
Cyanobacteria and their Toxins in Recreational Water: Guideline Technical Document for Public Consultation

Purpose of consultation
This guideline technical document evaluated the available information on cyanobacteria and their toxins with the intent of updating/recommending guideline value(s) for cyanobacteria toxins, total cyanobacteria cell counts, total cyanobacteria biovolume, and chlorophyll-a in recreational water. The purpose of this consultation is to solicit comments on the proposed guideline values, on the approach used for their development, and on the potential economic costs of implementing the guidelines.
The document was reviewed by external experts and subsequently revised. We now seek comments from the public. This document is available for a 90-day public consultation period.
Please send comments (with rationale, where required) to Health Canada via email at HC.water-eau.SC@canada.ca. If this is not feasible, comments may be sent by mail to:
Water and Air Quality Bureau, Health Canada,
269 Laurier Avenue West, A.L. 4903D,
Ottawa, Ontario K1A 0K9.
All comments must be received before November 20, 2020. Comments received as part of this consultation will be shared with the recreational water quality working group members, along with the name and affiliation of their author. Authors who do not want their name and affiliation shared with recreational water quality working group members should provide a statement to this effect along with their comments.
It should be noted that this guideline technical document will be revised following the evaluation of comments received, and the recreational water quality guideline will be updated, if required. This document should be considered as a draft for comment only.
Table of Contents
- Foreword
- Management of cyanobacteria and their toxins in recreational waters
- 1.0 Guideline values
- 2.0 Application of the guideline
- 3.0 Description and Health Effects
- 4.0 Routes of Exposure
- 5.0 Occurrence in the environment
- 6.0 Analytical methods
- 7.0 Rationale
- 8.0 References
- Appendix A: List of acronyms
- Appendix B: Flow chart for monitoring cyanobacteria and their toxins
Foreword
The Guidelines for Canadian Recreational Water Quality are comprised of multiple guideline technical documents that consider the various factors that could interfere with the safety of recreational waters from a human health perspective. They provide guideline values for specific parameters used to monitor water quality hazards, and recommend monitoring and risk management strategies. Recreational waters are considered to be any natural fresh, marine or estuarine bodies of water that are used for recreational purposes; this includes lakes, rivers, and human-made constructions (e.g., quarries, artificial lakes) that are filled with untreated natural waters. Jurisdictions may choose to apply these guidelines to other natural waters that are applying limited treatment (e.g., short-term disinfection for an athletic event). Recreational activities that could present a human health risk through intentional or incidental immersion and ingestion include primary contact activities (e.g., swimming, bathing, wading, windsurfing and waterskiing) and secondary contact activities (e.g., canoeing, boating or fishing).
Each guideline technical document has been established based on current, published scientific research related to health effects, aesthetic effects, and beach management considerations. The responsibility for recreational water quality generally falls under provincial and territorial jurisdiction, therefore the policies and approaches regarding the monitoring of cyanobacterial blooms and cyanotoxin concentrations, as well as the resulting management decisions, may vary between jurisdictions. The guideline technical documents are intended to guide decisions by provincial, territorial and local authorities that are responsible for the management of recreational waters.
For a complete list of the guideline technical documents available, please refer to the Guidelines for Canadian Recreational Water Quality summary document available on the Canada.ca website (in publication). For issues related to drinking water, please consult the Guidelines for Canadian Drinking Water Quality – Guideline Technical Document for Cyanobacterial toxins (Health Canada, 2017).
Management of cyanobacteria and their toxins in recreational waters
This document outlines guideline values and select strategies for managing health risks related to exposure to cyanobacteria (also known as blue-green algae) and their toxins. Most scientific studies on cyanobacterial toxins focus on microcystins, as they are regarded as the most prevalent and significant of the freshwater cyanotoxins. Other cyanotoxins, such as anatoxin-a and cylindrospermopsin, have more limited information available.
A risk management approach that focuses on the identification and control of water quality hazards and their associated risks before the point of contact with the recreational water user represents the best strategy for the protection of public health. More details on the risk management of recreational water quality are available in the Guidelines for Canadian Recreational Water Quality - Understanding and Managing Risks in Recreational Waters technical document (Health Canada, in preparation).
1.0 Guideline values
The guideline values for cyanobacteria and their toxins are divided into (1) direct measures for cyanotoxins (cyanotoxin guideline value) and (2) indicators of the potential presence of cyanotoxins (total cyanobacteria, total cyanobacterial biovolume, total chlorophyll a). The different measures offer a flexible approach to understanding potential bloom toxicity and can be used alone or in combination, depending on the jurisdiction and the recreational water quality management plan in place. Guidance on applying the guideline values can be found in Section 2.0.
1.1 Cyanotoxins
The proposed guideline value for total microcystins in recreational waters is a maximum concentration of 10 µg/L.
Total microcystins guideline value: 10 µg/L
When measuring microcystins, it is important to measure total microcystins. This includes microcystins that are both dissolved in the water (extracellular) and bound within the cyanobacterial cells (intracellular). In addition, although the guideline value is based on the toxicity of microcystin-LR (MC-LR), all measurable microcystin variants, not just MC-LR, should be included in the analysis.
Total microcystins is the only cyanotoxin guideline currently established in this document. Most scientific studies on cyanobacterial toxins focus on microcystins, as they are regarded as the most prevalent and significant of the cyanotoxins. Guideline values for other cyanotoxins, including anatoxin-a and cylindrospermopsin, have not been established in this document as health or exposure data on other toxins are limited.
1.2 Indicators of the potential presence of cyanotoxins
The indicators of the potential presence of cyanotoxins are based on measures of biomass for planktonic cyanobacteria. They are derived from the cyanotoxins guideline for microcystins and use the microcystins content of Microcystis. The indicators include total cyanobacteria, total cyanobacterial biovolume, and total chlorophyll a. The proposed guideline values are the following:
- Total cyanobacteria: 50 000 cells/mL
- Total cyanobacterial biovolume: 4.5 mm3/L
- Total chlorophyll a: 33 µg/L
These measures can be used alone or in combination. The choice of measure used can vary between recreational water locations and will be the decision of the responsible authority. See Section 2.0.
These guideline values can be used to indicate a planktonic bloom is present and that cyanotoxins, if they are present, may exceed the guideline value for total microcystins. Further details on using these indicators are provided in Section 2.0. Guideline values for indicators of benthic cyanobacteria proliferation have not yet been developed. Further information on benthic populations is available in Section 2.0.
2.0 Application of the guideline
The recommendations in this section are intended to provide flexibility for authorities responsible for the management of recreational waters to develop site-specific approaches for cyanobacteria management. Site-specific approaches allow public health protection while avoiding unnecessary closures of recreational areas. The assessment of risk and the resultant decision on how to manage cyanobacteria and their toxins should be detailed as part of a management plan for the recreational area. Further information on managing recreational water quality can be found in the accompanying document on Understanding and Managing Risks in Recreational Water (Health Canada, in preparation).
The guideline values for cyanobacteria and their toxins are divided into (1) direct measures for cyanotoxins and (2) monitoring for indicators of the potential presence of cyanotoxins. The only direct measure of health risk is the level of total microcystins in water from the suspected areas. Therefore, a health-based value (HBV) for total microcystins has been derived. The HBV for total microcystin is considered protective for all Canadians, including the most vulnerable populations.
Total cyanobacteriaand total cyanobacterial biovolume are measures of planktonic cyanobacteria biomass and total chlorophyll a is a measure of total phytoplankton biomass. These biomass values are derived based on their relationship with total microcystins, using conservative assumptions (see Section 7), and are used to indicate potential bloom toxicity. Applying these indicators may allow greater temporal and spatial coverage of a planktonic bloom as these methods may be more accessible than microcystin analysis in many recreational water areas. These guideline values are not intended as fixed values that should be applied to all recreational areas. The choice of biomass value used can vary between recreational water locations and will be the decision of the responsible authority. Information on their advantages and limitations is detailed in Section 5.5. The responsible authority can also set site-specific values for the indicator parameters using local knowledge to better reflect the risks encountered in the specific recreational environment. Responsible authorities may also use other methods to indicate potential bloom toxicity. For example, molecular methods can be used to determine if toxin-producing species are present. As mentioned earlier, site-specific values have the advantage of both protecting public health and promoting public health by avoiding unnecessary closures of recreational areas. Indicators have not yet been developed for benthic cyanobacteria, however, monitoring is still recommended (see section 2.1).
A flow chart overview of the general approach to monitoring planktonic cyanobacteria in recreational waters can be found in Appendix B. This flow chart is intended as a general guide. Site-specific knowledge and various local factors may influence the suitability of this general approach and therefore application of the cyanobacteria guideline values may vary between jurisdictions.
2.1 Monitoring
An appropriate monitoring program reduces the risk of user exposure to cyanobacterial blooms and their toxins. In Canada, there is an abundance of rivers and lakes that are used for recreational activities, and monitoring them all for cyanobacterial blooms is not feasible or recommended. Instead, responsible authorities should use criteria to help evaluate the risk of bloom formation to identify the areas that are at greater risk. Water quality characteristics (e.g., pH, total phosphorus concentrations) and historical information on cyanobacterial blooms in the watershed, including where scums have accumulated in the past, should be included as part of the assessment criteria. The types of recreational activities that are taking place in the area and the level of exposure individuals would have in the event of a cyanobacteria bloom need to be considered. This information can then be used to prioritize areas that should be monitored, and determine a monitoring approach (e.g., what to monitor, how often). Consideration should also be given to the intensity of sampling that is necessary for characterizing a waterbody and when this monitoring can be reduced based on an understanding of the site-specific conditions that may lead to cyanobacteria blooms. The recreational areas that are not selected for monitoring are generally those that represent a lower risk of human exposure to cyanobacteria blooms. In these areas, investigation of possible bloom occurrence could be triggered by citizens reporting unusual water quality characteristics.
Recreational water areas that are heavily used and that are suspected or are known to be susceptible to blooms should be routinely monitored as described in their monitoring plan (e.g., weekly or bi-weekly) and have an action plan in place for what measures to take in the event of a toxic bloom. It is very difficult to establish an action plan during a bloom event; prior discussion with local groups (e.g., other potentially affected parties, wildlife or agriculture agencies, analytical laboratories) is important to develop an appropriate action plan for use when/if it is needed.
Visually monitoring recreational water areas for accumulations of plankton, or bloom development is usually the first step in a monitoring program (see Figure 1). Visual inspection is extremely valuable as cyanobacteria are usually readily visible if they are at potentially hazardous concentrations. Cyanobacteria blooms may not be visually distinguishable from blooms of other phytoplankton, and confirmation that the samples contain cyanobacteria may require further testing (e.g., microscopy, molecular methods, cyanotoxin analysis). It is also not possible to determine if a cyanobacteria bloom contains toxins by visual inspection; samples must be sent to a laboratory for analysis. Some species of cyanobacteria can increase to high cell count values without producing surface scums. In clear shallow areas, the presence of benthic mats should also be visually assessed. Under certain environmental conditions, these mats can detach from the substrate or may be stranded when water recedes and accumulate along shores where they are more accessible to humans and animals. Publications are available that provide visual examples of cyanobacteria blooms and benthic cyanobacteria mats; these can be used to help assess local conditions (Huynh and Seredak, 2006; Blais, 2008; New Zealand Ministry for the Environment and Ministry of Health, 2009; Rosen and St Amand, 2015; Wood et al., 2015; California Water Quality Monitoring Council, 2019).
Figure 1: Monitoring steps for cyanobacteria blooms
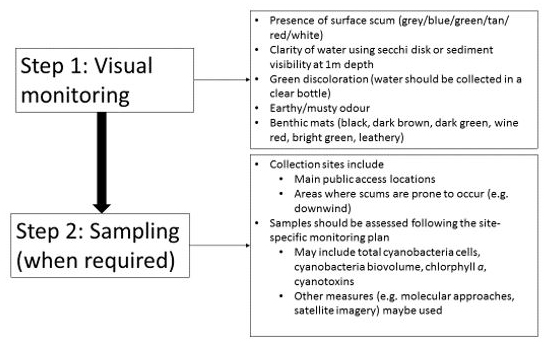
Text Description
This figure displays the two monitoring steps for identifying cyanobacteria blooms. The first step is visual monitoring. The visual monitoring step includes five things to look for: Presence of surface scum (grey/blue/green/tan/red/white); clarity of water using secchi disk or sediment visibility at 1 meter depth; green discoloration (water should be collected in a clear bottle); earty/musty odour; and benthic mats (black, dark brown, dark green, wine red, bright green, leathery). The second step is sampling (when required). The second step includes information on two topics: (1) Collection sites which includes the main public access locations and areas where scums are prone to occur (e.g. downwind); and (2) samples should be assessed following the site-specific monitoring plan which may include assessing total cyanobacteria cells, cyanobacteria biovolume, chlorophyll a, cyanotoxins, or using other measures such as molecular approaches or satellite imagery.
Samples may be collected for visual inspection (as described above), to assess the concentration of planktonic cyanobacteria (e.g., total cyanobacteria cells, cyanobacteria biovolume), of phytoplankton (e.g., chlorophyll a), to determine toxin levels, or some combination of these measures. Other measures, such as molecular approaches or using satellite imagery, may also be included in the monitoring plan, although these methods should first be validated for the site. It should be noted that not only cell density but also the level of toxicity within a bloom can vary considerably both temporally and spatially, particularly within large blooms, making it difficult to accurately characterize the toxin concentrations. In some locations, assessing samples for cyanobacteria or phytoplankton biomass and cyanobacteria toxins may present technical challenges, including lack of access to the necessary laboratory expertise and extended wait times for sampling results. In these situations, a conservative approach based only on visual inspection may be necessary. However, as mentioned previously, it is not possible to tell if a cyanobacteria bloom or a cyanobacteria benthic mat contains toxins by visual examination alone. Therefore, relying solely on visual examination assumes all blooms are toxic and can lead to unnecessary beach closures. By implementing a flexible approach to cyanobacteria management, responsible authorities should be able to address some of these challenges and continue to protect and promote public health. Further information on designing and implementing recreational water monitoring programs can be found in various publications (e.g., Chorus and Bartram, 1999; Newcombe, 2009).
A water body in which a toxic bloom has developed may contain toxins for a period after the bloom has dissipated. The length of time toxins remain a concern will be dependent on numerous factors, such as the dilution rate of the area, the type of toxin present, and the rate of biodegradation. In order to fully characterize the extent of the risk posed by the cyanobacterial population, authorities should conduct sampling during, and after the collapse of, the bloom in accordance with the recreational water management plan.2.2 Notifications
Waters in which a bloom has developed, or waters shown to exceed the total microcystin guideline value, may result in human exposure to cyanobacteria or cyanotoxins in amounts harmful to human health. Due to the difficulty in accurately characterizing the concentrations of toxins in a bloom, contact with recreational waters that contain visible blooms should be avoided as a precaution, and a swimming/contact advisory may be issued at the discretion of the responsible authority. In the case of benthic mats, the extent and location of the mats may prompt warning signs of the potential risks or result in advice to avoid the area for recreational activities. Similar to blooms, confirmation of the presence of toxins, or toxin producing cyanobacterial species in the benthic mats will require laboratory analysis.
