
Wolverine (Gulo gulo): COSEWIC assessment and status report 2014

Special Concern
2014
Table of Contents
- Document Information
- COSEWIC Assessment Summary
- COSEWIC Executive Summary
- Technical Summary
- Preface
- Wildlife Species Description and Significance
- Distribution
- Habitat
- Biology
- Population Sizes and Trends
- Threats and Limiting Factors
- Protection, Status, and Ranks
- Acknowledgements and Authorities Contacted
- Information Sources
- Biographical Summary of Report Writer(s)
- Collections Examined
List of Figures
- Figure 1. Illustration of the Wolverine. (Credit: Lee Mennell)
- Figure 2. Median-joining network of mtDNA control region haplotypes of samples originating from Eurasia (Sweden, Norway, Mongolia, Russia) and North America (see Zigouris et al. 2013 for jurisdictions used in key). Each branch represents one mutation step. (Source: Zigouris et al. 2013 [doi:10.1371/journal.pone.0083837.g004], used with permission)
- Figure 3. North American distribution of Wolverine. Adapted from COSEWIC (2003), Magoun et al. (2004), Ray (2004, 2012), Aubry et al. (2007), Thibault unpubl. data (2013). Map produced by Bonnie Fournier, NWT. Increased presence refers to observations of Wolverine on various islands, but it is not known if these are established or vagrant individuals
- Figure 4. Locations of Wolverine records based on aerial surveys conducted by Wildlife Conservation Society (WCS) and Ontario Ministry of Natural Resources (OMNR). The area west of the modelled core and peripheral range represents the main and peripheral locations of Wolverine before 2004. The large number of recent (>2009) records indicates an eastern expansion of the sub-population. (Source: Ontario Wolverine Recovery Team 2013; Ray unpubl. data.)
- Figure 5. Locations of Wolverine sighting reports in Québec, 2000 to 2012, according to their relative level of credibility. No sightings have been verified. Adapted from Thibault unpubl. data (2013)
List of Appendices
Document Information

Committee on the Status of Endangered Wildlife in Canada (COSEWIC) status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2014. COSEWIC assessment and status report on the Wolverine Gulo gulo in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xi + 76 pp. (Species at Risk Public Registry website).
Previous report(s):
COSEWIC. 2003. COSEWIC assessment and update status report on the wolverine Gulo gulo in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 41 pp.
Dauphiné, T.C. 1989. Update COSEWIC status report on the wolverine Gulo gulo in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 31 pp.
Kelsall, J.P. 1982. COSEWIC status report on the wolverine Gulo gulou in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 50 pp.
Production note:
COSEWIC would like to acknowledge Brian Slough for writing the status report on the Wolverine (Gulo gulo) in Canada, prepared under contract with Environment Canada. This status report was overseen and edited by Graham Forbes, Co-chair of the COSEWIC Terrestrial Mammals Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-953-3215
Fax: 819-994-3684
COSEWIC E-mail
COSEWIC web site
Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur le Carcajou (Gulo gulo) au Canada.
Cover illustration/photo:
Wolverine -- Illustration by Lee Mennell
©Her Majesty the Queen in Right of Canada, 2014.
Catalogue No. CW69-14/329-2014E-PDF
ISBN 978-1-100-23964-4

COSEWIC Assessment Summary
Assessment Summary - November 2014
- Common name
- Wolverine
- Scientific name
- Gulo gulo
- Status
- Special Concern
- Reason for designation
- This wide-ranging carnivore has an estimated Canadian population likely exceeding 10,000 mature individuals. Although population increases appear to be occurring in portions of the Northwest Territories, Nunavut, Manitoba and Ontario, declines have been reported in the southern part of the range, e.g. in British Columbia, and populations in a large part of the range (Quebec and Labrador) have not recovered. The species may be extirpated from Vancouver Island. Population estimates are very limited, and trends are not known. Most data are limited to harvest records, and harvest levels may be under-reported because many pelts used domestically are not included in official statistics. There is no evidence, however, of a decline in harvest over the last 3 generations. This species’ habitat is increasingly fragmented by industrial activity, especially in the southern part of its range, and increased motorized access increases harvest pressure. Climate change is likely impacting animals in the southern part of the range, and this impact is expected to increase northward. The species has a low reproductive rate, is sensitive to human disturbance, and requires vast secure areas to maintain viable populations.
- Occurrence
- Yukon, Northwest Territories, Nunavut, British Columbia, Alberta, Saskatchewan, Manitoba, Ontario, Quebec, Newfoundland and Labrador
- Status history
- The species was considered a single unit and designated Special Concern in April 1982. Split into two populations in April 1989 (Western and Eastern populations). The original designation was de-activated. In May 2014, the Eastern and Western populations were considered as a single unit across the Canadian range and was designated Special Concern.
COSEWIC Executive Summary
Wolverine
Gulo gulo
Wildlife Species Description and Significance
Wolverines are a stocky, medium-sized carnivore and the largest terrestrial member of the weasel family. They have long, glossy coarse fur, which varies from brown to black, often with a pale facial mask and stripes running laterally from the shoulders, crossing just above the tail. The skull structure is robust, allowing it to crush and consume bones and frozen carcasses. Adult males weigh 13 to 18 kg and adult females weigh 7.5 to 12.5 kg.
A single subspecies, Gulo gulo luscus, ranges across most of Canada. Further studies are required to determine if the Vancouver Island population is a separate subspecies, G. gulo vancouverensis. A single designatable unit is recognized for the Canadian population.
Wolverines may indicate ecosystem health, given their dependence on extensive connected ecosystems that support ungulates and large carnivores which create opportunities for scavenging. They are a valuable furbearer in the fur trade, and many furs that do not enter the fur trade are used locally, especially in the Arctic, where its frost-resistant fur is used for parka trim.
Distribution
Wolverines are found across northern Eurasia and North America. In Canada, they are found in northern and western forested areas, in alpine tundra of the western mountains, and in arctic tundra. It is not known whether Wolverines currently occupy Vancouver Island, Québec, or Labrador. Range reductions began in the 19th century, and subpopulations were extirpated from New Brunswick, southern Ontario, and from the aspen parkland of Manitoba, Saskatchewan and Alberta.
Habitat
A wide variety of forested and tundra vegetation associations are used by Wolverines. Habitats must have an adequate year-round supply of food, mainly consisting of smaller prey such as rodents and Snowshoe Hares, and the carcasses of large ungulates, like Moose, Caribou, and Muskox. Females den under snow-covered rocks, logs or within snow tunnels. Wolverines reproduce in areas where snow cover persists at least into April.
Biology
Most females breed after they are 2 or 3 years of age and produce on average 2 kits per litter. Wolverine home ranges are 50-400 km2 for females (smallest during denning periods) and 230-1580 km2 for males. Home ranges may overlap within and between sexes but home ranges of reproductive females do not overlap. Home range size in the eastern range is unknown. Wolverine densities are low and range from about 5 to 10/1,000 km2. Wolverines are scavengers and predators, often caching food for future use. Wolverines face mortality from predation and starvation. Anthropogenic sources of mortality include trapping, hunting, and road kill.
Population Sizes and Trends
Confidence regarding population size and trend is debated because most population data are derived from harvest records and the proportion of unreported harvest varies across their range. The Canadian population size is unknown but likely > 10,000 adults. Wolverine have been extirpated (or likely extirpated) in much of southern and eastern Canada. Wolverine observations continue to be reported in the range of the eastern sub-population (Québec and Labrador), but no observation has been verified since 1978. The last verified record on Vancouver Island was in 1991 and it is likely that they have been extirpated. There is concern that decline may be occurring in BC and parts of Alberta where Southern and Central Mountain Caribou, their primary prey, continue to decline and habitats are fragmented. Field studies since 2003 suggest Wolverines are more abundant in parts of the Northwest Territories (NWT) and Nunavut than previously thought. The sub-population in the NWT barren ground region may be decreasing while recent records in western Arctic islands suggest population increase, although it is unknown if these are resident or transient animals. Population trends are unknown, but based on numerous sources such as field studies, ATK, and trapper surveys, they are believed to have been stable over parts of the northern range for the last 3 generations (22.5 years). Wolverines in northern Manitoba and Ontario may be increasing; aerial surveys in northern Ontario have shown an eastward range reoccupation towards James Bay and Québec.
Threats and Limiting Factors
The variability in trap effort, the uncertainty on actual harvest levels in some jurisdictions, and increased access and efficiency of hunting using snowmobiles raises concerns over potential overharvest and the ability to document population size and trends. Transportation corridors, forestry, hydroelectric developments, oil and gas and mineral exploration and development increase access for harvest and contribute to permanent, temporary or functional habitat losses (sensitivity to disturbance), which may destabilize populations.
The decline in Caribou as a source of scavenged meat, particularly in Québec and Labrador where few Wolverines may persist, may limit population recovery. Other factors that may limit populations include harvest, disturbance of denning areas, threats to habitats, and population fluctuations in Wolves and other carnivores that provide scavenging opportunities. The Threats Calculator calculated an overall threat impact of medium.
Protection, Status, and Ranks
This species was assessed as Endangered by COSEWIC in 2003 and is listed on Schedule 1 of the Species at Risk Act(SARA). The Western Population (labelled western sub-population in this report) was assessed as Special Concern by COSEWIC in 2003, but was not listed under SARA due to concerns expressed by the Nunavut Wildlife Management Board. Provincial designations are Endangered in Labrador, and Threatened in Ontario and Québec (note: 'Threatened' is equivalent to Endangered in Québec). Remaining provincial designations range from no ranking to Sensitive or Special Concern. NatureServe (2013) rankings are Critically Imperilled (S1) in Québec and Newfoundland and Labrador, and Imperilled (S2) in Ontario. The Vancouver Island population is Imperilled. Wolverines are protected from non-Aboriginal harvest in Québec, Newfoundland and Labrador, and Ontario, although unreported harvest may be occurring. Aboriginal harvest would be in the northern part of the range (i.e.,James Bay and Northern Québec Agreement area). Wolverines are trapped and hunted in most other areas of their confirmed range.
Habitat Protection and Ownership
Numerous protected areas exist within the Wolverine’s range but refugia larger than 20,000 km2 may be required to maintain a Wolverine population. Many northern national, provincial and territorial parks allow trapping. In southern parks, population recovery may be impacted by road developments that can act as barriers to movements, and activities such as skiing and snowmobiling that may disturb denning females.
Technical Summary
Gulo gulo
Wolverine
Carcajou
- Range of occurrence in Canada (province/territory/ocean):
- Yukon, Northwest Territories, Nunavut, British Columbia, Alberta, Saskatchewan, Manitoba, Ontario, Québec, Newfoundland and Labrador
Demographic Information
- Generation time (estimated average age of breeding females in the population)
- 7.5 yrs
-
Is there an observed continuing decline in number of mature individuals?
Last verified record in eastern sub-population was in 1978, and Vancouver Island in 1991. - No; declines in south and east have occurred but some sub-populations may be increasing in north
-
Estimated percent of continuing decline in total number of mature individuals within 2 generations (15 years).
Harvest indices in much of northern range suggest stable populations but value of trap data uncertain. - Unknown, but possibly increasing in northern, and likely decreasing in southern range
- Observed percent reduction in total number of mature individuals over the last 3 generations (22.5 years)
- 0
- Projected percent reduction in total number of mature individuals over the next 3 generations (22.5 years).
- Unknown
-
Observed percent increase in total number of mature individuals over any 3 generations period, over a time period including both the past and the future.
There are reports of possible population increases in Nunavut and NWT. - Unknown
-
Are the causes of the decline clearly reversible and understood and ceased?
Declines in Vancouver Island and eastern sub-population are not understood and have not reversed. - No
- Are there extreme fluctuations in number of mature individuals?
- No
Extent and Occupancy Information
-
Estimated extent of occurrence
If unverified observations in eastern sub-population are used, EO is an additional 0.88 million km2. - >10 million km2
- Index of area of occupancy (IAO)
- >4,000 km2
- Is the total population severely fragmented?
- No
- Number of locations Extent and Occupancy Information Footnote1
- Many
- Is there an observed continuing decline in extent of occurrence?
- No
- Is there an observed continuing decline in index of area of occupancy?
- No
- Is there an observed continuing decline in number of populations?
- No
- Is there an [observed, inferred, or projected] continuing decline in number of locations Extent and Occupancy Information Footnote1?
- No
- Is there an observed continuing decline in quality of habitat?
- Yes, in some areas, notably in southern range, and areas of industrial development
- Are there extreme fluctuations in number of populations?
- No
- Are there extreme fluctuations in number of locations Extent and Occupancy Information Footnote1?
- No
- Are there extreme fluctuations in extent of occurrence?
- No
- Are there extreme fluctuations in index of area of occupancy?
- No
| Western sub-population (ON, MB, NU, NWT, YK, SK, AB, BC) | Unknown, likely >10,000 |
| Eastern sub-population (LAB, QU) | Unknown, likely near 0 |
| Total | Unknown, likely >10,000 mature individuals |
Quantitative Analysis
- Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years].
- Not conducted, but likely a high probability in eastern sub-population range
Threats (actual or imminent, to populations or habitats)
Potential overharvest associated with unknown harvest levels and facilitated by extensive access associated with snowmobile use. Habitat loss (permanent, temporary and/or functional) and fragmentation due to forestry, agriculture, oil and gas development, hydroelectric reservoirs, and roads. In southern range, functional habitat loss due to disturbance caused by recreational activities, such as ATVs, snowmobiles, hiking and skiing during the denning period, along roads and trails. Declining ungulate (esp. Caribou) populations represent loss of important source of food. Climate change concerns in southern range.
Rescue Effect (immigration from outside Canada)
-
Status of outside population(s)?
Secure in Alaska; Alaskan population is adjacent to northern BC, YT. Sensitive or At Risk in the conterminous U.S. states and a candidate species for federal listing as Threatened; southern population is adjacent to BC and Alberta. - Secure in Alaska; potentially Threatened in US states south of Canada
- Is immigration known or possible?
- Yes
- Would immigrants be adapted to survive in Canada?
- Yes
- Is there sufficient habitat for immigrants in Canada?
- Yes
-
Is rescue from outside populations likely?
Rescue possible from Alaska but unlikely from conterminous US. - Yes/No
Data-Sensitive Species
- Is this a data-sensitive species?
- No
Status History
- COSEWIC:
- The species was considered a single unit and designated Special Concern in April 1982. Split into two populations in April 1989 (Western and Eastern populations). The original designation was de-activated. In May 2014, the Eastern and Western populations were considered as a single unit across the Canadian range and was designated Special Concern.
Status and Reasons for Designation
- Status:
- Special Concern
- Alpha-numeric code:
- Not applicable
- Reason for Designation:
- This wide-ranging carnivore has an estimated Canadian population likely exceeding 10,000 mature individuals. Although population increases appear to be occurring in portions of the Northwest Territories, Nunavut, Manitoba and Ontario, declines have been reported in the southern part of the range, e.g. in British Columbia, and populations in a large part of the range (Quebec and Labrador) have not recovered. The species may be extirpated from Vancouver Island. Population estimates are very limited, and trends are not known. Most data are limited to harvest records, and harvest levels may be under-reported because many pelts used domestically are not included in official statistics. There is no evidence, however, of a decline in harvest over the last 3 generations. This species' habitat is increasingly fragmented by industrial activity, especially in the southern part of its range, and increased motorized access increases harvest pressure. Climate change is likely impacting animals in the southern part of the range, and this impact is expected to increase northward. The species has a low reproductive rate, is sensitive to human disturbance, and requires vast secure areas to maintain viable populations.
Applicability of Criteria
- Criterion A (Decline in Total Number of Mature Individuals):
- Unknown. Evidence for declines in some parts of its range, but may be increasing in others.
- Criterion B (Small Distribution Range and Decline or Fluctuation):
- Not Applicable. EO and IAO exceed thresholds.
- Criterion C (Small and Declining Number of Mature Individuals):
- Not Applicable. Population size likely exceeds 10,000 mature individuals.
- Criterion D (Very Small or Restricted Population):
- Not Applicable. Population exceeds 1,000 mature individuals.
- Criterion E (Quantitative Analysis):
- Not Applicable. Analysis has not been conducted for the Canadian population.
Preface
Notice: Some of the information used or referenced in this document is Government of the Northwest Territories Copyright, compiled on behalf of the Northwest Territories Species at Risk Committee under a contract with the Department of Environment and Natural Resources (Northwest Territories Species at Risk Committee (NWT SARC) 2013).
The original April 1982 COSEWIC status designation for Wolverines was Rare, which was synonymous with Special Concern prior to 1990 (Kelsall 1981). In April 1989, two populations were delineated, the Eastern Population of Québec and Labrador, and the Western Population of Ontario, Manitoba, Saskatchewan, Alberta, British Columbia, Yukon, Northwest Territories (NWT), and Nunavut (Dauphiné 1989). The justification for the separation was based on the very low, or extirpated, population found in Québec and Labrador, rather than the criteria of discreteness or significance used in present COSEWIC guidelines. Dauphiné (1989) suggested that the Eastern Population should be classified as Extirpated.
The COSEWIC status designations were Vulnerable for the Western Population (again, synonymous with Special Concern between 1990 and 1999), and Endangered for the Eastern Population. The status designations were confirmed in May 2003 for the Western Population (Special Concern) and Eastern Population (Endangered) (COSEWIC 2003). The Eastern Population is listed on Schedule 1 of the Species at Risk Act (SARA) as Endangered, but the Western Population was not added to the SARA list in order to further consult with the Nunavut Wildlife Management Board (NWMB) and the Nunavut government. Issues for the NWMB centred on the lack of ATK, inadequate consultation, issues with land claims agreement, and evidence of increasing population in Nunavut (Crystal pers. comm. 2014). The consultation process occurred in 2005; however, the Minister of Environment did not issue a recommendation.
A substantial number of Wolverine research projects underway at the time of writing the previous update status report on Wolverine (COSEWIC 2003) have since been completed. There have been publications on the effect of global warming on the Wolverines’ habitat requirement of late spring snow cover, genetic structuring at the southwest and eastern peripheries of the Wolverine’s range, distribution and abundance of Wolverine at the eastern edge of the species’ range in Ontario, and population density estimates in the southern Arctic ecozone in the NWT and Nunavut, and in montane and foothill habitats in western Alberta.
Ecozones and ecoregions discussed in this report are based on the scheme presented by the Ecological Stratification Working Group (1995). The 8 COSEWIC 'National Ecological Areas' are amalgamations of the 15 terrestrial ecozones.

COSEWIC History
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC Mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC Membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2014)
Wildlife Species
A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
- Special Concern (SC)Footnotea
- A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
- Not at Risk (NAR)Footnoteb
- A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
- Data Deficient (DD)Footnotec
- A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife Species Description and Significance
Name and Classification
- Family:
- Mustelidae
- Latin Name :
- Gulo gulo
- English Common Name:
- Wolverine
- French Common Name:
- Carcajou
- Aboriginal Names:
- ʔa¢̕pu (Ktunaxa), cišps (Salish), Kalvik / Qalviit (Inuktitut), Qavviit (Inuvialuit), kihkwahâkew (Plains Cree), Kuakuatsheu / kuekuatsheu (Innu), mámex-áy-shen (she shashishalhem), naghay (southern Tutchone), nàhtyä’ (Hän), Nehttryooh/Nehtryuh (Gwich’in – Gwich’in Settlement Area), Nehttryuh / nèhtrùh (Gwich’in – North Yukon), Nogha / Nághai (Dene), Noolh’utughih (Dakelh – Southern Carrier), noosik’ (Gitsenimx and Nisga’a), nóoskw (Tlingit), Noostel (Dakelh – Nadleh Whut’en), Noostel (Dakelh – Nak’azdli Dakelh), nòòwa (Kwadacha Tsek’ene) , nowa (Tse’K’hene), nuŝtil (Tsilhqot’in (Xeni Gwet’in)), nowe (Tsaa? Dene (Beaver People)), nustël (Wet’suwet’en), ogwiingwa’aage+g (Ojibwe), Piinotoyi (Blackfoot), Qavvik (Inuktitut – Inuit), quts’ik (Nuxalk), Qwílqwn (Secwepemc – Northern Státimcets), qwílqen ((Ĺiĺwat), shxwématsel (Upriver Halkomelem)).
Wolverines (Gulo gulo; Linnaeus 1758) in North America were formerly known as Gulo luscus until they were shown to be conspecific with Wolverine in Europe and Siberia (Kurtén and Rausch 1959). Up to four subspecies had been recognized in North America (Hall 1981), two of which occur in Canada; G. g. luscus, found across Canada, Alaska and the northwestern United States, and G. g. vancouverensis, found on Vancouver Island (see Designatable Units section). Taxonomists presently recognize only either a single subspecies (G. g. luscus in North America), or recognize G. gulo as a single Holarctic taxon (reviewed in Tomasik and Cook 2005).
Morphological Description
Wolverines are a medium-sized (i.e. 1-m long) carnivore and the largest terrestrial member of the weasel family in North America, appearing more like a small bear than a weasel (Figure 1; Inuvik Community Corporation (ICC) et al. 2006). It has long, glossy coarse fur, which varies from brown to black, often with a pale facial mask and yellowish or tan stripes running laterally from the shoulders, crossing just above the tail (Gwich’in Renewable Resources Board (GRRB) 2001; Cardinal 2004). Most individuals have a pale white or orange patch on the neck and chest. It has a large head, broad forehead, short stout neck, short stocky legs, and a heavy musculature. The feet are large, ears short and the tail is long and bushy. The skull structure is robust, allowing it to crush bones and consume frozen carcasses. Wolverines are sexually dimorphic with adult females ranging in size from 7.5 to 12.5 kg and males weighing 13 to 18 kg (Magoun 1985; Banci 1994; Copeland 1996; Mulders 2000; GRRB 2001; Lofroth 2001; Cardinal 2004; Golden unpubl. data 2013).
Figure 1 : Wolverine © Environment Canada
Population Spatial Structure and Variability
The amount of population genetic structuring varies by scale. At the continental scale, Wolverine have limited structuring in the core range but increasing structure at the southwestern and eastern range peripheries (Wilson et al. 2000; Kyle and Strobeck 2001; 2002; Chappell et al. 2004; Zigouris et al. 2012). These studies are based on nuclear DNA markers. The largely panmictic condition in the core is believed to be due to long-range dispersal behaviour by males (see Dispersal and Migration section). The increased structure at the peripheries is believed to be due to isolation, and possibly separate origins from multiple glacial refugia (see Designatable Units section). At the local scale, increased genetic structuring has been recorded using mitochondrial DNA (mtDNA) analyses (Tomasik and Cook 2005; Cegelski et al. 2006; Schwartz et al. 2007; Zigouris et al. 2012; 2013). This local structuring is believed to be due to a strong preference by females to remain near their natal territories (Tomasik and Cook 2005; Cegelski et al. 2006; Schwartz et al. 2007; Zigouris et al. 2012).
Designatable Units
A single subspecies is presently recognized for Wolverine in North America (Tomasik and Cook 2005). Earlier classifications had recognized a separate subspecies on Vancouver Island but its status is debated; the 1935 proposal was based on a sample of only two skulls (Cowan and Guiguet 1960) and Banci (1982) concludes there is little evidence for classifying the Vancouver Island population as a distinct subspecies, based on morphometrics. Adequate samples are not available for significant genetic analyses; however, the one sample that had been processed for the control region of mtDNA revealed a genetic type found throughout the contemporary range of wolverines (Zigouris pers. comm. 2013), suggesting a lack of uniqueness from mainland Wolverine. Until further work is done, this report does not recognize the Vancouver Island population as a designatable unit (DU). According to knowledge holders in the ATK report on Wolverine (Cardinal 2004), ATK across most of the range does not differentiate Wolverine into more than one type. However, there is ATK from the Kivalliq region of Nunavut noting two types; one variety that is larger and lighter-coloured (“greater wolverine”), and another variety that is smaller and darker (“lesser wolverine”), and more common towards Yellowknife. Some participants stated that the greater wolverine variety may just be older individuals and the lesser wolverines are younger ones. However, one knowledge holder in Arviat stated that the teeth and claws of some of the lesser variety that he had caught indicated that they were older individuals, suggesting two types. Cardinal (2004) notes that without more research, it is difficult to discern whether there are actually two different varieties, or whether the differences can be attributed to another factor such as age or sex.
COSEWIC had produced a Status Report (COSEWIC 2003) with two populations ('Eastern'; Québec and Labrador, and ‘Western’; Ontario westward and northward), but the reasons for separating those populations would not be relevant to present guidelines on DUs. The initial recognition by COSEWIC of the Eastern Population was based on its low population size and isolation from the Western population (Dauphiné 1989). The existing criteria for DU are discreteness and significance (Appendix F5 Operation and Procedures Manual), and the Eastern population, as presently delineated by population size, would not be recognized by COSEWIC.
Evidence for Discreteness
COSEWIC identifies discreteness, in part, by evidence of inherited traits (e.g., morphology, life history, behaviour) and/or neutral genetic markers (e.g.,allozymes, DNA microsatellites, DNA restriction fragment length polymorphisms (RFLPs), DNA sequences). Using mtDNA analyses on 183 specimens and nuclear DNA on 246 specimens from Ontario, Manitoba, Saskatchewan, Nunavut and NWT, Zigouris et al. 2012 recognized two main sub-populations in Canada. Wolverines from NWT, Nunavut and Saskatchewan are part of a large panmictic core population. Wolverines in the Manitoba/Ontario region were unique; the genetic composition of Wolverine (n = 40 mtDNA; n = 82 nuclear DNA) from the Manitoba/Ontario region was different from the western samples, potentially reflective of long-standing differences between regions (Zigouris et al. 2012). The Q values were relatively weak, however, indicating a mixing of haplotypes between the Manitoba/Ontario and remaining samples. Also, these analyses lacked samples from Quebec/Labrador and the southwestern US, which could influence the relative discreteness of the Manitoba/Ontario samples (Zigouris et al. 2012).
Additional mtDNA-based work on a larger, species-wide sample (n = 230 Eurasia; 148 Alaska; 188 contiguous US) identified 4 genetic clusters in Canada (Zigouris et al. 2013). The clusters, based mainly on haplotype frequencies, were in: 1) western Canada (BC, YK, AB, n = 161); 2) northern Canada (northern SK, NU, NWT, n = 151); 3) eastern Canada (ON, QC, Labrador, n = 67); and 4) Manitoba (n = 30). The Manitoba samples were distinct from all others. This expanded dataset provides more context for the relative amount of mtDNA control region genetic structure between ON, MB, NU, and NWT, where significant genetic structure was observed among other northern and western populations found to be nearly panmictic with nuclear DNA microsatellites.
