
Sable Island sweat bee (Lasioglossum sablense): COSEWIC assessment and status report 2014

Photograph taken by Jason Gibbs.
Sable Island Sweat Bee
2014
Table of contents
- Document information
- Assessment summary
- Executive summary
- Technical summary
- Preface
- Wildlife species description and significance
- Distribution
- Habitat
- Biology
- Population sizes and trends
- Threats and limiting factors
- Protection, status and ranks
- Acknowledgements and authorities contacted
- Information sources
- Biographical summary of report writers
- Collections examined
List of figures
- Figure 1. Lasioglossum sablense female, lateral view (modified from Gibbs 2010). Specimen collected on 8 August 2008 and held at Packer Collection York University. Photograph taken by Jason Gibbs.
- Figure 2. Lasioglossum sablense male, lateral view (modified from Gibbs 2010). Specimen collected on 8 August 2008 and held at Packer Collection York University. Photograph taken by Jason Gibbs.
- Figure 3. Neighbour-joining tree of cytochrome c oxidase subunit 1 sequences of L. sablense and related Lasioglossum species.
- Figure 4. North American map showing location of Sable Island, Nova Scotia.
- Figure 5. Map of Sable Island, Nova Scotia. Collection records of the Sable Island Sweat Bee are shown as black dots.
- Figure 6. Search effort applicable to the Sable Island Sweat Bee (Lasioglossum sablense). Each red dot represents one collection locality from which Lasioglossum specimens have been examined within the past ten years (Gibbs 2010, 2011, pers. data.). The black dots are Sable Island Sweat Bee search effort and records.
- Figure 7. A collection site of the Sable Island Sweat Bee, Lasioglossum sablense, near the Sable Island Station. Photograph taken July 2013 by Zoe Lucas.
List of tables
- Table 1. Summary of 2013 Sable Island bee collections. (Y = yellow bowl trap, P = pink bowl trap, B = blue bowl trap, G = green bowl trap).
- Table 2. Weather data for Sable Island (1971 – 2000) (climate.weather.gc.ca).
- Table 3. Threats assessment for Sable Island Sweat Bee based on the IUCN-CMP (World Conservation Union–Conservation Measures Partnership) unified threats classification system. For a detailed description of the threat classification system see the CMP website (CMP 2010) and Master et al. (2009).
Document information
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Cananda
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2014. COSEWIC assessment and status report on the Sable Island Sweat Bee Lasioglossum sablense in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. ix + 38 pp. (Species at Risk Public Registry).
Production note:
COSEWIC would like to acknowledge Jason Gibbs for writing the status report on the Sable Island Sweat Bee, Lasioglossum sablense, in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Jennifer Heron, Co-chair of the COSEWIC Arthropods Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
E-mail: COSEWIC E-mail
Website: COSEWIC
Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur L’halicte de l’île de Sable (Lasioglossum sablense) au Canada.
Cover illustration/photo:
Sable Island Sweat Bee -- Photograph taken by Jason Gibbs.
COSEWIC assessment summary
Assessment summary – November 2014
- Common name
- Sable Island Sweat Bee
- Scientific name
- Lasioglossum sablense
- Status
- Threatened
- Reason for designation
- This species is globally endemic to Sable Island, Nova Scotia, and occurs as one isolated population with a very small range and no possibility of rescue. The island has only about 13 km 2 of vegetated area that provides forage/nesting sites for this bee. Nesting likely occurs near or within this vegetated area and sweat bees are not known to travel large distances (i.e. > 200 m) for forage. Increased frequency and severity of storms, in addition to climate change and related sea level rise, are expected to drive change which will further decrease the quality and quantity of bee habitat on the island. Eco-tourism is also a potential future threat, which may also increase the introduction and spread of invasive species. Habitat on the island is also susceptible to invasive plant species, introduced horses, and seawater flooding.
- Occurrence
- Nova Scotia
- Status history
- Designated Threatened in November 2014.
COSEWIC executive summary
Sable Island Sweat Bee
Lasioglossum sablense
Wildlife species description and significance
The Sable Island Sweat Bee, Lasioglossum sablense Gibbs, is a small (5–6 mm), dull-metallic sweat bee in the family Halictidae. The species is endemic to Canada, occurring solely on Sable Island, Nova Scotia. Both sexes can be distinguished from the three other bee species (two of these sweat bees) on Sable Island by the combination of their small size and the dense lateral punctures on the dorsal part of the thorax.
Distribution
The global and Canadian distribution of the species is confined to Sable Island, Nova Scotia, which is approximately 34 km2 in area, excluding the intertidal zone. The island is isolated from mainland Nova Scotia by a distance of approximately 150 km.
Habitat
Sable Island is primarily composed of sand, with low levels of organic material in the sandy soil. Approximately 13 – 15 km2 of the island (39%) has vegetation and is considered potential bee habitat. Vegetated areas are composed of a few distinct plant communities, the largest of which are Marram-Forb grasslands, sparse grass lands and heath. Climatic conditions are cool and foggy with high winds during the summer and relatively warm conditions during the winter. Females dig underground nests. Flowering plants are visited for pollen and nectar resources.
Biology
The Sable Island Sweat Bee is a ground-nesting species and a generalist floral visitor. Inseminated females overwinter as adults and emerge in spring to form nests. Reproductive males and females are produced in the late summer. Adults fly from at minimum June 1st to September 11th. Related species are known to have social organization in nests, but the social behaviour of the Sable Island Sweat Bee remains unstudied.
Population size and trends
The population size of Sable Island Sweat Bee is not possible to estimate given the collection data available for the species. Of the four bee species occurring on Sable Island, the Sable Island Sweat Bee is the least commonly collected. Historical records are too sparse to effectively estimate historical trends. Relative proportions of the two sweat bees on the island, L. novascotiae (Mitchell) to the Sable Island Sweat Bee, collected with nets in 1966 –1967, 2008, and 2013 are comparable (3:1, 1:1, 2:1, respectively).
Population sizes have likely decreased over historical time due to decreases in the spatial vegetated area on Sable Island.
Threats and limiting factors
Loss of habitat due to the inundation of vegetation by sand or submersion of low-lying areas with rising sea levels would have negative impacts on population sizes of the Sable Island Sweat Bee. Harsh weather conditions could compound this effect while also reducing adult foraging activity.
Past human influence may have also reduced the extent and diversity of flowering vegetation. Current human activity is minimal due to the isolation of the island and the control of visitors. There is potential with increased future human visitation to the island to introduce non-native bee species.
Protection, status, and ranks
The Sable Island Sweat Bee is not protected under federal or provincial legislation. The species has not been assigned a conservation status rank. The species’ habitat is within Sable Island National Park Reserve, which is protected under the Parks Canada Act.
Technical summary
- Scientific name:
- Lasioglossum sablense
- English name:
- Sable Island Sweat Bee
- French name:
- Halicte de l’île de Sable
- Range of occurrence:
- Nova Scotia
Demographic information
Generation time
- 1 year to complete life cycle
Is there an [observed, inferred, or projected] continuing decline in number of mature individuals?
Historical data are too sparse for accurate estimate. Human influence and changing island geography suggest a long-term decline in suitable habitat and inferred decline in mature individuals.
- Inferred.
Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations]
Historical data are too sparse for accurate estimate.
- Unknown.
[Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations]
Historical data are too sparse for accurate estimate. Human influence and changing island geography suggest a long-term decline in suitable habitat and inferred decline in mature individuals.
- Inferred reduction.
[Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations]
Historical data are too sparse for accurate estimate. Human influence and changing island geography suggest a long-term decline in suitable habitat.
- Unknown.
[Observed, estimated, inferred or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future.
Historical data are too sparse for accurate estimate. Human influence and changing island geography suggest a long-term decline in suitable habitat and inferred decline in mature individuals.
- Inferred reduction.
Are the causes of the decline clearly reversible and understood and ceased?
- Not reversible; not fully understood; not ceased.
Are there extreme fluctuations in number of mature individuals?
Based on life cycle patterns of related Lasioglossum bee species.
- Not likely.
Extent and occupancy information
Estimated extent of occurrence (minimum convex polygon)
- 118 km2
Index of area of occupancy (IAO, 2 x 2 km2 grid values)
(Always report 2x2 grid value).
- 16 km2
Is the population severely fragmented?
- No
Number of locations
- < 5
Is there an observed continuing decline in extent of occurrence?
Projected habitat loss, extent and quality based on sand ecosystem erosion and sea level increases.
- Yes
Is there an observed continuing decline in index of area of occupancy?
Projected habitat loss, extent and quality based on sand ecosystem erosion and sea level increases.
- Yes
Is there an [observed, inferred, or projected] continuing decline in number of populations?
- No
Is there an [observed, inferred, or projected] continuing decline in number of locations?
- No
Is there an [observed, inferred, or projected] continuing decline in quality of habitat?
Projected habitat loss, extent and quality based on sand ecosystem erosion and sea level increases.
- Yes
Are there extreme fluctuations in number of populations?
- No
Are there extreme fluctuations in number of locations?
- No
Are there extreme fluctuations in extent of occurrence?
- No
Are there extreme fluctuations in index of area of occupancy?
- No
Number of mature individuals
Total SLE Population:
- Unknown
Quantitative analysis
Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years].
- Not calculated
Threats (actual or imminent, to populations or habitats)
Possible habitat loss due to sand ecosystem erosion caused by wind, wave action, storms, feral horses and human foot traffic. Loss of habitat due to rising sea levels and flooding due to storms. Potential for introductions of non-native vegetation and additional non-native (to the island) species leading to competition.
Rescue effect (immigration from outside Canada)
Status of outside population(s)?
- Endemic to Sable Island, no outside populations.
Is immigration known or possible?
- Not applicable
Would immigrants be adapted to survive in the SLE?
- Not applicable
Is there sufficient habitat for immigrants in the SLE?
- Not applicable
Is rescue from outside populations likely?
- Not applicable
Data-sensitive species
- Is this a data-sensitive species?
- No
Status history
- COSEWIC: Designated Threatened in November 2014.
Status and reasons for designation:
- Status:
- Threatened
- Alpha-numeric code:
- D2
- Reasons for designation:
- This species is globally endemic to Sable Island, Nova Scotia, and occurs as one isolated population with a very small range and no possibility of rescue. The island has only about 13 km 2 of vegetated area that provides forage/nesting sites for this bee. Nesting likely occurs near or within this vegetated area and sweat bees are not known to travel large distances (i.e. > 200 m) for forage. Increased frequency and severity of storms, in addition to climate change and related sea level rise, are expected to drive change which will further decrease the quality and quantity of bee habitat on the island. Eco-tourism is also a potential future threat, which may also increase the introduction and spread of invasive species. Habitat on the island is also susceptible to invasive plant species, introduced horses, and seawater flooding.
Applicability of Criteria
- Criterion A (Decline in total number of mature individuals):
- Not applicable. Population trends unknown. Historical data is too few (or sparse) since the island has few historic and recent intensive collection events. This species was only described in 2010, and few bee (or insect) surveys have been conducted due to isolation.
- Criterion B (Small distribution range and decline or fluctuation):
- Not applicable. The EO is less than 5,000 km 2 (118 km 2) and the IAO is less than 500 km 2 (16 km 2), but it does not meet other components of the B criterion. There is some decline in habitat quality, but number of locations could not be determined.
