Dusky Arion (Arion subfuscus) was found in the leaf litter and under logs, but especially on fungus in the forest (Figure 8). Dusky Arion and Proud Globelet could compete for fungi as a food source. Grey Fieldslug (Deroceras reticulatum), Grovesnail (Cepaea nemoralis) and Heath Snail (Xerolenta obvia), three other introduced gastropods, are also abundant and found in the same areas where shells of Proud Globelet were found. Remarkably, the snails in the former light industrial area appeared to be localized and restricted to a small area less than approximately 100 m2 but there were thousands of empty X. obvia shells and dozens to hundreds live X. obvia; this was also the first record for this species in southwestern Ontario, it previously only being known in south-central Ontario as early as 1969 (Grimm et al. 2010). It was discovered in a railway yard in nearby Detroit in 2001 (Grimm et al. 2010). As there is a railway line in and adjacent to the Black Oak Heritage Forest, it could be a recent introduction into southwestern Ontario. These other exotic species could be competitors for habitat and other food sources; however, it is difficult to estimate their actual direct impacts on Proud Globelet. The gastropod composition in and around the Black Oak Heritage Forest has definitely changed, including the probable recent disappearance of Proud Globelet and Whitelip Neohelix albolabris, but it is unclear if inter-specific competition or other environmental factors were the drivers of this change.
The high degree of industrialization surrounding the Black Oak Heritage Forest suggests some level of soil, water and air pollution. As xenobiotics are generally accumulated in snail tissues (Barker 2001), the pollution of the site (e.g., salts, heavy metals, artificial organic compounds) might have an impact on Proud Globelet. Acid rain in the area is likely to influence the calcium uptake by snails.
Additionally, garbage dumped in some parts of the forest can lead to local increases in organic and inorganic levels of pollution. Charrier et al. (2013) mentioned the ingestion of small plastic pieces as a potential threat for gastropods, because it could lead to increased mortality. Alternatively, cardboard and other garbage could increase the availability of moist refuges.
Climate change can have a large impact on ecosystem processes thereby changing habitat conditions for Proud Globelet, or directly affect snail survival. In temperate regions, climate change will involve increases in both average temperatures and the frequency of extreme weather events such as heat waves, drought and increased precipitation (Della Marta et al.2007). For instance, heat waves and drought could cause high mortality due to heat or dehydration stresses (Nicolai et al. 2011). Ongoing climate change will disproportionately alter winter conditions (IPCC 2007). The northern distributions of many species are currently thought to be set by winter conditions, so warmer winters could lead to range expansion if suitable habitat exists. Conversely, interactions between climate and microclimate could also result in more adverse conditions, for instance the absence of snow cover induces a decrease of soil temperature accompanied by day/night fluctuations that might affect the survival of species with low cold tolerance plasticity. High temperatures in early spring could lead to arousal, and the following low temperatures could increase mortality. During several years between 1996 and 2013, high temperatures were recorded in March and April followed by extreme low temperatures, e.g., March 1998: highest temperature 22.4°C, lowest temperature -15.9°C (Climate Canada 2014).These abnormal temperature extremes could have contributed to the population decline.
Limiting factors for terrestrial snails include (i) low dispersal or escape capacity, (ii) relatively long generation time, (iii) low physiological resistance to fluctuating environmental factors, such as temperature and humidity, (iv) susceptibility to bioaccumulation of toxic xenobiotics, and (v) limited genetic flow.
Canada’s only population of Proud Globelet is/was in and south of the Black Oak Heritage Forest of the City of Windsor. Given the small size of the occupied habitat where a single event could soon affect all individuals of a taxon present, there are only one or two locations following IUCN criteria (IUCN 2001): Black Oak Heritage Forest and the former light industrial area near the forest.
Proud Globelet is not protected by any legislation, regulations, customs or conditions nationally or worldwide. COSEWIC assessed this wildlife species as Endangered in May 2015. It is not listed under the U.S. Endangered Species Act or under any state acts (US Fish and Wildlife Service 2013). It is not listed under the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES 2013).
Proud Globelet is not listed by IUCN (2013). NatureServe (2013) provides the following ranks.
Global Rank: G4
National Rank (Canada): N1
National Rank (US): N4
Sub-national ranks (S-ranks) in Canada and the U.S. are as follows:
Canadian provinces where Proud Globelet occurs
Ontario: S1 (NHIC 2013). Acts of Ontario do not provide legal protection.