Evidence for Significance
An additional criterion for DU is significance, which is represented as: 1) a gap in the population's range, or (2) the need for genetic differences reflecting deep divergence. The Eastern population is on the edge of the species’ range and its removal as a DU would not be a gap within the population. Deep divergence implies genetic differences are the result of local evolutionary selection, typically because of long periods of isolation from other populations, often as a result of glacial refugia. The difficulty in interpreting present haplotype distribution is whether it is the product of a single refugia (which would not support deep divergence) with subsequent population bottlenecks and genetic drift, or multiple refugia (which would support deep divergence, and multiple DUs). The issue is compounded further by relative lack of genetic samples from the eastern population. Finally, uncertainty exists because deep phylogentic breaks cannot be assessed using only a small portion of the control region of the mitochondrial genome that is not under strong natural selection (see Zigouris et al. 2012; 2013; McKelvey et al. 2013). Data from other mitochondrial genes or mitogenomes and nuclear DNA analyses may bring additional clarity; however, these analyses will continue to be limited by sample sizes of this low-density and highly vagile creature in eastern regions.
The cause of the genetic structuring between regions is unknown. One model posits that Wolverine were isolated in multiple glacial refugia followed by long-range dispersal events, which allowed re-population of vacant areas and eventual contact between different genotypes (Zigouris et al. 2012; McKelvey et al. 2013). Unique haplotypes found in peripheral populations led McKelvey et al. (2013) to conclude that a population in the United States (as far south as California) persisted in refugia south of the last glacial sheet (approx. 18000 YBP) and then were extirpated in the 1800s. The unique 'California' haplotype is closely associated with genotype from Eurasia, and the authors consider this as evidence of common ancestry associated with early invasion by Wolverine into North America. Based on haplotypes common to Wolverine found today in western Canada and the northwestern US, McKelvey concludes Wolverine then restocked the northwestern US from Canada in the early 1900s. The different genotype reflects their origins as a separate, isolated population that moved south after persisting in refugia in Alaska. In this model, the discreteness of Wolverine haplotype in the Manitoba/Ontario region may represent the boundary where two genotypes from separate glacial refugia presently meet.
A second model posits that there was a single refugium and any subsequent genetic differences are due to local breeding dynamics (Zigouris et al. 2013). In this model, the discrete haplotype distribution in Manitoba/Ontario is an artifact of a severely reduced population in the east and a concurrent loss of haplotypes. It is possible that some genetic types were eliminated by genetic drift and others increased in frequency relative to the north and west; the unique haplotypes could be remnants in a biased sample relative to other regions that retain larger, more genetically diverse populations (Kyle pers. comm. 2014). Female philopatry would also result in creation of some degree of local genotype.
It is unlikely that the discrete genotype in the Manitoba/Ontario region is an adaptation to local environment or habitats. Zigouris et al. (2012) notes that the Manitoba/Ontario region is dominated by extensive wetlands of the Hudson Plains Ecozone, whereas the core western population inhabits Taiga and Boreal Shield Ecozones (CFS 2013). There is some evidence of biogeographic influences on Wolverine genotype; differences in spring snow cover along dispersal routes were shown to explain genetic distance in Wolverines in the US Rocky Mountains (Schwartz et al. 2009). However, Wolverines are highly vagile and are found in many habitat types, suggesting local conditions would not be reflected in a species with extensive movement by males (see Dispersal and Migration section)
Four hypotheses explaining genotype distribution of Wolverines in North America were tested by Zigouris et al. (2013). None of the models were strongly supported but the strongest was the model wherein Wolverines arrived in North America in a single event from Siberia, then colonized the continent from west to east. The pattern would be consistent with that of a single refugium and subsequent expansion during glacial retreat. Also, the pattern of haplotype distribution in North America (Figure 2) is more likely the result of a single refugium model than a multiple refugia model; in a multiple refugia model, haplotypes would be clustered in several groups, rather than being distributed in a star radiation pattern typical of rapid expansion from a single source across most of the range, as indicated (Zigouris et al. 2013).
[doi:10.1371/journal.pone.0083837.g004], used with permission).
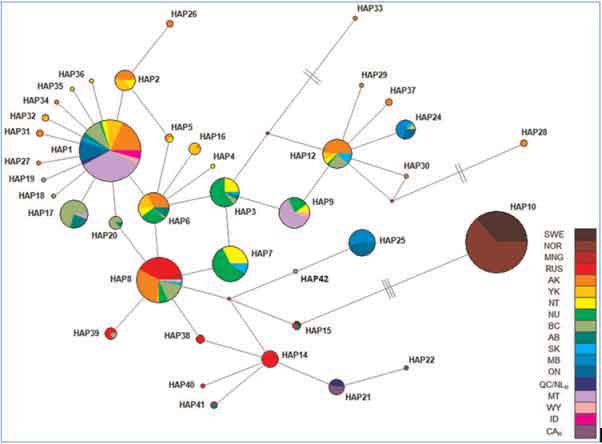
Figure 2: Network Map © Environment Canada
In summary, although the two populations in COSEWIC (2003) did not include genetic information, recent genetic analyses have established the presence of discrete genetic units. Essentially, there is a small fragment of mtDNA that evolves quickly and indicates female philopatry across the range, some evidence of two genetic groups, and slightly more differentiation of Manitoba/Ontario from the north and west. NuclearDNA shows near panmixia, with the exception of Manitoba/Ontario. The cause of the discrete genotype in the Manitoba/Ontario region is unknown.
The criteria of significance for a designatable unit has not been met. The Q values for discreteness are relatively weak. The discrete haplotypes in the Manitoba/Ontario region are more likely the product of a reduced population than an adaptation to local conditions. The mtDNA data largely represent the maternal genetic structure of Wolverines where clear breaks in frequency distributions of common haplotypes are found throughout their range, and, given the observation of near panmixia using neutral nuclear microsatellite loci for most of the range, the mtDNA results most likely are reflective of male-biased dispersal and female philopatry and not locally adapted subpopulations. Evidence to date suggests a single refugium model, and thus a lack of deep divergence. Also, a similar genotype distribution is apparent in other large Holarctic carnivores (i.e., Grizzly Bear, Polar Bear). Similar to Wolverine, these species possess high vagility in males and female philopatry and are considered a single DU across their Canadian range (COSEWIC 2008, 2012a). A single DU for Wolverine is recognized in this report.
Information is presented for the Canadian range, or when necessary, separated into western and eastern sub-populations. The eastern sub-population (Québec and Labrador) is isolated from the western sub-population and subject to different threats.
Special Significance
Wolverines are potential indicators of ecological integrity and disturbance due to biological characteristics and their dependence on large, connected and intact ecosystems (Carroll et al. 2001; GRRB 2001; Cluff and Paquet 2003; Cardinal 2004).Wolverines are viewed as one of several species of carnivores, which should be used in multi-species conservation planning in the Rocky Mountain region (Carroll et al. 2001) and northern Canada (Cluff and Paquet 2003). In Ontario, it is also suggested as an indicator for forest management practices and impacts from Far North development (Thompson pers. comm. 2013).
Wolverines are a much sought after and economically valuable furbearer (North Slave Metis Alliance (NSMA) 1999; WMAC(NS) and Aklavik HTC 2003; Cardinal 2004; Banci et al. 2005; ICC et al. 2006; GRRB 2014), and are trapped or hunted over much of their range in Canada (Table 1).The fur is frequently used for trim for garments due to its durability and frost-resistant characteristics (ICC et al. 2006).
| Season | ON | MB | SK | AB | BC | YT | NTFootnoteb.1 | NUFootnotec.1, Footnoted | Canada |
|---|---|---|---|---|---|---|---|---|---|
| 1992/93 | 4 | 48 | 2 | 44 | 236 | 176 | 93 | 34 | 637 |
| 1993/94 | 6 | 76 | 12 | 27 | 97 | 117 | 121 | 29 | 485 |
| 1994/95 | 8 | 52 | 11 | 23 | 186 | 145 | 119 | 15 | 559 |
| 1995/96 | 18 | 45 | 7 | 9 | 135 | 72 | 59 | 5 | 350 |
| 1996/97 | 14 | 46 | 14 | 27 | 225 | 161 | 86 | 26 | 599 |
| 1997/98 | 12 | 66 | 10 | 50 | 152 | 118 | 175 | 24 | 607 |
| 1998/99 | 4 | 33 | 4 | 40 | 123 | 104 | 62 | 15 | 385 |
| 1999/00 | 4 | 18 | 6 | 10 | 160 | 157 | 99 | 22 | 476 |
| 2000/01 | 7 | 53 | 23 | 37 | 162 | 188 | 56 | 19 | 545 |
| 2001/02 | 7 | 39 | 14 | 19 | 183 | 110 | 111 | 33 | 516 |
| 2002/03 | 8 | 39 | 29 | 35 | 120 | 131 | 106 | 29 | 497 |
| 2003/04 | 6 | 43 | 17 | 23 | 119 | 138 | 132 | 41 | 518 |
| 2004/05 | 11 | 47 | 18 | 29 | 161 | 104 | 118 | 24 | 512 |
| 2005/06 | 6 | 32 | 14 | 24 | 123 | 106 | 126 | 54 | 485 |
| 2006/07 | 2 | 24 | 18 | 43 | 120 | 106 | 154 | 30 | 497 |
| 2007/08 | 8 | 25 | 10 | 26 | 68 | 150 | 76 | 32 | 395 |
| 2008/09 | 7 | 52 | 18 | 26 | 123 | 142 | 133 | 39 | 540 |
| 2009/10 | 1 | 39 | 11 | 51 | 168 | 137 | 103 | 49 | 559 |
| Total | 133 | 777 | 243 | 543 | 2661 | 2362 | 1929 | 520 | 9162 |
Wolverines are important to the culture and mythology of numerous First Nations (i.e., Gwich’in stories on naming Nehtryuh (Wolverine); GRRB 2001, 2014). Wolverines often are given derogatory names, such as Devil Bear and Skunk Bear, because of their propensity to rob food caches and cabins and then mark the area with their musky scent (Fortin et al. 2005).They frequently consume animals in a trap, and trap-wise Wolverines can be difficult to catch, earning them a range of responses from respect to contempt from trappers (GRRB 2001; Thorpe et al. 2001; Cardinal 2004; Ray 2004; Paul F. Wilkinson and Associates 2009).They are rarely seen, especially in forested areas (Cardinal 2004).Legendary attributes, such as ferocity and cunning, are common in mythology and folklore surrounding Wolverines (Holbrow 1976; Cardinal 2004; ICC et al. 2006; Paul F. Wilkinson and Associates 2009).
Distribution
Global Range
Wolverines are found across the boreal regions of northern Eurasia and North America. The Eurasian range of the Wolverine includes Norway, Sweden, Finland, the Russian Federation, Estonia, Mongolia, and China. Range reductions have occurred in the European range south of 60°N.
Wolverine range in the conterminous United States has declined with human developments since the mid-19th century. They have been extirpated from the Great Lakes region, and from much of their range in the western mountains, where populations were naturally fragmented (Aubry et al. 2007). They ranged as far south as Colorado, Utah and California; however, small populations currently inhabit montane regions in Washington, Idaho, Montana and Wyoming (Figure 3; Aubry et al. 2007), with recent verified occurrences in Colorado (Inman et al. 2009), California (Moriarty et al. 2009), and Oregon (Magoun et al. 2013).
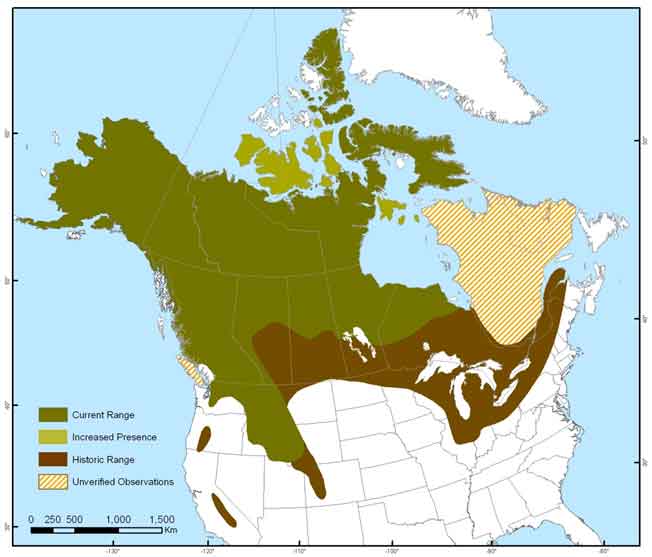
Map: North American distribution of Wolverine © Environment Canada.
Long description for Figure 3
North American distribution of Wolverine. Adapted from COSEWIC (2003), Magoun et al. (2004), Ray (2004, 2012), Aubry et al. (2007), Thibault unpubl. data (2013). Map produced by Bonnie Fournier, NWT. Increased presence refers to observations of Wolverine on various islands, but it is not known if these are established or vagrant individuals. Further details can be found in the preceding/next paragraph(s).
The historical range of Wolverines in North America was compiled from anecdotal evidence such as personal accounts and the interpretation of fur returns. The two major fur trading companies, Hudson’s Bay Company and the North West Company, traded over large areas (Novak et al. 1987; Obbard et al. 1987) and harvest locations cannot be confidently assigned to the community at which they were traded, or where they were trapped. Most of the southern areas from which Wolverines were presumably extirpated never produced significant numbers of Wolverine pelts (see Aubry et al. 2007). Human activities and habitat changes may have influenced the range contraction in the United States and Canada, but a northward shift in spring snow cover that persists through the reproductive denning period may be a critical factor as well (Copeland et al. 2010; see Habitat Trends section).
Canadian Range
Wolverine occur across much of the length of Canada, inhabiting forested areas, alpine tundra of the western mountains and arctic tundra (Figure 3). On the West Coast, Wolverines occur on Pitt (MacLeod 1950) and Princess Royale (Shardlow 2013) Islands, and were last confirmed on Vancouver Island in 1991 (BC Government 2013).Wolverines have not been reported on the island of Newfoundland, Nova Scotia, Prince Edward Island, Haida Gwaii, and some islands of the northwestern Arctic Archipelago in the NWT and Nunavut (Dauphiné 1989). Arctic islands supporting Wolverine include Victoria, Banks, Stefansson, Prince of Wales, Somerset, Devon, Cornwallis, Amund Ringnes, Ellesmere, Baffin, Bylot, Coates, Mansel (Community of Sachs Harbour et al. 2008; Environment Canada 2013; Carrière pers. comm. 2013), and it has recently been confirmed on Southampton Island (Awan pers. comm. 2013). Harvesters report a northwards expansion of wolverine (Cardinal 2004) with an increase in Wolverine on Victoria Island (Cardinal 2004; Banci et al. 2005), Baffin Island (Mallory et al. 2001), and Banks Island (Environment Canada 2013). Wolverine residency on some Arctic islands is unclear and is likely dependent on the availability of food resources.
Range reductions began in the mid-19th century, where it was extirpated from New Brunswick, and the southern half of Ontario and Québec, and from the aspen parkland of Manitoba, Saskatchewan and Alberta (Dauphiné 1989). In Labrador, range reduction began in the 1890s; between 1891-1910, 732 pelts were received at Fort Chimo and Davis Inlet, compared to 75 pelts between 1910-1930. The last verified record for Labrador was of two animals trapped in 1965 (Dagenais 1988). However, it is just as likely they persisted until the late 1970s because the last record for the entire eastern population was an animal trapped in 1978 in Schefferville, Québec, which is on the border with Labrador (Dagenais 1988; Fortin et al. 2005). Pelts were traded in Québec until 1980, but it is not known if they originated in Québec (Fortin et al. 2005).
Evidence from aerial surveys and trapper reports suggests a reoccupation of eastward range may be occurring in northern Ontario (Figure 4; Magoun et al. 2004; Ray et al. 2005; Koen et al. 2008; Ray 2012; Ontario Wolverine Recovery Team 2013; Ontario Ministry of Natural Resources, unpubl. data).
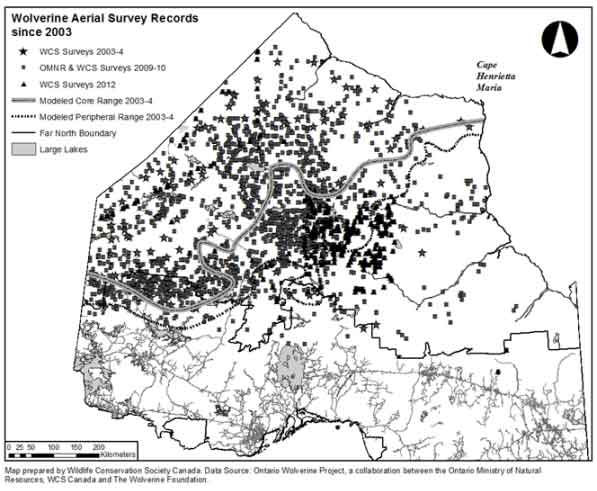
Map: Locations of Wolverine records © Environment Canada.
Long description for Figure 4
Locations of Wolverine records based on aerial surveys conducted by Wildlife Conservation Society (WCS) and Ontario Ministry of Natural Resources (OMNR). The area west of the modelled core and peripheral range represents the main and peripheral locations of Wolverine before 2004. The large number of recent (>2009) records indicates an eastern expansion of the sub-population. (Source: Ontario Wolverine Recovery Team 2013; Ray unpubl. data.)
Extent of Occurrence and Area of Occupancy
'Extent of occurrence' (EO) is defined as the area included in a convex polygon that encompasses the geographic distribution of all verified (scientific and ATK) occurrences of the Wolverine. The EO of the Western Population of the Wolverine in Canada is > 10 million km2. The EO of the eastern sub-population, based on recent, but unverified observations, would be 0.88 million km2.
'Index of area of occupancy' (IAO) is defined as the area within the EO that is occupied by Wolverines, excluding cases of vagrancy. It is calculated based on 2 km x 2 km grid cells that intersect the actual area occupied by Wolverines. This measure reflects the fact that the EO may contain some unsuitable or unoccupied habitats. The IAO of Wolverines in Canada is > 4,000 km2 for the western sub-population, and has not been determined for the unverified records in the eastern sub-population.
Search Effort
Wolverine distribution in Canada has been documented from several sources of information. Most data are derived from trapping activities. Registered trapping concessions or open trapping areas in BC, Alberta, and the Yukon correspond with area-specific harvest designation (Slough and Jessop 1996; Cardinal 2004; Jung et al. 2005; Arctic Borderlands Ecological Knowledge Coop Society (ABEKC) 2008; Slough 2009). In other jurisdictions, location of trap activity is recorded by nearest government office, trade centre, or through trapper surveys. ATK notes Wolverine are usually recorded during periods of snow cover but this likely relates to the period of trapping activities, which is biased to fall and winter. Most Wolverines are trapped incidentally (Cardinal 2004) and capture locations are rarely shared (ICC et al. 2006).
Techniques have been developed to determine the probability of Wolverine occurrence in a sample unit based on a hierarchical spatial modelling of Wolverine tracks in the snow (Schmelzer 2006; Magoun et al. 2007). Remote cameras have been used in many jurisdictions (Magoun et al. 2011a, b). Standard live-capture and radio-telemetry has been used in some area-specific studies to determine biological characteristics, such as home range size and movements, habitat use and density. Parks Canada and Department of National Defence maintain a database on Wolverine observations at their sites (Nantel pers. comm. 2012; Nernberg pers. comm. 2012).
Field studies to determine Wolverine population characteristics and/or density were conducted in the south-central Yukon from 1982 to 1985 (Banci 1987) and in the northern Yukon in 2004 (Golden et al. 2007). Wolverine DNA mark-recapture sampling has been undertaken in the diamond mining and exploration region northeast of Yellowknife since 2005 (Mulders et al. 2007; Boulanger and Mulders 2008, 2013a, b). Incidental observations are recorded at diamond mines in the NWT and Nunavut (NWT SARC 2013).
Diamond mining companies in the NWT (e.g., Diavik Diamond Mines Inc., BHP Billiton-Ekati, De Beers-Snap Lake) have Environmental Agreements and Wildlife Effects Monitoring Plans. The recording of observations and winter track-counts began at Snap Lake in 1999 (Golder Associates 2010). The track-count technique was modified in 2003 to use multiple, 4 km long transects. At Ekati, a regional DNA-based study replaced the Wolverine snow track survey that was conducted from 1997 to 2004 (Rescan 2012). Snow track surveys began at Diavik in 2003, and the methods were revised in 2008, using standard 4 km straight line transects (Diavik Diamond Mines Inc. (DDMI) 2012). Wolverine sightings have been documented at Kennady Lake since 1998, and winter track-counts began in 2004 (Golder 2010).
The survey effort in Nunavut is very similar to that in the NWT; Wolverines are monitored using fur harvest data, carcass collections, field studies, and ATK. Wolverine fur is prized in Nunavut, where most of the pelts are used locally.Wolverine densities are being monitored at diamond mining areas at High Lake (near Coronation Gulf) and Izok Lake (near the NWT border) using the DNA mark-recapture technique (Poole unpubl. data 2013). Both of these study areas are in the Southern Arctic ecozone.
British Columbia monitors Wolverines using harvest data and a compulsory reporting system for trappers and hunters, which receives low compliance (Krebs pers. comm. 2013). Two field studies of Wolverine ecology took place in the Omineca Mountains of north-central BC between 1995 and 2001, and the Columbia Mountains of the southeast (Lofroth and Krebs 2007).
Alberta has used opinion surveys of trappers (Petersen 1997) and fur harvest data analyses (Webb et al. 2013). Fisher et al. (2013) used genetic tagging to document Wolverine density in the Willmore Wilderness Park in the Rocky Mountains (2006 to 2008) and in the foothills near Hinton and Grande Cache (2005 to 2006). Genetic tagging has been used in Banff and Yoho National Parks since 2010 (Clevenger et al. 2011a). Camera trapping has been used in the central Rocky Mountains (Kananaskis Country) since 2010 (Fisher and Heim 2012) and in Banff and Yoho National Parks since 2009 (Clevenger et al. 2011b).
Wolverine distribution and abundance in Saskatchewan are monitored using fur harvest statistics (Siemens Worsley 2011), trapper questionnaire responses (Gollop 2012), and ATK (Berezanski 2004).
In 2000/01 a zero quota was placed on non-aboriginal trappers in Ontario; however, legal trapping continued until 2009 when the Wolverine season was closed. First Nation trappers may still harvest Wolverine via treaty rights but have not be able to sell the pelts since 2009. Most information on Wolverine distribution and abundance has come from fur harvest reports (incidental and Aboriginal harvest), ATK (Ray 2004) and aerial surveys (Ontario Wolverine Recovery Strategy). Recent aerial surveys have improved our knowledge of Wolverine range in Ontario (Figure 4) (Magoun et al. 2004, 2007; Ray et al. 2005; Koen et al. 2008; Ray 2012; Ontario Wolverine Recovery Team 2013). The 2005 Ontario Wolverine Project study area covered approximately 60,000 km2 in the Boreal Shield Ecozone in northern Ontario (Ray et al. 2005; Magoun et al. 2007; Bowman et al. 2010). Incidental Wolverine observations were recorded during a Caribou recruitment survey made across northern Ontario along a total transect length of 14,740 km in 2012 (Ray 2012). Knowledge of distribution is deemed robust, although incomplete along the range periphery (Ray pers. comm. 2013).
A study of Wolverine home range characteristics and ecology was conducted in lowland boreal forest near Red Lake, in northwestern Ontario in 2003-2004 (Dawson et al. 2010) along with a DNA mark-capture study, conducted in the same study area in 2004 by the Ontario Wolverine Project Team (Dawson et al. unpubl. data 2013). Data from these two studies have been used to estimate Wolverine density.
In Québec, Wolverine observations submitted since 2000 are numerous but unverified (Figure 5; Thibault unpubl. data 2013). An aerial survey for Wolverines in a 4,274 km2 area of the proposed Monts-Torngat-et-de-la-Rivière-Koroc National Park where Wolverine had once been present did not find evidence of the species (Fortin 2004). Another survey in the Abitibi and James Bay lowlands (100,000 km2) reported observations of two tracks in the area <100 km east of the Ontario border, but these have not been verified (Fortin 2006; Thibault unpubl. data 2013). A Wolverine camera trapping project initiated in the Abitibi lowlands and Laurentian Mountains in 2012 has not recorded Wolverine, to date (Thibault pers. comm. 2012).
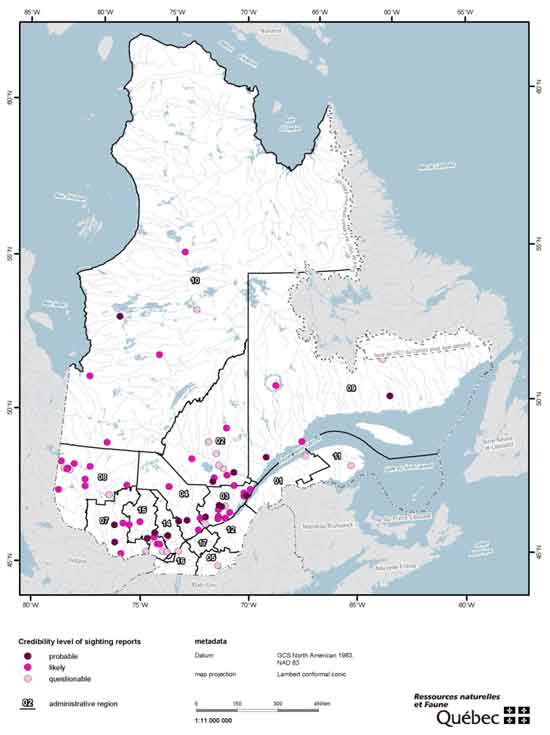
Map: Locations of Wolverine sighting reports in Québec © Environment Canada.
Long description for Figure 5
Locations of Wolverine sighting reports in Québec, 2000 to 2012, according to their relative level of credibility. No sightings have been verified. Adapted from Thibault unpubl. data (2013).
Johnson et al. (2012) assessed the Monts Groulx and Monts Otish in southeastern Québec for Wolverine denning habitat potential using GIS-based modelling of habitat variables, such as snow cover, summer snow persistence, the alpine-treeline ecotone, topographic ruggedness and distance from human activity. They concluded that the western massif of the Monts Groulx represented the highest quality reproductive habitat in sub-arctic Québec. A province-wide survey of Wolverine habitat potential in Québec also identified the Torngat Mountains, a large part of the Côte-Nord, the southeast section of the Nord-du-Québec, and part of the Charlevoix region as having potential to support Wolverines (Gallais and Messier 2012).
