
Recovery Strategy for the Greater Sage-Grouse phaios subspecies (Centrocercus urophasianus phaios) in Canada 2017
Greater Sage-Grouse phaios subspecies

- Figure 1. Estimated historical and known recent Greater Sage-Grouse distribution in, a) British Columbia and adjacent Washington State, and b) range-wide in North America (data from Connelly et al. 2004)
- Table 1. Conservation Status Greater Sage-Grouse phaios (NatureServe 2015; B.C. Conservation Data Center 2015; B.C. Conservation Framework 2015).
Recovery Strategy for the Greater Sage-Grouse phaios subspecies (Centrocercus urophasianus phaios) in Canada 2017

Environment and Climate Change Canada. 2017. Recovery Strategy for the Greater Sage-Grouse phaios subspecies (Centrocercus urophasianus phaios) in Canada. Species at Risk Act Recovery Strategy Series. Environment and Climate Change Canada, Ottawa. vii + 19 pp.
For copies of the recovery strategy, or for additional information on species at risk, including the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) Status Reports, residence descriptions, action plans, and other related recovery documents, please visit the Species at Risk (SAR) Public Registry.
Cover illustration: © United States Fish and Wildlife Service
Également disponible en français sous le titre
« Programme de rétablissement du Tétras des armoises de la sous-espèce phaios (Centrocercus urophasianus phaios) au Canada »
Content (excluding the illustrations) may be used without permission, with appropriate credit to the source.
The federal, provincial, and territorial government signatories under the Accord for the Protection of Species at Risk (1996) agreed to establish complementary legislation and programs that provide for effective protection of species at risk throughout Canada. Under the Species at Risk Act (S.C. 2002, c.29) (SARA), the federal competent ministers are responsible for the preparation of recovery strategies for listed Extirpated, Endangered, and Threatened species and are required to report on progress within five years after the publication of the final document on the SAR Public Registry.
The Minister of Environment and Climate Change is the competent minister under SARA for the Greater Sage-Grouse phaios and has prepared this recovery strategy, as per section 37 of SARA. To the extent possible, it has been prepared in cooperation with the Province of British Columbia, as per section 39(1) of SARA.
The recovery of the Greater Sage-Grouse phaios in Canada is not considered to be technically or biologically feasible. The species may, however, benefit from general conservation programs in the same geographic area and will receive protection through SARA and other federal, and provincial or territorial, legislation, policies, and programs.
The feasibility determination will be re-evaluated as part of the report on implementation of the recovery strategy, or as warranted in response to changing conditions and/or knowledge.
The recovery strategy sets the strategic direction to support recovery of the species, including identification of critical habitat to the extent possible. It provides all Canadians with information to help take action on species conservation. When critical habitat is identified, either in a recovery strategy or an action plan, SARA requires that critical habitat then be protected.
In the case of critical habitat identified for terrestrial species including migratory birds SARA requires that critical habitat identified in a federally protected area Footnote1 be described in the Canada Gazette within 90 days after the recovery strategy or action plan that identified the critical habitat is included in the public registry. A prohibition against destruction of critical habitat under ss. 58(1) will apply 90 days after the description of the critical habitat is published in the Canada Gazette.
For critical habitat located on other federal lands, the competent minister must either make a statement on existing legal protection or make an order so that the prohibition against destruction of critical habitat applies.
If the critical habitat for a migratory bird is not within a federal protected area and is not on federal land, within the exclusive economic zone or on the continental shelf of Canada, the prohibition against destruction can only apply to those portions of the critical habitat that are habitat to which the Migratory Birds Convention Act, 1994 applies as per SARA ss. 58(5.1) and ss. 58(5.2).
For any part of critical habitat located on non-federal lands, if the competent minister forms the opinion that any portion of critical habitat is not protected by provisions in or measures under SARA or other Acts of Parliament, or the laws of the province or territory, SARA requires that the Minister recommend that the Governor in Council make an order to prohibit destruction of critical habitat. The discretion to protect critical habitat on non-federal lands that is not otherwise protected rests with the Governor in Council.
This document was developed by Darcy Henderson, Matt Huntley, and Kella Sadler (Environment and Climate Change Canada, Canadian Wildlife Service – Pacific region (ECCC CWS-PAC). Due to the lack of adequate information for the Greater Sage-Grouse phaios much of the species information is based on the Amended Recovery Strategy for the Greater Sage-Grouse (Centrocercus urophasianus urophasianus) in Canada (Environment Canada 2014) prepared by Troy I. Wellicome and Joy Stevens. Valuable review and comments were provided by Marie-Andrée Carrière (ECCC CWS-National Capital Region) and Orville Dyer and Myke Chutter (B.C. Ministry of Environment).
The Greater Sage-Grouse (Centrocercus urophasianus) is a turkey-sized, ground-dwelling bird that inhabits temperate, semi-arid sagebrush (Artemisia spp.) steppe in Canada and the United States. These birds depend upon sagebrush as a source of food, particularly in winter, and as cover for nesting and avoiding predators. In spring, Greater Sage-Grouse congregate at the same location each year (known as leks) where males perform elaborate dancing displays to attract mates. In Canada two subspecies of Greater Sage-Grouse are recognized, with C. u. urophasianus in Alberta and Saskatchewan, and C. u. phaios in British Columbia. This recovery strategy applies only to the latter, the Greater Sage-Grouse phaios subspecies.
The Greater Sage-Grouse phaios was designated as Extirpated by the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) in 1997 and was subsequently listed as Extirpated in Canada on Schedule 1 of the federal Species at Risk Act as of 2003. The only Canadian distribution of this subspecies was the extreme southern end of the Okanagan and Similkameen valleys in south-central British Columbia. The last naturally occurring record of Greater Sage-Grouse phaios was 1918 near the community of Oliver, British Columbia. Reintroduction efforts in the late 1950s were unsuccessful. The British Columbia Conservation Data Centre ranks Greater Sage-Grouse SX (extirpated), and the species is protected from capture and killing under the B.C. Wildlife Act.
In the United States the Greater Sage-Grouse is not listed under the Endangered Species Act, but in Washington State the species is threatened and protected under State laws. Threats to Greater Sage-Grouse phaios in adjacent Washington State are primarily habitat loss and degradation. Irreversible loss of habitat was due to conversion for crop production and urban development. Habitat degradation has been due to livestock grazing, cheatgrass (Bromus tectorum) invasion, conifer encroachment, and increasing fire frequency in sagebrush. Increasing populations of native predators, and additive mortality from road collisions and West Nile virus are additional threats to populations. Washington State is using translocations from Nevada and Oregon to try and sustain a remnant viable population, just 100 km south of the British Columbia border. Threats in British Columbia were likely similar to those in Washington State.