- Criterion C (Small and declining number of mature individuals):
- Not applicable. Historical data are too sparse.
- Criterion D (Very small or restricted population):
- Meets Threatened D2 because the IAO is less than 20 km 2 and the species could become Endangered within a very short time period if there are extreme storm and flooding events.
- Criterion E(Quantitative analysis):
- Not applicable. Quantitative analysis has not been completed.
Preface
COSEWIC history
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2014)
- Wildlife species
- A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
- Special Concern (SC)
(Note: Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.) - A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
- Not at Risk (NAR)
(Note: Formerly described as “Not In Any Category”, or “No Designation Required.”) - A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
- Data Deficient (DD)
(Note: Formerly described as “Indeterminate” from 1994 to 1999 or “ISIBD” [insufficient scientific information on which to base a designation] prior to 1994. Definition of the [DD] category revised in 2006.) - A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife species description and significance
Name and classification
Kingdom Animalia
Phylum Arthropoda
Class Insecta
Order Hymenoptera
Superfamily Apoidea
Family Halictidae
Subfamily Halictinae
Tribe Halictini
Genus Lasioglossum Curtis
Subgenus Dialictus Robertson
Species Lasioglossum sablense Gibbs
English Common Name: Sable Island Sweat Bee
French Common Name: Halicte de l’île de Sable, Lasioglosse de l’île de Sable
Taxonomic synonyms: None
Type specimens:
The type locality of the species is Sable Island, Nova Scotia, Canada. The holotype specimen is deposited at the Packer Collection at York University (PCYU), Toronto. Paratypes are deposited at PCYU and the Canadian National Collection of Insects, Arachnids, and Nematodes (CNC), Ottawa.
Taxonomic background and similarities:
Lasioglossum sablense Gibbs was first described in a revision of the subgenus Lasioglossum for Canada (Gibbs 2010). Sable Island Sweat Bee was first recorded by Wright (1989) as “Lasioglossum sp.” [sic]. The original taxonomic description was based on 3 females and 5 males collected on the 8 August 2008 (PCYU) and 2 females and 2 males collected on the 11–15 September 1967 (CNC).
The Sable Island Sweat Bee is a member of the subfamily Halictinae and considered a ‘sweat bee’; some species are known to visit humans to drink their perspiration. Sweat bees are a diverse, widely distributed group of bees. Approximately 170 described species of sweat bee occur in Canada. The genus Lasioglossum has over 1800 described species worldwide (Ascher and Pickering 2014) and is present within most terrestrial ecosystems that have a component of flowering plants (Michener 2007). Lasioglossum sablense is the only sweat bee with such a restricted distribution in Canada.
Morphological description
The Sable Island Sweat Bee is a small, dull-metallic, sweat bee, approximately 5 to 6 mm in length. Females have pollen-collecting hairs on the hind legs and abdomen (Figure 1). Males are recognizable by their elongate antennae and slender abdomen (Figure 2). Both sexes have dense punctures on the dorsal surface of the thorax, adjacent to the wing bases. One other sweat bee, L. novascotiae, occurs on Sable Island, but can be distinguished by the sparse punctures on the dorsal surface of the thorax adjacent to the wing bases. The dense pale hairs on the end of the female’s abdomen can be used to distinguish Sable Island Sweat Bee females from those of the more sparsely pubescent L. novascotiae (Mitchell), even with the naked eye. Identification keys for adult forms are provided in Gibbs (2010) and Gibbs (2011).
Immature forms of the Sable Island Sweat Bee have not been studied, but are expected to appear similar to other Lasioglossum species. Michener (1953) described the larvae of Lasioglossum imitatum (Smith) and several other sweat bee species.

Long description for Figure 1
Photo of a female Sable Island Sweat Bee specimen (lateral view). The bee is a dull metallic colour, with pollen-collecting hairs on the hind legs and abdomen. It has dense punctures on the dorsal surface of the thorax, adjacent to the wing bases.

Long description for Figure 2
Photo of a male Sable Island Sweat Bee specimen (lateral view). The bee is a dull metallic colour and has dense punctures on the dorsal surface of the thorax, adjacent to the wing bases. Males are recognizable by their elongate antennae and slender abdomen.
Population spatial structure and variability
The known range of the Sable Island Sweat Bee is limited to Sable Island, a small geographic area without physical barriers sufficient to prevent gene flow between flying insects. The small size of the island and the known flight range of other sweat bees (Batra 1984; Greenleaf et al. 2007) would easily allow for gene flow to occur across the species’ range.
A short fragment (407 base pairs) of the mitochondrial gene cytochrome oxidase c subunit 1 was sequenced from five individuals of the Sable Island Sweat Bee (Gibbs unpublished data). There is evidence of allelic variation at two nucleotide positions. At one position this variation was only within individuals, suggesting heteroplasmy. This level of variation suggests a maximum genetic divergence of 0.25% within the population, but rigorous sampling has not been undertaken (Figure 3). P-distances from related species exceed 1%.
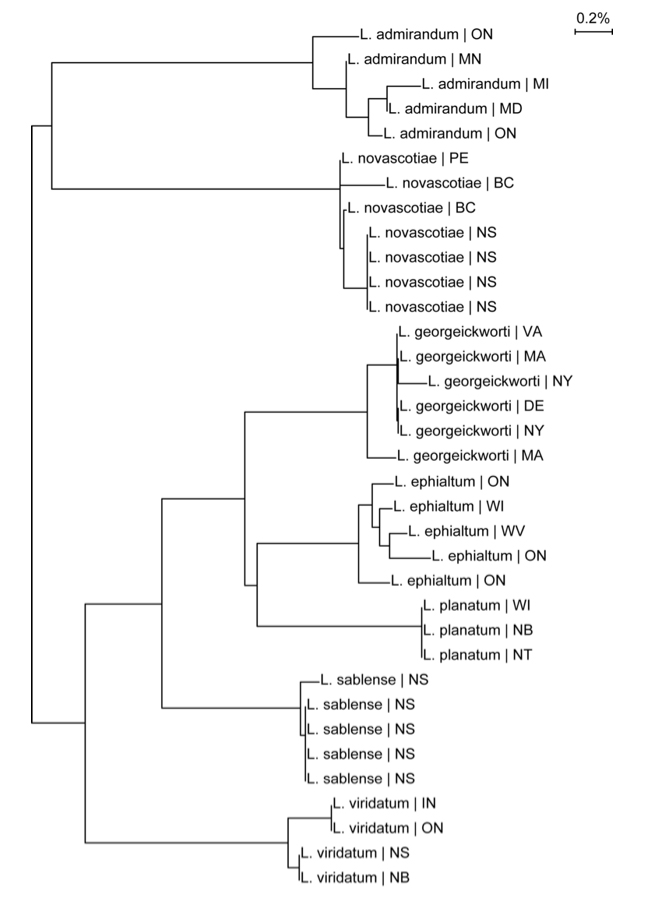
Long description for Figure 3
Neighbour-joining tree diagram of cytochrome c oxidase subunit 1 sequences of L. sablense and six related Lasioglossum species.
Designatable units
The Sable Island Sweat Bee is being assessed as one designatable unit. There is no information on discreteness or evolutionary significance among populations, although this is unlikely given the small global range extent of this species. The species occurs within the Atlantic Ecological Area (COSEWIC 2011).
Special significance
The Sable Island Sweat Bee is endemic to Canada, occurring solely on Sable Island, Nova Scotia. Three other bee species occur on the island: a sweat bee, Lasioglossum novascotiae, and two megachild bees, Megachile melanophaea Smith and Osmia simillima Smith. Only the Sable Island Sweat Bee is endemic to the island; the other three species are distributed across continental North America.
Sable Island is a unique ecosystem and includes several species or varieties of plants with restricted ranges (Catling et al. 1984). Some of these plants such as Hieracium scabrum (Michx.) var. leucocaule Fern. & St. John is visited by the Sable Island Sweat Bee. Pollination services by the Sable Island Sweat Bee and other bees on the island are likely crucial for the maintenance of these restricted plant populations on the island.
The bee is not known to have cultural significance to First Nations in the region. The first recorded humans to visit the island were Portuguese explorers in the 16th century. Human habitation over the next several centuries included the growing of vegetable crops and the exporting of wild cranberries. The Sable Island Sweat Bee would have contributed to the pollination of these crops.
Distribution
Global range and Canadian range
The global and Canadian range of the Sable Island Sweat Bee is restricted to Sable Island, Nova Scotia, Canada (Figure 4 and 5).
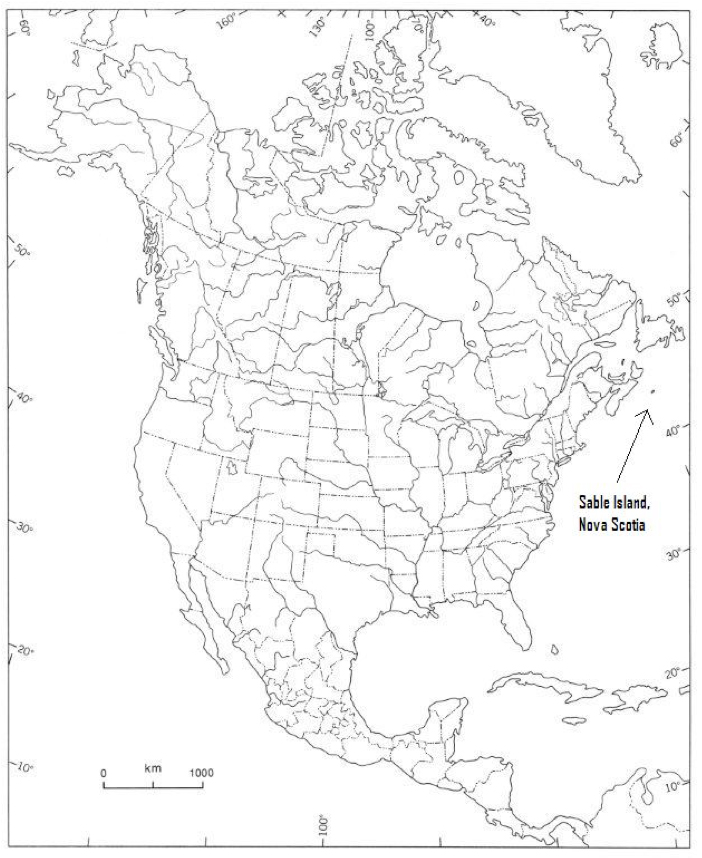
Long description for Figure 4
Map of North America showing the location of Sable Island, Nova Scotia, relative to mainland North America. Located at approximately 44°N and 60°W, Sable Island lies near the outer edge of the continental shelf, 300 km east of Halifax, Nova Scotia.

Extent of occurrence and area of occupancy
Sable Island is approximately 34 km2. Approximately 39% (13.26 km2) of the island is vegetated (Catling et al. 1984), and the Sable Island Sweat Bee requires the flowering plants that occur in these vegetated areas for foraging. The foraging range for a bee of this size is likely a distance less than 1 km (Greenleaf et al. 2007). The windy conditions of Sable Island make it less likely that nesting areas are separated from vegetated areas. The biological area of occupancy for the Sable Island Sweat Bee is likely 13 – 15 km2.