In Labrador, historical accounts are based on Hudson’s Bay Company fur trade data from posts in Davis Inlet and Moravian Missions of Hebron and Okak from 1884 to 1923 Schmelzer (unpubl. data 2012). The harvest of Wolverines was banned in Labrador (mainland Newfoundland and Labrador) in 1950. Aerial surveys, using 1,000-km2 hexagons, were conducted in 2005. The total transect length was 6,630 km, covering an area of 195,000 km2 between 54°N and 60°N and approximately 80% of Labrador and much of the range of the eastern population (Schmelzer 2006). Anecdotal observations continue to be reported; however, none have been verified by photographs or genetic analysis. A follow-up protocol, including search for hair samples, exists to attempt to verify observations (Moores pers. comm.). Inuit and Metis Aboriginal communities conduct ground detection monitoring using hair snag stations and remote cameras to try to verify reported sightings in the Cartwright area, and in northern Labrador (e.g., Habitat Stewardship Project by NunatuKavut Community Council; MacDonald pers. comm. 2013; Moores pers. comm.).
Anecdotal observations, including sightings of Wolverines and Wolverine tracks, are often ranked according to their reliability (e.g., Aubry and Houston 1992). A reliability ranking method is employed in the range of Eastern Wolverine Population in Québec (Thibault pers. comm. 2012).
Habitat
Habitat Requirements
Wolverines inhabit a variety of treed and treeless vegetation associations (ICC et al. 2006), at all elevations, although some Aboriginal harvesters suggest that they find more Wolverines in higher elevation areas (Wildlife Management Advisory Council (North Slope) (WMAC (NS)) and the Aklavik Hunters and Trappers Committee (HTC) 2003; Cardinal 2004; GRRB 2014) and fewer in open tundra (Cardinal 2004). They thrive in ecologically intact areas, where prey and other carnivore species are common and diverse. Wolverine habitat components include food (Cardinal 2004) and suitable physical features for natal and maternal den sites, and rendezvous sites, where kits are left during foraging periods by the female. In a multi-scale habitat use study in two multi-use regions of British Columbia, male Wolverines chose vegetation associations primarily based on food availability in summer and winter, while females were influenced by food, predation risk, and disturbance (Krebs et al. 2007).
Wolverine locations recorded by Mulders (2000) in the Southern Arctic ecozone in the NWT were correlated with vegetation density and patches, especially sedge density (Johnson et al. 2005). The reason for this association is unclear, but Wolverines were also associated with Wolves (Canis lupus), Caribou (Rangifer tarandus) and Grizzly Bears (Ursus arctos). Viable populations of large carnivores such as Grizzly Bears and Wolves are important as involuntary providers of ungulate carrion.
In mountainous areas, adult females used higher elevation alpine vegetation associations and steeper terrain more than males or other age classes (GRRB 2001; Cardinal 2004), while adult males and subadults of both sexes made extensive use of low elevation forested areas (Krebs et al. 2007). Lower elevations are used more in winter by males, and both sexes use all elevations in summer (Landa et al. 1998; Copeland et al. 2007; Krebs et al. 2007). Banci and Harestad (1990) found that Wolverines in the Kluane Game Sanctuary used vegetation associations according to availability, although males used coniferous subalpine areas more frequently than other vegetation associations in winter.
Den Sites
Wolverines have specific physical requirements for den sites. Dens are constructed either in talus boulders, along eskers, under deadfall, under logs in avalanche debris, or in snow tunnels at higher elevations and tundra (Copeland 1996; Magoun and Copeland 1998; GRRB 2001; WMAC and Aklavik HTC 2003; Cardinal 2004; Inman et al. 2007). Wolverine dens are also associated with large boulders and downed trees at lowland boreal sites in Ontario and Yukon (Dawson et al. 2010; Slough unpubl. data). In the eastern Arctic, Wolverines den in rocky areas with boulders, while in the western Arctic, knowledge holders describe finding dens in snow banks, under tree roots and along streams throughout the boreal region (GRRB 2001; Lutsël K’e Dene First Nation et al. 2001; WMAC(NS) and Aklavik HTC 2003; Cardinal 2004). It was reported that reproductive dens are often found in similar areas as dens used for other purposes (Cardinal 2004), although reproductive dens were usually less accessible and more isolated (GRRB 2001; ICC et al. 2006).
The availability of adequate insulating snow cover (i.e. ≥1.0 m deep) late into spring appears to be an important habitat feature for denning (Magoun and Copeland 1998). Copeland et al. (2010) found that 97.9% of 562 reproductive dens that they investigated in Fennoscandia and North America occurred at sites with persistent spring snow cover in at least 1 of 7 years. Female Wolverines must leave their kits for lengthy foraging trips, and often select natal den sites in talus or avalanche debris with snow cover late into spring (Copeland 1996; Inman et al. 2007) or under remnant snowdrifts (Magoun 1985). Such snow accumulation typically occurs in ravines and on leeward slopes. Dens with spring snow cover allow Wolverines to construct snow tunnels that provide thermoregulatory benefits for kits, are secure from excavation by predators, while dens are located in areas used by few other carnivores, and also provide an abundance of small-mammal prey for feeding kits (Magoun and Copeland 1998).
Individual Wolverines may reoccupy den sites or denning areas for several consecutive years (Magoun 1985; Inuit hunters, as reported by Lee and Niptanatiak 1996). Wolverine reliance on food caching in cold, structured microsites to inhibit competition with insects, bacteria, and other scavengers is likely a critical behavioural adaptation due to the limited availability of food resources. Inman et al. (2012) suggest that habitat features that facilitate caching, including low ambient temperatures, are likely important and related to the distributional limits of the Wolverine.
Habitat Trends
Considerable Wolverine habitat was permanently lost or fragmented with the extensive human development that occurred in the late 19th and 20th centuries at the southern edge of the range (van Zyll de Jong 1975). Much of the habitat lost may have been low quality habitat (as evidenced by low fur harvest returns), and numerical losses of Wolverines may have also been low. Losses were due primarily to human settlement and agriculture, with associated reductions due to hunting and trapping. Many Wolverines likely were non-target kills from poison baits used during predator control directed at wolves.
The reduction of ungulates, an important winter prey base, also likely contributed to their range contraction. Reduced numbers of prey remains a significant threat to Wolverine populations today, especially where Central and Southern Mountain, and southern Boreal Caribou herds are declining (Lofroth et al. 2007). Increases in Moose (Alces alces) and deer (Odocoileus spp.) populations in logged areas may, in part, compensate for losses in Caribou numbers. Wolverines may suffer from the recent declines of many Barren-ground Caribou herds (Vors and Boyce 2009). For example, the Bathurst herd declined from 203,800 to 16,400 breeding females from 1986 to 2009 (Boulanger et al. 2011). The Eastern Migratory George River herd of northern Québec and Labrador has declined from a high of > 600,000 in the 1980s to 27,600 animals in 2012 (Newfoundland and Labrador Department of Environment and Conservation 2012).
A positive correlation between Wolf and Wolverine harvests at trading posts in Québec and Labrador suggests that current-day Wolf population dynamics may be a useful tool to evaluate Wolverine recovery potential (Schmelzer unpubl. data 2012). However, Wolves in Labrador/Québec slowly recovered in the last century following the collapse and subsequent recovery of Eastern Migratory Caribou in the region, while Wolverines did not. The increase in Caribou that favoured the Wolf should also favour the Wolverine; reasons for the lack of a recovery by Wolverine in this region remain unknown.
Habitat fragmentation has resulted in isolated and threatened Wolverine populations in the western conterminous US (Banci 1994), and this process may be occurring in southern BC and Alberta, and in Manitoba and Ontario (Kyle and Strobeck 2002; Zigouris et al. 2012). Across parts of Wolverine range, forestry, oil and gas and mineral exploration and development, and large hydroelectric reservoirs threaten habitat with permanent, temporary and functional losses. Transportation corridors can act as barriers to movement and fragment habitats and isolate populations. Direct mortality may be a factor along transportation corridors in southern range, and elsewhere roads facilitate indirect mortality through improved motorized access for hunters, trappers and recreational users into remote areas. Travel and dispersal corridors of suitable vegetation associations must be maintained to facilitate connectivity of populations (e.g., Schwartz et al. 2009).
Kelsall (1981) and Dauphiné (1989) assumed that the relatively large number of parks and protected areas, which act as refugia from trapping and development in western Canada, had secured Wolverine populations. Trapping by Aboriginals, however, is allowed in most northern protected areas, and many Wolverines range outside protected area boundaries where they are vulnerable to trapping (Squires et al. 2007). Furthermore, human recreation, such as snowmobiling and other forms of snow travel allowed in some parks, may disturb Wolverines, particularly during the denning season in February-March (Heinemeyer and Squires 2012).
A study of the cumulative effects of developments on Arctic wildlife (Johnson et al. 2005) found that mines and other major developments had the largest negative effect on species occurrence, followed by exploration activities, and outfitter camps. The species most affected were Grizzly Bears and Wolves, followed by Barren-ground Caribou and Wolverines. The response by Wolverine was based on relocated Wolverine in the Slave lake area (Mulders 2000) which experienced a 2.4% decrease in use of summer habitat, likely because of a strong interaction between Wolverine and Caribou during the summer, when Caribou strongly avoided developments.
The extent of habitat loss from climate change is uncertain. Wolverine den in snowpack areas (see Den Sitessection) and spring snow cover during the denning period is closely related to historical Wolverine distribution in the contiguous United States (Aubry et al. 2007). Brodie and Post (2010a, 2010b) hypothesized that the declining snowpack in western Canada for the years 1974-2004 negatively affected Wolverine population dynamics, as evidenced by declining harvests. However, numerous biases could also explain declining harvests, such as trapping effort, declining pelt prices and licence sales, changes in trapping regulations, and trapper success, which raised questions about the validity of their conclusions (DeVink et al. 2010; McKelvey et al. 2010). A recent panel of experts generally agreed that snow is important at the scale of the den site, but did not agree if snow was limiting at the home-range, or species-range scales (Wolverine Science Panel 2014).
A climate-driven decline is not evident at this time in much of Wolverine range because Wolverine population trends in the northern range, while largely unknown, are possibly stable or increasing in many areas (COSEWIC 2003; Krebs et al. 2004; Lofroth and Krebs 2007; Slough 2007; see Abundance and Trends section). There is greater concern for southern areas of BC, and mountainous areas of the northern US (Weir pers. comm.; Wolverine Science Panel 2014); reductions in spring snow cover associated with climatic warming will likely reduce the extent of Wolverine habitat in the U.S. (McKelvey et al. 2011). Reductions in spring snow cover for lowland or mountain habitats in Canada have not been assessed specifically for Wolverine, but significant declines in sub-boreal and high altitude ecosystems are expected due to climate shifts (Wang et al. 2012).
Biology
Biological factors, such as a relatively low intrinsic rate of increase, low natural densities, and large home ranges limit population growth rates and affect the Wolverine’s ability to recolonize vacant habitats and recover from trapping (Banci and Proulx 1999). Re-population may take several decades but is possible in areas where factors (such as cessation of predator poisoning, trapping regulations, and ungulate population growth) favour Wolverine survival (Johnson 1990; Vangen et al. 2001).
Life Cycle and Reproduction
Wolverines have a polygamous mating system, with males mating with more than one female. Most Wolverines become sexually mature at 2 years of age, but few breed at this age (Banci 1994). Wolverines are thought to breed in the summer, from June through August (Magoun and Valkenburg 1983; Krott and Gardner 1985), and ATK suggests they breed during a few weeks in March and April during a period of active movement (GRRB 2001; Cardinal 2004; Deh Cho First Nation 2006). The proportion of adult female that were pregnant ranged from 74% (Banci and Harestad 1988) to 92% (Rausch and Pearson 1972) in studies in Yukon and Alaska. The pregnancy rate of subadults (1-2 years old) was 7% in the Yukon (Banci and Harestad 1988) and 50% for a combined Alaska/Yukon sample (Rausch and Pearson 1972). The latter study might have included some adults (Banci 1994). The pregnancy rate declined from 92% for 2-3 year olds to 53% for 5-6 year olds and 37% for combined 6 year old and older age classes. Recent (2005/06-2011/12) analyses of Wolverine carcasses from the Yukon showed a pregnancy rate of 0% for juveniles, 31% for yearlings, and 80% for adults. The highest pregnancy rates (90%) were in 3-5 year old animals (Jung and Kukka 2013).
The average number of fetuses ranged from 2.8 (2-3 year olds) to 3.4 (6+) in a 1980s Yukon study (Banci and Harestad 1988). Litter size was greatest for females over the age of 6 (Banci and Harestad 1988; Rauset 2013), but the pregnancy rate for that age class was lower. Overall reproductive rates observed in Alaska and Idaho were 0.69 and 0.89 kits per female per year, respectively (Magoun 1985; Copeland 1996); the low values reflect that females often breed every other year. Litter size was closely tied to primary productivity (Rauset 2013).
Active gestation takes 30-40 days (Rausch and Pearson 1972). Litters of four or less are born between January and April (Banci and Harestad 1988), or as late as June or July (GRRB 2001; Cardinal 2004). Wolverine kits reach adult body size by 7 months of age (Magoun 1985) and are weaned at 9-10 weeks (Banci 1994).
In this report, generation time is based on the average age of reproductive females in the population. The maximum age reported for Wolverines is 16 years in the Inuvik Region, NWT (Branigan and Pongracz unpubl. data 2012). Maximum ages of 11 were reported in the Kitikmeot Region, Nunavut (Lee 1998), and 12.9 in the Yukon (Jung and Kukka 2013). Most females do not breed until the age of 3, and the average age of females that are 3 or older is likely 7 or 8 years of age. The generation time for Wolverines is likely 7.5 years.
Physiology and Adaptability
Wolverines typically occupy home ranges that vary from about 50-400 km2 for females, and 230-1580 km2 for males (Hornocker and Hash 1981; Gardner 1985; Magoun 1985; Whitman et al. 1986; Banci and Harestad 1990; Copeland 1996; Krebs et al. 2007; Dawson et al. 2010; Persson et al. 2010). Lofroth (2001) documented an average home range of 1,230 km2 for subadult females, and a range of 3,500 km2 for dispersing subadult males. WMAC(NS) and Aklavik HTC (2003) report that fresh tracks of solitary Wolverine are observed in about every 40-80 km of snowmobiling in April and May. There may be home range overlap between members of the same and opposite sexes, with the latter being more common (Krebs and Lewis 2000). Cardinal (2004) reported that male home ranges would include the ranges of 4-5 females. Aboriginal and Inuit harvesters indicate that wolverine move extremely large distances (NSMA 1999; Cardinal 2004; ICC et al. 2006; Henry 2007) and that they are unclear if they have a home range or territory because they never seem to stay in the same area (Cardinal 2004). Some knowledge holders believe that older Wolverines establish home ranges, but younger individuals do not (Cardinal 2004; Henry 2007).
Wolverines show some adaptability to humans, being attracted to wilderness camps, traplines, and snowmobile trails by virtue of their scavenging habits. There is also a negative impact of disturbance, leading to the loss of functional habitat, possibly affecting the reproductive success of females (see Threats and Limiting Factors section).
Dispersal and Migration
A proportion of the population, typically yearlings, is transient at any given time. Yearling females tend to establish home ranges nearer their natal ranges than do yearling males, although both sexes are capable of long-distance movements. Male dispersal records include 200-874 km over several months (Gardner et al. 1986; Inman et al. 2004). A dispersal distance of 100 km was reported for a juvenile male in Ontario (Dawson et al. unpubl. data 2013). Maximum movements by females of 225-300 km have been recorded (Mulders 2000; Aubry et al. 2012).
Wolverines are able to traverse rugged terrain, including tundra and glaciers that would act as barriers to the dispersal of many other species of mammals. Long-distance dispersal abilities likely gave Wolverines the capacity to recolonize gaps in their distribution in Scandinavia (Vangen et al. 2001; Flagstad et al. 2004).
Interspecific Interactions
Wolverines are scavengers and predators, opportunistically feeding on abundant or readily procurable food (Cardinal 2004; Clément 2009; Benson 2011; Inman et al. 2012). Food is frequently cached for later use. Diets of Wolverine typically vary between seasons and years, as they switch between food sources depending on availability (Dalerum et al. 2009). Fresh prey are eaten more during summer and carrion, including cached items, is used more in winter (Magoun 1987). Many species of small mammal and ground-nesting bird are depredated. The most common sources of carrion are whichever large ungulate species are present (Gardner 1985; Banci 1987; Magoun 1987; Johnson and Ruttan 1993; NSMA 1999; Mulders 2000; GRRB 2001; Thorpe et al. 2001; Cardinal 2004; Ray 2004; Banci et al. 2005; Dumond 2007; Henry 2007; Lofroth et al. 2007; Clément 2009; Carriere 2010; Benson 2011).
In the Kitikmeot Region, Nunavut, Mulders (2000) reported that Caribou remains were found in 62% of stomachs of winter-trapped Wolverines, and Muskox were found in 11%. Minor food items included Arctic Ground Squirrels (Spermophilus parryii), voles and lemmings, and ptarmigan. Caribou were also the most common prey in the Inuvik Region, NWT (Branigan and Pongracz unpubl. data 2012), where they were found in 58% of Wolverine stomachs. Barren-ground Caribou were found in 56% of all stomachs in the North Slave Region, NWT (Mulders unpubl. data 2013), but were found in 12 to 32% of Wolverine stomachs from other regions. Snowshoe Hare (Lepus americanus) (and presumably Arctic Hare (L. arcticus) in tundra regions) is an important food for Wolverine in the north (Banci 1987; Branigan and Pongracz unpubl. data 2012) and a delayed numerical response of a 2-3 year time lag in Hare abundance has been documented (Bulmer 1975). Arctic Ground Squirrels are an important summer food in the Arctic (Magoun 1987). Several fish and seal species are scavenged (Golder Associates 2003; Cardinal 2004; Banci et al. 2005; Clement 2009). Porcupine (Erethizon dorsatum) are noted as a common food item in the Gwich’in Settlement region of the Mackenzie Delta (GRRB 2001, 2014).
Large carnivores generate carrion, which is an important food source for Wolverines, especially during the winter months (Wright and Ernst 2004). Carnivores also may compete with Wolverines at kill sites, and are a potential source of Wolverine mortality (White et al. 2002; Krebs et al. 2004; Jung and Kukka 2013). Johnson et al. (2005) found that Wolverines were positively associated with Wolves, Grizzly Bears and Caribou on the Slave study area, near Daring Lake, NWT, although a traditional knowledge study in the area of Wood Buffalo National Park did not report any cases of Wolverine preying on Caribou (Gunn 2009).
Wolverines are preyed on by Grizzly Bear, Polar Bear (Ursus maritimus), Wolves, Mountain Lions (Puma concolor), Golden Eagles (Aquila chrysaetos), and other Wolverines (Cardinal 2004). They may encounter large carnivores with greater frequency when scavenging carrion. Krebs et al. (2004) found that human-caused mortality from trapping and road/rail kill accounted for 46% of deaths in a review of 12 studies using radio-collared animals. Natural sources of mortality included predation by Wolves, Mountain Lions and conspecifics (30% of non-human causes), and starvation (49% of non-human causes).
Parasitic nematode roundworms (Trichinella spp.) were found in 88% of Wolverine samples tested from Nunavut (Reichard et al. 2008a) and 26% (n=43) of samples from the lower MacKenzie region (Larter pers. comm), and none (n=38) from the upper Mackenzie region (Addison and Boles 1978). Six other species of helminth parasites (trematodes, cestodes and nematodes) were recorded in the digestive tracts of Wolverine from the lower Mackenzie River valley (Addison and Boles 1978). Protozoan parasites (Sacrosystis spp.) infected 80% of Wolverines from Nunavut (Dubey et al. 2010). Another protozoan parasite, Toxoplasma gondii, was detected in 41% of Wolverines from Nunavut (Reichard et al. 2008b). The prevalence of clinical disease symptoms is unknown, but this parasite is transmissible to humans through skinning and fur handling. A rabid Wolverine (infected with a strain of Arctic Fox (Alopex lagopus) rabies) was recorded in Alaska (Beckman pers. comm. 2012). Rabies was not recorded in 71 Wolverine tested for rabies in the Yukon during recent years (Jung pers. comm. 2013).
Population Sizes and Trends
Sampling Effort and Methods
Establishing population abundance and trends is difficult because most numeric data are derived from harvest activity. It is unknown in large parts of the range whether harvest actually reflects total harvest levels. Fur harvest is susceptible to many factors other than the population size of furbearers, including environmental, social, cultural and economic factors (Slough and Jessup 1996). Trends are difficult to establish if trap effort and reporting varies over time and by jurisdiction. For example, Lee (1994a) believed that the harvest was underestimated by 50-90% in Nunavut/NWT. Some reports suggest that <20% of the harvest enters the fur trade (Government of Nunavut 2007; Spencer unpubl. data 2012). Based on carcass collection programs, the unreported harvest may be underestimated by 35% overall, and greater than 80% in Inuvialuit/Inuit coastal communities (Branigan and Pongracz unpubl. data 2012; Rossouw unpubl. data 2012; Mulders unpubl. data 2013; summarized in NWT SARC 2013). Lee (1994a) estimated that the actual harvest was underestimated by 50-90% in the Coppermine, Bay Chimo, and Bathurst Inlet area. (Note: although reporting is low, extra information on harvest is obtained with community-based carcass collections using cash incentives and may give an improved estimation of total area-specific Wolverine harvest (Lee 1994a, 1994b, 1998; Elliot and Dumond 2005; Branigan and Pongracz unpubl. data 2012; Mulders unpubl. data 2013)).
Although it is common to infer trend from harvest statistics, it may be problematic to base trend data upon the premise that current harvest is sustainable. In BC, for example, application of such a population model would have calculated population estimates in the 1970s near 10,000 Wolverine, rather than the 3,500 actually present (Lofroth pers. comm. 2012).
Provinces and territories which allow trapping and hunting of Wolverines maintain harvest records, but laws regarding the reporting of harvests vary greatly among jurisdictions. In the best cases, all harvested Wolverines are accounted for by one or more methods--mandatory pelt sealing, compulsory reporting, fur export permits or fur dealer records--and the harvest location of each animal is recorded. Jurisdictions using this type of harvest monitoring are BC, Alberta and Yukon.
Other jurisdictions monitor harvest using fur dealer records and export permits, which only capture furs exported to fur auctions. Depending on the local culture and traditions of the harvesters, varying proportions of the harvested pelts are retained for personal use, or for use in locally produced handicrafts. For example, much of the Inuvialuit/Inuit harvest of Wolverines across the Arctic (Yukon, NWT and Nunavut) is not sold at auction (Branigan and Pongracz unpubl. data 2012; Rossouw unpubl. data 2012; Mulders unpubl. data 2013).
Trapper questionnaire responses are sometimes converted to numerical values and compiled as Wolverine abundance and trend indices to provide a way to compare trappers’ interpretations of species abundance and trends in a given area over time (Slough 2009). Some of the data limitations originate from the lack of standardization of trappers’ perceptions, and animal behaviour patterns independent of population densities which affect perceptions of abundance (Jung et al. 2005). Verification with other data such as fur harvest and population surveys is recommended.
Abundance and Trends
ATK suggests that Wolverine are widespread, but not abundant (Cardinal 2004). The Gwich’in considered Wolverine to be scarce, both today, and in the past, and Wolverine do not appear to have the same population cycles seen in other furbearers (GRRB 2014). Most information from non-ATK sources is based on harvest data from within the last 200 years. The earliest Wolverine harvest data attributed to specific Canadian jurisdictions indicate that populations, based on harvest success, may have been declining in the Prairie Provinces in the 1920s and 1930s (Novak et al. 1987). Ontario, Québec and Labrador harvests were already low by that time. Wolverine harvests in northern Québec and Labrador declined in the 1914-1923 period (Schmelzer unpubl. data 2012). Wolverines had been extirpated from New Brunswick by the early 1800s.
