
Bumble bee (Bombus affinis): proposed recovery strategy 2016
Species at Risk Act
Recovery Strategy Series

2016
Table of Contents
- Document Information
- Preface
- Acknowledgements
- Executive Summary
- Recovery Feasibility Summary
- 1. COSEWIC Species Assessment Information
- 2. Species Status Information
- 3. Species Information
- 4. Threats
- 5. Population and Distribution Objectives
- 6. Broad Strategies and General Approaches to Meet Objectives
- 7. Critical Habitat
- 8. Measuring Progress
- 9. Statement on Action Plans
- 10. References
- Appendix A: Plant species used by the Rusty-patched Bumble Bee
- Appendix B: Effects on the environment and on other species
Document Information

Recommended citation:
Environment and Climate Change Canada. 2016. Recovery Strategy for the Rusty-patched Bumble Bee (Bombus affinis) in Canada [Proposed], Species at Risk Act Recovery Strategy Series, Environment and Climate Change Canada, Ottawa, vii + 56 p.
For copies of the recovery strategy, or for additional information on species at risk, including the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) Status Reports, residence descriptions, action plans, and other related recovery documents, please visit the Species at Risk (SAR) Public Registry.
Cover illustration: © Johanna James-Heinz
Aussi disponible en français sous le titre: « Programme de rétablissement du bourdon à tache rousse (Bombus affinis) au Canada [Proposition] »
Content (excluding the cover illustration) may be used without permission, with appropriate credit to the source.
Preface
The federal, provincial and territorial government signatories under the Accord for the Protection of Species at Risk (1996) agreed to establish complementary legislation and programs that provide for effective protection of species at risk throughout Canada. Under the Species at Risk Act (S.C. 2002, c. 29) (SARA), the federal competent ministers are responsible for the preparation of recovery strategies for listed Extirpated, Endangered, and Threatened species and are required to report on progress within five years after the publication of the final document on the Species at Risk Public Registry.
The Minister of Environment and Climate Change is the competent minister under SARA for the recovery of the Rusty-patched Bumble Bee and has prepared this strategy as per section 37 of SARA. To the extent possible, it has been prepared in cooperation with the Ontario Ministry of Natural Resources and Forestry and the Quebec Department of Forests, Wildlife and Parks.
Success in the recovery of this species depends on the commitment and cooperation of many different parties that will be involved in implementing the recommendations made in this strategy and will not be achieved by Environment and Climate Change Canada or any other jurisdiction alone. All Canadians are invited to join in supporting and implementing this strategy for the benefit of the Rusty-patched Bumble Bee and Canadian society as a whole.
This recovery strategy will be followed by one or more action plans that will provide information on recovery measures to be taken by Environment and Climate Change Canada and other jurisdictions and/or organizations involved in the conservation of the species. Implementation of this strategy is subject to appropriations, priorities, and budgetary constraints of the participating jurisdictions and organizations.
The recovery strategy sets the strategic direction to arrest or reverse the decline of the species, including identification of critical habitat to the extent possible. It provides all Canadians with information to help take action on species conservation. When the recovery strategy identifies critical habitat, there may be future regulatory implications, depending on where the critical habitat is identified. SARA requires that critical habitat identified within a national park named and described in Schedule 1 to the Canada National Parks Act, the Rouge National Urban Park established by the Rouge National Urban Park Act, a marine protected area under the Oceans Act, a migratory bird sanctuary under the Migratory Birds Convention Act, 1994 or a national wildlife area under the Canada Wildlife Act be described in the Canada Gazette, after which prohibitions against its destruction will apply. For critical habitat located on other federal lands, the competent minister must either make a statement on existing legal protection or make an order so that the prohibition against destruction of critical habitat applies. For any part of critical habitat located on non-federal lands, if the competent minister forms the opinion that any portion of critical habitat is not protected by provisions in or measures under SARA or other Acts of Parliament, or the laws of the province or territory, SARA requires that the Minister recommend that the Governor in Council make an order to prohibit destruction of critical habitat. The discretion to protect critical habitat on non-federal lands that is not otherwise protected rests with the Governor in Council.
Acknowledgements
This recovery strategy was prepared by Sylvain Giguère and Marie-José Ribeyron (Environment and Climate Change Canada, Canadian Wildlife Service – Quebec) based on the Government of Ontario's recovery strategy for the species (Colla and Taylor-Pindar 2011) and a draft federal addition to this strategy prepared by Talena Kraus (Artemis Eco-Works). The document was also made possible through the contributions of Michel Saint-Germain and Pierre-André Bernier (Environment and Climate Change Canada, Canadian Wildlife Service – Quebec), Angela Darwin, Lesley Dunn, Judith Girard, Elizabeth Rezek, Liz Sauer and Kathy St. Laurent (Environment and Climate Change Canada, Canadian Wildlife Service – Ontario), Isabelle Gauthier and Nathalie Desrosiers (Ministère des Forêts, de la Faune et des Parcs du Québec), Vivian Brownell, Jay Fitzsimmons, Anita Imrie, Dr. Brian Naylor, and Don A. Sutherland (Ontario Ministry of Natural Resources and Forestry) as well as André Payette and Maxim Larivée (Montreal Insectarium).
We would also like to thank everyone else who provided advice and input used to help inform the development of this recovery strategy, including various Aboriginal organizations and individuals, landowners, citizens and stakeholders who provided input and/or participated in consultation meetings.
Executive Summary
The Rusty-patched Bumble Bee is a medium to large bumble bee that has a rusty-coloured patch bordered by yellow on the first half of its abdomen. Like most bumble bees, it has an annual life cycle and requires a variety of habitats at different stages in this cycle.
In the 1970s, the Rusty-patched Bumble Bee was relatively common throughout its range which, in Canada, includes southern Ontario and southwestern Quebec. Since the mid-1990s, the species has suffered rapid, severe decline. It was listed as Endangered on Schedule 1 of the Species at Risk Act in 2012.
The primary threats to the Rusty-patched Bumble Bee are the use of pesticides, particularly neonicotinoids, pathogen transmission and spillover, climate change and severe weather events as well as intensive agriculture, urban and suburban development, and the road network development.
There are unknowns regarding the feasibility of recovery of the Rusty-patched Bumble Bee. Nevertheless, in keeping with the precautionary principle, this recovery strategy has been prepared as per subsection 41(1) of SARA, as would be done when recovery is determined to be feasible. This recovery strategy addresses unknowns surrounding the feasibility of recovery.
The population and distribution objectives for the Rusty-patched Bumble Bee in Canada are to ensure the viability of the local population in Pinery Provincial Park and of any other local population that might be discovered in the future, and, as needed, increase the number of viable local populations in the species' current and historical range, in order to form, in the long term and to the extent possible, a species' distribution that is not severely fragmented as a result of human activity.
Broad strategies as well as research and management approaches to achieve the objectives are presented in the Strategic Direction for Recovery section.
Critical habitat for the Rusty-patched Bumble Bee is partially identified in this recovery strategy. The critical habitat for the species is identified as any suitable habitat located within a 1,000 m radius of any valid sightings of the species since 2005. A schedule of studies outlines the activities required to complete the identification of critical habitat.
One or more action plans will follow this recovery strategy and will be posted on the Species at Risk Public Registry within five years of the posting of the final recovery strategy.
Recovery Feasibility Summary
Based on the following four criteria that Environment and Climate Change Canada uses to establish recovery feasibility, there are unknowns regarding the feasibility of recovery of the Rusty-patched Bumble Bee. In keeping with the precautionary principle, this recovery strategy has been prepared as per section 41(1) of SARA, as would be done when recovery is determined to be feasible. This recovery strategy addresses the unknowns surrounding the feasibility of recovery.
Individuals of the wildlife species that are capable of reproduction are available now or in the foreseeable future to sustain the population or improve its abundance.
Unknown. The presence of the Rusty-patched Bumble Bee in Canada was confirmed by surveys conducted in Ontario between 2002 and 2014. Three individuals were sighted in Pinery Provincial Park between 2005 and 2009, which confirmed the presence of at least one colony during that period. In Quebec, where no targeted survey has been undertaken since the mid-1970s, the species was documented sporadically from a variety of locations between 1990 and 2000. These few recent sightings, the species' generalist nature and its fairly broad historical range, all suggest that the species is still present in Canada. However, the best available evidence suggests that the Canadian population is very small and that a rescue effect from the United States is unlikely, given the population and range declines observed in states adjacent to Ontario and Quebec. For these reasons, there are uncertainties regarding the possibility of sustaining the Canadian population or increasing its abundance.
Enough suitable habitat is available to support the species, or could be made available through habitat management or restoration activities.
Yes. The Rusty-patched Bumble Bee is a generalist forager and fairly flexible in terms of its needs related to nesting and overwintering sites. It is therefore very likely that enough suitable habitat still exists in most of the species' historical range in Canada. The area of available suitable habitat could be increased through habitat management or restoration measures. The species' foraging habitat may be fragmented to the point where its survival is compromised; however, here again, management measures could be put in place to correct the situation.
The primary threats to the species or its habitat (including threats outside Canada) can be avoided or mitigated.
Unknown. Although the mechanisms responsible for the dramatic decline of the species are unknown, the main current threats to the species have been identified and are as follows: the use of pesticides, particularly neonicotinoids, pathogen transmission and spillover, climate change and severe weather events, as well as intensive agriculture, urban and suburban development, and the road network development. It may be possible to avoid or mitigate the impacts of nearly all these threats. However, there are doubts concerning the possibility of reducing or mitigating long-term impacts from climate change (e.g., species range contraction) and severe weather events which could eliminate colonies of the species.
Recovery techniques exist to achieve the population and distribution objectives or can be expected to be developed within a reasonable time frame.
Yes. Techniques for increasing the amount of suitable habitat or enhancing habitat exist, as do techniques for reducing or mitigating the main threats that have been identified (e.g., legislation regarding pesticide use, best management practices). The Rusty-patched Bumble Bee is also a good candidate species for population restoration.Footnote1 The feasibility of population restoration and the associated risks need to be evaluated, however, in order to determine the suitability of this approach. In addition, in the short term, severe weather events could cause the extirpation of the local population in Pinery Provincial Park, given that it is believed to be very small. Known climate change adaptation techniques (e.g., reduction and mitigation of other threats, establishment of connectivity corridors) could therefore be insufficient to address these impacts and, consequently, may not be sufficient to achieve the population and distribution objectives.
1. COSEWICNotea Species Assessment Information
- Date of Assessment:
- April 2010
- Common Name (population):
- Rusty-patched Bumble Bee
- Scientific Name:
- Bombus affinis
- COSEWIC Status:
- Endangered
- Reason for Designation:
- This species, which has a distinctive color pattern, was once commonly found throughout southern Ontario Noteb. Active searches throughout its Canadian range have detected only one small population over the past seven years which suggests a decline of at least 99% over the past 30 years. It is threatened by disease, pesticides, and habitat fragmentation, each of which could cause extirpation in the near future.
- Canadian Occurrence:
- Ontario and Quebec
- COSEWIC Status History:
- Designated Endangered in April 2010
2. Species Status Information
In the 1970s, the Canadian range of the Rusty-patched Bumble Bee accounted for less than 10% of the species’ global range (COSEWIC 2010). The species has since suffered rapid, severe decline causing global range losses of 70% to 87% (Cameron et al. 2011a; Colla et al. 2012). There are few data available to determine its current Canadian range (Evans et al. 2008; NatureServe 2015). The Rusty-patched Bumble Bee was listed as endangeredFootnote2 in Schedule 1 of the Species at Risk Act in 2012. It was listed as endangeredFootnote3 under Ontario’s Endangered Species Act (ESA), 2007 (S.O. 2007, c. 6) in 2007 and was included on the list of species likely to be designated threatened or vulnerable in Quebec (CQLR, c. E-12.01) in 2010.
Globally, the Rusty-patched Bumble Bee is listed as critically imperilled (G1) (NatureServe 2015). Its conservation status rank is N1 (critically imperiled) in Canada and NNR (status not yet assessed) in the United States, where the species is ranked “possibly extirpated” (SH) in 18 of the 23 states that make up its range in the United States. The subnational conservation status ranks for the species are presented in Table 1.
| Country | Provinces/States and NatureServe Conservation Status Ranks Notec |
|---|---|
| Canada | Ontario (S1), Quebec (SNR) |
| United States | North Carolina (SH), South Carolina (SH), Connecticut (SNR), North Dakota (SH), Georgia (SH), Illinois (SNR), Indiana (S1), Iowa (SH), Kentucky (SH), Maine (SH), Maryland (SH), Massachusetts (SH), Michigan (SH), Minnesota (SH), New Hampshire (SH), New Jersey (SH), New York (SH), Ohio (SH), Pennsylvania (SH), Tennessee (SH), Vermont (S1), Virginia (SH), Wisconsin (S1) |
The species is listed as critically endangeredFootnote4 (CR) on the International Union for Conservation of Nature (IUCN) Red List of Threatened Species (IUCN 2015).
3. Species Information
3.1 Species Description
The Rusty-patched Bumble Bee is a medium to large bumble bee with several distinguishing characteristics. Males and workers (11 to 16 mm in length) have a rusty brown spot on the otherwise yellow dorsal surface of their second abdominal segment (Mitchell 1962). This colour pattern explains the origin of the species' common name. Queens are larger (21 to 22 mm in length) and the dorsal surface of their second abdominal segment is completely yellow--a colour pattern characterizing many of the bumble bee species found in eastern North America (Mitchell 1962). The thorax is mostly yellow, usually with a black band between the wing bases (Laverty and Harder 1988). The species can be differentiated from other bumble bees in that it has a very short space between the eye and the mandible and an entirely black head and face (Colla and Taylor-Pindar 2011). For a more detailed description of the species, see the COSEWIC status report (2010).
3.2 Species Population and Distribution
The Rusty-patched Bumble Bee is an eastern North American species that reaches the northern limit of its range in Canada. Its historical range extended as far west as the Dakotas, and from southern Ontario and southwestern Quebec in the north to Georgia in the south (COSEWIC 2010).
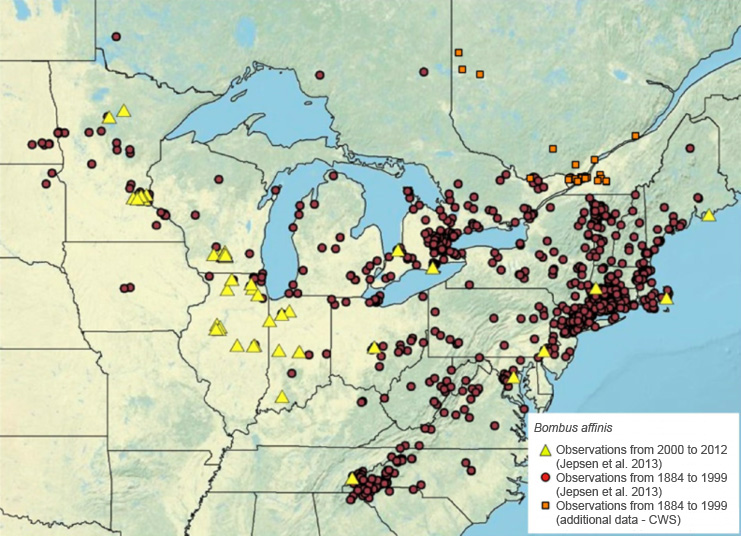
Prior to the 1990s, the Rusty-patched Bumble Bee was relatively common throughout its range compared to other bumble bee species (Evans et al. 2008; Williams and Osborne 2009; COSEWIC 2010; Cameron et al. 2011a). For example, in surveys conducted in southern Ontario in 1971-1973, it made up approximately 14% of the 3,622 bumble bees collected, making it the fourth most common of the 14 bumble bee species documented (Colla and Packer 2008). However, since at least 2002, the species, like many other bumble bee species in North America, has suffered rapid, severe decline throughout its entire range (Colla and Packer 2008; Evans et al. 2008; COSEWIC 2010; Cameron et al. 2011a; Colla et al. 2012; Bartomeus et al. 2013a). Range-wide studies have found relative abundance declines of up to 95% (Cameron et al. 2011a), and range losses of 70% to 87% (Cameron et al. 2011a; Colla et al. 2012). Not enough data are available on the species' presence to be able to determine its current range (NatureServe 2015). The Rusty-patched Bumble Bee was recorded repeatedly, but not commonly, from 2003 to 2012 in Illinois, Wisconsin, Minnesota and Ontario, which suggests that the species may still be present in one or two main areas (e.g., Dane County, Wisconsin) (NatureServe 2015). There are also isolated reports from Massachusetts (2009) and Pennsylvania (2007) (NatureServe 2015), as well as Virginia, where the species was considered extirpated until one individual was sighted in 2014 (Smithsonian Science 2014). These latter observations suggest that the species may still be present in areas of its historical range that have not been surveyed recently. Figure 1 shows the historical and recent global distribution of the species.