Extent of occurrence (EO) for the Sable Island Sweat Bee based on a minimum convex polygon is 118 km2, most of this larger area is ocean and unsuitable habitat. An index of area of occupancy (IAO) using a grid of 2 km X 2 km squares is 16 km2.
Search effort
The Sable Island Sweat Bee is considered endemic to Sable Island based on the examination of thousands of specimens from Nova Scotia (e.g. Sheffield et al. 2013), New Brunswick, Prince Edward Island and Newfoundland, as well as other provinces and states, during the course of revisionary studies of Canadian and American Lasioglossum (Halictidae) (Gibbs 2010, 2011). Since completion of these taxonomic revisions, thousands of additional Lasioglossum specimens have been examined from these four provinces and no Sable Island Sweat Bees have been found outside Sable Island (Gibbs pers. data; Sheffield pers. comm. 2013). See Figure 6 for a compilation of recent search effort for Lasioglossum species.
Long description for Figure 6
Map illustrating collection localities in eastern Canada and the eastern United States from which Lasioglossum species have been examined within the past 10 years. Sable Island Sweat Bee is recorded only on Sable Island.
The number of known specimens of the Sable Island Sweat Bee is very small due to limited sampling of bees on Sable Island. Additional specimens from field studies on Sable Island may be within unsorted samples housed in the Nova Scotia Museum (NSM, Halifax) and the CNC.
The Sable Island Sweat Bee specimens in the type series came from faunal surveys conducted in 1966 and 1967 (Howden et al. 1970) and 2008 (Catling et al. 2009). Approximately 800 specimens of unidentified Lasioglossum from Nova Scotia were identified from a recent loan of material from the CNC. A small proportion of these was from Sable Island and was composed of L. novascotiae (15 specimens) and the Sable Island Sweat Bee (5 specimens including the 4 CNC paratypes). These are the only two species of Halictidae confirmed for Sable Island (Catling et al. 2009; Gibbs 2010). Previous reports of L. lineatulum (Crawford) from Sable Island (Wright 1989) were probably misidentified specimens of L. novascotiae. Two additional species of bee, Megachile melanophaea and Osmia simillima, (both Megachilidae), are also known to occur on Sable Island (Wright 1989).
There are currently 106 bee specimens from Sable Island held at the NSM (Westby pers. comm. 2014). Six females (11-VIII-2011, 14-VIII-2011, 11-VII-2008, and 12-IX-1977) and 12 males (11-VIII-2011, 18-VIII-2008) of the Sable Island Sweat Bee were collected over the five collection dates. Few field observations accompany the specimens, including that some females were caught on cranberry (Vaccinium macrocarpon Aiton) and some males were caught within heath vegetation near a pond.
Bee collections in 2013 from Sable Island were completed using bowl traps and sweep netting. A total of 734 bees were collected; thirty-eight specimens were Sable Island Sweat Bees, including 19 females and 19 males. Search effort on Sable Island is summarized in Table 1.
| General survey location | Date | L. sablense detected |
Collection method |
Search effort |
Collector |
|---|---|---|---|---|---|
| Sable Island West Light | 01-Jun-2013 | Yes | 2Y,2P,2B,1G | 13 hours | Rob Ronconi |
| Sable Island West Light | 04-Jun-2013 | No | 1Y,1P,1B | 4 hours, 25 minutes |
Rob Ronconi |
| Sable Island West Light | 05-Jun-2013 | No | 1Y,1P,1B, 1G | 10 hours, 40 minutes |
Rob Ronconi |
| Sable Island West Light | 06-Jun-2013 | Yes | 1Y,1P,1B, 1G | 13 hours, 45 minutes |
Rob Ronconi |
| Sable Island East Light | 10-Jun-2013 | No | 2Y,1P,2B,2G | 8 hours, 5 minutes |
Rob Ronconi |
| Sable Island West Light | 13-Jun-2013 | No | 2Y,1P,2B,2G | 11 hours, 15 minutes |
Rob Ronconi |
| Sable Island West Light | 15-Aug-2013 | No | 2Y,1P,2B,2G | 11 hours, 5 minutes |
Rob Ronconi |
| Sable Island West Light | 16-Aug-2013 | No | 2Y,1P,2B,2G | 5 hours, 15 minutes |
Rob Ronconi |
| Sable Island West Light | 17-Aug-2013 | No | 2Y,1P,2B,2G | 12 hours, 20 minutes |
Rob Ronconi |
| Sable Island West Light | 18-Aug-2013 | Yes | 2Y,1P,2B,2G | 10 hours, 10 minutes |
Rob Ronconi |
| Sable Island East Light | 19-Aug-2013 | Yes | 2Y,1P,2B,2G | 4 hours | Rob Ronconi |
| Sable Island Station | 30-Jun-2013 | Yes | Netting from Rosa | 5 minutes | Zoe Lucas |
| Sable Island Station | 30-Jun-2013 | No | Netting from Rosa | 5 minutes | Zoe Lucas |
| Sable Island Station | 30-Jun-2013 | No | Sweep netting | untimed | Zoe Lucas |
| Sable Island Station | 30-Jun-2013 | Yes | Sweep netting | 10 minutes | Zoe Lucas |
| Sable Island Station | 22-Jul-2013 | No | Sweep netting | 10 minutes | Zoe Lucas |
| Sable Island Station | 22-Jul-2013 | No | 1Y | 1 hour | Zoe Lucas |
| Sable Island Station | 22-Jul-2013 | No | 1Y | 1 hour | Zoe Lucas |
| Sable Island Station | 22-Jul-2013 | No | 5Y | 1 hour | Zoe Lucas |
| Sable Island Calopo Pond | 25-Jul-2013 | Yes | Sweep netting | 3 minutes | Zoe Lucas |
| Sable Island Station | 25-Jul-2013 | No | 1Y | 1 hour | Zoe Lucas |
| Sable Island Station | 25-Jul-2013 | No | 3Y | 2 hours | Zoe Lucas |
| Sable Island Station | 26-Jul-2013 | No | 5Y | 3 hours 30 minutes |
Zoe Lucas |
| Sable Island Station | 26-Jul-2013 | No | 2Y | 3 hours 15 minutes |
Zoe Lucas |
| Sable Island Station | 26-Jul-2013 | No | 1Y | 3 hours 15 minutes |
Zoe Lucas |
| Sable Island Station | 26-Jul-2013 | No | 1Y | 3 hours | Zoe Lucas |
| Sable Island Station | 30-Jul-2013 | Yes | Sweep netting | 1 hour | Zoe Lucas |
| Sable Island Station | 30-Jul-2013 | No | Netting Hieracium | 1 hour | Zoe Lucas |
| Sable Island Station | 30-Jul-2013 | Yes | Netting Hieracium | 1 hour | Zoe Lucas |
| Sable Island Station | 30-Jul-2013 | No | 9Y | 4 hours | Zoe Lucas |
| Sable Island Station | 17-Aug-2013 | No | Netting Rosa | 10 minutes | Zoe Lucas |
Habitat
Habitat requirements
The Sable Island Sweat Bee is a ground-nesting bee with a likely nesting preference for sandy soils. Several mainland species of Lasioglossum are also limited to sand dune habitats (Gibbs 2010, 2011). Soils on the island have low organic matter content (Catling et al. 1984), which further supports the importance of sand as a nesting substrate. Vegetated areas are composed of a few distinct plant communities the largest of which are Marram-Forb grasslands, sparse grasslands and heath.
Sable Island Sweat Bees are present throughout most of the growing season, and thus foraging adults require a variety of floral resources from spring to fall. Females are active from at least June 1st to September 11th. Floral records come from six genera in five families - Asteraceae: Achillea (yarrow), Hieracium (hawkweed), Ericaceae: Vaccinium (blueberries, cranberries), Onagraceae: Oenothera (evening primrose), Polygonaceae: Polygonum (knotweed), and Rosaceae: Rosa (wild rose), demonstrating the species is polylectic (Gibbs 2010). Related species in the subgenus Lasioglossum have similar habitat requirements. A habitat photograph is shown in Figure 7.

Long description for Figure 7
Photo of a Sable Island Sweat Bee collection site near the Sable Island Station (weather station run by Parks Canada and Environment Canada). The image shows a flattish expanse of grass and shrubs with sandy patches and structures related to the weather station.
Habitat trends
Sable Island is estimated to be 14 to 16 thousand years old (King 2001). Over geological time, the island may have once been part of larger glacial refugia during the Wisconsinan ice age, when sea levels were 120 m lower than present (King and Fader 1986). The island is a constantly shifting sand bar, which is being gradually lost and rebuilt over time by waves. At present, Sable Island is part of a larger submerged sand mass near the edge of the continental shelf (Cameron 1965).
Sable Island does not appear to have decreased in size over the past few hundred years, although the shape and area of vegetation available to the bee may have changed. Cameron (1965) completed historical mapping and concluded the island is not overall rapidly eroding. Blowing winds and wave action cause the reduction in size in the western part of the island and build up on the eastern edge (Cameron 1965). Taylor (1982) notes the role of offshore, submarine sand bars in stabilizing the corresponding shoreline of the island. Apparently, more sand transport and instability occurs where such sand bars erode or are absent. Scott et al. (1984) found that while sea level has been rising steadily for the last 10,000 years, Sable Island has been stable over the past 7,500 years, continually aggrading as the sea rose. While the complete sinking of the island is not out of the question, Sable Island appears to also be rising, and this would prolong such a sinking event.
More recently Environment Canada (1998) summarized the above and more recent information, the island stability, and the long-term picture of dynamic weather including storms, erosion and accretion, impacts to vegetation and the conservation of the island.
With respect to Sable Island Sweat Bee, the extent and area of vegetation that provides the floral resources and nest sites for the species, and the habitat trends for these areas, are the key factors to its survival. There are challenges to assessing vegetative changes on Sable Island. In extreme winds, vegetated portions of dunes can become destroyed thus forming large blowouts (Cameron 1965). Freeman (1996) concluded there was little vegetation change between 1963, 1972, and 1981. However, in more recent years the spatial area may be changing and the extent of vegetated terrain may be decreasing as moving sand covers vegetation (Catling et al. 1984; Catling pers. comm. 2014).
Rising sea levels could shrink the island, and it is believed that this decrease would also result in a loss of vegetated area, which would have had a strong negative effect on populations of Sable Island Sweat Bee. For example, Lake Wallace, an inland lake on the south side of the island, has experienced numerous breaches of the dunes protecting the lake and subsequent influx of ocean water during storms (Environment Canada 1998).
Additional long-term decline in habitat quality has been ongoing since non-native horses were introduced to the island in the mid-1700s. The threats to the ecosystem from horses are discussed in the Threats section (IUCN Threat 8).
Biology
The Sable Island Sweat Bee belongs to a cosmopolitan (occurring on all continents except Antarctica) genus. Canadian members of the subgenus Lasioglossum were revised by Gibbs (2010). Species in this subgenus are predominately ground-nesting and form primitively eusocial colonies (annual colonies with reproductive division of labour) (Michener 1974; Gibbs et al. 2012). There are some exceptions (species that do not form annual colonies) in this genus, which are solitary or communal species (Michener 1974; Packer 1994; Yanega 1997). Some species in this genus are wood-nesting (Stockhammer 1967; Gibbs 2011) although the lack of trees on Sable Island precludes the Sable Island Sweat Bee as a wood-nester.