Wolverine densities vary across ecological areas, to a maximum of about 10 Wolverines/1,000 km2 in North America (Table 2). The highest densities occur in Northern Mountain, northern Boreal and eastern Southern Mountain ecological areas and in the Southern Arctic Ecozone (5 to 10/1,000 km2), where vegetation associations, prey species, ungulates, and large carnivores are most diverse and abundant. Moderate densities occur in the southern Boreal ecological area (1 to 3/1,000 km2). There have been no field studies in the Pacific and western Southern Mountain ecological areas and Northern Arctic Ecozone, where densities are presumed to be low.
| Density (per 1,000 km2) | Location | Study Area (km2) | Relative Habitat Quality | National Ecological Area or Ecozone | Methods | Reference |
|---|---|---|---|---|---|---|
| 6.5 (SE = 1.9) | Omineca Mountains, northern BC | 8,900 km2 | High | Northern and Southern Mountain | Mark-recapture and mark-resight (cameras) | Lofroth and Krebs 2007 |
| 5.8 (SE = 1.1) | Columbia Mountains, southern BC | 7,000 km2 | High | Southern Mountain | Mark-recapture and mark-resight (cameras) | Lofroth and Krebs 2007 |
| 0.3 - 2.0 - 4.1 | BC | na | Rare, Low and Moderate | Pacific, Boreal, and Southern Mountain (plateau regions) | Predictions based on habitat quality ratings | Lofroth and Krebs 2007 |
| 4.8 | Northeast BC | 51,200 | Moderate | Boreal | Trapper catch and snow tracking | Quick 1953 |
| 6.8 | Willmore Wilderness Park, AB | 4,600 | High | Southern Mountain | DNA mark-recapture (NGT, non-invasive genetic tagging) | Fisher et al. 2009 |
| 3.0 | Foothills, AB | 6,400 | Moderate | Boreal | DNA mark-recapture (NGT, non-invasive genetic tagging) | Fisher et al. 2009 |
| 5.6 | Kluane Wildlife Sanctuary, YT | 1,800 | High | Northern Mountain | Based on known residents only, identified by live capture and telemetry | Banci and Harestad 1990 |
| 10.8 | Kluane Wildlife Sanctuary, YT | 1,800 | High | Northern Mountain | Calculated from mean home range size and assuming saturated habitat | Banci and Harestad 1990 |
| 9.7 (CV 6.5%) Footnoteb.2 | Old Crow Flats, YT | 3,375 | High | Northern Mountain | Quadrat sampling of tracks in snow using sample-unit probability estimator (SUPE) | Golden et al. 2007 |
| Declining 10.73 (CV 10.9%) to 3.72 (CV 15.4%) | Daring Lake, NT | 2,556 | Moderate to high | Southern Arctic Ecozone | DNA mark-recapture | Mulders et al. 2013b |
| Declining 11.43 (CV 18%) to 3.87 (CV16%) | Diavik, NT | 1,269 | High | Southern Arctic Ecozone | DNA mark-recapture | Boulanger and Mulders 2013a |
| Declining 10.05 (CV 19%) to 6.14 (CV 15 %) | Ekati, NT | 1,062 to 1,647 | High | Southern Arctic Ecozone | DNA mark-recapture | Boulanger and Mulders 2013a |
| 4.6-5.2 (SE=1.3 F-2.4 M) Footnotea.2 | Kennady Lake, NT | 1,575 | Moderate to High | Southern Arctic Ecozone | DNA mark-recapture | Boulanger and Mulders 2008 |
| 6.85 (SE=1.05) 3.5 F, 3.3 M |
High Lake, NU | 3,000 | High | Southern Arctic Ecozone | DNA mark-recapture | Poole unpubl. data 2013 |
| 4.80 (SE=0.8) 1.5 F 3.3 M |
Izok Lake, NU | 3,000 | Moderate to High | Southern Arctic Ecozone | DNA mark-recapture | Poole unpubl. data 2013 |
| 0.7-1.4 | Red Lake, ON | 7,626 | Low to Moderate | Boreal | DNA mark-recapture and mean home range size | Dawson, Magoun, Ray and Bowman unpubl. data 2004 and 2013 |
| USA | - | - | - | - | - | - |
| 9.7 (5.9–15.0) | Southeast Alaska (AK) | 2,140 | High | Pacific | Camera trapping and spatial capture-recapture model | Royle et al. 2011 |
| 3.0 (CV 12.0%) Footnoteb.2 | South-central AK | 4,340 | Moderate | Maritime montane (N. Kenai Mtns) | SUPE | Golden et al. 2007 |
| 5.2 (CV 20.3%) Footnoteb.2 | South-central AK | 1,871 | High | Maritime montane (N. Chugach Mtns) | Transect intersect probability sampling (TIPS) | Becker 1991 |
| 4.7 (CV 13.0%) Footnoteb.2 | South-central AK | Moderate | Montane (E. Talkeetna Mtns) | TIPS | Becker and Gardner 1992 | |
| 4..0 – 7.4 | South-central AK | Moderate to high | Montane (Susitna River Basin) | Home range/telemetry | Gardner and Ballard 1982 | |
| 4.9 (CV 8.9%) Footnoteb.2 | South-central AK | 3,663 | Moderate | Maritime montane (W. Chugach Mtns) | SUPE | Becker and Golden 2008 |
| 4.62 (no variance est.) | South-central AK | 1,050 | Moderate | Maritime montane (W. Chugach and N. Kenai Mtns) | Total count of small study area | Golden 2010 |
| 5.0 (CV 17.1%) Footnoteb.2 | South-central AK | 1,939 | Moderate to high | Maritime montane (N. Kenai Mtns) | SUPE | Golden unpubl. data 2013 |
| 7.2 | Arctic AK | ~5,000 | High | Arctic, western Brooks Range | Home range/telemetry | Magoun 1985 |
| 20.8 (fall estimate) | Arctic AK | 2,400 | High | Arctic, western Brooks Range | Home range/telemetry | Magoun 1985 |
| 4.0-11.1 | Idaho | 8,000 | Moderate to High | Montane | Live capture, telemetry and reproductive potential | Copeland 1996 |
| 15.4 Footnotec.2 | Montana | 1,300 | High | Montane | Capture and snow tracking | Hornocker and Hash 1981 |
The following discussion of Wolverine population status is organized by jurisdiction, where sources of information from management programs and research projects vary greatly. Population estimates are provided but their accuracy may vary by jurisdiction.
Yukon Territory
All Wolverine pelts taken by Yukon trappers and hunters must be sealed, including pelts held over or used in the territory. Available evidence points to a stable population in all regions of the territory and a full recovery from the large-scale Wolf control programs (poisoning through the 1970s and aerial shooting through the 1990s).Trappers and/or Aboriginal knowledge holders report Wolverines to be generally common (Cardinal 2004; Henry 2004; ABEK 2008), and harvests are not declining relative to trapping pressure (Table 1). WMAC (NS) (2008) reported a harvest of about 10 Wolverines per year by Inuvialuit trappers from Aklavik, NWT.
Banci and Harestad (1990) estimated Wolverine density to be 10.75/1,000 km2 in continuous and saturated habitat in the south-central Yukon.Golden et al. (2007) estimated a population density of 9.7 Wolverines/1,000 km2 on the Old Crow Flats, Yukon. The estimate was 5.65/1,000 km2 when using known Wolverines (assuming variable habitat quality and not all habitat is saturated).They estimated the resident population for about 85% of the territory to be 2,503 Wolverines, and a fall population of 4,171, including juveniles and transients (Banci 1987).The estimate was based on habitat suitability estimates (area trapped, trapper-years and numbers of Wolverines harvested), which likely underestimated Wolverines in remote areas where trapping effort is less than near communities. A more realistic Wolverine population estimate for the Yukon, based on 100% coverage of the territory and more homogeneous densities in high quality habitats, is 3,500 to 4,000 residents.
Northwest Territories
Wolverines are described as widespread, but not numerous in the NWT (WMAC(NS) and Aklavik HTC 2003; Community of Inuvik et al. 2008; Community of Paulatuk et al. 2008; Community of Tuktoyaktuk et al. 2008). Large numbers of Barren-ground Caribou and Wolves in the taiga and tundra associations indicated that Wolverines may be as common as they are elsewhere (Cardinal 2004; Mulders et al. 2007).
Wolverine densities declined on three study areas in the NWT between 2004-2005 and 2011 (Boulanger and Mulders 2013a, b). At Daring Lake, the density declined 35% (10.73/1,000 km2to 3.72/1000 km2, between 2004 and 2011) (Boulanger and Mulders 2013b). At Diavik, the density declined 34% (11.43 to 3.87/1,000 km2, between 2005 and 2011), and at Ekati, the density declined 61% (10.05 to 6.14/1,000 km2 over the same period) (Boulanger and Mulders 2013a). Wolverine densities were stable at Kennady Lake between 2005 and 2006 where there were 4.6 to 5.2/1,000 km2 (Boulanger and Mulders 2008). Wolverine declines were concurrent with declines in the Bathurst Caribou herd (Boulanger et al. 2011; Mulders pers. comm. 2013). These densities, even at their reduced levels, are in the range of moderate to high densities relative to other areas where Wolverines have been studied in North America (Table 2).
Coastal communities exported fewer Wolverine pelts than other communities, with Paulatuk exporting none from 87 carcasses, and Tuktoyuktuk exporting 6.6% from 136 carcasses between 2004/05 and 2011/12 (Branigan and Pongracz 2012; Rossouw unpubl. data 2012; data summarized in NWT SARC 2013). The Inuvik regional average pelt export rate was 19.2%. The carcass collections from other regions (Dehcho, Sahtu, South Slave and North Slave) rarely exceeded reported harvests annually between 2001/02 and 2011/12, and the total pelt exports exceeded the carcass collections over that period (Rossouw unpubl. data 2012; Mulders unpubl. data 2013). An unknown proportion of furs from these regions may be retained for personal use and do not enter the fur trade. Another source of unreported Wolverine harvest occurs in the Rennie Lake, NWT area by trappers from Saskatchewan (Mulders unpubl. data 2013). Between 2000/01 and 2008/09, an annual average of 38.7 pelts, representing 25.6% of the combined harvest, was recorded on export permits by Saskatchewan Conservation Officers.
The mean annual harvest over 20 years in the NWT is 109, and long-term harvest data are stable (Table 1, 2). The mean annual harvest estimate is approximately 200 Wolverines (Note: data corrected for furs not entering the fur trade in the Inuvik and other regions, and in the Saskatchewan/Alberta border region (NWT SARC 2013). The unreported harvest from other regions is difficult to estimate from available data and so was not used to augment the harvest estimate further. At the NWT population estimate of 5,100, the harvest rate is well below (< 3.9%) the intrinsic rate of growth of 6.4% per year in untrapped populations (sustainable if refugia and trapped areas are equal in size; Krebs et al. 2004). Cardinal (2004) identified several refugia from trapping in the NWT.
According to ATK holders, Wolverine population trends are variable across space and time but there is no clear indication of decline. From 2002-2004 studies, the majority of knowledge holders across the Northwest Territories reported Wolverine as stable or increasing (Autsӱl K’e Dene First Nation 2002; Nagy et al. 2002; WMAC and Aklavik HTC 2003; ABEKC 2004; Cardinal 2004), with the possibility of a decline near Yellowknife in 2004 (Cardinal 2004). In 2005, Inuvialuit hunters from Aklavik, Inuvik and Tuktoyaktuk indicated that Wolverines were more difficult to find (ICC et al. 2006), but in 2006-2007, Wolverine in Aklavik were said to be more abundant (ABEK 2007). More recent ATK is unavailable. There is evidence from harvest patterns and local knowledge that Wolverines are increasing in numbers on Victoria Island (Cardinal 2004; Banci et al. 2005), and Banks Island (Environment Canada 2013).
The Wolverine population of the NWT in 2003 was estimated at 3,500 to 4,000 residents, based on the range of statistical and non-statistical density estimates, and expert opinion, from other North American studies, and from a crude assessment of habitat quality (COSEWIC 2003; Slough 2007). More recent data on Wolverine densities suggests that an estimated 3,430-7,325 Wolverines may be present in the NWT (NWT SARC 2013). An additional 220-470 juveniles, many being transients, are estimated to be present in the fall (pre-trapping) population (based on an annual growth rate of 6.4%; Krebs et al. 2004).
Nunavut
The fur trade statistics for Nunavut are based on furs exported to fur auction (Table 1), which is an underestimate of total harvest (Cardinal 2004).Wolverine harvest has also been monitored with the Nunavut Wildlife Harvest Study (Nunavut Wildlife Management Board (NWMB) 2004).
Wolverine densities are moderate in the west and low on the arctic islands and the northeast and eastern mainland, where numbers are believed to be stable or increasing (Awan pers. comm. 2012). Between 2002-2004, Wolverine populations in the Kitikmeot region of Nunavut were reported to be fairly stable and robust with a slight cyclic pattern related to local food availability (Cardinal 2004; Banci et al. 2005); comparatively, this region also reported some of the highest levels of harvest in Canada (Cardinal 2004; NWMB 2004; ICC et al. 2006). Similar comments were provided for Herschel Island, the Yukon North Slope (WMAC and Aklavik HTC 2003), North Slave Region, NWT (Cardinal 2004; Henry 2004), the Kakinyne region (Autsӱl K’e Dene First Nation 2002) and around Old Crow, Yukon (Cardinal 2004; Henry 2004) during the same time period. Comments provided to the Canadian Wildlife Service during 2005 community meetings on the SARA listing of Wolverine and the Government of Nunavut management planning process described an increasing Wolverine population (Han pers. comm. 2012). Communities involved included Gjoa Haven (King William Island), Kugaaruk, Cambridge Bay (Victoria Island), Arviat and Rankin Inlet. Awan et al. (2012) reported opinions of 65 Nunavut hunters; 58% believed that the Wolverine population in their area was increasing, 40% believed that it was stable, and 2% believed it was decreasing.
Wolverine densities at High Lake (near Coronation Gulf) and Izok Lake (near the NWT border), were estimated at 6.85/1,000 km2 and 4.80/1,000 km2, respectively, in 2008 (Poole unpubl. data 2013). Both of these study areas were 3,000 km2 and are in the Southern Arctic Ecozones.
The Nunavut Wildlife Harvest Study reported annual harvests from Nunavut communities in Wolverine range were < 5-20 Wolverines per year, except in Kugluktuk, where the harvest was estimated at 80-100 per year (NWMB 2004). In the Nunavut carcass collection program, the average annual reported harvest in Kugluktuk from 1986-2012 was 63 Wolverines.Wolverines were harvested (2009/10 to 2011/12) near most Nunavut communities including Coral Harbour (Southampton Island), Gjoa Haven, Cambridge Bay, and other locations on Victoria Island. No harvest was reported from Baffin Island, although Wolverine tracks were reported near Clyde River on northeastern Baffin Island in the winter of 2011/12 (Awan pers. comm. 2012). The only Wolverine harvest reported from the Qikiqtaaluk (Baffin) Region was from Hall Beach, Melville Peninsula, and Igloolik. The local overharvest of Wolverines may take place near communities and concentrations of Barren-ground and Dolphin and Union Caribou, depending on re-population from neighbouring refugia.
British Columbia
Lofroth and Krebs (2007) used a habitat quality rating system empirically derived from estimates of Wolverine density to predict Wolverine distribution and abundance at a provincial scale. Density and habitat association data were obtained from two study areas, the Columbia Mountains (7,000 km2, 1994-2002) and Omineca Mountains (8,900 km2, 1995-2001), where densities were estimated at 5.8 and 6.5/1000 km2, respectively. Wolverine numbers for the province were estimated at 3,530 (95% C.I. 2,700–4,760) (Lofroth and Krebs 2007). The lack of density data for low and rare areas could introduce error in the overall estimate. Mean densities were estimated at 0.3/1,000 km2 in rare-quality habitat, 4.1/1,000 km2 in moderate-quality, and 6.2 Wolverines/1,000 km2 in high-quality habitat. These estimates are consistent with 4.8/1,000 km2 reported by Quick (1953) for moderate quality habitat in northeast BC. The Shuswap Indian Band noted a decrease in Wolverine abundance in the Secwepemc Traditional Territory, which they attributed to overhunting and loss of important migration routes (Shuswap Indian Band 2008).
The Vancouver Island population is believed to be very low, or extirpated. Unverified sightings are reported almost annually to the British Columbia Ministry of Environment (Lofroth pers. comm. 2012). The last confirmed record was in 1992 (BC Government 2013) and it is possible that they have been extirpated.
Alberta
A population estimate for Wolverine in Alberta is unavailable. It is likely that the population was low in the 1950s during rabies control programs when non-selective poisoning killed 5,500 Wolves and an unreported number of Wolverine (Petersen 1997).Wolf control using aerial shooting and poisoning is ongoing in the west-central Alberta range of the Little Smoky Boreal Caribou herd (Campbell 2012), where at least 650 Wolves have been killed since 2005. A quota of one Wolverine harvested per trapline has been in place since 1989.
Opinion surveys of trappers in Alberta suggested a stable population existed north of 56°N latitude but was declining elsewhere in 1987, and in 1994, there was a province-wide declining population (Petersen 1997). An analysis of Wolverine harvest data from 1985-2011 has shown that the number of traplines harvesting Wolverines, and the average Wolverine harvest by trapline have increased since the early 1990s (Webb et al. 2013). Harvests increased significantly in the Northwest Boreal and Foothills natural regions, but declined in the Canadian Shield and Rocky Mountains regions. It is not known whether the increased or decreased harvest is due to changes in Wolverine populations, trapper effort, or other factors.
During 2005-2008, Fisher et al. (2013) used genetic tagging to conclude Wolverine densities of 6.8/1,000 km2 in the Willmore Wilderness Park, Rocky Mountains and 3/1,000 km2 in a foothills study area near Hinton and Grande Cache. Based on camera trap data, Fisher and Heim (2012) believe that the pattern of abundance is similar farther south in the montane Banff National Park, as compared with Alberta’s East Slopes. Northern boreal habitats have not been studied.
Saskatchewan
Harvests are low in the southern boreal forest (218,612 km2) and higher in the north (132,958 km2) (Siemens Worsley 2011), therefore Wolverine abundance is believed to follow the same pattern. The population is crudely estimated as <1,000, based on range and density assumptions. Trapper questionnaire responses from the South Saskatchewan Trapping Area and the Northern Fur Conservation Area reported that Wolverine were viewed as ‘scarce’ in both regions (2006/07 to 2011/12) (Gollop 2012).
Manitoba
Wolverines inhabit the northern part of the province, north of 53° latitude (Berezanski 2004).Highest densities appear to be in the northeast, but the northcentral and northwestern regions receive less trapping effort and may represent a population reservoir for surrounding regions. Wolverines were historically rare in southern Manitoba, where human developments forced the range limit north (Berezanski 2004).
Using harvest data, the Wolverine population in Manitoba was estimated at between 500 and 800 animals (Johnson 1990). Berezanski (2004) used a sustainable harvest rate of 6% (from Krebs et al. 2004), and the assumption that the harvest was sustainable in Manitoba, to estimate a population of 1100-1600, with 1100 more likely because of presumed lower productivity compared to other areas in western Canada (Berezanski 2004).
An increase in harvest in the 1980s was due to a number of factors, including the cessation of Wolf poisoning, an increase in carrion from Wolf kills, a trapping season closing date that offered some protection to females with kits (which may allow increased production), and an increase in fur prices and associated trapping activity (Johnson 1990). The Wolverine harvest rate since 1980 has been highly variable, but has not significantly changed (Berezanski pers. comm. 2013). The population is thought to be stable overall (Berezanski 2004) and based on harvest records, may be increasing in the Churchill area in the northeast and possibly the south (Berezanski pers. comm. 2013). Wolverine populations in northeastern Manitoba may be benefiting from the increase in the Pen Island (Eastern Migratory) Caribou herd, which is having the same beneficial effect in northwestern Ontario (Dawson 2000; Berezanski 2004).Keeyask Hydro Limited Partnership (2012) reported Wolverine populations to be increasing in the Keeyask region and lower Nelson River region of northern Manitoba, with increases first noted in 2009, albeit this evidence came from a single trapper.
Ontario
Some ATK indicates that Wolverines have always been rare, but have been consistently present with no discernible changes in relative abundance since the 1920s, yet others suggest increases since the late 1970s or early 1980s (Ray 2004). Wolverines reappeared in the Winisk-Peawanuck area in the mid-1990s (Dawson 2000).
Wolverine distribution in Ontario was summarized by the Ontario Wolverine Recovery Team (2013). Wolverines are currently found in northwestern Ontario following a range reduction in the 1800s due to habitat conversion during human settlement, logging and railroad construction, and the overharvest of ungulates (Ontario Wolverine Recovery Team 2013).
There is some evidence from aerial surveys and ATK that Wolverine have recently expanded their range east of the ‘peripheral’ range (Figure 4) following increases in the Pen Islands Caribou herd in the 1980s and 1990s (Dawson 2000), and changes in Boreal Caribou distribution in the Hudson Bay Lowland (Magoun et al. 2004, 2005a; Ray 2004). Ray (2012) recorded incidental Wolverine observations to < 200 km from the Québec border. A Wolverine was harvested in 2012 near the same limit (Dawson pers. comm. 2012). Kelsall (1981) believed that the Wolverines in Ontario were isolated from those in Québec and Labrador, but that may no longer be the case.
The population estimate for Wolverines in Ontario is 458 to 645 (Bowman pers. comm. 2013), based on an estimated minimum density of 1.4 Wolverines/1,000 km2 in a 7,626 km2 study area, where a DNA mark-capture study was undertaken (Dawson et al. unpubl. data). If Wolverines persist at similar densities across the province, the Ontario population estimate would be 645. However, results from aerial surveys (Ontario Wolverine Recovery Strategy) suggest that densities are not uniform and this estimate was assumed to be representative of the higher end of the spectrum in the province. Therefore, dividing Wolverine range into high (274,551 km2) and low density (172,563 km2) areas, and assuming the density in the latter are to be 0.7/1,000 km2, yields an estimate of 458 Wolverines. There are no confidence limits on this estimate.
A zero quota was applied to non-aboriginal trappers in 2001/02 and the season was closed in 2009, but occasional accidental kills of 1 - 4 occur annually (Dawson pers. comm. 2002; Bowman et al. 2010).Treaties in northern Ontario allow for Aboriginal peoples to continue harvesting Wolverines for their own use (Heydon pers. comm. 2002).
Québec
The historical harvests of Wolverine in Québec are not well documented. Historical Hudson’s Bay Company fur trade data from Fort Chimo (now Kuujjuak) from 1868 to 1923 documents a harvest of ≥1190 Wolverine pelts, with a sharp decline from 23 pelts/year, to 4 pelts/year in the 1914-1923 period (Schmelzer unpubl. data 2012). Harvests ranged between 0 and 24 per winter since Statistics Canada began keeping records in 1919 (Fortin et al. 2005). These data have led to the assumption that the species was never particularly abundant in the Eastern Population (Fortin et al. 2005). The harvest of Wolverines was banned in Québec in 1981, except in the James Bay and northern Québec Agreement area, which covers much of northern Québec. Current and past generations of Aboriginal knowledge holders from northern Québec report Wolverine as being very rare (COSEWIC 2012b).
Wolverines have not been confirmed to occur in Québec since the last specimen was captured in 1978. An individual captured in the Lanaudière region in 2004 was cited in Fortin et al. (2005) as evidence of remnant Wolverines but it has since been established that it had escaped from a zoo in Ontario (Ministère des Ressources naturelles et de la Faune du Québec 2013). There were about 60 unconfirmed reports from Québec and Labrador between 1965 and 2004 (Fortin et al. 2005). Over 70 observations were reported between 2000 and 2012 (Thibault unpubl. data 2013; Figure 5); 14 were considered ‘probable’, 35 'likely', and 22 'questionable', but none were verified. Unverified records based on tracks should be viewed with some skepticism because track size and shape varies with changing snow conditions (McKelvey et al. 2008).
Based on recent observations and research (see Search Effort section), Wolverines are either very rare, are at a level where they are not detectable using current survey effort, or are extirpated in Québec.
Labrador (mainland Newfoundland and Labrador)
A review of historical Wolverine fur trade data from Hudson’s Bay Company posts in Davis Inlet and Moravian Missions of Hebron and Okak in Labrador indicated 873 Wolverine pelts were traded between 1844 and 1923 (Schmelzer unpubl. data 2012). Some pelts traded at Fort Chimo, Québec likely originated from Labrador (Moores pers. comm.). Wolverine pelts continued to be traded until the mid-1900s (Schmelzer 2006).
Over 40 unconfirmed sightings of Wolverines were reported over the last 30 years (Schmelzer 2012). The lack of verified records from numerous surveys (see Search Effort section) led to a conclusion that Wolverines may be extirpated from their range in Labrador, and that populations in neighbouring Québec are too low, or absent, to support recolonization (Schmelzer unpubl. data 2012). Innu elders report knowing little about Wolverine at present and suggest they are very rare (Paul F. Wilkinson and Associates Inc. 2009), although they were relatively abundant around 1935-1940 in the Schefferville area of Québec and Labrador (Clement 2009).
Canadian Population Estimate
The population is unknown, but likely > 10,000 adults. If harvest data are accurate, and if population size can be adequately extrapolated from density estimates, the Wolverine population estimates (mature individuals) would be as follows: Yukon (3,500 to 4,000), NWT (3,430-7,325), BC (2,700 to 4,760), Saskatchewan (<1,000) and Manitoba (1,100 to 1,600). The available information on distribution and known densities in similar ecological areas suggests that populations in the remaining jurisdictions are approximately as follows: Alberta, 1,500 to 2,000 (Wolverine range is relatively extensive, with some high quality habitats); and Nunavut, 2,000 to 2,500.The estimate for Ontario is 458 to 645 Wolverines. The result is a total Western sub-population estimate of 15,688 to 23,830 adults.However, these estimates assume trapping effort is constant and that Wolverine densities are relatively uniform across the range.
There is no estimate for the Eastern Wolverine sub-population and the last verified record was in 1978. The lack of verified records among the many reported observations (Figure 4) implies that the population is very small (<100 individuals), and possibly extirpated.
Fluctuations and Trends
Wolverine population fluctuations and trends were discussed by jurisdiction in the Abundance section. To summarize, population estimates are uncertain but there are likely >10,000 Wolverine in the population. Trend is unknown but information from harvest and trapper survey indices suggests sub-populations are increasing in parts of the NWT, Nunavut, northern Alberta, Manitoba, and Ontario, but decreasing in the southern range. Extirpations (or near extirpations) have occurred in a substantial portion of the historical range, including eastern Canada, the Prairies, and Vancouver Island. It is unknown if past declines in the Southern Mountains National Ecological Area of BC and Alberta have continued since the last assessment (COSEWIC 2003).
There is no evidence of population changes in the range of the eastern sub-population (Québec-Labrador).
A minimum of 559 Canadian Wolverine were killed in 2009-2010, and over the last 17 years (1992-1993 to 2009-2010, there have been similar harvest levels (range 385-637) across Canada (Table 1).
Rescue Effect
Nuclear DNA evidence of genetic similarities suggests connectivity currently exists between Alaska and western Canada (Gardner et al. 1986; Kyle and Strobeck 2001, 2002). Wolverine populations in Alaska are continuous with those in the Yukon and BC, making rescue along the international boundary with Alaska likely. Wolverine density estimates in Alaska are presented in Table 2.
The likelihood of rescue of Canadian Wolverine populations from the conterminous United States is negligible. Increasingly fragmented populations currently inhabit montane regions in Washington, Idaho, western Montana and Wyoming (Aubry et al. 2007; Figure 3). The total number of Wolverines inhabiting this area may be 35 (credible limits 28-52) (Schwartz et al. 2009). Genetic sub-structuring has been documented in Wolverine populations in Idaho (Kyle and Strobeck 2002) and Montana (Cegelski et al. 2003, 2006), suggesting relatively low migration rates between adjacent mountain ranges.
Threats and Limiting Factors
An IUCN Threats Calculator was conducted among jurisdictions and COSEWIC members in order to document the scope and extent of major threats (Appendix 1). The overall Threat score was ‘medium’, due mainly to concerns with declining Caribou populations, response to disturbances such as road and infrastructure development, and access leading to potential unsustainable harvest levels.
Threats have been identified in several recent documents, including the 2003 COSEWIC Status Report (COSEWIC 2003), the Ontario Wolverine Recovery Plan (OWRT 2013), and the Eastern Population Recovery Plan (Fortin et al. 2005). Habitat loss, habitat fragmentation, and climate change continue to threaten Wolverine populations at the southern and eastern peripheries of their range. Habitat loss results from conversion of natural habitats for human land uses including agriculture, ranching, urban and suburban developments, mining, oil and gas development, hydroelectric reservoirs, and associated road networks that facilitate access for harvest and disturbance.
