
Western toad (Anaxyrus boreas): COSEWIC assessment and status report 2012
- non-calling population
- calling population
Special concern 2012

Description for cover photo
Photo of a Western Toad, Anaxyrus boreas, showing the face, abdomen, and much of the back and left side. The back and sides are pale olive green marked by reddish-brown and dark brown blotches. There are reddish-brown, circular "warts" on the back, sides and upper portions of the limbs. The throat and belly are pale with dark mottling.
Table of contents
- Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assessment summary
- COSEWIC executive summary
- Technical summary – non-calling population
- Technical summary – calling population
- Preface
- Wildlife species description and significance
- Distribution
- Habitat
- Biology
- Population sizes and trends
- Threats and limiting factors
- Protection, status, and ranks
- Acknowledgements and authorities contacted
- Information sources
- Biographical summary of report writer
- Collections examined
List of figures
- Figure 1. Adult male Anaxyrus boreas from northwestern British Columbia. Photograph by Brian G. Slough
- Figure 2. Western Toad eggs (left) and tadpoles (right) from Thompson-Nicola area, B.C. Photographs by K. Ovaska
- Figure 3. Global distribution of the Western Toad. From Environment Canada 2011. Map prepared by Rob Gau, Northwest Territories Department of Environment and Natural Resources; U.S. and Mexican range based on a map compiled by International Union for Conservation of Nature (IUCN)
- Figure 4. Canadian distribution of the Western Toad
- Figure 5. Mass breeding site of Western Toads in flooded shallows of Tutshi River Delta, Tutshi River, northwestern B.C. Photograph by Brian G. Slough
- Figure 6. Mass-breeding site of the Western Toad in the Thompson-Nicola area, B.C. Black band near the shoreline is composed of 1000s of tadpoles. Photograph by L. Sopuck
- Figure 7. Western Toad tadpoles on silty substrate of Lindeman Creek delta on Lindeman Lake, northwestern B.C. Photograph by Brian G. Slough
List of tables
List of appendices

Document information
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2012. COSEWIC assessment and status report on the Western Toad in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xiv + 71 pp.
Previous report(s)
COSEWIC. 2002. COSEWIC assessment and status report on the western toad (Bufo boreas) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 31 pp.
Wind, E.l. and L.A. Dupuis. 2002. COSEWIC status report on the western toad (Bufo boreas) in Canada, in COSEWIC assessment and status report on the western toad (Bufo boreas) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-31 pp.
Production note:
COSEWIC would like to acknowledge Brian G. Slough for writing the status report on the western toad, (Anaxyrus boreas), in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Kristiina Ovaska, Co-chair of the COSEWIC Amphibians and Reptiles Specialist Subcommittee.
For additional copies contact
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel. : 819-953-3215
Fax : 819-994-3684
COSEWIC E-mail
COSEWIC Website
Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur le crapaud de l'Ouest (Anaxyrus boreas) au Canada.
Cover illustration/photo
Western toad -- Cover photograph by Kristiina Ovaska
©Her Majesty the Queen in Right of Canada, 2013.
Catalogue No. CW69-14/346-2013E-PDF
ISBN 978-1-100-22121-2

COSEWIC assessment summary
Assessment summary – November 2012
Common name: Western Toad - Non-calling population
Scientific name: Anaxyrus boreas
Status: Special Concern
Reason for designation: This species has suffered population declines and population extirpations in the southern part of its range in British Columbia, as well as in the USA. The toads are particularly sensitive to emerging skin disease caused by the amphibian chytrid fungus, which has been linked to global amphibian declines. It is relatively intolerant of urban expansion, conversion of habitat for agricultural use, and habitat fragmentation resulting from resource extraction and road networks. Life history characteristics, including infrequent breeding by females, aggregation at communal, traditionally used breeding sites, and migrations to and from breeding sites, make populations vulnerable to habitat degradation and fragmentation. The species remains widespread, but declines are suspected and projected based on known vulnerabilities and threats.
Occurrence: Yukon, Northwest Territories, British Columbia, Alberta
Status history: The species was considered a single unit and designated Special Concern in November 2002. Split into two populations in November 2012. The Non-calling population was designated Special Concern in November 2012.
Assessment summary – November 2012
Common name: Western Toad - Calling population
Scientific name: Anaxyrus boreas
Status: Special Concern
Reason for designation: Almost the entire range of the calling population is within Canada. The toads are particularly sensitive to emerging skin disease caused by the amphibian chytrid fungus, which has been linked to global amphibian declines. This species is relatively intolerant of urban expansion, conversion of habitat for agricultural use, and habitat fragmentation resulting from resource extraction and road networks. Life history characteristics, including infrequent breeding by females, aggregation at communal, traditionally used breeding sites, and migrations to and from breeding sites, make populations vulnerable to habitat degradation and fragmentation. The species remains widespread throughout much of their historic range in Alberta and may be expanding their range eastwards. However, declines are suspected and projected based on known vulnerabilities and threats.
Occurrence: Alberta
Status history: The species was considered a single unit and designated Special Concern in November 2002. Split into two populations in November 2012. The Calling population was designated Special Concern in November 2012.

COSEWIC executive summary
Wildlife species description and significance
The Western Toad is a large toad with small round or oval “warts” on the back, sides and upper portions of the limbs. Large oblong parotoid or poison cheek glands are situated behind the eyes. Colour is typically brown or green but varies from olive green to almost reddish-brown or black; a creamy or white vertebral stripe often extends along the back. The “warts” and parotoid glands are often reddish-brown. There is a grey pelvic patch in the groin area that functions to absorb moisture from the environment.
Western Toads in most of Alberta are behaviourally and morphologically distinct from other populations of Western Toads in that males possess a vocal sac and produce loud advertisement calls during the breeding season. Preliminary phylogenetic analyses show that these populations are distinct. Consequently, two designatable units are recognized in this report: calling toads that occur in most of Alberta, extending into British Columbia in the Rocky Mountains, and non-calling toads within the remaining part of the species’ Canadian range.
The Western Toad contributes significantly to ecological processes over a wide range of wetlands and terrestrial habitats. The sheer volume of tadpoles and metamorphs (newly metamorphosed toadlets) at breeding sites implies that their conversion of biomass is significant, and the dispersal of metamorphs represents a significant transfer of energy from aquatic to terrestrial ecosystems.
Distribution
Globally, the Western Toad is found from Baja California (Mexico), Nevada, Utah, Colorado and New Mexico north through Canada and Alaska. In Canada, the Western Toad is found throughout British Columbia and western Alberta, entering Yukon and Northwest Territories in the Liard River basin. It appears to be the only amphibian native to Haida Gwaii.
Habitat
Western Toads use a wide variety of aquatic and upland habitats. They breed in a variety of wetlands including shallow, sandy margins of lakes, ponds, streams, river deltas, river backwaters, river estuaries, and geothermal springs. Following breeding, adults may remain to forage in the marshy or riparian edges of breeding sites, or they may disperse several kilometres to foraging areas in other wetlands, riparian areas along streams, or upland sites. Western Toads hibernate underground, often in spaces created or modified by small mammals.
Biology
Western Toads aggregate in spring to breed. Eggs are laid in long, intertwined strings in the shallow margins of lakes and ponds at communal breeding sites. Tadpoles are gregarious and form large aggregations. Metamorphosis is usually complete by August, but tadpoles have been observed later in colder areas at higher altitudes and latitudes. Newly emerged toadlets form large aggregations and migrate en masse away from ponds. Toads reach sexual maturity at 3-4 yrs for males and 4-6 yrs for females. Males may breed more than once per season, and in consecutive years, but females rarely do so and may breed only once in their lifetime. Western Toads are vulnerable to disturbance at breeding sites and during migrations to and from breeding sites.
Population sizes and trends
The Western Toad is apparently widespread, abundant, and persistent across much of its Canadian range. However, congregation at breeding sites and mass migrations of toadlets may give an appearance of abundance that is seldom substantiated. Numbers appear to be declining along the south coast of British Columbia and Vancouver Island, and localized declines or fluctuations have been noted elsewhere in British Columbia. In Alberta, the species may be expanding its range eastwards, but it is unclear whether the new records represent range expansion or simply reflect increased survey efforts associated with resource extraction. In all areas, the toads remain vulnerable to epidemics of emerging diseases, which have devastated Western Toad populations in the USA, and to increasing anthropogenic threats. The absence of long-term data sets is problematic for assessing trends.
Threats and limiting factors
Habitat loss, degradation, and fragmentation, including intersection of seasonally used habitats by roads, are issues near human population centres, agricultural settings, and areas of intensive resource development. Other, widespread threats to the Western Toad are amphibian chytrid fungus, which has been linked to global amphibian declines, and other infectious diseases. Co-stressors, such as habitat degradation, climate change, and increased UV-B, may increase vulnerability to disease. In southwest British Columbia, the introduced Bullfrog is a predator, competitor, and a reservoir for disease. Late maturity of females and their infrequent reproduction limit the ability of Western Toad populations to recover from declines.
Protection, status, and ranks
The Western Toad was designated as a species of Special Concern in 2002 by COSEWIC, and it is on Schedule 1 of the federal Species at Risk Act. In Canada, the general status of the Western Toad is Sensitive in all jurisdictions except Northwest Territories, where it is May Be at Risk. NatureServe rankings are Vulnerable to Apparently Secure in Canadian jurisdictions. The IUCN designation is Near Threatened.
Technical summary – non-calling population
Anaxyrus boreas
Western Toad - non-calling population
Crapaud de l’Ouest - population non-chantante
Range of occurrence in Canada: Yukon, Northwest Territories, British Columbia, Alberta
Demographic Information
Extent and occupancy information
* See definitions and abbreviations on COSEWIC website and IUCN 2010 for more information on this term.
Number of mature individuals (in each population)
| Population | Number of mature individuals |
|---|---|
| Populations not differentiated here, but there are potentially isolated regional populations. | Unknown, but >100,000 |
| Total | Unknown, but >100,000 |
Quantitative analysis
Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. Not conducted
Threats (actual or imminent, to populations or habitats)
- Amphibian chytrid fungus (introduced species such as Bullfrogs may act as reservoirs).
- Habitat loss and fragmentation due to human settlement and transportation corridors, which can isolate sub-populations, leading to increased risk of extinction.
- Road mortality during mass migrations to and from breeding sites.
- Other stressors such as chemical pollution pathogens, such as Saprolegnia (introduced with stocked fish), and increased UV-B radiation, which may act independently or synergistically to reduce populations.
The long time to maturity for females, and the fact that 95% of females breed only once in their lifetime, makes populations especially vulnerable to threats and declines.
Rescue effect (immigration from outside Canada)
Status history
COSEWIC: The species was considered a single unit and designated Special Concern in November 2002. Split into two populations in November 2012. The Non-calling population was designated Special Concern in November 2012.
Status and reasons for designation
Status: Special Concern
Alpha-numeric code: Not applicable
Reasons for designation: This species has suffered population declines and population extirpations in the southern part of its range in British Columbia, as well as in the USA. The toads are particularly sensitive to emerging skin disease caused by the amphibian chytrid fungus, which has been linked to global amphibian declines. It is relatively intolerant of urban expansion, conversion of habitat for agricultural use, and habitat fragmentation resulting from resource extraction and road networks. Life history characteristics, including infrequent breeding by females, aggregation at communal, traditionally used breeding sites, and migrations to and from breeding sites, make populations vulnerable to habitat degradation and fragmentation. The species remains widespread, but declines are suspected and projected based on known vulnerabilities and threats.
Applicability of criteria
- Criterion A (Decline in total number of mature individuals):
- Not applicable; population trends are unknown, although population declines have been reported from localized areas mainly from southwestern British Columbia.
- Criterion B (Small distribution range and decline or fluctuation):
- Not applicable; both EO and IAO are above threshold values.
- Criterion C (Small and declining number of mature individuals):
- Not applicable; population is larger than threshold values, and trends are unknown.
- Criterion D (Very small or restricted total population):
- Not applicable; population size is unknown but much larger than threshold values; number of locations and IAO exceed guidelines for D2 Threatened.
- Criterion E (Quantitative analysis):
- Insufficient data; not conducted.
Technical summary - calling population
Anaxyrus boreas
Western Toad - calling population
Crapaud de l’Ouest - population chantante
Range of occurrence in Canada: British Columbia, Alberta
Demographic information
Extent and occupancy information
* See Definitions and Abbreviations on COSEWIC website and IUCN 2010 for more information on this term.
Number of mature individuals (in each population)
| Population | Number of mature individuals |
|---|---|
| Populations not differentiated here, but there are potentially isolated regional populations. | Unknown, but >100,000 |
| Total | Unknown, but >100,000 |
Quantitative analysis
Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. Not conducted
Threats (actual or imminent, to populations or habitats)
- Amphibian chytrid fungus.
- Habitat loss and fragmentation due to human settlement, agriculture, forestry, oil and gas industry, and transportation corridors, which can isolate sub-populations, leading to increased risk of extinction.
- Road mortality during mass migrations to and from breeding sites.
- Several stressors including chemical pollution, pathogens such as Saprolegnia (introduced with stocked fish), and increased UV-B radiation, which may act independently or synergistically to reduce populations.
The long time to maturity for females, and the fact that 95% of females breed only once in their lifetime, makes populations especially vulnerable to threats and declines.
Rescue effect (immigration from outside Canada)
Status history
COSEWIC: The species was considered a single unit and designated Special Concern in November 2012. Split into two populations in November 2012. The Calling population was designated Special Concern in November 2012.
Status and reasons for designation
Status: Special Concern
Alpha-numeric code: Not applicable
Reasons for designation: Almost the entire range of the calling population is within Canada. The toads are particularly sensitive to emerging skin disease caused by the amphibian chytrid fungus, which has been linked to global amphibian declines. This species is relatively intolerant of urban expansion, conversion of habitat for agricultural use, and habitat fragmentation resulting from resource extraction and road networks. Life history characteristics, including infrequent breeding by females, aggregation at communal, traditionally used breeding sites, and migrations to and from breeding sites, make populations vulnerable to habitat degradation and fragmentation. The species remains widespread throughout much of their historic range in Alberta and may be expanding their range eastwards. However, declines are suspected and projected based on known vulnerabilities and threats.
Applicability of criteria
- Criterion A (Decline in total number of mature individuals):
- Not applicable; population trends are unknown.
- Criterion B (Small distribution range and decline or fluctuation):
- Not applicable; both EO and IAO are above threshold values.
- Criterion C (Small and declining number of mature individuals):
- Not applicable; population is larger than threshold values, and trends are unknown.
- Criterion D (Very small or restricted total population):
- Not applicable; population size is unknown but much larger than threshold values; number of locations and IAO exceed guidelines for D2 Threatened.
- Criterion E (Quantitative analysis):
- Insufficient data; not conducted.
Preface
This is an updated status report on the Western Toad, Anaxyrus (= Bufo) boreas(COSEWIC 2002). Frost et al.(2006, 2008) proposed that North American toads of the genus Bufo be placed in a separate genus, Anaxyrus.
New information indicates that Western Toads from the northeastern part of the species’ range, including most of Alberta, differ significantly from Western Toads from elsewhere in the species’ range in that they have a loud mating call and vocal sac (Pauly 2008). This is evidence for two designatable units in Canada. Surveys have improved knowledge of the extent of occurrence and area of occupancy.
Aboriginal Traditional Knowledge (ATK) was sought through COSEWIC’s ATK Subcommittee and relevant Wildlife Management Boards, but there is none to report.
COSEWIC history
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2012)
- Wildlife species
- A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
- Special concern (SC)*
- A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
- Not at risk (NAR)**
- A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
- Data deficient (DD)***
- A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
*Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.
**Formerly described as “Not In Any Category”, or “No Designation Required.”
***Formerly described as "Indeterminate" from 1994 to 1999 or "ISIBD" (insufficient scientific information on which to base a designation) prior to 1994. Definition of the (DD) category revised in 2006.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife species description and significance
Name and classification
The Western Toad, Anaxyrus(formerly Bufo) boreas, is part of the cosmopolitan family Bufonidae or True Toads (class Amphibia, order Anura). Frost et al. (2006) proposed that North American toads be removed from the genus Bufo and be placed in a separate genus, Anaxyrus (see also Frost et al. 2008). Though this proposal has been challenged by Pauly et al.(2009), who suggest that the change is unnecessary and destabilizing as no new clades were described, the nomenclature has been accepted (Crother 2012).
The Western Toad has two recognized subspecies: (1) Anaxyrus boreas boreas, the Boreal Toad, found from northern California, Nevada, Utah, Colorado and New Mexico north through Canada and Alaska, and (2) A. boreas halophilus, the California Toad, which ranges from California to Baja California and western Nevada. Goebel (2005) and Pauly (2008), however, found no genetic evidence to support this distinction.
Goebel et al. (2009) identified haplotype clades of Anaxyrus boreas from 288 individuals from 58 sites, 3 of which were in Canada (Surrey, Vancouver Island, and Little Tahltan River, all in British Columbia). No samples from Alberta were included in the analyses. Western Toads from the 3 sites in British Columbia belong to a major northwest mtDNA haplotype clade (Goebel et al. 2009). Goebel et al. (2009) recommended a taxonomic revision of the Anaxyrus boreas species group using nuclear DNA and morphological characteristics. A rearrangement of the taxonomy though, would not affect the subspecies found in Canada.
Pauly (2008) described behavioural and morphological uniqueness of Western Toads from Alberta, where males possess a vocal sac and produce advertisement calls. Preliminary phylogenetic analyses suggest that Western Toads inherited their call from a calling ancestor (as did the closely related Yosemite Toad, Anaxyrus canorus). The advertisement call was lost, however, in the lineage leading to the non-calling Western Toads throughout the remainder of the Canadian range (Pauly pers. comm. 2011).
Amplexus has been observed between mismatched pairs of the Western Toad and Canadian Toad (Anaxyrus hemiophrys) in west-central Alberta (Cook 1983; Eaton et al. 1999). One hybrid was identified by Cook (1983) based on morphology. The viability and fertility of hybrids are unknown, but these factors presumably contribute to reproductive isolation of the two species along with a low abundance of one of the two species typical at sites within the zone of overlap (Eaton et al. 1999). The appearance of vocal sacs and call productions was not due to hybridization between the Western and Canadian Toads, and there was no evidence of gene flow between the two species in Alberta (Pauly 2008).
Morphological description
The Western Toad is a large toad with small, round or oval glandular protuberances or “warts” on the back, sides and upper portions of the limbs (Figure 1; Russell and Bauer 2000; Matsuda et al.2006). Large oblong parotoid (cheek) glands are situated behind the eyes. Colour is typically brown or green but varies from olive green to almost reddish-brown or black with a creamy or white vertebral stripe extending, but sometimes broken or nearly absent, from snout to vent. The “warts” and parotoid glands are often reddish-brown and may be encircled by a ring of dark pigment. These structures are poison glands that excrete a noxious white liquid that deters predators. The throat and belly are pale with dark mottling. There is a grey pelvic patch in the groin area that is used to absorb moisture from the environment. The pupil is horizontal, and cranial crests are weakly developed. The limbs are relatively short, and the hind toes are partially webbed. Toads move on land by walking and hopping. Horny tubercles on the hind feet are used for digging backwards into the ground.
Figure 1. Adult male Anaxyrus boreas from northwestern British Columbia
Photograph by Brian G. Slough

Description of Figure 1
Photo of an adult male Western Toad showing the dorsal surfaces. This individual is olive green with brownish round protuberances on the back, sides and upper portions of the limbs. The pupil is horizontal, and cranial crests are weakly developed.Photo of an adult male Western Toad showing the dorsal surfaces. This individual is olive green with brownish round protuberances on the back, sides and upper portions of the limbs. The pupil is horizontal, and cranial crests are weakly developed.
Adult males are 60 to 110 mm in snout-vent length and weigh ≤ 80 g. Females are larger, reaching 75 to 125 mm in snout-vent length and weigh ≤ 115 g. Males develop black nuptial pads on their thumbs and first two toes of the forefeet during the breeding season. Male Western Toads are distinguished from females by the presence/absence of nuptial pads (usually in late May – early June), longer forelimbs, narrower heads, and a less prominent or discontinuous mid-dorsal stripe (Carstensen et al. 2003; Matsuda et al. 2006).
Eggs are black and are laid in long, intertwined paired strings (Figure 2). Tadpoles are jet-black or charcoal in colour and range from 9 to 42 mm in total length (Figure 2). Metamorphs are about 12 to 22 mm in snout-vent length and weigh ≤ 0.5 g.
Figure 2. Western Toad eggs (left) and tadpoles (right) from Thompson-Nicola area, B.C.
Photographs by K. Ovaska

Description of Figure 2
Two-part photo image showing Western Toad eggs (left image) and tadpoles (right image) in shallow, reedy water. The eggs are black and form long, paired strings. An observer has placed a hand beneath the eggs to make them easier to see against the dark background. The three tadpoles are black and photographed from above.
Vocalizations
The “release call” of the male is a quiet series of chirps like the peeping of a chick (Russell and Bauer 2000) and presumably prevents prolonged amplexus with other males. It may also be emitted without tactile stimulation, which is unusual among toads, and may function as an encounter call in other social contexts. It is not known whether the latter call is a signal to other males, or if it has some other purpose such as advertisement to females. True advertisement calls consist of relatively long and high-amplitude pulsed trills and have been documented only from northeastern portion of the species’ distribution, throughout much of Alberta (Pauly 2008; Long 2010) and from one site in the B.C. Rockies (McIvor per. comm. 2012). Advertisement calls have also been reported from a Montana population, but these calls are weak and dissimilar to the loud calls produced by Alberta toads (Pauly 2008). Pauly (2008) found that Alberta toads possessed vocal sacs necessary for producing loud advertisement calls, whereas Western Toads examined from elsewhere within the species’ range did not. A narrow zone of overlap between males with and without vocal sacs occurs in western and northwestern Alberta and northwestern Montana (Pauly 2008).