US states adjoining southwestern Ontario
Michigan: SNR and SC (Michigan Natural Features Inventory 2013)
Pennsylvania: S1S2 (Pennsylvania Natural Heritage Program 2013)
Ohio: SNR (Ohio Department of Natural Resources 2012)
New York: not present (Hubricht 1985; Schlesinger 2013)
Other U.S. states where Proud Globelet occurs
Iowa: SNR (Natural Resource Commission Iowa 2009)
Illinois: SNR (Cummings and Phillips 2013)
Indiana: SNR (Indiana Department of Natural Resources 2013)
Kentucky: SNR (Kentucky State Nature Preserve Commission 2013)
West Virginia: S1 (West Virginia Natural Heritage Program 2012)
Missouri: SNR (Missouri Department of Conservation 2014)
(G4 - apparently secure, N1 - critically imperilled nationally, N4 - apparently secure. SNR – not ranked sub-nationally, SC – special concern (at the state level), S1 - critically imperilled sub-nationally, S2 - imperilled sub-nationally, S3 – vulnerable sub-nationally, S4 – apparently secure sub-nationally)
The Black Oak Heritage Forest is owned and protected by the City of Windsor. Access is granted for recreational use. There is no particular management strategy. Changes to the forest are possible under future urban planning.
Although the former light industrial area south of the Black Oak Heritage Forest has remained undeveloped since at least 1999 (Figure 4), the land is available for industrial construction at any time. Therefore if any live Proud Globelet remain in the area, they would be susceptible to potential further habitat loss.
The authors thank Paul Pratt, Head Naturalist at the Ojibway Nature Centre, City of Windsor, for providing maps and information about the development of the Black Oak Heritage Forest. The Nature Conservancy of Canada (NCC) granted permission to access their properties on Pelee Island and provided accommodation at the Ivey Research Station. The report writers thank Mhairi MacFarlane and Dan Kraus of NCC. Tammy Dobbie, at Point Pelee National Park, assisted with fieldwork and provided background information on Middle Island and Point Pelee. Parks Canada granted permission to conduct surveys at Point Pelee and Middle Island. The Middle Sister Island landowner kindly allowed the authors to survey the island. Jane M. Bowles, Mykola Merkulov, Robert Foster, Litza Coello and Hiroko Udaka assisted with fieldwork. Allan Harris assisted with fieldwork and drawing the maps for area estimates. Ontario Parks is thanked by the authors for providing a collecting permit and permission to access provincial parks and protected areas. Robert Forsyth and Dwayne Lepitzki provided comments on proposed fieldwork and offered other support. The authors also thank Valérie Briand for assistance in literature search. The individuals and institutions listed below are thanked by the authors for assistance by supplying data and information.
Addison, J.A. 2008. Distribution and impacts of invasive earthworms in Canadian forest ecosystems. Biological Invasions 11:59-79.
Anderson, F.E. 2007. Population genetics of the carinate pillsnail, Euchemotrema hubrichti: genetic structure on a small spatial scale. Conservation Genetics 8:965-975.
Ansart, A., A. Guiller, O. Moine, M-C. Martin, and L. Madec. 2014. Is cold hardiness size-constrained? A comparative approach in land snails. Evolutionary Ecology 28:471-493.
Ansart, A., and P. Vernon. 2003. Cold hardiness in molluscs. Acta Oecologica 24:95-102.
Asami, T. 1993. Divergence of activity patterns in coexisting species of land snails. Malacologia 35:399-406.
Barker, G.M. 2001. The Biology of Terrestrial Molluscs. CABI Publishing New York, 558 pp.
Baur, A., and B. Baur. 1990. Are roads barriers to dispersal in the land snail Arianta arbustorum? Canadian Journal of Zoology 68:613-617.
Baur, A., and B. Baur. 2005. Interpopulation variation in the prevalence and intensity of parasitic mite infection in the land snail Arianta arbustorum. Invertebrate Biology 124(3):194-201
Blinn, W.C. 1963. Ecology of the land snails Mesodon thyroidus and Allogona profunda. Ecology 44:498-505.
Butt, S., P. Ramprasad, and A. Fenech. 2005. Changes in the landscape of Southern Ontario, Canada since 1750: Impacts of European colonization. Pp. 83-92. In A.Fenech, A., D. MacIver, H. Auld, and R. Hansell (eds.).Integrated Mapping Assessment. Environment Canada, Toronto, Ontario, Canada, 186 pp.