Life cycle, demographic parameters and reproduction
Lasioglossum are often primitively eusocial bees that form small colonies; however, the colonial nature of this genus is different from that of bees such as bumble bees or honey bees. Some Lasioglossum are solitary and some live communally in groups, although this may partially be attributed to suitable nest sites within a given area.
In temperate regions Lasioglossum typically have an annual life cycle (Michener 1974). Inseminated females overwinter and emerge in spring to initiate new nests. In eusocial colonies the first brood is composed of non-reproductive females (workers). The overwintering female becomes a queen and is primarily responsible for reproduction of the colony. Reproductive males and females are produced later in the summer. In solitary species there is no non-reproductive worker brood.
There are four life stages: egg, larva, pupa and adult. Adult females of Lasioglossum form simple underground burrows with lateral cells (Sakagami and Michener 1962). Females provision cells with a mixture of nectar and pollen, which are formed into a ball-shape. Nest construction and foraging activities are taken over by workers in eusocial colonies. A single egg is placed in each cell. Eggs hatch and the developing larvae feed on the nectar and pollen mass, which will be the total food provided to the immature bee. Developmental time from egg to adult is typically 3-4 weeks, developing through 5 larval instars and 1-2 weeks spent as pupae (Yanega 1997; Michener 2007). During pupation the larvae undergo complete metamorphosis.
Formation of eusocial colonies is likely the ancestral state for Lasioglossum (Danforth et al. 2003; Gibbs et al. 2012). The nesting biology of the Sable Island Sweat Bee has not been studied, but it could possibly be social based on phylogenetic relationships. Colony sizes are typically small and range from 2-84 females at maximum size depending on species and colony (Michener 1974). The cool weather conditions on Sable Island likely preclude the formation of large colony sizes. Some halictid bees are socially polymorphic, reverting to solitary behaviour when climatic conditions are poor (Packer 1990; Field et al. 2010). Further study of the nesting biology is needed to determine its social status and what proportion of females collected on flowers are reproductive.
Physiology and adaptability
Flight activity of foraging bees is greatest in warm (> 20°C), sunny weather with low winds (Tuell and Isaacs 2010). There is no information on the minimum flight temperature for the Sable Island Sweat Bee. However, studies on Lasioglossum ruidosensis in Alaska indicate that these sweat bees could fly only when the environmental temperature (not precisely the same as ambient temperature) reached 17.5°C. In general, this species and other small sweat bees studied were considered poor thermoregulators (Bishop and Armbruster 1999). Suboptimal temperatures reduce foraging activity and potentially productivity. Females return to nests at night and are protected from inclement weather. Male bees typically sleep outside and are capable of surviving nighttime temperatures.
Bees of the genus Lasioglossum occupy a wide range of climatic regions. The Sable Island Sweat Bee probably has similar adaptations to other temperate or boreal zone Lasioglossum. Sable Island has relatively cool summers for southern Canada (Table 2). Climate data from 1971–2000 records show mean maximum temperatures in June (18.4°C), July (20.6°C) and August (18.4°C) (Table 2). These low average temperatures could limit bee foraging activity. Winter weather is relatively mild on Sable Island relative to other regions of Canada (except areas of the Pacific coast).
The Sable Island Sweat Bee appears to be a floral generalist, based on the collection of specimens foraging from six genera in five plant families. This variety of floral resources suggests the Sable Island Sweat Bee is flexible in its pollen and nectar requirements. Many Lasioglossum show similar floral generalization, and have been recorded visiting more than two dozen plant families (Gibbs 2010).
| Variable | April | May | June | July | August | September | October |
|---|---|---|---|---|---|---|---|
| Average Temperature (°C) | 3.8 | 7.2 | 11.2 | 15.7 | 17.8 | 15.7 | 11.4 |
| Maximum Temperature (°C) | 6.2 | 9.8 | 14 | 18.4 | 20.6 | 18.4 | 14.1 |
| Minimum Temperature (°C) | 1.2 | 4.6 | 8.4 | 12.9 | 14.9 | 12.9 | 8.8 |
| Extreme Maximum Temperature (°C) | 13.9 | 17.8 | 21.7 | 29.6 | 27.8 | 27 | 22.8 |
| Days with Extreme Minimum Temperature (°C) | -8.9 | -8.3 | 0.6 | 3 | 4.4 | 0.6 | -1.2 |
| Days with temperatures above 0°C | 30 | 31 | 30 | 31 | 31 | 30 | 31 |
| Days with temperatures above 10°C | 2.2 | 14.6 | 28.3 | 31 | 31 | 30 | 27.6 |
| Days with temperatures above 20°C | 0 | 0 | 0.17 | 6.3 | 18.6 | 6.9 | 0.2 |
| Days with temperatures above 30°C | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Average wind speed (km/h) | 26.2 | 21.8 | 19.8 | 17.9 | 17.5 | 21.2 | 25.8 |
Dispersal and migration
There is little information on natural dispersal rates for Lasioglossum bees. The Sable Island Sweat Bee likely has limited dispersal capabilities. These are flying insects, but typical flight distances for bees of this size are probably less than 1 km (Greenleaf et al. 2007). Some sweat bees are known to return to natal nesting areas in spring, which limits dispersal rates (Yanega 1990). Dispersal from Sable Island to the nearest mainland would require abiotic factors (e.g. wind) to travel the greater distance (> 150 km). The most frequent wind direction during adult life stages of the bee is from the southwest. This would blow in the direction of Newfoundland, a distance of approximately 400 km. Ground-nesting of the Sable Island Sweat Bee makes human-mediated dispersal unlikely. Most bees transported by humans are cavity-nesting species (Cane 2003).
Interspecific interactions
The Sable Island Sweat Bee is dependent on the pollen and nectar resources of numerous flowering plants found on Sable Island. The long phenology of Lasioglossum colonies requires the use of multiple flowering species. No individual plant is likely to be an obligate requirement for the Sable Island Sweat Bee, but a diverse flowering community is necessary to maintain populations of the bee. Even with limited net collections, floral visitation records come from six genera in five families (Asteraceae: Achillea, Hieracium, Ericaceae: Vaccinium, Onagraceae: Oenothera, Polygonaceae: Polygonum, Rosaceae: Rosa). Lasioglossum species are regularly collected from other plant families (Gibbs 2010), including the following families, which are known from the island: Apiaceae, Brassicaceae, Fabaceae, Lamiceae, and Ranunculaceae (Catling et al. 1986). A list of the Sable Island vegetation is available in Catling et al. (1986).
In spring, the bee fauna is predominately composed of the two Lasioglossum species, the Sable Island Sweat Bee and L. novascotiae, with lower numbers of two megachilid bees. Osmia simillima is the first megachilid bee to emerge. It declines in abundance as spring progresses, becoming uncommon during the early summer and is absent later in the year. Megachile melanophaea first occurs in spring, but does not occur in abundance until summer.
Although there is potential for competition for floral resources among other bees, the generalist nature of the Sable Island Sweat Bee allows for the possibility of some niche partitioning.
Population sizes and trends
There is no information on population sizes and trends for the Sable Island Sweat Bee. To date, surveys have been opportunistic collection events during visits to the island by biologists studying other natural history questions. Historical specimens of the Sable Island Sweat Bee were made from sweep netting vegetation as part of general collections (Wright 1989; Catling et al. 2009).
Sampling effort and methods
Specimens collected during 2013 surveys used standardized bowl trapping and hand netting. Bowl trapping bees is a passive collection method that involves the use of coloured bowls (Droege et al. 2010). Although bowl trapping can give a biased perspective of bee community (Cane et al. 2000), it is known to collect Lasioglossum in high numbers (Droege et al. 2010). Bees were collected on Sable Island using coloured bowl traps (blue, pink, yellow and green) filled with dish soap water. Traps were deployed at two sites, the west (43.93208, -60.02316) and east (43.95876, -59.78298) end lighthouses, on opposite ends of the island. Collections were made during June and August, 2013 (Table 1).
Both the east and west end sampling sites near the lighthouses were sparse grassy areas with sandy soil. Flowering plants at these sites were primarily Sorrel (Rumex spp.), Wild Strawberry (Fragaria spp.) and Large False Solomon’s Seal (Maianthemum racemosum) during June and Yarrow (Achillea), Pearly Everlasting (Anaphalis margaritacea) and Smooth Hawksbeard (Crepis capillaris) during August.
Additional collections, primarily near the Sable Island Station (Figure 7) in late June and August (Table 1) were completed using an insect net and yellow bowl traps. Net collections included focused, timed sampling from Rugosa Rose (Rosa rugosa) and Hieracium sp. Sweep net samples from vegetation and bowl trap samples were in areas dominated by either clover (Fabaceae) or areas which included blooming individuals of Cranberry (Ericaceae), Tuberous Grass-pink (Calopogon sp.), Yellow Loosestrife (Lysimachia spp.) and/or Northern Bayberry Myrica spp. Most net collections were made for brief (5 – 10 min.) periods. Floral records of the Sable Island Sweat Bee were made from Rugosa Rose and Hieracium, which add another genus and another plant family to the known associations of the species. More than two thirds of the Sable Island Sweat Bee specimens netted from vegetation were found on Hieracium sp. Yellow bowl traps placed in areas with an abundance of Hieracium did not collect specimens of the Sable Island Sweat Bee, and in areas and times where Megachile melanophaea was collected or observed in great abundance, the Sable Island Sweat Bee was infrequently collected.
Abundance
There are four bee species recorded from Sable Island and the Sable Island Sweat Bee is the least abundant bee recorded during surveys. In bowl traps M. melanophaea (304 individuals) is the most commonly collected (337 individuals) followed by L. novascotiae (277 individuals), then O. simillima (10 individuals) and the Sable Island Sweat Bee (5 individuals). However, Sable Island Sweat Bees may not be well sampled by bowl-trapping. The large difference in the numbers of the two Lasioglossum species found in the bowl traps (ratio of 55:1) is not paralleled in the net captures of L. novascotiae (64 individuals) and the Sable Island Sweat Bee (33 individuals) (ratio approximately 2:1). Net collections of historical specimens at the CNC of these two Lasioglossum found a similar ratio (3:1). Although sample sizes are small, these data suggest a stable proportion of these two species.
Fluctuations and trends
Results from sweep netting show comparable proportions of the two Lasioglossum species on the island. Twenty specimens of Lasioglossum collected from Sable Island in 1966 and 1967 were examined and found to be in a ratio of 3:1 (15 L. novascotiae and 5 Sable Island Sweat Bees). Seventeen specimens of Lasioglossum collected in 2008 were found to be in nearly equal proportions of the two species (8 Sable Island Sweat Bees and 9 L. novascotiae). Net collections made in 2013 found a similar proportion of individuals of these two species (2:1).
Comparison of the historical collections to bowl trap numbers gives a dramatically different picture because L. novascotiae is collected 55 times more frequently than the Sable Island Sweat Bee in bowls.