Population spatial structure and variability
Pond breeding amphibians such as the Western Toad typically are assumed to have strong breeding site fidelity, high vagility within home ranges, limited dispersal abilities, and spatially disjunct breeding sites (Smith and Green 2005). The authors noted, however, that caution should be exercised when applying the metapopulation approach to conservation, because not all species are structured as metapopulations. Known breeding sites of Western Toads may be separated by distances of up to 30 km (Slough 2004). As a result, the breeding sub-populations may be subject to local extinction and recolonization, and hence probably form metapopulations (Marsh and Trenham 2001; Smith and Green 2005). Local and regional population persistence depends on breeding site distribution and connectivity.
Western Toads’ vagility appears to be high among pond-breeding amphibians (see Marsh and Trenham 2001 and references therein). Western Toads have demonstrated an affinity for movements along streams and riparian areas (Adams et al. 2005b; Bull 2009) and may also occur in open habitats such as burns (Guscio et al.2008), agricultural fields (Browne 2010), and upland sites (Schmetterling and Young 2008). Schmetterling and Young (2008) documented mean in-stream summer movements of 2.9 km, with a maximum movement of 13 km, in an average of 23 days.
A phylogeographic study of Western Toads from the islands and mainland of southeast Alaska and interior watersheds in British Columbia that connect to the coast is currently underway (2009 – 2011) to assess genetic diversity in the area (Payare pers. comm. 2011).
Designatable units
There is evidence for two designatable units (DUs): (1) population where males produce an advertisement call and possess vocal sacs (Calling population) and (2) population where males lack an advertisement call and vocal sac (Non-calling population). The Calling population occurs in most of Alberta, entering into British Columbia in the Rocky Mountains, whereas the Non-calling population occurs in most of British Columbia, Yukon, Northwest Territories, and extreme western and southwestern Alberta. Divergence in a primary mating signal is significant in anurans where species and mate recognition is typically accomplished by acoustic communication. Such divergence is usually associated with species-level differentiation (e.g., Red-legged Frog, Rana aurora, complex, Shaffer et al. 2004).
The non-calling and calling toads occupy discrete geographic areas. The calling toads occur in the northeastern portion of the species’ range, mostly in Alberta, but the Rocky Mountains are not an absolute barrier; there are a few sites with non-calling toads in western Alberta, east of the Rocky Mountains, and at least one site with calling toads west of the continental divide in the Rocky Mountains (see Canadian Range). A map produced by Pauly (2008), based on specimens he examined for the presence of vocal sacs, provides an approximate overview of the distribution of the two populations (Figure 2.1 in Pauly 2008, reproduced here as Appendix 1), but the boundary zone is incompletely known. The Calling population occurs mainly in the Prairie/Western Boreal Amphibian and Reptile faunal province, whereas the Non-calling population occurs in the Pacific Coast, Intermountain, and Prairie/Western Boreal Amphibians and Reptile faunal provinces. Both populations enter into the Rocky Mountain faunal province.
Special significance
The Western Toad contributes significantly to ecological processes over a wide range of wetlands and terrestrial habitats. Eggs, tadpoles, and metamorphs are preyed on by a variety of aquatic invertebrates, fish, birds, reptiles, mammals and other amphibian species. Tadpoles consume algae and detritus. The sheer volume of tadpoles and metamorphs at some breeding sites ensures that their conversion of biomass is significant, and the dispersal of metamorphs represents a significant transfer of energy from aquatic to terrestrial systems. Adults and juveniles consume invertebrates, many of which are insect pests, and are in turn preyed on by birds, mammals and reptiles.
Distribution
Global range
The Western Toad ranges from coastal Alaska in Prince William Sound and northwestern Canada in the north to Baja California, Mexico in the southwest and northern New Mexico, Colorado, and Wyoming in the east (Figure 3; Stebbins 2003).
Figure 3. Global distribution of the Western Toad. From Environment Canada 2011
Map prepared by Rob Gau, Northwest Territories Department of Environment and Natural Resources; U.S. and Mexican range based on a map compiled by IUCN, Conservation International, NatureServe, and collaborators, 2004 (NatureServe 2012).
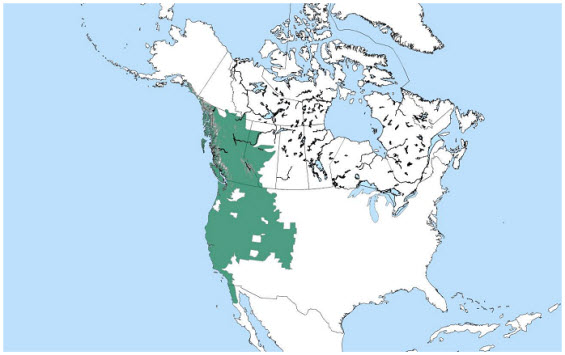
Description of Figure 3
Map of the global distribution of the Western Toad. The species ranges from coastal Alaska in Prince William Sound and northwestern Canada in the north to Baja California, Mexico, in the southwest and northern New Mexico, Colorado, and Wyoming in the east.
Canadian range
The Canadian range of the Western Toad includes most of British Columbia (Matsuda et al. 2006; Friis unpubl. data 2007; Leaver unpubl. data 2007), though the species appears to be absent from the Teslin River basin in the northwest (Slough and Mennell unpubl. data 2007) (Figure 4). There is an acoustic record of a release call from 2001 east of Teslin Lake (shown in Matsuda et al. 2006) that remains unconfirmed after 4 subsequent surveys (Slough unpubl.data 2001, 2004, 2005, 2007). Western Toads are likely absent from extreme northeast British Columbia.
Figure 4. Canadian distribution of the Western Toad. See text for data sources.

Description of Figure 4
Map of the Canadian distribution of the Western Toad. The range includes most of British Columbia, though the species appears to be absent from the Teslin River basin in the northwest. The species has been found in southeastern Yukon and in the southwestern Northwest Territories. In Alberta, the Western Toad ranges from the forested regions of the southwest to central and northern Alberta. It also enters the short-grass prairie and aspen parkland. Western Toad observations are indicated on the map as follows: solid black circle equals recent observations (after 2000); black circle with grey fill equals older observations (1980 to 1999); hollow black circle equals historical (before 1980).
The Western Toad has been found in the southeast Yukon (Slough and Mennell 2006) and in the southwest Northwest Territories (Northwest Territories Department of Environment and Natural Resources 2006; Schock et al.2009) in the Liard River Basin.
In Alberta, the Western Toad ranges from the forested regions of the southwest to central and northern Alberta. It also enters the short-grass prairie and aspen parkland. Based on widely separated occurrence records (Figure 4), the distribution in northern Alberta may be more extensive than currently documented, but lack of access has hindered survey efforts (Russell and Bauer 2000; Paszkowski pers. comm. 2012). The Western Toad might be expanding its range eastward in northern Alberta or, alternatively, it may have been always present, but increased survey efforts associated with oil and gas drilling and forestry activities have only recently detected its presence (Paszkowski pers. comm. 2012).
The Calling population occurs mainly to the east and the Non-calling population to the west of the Rocky Mountains, but the boundary is incompletely known and can be determined on the ground, as needed. Western Toads from Banff National Park reportedly call (Lepitzki pers. comm.2012). Calling toads also occur in Kootenay National Park, British Columbia, west of the continental divide based on a photograph of a calling toad taken by Diane McIvor in 2004, which shows the vocal sac (McIvor pers. comm.2012). Minor overlap occurs along the western Alberta border, where a small number of populations contain both morphotypes (Appendix 1).
Extent of occurrence and area of occupancy
Using the minimum polygon method, the EO for the Non-calling and Calling populations is 1,252,000 km² and 348,400 km², respectively. The IAO is difficult to calculate with any accuracy because of incomplete survey coverage and paucity of distributional information from wide areas, especially from central and northern British Columbia. Ideally, only breeding sites rather than all site records would be included in the IAO calculation for this species that congregate from the surrounding landscape at often traditionally used communal breeding sites, but adequate information from across the species’ range is lacking. Using all known records (breeding and non-breeding), the discrete IAO is 2,536 km² for the Calling population and 3,024 km² for the Non-calling DU. Both values almost certainly underestimate the actual IAO for both populations.
Search effort
Occurrence records for British Columbia are from the BC Ministry of Environment and from various researchers that were contacted individually. British Columbia records collected in 2008-2011 by government officials, consultants and naturalists have not been collated into a central database (Gelling pers. comm. 2011; Ramsay pers. comm. 2011) and so did not contribute to distribution mapping in this report. Additional occurrence data for British Columbia and other jurisdictions were provided by the Canadian Museum of Nature (Steigerwald unpubl. data 2011), Parks Canada (Howes pers. comm. 2011), and the Canadian Department of National Defence (Nernberg unpubl. data 2011). Occurrence records for Alberta were obtained from the Fisheries and Wildlife Management Information System (Berg unpubl. data 2011) and the Alberta Volunteer Amphibian Monitoring Program (Kendell unpubl. data 2011). Occurrence data sets are maintained by the two territories (Carrière unpubl. data 2011; Mulder unpubl. data 2011).
In the past 18 years, significant range extensions have been documented for the Western Toad and other anurans in Yukon (Slough and Mennell 2006) and in Northwest Territories (Northwest Territories Department of Environment and Natural Resources 2006). These records reflect increased survey effort rather than range expansion. Northern Alberta and northern British Columbia remain inadequately surveyed.
Habitat
Habitat requirements
Western Toads use a wide variety of aquatic habitats for breeding. Typical breeding sites include shallow, sandy margins of lakes, ponds, streams, river deltas, river backwaters, river estuaries, and geothermal springs (Dupuis unpubl. data 2002; Wind unpubl. data 2002; Jones et al. 2005; Slough unpubl. data 2011; see Figures 5 and 6 for examples of breeding sites). Human-made water bodies such as ditches, road ruts, tailings ponds, and borrow pits are also used. The bottom substrate is often silty or sandy (Figure 7). Beaver ponds are used extensively in the northern part of the species’ range (Slough and Mennell 2006; Stevens et al. 2007). Oviposition sites may be in water depths ≤ 2 m, but shallow water <1 m is preferred (Corn 1998). Browne et al.(2009) found that toads were found in shallow ponds with higher daytime temperatures and higher dissolved oxygen, which facilitate tadpole growth, than wetlands that lacked toads. Some human-made habitats may be reproductive sinks that fail to produce metamorphs and result in wasted reproductive effort (Stevens and Paszkowski 2006). Permanent water bodies are usually preferred over ephemeral ponds, which may also be reproductive sinks in times of drought. Water may be clear, silty or even brackish (Storer 1925; Slough and Mennell 2006). Breeding sites may be extremely open and unprotected by riparian or aquatic vegetation, or by other structural components such as woody debris, rocks or undercut banks. A key feature of breeding sites is water temperature, with higher temperatures accelerating tadpole growth (Ultsch et al.1999).
Figure 5. Mass breeding site of Western Toads in flooded shallows of Tutshi River Delta, Tutshi River, northwestern B.C.
Photograph by Brian G. Slough

Description of Figure 5
Photo of a mass breeding site of Western Toads in flooded shallows of the Tutshi River Delta in northwestern British Columbia. A watery area occupies the foreground with clumps of vegetation emerging above the surface. Farther away, about half way up the image, is the shoreline with trees in the background. Steep valley sides are visible to the left and right of the image. In the distance are mountains with some snow covering.
Figure 6. Mass-breeding site of the Western Toad in the Thompson-Nicola area, B.C. Black band near the shoreline is composed of 1000s of tadpoles. Photograph by L. Sopuck.

Description of Figure 6
Photo of a mass-breeding site of the Western Toad in the Thompson-Nicola area, British Columbia. In the foreground, low vegetation gives way to a sandy shore and water body, which runs from the bottom right to upper left third of the image. A black band in the water parallel to the shoreline is composed of thousands of tadpoles. A wooded hillside occupies the top portion of the image.
Figure 7. Western Toad tadpoles on silty substrate of Lindeman Creek delta on Lindeman Lake, northwestern B.C.
Photograph by Brian G. Slough

Description of Figure 7
Photo of Western Toad tadpoles on silty substrate of the Lindeman Creek delta on Lindeman Lake, northwestern British Columbia. There are dozens of tadpoles dispersed across the image.
Western Toads exhibit breeding site fidelity, returning to the same wetlands in successive years (Bull and Carey 2008). They use an “explosive” breeding strategy at communal breeding sites, where large numbers of adults aggregate over a 1- or 2-week period. Aggregate breeding reduces energy costs associated with pair-bonding, it takes advantage of a brief summer and temporary breeding ponds available for larval development, and ensures genetic mixing (Myers and Zamudio 2004). Site fidelity and communal breeding may result in the selection of only one or a few of the potential breeding sites in a relatively large area (Slough 2004).
Western Toad tadpoles aggregate in warm shallow margins of lakes and streams during the day (Figure 6), a behaviour that accelerates their rate of development and provides cover from predators in emergent and flooded shoreline vegetation. They may disperse to deeper waters at night.
Following breeding, adults may remain and forage in adjacent marshes or riparian edges of breeding sites, or they may travel up to several kilometres to other wetlands, riparian areas along streams, or upland sites, such as forests, meadows, shrub lands, or subalpine or alpine meadows. Females tend to travel farther than males to reach foraging grounds (Muths 2003; Bartelt et al. 2004; Bull 2006, Browne 2010). Males are more closely associated with water and move shorter distances than females (Bull 2006). In a boreal mixed-wood forest of Alberta, Macdonald et al.(2006) found that Western Toads were more abundant in forests ≤ 100 m from lakes than they were 400-1200 m away. Open, warm areas with abundant prey, such as wet shrub and hay/crop fields were selected in Alberta during the breeding and pre-hibernation seasons in boreal forest and aspen parkland sites (Browne 2010). Closed deciduous forest cover was preferred in the boreal region, and low shrub cover was avoided during the foraging season (Browne et al. 2009). Clearcuts and edges of clearcuts are used by Western Toads (Ward and Chapman 1995; Gyug 1996; Davis 2000; Deguise and Richardson 2009a) depending on the seasonal risk of desiccation. Toads preferred and increased their breeding activity in recent burns in Montana (Guscio et al. 2008), providing further evidence of use of open terrestrial habitats.
Western Toads seek overhead cover, such as shrubs, dense herb layers, coarse woody debris, boulders or mammal burrows, presumably for protection from desiccation and predation (Davis 2000; Bartelt et al. 2004). They may also dig shallow scrapes or burrows in loose soils or sand.
Western Toads hibernate underground, below the frost line to prevent freezing, and near water to prevent desiccation. In Alberta, Western Toads used cavities in peat hummocks, Red Squirrel (Tamiasciurus hudsonicus) middens, natural crevices, decayed root channels, cavities under spruce trees, abandoned Beaver (Castor canadensis) lodges, and Muskrat (Ondatra zibethicus) tunnels for hibernation (Browne and Paszkowski 2010a). These sites were in a wide variety of forested and unforested (shrub lands, marshes and meadows) habitats, but there was a strong selection for spruce forests. Most (68%) hibernation sites were communal at variable distances (146-1936 m) from breeding sites. Larger toads arrived at hibernation sites later and along straighter routes than smaller toads, suggesting that they had superior navigational skills or sufficient energy or water reserves to accomplish rapid and longer movements (Browne and Paszkowski 2010b). Hibernation sites were 180-6230 m from breeding sites in Oregon (Bull 2006) and were in rodent burrows, under large rocks, logs, root wads, or under stream or lake shore banks. Montane populations in Colorado used upland areas near seeps, stream banks, and mammal burrows (Jones et al. 1998). The Western Toad is not freeze-tolerant (Browne and Paszkowski 2010a); therefore hibernacula must be below the frost line. It has been suggested that deep snow accumulation is a requirement for survival of Western Toads in northern British Columbia (Cook 1977).
Habitat trends
Habitat alteration from resource industries, including forestry, mining, and oil and gas exploration and development, is widespread throughout much of the species’ Canadian range.
Forestry continues to modify landscapes. An average of 700 km² /year were logged on public lands in Alberta from 2006 – 2009, while in British Columbia, the average logged was 77 km² /year from 1996 – 2006 (summarized in Environment Canada 2011). Western Toads are relatively tolerant of logging, but it is unclear what the long-term effects of forest harvesting might be on population dynamics. Davis (2000) suggested that the increased proportion of closed canopy, young second-growth stands could decrease suitable toad habitat over the long term, although toads preferentially used recent clearcuts in his study on Vancouver Island. Gyug (1996) suggested that the major impact of forest harvesting on pond-breeding amphibians might be the creation of breeding ponds in clearcuts that act as population sinks due to short hydroperiods.
Oil and gas exploration and development is another source of habitat disturbance for the Western Toad. Impacts of oil and gas extraction include ecosystem conversion, fragmentation, and environmental contamination. Secondary impacts such as the construction of roads and seismic lines can also impact amphibian habitat. The oil and gas industry is most active in northeast British Columbia and northwest Alberta, overlapping the Western Toad’s range (Austin et al.2008). The Alberta Oil Sands region is at the northern extent of the Western Toads range in Alberta and will have little overlap with the species’ range. The footprint of pipelines in Alberta, where the industry has the largest footprint, is only 1.1% (Alberta Biodiversity Monitoring Institute 2012). However, fragmentation of habitats and the expansion of a network of roads associated with oil and gas exploration and other resource extraction are of concern for the Western Toad, as they affect populations over a much greater area than the actual footprint of habitat disturbance. Habitat fragmentation could potentially lead to disruption of metapopulation dynamics, population isolation, and mortality from road kill during mass migrations.
Road length in British Columbia increased 82% to over 700,000 km between 1988 and 2005 (Austin et al. 2008). The greatest density of roads is in the northeast, central interior, southwest (including Vancouver Island) and southern interior. Density of roads and other linear developments is greater than 1km / km² over much of these regions (Austin et al. 2008). While terrestrial and aquatic habitats remain or have the potential to recover in areas subject to resource use, anthropogenic activities in urban and rural areas result in permanent habitat loss. The human population in the Western Toad’s range continues to grow (5.3% in British Columbia between 2001 and 2006), and in British Columbia is concentrated in the lower mainland, the east and south coasts of Vancouver Island, and the low-elevation lake and river valleys of the southern interior (Austin et al. 2008). Wetlands and adjacent upland habitats continue to be converted to agriculture, transportation corridors and urban development in southern British Columbia and southern and central Alberta. As wetlands become more isolated, remaining populations may be unable to maintain their metapopulation structure through migration and dispersal movements, thereby becoming more vulnerable to stochastic events (Gibbs 2000).
Within the Western Toad’s range in Canada, agricultural croplands and pastures are most prevalent in Alberta, where they cover 21.6% of the province, combined (Alberta Biodiversity Monitoring Institute 2012). In British Columbia, croplands are found in the southwest, southern interior, and Peace River regions. Croplands and pastures may provide marginal habitats including breeding ponds. Wetlands used by toads for breeding are often drained for use in agriculture. In the aspen parklands of Alberta, Eaves (2004) found that Western Toads used ponds on agricultural lands less than ponds under other land uses or in undisturbed habitat; the toads occasionally used pasture ponds but not those in row crops. Toads will use cultivated areas for foraging; however, these uniform habitats may not provide hibernation sites (Browne 2010). Ponds created on farms for irrigation, borrow pits, and ponds that result from road construction, may be used by toads for breeding (Stevens and Paszkowski 2006). Pesticides used in agriculture can increase mortality rates and deformities, and reduce growth rates of larval amphibians (Bridges 2000).
Livestock grazing is widespread in the interior of British Columbia and in Alberta. Cattle congregate in riparian areas and ponds, causing extensive damage from trampling (Bartelt 1998) and eutrophication of wetlands. Western Toad life stages from eggs to adult may be trampled. Communal breeding sites and aggregations of metamorphs are especially vulnerable (Fleischner 1994). On the other hand, human-made cattle ponds may provide additional breeding sites.
Human-made breeding habitats that act as population sinks may have a negative effect on population persistence. These include ponds in clearcuts, ditches and wheel ruts, which are ephemeral (Gyug 1996; Waldick et al.1999), borrow pits, and tailings ponds where heavy metals such as cadmium reduce growth (Brinkman 1998). Borrow pits are attractive to and widely used as breeding sites by Western Toads in Alberta, but some may be unfavourable for developing embryos and larvae due to lack of thermal cover and ponds drying up before metamorphosis is completed (Stevens and Paszkowski 2006). Human-made ponds in pastures in close proximity to forests apparently compensated for the loss of natural breeding ponds in the aspen parkland of Alberta (Garrett 2005). Pearl and Bowerman (2006) documented the rapid colonization of new human-made ponds, but productivity varied among sites and overall benefit to the local population was unknown. Dug-out ponds constructed for fish research near Athabasca, Alberta in the 1960s, were rapidly colonized by Western Toads, and consistently produced large numbers of metamorphs (Paszkowski pers. comm. 2012).
Biology
Life cycle and reproduction
Western Toads congregate to breed in the spring, when minimum and maximum temperatures rise above 0°C and 10°C, respectively (Okanagan Highlands; Gyug 1996), or shortly after ice breakup (northwestern British Columbia; Slough and Mennell 2006), which occurs from late April to late May depending on latitude and elevation. Toads in central Alberta (near Whitecourt) began calling on a hot, windless day in mid-May (Long pers. comm. 2012). At Elk Island National Park, they began calling in mid- to late May and extended calling into June (Browne pers. comm. 2012). Western Toad calling was observed April 19-May 14 in Jasper National Park (Shepherd and Hughson 2012). Toads at the Atlin Warm Springs of northwestern British Columbia breed in late-February – early March (Slough and Mennell 2006). Thompson (2004) also reported early breeding at warm springs in Utah, where one of the populations did not hibernate.