Contact with waters where an advisory has been issued should be avoided until the advisory has been rescinded. Educational materials outlining steps the public may take to reduce their personal risk in the event of a bloom should be provided. Pets should also not swim in, or drink from, areas where the water has taken on an abnormal discolouration consistent with that of a bloom, or where accumulations of cyanobacterial material, including benthic mats, are visible. In the case of accidental contact with cyanobacterial material, users should shower or wash themselves, as well as any items that may have come into contact with the cyanobacterial material, as soon as is practical upon exiting the water. Any user experiencing adverse health effects from recreational water activity should consult a medical professional and, if necessary, alert the appropriate authorities.
The swimming/contact advisory should remain in place until the area has been determined to be safe for recreational activities. The dissipation of the bloom along with a determination that the cyanobacteria toxin is below the guideline value can be used to remove the swimming/contact advisory. In the absence of toxin testing, the swimming/contact advisory should remain in place for a period after the bloom has dissipated to allow any toxin present to be diluted or degraded. The length of time required for toxin dissipation will depend on numerous factors such as dilution and biodegradation rate and will need to be determined on a site-specific basis. The conditions required to remove the contact advisory should be determined by the responsible authority based on the cyanobacteria management plan in place for the recreational area, or, in the absence of a recreational monitoring plan, based on site-specific information from the recreational area (e.g., dilution, historical occurrence of blooms).
3.0 Description and Health Effects
3.1 Cyanobacteria
Cyanobacteria are bacteria that share features with algae, such as oxygen-producing photosynthesis using their blue-green photosynthetic pigments; hence, historically they have been termed blue-green algae (WHO, 2003). Blooms with intact, active cells are generally more green than blue in appearance, although other colors ranging from tan to bright red or wine color can also occur. Blooms with dying cells can appear bluer. This is because bacteriochlorophyll, the pigment responsible for the green colour, is rapidly bleached by sunlight following cell lysis while the blue pigment (phycocyanin) persists (Newcombe, 2009). Under the microscope, most planktonic cyanobacteria, including the species found in Canadian lakes, appear as regular or irregular groupings of cells or as filamentous chains that can be straight, coiled or branched (Chorus and Bartram, 1999; Falconer, 2005). In a typical summer, a lake water sample can contain numerous species of cyanobacteria, both toxic and non-toxic strains, along with species of algae. The conditions that favour bloom formation include eutrophic waters and higher water temperatures (leading to a more stable stratification of the water column) (Huisman et al., 2018).
Many cyanobacterial cells can alter position within the water column through changes in cell buoyancy or mixing, altering their access to sunlight and nutrients; light intensity is highest at the surface and macronutrients are generally higher near the bottom sediments (Falconer, 2005). Under calm conditions, there can be intense proliferations, creating a visible discoloration and accumulation of cells known as a cyanobacteria bloom (Chorus and Bartram, 1999; Falconer, 2005). Cyanobacteria blooms may increase their density by a factor of 1000 or more in a very short period under calm conditions (Chorus et al., 2000). Offshore winds may then drive these scums towards the shore where they can accumulate (Chorus and Bartram, 1999; Falconer, 2005). These blooms can be very dense and can have the appearance of being gelatinous or resemble a collection of fine grass clippings, and may appear as a homogeneous, soupy mass, as if green paint has been spilled into the water (WHO, 2003; Falconer, 2005). As mentioned earlier, scums can also appear in other colours, including red-blue and tan.
Cyanobacteria blooms are a public health concern as they can contain intracellular cyanotoxins, cell-surface endotoxins, and extracellular cyanotoxins (see Section 3.2). In Canada, the most troublesome planktonic toxic cyanobacteria genera are also those most frequently encountered worldwide: Dolichospermum (formerly called Anabaena), Aphanizomenon, Gloeotrichia, Microcystis, Planktothrix, Pseudoanabaena and Woronichinia (Winter et al., 2011; MDDEP, 2012; Ontario Ministry of the Environment, 2012). Toxins are usually associated with the cyanobacterial cells - either bound within membranes or occurring freely within the cells. Cyanotoxin release to the surrounding waters can occur as the cells die or are damaged and leak their contents (Chorus and Bartram, 1999). Cyanobacteria blooms can consist of a mix of species and strains, each of which may or may not produce toxins. The bulk of the toxicity, if present, generally lasts as long as the bloom. While some toxins may persist for a period after the bloom has dissipated (Chorus and Bartram, 1999; Falconer, 2005), concentrations may quickly fall to below a level that represents a health risk through recreational water exposure. The time necessary for this to occur needs to be determined on a site-specific basis. Health effects have also been associated with contact with cyanobacteria cells. A prospective study of three Canadian lakes with a history of cyanobacterial presence reported gastrointestinal symptoms associated with cyanobacteria cell densities, not with cyanotoxin concentrations (Lévesque et al., 2014).
Warning signs of cyanobacterial toxins may be observed, such as the presence of dead waterfowl or other wildlife along the shoreline or reports of domestic animal poisonings, specifically cattle and dogs (Chorus and Bartram, 1999). Still, toxic blooms can occur without any noticeable effect on the local animal populations. As a result, any bloom encountered should be treated as potentially toxic.
Benthic cyanobacteria, or those that grow on the bottom surfaces, can also be found in freshwater habitats in Canada. Genera of these cyanobacteria are capable of producing toxins such as: Oscillatoria, Phormidium and Lyngbya (Vis et al., 2008; Lajeunesse et al., 2012; Quiblier et al., 2013). The information database for toxic benthic cyanobacterial populations, however, is comparatively sparse (Quiblier et al., 2013; Gaget et al., 2017a; Burford et al., 2019). Benthic mats are generally considered a lower risk to human health due to limited contact with the attached material. However, domestic pets and livestock may still have access to mats, particularly as they dry on shorelines after water recession, and be at risk (WHO 2003). If the benthic mats are in recreational areas, or if benthic cyanobacteria become detached from bottom surfaces and subsequently rise to the water surface and accumulate along shorelines, human exposure to benthic cyanobacteria may be of concern (Quiblier et al., 2013; Gaget et al., 2017a). Benthic mats are also being increasingly reported (Wood and Puddick, 2017; Burford et al., 2019), and as more becomes known about the toxicity of benthic cyanobacteria, more information should become available on their significance to human health.
There are known toxic marine algal species that are capable of forming blooms and producing toxins (e.g., Alexandrium spp. and the phenomenon known as “red tide”) (Chorus and Bartram, 1999). However, as the focus of this section is the human health risks from exposure to toxic cyanobacteria through recreational water activities, they will not be discussed here.
3.2 Cyanobacterial toxins
As mentioned earlier, cyanobacterial blooms are a public health concern as they can produce cell surface endotoxins as well as intracellular cyanotoxins. The cell-surface endotoxins are not well understood. They may elicit an irritant or allergic response in humans following dermal contact (irritant toxins), as well as potential illnesses from ingestion and inhalation (Lévesque et. al., 2014, 2016; Ohkouchi et al., 2015; Otten and Paerl, 2015). It is also possible that these health effects are related to other unknown substances in cyanobacteria or to other bacteria that are associated with cyanobacteria blooms. More research in this area is needed.
Intracellular cyanotoxins are produced by a variety of cyanobacteria (although not all cyanobacteria) and are associated with various harmful effects on humans (Chorus and Bartram, 1999; Otten and Paerl, 2015; Carmichael and Boyer, 2016). These toxins are usually contained within intact cyanobacteria cells and released when cells are lysed, although some intracellular toxins can be released naturally without cell lysis (e.g., cylindrospermopsin).There are several known intracellular cyanotoxins, including microcystins, nodularins, anatoxins, cylindrospermopsins, saxitoxins, and dermatotoxins. Microcystins and nodularins are cyclic peptides that affect the liver (hepatotoxins). Anatoxins are alkaloids that target the nervous system (neurotoxins). Saxitoxins also target nerve and muscle cells (neurotoxins). Cylindrospermopsins are an alkaloid that affects the liver, although it has demonstrated an ability to also affect a wide range of organs (cytotoxic properties), especially the kidneys (Chorus and Bartram, 1999; Falconer, 2005). Although these toxins can result in serious illness, the predominant health effects encountered from accidental ingestion of cyanobacteria may be gastrointestinal or flu-like in nature and may often go unreported or are attributed to other causes (Falconer, 2005; Lévesque et al., 2014; Otten and Paerl, 2015).
3.2.1 Microcystins
Microcystins (MC) are hepatotoxins that are generally regarded as the most prevalent and significant of the freshwater cyanotoxins owing to their stability and resistance to biological and chemical breakdown, their widespread occurrence and their potential to reach high concentrations in blooms and scums (Boyer, 2007; Williams et al., 2007; Winter et al., 2011). Microcystins are largely cell-bound (i.e., contained with intact cells) until cell death and lysis. More than 200 microcystin variants have been identified (Spoof and Arnaud, 2017; Bouaïcha et al., 2019). MC-LR is the most commonly measured and one of the most toxic variants worldwide (Graham et al., 2010), although accounts of other variants dominating blooms and/or co-occurring with MC-LR have been documented (Kemp and John, 2006; Graham et al., 2010; Li et al., 2010; Sabart et al., 2010; B.C. Ministry of Health, 2012; MDDEP, 2012; Srivastava et al., 2012). A number of cyanobacterial genera have been identified as microcystin producers (Kotak and Zurawell, 2007; Funari and Testai, 2008; Pearson et al., 2010; Martins and Vasconcelos, 2011; Carmichael and Boyer, 2016; Bernard et al., 2017). Those most commonly observed in North America are Dolichospermum (Anabaena), Microcystis, Planktothrix and Pseudoanabaena (Williams et al., 2007; Winter et al., 2011). Planktothrix species appear to produce only demethylated microcystin variants (Fastner et al., 1999; Briand et al., 2005; Kurmayer et al., 2004; Cerasino et al, 2016), which may not be detected depending on the analytical method used.
The health effects associated with recreational exposure to waters contaminated by blooms of Microcystis and Dolichospermum (Anabaena) have included headaches, nausea, vomiting, diarrhea, abdominal pain, muscle aches, fever, mouth ulcers, blistering of the lips, sore throat, skin rashes and ear and eye irritations (Chorus and Bartram, 1999; Otten and Paerl, 2015; Gaget et al., 2017a). In Argentina, accidental contact with a cyanobacterial bloom containing microcystins resulted in symptoms of fever, nausea, and abdominal pain followed by atypical pneumonia and impacts on the liver (Giannuzzi et al., 2011). There has also been a single case of acute hepatic failure linked to recreational exposure to microcystins (Vidal et al., 2017). Outbreaks in recreational water have also occasionally been associated with exposure to cyanobacterial blooms containing microcystins. In the United States, two outbreaks were reported in 2003-2004 (Dziuban et al., 2006) and eight outbreaks were reported in 2009-2010 (Hilborn et al., 2014). The symptoms reported during these outbreaks included abdominal cramps, diarrhea, nausea, vomiting, fever, headache, rashes, eye irritation, earache, neurologic symptoms, tingling, confusion, and respiratory symptoms (Hilborn et al., 2014). In addition to acute effects, there is also some evidence that microcystin can act as a tumour promoter, and IARC (2010) has classified this cyanotoxin as possibly carcinogenic to humans. For more information on microcystins, see the Guidelines for Canadian Drinking Water Quality – Guideline Technical Document for cyanobacterial toxins (Health Canada, 2017).
3.2.2 Anatoxins
The anatoxins (anatoxin-a, anatoxin-a(S), homoanatoxin-a) can be produced by species of Dolichospermum (Anabaena) (anatoxin-a, anatoxin a(S)), Aphanizomenon (anatoxin-a), Microcystis (anatoxin-a) and Oscillatoria (anatoxin-a, homoanatoxin-a) (Chorus and Bartram, 1999; Funari and Testai, 2008). Cuspidothrix issatschenkoi (formerly Aphanizomenon issatschenkoi) is also a recognized anatoxin-a producer in Europe and Japan (Hodoki et al., 2012). In waterbodies sufficiently clear for the growth of macrophytes and associated cyanobacteria, Tychonema sp. may also be a producer of anatoxin-a (Fastner at al. 2016). Similar to microcystins, anatoxins are intracellular toxins. When they are released from cells (dissolved in water), they are unstable (Stevens and Krieger, 1991; Chorus and Bartram, 1999, Bownik, 2010).
Anatoxins are neurotoxins that interfere with the activity of the nerve transmitter acetylcholine. This, in turn, affects the functioning of the nervous system by disrupting communication between nerves and muscle cells. Health effects associated with anatoxin exposure include paralysis of both the skeletal and respiratory muscles, resulting in tremors, convulsions and, ultimately, death due to respiratory failure (Rogers et al., 2005). Non-lethal human poisonings, with symptoms of acute gastrointestinal disorders such as nausea, vomiting and diarrhea, were reported following the ingestion of water with unspecified species of Microcystis and Dolichospermum (Anabaena) (producers of anatoxin-a); allergic reactions (such as skin papulo-vesicular eruptions) have also been related to swimming in water containing a bloom of Dolichospermum (Anabaena) (Schwimmer and Schwimmer, 1968). Detection of the actual anatoxin-a toxin, however, was not reported. Only one human fatality has been potentially associated with exposure to cyanobacterial neurotoxins in natural waters; exposure occurred through ingestion of contaminated water during accidental immersion at a location where swimming was not permitted (Falconer, 2005). Anatoxin-a has been associated with poisonings and deaths of various animals following exposure to cyanotoxin-contaminated water (Carmichael and Gorham, 1978; Edwards et al., 1992; Gunn et al., 1992; Puschner et al., 2008; Stewart et al., 2008; Backer et al., 2013). However, exposure levels were not reported. Clinical symptoms were mostly neurologic with deaths attributed to rapid onset of respiratory paralysis (a characteristic adverse effect of anatoxin-a). The high number of reported deaths of animals (in contrast to nearly no known human fatalities) due to anatoxin-a is attributable to the much larger volume animals will ingest as compared to humans exposed during recreation. For more information on anatoxins, see the Guidelines for Canadian Drinking Water Quality – Guideline Technical Document for Cyanobacterial toxins (Health Canada, 2017).