Long description for Figure 1
Figure 1 shows the historical and recent distribution of the species in North-America. From 1884 to 1999 records of the species were found along the northeastern coast of the United States from Maine to Northern Alabama and from Massachusetts to North Dakota. The greatest densities of records are near Connecticut, Toronto and Chattanooga. The data provided by CWS for the same timeline adds records of the species in southern Quebec. The records from 2000 to 2012 are much less dense but equally dispersed as those from 1884 to 1999. One or two records are found in each of the high density areas from 1884 to 1999. The highest density for this time period is in the states of Illinois and Indiana where a dozen or so records are found.
The current size of the Rusty-patched Bumble Bee population in Canada (Ontario and Quebec) is unknown. In Ontario, surveys were carried out in historical Rusty-patched Bumble Bee locations between 2002 and 2010 (Colla and Packer 2008; COSEWIC 2010; Colla and Taylor-Pindar 2011). Despite over 600 hours of targeted searches, only three individualsFootnote5 (one male in 2005 and two workers in 2009Footnote6) were sighted, all in Pinery Provincial Park, Ontario. The species was not re-located in subsequent surveys conducted in Pinery Provincial Park up to 2014 (A. MacKenzie pers. comm. 2015). Locations that had records for this species in the 1990s but where surveys since 2000 have not produced any records include Manestar Tract (Norfolk County)Footnote7; the University of Western Ontario campus, in London (Middlesex County); High Park, in Toronto; Darlington Provincial Park; Guelph (Wellington County); and the Humber River, in Toronto (Colla and Packer 2008; Colla and Taylor-Pindar 2011). A specimen collected close to Pinery Provincial Park in 2014 and originally identified as Rusty-patched Bumble Bee (Patterson 2014) was later confirmed to be another species (A. MacDougall pers. comm. 2015). In Quebec, where no targeted surveys have been documented in the species' historical range, the available data (1920-1996) indicate the presence of the Rusty-patched Bumble Bee in the southwestern part of the province, specifically in Montérégie, the Greater Montreal Area, the Outaouais, the Laurentian Mountains, and the Quebec Capitale Nationale region, as well as farther north, in the Abitibi-Témiscamingue region (Savard 2009; Duquette and Boutin 2015) (Figure 1). Many specimens of the species were collected in Oka (Ouellet-Robert entomological collection, University of Montreal) and Sainte-Anne-de-Bellevue (Lyman Museum, McGill University). The most recent confirmed records date from 1993 (Luskville) and 1996 (Mont Rigaud) (A. Payette pers. comm. 2015). These two sites are located in conservation areas and have not been surveyed since.
The information presented earlier in this section indicates that the Rusty-patched Bumble Bee may still be present in Canada. It also demonstrates that there are significant knowledge gaps regarding the species' population and distribution in Canada and that additional surveys and monitoring activities need to be carried out in order to fill these gaps.
The fact that only three individuals were sighted during the 600 hours of surveys conducted in Ontario between 2005 and 2009 suggests that the Rusty-patched Bumble Bee population is very small (COSEWIC 2010). This deduction is confirmed and validated by current knowledge about the demographic characteristics of social insects. Although a bumble bee colony contains a large number of individuals, it cannot be considered a population. In fact, each colony contains only one reproducing female (the queen) and she usually mates only once; consequently, each colony essentially represents only one breeding pair (Goulson 2010) and this pair dies at the end of the season. Thus, the observation of one male in 2005 and two workers located about three kilometres apart in 2009 represents only an indication of the presence of one breeding pair in 2005 and possibly two breeding pairs in 2009.
Given its small size, the only known local population of the Rusty-patched Bumble Bee in Canada is more likely to become extinct due to demographicFootnote8 or environmental stochasticityFootnote9 (Chapman and Bourke 2001; Frankham et al. 2002; Goulson 2008; Evans 2009). It is therefore at very high risk of extinction. If a small local population, like the Pinery Provincial Park population, is part of a group of interrelated local populations (i.e., movements of breeding individuals between habitat fragments are not prevented by the level of habitat fragmentation), local extinctions may be offset by subsequent recolonization by individuals dispersing from other local populations, thereby ensuring persistence of the species at the local scale (Goulson 2008; Dempster and McLean 2012) as well as genetic cohesion of the species (Goulson 2008). However, there is no evidence indicating that recolonization is a possibility for the Rusty-patched Bumble Bee in Canada.
The Rusty-patched Bumble Bee is relatively cold-tolerant, and evidence from other bumble bees (Bombus spp.) shows limited genetic variation across the range (Lozier et al. 2011), suggesting it may be possible that individuals from the United Sates would be adapted to live in Canada. However, it is unlikely that the Canadian population would benefit from a rescue effect from the United States population because the species has experienced the same significant decline there as it has in Canada. Furthermore, the distances separating the existing populations in the United States and Canada are too great.
3.3 Needs of the Rusty-patched Bumble Bee
The Rusty-patched Bumble Bee is a generalist species that uses open habitats; it has been found in a variety of habitats, such as mixed farmland (cropping and livestock use), savannah, sand dunes, marshes, and urban and wooded areas (COSEWIC 2010).
The Rusty-patched Bumble Bee is more cold-tolerant than the other bumble bee species of North America, allowing it to occur at higher elevations and emerge earlier in the spring (Colla and Dumesh 2010). It is one of the first bee species to emerge in the spring (mid-April) and one of the latest to cease foraging in the fall (late October) (COSEWIC 2010). New queens and males are produced from mid-July or early August to September (Jepsen et al. 2013). The timing of the different stages in the colony cycle can vary from year to year with seasonal variation and latitude (Colla and Taylor-Pindar 2011), but the species requires habitats with rich supplies of floral resources with continuous blooming from spring to fall (Evans 2009), i.e., a variety of species blooming one after the other from April to October. The Rusty-patched Bumble Bee may be particularly susceptible to stressors (e.g., extreme temperatures, pathogens) because it emerges so early in the spring when few flowers are in bloom and because it does not produce the next generation until late in the summer (Jepsen et al. 2013).
The habitat needs of the Rusty-patched Bumble Bee are closely linked to its life cycle or colony cycle, which, as is the case for most bumble bees, is annual and consists of the following stages:
- in spring, the queen produced the previous year emerges from its overwintering site to forage and locate a suitable nest site;
- after selecting a nest site, the queen lays eggs and continues to forage to provision its first brood, which will give rise to female workers;
- once the workers hatch, they take over nest care and foraging, and the queen focuses on egg-laying. Towards the end of the cycle, the colony begins producing new queens and males, instead of workers;
- the new queens mate with males;
- newly mated queens locate suitable overwintering sites; the existing colony declines until late fall, when the males, workers and the old queen die (Williams et al. 2014).
Early spring is a critical time for bumble bees because this is when the queens emerge from hibernation and initiate nests (Jepsen et al. 2013).
Bumble bees use three different types of habitat, which correspond to the needs associated with the different stages in their life cycle.
Overwintering habitat
There are no data on the overwintering habitat of the Rusty-patched Bumble Bee, but its requirements are assumed to be similar to those of other Bombus species, including sites suitable for underground burrows in loose soil or the presence of fallen dead wood (Macfarlane 1974). Queens of other bumble bee species dig a few centimetres into the soil and form an oval-shaped chamber in which they spend the entire winter (Jepsen et al. 2013). Compost in gardens or mole hills may provide suitable sites for queens to overwinter (Goulson 2010).
Nesting habitat
All members of the subgenus Bombus sensu stricto usually nest underground (Macfarlane 1974; Laverty and Harder 1988). The Rusty-patched Bumble Bee generally nests one to four feet (30 to120 cm) below ground in abandoned rodent nests or other cavities (Jepsen et al. 2013). When suitable burrows are not available, the species may nest in hollow tree stumps or, occasionally, above ground, in dead wood or clumps of grasses (Macfarlane 1974; Evans et al. 2008; Jepsen et al. 2013). The number of nesting sites may thus be limited by a decrease in the abundance of rodents and the presence of undisturbed grassland (Evans 2009; Jepsen et al. 2013). The nest is annual in that the colony that occupies it over the summer will perish with the onset of winter and the new queens that mate in the fall will emerge the following spring and establish nests for new colonies. Lepais et al. (2010) reported that queens of the genus Bombus may travel a few kilometres (3-5 km) to find a suitable nest site. The location of nests thus changes from year to year, and the new nest may be a considerable distance from the previous one.
Foraging habitat
Foraging habitat for the Rusty-patched Bumble Bee typically consists of an abundance of wildflowers (e.g., Wild Bergamot [Mondarda fistulosa], Virginia Mountain-Mint [Pycnanthemum virginianum], and Wild Mint [Mentha arvensis]) growing in the forest understory or in open habitats. The plant species used by the Rusty-patched Bumble Bee are listed in Appendix A. Bumble bees are generalist foragers, collecting nectar and pollen from a wide variety of plant species. Nectar provides the bees with carbohydrates and pollen provides proteins. The amount of pollen available to bumble bee colonies has a direct effect on the number of new queens that can be produced (Burns 2004 in Jepsen et al. 2013) and, consequently, on future population levels. The loss of early flowering plant species can compromise the survival of queens emerging in the spring (COSEWIC 2010). Given the diversity of nectar and pollen sources it uses, the Rusty-patched Bumble Bee forages in a wide variety of habitats, including mixed farmland, sand dunes, marshes, and urban and lightly wooded areas (Colla and Dumesh 2010). Workers may fly considerable distances while foraging. For the Buff-tailed Bumble Bee (Bombus terrestris), a closely related species, maximum foraging distance was estimated to range from 100 to 2,300 m (Osborne et al. 1999; Walther-Hellwig and Frankl 2000; Chapman et al. 2003; Darvill et al. 2004; Knight et al. 2005; Kraus et al. 2008; Osborne et al. 2008; Wolf and Moritz 2008). In Walther-Hellwig and Frankl (2000), a maximum distance of 500 m was recorded for 43% of the sample size, while 25% of the sample size travelled a maximum distance of 1,750 m. More recent studies (Hagen et al. 2011; Rao and Strange 2012) indicate that a number of Bombus species may travel more than 2.5 km.
It has been shown that habitat quality at the landscape scale has an effect on the diversity and abundance of bumble bee species and that isolated patches of habitat are not sufficient to support bumble bee populations (Hatfield and LeBuhn 2007 and Öckinger and Smith 2007 in Evans et al. 2008). Darvill et al.(2012) found that bumble bee populations limited to less than 15 km2 of suitable habitat are more likely to show signs of inbreeding. Goulson (2010) suggested that a viable population of bumble bees probably requires 3.3 to 10 km2 of suitable habitat.
Limiting factors
The Rusty-patched Bumble Bee is at the northern limit of its range in Canada. Climatic variables such as snow cover, precipitation, and growing season length are likely important determinants of suitable habitat for the species. Given that the Rusty-patched Bumble Bee is found only at high elevations in the southern part of its range, it is likely that the species is restricted to a narrow climatic niche. Williams et al. (2009) found that bumble bees with narrow climatic niches are more vulnerable to extinction.
Zayed and Packer (2005) reported that as a bumble bee population decreases in size, the frequency of diploid malesFootnote10 increases. As diploid males are usually sterile or non-viable, an increase in their number in small populations increases the rate of population decline, causing a special case of the extinction vortex called “the diploid male extinction vortex” (Zayed and Packer 2005). For this reason, bumble bee populations could be particularly vulnerable when small population size is combined with habitat fragmentation (Evans 2009; Goulson 2010; Jepsen et al. 2013).
4. Threats
COSEWIC (2010) identified and assessed the threats to the Rusty-patched Bumble Bee on the basis of the contribution of those threats to the decline of the species in recent decades. As the causes and mechanisms of this decline are still unknown, all identified threats were considered potential threats in the COSEWIC status report (2010). Since this recovery strategy is based on the IUCN/CMP unified threats classification scheme, only current and future threats are considered, with no attempt being made to understand the causes of past declines. Given the lack of direct evidence of threats to the Rusty-patched Bumble Bee in Canada, this recovery strategy considers the threats to the species to be inferred threats.Footnote11 Two potential threats are identified and described below.
4.1 Threat Assessment
The threats have been assessed using the entire historical range of the species in Canada, taking into account that the species may be present elsewhere than within Pinery Provincial Park, i.e., in parts of the range where targeted surveys have not been conducted recently.
The assessment of the threats to the Rusty-patched Bumble Bee is based on the IUCN/CMP (International Union for the Conservation of Nature/Conservation Measures Partnership) unified threats classification scheme. Limiting factors are not considered during this assessment process. Threats are defined as the proximate activities or processes that have caused, are causing, or may cause the destruction, degradation, and/or impairment of the entity being assessed (population, species, community, or ecosystem) in the area of interest (global, national, or subnational). For information on how the values are assigned in the table (Table 2), see the table footnotes. Historical threats, indirect or cumulative effects of the threats, and any other relevant information that would help to understand the nature of the threats are presented in section 4.2 (Description of Threats).
| Threat | Threat Description | ImpactNoted | ScopeNotee | SeverityNotef | TimingNoteg | Threat Details |
|---|---|---|---|---|---|---|
| 1 | Residential and commercial development | - | - | - | - | - |
| 1.1 | Housing and urban areas | High - Low | Large - Restricted (11-70%) | Serious - Slight (1-70%) | High | Urban and suburban development |
| 2 | Agriculture and aquaculture | - | - | - | - | - |
| 2.1 | Annual and perennial non-timber crops | High - Low | Large - Restricted (11-70%) | Extreme - Moderate (11-100%) | High | Intensive agriculture |
| 2.3 | Livestock farming and ranching | Low | Restricted -Small (1-30%) | Moderate - Slight (1-30%) | High | Livestock grazing |
| 4 | Transportation and service corridors | - | - | - | - | - |
| 4.1 | Roads and railroads | High - Low | Pervasive - Large (31-100%) | Serious - Slight (1-70%) | High | Road network Development |
| 7 | Natural system modifications | - | - | - | - | - |
| 7.1 | Fire and fire suppression | Low | Restricted -Small (1-30%) | Moderate - Slight (1-30%) | Moderate - Insignificant/ Negligible |
Fire suppression |
| 8 | Invasive and other problematic species and genes | - | - | - | - | - |
| 8.1 | Invasive non-native/alien species | Very High -Medium | Pervasive - Large (31-100%) | Extreme - Moderate (11-100%) | High | Pathogen transmission and spillover |
| 8.1 | Invasive non-native/alien species | Medium - Low | Large - Restricted (11-70%) | Moderate - Slight (1-30%) | High | Competition with honey bees |
| 9 | Pollution | - | - | - | - | - |
| 9.3 | Agricultural and forestry effluents | Very High -Medium | Pervasive - Large (31-100%) | Extreme – Moderate (11-100%) | High | Pesticide use |
| 11 | Climate change and severe weather | High - Medium | Pervasive - Large (31-100%) | Serious - Moderate (11-70%) | High | - |
4.2 Description of Threats
The threats to the Rusty-patched Bumble Bee are presented below in decreasing order of impact.
Agricultural and forestry effluents – Pesticide use
Bumble bees are non-target species that can be adversely affected by pest and weed control programs implemented in the agriculture and forestry sectors. They can be poisoned by pesticides when they absorb toxins directly through their exoskeleton, drink contaminated nectar or gather contaminated pollen, or when the larvae consume contaminated pollen (Jepsen et al. 2013). Various life-history traits of the Rusty-patched Bumble Bee (such as large body size, early emergence and long colony cycle) may make it especially vulnerable to accumulation of pesticides in the colony (COSEWIC 2010). Since males and queens are produced at the end of the colony cycle, pesticides applied at any time during the bumble bee life cycle can have substantial adverse effects on subsequent generations (Jepsen et al. 2013). However, the application of pesticides in the spring when the queens are foraging and when nests are small is likely to have the greatest impact on bumble bee populations (Goulson et al. 2008).