Recovery in Canada is not considered biologically and technically feasible at this time. There is insufficient quantity and unsuitable quality of habitat in Canada, very low likelihood of restoring brood-rearing habitat in Canada, and very low likelihood of agreement with the United States to supply birds for translocation into these sub-optimal conditions and contrary to their existing priorities under the Sage Grouse Initiative. Recovery feasibility may be revisited if relict populations are discovered.
Based on the following four criteria that Environment and Climate Change Canada uses to establish recovery feasibility, recovery of the Greater Sage-Grouse phaios in Canada has been determined not to be biologically or technically feasible at this time. Recovery is considered not feasible when the answer to any of the following questions is “no”. The feasibility of recovery may be revised if relict population(s) are discovered in Canada, or if translocation from U.S. populations becomes appropriate.
i COSEWIC (Committee on the Status of Endangered Wildlife in Canada)
ii Note that the subspecies has not naturally occurred in the former range since 1918, and an attempted reintroduction in 1958 was unsuccessful. This accounts for why the species has not been reported since the 1960s in the statement above.
Legal Status: SARA Schedule 1 (Extirpated) (2003).
| Global (G) Ranka | National (N) Ranka | Sub-national (S) Ranka | COSEWIC Designation | B.C. List | B.C. Conservation Framework |
|---|---|---|---|---|---|
| G3G4T3Q | Canada (N1)b; United States (N3) |
Canada: British Columbia (SX); United States: Oregon (S3), Washington (S1) |
Extirpated (2008) | Red | Highest priority: 6 under goal 1,2,3c |
a The conservation status of a species is designated by a number from 1 to 5, preceded by a letter reflecting the appropriate geographic scale of the assessment (G = Global, N = National, and S = Subnational). The numbers have the following meaning: 1 = critically imperiled, 2 = imperiled, 3 = vulnerable, 4 = apparently secure, 5 = secure. SX = Presumed Extirpated, SNR = Unranked. Additional rank information includes Q = questionable taxonomic validity and T = status of infraspecific taxa (i.e. the phaios subspecies).
b The sub-species phaios is not ranked separately from subspecies urophasianus at the National level, so the N1 ranking reflects the two combined despite the long-term extirpation of subspecies phaios from Canada. Only states with the subspecies phaios have S ranks reported here.
c The three goals of the B.C. Conservation Framework are: 1. Contribute to global efforts for species and ecosystem conservation; 2. Prevent species and ecosystems from becoming at risk; 3. Maintain the diversity of native species and ecosystems
As per COSEWIC (2008), this recovery strategy seperates the Greater Sage-Grouse (Centrocercus urophasianus) into the eastern (C. u. urophasianus) and western C. u. phaios). Recent genetic evidence does not support delineation of Greater Sage-Grouse into these subspecies (Benedict et al. 2003; Oyler-McCance et al. 2005). However, in Canada COSEWIC still considers the two subspecies designatable units with geographically discrete breeding populations and in two significantly different ecological areas (see Figure 1). The subspecies phaios historically occupied grazing-sensitive vegetation dominated by the evergreen basin big sagebrush (Artemisia tridentata ssp. tridentata) and bluebunch wheatgrass (Pseudoregneria spicata) in the Southern Mountain Ecological Area of British Columbia. Meanwhile, the subspecies urophasianus occupies grazing-resilient vegetation dominated by mixedgrass prairie with the deciduous silver sagebrush (Artemisia cana) in the Prairie Ecological Area of southeastern Alberta and southwestern Saskatchewan.
The Greater Sage-Grouse phaios was formerly a range extension of the Columbia Basin Distinct Population Segment (DPS) that is currently critically imperiled in Washington State (USFWS 2003).
Greater Sage-Grouse are ground-dwelling birds and the largest species of grouse in North America . They have brownish-grey upperparts with a blackish belly, the undersides of the rounded-wings are whitish and its long black-and-white tail has distinctive tips. Adult males have a white band on a black throat and a large ruff of pointed white feathers concealing the yellowish air sacs that inflate during courtship displays. Other male characteristics include fleshy yellow combs above the eyes and long, hairlike feathers arising from the nape. Both sexes have a diagnostic black patch on the belly (it is larger on the male). The female has more cryptic plumage and an inconspicuous comb above the eye and is smaller than the male, which can reach 75 cm in length.
Greater Sage-Grouse are polygynous, as individual males typically mate with several females. Males perform ritualistic displays, called strutting, on communal leks Footnote2 to attract females, who select a male from the group with which to mate (Bergerud 1988; Connelly et al. 2004). Males begin displaying at leks in early spring, and strutting commences before sunrise each morning and continues until about a half-hour after sunrise (Jenni and Hartzler 1978; Aldridge 2000).
Greater Sage-Grouse require sagebrush (Artemisia spp.) and are year-round residents in the sagebrush-grasslands of the semi-arid mixed-grass prairie of southeastern Alberta and southwestern Saskatchewan, and formerly of the semi-arid bunchgrass biogeoclimatic zone in southcentral British Columbia. This Canadian distribution is the northern limit of the Greater Sage-Grouse range in North America (Aldridge 1998; Braun 1998; Connelly et al. 2000; Connelly et al. 2004).
By the year 2000, the range of Greater Sage-Grouse in North America had been reduced to half its historical range (668 412 km2, down from 1 200 483 km2; Schroeder et al. 2004), now occurring in eleven states and the provinces of Alberta and Saskatchewan (Figure 1) (Aldridge and Brigham 2003). The current Greater Sage-Grouse range in Canada (~7370 km2; Figure 1) occupies about 7% of the historical Canadian range (~100 000 km2), and now only includes the urophasianus owing to extirpation of the phaios from British Columbia (Aldridge 2000; Aldridge and Brigham 2003). In Washington State, Greater Sage-Grouse phaios only occupy 8% of their former range (Stinson et al. 2004).
The area historically occupied by Greater Sage-Grouse phaios in Canada is approximately 200 km2 at the extreme northern extent of its range, and at the extreme southern end of two deep and narrow valleys, the Okanagan and Similkameen, that open to a broader plain southward into Washington State (Cannings et al. 1987). The big sagebrush on which Greater Sage-Grouse phaios depend is limited to lower elevation grasslands within the Bunchgrass and Ponderosa Pine biogeoclimatic zones (Meidinger and Pojar 1991).