Cain, A.J. 1983. Ecology and ecogenetics of terrestrial molluscan populations. Pp. 597-647 In W.D. Russel Hunter (ed.). The Mollusca, Volume VI. Academic Press, New York. 695 pp.
Charrier, M., A. Nicolai, M.-P. Dabard, and A. Crave. 2013. Plan National d’Actions de Tyrrhenaria ceratina, escargot terrestre endémique de Corse. National Recovery Plan, Ministry of Ecology, Energy, Sustainable Development and the Ocean(ed.), 92 pp.
Churchfield, S. 1984. Dietary separation in three species of shrew inhabiting water-cress beds. Journal of Zoology 204:211–228.
CITES. 2013. Convention on international trade in endangered species of wild fauna and flora. [Accessed November 14, 2013].
Clapp, G.H. 1916. Notes on the land shells of the islands at the western end of Lake Erie and description of new varieties. Annals of the Carnegie Museum 10:532-540.
Climate Canada. 2014. Monthly Data Report for Windsor 1940-2013. [Accessed December 22, 2014].
Coppolino, M.L. 2009. Land Snails of Southern Illinois. Southern Illinois University, Carbondale, Illinois, USA. [Accessed December 18, 2013].
COSEWIC. 2011. Guidelines for recognizing designatable units. [Accessed December 06, 2013].
COSEWIC. In press. COSEWIC assessment and status report on the Broad-banded Forestsnail Allogona profunda in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xx pp.
Crabb E.D. 1928. A predatory Polygyra. Nautilus 42:35-36.
Cummings, K., and C. Phillips. 2013. Mollusca of Illinois. Illinois Natural History Survey. Prairie Research Institute. University of Illinois at Urbana-Champaign. [Accessed November 20, 2013].
Della Marta, P.M., J. Luterbacher, H. von Weissenfluh, E. Xoplaki, M.Brunet, and H. Wanner. 2007. Summer heat waves over western Europe 1880-2003, their relationship to large-scale forcings and predictability. Climate Dynamics 29:251-275.
Douglas, D.D., D.R. Brown, and N. Pederson. 2013. Land snail diversity can reflect degrees of anthropogenic disturbance. Ecosphere 4:28.
Edworthy, A.B., K.M.M. Steensma, H.M. Zandberg, and P.L. Lilley. 2012. Dispersal, home-range size, and habitat use of an endangered land snail, the Oregon forestsnail (Allogona townsendiana). Canadian Journal of Zoology 90(7):875-884.
Emberton, K.C. 1991. The genitalic, allozymic and conchological evolution of the tribe Mesodontini (Pulmonata: Stylommatophora: Polygyridae). Malacologia 33(1–2): 71–178.
ERCA (Essex Region Conservation Authority). 2002. Essex Region Biodiversity Conservation Strategy - Habitat Restoration and Enhancement Guidelines (Comprehensive Version). Dan Lebedyk, Project Co-ordinator. Essex, Ontario. 181 pp.
Forsyth, R.G. 2012. General status of terrestrial molluscs of Canada. Spreadsheet containing a list of and Canadian and provincial or territorial general status ranks. Prepared for the General Status of Wildlife Species 2015 project. 11 December 2012.
Forsyth, R. 2013. Terrestrial molluscs of Canada online database: Bibliography of the terrestrial molluscs of Canada. [Accessed June 10, 2013].
Frest, T.J., and E.J. Johannes. 1995. Interior Columbia Basin mollusk species of special concern. Deixis Consultants, Seattle, Washington. Prepared for the U.S. Department of Agriculture, Forest Service; U.S. Department of the Interior, Bureau of Land Management, Upper Columbia River Basin Ecosystem Management Project. 274 pp. + appendices.
GBIF. 2013. Global Biodiversity Information Facility. [Accessed November 10, 2013].
Goodrich, C. 1916. A trip to the islands in Lake Erie. Annals of the Carnegie Museum 10:527-531.
Grimm, F.W. 1996. Terrestrial molluscs. In I.M. Smith, Assessment of species diversity in the Mixedwood Plains ecosystem. Ecological Monitoring and Assessment Network. [Accessed November 20, 2013].