Systemic insecticides
Systemic insecticides are taken up by the roots and spread to all parts of the plant, including the nectar and pollen. In the late 1990s, Canada approved the use of several neonicotinoid insecticides (COSEWIC 2010). These insecticides are widely used on agricultural crops that are attractive to insect pollinators (vegetable crops, orchards), as well as on horticultural plants and lawns in urban and suburban areas (Jepsen et al. 2013). They are also used in forestry and aquaculture as well as in veterinary products (Simon-Delso et al. 2015). The metabolitesFootnote12 that form after neonicotinoids are absorbed and metabolized by plants can have effects on various non-target species (e.g., soil bacteria, tomatoes, honey bees, mice, chickens) (Simon-Delso et al. 2015). They can also persist and accumulate in soils; they are water soluble and prone to leaching into waterways (Goulson 2013). Studies using colonies of bumble bees fed a sugar solution containing sublethal doses of neonicotinoids (Gill et al. 2012; Whitehorn et al. 2012), along with studies of field-foraging colonies (Gill and Raine 2014; Rundlöf et al. 2015), showed that neonicotinoids can modify the behaviour of bumble bees (e.g., pollen foraging ability) and reduce the colony growth rate as well as the production of queens. In a recent study, Kessler et al. (2015) reported that the honey bee (Apis mellifera) and the Buff-tailed Bumble Bee (Bombus terrestris) did not avoid sugar solutions containing neonicotinoids. Instead, the two species preferred sucrose solutions laced with two neonicotinoids (imidacloprid, thiamethoxam) to sucrose alone. The authors concluded that treating flowering crops with these two neonicotinoids presents a sizeable hazard to foraging bees. See Pisa et al. (2015) for more information on the impact of neonicotinoids on non-target invertebrates such as the Rusty-patched Bumble Bee.
Non-systemic (or contact) insecticides
Contact insecticides have a toxic effect when the target pest comes into direct contact with them. They can be inorganic, usually derived from a metal (e.g., sulphur), organic (i.e., synthetically produced organic chemical compounds), or of natural origin (e.g., pyrethrum, neem oil). Contact insecticides have little to no residual activity (non-persistent) so their effects are temporary. However, contact insecticides can cause direct bumble bee mortality. For example, a study carried out in New Brunswick (reviewed in Kevan and Plowright 1995) documented drastic declines in bumble bee populations following the application of fenitrothion.Footnote13 Thompson (2001) reported bumble bee deaths after a pyrethroidFootnote14 insecticide was applied to oilseed rape. Exposure to low doses of SpinosadFootnote15 has been reported to cause queen mortality in native bumble bees in Oregon (Skyrm et al. 2011 in Jepsen et al. 2013). Contact insecticides can also have important sublethal effects. For example, at concentrations that are likely to be encountered on pollen in the wild, Spinosad decreased the foraging efficiency of adult workers that were exposed to this pesticide during the larval stage (Morandin et al. 2005). Mommaerts et al. (2006) also reported a dramatic reduction in brood production of Bombus terrestris when these bumble bees were fed pollen treated with chitinFootnote16 synthesis inhibitors (e.g., diflubenzuron and teflubenzuron) at the maximum field recommended concentration. Adjuvants and other inert ingredients used with the active ingredients in contact insecticides (and, more broadly, pesticides) can have sublethal effects as well. For example, Mullin et al. (2015) showed that honey bees are sensitive to a surfactant and a solvent that are widespread ingredients in agricultural chemicals such as insecticides and other pesticides.
Herbicides
Herbicides, particularly broad-spectrum herbicides used to control weeds, can indirectly affect pollinators by reducing the quality of suitable habitat through the elimination of flowering plants that provide pollen and nectar for bumble bees (Williams 1986; Shepherd et al. 2003; Pleasants and Oberhauser 2012 in Jepsen et al. 2013). Jepsen et al. (2013) state that the reduction in these resources due to herbicides use can cause a decline in bumble bee reproductive success and/or survival rates. Pleasants and Oberhauser (2012) report that the increased use of the herbicide glyphosate (RoundupTM) in agricultural areas has likely led to the reduced availability of wildflowers in field margins, which otherwise could have been an important resource for the Rusty-patched Bumble Bee. Kearns et coll (1998) also state that, in some circumstances, herbicides appear to have a greater effect than insecticides on wild bee populations.
Fungicides
Until recently, fungicides were viewed as non-toxic or only slightly toxic to bees (Johansen et al. 1983; Mayer and Lunden 1986). However, it has been shown that the intensive use of fungicides can be toxic to honey bees and native bees (Mullin et al. 2010; Park et al. 2015) and that frequent exposure to high doses of fungicides can have a significant impact on populations (Park et al. 2015). With respect to native bees, the precise mode of action of fungicides remains unclear. However, reduced fitness (e.g., decreased foraging efficiency or nest recognition) and a reduction in beneficial fungi in pollen or “bee bread” are among the suspected mechanisms (Yoder et al. 2013; Park et al. 2015). Fungicides can also have a synergistic toxic effect in conjunction with other pesticides (Pilling and Jepson 1993; Sanchez-Bayo and Goka 2014), or they may be combined with ingredients that are considered inert but can have toxic effects on bees (Ciarlo et al. 2012; Mullin et al. 2015).
Invasive non-native/alien species – Pathogen transmission and spillover
Commercial non-native bumble bee species and other bee species that are used in Canada for honey production and for pollination of fruit and vegetable crops can act as vectors of pathogens that are harmful to the Rusty-patched Bumble Bee. The increased use of imported bumble bees for pollination in greenhouses has been linked to the decline of Bombus species, including the Rusty-patched Bumble Bee (Colla et al. 2006; Berenbaum et al. 2007; Cameron et al. 2011a). The transmission of pathogens by exotic bees and bumble bees can occur through various mechanisms, i.e., through the introduction and spread of exotic pathogens (spillover mechanism), through an increase in the prevalence of naturally occurring parasites (spillback mechanism), or through facilitation of pathogen transmission associated with resource sharing or competition for resources (facilitation mechanism) (Graystock et al. in press). Pathogens may be transmitted directly through contact as well as indirectly through a shared resource that is contaminated. For example, host insects infected by pathogens can contaminate flowers as they feed on them. Another pollinator can in turn become infected when it forages on the same flower. Non-host insects can likewise act as vectors by spreading parasites from flower to flower (Graystock et al. 2015).
Three pathogens found in Canada to date could be especially harmful to the Rusty-patched Bumble Bee. A study conducted in southern Ontario showed that commercial bumble bees transmitted pathogens, specifically the unicellular fungus Nosema bombi and the protozoan Crithidia bombi, to wild bumble bees foraging nearby (Colla et al. 2006). This type of transmission has since been confirmed by other studies (e.g., Graystock et al. 2014). The unicellular fungus N. bombi can reduce colony fitness as well as reduce individual reproduction rate and life span in bumble bees (Jepsen et al. 2013). C. bombi can dramatically reduce bumble bee longevity and colony fitness (Brown et al. 2003; Otterstatter and Whidden 2004 in Jepsen et al. 2013), interfere with learning among bumble bee foragers (Otterstatter et al. 2005 in Jepsen et al. 2013) and decrease pollen loads carried by workers (Shykoff and Schmid-Hempel 1991). The scope of the threat that these two pathogens pose to the species is closely tied to the presence of greenhouses that use insect pollinators. Graystock et al. (2013b) demonstrated that pathogens were present in bumble bee colonies imported to Great Britain for pollination purposes, in spite of control measures designed to ensure that the colonies would be pathogen-free. Honey bees (Apis mellifera) appear to be another vector allowing the transmission of N. bombi towild bumble bees. Graystock and Hugues (2014) showed that, in Great Britain, the prevalence of N. bombi was 18% greater in bumble bees near an apiary than in those farther away from it. No research has been done so far to check whether this type of spillover occurs in the Canadian population of the Rusty-patched Bumble Bee. The fungus Nosema ceranea, an Asian parasite of honey bees, was detected in bees for the first time in Canada in 2006 (Geoffrey et al. 2008), raising new concerns. A study done in Great Britain showed that wild bumble bees have been infected with N. ceranea. Laboratory tests indicate that this parasite can reduce the survival of bumble bees by 48% and can affect their behaviour (Graystock et al. 2013a). There have been no studies to detect the presence of N. ceranea in the Canadian population of the Rusty-patched Bumble Bee.
Yet other pathogens documented in other parts of the world could have an impact on the Rusty-patched Bumble Bee in Canada. The protozoan Apicystis bombi may have caused the population collapse observed in a native bumble bee species in Argentina, where it appears to have been introduced from Europe with imported colonies of the Buff-tailed Bumble Bee (Arbetman et al. 2013). The presence of this parasite has been recorded in populations of over 20 bumble bee species in Europe and North America as well as in the honey bee (Arbetman et al. 2013). Many other pathogens, including RNA viruses (Manley et al. 2015; McMahon et al. 2015), should also be studied.
Climate change and severe weather
Climatic parameters, such as temperature and precipitation, have strong effects on the distribution and composition of bumble bee fauna (e.g., Iserbyt and Rasmont 2012; Lecocq et al. 2013). Many studies worldwide have indicated that climate change is contributing to the current declines in bumble bee species (e.g., Bartomeus et al. 2013a). The Rusty-patched Bumble Bee, which is believed to have a restricted climatic niche (Colla and Taylor-Pindar 2011), may be especially sensitive to the effects of climate change in Canada since bumble bee species tend to be more vulnerable to extinction if they have narrow climatic tolerances and are close to the limit of their range (Williams et al. 2009). Climate change may cause adverse effects on bumble bees either through an increase in the frequency of extreme events (severe weather) or through gradual changes in average weather conditions (Jepsen et al. 2013; Rasmont et al. 2015).
Given their nature (e.g., intense, sporadic), severe weather events mostly affect bumble bee populations through local extinctions (Rasmont et al. 2015). For example, an analysis of data from four European countries has shown that bumble bees are sensitive to heat waves and droughts (Rasmont and Iserbyt 2012). Results of a study conducted by Harder (1986) on Amherst Island, Ontario indicated that underground nesting bumble bee species (including B. affinis) are particularly vulnerable to abnormally wet spring weather. Jepsen et al. (2013) reported that extreme precipitation and temperature events could be detrimental to the Rusty-patched Bumble Bee. Aside from direct mortality, such changes may also lead to a higher prevalence of pathogens and a decrease in resource availability (floral resources, and nesting and overwintering sites) (Cameron et al. 2011b).
Gradual change in average weather can likewise have significant impacts on bumble bees. The meta-analysis carried out by Kerr et al. (2015), using observations for 67 bumble bee species collected from Europe and North America over 110 years, revealed a cross-continentally consistent trend of range losses from the southern limits of their historical range (up to 300 km since 1974). The study also found that species with southern geographical ranges retreated to higher elevations where possible, and that in the case of more northerly species, losses from their southern range limits tend not to be offset by a northward expansion. None of the currently available data make it possible to determine what proportion of the reduction in the Rusty-patched Bumble Bee’s range is attributable to climate change. Kerr et al. (2015) suggested that climate change represents a long-term threat to the species. In addition, it has been hypothesized that climate-driven changes in phenologyFootnote17 could have a negative impact on the Rusty-patched Bumble Bee (COSEWIC 2010; Colla and Taylor-Pindar 2011). Bartomeus et al. (2011) observed significant phenological changes in 10 generalist bee species of northeastern North America. Their data show that the phenology of these bee species has advanced by a mean of just over 10 days in the past 130 years, with most of this advance occurring since the 1970s. This shift has been accompanied by parallel advances in the phenology of their plant resources, suggesting that bee emergence (generalist bees, at least) has kept pace with host-plant flowering. For the recovery of the Rusty-patched Bumble Bee, the diversity of flowering plants that flower at different times of year allows the protection of the species from potential mismatches between the timing of flowering and periods when the species needs to feed on flowers (Bartomeus et al. 2013b; Solga et al. 2014).
Annual and perennial non-timber crops – Intensive agriculture
The increased reliance on intensive agricultureFootnote18 has resulted in decreased quality foraging habitat for bumble bees in many countries (COSEWIC 2010). In Europe, declines in bumble bee populations over the past 60 years have been driven primarily by habitat loss and decreases in floral abundance and diversity resulting from agricultural intensification (Goulson et al. 2008).
Canada experienced an intensification of agricultural operations over the same time period (Agriculture and Agri-Food Canada 2013). Intensive farming monopolizes large areas of land to grow plant species which, for the most part, have a very brief flowering period that does not meet the needs of bumble bees, which require a continuous succession of flowers throughout the colony cycle (Goulson et al. 2008). Spring is the most critical time because the queen has to find enough pollen and nectar to feed the first brood of workers, on which the colony’s survival will depend. However, very few cultivated plant species flower early in the season (Bohart and Knowlton 1953; Alford 1975 in Goulson et al. 2008). Goulson et al. (2008) reported that farms that have large areas devoted to crops but no field margins or other areas with wildflowers cannot support the development and maintenance of bumble bee colonies.
Owing to the particular foraging strategy adopted by certain bumble bee species, the impacts of intensive farming may vary with the type of crop plants grown. For example, the Buff-tailed Bumble Bee, which is closely related to the Rusty-patched Bumble Bee, prefers to use large expanses of flowering plants offering abundant food resources rather than isolated patches of forage crops in agricultural landscapes (Walther-Hellwig and Frankl 2000). This species may even forego resources located near the nest and travel considerable distances to exploit specific crops of flowering plants (e.g., sunflower [Helianthus sp.], mustard [Synapis arvensis], and bean [Vicia sp.]) (Westphal et al. 2006). Large areas of crops that are not used by the species (e.g., corn [Zea mays) have significant greater adverse effects. Conservation or development of uncropped areas (e.g., hedgerows, field margins and borders of streams) in agricultural landscapes where the species can forage on flowering plants is of paramount importance for maintaining bumble bee colonies (Goulson et al. 2008; Jönsson et al. 2015; Williams et al. 2015; Wood et al. 2015). However, these floral resources growing near intensively farmed crops may pose a risk to the Rusty-patched Bumble Bee, because their pollen may be contaminated with pesticides (David et al. 2016). Intensive agriculture has significant indirect effects, considering the widespread use of pesticides in intensive cropping systems (see the threat “Agricultural and forestry effluents – Pesticide use”).
Housing and urban areas – Urban and suburban development
The Rusty-patched Bumble Bee’s Canadian range is located in the most highly populated and urbanized regions of the country. Because of habitat loss and fragmentation resulting from urban development, the species may have greater difficulty finding the various types of habitat (nesting, overwintering, and foraging) it needs during its life cycle (COSEWIC 2010). Conversion of land for urban and suburban development continues to transform and fragment habitat, a situation that has likely had an adverse effect on populations of bumble bee species, including the Rusty-patched Bumble Bee (Jepsen et al. 2013).
However, it is important to add that a variety of studies mention the existence of large populations of bumble bees in components of the urban landscape where there is an abundance of flowering plants (e.g., community and botanical gardens, urban wild areas, flower beds containing native or non-native flower species, managed or natural urban parks, green roofs) (Chapman et al. 2003; Tommasi et al. 2004; Colla et al. 2009; Matteson and Langellotto 2009). Urban habitats could support large populations of bumble bees and could play a valuable role in bumble bee conservation (Chapman et al. 2003), even if they cannot ensure maintenance of the species diversity that was historically present (McFrederick and LeBuhn 2006). A lack of nest sites may, however, be a limiting factor for bumble bee populations in the urban environment (Goulson et al. 2008). At present, there is no specific information on the Rusty-patched Bumble Bee in urban areas, except some reports of the species’ presence in several Canadian cities during the 20th century (COSEWIC 2010).
Roads and railroads – Road network development
Road construction leads to fragmentation of insect populations and, for a number of species including bumble bees, creates barriers impeding their access to available food resources (Reck and van der Ree 2015; Bhattacharya et al. 2003). Creation of roadside habitat for pollinators along roads and highways is considered by many experts as a way to mitigate this threat (Reck and van der Ree 2015; US Department of Transportation 2015).
The road network also affects populations of insect pollinators through accidental mortality. A recent study conducted on a stretch of highway in Ontario examined the significance of this threat. The results showed that, among Hymenopterans,Footnote19 accidental mortality amounted to about 27 individuals, on average, per kilometre of road every day (Baxter-Gilbert et al. 2015). By extrapolating this finding to southern Ontario, the authors estimated that more than 133 million Hymenopterans are accidentally killed by vehicles every summer. They indicated that these values likely represent an underestimation, since they did not include insects that were ricocheted off the road, adhered to vehicles during the collision or were scavenged after the collision on the road. The authors of the study concluded that accidental road mortality can pose a significant threat to populations of Hymenopterans. The population of the Rusty-patched Bumble Bee, whose historical range encompasses some of the most densely populated and urbanized regions in the country, is very likely to be affected by this threat. For example, there were more than 35,000 km of roads in southern Ontario in 1995, which represents an increase of 400% compared with the road density in 1935 (Fenech et al. 2005).