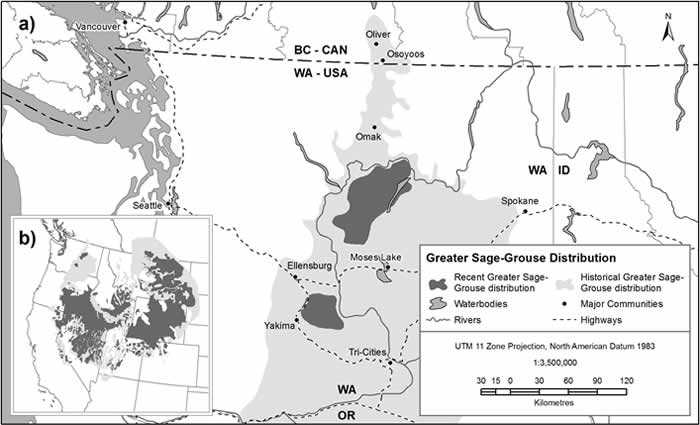
Long description for Figure 1
Figure 1 contains 2 figures; a and b. Figure a) is a zoomed in map of the estimated historical and known recent Greater Sage-Grouse distribution in British Columbia and Washington State. There is a historical Greater Sage-Grouse distribution which extends from just north of Oliver, B.C. well into Oregon and in the eastern direction until Spokane, WA. Two Recent Greater Sage-Grouse distributions are noted just south of Omak, WA and just east of Yakima, WA. Figure b) is a zoomed out map of the range in North America.
In Canada, the Greater Sage-Grouse phaios was historically at the periphery of its North American range (Figure 1). Infrequent occurrences in the south Okanagan Valley were documented from 1864 to 1918 and during this period there is no documented reference of Greater Sage-Grouse phaios breeding. The last naturally occurring Greater Sage-Grouse phaios in British Columbia was shot in 1918 (Cannings et al. 1987). In 1958, 57 birds from Oregon were translocated to British Columbia near Richter Lake, 10 km west of Osoyoos and between the Okanagan and Similkameen valleys (Campbell and Ryder 2010). Most did not survive the winter or crossed the border into Washington, but two independant breeding records were documented in May and July of the following year near Kilpoola Lake, 5 km west of Osoyoos and 5 km southeast of the release site (Campbell and Ryder 2010). Four post-translocation records of Greater Sage-Grouse observations were recorded between 1962 and 1966 including the last report of Greater Sage-Grouse in British Columbia, of a dead bird observed August 14, 1966 on the "Osoyoos side of Anarchist Mtn" (Barkley 1966 as cited in Campbell and Ryder 2010) perhaps 15 km east of the original release site. Greater Sage-Grouse phaios has not been recorded in Canada in 50 years and was designated as Extirpated by COSEWIC in 1997 (Hyslop 1998).
Greater Sage-Grouse have specific habitat requirements for mating, nesting, brood-rearing, and wintering that appear consistent across their range (see Habitat reviews in Aldridge 2000; Connelly et al. 2004; Connelly et al. 2011). For that reason, reference to research throughout the range of Greater Sage-Grouse should reasonably apply to the extirpated Greater Sage-Grouse phaios in British Columbia.
Generally, sagebrush is important for both cover and food (Patterson 1952; Braun et al. 1977; Connelly et al. 2000; Connelly et al. 2004), with leaves comprising <60% of Greater Sage-Grouse summer diet but virtually 100% of their winter diet (Patterson 1952; Wallestad et al. 1975; Hanf et al. 1994; Connelly et al. 2004). Broadleaf non-woody plants (forbs) and insects are dietary requirements during all seasons except winter (Wallestad et al. 1975; Drut et al. 1994a and 1994b). Forbs are a rich source of protein and provide habitat that enhances the availability of insects (Huwer 2004), which in turn can influence nest initiation, clutch size, and reproductive success (Barnett and Crawford 1994; Coggins 1998; Connelly et al. 2004), including growth and survival of the chicks (see summary in Lungle and Pruss 2008).
In Washington State, Greater Sage-Grouse phaios populations occur on low-relief topography with slopes <16°, with 10-35% canopy cover of sagebrush interspersed with rabbitbrush (Ericameria nauseosus) and antelope bitterbush (Purshia tridentata), a dense understory of native bluebunch wheatgrass and Idaho fescue (Festuca idahoensis), and few or no trees (Stinson et al. 2004). Research throughout the range of Greater Sage-Grouse indicates sagebrush density at northern peripheries of their range is near or below the threshold required further south (Doherty et al. 2016). Further, Greater Sage-Grouse are known to remain in proximity to riparian areas (Blomberg et al. 2014), and avoid croplands, grasslands without sagebrush, trees, rough terrain and steep slopes (Patterson 1952, Fedy et al. 2014, Doherty et al. 2008, Doherty et al. 2016). In British Columbia, suitable habitat occurred in narrow strips along valley bottoms of the south Similkameen and Okanagan valleys. It is likely the steep rocky slopes of the valleys and increasing tree cover with elevation limited habitat availability. In comparison, the Okanagan valley immediately south of the Canadian border is broader, gentler in topography, and has fewer trees.
In spring, Greater Sage-Grouse congregate at leks upon which male birds dance and female birds select mates. Leks vary in area from 0.04–16 ha (Scott 1942; Patterson 1952; Dalke et al. 1963; Parks Canada Agency 2009), and can occur on natural or anthropogenically disturbed locations (Stinson et al. 2004). Most leks have a central area of matted grass and bare ground free of shrubs, to improve visibility of the display and for predator vigilance. Males will occupy surrounding stands of dense sagebrush for feeding and roosting during the mating season.
Greater Sage-Grouse nesting habitat is typically a broad area of sagebrush and grassland surrounding leks (Aldridge 2000). Nests are on the ground, usually under the canopy of a sagebrush, and surrounded by tall grass cover to help conceal nests from mammalian and avian predators (DeLong et al. 1995; Connelly et al. 2011).
In summer, Greater Sage-Grouse brood-rearing habitat changes over time (Stinson et al. 2004). Initially the brood remains close to the nest site to forage. As the dry heat of summer cures the upland vegetation, broods move to progressively moister meadows in search of insects and green and moisture-rich forbs on which to feed (Crawford et al. 2004). Foraging occurs in patches of low sagebrush cover and high ratios of forbs to grasses, whereas roosting and loafing occurs in adjacent dense patches of sagebrush. Males remain in upland sagebrush cover throughout the summer (Hagen 1999). Hens and broods move into dense sagebrush in late-summer and fall before moving to wintering grounds (Patterson 1952; Wallestad 1971; Drut et al. 1994a).