Eggs are laid in long, intertwined paired strings on vegetation or branches, or at the bottom of the shallow margins of lakes and ponds at communal oviposition sites. Tadpoles hatch in 3-12 days, depending on water temperature (Hengeveld 2000; Jones et al. 2005). Western Toad tadpoles are highly gregarious and often form large aggregations (Dehn 1990). The period from egg to metamorphosis takes 4-12 weeks, depending on water temperature. Metamorphosis is usually complete by late July or early August; tadpoles have been observed into the late summer and fall in colder areas at high altitudes and latitudes, but their survival is doubtful. There are no reports of overwintering of larvae. Metamorphosis is complete by early April at the Atlin Warm Springs (Slough 2009). Toadlets form large post-metamorphic aggregations at edges of breeding sites and during migrations away from these sites (Livo 1998; Black and Black 1969).
Western Toads may aggregate at all life stages (adults at breeding, foraging or hibernation sites, eggs laid communally, tadpoles and metamorphs) making them vulnerable to predation, or other agents of mass mortality such as disease, desiccation, trampling by humans, livestock, or wildlife, or roadkill (Bartelt 1998).
Males reach sexual maturity in 3-4 years and females in 4-6 years (Olson 1988; Carey 1993; Blaustein et al. 1995; Matsuda et al. 2006). Males may mate more than once per season, and in consecutive years. In Oregon, females reached sexual maturity at 4-5 years of age and only approximately 5% mated a second time in their lives, approximately 2-4 years after their first mating (Olson 1988; Blaustein et al.1995). Bull and Carey (2008) found that 8.5% of 844 females returned to a breeding site within 5 years, and 2.5% bred in 2-3 consecutive years. There were no cases of consecutive-year breeding at a high elevation site for females. Consecutive-year breeding was not observed for females in Colorado (Carey et al. 2005). Five to 35% of males bred in consecutive years, resulting in a male-biased sex ratio that ranges from 1.5:1 (Olson et al. 1986) to 20:1 (Muths and Nanjappa 2005). The high energy costs of reproduction appear to limit most females to breed once in their lifetimes (Olson 1988). In the wild, males may live to 11 years and females to 9 years of age in Colorado and Oregon (Campbell 1970; Olson 1988; Carey 1993). In Alberta, female toads from pasture and forest populations reached 6 and 8 years of age, respectively, whereas males from both populations reached 6 years (Paszkowski pers. comm. 2012).
Generation time
Generation time is the average age of parents of a cohort. Using average age at maturity and maximum ages of Western Toads compiled from the literature, the generation time for males is (11-3.5) = 7.5 years, and for females (9-5) = 4 years, or an average of = 5.75 years.
Physiology and adaptability
Like other amphibians, Western Toads are ectotherms, exchanging heat with their surroundings rather than producing body heat internally. Voluntary thermal minima and maxima are 3.0°C and 29.5°C, respectively (Brattstrom 1963; Davis 2000). Western Toads thermoregulate behaviourally, by moving to habitats or micro-sites with suitable ambient temperatures, by basking, and by evaporative cooling from the skin and lungs (Stebbins and Cohen 1995). This wide thermal tolerance allows toads to exploit a wide range of habitats.
Western Toads are moderately resistant to desiccation with a relatively dry, thick and “warty” skin (Stebbins and Cohen 1995). Western Toads reached a critical activity point, losing the ability to right themselves, when dehydrated to 41.4% of their initial hydrated body mass (Hillman 1980). Western Toads are frequently found far from standing water in relatively xeric habitats, but moist micro-sites are required for rehydration by absorbing moisture through the pelvic patch. Metamorphs and smaller juveniles have a higher surface area to volume ratio than adults and are more vulnerable to desiccation (Livo 1998).
Dispersal and migration
Western Toads occupy home ranges and show strong fidelity to breeding sites, summer foraging areas, and probably hibernacula (implied by limited availability and communal use of hibernating sites; Browne and Paszkowski 2010a). Micro-sites providing thermal or protective cover and moist soil patches are used repeatedly (Carpenter 1954; Jones and Goettl 1998; Davis 2000; Bartelt et al. 2004). Summer home ranges were 0.1 ha or smaller on Vancouver Island (Davis 2000). Muths (2003) found that mean home ranges of males in Colorado were 0.58 km² (maximum 2.64 km²), while those of females were 2.46 km² (maximum 7.02 km²). The greatest seasonal movement between a breeding site and summer home range by a male was 0.97 km and 2.3 km by a female, similar to 0.94 and 2.44 km, respectively, found by Bartelt et al. (2004) in Idaho. Adams et al. (2005b) reported that Western Toads used streams to move within their home ranges in Montana. The largest along-stream movement was 1.5 km in 6 days, and maximum movement rates were 500 m/day. In Alberta, Western Toads moved overland at rates of 782 m in two days (Browne et al. 2004).
Western Toads are capable of directional long-distance dispersal movements (up to 7.2 km in <24 hours in spring on Vancouver Island; Davis pers. comm. 2004). Schmetterling and Young (2008) documented movements up to 13 km. Adult Western Toads are frequently found across the landscape, often far from known breeding sites. Habitats and landscapes that are unsuitable for foraging or dispersal include xeric uplands and high alpine passes (Haines Triangle, northern British Columbia, Cannings pers. comm. 2010; Northern Rockies, British Columbia, Barichello pers. comm. 2010). Recent clearcuts <5 ha were not impediments to movements by toads in spring (Deguise and Richardson 2009a). Larger clearcuts, and smaller clearcuts later in summer when temperatures are relatively high, may be inhospitable to Western Toads (Deguise and Richardson 2009a). In studies in northwestern British Columbia (Mennell and Slough 1998; Slough 2004, 2005), adult toads were found along lake and stream corridors and in upland sites throughout the region up to 30 kmfrom known breeding sites.
Movements and habitat use by juvenile Western Toads are poorly known. Bull (2009) studied dispersal by metamorphs and yearling Western Toads in Oregon and found that metamorphs travelled ≤ 2.7 km from breeding sites within 8 weeks of metamorphosis at an average rate of 84 m/d. Drainages were used as dispersal corridors. Yearling juveniles were found 1.1-2.7 km from breeding sites. Dispersal distances were limited by the availability of moist habitats and by time between metamorphosis and hibernation. Davis (2000) found metamorphs within 300 m of breeding sites on Vancouver Island. Western toads have also been observed using grass/dirt roads and seismic lines, which facilitate movements, in Alberta (Long pers. comm. 2012).
In the southern part of their range in Canada, Western Toads are nocturnal (Davis 2000), but at higher elevations and higher latitudes (where nights are shorter in summer), they are diurnal (Carey 1978; Russell and Bauer 2000). Activity patterns are likely behavioural adaptations to ambient temperatures because some toads switch between diurnal activity in spring, to nocturnal activity in summer, and back to diurnal activity in fall (Sullivan 1994).
Interspecific interactions
Western Toad tadpoles eat filamentous algae and organic detritus and opportunistically scavenge carrion. Adult toads are primarily ambush predators and feed on a wide variety of invertebrates, including worms (Clitellata), slugs (Gastropoda), spiders (Arachnida), bees (Hymenoptera), beetles (Coleoptera), sow bugs (Isopoda), grasshoppers (Orthoptera), caddisflies (Trichoptera), moths and butterflies (Lepidoptera), (Diptera), true bugs (Hemiptera), and ants (Formicidae) (Sullivan 1994; Jones et al. 2005; Bull 2006; Bull and Hayes 2009).
Egg predators are largely unknown. In the family Bufonidae, antipredator bufotoxin is secondarily deposited in the eggs before it develops, although the concentration diminishes over time (Brodie et al. 1978). Western Toads may avoid spawning in wetlands that are dominated by Wood Frogs (Lithobates sylvatica) to reduce predation on eggs and larvae (Long pers. comm. 2012). Western Toad tadpoles have been known to resort to cannibalism in times of stress during times of high larval densities and scarce food resources associated with low water levels (Jordon et al. 2004).
Western Toad tadpoles possess bufotoxins and are unpalatable to fish (Formanowicz and Brodie 1982) and newts (Salamandridae); however they are consumed by birds including Common Ravens (Corvus corax) and crows (Corvus spp.), Spotted Sandpipers (Actitis macularius), Mallards (Anas platyrhynchos) and likely other species of waterfowl, Great Blue Herons (Ardea herodias), reptiles (e.g., gartersnakes, Thamnophis spp.), amphibians (Bullfrogs, Lithobates catesbeianus) and invertebrates (backswimmers, Notonecta spp.; Giant Waterbugs, Lethocerus americanus) (Olson 1989; Jones et al. 2005). Western Toad abundance was not affected by the presence of native or non-native fish in the foothills of Alberta (Schank 2008). McGarvie Hirner and Cox (2007) found higher Western Toad abundance in lakes with Rainbow Trout (Oncorhynchus mykiss) than in lakes without trout in the southern interior of British Columbia. The presence of fish may help to depress the abundance of invertebrate tadpole predators (Eaton et al. 2005; McGarvie and Cox 2007). Predator recognition is based primarily on chemical cues given off by the predators and recognized by the tadpoles (Kiesecker et al. 1996). Anti-predator behaviours include foraging in aggregations (Brodie and Formanowicz 1987), decreased movements and the use of shelter (Kiesecker et al. 1996). In Colorado predaceous diving beetle larvae (Dytiscus spp.) had a greater impact on Western Toad tadpoles than did adult beetles, Western Tiger Salamander larvae (Ambystoma mavortium) or Western Terrestrial Gartersnakes (Thamnophis elegans). Predaceous diving beetles occur throughout the range of the Western Toad (Livo 1998). Western Toad larvae occasionally practise cannibalism and predation on other anuran larvae (Jordan et al.2004).
Western Toads are particularly vulnerable to predation when they are transforming or newly metamorphosed, and in post-metamorphic aggregations; they are taken by birds (Gyug 1996) and Gartersnakes at this time (Davis 2000; Wind unpubl. data 2002). Pearl (2002) observed adult Columbia Spotted Frogs (Rana luteiventris) preying on Western Toad tadpoles and metamorphs in Oregon. Synchronous metamorphosis and aggregation may be anti-predator adaptations and satiate predators during this vulnerable period. Devito et al. (1998) found that metamorphosis occurred earlier (and at a smaller size) when larval density was higher, and more synchronously in the presence of a predator, the Common Gartersnake (T. sirtalis). Emergence at a smaller size may compromise long-term survival (Berven 1990).
Adult and metamorphosed juvenile Western Toads excrete bufotoxin, among other amines and alkaloids, from their parotoid glands to deter predators; however, many species of birds, snakes, mammals, and amphibians such as corvids, Gartersnakes, Coyotes (Canis latrans), Raccoons (Procyon lotor), Skunks (Striped Skunk, Mephitis mephitis, and Western Spotted Skunk, Spilogale gracilis), and Red Foxes (Vulpes vulpes), prey on them (Olson 1989; Jones et al.2005). Many species eviscerate toads to avoid their toxic skin. The highest predation pressure on adult toads occurs at breeding aggregations when they are exposed in shallow water (Olson 1988). Juvenile Western Toads avoided chemical cues from snakes fed juvenile toads (Belden et al.2000), demonstrating the importance of the diet of predators in mediating anti-predator behaviour.
Eaton et al. (2008) reported the infection of two adult and 10 juvenile Western Toads in boreal Alberta by parasitic green blowfly (Lucilia silvarum (Calliphoridae)) larvae. One adult toad survived myiasis. The parasite was detected in 2 of 4 years of observation, and maximum rate of infection was 12.8% of juveniles.
Population sizes and trends
Sampling effort and methods
There have been few dedicated population surveys for Western Toads over most of their range (Table 1). Most observations have been collected opportunistically and relatively few sites have been revisited to determine population persistence or trends.
| Agency or person | Site or area | Population trends/status | Comments |
|---|---|---|---|
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
Rocky Mountain National Army Cadet Summer Training Centre, Alta. | Unknown | Breeding site, 2009. |
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
Slesse Creek Demolition Area, B.C. | Unknown | Present. |
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
Columbia Valley, B.C. | Unknown | Present, probable breeding site, 2008. |
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
Chilcotin Training Area, B.C. | Unknown | Breeding site, 2009. |
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
CFS Holberg, B.C. | Unknown | Unconfirmed. |
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
CFB Esquimalt, B.C. | Unknown | Present. |
| DND (Nernberg unpubl. data 2011; Hawkes et al. 2009, 2010) |
CFS Leitrum Detachment, Masset, B.C. | Unknown | Present. |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Banff National Park, Alta. | Vulnerable | State of the Park Report in 2008 states “fair and stable” for all amphibians in the park, utilizing the same data but a different assessment method (see Lepitzki 2008 for compendium of data in the park for the Western Toad). |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Cave and Basin National Historic Site, Alta. | Unknown | Present in sections of the site, including hot spring pools (Lepitzki pers. comm.2012) |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Ya-Ha-Tinda Ranch, Banff National Park, Alta. | Unknown | Potential, but unconfirmed. |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Elk Island National Park, Alta. | Vulnerable | Species was first reported in 1999 (Paszkowski pers. comm. 2012). Seems to have replaced CDN toad in parks in previous 10 years. It occurred in about 33% of wetlands sampled. |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Jasper National Park, Alta. | Apparently Secure | Ongoing monitoring program. Occupancy of sites remained stable 2007-2011 (Shepherd and Hughson 2012). |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Waterton Lakes National Park, Alta. | Vulnerable | Uncommon in park. 10% of historical sites were occupied in 2003. Population stable 1998-2011, but absent from 3 early 1990s breeding sites (Taylor and Smith 2003; Johnston and Crowshoe 2011). |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Nahanni National Park, NT | Unknown | Potential, but unconfirmed. |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Chilkoot Trail National Historic Site, B.C. | Imperiled | Breeding sites used consistently for >20 years were monitored in 2004 (Slough 2004) and remain active to 2011 (Rivard pers. comm. 2011). Bd swabs negative in 2008, 2009 (Wong unpubl. data 2011). |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Fort Langley National Historic Site, B.C. | Unknown | |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Glacier National Park, B.C. | Vulnerable-Apparently Secure / population stable | See Mount Revelstoke National Park. |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Gulf Islands National Park Reserve, B.C. | Not Assessed | Potential, but unconfirmed. |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Gwaii Haanas National Park Reserve and Haida Heritage Site, B.C. | Vulnerable-Apparently Secure | At least 6 breeding sites and 14 other sites were monitored 2005-2011 (Wojtaszek unpubl. data 2011) |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Kootenay National Park, B.C. | Apparently Secure | |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Mount Revelstoke National Park, B.C. | Imperiled-Vulnerable / population stable | Recorded at 30 sites in Mount Revelstoke and Glacier National Parks in 1982-83 and 24 sites in 2003-04. Breeding at 15 sites in both periods (Van Tighem and Gyug 1984; Dykstra 2004; Adama and Ohanjanian 2005). |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Pacific Rim National Park Reserve, B.C. | Critically Imperiled-Imperiled | Breeding in 2 of 25 ponds surveyed 2009-2011 (Beasley unpubl. data 2011). |
| Parks Canada (Parks Canada Agency 2009 – 2011; data provided by Howes pers. comm. 2011, 2012) |
Yoho National Park, B.C. | Vulnerable-Apparently Secure | |
| Alberta Wilkinson and Hanus (2003) |
Trapping at 5 sites, 179 ponds at 8 sites within Western Toad range | Unknown | Western Toads were found in 34 ponds, in all areas except Banff National Park and east of the park. Other surveys have confirmed the species from Banff National Park. |
| Wilkinson and Berg (2006) | Trapping at 4 sites, 101 ponds searched at 3 sites | Increasing | Large population fluctuations 1997-2005, but persistence at breeding sites increased 2001-2005. |
| Eaves (2004) | Beaver Hills, east of Edmonton | Increasing | May be colonizing the aspen parklands. |
| Schank (2008) | 11 lakes in boreal foothills, near Rocky Mountain House | Stable | Toads called on all 11 lakes annually; toadlets found on 18-54% of lakes, 2005-07. |
| British Columbia: Beasley unpubl. data (2011) |
Clayoquot Sound | Unknown | Breeding in 2 of 148 ponds surveyed in 1998. |
| British Columbia: Beasley unpubl. data (2011) |
Rae Lake, Hesquilaht Harbour | Apparently Stable | Breeding from 2005 to present. |
| British Columbia: Beasley unpubl. data (2011) |
Frederick and Rousseau lakes, near Bamfield | Unknown | Breeding sites 2009-10. |
| British Columbia: Beasley unpubl. data (2011) |
Penrose Island, near Rivers Inlet | Unknown | Breeding in wetland 2011. |
| Kinsey unpubl. data (2011) | Prince George and Peace River forest districts | Apparently Stable | Auditory, road and systematic pond surveys, Prince George 1996-97, Dawson Creek 2006-09. |
| Wind unpubl. data (2011) | 28 breeding sites identified in southern B.C. and Vancouver Island | Apparently stable | 2001-2011; may be declining at Eva Lake, Whistler, where no breeding was observed in 2009. |
| Wind unpubl. data (2011) | Vancouver Island, Morrell Lake, Nanaimo | Apparently stable | Breeding monitored 2006-2011. |
| Thompson unpubl. data (2011) | Pond in Prince George city park | Unknown | Mass mortality of toadlets due to trampling by trail users. |
| Tayless (2011) | Lost Lake, Whistler | Apparently stable | Monitored 2005-2011. Road and trail mortality a problem. Toadlets transported across road and trail in 2007. |
| Ovaska, Nicola Naturalist Society and BC FrogWatch, (Ovaska et al.2011 and unpubl. data 2012) | Thompson-Nicola area | Widespread (found in 20 of 42 10x10 km grid cells), but population sizes and trends are unknown | Monitoring of toad breeding sites initiated in 2011 as part of a community-based amphibian monitoring program by Nicola Naturalists Society. 93 wetlands and wetland complexes were surveyed in 2011-2012; 16 breeding sites were identified. Road mortality is a problem at least at two of these sites. |
| Dulisse (2007) | Near Kaslo | Unknown | No breeding sites located. |
| Dulisse and Hausleitner (2009) | West Kootenay | Unknown | 9 breeding sites located. |
| Dulisse and Hausleitner (2010) | Columbia Forest District | Unknown | 12 breeding sites located. |
| Ohanjanian and Beaucher (2000); Dulisse et al. (2011) | Summit Lake | Stable | 6 breeding sites on lake. |
| RESCAN (2008) | Near Smithers | Unknown | 1 breeding site identified. |
| Fraker and Hawkes (2000); Hawkes et al. (2006) | Peace River, from Lynx Creek to Alberta border | Stable | 1 breeding site identified, overall population was “healthy”. |
| Hawkes and Tuttle (2010); Hawkes et al. (2011) | Kinbasket and Arrow reservoirs | Stable or increasing | 16 breeding sites identified, 6 sites with 10-20,000 tadpoles at Kinbasket, over ½ million tadpoles at Arrow reservoir. |
| Hengeveld (1999) | Williston Reservoir watershed | Unknown | Breeding sites identified. |
| Ohanjanian et al.(2006) | East Kootenay | Possible decline | 11 breeding sites found, 7 of 23 historic breeding sites occupied. |
| Rach (2008) | Taku River | Unknown | 6 breeding sites located. |
| Rescan Tahltan Environmental Consultants (2008) | Schaft Creek, near Telegraph Creek | Unknown | 5 breeding sites located. |
| Slough (2009); Slough unpubl.data | Atlin Warm Springs | Declining | Consistent breeding 1924 to 2005; however, only limited breeding two years between 2006 and 2012 (in 2008 and 2012). |
| Slough unpubl. data | Near Griffith Island, Atlin Lake | Unknown | Tadpoles here in 2001, 2003, not in 2007. |
| Mennell and Slough (1998); Slough unpubl. data | Tagish Lake | Stable | 2 large breeding sites at Swanson River (2001) and Fantail River (1998). |
| Slough unpubl. data | Tutshi Lake | Stable | Consistent breeding at Tutshi River estuary 2005, 2007 and 2009. |
| Yukon: Slough (2005); Slough and Mennell (2006); Slough unpubl. data |
Coal River and Coal River Springs | Stable | Consistent reproduction at Coal River Springs at least between 1977 and 2008. Also at least 5 other breeding sites in vicinity on Coal River. |
Table summarizing Western Toad population status, trends, and monitoring information at various locations grouped by jurisdication: Department of National Defence, Parks Canada, Alberta, British Columbia, and Yukon.
The greatest effort to monitor amphibian populations has been in Alberta, where the RANA program (Researching Amphibian Numbers in Alberta) has been monitoring amphibians at up to 7 sites per year since 1997 (Wilkinson and Berg 2006). Methods include pitfall trapping, pond surveys, and road transects (calling surveys). Pond surveys typically use visual encounter surveys for the various life stages, which involve the searching and dipnetting of terrestrial, semi-terrestrial (wetland) and aquatic habitats (Thoms et al. 1997). Calling surveys are effective for detecting Western Toads in much of Alberta, where the species uses advertisement calls during the breeding season (Pauly 2008; Long 2010).
The British Columbia Ministry of Environment (2009) initiated a Western Toad population monitoring program with a pilot study in 2009. Efforts have been restricted to a few sites on Vancouver Island and in the south of the province. Methods include call surveys (which are not effective for Western Toads in British Columbia, where toads do not produce true advertisement calls) and visual encounter surveys. This program relies on volunteers and does not employ expert observers as in the Alberta RANA program. Most program results were not available for this status report; however, some individual participants provided data (Beasley unpubl. data 2011; Kinsey unpubl. data 2011; Thompson unpubl. data 2011; Wind unpubl. data 2011). Reports are available for the Whistler (Tayless 2011) and Merritt (Ovaska et al. 2011) areas.