Invasive non-native/alien species – Competition with honey bees
A number of studies have reported that the presence of honey bee hives reduces the availability of nectar and pollen for native bees due to competition between honey bees and native bumble bee species for floral resources (Anderson 1989; Paton 1990; Wills et al. 1990; Dafni and Shmida 1996; Paton 1996; Horskins and Turner 1999; Dupont et al. 2011 cited in Jepson et al. 2013; Torné-Noguera et al. 2015). The negative effects of this competition on bumble bees include reduced production of males and queens, smaller body size, male-biased sex ratio, and reduced pollen collection (Thomson 2004; Thomson 2006; Goulson and Sparrow 2009; Elbgami et al. 2014). Honey bees may also displace some bumble bee species when they are foraging in the same area (Walther-Hellwig et al. 2006). It appears that larger Hymenoptera (such as the Rusty-patched Bumble Bee) are more likely to be adversely affected by honey bee competition (Torné-Noguera et al. 2015). A high density of honey bee hives near a local Rusty-patched Bumble Bee population could threaten that population’s long-term survival (Jepsen et al. 2013).
Livestock farming and ranching – Livestock grazing
Studies have shown that livestock farming can have both direct and indirect effects on bumble bee populations. Indirect effects include the removal of floral resources (Jepsen et al. 2013) and the potential reduction of rodent populations (Bueno et al. 2011), which can lead to a decrease in the number of nest sites available for bumble bees. Livestock can also have direct effects by trampling above-ground nests (Sugden 1985). The type of habitat, the diversity of flowering plants, the livestock species as well as the timing, intensity and duration of grazing are all factors that can influence how this practice affects flora and fauna (Gibson et al.1992; Carvell 2002; Sjodin 2007 in Jepsen et al. 2013; Orford et al. 2016), including the Rusty-patched Bumble Bee.
Fire and fire suppression – Fire suppression
Occasional natural fires play a role in the maintenance of herbaceous vegetation in fields and grasslands and help prevent the establishment of trees and shrubs. Fire suppression can lead to major changes in vegetation structure. In addition to reducing the amount of habitat available for bumble bees, encroaching forest can block corridors between fields, thereby interfering with bumble bee dispersal and foraging activities (Roland and Matter 2007). Prescribed fire, which is used to restore native prairie and meadow vegetation, can be beneficial for bumble bees, provided that burns are planned and implemented carefully, taking into account the life-history needs of bumble bees (Jepsen et al. 2013). The prescribed burn activities carried out annually in Pinery Provincial Park (Farrel et al. 2011) should respect these conditions.
Potential threats
Atmospheric pollutant – aluminum
Aluminum is the most abundant metal in the lithosphereFootnote20 and a major environmental contaminant (Exley 2013; Exley et al. 2015). It is extracted from the lithosphere and released into the biosphere either directly (e.g., extraction and processing of aluminum ore) or indirectly (e.g., acid depositionFootnote21 resulting from coal burning, and cultivation of acid sulphate soilsFootnote22) (Sammut and Lines-Kelly 2004; Agency for Toxic Substances and Disease Registry 2008; Exley 2013). A recent study in the United Kingdom revealed the presence of elevated concentrations of aluminum in pupae of the Buff-tailed Bumble Bee, which points to significant exposure of the species to this metal (Exley et al. 2015). These authors indicate that bees are likely exposed to aluminum that is present in pollen (Morgano et al. 2010) and possibly nectar (Meindl and Ashman 2013). Exley et al. (2015) state that it is conceivable that the high aluminum levels measured in bumble bee pupae could interfere with the development or functioning of cognitive performance in adults. They conclude that this finding points to the possibility that cognitive dysfunction is playing a role in the decline of bee populations.
Diesel exhaust
Predictions from theoretical models indicate that certain atmospheric pollutants could reduce the distance from the source at which pollinators can detect floral scent signals (McFrederick et al. 2008). Girling et al. (2013) conducted a laboratory study on the effects of diesel exhaust pollution on honey bees and found that exhaust gases reduce the bees’ ability to recognize floral odours. They showed that the mono-nitrogen oxide (NOx) fraction of the exhaust gases could play a key role in this odour degradation. The authors concluded that the interference with communication signals caused by the compounds in exhaust gases may be harmful to a number of insect species and that, in the case of pollinating insects, there could be major economic and ecological repercussions, particularly if the effects are combined with other factors harmful to pollinator health.
5. Population and Distribution Objectives
The Canadian population of the Rusty-patched Bumble Bee is very small, according to the current knowledge. In addition, the possibility of rescue effect through immigration from the United States population is very limited. For these reasons and based on the criteria that Environment and Climate Change Canada uses to determine recovery thresholds, it would be unrealistic to set population and distribution objectives that go beyond the minimum recovery threshold. This threshold represents the best achievable scenario for the Rusty-patched Bumble Bee.
The population and distribution objectives for the Rusty-patched Bumble Bee in Canada are to:
- ensure the viabilityFootnote23 of the local population in Pinery Provincial Park and of any other local population that might be discovered in the future, and
- as needed, increase the number of viable local populations in the species' current and historical range, in order to form in the long term and to the extent possible a species' distribution that is not severely fragmented as a result of non-natural causes.
The only evidence of the recent presence of the Rusty-patched Bumble Bee in Canada consists of sightings of the species in Pinery Provincial Park in Ontario. These sightings show that the park contained one colony of the species in 2005 and at least one colony, and possibly two colonies, in 2009. They also indicate that the local population in Pinery Provincial Park is very small, and therefore at high risk of extinction. Since this local population is the only one that confirms the species' recent presence in Canada, its viability should be ensured by increasing the population size (i.e., increase in number of colonies), for example, through threat reduction and mitigation and through habitat management (for more information, see sections 6.2 and 6.3).
Ensuring the viability of any other local population that might be discovered in the future is also necessary in order to ensure redundancyFootnote24 of the Canadian population. Although currently it is not certain that the Rusty-patched Bumble Bee is in fact present anywhere else in Canada other than in Pinery Provincial Park in Ontario, the species could be present in other parts of its historical range which have not been recently surveyed, especially in Quebec.
The objectives also include, as needed, increasing the number of viable local populations in the species' current and historical range. This part of the objectives includes the species' distribution objective. The available information indicates that the Canadian population of the species may currently have very low resilience.Footnote25 The discovery of other local populations might be sufficient to correct this situation; however, if this does not occur, it will be necessary to increase the number of viable local populations through natural expansion (e.g., through threat reduction and mitigation and through habitat management; for more information, see sections 6.2 and 6.3). This approach will also contribute to redundancy and help to ensure that the species' representation is in keeping with its historical range by endeavouring to capture the full range of its genetic and ecological diversity.
The last part of the objectives specifies that increasing the number of local populations is aimed at obtaining, over the long term and to the extent possible, a species' distribution that is not severely fragmented as a result of non-natural causes. This characteristic is necessary to ensure the persistence of local populations and maintenance of the species' genetic cohesion.
Given that the recovery feasibility for the Rusty-patched Bumble Bee is affected by uncertainty with regard to the possibility of maintaining or increasing population size and preventing or mitigating the main threats to the species and its habitat, the population and distribution objectives may not be attainable.
6. Broad Strategies and General Approaches to Meet Objectives
6.1 Actions Already Completed or Currently Underway
The only confirmed Canadian population of the Rusty-patched Bumble Bee is located in a Natural Environment class park, designated by the Government of Ontario, whose mandate includes “protection of natural heritage.” A long-term strategy aimed at rehabilitating the savannah habitats within the park has been implemented. The actions taken include prescribed burns, some of which were carried out between 1986 and 1994 (Rodger 1998), and again in 2001. Farrell et al. (2011) reported that prescribed burns are now carried out annually in the park to re-establish and manage savannah habitat.
Since the publication of the COSEWIC status report, surveys have been carried out annually in Pinery Provincial Park, but no Rusty-patched Bumble Bees have been detected (A. MacKenzie pers. comm. 2015). This monitoring is supported by the Wildlife Preservation Canada Native Pollinator Program, which initiated an Ontario Native Pollinators program in 2012. The project, undertaken in collaboration with York University, aims to monitor the Rusty-patched Bumble Bee and assess habitat quality, parasite levels and pesticide use across southern Ontario. Under this program, captive rearing and releases of another declining bumble bee species, the Yellow-banded Bumble Bee (Bombus terricola), began in Ontario in 2014, partly to provide protocols and techniques that could be beneficial for the Rusty-patched Bumble Bee. Other reintroduction initiatives have been launched elsewhere in the world (e.g., The Short-haired Bumble Bee Project).
In 2011, Farms at Work initiated a Rusty-patched Bumble Bee Project to raise awareness about native pollinator conservation issues and support surveys for the species in Ontario. This organization has created a Landowners Guide to Conserving Native Pollinators in Ontario (Chan 2012), initiated a Pollinator Habitat on Farms Project, and offers pollinator consultation services (see website for details).
Best management practices for agriculture (Agriculture, pêcheries et al.mentation Québec 2013; Health Canada 2013a and 2013b), and for creating and managing bumble bee habitat (Colla et al. 2009; Hatfield et al. 2012; Blackmore and Goulson 2014) have been developed.
In 2014, the Xerces Society for Invertebrate Conservation launched the Bumble Bee Watch website). This is a citizen science project that was initiated to track and conserve native North American bumble bees. The website allows users to upload photographs of bumble bee sightings to be identified and verified by experts. This project also helps researchers determine the status and conservation needs of species, helps locate rare or endangered populations and engages the public in learning about bumble bee ecology.
A recovery strategy for the Rusty-patched Bumble Bee in Ontario was developed (Colla and Pindar-Taylor 2011) and the Government of Ontario published a response statement, which constitutes its policy response to the scientific advice set out in the strategy (OMNR 2011). In January 2014, a section was also added to the ESA Ontario regulation 242/08. This section prescribes the areas that are protected as the species' habitat (Ontario Regulation 242/08).
In the spring of 2014, Health Canada implemented measures to reduce honey bee exposure to pesticides from corn and soybean (Glycine max) seed dust. A complete analysis is not yet available, but information gathered to date indicates that the numbers and severity of incident reports associated with neonicotinoid pesticide use during the planting period in 2014 are 70% lower than in 2013. Health Canada's Pest Management Regulatory Agency (PMRA) continues to work with the provinces and stakeholder groups to reduce honey bee exposure during planting of corn and soybeans. Pending the results of this work, additional regulatory measures may be taken if warranted and if supported by the available science. The PMRA is also continuing its re-evaluation of various classes of pesticides, including the neonicotinoid class, in collaboration with the United States Environmental Protection Agency and the California Department of Pesticide Regulation. As a part of this evaluation, the potential for both acute and sublethal effects on pollinators will be assessed by considering available information from scientists and researchers as well as new studies being generated to specifically address these questions (Health Canada 2014). Although they target honey bees, these measures are beneficial for all native pollinators, including the Rusty-patched Bumble Bee.
In May 2015, the Standing Senate Committee on Agriculture and Forestry published a report outlining a strategy for preserving the health of insect pollinators in Canada (Government of Canada 2015). The report first discusses honey bees because of their critical role in the pollination of crops, which makes their contribution to human food resources extremely important. It also describes native pollinators (800 species in Canada) and points out that they often have unique characteristics which make them specialist pollinators of particular plant species. They therefore play a vital role in the reproduction of the native plant species with which they are linked. The report also mentions that wild pollinators, owing to their morphological characteristics, are sometimes much more effective than honey bees at pollinating certain crops; however, there are no data available on this topic. Recommendations for preserving the health of pollinators (e.g., implementation of the biosecurity standard,Footnote26 re-evaluation of neonicotinoid insecticides by the Pest Management Regulatory Agency, and funding of research on insect pollinator health) are provided.
During the summer of 2015, a Quebec organization called Éco-NatureFootnote27 began a project dedicated to the Rusty-patched Bumble Bee. The first stage, undertaken that summer, involved developing and testing tools for carrying out surveys (capture, identification, etc.). Over the coming years, surveys (historical sites and potential habitat) in Éco-Nature's area of intervention will be carried out, along with public awareness activities (A. Boutin pers. comm. 2015).
In July 2015, the Ontario government announced measures aimed at bringing about an 80% reduction in the number of acres planted with neonicotinoid-treated seed by 2017. In 2016, farms will be able to use neonicotinoid-treated seed on up to 50% of their crop area; however, they will have to prove they have a pest problem requiring this type of treatment. In Quebec, the sale and use of pesticides is regulated under the Pesticides Act (CQLR, c. P-9.3) and, as a complement, under the Environment Quality Act (CQLR, c. Q-2). In 2015, the Quebec Department of Sustainable Development, Environment and the Fight Against Climate Change (MDDELCC) developed the Quebec Pesticide Strategy 2015-2018 (MDDELCC 2015) to guide the Quebec government's legislative and regulatory amendments over the coming years, in part, in order to conserve populations of pollinators. The objectives of the strategy include banning the use of neonicotinoids for ornamental purposes (e.g., lawns, flower beds), requiring that an agronomist approve all uses of these insecticides for agriculture, and promoting the use of non-neonicotinoid seed through the introduction of economic incentives.
6.2 Strategic Direction for Recovery
| Threat or Limiting Factor | Broad Recovery Strategy | PriorityNoteh | General Description of Research and Management Approaches |
|---|---|---|---|
| Knowledge gaps | Surveys and monitoring | High |
|
| All threats and Knowledge gaps | Management of individuals and habitat | High |
|
| All threats and knowledge gaps | Research | Medium to High |
|
| All threats and knowledge gaps | Research | Medium |
|
| All threats | Partnership, communication, outreach and education | Medium |
|
6.3 Narrative to Support the Recovery Planning Table
Surveys and monitoring
There are significant knowledge gaps related to the population and distribution of the Rusty-patched Bumble Bee in Canada. Survey activities undertaken to fill these gaps are essential for assessing the current population size, for determining the distribution of the population, and for confirming whether the number of local populations needs to be increased. A survey protocol and monitoring plan to be developed and implemented in suitable habitat within the species' historical range based on the data collected in Canada since 1975, taking care not to duplicate the recent surveys carried out in Ontario. Special attention should be given to the portion of the range located in Quebec, where no targeted surveys have been carried out since the mid-1970s. It should be mentioned that current colonies of the species may be relatively far from the sites where the historical observations were made. The Rusty-patched Bumble Bee is a generalist forager species that uses a wide variety of habitats. Furthermore, queens that emerge in the spring can travel several kilometres to found new colonies (Lepais et al. 2010). Therefore, surveys must be planned at the scale of the region surrounding the sites where historical observations were made, rather than at the scale of the sites themselves. A standardized monitoring method should also be developed and implemented periodically. These monitoring activities are necessary to determine the population trends for the species in Canada and to measure progress toward recovery. Survey and monitoring activities should also permit the population modelling needed to determine the viability of local populations of the species and of the Canadian population.
Management of individuals and habitat
The development and implementation of a habitat management plan taking into account all local populations of the Rusty-patched Bumble Bee is a key element for reaching the population and distribution objectives of this recovery strategy. This plan, which could be an existing habitat management plan (e.g., Pinery Provincial Park), should include maintaining or increasing the availability of the three types of habitat required to meet the species' life-history needs, with a focus on maintaining foraging habitat, which is under the greatest threat. The plan should also provide for the availability of suitable habitat needed for the expansion of the local population in Pinery Provincial Park or any other local population.
A threat reduction and mitigation plan should also be developed and implemented, taking into account the specific context of each local population. It will focus on the main threats to the Rusty-patched Bumble Bee (i.e., pesticide use, pathogen transmission and spillover, climate change and severe weather events, intensive agriculture, urban and suburban development, and the road network) and will include all appropriate measures (e.g., legislative, stewardship, best management practices) to reduce and mitigate these threats. The implementation of best management practices by landowners, land-use planners, and other stakeholders (e.g., farmers, greenhouse and nursery owners, urban planners, retail outlets that sell plants) located within important areas for the species' recovery is of key importance for reducing and mitigating these threats as well as threats of lower concern, such as livestock farming, fire suppression and competition with honey bees.
This strategy also includes an assessment of the feasibility of restoring the Canadian population of the species and the associated risks. Restoration of the Canadian population is included based on the following facts:
- the Canadian population, currently represented solely by the local population in Pinery Provincial Park, faces a high risk of extinction because of its very small size;
- natural expansion may not be sufficient to increase the size of this local population or any other local population to a level where its viability will be ensured, or to increase the number of local populations to the point where they are interconnected.
Based on the IUCN's Guidelines for Reintroductions and Other Conservation Translocations (IUCN 2013), the Rusty-patched Bumble Bee may be a good candidate for population restoration considering its high level of risk and its ecological role (pollinator). However, taking into account these guidelines and current conditions in Canada, population restoration is not recommended in the short term, for the following reasons:
- there is no evidence that the threats that caused past extinctions have been reduced sufficiently;
- the risks associated with such a restoration have not been assessed;
- there are gaps in the knowledge required to support decision making (e.g., threats, suitable habitat characteristics and availability).