Threats to the eastern sub-population relate mainly to loss of carrion associated with declining Caribou herds, hydroelectric developments, and road density in areas with increased resource development. Threats in the southwestern range relate mainly to declining Caribou, roads, increased access and potential overharvest, and climate change. Threats in the northern range relate mainly to resource development sites, harvest, and declining Caribou in some areas. Details are discussed below.
Harvest
Harvest is presently the human activity with the greatest potential to directly reduce Wolverine populations to levels which could have a detrimental effect on metapopulation dynamics and cause extirpation (Lofroth and Ott 2007). In BC, the harvest of Wolverine was stable at the provincial level, but was unsustainable in 15 of 71 population units (Lofroth and Ott 2007). Low densities, large home range size, and long-distance movements by dispersing individuals contribute to the Wolverines’ vulnerability to trapping and hunting. An increased vulnerability of Wolverines to trapping after Snowshoe Hare population crashes leads to the false perception that they are more abundant (Hatler 1989; Jung et al. 2005; Slough 2009). Banci (1987) noted an increase in the harvest of adult males in March 1983 following a Snowshoe Hare population crash.
Harvest levels have remained stable in western jurisdictions (Table 1). Harvest management, including trapping closures, limited seasons, quotas, limited entry, and registered trapping concessions have reduced these threats in most jurisdictions (Slough et al. 1987; Johnson 1990). Beneficiaries of the Nunavut Land Claims Agreement are not restricted to these harvest rules (although members may have local restrictions; Crystal pers comm. 2014) and harvest rates by beneficiaries are not as well documented. Exported Wolverine furs require permits but a high proportion of Wolverines from the Arctic are used locally and not exported to fur auctions, so there is a potential for overharvest going unnoticed. The estimated annual harvest of Wolverines (including unrecorded harvests from Yukon, NWT and Nunavut) is likely <1,000.
Survival was <0.75 among all age/sex classes in trapped areas, and >0.84 in areas where trapping does not occur (Krebs et al. 2004). Intrinsic rates of increase (λ) were estimated at 0.88 in trapped populations and 1.06 in untrapped populations. Survival was highest among adult females (0.88 in untrapped areas, 0.73 in trapped areas) and lowest among subadult males (0.45 in trapped areas) (Krebs et al. 2004). This evidence suggests that most trapped populations would decline without immigration from refugia. Local overharvest of Wolverines may occur in some areas (Krebs et al. 2004), but this harvest is largely replenished by immigrants from untrapped refugia. Cardinal (2004) identified several refugia from trapping across northern Canada. The consideration of use of untrapped areas as functional refugia in trapline management is a strategy practised by many trappers and promoted in trapline education (e.g., Yukon Department of Renewable Resources, no date). Many refugia are de facto, where trapping areas are simply large and inaccessible. Krebs et al. (2004) pooled data from 12 North American studies, and estimated that at an average harvest rate of 12.2%, and an intrinsic rate of increase of 6.4%, refugia from trapping would need to cover twice as much area as harvested areas. Self-management by trappers and inaccessibility are the primary sources of refugia. The percentage of land not subject to harvest is unknown but expert opinion during the Threats Calculator exercise estimated that 70-100% of the range (in jurisdictions where trapping is allowed) would be exposed to potential harvest. It is noted that refugia are not regulated and the reliance on refugia to maintain a sustainable harvest is at risk of failure if incentives for harvest increase and refugia become trapped.
Refugia can quickly become accessible; in northeastern BC, 279,595 km of forest was cleared for seismic lines between 1996-2008, which is approximately an extra 2.54 km/km2of access for trappers (Weir pers. comm. 2014). The use of snowmobiles to run down Wolverine, or to get close and shoot one, is common in some jurisdictions (e.g., Yukon; Jung pers. comm.) and there is concern that harvest by snowmobile, rather than traditional set traps increases the range of harvest and minimizes actual trap refugia.
Roads and Transportation Corridors
The increased density of roads in logged areas and elsewhere may have a direct effect on Wolverine and their prey (e.g., Caribou), through disturbance (Krebs et al. 2007; Bowman et al. 2010). Roads also place Wolverines vulnerable to trapping and road mortality. Gwich’in in the Mackenzie River region suspect the Mackenzie Valley Highway will increase access to harvest, and roadkill of Wolverine (GRRB 2014). May et al. (2006) found that Wolverines in Scandinavia selected home ranges based on degree of human development and, to a lesser extent, habitat quality, and in Ontario, road densities had an apparent influence on home range selection (Dawson et al. 2010). Areas with roads or human structures were avoided or selected less than large roadless areas. A similar relationship was found in the northwestern conterminous US where road density or human population density were more important than quantity or quality of vegetation in determining Wolverine counts (Rowland et al. 2003). Their watershed-scale models predicted lower Wolverine counts at higher road densities (road densities of 0.44 to 1.06 km/km2). In the Rocky Mountain region, Carroll et al. (2001) predicted a decline in the occurrence of Wolverines when road densities exceeded approximately 1.7 km/km2. The mean road density in Wolverine home ranges was 0.43 km/km2 in Ontario, and individuals whose home ranges had a higher road density than the mean had a higher risk of mortality due to anthropogenic factors (Bowman et al. 2010). Wolverines were less abundant in areas with human activities including roads (mean road density of 0.37 km/km2) and logging (i.e., temporary and functional habitat losses) (Bowman et al. 2010). Resource development is often accompanied by road access, so road density serves as a proxy for the extent of the human footprint (Bowman et al. 2010). Road density is not only associated with avoidance behaviour by Wolverine, but there is an increased mortality risk due to trapping, hunting, and collisions with vehicles.
Large highways and other transportation corridors such as those in southern Alberta and BC may act as barriers to movements and dispersal, and may be a source of mortality (Gibeau and Heuer 1996). Disturbance by noise (Golder Associates 2003) and traffic may lead to avoidance of transportation corridors. Gibeau and Heuer (1996) documented two highway mortalities of Wolverine in Banff National Park, where they also noted patterns of approach and retreat behaviour before successfully crossing roads during busy periods. Austin (1998) found that Wolverines selected narrow crossings (i.e. <100 m wide), avoided areas within 100 m of the Trans-Canada Highway, and preferred areas more than 1,100 m away. Alexander et al. (2005) studied the effect of traffic volume on the carnivore guild, which included Wolverines, in the same area. They suggest that a carnivore movement threshold exists between 300 and 500 vehicles per day, or an average annual daily traffic volume (AADT) of 3000-5000, assuming a 10:1 annual to winter traffic ratio. No wolverine crossings (n=39) were detected at that traffic volume (Alexander pers. comm. 2013). The AADT in Banff National Park is 14,000, and similar volumes are likely for the Trans-Canada highway westward through Wolverine range.
Wolverine gene flow and movements across highway crossing structures (underpasses/overpasses along fenced sections of highway) are being assessed on an additional 30 km of the Trans-Canada Highway (TCH) twinned in Banff National Park (Clevenger et al. 2011a). A larger study is monitoring the occurrence and rate of all wildlife-vehicle collisions and use of crossing structures (Clevenger et al. 2011b). In the 17 years (1996-2014) of monitoring of the TCH, there have been 10 recorded crossings (9 using underpasses, 1 using overpass) by at least two different Wolverines (Clevenger, 2013; Clevenger pers. comm. 2014). Genetic typing from hair samples indicates five males and two females crossed the TCH over a three-year period, though they were most likely crossing at unfenced sections. There have been four mortalities since 1980 at unfenced parts of the TCH. Sample size is low but early results suggest that the TCH is not a barrier to males, and may be for females (Clevenger pers. comm. 2014).
Biological Factors
Biological factors that limit Wolverine populations include the species’ low intrinsic rate of increase, low natural densities and large home ranges (Banci and Proulx 1999). Conversely, long-distance dispersal abilities give Wolverines the capacity to recolonize vacant habitats (Vangen et al. 2001; Flagstad et al. 2004). Re-population may take several decades but is possible where factors favour Wolverine survival (Johnson 1990; Vangen et al. 2001).
Indirect effects on the prey base and large carnivores, which impact scavenging opportunities, will also impact Wolverine populations. Such effects include overhunting of ungulates and carnivores, and population declines of prey and carrion due to loss and fragmentation of their habitats (Cardinal 2004). A recent recolonization of southern Scandinavia by Wolves led to an increase in scavenged Moose in the diet of Wolverine, and a decrease in the occurrence of Wolverine-killed Caribou and rodents (van Dijk et al. 2008).
The availability of prey and carrion is another biological limiting factor (see Habitat Requirementssection). Wolverines rarely prey on livestock in North America (as they do in Europe; Landa et al. 1997) and so are not directly targeted for predator control. Wolf poisoning programs in western Canada that killed Wolverines as by-catch ended in the 1970s, but were reinstated in west-central Alberta by 2005. Wolverines may be impacted by Wolf reduction programs designed to reduce predation on declining Boreal Caribou herds.
Climate Change
Spring snow cover during the denning period is an important habitat requirement of Wolverines (Aubry et al. 2007; see Habitat section). Climate models predict increases in temperature and precipitation in Canada (IPCC 2007), with the largest warming projected for northern Canada. Precipitation is likely to increase in winter and spring, but decrease in summer. Snow season length is predicted to decrease, but a net increase in snowfall should make up for the shorter snow season, resulting in a net increase in snow accumulation. Spring snow cover in the Arctic has melted about 4-7 days earlier since the late 1980s (Foster et al. 2008), and earlier snowmelt is also occurring in the western mountains (Stewart 2009).
The impact of reduction of snow on Wolverine is uncertain (Wolverine Science Panel 2014). In northern range, earlier snowmelt may not be an issue because snow persists during the natal denning period (Magoun and Copeland 1998). However, in mountainous regions, particularly in BC and Alberta, decreasing spring snow is considered a serious threat. Productivity could decrease, range could shift and habitats and populations could become more fragmented (Copeland et al. 2010; McKelvey et al. 2011; Weir pers. comm. 2013).
Public Attitude
Societal views on Wolverine vary from dislike due to the damage they cause to traps, animals, cabins, and food caches, to respect for their abilities (see Special Significancesection). Attitudes could be an impediment to recovery of Wolverine population in Québec and Labrador where even low mortality levels would significantly impact the population (Fortin et al. 2005).
Recreational Activities
Wolverines prefer large roadless areas, but home ranges frequently overlap active traplines, cross-country ski trails, busy roads such as logging roads, seismic lines (Krebs et al. 2007), and the edges of communities. Winter recreation in the form of helicopter skiing and backcountry skiing, snowmobiling (e.g., high-marking) and the presence of roads reduced habitat values (i.e., functional losses) for Wolverines in the Columbia Mountains, BC (Krebs et al. 2007).
Habitat avoidance results from human activities such as backcountry recreation, which may impact Wolverine behaviour patterns such as denning, kit rearing, travel and foraging. Disturbance of Wolverine maternal den sites may lead to den relocation or litter abandonment (Myrberget 1968; Pulliainen 1968). Evidence for direct impacts of winter recreation on denning Wolverines is conflicting (Copeland 2009; Heinemeyer and Squires 2012), but helicopter skiing and backcountry skiing in western Canada have been shown to reduce functional habitat values for Wolverines, especially adult females (Copeland et al. 2007; Krebs et al. 2007; May et al. 2012), whose reproductive success may be affected (Carroll et al. 2001; Rowland et al. 2003; Copeland et al. 2007; Krebs et al. 2007; Copeland 2009). Aboriginal knowledge holders feel that the use of snow machines has changed harvesting practices and patterns, allowing people to travel further and faster, which has resulted in increased Wolverine harvest (WMAC and Aklavik HTC 2003; Cardinal 2004; Banci et al. 2005). Wolverines opportunistically use inactive snowmobile trails for travel and scavenging trapped animals and hunter kills.
Conflicts at Resource Development Sites
Wolverine-human conflicts at mining camps have recently been identified as an potential threat to Wolverine populations in the NWT and Nunavut, where diamond mines in particular are becoming more common in the Southern Arctic ecozone. Wolverines generally avoid areas of human activity but are curious, and will investigate campsites, food caches, garbage dumps, cabins, traplines, and hunter kills, usually when humans are not present, and scavenge for food. Occupied mining camps are approached in the NWT and Nunavut for food and shelter.
Wolverines usually maintain a separation distance of at least 500 m from human activity (AXYS 1998), but attraction to food sources at camps and other areas of human activity (see Physiology and Adaptability section) may result in the habituation of animals, and increased vulnerability to problem wildlife control, hunting and trapping, and collisions with vehicles (NSMA 1999; Golder Associates 2003). Adaptive mitigative measures are used to reduce Wolverine occurrences at diamond mines (Golder Associates 2012a, b; Rescan 2012). There were one nuisance-control mortality and four relocations at the Ekati diamond mine between 2003-2011 (Rescan 2012) and one accidental human-related Wolverine mortality at Snap Lake in 2011 (Golder Associates 2012a). There were two incidents of Wolverines being deterred from the camp prior to 2008, and 8 cases in 2008 (Golder Associates 2010). There were 47 incidents involving Wolverine deterrence from 2000 to 2011 at Diavik mine. There were also two relocations and two mortalities (DDMI 2012). There were 30 incidents involving Wolverine deterrence and two reported mortalities at Meadowbank gold mine in the Kivalliq region, Nunavut, in 2011 and 2012 (Lecomte pers. comm. 2013). ConocoPhillips (2006) waste management plan recognizes that certain petroleum-based products such as lubricants may attract Wolverine. Wolverine mortalities and relocations as a result of diamond mining activities appear minor, but in conjunction with trapping and hunting, may be contributing to local population declines (Boulanger and Mulders 2013a).
Responses of Wolverines to flying aircraft are variable, ranging from no response to running away and hiding (AXYS 1998). Generally, Wolverines are reluctant to abandon carrion, even when disturbed. A lack of response was most common (38 of 40 encounters) in fly-overs in the Yukon (Jung pers. comm. 2013).
Number of Locations
The term 'location' defines a geographically or ecologically distinct area in which a single threatening event can rapidly affect all Wolverines present. As such, there are 'many' locations in Canada.
Protection, Status, and Ranks
Legal Protection and Status
In Canada, Wolverines are harvested in all northern and western jurisdictions (Manitoba, Saskatchewan, Alberta, BC, Yukon, NWT, and Nunavut).There is no non-aboriginal harvest permitted in Québec, Labrador or Ontario (since 2001/02).Aboriginal harvest is permitted in the northern part of the range in Québec, as per the James Bay and Northern Québec Agreement: Fortin et al. 2005). The incidental harvest of Wolverine still occurs in Ontario (Ontario Wolverine Recovery Team 2013). In the United States, Wolverine harvest is permitted in Alaska and Montana, although it has been temporarily closed since November 2012 in Montana.
Harvests are managed with spatial and temporal restrictions including variations in season length, quotas, limited entry (e.g., the use of registered trapping concessions, group and community trapping areas), and trapline management by the trapper (Slough et al. 1987).Harvests are monitored either through mandatory pelt sealing, year-end harvest reporting, or by monitoring fur exports.Local use of Wolverine pelts is common, particularly in Inuvialuit and Inuit communities in the NWT and Nunavut, where harvests are monitored using carcass collection programs (see Sampling Effort and Methods section).
Wolverines are not listed by the Convention on International Trade in Endangered Species; international trade in Wolverines is not monitored or restricted.
Non-Legal Status and Ranks
Wolverines were ranked Least Concern by IUCN in 2009 (IUCN 2012) due to their wide distribution and remaining large populations. Although there is an overall continued decline due to human persecution and land-use change, it is not at a rate sufficient to qualify for listing at this time. Previous listings by IUCN were Near Threatened (2008) and Vulnerable (1996).
The provincial/territorial status rankings for Wolverines are given in Table 3.Most rankings are similar to the COSEWIC (COSEWIC 2003) rankings for the Eastern and Western populations (Endangered and Special Concern, respectively). Provincial/territorial rankings are Endangered or Threatened for the Eastern Population, and Sensitive or Special Concern for the Western Population, although Wolverines are ranked Threatened in Ontario.
| Jurisdiction | COSEWIC / SARA | General Status Footnotea.3 | Provincial/ Territorial |
NatureServe Footnoteb.3 |
|---|---|---|---|---|
| Global IUCN: Least Concern |
- | - | - | G4 |
| U.S.A | - | - | - | N4 |
| WA | - | - | - | S1 |
| CA | - | - | - | S1 |
| CO | - | - | - | S1 |
| Idaho | - | - | - | S2 |
| WY | - | - | - | S2 |
| OR | - | - | - | S2 |
| MT | - | - | - | S3 |
| AK | - | - | - | S4 |
| Canada | - | 3 | - | N3N4 |
| NL | Endangered; Schedule 1 | 1 | Endangered | S1 |
| QC | Endangered; Schedule 1 | 1 | Threatened | S1 |
| ON | Special Concern; No Status | 1 | Threatened | S2S3 |
| MB | Special Concern; No Status | 3 | Special Concern | S3S4 |
| SK | Special Concern; No Status | 1 | No Provincial Ranking | S3S4 |
| AB | Special Concern; No Status | 2 | Data Deficient | S3 |
| BC (luscus ssp.) | Special Concern; No Status | 3 | Blue (Special Concern) | S3 |
| BC (vancouverensis ssp.) | Special Concern; No Status | No Status | Red (Extirpated, Endangered, or Threatened) | G4T1QSH |
| YT | Special Concern; No Status | 3 | No Territorial Ranking | S3 |
| NT | Special Concern; No Status | 3 | No Territorial Ranking | S3 |
| NU | Special Concern; No Status | 4 | Sensitive | SNR |
General status ranks (Canadian Endangered Species Conservation Council (CESSC) 2011) (Table 3) are Secure (NU), Sensitive (Canada, YT, NT, BC, MB), May Be at Risk (AB) and At Risk (SK, ON, QC, NL).
State listings for Wolverines are: Washington (Protected Wildlife; candidate), Idaho (Protected Nongame Species), Wyoming (Vulnerable), and Montana (Candidate). There is no state listing for Alaska. NatureServe ranks are provided in Table 3.
The U.S. Fish and Wildlife Service (2013) has proposed listing Wolverines in the conterminous United States as Threatened under the Endangered Species Act. The effect of climate warming on the snow pack is listed as the major threat, while backcountry recreation, timber harvesting and infrastructure development are not considered significant threats and therefore will be allowed to continue.
Habitat Protection and Ownership
Most of the Wolverine’s range is on federal land administered by Aboriginal Affairs and Northern Development Canada. Modern land claim and self-government programs have resulted in the ownership of settlement lands by Aboriginal governments and authorities in Yukon, NWT, Nunavut, and BC.Land use activities on both Aboriginal and non-aboriginal lands, not defined as settlement lands, are administered by the applicable land manager in consultation with First Nations, Inuvialuit, and Inuit.
Six communities in the Inuvialuit Settlement Region (Aklavik, Ulukhaktok, Inuvik, Paulatuk, Sachs Harbour, Tuktoyaktuk) have adopted community conservation plans, updated in 2008 (Joint Secretariat). Recommended Wolverine conservation measures include the identification and protection of important habitats from disruptive land uses, avoidance of den disturbance, discouraging hunting in summer, and forbidding the use of poison. Some conservation plans specify winter Wolverine harvesting areas.
Department of National Defence establishments in some parts of the country are used by Wolverine, including NWS radar sites in the Arctic (Shingle Point, Yukon; Horton River, NWT; Hall Beach, Nunavut), and properties in the Chilcotin (west of Williams Lake, BC) and Pierce Creek (south of Chilliwack, BC) (Nernberg pers. comm. 2012). Wolverines are suspected to be present at CFB Cold Lake, Alberta.
Kelsall (1981) and Dauphiné (1989) considered parks to be refugia from trapping and resource developments but some caution seems necessary in parts of the range because Wolverine trapping is permitted in most northern National and Provincial Parks of the western range. Also, Wolverines have large spatial requirements (see Habitat section) and sub-populations within the parks are not buffered from trapping activities around their peripheries. Wolverines that reside partly in refugia are susceptible to harvest mortality. Recreational activities, like snowmobiling and skiing, during the late winter denning period may result in disturbance to females and their litters, leading to relocation or abandonment (Heinemeyer and Squires 2012). The size requirement for an effective refugium from development has not been determined, but refugia larger than 20,000 km2 may be required for an isolated habitat to maintain a viable population (Magoun et al. 2005b).
Trapping is permitted in many of the BC provincial parks, and by First Nations, Inuvialuit and Inuit Beneficiaries in national parks in Yukon, NWT, and Nunavut Territory, and in Wood Buffalo (NWT and Alberta) and Wapusk (Manitoba) National Parks. Most types of resource extraction or activities contributing to habitat losses and fragmentation are not permitted in protected areas.However, recreational activities, such as snowmobiling and skiing, which may disturb denning Wolverines, are generally not restricted and transportation corridors bisect and penetrate parks.
In Ontario, the habitat of Wolverine is protected from damage and destruction under the Endangered Species Act, 2007, and forest management operations are modified to maintain buffers around known Wolverine dens (OMNR 2010).
Acknowledgements and Authorities Contacted
The report writer would like to acknowledge the contributions of persons listed under 'Authorities Contacted', who provided much of the information for this report as well as Donna Bigelow, Jeff Bowman, Marsha Branigan, Tony Clevenger, Robert Craig, Jason Fisher, Clément Fortin, Howard Golden, Bob Inman, Lois Koback, Chris Kyle, Benoit Landry, Mireille Pilotte, Kim Poole, Justina Ray, Francois Rossouw, Isabelle Schmelzer, Sarah Spencer, Isabelle Thibault and Joanna Zigouris for providing unpublished papers, reports and data for use in this status assessment.
The report writer thanks Graham Forbes, Co-chair, COSEWIC Terrestrial Mammals Specialist Group for facilitating the project in many ways.I am indebted to the Aboriginal Traditional Knowledge (ATK) subcommittee who compiled and integrated Aboriginal Traditional Knowledge into this report.
Previous COSEWIC status reports on the Wolverine were prepared by John P. Kelsall (1982), T. Charles Dauphiné (1989), and Brian G. Slough (COSEWIC 2003).
The pen and ink drawing of the Wolverine is by Yukon artist and illustrator Lee Mennell.Jenny Wu, Scientific Project Officer, COSEWIC Secretariat, coordinated mapping and area calculations.
Authorities Contacted
Anderson, Robert. June 2012. Research Division, Canadian Museum of Nature, Ottawa, ON.
Awan, Malik. October 2013. Wildlife Biologist - Carnivores, Department of Environment, Igloolik, NU.
Berezanski, Dean. June 2012. Furbearer and Problem Wildlife Management Unit, Wildlife and Ecosystem Protection Branch, Manitoba Conservation and Water Stewardship, Winnipeg, MB.
Bigelow, Donna. June 2012. Species at Risk Biologist, Canadian Wildlife Service, Environment Canada, Yellowknife, NWT.
Bilyk, Lonnie. June 2012. Resource Data Biologist, Alberta Fisheries and Wildlife Management Information System (FWMIS), Fish and Wildlife Division, Alberta Sustainable Resource Development, Edmonton, AB.
Blaney, Sean. June 2012. Botanist / Assistant Director, Atlantic Canada Conservation Data Centre, Sackville, NB.
Bradley, Alex. June 2012. Resource Biologist, Wildlife Management Advisory Council – Northwest Territories, Inuvik, NWT.
Branchaud, Alain. June 2012. Wolverine Recovery Team (Eastern Population) Chair, Species at Risk Recovery Biologist, Canadian Wildlife Service, Environment Canada, Montréal, QC.
Branigan, Marsha. December 2012. Manager, Wildlife Management, Environment and Natural Resources, Inuvik Region. Inuvik, NWT.
Brown, Rosa. January 2013. Secretariat, Wildlife Management Advisory Council (North Slope), Whitehorse, YT.
Brownell, Vivian. June 2012. Senior Species at Risk Biologist, Species at Risk Branch, Ministry of Natural Resources, Peterborough, ON.
Carrière, Suzanne. October 2012. Biologist – Biodiversity, Wildlife Division, Environment and Natural Resources, Yellowknife, NWT.
Clark, Karin. June 2012. Wildlife Management Biologist, Wek’èezhìi Renewable Resources Board, Yellowknife, NWT.
Clevenger, Tony. April 2014. Senior Wildlife Research Scientist. Western Transportation Institute, University of Montana, Bozeman, Montana.
Corrigan, Rob. October 2012. Big Game Allocation Biologist, Game and Priorities Species, Sustainable Resource Development, Edmonton, AB.
Court, Gordon. September 2012. Provincial Wildlife Status Biologist, Fish and Wildlife Division, Sustainable Resource Development, Edmonton, AB.
Craig, Robert. June 2012. NHIC Information Analyst, Natural Heritage Information Centre, Ministry of Natural Resources, Peterborough, ON.
Crowley, Shannon. June 2012. Senior Biologist, Wildlife Division, Department of Environment and Conservation, Happy Valley-Goose Bay, NL.
Crystal, Danica. April 2014. Wildlife Management Biologist. Nunavut Wildlife Management Board, Iqaluit, NU.
Dawson, Neil. September 2012. Wildlife Assessment Program Leader, Ministry of Natural Resources, Northwest Science and Information, Rosslyn, ON.
Dumond, Mathieu. October 2012. Manager of Wildlife Research and Kitikmeot Regional Biologist Kitikmeot, Department of Environment, Kugluktuk, NU.
Duncan, Dave. June 2012. Manager, Population Conservation Section, Canadian Wildlife Service, Environmental Stewardship Branch, Prairie and Northern Region, Environment Canada, Edmonton, AB.
Duncan, James. October 2012. Director, Wildlife Branch, Manitoba Conservation and Water Stewardship, Winnipeg, MB.
Durocher, Adam. June 2012. Data Manager, Atlantic Canada Conservation Data Centre, Corner Brook, NL.
Eaton, Samara. November 2012. Wildlife Biologist, Species at Risk Recovery, Canadian Wildlife Service, Environmental Stewardship Branch, Environment Canada, Atlantic Region, Sackville, NB.
Falardeau, Gilles. June 2012. Liaison Officer COSEWIC and SAR biologists, Canadian Wildlife Service, Environment Canada, Ste-Foy, QC.