There are no organized efforts to monitor Western Toads in either Yukon or Northwest Territories, although some sites in Yukon and northern British Columbia have been systematically resurveyed by volunteers (Slough 2009, 2012).
Parks Canada maintains presence/absence type of data and current population status assessments for species at risk such as the Western Toad in most parks and park reserves (Table 1). Breeding sites have been monitored in Gwaii Haanas National Park Reserve and Haida Heritage Site since 2005 (Wojtaszek unpubl. data 2011). Breeding sites in the Chilkoot Trail National Historic Site (Slough 2004) are monitored on ad hoc basis by park wardens (Rivard pers. comm.2011). Western Toads in Mount Revelstoke and Glacier National Parks were monitored at historical breeding sites (1982-1983) in 2003-2004 (Adama and Ohanjanian 2005). Western Toads have been monitored in Waterton Lakes National Park since 1998 (Taylor and Smith 2003; Johnston and Crowshoe 2011). Amphibian monitoring has been carried out in Banff National Park, where distributional data from various programs and volunteer observations have been compiled into a BNP Microsoft Access® Herptile Database (Lepitzki 2008). Monitoring of Western Toad abundance at selected wetlands was conducted as part of the Banff-Bow Valley Amphibian Monitoring Program (2001 – 2005) and of the Fairholme Bench Ecologically Sensitive Site Amphibian project (2004 – 2006), which examined the effects of prescribed fire and thinning on amphibian abundance and distribution (Lepitzki 2008). Amphibian monitoring is also ongoing at Elk Island National Park (since 1999; Paszkowski pers. comm. 2012) and Jasper National Park (Shepherd and Hughson 2012).
The Department of National Defence monitors Western Toad occurrences and has conducted breeding site surveys on DND lands in British Columbia and Alberta in 2009 (Hawkes et al. 2009, 2010). BC Hydro has monitored Western Toad populations in the East and West Kootenays as part of its Columbia Basin Fish and Wildlife Compensation Program (Ohanjanian et al. 2006; Dulisse and Hausleitner 2009, 2010; Dulisse et al. 2011) and environmental monitoring projects (Hengeveld 1999; Hawkes et al. 2006; Hawkes et al.2011). Field studies for BC Hydro generally involved time-constrained searches (visual encounter surveys) conducted at regular intervals from the breeding season to the end of summer.
Abundance
The Western Toad is apparently widespread, abundant and persistent across a large portion of its Canadian range (Table 1), but little information exists on population sizes or densities, and few populations have been systematically monitored. Large breeding populations, tadpole aggregations, and post-metamorphic aggregations are frequently reported. Aggregations of metamorphs are often reported in the 10s to 100s of thousands (e.g., Hawkes and Tuttle 2010; Hawkes et al. 2011). It should be noted, however, that the propensity of Western Toads to gather at breeding sites, often from a wide area, can give an exaggerated impression of abundance. Furthermore, in anurans, tadpole and metamorph abundance is a poor indicator of adult population size because of fluctuations in breeding success and high early mortality rates. Numerous extant breeding populations have been reported from the south coast of British Columbia (Beasley unpubl. data 2011; Tayless 2011; Wind unpubl. data 2011), an area where numbers were declining (COSEWIC 2002). Nevertheless, the number of breeding sites is believed to have declined in the past 20 years. The Western Toad appears to be not as abundant north of 58°N where aggregations of tadpoles and metamorphs have been reported only in the hundreds or thousands (Slough 2004, 2005, 2009). Large aggregations have been observed near Telegraph Creek, British Columbia, 57° 55’N (Slough, unpubl.data 2001) and at Liard Hot Springs, British Columbia (Bennett, pers. comm. 2012).
Fluctuations and trends
Population trends are unknown for most of the species’ Canadian range. Monitoring has been initiated in a number of areas, but few data are available to date or the time-series are too short to draw reliable conclusions (Table 1). A local extirpation has been documented from a large wetland complex, Jordan Meadows, on southern Vancouver Island (Davis and Gregory 2003). The extirpation has several potential causes, most of which are anthropogenic in nature. Habitat fragmentation, road mortality, pollution, and the introduction of Saprolegnia fungus (during fish stocking) were proposed causes (Davis and Gregory 2003), in addition to the more global causes of amphibian declines such as UV-B radiation and chytrid fungus. The Atlin Warm Springs population has been documented by numerous observations since 1924, but the species was absent in 5 of 7 years since 2005, suggesting temporary extirpation followed by recolonization (Slough 2009; Slough unpubl. data 2010 – 2012). Disease caused by chytrid fungus is a potential cause of the decline (see Threats and Limiting Factors). Additional population declines have been documented from the Lower Fraser Valley, most of which are associated with local anthropogenic habitat loss in heavily settled rural and urban areas (e.g., Haycock 1998 and review in COSEWIC 2002) and from the East Kootenays (Ohanjanian et al.2006). Ohanjanian et al.(2006) visited 87 wetlands known to have Western Toads historically and found Western Toads breeding at less than a third of these sites.
In Alberta, Western Toads may have declined in Waterton Lakes National Park between 1998 and 2003 but appear to have remained relatively stable since (Taylor and Smith 2003; Johnston and Crowshoe 2011). During surveys of 120 historical and potential breeding sites in 2003, only 10% of the historical sites were occupied. From 1998 – 2011, Western Toads have been recorded at 14 of 20 long-term amphibian monitoring sites. Other recent reports and unpublished data indicate that the Western Toad populations studied might be stable or even increasing in some areas of Alberta (Table 1; Eaves 2004; Wilkinson and Berg 2006; Paszkowski pers. comm. 2012). There are reports of a possible population expansion in the aspen parklands (Eaves 2004), possibly at the expense of the Canadian Toad (Browne et al. 2003; Paszkowski pers. comm.2012).
Detecting trends in amphibian numbers requires long-term data, because amphibian populations are characterized by inherent fluctuations and are vulnerable to stochastic events (Marsh and Trenham 2001). Single surveys can greatly bias apparent trends (Skelly et al. 2003), and restricting surveys to historical breeding sites does not distinguish between population losses and site switching (Petranka et al. 2004; Pearl et al. 2009b), or the occupation of new habitats (Wente et al.2005).
Western Toad populations in the U.S. are severely threatened by activities that destroy or modify habitat including timber management, livestock grazing, pesticide application, water management, recreation, residential and commercial development, and road building (Center for Biological Diversity et al. 2011). Chytridiomycosis has devastated populations in the southern Rocky Mountains and is also considered a threat elsewhere in the U.S.
Rescue effect
The Canadian population of Western Toads could be rescued by dispersal of toads from Montana, Idaho, Washington and Alaska. Immigrants from the U.S. would be expected to survive in Canada as environmental and habitat features are largely similar. However, threats to Western Toad populations in the U.S. are similar, and these populations may be experiencing declines that exceed those in Canada, reducing the likelihood and rate of rescue.
Threats and limiting factors
Amphibians face many threats globally and are declining more rapidly than either birds or mammals (Stuart et al. 2004). The main threats to the Western Toad in Canada are the emerging infectious disease chytridiomycosis and habitat fragmentation and mortality associated with roads. Habitat loss due to logging, agriculture and the oil and gas industry is also of concern.
The IUCN Threats Calculator was applied to the Non-calling and Calling populations of the Western Toad separately as part of the preparation of this status report (Appendices 2 and 3). The assessment built on previous identification of threats in management plans for the species by the Provincial Western Toad Working Group (2011) and by Environment Canada (2011). The overall threat impact ranged from high to medium for the Non-calling population and from very high to medium for the Calling population.
Threats with the greatest levels of concern are summarized below. Apart from emerging diseases, most of these threats are localized to human population centres and agricultural areas, where habitat loss and degradation are issues, and to areas of intensive resource development, where habitat fragmentation is of main concern.
Emerging diseases: Chytridiomycosis
A global threat to the Western Toad in Canada is the emerging infectious disease chytridiomycosis, caused by the fungus Batrachochytrium dedrobatidis(Bd). First described in the late 1990s (Berger et al. 1998; Longcore et al. 1999), the pathogen has been found in Western Toads across the Canadian range of the species from Vancouver Island and southern British Columbia, to the northern range limit in northern British Columbia, Alberta, Yukon, and Northwest Territories (Raverty and Reynolds 2001; Adams et al. 2007; Deguise and Richardson 2009b; Schock et al. 2009; Slough 2009; Govindarajulu unpubl. data 2011; Stevens et al. 2012). Bd has also been isolated from numerous sympatric native anurans and salamanders and from introduced anurans during these surveys. The occurrence of Bd is widespread but patchy (Govindarajulu unpubl.data 2011). Slough (2009) reported Bd -positive samples at 6 of 9 sites in northwestern BC, and Schock et al. (2009) found chytrid fungus at 1 of over 20 sites in Northwest Territories. Western Toads tested positive at 8 of 16 sites in Alberta (Stevens et al. 2012); however, other amphibian species tested positive at most of the negative sites, suggesting that Western Toads are widely exposed to Bd.
Bd in Western Toads (from Colorado) and Bullfrogs (from Quebec and California) from North America has been identified as a hyper-virulent recombinant lineage that resulted from the anthropogenic mixing of two other lineages and subsequent anthropogenic spread (probably through global trade in amphibians) across North America, Central America, Caribbean, Australia and Europe (Farrer et al. 2011). It may require other environmental co-stressors, such as increased UV-B radiation, for the disease to become pathogenic. Diseased toads are rarely reported; however, rapid mortality may quickly remove them from the environment. The disease has been linked to Western Toad population declines in the U.S. (Kiesecker and Blaustein 1995; Daszak et al.1999; Kiesecker et al. 2001a; Muths et al. 2003). However, population monitoring and Bd testing have rarely, if at all, been conducted concurrently. Nonetheless, Western Toad population declines have been noted at the Atlin Warm Springs in northern British Columbia, and Bd was isolated from nearby populations; no toads were found at the springs during Bd sampling in the area (Slough 2009).
There is concern that humans may be agents of Bd transmission between wetland sites on recreational gear such as waders and research equipment (British Columbia Ministry of Environment 2008; Mendez et al. 2008; Vredenburg et al. 2010). If this is true, then increased access across the Western Toads range will facilitate the spread of the pathogen.
There is some evidence that some amphibian species are able to survive Bdepidemics, either through increased immunity which allows them to live with low-level fungal infections, the evolution of less pathogenic strains of Bd, environmental conditions, infection intensities (Briggs et al.2010), or through adaptation and the evolution of better defences (such as anti-microbial peptides shown to be released by the skin of 4 Australian anurans; Woodhams et al. 2010). While there is evidence that Bdis a spreading pathogen that can have negative consequences for amphibian populations (Skerratt et al.2007), there is also evidence that Bd is widespread in areas where there is little evidence of harm (Longcore et al. 2007; Pearl et al. 2007) or where Bd has become endemic in apparently stabilized populations (Ouellet et al. 2005; Pearl et al. 2009a,b; Pilliod et al. 2010). Most Western Toad populations in Colorado became extinct in the late 1970s through early 1980s (Carey 1993), but their continued survival in relict populations is not believed to be due to increased immunity, because these populations remain susceptible to the pathogen (Carey et al. 2006). Past and future declines of Western Toads due to Bd cannot be ruled out. However, at this time there is no evidence of a wavelike spread of the pathogen and population declines northward, as feared might be the case at the time of the previous status assessment for the Western Toad.
In laboratory experiments, Western Toad tadpoles exposed to Bd experienced increased mortality and lethargy as compared with unexposed control tadpoles (Blaustein et al. 2005b). Infection must reach a threshold number of zoospores before death results (Carey et al. 2006). Adult females are less likely to reach this threshold because they spend less time at aquatic breeding sites than males (where they can be re-infected through contact with water containing Bd zoospores), they do not breed every year, and they are more likely to hibernate as solitary individuals (Carey et al. 2006).
Other co-stressors such as habitat degradation, climate change, Saprolegnia, and increased UV-B may act synergistically to cause immunosuppression and vulnerability to Bd (Carey 1993; Kiesecker and Blaustein 1995; Kiesecker et al. 2001a). As climate warming progresses, outbreaks of chytriomycosis could increase at northern latitudes. Muths et al. (2008) found that maximum daily temperatures explained much of the variation in Bd occurrence in Western Toad populations in the U.S. Rocky Mountains and suggested that low temperatures at higher latitudes and altitudes may limit Bd occurrence at present.
Emerging diseases: Ranavirus
Ranavirus (Family: Iridoviridae) was detected in Wood Frogs but not in Western Toads in Northwest Territories (Schock et al. 2009). Infections and mortalities due to ranavirus have been reported for Western Toads in captive and wild populations (Miller et al. 2011). Co-stressors may have an effect on ranavirus dynamics as well (Gray et al.2009).
Invasive species
While Western Toads are unpalatable to most fish species due to skin toxins (Kats et al.1988), they are threatened by diseases associated with stocked fish, which are not screened for prior to fish release. Examples are the bacterium Aeromonas(Carey 1993), the fungus Saprolegnia (Blaustein et al. 1994b; Kiesecker and Blaustein 1997; Kiesecker et al. 2001b), and ranavirus (Miller et al. 2011).
The introduced Bullfrog in southwestern British Columbia is a direct threat as a predator and may act as a reservoir for Bd. Racoons (Procyon lotor) have been introduced to Haida Gwaii and have apparently depleted some Western Toad populations (Reimchen 1991). Northern Pacific Tree Frogs (Pseudacris regilla) were also introduced there and may adversely affect the Western Toad through competition for breeding sites or changes to the ecosystem (Reimchen 1991). Predators that have increased in abundance in human-modified landscapes, such as Raccoons, rats (Rattus spp.), and Common Ravens, may be a threat.
Transportation corridors
Roads and other transportation corridors are a threat to Western Toads. Roads remove woodland habitat and fragment habitats, but more importantly they represent barriers to dispersal and migration and result in road kill (Fahrig et al. 1995; Eigenbrod et al. 2008). Adults and toadlets are vulnerable near breeding, foraging and hibernation sites, especially during mass movement events (Carr and Fahrig 2001). Recent mass mortality events have been reported for toadlets and adults at several areas in British Columbia: near Prince George (Thompson pers. comm. 2011), Dawson Creek (Kinsey pers. comm. 2011), Chilliwack (Clegg 2011), and Summit Lake (Dulisse et al.2011). The 1998 mass mortality of toadlets on the edge of the Stewart-Cassiar Highway in northern British Columbia (near Bell-Irving River) was attributed to extreme heat at the time of post-metamorphic aggregation and not to road mortality (M. Inniss unpubl. manuscript, reproduced in COSEWIC 2002). Under-road amphibian passages have been installed to alleviate amphibian mortality and are used by Western Toads in Waterton Lakes National Park, AB (Pagnucco et al.2011). Amphibian tunnel systems have also been installed on Vancouver Island, where a highway intersects a wetland near Courtney, and on an amphibian migration route near Qualicum (Fitzgibbon 2001). However, mitigation has been undertaken at a minute proportion of sites where roadkill is a problem.
Habitat loss and degradation
Factors affecting Western Toad habitat loss, alteration and fragmentation were discussed in Habitat trends. A synopsis follows.
Loss of aquatic and terrestrial habitat due to residential developments is ongoing in localized areas around population centres in the southwestern and southern interior of British Columbia and Calgary, Edmonton, and Peace River areas of Alberta. Western Toads are largely intolerant of urbanization, and declines in the Lower Fraser Valley in British Columbia have been attributed mainly to habitat loss (COSEWIC 2002).
Habitat loss also occurs in association with agricultural developments, as wetlands are drained or degraded, and surrounding terrestrial habitat is modified and may become unsuitable for foraging or hibernation. Row crops, in particular, provide poor habitat for toads (Eaves 2004). Conversion of land to agricultural uses is largely historical, but where agricultural areas are expanding, such as in the Peace River region in British Columbia and Alberta and in Aspen Parkland in central Alberta, or where the use of existing areas is intensified, habitat loss is of concern.
Although significant to local populations, direct habitat loss from residential and agricultural developments is considered a low threat to the overall population of toads because of their relatively small spatial extent.
Some of the habitat threats are more significant in Alberta where their footprint on the landscape is much greater. The oil and gas industry (drilling) and agriculture (row crops) are major land uses that pose threats to Western Toad habitat. Impacts of oil and gas extraction include ecosystem conversion, fragmentation, and environmental contamination. Secondary impacts include the construction of roads and seismic lines. It is these secondary impacts associated with development and resource extraction that pose greatest threats to Western Toad populations due to disruption of migration and dispersal corridors and the potential for mass mortality events.
Pollution
While amphibians are known to be sensitive to chemical contaminants from industrial wastes, agricultural effluents and other sources that are transported by water and air, very little research has been undertaken on the effects of specific contaminants on the Western Toad. The effects of contaminants on amphibians in general are reviewed by Bishop (1992) and Biolinx Environmental Research Ltd. and E. Wind Consulting (2004).
The acidification of wetlands from airborne sources may be a source of developmental abnormalities and increased mortality of embryos and larvae (Vertucci and Corn 1996). Acidification from airborne sulphur is associated with oil and gas extraction in northeast British Columbia and Alberta (Austin et al. 2008). Heavy metals are also transported by air. UV-Bradiation may not be a serious threat to Western Toads (see Ultraviolet Radiationsubsection below), but heavy metals and UV radiation may act synergistically with other environmental stressors and depress the immune system of Western Toads, making them vulnerable to pathogens (Carey 1993). Heavy metals including zinc, cadmium and copper can have negative effects on amphibian growth, development and survival (Glooschenko et al. 1992; Brinkman 1998).
Western Toad larvae are vulnerable to rotenone-containing piscicides, as the lethal dose for fish and amphibians overlaps (Fontenot et al. 1994). Bishop (1992) reported that amphibians are vulnerable to other environmental contaminants including pesticides, herbicides and fertilizers. The pesticide malathion kills the plankton that tadpoles feed on (Relyea and Diecks 2008). Many compounds such as atrazine, DDT, dieldrin, and acids cause immunosuppression in amphibians in low concentrations. Atrazine can disrupt sexual development (Hayes 2004). Air transported organochlorine compounds were in higher concentrations at higher elevations in the Canadian Rockies, and most were associated with decreased growth rate and age of trout species (Demers et al. 2007).
Eutrophication of water from fertilizers and livestock waste causes algal blooms and lowers dissolved oxygen levels. Agricultural runoffs can also cause limb deformities (Kiesecker 2002) and mortality (Rouse et al. 1999) in amphibians. Exposure of Western Toad metamorphs to a forest fertilizer, urea, resulted in reduced prey consumption and significant increased mortality (Hatch et al. 2001). Glyphosate herbicides used for conifer release in British Columbia also produce lethal sublethal effects on amphibians (Govindarajulu 2008). Road salts have lethal and sublethal effects on amphibians (Harfenist et al. 1989). Sublethal effects on Wood Frogs included reduced tadpole activity and weight, and physical abnormalities (Sanzo and Hecnar 2005).
Climate change and severe weather
The potential impacts of climate change on the Western Toad over the next decade are unknown or ambiguous at best (Ovaska 1997). However, Western Toads are adaptable to a wide range of habitats and temperature/moisture conditions; therefore climate change over the next 10-20 years may not be a serious threat to this species. Climate models predict increases in temperature and precipitation in Canada (IPCC 2007), with the largest warming projected for northern Canada. Precipitation is likely to increase in winter and spring, but decrease in summer. Snow season length is predicted to decrease, but increased snowfall will more than make up for the shorter snow season, resulting in increased snow accumulation.
Positive impacts may be earlier breeding and range expansion in the north. Corn (2003) documented earlier breeding by Western Toads in the Cascade Mountains of Oregon concurrent with reduced winter precipitation. On the other hand, a decrease in summer precipitation might increase the frequency and duration of droughts, affecting the persistence of smaller wetlands used for breeding, decrease connectivity across the landscape, and decrease the availability of moist micro-sites used for rehydration (Provincial Western Toad Working Group 2011). Increased snowpack would favour hibernating toads, whereas a diminished snow pack would lead to increased depth of ground freeze, endangering overwintering individuals (Alaska Department of Fish and Game 2006).
An increase in wildfires could result in a loss of forest habitat, though open areas preferred by Western Toads are created by wildfire (Guscio et al.2008). Hossack and Corn (2007) found that Western Toads were resistant to wildfire and benefited over the short-term, by colonizing wetlands after burns. UV-B radiation could increase, as well as the ranges of other competitors and predators. Increased maximum temperatures may decrease environmental constraints on Bd and permit it to expand into higher elevations (Muths et al. 2008). However, a warmer climate with greater amounts of atmospheric moisture would benefit toads by reducing physiological costs of crossing landscapes (Bartelt et al. 2010) and possibly reduce the incidence of Bd which can be cleared from infected individuals at elevated body temperatures (Woodhams et al. 2003).
Amphibian malformations
Amphibian malformations have received a lot of attention over the past 20 years (Ballengée and Sessions 2009). There have been a few cases of malformed Western Toads reported from British Columbia (COSEWIC 2002). The cause of the malformations may be a trematode parasite (Ribeiroia sp.) which infects developing embryos, causing multiple limbs to develop and reducing tadpole survival (Johnson et al. 1999, 2001). An aquatic snail, (Planorbella tenuis) is the first host of the parasite, and environmental changes, such as eutrophication due to organic pollution, may have caused snail populations to increase (Johnson et al. 1999). Missing limbs and other limb deformities are frequently the result of physical trauma such as failed predation attempts of tadpoles by invertebrates such as dragonfly larvae (Eaton et al. 2004; Ballangée and Sessions 2009). Malformed toadlets would have compromised dispersal abilities, and ultimately lower survival (Davis 2000).