During the period required to ensure that the threats to the habitat and individuals are sufficiently reduced, it will be imperative to assess the feasibility of population restoration and the associated risks. The results of this assessment will be used to determine whether population restoration can be undertaken, in the event that it is shown that natural expansion is insufficient. The feasibility assessment should include aspects such as availability of suitable habitat, climatic requirements, the availability and source of mated queens, captive rearing and release techniques, alternative recovery approaches, etc. Risk assessment should consider the benefits and the potential negative impacts related to ecological (e.g., risks to source populations or ecosystems), social (e.g., multiple jurisdictions), and economic aspects. The IUCN's Guidelines for Reintroductions and Other Conservation Translocations (IUCN 2013) should be used as a basis for these evaluations, along with information available from programs already underway elsewhere in the world on endangered bumble bee species (see section 6.1).
Research
Studies should be carried out to develop a better understanding of the threats to the species, with emphasis on the most significant threats. This includes research to determine the extent of contamination by pesticides (particularly neonicotinoids) in areas occupied and used by the species and to identify the lethal and sublethal effects of these substances on native bumble bee species. This research should be carried out only on species that are not currently declining.Footnote28 Research on pathogen transmission to native bees by bumble bees used for greenhouse pollination is also needed to better understand the importance of this threat to the Rusty-patched Bumble Bee and other native species. The modes of transmission and causal effects of these pathogens should also be examined. As well, research on the effects of climate change on the species is required, in particular to assess the contraction of the species' Canadian range. Studies aimed at developing best management practices for native bumble bees in agricultural, urban and suburban environments are needed to develop the threat reduction and mitigation plan mentioned above.Knowledge of the species' ecology and population dynamics should also be improved (e.g., foraging habitat needs, spatial ecology and response to restoration practices).
Partnership, communication, outreach and education
A communication strategy is needed to improve the efficiency of management approaches. This strategy will include means of communication and outreach aimed at specific target audiences (e.g., farmers, greenhouse owners). It would be beneficial to maintain or establish partnerships with local, regional and national organizations in order to capitalize on conservation programs and initiatives targeting the agroenvironment in general or native pollinators in particular. Partnerships with groups of field naturalists possessing the skills needed to assist with surveys could also be developed. It would also be beneficial to develop outreach tools (e.g., media, brochures, and websites) in order to raise awareness among the general public about the situation of pollinators in Canada.
This strategy also includes collaboration with the United States with regard to conservation of the species. The species' global situation is very precarious (NatureServe: critically imperiled – IUCN: critically endangered) and it falls to the two countries in its range--Canada and the United States--to improve this situation. Collaboration between Canada and the United States (e.g., exchange of information, coordinated and complementary efforts) will ensure greater efficiency and help to maximize the success of the conservation efforts devoted to the species.
7. Critical Habitat
Paragraph 41(1)(c) of SARA requires that recovery strategies include an identification of the species’ critical habitat, to the extent possible, as well as examples of activities that are likely to result in its destruction. Under SARA, critical habitat is “the habitat that is necessary for the survival or recovery of a listed wildlife species and that is identified as the species’ critical habitat in the recovery strategy or in an action plan for the species.”
The critical habitat of the Rusty-patched Bumble Bee is partially identified in this recovery strategy, to the extent possible, based on the best available information. This identification currently applies only to the population in Pinery Provincial Park in Ontario. It is acknowledged that the present identification of critical habitat does not allow the population and distribution objectives for the species to be attained. A schedule of studies (section 7.2; Table 5) has been developed which describes the activities required for the identification of additional critical habitat necessary to reach the population and distribution objectives. Additional critical habitat may be identified in the future as new information becomes available.
7.1 Identification of the Species' Critical Habitat
The critical habitat of the Rusty-patched Bumble Bee is identified on the basis of two criteria: habitat occupancy and habitat suitability.
7.1.1 Habitat occupancy
Valid sightings (i.e., made or validated by a species' expert) of individuals (including those indicating the presence of an occupied nest or overwintering site) form the basis for determining habitat occupancy by the Rusty-patched Bumble Bee. Only sightings made since 2005 will be used. This period is consistent with the requirements of Ontario Regulation 242/08 adopted under the ESA, and is considered relevant considering the species' annual life cycle, the dramatic range-wide declines in species abundance and distribution since the early 2000s, and survey results obtained during this period in Ontario.
7.1.2 Habitat suitability
Habitat suitability refers to the biophysical attributes of the habitat used by the species. The Rusty-patched Bumble Bee requires habitats with varying characteristics during its different life cycle stages. Little information is available on the biophysical attributes of the species' nesting and overwintering habitats. Consequently, the biophysical attributes of habitat for nesting and overwintering sites are not limited to the characteristics described below and could include any other type of structures or habitat used by the species.
- Nesting habitat includes, but is not limited to:
- old rodent burrows and nests;
- hollow tree stumps;
- fallen dead wood.
- Overwintering habitat includes, but is not limited to:
- sites suitable for underground burrows in loose soil;
- fallen dead wood.
- Foraging habitat includes:
- foraging opportunities (e.g., abundance of suitable wild flowering plant species) in the following community classes:Footnote29 savannah, woodland, forest, prairie, marsh, bog, sand dune, cultural (e.g., old field);
- known plant species used as food sources (a list of these species is provided in Appendix A of this recovery strategy).
The most recent sightings (since 2005) of the Rusty-patched Bumble Bee have been made in oak savannah habitat (Tallgrass SavannahFootnote29) which possesses the following characteristics:
- widely spaced, open-grown trees with a floristically diverse understory of prairie grasses and forbs;
- moderately open tree canopy (between 25% and 35%);
- well-drained sandy soils.
Suitable habitat is located within a 1,000-m radius around each valid sighting. The 1,000-m distance is consistent with Ontario Regulation 242/08 made under the ESA. It comes from an analysis performed by the Ontario Ministry of Natural Resources and Forestry based on the foraging distance reported for workers of the Buff-tailed Bumble Bee (Bombus terrestris) (Osborne et al. 1999; Walther-Hellwig and Frankl 2000; Chapman et al. 2003; Darvill et al. 2004; Knight et al. 2005; Kraus et al. 2009; Osborne et al. 2008; Wolf and Moritz 2008; Hagen et al. 2011). The Buff-tailed Bumble Bee was used since no data are available on the foraging distance of the Rusty-patched Bumble Bee. This species was chosen as a model because it is in the same subgenus as the Rusty-patched Bumble Bee, and it has been the object of numerous studies in North America and Europe. However, since no data are available on how much suitable habitat is needed to enable a colony of the Rusty-patched Bumble Bee to meet all its life cycle needs, it is unclear whether the amount of suitable habitat contained within a 1,000-m radius is sufficient to meet the population and distribution objectives. Other information which has become availableFootnote30 since this analysis (e.g., distance travelled by males and queens of other species, vulnerability to inbreeding depression) suggests that this distance may not be sufficient to meet these objectives. An activity has been included in the schedule of studies in order to determine the validity of this distance.
Given the key importance of nesting and overwintering sites for the Rusty-patched Bumble Bee, any habitat located within a 30-m radius of an occupied nesting or overwintering site is also considered suitable habitat. This area represents the site's critical function zone, or the minimum habitat area required to maintain the elements that characterize the microhabitat (e.g., soil drainage, heat absorption, availability of cover). The 30-m radius is in keeping with Ontario Regulation 242/08 adopted under the ESA. It is not certain that the 30-m radius is sufficient to circumscribe the minimum area required to maintain the microhabitat characteristics of Rusty-patched Bumble Bee nesting sites or overwintering sites. An activity has been included in the schedule of studies in order to determine the validity of this distance.
7.1.3 Application of the criteria for identifying critical habitat for the Rusty-patched Bumble Bee
Critical habitat for the Rusty-patched Bumble Bee corresponds to any habitat with the biophysical attributes described in section 7.1.2 that is located within a 1,000-m radius of any record of the species meeting occupancy criterion 7.1.1. Anthropogenic structures (e.g., homes, paved surfaces) and areas such as open-water areas and manicured lawns, which do not possess the biophysical attributes of suitable habitat for the Rusty-patched Bumble Bee, are not identified as critical habitat, except with respect to the critical function zone located within a 30-m radius of any valid record of an occupied overwintering or nesting site. In this critical function zone, anthropogenic structures and suitable and unsuitable habitat zones are identified as critical habitat.
When the critical habitat identification criteria were applied to the data available on the species (current to August 2014), the critical habitat for the three recent sightings (since 2005) of the Rusty-patched Bumble Bee in Canada was identified and its total area was estimated at ~498 haFootnote31 (Figure 2; see also Table 4). This identification of critical habitat is considered partial as the identified critical habitat is insufficient to meet the population and distribution objectives. Section 7.2 presents the schedule of studies which lists the activities to be undertaken to gather the information required to complete the identification of critical habitat for the Rusty-patched Bumble Bee. If new sites meeting the above-mentioned criteria are found, additional critical habitat will be identified in an updated recovery strategy or an action plan.
Critical habitat for the Rusty-patched Bumble Bee is presented using 1 x 1 km standardized UTM grid squares (Figure 2; Table 4). The UTM grid squares are part of a standardized grid system that indicates the general geographic areas containing critical habitat, which can be used for land-use planning and/or environmental assessment purposes. In addition, the grid complies with provincial data-sharing agreements in Ontario. The areas of critical habitat within each grid square occur where the description of critical habitat is met. More detailed information on critical habitat may be requested by contacting Environment and Climate Change Canada – Canadian Wildlife Service at ec.planificationduretablissement-recoveryplanning.ec@canada.ca.
Long description for Figure 2
Figure 2 shows the location of critical habitat for the species in Ontario. Sixteen grid squares are found along the coast of Lake Huron in Pinery Provincial Park.
| 1 x 1 km standardized UTM Grid Square IDNotei | Province/ Territory | UTM Grid Square CoordinatesNotej Easting |
UTM Grid Square CoordinatesNotej |
Land tenureNotek |
|---|---|---|---|---|
| 17TMH3818 | Ontario | 431000 | 4788000 | Non-federal |
| 17TMH3819 | Ontario | 431000 | 4789000 | Non-federal |
| 17TMH3828 | Ontario | 432000 | 4788000 | Non-federal |
| 17TMH3829 | Ontario | 432000 | 4789000 | Non-federal |
| 17TMH3838 | Ontario | 433000 | 4788000 | Non-federal |
| 17TMH3839 | Ontario | 433000 | 4789000 | Non-federal |
| 17TMH3910 | Ontario | 431000 | 4790000 | Non-federal |
| 17TMH3920 | Ontario | 432000 | 4790000 | Non-federal |
| 17TMH3930 | Ontario | 433000 | 4790000 | Non-federal |
| 17TMH3951 | Ontario | 435000 | 4791000 | Non-federal |
| 17TMH3941 | Ontario | 434000 | 4791000 | Non-federal |
| 17TMH3931 | Ontario | 433000 | 4791000 | Non-federal |
| 17TMH3952 | Ontario | 435000 | 4792000 | Non-federal |
| 17TMH3942 | Ontario | 434000 | 4792000 | Non-federal |
| 17TMH3932 | Ontario | 433000 | 4792000 | Non-federal |
| 17TMH3943 | Ontario | 434000 | 4793000 | Non-federal |
7.2 Schedule of Studies to Identify Critical Habitat
| Description of Activity | Rationale | Timeline |
|---|---|---|
Conduct surveys:
Surveys should be prioritized at the provincial level on the basis of habitat suitability, proximity to already identified critical habitat and the characteristics of the available data (e.g., year, precision of location), and such work should not be done at sites where recent research effort is considered sufficient. |
Since recent inventories were insufficient to determine the current population size and distribution of the species, it is important to carry out surveys in portions of the suitable habitat in locations within the historical range that have not been covered in recent surveys. These surveys will make it possible to determine if and where other local populations of the species exist. Locating extant populations that have not yet been surveyed and identifying their critical habitat is necessary in order to achieve the population and distribution objectives. This activity is needed to delineate the species' distribution (area of occupancy of the local population) in Pinery Provincial Park, which will make it possible to complete the identification of critical habitat in the area adjacent to the park, as necessary. |
2016-2021 |
| Determine the amount of suitable habitat that is required to enable a colony of the species to meet its life cycle needs and the spatial scale within which this habitat must be available to meet these needs. | This study is needed to ensure that the boundaries of critical habitat (see section 7.1.2) are adequate to ensure that each colony of the species can meet the needs of its different life cycle stages. The results of this study could lead to a need to increase the amount of critical habitat identified. | 2016-2021 |
| Within the Rusty-patched Bumble Bee's current and historical range, determine whether it is necessary to increase the number of local populations and, where applicable, identify the suitable habitat needed for the natural expansion of local populations. | After surveys are conducted (see first activity in table), it will be possible to determine whether it is necessary to increase the number of local populations. If this is the case, additional suitable habitat and, ultimately, additional critical habitat will be required to meet the population and distribution objectives. | 2016-2023 |
7.3 Activities Likely to Result in the Destruction of Critical Habitat
Understanding what constitutes destruction of critical habitat is necessary for the protection and management of critical habitat. Destruction is determined on a case-by-case basis. Destruction would result if part of the critical habitat were degraded, either permanently or temporarily, such that it would not serve its function when needed by the species. Destruction may result from a single activity or multiple activities at one point in time or from the cumulative effects of one or more activities over time. It should be noted that not all activities that occur in or near critical habitat are likely to cause its destruction.
Critical habitat for the Rusty-patched Bumble Bee may be destroyed by any alteration that adversely modifies any biological, chemical or physical feature to the extent that individuals can no longer use their environment for one of their life processes, such as overwintering, nesting or foraging. Within the critical habitat boundaries, activities that could ultimately alter the structure and composition of open habitats where suitable flowering plant species are available can destroy Rusty-patched Bumble Bee critical habitat. Overwintering and nesting habitats (see section 7.2 – Habitat suitability) are of prime importance because they restrict important population segments within a microhabitat for a substantial period of time. Little information exists on the biophysical attributes of nesting and overwintering habitat, and all occupied nesting and overwintering sites deserve special attention. Considering the species' ecological traits, the timing of activities has a particularly significant influence in relation to the likelihood of destruction.
Table 6 presents examples of activities that could result in the destruction of critical habitat. This list of activities is based on the threats assessed and described in section 4 (Threats) and should not be considered exhaustive or exclusive. For some activities, the identification of thresholds may lead to a refinement or more precise description of the aspects of a given activity that are likely to destroy critical habitat.
| Description of Activity | Description of Effect (biophysical or other characteristics) in Relation to Function Loss | Details of Effect |
|---|---|---|
| Application of insecticides (particularly neonicotinoids) within or near critical habitat, including areas where drift into critical habitat may occur. | Application of systemic insecticides (e.g., neonicotinoids) may result in direct loss or degradation of foraging habitat because they spread throughout the plant, from roots up through the vascular system to the floral parts (e.g., pollen and nectar) that are used by bumble bee workers. The reduced quality of foraging resources could result in a decrease in reproductive success and survival rates. The use of non-systemic (or contact) insecticides may result in the temporary destruction of critical habitat because the chemicals may drift onto the pollen and nectar of surrounding flowers, making them unsuitable as forage for Bumble Bees. |
If this activity were to occur within the boundaries of critical habitat at any time of year, it is likely that the effects on critical habitat would be direct and cumulative. If this activity were to occur outside the boundaries of critical habitat, it could destroy critical habitat, as the chemicals could drift or leach into critical habitat. The effects of this activity apply year-round because many pesticides are persistent; however, more serious effects could result when products are applied during the active colony period (March/April to October). |
| Application of herbicides within or near critical habitat, including areas where drift into critical habitat may occur. | Application of herbicides can directly eliminate suitable flowering plants used as forage or reduce the abundance of such plants. The reduction in foraging resources could lead to a decrease in reproductive success and survival rates. The use of broad-spectrum herbicides (e.g., glysophate) increases the likelihood of destruction of foraging habitat. | If this activity were to occur during the active colony period (March/April to October) within the bounds of critical habitat, it is likely that the effects on critical habitat would be direct and cumulative. If this activity were to occur outside the boundaries of critical habitat, it could destroy critical habitat, as the products used could drift or leach into critical habitat. |
| Activities that alter soil characteristics (e.g., removal, of woody debris, compaction, modification of drainage). | Activities that alter soil characteristics may cause habitat loss or permanent or temporary degradation of nesting and overwintering habitat, if the extent of alteration exceeds a critical threshold (this threshold remains to be determined). The risk of destruction of critical habitat is increased if the activities are carried out in the critical function zone of a nesting or overwintering site. | If this activity were to occur within the boundaries of critical habitat at any time of year, it is likely that the effects on critical habitat would be direct and cumulative. The effects of this activity are applicable at all times of the year. The effects of this type of alteration of nesting habitat would be more severe during the active colony period (March/April to October). By contrast, in overwintering habitat, the effects would be more severe during the period when queens are hibernating (November to February/March). |
| Mowing, brush clearing, tree cutting and livestock grazing. | Removal of plant species whose flowers are used as a food source by the Rusty-patched Bumble Bee can result in the loss or degradation of foraging habitat for variable periods of time, if the quantity removed exceeds a critical threshold (which remains to be determined). | If this activity were to occur within the boundaries of critical habitat during the active colony period (March/April to October), it is likely that the effects on critical habitat would be direct and cumulative, even if temporary in nature. |
| Large-scale construction such as housing or road network development. | This activity would eliminate foraging, nesting and overwintering habitat, resulting in complete and permanent loss of critical habitat or the fragmentation of remaining critical habitat. | If this activity were to occur within the boundaries of critical habitat at any time of year, it is likely that the effects on critical habitat would be direct and cumulative, as construction would result in the complete loss or fragmentation of critical habitat on which the species depends. |
8. Measuring Progress
Every five years, success of recovery strategy implementation will be measured against the following performance indicators:
- The local population of the Rusty-patched Bumble Bee in Pinery Provincial Park is viable;
- Other local populations of the species that have been discovered are viable;
- The number of viable local populations in the species' current and historical range has been increased, as needed;
- In the long term, and to the extent possible, the Canadian distribution of the species is not severely fragmented by human activity.