Grimm, F.W., R.G. Forsyth, F.W. Schueler, and A. Karstad. 2010. Identifying Land Snails and Slugs in Canada: Introduced Species and Native Genera. Ottawa: Canadian Food Inspection Agency. 168 pp.
Heller, J. 2001. Life history strategies. Pp. 413-445. In G.M. Barker (ed.). The Biology of Terrestrial Molluscs. CABI Publishing New York, 558 pp.
Hubricht, L. 1985. The distributions of the native land mollusks of the Eastern United States. Fieldiana Zoology 24:47-171.
Indiana Department of Natural Resources. 2013. Indiana’s state endangered species [PDF; ]. [Accessed November 20, 2013].
IPCC. 2007. Climate Change 2007: Synthesis Report. Core Writing Team, R.K. Pachauri. and A. Reisinger (eds.). International Panel on Climate Change, Geneva, Switzerland. 104 pp.
IUCN. 2001. The IUCN Red List of Threatened Species - 2001 Categories & Criteria (version 3.1). [Accessed January 08, 2014].
IUCN. 2013. The IUCN Red List of Threatened Species. [Accessed November 14, 2013].
IUCN and CMP (International Union for Conservation of Nature and Conservation Measures Partnership). 2006. IUCN – CMP unified classification of direct threats, version 1.0 – June 2006. Gland, Switzerland. 17 pp.
Jennings, T.J., and J.P. Barkham. 1979. Litter decomposition by slugs in mixed deciduous woodland. Holarctic Ecology 2:21-29.
Jordan S.F., and S.H. Black. 2012. Effects of Forest Land Management on Terrestrial Mollusks: A Literature Review. USDA Forest Service, Region 6 USDI Oregon/Washington, Bureau of Land Management. 87 pp.
Kentucky State Nature Preserve Commission. 2013. Endangered, threatened, and special concern plants, animals, and natural communities of Kentucky with habitat description [PDF; 212 KB]. [Accessed December 06, 2013].
Martin, R.J., and E.A. Bergey. 2013. Growth plasticity with changing diet in the land snail Patera appressa (Polygyridae). Journal of Molluscan Studies 79:364-368.
Martin, S.M. 2000. Terrestrial snails and slugs (Mollusca: Gastropoda) of Maine. Northeastern Naturalist 7:33–88.
Mason, C.F. 1970a. Food, feeding rates and assimilation in woodland snails. Oecologia 4:358–373.
Mason, C.F. 1970b. Snail populations, beech litter production, and the role of snails in litter decomposition. Oecologia 5:215–239.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay, and A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk [PDF]. NatureServe, Arlington, Virginia. 57 pp.
Michigan Natural Features Inventory. 2013. Michigan’s special animals. Michigan State University Extension (ed.) Lansing, Michigan. 16 pp.
Missouri Department of Conservation. 2014. Missouri species and communities of conservation concern [PDF]. [Accessed January 06, 2014].
Natural Resource Commission Iowa. 2009. Endangered and threatened plant and animal species [PDF]. IAC 571, Chapter 77. [Accessed January 07, 2014].
NatureServe. 2013. NatureServe Explorer: An online encyclopedia of life. Version 7.1. NatureServe, Arlington, Virginia. [Accessed: November 20, 2013].
Nekola, J.C. 2005. Geographic variation in richness and shell size of eastern North American land snail communities. Records of the Western Australian Museum Supplement No. 68:39–51.
NHIC. 2013. Mollusc species list. Website: Natural Heritage Information Centre, Ontario Ministry of Natural Resources. [Accessed December 06, 2013].
Nicolai, A., J. Filser, R. Lenz, C. Bertrand, and M. Charrier. 2011. Adjustment of metabolite composition in the haemolymph to seasonal variations in the land snail Helix pomatia. Journal of Comparative Physiology B 181:457-466.
Nicolai, A., J. Filser, R. Lenz, V. Briand, and M. Charrier. 2012. Composition of body storage compounds influences egg quality and reproductive investment in the land snail Cornu aspersum. Canadian Journal of Zoology 90:1161-1170.
Nicolai, A., P. Vernon, R. Lenz, J. Le Lannic, V. Briand, and M. Charrier. 2013. Well wrapped eggs: Effects of egg shell structure on heat resistance and hatchling mass in the invasive land snail Cornu aspersum. Journal of Experimental Zoology A319:63-73.