Ultraviolet radiation
UV-B radiation has been shown to reduce the survival of Western Toad embryos and tadpoles in Oregon (Blaustein et al. 1994a; Hays et al. 1996; Blaustein et al. 2005a), whereas studies in Colorado did not detect a UV-B effect (Corn 1998). The Western Toad may be vulnerable to UV-Bradiation because it lays eggs in open shallow water subjected to solar radiation and has a poor ability to repair UV-induced DNA damage (Blaustein et al.1998). Other stressors may act in combination with UV-B to encourage infection by pathogens. For example, Western Toad embryos and larvae were susceptible to a complex interaction between UV-Bradiation, water mould (Saprolegnia spp.), and low water levels caused by lower precipitation (Kiesecker et al. 2001a). Western Toad juveniles also experience elevated mortality when exposed to ambient levels of UV-B radiation (Blaustein et al. 2005a). Garcia et al. (2006) exposed the metamorphs to a similar level of UV-B for a reduced period and found resultant mortality. UV radiation and other environmental stressors may interact synergistically to induce lethal and sublethal effects (Blaustein et al. 2003; Bancroft et al. 2008).
Adams et al. (2005a) found no effect of UV-Bradiation on Western Toad distribution in western North America. Sub-lethal effects of UV-Bradiation may include reduced anti-predator behaviour of juvenile Western Toads (Kats et al. 2000).
Number of locations
The amphibian chytrid fungus is considered the major plausible threat to the Western Toad in Canada. Although it is found throughout the species range in Canada, it is highly unlikely that this threat could rapidly affect all Western Toads. Because of the wide extent and elevational range of the species across different terrain and landscape features, it is more likely that the disease would spread from multiple centres with some areas escaping infection, resulting in an unknown number of multiple locations. Other threats affecting populations, including habitat loss and modification from various sources and road mortality, could rapidly affect populations within localized areas, resulting in an unknown but large number (100s) of locations.
Protection, status, and ranks
Legal protection and status
Amphibians are classified as wildlife under the various Wildlife Acts in British Columbia, Alberta, Northwest Territories and Yukon. Permits are required for harvesting, studying, or keeping them in captivity. Habitat is not protected under these acts.
The Western Toad was designated as a species of Special Concern in 2002 by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC 2002). It was listed on Schedule 1 of the federal Species at Risk Act (SARA), the official list of wildlife species at risk in Canada, in 2005. Under the national Accord for the Protection of Species at Risk, the federal, provincial and territorial governments agreed to develop management plans for species of Special Concern within 3 years of listing. Management plans are being prepared by Environment Canada (2011), British Columbia (Provincial Western Toad Working Group 2011), and Yukon (Yukon Department of Environment 2011).
The U.S. Fish and Wildlife Service (2005) determined that listing the Southern Rocky Mountains Population of the Western Toad, contiguous with Canadian populations, at this time is not warranted because it does not constitute a distinct population segment. The Service will continue to seek new information on threats to the species. A new petition to list this Distinct Population Segment or the Eastern Population (Southern Rocky Mountains, Utah, northeastern Nevada, and southern Idaho, based on the evolutionarily significant “Eastern clade” identified by Goebel et al. 2009) was submitted to the Secretary of the Interior, through the United States Fish and Wildlife Service 2011 (Center for Biological Diversity et al. 2011).
The Convention on International Trade in Endangered Species of Wild Flora and Fauna (CITES 2012) does not list the Western Toad.
Non-legal status and ranks
Two non-legal status ranking systems are applied by Canadian jurisdictions, the general status of species (Canadian Endangered Species Conservation Council [CESCC] 2011) and NatureServe (2012). NatureServe rankings are further simplified by grouping species into coloured ‘lists’ in British Columbia. The use of colour-coded lists was dropped in Alberta.
The general status of the Western Toad is Sensitive in Canada, Yukon, British Columbia and Alberta (Alberta Environment and Sustainable Resource Development 2010; CESCC 2011) (Table 2). The general status of the Western Toad in Northwest Territories is May be at Risk (CESCC 2011; Working Group on General Status of NWT Species 2011).
| Jurisdiction | General status rank | NatureServe rank | Other |
|---|---|---|---|
| Global (Species Range) | G4 | IUCN : Near Threatened | |
| Canada | Sensitive | N4 | |
| BC | Sensitive | S3S4 | |
| AB | Sensitive | S4 | |
| YT | Sensitive | S3 | |
| NT | May be at risk | S1S2 | |
| U.S.A. | State status | N4 | |
| AK | S3S4 | ||
| CA | S5 | ||
| ID | Protected nongame species | S3 | USFS : Sensitive (Region 1) |
| MO | Species of concern | S2 | USFS : Sensitive |
| NE | S4 | ||
| OR | Vulnerable | S3 | |
| UT | Species of concern | S2S3 | |
| WA | Candidate | S3 | |
| WY (northern) | Special Concern | S1 | |
| Southern Rocky Mountains population | T1Q | USFWS : No longer candidate for listing | |
| CO | Endangered | S1 | USFS : Sensitive |
| NM | Threatened | SH | USFS : Sensitive |
| WY (southern) | Special Concern | S1 | USFS : Sensitive (Region 2) |
Table summarizing Western Toad status ranks in Canada and the United States, nationally and by province or state. Ranks given are General Status rank, NatureServe rank, and “other.”
G=Global, N=national, S=Subnational, T=Infraspecific taxon.
H=Presumed Extirpated, 1=Critically Imperiled, 2=Imperiled, 3=Vulnerable, 4=Apparently Secure, 5= Secure.
Recent sources different from NatureServe (2012): Idaho Department of Fish and Game (2005), Washington Department of Fish and Wildlife (2008).
NatureServe rankings are G4 - Apparently Secure globally and N4 - Apparently Secure nationally, S3S4 - Vulnerable/Apparently Secure (i.e., Blue List) in British Columbia (British Columbia Conservation Data Centre 2012), S4 - Apparently Secure in Alberta (NatureServe 2012), S3 - Vulnerable in Yukon (NatureServe 2012), and S1S2 - Imperilled/Critically Imperilled in Northwest Territories (Table 2). Several NatureServe (2012) rankings for the Western Toad in Canada and the U.S. have been upgraded to higher levels of threat since the previous assessment (COSEWIC 2002).
NatureServe (2012), state and federal rankings for the U.S. are listed in Table 2. NatureServe ranking for populations bordering Canada are Washington S3 (candidate for listing), Idaho S3 (protected nongame species), Montana S2 (species of concern), and Alaska S3S4 (no state ranking program).
The Western Toad is on the IUCN Red List as Near Threatened (Hammerson et al. 2004).
Habitat protection and ownership
Most of the Western Toad’s range is on Crown Lands, including many protected areas, and some of the range is on private lands, municipal lands, Department of National Defence lands, and First Nation settlement lands. There are numerous national parks, provincial parks, Ecological Reserves, Wildlife Habitat Areas and private conservancies that support Western Toad populations. (see Table 1 for National Parks and DND lands).
Western Toad habitat in Yukon is protected in the 16 km² Coal River Ecological Reserve in the Liard River basin, and there are scattered interim protected lands, protected for future First Nation Settlement Lands. Some of the toad’s range in Northwest Territories, along the Liard River to Nahanni Butte, is within the Deh Cho First Nations Interim Measures Agreement Area and the Acho Dene Koe First Nation (Fort Liard) comprehensive land claim area that is currently being negotiated with the federal government.
The Western Toad occurs in national and provincial parks in British Columbia totalling almost 150,000 km² (135,000 km² in provincial parks). The largest national parks and park reserves in British Columbia are Glacier, Kootenay and Gwaii Haanas, all over 1000 km². There are almost 1000 provincial parks, the most significant in area being Tweedsmuir (98810 km²), Tatshenshini-Alsek (9580 km²), Spatsizi-Plateau Wilderness (6951 km²), Northern Rocky Mountains (6657 km²) and Wells Gray-Caribou Mountains-Bowron Lakes (total 8000 km²).
The most significant national parks in the Western Toad’s range in Alberta are Jasper (10,878 km²) and Banff (6,641 km²). The Willmore Wilderness Park (4,600 km²) is adjacent to Jasper. Numerous other protected areas encompass Western Toad range, particularly along the Rocky Mountains and foothills.
Acknowledgements and authorities contacted
K. Ovaska and members of the Amphibian and Reptile Specialist Subcommittee provided constructive reviews of the draft report and guidance throughout the process. B. Eaton and G. Pauly provided comments on an earlier draft of the report. A. Filion of the COSEWIC Secretariat drafted the distribution map and provided calculations for the species’ extent of occurrence and index of area of occupancy.
Authorities contacted
Doug Adama, Natural Resource Specialist, BC Hydro - Water License Requirements, Golden, BC.
Lorna Allen, Coordinator, Alberta Conservation Information Management System, Alberta Tourism, Parks and Recreation, Edmonton, AB.
Robert Anderson, Research Scientist, Canadian Museum of Nature, Ottawa, ON.
Barbara Beasley, Coastal Ecologist, Association of Wetland Stewards for Clayoquot and Barkley Sounds, Ucluelet, BC.
Gavin Berg, Species at Risk Biologist, Non-game and Wildlife Disease, Wildlife Management Branch, Alberta Environment and Sustainable Resource Development, Edmonton, AB.
Constance (Connie) Browne, Postdoctoral Fellow, University of New Brunswick, Saint John, NB.
Suzanne Carrière, Biologist (Biodiversity), Wildlife Division, Northwest Territories Department of Environment and Natural Resources, Yellowknife, NT.
Gordon Court, Provincial Wildlife Status Biologist, Fish and Wildlife Division, Alberta Environment and Sustainable Resource Development, Edmonton, AB.
Andrew Didiuk, Canadian Wildlife Service, Environment Canada, Prairie and Northern Region, Saskatoon, SK.
Dave Duncan, A/Regional Director, Canadian Wildlife Service, Environment Canada, Prairie and Northern Region, Edmonton, AB.
Linda Dupuis, Wildlife Consultant, Brackendale, BC.
Orville Dyer, Ecosystems Biologist, Ministry of Natural Resource Operations, Penticton, BC.
Brian Eaton, Senior Research Scientist – Aquatic Ecology, Ecological Conservation - Ecosystem Management, Alberta Innovates - Technology Futures, Vegreville, AB.
Alain Filion, Scientific and GIS Project Officer, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, QC.
David F. Fraser, Scientific Authority Assessment, A/Manager BC CDC, Ecosystem Branch, Conservation Planning Section, BC Ministry of Environment, Victoria, BC.
Lea Gelling, Zoologist, BCConservation Data Centre, BCMinistry of Environment, Victoria, BC.
Monique Goit, Scientific Project Officer, COSEWIC Secretariat, Canadian Wildlife Service Environment Canada, Gatineau, QC.
Purnima Govindarajulu, Small Mammal and Herpetofauna Specialist, Ecosystems Branch, Wildlife Science Section, Victoria, BC.
Mike Gravel, Territorial Forest Ecologist, Northwest Territories Department of Environment and Natural Resources, Hay River, NT.
Virgil C. Hawkes, Vice-president & Senior Wildlife Biologist, LGL Limited, Sidney BC.
Briar Howes, Science Support, Species at Risk Program, Parks Canada, Gatineau, QC.
Neil Jones, Scientific Project Officer & ATK Coordinator, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, QC.
Thomas Jung, Senior Wildlife Biologist, Fish and Wildlife Branch, Environment Yukon, Whitehorse, YT.
Kris Kendell, Senior Biologist, Alberta Conservation Association, Sherwood Park, AB.
Kamal Khidas, Chief Collection Manager, Vertebrate Collection, Canadian Museum of Nature, Ottawa, ON.
Sandra Kinsey, Wildlife Consultant, Prince George, BC.
John Krebs, Wildlife Biologist, Columbia Basin Fish and Wildlife Compensation Program, Nelson, BC.
Zachary Long, PhD Candidate, Faculty of Natural Resources Management, Lakehead University, Thunder Bay, ON.
Randi Mulder, Biodiversity Information Specialist, Biodiversity Programs, Environment Yukon, Whitehorse, YT.
Patrick Nantel, Conservation Biologist, Species at Risk Program, Ecological Integrity Branch, Parks Canada, Gatineau, QC.
Dean Nernberg, Species at Risk Officer, Director General of Environment, Directorate of Environmental Stewardship, National Defence, Ottawa, ON.
Deanna (Dede) H. Olson, Supervisory Research Ecologist, Team Leader: Aquatic Ecology and Management Team, Land and Watershed Management Research Program, Pacific Northwest Research Station, US Forest Service, Corvallis, OR, U.S.A.
Kristiina Ovaska, Biolinx Environmental Research Ltd., Victoria, BC, and Co-chair, Amphibians and Reptiles Specialist Subcommittee, COSEWIC.
Cynthia (Cindy) Paszkowski, Professor & Associate Chair Research, Department of Biological Sciences, University of Alberta, Edmonton AB.
Greg Pauly, Assistant Curator, Herpetology, Los Angeles County Natural History Museum, Los Angeles, California, U.S.A.
Sanjay Payare, Assistant Professor, Environmental Science & Geography Program, University of Alaska Southeast, Juneau, AK.
Jason Pitre, Scientific Project Officer, COSEWIC Secretariat, Canadian Wildlife Service, Environmental Stewardship Branch, Environment Canada, Gatineau, QC.
Dave Prescott, Species at Risk Biologist, Fish and Wildlife Division, Alberta Environment and Sustainable Resource Development, Red Deer, AB.
Troy Pretzlaw, Kluane Regional Biologist, Kluane Region, Environment Yukon, Whitehorse, YT.
Richard Quinlan, Provincial Species at Risk Specialist, Section Head: Non-game Wildlife, Species at Risk and Wildlife Disease, Fish and Wildlife Division, Alberta Environment and Sustainable Resource Development, Lethbridge, AB.
Lis Rach, TerraNiche Environmental Solutions, Smithers, BC.
Leah Ramsay, Program Zoologist, BC Conservation Data Centre, BC Ministry of Environment, Victoria, BC.
Lucy Reiss, Habitat Planner, Ecosystem Conservation, Environment Canada, Pacific and Yukon Region, Kelowna, BC.
René Rivard, Chilkoot Trail Warden Patrolperson, Yukon Field Unit, Parks Canada, Whitehorse, YT.
Michèle Steigerwald, Assistant Collections Manager, Amphibian & Reptile Collection, Vertebrate Section, Canadian Museum of Nature, Ottawa, ON.
Katrina Stipec, BCConservation Data Centre, Wildlife Inventory Section, Resources Inventory Branch, BC Ministry of Environment, Lands and Parks, Victoria, BC.
Emma Tayless, Fish and Wildlife Technician, Resort Municipality of Whistler, Whistler, BC.
David Tessler, Regional Wildlife Biologist, Wildlife Diversity Program, Alaska
Department of Fish and Game, Division of Wildlife Conservation, Anchorage, Alaska.
Mark Thompson, President, NAMOS BC (Northern Amphibian Monitoring Outpost Society), University of Northern British Columbia, Prince George, BC.
Graham Van Tighem, Executive Director, Yukon Fish and Wildlife Management Board, Whitehorse, YT.
Drajs Vujnovic, Zoologist, Alberta Conservation Information Management System, Alberta Tourism, Parks and Recreation, Edmonton, AB.
Leah Westereng, Recovery Planning Coordinator, Species and Ecosystems at Risk Section, Ecosystems Protection and Sustainability Branch. BCMinistry of Environment, Victoria, BC.
Elke Wind, E. Wind Consulting, Nanaimo, BC.
Barbara (Basia) F. Wojtaszek, Monitoring Ecologist, Gwaii Haanas National Park Reserve, National Marine Conservation Area Reserve, and Haida Heritage Site, Parks Canada, Skidegate, BC.
Carmen Wong, Ecosystem Scientist, Yukon Field Unit, Parks Canada, Whitehorse, YT.
Information sources
Adama, D., and P. Ohanjanian. 2005. A survey of western toads, Bufo boreas, and other amphibians in Glacier and Mount Revelstoke National Parks. Prepared by Adama Wildlife Consulting, Golden, British Columbia., for Parks Canada, Revelstoke, British Columbia. iii + 52 pp.
Adams, M.J., S.K. Galvan, D. Reinitz, R.A. Cole, S. Payre, M. Hahr, and P. Govindarajulu. 2007. Incidence of the fungus, Batrachochytrium dendrobatidis, in amphibian populations along the northwest coast of North America. Herpetological Review 38:430-431.
Adams, M.J., B.R. Hossack, R.A. Knapp, P.S. Corn, S.A. Diamond, P.C. Trenham, and D.B. Fagre. 2005a. Distribution patterns of lentic-breeding amphibians in relation to ultraviolet radiation exposure in western North America. Ecosystems 8:488-500.
Adams, S.B., D.A. Schmetterling, and M.K. Young. 2005b. Instream movements by boreal toads (Bufo boreas boreas). Herpetological Review 36:27-33.
Alaska Department of Fish and Game. 2006. Native amphibians – introduction. Appendix 4, pp. 127-145, in Our Wealth Maintained: a strategy for conserving Alaska’s diverse wildlife and fish resources. Alaska Department of Fish and Game, Juneau, Alaska. xviii + 824 pp.
Alberta Biodiversity Monitoring Institute. 2012. Human footprint summary. [accessed 4 September 2012].
Alberta Environment and Sustainable Resource Development. 2010. General status of Alberta wild species 2010. Web site: http://www.srd.alberta.ca/FishWildlife/SpeciesAtRisk/GeneralStatusOfAlbertaWildSpecies/GeneralStatusOfAlbertaWildSpecies2010/Default.aspx", accessed 6 September 2012.
Austin, M.A., D.A. Buffett, D.J. Nicolson, G.G.E. Scudder, and V. Stevens (eds.). 2008. Taking nature’s pulse: the status of biodiversity in British Columbia. Biodiversity BC, Victoria, B.C. 268 pp. [accessed 25 February 2012].
Ballengée, B., and S.K. Sessions. 2009. Explanation for missing limbs in deformed amphibians. Journal of Experimental Zoology Part B, Molecular and Developmental Evolution 312:770-779.
Bancroft, B.A., N.J. Baker, and A.R. Blaustein. 2008. A meta-analysis of the effects of ultraviolet B radiation and its synergistic interactions with pH, contaminants, and disease on amphibian survival. Conservation Biology 22:987-996.
Barichello, N., pers. comm. 2010. Conversation with B. Slough. Wildlife Consultant, Whitehorse, Yukon.
Bartlelt, P.E. 1998. Natural history notes: Bufo boreas mortality. Herpetological Review 29:96.
Bartelt, P.E., R.W. Klaver, and W.P. Porter. 2010. Modeling amphibian energetics, habitat suitability, and movements of western toads, Anaxyrus (=Bufo) boreas, across present and future landscapes. Ecological Modelling 221:2675-2686.
Bartelt, P.E., C.R. Peterson, and R.W. Klaver. 2004. Sexual differences in the post-breeding movements and habitats selected by western toads (Bufo boreas) in southeastern Idaho. Herpetologica 60:455-467.
Beasley, B., unpubl. data. 2011. Coastal Ecologist, Association of Wetland Stewards for Clayoquot and Barkley Sounds, Ucluelet, British Columbia.
Belden, L.K., E.L. Wildy, A.C. Hatch, and A.R. Blaustein. 2000. Juvenile western toads, Bufo boreas, avoid chemical cues of snakes fed juvenile, but not larval, conspecifics. Animal Behaviour 59:871-875.
Bennett, B., pers. comm. 2012. Email correspondence to K. Ovaska. November 2012. Biologist. Whitehorse, Yukon.
Berg, G., unpubl. data. 2011. Species at Risk Biologist, Non-game and Wildlife Disease, Wildlife Management Branch, Alberta Environment and Sustainable Resource Development, Edmonton, Alberta.
Berger, L., R. Speare, P. Daszak, D.E. Green, A.A. Cunningham, C.L. Goggin, R. Slocombe, M.A. Ragan, A.D. Hyatt, K.R. McDonald, H.B. Hines, K.R. Lips, G. Marantelli, and H. Parks. 1998. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proceedings of the National Academy of Sciences of the U.S.A.95:9031–9036.
Berven, K.A. 1990. Factors affecting population fluctuations in larval and adult stages of the wood frog (Rana sylvatica). Ecology 71:1599-1608.
Biolinx Environmental Research Ltd.and E. Wind Consulting. 2004. Best management practices for amphibians and reptiles in urban and rural environments in British Columbia. Prepared for B.C.Ministry of Water, Land and Air Protection, Nanaimo, British Columbia. viii + 151 pp.
Bishop, C.A. 1992. The effects of pesticides on amphibians and the implications for determining causes of declines in amphibian populations. pp. 67-70. in C.A. Bishop and K.E. Pettit (eds.). Declines in Canadian Amphibian Populations: Designing a National Monitoring Strategy. Canadian Wildlife Service, Occasional Paper No. 76.
Black, J.H., and J.N. Black. 1969. Postmetamorphic basking aggregations of the boreal toad, Bufo boreas boreas. Canadian Field-Naturalist 83:155-156.
Blaustein, A.P., J.J. Beatty, D.H. Olson, and R.M. Storm. 1995. The biology of amphibians and reptiles in old-growth forests in the Pacific Northwest. Gen. Tech. Rep. PNW-GTR-337. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station, Portland, Oregon.
Blaustein, A.R., P.D. Hoffman, D.G. Hokit, J.M. Kiesecker, S.C. Walls, and J.B. Hays. 1994a. UV repair and resistance to solar UV-B in amphibian eggs: a link to amphibian population declines? Proceedings of the National Academy of Sciences of the U.S.A. 91:1791-1795.
Blaustein, A.R., D.G. Hokit, R.K. O’Hara, and R.A. Holt. 1994b. Pathogenic fungus contributes to amphibian losses in the Pacific Northwest. Biological Conservation 67:251-254.