Fisher, Jason. June 2012. Research Scientist – Wildlife Ecology, Alberta Innovates - Technology Futures, Vegreville, AB, and Adjunct Assistant Professor, School of Environmental Studies, University of Victoria, Victoria, BC
Firlotte, Nicole. October 2012. Conservation, Wildlife and Ecosystem Protection Branch, Conservation Data Centre Acting Coordinator, Conservation and Water Stewardship, Winnipeg, MB.
Forbes, Graham. July 2012, January 2013. Co-Chair, Terrestrial Mammals Specialist Subcommittee and Faculty of Forestry and Environmental Management. University of New Brunswick, Fredericton, NB,
Fortin, Clément. June 2012. Président, Carcajou Québec, Tewkesbury, QC.
Fournier, François. June 2012. Manager – Wildlife Research Science and Technology Branch, Environment Canada, Québec, QC.
Fraser, Dave. June 2012. Scientific Authority Assessment, A/Manager BC CDC, Ecosystem Branch, Conservation Planning Section, Ministry of Environment, Victoria, BC.
Gallais, Sophie. October 2012. Chargée de projet Aires protégées, Nature Québec, Québec, QC.
Gauthier, Isabelle. June 2012. Biologiste en conservation, Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Direction générale de l’expertise sur la faune et ses habitats, Québec, QC.
Giasson, Pascal. June 2012. Manager, Species at Risk and Protected Natural Areas, NB Dept. of Natural Resources, Fredericton, NB.
Gignac, Hilary. July 202. Stewardship Coordinator, Thunder Bay District Stewardship Council, Thunder Bay, ON.
Goudie, James. June 2012. Wildlife Manager, Nunatsiavut Government, Postville, Nunatsiavut, NL.
Gougeon, Nicole. June 2012. Secretary-Treasurer, Hunting, Fishing and Trapping Coordinating Committee, Montréal, QC.
Han, Siu-ling. June 2012. Head, CWS Eastern Arctic Unit, Iqaluit, NU.
Herdman, Emily. September 2012. Senior Wildlife Biologist, Endangered Species and Biodiversity, Wildlife Division, Department of Environment and Conservation, Corner Brook, NL.
Hurlburt, Donna. July 2012. Co-chair, COSEWIC ATK Subcommittee, Annapolis Royal, NS.
Jessup, Harvey. May 2013. Chair, Yukon Fish and Wildlife Management Board, Whitehorse, YT.
Jones, Neil. July 2012. Scientific Project Officer and ATK Coordinator, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, QC.
Jung, Thomas. June 2012. Senior Wildlife Biologist, Biodiversity, Fish and Wildlife Branch, Environment Yukon, Whitehorse, YT.
Keith, Jeff. October 2012. Biodiversity Biologist, Ministries of Environment, Management and Resource Compliance, Fish and Wildlife Branch, Science and Assessment Unit, Regina, SK.
Khidas, Kamal. October 2012. Curator, Vertebrate Zoology, Research and Collections, Canadian Museum of Nature, Ottawa, ON.
Koback, Lois. June 2012. FWDF/Fur and Problem Wildlife Support, Saskatchewan Ministry of Environment, Fish and Wildlife Branch, Saskatoon, SK.
Kyle, Chris. March 2014. Conservation geneticist. Department of Biology, Trent University, Peterborough, ON.
Landry, Benoit. June 2012. Technicien en cartographie, Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Direction générale de l’expertise sur la faune et ses habitats, Québec, QC.
Lecomte, Nicolas. June 2012. Ecosystem Biologist, Department of Environment, Igloolik, NU.
Levesque, Annie. June 2012. Biologiste en conservation, Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Direction générale de l’expertise sur la faune et ses habitats Québec, QC.
Lofroth, Eric. October 2012. Manager, British Columbia Conservation Data Centre, BC Ministry of Environment, Victoria, BC.
MacDonald, Bruce. June 2012. Chief, Northern Conservation Section, Canadian Wildlife Service, Environment Canada, Yellowknife, NWT.
McDonald, Lauren. June 2012. A/Regional Species at Risk Biologist, Ontario Ministry of Natural Resources, Regional Planning Unit, Northeast Region, Kirkland Lake, ON.
Milliken, Rhonda. June 2012. A/Head Population Assessment, Pacific Wildlife Research Centre, Canadian Wildlife Service, Pacific and Yukon Region, Environment Canada, Delta, BC.
Mitchell Foley, Jennifer. June 2012. Research Program Manager – Wildlife and Plants, Torngat Wildlife, Plants and Fisheries Secretariat, Torngat Wildlife and Plants Co-Management Board, Happy Valley-Goose Bay, NL.
Mulders, Robert. August 2012. Wildlife Biologist, Carnivores/Furbearers, Wildlife Division, Environment and Natural Resources, Yellowknife, NWT.
Nantel, Patrick. June 2012. Science Advisor, Parks Canada, Gatineau, QC.
Nernberg, Dean. August 2012. Species at Risk Officer, Director General of Environment, Directorate of Environmental Stewardship, National Defence, Ottawa, ON.
Nyce, Harry. June 2012. Director of Fisheries and Wildlife, Nisga’a Wildlife Committee, Nisga’a Lisims Government, New Aiyansh, BC.
O'Connor, Mark. October 2012. Director of Wildlife Management, Nunavik Marine Region Wildlife Board, Inukjuak, QC.
Oldham, Michael. June 2012. Ontario Natural Heritage Information Centre (NHIC), Ministry of Natural Resources, Peterborough, ON.
Otto, Robert. October 2012. Labrador Wolverine Working Group Chair, Director, Institute for Biodiversity, Ecosystem Science, and Sustainability, Department of Environment and Conservation, Government of Newfoundland and Labrador, St. John’s, NL.
Paquet, Annie. July 2012. Technicienne de la Faune, Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Direction générale de l’expertise sur la faune et ses habitats, QC.
Pendergast, Sean. October 2012. Senior Wildlife Biologist, Ministry of Forests, Lands and Natural Resource Operations, West Coast Region, Nanaimo, BC.
Moores, Shelley. June 2012. Senior Manager, Endangered Species and Biodiversity, Wildlife Division, Department of Environment and Conservation, Corner Brook, NL.
Phillips, Frank. June 2012. Department of Natural Resources, Happy Valley-Goose Bay, NL.
Pilotte, Mireille. June 2012. Environmental and Social Affairs, New Millennium Iron Corp., and Paul F. Wilkinson and Associates Inc., Montréal, QC.
Pittoello, Gigi. October 2012. Habitat Ecologist, Fish and Wildlife Branch, Saskatchewan Ministry of Environment, Regina, SK.
Poole, Kim. September 2012. Independent Environmental Monitoring Agency (BHP Biliton, Ekati Diamond Mine), Wildlife Research Biologist, Aurora Wildlife Research, Nelson, BC.
Ray, Justina. June 2012. Executive Director/Senior Scientist, Wildlife Conservation Society Canada, Toronto, ON.
Rossouw, Francois. September 2012. Traditional Economy and Fur Management, Industry, Tourism, and Investment, Yellowknife, NWT.
Santiapillai, Crista. December 2012. Statistical Information Service, Statistics Canada. Government of Canada, Ottawa, ON.
Sawa, Ben. June 2012. Saskatchewan Conservation Data Centre, Fish and Wildlife Branch, Saskatchewan Ministry of Environment, Regina, SK.
Schmelzer, Isabelle. September 2012. Senior Wildlife Biologist, Terrestrial, Research, Wildlife Division, Department of Environment and Conservation, Corner Brook, NL.
Schnobb, Sonia. June 2012. Administrative Assistant, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, QC.
Siemens Worsely, Allison. October 2012. Zoologist, Saskatchewan Conservation Data Centre, Fish and Wildlife Branch, Regina, SK.
Simmons, Deborah. June 2012. Executive Director, Sahtú Renewable Resources Board, Tulı́t’a, NT.
Slezak, Barbara. June 2012. Species at Risk Biologist, Canadian Wildlife Service, Environmental Stewardship Branch, Environment Canada, Toronto, ON.
Smith, Jennifer. October 2012. Wildlife Management Advisory Council – North Slope, Whitehorse, YT.
Spencer, Sarah. June 2012. Terrestrial Wildlife Management Biologist, Nunavut Wildlife Management Board, Iqaluit, NU.
Staples, Lindsay. July 2013. Chair, Wildlife Management Advisory Council (North Slope), Whitehorse, YT.
Stipec Katrina. June 2012. BC Conservation Data Centre, Ministry of Environment, Victoria, BC.
Thibault, Isabelle. June 2013. Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Direction générale de l’expertise sur la faune et ses habitats, Québec, QC.
Thomas, Peter. December 2012. Species at Risk Biologist, Canadian Wildlife Service, Environment Canada, Mount Pearl, NL.
Thompson, Amy. June 2012. Executive Director, Gwich’in Renewable Resources Board, Inuvik, NWT.
Trumbley, Dean. COSEWIC ATK Subcommittee representative on Terrestrial Mammals Subcommittee, Salmon Arm, BC.
Tuininga, Ken. June 2012. Senior Species at Risk Biologist, Canadian Wildlife Service, Environment Canada, Ontario Region, Toronto, ON.
Wilson, Joanna. NWT Species at Risk Secretariat, Yellowknife, NWT.
Van Tighem, Graham. June 2012. Executive Director, Yukon Fish and Wildlife Management Board, Whitehorse, YT.
Watkins, Bill. June 2012. Biodiversity Conservation Zoologist, Wildlife Branch, Manitoba Conservation and Water Stewardship, Winnipeg, MB.
Webb, Nathan. October 2012. Carnivore Specialist, Wildlife Management Branch, Environment and Sustainable Resource Development, Edmonton, AB.
Weir, Richard. June 2012. Carnivore Conservation Specialist, Ecosystems Protection and Sustainability Branch, Ministry of Environment, Victoria, BC.
Wu, Jenny. June 2012. Scientific Project Officer, COSEWIC Secretariat, Canadian Wildlife Service, Environment Canada, Gatineau, QC.
Information Sources
Addison, E.M., and B. Boles. 1978. Helminth parasites of wolverine, Gulo gulo, from the District of Mackenzie, Northwest Territories. Canadian Journal of Zoology 56:2241-2242.
Alexander, S.M. pers. comm. 2013. Email correspondence to B. Slough. September 2013. Associate Professor, Department of Geography, University of Calgary, Calgary, AB.
Alexander, S.M., N.M. Waters, and P.C. Paquet. 2005. Traffic volume and highway permeability for a mammalian community in the Canadian Rocky Mountains. The Canadian Geographer / Le Géographe canadien 49:321–331.
Arctic Borderlands Ecological Knowledge Coop (ABEKC). 2004. Community reports, 2003-2004. Arctic Borderlands Ecological Society. Whitehorse, Yukon. 70 pp.
Arctic Borderlands Ecological Knowledge Coop (ABEKC). 2007. Community reports, 2005-2006. Arctic Borderlands Ecological Society. Whitehorse, Yukon. 40 pp.
Arctic Borderlands Ecological Knowledge Coop (ABEKC). 2008. Community reports, 2006-2007. Arctic Borderlands Ecological Society. Whitehorse, Yukon. 58 pp.
Aubry, K.B., and D.B. Houston. 1992. Distribution and status of the fisher (Martes pennanti) in Washington. Northwestern Naturalist 73:69–79.
Aubry, K.B., K.S. McKelvey, and J.P. Copeland. 2007. Distribution and broadscale habitat relations of the wolverine in the contiguous United States. Journal of Wildlife Management 71:2147-2158.
Aubry, K.B., J. Rohrer, C.M. Raley, R.D. Weir, and S. Fitkin. 2012. Wolverine distribution and ecology in the North Cascades ecosystem: 2012 annual report. U.S. Forest Service, Olympia, WA. 44 pp.
Austin, M. 1998. Wolverine Winter Travel Routes and Response to Transportation Corridors 1848 in Kicking Horse Pass between Yoho and Banff National Parks. M.E.Des. Thesis, 1849 University of Calgary, Calgary, Alta. 40 pp. + iv.
Autsӱl K’e Dene First Nation. 2002. Traditional knowledge in the Kache Tué study region. Phase three – towards a comprehensive environmental monitoring program in the Kakinӱne region. Final Report. West Kitikmeot Slave Study Society. Yellowknife, NWT. 89 pp.
Awan, M., pers. comm. 2012. Email correspondence to B. Slough, August 2012. Wildlife Biologist – Carnivores, Department of Environment, Igloolik, NU.
Awan, M., G. Szor, N. Lecomte, V.L. Hérault, and D. Berteaux. 2012. Wolverine harvest in Nunavut. Poster. Government of Nunavut, Department of Environment.
AXYS Environmental Consulting Ltd. (AXYS). 1998. Diavik Diamonds project: Environmental effects report, wildlife. Prepared for Diavik Diamonds Mines Inc. by AXYS Environmental Consulting Ltd. and Penner and Associates Ltd.
Banci, V. 1982. The Wolverine in British Columbia: distribution, methods of determining age and status of Gulo gulo vancouverensis. B.C. Ministries of Environment and Forests, Rep. No. IWIFR-15. Victoria, BC. 90 pp.
Banci, V. 1987. Ecology and Behaviour of Wolverine in Yukon. M.Sc. Thesis, Simon Fraser University, Burnaby, B.C. l78 pp.
Banci, V. 1994. Wolverine. pp. 99-127. in L.F. Ruggiero, K.B. Aubry, S.W. Buskirk, L.J. Lyon and W.J. Zielinski (eds.). The Scientific Basis for Conserving Forest Carnivores: American Marten, Fisher, Lynx and Wolverine in the Western United States. U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station, Ft. Collins, Colo. General Technical Report RM-254.
Banci, V., and A. Harestad. 1988. Reproduction and natality of wolverine (Gulo gulo) in Yukon. Annals Zoologic Fennici 25:265-270.
Banci, V., and A. Harestad. 1990. Home range and habitat use of Wolverines Gulo gulo in Yukon Canada. Holarctic Ecology 13:195-200.
Banci, V., C. Hanks, R. Spicker, and Kitikmeot Inuit Association. 2005. Inuit traditional knowledge of kalvik (wolverine) in Nunavut, Canada. Presentation at 1st International Symposium on Wolverine Research and Management. Swedish University of Agricultural Sciences, 13-15 June, 2005. Jokkmokk, Sweden.
Banci, V., and G. Proulx. 1999. Resiliency of furbearers to trapping in Canada. pp. 175-204. in G. Proulx (ed.). Mammal Trapping. Alpha Wildlife Research and Management Ltd., Sherwood Park, Alberta.
Becker, E.F. 1991. A terrestrial furbearer estimator based on probability sampling. Journal of Wildlife Management 55:730-737.
Becker, E.F., and C. Gardner. 1992. Wolf and wolverine density estimation techniques. Alaska Department of Fish and Game and Federal Aid in Wildlife Restoration, Research Progress Report, Grant W-23-5, Juneau, Alaska. 31 pp.
Becker, E.F., and H.N. Golden. 2008. Results of recent wolverine survey of GMU 14C. Alaska Department of Fish and Game, Anchorage, Alaska. 10 pp.
Beckman, K., pers. comm. 2012. Email correspondence to A. Magoun. Wildlife Veterinarian, Alaska Department of Fish and Game, Fairbanks, AK.
Benson, K. 2011. Gwich’in traditional knowledge: Woodland caribou boreal population. Gwich’in Social and Cultural Institute. Tsiigehtchic, Northwest Territories. 52 pp.
Berezanski, D. 2004. Status summary: Wolverine, Gulo gulo. Prepared for the Manitoba Endangered Species Advisory Committee. Manitoba Conservation, Fur Management Unit, Manitoba Wildlife and Ecosystem Protection Branch, Winnipeg, Man. 13 pp.
Berezanski, D., pers. comm. 2012. Email correspondence to B. Slough, June 2012. Furbearer and Problem Wildlife Management Unit, Wildlife and Ecosystem Protection Branch, Manitoba Conservation and Water Stewardship, Winnipeg, MB.
Boulanger, J., A. Gunn, J. Adamczewski, and B. Croft. 2011. A data-driven demographic model to explore the decline of the Bathurst caribou herd. The Journal of Wildlife Management 75:883-896.
Boulanger, J., and R. Mulders. 2008. Analysis of 2005 and 2006 Wolverine DNA mark-recapture sampling at Daring Lake, Ekati, Diavik, and Kennady Lake, Northwest Territories. Integrated Ecological Research, Nelson, B.C. 30 pp.
Boulanger, J., and R. Mulders. 2013a. Analysis of wolverine DNA mark-recapture sampling at Daring Lake, Diavik, and Ekati, Northwest Territories from 2005 to 2011. Integrated Ecological Research, Nelson, BC, and Environment and Natural Resources, Government of the Northwest Territories, Yellowknife, NT. 28 pp. [Draft 7 Feb. 2013].
Boulanger, J., and R. Mulders. 2013b. Analysis of wolverine DNA mark-recapture sampling at Daring Lake from 2004 to 2011. Integrated Ecological Research, Nelson, BC, and Environment and Natural Resources, Government of the Northwest Territories, Yellowknife, NT. 34 pp. [Draft 17 May 2013].
Bowman, Jeff., pers. comm. 2013. Email correspondence to B. Slough, July 2013. Research Scientist, Wildlife Research and Development Section, Ontario Ministry of Natural Resources, Peterborough, Ontario.
Bowman, J., J.C. Ray, A.J. Magoun, D.S. Johnson. 2010. Roads, logging, and the large-mammal community of an eastern Canadian boreal forest. Canadian Journal of Zoology 88:454-467.
Branigan, M., and J. Pongracz, unpubl. data. 2012. Wolverine carcass analysis report, Inuvik region. Wildlife Management, Environment and Natural Resources, Inuvik, NWT.
British Columbia Government. 2013. Species at Risk webpage. [www http://www.speciesatrisk.bc.ca/node/7754]
Brodie, J.F., and E. Post. 2010a. Non-linear responses of Wolverine populations to declining winter snowpack. Population Ecology 52:279-287.
Brodie, J.F., and E. Post. 2010b. Wolverines and declining snowpack: response to comments. Population Ecology 53:267-269.
Bulmer, M. 1975. Phase relations in the 10-year cycle. Journal of Animal Ecology 44:609-621.
Campbell, C. 2012. Caribou habitat protection: It’s urgent to reduce industry’s bootprint. Wild Lands Advocate 20 (2):6-9.
Canadian Forest Service. 2013. Forest Ecozones of Canada. http://cfs.nrcan.gc.ca/pages/125
Cardinal, N. 2004. Aboriginal traditional knowledge COSEWIC status report on wolverine Gulo gulo Qavvik. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ont. ix + 40 pp.
Carriere, N. 2010. Distribution Patterns and General Biology of Woodland Caribou based on Collection of Local and Traditional Knowledge in North-central Saskatchewan. Master’s Thesis, University of Saskatchewan, Saskatoon, Saskatchewan. 133 pp.
Carrière, S., pers. comm. 2013. Email correspondence to B. Slough, August 2013. Ecosystem Management Biologist, Biodiversity, Environment and Natural Resources, Yellowknife, NT.
Carroll, C., R.F. Noss, and P.C. Paquet. 2001. Carnivores as focal species for conservation planning in the Rocky Mountain region. Ecological Applications 11:961-980.
Cegelski C.C., L.P Waits, and J. Anderson. 2003. Assessing population structure and gene flow in Montana wolverines (Gulo gulo) using assignment-based approaches. Molecular Ecology 12:2907-2918.
Cegelski C.C., L.P. Waits, N.J. Anderson, O. Flagstad, C. Strobeck, and C.J. Kyle. 2006. Genetic diversity and population structure of wolverine (Gulo gulo) populations at the southern edge of their current distribution in North America with implications for genetic viability. Conservation Genetics 7:197-211.
Canadian Endangered Species Conservation Council (CESCC). 2011. Wild Species 2010: The General Status of Species in Canada. National General Status Working Group: 302 pp.
Chappell D.E., R.A. Van Den Bussche, J. Krizan, and B. Patterson. 2004. Contrasting levels of genetic differentiation among populations of wolverines (Gulo gulo) for northern Canada revealed by nuclear and mitochondrial loci. Conservation Genetics 5:759-767.
Clément, D. 2009. Direct-shipping ore project. Appendix D. Innu use of the territory and knowledge of its resources. Final Report. New Millenium Capital Corp. Calgary, Alta. 175 pp.
Clevenger, T. pers. comm. 2014. Email correspondence with Dwayne Lepitzki, April 2014. Senior Wildlife Research Scientist. Western Transportation Institute, University of Montana, Bozeman, Montana.
Clevenger, T. 2013. Mitigating highways for a ghost: Data collection challenges and implications for managing wolverines and transportation corridors. Northwest Science Notes 87:297-304.
Clevenger, AP, B Dorsey and N Heim. 2011a. Non-invasive genetic survey of wolverines and their response to the Banff-Yoho transportation corridor. Annual report: Year 1, 2010-11. Western Transportation Institute – Montana State University, Parks Canada, Miistakis Institute for the Rockies. 20 pp.
Clevenger, A.P., D. Duke, R. Haddock, and R. Ament. 2011b. Trans-Canada Highway Wildlife Monitoring and Research, Annual Report, Year 2 - 2010-2011. Prepared for Parks Canada, Radium Hot Springs, B.C. 30 pp.
Cluff, D., and P. Paquet. 2003. Large carnivores as umbrellas for reserve design and selection in the north. inDesigning Protected Areas: Wild Places for Wild Life. Proceedings Summary of the 2003 Canadian Council on Ecological Areas (CCEA) and Circumpolar Protected Areas Network (CPAN) Workshop, September 9-10, 2003, Yellowknife, N.W.T.
Community of Inuvik, Wildlife Management Advisory Council (NWT), and Joint Secretariat. 2008. Inuvik Inuvialuit community conservation plan. Joint Secretariat. Inuvik, NWT. 149 pp.
Community of Paulatuk, Wildlife Management Advisory Council (NWT), and Joint Secretariat. 2008. Paulatuk community conservation plan. Joint Secretariat. Inuvik, NWT. 142 pp.
Community of Sachs Harbour, Wildlife Management Advisory Council (NWT), and Joint Secretariat. 2008. Sachs Harbour community conservation plan. Joint Secretariat. Inuvik, NWT. 109 pp.
Community of Tuktoyaktuk, Wildlife Management Advisory Council (NWT), and Joint Secretariat. 2008. Tuktoyaktuk community conservation plan. Joint Secretariat. Inuvik, NWT. 169 pp.
ConocoPhillips Canada. 2006. Grizzly bear and wolverine protection plan for the Parsons Lake Field development: discussion draft. 19 pp. + appendices.
Copeland, J. 1996. Biology of the Wolverine in Central Idaho. M.Sc. Thesis, University of Idaho, Boise, Idaho. 138 pp.
Copeland, J. 2009. Investigating the relationship between winter recreation and wolverine spatial use in Central Idaho. Rocky Mountain Research Station, Missoula, Montana. Progress Report. 9 pp.
Copeland, J.P., K.S. McKelvey, K.B. Aubry, A. Landa, J. Persson, R.M. Inman, J. Krebs, E. Lofroth, H. Golden, J.R. Squires, A. Magoun, M.K. Schwartz, J. Wilmot, C.L. Copeland, R.E. Yates, I. Kojola, and R. May. 2010. The bioclimatic envelope of the wolverine (Gulo gulo): do climatic constraints limit its geographic distribution? Canadian Journal of Zoology 88:233-246.
Copeland, J.P., J.M. Peek, C.R. Groves, W.E. Melquist, K.S. McKelvey, G.W. McDaniel, C.D. Long, and C.E. Harris. 2007. Seasonal habitat associations of the wolverine in central Idaho. Journal of Wildlife Management 71:2201-2212.
COSEWIC. 2003. COSEWIC Assessment and Update Status Report on the Wolverine Gulo gulo in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 41 pp.
COSEWIC. 2008. COSEWIC Assessment and Update Status Report on the Polar Bear (Ursus maritimus) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 75 pp.
COSEWIC. 2012a. COSEWIC Assessment and Update Status Report on Grizzly Bear (Ursus arctos) in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ont. 85 pp.
COSEWIC. 2012b. Aboriginal traditional knowledge assessment report on Wolverine Gulo gulo in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ont. 50 pp.
Cowan, I. M., and C. Guiguet. 1960. The Mammals of British Columbia. British Columbia Provincial Museum, Victoria, BC. 413 p.
Crystal, D., pers. comm. 2014. Email correspondence with G. Forbes, February 2014. Wildlife Management Biologist. Nunavut Wildlife Management Board, Iqaluit, Nunavut.
Dagenais, J. 1988. Rapport sur la situation du carcajou (Gulo gulo) au Québec et dans les Maritimes. Internal report, Ministère du Loisir, de la Chasse et de la Pêche du Québec. 33 pp.
Dalerum, F., K. Kunkel, A. Angerbjörn, and B.S. Shults. 2009. Diet of wolverines (Gulo gulo) in the western Brooks Range, Alaska. Polar Research 28:246-253.
Dauphiné, C. 1989. Update COSEWIC status report on the Wolverine Gulo gulo in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ontario. 31 pp.
Dawson, N. 2000. Report on the status of Wolverine (Gulo gulo) in Ontario. Committee on the Status of Species at Risk in Ontario, Ontario Ministry of Natural Resources, Thunder Bay, Ont. 38pp. + iii.
Dawson, N., pers. comm. 2012. Email correspondence with B. Slough, May 2012. Wildlife Assessment Program Leader, Ministry of Natural Resources, Northwest Science and Information, Rosslyn, Ont.
Dawson, F.N., A.J. Magoun, J. Bowman, and J.C. Ray, unpubl. data. 2013. Ontario Ministry of Natural Resources, RR#1, Thunder Bay, Ontario, Wildlife Research and Management (WRAM), Fairbanks, Alaska, and Wildlife Conservation Society Canada, Toronto, Ontario.
Dawson, F.N., A.J. Magoun, J. Bowman, and J.C. Ray. 2010. Wolverine, Gulo gulo, home range size and denning habitat in lowland boreal forest in Ontario. Canadian Field-Naturalist 124:139-144.
DeVink, J.-M., D. Berezanski, and I. Devinet. 2010. Comments on Brodie and Post: Harvest effort: the missing covariate in analyses of furbearer harvest data. Population Ecology 53:261-262.
Diavik Diamond Mines Inc. (DDMI) 2012. Wildlife monitoring program report – 2011. Diavik Diamond Mine, Health, Safety, Environment and Training Department. 116 pp.