Norden, A.W. 2010. Invasive earthworms: a threat to eastern North American forest snails? Tentacle 18:29-30.
Nyffeler, M., and W.O.P. Symondson. 2001. Spiders and harvestmen as gastropod predators. Ecological Entomology 26:617-628.
Ohio Department of Natural Resources. 2012. Wildlife that are considered to be endangered, threatened, species of concern, special interest, extirpated, or extinct in Ohio. Publication 5356 (R1012). 10 pp.
Oughton, J. 1948. A Zoogeographical Study of the Land Snails of Ontario. University of Toronto Press, Toronto. 128 pp. plus maps, charts, diagrams.
Peake, J. 1978. Distribution and Ecology of the Stylommatophora. Pp. 429-526 In V. Fretter, and J. Peake (eds.). Pulmonates. Academic Press, London. 540 pp.
Pearce T.A. 1990. Spooling and line technique for tracing field movements of terrestrial snails. Walkerana4(12):307-316.
Pearce, T.A. 2008. When a snail dies in the forest, how long will the shell persist? Effect of dissolution and micro-bioerosion.American Malacological Bulletin26:111-117.
Pearce, T.A., and A. Örstan. 2006. Terrestrial gastropoda. Pp. 261-285. In C.F. Sturm, T.A. Pearce, and A. Valdés (eds.). The Mollusks: A Guide to Their Study, Collection, and Preservation. American Malacological Society, Pittsburgh, Pennsylvania. 445 pp.
Pennsylvania Natural Heritage Program. 2013. Species of Special Concern Lists. [Accessed December 06, 2013].
Pilsbry, H.A. 1940. Land Mollusca of North America (North of Mexico). Volume 1. Part 2. The Academy of Natural Sciences of Philadelphia, Philadelphia. 1113 pp.
Pratt, P.D. 2012. Provincially Rare Vascular Plants and Wildlife of the Ojibway Prairie Complex (Version nov2012). Ojibway Nature Centre. Department of Parks, Windsor, Ontario. [Accessed April 20, 2013].
Pratt, P.D., pers. comm. 2013. Email correspondence to A. Nicolai. August 2013. Head Naturalist, Ojibway Nature Centre. Department of Parks, Windsor, Ontario Robinson D.G. 1999. Alien invasions: the effects of the global economy on non-marine gastropod into the United States. Malacologia 41:413-438.
Robinson, D., and J. Slapcinsky. 2005. Recent introductions of alien gastropods into North America. American Malacological Bulletin 20:89-93.
Rosenzweig, C., G. Casassa, D.J. Karoly, A. Imeson, C. Liu, A. Menzel, S. Rawlins, T.L. Root, B. Seguin, and P. Tryjanowski. 2007. Assessment of observed changes and responses in natural and managed systems. Pp. 79-131. In M.L. Change, O.F. Parry, J.P. Canziani, P.J. Palutikof van der Linden, and C.E. Hanson (eds.). Climate Change 2007: Impacts, Adaptation and Vulnerability. Fourth Assessment Report of the Intergovernmental Panel on Climate, Cambridge University Press, Cambridge, UK. 986 pp.
Rowley M.A., E.S. Loker, J.F. Pagels, and R.J. Montali. 1987. Terrestrial gastropod hosts of Parelaphostrongylus tenuisat the National Zoological Park's Conservation and Research Center, Virginia. Journal of Parasitology 73:1084-1089.
Samuel P. 2009. US Gov freezes permitting for new span of Ambassador Bridge Detroit-Windsor. [Accessed December 18, 2014].
Schlesinger, M.D. 2013. Rare Animal Status List January 2013. New York Natural Heritage Program. [Accessed December 06, 2013].
Simpson, G.B. 1901. Anatomy and physiology of Polygyra albolabris and Limax maximus. Bulletin of the New York State Museum 40(8):237-313
South, A. 1980. A technique for the assessment of predation by birds and mammals on the slug Deroceras reticulatum(Müller) (Pulmonata: Limacidae). Journal of Conchology30:229–234.
Steensma, K.M.M., P.L. Lilley and H.M. Zandberg. 2009. Life history and habitat requirements of the Oregon forestsnail, Allogona townsendiana (Mollusca, Gastropoda, Pulmonata, Polygyridae), in a British Columbia population. Invertebrate Biology 128:232-242.