In winter, Greater Sage-Grouse usually congregate in sexually-segregated flocks (Beck 1977; Eng and Schladweiler 1972; Connelly et al. 1988). Winter survival of Greater Sage-Grouse is typically high (Connelly et al. 2004; Aldridge et al 2004). For food and shelter during winter, Greater Sage-Grouse rely almost exclusively on taller sagebrush exposed above snow (Tack 2009; Connelly et al. 2011), and they will move long distances to avoid deep snow and seek out dense sagebrush for thermal cover in severe weather (Moynahan et al. 2006). On a local scale, Greater Sage-Grouse usually select low elevation, south- or southwest-facing slopes with tall and dense sagebrush (see detailed review in Connelly et al. 2011). Ideal winter habitats were likely very limiting in British Columbia compared to southward in Washington State.
Threats are defined as the proximate activities or processes that have caused, are causing, or may cause in the future the destruction, degradation, and/or impairment of the entity being assessed (population, species, community, or ecosystem) in the area of interest (global, national, or subnational) (Salafsky et al. 2008). Threats presented here do not include biological features of the species or population which are considered limiting factors.
The threat classification used in this document is based on the IUCN-CMP (World Conservation Union–Conservation Measures Partnership) unified threats classification system and is consistent with methods used by the B.C. Conservation Data Centre and the B.C. Conservation Framework. For a detailed description of the threat classification system, see the Conservation Measures Partnership website (CMP 2010).
A threat assessment was not completed for the Greater Sage-Grouse phaois as no extant locations in Canada are known. As no locations are known, threats cannot be scored for scope Footnote3 or severity Footnote4 to determine individual threat impacts Footnote5 nor is it possible to calculate the overall threat Impact Footnote6 for this species at this time. Instead, threats were examined and described using the IUCN-CMP threat categories.
Possible threats to Greater Sage-Grouse phaios in Canada are discussed below under the Threat Level 1 headings (Threat Level 2 headings provided in brackets). Some of these threats (particularly habitat loss, livestock grazing, and West Nile virus) may be ongoing should a relict population be discovered in Canada or if reintroduced populations were established in the future.
Habitat loss and degradation due to urban and commercial developments were likely significant threats to Greater Sage-Grouse phaios in Canada. It is likely the development of Osoyoos and Oliver in valley bottoms (see Lea 2008) interfered with movements of individuals historically. Now the footprint of those communities is growing to span the full width of the Okanagan valley, effectively fragmenting the available habitat. Anthropogenic areas are believed to contribute to cumulative landscape effects that suppress populations (Braun et al. 2002; Naugle et al. 2011). It is unclear whether year-round avoidance of anthropogenic sites is primarily due to noise or to other influences, such as associated habitat conversion or elevated vehicle traffic at such sites (Environment Canada 2014).
Habitat loss due to conversion of native vegetation to irrigated croplands was likely a significant historical threat. Although much of the upland sagebrush on sloped alluvial fans and glacio-lacustrine benches remain in British Columbia, the brood-rearing habitat of moister valley bottom grasslands has all but been eliminated in the Okanagan valley (Lea 2008, Bezener et al. 2006)). Although the rate of conversion has slowed because much of the arable land is already under cultivation, this threat does continue and is unlikely to be reversed.
Habitat degradation due to certain livestock grazing practices is recognized as a historical, current, and future threat throughout the Greater Sage-Grouse range in Canada and the United States (see review in Environment Canada 2014). High stocking-rates can adversely impact habitat by decreasing vegetation height for nesting cover, and decreasing forb and insect biomass upon which broods forage (Beck and Mitchell 2000). Grazing has also been shown to increase the probability of invasion by the invasive annual cheatgrass (Reisner et al. 2013), with consequences for fire frequency and sagebrush habitat loss (see Threat 7 below for additional information).
Direct impacts from roads on Greater Sage-Grouse may include direct habitat loss from road construction, direct mortality due to roadkill, fragmenting potential habitat, and facilitating the movement of predators and the spread of invasive exotic plant species (Aldridge 1998; Braun 1998). Indirect influences such as noise and motion from vehicles can disrupt local breeding activities and Greater Sage-Grouse tend to avoid these areas (Braun 1998). A major Provincial highway #97, and National highway #3, with associated high traffic volumes, bisect the historic range of Greater Sage-Grouse in British Columbia.
Fire is a significant threat to recovery efforts in the Columbia Basin population in central Washington (Stinson 2014), which is the closest analogue to the extirpated British Columbia population. Basin big sagebrush can take decades to centuries for cover and density to fully recover following fires (Baker 2011). Following fires, Greater Sage-Grouse are slow to recolonize burned areas even if structural features of the shrub community have recovered (Connelly et al. 2011). Fire is a natural disturbance in sagebrush steppe, and historically Greater Sage-Grouse could simply move to more suitable unburned habitat. Currently, the population and habitat has been reduced and fragmented such that a fire in an isolated fragment could further reduce the population and distribution. Fire frequency has also increased over time in habitats further south in Washington (Stinson 2014), but not yet in British Columbia.
Conversely, after a century of successful fire suppression coniferous trees have expanded from higher elevations into lower elevation grasslands and sagebrush steppe in British Columbia (Bai et al. 2005; Gyug and Martens 2002; Turner and Krannitz 2000). Conifer encroachment also contributes to Greater Sage-Grouse habitat loss independently of the other factors, as proximity to trees negatively affects Greater Sage-Grouse behavior and vulnerability to predation (Baruch-Mordo et al. 2013; Prochazka et al. 2016; see Threat 8 below for more information).
Another complex natural system modification on-going and increasing in future, involves the invasion of cheatgrass facilitated by livestock grazing (Reisner et al. 2013), which then increases fuel loads and the frequency of fire (Baker 2011), which reduces sagebrush habitat for Greater Sage-Grouse (West and Yorks 2002; Connelly et al. 2004). Ironically though, after cheatgrass invasion has occurred, livestock grazing is one of the few tools available to reduce fuel-loads and the risk of wildfire while also protecting adjacent sagebrush habitat (Reisner et al. 2013). This natural system modification appears to be irreversible in the foreseeable future.
Invasion by non-native exotic plant species has changed the composition of sagebrush communities in the Okanagan Valley and across western North America. Replacement of native perennial bunchgrasses by cheatgrass has been shown to negatively correlate with habitat selection by Greater Sage-Grouse (Kirol et al. 2012). Invasion by cheatgrass has also resulted in dramatic increases in the number and frequency of fires that results in replacement of sagebrush by annual species, which provides little habitat value for Greater Sage-Grouse (Baker 2011; Condon et al. 2011). Cheatgrass is widespread in the Bunchgrass biogeoclimatic zone of British Columbia, and appears more common where soil disturbance has occurred in the past.