3.2.3 Cylindrospermopsins
Cylindrospermopsins are primarily categorized as a hepatotoxin, although they have also been shown to exert cytotoxicity in other organs such as the kidney, spleen, thymus, heart and gastrointestinal tract (Chorus and Bartram, 1999; Chong et al., 2002; Falconer, 2005; Funari and Testai, 2008). Unlike microcystins, a substantial amount of cylindrospermopsin is released into the water column during bloom growth as opposed to being cell-bound. It is also quite stable in the environment compared to other toxins (Wörmer et al., 2008, 2009). Cylindrospermopsin is more commonly encountered in tropical and subtropical regions of the globe (Williams et al., 2007), however, there have been increasing reports of potential toxin-producing species in temperate fresh waters, suggesting that the geographical range of cylindrospermopsin producing species may be expanding (Graham et al., 2010; Xie et al., 2011; Sinha et al., 2012). The first recorded incident of human cylindrospermopsin poisoning occurred in 1979, off the coast of Queensland, Australia, and was attributed to a bloom of Raphisiopsis (previously known as Cylindrospermopsis) raciborskii. Symptoms associated with the outbreak included vomiting, malaise, headache and constipation, later followed by bloody diarrhea and evidence of liver and kidney damage (Chorus and Bartram, 1999). At present, there have been no human fatalities associated with cylindrospermopsins, and there have been no other recorded poisonings from either drinking or recreational water.
Numerous cyanobacterial species are capable of producing cylindrospermopsins such as Raphisiopsis (formerly Cylindrospermopsis) raciborskii, Chrysosporum ovalisporum (formerly Aphanizomenon ovalisporum), Aphanizomenon gracile, Umezakia natans, Anabaena bergii, Anabaena lapponica, Dolichospermum (Anabaena) planctonica, Lyngbya wollei, Rhaphidiopsis curvata, and Rhaphidiopsis mediterranea (U.S. EPA, 2015). Aphanizomenon flos-aquae may also produce the toxin (U.S. EPA, 2015), however, further research is needed to explore the conditions under which this may occur (Lyon-Colbert et al., 2018). For more information on cylindrospermopsin, see the Guidelines for Canadian Drinking Water Quality – Guideline Technical Document for Cyanobacterial toxins (Health Canada, 2017).
3.2.4 Nodularins
Nodularins are hepatotoxins usually caused by strains of the brackish-water cyanobacterial genus Nodularia although additional species, such as cyanobacteria from the genus Nostoc, have been reported to be capable of producing the toxin (Gehringer et al., 2012; Wood et al., 2012). Multiple variants of nodularins have been identified, with nodularin-R being the most abundant form of the cyanotoxin (Mazur-Marzec et al., 2006). The toxins are closely related to microcystins in both structure and function (Chorus and Bartram, 1999). Data derived from experimental studies, although limited, have suggested that nodularins exhibit toxicity similar to that of microcystin-LR. Results obtained from chronic toxicity studies using animal models have suggested that nodularins may be a more potent tumour promoter than microcystins (Chorus and Bartram, 1999).
3.2.5 Saxitoxins
Saxitoxin and its nearly 60 related analogs are a group of toxins that include saxitoxin, neosaxitoxin, the gonyautoxins and C-toxins (Codd et al., 1999; Wiese et al., 2010). These toxins act by blocking sodium channels in nerves and muscle cells, preventing the transmission of electrical impulses. They are also called paralytic shellfish poisoning (PSP) toxins because marine shellfish accumulate the toxins by feeding on blooms of the marine plankton Alexandrium (Codd et al., 1999). There have been fatal cases of PSP reported in North and Central America from consuming shellfish (Chorus and Bartram, 1999), although to date there have been no saxitoxin-related illnesses reported for humans through drinking or recreational water exposure. Animal deaths have also been linked to contact with cyanobacteria blooms containing saxitoxins (Negri et al., 1995). Some members of the cyanobacteria genera Dolichospermum (Anabaena), Aphanizomenon, Raphisiopsis (Cylindrospermopsis) and the benthic cyanobacterium Lyngbya have been reported to produce saxitoxins (Aráoz et al., 2010; Carmichael and Boyer, 2016). For more information on saxitoxins, please refer to Guidelines for Canadian Drinking Water Quality – Guideline Technical Document for Cyanobacterial toxins (Health Canada, 2017).
3.2.6 Dermatotoxins and other irritant toxins
Certain marine cyanobacteria, such as species of Lyngbya, Oscillatoria and Schizothrix, can produce toxins called aplysiatoxins and lyngbyatoxins, which have been reported to cause severe dermatitis.In addition, aplysiatoxins are considered potent tumour promoters and are thought to demonstrate other properties that may be linked to carcinogenesis (Chorus and Bartram, 1999). Some Lyngbya species alsoproduce debromoaplysiatoxin and apratoxin A, the latter of which is highly cytotoxic and can induce apoptosis in cells (Luesch et al,2001). First aid records from Fraser Island, Australia, the world's largest sand island, revealed that during a seven-week period where Lyngbya majuscula was identified in the waters there was an increase in the number of marine recreational water users reporting symptoms consistent with exposure to L. majuscula (Osborne and Shaw, 2008). Symptoms typically included painful rashes, inflammation, itching, and irritation of the nose, eyes, and throat. The majority of people who reported symptoms had come into direct contact with Lyngbya via swimming, although two individuals reported symptoms after inhaling sea spray while driving along the beach.
Freshwater mat-forming Lyngbya species, although not as thoroughly studied as marine species, have been reported to cause skin irritation and dermatitis in scuba divers (Florida) as well as in individuals cleaning up beach wrack from a Lyngbya-infested bay (Lake Ontario) (Carmichael and Boyer, 2016).Therefore, although dermatotoxins are primarily produced by cyanobacterial marine species, they may still be of concern for freshwater lakes and rivers.
Although not well understood, the cell-surface endotoxins (i.e., the lipopolysaccharide component of the cyanobacterial cell wall) may elicit an irritant or allergenic response in humans following dermal contact (Chorus and Bartram, 1999), as well as result in symptoms of gastrointestinal illness (Lévesque et al., 2014). Lipopolysaccharides have been known to exhibit inflammatory, pyrogenic (fever-inducing), and toxic properties. However, it is generally regarded that lipopolysaccharides from cyanobacteria are considerably less toxic than those of other Gram-negative bacteria, such as Salmonella (Chorus and Bartram, 1999). Therefore, it is also possible that these health effects are related to other unknown substances in cyanobacteria or to other bacteria that are associated with cyanobacteria blooms.
3.2.7 Compound of interest: Β-methylamino-L-alanine
A topic of interest involves the unusual amino acid β-methylamino-L-alanine (BMAA), its links to cyanobacteria, and the research findings concerning its potential neurotoxic capabilities. BMAA can be found in virtually all groups of cyanobacteria, including the notable freshwater genera Dolichospermum (Anabaena), Aphanizomenon, Microcystis, Nodularia, and Oscillatoria, as well as the marine cyanobacteria genus Nostoc (Cox et al., 2005; Banack et al, 2007; Metcalf et al., 2008). BMAA can co-occur with other cyanotoxins (Metcalf et al., 2008).
BMAA has been reported from in the brain tissues of patients with amyotrophic lateral sclerosis/parkinsonism–dementia complex (ALS/PDC), ALS, and Alzheimer's disease (Cox et al., 2003; Murch et al., 2004; Pablo et al., 2009), although a more recent large study did not find BMAA present in Alzheimer's patients (Meneenly et al., 2016). Studies have also investigated whether cyanobacterial blooms, as a source of BMAA, could lead to biomagnification through the food chain and result in an increased risk of illness in the exposed population (Cox et al., 2003; Banack et al., 2015). More work is needed before a cause-and-effect relationship between BMAA and neurological disease can be established or discounted (Holtcamp, 2012; ANSES, 2017). Similarly, there is insufficient evidence at this time to suggest that water or dietary sources could constitute a significant source of BMAA exposure. Developments on this topic will continue to be monitored.
4.0 Routes of Exposure
The three main routes of human exposure to cyanobacteria and their toxins in recreational waters are ingestion, inhalation and direct body (dermal) contact (Chorus and Bartram, 1999; NHMRC, 2008). Cyanobacteria and their toxins can occur in both the water column and in benthic mats.
Ingestion is the most frequently documented route of exposure for cyanobacteria and their toxins. Cases of illness have been reported following the accidental swallowing of bloom-impaired waters (Chorus and Bartram, 1999; WHO, 2003; Stewart et al., 2006). Activities involving sudden or repeated immersion of the head (e.g., windsurfing or kayaking) may also lead to ingestion or inhalation exposure via water forced into the mouth and/or nasal passages. Although not related directly to recreational water exposure, ingestion of foods, such as fish and shellfish, and food supplements such as algal supplements, can also be a potential source of cyanotoxins. Further information on ingestion exposure through food can be found in the Guidelines for Canadian Drinking Water – Guideline Technical Document on Cyanobacterial toxins (Health Canada, 2017).
Cyanobacteria and their toxins can also be present as aerosols generated by wind, waterskiing or swimming, thereby potentially providing an inhalation route of exposure. In a study of recreational exposure to aerosolised microcystins from two California lakes, Backer et al. (2010) reported detectable levels of microcystins in personal air samples and nasal swabs from 81 children (12 years and older) and adults following recreational activities (waterskiing, operating personal watercraft, swimming or wading). Microcystins, however, were not detected in blood samples indicating that aerosolized toxins did not enter the lungs deep enough to be absorbed into the blood stream. Although this evidence indicates a potential for inhalation exposure, more research is required to determine whether aerosolised toxins can reach the lower respiratory tract for it to be absorbed by the lungs and enter the blood.
Direct contact with cyanobacteria cells has been known to cause irritation of varying severity, although the exact mechanisms for this are not fully understood. Allergic reactions are more commonly reported in sensitive individuals. It has been suggested that the irritations are due to unknown cyanobacterial components, separate from the toxins (Chorus and Bartram, 1999). Bathing suits and wet suits may also exacerbate the potential for skin irritations by trapping the cells and then disrupting their contents as a result of the friction created between the suit material and the user's skin (Chorus and Bartram, 1999).
In general, the likelihood of exposure to cyanobacterial toxins in sufficient amounts to constitute a chronic or acute health risk is considered relatively low. This is because of circumstances such as the seasonality and localized nature of blooms, their unappealing aesthetic properties and the way drinking water supplies and monitored recreational water areas are managed. Under circumstances where a recreational area is experiencing prolonged and persistent blooms and where intensive recreational activities are continuing (as may be the case in recreational areas that are not monitored or managed), the risks of acute exposure may be greater (Funari and Testai, 2008).
5.0 Occurrence in the environment
Cyanobacteria are a normal component of aquatic phytoplankton and of the benthic community, with many species occurring in fresh waters. Blooms of these microorganisms in surface waters are not a new phenomenon. There are reports of cyanobacteria blooms, linked to animal poisonings, dating back to the early 1900s in Canada. However, the nutrient enrichment (eutrophication) of surface waters by nitrogen and phosphorus has substantially increased the amount of cyanobacteria that can occur and thus, has had a significant impact on the frequency and severity of cyanobacteria blooms (Chorus and Bartram, 1999; Falconer, 2005; Newcombe, 2009; Chorus & Niesel, 2011). The amount of biomass that can occur in a given waterbody depends on the concentration of nutrients needed to sustain it, and upper limits can be estimated from the concentrations of total phosphorus and total nitrogen. Studies in Europe have identified total phosphorus thresholds in the range of 25 to 100 µg/L; below this level, cyanobacteria become irrelevant for recreational water exposure (Carvalho et al., 2013; Phillips et al., 2008; Chorus & Niesel, 2011). This range is somewhat influenced by the depth of the mixed water layer (epilimnion; Fastner et al., 2016). Other factors that can favour cyanobacteria growth include a low water exchange rate, persistent thermal stratification, and for some species, high turbidity. Blooms are not likely to occur in acidified water with a pH below 6-7 (Chorus and Nielsel, 2011). Cyanobacteria can also grow under a range of temperatures (Chorus and Bartram, 1999; Falconer, 2005), although they have relatively low growth rates compared to many eukaryotic algae, which may help explain why blooms typically occur in the late summer months after having had sufficient time to build a large population. However, this historical trend towards late summer blooms is no longer as clear-cut (Jacoby and Kann, 2007). Canadian data demonstrates that blooms are appearing earlier in the spring and extending later into the year (Health Canada, 2017).
The factors responsible for the dominance of toxin-producing strains in a cyanobacteria bloom are not well understood (Chorus and Bartram, 1999; Falconer, 2005). Variations in toxicity within a bloom are primarily due to the rise and fall of subpopulations of strains having different toxicities. Environmental factors (e.g., conditions of waterbody mixing, sunlight and temperature) also contribute to the amount of toxin produced (by strains with toxin production genes), but to a lesser extent (Chorus and Bartram, 1999). Consequently, toxin formation from cyanobacteria is less predictable than the cyanobacterial blooms themselves. Lakes that have had cyanobacteria blooms with low toxin contents per cell can develop blooms that may contain high amounts of toxins. Conversely, even if lakes have had toxic blooms in the past, it does not necessarily mean they will experience blooms in the future. It has been suggested that worldwide, an average of 60% of the cyanobacterial bloom samples investigated have been positive for cyanobacterial toxins (range 10–90%) (Chorus et al., 2000; WHO, 2003). As a result of the interplay of the factors affecting bloom development, there may be large year-to-year fluctuations in the levels of cyanobacteria and their toxins (Health Canada, 2017).
The dissipation of cyanotoxins after the collapse of a cyanobacteria bloom will depend on numerous factors, such as the amount of dilution, the type of toxin, and the rate of biodegradation. Biodegradation rates usually encompass a lag phase, where no degradation is occurring. This lag phase may reflect the time required for the microbial population (responsible for degrading the cyanotoxins) to reach a sufficient density or to use up other nutrient sources (Smith et al., 2008). This lag phase can vary from none to as much as 3 weeks (Jones and Orr, 1994; Grutzmacher et a., 2010; Klitzke and Fastner, 2012); however, the lag phase has been shown to be reduced in waters with repeated exposure to cyanotoxins (Christoffersen et al., 2002; Smith et al., 2008). After the lag phase (if present), biodegradation can then occur quite rapidly. For example, for dissolved microcystin-LR, 90-95% degradation occurred within 3-4 days (Jones and Orr, 1994) although longer time periods (1 to 2 weeks) have also been reported (Lam et al., 1995). Due to the number of variables that affect cyanotoxin dissipation, the time necessary for the complete disappearance of toxin will vary between water sources.
Less information is known about the occurrence of benthic cyanobacteria (e.g., Lyngbya spp.). These cyanobacteria can grow to form dense, bottom-covering mats of cyanobacterial material (Chorus and Bartram, 1999; WHO 2003; New Zealand Ministry for the Environment and Health, 2009). These mats typically occur in clear, shallow waters where sunlight can penetrate to the bottom, although they may also be present under other environmental conditions. The mats can occasionally be dislodged and washed ashore, where they may be scavenged by animals. In 2017, isolates purified from benthic mats at three different drinking water reservoirs in Australia tested positive for the production of cyanotoxins (Gaget et al., 2017a). Additionally, benthic mats have been increasing in abundance in fluvial lakes along the St. Lawrence River in Québec and in lakes at Whiteshell Provincial Park in Manitoba (Macbeth, 2004; Lajeunesse et al., 2012; Hudon et al., 2016). As waterbodies become clearer due to the reduction of eutrophication, benthic cyanobacteria as well as those growing on macrophytes (e.g., Tychonema spp.) may become more frequent (Fastner at al. 2016).