9. Statement on Action Plans
One or more action plans detailing the measures to be taken to implement this recovery strategy for the Rusty-patched Bumble Bee will be posted on the Species at Risk Public Registry within five years of the posting of the final version of this recovery strategy.
10. References
Agency for Toxic Substances and Disease Registry. 2008. Public Health Statement for Aluminum. Website: http//www.atsdr.cdc.gov/phs.asp?id=1076&tid=34 [accessed December 2015].
Agriculture and Agri-Food Canada. 2013. Environmental Sustainability of Canadian Agriculture. Environmental Indicator Report Series – Report No. 3. Website: http://agriculture.canada.ca/en/?id=1295901472640 [accessed August 2015].
Agriculture, pêcheries et al.mentation Québec. 2013. Protégeons les abeilles des applications de pesticides. Centre de recherche en sciences animales de Deschambault (CRSAD). 18 p.
Alford, D.V. 1975. Bumblebees. Davis-Poynter, London, United Kingdom. 352 p.
Anderson, G.J., and M.K. Anderson. 1989. Assaying pollinator visitation to Solanum flowers. Solanaceae Newsletter 3(1):71.
Arbetman, M.P., I. Meeus, C.L. Morales, M.A. Aizen, and G. Smagghe. 2013. Alien parasite hitchhikes to Patagonia on invasive bumblebee. Biological Invasions 15(3):489-494.
Bartomeus, I., J.S. Ascher, D. Wagner, B.N. Danforth, S. Colla, S. Kornbluth, and R. Winfree. 2011. Climate-associated phenological advances in bee pollinators and bee-pollinated plants. Proceedings of the National Academy of Sciences of the United States of America 108(51):20645-20649.
Bartomeus, I., J.S. Ascher, J. Gibbs., B.N. Danforth, D.L. Wagner, S.M. Hedtke, and R. Winfree. 2013a. Historical changes in northeastern US bee pollinators related to shared ecological traits. Proceedings of the National Academy of Sciences of the United States of America 110(12):4656-4660.
Bartomeus, I., M. Park, J. Gibbs, B. Danforth, A. Lasko, and R. Winfree. 2013b. Biodiversity ensures plant-pollinator phenological synchrony against climate change. Ecology Letters 16:1331-1338.
Baxter-Gilbert, J.H., J.L. Riley, C.J.H. Neufeld, J.D. Litzgus, and D. Lesbarrères. 2015. Road mortality potentially responsible for billions of pollinating insect deaths annually. Journal of Insect Conservation 19(5):1029-1035.
Berenbaum, M., P. Bernhardt, S. Buchmann, N.W. Calderone, P. Goldstein, D.W. Inouye, P.G. Kevan, C. Kremen, R.A. Medellin, T. Ricketts, G.E. Robinson, A.A. Snow, S.M. Swinton, L.B. Thien, and F.C. Thompson. 2007. Status of pollinators in North America. The National Academies Press, Washington, DC. 312 p.
Bhattacharya M., R.B. Primack, and J. Gerwein. 2003. Are roads and railroads barriers to bumblebee movement in a temperate suburban conservation area? Biological Conservation 109:37-45.
Blackmore, L., and D. Goulson. 2014. Evaluating the effectiveness of wildflower seed mixes for boosting floral diversity and bumblebee and hoverfly abundance in urban areas. Insect Conservation and Diversity 7(5):480-484.
Bohart, G.E., and G.F. Knowlton. 1953. Yearly population fluctuations of Bombus morrisoni at Fredonia, Arizona. Journal of Economic Entomology 45:890.
Brown, M.J.F., R. Schmid-Hempel, and P. Schmid-Hempel. 2003. Strong context-dependent virulence in a host-parasite system: reconciling genetic evidence with theory. Journal of Animal Ecology 72:994-1002.
Bueno, C, K.E. Ruckstuh, N. Arrigo, A.N. Aivaz, and P. Neuhaus. 2011. Impacts of cattle grazing on small-rodent communities: an experimental case study. Canadian Journal of Zoology 90:22-30.
Burns, I. 2004. Social development and conflict in the North American bumblebee Bombus impatiens Cresson. Ph.D. Thesis. University of Minnesota, Minneapolis, Minnesota.
Cameron, S.A., J.D. Lozier, J.P. Strange, J.B. Koch, N. Cordes, L.F. Solter, and T.L. Griswold. 2011a. Patterns of widespread decline in North American bumble bees. Proceedings of the National Academy of Sciences of the United States of America.108:662-667.
Cameron, S., S. Jepsen, E. Spevak, J. Strange, M. Vaughan, J. Engler, and O. Byers. 2011b. North American Bumble Bee Species Conservation Planning Workshop Final Report (PDF; 3.8MB). IUCN/SSC Conservation Breeding Specialist Group, Apple Valley, Minnesota. Website: [accessed September 2015]
Carvell, C. 2002. Habitat use and conservation of bumblebees (Bombus spp.) under different grassland management regimes. Biological Conservation 103:33-49.
Chan, S. 2012. A Landowners Guide to Conserving Native Pollinators in Ontario. Farms at Work and the Peterborough County Stewardship Council. 40 p. [accessed August 2014].
Chapman, R.E., and A.F.G. Bourke. 2001. The influence of sociality on the conservation biology of social insects. Ecology Letters 4:650-662.
Chapman, R.E., J. Wang, and A.F.G. Bourke. 2003. Genetic analysis of spatial foraging patterns and resource sharing in bumble bee pollinators. Molecular Ecology 12:2801-2808.
Ciarlo, T.J., C.A. Mullin, J.L. Frazier, and D.R. Schmehl. 2012. Learning impairment in honey bees caused by agricultural spray adjuvants. PLoS ONE 7:e40848.
Colla, S.R., M.C. Otterstatter, R.J. Gegear, and J.D. Thomson. 2006. Plight of the bumblebee: Pathogen spillover from commercial to wild populations. Biological Conservation 129:461-467.
Colla, S.R., and L. Packer. 2008. Evidence for the decline of Eastern North American Bumblebees, with special focus on Bombus affinis Cresson. Biodiversity and Conservation 17:1379-1391.
Colla, S.R., E. Willis, and L. Packer. 2009. Can Green Roofs Provide Habitat for Urban Bees (Hymenoptera: Apidae)? Cities and the Environment 2(1): article 4, 12 p.
Colla, S.R., and S. Dumesh. 2010. The Bumble Bees of Southern Ontario: Notes on Natural History and Distribution. Journal of the Entomological Society of Ontario 141:38-67.
Colla, S.R., and A. Taylor-Pindar. 2011. Recovery Strategy for the Rusty-patched Bumble Bee (Bombus affinis) in Ontario. Ontario Recovery Strategy Series. Prepared for the Ontario Ministry of Natural Resources, Peterborough, Ontario, vi + 21 p.
Colla, S.R., L. Richardson, and P. Williams. 2011. Bumble Bees of the Eastern United States. United States Department of Agriculture – Forest Service, Pollinator Partnership, 104 p.
Colla, S.R. 2012. The Ecology and Conservation of North American Bumblebees. Ph.D. Thesis, York University, Toronto, Ontario.
Colla, S. R., F. Gadallah, L. Richardson, D. Wagner, and L. Gall. 2012. Assessing declines of North American bumble bees (Bombus spp.) using museum specimens. Biodiversity and Conservation 2012(21):3585-3595.
Committee on the Status of Endangered Wildlife in Canada (COSEWIC). 2010. COSEWIC Assessment and Status Report on the Rusty-patched Bumble Bee (Bombus affinis) in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, Ontario. vi + 34 p.
Corbet, S.A., M. Fussell, R. Ake, A. Fraser, C. Gunson, A. Savage, and K. Smith. 1993. Temperature and the pollinating activity of social bees. Ecological Entomology 18:17-30.
Dafni, A., and A. Shmida. 1996. The possible ecological implications of the invasion of Bombus terrestris (L.) (Apidae) at Mt. Carmel, Israel. p. 183-200 in A. Matheson, S.L. Buchmann, C. O'Toole, P. Westrich and I.H. Williams (eds.). The Conservation of Bees. Academic Press, London, United Kingdom. 254 p.
Darvill, B., M.E. Knight, and D. Goulson. 2004. Use of genetic markers to quantify bumblebee foraging range and nest density. Oikos 107:471-478.
Darvill, B., O. Lepais, L.C. Woodall, and D. Goulson. 2012. Triploid bumblebees indicate a direct cost of inbreeding in fragmented populations. Molecular Ecology 21(16):3988-3995.
David, A., C. Botías, A. Abdul-Sada, E. Nicholls, E.L. Rotheray, E.M. Hill, and D. Goulson. 2016. Widespread contamination of wildflower and bee-collected pollen with complex mixtures of neonicotinoids and fungicides commonly applied to crops. Environment International 88:169-178.
Dempster, J.P., and I.F.G. McLean. 2012. Insect populations in theory and in practice: 19th Symposium of the Royal Entomological Society, 10–11 September 1997 at the University of Newcastle. Springer Science and Business Media, New York, New York. 486 p.
Dupont, Y.L., C. Damgaard, and V. Simonsen. 2011. Quantitative Historical Change in Bumblebee (Bombus spp.) Assemblages of Red Clover Fields. PLoS ONE 6(9): e25172. doi:10.1371/journal.pone.0025172.
Duquette, M., and A. Boutin. 2015. Développement de méthodes d'identification et d'inventaire du bourdon à tache rousse (Bombus affinis) à la rivière des Mille Îles. Report submitted to Environment Canada's Habitat Stewardship Program for Species at Risk. 10 p.
Elbgami, T., W.E. Kunin, W.O.H. Hughes, and J.C. Biesmeijer. 2014. The effect of proximity to a honeybee apiary on bumblebee colony fitness, development, and performance. Apidologie 45:504-513.
Evans, E., R. Thorp, S. Jepson, and S.H. Black. 2008. Status review of three formerly common species of bumblebee in the subgenus Bombus. Report prepared for The Xerces Society for Invertebrate Conservation. 63 p.
Evans, E. 2009. Red list assessment (PDF; 139KB) for Bombus affinis. [accessed September 2015]
Exley, C. 2013. Human exposure to aluminium. Environmental Science: Processes & Impacts 15:1807-1816.
Exley C., E. Rotheray, and D. Goulson. 2015. Bumblebee Pupae Contain High Levels of Aluminium. PLoS ONE 10(6):e0127665. doi:10.1371/journal.pone.0127665.
Farrell, T., C. Copeland, and C. Bahlai. 2011. Recovery Strategy for the Northern Barrens Tiger Beetle (Cicindela patruela) in Ontario. Ontario Recovery Strategy Series. Prepared for the Ontario Ministry of Natural Resources, Peterborough, Ontario. vi +18 p.
Fenech, A., B. Gaylor, R. Hansell, and G. Whitelaw. 2005. Changes in the major roads of southern Ontario, Canada 1935-1995: Implications for protected areas. P. 93-113 in A. Fenech, D. MacIver and H. Auld (eds.). Integrated Mapping Assessment. Meteorological Service of Canada, Environment Canada, Toronto, Ontario.
Frankham, R., J.D. Ballou, and D.A. Briscoe. 2002. Introduction to Conservation Genetics. Cambridge University Press, Cambridge, United Kingdom. 617 p.
Geoffrey, R.W., A.B.A. Shafer, R.E.L. Rogers, D. Shutler, and D.T. Stewart. 2008. First detection of Nosema ceranae, a microsporidian parasite of European honey bees (Apis mellifera), in Canada and central USA. Journal of Invertebrate Pathology 97:189-192.
Gibson, C., V. Brown, L. Losito, and G. McGavin. 1992. The response of invertebrate assemblies to grazing. Ecography 15:166-176.
Gill, R.J., O. Ramos-Rodriguez, and N.E. Raine. 2012. Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 491(7422):105-108.
Gill, R. J., and N.E. Raine. 2014. Chronic impairment of bumblebee natural foraging behaviour induced by sublethal pesticide exposure. Functional Ecology (28)(6): 1459-1471.
Girling, R.D., I. Lusebrink, E. Farthing, T.A. Newman, and G.M. Poppy. 2013. Diesel exhaust rapidly degrades floral odours used by honeybees. Scientific Reports 3:2779.
Goulson, D., and K. Sparrow. 2009. Evidence for competition between honeybees and bumblebees; effects on bumblebee worker size. Journal of Insect Conservation 13(2):177-181.
Goulson, D., G.C. Lye, and B. Darvill. 2008. Decline and conservation of bumblebees. Annual Review of Entomology 53:191-208.
Goulson, D. 2010. Bumblebees: Behaviour, ecology and conservation. 2nd edition. Oxford University Press, Oxford, United Kingdom. 317 p.
Goulson, D.C. 2013. An overview of the environmental risks posed by neonicotinoid insecticides. Journal of Applied Ecology 50:977-987.
Government of Canada. 2015. The Importance of Bee Health to Sustainable Food Production in Canada. The Standing Senate Committee on Agriculture and Forestry, Ottawa, Ontario. 41 p.
Graystock P., K. Yates, B. Darvill, D. Goulson, and W.O.H. Hughes. 2013a. Emerging dangers: deadly effects of an emergent parasite in a new pollinator host. Journal of Invertebrate Pathology 114:114-119.
Graystock, P., K. Yates, S. Evison, B. Darvill, D. Goulson, and W.O.H. Hughes. 2013b. The Trojan hives: pollinator pathogens, imported and distributed in bumblebee colonies. Journal of Applied Ecology 50:1207-1215.
Graystock, P., D. Goulson, and W.O.H. Hughes. 2014. The relationship between managed bees and the prevalence of parasites in bumblebees. PeerJ2:e522. doi:10.771/peerj.522.
Graystock, P., D. Goulson, and W.O.H. Hughes. 2015. Parasites in bloom: flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proceedings of the Royal Society B 282:20151371.
Graystock, P., E.J. Blane, Q.S. McFrederick, D. Goulson, and W.O.H. Hughes. In press. Do managed bees drive parasite spread and emergence in wild bees? International Journal for Parasitology: Parasites and Wildlife. doi:10.1016/j.ijppaw.2015.10.001.
Hagen, M., M. Wikeslki, and W.D. Kissling. 2011. Space use of bumblebees (Bombus spp.) revealed by radio-tracking. PLoS One 6(5):e19997. doi:10.1371/journal.pone.0019997.
Harder, L.D.1986. Influences on the density and dispersion of bumble bee nests (Hymenoptera, Apidae). Holarctic Ecology 9:99-103.
Hatfield, R.G., and G. LeBuhn. 2007. Patch and landscape factors shape community assemblage of bumble bees, Bombus spp. (Hymenoptera: Apidae), in montane meadows. Biological Conservation 139:150-158.
Hatfield, R., S. Jepson, E. Mader, S. Hoffman Black, and M. Shepherd. 2012. Conserving Bumble Bees. Guidelines for Creating Habitat for America's Declining Pollinators. The Xerces Society for Invertebrate Conservation, Portland, Oregon. 32 p.
Horskins, K., and V.B. Turner. 1999. Resource use and foraging patterns of honeybees, Apis mellifera, and native insects on flowers of Eucalyptus costata. Australian Journal of Ecology 24:221-27.
Health Canada. 2013a. Pollinator Protection: Reducing Risk from Treated Seed, Health Canada, Pest Management Regulatory Agency. [accessed August 2014].