The low number of historical specimens of the Sable Island Sweat Bee makes it difficult to determine if the populations have changed over time. Habitat changes caused by anthropogenic actions (e.g., historical farming) and natural processes have influenced habitat availability, and the likely drivers for population fluctuations (see Habitat Trends and Threats).
Rescue effect
The Sable Island Sweat Bee is endemic to Sable Island and rescue effect is not possible.
Threats and limiting factors
The International Union for Conservation of Nature-Conservation Measures Partnership (2006) (IUCN-CMP) threats calculator was used to classify and list threats to the Sable Island Sweat Bee (Salafsky et al. 2008; Master et al. 2009).
Most threats to the Sable Island Sweat Bee are negligible (explained in Table 3), except for low threats from invasive and other problematic native species and from climate change and severe weather. Non-native horses on the island can cause substantial damage to the integrity of the bee’s habitat by increasing erosion and decreasing available forage plants. Increasing populations of native gray seals, which spend time on the island, could also have an impact on the bee. There are medium to low threats from periodic storms and flooding, which inundate the island, damaging the bee’s habitat. Multiple sites on the island are at risk of being breached by sea levels, which could have a significant impact on the structure of the vegetated surface and the bee’s habitat. Future eco-tourism may pose a future threat to bee habitat.
The threats below are discussed according to the IUCN-CMP threat classification number and are considered equally applicable. The overall Threat Impact for the Sable Island Sweat Bee is low (Table 3).
- Species or ecosystem scientific name:
- Sable Island Sweat Bee, Lasioglossum sablense
- Date:
- 2014-01-22; updated 2014-08-22
- Assessor(s):
- Jason Gibbs (status report author), Paul Catling (Agriculture and Agri-Food Canada), Heather Stewart (Parks Canada Agency), Cory Sheffield (Royal Saskatchewan Museum), Ruben Boles (Canadian Wildlife Service), Syd Cannings (Canadian Wildlife Service), Julie McKnight (Canadian Wildlife Service), Mary Sabine (New Brunswick), Bill Freedman (Dalhousie University), Jennifer Heron (Arthropods SSC Co-chair).
| Threat impact | Threat impact (descriptions) | Level 1 Threat impact counts: high range |
Level 1 Threat impact counts: low range |
|---|---|---|---|
| A | Very High | 0 | 0 |
| B | High | 0 | 0 |
| C | Medium | 0 | 0 |
| D | Low | 2 | 2 |
| - | Calculated overall threat impact: | Low | Low |
| # | Threat | Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | Negligible | Negligible (<1%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | There is a large footprint of existing buildings currently on the island; however, most of these are abandoned and in various stages of collapse and decay. Parks Canada plans to take inventory of these buildings and, in time, decommission those deemed to be a safety hazard (Stewart pers. comm. 2014). Soil compaction and trampling of floral resources and nest sites may impact the bee during decommissioning, if and when this occurs. |
| 1.1 | Housing & urban areas | Negligible | Negligible (<1%) | Negligible (<1%) | Low (Possibly in the long term, >10 yrs) | There aren't many buildings in use for housing on the island. Currently not applicable, this is considered a historic or past threat. Historically, small communities of people have lived on the island, but there is currently limited housing on the island. |
| 1.2 | Commercial & industrial areas | Negligible | Negligible (<1%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | There are several buildings on the island, which are used to store maintenance and research equipment and supplies, including fuel. Future development is unlikely given the island’s protection as a national park; however, future decommissioning of existing buildings or their replacement and/or upgrades with new structures could lead to increased disturbance in the vicinity of the structures. There are likely to be upgrading and construction of maintenance buildings on the island and the footprint will likely expand within the next ten years (Stewart pers. comm. 2014). |
| 1.3 | Tourism & recreation areas | Negligible | Negligible (<1%) | Slight (1-10%) | Low (Possibly in the long term, >10 yrs) | Sable Island was established as a National Park Reserve in June 2013. Until the development of a management plan for the Sable Island National Park Reserve, it is difficult to predict how expected increases in tourism will affect the bee and its habitat. It is not expected that there will be substantial infrastructure development to facilitate tourism. Marina and boat launch facilities are not likely to be constructed due to the shifting sands and soft substrate on the island. It is unlikely camping facilities or overnight housing for tourists will be constructed. |
| 2 | Agriculture & aquaculture | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/ Negligible (Past or no direct effect) |
- |
| 2.1 | Annual & perennial non-timber crops | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/ Negligible (Past or no direct effect) |
Currently not applicable, this is considered a historic or past threat. It is likely that habitat changes on the island from anthropogenic disturbance since the 1500s have had a negative impact on Sable Island Sweat Bee nesting and foraging habitat. Historically, large areas of vegetated habitat on the island were lost during periods of intensive agriculture (see Catling et al. 1984). |
| 2.2 | Wood & pulp plantations | - | - | - | - | Not applicable. |
| 2.3 | Livestock farming & ranching | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/ Negligible (Past or no direct effect) |
Currently not applicable and considered a historic threat. Various livestock including cattle, horses and pigs, were introduced at various times during human settlement on Sable Island. Today, only feral horses remain on the island. The presence of feral horses on the island likely increases erosion and reduces floral diversity through trampling, soil compaction, urination and defecation. These cumulative effects likely impact bee foraging sites and nesting sites and ultimately negatively impact bee populations. The horses are considered naturalized; they are not fed or cared for nor is there any other form of intervention in population management. It is possible there may be some stewardship measures considered in the future, but at present there are none. This threat is categorized under Threat 8.1. |
| 2.4 | Marine & freshwater aquaculture | - | - | - | - | Not applicable. |
| 3 | Energy production & mining | Not a Threat (in the assessed timeframe) |
Small (1-10%) | Moderate (11-30%) | Low (Possibly in the long term, >10 yrs) | - |
| 3.1 | Oil & gas drilling | Not a Threat (in the assessed timeframe) |
Small (1-10%) | Moderate (11-30%) | Low (Possibly in the long term, >10 yrs) | The Sable Offshore Energy Project includes a central processing facility near Sable Island. Current regulations prevent oil or gas drilling within one nautical mile of Sable Island. The current nearest drilling facility is 12 km from the island. Drilling facilities occur to the east, south and west of the island (Lucas et al. 2012). The Offshore Accord Implementation Act takes precedent over the Canada National Parks Act so future energy production on or near the island is possible. Subsurface horizontal drilling beneath the island is permitted although not currently underway. It is unclear what effect subsurface drilling would have on the island, but any disturbance or collapse of material beneath the island could be devastating if it causes the low-lying island to sink. A catastrophic release of liquid gas from a nearby oil facility could potentially damage the island’s ecosystem. Local ocean currents result in a counterclockwise gyre, which occasionally deposits floating material, including oil or oil-contaminated objects on Sable Island (Hannah et al. 2001; Lucas et al. 2012). The frequency of oil contaminated birds reaching the island has decreased in recent years, which may be a product of increased regulation and enforcement (Lucas et al. 2012). These floating objects may act as attractants to bees, but there are no studies to confirm if Sable Island Sweat Bee lands on these objects. |
| 3.2 | Mining & quarrying | - | - | - | - | Not applicable. |
| 3.3 | Renewable energy | Not a Threat | Negligible (<1%) | Neutral or Potential Benefit | Insignificant/ Negligible (Past or no direct effect) |
There is small-scale solar and wind energy generation on the island for local use only. Although there may be replacement of equipment in the future, it is not expected the footprint of these structures will increase in the future. |
| 4 | Transportation & service corridors | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | - |
| 4.1 | Roads & railroads | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Vehicular transport on the island is limited to park administration and researchers. There are currently 33 vehicles on the island, including but not limited to: all-terrain vehicles, service vehicles for plane traffic, a truck, and a tractor. Vehicle traffic is restricted to barren beaches, limited roads near the island station and other predefined routes on the island. Designated vehicle traffic areas on the island are not severely compacted and are not likely to impact bee nesting. Driving on vegetation and dune slopes is prohibited, which should limit direct impact on floral hosts and nesting sites of Sable Island Sweat Bee. This threat is considered negligible. |
| 4.2 | Utility & service lines | Negligible | Negligible (<1%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | There are existing electrical communication lines running underground on the island. Some of these cables and lines are linked to live lines and may be active. Future hazard and electrical activity assessments are planned. If deemed a physical or electrical safety hazard, then these lines will be removed. Removal would likely result in minimal invasive methods and involve small-scale digging with small machinery or by hand. This could have small impacts on bee nests, but is not considered a significant threat. |
| 4.3 | Shipping lanes | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Passing ships could lead to oil discharge, which due to ocean currents could reach the island in small amounts. The occurrence of oil slicks or tar particles reaching the island is infrequent and was not recorded during a four-year survey from 2006 to 2009 (Lucas et al. 2012). |
| 4.4 | Flight paths | - | - | - | - | Not applicable. |
| 5 | Biological resource use | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | - |
| 5.1 | Hunting & collecting terrestrial animals | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | This is limited to permitted research activities on the island. General insect collections on the island could potentially impact the population size if too many specimens are taken. To date the total number of Sable Island Sweat Bee collected is quite low, but future insect collections particularly using passive sampling methods such as bowl traps and Malaise traps could potentially kill large numbers of individuals in a short period. Malaise trap-use is limited to certain fenced areas to protect feral horses. Illegal insect or flower collections by visitors on the island would negatively impact Sable Island Sweat Bee, but these activities should be minimized by the limited traffic on the island. Bycatch from other research activities (non-entomologically related) would also consider impacts to the bee and its habitat. |
| 5.2 | Gathering terrestrial plants | - | - | - | - | Not applicable. |
| 5.3 | Logging & wood harvesting | - | - | - | - | Not applicable. |
| 5.4 | Fishing & harvesting aquatic resources | - | - | - | - | Not applicable. |
| 6 | Human intrusions & disturbance | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | - |
| 6.1 | Recreational activities | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Sable Island is protected under the Canada National Parks Act. Direct human impacts on the island are limited by its national park status (see Habitat Protection and Ownership) and its isolation from the mainland. Sable Island has a limited number of regular occupants and research activities. It is occasionally visited by tourists, either by chartered flight or cruise ship. Many activities are prohibited on the island, but human foot traffic can potentially increase erosion on dune slopes and vegetated areas and ultimately damage sensitive bee habitat. It is expected that tourism on the island will increase with its establishment as a National Park. There are plans for tourist cruise ships to visit the island. Each ship could bring 100 or more visitors to the island, which represents an increase over recent tourism. Visitors are monitored and given an orientation to the island by park staff, which should further limit impacts of increased tourism. Increased visitation will increase the probability of introduction of invasive species. |
| 6.2 | War, civil unrest & military exercises | - | - | - | - | Not Applicable. |
| 6.3 | Work & other activities | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | There may be limited impact resulting from research activities on the island related to vehicle travel and transects for monitoring wildlife. By-catch of bees as part of non-specific trapping methods could occur. These are considered minor or insignificant threats to the bee. |