5.1 Microcystin
Microcystin can be produced by both planktonic and benthic cyanobacteria, and both toxic and non-toxic species exist for all of the predominant microcystin-producing genera (Chorus and Bartram, 1999; Carillo et al., 2003; Quiblier et al., 2013; Ngwa et al., 2014). Detection of the genes responsible for microcystin production (mcy genes) can be used as a tool to discriminate between toxic and non-toxic strains of Microcystis, Dolichospermum (Anabaena) and Planktothrix that are otherwise indistinguishable (Davis et al, 2009; Ngwa et al., 2014). Numerous studies have investigated the occurrence of cyanobacteria and their toxins in Canada. A selection of these studies is presented in Table 1.
| Location | Summary findings | Reference |
|---|---|---|
Freshwaters across Canada (2001-2011) |
|
Orihel et al. (2012) |
Freshwaters in QC |
|
Bourbonnais and Robert (2014) |
Missisquoi Bay, QC |
|
Fortin et al. (2010); Blais (2014, 2015, 2019); Bowling et al. (2014) |
Freshwater lakes, QC |
|
Giani et al. (2005) |
The lower Great Lakes, ON |
|
Carmichael and Boyer (2016) |
Lake Erie, ON |
|
Millie et al. (2009) |
Lake of the Woods, ON/MB |
|
Chen et al. (2007) |
Lake Erie, ON |
|
Rinta-Kanto et al. (2005) |
Recreational lakes, MB |
|
Jones et al. (1998) |
Freshwater lakes, AB |
|
Kotak et al. (1996) |
| TEQ – Toxic equivalent includes the total concentration of both microcystin-LR and non-microcystin-LR variants. Non-microcystin-LR variants are included in the total only if a toxicity equivalent factor is available and applied. | ||
5.2 Anatoxin
Blooms of anatoxin-producing species are not routinely reported in Canadian waters, although they have been detected (Kotak and Zurawell, 2007; Bourbonnais and Robert, 2014). Although they occur less frequently than microcystin-producing cyanobacteria, they have been reported as the cause of animal poisonings (Hoff et al., 2007; Backer et al., 2013). In Canada, a low detection frequency and limitations of the analytical methods mean that there are few data available on the levels of anatoxins in natural waters affected by cyanobacterial blooms. In addition, anatoxins dissolved in water are relatively unstable and, as such, are not considered to be as widespread as microcystins in water supplies (Chorus and Bartram, 1999). As a result, anatoxins are currently considered to be of lesser concern than microcystins for Canadian recreational waters. The ecology of benthic cyanobacteria is increasingly a focus of research. Studies have documented the detection of anatoxins (along with saxitoxins and most of the other known cytotoxins) in benthic cyanobacterial populations (Vis et al., 2008; Lajeunesse et al., 2012; Quiblier et al., 2013; Hudon et al., 2016). Although the risks to human health from anatoxins in benthic material are likely very low, more research is required to determine how such benthic species affects recreational waters and what level of risk they pose for human health.
5.3 Cylindrospermopsin
Cylindrospermopsin is more commonly encountered in tropical and subtropical regions of the globe (Williams et al., 2007). Australia and the state of Florida in particular have reported multiple instances of cylindrospermopsin detection in lakes, rivers and drinking water reservoirs (Falconer and Humpage, 2006; de la Cruz et al., 2013). Raphisiopsis (formerly Cylindrospermosis) raciborskii has long been recognized as the most widely distributed species capable of producing cylindrospermopsin. There have been increasing reports of potential toxin-producing species in temperate fresh waters including the northern United States and Canada: in Ohio (Conroy et al., 2007), Michigan (Hong et al., 2006), Minnesota (Sinha et al., 2012), Manitoba (Kling, 2009) and Ontario (Hamilton et al., 2005). This may suggest that the geographical range of these species is expanding (Graham et al., 2010; Xie et al., 2011; Sinha et al., 2012), however, the Raphisiopsis (Cylindrospermopsis) raciborskii strains isolated in North America are usually non-toxic (Burford and Davis, 2011; Yilmaz and Phlips, 2011). Cylindrospermopsin has been detected, albeit rarely, in Canadian surface water sources. In an investigation in the province of Québec, cylindrospermopsin was detected in two of twelve bloom samples at concentrations of 0.1 μg/L and 0.2 μg/L (Roy-Lachapelle et al., 2015). However, it was not detected in a subsequent investigation of bloom samples (Fayad et al., 2015). Cylindrospermopsin was also detected in a Québec drinking water treatment plant study in scum that had accumulated in the filtration system. The dominant cyanobacteria species in the source water and in the sludge bed were M. aeruginosa and Dolichospermum (Anabaena). (Zamyadi et al., 2012a). Most surface waters that are monitored for cylindrospermopsin in Canada have been negative for the toxin (Carmichael and Boyer, 2016). Cylindrospermopsin has been detected occasionally in surface waters in the United States (Boyer, 2007; U.S. EPA, 2015), usually isolated from blooms that were dominated by Chrysosporum (Aphanizomenon)or Dolichospermum (Anabaena)and Microcystis (Yilmaz and Phlips, 2011; U.S. EPA, 2015).
5.4 Other cyanobacterial toxins
Nodularia blooms have been encountered in brackish lakes in Australia and New Zealand, as well as in the Baltic Sea. In general, this species prefers brackish and saline waters, although blooms have been identified in freshwater lakes in Turkey (Akcaalan et al., 2009). Nodularia was found in two American lakes (Beutel et al., 2001). To date, there have been no recorded occurrences of Nodularia or Nostoc blooms in Canadian waters. As a result, nodularins are not considered a significant public health threat in Canadian recreational waters.
Lyngbya has a wide geographic distribution, as the genus comprises both fresh and marine water species of cyanobacteria. Benthic blooms of Lyngbya tend to occur in warmer climates, including Florida and Hawaii, although certain species are common in North American lakes and large mats are emerging in Lake Erie and the St. Lawrence River (Osborne et al., 2001; Bridgeman and Penamon, 2010; Hudon et al., 2014). The dermatotoxins produced by Lyngba are primarily produced by cyanobacterial marine species; however, they may still be of concern for freshwater lakes and rivers.
Saxitoxin-containing blooms are widespread in marine waters in Australia (Osborne et al., 2001), and toxic blooms have also been detected in freshwaters in many parts of the world, including (but not limited to) Brazil, Europe, and the United States (Teneva et al., 2003; dos Anjos et al., 2006; Vijayavel et al., 2013). To date, saxitoxins are not considered a significant concern in Canadian recreational waters. However, the detection of saxitoxin analogues in Lyngbya benthic blooms along the St. Laurence River and two of its fluvial lakes suggests that this issue should continue to be monitored (Lajeunesse et al., 2012; Hudon et al., 2016). When Lyngbya benthic blooms are found in recreational areas, submitting bloom material for toxin testing will provide a better idea of potential health risks.
5.5 Indicators of cyanobacteria and their toxins
Various indicators can be used to help assess health risks from cyanotoxins in recreational areas, as well as to collect information on planktonic cyanobacteria occurrence to aid in risk assessment and risk management. The array of methods included in this guideline offer a flexible approach to detecting and understanding potential bloom toxicity. They can be used alone or in combination, depending on the jurisdiction and the recreational water-quality management plan in place. The choice of indicators used will also depend on local access to methods. There are advantages and limitations to using indicators for assessing potential health risks (described below). Of particular note is that the conservative nature of indicators means they are likely to overestimate risk.
5.5.1 Total cyanobacteria cell counts
Total cyanobacteria cell counts are a measure of planktonic cyanobacteria biomass that can be used to indicate that a bloom is developing. As all blooms should be treated as potentially toxic unless otherwise known, cyanobacteria cell counts can be used as an indicator of potential health effects. As discussed above, there is also evidence of direct health impacts from contact with cyanobacteria cells, in the form of allergic or irritative effects (Backer et al., 2015) and potential gastrointestinal symptoms (Lévesque et al., 2014, 2016). The concentration of cyanobacteria cells at which these health impacts are seen is highly variable, dependent on factors such as the individuals exposed (i.e., those with an allergic predisposition) and the composition of the cyanobacteria present. Accounting for this variability would result in the development of guideline values that are unnecessarily conservative for most situations, therefore the indicator values given here are based on the potential presence of microcystins. Further information on the derivation of the total cyanobacteria cell count value can be found in Section 7.2.
A significant drawback of the cyanobacteria cell count measurement is the diversity in the range of shapes and sizes of cyanobacteria cells (Wood et al., 2008). Depending on the types of cyanobacteria present, cyanobacteria cell concentrations could exceed of the guideline value with no visual evidence of a planktonic bloom. For example, water bodies that contain high concentrations of picocyanobacteria, which are small cyanobacteria of less than 2 μm in diameter, could exceed the guideline value with no evidence of a bloom and no increased risk to human health. Therefore, when using total cyanobacteria cell counts, it is important to also consider the types of cyanobacteria that are being identified and where possible, their potential for toxin production. Determining the types of cyanobacteria present is also the first step to calculating cyanobacteria biovolume, which has been shown to have a closer relationship to cyanotoxin concentrations (see section 5.5.2). It is also important to note that when total cell counts are decreasing during the dissipation of a bloom, there may still be high levels of cyanotoxins present as the intracellular toxins are released from the dying cells into the surrounding waters. This is important for toxins that are usually contained within intact cells, such as microcystins, but is less of a concern for other toxins, such as cylindrospermopsin, that are released naturally from cells irrespective of cell lysis.
5.5.2 Cyanobacteria biovolume
Cyanobacteria biovolume is a measure of the planktonic cyanobacteria biomass in a water sample. It can be used to indicate a bloom is forming and may pose a risk to human health. Biovolume is a more accurate assessment of the cyanobacteria biomass than total cyanobacteria cell counts since this measurement accounts for the surface area of the cell, as well as the mass of all cellular material, or cellular biomass (Saccà, 2016). Using a biovolume measurement, as opposed to total cyanobacteria cell counts, means that small cyanobacteria cells such as the aforementioned picocyanobacteria, do not have a large impact on the calculated concentration. Similar to total cell counts, the types of cyanobacteria that are identified, as well as their potential for toxin production, should be considered to help avoid issuing unnecessary swimming/contact advisories. Cyanotoxin concentrations have been found to relate more directly to cellular biomass than to cell numbers (Ibelings et al., 2014; Dong et al., 2016). Similar to total cyanobacteria cell counts, depending on the cyanotoxins present, the cyanotoxin concentrations may be high during and immediately following the dissipation of a bloom but the biovolume measurements will be low. Further information on the derivation of the biovolume guideline value can be found in Section 7.3.
5.5.3 Chlorophyll a
Chlorophyll a is a photosynthetic green pigment present in cyanobacteria and other phytoplankton (Fiedor et al., 2008; Søndergaard et al., 2011). It is frequently used as an index for eutrophication and can be used as part of a cyanobacteria alert system to trigger further investigation and actions (Bartram et al., 1999). Chlorophyll a is particularly useful if it can be combined with brief qualitative microscopy to assess whether or not the majority of the phytoplankton is cyanobacteria. This would require identification of cyanobacteria at the genus level, which can be readily learned by microbiologists with some training in microscopy. Chlorophyll a measurements have an advantage over other biomass indicators in that the method for detection is simpler and in-situ methods are available (see Section 6.0). As other phytoplankton also contain chlorophyll a, the relationship between chlorophyll a and cyanobacteria biomass is stronger when cyanobacteria are the main or dominant organisms present. Although positive correlations have been found between chlorophyll a concentrations and cyanobacterial biomass or cyanotoxin concentrations in various studies (Huot, et al., 2007; Izydorczyk et al., 2009; Du et al., 2014; Yuan et al., 2014), the main advantage of monitoring for chlorophyll a as part of a cyanobacteria alert system is the greater temporal and spatial coverage that is possible with less expense and effort.
Phycocyanin, a photosynthetic accessory pigment to chlorophyll a, has also been investigated as a possible parameter for cyanobacteria monitoring. Concentrations of these two pigments are highly correlated,and similar to chlorophyll a, positive correlations have been observed between phycocyanin content and cyanobacterial biomass (Brient et al., 2008; McQuaid et al., 2011; Kasinak, 2015; Pace et al., 2017). An important advantage for phycocyanin is that this pigment is more specific to cyanobacteria, and various studies have shown that it is an accurate predictor of cyanobacterial abundance (Gregor et al., 2007; Brient et al., 2008; Ibelings et al., 2014; Macario et al., 2015). The presence of known microcystin producers has been shown to correlate strongly with phycocyanin concentrations (Oh et al., 2001); however, it does not directly relate to cellular microcystin content as all cyanobacteria possess this pigment. Although no guideline value has been provided for phycocyanin, this pigment could also be used as part of an alert system for cyanobacteria bloom development. Its use would require the development of site-specific ratios between phycocyanin and the measure of interest (e.g., microcystin, cyanobacteria biovolume).
5.5.4 Molecular methods
There are other measures that can be used for indicating the presence of cyanobacteria and their potential toxins. Foremost among these are the variety of molecular methods that can be used to detect specific genes that identify various cyanobacteria species as well as the presence of the toxin-producing genes. The relationship between molecular methods results and detection using more traditional methods (i.e., microscopy, enzyme-linked immunosorbent assay [ELISA], physicochemical analysis) is not always clear. In a study by Chiu et al., (2017), cell enumeration and microcystin concentration results obtained via microscopy and ELISA methods, respectively, were found to correlate well with their corresponding gene copies. However, Beversdorf et al. (2015) found that mcy gene concentrations were highly variable and showed a negative correlation with toxin concentrations at some locations. Therefore, although the relationship between molecular method results (e.g., mcy gene detection) and more traditional methods (e.g., physicochemical methods or ELISA) may be variable, molecular methods can be used as a screening tool to determine the presence of cyanobacteria species and to provide an indication of the potential for toxin production. Further information on molecular methods can be found in Section 6.0.
5.6 Control of cyanobacterial blooms
The most effective component of a long-term strategy for reducing the incidence of planktonic cyanobacterial blooms is to control the input of nutrients into the water body, specifically the input of phosphorus and nitrogen as their availability controls cyanobacterial growth (Downing et al., 2001; Jančula and Maršálek, 2011; Paerl et al., 2011a; Matthijs et al., 2012; Merel et al., 2013; Glibert et al., 2016; Hamilton et al., 2016). However, this may not be possible in all locations. In some water bodies, a major source of nutrients may be from internal loading of phosphorus from the sediment. It is important to differentiate whether this phosphorus load is from the degradation of recently sedimented organic material and is likely to decrease once the external load decreases, or if it is likely to be a long-term problem requiring specific remediation measures (e.g., in waterbodies with a low water exchange rate). Where nutrient inputs are the result of external influences, one way in which nutrient control can be achieved is through effective control of agricultural, municipal sewage, and residential waste disposal practices in the watershed. Nutrient input via wastewater and industrial effluent, and runoff from urban or agricultural/ deforested areas can be influenced by rainfall events. As a result, a nutrient control strategy that addresses the connection between climate change and nutrient loading will be important (Paerl et al., 2011b; Carey et al., 2012). Other efforts to develop strategies that are aware and adaptive to increasing climatic variability and extremes are continuing (Hamilton et al., 2016; Paerl, 2017). In addition to nutrient input, it has been reported that herbicides from agricultural activities may improve the growth conditions for cyanobacteria by decreasing the eukaryotic algae, thereby removing this competitor (Beaulieu et al., 2014).