Health Canada. 2013b. Protecting Pollinators during Pesticide Spraying, Health Canada, Pest Management Regulatory Agency. [accessed August 2014].
Health Canada. 2014. Update on Neonicotinoid Pesticides and Bee Health. [accessed August 2015]
International Union for Conservation of Nature (IUCN). 2013. Guideline for reintroduction and other conservation translocations. Version 1.0. IUCN Species Survival Commission, Gland, Switzerland. 57 p.
International Union for Conservation of Nature (IUCN). 2015. The IUCN Red List of Threatened Species. [accessed August 2015].
Iserbyt, S., and P. Rasmont. 2012. The effect of climatic variation on abundance and diversity of bumblebees: A ten years survey in a mountain hotspot. Annales de la Société Entomologique de France 48:261-273.
Jepsen, S., E. Evans, R. Thorp, R. Hatfield, and S.H. Black. 2013. Petition to list the rusty patched bumble bee Bombus affinis (Cresson), 1863, as an Endangered species under the U.S. Endangered Species Act. The Xerces Society for Invertebrate Conservation, Portland, Oregon. 42 p.
Johansen, C.A., D.F. Mayer, J.D. Eves, and C.W. Kiuos. 1983. Pesticides and bees. Environmental Entomology 12:1513-1518.
Jönsson, A., J. Ekroos, J. Dänhardt, G.K.S. Andersson, O. Olsson, and H.G. Smith. 2015. Sown flower strips in southern Sweden increase abundances of wild bees and hoverflies in the wider landscape. Biological Conservation 184:51-58.
Kearns, C.A., D.W. Inouye, and N. Waser. 1998. Endangered mutualisms: the conservation of plant-pollinator interactions. Annual Review of Ecology and Systematics 29:83-112.
Kells, A.R., and D. Goulson. 2003. Preferred nesting sites of bumblebee queens (Hymenoptera: Apidea) in agroecosystems in the UK. Biological Conservation 109: 165-174.
Kerr, J. T., A. Pindar, P. Galpern, L. Packer, S.G. Potts, S.M. Roberts, P. Rasmont, O. Schweiger, S.R. Colla, L.L. Richardson, D.L. Wagner, L.F. Gall, D.S. Sikes, and A. Pantoja. 2015. Climate change impacts on bumblebees converge across continents. Science 349(6244):177-180.
Kessler, S.C., E.J. Tiedeken, K.L. Simcock, S. Derveau, J. Mitchell, S. Softley, J.C. Stout, and G.A. Wright. 2015. Bees prefer foods containing neonicotinoid pesticides. Nature 521:74-76.
Kevan, P.G., and R.C. Plowright. 1995. Impact of pesticides on forest pollination. p. 607-618 in J.A. Armstrong and W.G.H. Ives (eds.) Forest Insect Pests in Canada. Natural Resources Canada, Canadian Forest Service, Science and Sustainable Development Directorate, Ottawa, Ontario.
Knight, M.E., A.P. Martin, S. Bishop, J.L. Osborne, R.J. Hale, A. Sanderson, and D. Goulson. 2005. An interspecific comparison of foraging range and nest density of four bumblebee (Bombus) species. Molecular Ecology 14:1811-1820.
Kraus, F.B., S. Wolf, and R.F.A. Moritz. 2009. Male flight distance and population substructure in the bumblebee Bombus terrestris. Journal of Animal Ecology 78:247-252.
Laverty, T., and L.D. Harder. 1988. The bumble bees of eastern Canada. Canadian Entomologist 120:965-987.
Lecocq, T., S. Dellicour , D. Michez, P. Lhomme, M. Vanderplanck, I. Valterova, J.-Y. Rasplus, and P. Rasmont. 2013. Scent of a break-up: Phylogeography and reproductive trait divergences in the red-tailed bumblebee (Bombus lapidarius). BMC Evolutionary Biology 13:263.
Lee, H., W. Bakowsky, J. Riley, J. Bowles, M. Puddister, P. Uhlig, and S. McMurray. 1998. Ecological Land Classification for Southern Ontario: First Approximations and Its Application. SCSS Field Guide FG-02, Ontario Ministry of Natural Resources.
Lepais, O., B. Darvill, S. O'Connor, J.L. Osborne, R.A. Sanderson, J. Cussans, L. Goffe, and D. Goulson. 2010. Estimation of bumblebee queen dispersal distances using sibship reconstruction method. Molecular Ecology 19: 819-831.
Lozier, J.D., J.P. Strange, I.J. Stewart and S.A. Cameron. 2011. Patterns of range-wide variation in six North American bumble bee (Bombus) species. Molecular Ecology 20: 4870-4888.
Macfarlane, R.P. 1974. Ecology of Bombinae (Hymenoptera: Apidae) of Southern Ontario, with emphasis on their natural enemies and relationships with flowers. Ph.D. Thesis, University of Guelph, Guelph, Ontario. 210 p.
Manley, R., M. Boots, and L. Wilfert. 2015. Emerging viral disease risk to pollinating insects: ecological, evolutionary and anthropogenic factors. Journal of Applied Ecology 52:331-340.
Master, L.L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, L. Ramsay, K. Snow, A. Teucher, and A. Tomaino. 2012. NatureServe Conservation Status Assessments: Factors for Evaluating Species and Ecosystem Risk. (PDF; 2.1MB) NatureServe, Arlington, Virginia. 45 p. + 5 appendices. [accessed February 2016].
Matteson, K.C., and G.A. Langellotto. 2009. Bumblebee abundance in New York City community gardens: implications for urban agriculture. Cities and the Environment 2(1): article 5, 12 p.
Mayer, D.F., and J.D. Lunden. 1986. Toxicity of fungicides and an acaricide to honey bees (Hymenoptera: Apidae) and their effects on bee foraging behavior and pollen viability on blooming apples and pears. Environment Entomology 15:1047-1049.
McFrederick, Q.S., and G LeBuhn. 2006. Are urban parks refuges for bumble bees Bombus spp. (Hymenoptera: Apidae)? Biological Conservation 129:372-382.
McFrederick, Q.S., J.C. Kathilankal, and J.D. Fuentes. 2008. Air pollution modifies floral scent trails. Atmospheric Environment 42(10):2336-2348.
McMahon, D.P., M.A. Fürst, J. Caspar, P. Theodorou, M.J.F. Brown, and R.J. Paxton. 2015. A sting in the spit: widespread cross-infection of multiple RNA viruses across wild and managed bees. Journal of Animal Ecology 84:615-624.
Meindl, G.A., and T-L. Ashman. 2013. The effects of aluminium and nickel in nectar on the foraging behaviour of bumblebees. Environmental Pollution 177:78-81.
Memmott, J., N.M. Waser, and M.V. Price. 2004. Tolerance of pollination networks to species extinctions. Proceedings of the Royal Society B 271:2605-2611.
Ministère du Développement durable, de l'Environnement et de la Lutte contre les changements climatiques (MDDELCC). 2015. Stratégie québécoise sur les pesticides 2015-2018. Ministère du Développement durable, de l'Environnement et de la Lutte contre les changements climatiques, Quebec City, Quebec. 24 p.
Mitchell, T.B. 1962. Bees of the Eastern United States. North Carolina Agricultural Experiment Station Technical Bulletin 152:1-557.
Mommaerts, V., G. Sterk, and G. Smagghe. 2006. Hazards and uptake of chitin synthesis inhibitors in bumblebees Bombus terrestris. Pest Management Science 62:752-758.
Morandin, L.A., M.L. Winston, M.T. Franklin, and V.A. Abbott. 2005. Lethal and sub-lethal effects of spinosad on bumble bees (Bombus impatiens Cresson). Pest Management Science 61:619-626.
Morgano, M.A., M.C.T. Martins, L.C. Rabonato, R.F. Milani, K. Yotsuyanagi, and D.B. Rodriguez-Amaya. 2010. Inorganic contaminants in bee pollen from southeastern Brazil. Journal of Agricultural and Food Chemistry 58:6876-6883.
Mullin, C.A., M. Frazier, J.L. Frazier, S. Ashcraft, R. Simonds, D. Vanengelsdorp, and J.S. Pettis. 2010. High levels of miticides and agrochemicals in North American apiaries: implications for honey bee health. PLoS ONE 5:e9754. doi:10.1371/journal.pone.0009754.
Mullin, C.A., J. Chen, J.D. Fine, M.T. Frazier and J.L. Frazier. 2015. The formulation makes the honey bee poison. Pesticide Biochemistry and Physiology 120:27-35.
NatureServe. 2015. NatureServe Explorer: An online encyclopedia of life [web application], version 7.1. [accessed April 2015].
Öckinger, E., and H.G. Smith. 2007. Semi-natural grasslands as population sources for pollinating insects in agricultural landscapes. Journal of Applied Ecology 44:50-59.
Ontario Ministry of Natural Resources (OMNR). 2011. Rusty-patched Bumble Bee. Ontario Government Response Statement. (PDF; 202.5KB) Government of Ontario. 4 p. [accessed September 2015].
Orford, K.A., P.J. Murray, I.P. Vaughan, and J. Memmott. 2016. Modest enhancements to conventional grassland diversity improve the provision of pollination services. Journal of Applied Ecology: in press. doi: 10.1111/1365-2664.12608.
Osborne, J., S.J. Clark, R.J. Morris, I.H. Williams, J.R. Riley, A.D. Smith, D.R. Reynolds, and A.S. Edwards. 1999. A landscape-scale study of bumble bee foraging range and constancy, using harmonic radar. Journal of Applied Ecology 36(4):519-533.
Osborne, J.L., A.P. Martin, N.L. Carreck, J.L. Swain, M.E. Knight, D. Goulson, R.J. Hale, and R.A. Sanderson. 2008. Bumblebee flight distances in relation to the forage landscape. Journal of Animal Ecology 77(2):406-415.
Otterstatter, M.C. and T.L. Whidden. 2004. Patterns of parasitism by tracheal mites (Locustacarus buchneri) in natural bumble bee populations. Apidologie 35:351-357.
Otterstatter, M.C., R.J. Gegear, S.R. Colla, and J.D. Thomson. 2005. Effects of parasitic mites and protozoa on the flower constancy and foraging rate of bumble bees. Behavioral Ecology and Sociobiology 58:383-389.
Park, M.G., E.J. Blitzer, J. Gibbs, J.E. Losey, and B.N. Danforth. 2015. Negative effects of pesticides on wild bee communities can be buffered by landscape context. Proceedings of the Royal Society B 282:20150299.
Paterson, C. 2014. Small restoration, big impacts: how habitat influences native pollinators in intensive agricultural landscapes. Master's Thesis. University of Guelph, Ontario. 100 p.
Paton, D.C. 1990. Budgets for the use of floral resources in mallee heath. p. 189-193 in J.C. Noble, P.J. Joss, G.K. Jones (eds.). The Mallee Lands: A Conservation Perspective – proceedings of the National Mallee Conference. CSIRO, Melbourne, Australia. 322 p.
Paton, D.C. 1996. Overview of feral and managed honeybees in Australia: distribution, abundance, extent of interactions with native biota, evidence of impacts and future research. Australian Nature Conservation Agency, Canberra, Australia. 76 p.
Pilling, E.D., and P.C. Jepson. 1993. Synergism between EBI fungicides and a pyrethroid insecticide in the honeybee (Apis mellifera). Pesticide Management Science 39:293-297.
Pisa, L.W., V. Amaral-Rogers, L.P. Belzunces, J.M. Bonmatin, C.A. Downs, D. Goulson, D.P. Kreutzweiser, C. Krupke, M. Liess, M. McField, C.A. Morrissey, D.A. Noome, J. Settele, N. Simon-Delso, J.D. Stark, J.P. Van der Sluijs, H. Van Dyck, and M. Wiemers. 2015. Effects of neonicotinoids and fipronil on non-target invertebrates. Environmental Science and Pollution Research 22:68-102.
Pleasants, J.M., and K.S. Oberhauser. 2012. Milkweed loss in agricultural fields because of herbicide use: effect on the monarch butterfly population. Insect Conservation and Diversity 6(2):135-144.
Potts, S.G., J.C. Biesmeijer, C. Kremen, P. Neumann, O. Schweiger, and W.E. Kunin. 2010. Global pollinator declines: trends, impacts and drivers. Trends in Ecology and Evolution 25:345-353.
Rao, S., and J.P. Strange. 2012. Bumble bee (Hymenoptera: Apidae) foraging distance and colony density associated with a late-season mass flowering crop. Environmental Entomology 41:905-915.
Rasmont, P., and S. Iserbyt. 2012. The Bumblebees Scarcity Syndrome: Are heat waves leading to local extinctions of bumblebees (Hymenoptera: Apidae: Bombus)? Annales de la Société Entomologique de France 48:275-280.
Rasmont, P., M. Franzén, T. Lecocq, A. Harpke, S. Roberts, J.C. Biesmeijer, L. Castro, B. Cederberg, L. Dvorak , Ú. Fitzpatrick, Y. Gonseth, E. Haubruge, G. Mahé, A. Manino, D. Michez, J. Neumayer, F. Ødegaard , J. Paukkunen, T. Pawlikowski, S. Potts, M. Reemer, J. Settele, J. Straka, and O. Schweige. 2015. Climatic Risk and Distribution Atlas of European Bumblebees. Biorisk 10 (special issue): 1-236.
Reck, H., and R. van der Ree. 2015. Insects, snails and spiders: the role of invertebrates in road ecology. p. 247-257 in R. van der Ree, D.J. Smith and C. Grilo (eds.). Handbook of Road Ecology. John Wiley and Sons, Chichester, United Kingdom.
Rodger, L. 1998. Tallgrass communities of southern Ontario: A recovery plan. World Wildlife Fund Canada and the Ontario Ministry of Natural Resources. 66 p.
Roland, J., and S.F. Matter. 2007. Encroaching forests decouple alpine butterfly population dynamics. Proceedings of the National Academy of Sciences of the United States of America. 104:13702-13704.
Rundlöf, M., G.K.S. Andersson, R. Bommarco, I. Fries, V. Hederström, L. Herbertsson, O. Jonsson, B.K. Klatt, T.R. Pedersen, J. Yourstone, and H.G. Smith. 2015. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 521:77-80.
Sammut, J., and R. Lines-Kelly. 2004. An introduction to acid sulfate soils. Department of Primary Industry, New South Wales Government. [accessed December 2015].
Sanchez-Bayo, F., and K. Goka. 2014. Pesticide residues and bees -- a risk assessment. PLoS ONE 9:e94482. doi:10.1371/journal.pone.0094482.
Savard, M. 2009. Les spécimens du Bombus affinis conservés dans la Collection André Francoeur à Saguenay (Québec). Bulletin de l'entomofaune 38:7-9.
Shepherd, M., S.L. Buchmann, M. Vaughan, and S.H. Black. 2003. Pollinator Conservation Handbook. The Xerces Society, Portland, Oregon.
Shykoff, J.A., and P. Schmid-Hempel. 1991. Incidence and effect of four parasites in natural populations of bumble bees in Switzerland. Apidologie 22(2):117-125.
Simon-Delso, V. Amaral-Rogers, L.P. Belzunces, J.M. Bonmatin, M. Chagnon, C. Downs, L. Furlan, D.W. Gibbons, C. Giorio, V. Girolami, D. Goulson, D.P. Kreutzweiser, C.H. Krupke, M. Liess, E. Long, M. McField, P. Mineau, E.A.D. Mitchell, C.A. Morrissey, D.A. Noome, L. Pisa, J. Settele, J.D. Stark, A. Tapparo, H. Van Dyck, J. Van Praagh, J.P. Van der Sluijs, P.R. Whitehorn, and M. Wiemers. 2015. Systemic insecticides (neonicotinoids and fipronil): trends, uses, mode of action and metabolites. Environmental Science and Pollution Research 22:5-34.
Sjodin, N.E. 2007. Pollinator behavioural responses to grazing intensity. Biodiversity Conservation 16:2103-2121.
Skyrm, K.M. 2011. Environmental impacts on native bumble bee pollinators in an agricultural landscape of western Oregon. Ph.D. Dissertation in Entomology. Oregon State University, Corvallis, Oregon. 151 p.
Smithsonian Science. 2014. Rare Rusty-patched Bumble Bee Discovered in Virginia Survey. Smithsonian Science News. [accessed July 2015].
Solga, M.J., J.P. Harmon, and A.C. Ganguliet. 2014. Timing is Everything: An Overview of Phenological Changes to Plants and Their Pollinators. Natural Areas Journal 34:227-234.
Sugden, E.A. 1985. Pollinators of Astragalus monoensis Barneby (Fabaceae): New host records; potential impact of sheep grazing. Great Basin Naturalist 45:299-312.
Thompson, H.M. 2001. Assessing the exposure and toxicity of pesticides to bumblebees (Bombus sp.). Apidologie 32:305-321.