| 7 | Natural system modifications | Negligible | Negligible (<1%) | Negligible (<1%) | Low (Possibly in the long term, >10 yrs) | - |
| 7.1 | Fire & fire suppression | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/ Negligible (Past or no direct effect) |
Fire is not a component of the island’s natural ecosystem. There are no fire suppression activities. There is a minor threat of accidental fire near buildings, but this is unlikely to impact the bees or their host vegetation. |
| 7.2 | Dams & water management/use | - | - | - | - | There is not yet a management plan developed for the park. Some water is drawn for use in structures and waste water is released into a septic system (see Threat 9.1). There may be some future assessment of waste water, but it is not expected to impact bees. |
| 7.3 | Other ecosystem modifications | Negligible | Negligible (<1%) | Negligible (<1%) | Low (Possibly in the long term, >10 yrs) | There are likely to be future plans and recommendations for ecosystem restoration or enhancement on the island. These may include modifications of existing vehicle routes to decrease erosion, by making these routes sinuous rather than the current straight-line routes. In general, these ecosystem modifications are not likely to impact bees, and modifications that prevent erosion of vegetated areas may benefit Sable Island Sweat Bee habitat. |
| 8 | Invasive & other problematic species & genes | Low | Pervasive - Restricted (11-100%) | Slight (1-10%) | High (Continuing) | - |
| 8.1 | Invasive non-native/alien species | Low | Pervasive (71-100%) | Slight (1-10%) | -High (Continuing) | Sable Island has a resident population of naturalized horses. The original source population of these horses is unknown; however, horses have been on the island for at least 500 years. The impact of the feral horse population on Sable Island Sweat Bee is unclear and requires additional study. Trampling of vegetation or nest sites by the horses is potentially damaging either directly or through increased erosion (Freedman et al. 2011). Horses preferentially consume Marram grass (Ammophila breviligulata), which is the primary plant sand-binding species responsible for preventing erosion. This large herbivore has the potential to also consume valuable floral hosts of the bee. However, feral horses may also help structure the sand ecosystem, allowing forbs to compete with grasses on the island and enable the open sand ecosystems to remain as such. Since the bees and most plants likely preceded the presence of the horses by several thousand years this possible benefit may not be required. Population sizes vary from 150 to 400 to horses (Lucas et al. 1991) so there may significant year-to-year variation in the impact of the horses. Recent estimates put the horse population near its maximum (Lucas et al. 2009). Additional research on the effect of horses on Sable Island Sweat Bee and its floral hosts using horse enclosures are needed (Freedman et al. 2011). If other bees were introduced to the island then they could potentially impact Sable Island Sweat Bee through competition or introduced disease. In the past, bees have been intentionally and unintentionally transported during human activities, but this has primarily been cavity-nesting species (Cane 2003). Nest sites for cavity-nesting bees would be limited to areas near buildings or other structures, which would minimize the impact and likelihood of establishment of these bees. Transportation of ground-nesting species is unlikely since it would require movement of large amounts of soil. The broad floral host range of Sable Island Sweat Bee also may help it escape competition from other species. Other invasive insects, such as ants or bee-attacking insects, could potentially impact Sable Island Sweat Bee. Additional surveys of the insect fauna to monitor any changes are needed. The last extensive faunal list for Sable Island was published by Wright (1989), and much of this was based on earlier literature (e.g. Howden et al. 1970). Introduction of an invasive plant species to the island could be problematic to Sable Island Sweat Bee and its habitat. These could be transported through numerous pathways, including seeds being moved in clothing and shoes of tourists. The harsh conditions of the island (e.g. cool summer climate, high sand content, and heavy inundation of salt water) is likely to limit the number of invasive species that could successfully establish themselves on the island. Nevertheless, any invasive plant that interferes with the natural ecosystem of the island could be potentially harmful to Sable Island Sweat Bee. Some of the plant species on the island are non-native (Catling et al. 1984) and some such as Calluna vulgaris (L.) (ERICACEAE) Hull may be increasing in population size. |
| 8.2 | Problematic native species | Low | Restricted (11-30%) | Slight (1-10%) | High (Continuing) | Grey seals (Halichoerus grypus) on the island have the potential to damage vegetated areas during the winter months when they come inland. Grey seal populations increased exponentially over several decades during the late 20th century (Bowen et al. 2003), although this population growth has declined (Bowen et al. 2007). During winter months, the seals move inland and occupy the lowland terrestrial areas of the island, although it is unknown how this impacts the bee overwintering sites and dormant floral resources. |
| 8.3 | Introduced genetic material | - | - | - | - | Not applicable. |
| 9 | Pollution | Negligible | Small (1-10%) | Unknown | High (Continuing) | - |
| 9.1 | Household sewage & urban waste water | Not a Threat | Negligible (<1%) | Neutral or Potential Benefit | High (Continuing) | There are septic systems in place on the island for waste water and sewage. These are unlikely to directly impact Sable Island Sweat Bee. |
| 9.2 | Industrial & military effluents | - | Small (1-10%) | Unknown | High (Continuing) | There may be potential impacts from shipping lanes and offshore oil rigging (see above.) |
| 9.3 | Agricultural & forestry effluents | Not a Threat (in the assessed timeframe) | Unknown | Unknown | Insignificant/ Negligible (Past or no direct effect) |
These are prohibited due to the island’s status as a national park. Historical use of chemicals is uncertain. |
| 9.4 | Garbage & solid waste | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | There is garbage from past human habitation on the island. Garbage includes oil drums, containers, and structural remains from buildings, farming equipment, and other unknown materials. Most garbage is inert and unlikely to impact bee populations. Some chemical leaching occurs, including mercury from old light bulbs, in the vicinity of the lighthouses and oil from various sources (see above). These likely impact the sand ecosystem in the vicinity of leaching and may have minor impacts on Sable Island Sweat Bee. |
| 9.5 | Air-borne pollutants | - | - | - | - | There are occasional aerosol pollutants, which reach the island from offshore drilling facilities. The impact of these is uncertain. |
| 9.6 | Excess energy | - | - | - | - | Not applicable. |
| 10 | Geological events | - | Small (1-10%) | Serious (31-70%) | Unknown | - |
| 10.1 | Volcanoes | - | - | - | - | Not applicable. |
| 10.2 | Earthquakes/ tsunamis | Low | Small (1-10%) | Serious (31-70%) | Unknown | Tsunamis are uncommon in the Atlantic Ocean, and are an unlikely threat in the short term. In the event of a tsunami there would likely be significant damage to floral vegetation, erosion of habitat, and potentially damaging submersion of nests. |
| 10.3 | Avalanches/landslides | - | - | - | - | Not applicable. |
| 11 | Climate change & severe weather | Medium-low | Large-restricted (11-70%) | Moderate (11-30%) | High (Continuing) | - |
| 11.1 | Habitat shifting & alteration | Not a Threat (in the assessed timeframe) |
Small (1-10%) | Moderate - Slight (1-30%) | Low (Possibly in the long term, >10 yrs) | Since the last ice age, rising sea levels have caused a gradual decrease in the size of Sable Island (Cameron 1965; Wright 1989). The dune systems of the island may be more unstable in recent times than they were prior to human colonization (McCann and Byrne 1994). Estimates of the island’s size over the last two and a half centuries suggests a loss of as much as 35 km2 (Wright 1989). The potential range of the bee would have been reduced greatly over this time period. Climate change is likely to increase sea level rises in the future (Houghton et al.1996; Shaw 2001; Kont et al. 2003; Environment Canada 2006). The island has a low-elevation and sea level rises will shrink the island and reduce available habitat. Global estimates of sea level rises are as great as 120 cm by 2100 (Rahmstorf 2007). In the 20th century sea levels rose by approximately 17 cm, but the rate or increase has nearly doubled in recent years (Church and White 2006). The island is continually shifting as wave action reduces material from one end of the island and replaces it on the opposite side of the island. This is part of the natural process of the island. Increasing inundation of ponds on the island by sand and sea water could lead to a loss of floral species such as Polygonum in these regions. |
| 11.2 | Droughts | - | - | - | - | - |
| 11.3 | Temperature extremes | Not a Threat (in the assessed timeframe) |
Unknown | Unknown | Low (Possibly in the long term, >10 yrs) | The cool climate of Sable Island (Table 2) likely limits the productivity of bees, which prefer warm temperatures (Tuell and Isaacs 2010). Extreme cold temperatures could potentially impact Sable Island Sweat Bee directly or indirectly by damaging floral hosts. Climate models for the Maritime regions suggest that the climate will become even cooler and wetter, which may put pressure on the physiological tolerances of bees on the island. |
| 11.4 | Storms & flooding | Medium - Low | Large - Restricted (11-70%) | Moderate - Slight (1-30%) | High (Continuing) | Sable Island regularly experiences strong winds (Table 2) and is subject to storms and flooding. Climate change is expected to increase storm frequency and severity in the region (Keim et al. 2004). Extreme weather conditions increase the rate of erosion and loss of vegetated areas. The last hurricane resulted in the largest documented flooding of the habitat by sea water. The effect of this flooding on vegetation or the impact of submersion of Sable Island Sweat Bee nests requires study. There are at least three areas on the island that are at risk of being breached if sea levels and flooding continue. Breaching would result in separation of Sable Island into multiple land masses and could potentially significantly reduce the amount of available habitat for the bee. |
Invasive and other problematic species and genes
Invasive non-native/alien species (8.1)
Sable Island has a resident population of naturalized horses. The current population of horses may have been introduced in approximately 1760 (Christie 1995). The impact of the feral horse population on Sable Island Sweat Bee is unclear and requires additional study. Trampling of vegetation or nest sites by the horses is potentially damaging either directly or through increased erosion (Freedman et al. 2011). Horses preferentially consume Marram Grass (Ammophila breviligulata), which is the primary plant sand-binding species responsible for preventing erosion. This large herbivore has the potential to also consume valuable floral hosts of the bee. However, feral horses may also help structure the sand ecosystem, allowing forbs to compete with grasses on the island and enable the open sand ecosystems to remain as such. Since the bees and most plants likely precede the presence of the horses by several thousand years this possible benefit may not be required. Population sizes vary from 150 to 400 to horses (Lucas et al. 1991) so there may be significant year-to-year variation in the impact of the horses. Horses have not been culled since 1960 (Plante et al. 2007) so their populations may be on the rise. Recent estimates put the horse population near its maximum (Lucas et al. 2009). Additional research on the effect of horses on the Sable Island Sweat Bee and its floral hosts using horse exclosures is needed (Freedman et al. 2011).
If other bees were introduced to the island then they could potentially impact the Sable Island Sweat Bee through competition or introduced disease. In the past, bees have been intentionally and unintentionally transported during human activities, but this has primarily been cavity-nesting species (Cane 2003). Nest sites for cavity-nesting bees would be limited to areas near buildings or other structures, which would minimize the impact and likelihood of establishment of these bees. The megachilid bees on the island are both ground-nesting species although cavity-nesting is common in this family, which may be indicative of the few nesting opportunities for cavity-nesting bees. Transportation of ground-nesting species is unlikely since it would require movement of large amounts of soil. Nevertheless, there are cases of ground-nesting bees becoming established outside their native range (e.g. Magnacca et al. 2013). The broad floral host range of the Sable Island Sweat Bee also may help it escape competition from other species.
Other invasive insects, such as ants or bee-attacking insects, could potentially impact the Sable Island Sweat Bee. Additional surveys of the insect fauna to monitor any changes are needed. The last extensive faunal list for Sable Island was published by Wright (1989), and much of this was based on earlier literature (e.g. Howden et al. 1970).