Alternative approaches have also been used for controlling cyanobacteria blooms, usually when an immediate action is required, or nutrient management is not feasible. This may involve both direct and indirect control of cyanobacteria. Direct control methods, such as the addition of copper sulphate or other algaecides to mature toxic blooms, have been used, however this approach is not usually recommended. Although it will destroy the cyanobacteria cells, it will also cause the release of intracellular cyanotoxins into the surrounding waters if present within the cells. Jones and Orr (1994) reported that microcystin-LR could be detected up to 21 days after algaecide treatment of a toxic Microcystis aeruginosa bloom that had developed in a recreational lake. There is also some evidence that repeated treatments with algaecides may lead to the development of algaecide-resistant cells (Garcia-Villada et al., 2004). Algaecides may be recommended as an emergency measure in the early stages of a bloom when the resultant toxin concentrations released from the lysed cells would be low. Environmental concerns have also been cited as additional reasons for not pursuing this approach, as the algaecides can be detrimental to the healthy functioning of the aquatic ecosystem. The addition of hydrogen peroxide (H2O2) has also been used to suppress cyanobacteria blooms as cyanobacteria are more sensitive to H2O2 than other phytoplankton (e.g., green algae, diatoms) (Barroin and Feuillade, 1986; Barrington and Ghadouani, 2008). H2O2 has the advantage that it rapidly degrades to hydrogen and oxygen in the aquatic ecosystem, and because it is a strong oxidant, it may also help degrade any cyanotoxins present in the bloom (Matthijs et al., 2012). Indirect methods for cyanobacteria bloom control may also be used. This could include artificial mixing, flushing, and treatment of the water with phosphate-binding clays. Before direct or indirect measures are used, consideration needs to be given to the chemical, physical and biological characteristics of the watershed, as well as the costs and the environmental and social acceptability of the approach being considered. Only after a thorough scientific evaluation should a decision be made on the best solution for cyanobacterial control. Further information on control measures can be found in the Guidelines for Canadian Drinking Water – Guideline Technical Document on Cyanobacterial toxins (Health Canada, 2017).
6.0 Analytical methods
A comprehensive review of the analytical methods and limitations associated with each measurement can be found in the Guidelines for Canadian Drinking Water Quality – Guideline Technical Document for Cyanobacterial toxins (Health Canada, 2017). Information on monitoring methods used in recreational waters is also available in U.S. EPA (2017) and Meriluoto et al., (2017). There is no single method available to identify and quantify all of the different types of cyanotoxins and their variants simultaneously (Merel et al., 2013). Below is a brief overview of commonly used methods.
6.1 Microcystins
In order to compare the concentration of microcystins in a sample to the guideline value, it is necessary to determine the concentration of total microcystins – this includes both free and cell-bound microcystins. Initial processing steps are required to extract cell-bound toxins and to concentrate the dissolved toxins in the sample. These may include concentration of the cyanobacterial cells, cell lysis and toxin extraction and purification.
The analytical methods for microcystins that are currently being used in commercial and research laboratories include:
- Enzyme-linked immunosorbent assay (ELISA);
- physicochemical analysis by chromatographic separation (i.e., liquid chromatographic (LC) methods, such as high-performance liquid chromatography (HPLC) and ultra-high-performance liquid chromatography (UHPLC) and detection by either ultraviolet (UV) absorbance (photodiode array detector (PDA)) or mass spectrometry (MS); and
- protein phosphatase inhibition assays (PPIA).
Liquid chromatography (LC) coupled with mass spectroscopy (MS) is the most commonly used laboratory method for the identification and quantification of microcystin variants and represents the reference standard against which other methods are judged. Standardized procedures for this method have been described, and many analytical laboratories possess the necessary instrumentation for this time consuming and technically demanding analysis. A limitation of MS is the need for certified reference standards for quantitative results. These standards are not available for all of the relevant microcystin variants. In contrast, UV-PDA detection can use a small number of microcystins to quantify the peaks for all other microcystins (identified by their characteristic adsorption spectra) in a sample. This method is significantly less sensitive than LC-MS, with a detection limit near 1μg/L (Health Canada, 2017).
ELISA and PPIA field test kits are useful as they provide an estimate of the level of dissolved microcystins in a sample (lysis is needed to measure intracellular microcystins) within certain concentration ranges or above/below a specified concentration. ELISA and PPIA methods are rapid and sensitive. While they are not specific enough to distinguish between individual microcystin variants, in practice they may be sufficient for current risk assessments where all microcystins are assumed to be as toxic as microcystin-LR. Although these kits do not give quantitative results for determining if water samples meet the guideline value, they can be used as a screening tool for determining the presence or absence of toxin in recreational waters. The responsible authority should also be aware that some test kits detect both microcystins and nodularins.
6.2 Cyanobacteria cells, biovolumes, and chlorophyll a
Cyanobacteria cell concentrations are determined by direct microscopic count with a counting chamber of known dimensions and then back-calculating to the volume of the original sample (APHA, 2017; Chorus and Bartram, 1999). Cyanobacteria cells come in different shapes and sizes (from round to filamentous), groups of cells can exist as dense colonies or as long filaments, and populations can be composed of a mixture of these cell types. Identifying cyanobacteria to the species level can be difficult and may be further hampered as the key morphological features used to identify the individual taxa can change depending on the environment and growth stage (Chorus and Bartram, 1999; Yoshida et al., 2008; Carmichael and Boyer, 2016). However, for risk assessment, identification to the genus level is often sufficient. Individuals can learn to count and identify cyanobacteria cells if they have some experience in microscopy and receive specific training for identifying and quantifying cyanobacteria.
Similar to cyanobacteria cell counts, biovolumes are determined by first counting the number of each species of cyanobacteria present in the sample using a microscope, and then using the average cell volume to calculate the total biovolume concentrations (CEAEQ, 2012a, 2012b). Average cell volumes for many of the common cyanobacteria species found have been published (Wood et al., 2008; CEAEQ, 2012a, 2012b). It is also possible to calculate the biovolume of individual cells, as opposed to using average cell volumes (Zohary et al., 2016). Biovolume estimates share many of the same methodological difficulties as cell counts, with the additional complication that cell volumes can vary greatly within the same species, as well as between watersheds and regions.
Chlorophyll a measurements may be easier to carryout than cyanobacteria cell counts or biovolumes. This pigment is easily detected using spectrophotometric methods, fluorometric methods, or high-performance liquid chromatographic methods (Millie et al., 2010; APHA, 2017). In-situ monitoring using submersible probes has also been used (Zamyadi et al., 2012b). It should be noted that chlorophyll fluorescence measurements are influenced by environmental conditions and the physiological state of the cells (Ibelings et al., 2014), and in some cases, corrections need to be applied to get accurate measurements (Bertone et al., 2019).
6.3 Molecular methods
Molecular methods are being widely used to detect genetic material in environmental samples. Numerous molecular tools have been developed to detect cyanobacteria (Kurmayer et al., 2017). For example, the use of qPCR methodologies is routinely reported in the literature. This method uses primers targeting species-specific gene fragments to differentiate between various species in a cyanobacterial population. Primers have also been developed to detect toxin genes, providing an indication as to whether cells are capable of producing toxins (Rinta-Kanto et al., 2005). For example, Microcystis cell numbers and the microcystin production gene can be detected using a 16S rRNA region unique to all Microcystis cells and the mcy gene, respectively (Tillett et al., 2000; Kurmayer and Kutzenberger, 2003; Fortin et al., 2010; Chiu et al., 2017). Primers have also been published for Cylindrospermopsis (recently renamed to Raphisiopsis), cylindrospermopsin-producing cyanobacteria, saxitoxins, and nodularins (Chiu et al., 2017; Gaget et al., 2017b), and new primers are continually being investigated. Strengths and weaknesses of PCR methods have been published elsewhere (Gaget et al., 2017b). The development of molecular tools is a continually evolving field of research that holds a lot of promise for improving the methods available to responsible authorities for making public health decisions. Advances in this field will continue to be monitored.
7.0 Rationale
The recommended guideline value for total microcystins in Canadian recreational waters is based upon the approach used in the derivation of the maximum acceptable concentration (MAC) for microcystin-LR in the Guidelines for Canadian Drinking Water – Guideline Technical Document on Cyanobacterial Toxins (Health Canada, 2017). In brief, exposure to microcystin is expected to be for short durations, and hence the best available study to represent this type of exposure was determined to be the short-term exposure study conducted by Heinze et al., (1999). This study used a duration of 28 days and the exposure route was ingestion via drinking water. This is a change from the 2012 Guidelines for Canadian Recreation Water Quality total microcystins guideline which used the longer-term exposure study (13 weeks) published by Fawell et al., 1999. The complete health risk assessment, including how the key study was selected, is described in Health Canada (2017). The change in the key study, along with changes to the uncertainty factors and the exposure factors has resulted in a lower guideline value for total microcystins. The guideline values for total cyanobacteria cells, total cyanobacteria biovolume, and total chlorophyll a are based on their relationship to microcystin. Guidelines are not established for other cyanotoxins, including anatoxin-a and cylindrospermopsin, as health or exposure data on these toxins are limited.
The use of single guideline values for each parameter is the preferred approach to setting guidelines for Canadian recreational waters at this time. They are intended to protect against both the risk of exposure to microcystins through inadvertent ingestion of water as well as from other harmful effects that may be possible after exposure to high densities of cyanobacterial material. The guideline values were derived based on children as they are more likely to accidentally swallow contaminated water and often spend more time in water than adults.
7.1 Total microcystins
The guideline value for total microcystins is calculated based on the toxicity of microcystin-LR, and is intended to be protective against exposure to other microcystin variants that may be present. For microcystin-LR, the tolerable daily intake (TDI) for recreational water exposures is derived as follows:
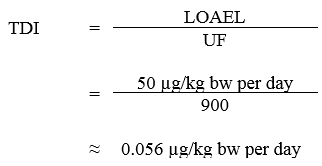
Text description
Equation for TDI:
This equation calculates the tolerable daily intake for microcystin-LR. The tolerable daily intake is calculated by dividing the lowest observed adverse effect level for microcystin-LR (50 micrograms per kilogram of body weight per day) by the uncertainty factor of 900 which equals 0.056 micrograms per kilogram of body weight per day.
Equation 1
where:
- 50 μg/kg body weight (bw) per day is the lowest observed adverse effect level (LOAEL) for increased liver weight and slight to moderate liver lesions with hemorrhages in rats as reported by Heinze (1999); and
- 900 is the uncertainty factor (UF): ×10 for intraspecies variability, ×10 for interspecies variability, ×3 for deficiencies in the health effects database and ×3 for the use of a LOAEL instead of a no observed adverse effect level (NOAEL).
Using this TDI, the health based value (HBV) for total microcystins can be derived as follows:
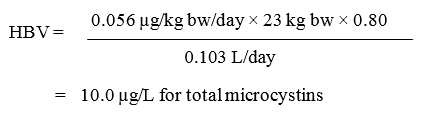
Text description
Equation for HBV:
This equation calculates the health based value for microcystin-LR. The health based value is calculated by multiplying the tolerable daily intake of 0.056 micrograms per kilogram of body weight per day by a body weight of 23 kilograms and by an allocation factor of 0.80 and then dividing by a volume of water ingested per day of 0.103 litres. This equals a health based values of 10.0 micrograms per litre for total microcystins.
Equation 2
where:
- 0.056 µg/kg bw/day is the TDI as derived above;
- 23 kg bw is the average weight of a Canadian child aged 4 to 8 years (Health Canada, unpublished);
- 0.80 is the “ceiling value” allocation factor, as the majority of exposure to microcystins is expected to be through water ingestion during recreational activities; the remaining 0.20 allows for allocation to other non-negligible exposures from other media (Krishnan and Carrier, 2013); and
- 0.103 L/day (103 mL/day) is the estimated amount accidentally ingested per day during recreational water activities by a child aged 6 to 10 years (38 mL/h x 2.7 h/day)
The amount of water accidentally ingested per day uses ingestion rates based on an American study published by Dufour et al. (2017). The study enrolled 549 participants from the ages of 6 to adult, recreating in a pool setting. The use of the pool environment allowed the researchers to determine the amount of water ingested per participant by conducting a 24-hour urinalysis for a chemical compound found in the chlorinated pool environment. The average ingestion rate from all study participants was 32 mL/h. Since children are the target population due to their increased exposure (see below) to recreational activities in water, the average ingestion rate for children was used as opposed to the general population ingestion rate. The average ingestion rates from Dufour (2017), broken down by age categories, are published in United States Environmental Protection Agency (U.S. EPA, 2019). For children aged 6 to 10, this corresponds to an average incidental ingestion rate of 38 mL/h. The average ingestion rate for teens (11-17 years) was slightly higher (40 mL/h), however, as their average body weight is also greater (42 kg for ages 9-13 years; Health Canada, unpublished), the younger children are still the more susceptible age group. There is no ingestion data available from this study for children younger than 6 years. The ingestion rates used are supported by those previously obtained by Dufour et al. (2006) in a similar pilot study (average 21 mL/h for adults; 49 mL/h for children), and are more conservative than results obtained in a separate small pool study by Suppes et al. (2014) (average 3.5 mL/h for adults, 25.7 mL/h for children). It is not surprising that children consume the greatest amount of water while swimming as they often engage in playful behaviours that increase the odds of them swallowing water such as spouting water for fun, “bobbing” in the water, and splashing.
The duration of exposure to recreational water is based on data collected during the National Human Activity Pattern Survey, conducted in the United Sates and reported in the U.S. EPA Exposure Factors Handbook. Data from the United States has been used, as this information has not been collected for the Canadian population. The data shows that children aged 5-11 years spend the greatest amount of time (in outdoor pools) with an average exposure of 2.7 hours/day (164.2 minutes/day), whereas children aged 1-4 years spent 1.4 hours/day (85.6 minutes/day) and 12-17 years spent1.6 hours/day (97.0 minutes/day) (U.S. EPA, 2011). Another large study, conducted in the Netherlands, reported average swimming event durations for children aged 0-14 years to be 1.4 hours (81 minutes), 1.3 hours (79 minutes) and 1.1 hours (65 minutes) for pools, fresh waters and marine waters, respectively (Schets et al., 2011). The average body weight for children in Canada corresponding to the greatest exposure duration is 23 kg (based on ages 4- 8 years) (Health Canada, unpublished). As younger children (aged 1-4 years) spend approximately half the amount of time in the pool, the guideline value derived using older children is also protective of this younger age group.