West Nile virus (WNv) is a current and future threat to Greater Sage-Grouse populations throughout North America. WNv was introduced into North America in 1999 (Reisen 2013) and was first documented in Greater Sage-Grouse in 2002 (Walker and Naugle 2011). Greater Sage-Grouse are highly susceptible to WNv and suffer high rates of mortality (Clark et al. 2006; McLean 2006). The primary vector of WNv in sagebrush ecosystems is the mosquito Culex tarsalis (Walker and Naugle 2011) and WNv persists through a mosquito-bird-mosquito infection cycle (McLean 2006), although bird-to-bird transmission has been observed (McLean 2006; Walker and Naugle 2011). Although mortality from WNv has been observed in Alberta (Naugle et al.2004), cooler ambient temperatures in more northerly locations may reduce the exposure risk of Greater Sage-Grouse to WNv in Canada. In British Columbia, WNv surveillance between 2005 and 2014 in the Okanagan Valley detected the virus in 2009, 2010, 2011 and 2013 (BC Centre for Disease Control 2014) demonstrating the virus occurs within the former range.
Many small and medium-sized predator populations have increased over the past century with the removal of larger predators like Grey Wolves (Canis lupus) and Grizzly Bears (Ursus arctos), and expansion of trees and irrigated agriculture. Now Coyote (Canis latrans), Red Fox (Vulpes vulpes), Raccoon (Procyon lotor), Great-horned Owl (Bubo virginianus), Common Raven (Corvus corax) and American Crow (Corvus brachyrynchos) are more abundant, and are known to be predators of Greater Sage-Grouse birds and eggs (Stinson et al. 2004; Environment Canada 2014). Conover and Roberts (2016) reviewed the evidence though, and suggest only Common Raven depredation of nests appears to be a range-wide threat.
Section 41 (2) of SARA requires that if the recovery of a listed wildlife species is not feasible, the recovery strategy must include an identification of the species' critical habitat to the extent possible. Critical habitat identification for the Greater Sage-Grouse phaios is not possible at this time. The former distribution of the species in British Columbia prior to 1918 is not well-known, and habitat sufficient and suitable for management or restoration does not appear to be available to support the species. Critical habitat for the Greater Sage-Grouse phaios in Canada is therefore not identified in this federal recovery strategy.
Recovery of Greater Sage-Grouse phaios is not considered technically and biologically feasible at the present time. Although the Province of British Columbia, Nature Conservancy of Canada, and The Nature Trust of British Columbia have substantial land holdings protected for conservation surrounding Osoyoos and along the border with the U.S.A., there remain many limitations. It is highly unlikely that habitat in Canada was ever sufficient in area or suitable in quality to sustain a population independently of a contiguous population in Washington State. More valley-bottom moist meadow habitat cannot feasibly be restored to support brood-rearing habitat. Translocation of 3200 birds into southern British Columbia and along a 100 km length of the Okanagan valley in northern Washington State to create a self-sustaining population is not part of the on-going U.S. Sage Grouse Initiative (NRCS 2015) nor the comprehensive conservation plan (Stinson et al. 2006). While translocations of Greater Sage-Grouse to supplement existing populations has met limited success (Baxter et al. 2008), translocation efforts for reintroduction of extirpated populations are most often met with failure (Schroeder and Vander Haegen 2011). Recovery of the species may become biologically and technically feasible if relict populations are discovered in Canada, and/or if translocation from viable populations in the United States becomes feasible and appropriate.
Any discovered or reintroduced Greater Sage-Grouse phaios populations in Canada would face several threats and limiting factors, as outlined previously in this document. Anthropogenic threats would need to be identified and addressed in survival and recovery habitats through habitat protection, restoration, and management; and continued management intervention would likely be required over the long term.
A strategic environmental assessment (SEA) is conducted on all SARA recovery planning documents, in accordance with the Cabinet Directive on the Environmental Assessment of Policy, Plan and Program Proposals. The purpose of a SEA is to incorporate environmental considerations into the development of public policies, plans, and program proposals to support environmentally sound decision-making and to evaluate whether the outcomes of a recovery planning document could affect any component of the environment or any of the Federal Sustainable Development Strategy's (FSDS) goals and targets.
Recovery planning is intended to benefit species at risk and biodiversity in general. However, it is recognized that strategies may also inadvertently lead to environmental effects beyond the intended benefits. The planning process based on national guidelines directly incorporates consideration of all environmental effects, with a particular focus on possible impacts upon non-target species or habitats. The results of the SEA are incorporated directly into the strategy itself, but are also summarized below in this statement.
The historic range of Greater Sage-Grouse phaios overlaps with that of several other rare species occurring in similar habitats of the south Okanagan valley in British Columbia, such as: American Badger (Taxidea taxus jeffersonii), Behr's Hairstreak (Satyrium behrii), Burrowing Owl (Athene cunicularia), Common Nighthawk (Chordeiles minor), Desert Nightsnake (Hypsiglena chlorophaea), Grand Coulee Owl-clover (Orthocarpus barbatus), Great Basin Gophersnake (Pituophis catenifer deserticola), Great Basin Spadefoot (Spea intermontana), Half-moon Hairstreak (Satyrium semilunar), Lyall's Mariposa Lily (Calochortus lyallii), Morman Metalmark (Apodemia mormo), Northern Rubber Boa (Charina bottae), Nuttall's Cottontail (Sylvilagus nuttallii nuttallii), Okanagan Efferia (Efferia okanagana), Sage Thrasher (Oreoscoptes montanus), Western Harvest Mouse (Reithrodontomys megalotis megalotis), Western Rattlesnake (Crotalus oreganus), Western Skink (Plestiodon skiltonianus), Western Tiger Salamander (Ambystoma mavortium), Western Toad (Anaxyrus boreas), Western Yellow-bellied Racer (Coluber constrictor mormon), and Yellow-breasted Chat (Icteria virens). If a relictual population of Greater Sage-Grouse phaios is discovered and/or if reintroduction of the species is considered, recovery planning impacts on non-target recovery species in British Columbia will need to be taken into account. Any recovery planning activities for the Greater Sage-Grouse phaios will be implemented with consideration of all co-occurring species at risk, to avoid or minimize negative impacts to these species and their habitats.
Aldridge, C.L. 1998. Status of Sage-Grouse (Centrocercus urophasianus urophasianus) in Alberta. Wildlife Status Report No. 13. Alberta Environmental Protection, Wildlife Management Division, and Alberta Conservation Association, Edmonton, Alberta. 23 pp.