Introduction of an invasive plant species to the island could be problematic to the Sable Island Sweat Bee and its habitat. These could be transported through numerous pathways, including seeds being moved in clothing and shoes of tourists. The harsh conditions of the island (e.g. cool summer climate, high sand content, and heavy inundation of salt water) is likely to limit the number of invasive species that could successfully establish themselves on the island. Nevertheless, any invasive plant that interferes with the natural ecosystem of the island could be potentially harmful to the Sable Island Sweat Bee. Some of the plant species on the island are non-native (Catling et al. 1984) and some such as Scottish Heather - Calluna vulgaris (L.) (Ericaceae) Hull may be increasing in population size.
Problematic native species (8.2)
Grey seals (Halichoerus grypus) on the island have the potential to damage vegetated areas during the winter months when they come inland. Grey seal populations increased exponentially over several decades during the late 20th century (Bowen et al. 2003), although this population growth has declined (Bowen et al. 2007). During winter months, the seals move inland and occupy the lowland terrestrial areas of the island, although it is unknown how this impacts the bee overwintering sites and dormant floral resources.
Climate change and severe weather
Habitat shifting and alteration (11.1)
Since the last ice age, rising sea levels have caused a gradual decrease in the size of Sable Island (Cameron 1965; Wright 1989). The dune systems of the island may be more unstable in recent times than they were prior to human colonization (McCann and Byrne 1994). Estimates of the island’s size over the last two and a half centuries suggests a loss of as much as 35 km2 (Wright 1989). The potential range of the bee would have been reduced greatly over this time period.
Climate change is likely to increase sea level rises in the future (Houghton et al.1996; Shaw 2001; Kont et al. 2003; Environment Canada 2006). The island has a low elevation and sea level rises will shrink the island and reduce available habitat. Global estimates of sea level rises are as great as 120 cm by 2100 (Rahmstorf 2007). In the 20th century sea levels rose by approximately 17 cm, but the rate or increase has nearly doubled in recent years (Church and White 2006).
The island is continually shifting as wave action reduces material from one end of the island and replaces it on the opposite side of the island. This is part of the natural process of the island. Increasing inundation of ponds on the island by sand and sea water could lead to a loss of floral species such as Polygonum in these regions.
Storms and flooding (11.4)
Sable Island regularly experiences strong winds (Table 2) and is subject to storms and flooding. Climate change is expected to increase storm frequency and severity in the region (Keim et al. 2004). Extreme weather conditions increase the rate of erosion and loss of vegetated areas. The last hurricane resulted in the largest documented flooding of the habitat by sea water. The effect of this flooding on vegetation or the impact of submersion of the Sable Island Sweat Bee nests requires study.
There are at least three areas on the island that are at risk of being breached if sea levels and flooding continue. Breaching would result in separation of Sable Island into multiple land masses and could potentially significantly reduce the amount of available habitat for the bee.
Human intrusions and disturbance
Recreational activities (6.1)
Sable Island is protected under the Canada National Parks Act. Direct human impacts on the island are limited by its national park status (see Habitat Protection and Ownership) and its isolation from the mainland. Sable Island has limited number of regular occupants and research activities. It is occasionally visited by tourists, either by chartered flight or cruise ship. Many activities are prohibited on the island, but human foot traffic can potentially increase erosion on dune slopes and vegetated areas and ultimately damage sensitive bee habitat. It is expected that tourism on the island will increase with its establishment as a National Park. There are plans for tourist cruise ships to visit the island. Each ship could bring 100 or more visitors to the island, which represents an increase over recent tourism. Visitors are monitored and given an orientation to the island by park staff, which should further limit impacts of increased tourism. Increased visitation will increase the probability of introduction of invasive species.
Geological events (IUCN Threats 10)
Earthquakes/tsunamis (10.2)
Tsunamis are uncommon in the Atlantic Ocean, and are an unlikely threat in the short term (Clague et al. 2003), but their occurrence is unpredictable. An earthquake or landslide on the Laurentian Shelf could cause a tsunami with waves that could reach Sable Island (Fine et al. 2005). In the event of a tsunami there would likely be significant damage to floral vegetation, erosion of habitat, and potentially damaging submersion of nests.
Limiting factors
Harsh weather could potentially limit foraging activity of the Sable Island Sweat Bee. Typical weather conditions during summer on Sable Island are often relatively cool and foggy, which is suboptimal for most bees. Wind speeds average nearly 25 km/h, which is above what is considered optimal for bee foraging (Tuell and Isaacs, 2010). Year-to-year variation in weather patterns could affect population levels due to decreased foraging activity in extreme conditions. Strong winds can also lead to increased erosion levels and the burying of flowering vegetation under sand.
Number of locations
Sable Island Sweat Bee is considered to have fewer than five locations on the isolated Sable Island. The most likely threat to this location is the inundation of the island due to severe weather or rising sea levels. The low elevation of the island means that rising sea levels expected due to climatic changes could greatly decrease the amount of available habitat. Severe storms, including tsunamis caused by earthquakes on the Laurentian Shelf, can potentially inundate the island with sea water. Frequent and continued submersion of underground nests by seawater could potentially have severe impacts on the Sable Island Sweat Bee.
Protection, status and ranks
Legal protection and status
There are no federal or provincial laws that specifically protect the Sable Island Sweat Bee, mitigate threats to this group or protect the species’ nest sites or habitat.
Non-legal satus and ranks
The Sable Island Sweat Bee has not been assigned a global status rank (NatureServe 2014). Provisional conservation status ranks for this species using NatureServe (2014) methodology recommend a global rank of G1 (globally critically imperiled), national rank of N1 (nationally critically imperiled) and provincial rank of S1 (critically imperiled).
Habitat protection and ownership
The global and Canadian range of the Sable Island Sweat Bee is entirely within the Sable Island National Park Reserve, which was established in July 2013. Parks Canada will develop a Management Plan for Sable Island National Park Reserve within five years following legal designation under the Canada National Parks Act. Interfering with the native flora and fauna is prohibited. Vehicular travel is limited to researchers and park staff and is restricted to certain areas. Various other activities are prohibited such as walking on steep dunes, unstable vegetation, fires, pets, littering and camping. In practice, some prohibited activities (e.g. walking on dune slopes) would be difficult to enforce due to the limited number of park staff.
Direct human impacts on the island are limited by its national park status and its isolation from the mainland. The island is approximately 280 km from Halifax. Visitation to the island requires registration, but is not strictly controlled. Visits to the island are primarily through chartered flights, cruise ships and private boat tours. The expense of flights and the frequent interference of extreme weather conditions tend to limit travel to the island. The recent conversion to a national park reserve means access to the island is now under the control of a different agency. It is possible that its new status as a park may lead to increased visitor traffic due to ecotourism. Should Parks Canada seek to accommodate tourism on the island through construction of boardwalks or other facilities it could have negative impacts on nesting sites.
Acknowledgements and authorities contacted
The efforts of Rob Ronconi (Acadia University) and Zoe Lucas (Greenhorse Society) in sampling Sable Island for bees are greatly appreciated. Sophie Cardinal (CNC) prepared a loan of historical material for this study. John Klymko (Atlantic Canada Conservation Data Centre) and Darien Ure (Parks Canada) helped with contact information of potential Sable Island researchers. Chris McCarthy (Parks Canada) assisted with collection permits. Paul Catling (Agriculture and Agri-food Canada) provided details about previous collections of the Sable Island Sweat Bee. Productive discussions of potential threats to the Sable Island Sweat Bee involved Jenny Heron (COSEWIC), Syd Cannings (Canadian Wildlife Service), Paul Catling, Ruben Boles (Canadian Wildlife Service), Bill Freedman (Dalhousie), Cory Sheffield (Royal Saskatchewan Museum) and Heather Stewart (Parks Canada). Jenny Heron and Angèle Cyr (COSEWIC Secretariat) provided invaluable assistance in the preparation of the field and status reports.
Authorities contacted
Ruben Boles, Biologist, Canadian Wildlife Service, Gatineau, QC
Syd Cannings, Species at Risk Biologist, Canadian Wildlife Service, Whitehorse, YK
Sophie Cardinal, Research Scientist, Agriculture and Agri-Food Canada, Canadian National Collection of Insects, Arachnids and Nematodes, Ottawa, ON
Paul Catling, Research Scientist, Agriculture and Agri-Food Canada, Ottawa, ON
Bill Freedman, Professor, Dalhousie University, Halifax, NS
Andrew Hebda, Curator of Zoology, Nova Scotia Museum, Halifax, NS
Rémi Hébert, Scientific Project Coordinator, Canadian Wildlife Service, Gatineau, QC
Steve Javorek, Research Biologist, Agriculture and Agri-Food Canada, Kentville, NS
John Klymko Atlantic Canada Conservation Data Centre, Sackville, NB
Zoe Lucas, Greenhorse Society, NS
Chris McCarthy, Ecologist Team Leader, Parks Canada, Maitland Bridge, NS
Laurence Packer, Professor, York University, Toronto, ON
Rob Ronconi, Acadia University, Wolfville, NS
Cory Sheffield, Curator of Invertebrate Biology, Royal Saskatchewan Museum, Regina, SK
Heather Stewart, Ecologist Team Leader, Parks Canada, NS
Sue Westby, Entomology Technician (ret.), Agriculture and Agri-Food Canada, Halifax, NS
Darien Ure, Ecologist, Parks Canada, Caledonia, NS
Information sources
Ascher, J.S. and J. Pickering. 2014. Discover Life bee species guide and world checklist (Hymenoptera: Apoidea: Anthophila). Draft 40. 26 July 2014.
Batra, S.W.T. 1984 Solitary bees. Scientific American 250:120–127.
Bishop, J.A., and W.S. Armbruster. 1999. Thermoregulatory abilities of Alaskan bees: effects of size, phylogeny and ecology. Functional Ecology 13:711-724.
Bowen, W.D., J.I. McMillan, and R. Mohn. Reduced population growth of gray seals at Sable Island: evidence from pup production and age of primiparity. Marine Mammal Science 23(1):48–64.
Bowen, W.D., J.I. McMillan, and W. Blanchard. 2007. Sustained exponential population growth of the grey seal on Sable Island. ICES. Journal of Marine Sciences 60:1265–1374.
Cameron, H.L. 1965. The shifting sands of Sable Island. The Geographical Review 55(4):463–476.
Cane, J.H. 2003. Exotic nonsocial bees (Hymenoptera: Apiformes) in North America: ecological implications. Pp. 113–126. in K. Strickler and J.H. Cane (eds.). For Non-native Crops, Whence Pollinators of the Future? Thomas Say Publications in Entomology: Proceedings, Entomological Society of America, Lanham, MD.
Cane, J.H., R.L. Minckley, and L.J. Kervin. 2000. Sampling bees (Hymenoptera: Apiformes) for pollinator community studies: pitfalls of pan-trapping. Journal of the Kansas Entomological Society 73(4):225–231.
Catling, P.M., B. Freedman, and Z. Lucas. 1984. The vegetation and phytogeography of Sable Island, Nova Scotia. Proceedings of the Nova Scotian Institute of Science 34:181–245.