Dubey, J.P., V. Mason, L.T. Reichard, J.M. Garvon, N. Sundar, and M.E. Grigg. 2010. Two new species of Sarcocystis (Apicomplexa: Sarcocystidae) infecting the wolverine (Gulo gulo) from Nunavut, Canada. Journal of Parasitology 96:972–976.
Dumond, M. 2007. Western Kitikmeot caribou workshop. Government of Nunavut, Department of Environment, Final Wildlife Report: 19, Iqaluit, NU. 47 pp.
Ecological Stratification Working Group. 1995. A national ecological framework for Canada. Agriculture and Agri-Food Canada, Research Branch, Centre for Land and Biological Resource Research and Environment Canada, State of the Environment Directorate, Ecozone Analysis Branch, Ottawa/Hull. 125 pp. Map at 1:7,500,000.
Environment Canada. 2013. Meeting notes from Peary Caribou Recovery Strategy Development Community Technical Meeting - Public Meetings in Sachs Harbour – 05 March 2013. Prepared by Donna Bigelow, Canadian Wildlife Service, Yellowknife, NT. 11 pp.
Elliott, J., and M. Dumond. 2005. Harvested wolverine (Gulo gulo) age structure, sex ratio, body condition, and reproduction in the Kitikmeot Region (Nunavut) 1985-2004. [Working Draft]
Fisher, J.T., S.M. Bradbury, A.C. Fisher, and L. Nolan. 2009. Wolverines on the edge of Alberta's Rockies. Alberta Research Council, Edmonton, Alta. 80 pp.
Fisher, J.T., S. Bradbury, M. Wheatley, B. Anholt, L. Roy, J.P. Volpe, and L. Nolan. 2013. Wolverine (Gulo gulo luscus) on the Rocky Mountain slopes: natural heterogeneity and landscape alteration. Canadian Jounral of Zoology 91:706-716.
Fisher, J.T., and N. Heim. 2012. East Slopes predators: mammalian spatial distribution and habitat selection in a heterogeneous mountain landscape, year-end report 2011-2012. East Slopes Predators Project. Alberta Innovates - Technology Futures. Vegreville, Alberta. 24 pp.
Flagstad, O., E. Hedmark, A. Landa, H. Brøseth, J. Persson, R. Andersen, P. Segertröm, and H. Ellegren. 2004. Colonization history and non-invasive monitoring of a re-established Wolverine population. Conservation Biology 18:676-688.
Fortin, C. 2004. Inventaire de la faune en période hivernale dans le parc national projeté des Mont-Torngat-et-de-la-Rivière-Koroc (Nunavik). Rapport préparé pour l’Administration Régionale Kativik. 18 pp.
Fortin, C. 2006. Inventaire aérien du carcajou dans les basses-terres de l’Abitibi et de la baie James à l’hiver 2006. 11 pp.
Fortin, C., V. Banci, J. Brazil, M. Crête, J. Huot, M. Huot, R. Lafond, P. Paré, J. Shaefer, and D. Vandal. 2005. National Recovery Plan for the Wolverine (Gulo gulo) [Eastern Population]. National Recovery Plan No. 26. Recovery of Nationally Endangered Wildlife (RENEW). Ottawa, Ontario. 33 pp.
Foster, J.L., D.A. Robinson, D.K. Hall, and T.W. Estilow. 2008. Spring snow melt timing and changes over Arctic lands. Polar Geography 31:145-157.
Fraser, D., pers. comm. 2013. Email correspondence to B. Slough, July 2013. Scientific Authority Assessment, A/Manager BC CDC, Ecosystem Branch, Conservation Planning Section, Ministry of Environment, Victoria, BC.
Gallais, S., and F. Messier. 2012. Projet carcaj’ou ? : Habitat potentiel du carcajou (Gulo gulo) au Québec. Rapport présenté à la Fondation de la faune de Québec. Nature Québec. 45 pp.
Gardner, C.L. 1985. The Ecology of Wolverines in Southcentral Alaska. M.Sc. Thesis. University of Alaska, Fairbanks, Alaska. 82 pp.
Gardner, C.L., and W.B. Ballard. 1982. Susitna hydro electric project. Phase I final report. Vol. VII: wolverine. Alaska Department of Fish and Game, Anchorage, AK. 43 pp.
Gardner, C.L., W.B. Ballard and R.H. Jessup. 1986. Long distance movement by an adult wolverine. Journal of Mammalogy 67:603.
Gibeau, M.L., and K. Heuer. 1996. Effects of transportation corridors on large carnivores in the Bow River Valley. inG.L. Evink, P. Garrett, D. Zeigler, and J. Berry (eds). Trends in Addressing Transportation-Related Wildlife Mortality. State of Florida Department of Transportation, Environmental Management Office, Tallahassee, Florida. FL-ER-58-96.
Golden, H.N. 2010. Wolverine abundance in upper Turnagain Arm and the Kenai Mountains with emphasis on helicopter-skiing permit areas. Alaska Department of Fish and Game and Chugach National Forest. Interagency Collaborative Project Progress Report, Anchorage, Alaska. 9 pp.
Golden, H.N., unpubl. data 2013. Wildlife Research Biologist, Alaska Department of Fish and Game, Division of Wildlife Conservation, Anchorage, Alaska.
Golden, H.N., J.D. Henry, E.F. Becker, M.I. Goldstein, J.M. Morton, D. Frost, Sr., and A.J. Poe. 2007. Estimating wolverine Gulo gulo population size using quadrat sampling of tracks in snow. Wildlife Biology 13 (Suppl. 2):52-61.
Golder Associates. 2003. Report on Inuit qaujimajatuqangit literature review, gap analysis and workshop results related to the Doris North project, Hope Bay Belt, Nunavut. Report prepared for Miramar Hope Bay Ltd., North Vancouver, BC. 214 pp.
Golder Associates. 2010. Snap Lake Mine: Environmental agreement, 2009 Annual Report. Prepared for De Beers Canada Inc. by Golder Associates, Yellowknife N.W.T. iv + 90 pp.
Golder Associates. 2012a. De Beers Snap Lake Mine wildlife effects monitoring program. Prepared for De Beers Canada Inc. by Golder Associates, Yellowknife N.W.T. viii + 37 pp.
Golder Associates. 2012b. Gahcho Kué Project: 2011 wildlife supplemental monitoring project. Prepared for De Beers Canada Inc. by Golder Associates, Yellowknife, N.W.T. ii + 23 pp.
Gollop, M. 2012. Annual status of furbearers survey and results of previous trapper surveys. Fish and Wildlife Branch, Saskatchewan Ministry of Environment, Saskatoon, SK. 2 pp.
Government of Nunavut. 2007. Statutory report on wildlife to the Nunavut Legislative Assembly, Section 176 of the Wildlife Act. vii + 125 pp.
Gunn, F.E. 2009. Traditional Ecological Knowledge of Boreal Woodland Caribou in Western Wood Buffalo National Park. M.A. dissertation, Royal Roads University, Victoria, British Columbia. Publication No. AAT MR55878. 177 pp.
Gwich’in Renewable Resource Board (GRRB). 2001. More Gwich’in words about the land. Gwich’in Renewable Resource Board. Inuvik, Northwest Territories. 184 pp.
Gwich’in Renewable Resource Board (GRRB). 2014. Gwich’in Traditional Knowledge: Nèhtrùh (Wolverine). Gwich’in Renewable Resource Board. Inuvik, Northwest Territories. 56 pp.
Hall, E.R. 1981. Wolverine. pp. 1006-1009. in The Mammals of North America, Volume II, 2nd Edition. John Wiley and Sons, New York, N.Y.
Han, S., pers. comm. 2012. Email correspondence to B. Slough, June 2012. Head, Canadian Wildlife Service, Environment Canada, Eastern Arctic Unit, Iqaluit, NU.
Hatler, D.F. 1989. A wolverine management strategy for British Columbia. Wildlife Bulletin No. B-60, Ministry of Environment, Victoria, B.C. 124 pp.
Heinemeyer, K., and J. Squires. 2012. Idaho wolverine – winter recreation research project: Investigating the interactions between wolverines and winter recreation. 2011-2012 Progress Report. Round River Conservation Studies and the USFS Rocky Mountain Research Station. iv + 20 pp.
Henry, J.D. 2004. Track transects for monitoring changes in wolverine and other mustelid populations in the Old Crow Flats: Third annual report (winter 2003-2004). Parks Canada, Haines Junction, Yukon. 36 pp.
Henry, J.D. 2007. Monitoring predator populations in the Old Crow Flats, 2001-2006. Parks Canada, Old Crow, Yukon. 132 pp.
Heydon, C. pers. comm. 2002. Email correspondence to B. Slough, September 2002. Fur Program Biologist, Fish and Wildlife Branch, Ontario Ministry of Natural Resources, Peterborough, Ont.
Holbrow, W.C. 1976. The Biology, Mythology, Distribution and Management of the Wolverine in Western Canada. M.N.R.M. Practicum, Natural Resources Institute, University of Manitoba, Winnipeg, Man. 214 pp.
Hornocker, M.G. and H.S. Hash. 1981. Ecology of the wolverine in northwestern Montana. Canadian Journal of Zoology 59:1286-1301.
Inman, R.M., K.H. Inman, M.L. Packila, and A.J. McCue. 2007. Chapter 4: Wolverine reproductive rates and maternal habitat in Greater Yellowstone. in Wildlife Conservation Society (2007). Greater Yellowstone Wolverine program: cumulative report, May 2007. Ennis, Montana.
Inman, R.M., A.J. Magoun, J. Persson, and J. Mattisson. 2012. The wolverine’s niche: linking reproductive chronology, caching, competition and climate. Journal of Mammalogy 93:634-644.
Inman, R. M., M.L. Packila, K.H. Inman, B. Aber, R. Spence, and D. McCauley. 2009. Greater Yellowstone Wolverine Program, Progress Report – December 2009. Wildlife Conservation Society, North America Program, General Report, Bozeman, Montana. v + 32 pp.
Inman, R.M., R.R. Wigglesworth, K.H. Inman, M.K. Schwartz, B.L. Brock, and J.D. Reick. 2004. Wolverine makes extensive movements in Greater Yellowstone area. Northwest Science 78:261-266.
Inuvik Community Corporation (ICC), Tuktuuyaqtuuq Community Corporation, and Aklavik Community Corporation. 2006.Inuvialuit Settlement Region traditional knowledge report. Mackenzie Project Environmental Group. Calgary, Alberta. 200 pp.
IPCC. 2007. Climate Change 2007: The physical science basis. in Contribution of Working Group I to the Fourth Assessment. Report of the Intergovernmental Panel on Climate Change. Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. Tignor and H.L. Miller (eds.). Cambridge University Press, Cambridge, U.K. and New York, N.Y. 996 pp.
IUCN 2012. IUCN red list of threatened species. Version 2012.2. Web site: www.iucnredlist.org. [accessed January 2013].
Johnson, C.J., M.S. Boyce, R.L. Case, H.D. Cluff, R.J. Gau, A. Gunn, and R. Mulders. 2005. Cumulative effects on human developments on Arctic wildlife. Wildlife Monographs 160:1-36.
Johnson, C.S. 1990. Re-evaluation of the status of the wolverine in Manitoba. Wildlife Biological Services Technical Report No. 90-01, Manitoba Natural Resources, Winnipeg, Man. 25 pp.
Johnson, M. and R. Ruttan. 1993. Traditional Dene Environmental Knowledge: A Pilot Project conducted in Fort Good Hope and Colville Lake, NT 1989-1993. Dene Cultural Institute, Hay River, NT. 20 pp.
Johnson, L.M., L.L. Willey, M.T. Jones, and F. Sangermano. 2012. Predicting core reproductive habitat for wolverine (Gulo gulo) in Québec, Canada using a GIS-based deductive modeling approach. Prepared for Ministère des Ressources Naturelles et de la Faune et Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs, Province du Québec. 28 pp.
Joint Secretariat – Inuvialuit Settlement Region web site: http://www.jointsecretariat.ca/documents.html
Jung, T.S., pers. comm. 2013. Senior Wildlife Biologist, Biodiversity Programs, Fish and Wildlife Branch, Environment Yukon, Whitehorse, Yukon.
Jung, T.S., and P. Kukka. 2013. Wolverine carcass collection program: 2013 progress report. Yukon Fish and Wildlife Branch Report PR-13-04. Whitehorse, Yukon. 39 pp.
Jung, T. S., B. G. Slough, B. L. Smith, H. Slama, and R. H. Jessup. 2005. Using trapper knowledge to monitor population trends of wolverine, Gulo gulo, in the Yukon. Poster presented at 1st International Symposium on Wolverine Research and Management, June 13-15, 2005, Jokkmokk, Sweden.
Keeyask Hydro Limited Partnership (HLP). 2012. Keeyask generation project environmental impact statement. Volume 7: Terrestrial environment. Keeyask HLP, Winnipeg, Manitoba. 302 pp.
Kelsall, J.P. 1981. Status report on the wolverine, Gulo gulo, in Canada in 1981. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 50 pp.
Koback, L. 2012. Saskatchewan wild fur harvest and cash values 2010-11. Fish and Wildlife Branch Summary Report. Saskatchewan Ministry of Environment, Saskatoon, Sask. 12 pp. + append.
Koen, E.L., J.C. Ray, J. Bowman, F.N. Dawson, and A.J. Magoun. 2008. Surveying and monitoring wolverines in Ontario and other lowland, boreal forest habitats: Recommendations and protocols. Ontario Ministry of Natural Resources, Northwest Science and Information. Thunder Bay, Ont. NWSI Field Guide FG-06. 94 pp. + append.
Krebs, J.A., pers. comm. 2013. Email correspondence to G. Forbes. Senior Biologist, Fish and Wildlife Compensation Program, BC Hydro, Nelson, BC.
Krebs, J.A., and D. Lewis. 2000. Wolverine ecology and habitat use in the North Columbia Mountains: Progress Report. pp. 695-703. in L.M. Darling (ed.). Proceedings of a Conference on the Biology and Management of Species and Habitats at Risk, Kamloops, B.C., 15-19 Feb., 1999. Volume Two. BC. Ministry of Environment, Lands and Parks, Victoria, B.C. and University College of the Cariboo, Kamloops, B.C.
Krebs, J., E. Lofroth, J. Copeland, V. Banci, D. Cooley, H. Golden, A. Magoun, R. Mulders and B. Shults. 2004. Synthesis of survival rates and causes of mortality in North American wolverines. Journal of Wildlife Management 68:493-502.
Krebs, J., E.C. Lofroth, and I. Parfitt. 2007. Multiscale habitat use by wolverines in British Columbia, Canada. Journal of Wildlife Management 71:2180-2192.
Krott, P., and C. Gardner. 1985. Wolverine breeding behavior in Alaska. Säugetierkundliche Mitteilungen 32:87.
Kurtén, B., and R. Rausch. 1959. Biometric comparisons between North American and European mammals. I. A comparison between Alaskan and Fennoscandian Wolverine (Gulo gulo Linnaeus). Acto Arctica 11:1-21.
Kyle, C. March 2014. Email correspondence to Graham Forbes. Department of Biology, Trent University, Peterborough, ON.
Kyle, C.J., and C. Strobek. 2001. Genetic structure of North American wolverine (Gulo gulo) populations. Molecular Ecology 10:337-347.
Kyle, C.J., and C. Strobek. 2002. Connectivity of peripheral and core populations of North American wolverines. Journal of Mammalogy 83:1141-1150.
Landa, A., O. Strand, J.D.C. Linnell, and T. Skogland. 1998. Home-range sizes and altitude selection for arctic foxes and wolverines in an alpine environment. Canadian Journal of Zoology 76:448-457.
Landa, A., O. Strand, J.E. Swenson, and T. Skogland. 1997. Wolverines and their prey in southern Norway. Canadian Journal of Zoology 75:1292-1299.
Larter, N. and D. Allaire. 2013. MacKenzie Mountain non-resident and non-resident alien hunter harvest summary 2012. Environment and Natural Resources. Government of NWT Manuscript Report # 234. 88 pp.
Lecomte, N., pers. comm. 2013. Email correspondence with G. Forbes, July 2013. Ecosystem Biologist, Department of Environment, Igloolik, NU.
Lee, J. 1994a. Wolverine harvest and carcass collection, Coppermine, Bay Chimo and Bathurst Inlet, 1992/93. Wildlife and Fisheries Division, Department of Resources, Wildlife, and Economic Development, Yellowknife, NT. 15 pp.
Lee, J. 1994b. Wolverine harvest and carcass collection, Coppermine, Bay Chimo and Bathurst Inlet, 1993/94. Wildlife and Fisheries Division, Department of Resources, Wildlife, and Economic Development, Yellowknife, NT. 14 pp.
Lee, J. 1998. Database description and partial data summary and analysis of the wolverine harvest from Kugluktuk, Umingmaktok and Bathurst Inlet, Northwest Territories, 1985/86 to 1996/97. Department of Resources, Wildlife and Economic Development, Yellowknife, NWT. 42 pp. [Draft].
Lee, J. and A. Niptanatiak. 1996. Observation of repeated use of a wolverine, Gulo gulo, den on the tundra of the Northwest Territories. Canadian Field-Naturalist 110:349-350.
Lofroth, E. 2001. Wolverine ecology in plateau and foothill landscapes, 1996-2001. 2000/01 Year-end report, northern Wolverine project. Forest Renewal Activity No. 712260. Ministry of Environment, Lands and Parks, Victoria, B.C. 98 pp.
Lofroth, E., pers. comm. 2012. Email correspondence to B. Slough, October 2012. Manager, British Columbia Conservation Data Centre, BC Ministry of Environment, Victoria, B.C.
Lofroth, E.C., and J. Krebs. 2007. The abundance and distribution of wolverines in British Columbia, Canada. Journal of Wildlife Management 71:2159-2169.
Lofroth, E.C., J.A. Krebs, W.L. Harrower, and D. Lewis. 2007. Food habits of wolverine (Gulo gulo) in montane ecosystems of British Columbia, Canada. Wildlife Biology 13 (Suppl. 2):31-37.
Lofroth, E.C., and P.K. Ott. 2007. Assessment of the sustainability of wolverine harvest in British Columbia., Canada. Journal of Wildlife Management 71:2193-2200.
Lutsël K’e Dene First Nation, B. Parlee, M. Basil, and N. Casaway. 2001. Traditional ecological knowledge in the Kaché Tué Study Region. Final Report. West Kitikmeot Slave Study Society. Yellowknife, NWT. 88 pp.
MacLeod, C.F. 1950. The Productivity and Distribution of Fur-bearing Species of the Coast of British Columbia in Relation to Some Environmental Factors. M.A. Thesis, University of British Columbia, Vancouver, B.C. 105 pp.
MacDonald, P., pers. comm. 2013. E-mail correspondence to G. Forbes, July 8, 2013. Wildlife Biologist, Canadian Wildlife Service, Atlantic Region, Goose Bay, NL.
Magoun, A.J. 1985. Population Characteristics, Ecology and Management of Wolverines in Northwestern Alaska. Ph.D. dissertation, University of Alaska, Fairbanks, Alaska. 197 pp.
Magoun, A.J. 1987. Summer and winter diets of wolverines, Gulo gulo, in Arctic Alaska. Canadian Field-Naturalist 101:392-397.
Magoun, A.J., K.F. Abraham, J.E. Thompson, J.C. Ray, M.E. Gauthier, G.E. Brown, G. Woolmer, C.J. Chenier, and F. N. Dawson. 2005a. Distribution and relative abundance of caribou in the Hudson Plains Ecozone of Ontario. Rangifer special issue 16:105-121.
Magoun, A.J., and J.P. Copeland. 1998. Characteristics of wolverine reproductive den sites. Journal of Wildlife Management 62:1313-1320.
Magoun A., N. Dawson, J. Ray, and J. Bowman. 2005b. Forest management considerations for wolverine populations in areas of timber harvest in Ontario: preliminary recommendations. Ontario Boreal Wolverine Project, The Wolverine Foundation, Ontario Ministry of Natural Resources. 17 pp.
Magoun, A., N. Dawson, J. Ray, J. Bowman, C. Lipsett-Moore, and G. Lipsett-Moore. 2004. Boreal wolverine: a focal species for land use planning in Ontario's northern boreal forest. Ontario Wolverine Project Report. 27 pp.
Magoun, A.J., C.D. Long, M.K. Schwartz, K.L. Pilgrim, R.E. Lowell, and P. Valkenburg. 2011a. Integrating motion-detection cameras and hair snags for wolverine detection. Journal of Wildlife Management 75:731-739.
Magoun, A.J., J.C. Ray, D.S. Johnson, P. Valkenburg, F.N. Dawson, and J. Bowman. 2007. Modeling wolverine occurrence using aerial surveys of tracks in snow. Journal of Wildlife Management 71:2221-2229.
Magoun, A.J., and P. Valkenburg. 1983. Breeding behavior of free-ranging wolverines (Gulo gulo). Acta Zoologica Fennici. 174:175-177.
Magoun, A.J., P. Valkenburg, C.D. Long, and J.K. Long. 2013. Monitoring wolverines in northeast Oregon, January 2011-December 2012. Final Report. The Wolverine Foundation, Inc., Kuna, Idaho, USA.
Magoun, A.J., P. Valkenburg, D.N. Pedersen, C.D. Long, and R.E. Lowell. 2011b. Wolverine images - using motion-detection cameras for photographing, identifying, and monitoring Wolverines. v + 155 pp. Web site: http://www.blurb.com/bookstore. [Accessed 14 January 2013].
Mallory, M.L., J.A. Akearok, and A.J. Fontaine. 2001. Community knowledge on the distribution and abundance of species at risk in southern Baffin Island, Nunavut, Canada. Canadian Wildlife Technical Report Series No. 363. Canadian Wildlife Service, Prairie and Northern Region. Iqaluit, Nunavut. 68 pp.
May, R., L. Gorini, J. van Diijk, H. Brøseth, J.D.C. Linnell, and A. Landa. 2012. Habitat characteristics associated with wolverine den sites in Norwegian multiple-use landscapes. Journal of Zoology 287:195-204.
May, R., A. Landa, J. van Dijk, J.D.C. Linnell, and R. Andersen. 2006. Impact of infrastructure on habitat selection of wolverines Gulo gulo. Wildlife Biology 12:285-295.
McKelvey, K.S., K.B. Aubry, and M.K. Schwarz. 2008. Using anecdotal occurrence data for rare or elusive species: the illusion of reality and a call for evidentiary standards. BioScience 58:549-555.
McKelvey, K.S., J.P. Copeland, M.K. Schwartz, J.S. Littell, K.B. Aubry, J.R. Squires, S.A. Parks, M.A. Elsner, and G.S. Guillaume. 2011. Climate change predicted to shift wolverine distributions, connectivity, and dispersal corridors. Ecological Applications 21: 2882-2897.
McKelvey, K. S., E.C. Lofroth, J.P. Copeland, K.B. Aubry, and A.J. Magoun. 2010. Comments on Brodie and Post: Climate-driven declines in wolverine populations: causal connection or spurious correlation? Population Ecology. doi:10.1007/s10144-010-0242-5.
McKelvey, K.S. (and 10 co-authors). 2013. Recovery of wolverines in the western United States: Recent extirpation and recolonization, or range retraction and expansion? Journal of Wildlife Management. DOI:10.1002/jwm.649.
Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs du Québec. 2013. Liste des espèces fauniques menaces ou vulnérables au Québec: Carcajou. Web site: http://www.mddefp.gouv.qc.ca/faune/especes/menacees/fiche.asp?noEsp=4. [accessed July 2013].
Moores, S. June 2012. Email correspondence to Brian Slough. Senior Manager, Endangered Species and Biodiversity, Wildlife Division, Department of Environment and Conservation, Corner Brook, NL.
Moriarty, K.M., W.J. Zielinski, A.G. Gonzales, T.E Dawson, K.M. Boatner, C.A. Wilson, F.V. Schlexer, K.L. Pilgrim, J.P. Copeland, and M.K. Schwartz. 2009. Wolverine confirmation in California after nearly a century: Native or long-distance immigrant? Northwest Science 83:154-162.
Mulders, R. 2000. Wolverine ecology, distribution and productivity in the Slave Geological Province. Final Report to the West Kitikmeot/Slave Study Society. Dept. of Resources, Wildlife and Economic Development, Government of the Northwest Territories, Yellowknife, N.W.T. 92 pp.
Mulders, Robert. August 2013. Email correspondence with Brian Slough. Wildlife Biologist, Carnivores/Furbearers, Wildlife Division, Environment and Natural Resources, Yellowknife, NWT.
Mulders, R., unpubl. data. 2013. Wolverine carcass analysis report, Kitikmeot, Sahtu, Dehcho, South Slave and North Slave regions, and Border A/B expert of wolverine from South Slave to Saskatchewan. Wildlife Biologist, Carnivores/Furbearers, Wildlife Division, Environment and Natural Resources, Yellowknife, NWT.
Mulders, R., J. Boulanger, and D. Paetkau. 2007. Estimation of population size for wolverines at Daring Lake, Northwest Territories, using DNA based mark-recapture methods. Wildlife Biology 13 (Suppl. 2):38-51.
Myrberget, S. 1968. Jervens ynglehi [The breeding den of the wolverine, Gulo gulo]. Fauna (Oslo) 21:108-115.
Nagy, J. A., T. Creighton, T. Slack and W. Wright. 2002. Local knowledge about boreal woodland caribou in the Inuvialuit Settlement Region. Unpublished report prepared for the Department of Resources, Wildlife, and Economic Development, GNWT, Inuvik Region, Inuvik, Northwest Territories. 34 pp.
Nantel, P., pers. comm. 2012. E-mail correspondence to B. Slough, June 2012. Science Advisor, Parks Canada, Gatineau, QC.
NatureServe. 2013. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. Available http://www.natureserve.org/explorer. [accessed: July 9, 2013].
Nernberg, D., pers. comm. 2012. Email correspondence to B. Slough, August 2012. Species at Risk Officer, Director General of Environment, Directorate of Environmental Stewardardship, National Defence, Ottawa, ON.
Newfoundland and Labrador Department of Environment and Conservation. 2012. George River caribou herd continues to decline. Press Release. Web Site: http://www.releases.gov.nl.ca/releases/2012/env/0816n03.htm. [accessed January 2013].
North Slave Metis Alliance (NSMA). 1999. Can't live without work. North Slave Metis Alliance environmental, social, economic and cultural concerns: a companion to the Comprehensive Study Report on the Diavik Diamond Project. Yellowknife, NWT. 300 pp.