Thomson, D. 2004. Competitive interactions between the invasive European honey bee and native bumble bees. Ecology 85:458-470.
Thomson. D.M. 2006. Detecting the effects of introduced species: a case study of competition between Apis and Bombus. Oikos 114:407-418.
Tommasi, D., A. Miroa, H.A. Higoa, and M.L. Winstona. 2004. Bee diversity and abundance in an urban setting. The Canadian Entomologist 136(6): 851-869.
Torné-Noguera, A., A. Rodrigo, S. Osorio, and J. Bosch. 2015. Collateral effects of beekeeping: impacts on pollen-nectar resources and wild bee communities. Basic and Applied Ecology: in press. doi:10.1016/j.baae.2015.11.004.
US Department of Transportation. 2015. FHWA Encourages States to Take Action in Achieving Pollinator Health. (PDF; 199KB) Federal Highway Administration. 3 p. [accessed January 2016].
Walther-Hellwig, K., and R. Frankl. 2000. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidea), in an agriculture landscape. Journal of Applied Entomology 124: 299-306.
Walther-Hellwig, K., G. Fokul, R. Frankl, R. Buchler, K. Ekschmitt, and V. Wolters. 2006. Increased density of honeybee colonies affects foraging bumblebees. Apidologie 37:517-532
Westphal, C., I. SteVan-Dewenter, and T. Tscharntke. 2006. Bumblebees experience landscapes at different spatial scales: possible implications for coexistence. Oecologia 149: 289-300.
Whitehorn, P.R., S. O'Connor, F.L. Wackers, and D. Goulson. 2012. Neonicotinoid Pesticide Reduces Bumble Bee Colony Growth and Queen Production.Science 336 (6079):351-352.
Williams, P.H. 1986. Environmental change and the distribution of British bumble bees (Bombus Latr.). Bee World 67:50-61.
Williams, P.H., and J.L. Osborne, 2009. Bumblebee vulnerability and conservation world-wide. Apidologie 40: 367-387.
Williams, P.H., S.R. Colla, and Z. Xie. 2009. Bumble bee vulnerability: common correlates of winners and losers across three continents. Conservation Biology 23:931-940.
Williams, P.H., R.W. Thorp, L.L. Richardson, and S.R. Colla. 2014. Bumble Bees of North America: An Identification Guide. Princeton University Press, Princeton, New Jersey. 208 p.
Williams, N. M., K. L. Ward, N. Pope, R. Isaacs, J. Wilson, E. A. May, J. Ellis, J. Daniels, A. Pence, K. Ullmann, and J. Peters. 2015. Native wildflower plantings support wild bee abundance and diversity in agricultural landscapes across the United States. Ecological Applications 25(8):2119-2131.
Wolf S., and R.F.A. Moritz. 2008. Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 39(2008): 419-427.
Wood, T.J., J.M. Holland, and D. Goulson. 2015. Pollinator-friendly management does not increase the diversity of farmland bees and wasps. Biological Conservation 187:120-126.
Yoder, J.A., A.J. Jajack, A.E. Rosselot, T.J. Smith, M.C. Yerke, and D. Sammataro. 2013. Fungicide contamination reduces beneficial fungi in bee bread based on an area-wide field study in honey bee, Apis mellifera, colonies. Journal of Toxicology and Environmental Health A 76(10):587-600.
Zayed, A., and L. Packer. 2005. Complementary sex determination substantially increases extinction proneness of haplodiploid populations. Proceedings of the National Academy of Sciences of the United States of America 102(30):10742-10746.
Personal communications
Boutin, A., pers. comm. 2015. Communication with M.-J. Ribeyron. August 2015. Project manager at Éco-Nature, Laval, Quebec.
MacDougall, A., pers. comm. 2015. Communication with J. Girard. December 2015. Associate Professor, Department of Integrative Biology, University of Guelph, Ontario.
MacKenzie, A., pers. comm. 2015. Communication with J. Girard. August 2015. Project manager at Pinery Provincial Park, Ontario.
Payette, A., pers. comm. 2015. Communication with M. Saint-Germain. June 2015. Entomologist at Montreal Insectarium, Montreal, Quebec.
Appendix A: Plant species used by the Rusty-patched Bumble Bee
The list of plant species provided in this appendix is based on available published records. It is likely that the Rusty-patched Bumble Bee uses other food sources as well.
A.1. Known plant food sources for the Rusty-patched Bumble Bee in southern Ontario (Colla and Dumesh 2010).
- From Macfarlane (1974)
- Arctium minus Notel - A Notem
- Asclepias syriaca - P
- Berberis thunbergii Notel - P
- Carduus nutans Notel - P
- Centaurea cyanus Notel - P
- Cotoneaster adpressa Notel - P
- Crataegus - P
- Cucumis melo Notel - A
- Deutzia gracilis Notel - P
- Echium vulgare Notel - A
- Hydrophyllum virginianum - P
- Hypericum perforatum Notel - P
- Impatiens capensis - A
- Kalmia latifolia - P
- Linaria vulgaris Notel - P
- Lonicera caerulea Notel - P
- Lonicera periclymenum Notel - P
- Lonicera tatarica Notel - P
- Lotus corniculatus Notel - P
- Melilotus alba Notel - A
- Medicago sativa Notel - P
- Nepeta cataria Notel - P
- Prunus Americana - P
- Prunus cerasus Notel - P
- Prunus tomentosa Notel - P
- Pyrus malus Notel - P
- Rhododendron Notel - P
- Ribes grossularia Notel - P
- Ribes nigrum Notel - P
- Robinia fertilis Notel - P
- Rubus - P
- Salix - P
- Silene dichotoma Notel - P
- Solanum dulcamara Notel - P
- Solidago canadensis - P
- Solidago flexicaulis - P
- Sonchus oleraceus Notel - A
- Stachys palustris Notel - P
- Symphyotrichum (Aster) ericoides - P
- Symphyotrichum (Aster) lateriflorum - P
- Symphyotrichum novae-angliae - P
- Symphytum officinale Notel - P
- Syringa vulgaris Notel - P
- Taraxacum officinale Notel - P
- Trifolium pratense Notel - P
- Trifolium repens Notel - P
- Vicia cracca Notel - P
- Vinca minor Notel - P
- Weigelia florida Notel - P
- Museum Archives and Field Data
- Eupatorium maculatum - P
- Eupatorium perfoliatum - P
- Eupatorium rugosum - P
- Euthamia graminifolia - P
- Helianthus decapetalus - P
- Helianthus divaricatus - P
- Kalmia – P
- Onopordum acanthium Notel - P
- Rhexia virginica - P
- Rhus - P
- Spiraea - P
- Vaccinium angustifolium - P
- From Colla et al. (2011)
- Centaurea stoebe (syn. C. biebersteinii, C. maculosa) Notel - P
A.2 Wild plant species visited by the Rusty-patched Bumble Bee (Evans et al. 2008).
- Abelia grandiflora (Speight, 1967)
- Aesculus spp. (Dieringer, 1982; Macfarlane, 1974)
- Agastache foeniculum (C. Reed pers. comm. with E. Evans, July 2008)
- Amorpha canadense (C. Reed pers. comm. with E. Evans, July 2008)
- Asclepias syriaca, A. incarnata, A. verticillata (Frost, 1965; Macior,1965)
- Aralia spp. (Mitchell, 1962),
- Aster spp. (Costelloe, 1988),
- Aquilegia canadensis (Macior, 1978a),
- Aureolaria pedicularia (Stiles, 1977)
- Berberis spp. (Macior ,1965),
- Camassia scilloides (Macior, 1978b)
- Carduus sp. (Macior 1965)
- Ceanothus americanus (Bequaert, 1920),
- Cercis canadensis (Fye et Medler, 1954)
- Chamaedaphne calyculata (Judd, 1966),
- Coreopsis major (Speight, 1967)
- Crataegus spp. (Macior, 1968)
- Dalea purpurea (C. Reed pers. comm. with E. Evans, July 2008)
- Delphinium tricorne (Macior, 1975)
- Dicentra canadensis, D. cucullaria (Macior, 1978b)
- Echium vulgare (Macfarlane, 1974),
- Helianthus spp. (Fye and Medler,1954; Colla and Packer 2008)
- Hydrangea spp. (Mitchell, 1962),
- Hydrophyllum spp. (Macior 1978b, Macfarlane 1974)
- Impatiens capensis (R. Gegear pers. comm. with E. Evans, May 2008)
- Lamium purpureum (Macior, 1978a)
- Laportea spp. (Speight, 1967)
- Leonurus sp. (Macior, 1965)
- Linaria sp. (Macior, 1965)
- Lonicera spp. (Macior, 1968)
- Lotus corniculatus (Fye et Medler, 1954)
- Medicago sativa (Fye et Medler, 1954)
- Mertensia virginica (Macior, 1978b)
- Monarda sp. (Macior, 1965)
- Nepeta spp. (Macior, 1965)
- Pedicularis canadensis (Macior, 1978b; Dieringer, 1982)
- Pedicularis lanceolata (Costelloe, 1988; Macior, 1969)
- Penstemon grandiflorus (C. Reed, pers. comm. with E. Evans, July 2008)
- Philadelphus spp. (Speight, 1967)
- Polymnia spp. (Speight, 1967)
- Prunella vulgaris (Speight, 1967)
- Prunus spp. (Fye et Medler, 1954)
- Pyrus ioensis (Macior, 1968)
- Pyrus malus (Macior, 1968)
- Ratibida pinnata (C. Reed, pers. comm. with E. Evans, July 2008)
- Rhododendron spp. (Macfarlane, 1974)
- Rhus spp. (Speight, 1967)
- Ribes spp. (Macfarlane, 1974)
- Robinia spp. (Mitchell, 1962)
- Rosa spp. (Macior ,1965)
- Rubus spp. (Macfarlane, 1974)
- Salix spp. (Medler et Carney, 1963)
- Sarracenia purpurea (Ne'eman et coll., 2006)
- Solanum sp. (Macior, 1965)
- Solidago spp. (Mitchel,l 1962),
- Symphytum officinale (Macfarlane, 1974)
- Syringia spp.(Macior, 1968)
- Syringia vulgaris (Fye et Medler, 1954)
- Taraxacum spp. (Macior, 1968)
- Trifolium spp. (Fye et Medler, 1954; Macfarlane, 1974)
- Vaccinium spp. (Mitchell, 1962)
- Verbascum spp. (Macior, 1965)
- Verbesina occidentalis (Speight, 1967)
- Vicia spp. (Fye et Medler, 1954; Macfarlane, 1974)
References
Bequaert, J. 1920. Hymenoptera collected near Boston, Mass., with description of a variety of Bombus affinis. Psyche 27:6-12.
Costelloe, B. H. 1988. Pollination ecology of Gentiana andrewsii. Ohio. Journal of Science 88:132- 138.
Dieringer, G. 1982. The pollination ecology of Orchis spectabilis L. (Orchidaceae). Ohio Academy of Science 82:218-225.
Frost, S.W. 1965. Insects and pollinia. Ecology 46:556-558.
Fye, R.E. et J.T. Medler. 1954. Spring emergence and floral hosts of Wisconsin bumblebees. Transactions of the Wisconsin Academy of Science, Arts and Letters 43:75-82.
Gegear, R., comm. pers. 2008. Communication avec E. Evans. Mai 2008. Citée Evans et coll. (2008).
Macfarlane, R.P. 1974. Ecology of Bombinae (Hymenoptera: Apidae) of Southern Ontario, with emphasis on their natural enemies and relationships with flowers. Thèse de doctorat, University of Guelph, Guelph, Ontario.
Macior, L.W. 1965. Insect adaptation and behavior in Asclepias pollination. Bulletin of the Torrey Botanical Club 92:114-126.
Macior. L.W. 1968. Bombus (Hymenoptera, Apidae) queen foraging in relation to vernal pollination in Wisconsin. Ecology 49:20-25.
Macior, L. W. 1975. The pollination ecology of Delphinium tricorne (Ranunculaceae). American Journal of Botany 62:1009-1016.
Macior, L.W. 1978a. Pollination interactions in sympatric Dicentra species. American Journal of Botany65:57-62.
Macior, L.W. 1978b. Pollination ecology of vernal angiosperms. Oikos 30:452-460.
Medler, J. T. and D. W. Carney. 1963. Bumblebees of Wisconsin (Hymenoptera: Apidae). Research Bulletin, University of Wisconsin, Agricultural Experiment Station 240:1-47.
Mitchell, T.B. 1962. Bees of the Eastern United States. Vol. II. North Carolina Agricultural Experiment Station Technical Bulletin 152:1-557.
Ne'eman, G., R. Ne'eman et A.M. Ellison. 2006. Limits to reproductive success of Sarracenia purpurea (Sarraceniaceae). American Journal of Botany 93:1660-1666.
Reed, C., comm. pers. 2008. Communication avec E. Evans. Juillet 2008. Citée Evans et coll. (2008).
Speight, D.L. 1967. The bumblebees (Hymenoptera, Apidae, Bombinae) of east Tennessee. Mémoire de maîtrise, University of Tennessee, Knoxville, Tennessee.
Stiles, E.W. 1977. Foraging behavior of bumblebees (Bombus impatiens, Bombus vagans, Bombus affinis) on false foxglove (Aureolaria pedicularia). Journal of the New York Entomological Society 85:249-252.
Appendix B: Effects on the environment and on other species
A strategic environmental assessment (SEA) is conducted on all SARA recovery planning documents, in accordance with the Cabinet Directive on the Environmental Assessment of Policy, Plan and Program Proposals. The purpose of a SEA is to incorporate environmental considerations into the development of public policies, plans, and program proposals to support environmentally sound decision making and to evaluate whether the outcomes of a recovery planning document could affect any component of the environment or any of the Federal Sustainable Development Strategy's (FSDS) goals and targets.
Recovery planning is intended to benefit species at risk and biodiversity in general. However, it is recognized that strategies may also inadvertently lead to environmental effects beyond their intended benefits. The planning process based on national guidelines directly incorporates consideration of all environmental effects, with a particular focus on possible impacts upon non-target species or habitats. The results of the SEA are incorporated directly into the strategy itself, but are also summarized below in this statement.
Conservation of the Rusty-patched Bumble Bee and its habitat will benefit other species at risk in Canada, including the Gypsy Cuckoo Bumble Bee (Bombus bohemicus), which has a close relationship with the Rusty-patched Bumble Bee (parasite-host), as well as other insect pollinators (e.g., the Yellow-banded Bumble Bee [Bombus terricola]), the Monarch [Danaus plexippus] and species that use the same type of habitat or are present in the area of Pinery Provincial Park (e.g., the Eastern Hog-nosed Snake [Heterodon platirhhinos]), the Northern Barrens Tiger Beetle [Cicindela patruela], and the Aweme Borer [Papaipema awane]. It will also favour the maintenance or restoration of certain plant communities which are currently rare in Canada (e.g., Black/White Oak savanna, tallgrass prairie) as well as the wildlife they support. The approaches presented in Table 3 will likely benefit other species by reducing pathogen transmission as well as pesticide use--which contributes to water and soil contamination.
Bumble bees, in general, are important pollinators of many native flowering plants and crops (COSEWIC 2010). They have several characteristics that contribute to their effectiveness as pollinators of crop plant species (Corbet et al. 1993). For example, they are able to fly at lower temperatures than other bees, which allows for a longer work day and improves pollination of crops during inclement weather. They also have the capacity to “buzz pollinate” (vibratile pollination)Note32, which can increase the rate of pollination of plants. Some cultivated plants, such as tomato and pepper, benefit from buzz pollination (Jepsen et al. 2013). The Rusty-patched Bumble Bee has been shown to be an excellent pollinator of cranberry and other important food crops such as plum and apple, alfalfa and onion (COSEWIC 2010). It is likely the primary pollinator for many ecologically and economically important plants (including apple, raspberry, lilac, honeysuckle, hawthorn, nightshade, clover, milkweed, goldenrod, and aster) (COSEWIC 2010). The Rusty-patched Bumble Bee plays a vital role as a generalist pollinator of native flowering plants, and its loss could have far-ranging impacts (Jepsen et al. 2013). It has been shown that the loss of generalist pollinators, especially bumble bees, would cause a greater number of plant extinctions than would the loss of specialist pollinators (Memmott et al. 2004). Finally, some of the plants visited by the Rusty-patched Bumble Bee are prized by First Nations communities for their important medicinal properties (e.g., Aralia, Rosa, Rubus and Spiraea) (COSEWIC 2010).
The potential for this recovery strategy to inadvertently lead to adverse effects on other species was considered. None of the measures proposed include activities that would negatively affect other species. The SEA concluded that this strategy will clearly benefit the environment and will not entail significant adverse effects.