Catling P., Z. Lucas, and B. Freedman. 2009. Plants and insects new to Sable Island, Nova Scotia. The Canadian Field-Naturalist 123:141–145.
Church, J.A. and N.J. White. 2006. A 20th century acceleration in global sea level rise, Geophysical Research Letters, 33, L01602, doi:10.1029/2005GL024826.
Christie, B.J. 1995. The horses of Sable Island. 2nd ed. Pottersfield Press, Porters Lake, NS, pp. 111.
Clague, J.J., A. Munro, and T.S. Murty. 2003. Tsunami hazard and risk in Canada. Natural Hazards 28:433–461.
Danforth, B.N., L. Conway, and S. Ji. 2003. Phylogeny of eusocial Lasioglossum reveals multiple losses of eusociality within a primitively eusocial clade of bees (Hymenoptera: Halictidae). Systematic Biology 52:23–36.
Droege, S., V.J. Tepedino, G. Lebuhn, W. Link, R.L. Minckley, Q. Chen, and C. Conrad. 2010. Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conservation and Diversity 3(1):15–23.
Environment Canada. 1998. Towards a conservation strategy for Sable Island. Environment Canada, Canadian Wildlife Service, Atlantic Region. 45 pages.
Environment Canada. 2006. The impacts of sea level rise and climate change on the coastal zone of southeastern New Brunswick. (Daigle, R., project lead).
Environment Canada. 611 pp.
Field, J., R.J. Paxton, A. Soro, and C. Bridge. 2010. Cryptic plasticity underlies a major evolutionary transition. Current Biology 20:2028–2031.
Fine, I.V., A.B. Rabinovich, B.D. Bornhold, R.E. Thomson, and E.A. Kulikov. 2005. The Grand Banks landslide-generated tsunami of November 18, 1929: preliminary analysis and numerical modeling. Marine Geology 215:45–57.
Freedman, B., P.M. Catling, and Z. Lucas. 2011. Effects of feral horses on vegetation of Sable Island, Nova Scotia. The Canadian Field-Naturalist 125(3):200-212.
Gibbs, J. 2010. Revision of the metallic Lasioglossum (Dialictus) of Canada (Hymenoptera, Halictidae, Halictini). Zootaxa 2591:1–382.
Gibbs, J. 2011. Revision of the metallic Lasioglossum (Dialictus) of eastern North America (Hymenoptera: Halictidae: Halictini). Zootaxa 3073:1–216.
Gibbs, J. and C.S. Sheffield. 2009. Rapid range expansion of the Wool-Carder Bee, Anthidium manicatum (Linnaeus) (Hymenoptera: Megachilidae), in North America. Journal of the Kansas Entomological Society 82(1):21–29.
Gibbs, J., S. Brady, K. Kanda, and B.N. Danforth. 2012. Phylogeny of halictine bees supports a shared origin of eusociality for Halictus and Lasioglossum (Apoidea: Anthophila: Halictidae). Molecular Phylogenetics and Evolution 65:926–939.
Greenleaf, S.S., N.M. Williams, R. Winfree, and C. Kremen. 2007. Bee foraging ranges and their relationship to body size. Oecologia 153:589–596.
Hannah, C.G., J.A. Shore, J.W. Loder, and C.E. Naimie. 2001. Seasonal circulation on the Eastern and Western Scotian Shelf. Journal of Physical Oceanography 31:591–615.
Houghton, J.T., et al., eds., 1996. Climate Change 1995--the Science of Climate Change, Cambridge University Press, Cambridge, U.K., Chap. 7, Changes in Sea Level, pp. 359–405.
Howden H.F., J.E.H., Martin, E.L Bousfield, and D.E. McAllister. 1970. Fauna of Sable Island and its zoogeographic affinities – a compendium. National Museum of Natural Sciences Publications in Zoology, No. 4. Queen’s Printer for Canada, Ottawa, pp. vi + 45.
Hubert, R. 2014. Personal communication to J. Gibbs. Canadian Wildlife Service, Environment Canada, Gatineau, QC.
Keim, B.D., R.A. Muller, and G.W. Stone. 2004. Spatial and temporal variability of coastal storms in the North Atlantic Basin. Marine Geology 210:7–15.
King, E.L. 2001. A glacial origin for Sable Island: ice and sea-level fluctuations from seismic stratigraphy on Sable Island Bank, Scotian Shelf, offshore Nova Scotia. Geological Survey of Canada, Current Research 2001-D19, 18 p.
King, L.H. and G.B.J. Fader. 1986. Wisconsinan glaciation of the continental shelf-southeast Atlantic Canada. Geological Survey of Canada, Bulletin 363, 72 p.
Kont, A., J. Jaagus, and R. Aunap. 2003. Climate change scenarios and the effect of sea-level rise for Estonia. Global and Planetary Change 36:1–15.
Lucas, Z., J.R. Raeside, and K.G. Betteridge. 1991. Non-invasive assessment of the incidences of pregnancy and pregnancy loss in the feral horses of Sable Island. Journal of Reproductive Fertility 44(Suppl.):479–488.
Lucas, Z.L., P.D. McLoughlin, D.W. Coltman, and C. Barber. 2009 Multiscale analysis reveals restricted gene flow and a linear gradient in heterozygosity for an island population of feral horses. Canadian Journal of Zoology 87:310–316.
Lucas, Z., A. Horn, and B. Freedman. 2012. Beached bird surveys on Sable Island, Nova Scotia, 1993 to 2009, show a decline in the incidence of oiling. Proceedings of the Nova Scotian Institute of Science 47(1):91–129.
Magnacca, K.N, J. Gibbs, and S. Droege. 2012. Notes on alien and native bees (Hymenoptera: Apoidea) from the Hawaiian Islands. Bishop Museum Occasional Papers 114: 61–65.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay, and A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk. NatureServe, Arlington, Virginia, USA. Web site: http://www.NatureServe.org/publications/ConsStatusAssess_StatusFactors.pdf [accessed January 15, 2014].
McCann, S.B. and M.L. Byrne. 1994. Dune morphology and the evolution of Sable Island, Nova Scotia, in historic times. Physical Geography 15(4):342–357.
Michener, C.D. 1953. Comparative morphological and systematic studies of bee larvae with a key to the families of hymenopterous larvae. University of Kansas Science Bulletin 35:987–1102.
Michener, C.D. 1974. The Social Behavior of the Bees. Belknap Press, Cambridge, Massachusetts. xii + 404 pp.
Michener, C.D. 2007. The Bees of the World, 2nd Ed. Johns Hopkins University Press, Baltimore, Maryland. xvi + [1] + 953 pp.
NatureServe. 2014. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. Web site: [accessed January 15, 2014].
Packer, L., 1990. Solitary and eusocial nests in a population of Augochlorella striata (Provancher) (Hymenoptera; Halictidae) at the northern edge of its range. Behavioral Ecology and Sociobiology 27: 339–344.
Packer, L. 1994. Lasioglossum (Dialictus) tenax (Sandhouse) (Hymenoptera; Halictidae) as a solitary sweat bee. Insectes Sociaux 41:309–313.
Plante Y., J.L. Vega-Pla, Z. Lucas, D. Colling, B. de March, and F. Buchanan. 2007. Genetic diversity in a feral horse population from Sable Island, Canada. Journal of Heredity 98:594–602.
Rahmstorf, S. 2007. A semi-empirical approach to projecting future sea-level rise. Science 315:368–370.
St. John, H. 1921. Sable Island, with a catalogue of its vascular plants. Proceedings of the Boston Society of Natural History 36(1): 1–103, pls 1, 2.
Sakagami, S.F. and C.D. Michener. 1962. The Nest Architecture of the Sweat Bees (Halictinae), a Comparative Study of Behavior. The University of Kansas Press, Lawrence, Kansas, [v] + 135 pp.
Salafsky, N., D. Salzer, A.J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O’Connor, and D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conservation Biology 22:897–911.
Shaw, R.W. and the Climate Change Action Fund (CCAF A041) project team. 2001. Coastal impacts of climate change and sea-level rise on Prince Edward Island, synthesis report. Prepared for Environment Canada, Natural Resources Canada and Fisheries and Oceans Canada, Darthmouth, Nova Scotia. 74 pp.
Sheffield, C.S. 2013. Personal communication with J. Gibbs. Royal Saskatchewan Museum, Regina, SK.
Sheffield C.S., P.G. Kevan, A. Pindar, and L. Packer. 2013. Bee (Hymenoptera: Apoidea) diversity within apple orchards and old fields in the Annapolis Valley, Nova Scotia, Canada. The Canadian Entomologist 145:94–114.
Stockhammer, K. A. 1967. Some notes on the biology of the blue sweat bee, Lasioglossum coeruleum. Journal of the Kansas Entomological Society 40:177–189.
Tuell, J.K. and R. Isaacs. 2010. Weather during bloom affects pollination and yield of highbush blueberry. Journal of Economic Entomology 103:557–562.
Vasseur L. and N. Catto. 2008. Atlantic Canada. In: Lemmen, D.S., F.J., Warren, J,Lacroix, and E. Bush,E. (eds.) From Impacts to Adaptation: Canada in a Changing Climate 2007; Government of Canada, Ottawa, ON, 448 pp.
Westby, S. 2014. Personal communication to Cory Sheffield. Entomology technician (retired). Agriculture and Agri-Foods Canada, Halifax, Nova Scotia.
Wright, B. 1989. The fauna of Sable Island. Curatorial Report Number 68, Nova Scotia Museum, Halifax, pp. 93.
Yanega, D. 1990. Philopatry and nest founding in a primitively social bee, Halictus rubicundus. Behavioral Ecology and Sociobiology 27:37–42.
Yanega, D. 1997. Demography and sociality in halictine bees (Hymenoptera: Halictidae). Pp. 293–315. in Choe, J.C. and B.J. Crespi (eds.). The Evolution of Social Behaviour in Insects and Arachnids. Cambridge University Press, Cambridge, U.K.
Biographical summary of report writers
Jason Gibbs completed his PhD at York University in 2009. His main research area is the systematics of bees with a focus on the genus Lasioglossum in the family Halictidae. He has completed three large-scale revisions of Lasioglossum species in North America including the species of Lasioglossum in Canada. He described Lasioglossum sablense as a new species in 2010. As a post-doctoral researcher at Cornell University Dr. Gibbs completed, with help from colleagues, the most comprehensive phylogeny of Lasioglossum published to date. In 2013 he began a new position as Research Associate in the Department of Entomology at Michigan State University. He is currently involved in a large-scale project studying pollinators of specialty crops. He is still involved in ongoing efforts to study systematics of Lasioglossum.
Collections examined
Material from the Canadian National Collection of Insects, Arachnids and Nematodes (CNC) was examined specifically for this report. Prior to preparation of this report tens of thousands of specimens of Lasioglossum were examined from more than two dozen collections in Canada and the United States, including material from Maritime Provinces and New England states as part of ongoing taxonomic studies of halictid bees. The type series, deposited at Packer Collection York University (PCYU) and the CNC was examined during the original species description (Gibbs 2010). Relevant material examined includes specimens from PCYU, CNC, and collections from Agriculture and Agri-Food Canada Atlantic Food and Horticulture Research Centre in Kentville, Nova Scotia.