Stiven, A.E., and B.A. Foster. 1996. Density and adult size in natural populations of a southern Appalachian low-density land snail, Mesodon normalis (Pilsbry). American Midland Naturalist 136 (2):287-299.
Stoll, P., K. Gatzsch, H. Rusterholz, and B. Baur. 2012. Response of plant and gastropod species to knotweed invasion. Basic and Applied Ecology 13:232-240.
Turgeon, D.D., J.F. Quinn, Jr., A.E. Bogan, E.V. Coan, F.G. Hochberg, W.G. Lyons, P. Mikkelsen, R.J. Neves, C.F.E. Roper, G. Rosenberg, B. Roth, A. Scheltema, F.G. Thompson, M. Vecchione, and J.D. Williams. 1998. Common and Scientific Names of Aquatic Invertebrates from the United States and Canada: Mollusks, Second Edition. American Fisheries Society Special Publication. 26. Bethesda, Maryland. 526 pp.
US Fish and Wildlife Service. 2013. Endangered Species. [Accessed November 14, 2013].
Van Cleave, H.J. and T.D. Foster. 1937. The seasonal life history of land snail Polygyra thyroidus (Say). Nautilus 51:50-54.
Walsh, G.E., and B.F. Coles. 2006. Daedalochila lithicaand Daedalochila dorfeuilliana (Gastropoda : Polygyridae) in Arkansas, USA: morphology, distribution, and habitat. Nautilus 120(4):131-138.
Webb, G.R. 1954 Pulmonata, Polygyridae, Polygyrinae: the sexology and taxonomy of seven species of land-snails of the genus Mesodon. Gastropodia 1:19-20.
West Virginia Natural Heritage Program. 2012. Rare, threatened, and endangered animals [PDF; 391 KB]. [Accessed December 06, 2013].
Whitson, M. 2005. Cepaea nemoralis (Gastropoda, Helicidae): The Invited Invader. Journal of the Kentucky Academy of Science 66:82–88.
Annegret Nicolai is a biologist at the Western Catholic University in Angers, France. She has a Ph.D. from the University of Bremen in Germany and from the University of Rennes 1 in France. Her research involves investigating eco-physiological questions in terrestrial snails, specifically about the impact of climate change and resource availability on the physiology and reproduction in endangered and invasive species. She also co-supervises a Ph.D. thesis analyzing the pathways of invasion of terrestrial gastropods and adaptation to climate change. She has very specific knowledge about the biology, anatomy, physiology and ecology of terrestrial gastropods. In Germany she developed a captive-breeding program for the protected Helix pomatia and in France she was co-author of the Species Report and the Recovery Strategy in Tyrrhenaria ceratina in Corsica. In the Sinclair lab at Western University, Ontario, she investigated the overwintering strategy of Cepaea nemoralis. Since 2012 she has been surveying terrestrial gastropods in Ontario and participating in the “barcoding of life” project at the University of Guelph. She became a member of the mollusc subcommittee of COSEWIC in 2014.
Michael J. Oldham has been a biologist with the Natural Heritage Information Centre, Ontario Ministry of Natural Resources (OMNR), for the past 20 years. Previously he worked for the OMNR and conservation authorities in Aylmer, Chatham, Essex, London, Richmond Hill, and Toronto. He has a B.Sc. in Biology from the University of Guelph and is a former member of COSEWIC and the COSEWIC Amphibians and Reptiles SSC and a current member of the Vascular Plants SSC. He also served for more than a decade on the provincial Committee on the Status of Species At Risk in Ontario (COSSARO). He has authored or co-authored more than a dozen COSEWIC status reports. He has been interested in terrestrial gastropods for more than 20 years and has studied and collected them throughout Ontario.
The Global Biodiversity Information Facility, GBIF, database was used to examine several collections worldwide: FLMNH - Florida Museum of Natural History, MCZ - Museum of Comparative Zoology at Harvard University, LI - Biologiezentrum Linz Oberösterreich, NMNH - National Museum of Natural History, Smithsonian Institution, ANSP - Academy of Natural Sciences Philadelphia, NMR - Natural History Museum Rotterdam, FMNH - Field Museum of Natural History, NTSRV - NatureServe Central Databases, MACN - Museo Argentino de Ciencias Naturales (Appendix 3). Collections in Canadian Museums were examined: CMN – Canadian Museum of Nature, ROM – Royal Ontario Museum (Appendix 1) and in Bishops Mills Natural History Centre.