Blaustein, A.R., J.M. Kiesecker, D.P. Chivers, D.G. Hokit, A. Marco, L.K. Belden, and A. Hatch. 1998. Effects of ultraviolet radiation on amphibians: field experiments. American Zoologist 38:799-812.
Blaustein, A.R., J.M. Romansic, J.M. Kiesecker, and A.C. Hatch. 2003. Ultraviolet radiation, toxic chemicals and amphibian population declines. Diversity and Distributions 9:123-140.
Blaustein, A.R., J.M. Romansic, and E.A. Scheessele. 2005a. Ambient levels of ultraviolet-B radiation cause mortality in juvenile western toads, Bufo boreas. American Midland Naturalist 154:375-382.
Blaustein, A.R., J.M. Romansic, E.A. Scheessele, B.A. Han, A.P. Pessier, and J.E. Longcore. 2005b. Interspecific variation in susceptibility of frog tadpoles to the pathogenic fungus Batrachochytrium dendrobatidis. Conservation Biology 19:1460-1468.
Brattstrom, B.H. 1963. A preliminary review of the thermal requirements of amphibians. Ecology 44:238-255.
Bridges, C.M. 2000. Long-term effects of pesticide exposure at various life stages of the southern Leopard frog (Rana sphenocephala). Archives of Environmental Contamination and Toxicology 39:91-96.
Briggs, C.J., R.A. Knapp, and V.T. Vredenburg. 2010. Enzootic and epizootic dynamics of the chytrid fungal pathogen of amphibians. Proceedings of the National Academy of Sciences of the U.S.A.107:9695-9700.
Brinkman, S. 1998. Boreal toad toxicology studies. pp. 83-114, in M.S. Jones, J.P. Goettl, K.L. Scherff-Norris, S. Brinkman, L.J. Livo, and A.M. Goebel (eds.). Boreal toad research progress report 1995-1997. Colorado Division of Wildlife, Fort Collins, Colorado.
British Columbia Conservation Data Centre. 2012. BC Species and Ecosystems Explorer. British Columbia Ministry of Environment, Victoria, British Columbia. [accessed 6 September 2012].
British Columbia Ministry of Environment. 2008. Standard operating procedures: Hygiene protocols for amphibian fieldwork, 2008. Victoria, British Columbia. 8 pp.
British Columbia Ministry of Environment. 2009. Western toad / amphibian monitoring program survey protocol. Victoria, British Columbia. 15 pp.
Brodie, E.D., Jr., and D.R. Formanowicz. 1987. Antipredator mechanisms of larval anurans: protection of palatable individuals. Herpetologica 43:369–373.
Brodie, E.D., Jr., D.R. Formanowicz, Jr., and E.D. Brodie, III. 1978. The development of noxiousness of Bufo americanus tadpoles to aquatic insect predators. Herpetologica 34:302–306.
Browne, C.L. 2010. Habitat use of the western toad in north-central Alberta and the influence of scale. Ph.D. dissertation, University of Alberta, Edmonton, Alberta. 245 pp.
Browne, C.L. 2012. pers. comm. Email correspondence to B. Slough. March 2012. Postdoctoral Fellow, University of New Brunswick, Saint John, Newbrunswick.
Browne, C., R. Chapman, L. Foote, and C. Paszkowski. 2004. Tracking western toads. The Trumpter (Special Edition, Research Review), Friends of Elk Island Society. 2 pp.
Browne, C., B. Eaton, C. Paszkowski, and R. Chapman. 2003. Toads in trouble. Research Links (Parks Canada, Ottawa) 11:12.
Browne, C.L., and C.A. Paszkowski. 2010a. Hibernation sites of western toads (Anaxyrus boreas): characterization and management implications. Herpetological Conservation and Biology 5:49-63.
Browne, C.L., and C.A. Paszkowski. 2010b. Factors affecting the timing of movements to hibernation sites by western toads (Anaxyrus boreas). Herpetologica 66:250-258.
Browne, C.L., C.A. Paszkowski, A.L Foote, A. Moenting, and S.M. Boss. 2009. The relationship of amphibian abundance to habitat features across spatial scales in the boreal plains. Écoscience 16:209-223.
Bull, E.L. 2006. Sexual differences in the ecology and habitat selection of western toads (Bufo boreas) in northeastern Oregon. Herpetological Conservation and Biology 1:27-38.
Bull, E.L. 2009. Dispersal of newly metamorphosed and juvenile western toads (Anaxyrus boreas) in northeastern Oregon, USA. Herpetological Conservation and Biology 4:236-247.
Bull, E.L., and C. Carey. 2008. Breeding frequency of western toads (Bufo boreas) in northeastern Oregon. Herpetological Conservation and Biology 3:282-288.
Bull, E.L., and J.L. Hayes. 2009. Selection of diet by metamorphic and juvenile western toads (Bufo boreas) in northeastern Oregon. Herpetological Conservation and Biology 4: 85-95.
Campbell, J.B. 1970. Life history of Bufo boreas boreas on the Colorado Front Range. Ph. D. dissertation, University of Colorado, Boulder, Col. 124 pp.
Canadian Endangered Species Conservation Council (CESCC). 2011.Wild Species 2010: the general status of species in Canada. 302 pp. + database. (PDF, 4.81MB) [accessed 6 September 2012].
Cannings, S., pers. comm. 2010. Conversation with B. Slough. Endangered Species Biologist, Canadian Wildlife Service, Environment Canada, Whitehorse, Yukon.
Carey, C. 1978. Factors affecting body temperatures of toads. Oecologia 35:197-219.
Carey, C. 1993. Hypothesis concerning the causes of the disappearance of boreal toads from mountains of Colorado. Conservation Biology 7:355-362.
Carey, C., J.E. Bruzgul, L.J. Livo, M.L. Walling, K.A. Kuehl, B.F. Dixon, A.P. Pessier, R.A. Alford, and K. B. Rogers. 2006. Experimental exposures of boreal toads (Bufo boreas) to a pathogenic chytrid fungus (Batrachochytrium dendrobatidis). EcoHealth 3:5-21.
Carey, C., P.S. Corn, M.S. Jones, L.J. Livo, E. Muths, and C.W. Loeffler. 2005. Factors limiting the recovery of boreal toads (Bufo b. boreas). pp. 222-236. in M.J. Lannoo (ed.). Amphibian declines: the conservation status of United States Species. University of California Press, Berkeley, CA
Carpenter, C.C. 1954. A study of amphibian movement in the Jackson Hole Wildlife Park. Copeia 1954:197-200.
Carr L. W., and L. Fahrig. 2001. Effect of road traffic on two amphibian species of differing vagility. Conservation Biology 15:1071-1078.
Carrière, S., unpubl. data. 2011. Biologist (Biodiversity), Wildlife Division, Northwest Territories Department of Environment and Natural Resources, Yellowknife, Northwest Territories.
Carstensen, R, M. Willson, and R. Armstrong. 2003. Habitat use of amphibians in northern Southeast Alaska. Report to the Alaska Department of Fish and Game. Discovery Southeast, Juneau, Alaska. 75 pp.
Center for Biological Diversity, Center for Native Ecosystems, and Biodiversity
Conservation Alliance. 2011. Before the Secretary of the Interior: petition to list a distinct population segment of the boreal toad (Anaxyrus boreas boreas) as endangered or threatened under the Endangered Species Act. 72 pp.
CITES. 2012. Appendices I, II and III: valid from 25 September 2012. Convention on International Trade in Endangered Species of Wild Fauna and Flora. 46 pp. Web Site: Convention on International Trade in Endangered Species of Wild Fauna and Flora - Appendices I, II and III (PDF, 391 KB) [accessed 6 September 2012].
Clegg, S. 2011. Assessing the impacts of vehicular mortality of migrating amphibians near Ryder Lake, British Columbia. Fraser Valley Conservancy. 11 pp.
Cook, F.R. 1977. Records of the boreal toad from
Cook, F.R. 1983. An analysis of toads of the Bufo americanus group in a contact zone in central northern North America. National Museums of Canada, Publications in Natural Sciences 3:1-89.
Corn, P.S. 1998. Effects of ultraviolet radiation on boreal toads in Colorado. Ecological Applications 8:18-26.
Corn, P.S. 2003. Amphibian breeding and climate change: Importance of snow in the mountains. Conservation Biology 17:622-625.
COSEWIC. 2002. COSEWIC assessment and status report on the western toad Bufo boreas in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 31 pp.
Crother, B.I. (ed.). 2012. Scientific and standard English names of amphibians and reptiles of North America North of Mexico, with comments regarding confidence in our understanding. 7th Edition. Herpetological Circulars. Society for the Study of Amphibians and Reptiles, St. Louis, Missouri.
Davis, T.M. 2000. Ecology of the western toad (Bufo boreas) in forested areas on Vancouver Island. Final Report. Forest Renewal Research Project. B.C. Ministry of Forests, Victoria, British Columbia. 28 pp.
Davis, T.M., pers. comm. 2004. Email correspondence to B. Slough. September 2004. Instructor, Biology and the Environmental Technology Departments, Camosun College, Victoria, British Columbia.
Davis, T.M., and P.T. Gregory. 2003. Decline and local extirpation of the western toad, Bufo boreas, on southern Vancouver Island, British Columbia, Canada. Herpetological Review 34:350-352.
Daszak, P., L. Berger, A.A. Cunningham, A.D. Hyatt, D.E. Green, and R. Speare. 1999. Emerging infectious diseases and amphibian population declines. Emerging Infectious Diseases 5:735-748.
Deguise, I., and J.S. Richardson. 2009a. Movement behaviour of adult western toads in a fragmented, forest landscape. Canadian Journal of Zoology 87:1184-1194.
Deguise, I., and J.S. Richardson. 2009b. Prevalence of the chytrid fungus (Batrachochytrium dendrobatidis) in western toads in southwestern British Columbia, Canada. Northwestern Naturalist 90:35-38.
Dehn, M.M. 1990. Vigilance for predators: detection and dilution effects. Behavioral Ecology and Sociobiology 26:337–342.
Demers, M.J., E.N. Kelly, J.M. Blais, F.R. Pick, V.L. St. Louis, and D.W. Schindler. 2007. Organochlorine compounds in trout from lakes over 1600 meter elevation gradient in the Canadian Rocky Mountains. Environmental Science and Technology 41:2723-2729.
Devito, J., D.P. Chivers, J.M. Kiesecker, A. Marco, E.L. Wildy, and A.R. Blaustein. 1998. The effects of snake predation on metamorphosis of western toads, Bufo boreas (Amphibia, Bufonidae). Ethology 104:185-193.
Dulisse, J. 2007. Western toad surveys for proposed Glacier - Howser Hydroelectric Project. Nelson, BC. Prepared for AXOR Group Inc., Montréal, Québec. 13 pp.
Dulisse, J., and D. Hausleitner. 2009. 2008 West Kootenay amphibian survey. Prepared for Columbia Basin Fish & Wildlife Compensation Program, Nelson, British Columbia. iv + 33 pp.
Dulisse, J., and D. Hausleitner. 2010. 2009 amphibian survey, Columbia Forest District. Prepared for Columbia Basin Fish & Wildlife Compensation Program, Nelson, British Columbia. iv + 30 pp.
Dulisse, J., D. Hausleitner and J. Boulanger. 2011. Western toads at Summit Lake, 2010 field season. Prepared for Columbia Basin Fish & Wildlife Compensation Program, Nelson, British Columbia. vi + 45 pp.
Dupuis, L., unpubl. data. 2002, 2011. Wildlife Consultant, Brackendale, B.C.
Dykstra, J. 2004. A survey of the status of the western toad (Bufo boreas) and other amphibians in Mount Revelstoke and Glacier National Parks, British Columbia, Canada. B.Sc. Honours Thesis, Okanagan University College, Kelowna, British Columbia. 58 pp.
Eaton, B.R., S. Eaves, C. Stevens, A. Puchniak, and C.A. Paszkowski. 2004. Deformity levels in wild populations of the wood frog (Rana sylvatica) in three ecoregions of western Canada. Journal of Herpetology 38:283–287.
Eaton, B.R., C. Grekul, and C. Paszkowski. 1999. An observation of interspecific amplexus between boreal, Bufo boreas, and Canadian, B. hemiophrys, toads, with a range extension for the boreal toad in central Alberta. Canadian Field-Naturalist 113:512-513.
Eaton, B.R., A.E. Moenting, C.A. Paszkowski and D. Shpeley. 2008. Myiasis by Lucilia silvarum (Calliphoridae) in amphibian species in boreal Alberta, Canada. Journal of Parasitology 94:949–952.
Eaton, B.R., W. M. Tonn, C.A. Paszkowski, A.J. Danylchuk, and S.M. Boss. 2005. Indirect effects of fish winterkills on amphibian populations in boreal lakes. Canadian Journal of Zoology 83:1532-1539.
Eaves, S.E. 2004. The distribution and abundance of amphibians across land-use types in Alberta's aspen parkland. M.Sc. Thesis, University of Alberta, Edmonton, Alberta. 126 pp.
Eigenbrod, F., S. Hecnar, and L. Fahrig. 2008. The relative effects of road traffic and forest cover on anuran populations. Biological Conservation 141:35-46.
Environment Canada. 2011. Management plan for the Western Toad (Anaxyrus boreas) in Canada [Draft]. Species at Risk Act Management Plan Series. Environment Canada, Ottawa. iii + 20 pp.
Fahrig, L., J.H. Pedlar, S.E. Pope, P.D. Taylor, and J.F. Wegner. 1995. Effect of road traffic on amphibian density. Biological Conservation 73:177-182.
Farrer, R.A., L.A. Weinert, J. Bielby, T.W.J. Garner, F. Balloux, F. Claire, J. Bosch, A.A. Cunningham, C. Weldon, L.H. du Preez, L. Anderson, S.L. Kosakovsky Pond, R. Sharar-Golan, D.A. Henk, and M.C. Fisher. 2011. Multiple emergences of genetically diverse amphibian-infected chytrids include a globalized hypervirulent recombinant lineage. Proceedings of the National Academy of Sciences of the U.S.A. 108:18732-18736.
Fitzgibbon, K. 2001. An evaluation of corrugated steel culverts as transit corridors for amphibians and small mammals at two Vancouver Island wetlands and comparative culvert trials. M.A. thesis, Royal Roads University, Victoria, British Columbia. 112 pp.Fleischner, T.L. 1994. Ecological costs of livestock grazing in western North America. Conservation Biology 8:629-644.
Fontenot, L.W., G.P. Noblet, and S.G. Platt. 1994. Rotenone hazards to amphibians and reptiles. Herpetological Review 25: 50-156.
Formanowicz, D.R., Jr., and E.D. Brodie, Jr. 1982. Relative palatabilities of members of a larval amphibian community. Copeia 1982:91–97.
Fraker, M.A., and V.C. Hawkes. 2000. Peace River wildlife surveys: 1999. Prepared by TerraMar Environmental Research Ltd., North Saanich, British Columbia., for BC Hydro, Burnaby, British Columbia. 85 pp.
Friis, L., unpubl. data. 2007. Small Mammal and Herpetofauna Specialist, Ecosystems Branch Ministry of Environment, Victoria, British Columbia.
Frost, D.R., T. Grant, J. Faivovick, R.H. Bain, A. Haas, C.F.B. Haddad, R.O. De Sá, A. Channing, M. Wilkinson, S.C. Donnellan, C. Raxworthy, J.A. Campbell, B.L. Blotto, P. Moler, R.C. Drewes, R.A. Nussbaum, J.D. Lynch, D.M. Green, and W.C. Wheeler. 2006. The amphibian tree of life. Bulletin of the American Museum of Natural History 297:1–370.
Frost, D.R., R.W. McDiarmid, and J.R. Mendelson III. 2008. Anura: Frogs. Pp. 2-12, in B.I. Crother (ed.). Scientific and standard English names of amphibians and reptiles of North American North of Mexico. SSAR Herpetological Circular 37.
Garcia, T.S., J.M. Romansic, and A.R. Blaustein. 2006. Survival of three species of anuran metamorphs exposed to UV-B radiation and the pathogenic fungus Batrachochytrium dendrobatidis. Diseases of Aquatic Organisms 72:163-169.
Garrett, C. 2005. The influence of habitat on the growth and longevity of the western toad (Bufo boreas) in the aspen parkland of Alberta. unpubl. manuscript. University of Alberta, Edmonton, Alberta. 35 pp.
Gelling, L., pers. comm. 2011. Email correspondence to B. Slough. July 2011. Zoologist, Conservation Data Centre, BC Ministry of Environment, Victoria, British Columbia.
Gibbs, J.P. 2000. Wetland loss and biodiversity conservation. Conservation Biology 14:314-317.
Glooschenko, V., W.F. Weller, P.G.L. Smith, L. Alvo, and J.H.G. Archbold. 1992. Amphibian distribution with respect to pond and water chemistry near Sudbury, Ontario. Canadian Journal of Fisheries & Aquatic Sciences 49 (Suppl. 1):114-121.
Goebel, A.M. 2005. Conservation systematics: the Bufo boreas species group. Chapter 30, Pp. 210-221. in M.J. Lannoo (ed.). The Conservation Status of United States Species. University of California Press, Berkeley, California.
Goebel, A.M., T.A. Ranker, P.S. Corn, and R.G. Olmstead. 2009. Mitochondrial DNAevolution in the Anaxyrus boreas species group. Molecular Phylogenetics and Evolution 50:209-225.
Govindarajulu, P. 2008. Literature review of impacts of glyphosate herbicide on amphibians: what risks can the silvicultural use of this herbicide pose for amphibians in B.C. ? Wildlife Report No. R-28. B.C. Ministry of Environment, Victoria, B.C. v + 79 pp.
Govindarajulu, P., unpubl.data. 2011. Small Mammal and Herpetofauna Specialist, Ecosystems Branch, Wildlife Science Section, Victoria, British Columbia.
Gray, M.J., D.L. Miller, and J.T. Hoverman. 2009. Ecology and pathology of amphibian ranaviruses. Diseases of Aquatic Organisms 87:243-266.
Guscio, C.G., B.R. Hossack, L.A. Eby, and P.S. Corn. 2008. Post-breeding habitat use by adult boreal toads (Bufo boreas) after wildfire in Glacier National Park, USA. Herpetological Conservation and Biology 3:55-62.
Gyug, L.W. 1996. Timber-harvesting effects on riparian wildlife and vegetation in the Okanagan Highlands of British Columbia. British Columbia Ministry of Environment, Lands and Parks, Wildlife Branch, Victoria, British Columbia. Wildlife Bulletin No. B-97. 112 pp.
Hammerson, G., G. Santos-Barrera, and E. Muths. 2004. Anaxyrus boreas in: IUCN 2012. IUCN Red List of Threatened Species. Version 2012.1. Web site: IUCN Red List of Threatened Species [accessed 6 September 2012].
Harfenist, A., T. Power, K.L. Clark, and D.B. Peakall. 1989. A review and evaluation of the amphibian toxicological literature. Canadian Wildlife Service Technical Report Series 61.
Hatch, A.C., L.K. Belden, E. Scheellele, and A.R. Blaustein. 2001. Juvenile amphibians do not avoid potentially lethal levels of urea on soil substrate. Environmental Toxicology and Chemistry 20:2328-2335.
Hawkes, V.C., J.D. Fenneman, and K.N. Tuttle. 2010. Natural Resources Inventory (NRI), Department of National Defence Canada, ASU Chilliwack – Chilcotin Training Area, British Columbia. LGL Report EA3158. Prepared by LGL Limited environmental research associates for Public Works and Government Services Canada, Vancouver, B.C. 134 pp. + Appendices.
Hawkes, V.C., G.F. Searing, M. Todd, M. Demarchi, J. Muir, and M.K. McNicholl. 2006. Peace River wildlife surveys: 2005 habitat suitability modelling and wildlife inventory. Draft final report. Prepared by LGL Limited environmental research associates, Sidney, B.C., for BC Hydro, Burnaby, B.C. xiii + 127pp.
Hawkes, V.C., K. Tuttle, J.D. Fenneman, and M.W. Demarchi. 2009. Natural Resources Inventory (NRI), Department of National Defence Canada, ASU Chilliwack, British Columbia. LGL Report EA3005. Prepared by LGL Limited Environmental Research Associates for Public Works and Government Services Canada, Vancouver, British Columbia. 375pp.
Hawkes, V.C., and K.N. Tuttle. 2010. Kinbasket and Arrow Lakes reservoirs: Amphibian and reptile life history and habitat use assessment. Annual Report – 2009. LGL Report EA3075. Prepared by LGL Limited environmental research associates, Sidney, B.C., for BC Hydro, Burnaby, British Columbia. 70 pp + App.
Hawkes, V.C., K.N. Tuttle, and P. Gibeau. 2011. CLBMON-37. Kinbasket and Arrow Lakes Reservoirs: amphibian and Reptile Life History and Habitat Use Assessment. Year 3 Annual Report – 2010. LGL Report EA3075A. Prepared by LGL Limited environmental research associates, Sidney, British Columbia, for BC Hydro Generations, Water License Requirements, Burnaby, British Columbia. 90 pp + App.
Haycock, R.D. 1998. Amphibian survey with special emphasis on the Oregon spotted frog Rana pretiosa. Selected wetland sites: Fraser River Lowlands and corridors to the Interior Plateau. Report to Ministry of Environment, Wildlife Branch, British Columbia, Canada. 261 pp.
Hayes, T.B. 2004. There is no denying this: defusing the confusion about atrazine. BioScience 54:1138-1149.
Hays, J.B., A.R. Blaustein, J.M. Kiesecker, P.D. Hoffman, I. Padelova, D. Coyle, and T. Richardson. 1996. Developmental responses of amphibians to solar and artificial UVB sources: a comparative study. Photochemistry and Photobiology 64:449-456.