Aldridge, C.L. 2000. Reproduction and habitat use by Sage-Grouse (Centrocercus urophasianus) in a northern fringe population. M.Sc. thesis. University of Regina. Regina, Saskatchewan. 109 pp.
Aldridge, C.L. and R.M. Brigham. 2003. Distribution, status and abundance of Greater Sage-Grouse, Centrocercus urophasianus, in Canada. Canadian Field Naturalist 117:25–34.
Bai, Y., B. Roddan, D.A. Hill, K. Broersma, and D. Thompson. 2005. Quantifying tree cover in the forest-grassland ecotone of British Columbia using crown delineation and pattern detection. Forest Ecology and Management 212(1–3):92–100.
Baker, W.L., 2011, Pre- Euro-American and recent fire in sagebrush ecosystems, in Knick, S.T. and Connelly, J.W., eds., Greater sage-grouse: ecology and conservation of a landscape species: Berkeley, Calif., University of California Press, p. 185–202 .
Barnett, J.F. and J.A. Crawford. 1994. Pre-laying nutrition of Sage-Grouse hens in Oregon. Journal of Range Management 47: 114–118.
Baruch-Mordo, S., Evans, J.S., Severson, J.P., Naugle, D.E.,Maestas, J.D., Kiesecker, J.M., Falkowski, M.J., Hagen, C.A., and K.P. Reese. 2013. Saving sage-grouse from the trees: a proactive solution to reducing a key threat to a candidate species. Biological Conservation 167: 233–241.
Baxter, R.J., J.T. Flinders, and D.L. Mitchell. 2008. Survival, movements, and reproduction of translocated Greater Sage-Grouse in Strawberry Valley, Utah. Journal of Wildlife Management 72:179–186.
Beck, T.D.I. 1977. Sage-Grouse flock characteristics and habitat in winter. Journal of Wildlife Management 41: 18–26.
Beck, J.L. and D.L. Mitchell. 2000. Influences of livestock grazing on sage grouse habitat. Wildl. Soc. Bull. 28:993–1002
Benedict, N.G., S.J. Oyler-McCance, S.E. Taylor, C.E. Braun, and T.W. Quinn. 2003. Evaluation of the eastern (Centrocercus urophasianus urophasianus) and western (Centrocercus urophasianus phaios) subspecies of sage-grouse using mitochondrial control-region sequence data. Conservation Genetics 4: 301-310.
Bergerud, A.T. 1988. Population ecology of North American grouse. Pages 578–648 in A.T. Bergerud and M.W. Gratson [eds.], Adaptive strategies and population ecology of northern grouse. University of Minnesota Press, Minneapolis, Minnesota.
Bezener, A.M., M.Dunn, O. Dyer, R. Hawes, T. Hayes, H. Richardson, and B. White. 2006. Draft Recovery Strategy for Species At Risk in the South Okanagan and Lower Similkameen Valleys of British Columbia, Canada: Towards Integrating the Landscape-Level and Single-Species Approaches to Conservation, Vol 1. South Okanagan-Similkameen Conservation Program, Penticton, BC. 67 pp.
Blomberg, E. J., J. S. Sedinger, D. Gibson, P. S. Coates, and M. L. Casazza. 2014. Carryover effects and climatic conditions influence the postfledging survival of Greater Sage-Grouse. Ecology and Evolution 4:4488–4499.
Braun, C.E. 1998. Sage-Grouse declines in western North America: What are the problems? Proceedings of the Western Association of State Fish and Wildlife Agencies 78: 139–156.
Braun, C.E., T. Britt, and R.O. Wallestad. 1977. Guidelines for maintenance of Sage-Grouse habitats. Wildlife Society Bulletin 5(3): 99–106.
Braun, C.E., O.O. Oedekoven, and C.L. Aldridge. 2002. Oil and gas development in western North America: Effects on sagebrush steppe avifauna with particular emphasis on Sage-Grouse. Transactions of the North American Wildlife and Natural Resources Conference 67: 337–349.
B.C. Centre for Disease Control. 2014. West Nile Virus Surveillance Reports (2004 to 2014). Vancouver. B.C. [Accessed August 9, 2016].
B.C. Conservation Data Centre. 2015. B.C. Species and Ecosystems Explorer. B.C. Ministry of Environment, Victoria, B.C. [Accessed June 18, 2015].
B.C. Conservation Framework. 2015. Conservation Framework Summary: Pituophis catenifer catenifer. B.C. Ministry of the Environment. Available: http://a100.gov.bc.ca/pub/eswp/ [Accessed June 18, 2015].
Campbell, R.W. and G.R. Ryder. 2010. Greater Sage-Grouse (Centrocercus urophasianus) in British Columbia: History, Translocation, Breeding, and Current Status. Wildlife Afield 7(1):3-11.
Cannings, R.A., R.J. Cannings and S.G. Cannings. 1987. Birds of the Okanagan Valley, British Columbia. Royal British Columbia Museum, Victoria, BC.
Clark, L., J. Hall, R. McLean, M. Dunbar, K. Klenk, R. Bowen, and C.A. Smeraski. 2006. Susceptibility of Greater Sage-Grouse to experimental infection with West Nile virus: Journal of Wildlife Diseases, 42: 14–22.
Coggins, K.A. 1998. Relationship between habitat changes and productivity of Sage-Grouse at Hart Mountain National Antelope Refuge, Oregon. M.Sc. thesis, Oregon State University, Corvallis, Oregon. 61 pp.
Condon, L., P.J. Weisberg, and J.C. Chambers. 2011, Abiotic and biotic influences on Bromus tectorum invasion and Artemisia tridentata recovery after fire: International Journal of Wildland Fire, v. 20, p. 597–604.
Connelly, J.W., H.W. Browers, and R.J. Gates. 1988. Seasonal movements of Sage-Grouse in southeastern Idaho. Journal of Wildlife Management 52(1): 116–122.
Connelly, J.W., M.A. Schroeder, A.R. Sands, and C.E. Braun. 2000. Guidelines to manage Sage-Grouse populations and their habitats. Wildlife Society Bulletin 28(4): 967–985.
Connelly, J.W., S.T. Knick, M.A. Schroeder, and S.J. Stiver. 2004. Conservation assessment of greater Sage-Grouse and sagebrush habitats. Western Association of Fish and Wildlife Agencies. Unpublished Report, Cheyenne, Wyoming. 610 pp.
Connelly, J.W., E.T. Rinkes, and C.E. Braun. 2011. Characteristics of Greater Sage grouse habitats: a landscape species at micro- and macro scales. Pages 69–84 in S.T. Knick and J.W. Connelly (eds). Ecology and Conservation of Greater Sage grouse: A Landscape Species and its Habitats. Studies in Avian Biology (38). University of California Press, Berkeley, California.