7.2 Total cyanobacteria cells
The guideline value for total cyanobacteria cells represents a general indication of the potential for bloom development and is intended to be protective against exposure to high levels of cyanobacterial toxins and high densities of cyanobacterial material. It is calculated based on the microcystin guideline value using a reference value for the average toxin quota for Microcystis cells.
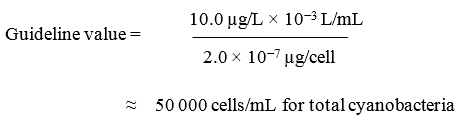
Text description
Equation for Guideline value:
This equation calculates the guideline value for total cyanobacteria cells. The health-based value for total microcystins of 10.0 micrograms per litre is multiplied by 10-3 litres per milliliter and then divided by a toxin cell quota for Microcystis of 2.0 times 10-7 micrograms per cell. This equals a guideline value of 50,000 cells per millilitre.
Equation 3
where:
- 10.0 µg/L is the health based value for total microcystins in recreational waters, as derived in the previous section;
- 2.0 × 10−7 µg/cell is the toxin cell quota for total microcystins per Microcystis cell (Fitzgerald et. al., 1999; WHO, 2003) and
- 10−3 L/mL is the factor converting litres to millilitres.
The average toxin quota used in this guideline value has also been used internationally for the development of total cyanobacteria cell count guideline values (NHMRC, 2008; WHO, 2003). Other average toxin quotas reported for Microcystis cells from natural environments are lower, and range from 1.9× 10−9 µg/cell to 1.4 × 10−7 µg/cell (Kurmayer et al., 2003; Lyck et al., 2003; Fahnenstiel et al., 2008). This approach assumes all cells in the bloom are producing microcystin, providing a conservative estimate for cyanobacteria cell counts. This approach is also intended to provide protection against exposure to blooms of other cyanobacteria with toxic potential, not just the microcystin-producing species.
Although the total cyanobacteria cell count guideline is derived based on the potential for toxin production, cyanobacteria cells themselves have been implicated in adverse health reactions. There is significant variability in the concentration of cyanobacteria cells necessary to negatively impact health. Backer et al. (2015) reviewed harmful algal bloom data from 2007-2011 and reported cyanobacteria cell counts from samples collected in response to health events. The cell count values ranged over several orders of magnitude from as low as approximately one hundred cells per millilitre to as high as several million cells per millilitre. Earlier studies by Pilotto et al. (1997, 2004) reported that rates of gastrointestinal symptom occurrence were significantly higher after exposure for more than an hour to cyanobacteria cell concentrations ranging from 5000 to 20 000 cells/mL, however, only a small number of people were affected, and only with mild irritation. A study by Lévesque et al., (2014) also showed a link between cyanobacteria densities and gastrointestinal health impacts. A prospective epidemiological study, using cell surface area as the principal exposure variable, observed that symptoms were more likely to be reported among persons exposed to moderate or high (approximately 20 000 cells/mL to approximately 100 000 cells/mL) versus low (< approximately 20 000 cells/mL) levels of cyanobacteria (Stewart, 2004; Stewart et al., 2006). Respiratory symptoms were recorded with the greatest frequency, albeit with mild severity (e.g., some difficulty breathing, dry cough, sore throat, sneezing, and runny nose) when cyanobacteria levels were high. Due to this variability, it was not possible to derive a guideline value based on the risk of allergenic or other irritative effects caused by levels of unknown cyanobacterial material. The guideline value based on the potential for microcystin production is intended to be protective of exposure to both high levels of toxins as well as high levels of cyanobacterial material.
7.3 Cyanobacteria biovolume
The biovolume guideline value for cyanobacteria is calculated using the cyanobacteria cell count guideline (from Section 7.2) combined with the average cell volume of Microcystis, as they reflect the highest likely water quality hazard scenario of a toxic bloom containing high levels of microcystin (Kemp and John, 2006). The guideline value for cyanobacteria biovolume can be calculated as follows:
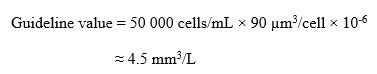
Text description
Equation for Guideline value:
This equation calculates the guideline value for cyanobacteria biovolume. The guideline value for total cyanobacteria cells of 50,000 cells per milliliter is multiplied by the average cell volume of 90 micrometers cubed per cell multiplied by a 10-6 unit conversion factor to give a value of 4.5 millimeters cubed per liter.
Equation 4
where:
- 50 000 cells/mL is the guideline value for total cyanobacteria cells in recreational waters, as derived above;
- 90 µm3/cell is the average cell volume for large Microcystis cells (Wood et al., 2008; NHMRC, 2011)
- 10-6 is a units conversion from µm3/mL to mm3/L.
7.4 Chlorophyll a
The guideline value for total chlorophyll a is calculated based on the observed ratio between microcystin and chlorophyll a in blooms where Microcystis dominates the population, as this represents a water-quality hazard scenario of potentially elevated microcystin concentrations (Carrasco et al., 2006; Kemp and John, 2006):
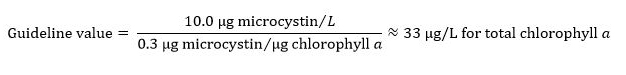
Text description
Equation for Guideline value:
This equation calculates the guideline value for chlorophyll a. The health based value for total microcystins of 10.0 micrograms per litre is divided by an average microcystin content per microgram of chlorophyll a of 0.3 to produce a guideline value of 33 micrograms per litre for total chlorophyll a.
Equation 5
where:
- 10.0 µg/L is the HBV for total microcystin in recreational waters, as derived above; and
- 0.3 µg is the average microcystin content of per µg of chlorophyll a (Carrasco et al., 2006)
Chlorophyll a is frequently used as an index for eutrophication and has been used as an indicator of cyanobacterial biomass or cyanotoxins. The guideline value derived for chlorophyll a is based on conservative assumptions and therefore, at the guideline value, the concentration of microcystin (if present) is unlikely to exceed the MAC of 10 μg/L.
The relationship between chlorophyll a and both cyanobacteria biovolume and cyanotoxin concentrations is inconsistent and dependent on the environmental conditions, with both positive and negative associations found (Hartshorn et al., 2016; Zastepa et al., 2017). Other phytoplankton also contain chlorophyll a so this measurement works best when cyanobacteria are the dominant organisms present. In an analysis of data on a national scale in the US, threshold levels of chlorophyll a associated with a greater than 50% probability of exceeding a health advisory level for microcystin were reported (i.e., microcystin levels between 0.3 and 2.0 µg/L were associated with chlorophyll a concentrations between 23 and 104 µg/L) (Hollister and Kreakie, 2016). Although there is variability in the relationship between cyanobacteria and chlorophyll a, this indicator can be used as part of a cyanobacteria alert system to trigger further investigation and actions (see Section 2.0).
7.5 Other cyanobacterial toxins
Currently, the data available for anatoxin-a are insufficient for deriving a recreational water quality guideline value. A guideline value for cylindrospermopsin is also not established, since the toxin is infrequently detected in Canadian surface waters, according to available occurrence data. A cylindrospermopsin guideline value has been established by the U.S. EPA (2019). Further information on cylindrospermopsin can be found in Health Canada (2017).
7.6 International considerations
Numerous other countries worldwide include recommended guideline values for cyanobacteria and their toxins (see Table 2). These guidelines or standards may vary from the guidelines developed by Health Canada due to their individualized assessments as well as differing policies and approaches. In general, the health risk assessments for microcystin concentrations have resulted in similar values worldwide. Only one jurisdiction has developed a guideline value for cylindrospermopsin (U.S. EPA, 2019). The parameters used as indicators of potential health risk differ as many jurisdictions use alert levels as opposed to a single value approach. In general, the Health Canada values align with the alert/action modes or moderate risk values provided by the WHO.
| Country/ organization | Guideline values | Reference | ||||
|---|---|---|---|---|---|---|
| Microcystin concentration | Cell Counts (cells/mL)Table 2 Footnote b |
BiovolumeTable 2 Footnote b | Chlorophyll aTable 2 Footnote b | |||
| U.S EPATable 2 Footnote a | 8 µg/L | No value included | No value included | No value included | U.S EPA, 2019 | |
| WHO | Low Risk |
No value included | Total cyanobacteria: < 20 000 |
No value included | Total cyanobacteria: 10 µg/L |
WHO, 2003 |
| Moderate Risk |
20 000 – 100 000 |
10-50 µg/L | ||||
| High Risk | Scum formation in areas with risk ingestion/aspiration | >50 µg/L | ||||
| Australia | Surveillance Mode | — | Microcystis: 500 – 5000 |
Microcystis: 0.04 – 0.4 mm3/L |
No value included | NHMRC, 2008 |
| Alert Mode | — | 5000 – 50 000 | 0.4 – 4 mm3/L | |||
| Action Mode | ≥10 µg/L | ≥50 000 | ≥4 mm3/L If known toxin producers are not present, alert mode is 4 – 10 mm3/L, action mode is ≥10 mm3/L | |||
| European Union | No value included | Total cyanobacteria: 100 000 |
No value included | Total cyanobacteria: 50 µg/L |
European Commission, 2009 | |
| New Zealand | Surveillance Mode | — | Total cyanobacteria : 500 |
Total cyanobacteria : 0.5 mm3/L |
No value included | New Zealand Ministry for the Environment and Ministry of Health, 2009 |
| Alert Mode | — | 500 –19 000 | 0.5 – 10 mm3/L | |||
| Action Mode | ≥12 µg/L | ≥19 000 | ≥10 mm3/L If known toxin producers are present, alert mode is 0.5 – 1.8 mm3/L , action mode is ≥ 1.8 mm3/L | |||
|
||||||
8.0 References
- Akcaalan, R., Mazur-Marzec, H., Zalewska, A. and Albay, M. (2009). Phenotypic and toxicological characterization of toxic Nodularia spumigena from a freshwater lake in Turkey. Harmful Algae, 8(2): 273-278.
- ANSES (2017). Données relatives à la toxicité aiguë et chronique de la bêta-méthylamino-L-alanine (BMAA). Avis de l'Anses, Rapport d'expertise collective. Agence nationale de sécurité sanitaire de l'alimentation, de l'environnement et du travail. France.
- APHA (2017). Standard methods for the examination of water and wastewater. American Public Health Association, American Water Works Association, and Water Environment Foundation. Washington, DC.
- Aráoz, R., Molgó, J. and Tandeau de Marsac, N. (2010). Neurotoxic cyanobacterial toxins. Toxicon, 56(5): 813-828.
- Backer, L.C., McNeel, S.V., Barber, T., Kirkpatrick, B., Williams, C., Irvin, M., Zhou, Y., Johnson, T.B., Nierenberg, K., Aubel, M., LePrell, R., Chapman, A., Foss, A., Corum, S., Hill, V.R., Kieszak, S.M. and Cheng, Y.S. (2010). Recreational exposure to microcystins during algal blooms in two California lakes. Toxicon, 55(5): 909-921.
- Backer, L.C., Landsberg, J.H., Miller, M., Keel, K. and Taylor, T.K. (2013). Canine cyanotoxin poisonings in the United States (1920-2012): Review of suspected and confirmed cases from three data sources. Toxins, 5(9): 1597-1628.
- Backer, L.C., Manassaram-Baptiste, D., LePrell, R. and Bolton, B. (2015). Cyanobacteria and algae blooms: Review of health and environmental data from the harmful algal bloom-related illness surveillance system (HABISS) 2007-2011. Toxins, 7(4): 1048-1064.
- Banack, S.A., Johnson, H.E., Cheng, R. and Cox, P.A. (2007). Production of the neurotoxin BMAA by a marine cyanobacterium. Marine Drugs, 5(4): 180-196.
- Banack, S.A., Caller, T., Henegan, P., Haney, J., Murby, A., Metcalf, J.S., Powell, J., Cox, P.A., and Stommel, E. (2015). Detection of cyanotoxins, β-N-methylamino-L-alanine and microcystins, from a lake surrounded by cases of amyotrophic lateral sclerosis. Toxins, 7:322-336.
- Barrington, D.J., and Ghadouani, A. (2008). Application of hydrogen peroxide for the removal of toxic cyanobacteria and other phytoplankton from wastewater. Environmental Science and Technology, 42(23): 8916-8921.
- Barroin, G., and Feuillade, G. (1986). Hydrogen peroxide as a potential algicide for Oscillatoria rubescens D.C., Water Research, 20(5): 619-623.
- Bartram, J., Burch, M., Falconer, I.R., Jonem, G., KuiperGoodman, T., 1999. Situation assessment, planning and management. In: Chorus, I., Bartram, J, (Eds.), Toxic Cyanobacteria in Water. A Guide to their Public Health Consequences, Monitoring and Management. E&FN Spon on behalf of the World Health Organization, London, pp. 179–209.
- Beaulieu, M., Trudel, M., Marcil, G-K., Huot, Y., Lacey, J., Leconte, R., and Cabana, H. (2014). Pesticides et phytoplancton au Québec : Conditions propices aux efflorescences cyanobactériennes? Revue de littérature. Université de Sherbrooke, Sherbrooke, Canada (http://environnement.recherche.usherbrooke.ca/Revue_de_Litte%CC%81rature_Pesticides_et_Phytoplancton_au_Que%CC%81bec_8jan2014.pdf)
- Bernard, C., Ballot, A., Thomazeau, S., Maloufi, S., Furey, A., Mankiewicz-Boczek, J., Pawlik-Skowrońska, Capelli, C., and Salmaso, N. (2017). Cyanobacteria associated with the production of cyanotoxins. In: Meriluoto, J., Spoof, L., and Codd, G.A. (Eds). Handbook of cyanobacterial monitoring and cyanotoxin analysis. Wiley, European Cooperation in Science and Technology, Cyanocost, United Kingdom.
- Bertone, E., Chuang, A., Burford, M.A., and Hamilton, D.P. (2019). In-situ fluorescence monitoring of cyanobacteria: laboratory-based quantification of species-specific measurement accuracy. Harmful Algae, 87: 101625.
- Beutel, M.W., Horne, A.J., Roth, J.C. and Barratt, N.J. (2001). Limnological effects of anthropogenic desiccation of a large, saline lake, Walker Lake, Nevada. Hydrobiologia, 466(1): 91-105.
- Beversdorf, L.J., Chaston, S.D., Miller, T.R. and McMahon, K.D. (2015). Microcystin mcyA and mcyE gene abundances are not appropriate indicators of microcystin concentrations in lakes. PLoS One, 10(5): e0125353.
- Blais, S. (2008). Guide d'identification des fleurs d'eau de cyanobactéries. Comment les distinguer des végétaux observés dans nos lacs et nos rivières, 3e édition, Direction du suivi de l'état de l'environnement, ministère du Développement durable, de l'Environnement et des Parcs, ISBN 978-2-550- 52408-3.
- Blais, S. (2014). État de situation sur les cyanobactéries à la baie Missisquoi de 2000 à 2008 en lien avec les seuils provisoires pour les eaux récréatives. Québec, ministère du Développement durable, de l'Environnement et de la Lutte contre les changements climatiques, Direction du suivi de l'état de l'environnement, ISBN 978-2-550-71485-9.