Northwest Territories Species at Risk Committee. 2013. Draft species status report (scientific knowledge component) for wolverine [Gulo gulo] in the Northwest Territories. Environment and Natural Resources, Yellowknife, NWT. [Draft, 26 April 2013].
Novak, M., M.E. Obbard, J.G. Jones, R. Newman, A. Booth, A.J. Sattherwaite, and G. Linscombe. 1987. Furbearer harvests in North America, 1600-1984. Ontario Ministry of Natural Resources, Toronto, Ontario and Ontario Trappers Association, North Bay, ON. 270 pp.
Nunavut Wildlife Management Board (NWMB). 2004. The Nunavut wildlife harvest study. Nunavut Wildlife Management Board. Iqaluit, NU. 816 pp.
Obbard, M.E., J.G. Jones, R. Newman, A. Booth, A.J. Sattherwaite, and G. Linscombe. 1987. Furbearer harvests in North America. pp. 1007-1034. in M. Novak, J.A. Baker, M.E. Obbard and B. Malloch (eds). Wild Furbearer Management and Conservation in North America. Ontario Trappers Association, North Bay, ON.
OMNR. 2010. Forest management guide for conserving biodiversity at the stand and site scales. Ontario Ministry of Natural Resources. Queen’s Printer for Ontario, Toronto, ON. 211 pp.
Ontario Wolverine Recovery Team. 2013. Recovery Strategy for the Wolverine (Gulo gulo) in Ontario. Ontario Recovery Strategy Series. Prepared for the Ontario Ministry of Natural Resources, Peterborough, ON. vi + 66 pp.
Paul F. Wilkinson and Associates Inc. on behalf of the Naskapi Nation of Kawawachikamach. 2009. Wolverine workshop. 26 and 27 March, 2009. Activity Report. Paul F. Wilkinson and Associates Inc. Montreal, QC. 32 pp.
Persson, J., P. Wedholm, and P. Segerström. 2010. Space use and territoriality of wolverines (Gulo gulo) in northern Scandinavia. European Journal of Wildlife Research 56:49-57.
Petersen, S. 1997. Status of the wolverine (Gulo gulo) in Alberta. Alberta Environmental Protection, Wildlife Management Division, Wildlife Status Report No. 2, Edmonton, Alta. l7 pp.
Poole, K., unpubl. data. 2013. Aurora Wildlife Research, Nelson, BC.
Pulliainen, E. 1968. Breeding biology of the wolverine (Gulo gulo L.) in Finland. Annales Zoologici Fennici 5:338-344.
Quick, H.F. 1953. Wolverine, fisher and marten studies in a wilderness region. Transactions of the North American Wildlife Conference 18:513-532.
Rausch, R.A., and A.M. Pearson. 1972. Notes on the wolverine in Alaska and the Yukon Territory. Journal of Wildlife Management 36:249-268.
Rauset, G.R. 2013. Life and Death in Wolverines: Linking Demography and Habitat for Conservation. Doctoral Thesis. Swedish University of Agricultural Sciences, Uppsala, Sweden. 41 pp.
Ray, J. 2004. Collecting ATK on wolverines in northern Ontario. pp. 94-98 in N. Cardinal. Aboriginal Traditional Knowledge and the COSEWIC Species Assessment Process: a Case Study of Northern Canada Wolverines, Environment Canada, Ottawa, ON.
Ray, J.C. 2012. Caribou recruitment and baseline surveys in the Ring of Fire: informing range assessment and recovery. Final Report to Ontario Ministry of Natural Resources Species at Risk Stewardship Fund 2011-2 (#159-11-WCSC). Wildlife Conservation Society Canada. 19 pp.
Ray, J.C., pers. comm. 2013. Email correspondence to G. Forbes, May 2013. Executive Director/Senior Scientist, Wildlife Conservation Society Canada, Toronto, ON.
Ray, J.C., A.J. Magoun, N. Dawson, and J. Bowman. 2005. Ontario Wolverine Project – Fieldwork 1 Jan - 31 March 2005. Progress report to Ontario Ministry of Natural Resources, Species-at-Risk Section. Ontario Wolverine Project Report. 5 pp.
Reichard, M.V., L. Toretti, T.A. Snider, J.M. Garvon, G., G. Marucci, and E. Pozio. 2008a. Trichinella T6 and Trichinella nativa in wolverines (Gulo gulo) from Nunavut, Canada. Parasitology Research 103:657–661.
Reichard, M.V., L. Toretti, J.M. Garvon, and J.P. Dubey. 2008b. Prevalence of antibodies to Toxoplasma gondii in wolverines from Nunavut, Canada. Journal of Parasitology 94:764–765.
Rescan. 2012. EKATI Diamond Mine: 2011 Wildlife Effects Monitoring Program. Prepared for BHP Billiton Canada Inc. by Rescan Environmental Services Ltd., Yellowknife, NWT. 158 pp.
Rossouw, F., unpubl. data 2012. Fur harvest data. Traditional Economy and Fur Management, Industry, Tourism, and Investment, Yellowknife, NWT.
Rowland, M.M., M.J. Wisdom, D.H. Johnson, B.C. Wales, J.P. Copeland, and F.B. Edelmann. 2003. Evaluation of landscape models for wolverines in the Interior Northwest, United States of America. Journal of Mammalogy 84:92-105.
Royle, J.A., A.J. Magoun, B. Gardner, P. Valkenburg, and R.E. Lowell. 2011. Density estimation in a wolverine population using spatial capture-recapture models. Journal of Wildlife Management 75:604-611.
Schmelzer, I. 2006. Occurrence and distribution of wolverines in northern Labrador: an aerial survey to clarify status and focus recovery. Department of Environment and Conservation, Corner Brook, NL. vii + 32 pp.
Schmelzer, I., unpubl. data 2012. Occurrence and distribution of wolverines in Labrador. Senior Wildlife Biologist, Terrestrial, Research, Wildlife Division, Department of Environment and Conservation, Corner Brook, NL.
Schwartz M.K., K.B. Aubry, K.S. McKelvey, K.L. Pilgrim, J.P. Copeland, J.R. Squires, R.M. Inman, S.M. Wisely, and L.F. Ruggiero. 2007. Inferring geographic isolation of wolverines in California using historical DNA. Journal of Wildlife Management 71:2170-2179.
Schwartz M.K., J.P. Copeland, N.J. Anderson, J.R. Squires, R.M. Inman, K.S. McKelvey, K.L. Pilgrim, L.P. Waits, and S.A. Cushman. 2009. Wolverine gene flow across a narrow climatic niche. Ecology 90:3222-3232.
Shardlow, T. 2013. Isotopic evidence of Salmon, Oncorhynchus spp., in the diet of wolverine, Gulo gulo, on Princess Royal Island, British Columbia. Canadian Field-Naturalist 127:338-342.
Shuswap Indian Band. 2008. Re Tsqwátstens-kucw ne Csaliken’, our people between the two mountain ranges. Shuswap Indian Band Traditional Land Use Study. Shuswap Indian Band. Kamloops, BC. 224 pp.
Siemens Worsley, A. 2011. Status assessment: Wolverine (Gulo gulo luscus) in Saskatchewan. Saskatchewan Conservation Data Centre, Fish and Wildlife Branch, Regina, Sask. iii + 12 pp.
Slough, B.G. 2007. Status of the wolverine Gulo gulo in Canada. Wildlife Biology 13 (Suppl. 2):76-82.
Slough, B.G. 2009. A Yukon Trappers’ Perspective on Wildlife Populations: A Preliminary Analysis of Trapper Questionnaires, 1992-93 to 2007-08. Fish and Wildlife Branch, Yukon Department of Environment, Whitehorse, Y.T. v + 79 pp.
Slough, B.G., unpubl. data. Independent Wildlife Biologist, Whitehorse, YU.
Slough, B.G., and R.H. Jessup. 1996. Furbearer trapping in the Yukon, Canada. pp.30-36. in R. Prescott-Allen and Prescott-Allen (eds). Assessing the Sustainability of Uses of Wild Species: Case Studies and Initial Assessment Procedure. Occasional paper of the IUCN Species Survival Commission No. 12. IUCN, Gland, Switzerland.
Slough, B.G., R.H. Jessup, D.I. McKay, and A.B. Stephenson. 1987. Wild furbearer management in western and northern Canada. pp. 1062-1076. in M. Novak, J.A. Baker, M.E. Obbard and B. Malloch (eds). Wild Furbearer Management and Conservation in North America. Ontario Trappers Association, North Bay, ON.
Spencer, S., unpubl. data. 2012. Wolverine Harvest Study Data 1996-2001. Terrestrial Wildlife Management Biologist, Nunavut Wildlife Management Board, Iqaluit, NU.
Squires, John R., J.P. Copeland, T.J. Ulizio, M.K. Schwartz, and L.F. Ruggiero. 2007. Sources and patterns of wolverine mortality in western Montana. Journal of Wildlife Management 71: 2213-2220.
Stewart, I.T. 2009. Changes in snowpack and snowmelt runoff for key mountain regions. Hydrological Processes 23:78–94.
Thibault, I., pers. comm. 2012. Email correspondence to B. Slough, December 2012. Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs du Québec, Direction générale de l’expertise sur la faune et ses habitats, Québec, QC.
Thibault, I., unpubl. data. 2013. Ministère du Développement durable, de l’Environnement, de la Faune et des Parcs du Québec, Direction générale de l’expertise sur la faune et ses habitats, Québec, QC.
Thompson, I., pers. comm. 2013. Email correspondance to G. Forbes, May 2013. Research Scientist, Ecology and Biodiversity, Great Lakes Forestry Centre, Sault Ste. Marie, ON.
Thorpe, N., Hakongak, N., Eyegetok, S., and Qitirmiut Elders. 2001. Tuktu and Nogak Project. A Caribou Chronicle. West Kitikmeot Slave Study Society. Yellowknife, Northwest Territories. 198 pp.
Tomasik, E., and J.A. Cook. 2005. Mitochondrial phylogeography and conservation genetics of wolverine (Gulo gulo) of northwestern North America. Journal of Mammalogy 86:386-396.
U.S. Fish and Wildlife Service. 2013. Endangered and threatened wildlife and plants; threatened status for the distinct population segment of the North American Wolverine occurring in the contiguous United States. Federal Register Vol. 78, No. 23:7864-7890.
Vors, L.,and M. Boyce. 2009. Global declines of caribou and reindeer. Global Change Biology 15: 2626–2633.
Vangen, K.M., J. Persson, A. Landa, R. Anderson, and P. Segerström. 2001. Characteristics of dispersal in wolverines. Canadian Journal of Zoology 79:1641-1649.
van Dijk, J., L. Gustavsen, A. Mysterud, R. May, Ø. Flagstad, H. Brøseth, Roy Andersen, Reider Andersen, H. Steen, and A. Landa. 2008. Diet shift of a facultative scavenger, the wolverine, following recolonization of wolves. Journal of Animal Ecology 77:1183–1190.
Van Zyll de Jong, C.G. 1975. The distribution and abundance of the wolverine (Gulo gulo) in Canada. Canadian Field-Naturalist 89:431-437.
Wang, T., E. Campbell, G. O’Neill, and S. Aitken. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. Forest Ecology and Management 279 (2012) 128–140).
Webb, S., D. Manzer, R. Anderson, and M. Jokinen. 2013. Wolverine harvest summary from registered traplines in Alberta, 1985-2011. Technical Report, T-2013-001, Alberta Conservation Association, Sherwood Park, AB. 37 pp +App.
Weir, R., unpubl. data. 2012. Fur harvest data. Carnivore Conservation Specialist, Ecosystems Protection and Sustainability Branch, Ministry of Environment, Victoria, BC.
White, K.S., H.N. Golden, K.J. Hundertmark, and G.R. Lee. 2002. Predation of wolves, Canis lupus, on wolverines, Gulo gulo, and an American marten, Martes americana, in Alaska. Canadian Field-Naturalist 116:132-133.
Whitman, J.S., W.B. Ballard, and C.L. Gardner. 1986. Home range and habitat use by wolverines in southcentral Alaska. Journal of Wildlife Management 50:460-462.
Wildlife Management Advisory Council (North Slope). 2008. Species status reports for the Yukon North Slope. Update: January 2008. 142 pp.
Wildlife Management Advisory Council (North Slope) (WMAC(NS)) and the Aklavik Hunters and Trappers Committee (HTC). 2003. Aklavik Inuvialuit describe the status of certain birds and animals on the Yukon North Slope. March 2003. Final Report. Wildlife Management Advisory Council (North Slope). Whitehorse, YU. 60 pp.
Wilson, G.M., R.A. Van Den Bussche, P.K. Kennedy, A. Gunn, and K. Poole. 2000. Genetic variability of wolverines (Gulo gulo) from the Northwest Territories, Canada: conservation implications. Journal of Mammalogy 81:186-196.
Wolverine Science Panel. 2014. Wolverine Science Panel Workshop. Spokane, Washington, April 2014. 47pp.
Wright, J.D., and J. Ernst. 2004. Wolverine, Gulo gulo luscus, resting sites and caching behavior in the boreal forest. Canadian Field-Naturalist 118:61-64.
Yukon Department of Renewable Resources. No date. Managing your wolverine trapline. Yukon Trapline Management Series. Fish and Wildlife Branch, Whitehorse, YT. 4 pp.
Zigouris, Joanna. 2014. Email correspondence with Graham Forbes. Doctoral student, Trent University, Peterborough, ON.
Zigouris, J., F.N. Dawson, J. Bowman, R.M. Gillett, J.A. Schaefer, and C.J. Kyle. 2012. Genetic isolation of wolverine (Gulo gulo) populations at the eastern periphery of their North American distribution. Conservation Genetics 13:1543–1559.
Zigouris, J., J.A. Schaefer, C. Fortin, and C.J. Kyle, 2013. Phylogeography and post-glacial recolonization in wolverines (Gulo gulo) from across their circumpolar distribution. PLoS ONE 8(12) e83837.doi:10.1371/journal.pone.0083837.
Biographical Summary of Report Writer(s)
Brian G. Slough obtained an M.Sc. from Simon Fraser University in 1976. His thesis on Beaver ecology led him to a 15-year career as furbearer management biologist with the Yukon Fish and Wildlife Branch. He has published work on furbearer species including Beaver, Arctic Fox, American Marten, and Canada Lynx, and has also written about trapline and furbearer management in northern and western Canada. He prepared COSEWIC status reports on Wolverine (2003), American Marten, Newfoundland population (2007), and Western Toad (2012). Mr. Slough has recently prepared NWT Species at Risk Committee status reports on the Northern Leopard Frog, Western Toad, and Wolverine.
Since leaving the Yukon government in 1996, Mr. Slough has conducted environmental assessments, protected areas research, and research on rare amphibians and mammals, including rodents, shrews and bats. He served two terms as a member of the Terrestrial Mammals Specialist Subcommittee of COSEWIC (2006 to 2013).
Collections Examined
No collections were examined during the preparation of this report.
Appendix 1. Threats calculator summary for Wolverine.
Threats Assessment Worksheet
- Species or Ecosystem Scientific Name
- Wolverine (Gulo gulo)
- Date:
- 16/01/2014
- Assessor(s):
-
Brian Slough (report writer), Graham Forbes (TM SSC Co-chair), Dave Fraser (BC), Gord Court (AB), Suzanne Carrière (NT), Shelly Moores (NL), Vivian Brownell (ON), Isabelle Gauthier (QC), Thomas Jung (YK), Ruben Boles (CWS), Patrick Nantel (Parks), Donna Hurlburt (ATK Co-chair)
Jeff Bowman (TM SSC member); Nicolas Lecomte (TM SSC member), Neil Dawson (ON), Dean Berezanski (MB), Robert Mulders (NT), Janet Winbourne (TK Researcher), Emily Herdman (NL), Malik Awan (NU), Isabelle Thibault (QC), Vincent Carignan (CWS-QC), Victoria Snable (CWS-Prairie), Paul MacDonald (CWS-NL), Donna Begelow (CWS-NT), Rich Weir (BC)
- References:
- 2014 Draft COSEWIC status report
| Threat Impact | Threat Impact (descriptions) | Level 1 Threat Impact Counts high range |
Level 1 Threat Impact Counts low range |
|---|---|---|---|
| A | Very High | 0 | 0 |
| B | High | 0 | 0 |
| C | Medium | 0 | 0 |
| D | Low | 4 | 4 |
| Calculated Overall Threat Impact: | Medium | Medium |
| # | Threat | Impact (calculated) Criterion |
Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential & commercial development | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 1.1 | Housing & urban areas | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 1.2 | Commercial & industrial areas | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 1.3 | Tourism & recreation areas | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 2 | Agriculture & aquaculture | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 2.1 | Annual & perennial non-timber crops | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 2.2 | Wood & pulp plantations | - | Negligible | Negligible(<1%) | Negligible (<1%) | Low (Possibly in the long term > 10 yrs) | - |
| 2.3 | Livestock farming & ranching | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 3 | Energy production & mining | - | Negligible | Negligible(<1%) | Slight (1-10%) | High (Continuing) | - |
| 3.1 | Oil & gas drilling | - | Negligible | Negligible(<1%) | Slight (1-10%) | High (Continuing) | Increased access may lead to disturbance and mortality from road kill, trapping and nuisance control mortality at camps. BC: increase in access will increase harvest/trapping (see 5.1) Note: road networks are excluded in this calculation (see 4.1) |
| 3.2 | Mining & quarrying | - | Negligible | Negligible(<1%) | Slight (1-10%) | High (Continuing) | Increased access may lead to disturbance and mortality from road kill, trapping and nuisance control mortality at camps. BC: increase in access will increase harvest (see 5.1) NT: There are 4 mines in NT that increase the disturbance and increase a number of related issues (see 5.1) Note: road networks are excluded in this calculation (see 4.1) |
| 3.3 | Renewable energy | - | Negligible | Negligible(<1%) | Negligible (<1%) | High (Continuing) | - |
| 4 | Transportation & service corridors | D | Low | Restricted(11-30%) | Slight (1-10%) | High (Continuing) | (This would include habitat that are rendered ineffective from (ex.) mines and would include road kills as well) |
| 4.1 | Roads & railroads | D | Low | Negligible(<1%) | Slight (1-10%) | High (Continuing) | Transportation corridors may act as barriers to movements and dispersal, and may be a source of mortality (based on road mortality and the effects of access roads. This does not deal with active trapping but creating movement and avoidance) Scope BC: half the population would encounter roads YK: 30-70% of the population would encounter roads NT: the avoidance of roads is low but increase in access will increase hunting; believes that 11-30% of the Canadian population would experience roads Severity YK: there has only been 1 road kill in the past 10 years ON: 6-7 road kills documented BC: there have been a number of documented cases along TransCanada highway; mortality from road evident. There are also about 6-7 deaths associated with logging roads and highways in the southern portion of the range. |
| 4.2 | Utility & service lines | - | Negligible | Negligible (<1%) | Slight (1-10%) | High (Continuing) | Transportation corridors may act as barriers to movements and dispersal, and may be a source of mortality |
| 5 | Biological resource use | D | Low | Pervasive (71-100%) | Slight (1-10%) | High (Continuing) | - |
| 5.1 | Hunting & collecting terrestrial animals | D | Low | Pervasive (71-100%) | Slight (1-10%) | High (Continuing) | Wolverine harvest is restricted and monitoring is mandatory in most jurisdictions. Monitoring in NT and NU is by fur exports and voluntary carcass collection programs. The efficacy of harvest monitoring using carcass collections, in Inuit communities where most pelts are used locally, is uncertain. Harvest of ungulates and large carnivores may impact Wolverine prey and scavenging opportunities. Discussion: what % of Canadian pop. are exposed to trapping/hunting/bycatch? The only area that is excluded is where animals aren’t accessible. Is untrappable area more than 30% in Canada? YK: most animals are exposed to hunting and trapping in YK. Even most parks north of 60° allow trapping (there are no legal barriers) ON: high but not large NL: none in NL NU: NU has more than 90% of the population and gov’t has no jurisdictions BC: increase in road access (due to oil/gas/mining) will increase harvest Severity of the impact QC: Eastern situation is very different than elsewhere. Any mortality will have a high impact on the population to the east. MB: Wolverines are only hunted/trapped for a certain part of the year. Wolverine numbers are increasing in the province AB: has no information ON: 4-5 trapping this year. Similar to MB, Wolverine numbers are expanding in the province (eastward) YK: does not believe there is a decline in Wolverine numbers NU: same, range is also expanding eastward and towards the southern portion NT: agrees that severity is 1-10%; they are taking upwards of 200 and there is a slight range expansion up north BC: harvest is stable and range does not look like it is contracting The situation is more dire in the east but probably negligible for the whole country. |
| 5.3 | Logging & wood harvesting | D | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Road density in logged areas leads to disturbance and mortality from trapping and road kill. Ecological changes due to logging are not permanent, and not necessarily negative Scope BC: all of BC has logging; about 80% of the range QC: we need to look at the connectivity b/w QC and ON ON: most of the logging is in the southern end and the Wolverine population is located north of that. The effect is relatively small; about 1-10% in ON YK: fuel wood harvesting is higher here NT: logging is small but more than people might think. A plan in NT is a move towards oil harvesting. Comfortable with 1% for Canada's range Severity - question "if there is more logging, then wouldn't that mean better habitat for moose = better for Wolverine?" ON: around Red Lake it really depends on the forest. If forest is deciduous, then there is an avoidance by Wolverine, even though there are more moose Concern is that we cannot include cumulative effects of logging, which will change the hunting and trapping of Wolverines. That must be taken into account in 5.1 |
| 6 | Human intrusions & disturbance | D | Low | Restricted (11-30%) | Slight (1-10%) | High (Continuing) | - |
| 6.1 | Recreational activities | D | Low | Restricted (11-30%) | Slight (1-10%) | High (Continuing) | Disturbance of denning females may lead to litter abandonment and kit mortality Discussion that 6.1 and 6.3 should be similar - difficult to separate recreational activity and other activities. |
| 6.2 | War, civil unrest & military exercises | - | Negligible | Negligible (<1%) | Unknown | High (Continuing) | - |
| 6.3 | Work & other activities | D | Low | Restricted (11-30%) | Slight (1-10%) | High (Continuing) | Discussion that 6.1 and 6.3 should be similar - difficult to separate recreational activity and other activities. |
| 7 | Natural system modifications | D | Low | Small(1-10%) | Slight(1-10%) | High (Continuing) | - |
| 7.1 | Fire & fire suppression | - | Not a threat | Restricted - Small (1-30%) | Neutral or Potential Benefit | High (Continuing) | Fires are a natural occurrence in boreal forests and ecosystem effects are positive for Wolverine. Forest fires definitely play a factor in the Western Boreal; scope is larger than 10%. |
| 7.2 | Dams & water management/use | - | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | - |
| 7.3 | Other ecosystem modifications | D | Low | Small(1-10%) | Slight(1-10%) | Moderate (Possibly in the short term, < 10 yrs) | Discussion on the declining caribou population * BC: there is a decline in the Southern Mountain pop. but the effec. is not a large factor; Wolverines have other ungulates to choose from (ex. moose) * YK: similar situation as BC; there is more diversity and Wolverines rely on more than just caribou * MB: unsure, but one staff certainly thought that moose could play a factor. It has been over 25 years since the big forest fire and there is an increase in moose habitat now; particularly in the northern part of MB. NT: Bathurst has a decline of 90% in herd size and about 70% decline in the central herd |
| 8 | Invasive & other problematic species & genes | - | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/Negligible (Past or no direct effect) | - |
| 8.1 | Invasive non-native/alien species | - | - | - | - | - | MB: getting an increase in White-tailed Deer; another potential prey that could affect the Wolverine population Noted that forest pests are dealt with under ecosystem change (7.3) |
| 9 | Pollution | - | Negligible | Negligible (<1%) | Negligible (<1%) | Low (Possibly in the long term, >10 yrs) | - |
| 9.2 | Industrial & military effluents | - | Not Calculated (outside assessment timeframe) | Unknown | Unknown | Insignificant/Negligible (Past or no direct effect) | This could be an issue, but not enough information exists YK: there was a paper done but cannot recall the findings |
| 9.3 | Agricultural & forestry effluents | - | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/Negligible (Past or no direct effect) | - |
| 9.4 | Garbage & solid waste | - | - | - | - | - | May result in habituation of Wolverine and resultant nuisance control mortality. Of the lone Wolverine found, garbage did not contribute to the death, it was a capture attempt and therefore would fall under hunting and trapping (5.1) |
| 9.5 | Air-borne pollutants | - | Unknown | Unknown | Unknown | High (Continuing) | - |
| 9.6 | Excess energy | - | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/Negligible (Past or no direct effect) | BC: there could be threats around oil/gas exploration or maintenance |
| 10 | Geological events | - | Not a threat | Negligible (<1%) | Neutral or Potential Benefit | Moderate (Possibly in the short term, < 10 yrs) |
- |
| 10.3 | Avalanches/landslides | - | Not a threat | Negligible (<1%) | Neutral or Potential Benefit | Moderate (Possibly in the short term, < 10 yrs) |
Minor source of mortality Mortality of Wolverine might be insignificant, but the carrion and debris from avalanches might provide scavenging opportunities and den sites. |
| 11 | Climate change & severe weather | - | Unknown | Pervasive (71-100%) |
Unknown | High (Continuing) | - |
| 11.1 | Habitat shifting & alteration | - | Unknown | Pervasive (71-100%) |
Unknown | High (Continuing) | Reductions in the extent of Wolverine habitat due to reductions in spring cover is disputable. It is likely that a decrease in snow season length would not impact natal denning requirements, because a net increase in snowfall should make up for the shorter snow season, resulting in increased snow accumulation. Any impact on Canadian Wolverine populations should be first detected in the Southern Mountain ecological area of BC and Alberta. A shorter snow season may increase primary productivity to the benefit of Wolverine. Discussion on snow conditions (US study) and if it’ll happen in 10 yrs BC: the model forecast is within 25 yrs; more of a longer term issue. However, in the last 10 yrs, southern portion of BC had noticeable snow shrinkage NT: impact of snow is minimal in northern latitude. What would be the bigger issue is how climate change affects larger ungulates, thus having an effect on the scavenger species. Both a direct and indirect effect. YK: Unknown, but reduced snow length season would benefit Wolverines. The whole issue of the US study is controversial but most like the model that US has proposed. Expecting that the suitable snow conditions will shrink substantially over the long term; this will have an effect, but not sure by how much. |
| 11.3 | Temperature extremes | - | Unknown | Unknown | Unknown | Unknown | - |