Conover, M.R. and A.J. Roberts. 2016. Predators, predator removal, and sage-grouse: a review. Journal of Wildlife Management. 80: 10.1002.
CMP (Conservation Measures Partnership). 2010. Threats taxonomy. [accessed August, 2015].
COSEWIC. 2008. COSEWIC assessment and update status report on the Greater Sage-Grouse Centrocercus urophasianus, Phaios subspecies and Urophasianus subspecies, Centrocercus urophasianus urophasianus, in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 38 pp. (www.sararegistry.gc.ca/status/status_e.cfm).
Crawford, J.A., R.A. Olson, N.E. West, J.C. Mosley, M.A. Schroeder, T.D. Whitson, R.F. Miller, M.A. Gregg, and C.S. Boyd. 2004. Ecology and management of sage-grouse and sage-grouse habitat. Rangeland Ecology and Management. 57: 2-19.
Dalke, P.D., D.B. Pyrah, D.C. Stanton, J.E. Crawford, and E.F. Schlatterer. 1963. Ecology, productivity and management of Sage-Grouse in Idaho. Journal of Wildlife Management 27(4): 811–841.
DeLong, A.K., J.A. Crawford, and D.C. DeLong Jr. 1995. Relationship between vegetational structure and predation of artificial Sage-Grouse nests. Journal of Wildlife Management 59(1): 88–92.
Doherty, K.E., J.S. Evans, P.S. Coates, L.M. Juliusson, and B.C. Fedy. 2016. Importance of regional variation in conservation planning: a rangewide example of the Greater Sage-Grouse. Ecosphere. 7(10): e01462.
Doherty, K.E., D.E. Naugle, B.L. Walker, and J.M. Graham. 2008. Greater sage-grouse winter habitat selection and energy development. Journal of Wildlife Management. 72: 187-195.
Drut, M.S., J.A. Crawford, and M.A. Gregg. 1994a. Brood habitat use by Sage-Grouse in Oregon. Great Basin Naturalist 54(2): 170–176.
Drut, M.S., W.H. Pyle, and J.A. Crawford. 1994b. Diets and food selection of Sage-Grouse chicks in Oregon. Journal of Range Management 47: 90–93.
Eng, R.L. and P. Schladweiler. 1972. Sage-Grouse winter movements and habitat use in central Montana. Journal of Wildlife Management 36(1): 141–146.
Environment Canada. 2014. Amended Recovery Strategy for the Greater Sage-Grouse (Centrocercus urophasianus urophasianus) in Canada. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa.vi + 53pp.
Fedy, B.C., K.E. Doherty, C.L. Aldridge, M. O'Donnell, J.L. Beck, B. Bedrosian, D. Gummer, J.J. Holloran, G.D. Johnson, and N.W. Kaczor. 2014. Habitat prioritization across large landscapes, multiple seasons, and novel areas: an example using Greater Sage-Grouse in Wyoming. Wildlife Monographs. 190: 1-39.
Gyug, L., and G. Martens. 2002. Forest canopy changes from 1947 to 1996 in the Lower Similkameen, British Columbia. Forest Renewal BC, Victoria, B.C.
Hagen, C.A. 1999. Sage-Grouse habitat use and seasonal movements in a naturally fragmented landscape, Northwestern Colorado. M.Sc. thesis, University of Manitoba, Winnipeg, Manitoba. 136 pp.
Hamann, A., and T. Wang. 2006. Potential effects of climate change on ecosystem distribution and tree species distribution in British Columbia. Ecology 87:2773–2786.
Hanf, J.M., P.A. Schmidt, and E.B. Groshens. 1994. Sage-Grouse in the High Desert of central Oregon: Results of a study, 1988–1993. U.S. Department of the Interior, Bureau of Land Management, Prineville, Oregon. 57 pp.
Huwer, S.L. 2004. Evaluating greater Sage-Grouse brood habitat using human-imprinted chicks. M.Sc. thesis. Colorado State University, Fort Collins, Colorado. 85 pp.
Hyslop, Colleen (ed.). 1998 (based on draft 1997 report). COSEWIC status report on the Sage-Grouse Centrocercus urophasianus urophasianus (Prairie population) Centrocercus urophasianus phaios (B.C. population) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-34 pp.
Iverson, K., D. Curran, T. Fleming, and A. Haney. 2008. Sensitive ecosystems inventory Okanagan Valley: Vernon to Osoyoos 2000-2007, Methods, ecological descriptions, results and conservation tools. Canadian Wildlife Service, Technical Report Series No. 495. Environment Canada, Pacific and Yukon Region. Delta BC.
Jenni, D.A. and J.E. Hartzler. 1978. Attendance at a Sage-Grouse lek: Implications for spring censuses. Journal of Wildlife Management 42: 46–52.
Kirol, C.P., J.L. Beck, J.B. Dinkins, and M.R. Conover. 2012, Microhabitat selection for nesting and brood-rearing by the Greater Sage-Grouse in xeric big sagebrush: Condor, v. 114, p. 75–89.
Lea, T. 2008. Historical (pre-European settlement) ecosystems of the Okanagan Valley and Lower Similkameen Valley of British Columbia – pre-European contact to the present. Davidsonia 19: 3-36.
Lungle, K. and S. Pruss. 2008. Recovery Strategy for the Greater Sage-Grouse (Centrocercus urophasianus urophasianus) in Canada. In Species at Risk Act Recovery Strategy Series. Parks Canada Agency. Ottawa. vii + 43 pp.
McLean, R.G. 2006. West Nile virus in North American birds. Ornithological Monographs 60: 44–64.
Meidinger, D and J. Pojar. 1991. Ecosystems of British Columbia, B.C. Ministry of Forests, Victoria, BC. Special Report No. 6. 330 pp.
Moynahan, B.J., M.S. Lindberg, and J.W. Thomas. 2006. Factors contributing to process variance in annual survival of female greater Sage-Grouse in Montana. Ecological Applications 16(4): 1529–1538.
NatureServe. 2015. NatureServe Explorer: an online encyclopedia of life [web application]. Version 7.1. Arlington, V.A. [Accessed June 18, 2015].
Naugle, D.E., C.L. Aldridge, B.L. Walker, T.E. Cornish, B.J. Moynahan, M.J. Holloran, K.Brown, G.D. Johnson, E.T. Schmidtman, R.T. Mayer, C.Y. Kato, M.R. Matchett, T.J. Christiansen, W.E. Cook, T. Creekmore, R.D. Falise, E.T. Rinkes, and M.S. Boyce. 2004. West Nile virus: Pending crisis for Greater Sage-Grouse. Ecology Letters 7:704-713.