Hengeveld, P.E. 1999. Amphibian reconnaissance surveys in the Williston Reservoir watershed, May 1998. Peace/Williston Fish and Wildlife Compensation Program, Report No. 207. 19 pp. + app.
Hengeveld, P.E. 2000. Presence and distribution of amphibians in the Williston and Dinosaur Reservoir watersheds. Peace/Williston Fish and Wildlife Compensation Program Report No. 212.
Hillman, S.S. 1980. Physiological correlates of differential dehydration tolerance in anuran amphibians Copeia 1980:125-129.
Hossack, B.R., and P.S. Corn. 2007. Responses of pond-breeding amphibians to wildfire: Short-term patterns in occupancy and colonization. Ecological Applications 17:1403-1410.
Howes, B., pers. comm. Email correspondence to B. Slough and to K. Ovaska. 2011 and 2012. Science Support, Species at Risk Program, Parks Canada, Gatineau, Québec.
Idaho Department of Fish and Game. 2005. Idaho Comprehensive Wildlife Conservation Strategy. Idaho Conservation Data Center, Idaho Department of Fish and Game, Boise, Idaho. Web Site: https://fishandgame.idaho.gov/public/wildlife/cwcs/ [accessed 6 September 2012].
IPCC. 2007. Climate Change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment. Report of the Intergovernmental Panel on Climate Change, Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller (eds.). Cambridge University Press, Cambridge, U.K. and New York, N.Y., USA. 996 pp.
IUCN Standards and Petitions Subcommittee. 2010. Guidelines for using the IUCN red list categories and criteria. Version 8.1. Prepared by the Standards and Petitions Subcommittee in March 2010. Web site: http://intranet.iucn.org/webfiles/doc/SSC/RedList/RedListGuidelines.pdf [accessed 25 February 2012].
International Union for Conservation of Nature and Conservation Measures Partnership (IUCN and CMP). 2006. IUCN -CMP unified classification of direct threats, ver. 1.0 – June 2006. Gland, Switzerland. 17 pp. Web site: The Conservation Measures Partnership [accessed 6 September 2012].
Jones, M.S., and J.P. Goettl. 1998. Henderson/Urad boreal toad studies. Pp. 21-82, in M.S. Jones, J.P. Goettl, K.L. Scherff-Norris, S. Brinkman, L.J. Livo, and A.M. Goebel (eds.). Boreal toad research progress report 1995-1997. Colorado Division of Wildlife, Fort Collins, Colorado.
Jones, M.S., J.P. Goettl, K.L. Scherff-Norris, S. Brinkman, L.J. Livo, and A.M. Goebel. 1998. Boreal toad research progress report 1995-1997. Colorado Division of Wildlife, Ft. Collins, Colorado. 171 pp.
Jones, L.C., W.P. Leonard, and D.H. Olson (eds.). 2005. Amphibians of the Pacific Northwest. Seattle Audubon Society, Seattle, Washington. 227 pp.
Johnson, P.T.J., K.B. Lunde, R.W. Haight, J. Bowerman, and A.R. Blaustein. 2001. Ribeiroia ondatrae (Trematoda: Digenea) infection induces severe limb malformations in western toads (Bufo boreas). Canadian Journal of Zoology 79:370-379.
Johnson, P.T.J., K.B. Lunde, E.G. Ritchie, and A.E. Launer. 1999. The effect of trematode infection on amphibian limb development and survivorship. Science 284:802–804.
Johnston, B., and S. Crowshoe. 2011. Amphibian Monitoring in Waterton Lakes National Park, 1997-2011. Parks Canada unpublished report. 10 pp.
Jordan, D.J., C.J. Rombourgh, C.A. Pearl, and B. McCreary. 2004. Cannibalism and predation by western toad (Bufo boreas boreas) larvae in Oregon, USA. Western North American Naturalist 64:403-405.
Kats, L.B., J.M. Kiesecker, D.P. Chivers, and A.R. Blaustein. 2000. Effects of UV-B radiation on anti-predator behavior in three species of amphibians. Ethology 106:921-931.
Kats, L.B., J.W. Petranka, and A. Sih. 1988. Antipredator defenses and persistence of amphibian larvae with fishes. Ecology 69:1865-1870.
Kendell, K., unpubl. data. 2011. Senior Biologist, Alberta Conservation Association, Sherwood Park, Alberta.
Kiesecker, J.M. 2002. Synergism between trematode infection and pesticide exposure: a link to amphibian limb deformities in nature? Proceedings of Natural Academy of Sciences 99:9900-9904.
Kiesecker, J.M., and A.R. Blaustein. 1995. Synergism between UV-B radiation and a pathogen magnifies amphibian embryo mortality in nature. Proceedings of the National Academy of Sciences of the U.S.A. 92:11049–11052.
Kiesecker, J.M., and A.R. Blaustein. 1997. Influences of egg laying behavior on pathogenic infection of amphibian eggs. Conservation Biology 11:214-220.
Kiesecker, J.M., D.P. Chivers, and A.R. Blaustein. 1996. The use of chemical cues in predator recognition by western toad tadpoles. Animal Behaviour 52:1237-1245.
Kiesecker, J.M., A.R. Blaustein, and L.K. Belden. 2001a. Complex causes of amphibian population declines. Nature 410:681–684.
Kiesecker, J.M., A.R. Blaustein, and C.L. Miller. 2001b. Transfer of a pathogen from fish to amphibians. Conservation Biology 15:1064-1070.
Kinsey, S., unpubl. data. 2011. Wildlife Consultant, Prince George, British Columbia.
Kinsey, S., pers. comm. 2011. Email correspondence to B. Slough. October and November 2011. Wildlife Consultant, Prince George, British Columbia.
Leaver, S., unpubl. data. 2007. Small Mammal and Herpetofauna Technician, Ecosystems Branch, Ministry of Environment, Victoria, British Columbia.
Lepitzki, D., pers. comm. 2012. Email correspondence to K. Ovaska. Wildlife Systems Research, Banff, and co-chair, Molluscs SSC of COSEWIC, Banff, Alberta.
Lepitzki, D. 2008. Synopsis and technical compendium – amphibian abundance and Boreal Toad. Unpublished reports prepared for the the Aquatics Section of Banff National Park for the 2008 Banff National Park State of the Park Report.
Livo, L.J. 1998. Investigation of boreal toad tadpole ecology. Pp. 115-146, in M.S. Jones, J.P. Goettl, K.L. Scherff-Norris, S. Brinkman, L.J. Livo, and A.M. Goebel (eds.). Boreal toad research progress report 1995-1997. Colorado Division of Wildlife, Fort Collins, Colorado.
Long, Z. 2010. Anaxyrus boreas (Western Toad): Advertisement vocalization. Herpetological Review 41:332-333.
Long, Z., pers. comm. 2012. Email correspondence to B. Slough. March and July 2012. PhD Candidate, Faculty of Natural Resources Management, Lakehead University, Thunder Bay, Ontario.
Longcore, J.R., J.E. Longcore, A.P. Pessier, and W.A. Halteman. 2007. Chytridiomycosis widespread in anurans of northeastern United States. Journal of Wildlife Management 71:435–444.
Longcore,.J.E., A.P. Pessier, and D.K. Nichols. 1999. Batrachochytrium dendrobatidisgen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia 91:219–227.
Macdonald, S.E., B. Eaton, C.S. Machtans, C. Paszkowski, S. Hannon, and S. Boutin. 2006. Is forest close to lakes ecologically unique? Forest Ecology and Management 223:1-17.
McGarvie Hirner, J.L., and S.P. Cox. 2007. Effects of rainbow trout (Oncorhynchus mykiss) on amphibians in productive recreational fishing lakes of British Columbia. Canadian Journal of Fisheries and Aquatic Sciences 64:1770-1780.
Marsh, D. M., and P. C. Trenham. 2001. Metapopulation dynamics and amphibian conservation. Conservation Biology 15:40-49.
Matsuda, B.M., D.M. Green, and P.T. Gregory. 2006. Amphibians and reptiles of British Columbia. Royal BC Museum Handbook, Victoria, British Columbia. 266 pp.
McIvor, M. and D. Lepitzki, pers. comm. 2012. Email and photo of a calling Western Toad from Kootenay National Park sent to D. Lepitzki. 20 November 2012. Bow Valley Naturalists, Banff, Alberta.
Mendez, D., R. Webb, L. Berger, and R. Speare. 2008. Survival of the amphibian chytrid fungus Batrachochytrium dendrobatidis on bare hands and gloves: hygiene implications for amphibian handling. Diseases of Aquatic Organisms 82:97-104.Mennell, L., unpubl.data. 2007. Wildlife Consultant, Lewes Lake, Yukon.
Mennell, R.L., and B.G. Slough. 1998. Amphibian and biodiversity inventories of ecoregions in northwestern British Columbia. Habitat Conservation Trust Fund Project TF28056. British Columbia Ministry of Environment, Lands and Parks, Victoria, British Columbia. 86 pp.
Miller, D., M. Gray, and A. Storfer. 2011. Ecopathology of ranaviruses infecting amphibians. Viruses 2011:2351-2373.
Mulder, R., unpubl. data. 2011. Biodiversity Information Specialist, Biodiversity Programs, Yukon Conservation Data Centre, Environment Yukon, Whitehorse, Yukon.
Muths, E. 2003. Home range and movements of boreal toads in undisturbed habitat. Copeia 2003:160-165.
Muths, E., P.S. Corn, A.P. Pessier, and D.E. Green. 2003. Evidence for disease-related amphibian decline in Colorado. Biological Conservation 110:357-365.
Muths, E., and P. Nanjappa. 2005. Bufo boreas Baird and Girard, 1852(b): Western Toad. Pp 392-396, in M. Lannoo (ed.). Amphibian declines: The conservation status of United States Species. University of California Press, Berkeley, California.
Muths, E., D. Pilliod, and L. Livo. 2008. Distribution and environmental limitations of an amphibian pathogen in the Rocky Mountains, USA. Biological Conservation 141:1484-1492.
Myers, E.M., and K.R. Zamudio. 2004. Multiple paternity in an aggregate breeding amphibians: the effect of reproductive skew on estimates of male reproductive success. Molecular Ecology 13:1951-1963.
NatureServe. 2012. NatureServe Explorer: an online encyclopedia of life. Version 7.1. NatureServe, Arlington, VA. [accessed 6 September 2012].
Nernberg, D., unpubl. data. 2011. Species at Risk Officer, Director General of Environment, Directorate of Environmental Stewardship, National Defence, Ottawa, Ontario.
Northwest Territories Department of Environment and Natural Resources. 2006. 2006 update: amphibians and reptiles. Web site: http://www.enr.gov.nt.ca/_live/documents/content/amphibians_reptiles_2006Update.pdf, (PDF, 86.0 KB) [accessed 24 October 2011].
Ohanjanian, P., D. Adama, and A. Davidson. 2006. An amphibian inventory of the East Kootenays with an emphasis on Bufo boreas, 2005. Prepared for Columbia Basin Fish and Wildlife Compensation Program, Nelson, British Columbia. iii + 31 pp.
Ohanjanian, I.A., and M-A. Beaucher. 2000. Faunal and habitat diversity at Summit Lake Provincial Park with special emphasis on red and blue-listed species, amphibians, and ground-nesting birds. Report prepared for B.C.Parks, Nelson, British Columbia. iv + 24 pp.
Olson, D.H. 1988. The ecological and behavioral dynamics of breeding in three sympatric anuran amphbians. Ph.D. Dissertation, Oregon State University, Corvallis, Oregon. xi + 260 pp.
Olson, D.H. 1989. Predation of breeding western toads. Copeia 1989:391-397.
Olson, D.H., A.R. Blaustein, and R.K. O’Hara. 1986. Mating pattern variability among western toad (Bufo boreas) populations. Oecologia 70:351-356.
Ouellet, M., I. Mikaelian, B.D. Pauli, J. Rodrigue, and D. M. Green. 2005. Historical evidence of widespread chytrid infection in North American amphibian populations. Conservation Biology 19:1431–1440.
Ovaska, K.E. 1997. Vulnerability of amphibians in Canada to global warming and increased ultraviolet radiation. Herpetological Conservation 1:206-225.
Ovaska, K.], unpubl. data. 2012. Biolinx Environmental Research Ltd., Victoria, British Columbia.
Ovaska, K., L. Sopuck, and C. Engelstoft. 2011. Community-based amphibian monitoring program in multi-use landscapes in south-central B.C. : progress report, 2011. Biolinx Environmental Research Ltd., North Saanich, B.C. 19 pp. [Website: http://www.nicolanaturalists.ca/files/Nicola-amphibian-monitoring-rep-website-version-Dec-11.pdf, accessed November 2012].
Pagnucco, K.S., C.A. Paszkowski, and G.J. Scrimgeour. 2011. Using cameras to monitor tunnel use by long-toed salamanders (Ambystoma macrodactylum): an informative, cost-efficient technique. Herpetological Conservation and Biology 6:277-286.
Parks Canada Agency. 2009 – 2011. Draft Managed Area Element Status Reports for the Western Toad. Separate reports are available for each park, summarized by B. Howes for this report.
Paszkowski, C., pers. comm. 2012. Email correspondence to B. Sloughand to K. Ovaska. February, March, and November 2012. Professor and Associate Chair Research, Department of Biological Sciences, University of Alberta, Edmonton, Alberta.
Pauly, G.B. 2008. Phylogenetic systematics, historical biogeography, and the evolution of vocalizations in Nearctic toad (Bufo). Ph.D. Dissertation, University of Texas, Austin, Texas. xvi + 165 pp.
Pauly, G.B., pers. comm. 2011. Email correspondence to B. Slough. March and October 2011. Assistant Curator, Herpetology, Los Angeles County Natural History Museum, Los Angeles, California.Pauly, G.B., Hillis, D.M., and D.C. Cannatella. 2009. Taxonomic freedom and the role of official lists of species names. Herpetologica 65:115-128.
Payare, S., pers. comm. 2011. Email correspondence to B. Slough. May 2011. Assistant Professor, Environmental Science & Geography Program, University of Alaska Southeast, Juneau, Alaska.
Pearl, C.A. 2002. Bufo boreas (western toad) predation. Herpetological Review 31:233-234.
Pearl, C.A., M.J. Adams, R.B. Bury, W.H. Wente, and B. McCreary. 2009b. Evaluating amphibian declines with site revisits and occupancy models: status of montane anurans in the Pacific Northwest USA. Diversity 1:166-181.
Pearl, C.A., and J. Bowerman. 2006. Observations of rapid colonization of constructed ponds by western toads (Bufo boreas) in Oregon, USA. Western North American Naturalist 66:397-401.
Pearl, C.A., J. Bowerman, M.J. Adams, and N.D. Chelgren. 2009a. Widespread occurrence of the chytrid fungus Batrachochytrium dendrobatidis on Oregon spotted frog (Rana pretiosa). EcoHealth 6:209-218.
Pearl, C.A., E.L. Bull, D.E. Green, J. Bowerman, M.J. Adams, A. Hyatt, and W.H. Wente. 2007. Occurrence of the amphibian pathogen Batrachochytrium dendrobatidisin the Pacific Northwest. Journal of Herpetology 41:145-149.
Petranka, J.W., C.K. Smith, and A.F. Scott. 2004. Identifying the minimal demographic unit for monitoring pond-breeding amphibians. Ecological Applications 14:1065–1078.
Pilliod, D.S., E. Muths, R.D. Scherer, P.E. Bartelt, P.S. Corn, B.R. Hossack, B.A. Lambert, R. McCaffery, and C. Gaughan. 2010. Effects of amphibian chytrid fungus on individual survival probability in wild boreal toads. Conservation Biology 24:1259-1267.
Provincial Western Toad Working Group. 2011. Management plan for the Western Toad (Anaxyrus boreas; formerly Bufo boreas) in British Columbia. Prepared for the British Columbia Ministry of Environment, Victoria, British Columbia. 25 pp.
Rach, L. 2008. Amphibian assessments in the lower Taku River for the Tulsequah Chief Project. Gartner Lee Limited for Redfern Resources Ltd. iii + 17 pp.
Ramsay, L., pers. comm. 2011. Email correspondence to B. Slough. April 2011. Program Zoologist, BC Conservation Data Centre, BC Ministry of Environment, Victoria, British Columbia.
Raverty, S., and T. Reynolds. 2001. Cutaneous chytridiomycosis in dwarf aquatic frogs (Hymenochirus boettgeri) originating from Southeast Asia and in a Western Toad (Bufo boreas) from northeastern British Columbia. Canadian Veterinary Journal 42:385-386.
Reimchen, T.E. 1991. Introduction and dispersal of the Pacific treefrog, Hyla regilla, on the Queen Charlotte Islands, British Columbia. Canadian Field-Naturalist 105:288-290.
Relyea, R.A., and N. Diecks. 2008. An unforeseen chain of events: Lethal effects of pesticides on frogs at sublethal concentrations. Ecological Applications 18:1728-1742.
Rescan. 2008. Morrison Copper/Gold Project Amphibian Baseline Report. Prepared by Rescan Environmental Services Ltd., Vancouver, B.C. for Pacific Booker Minerals Inc., Vancouver, British Columbia. 43 pp.
Rescan Tahltan Environmental Consultants. 2008. Schaft Creek western toad baseline 2007. Prepared by Rescan Tahltan Environmental Consultants, Vancouver, B.C. for Copper Fox Metals Inc., Calgary, Alberta. vi + 52 pp.
Rivard, R., pers. comm. 2011. Email correspondence to B. Slough. April 2011. Chilkoot Trail Warden-Patrolperson, Yukon Field Unit, Parks Canada, Whitehorse, Yukon.
Rouse, J.D., C.A. Bishop, and J. Struger. 1999. Nitrogen pollution: an assessment of its threats to amphibian survival. Environmental Health Perspectives 107:799-803.
Russell, A.P., and A.M. Bauer. 2000. The amphibians and reptiles of Alberta: A field guide and primer of boreal herpetology. Second edition. University of Calgary Press, Calgary, Alberta. 279 pp.
Salafsky, N., D. Salzer, A.J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O’Connor, and D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classification of threats and actions. Conservation Biology 22:897-911.
Sanzo, D., and S.J. Hecnar. 2005. Effects of road de-icing salt (NaCl) on larval wood frogs (Rana sylvatica). Environmental Pollution 140:247-256.
Schank, C.M.M. 2008. Assessing the effects of trout stocking on native amphibian communities in small boreal foothill lakes of Alberta. M.Sc. Thesis, University of Alberta, Edmonton, Alberta. 112 pp.
Schmetterling, D. A., and M. K. Young. 2008. Summer movements of boreal toads (Bufo boreas boreas) in two western Montana basins. Journal of Herpetology 42:111-123.
Schock, D.M., G.R. Ruthig, J.P. Collins, S.J. Kutz, S. Carrière, R.J. Gau, A.M. Veitch, N.C. Larter, D.P. Tate, G. Guthrie, D.G. Allaire, and R.A. Popko. 2009. Amphibian chytrid fungus and ranaviruses in the Northwest Territories, Canada. Diseases of Aquatic Organisms 92:231-240.
Shaffer, H.B., G.M. Fellers, S.R. Voss, J.C. Oliver, and G.B. Pauly. 2004. Species boundaries, phylogeography and conservation genetics of the red-legged frog (Rana aurora/draytonii) complex. Molecular Ecology 13:2667-2677.
Shepherd, B., and W. Hughson. 2012. 2011 Amphibian monitoring project. Jasper National Park. Parks Canada unpubl. report. 11 pp.
Skelly, D.K., K.L. Yurewicz, E.E. Werner, and R.A. Relyea. 2003. Estimating decline and distributional change in amphibians. Conservation Biology 17:744-751.
Skerratt, L.F., L. Berger, R. Speare, S. Cashins, K.R. McDonald, A.D. Phillott, H. B. Hines, and N. Kenyon. 2007. Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. EcoHealth 4:125–134.
Slough, B.G., unpubl. data. 2001, 2004, 2005, 2007, 2011. Wildlife Consultant, Whitehorse, Yukon.
Slough, B.G. 2004. Western toad inventory in the Chilkoot Trail National Historic Site, July-August 2004. Parks Canada Species at Risk Inventory Fund Project SARINV04-30. 54 pp.
Slough, B.G. 2005. Western toad, Bufo boreas, stewardship in the Yukon. NatureServe Yukon, Whitehorse, Yukon. 26 pp.
Slough, B.G. 2009. Amphibian chytrid fungus in western toads (Anaxyrus boreas) in British Columbia and Yukon, Canada. Herpetological Review 40:319-321.
Slough, B.G. 2012. Amphibians. Pp. 130-137, in Southern Lakes Wildlife Coordinating Committee. Regional Assessment of Wildlife in the Yukon Southern Lakes Region: Volume 2: Species Status Assessment. Environment Yukon, Whitehorse, Yukon.
Slough, B.G., and R.L. Mennell. 2006. Diversity and Range of Amphibians of the Yukon Territory. Canadian Field-Naturalist 120:87-92.
Smith, M.A. and D.M. Green. 2005. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: are all amphibian populations metapopulations? Ecography 28:110-128.
Stebbins, R.C. 2003. A field guide to western reptiles and amphibians. 3rd edition. The Peterson Field Guide Series. Houghton Mifflin Company, Boston, New York. 533 pp.
Stebbins, R.C., and N.W. Cohen. 1995. A natural history of amphibians. Princeton University Press, Princeton, New Jersey. 316 pp.
Steigerwald, M., unpubl. data. 2011. Assistant Collections Manager, Amphibian & Reptile Collection, Vertebrate Section, Canadian Museum of Nature, Ottawa, Ontario.