- Blais, S. (2015). Faits saillants – État de situation sur les cyanobactéries à la baie Missisquoi de 2000 à 2008 en lien avec les seuils provisoires pour les eaux récréatives. Québec, ministère du Développement durable, de l'Environnement et de la Lutte contre les changements climatiques, Direction du suivi de l'état de l'environnement, ISBN 978-2-550-72639-5.
- Blais, S. (2019). État de situation sur les cyanobactéries et les algues eucaryotes à la baie Missisquoi en 2014. Québec, ministère de l'Environnement et de la Lutte contre les changements climatiques, Direction générale du suivi de l'état de l'environnement, ISBN 978-2-550-77638-3.
- Bouaïcha, N., Miles, C.O., Beach, D.G., Labidi, Z., Djabri, A., Benayache, N.Y., and Nguyen-Quang, T. (2019). Structural diversity, characterization and toxicology of microcystins. Toxins, 11(12): 714.
- Bourbonnais, N. and Robert, C. (2014). Bilan de la gestion des épisodes de fleurs d'eau d'algues bleu-vert au Québec, de 2007 à 2012. Québec, Ministère du Développement durable, de l'Environnement, de la Faune et des Parcs, Direction du suivi de l'état de l'environnement, ISBN 978-2-550-70347-1.
- Bowling, L., Blais, S., and Sinotte, M. (2014). An analysis of cyanobacterial bloom occurrence in Missisquoi Bay (Québec, Canada) between 2000 and 2008, and possible environmental factors underlying them. New South Wales Government, Department of Primary Industries, Office of Water, Australia.
- Bownik, A. (2010). Harmful algae: effects of alkaloid cyanotoxins on animal and human health. Toxin Rev., 29: 99–114.
- Boyer, G.L. (2007). The occurrence of cyanobacterial toxins in New York lakes: Lessons from the MERHAB-lower great lakes program. Lake Reserv. Manage., 23(2): 153-160.
- Briand, J.F., Jacquet, S., Flinois, C., Avois-Jacquet, C., Maisonnette, C., Leberre, B., and Humbert, J.F. (2005). Variations in the microcystin production of Planktothrix rubescens (Cyanobacteria) assessed from a four-year survey of Lac du Bourget (France) and from laboratory experiments. Microb. Ecol., 50:418-28.
- Bridgeman, T.B. and Penamon, W.A. (2010). Lyngbya wollei in Western Lake Erie. Journal of Great Lakes Research, 36(1): 167-171.
- Brient, L., Lengronne, M., Bertrand, E., Rolland, D., Sipel, A., Steinmann, D., Baudin, I., Legeas, M., Le Rouzic, B. and Bormans, M. (2008). A phycocyanin probe as a tool for monitoring cyanobacteria in freshwater bodies. J. Environ. Monit., 10(2): 248-255.
- British Columbia Ministry of Health (2012). Personal communication with B. Boettger.
- Burford, M.A. and Davis, T.W. (2011). Physical and chemical processes promoting dominance of the toxic cyanobacterium Cylindrospermopsis raciborskii. Chin. J. Oceanol. Limnol., 29(4): 883-891.
- Burford, M.A., Carey, C.C., Hamilton, D.P., Huisman, J., Paerl, H.W., Wood, S.A., and Wulff, A. (2019). Perspective: Advancing the research agenda for improving understanding of cyanobacteria in a future of global change. Harmful Algae, https://doi.org/10.1016/j.hal.2019.04.004.
- California Water Quality Monitoring Council (2019). SWAMP – HAB field guide – visual guide to observing blooms. Available at: https://mywaterquality.ca.gov/habs/resources/field.html.
- Carey, C.C., Ibelings, B.W., Hoffmann, E.P., Hamilton, D.P. and Brookes, J.D. (2012). Eco-physiological adaptations that favour freshwater cyanobacteria in a changing climate. Water Research, 46(5): 1394-1407.
- Carillo, E., Ferrero, L., M., Alonso-Andicoberry, C., Basanta, A., Martin, A., López-Rodas, V. and Costas, E. (2003). Interstrain variability in toxin production in populations of the cyanobacterium Microcystis aeruginosa from water-supply reservoirs of Andalusia and lagoons of Doñana National Park (Southern Spain). Phycologia, 42(3): 269-274.
- Carmichael, W.W. and Boyer, G.L. (2016). Health impacts from cyanobacteria harmful algae blooms: Implications for the North American Great Lakes. Harmful Algae, 54:194-212.
- Carmichael, W.W. and Gorham, P.R. (1978). Anatoxins from clones of Anabaena flos-aquae isolated from lakes of Western Canada. Mitt. Int. Ver. Limnol, 21(1): 285-295.
- Carrasco, D., Moreno, E., Sanchis, D., Wörmer, L., Paniagua, T., Del Cueto, A. and Quesada, A. (2006). Cyanobacterial abundance and microcystin occurrence in Mediterranean water reservoirs in central Spain: Microcystins in the Madrid area. Eur. J. Phycol., 41(3): 281-291.
- Carvalho L., McDonald, C., de Hoyos, C., Mischke, U., Philipps, G., Borics, G., Poikane, S., Skjelbred, B., Lyche Solheim, A., van Wichelen, J., and Cardoso, A.C. (2013). Sustaining recreational quality of European lakes: minimizing the health risks from algal blooms through phosphorus control. J. Appl. Ecol., 50:315–323.
- CEAEQ (Centre d'expertise en analyse environnementale du Québec). (2012a). Identification, dénombrement et estimation du biovolume des cyanobactéries et des algues. MA. 800 – Cya.alg 1.0, Québec, ministère du Développement durable, de l'Environnement et des Parcs.
- CEAEQ (Centre d'expertise en analyse environnementale du Québec). (2012b). Dépistage des cyanobactéries. MA. 800 – Cya.dep 1.0, Québec, ministère du Développement durable, de l'Environnement et des Parcs.
- Cerasino, L., Shams, S., Boscaini, A., and Salmaso, N. (2016). Multiannual trend of microcystin production in the toxic cyanobacterium Planktothrix rubescens in Lake Garda (Italy). Chemistry and Ecology, 32(5), 492-50.
- Chen, H., Burke, J., Paul Dinsmore, W., Prepas, E.,E. and Fedorak, P.,M. (2007). First assessment of cyanobacterial blooms and microcystin-LR in the Canadian portion of Lake of the Woods. Lake and Reservoir Management, 23(2): 169-178.
- Chiu, Y.T., Chen, Y.H., Wang, T.S., Yen, H.K. and Lin, T.F. (2017). A qPCR-based tool to diagnose the presence of harmful cyanobacteria and cyanotoxins in drinking water sources. Int. J. Environ. Res. Public. Health., 14(Article Number: 547): 1-17.
- Chong, M.W.K., Wong, B.S.F., Lam, P.K.S., Shaw, G.R. and Seawright, A.A. (2002). Toxicity and uptake mechanism of Cylindrospermopsin and Lophyrotomin in primary rat hepatocytes. Toxicon, 40(2): 205-211.
- Chorus, I. and Bartram, J. (eds.). (1999). Toxic cyanobacteria in water: A guide to their public health consequences, monitoring and management. E & FN Spon / Chapman & Hall, London, United Kingdom.
- Chorus, I., Falconer, I.R., Salas, H.J. and Bartram, J. (2000). Health risks caused by freshwater cyanobacteria in recreational waters. J. Toxicol. Environ. Health B Crit. Rev., 3(4): 323-347.
- Chorus, I. and Niesel, V. (2011). Steps towards a statistical model to predict phytoplankton responses to changes in trophic state. Chorus, I., Schauser, I., (Eds), In: Oligotrophication of Lake Tegel and Schlachtensee, Berlin Analysis of system components, causalities and response thresholds compared to responses of other waterbodies. Umweltbundesamt, Dessau, DE. 109-39.
- Christoffersen, K., Lyck, S., and Winding, A. (2002). Microbial activity and bacterial community structure during degradation of microcystins. Aquat. Microb. Ecol., 27: 125-136.
- Codd, G., Bell, S., Kaya, K., Ward, C., Beattie, K. and Metcalf, J. (1999). Cyanobacterial toxins, exposure routes and human health. Eur. J. Phycol., 34(4): 405-415.
- Conroy, J.D., Quinlan, E.L., Kane, D.D. and Culver, D.A. (2007). Cylindrospermopsis in Lake Erie: Testing its association with other cyanobacterial genera and major limnological parameters. Journal of Great Lakes Research, 33(3): 519-535.
- Cox, P.A., Banack, S.A. and Murch, S.J. (2003). Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA, 100(23): 13380-13383.
- Cox, P.A., Banack, S.A., Murch, S.J., Rasmussen, U., Tien, G., Bidigare, R.R., Metcalf, J.S., Morrison, L.F., Codd, G.A. and Bergman, B. (2005). Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA, 102(14): 5074-5078.
- Davis, T.W., Berry, D.L., Boyer, G.L. and Gobler, C.J. (2009). The effects of temperature and nutrients on the growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae, 8(5): 715-725.
- de la Cruz, A.A., Hiskia, A., Kaloudis, T., Chernoff, N., Hill, D., Antoniou, M.G., He, X., Loftin, K., O'Shea, K., Zhao, C., Pelaez, M., Han, C., Lynch, T.J. and Dionysiou, D.D. (2013). A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process. Impacts, 15(11): 1979-2003.
- Dong, X., Zeng, S., Bai, F., Li, D. and He, M. (2016). Extracellular microcystin prediction based on toxigenic Microcystis detection in a eutrophic lake. Sci. Rep., 6(Article Number: 20886): 1-8.
- dos Anjos, F.M., Bittencourt-Oliveira, M.C., Zajac, M.P., Hiller, S., Christian, B., Erler, K., Luckas, B. and Pinto, E. (2006). Detection of harmful cyanobacteria and their toxins by both PCR amplification and LC-MS during a bloom event. Toxicon, 48(3): 239-245.
- Downing, J.A., Watson, S.B. and McCauley, E. (2001). Predicting cyanobacteria dominance in lakes. Can. J. Fish. Aquat. Sci., 58(10): 1905-1908.
- Du, S., Huang, S., Pan, H., Zang, C., Wu, M., Lin, C., Guo, Y., Gao, F. and Scholz, M. (2014). Monitoring algal bloom through relationships between chlorophyll a and phytoplankton. Environ. Eng. Manage. J., 13(4): 805-815.
- Dufour, A.P., Evans, O., Behymer, T.D. and Cantu, R. (2006). Water ingestion during swimming activities in a pool: A pilot study. J. Water. Health., 4(4): 425-430.
- Dufour, A., Behymer, T.D., Cantu, R., Magnuson, M. and Wymer, L. (2017). Ingestion of swimming pool water by recreational swimmers. Journal of Water and Health, 15(3): 429-437.
- Dziuban, E.J., Liang, J.L., Craun, G.F., Hill, V., Yu, P.A., Painter, J., Moore, M.R., Calderon, R.L., Roy, S.L. and Beach, M.J. (2006). Surveillance for waterborne disease and outbreaks associated with recreational water-United States, 2003-2004. MMWR Surveill. Summ., 55(12): 1-30.
- Edwards, C., Beattie, K.A., Scrimgeour, C.M. and Codd, G.A. (1992). Identification of anatoxin-A in benthic cyanobacteria (blue-green algae) and in associated dog poisonings at Loch Insh, Scotland. Toxicon, 30(10): 1165-1175.
- European Commission (2009). Bathing water profiles: Best practice and guidance. European Commission, Directorate General Environment, B-1049 Brussels.
- Fahnenstiel, G.L., Millie, D.F., Dyble, J., Litaker, R.W., Tester, P.A., McCormick, M.J., Rediske, R. and Klarer, D. (2008). Microcystin concentrations and cell quotas in Saginaw Bay, Lake Huron. Aquat. Ecosyst. Health Manage., 11(2): 190-195.
- Falconer, I.R. (2005). Cyanobacterial toxins of drinking water supplies: Cylindrospermopsins and microcystins. CRC Press, Boca Raton, FL.
- Falconer, I.R. and Humpage, A.R. (2006). Cyanobacterial (blue-green algal) toxins in water supplies: Cylindrospermopsins. Environ. Toxicol., 21(4): 299-304.
- Fastner, J., Erhard, M., Carmichael, W.W., Sun, F., Rinehart, K.L., Rönicke, H., and Chorus, I. (1999). Characterization and diversity of microcystins in natural blooms and strains of the genera Microcystis and Planktothrix from German freshwaters. Arch. Hydrobiol., 145(2):147-63.
- Fastner, J., Abella, S., Litt, A., Morabito, G., Vörös, L., Pálffy, K., Straile, D., Kümmerlin, R., Matthews, D., Phillips, M.G., and Chorus, I. (2016). Combating cyanobacterial proliferation by avoiding or treating inflows with high P load—experiences from eight case studies. Aquatic Ecology, 50(3): 367-383.
- Fayad, P.B., Roy-Lachapelle, A., Duy, S.V., Prévost, M. and Sauvé, S. (2015). On-line solid-phase extraction coupled to liquid chromatography tandem mass spectrometry for the analysis of cyanotoxins in algal blooms. Toxicon, 108: 167-175.
- Fiedor, L., Kania, A., Myśliwa-Kurdziel, B., Orzeł, Ł. and Stochel, G. (2008). Understanding chlorophylls: Central magnesium ion and phytyl as structural determinants. Biochimica Et Biophysica Acta (BBA) - Bioenergetics, 1777(12): 1491-1500.
- Fitzgerald, D.J., Cunliffe, D.A. and Burch, M.D. (1999). Development of health alerts for cyanobacteria and related toxins in drinking water in South Australia. Environ. Toxicol., 14(1): 203-209.
- Fortin, N., Aranda-Rodriguez, R., Jing, H., Pick, F., Bird, D. and Greer, C.W. (2010). Detection of microcystin-producing cyanobacteria in Missisquoi Bay, Québec, Canada, using quantitative PCR. Appl. Environ. Microbiol., 76(15): 5105-5112.
- Funari, E. and Testai, E. (2008). Human health risk assessment related to cyanotoxins exposure. Crit. Rev. Toxicol., 38(2): 97-125.
- Gaget, V., Humpage, A.R., Huang, Q., Monis, P. and Brookes, J.D. (2017a). Benthic cyanobacteria: A source of cylindrospermopsin and microcystin in Australian drinking water reservoirs. Water Research, 124: 454-464.
- Gaget, V., Lau, M., Sendall, B., Froscio, S., and Humpage, A.R. (2017b). Cyanotoxins: which detection technique for an optimum risk assessment? Water Research, 118: 227-238.
- Garcı́a-Villada, L., Rico, M., Altamirano, M., Sánchez-Martı́n, L., López-Rodas, V., and Costas, E. (2004). Occurrence of copper resistant mutants in the toxic cyanobacteria Microcystis aeruginosa: characterisation and future implications in the use of copper sulphate as algaecide. Water Research, 38(8): 2207-2213.