Naugle, D.E., K.E. Doherty, B.L. Walker, M.J. Holloran, and H.E. Copeland. 2011. Energy development and greater sage grouse. Pages 489–503 in S.T. Knick and J.W. Connelly (eds). Ecology and Conservation of Greater Sage Grouse: A Landscape Species and its Habitats. Studies in Avian Biology (38). University of California Press, Berkeley, California.
NRCS (Natural Resource Conservation Service). 2015. Sage Grouse Initiative 2.0 Investment Strategy, FY 2015-2018. United States Department of Agriculture, Natural Resource Conservation Service. Washington D.C.
Oyler-McCance, S.J., S.E. Taylor, and T.W. Quinn. 2005. A multilocus population genetic survey of the Greater Sage-Grouse across their range. Molecular Ecology 14:1293–1310.
Parks Canada Agency. 2009. "Replacement of Section 2.6 of the Recovery Strategy for the Sage Grouse (Centrocercus urophasianus urophasianus) in Canada" as found in: Lungle, K. and S. Pruss. 2008. Recovery Strategy for the Greater Sage-Grouse (Centrocercus urophasianus urophasianus) in Canada. In Species at Risk Act Recovery Strategy Series. Parks Canada Agency. Ottawa. vii + 43 pp.
Patterson, R.L. 1952. The Sage-Grouse in Wyoming. Sage Books, Denver, Colorado. 399 pp.
Prochazka, B.G., P.S. Coates, M.A. Ricca, M.L. Casazza, K.B. Gustafson, and J.M. Hull. 2016. Encounters with pinyon-juniper influence riskier movements in Greater Sage-Grouse across the Great Basin. Rangeland Ecology and Management.
Reisen, W.K. 2013. Ecology of West Nile Virus in North America. Viruses 5(9):2079–2105.
Reisner, M.D., J.B. Grace, D.A. Pyke, and P.S. Doescher. 2013. Conditions favouring Bromus tectorum dominance of endangered sagebrush steppe ecosystems. Journal of Applied Ecology. 50: 1039-1049.
Salafsky, N., D. Salzer, A. J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S. H. M. Butchart, B. Collen, N. Cox, L. L. Master, S. O'Connor, and D. Wilkie. 2008. A Standard Lexicon for Biodiversity Conservation: Unified Classifications of Threats and Actions. Conservation Biology 22:897–911.
Scott, J.W. 1942. Mating behavior of the sage grouse. Auk 59:477–498.
Schroeder, M.A., J.R. Young, and C.E. Braun. 1999. Sage-Grouse, Centrocercus urophasianus. In The Birds of North America. No. 425 (A. Poole and F. Gills eds.). The Birds of North America Inc., Philadelphia, PA.
Schroeder, M.A., C.L. Aldridge, A.D. Apa, J.R., Bohne, C.E. Braun, S.D. Bunnell, J.W. Connelly, P.A. Diebert, S.C. Gardner, M.A. Hilliard, G.D. Kobriger, S.M. McAdam, C.W. McCarthy, J.J. McCarthy, D.L. Mitchell, E.V. Rickerson, and S.J. Stiver. 2004. Distribution of Sage-Grouse in North America. Condor 106(2): 363–376.
Schroeder, M. A., and W. M. Vander Haegen. 2011. Response of Greater Sage-Grouse to the conservation reserve program in Washington State. Pp. 517–529 in S. T. Knick and J. W. Connelly (editors). Greater Sage-Grouse: ecology and conservation of a landscape species and its habitats. Studies in Avian Biology (vol. 38), University of California Press, Berkeley, CA.
Stinson, C.M., D.W. Hays, and M.A. Schroeder. 2004. Washington State recovery plan for the Greater Sage-Grouse. Washington Department of Fish and Wildlife. Olympia WA.
Stinson, C.M. 2014. Report on Conservation Efforts in Response to Threats to Greater Sage-grouse in Washington: An evaluation of Washington State's efforts to address threats to the viability of Sage-grouse listed in the Conservation Objective Team (COT) report (USFWS 2013). Washington Department of Fish and Wildlife, Olympia, Washington. 26 pp.
Stiver, S., A. Apa, J. Bohne, S. Bunnell, P. Deibert, S. Gardner, M. Hilliard, C. McCarthy, and M. Schroeder. 2006. Greater sage-grouse comprehensive conservation strategy. Western Association of Fish and Wildlife Agencies. Cheyenne WY.
Tack, J.D. 2009. Sage-grouse and the human footprint: implications for conservation of small and declining populations. MSc. Thesis. University of Montana, Missoula, Montana. 96 pp.
Turner, J., and P. Krannitz. 2000. Tree encroachment in the south Okanagan and lower Similkameen valleys of British Columbia. In: Proceedings, From science to management and back: A science forum for southern interior ecosystems of British Columbia. C. Hollstedt, K. Sutherland, and T. Innes (editors). Southern Interior Forest Extension and Research Partnership, Kamloops, B.C. SIFERP Series No. 1, pp. 81–83.
U.S. Fish and Wildlife Service (USFWS). 2003. Candidate assessment and listing priority assingment form: Centrocercus urophasianus, greater sage grouse (Columbia Basin distinct population segment). 27 pp.
Walker, B.E. and D.E. Naugle. 2011. West Nile Virus ecology in sagebrush habitat and impacts on Greater Sage-Grouse populations. In: Knick ST, Connelly JW, editors. Greater Sage-Grouse: ecology and conservation of a landscape species and its habitats. Berkeley, CA.: University of California Press
Wallestad, R.O. 1971. Summer movements and habitat use by Sage-Grouse broods in central Montana. Journal of Wildlife Management 35: 129–136.
Wallestad, R., J.G. Peterson, and R.L. Eng. 1975. Foods of adult Sage-Grouse in central Montana. Journal of Wildlife Management 39(3): 628–630.
Walsh, D.P., J.R., Stiver, G.C. White, T.E. Remington, and A.D. Apa. 2010. Population estimation techniques for lekking species. Journal of Wildlife Management 74(7): 1607-1613.
Wang, T., E.M. Campbell, G.A. O'Neill, and S.N. Aitken. 2012. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. Forest Ecology and Management 279:128–140.
West, N.E., and T.P. Yorks. 2002, Vegetation responses following wildfire on grazed and ungrazed sagebrush semi-desert: Journal of Range Management, v. 55, p. 171–181.