Stevens, C.E., and C.A. Paszkowski. 2006. Occurrence of the western toad and its use of 'borrow pits' in west-central Alberta. Northwestern Naturalist 87:107-117.
Stevens, C., C. Paszkowski, and A. Foote. 2007. Beaver (Castor canadensis) as a surrogate species for conserving anuran amphibians on boreal streams in Alberta, Canada. Biological Conservation 134:1-13.
Stevens, S.D., D.R.C. Prescott, and D.P. Whiteside. 2012. Occurrence and prevalence of chytrid fungus (Batrochochytrium dendrobatidis) in amphibian species of Alberta. Alberta Sustainable Resource Development, Fish and Wildlife Division, Alberta Species at Risk Report No. 143, Edmonton, Alberta. 24 pp.
Storer, T.I. 1925. A synopsis of the amphibia of California. University of California Publications in Zoology 27:1-342.
Stuart, S.N., J.S. Chanson, N.A. Cox, B.E. Young, A.S. Rodrigues, D.L. Fischman, and R.W. Waller. 2004. Status and trends of amphibian declines and extinctions worldwide. Science 306: 1783-1786.
Sullivan, J. 1994. Bufo boreas, in: Fire Effects Information System (U.S.Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory). Web site: http://www.fs.fed.us/database/feis/animals/amphibian/bubo/all.html [accessed 6 September 2012].
Tayless, E. 2011. Western toad population summary 2005-2011 (May 31), Lost Lake population, Whistler, B.C. Resort Municipality of Whistler, British Columbia. 6 pp.
Taylor, M. and C. M. Smith. 2003. Northern leopard frog and western toad inventory in Waterton Lakes National Park, Alberta, in 2003. Unpublished Technical Report. Parks Canada, Waterton Park, Alberta. 82 pp.
Thompson, M., unpubl. data. 2011. President, NAMOS BC(Northern Amphibian Monitoring Outpost Society), University of Northern British Columbia, Prince George, British Columbia.
Thompson, M., pers. comm. 2011. Email correspondence to B. Slough. May 2011. President, NAMOS BC (Northern Amphibian Monitoring Outpost Society), University of Northern British Columbia, Prince George, British Columbia.
Thompson, P.D. 2004. Observations of boreal toad (Bufo boreas) breeding populations in northwestern Utah. Herpetological Review 35:342-344.
Thoms, C., C.C. Corkran, and D.H. Olson. 1997. Basic amphibian survey for inventory and monitoring in lentic habitats. Pp. 35-46, in D.H. Olson, W.P. Leonard and R.B. Bury (eds). Sampling amphibians in lentic habitats. Northwest Fauna 4:1-134. Society for Northwestern Vertebrate Biology, Olympia, Washington.
Ultsch, G.R., D.F. Bradford, and J. Freda. 1999. Physiology: Coping with the environment. Pp. 189-214 in R.W. McDiarmid, and R. Altig (eds). Tadpoles: The biology of anuran larvae. The University of Chicago Press, Chicago, Illinois.
U.S. Fish and Wildlife Service. 2005. Revised 12-month finding for the Southern Rocky Mountain distinct population segment of the Boreal Toad (Bufo boreas boreas). Federal Register 70:56880–56884.
Van Tighem, K.J., and L.W. Gyug. 1984. Ecological land classification of Mount Revelstoke and Glacier National Parks, British Columbia. Vol. II, Wildlife Resource. Prepared by Canadian Wildlife Service, Edmonton, Alberta, for Parks Canada, Western Region. 254 pp.
Vertucci, F.A., and P.S, Corn. 1996. Evaluation of episodic acidification and amphibian declines in the Rocky Mountains. Ecological Applications 6:447-453.
Vredenburg, V.T., R.A. Knapp, T.S. Tunstall, and C.J. Briggs. 2010. Dynamics of an emerging disease drive large-scale amphibian population extinctions. Proceedings of the National Academy of Sciences of the U.S.A. 107:9689-9694.
Waldick, R.C., B. Freedman, and R.J. Wassersug. 1999. The consequences for amphibians of the conversion of natural, mixed-species forests to conifer plantations in southern New Brunswick, Canada. Canadian Field-Naturalist 113:404-418.
Ward, K., and B. Chapman. 1995. The Cariboo underworld: amphibians in clearcuts and forests. B.C. Nature Federation. Cordillera Winter 1995:32-41.
Washington Department of Fish and Wildlife. 2008. Priority Habitat and Species List. Olympia, Washington. 177 pp.
Wente, W.H., M.J. Adams, and C.A. Pearl. 2005. Evidence of decline for Bufo boreas and Rana luteiventris in and around the northern Great Basin, western USA. Alytes 22:95-108.
Wilkinson, L., and G. Berg. 2006. Researching amphibian numbers in Alberta (RANA): 2005 provincial summary. Alberta Species at Risk Report No. 110. Alberta Sustainable Resource Development, Fish and Wildlife Division, Edmonton, Alberta. 29 pp.
Wilkinson, L, and S. Hanus. 2003. Researching amphibian numbers in Alberta (RANA): 2002 provincial summary. Alberta Species at Risk Report No. 74. Alberta Sustainable Resource Development, Fish and Wildlife Division, Edmonton, Alberta. 19 pp.
Wind, E., unpubl. data. 2002, 2011. E. Wind Consulting, Nanaimo, British Columbia.
Wojtaszek, B.F., unpubl. data. 2011. Monitoring Ecologist, Gwaii Haanas National Park Reserve, National Marine Conservation Area Reserve, and Haida Heritage Site, Parks Canada, Skidegate, British Columbia.
Wong, C., unpubl. data. 2011. Ecosystem Scientist, Yukon Field Unit, Parks Canada, Whitehorse, Yukon.
Woodhams, D.C., R.A. Alford, and G. Marantelli. 2003. Emerging disease of amphibians cured by elevated body temperature. Diseases of Aquatic Organisms 55:65–67.
Woodhams, D.C., N. Kenyon, S.C. Bell, R.A. Alford, S. Chen, D. Billheimer, Y. Shyr, and L.A. Rollins-Smith. 2010. Adaptations of skin peptide defences and possible response to the amphibian chytrid fungus in populations of Australian green-eyed treefrogs, Litoria genimaculata. Diversity and Distributions 16:703-712.
Working Group on General Status of NWT Species. 2011. NWT species 2011-2015 – general status ranks of wild species in the Northwest Territories. Department of Environment and Natural Resources, Government of the Northwest Territories, Yellowknife, Northwest Territories. 172 pp.
Yukon Department of Environment. 2011. Management Plan for Amphibians in the Yukon. Fish and Wildlife Branch Technical Report. Whitehorse, Yukon. 24 pp. Draft.
Biographical summary of report writer
Brian G. Slough obtained an M.Sc. in Biological Sciences from Simon Fraser University in 1976. His thesis on beaver Castor canadensis ecology led him to a 15-year career as furbearer management biologist with the Yukon Fish and Wildlife Branch. He has published work on several furbearer species including beaver, arctic fox, Alopex lagopus, American marten, Martes americana, and Canada lynx, Lynx canadensis, and has also written about trapline and furbearer management in northern and western Canada. He prepared COSEWIC status reports on wolverine, Gulo gulo (2003) and American marten, Newfoundland population (M. americana atrata) (2007).
Since leaving the Yukon government in 1996, Mr. Slough has conducted environmental assessments, protected areas research, and research on rare amphibians and mammals, including rodents, shrews and bats. He has conducted extensive amphibian surveys in the Yukon and northern British Columbia and has sampled the region for amphibian chytrid fungus. He is currently serving a second term as a member of the Terrestrial Mammals Specialist Subcommittee of COSEWIC.
Collections examined
No collections were examined for the preparation of this status report.
Appendix 1. Sites verified to contain either non-calling or calling males of the Western Toad
Reproduction of Figure 2.1 from Pauli (2008)
Original figure caption: “Range map of Bufo boreas (modified from Stebbins, 1985) with localities of museum specimens examined for the occurrence of vocal sacs depicted (n = 419). At each site, 1–31 males were examined, and either all males lacked vocal sacs (white), all males had vocal sacs (black), or males with and without vocal sacs were present (gray). Numbered sites in Alberta are localities where males were recorded. The in northern Alberta is a site where individuals were observed to have vocal sacs and produce long, pulsed calls but no specimens or recordings were taken”.
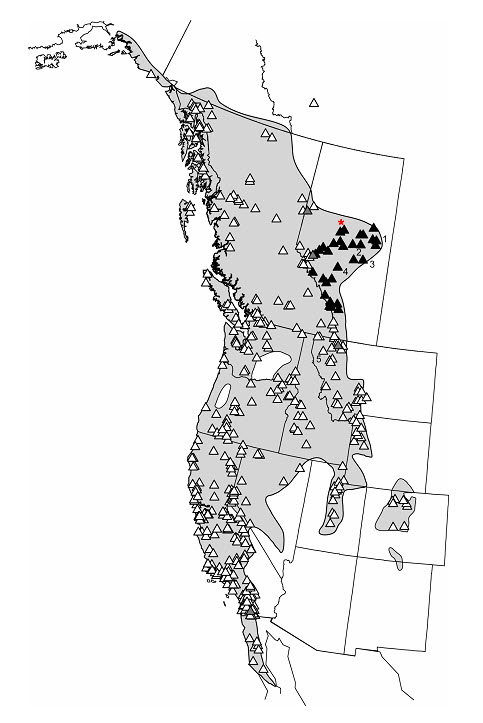
Description for Appendix 1
Map of sites confirmed as containing either non-calling or calling males of the Western Toad. Sites where all males lacked vocal sacs are shown by white triangles; sites where all males had vocal sacs are shown by black triangles; and sites where males with and without vocal sacs were present are shown by grey triangles.
Appendix 2. Threats calculator results for the non-calling population of the Western Toad
Assessment conducted on 27 February 2012. Participants: Dave Fraser, Syd Cannings, Brian Slough, Cindy Paszkowski, Kristiina Ovaska; Angele Cyr (COSEWIC Secretariat; minutes); modified from initial assessment for the B.C. population for Western Toad in 2010 (which was revised by Dave Fraser & Orville Dyer on 7 Feb 2012). Generation time: 6 yrs ; hence severity assessed for 18 years.
| Threat impact | High range | Low range |
|---|---|---|
| A = Very High | 0 | 0 |
| B = High | 0 | 0 |
| C = Medium | 2 | 0 |
| D = Low | 2 | 4 |
| Calculated overall threat impact: | High | Medium |
| Threat | Impact (calculated) | Scope (next 10 years) | Severity (10 years or 3 generations) | Timing | Comments |
|---|---|---|---|---|---|
| 1 Residential & commercial development | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | |
| 1.1 Housing & urban areas | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | |
| 1.2 Commercial & industrial areas | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | |
| 1.3 Tourism & recreation areas | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 2 Agriculture & aquaculture | Negligible | Large (31-70%) | Negligible (<1%) | High (Continuing) | |
| 2.1 Annual & perennial non-timber crops | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | |
| 2.2 Wood & pulp plantations | |||||
| 2.3 Livestock farming & ranching | Negligible | Large (31-70%) | Negligible (<1%) | High (Continuing) | Livestock as far as Prince George & beyond to Fort St. James and little beyond in the north; mountain & coastal forests are also excluded from scope. Scope reduced to large from large-pervasive for BC only. Negative effects from trampling & shoreline modification. Possibility of creating habitat; toads known to breed successfully in dugouts & in cattle pastures; not heavy impact |
| 2.4 Marine & freshwater aquaculture | |||||
| 3 Energy production & mining | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | |
| 3.1 Oil & gas drilling | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | |
| 3.2 Mining & quarrying | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | |
| 3.3 Renewable energy | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 4 Transportation & service corridors | CD = Medium - Low | Restricted (11-30%) | Extreme - Moderate (11-100%) | High (Continuing) | |
| 4.1 Roads & railroads | CD = Medium - Low | Restricted (11-30%) | Extreme - Moderate (11-100%) | High (Continuing) | Roadkill risk during mass migrations; mortality of females that breed only once in their life time is significant |
| 4.2 Utility & service lines | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 4.3 Shipping lanes | |||||
| 4.4 Flight paths | |||||
| 5 Biological resource use | D = Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | |
| 5.1 Hunting & collecting terrestrial animals | |||||
| 5.2 Gathering terrestrial plants | |||||
| 5.3 Logging & wood harvesting | D = Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | |
| 5.4 Fishing & harvesting aquatic resources | |||||
| 6 Human intrusions & disturbance | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 6.1 Recreational activities | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 6.2 War, civil unrest & military exercises | |||||
| 6.3 Work & other activities | |||||
| 7 Natural system modifications | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 7.1 Fire & fire suppression | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 7.2 Dams & water management/use | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 7.3 Other ecosystem modifications | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 8 Invasive & other problematic species & genes | CD = Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | |
| 8.1 Invasive non-native/alien species | CD = Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | Incorporates threats from disease (chytrid, rana virus, Saprolegnia), fish, bullfrogs (in southwest); scope scored pervasive mainly because of chytrid (Bd). Severity: Pathogenity could increase through introduction of new strains and their hybridization (Farrer et al. 2011) and sweep across the range. Interactions with climate change: higher temperature could increase suitability of higher elevation habitats to Bdand facilitate its spread (Muths et al. 2008); introduced fish: ranavirus spread by fish. Purnima Govindarajulu's review of B.C. threats assessment indicated that severity due to chytrid should be higher than small as initially assessed. |
| 8.2 Problematic native species | |||||
| 8.3 Introduced genetic material | |||||
| 9 Pollution | D = Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | |
| 9.1 Household sewage & urban waste water | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 9.2 Industrial & military effluents | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 9.3 Agricultural & forestry effluents | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 9.4 Garbage & solid waste | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 9.5 Air-borne pollutants | D = Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Pollutants transported far from source of origin and end up in water bodies, even at high elevation lakes. Detected at higher elevation sites in Okanagan & also in Alberta. |
| 9.6 Excess energy | |||||
| 10 Geological events | |||||
| 10.1 Volcanoes | |||||
| 10.2 Earthquakes/tsunamis | |||||
| 10.3 Avalanches/landslides | |||||
| 11 Climate change & severe weather | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 11.1 Habitat shifting & alteration | This threat can become an issue over longer term but not considered so over the next 10 years. | ||||
| 11.2 Droughts | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | This threat can become more of an issue over longer term; some changes are already happening. Some climate change effects may be beneficial, such as warmer, wetter spring that permits earlier breeding, but shorter hydro-periods of breeding sites (ponds, shallows of larger water bodies) would be detrimental. |
| 11.3 Temperature extremes | |||||
| 11.4 Storms & flooding |
Classification of Threats adopted from IUCN and CMP (2006), Salafsky et al.(2008).
Appendix 3. Threats calculator results for the calling population of the Western Toad
Assessment conducted on 27 February 2012. Participants: Dave Fraser, Syd Cannings, Brian Slough, Cindy Paszkowski, Kristiina Ovaska; Angele Cyr (COSEWIC Secretariat; minutes); modified from initial assessment for the B.C. population for Western Toad in 2010 (which was revised by Dave Fraser & Orville Dyer on 7 Feb 2012). Generation time: 6 yrs ; hence severity assessed for 18 years.
| Threat impact | High range | Low range |
|---|---|---|
| A = Very High | 0 | 0 |
| B = High | 1 | 1 |
| C = Medium | 3 | 0 |
| D = Low | 3 | 6 |
| Calculated overall threat impact: | Very high | High |
| Threat | Impact (calculated) | Scope (next 10 years) | Severity (10 years or 3 generations) | Timing | Comments |
|---|---|---|---|---|---|
| 1 Residential & commercial development | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | |
| 1.1 Housing & urban areas | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | Housing expanding around Edmonton and Calgary but is still negligible when the entire range is considered. |
| 1.2 Commercial & industrial areas | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | |
| 1.3 Tourism & recreation areas | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 2 Agriculture & aquaculture | CD - Medium - Low | Restricted - Small (1-30%) | Serious - Moderate (11-70%) | High (Continuing) | |
| 2.1 Annual & perennial non-timber crops | CD - Medium - Low | Restricted - Small (1-30%) | Serious - Moderate (11-70%) | High (Continuing) | Row crops such as alfalfa and canola are prevalent in some inhabited zones away from the mountains. Toads occur in these areas but likely suffer increased mortality (e.g., encounters with farm machinery) and destruction of breeding ponds and hibernating sites. |
| 2.2 Wood & pulp plantations | |||||
| 2.3 Livestock farming & ranching | Negligible | Large (31-70%) | Negligible (<1%) | High (Continuing) | Negative effects from trampling & shoreline modification. Possibility of creating habitat; toads known to breed successfully in dugouts & natural ponds in cattle pastures; not heavy impact but could be affected by habitat modification. |
| 2.4 Marine & freshwater aquaculture | |||||
| 3 Energy production & mining | B - High | Large (31-70%) | Serious (31-70%) | High (Continuing) | |
| 3.1 Oil & gas drilling | B - High | Large (31-70%) | Serious (31-70%) | High (Continuing) | Everywhere, scope large. Severity: gas pads & structures may be next to breeding sites or hibernating sites. Compressor noise may affect breeding choruses. |
| 3.2 Mining & quarrying | Negligible | Negligible (<1%) | Serious (31-70%) | High (Continuing) | Coal mining and gravel extraction in some areas but scope is small. Peat mining could destroy hibernating sites. |
| 3.3 Renewable energy | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | Wind farms |
| 4 Transportation & service corridors | CD - Medium - Low | Restricted - Small (1-30%) | Extreme - Moderate (11-100%) | High (Continuing) | |
| 4.1 Roads & railroads | CD - Medium - Low | Restricted - Small (1-30%) | Extreme - Moderate (11-100%) | High (Continuing) | Scope: Paved roads through toad habitat are relatively few, and few problem areas for roadkill identified; however, numerous gravel resource roads are present through much of the range. Severity: Roadkill risk during mass migrations; mortality of females that breed only once in their life time is significant. Road construction can create potential breeding sites through the excavation of borrow pits. |
| 4.2 Utility & service lines | D - Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Numerous seismic lines associated with oil & gas. Fragmentation of natural vegetation. |
| 4.3 Shipping lanes | |||||
| 4.4 Flight paths | |||||
| 5 Biological resource use | D = Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | |
| 5.1 Hunting & collecting terrestrial animals | |||||
| 5.2 Gathering terrestrial plants | |||||
| 5.3 Logging & wood harvesting | D = Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | Similar to B.C. in scope. Harvested areas for pulp & paper and timber in foothills and boreal forest. Toads are relatively tolerant of logging and use opened up (warmer) areas and shallow ponds resulting from reduced surface water loss via transpiration. |
| 5.4 Fishing & harvesting aquatic resources | |||||
| 6 Human intrusions & disturbance | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 6.1 Recreational activities | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | Skiing, snowmobiling, hiking, camping |
| 6.2 War, civil unrest & military exercises | |||||
| 6.3 Work & other activities | |||||
| 7 Natural system modifications | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 7.1 Fire & fire suppression | Negligible | Negligible (<1%) | Moderate (11-30%) | High (Continuing) | |
| 7.2 Dams & water management/use | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 7.3 Other ecosystem modifications | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 8 Invasive & other problematic species & genes | C = Medium | Pervasive (71-100%) | Moderate – Slight (1-30%) | High (Continuing) | |
| 8.1 Invasive non-native/alien species | C = Medium | Pervasive (71-100%) | Moderate – Slight (1-30%) | High (Continuing) | Incorporates threats from disease (chytrid, rana virus, Saprolegnia), fish, bullfrogs (in southwest); scope scored pervasive mainly because of chytrid (Bd). Severity: Pathogenity could increase through introduction of new strains and their hybridization (Farrer et al. 2011) and sweep across the range. Interactions with climate change: higher temperature could increase suitability of higher elevation habitats to Bd and facilitate its spread (Muths et al. 2008); introduced fish: ranavirus spread by fish. Purnima Govindarajulu's review of B.C. threats assessment indicated that severity due to chytrid should be higher than small as initially assessed. |
| 8.2 Problematic native species | Some evidence of hybridization with Canadian Toad, but narrow zone of known overlap in small area. Could suffer increased predation from subsidized predators, e.g., corvids, skunks. | ||||
| 8.3 Introduced genetic material | |||||
| 9 Pollution | D = Low | Restricted - Small (1-30%) | Moderate - Slight (1-30%) | High (Continuing) | |
| 9.1 Household sewage & urban waste water | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 9.2 Industrial & military effluents | D = Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Fracking & other effluents from oil & gas extraction; surface fill |
| 9.3 Agricultural & forestry effluents | D = Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Includes run-off from forestry roads and agricultural effluents |
| 9.4 Garbage & solid waste | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | |
| 9.5 Air-borne pollutants | D = Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Pollutants transported far from source of origin and end up in water bodies, even at high elevation lakes. Detected at higher elevation sites in Okanagan & also in Alberta. Both scope and severity are higher than individual threat scores for sub-categories because they are additive. |
| 9.6 Excess energy | |||||
| 10 Geological events | |||||
| 10.1 Volcanoes | |||||
| 10.2 Earthquakes/tsunamis | |||||
| 10.3 Avalanches/landslides | |||||
| 11 Climate change & severe weather | D = Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | |
| 11.1 Habitat shifting & alteration | This threat can become an issue over longer term but not considered so over the next 10 years. | ||||
| 11.2 Droughts | D = Low | Small (1-10%) | Moderate (11-30%) | High (Continuing) | Reality for parklands and amphibian breeding sites there; predicted to get worse. Some climate change effects may be beneficial, such as warmer, wetter spring that permits earlier breeding, but shorter hydro-periods of breeding sites (ponds, shallows of larger water bodies) would be detrimental. |
| 11.3 Temperature extremes | |||||
| 11.4 Storms & flooding |
Classification of Threats adopted from IUCN and CMP (2006), Salafsky et al.(2008).