- Gehringer, M.M., Adler, L., Roberts, A.A., Moffitt, M.C., Mihali, T.K., Mills, T.J., Fieker, C. and Neilan, B.A. (2012). Nodularin, a cyanobacterial toxin, is synthesized in planta by symbiotic Nostoc sp. Isme j., 6(10): 1834-1847.
- Glibert, P.M., Wilkerson, F.P., Dugdale, R.C., Raven, J.A., Dupont, C.L., Leavitt, P.R., Parker, A.E., Burkholder, J.M., and Kana, T.M. (2016). Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions. Limnol. Oceanogr., 61(1): 165-197.
- Giani, A., Bird, D.F., Prairie, Y.T. and Lawrence, J.F. (2005). Empirical study of cyanobacterial toxicity along a trophic gradient of lakes. Can. J. Fish. Aquat. Sci., 62(9): 2100-2109.
- Giannuzzi, L., Sedan, D., Echenique, R. and Andrinolo, D. (2011). An acute case of intoxication with cyanobacteria and cyanotoxins in recreational water in Salto Grande Dam, Argentina. Mar. Drugs, 9(11): 2164-2175.
- Graham, J.L., Loftin, K.A., Meyer, M.T. and Ziegler, A.C. (2010). Cyanotoxin mixtures and taste-and-odor compounds in cyanobacterial blooms from the Midwestern United States. Environ. Sci. Technol., 44(19): 7361-7368.
- Gregor, J., Maršálek, B. and Šípková, H. (2007). Detection and estimation of potentially toxic cyanobacteria in raw water at the drinking water treatment plant by in vivo fluorescence method. Water Research, 41(1): 228-234.
- Grutzmacher, G., Wessel, G., Klitzke, S., and Chorus, I. (2010). Microcystin elimination during sediment contact. Environ. Sci. Technol., 44: 657-662.
- Gunn, G.J., Rafferty, A.G., Rafferty, G.C., Cockburn, N., Edwards, C., Beattie, K.A. and Codd, G.A. (1992). Fatal canine neurotoxicosis attributed to blue-green algae (cyanobacteria). Vet. Rec., 130(14): 301-302.
- Hamilton, P.B., Ley, L.M., Dean, S. and Pick, F.R. (2005). The occurrence of the cyanobacterium Cylindrospermopsis raciborskii in Constance Lake: An exotic cyanoprokaryote new to Canada. Phycologia, 44(1): 17-25.
- Hamilton, D.P., Salmaso, N., and Paerl, H.W. (2016). Mitigating harmful cyanobacterial blooms: strategies for control of nitrogen and phosphorus loads. Aquat. Ecol., 50: 351-366.
- Hartshorn, N., Marimon, Z., Xuan, Z., Cormier, J., Chang, N. and Wanielista, M. (2016). Complex interactions among nutrients, chlorophyll-a, and microcystins in three stormwater wet detention basins with floating treatment wetlands. Chemosphere, 144: 408-419.
- Health Canada (unpublished). Canadian exposure factors used in human health risk assessments.
- Health Canada (in preparation). Guidelines for Canadian Recreational Water Quality – understanding and managing risk in recreational waters. Prepared by the Federal-Provincial-Territorial Working Group on Recreational Water Quality of the Federal-Provincial-Territorial Committee on Health and the Environment, Ottawa, ON.
- Health Canada (2017). Guidelines for Canadian drinking water quality - cyanobacterial toxins. Prepared by the Federal-Provincial-Territorial Committee on Drinking Water of the Federal-Provincial-Territorial Committee on Health and the Environment, Ottawa, ON.
- Heinze, R. (1999). Toxicity of the cyanobacterial toxin microcystin-LR to rats after 28 days intake with the drinking water. Environ. Toxicol., 14(1): 57-60.
- Hilborn, E.D., Roberts, V.A., Backer, L., Deconno, E., Egan, J.S., Hyde, J.B., Nicholas, D.C., Wiegert, E.J., Billing, L.M., Diorio, M., Mohr, M.C., Joan Hardy, F., Wade, T.J., Yoder, J.S. and Hlavsa, M.C. (2014). Algal bloom-associated disease outbreaks among users of freshwater lakes - United States, 2009-2010. Morb. Mortal. Wkly. Rep., 63(1): 11-15.
- Hodoki, Y., Ohbayashi, K., Kobayashi, Y., Okuda, N. and Nakano, S. (2012). Detection and identification of potentially toxic cyanobacteria: Ubiquitous distribution of Microcystis aeruginosa and Cuspidothrix issatschenkoi in Japanese lakes. Harmful Algae, 16: 49-57.
- Hoff, B., Thomson, G., and Graham, K. (2007) Ontario: Neurotoxic cyanobacterium (blue-green alga) toxicosis in Ontario. Can. Vet. J., 48(2): 147.
- Hollister, J.W. and Kreakie, B.J. (2016). Associations between chlorophyll a and various microcystin health advisory concentrations. F1000research, 5: 151.
- Holtcamp, W. (2012). The emerging science of BMAA: Do cyanobacteria contribute to neurodegenerative disease? Environ. Health Perspect., 120(3): a110-a116.
- Hong, Y., Steinman, A., Biddanda, B., Rediske, R. and Fahnenstiel, G. (2006). Occurrence of the toxin-producing cyanobacterium Cylindrospermopsis raciborskii in Mona and Muskegon Lakes, Michigan. Journal of Great Lakes Research, 32(3): 645-652.
- Hudon, C., Gagnon, P., Poirier Larabie, S., Gagnon, C., Lajeunesse, A., Lachapelle, M. and Quilliam, M.A. (2016). Spatial and temporal variations of a saxitoxin analogue (LWTX-1) in Lyngbya wollei (cyanobacteria) mats in the St. Lawrence River (Québec, Canada). Harmful Algae, 57(Part A): 69-77.
- Hudon, C., De Sève, M. and Cattaneo, A. (2014). Increasing occurrence of the benthic filamentous cyanobacterium Lyngbya wollei: A symptom of freshwater ecosystem degradation. Freshwater Science, 33(2): 606-618.
- Huisman, J., Codd, G.A., Paerl, H.W., Ibelings, B.W., Verspagen, J.M.H., and Visser, P.M. (2018). Cyanobacterial blooms. Nature Reviews – Microbiology, 16: 471- 483.
- Huot, Y., Babin, M., Bruyant, C., Grob, C., Twardowski, M.S. and Claustre, H. (2007). Does chlorophyll-a provide the best index of phytoplankton biomass for primary productivity studies? Biogeosciences, 4(2): 707-745.
- Huynh, M., and Serediak, N. (2006). Algae identification field guide. Agriculture and Agri-Food Canada. ISBN: 978-1-100-18309-1. Available at: http://publications.gc.ca/collections/collection_2011/agr/A125-8-2-2011-eng.pdf.
- IARC (2010). Ingested nitrate and nitrite and cyanobacterial peptide toxins. International Agency for Research on Cancer, IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, Vol. 94, Lyon, France.
- Ibelings, B.W., Backer, L.C., Kardinaal, W.E.A. and Chorus, I. (2014). Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae, 40: 63-74.
- Izydorczyk, K., Carpentier, C., Mrowczynski, J., Wagenvoort, A., Jurczak, T. and Tarczynska, M. (2009). Establishment of an alert level framework for cyanobacteria in drinking water resources by using the algae online analyser for monitoring cyanobacterial chlorophyll a. Water Res., 43(4): 989-996.
- Jacoby, J.M. and Kann, J. (2007). The occurrence and response to toxic cyanobacteria in the Pacific Northwest, North America. Lake Reserv. Manage., 23(2): 123-143.
- Jančula, D. and Maršálek, B. (2011). Critical review of actually available chemical compounds for prevention and management of cyanobacterial blooms. Chemosphere, 85(9): 1415-1422.
- Jones, G., Gurney, S. and Rocan, D. (1998). Water quality in farm and recreational surface water supplies of Southwestern Manitoba: 1995 sampling results. . Manitoba Environment, Report Number: 98-05, Winnipeg, MB.
- Jones, G.J. and Orr, P.T. (1994). Release and degradation of microcystin following algicide treatment of a Microcystis aeruginosa bloom in a recreational lake, as determined by HPLC and protein phosphatase inhibition assay. Water Res., 28(4): 871-876.
- Kasinak, J.E., Holt, B.M., Chislock, M.F. and Wilson, A.E. (2015). Benchtop fluorometry of phycocyanin as a rapid approach for estimating cyanobacterial biovolume. Journal of Plankton Research, 37(1): 248-257.
- Kemp, A. and John, J. (2006). Microcystins associated with Microcystis dominated blooms in the southwest wetlands, Western Australia. Environ. Toxicol., 21(2): 125-130.
- Kling, H. (2009). Cylindrospermopsis raciborskii (nostocales, cyanobacteria): A brief historic overview and recent discovery in the Assiniboine River (Canada). Fottea, 9(1): 45-47.
- Klitzke, S., and Fastner, J. (2012) Cylindrospermopsin degradation in sediments – the role of temperature, redox conditions, and dissolved organic carbon. Wat. Res., 46: 1549-1555.
- Kotak, B.G., Zurawell, R.W., Prepas, E.E. and Holmes, C.F. (1996). Microcystin-LR concentration in aquatic food web compartments from lakes of varying trophic status. Can. J. Fish. Aquat. Sci., 53(9): 1974-1985.
- Kotak, B.G. and Zurawell, R.W. (2007). Cyanobacterial toxins in Canadian freshwaters: A review. Lake Reserv. Manage., 23(2): 109-122.
- Krishnan, K. and Carrier, R. (2013). The use of exposure source allocation factor in the risk assessment of drinking-water contaminants. J. Toxicol. Environ. Health B Crit. Rev., 16(1): 39-51.
- Kurmayer, R., Christiansen, G., Fastner, J., and Börner, T. (2004). Abundance of active and inactive microcystin genotypes in populations of the toxic cyanobacterium Planktothrix spp. Environ. Microbiol., 6(8):831-41
- Kurmayer, R., Christiansen, G. and Chorus, I. (2003). The abundance of microcystin-producing genotypes correlates positively with colony size in Microcystis sp. and determines its microcystin net production in Lake Wannsee. Appl. Environ. Microbiol., 69(2): 787-795.
- Kurmayer, R. and Kutzenberger, T. (2003). Application of real-time PCR for quantification of microcystin genotypes in a population of the toxic cyanobacterium Microcystis sp. Appl. Environ. Microbiol., 69(11): 6723-6730.
- Kurmayer, R., Sivonen, K., Wilmotte, S., and Salmaso, N (Eds.) (2017) Molecular tools for the detection and quantification of toxigenic cyanobacteria. John Wiley and Sons Ltd., USA.
- Lam, A.K.Y., Prepas, E.E., Spink, D., and Hrudey, S.E. (1995). Chemical control of hepatotoxic phytoplankton blooms: implication for human health. Wat. Res., 29(8): 1845-1854.
- Lajeunesse, A., Segura, P.A., Gélinas, M., Hudon, C., Thomas, K., Quilliam, M.A. and Gagnon, C. (2012). Detection and confirmation of saxitoxin analogues in freshwater benthic Lyngbya wollei algae collected in the St. Lawrence River (Canada) by liquid chromatography–tandem mass spectrometry. Journal of Chromatography A, 1219: 93-103.
- Lévesque, B., Gervais, M., Chevalier, P., Gauvin, D., Anassour-Laouan-Sidi, E., Gingras, S., Fortin, N., Brisson, G., Greer, C. and Bird, D. (2014). Prospective study of acute health effects in relation to exposure to cyanobacteria. Science of the Total Environment, 466-467: 397-403.
- Lévesque, B., Gervais, M., Chevalier, P., Gauvin, D., Anassour-Laouan-Sidi, E., Gingras, S., Fortin, N., Brisson, G., Greer, C. and Bird, D. (2016). Exposure to cyanobacteria: acute health effects associated with endotoxins. Public Health, 134:98-101.
- Li, Z., Yu, J., Yang, M., Zhang, J., Burch, M.D. and Han, W. (2010). Cyanobacterial population and harmful metabolites dynamics during a bloom in Yanghe Reservoir, North China. Harmful Algae, 9(5): 481-488.
- Luesch, H., Yoshida, W.Y., Moore, R.E., Paul, V.J. and Corbett, T.H. (2001). Total structure determination of apratoxin A, a potent novel cytotoxin from the marine cyanobacterium Lyngbya majuscula. J. Am. Chem. Soc., 123(23): 5418-5423.
- Lyck, S. and Christoffersen, K. (2003). Microcystin quota, cell division and microcystin net production of precultured Microcystis aeruginosa CYA 228 (chroococcales, cyanophyceae) under field conditions. Phycologia, 42(6): 667-674.
- Lyon-Colbert, A., Su, S., and Cude, D. (2018). A systematic literature review for evidence of Aphanizomenon flos-aquae toxigenicity in recreational waters and toxicity of dietary supplements: 2000-2017. Toxins (Basel), 10(7):254.
- Macario, I.P.E., Castro, B.B., Nunes, M.I.S., Antunes, S.C., Pizarro, C., Coelho, C.M., Goncalves, F. and de Figueiredo, D.R. (2015). New insights towards the establishment of phycocyanin concentration thresholds considering species-specific variability of bloom-forming cyanobacteria. Hydrobiologia, 757: 155-165.
- Macbeth, A.J. (2004). Investigation of an introduced subtropical alga (Lyngbya wollei) in Whiteshell Provincial Park, Manitoba. MSc Thesis, University of Manitoba. Winnipeg, Manitoba.
- Martins, A. and Vasconcelos, V. (2011). Use of qPCR for the study of hepatotoxic cyanobacteria population dynamics. Arch. Microbiol., 193(9): 615-627.
- Matthijs, H.C., Visser, P.M., Reeze, B., Meeuse, J., Slot, P.C., Wijn, G., Talens, R. and Huisman, J. (2012). Selective suppression of harmful cyanobacteria in an entire lake with hydrogen peroxide. Water Res., 46(5): 1460-1472.
- Mazur-Marzec, H., Meriluoto, J., Plinski, M. and Szafranek, J. (2006). Characterization of nodularin variants in Nodularia spumigena from the Baltic Sea using liquid chromatography/mass spectrometry/mass spectrometry. Rapid Commun. Mass Spectrom., 20(13): 2023-2032.
- McQuaid, N., Zamyadi, A., Prevost, M., Bird, D.F. and Dorner, S. (2011). Use of in vivo phycocyanin fluorescence to monitor potential microcystin-producing cyanobacterial biovolume in a drinking water source. J. Environ. Monit., 13(2): 455-463.
- MDDEP (2005). Cyanobactéries et cyanotoxines au québec : Suivi à six stations de production d'eau potable (2001-2003). Ministère du Développement durable, de l'Environnement et des Parcs du Québec, Direction des politiques de l'eau, Direction générale des politiques, Envirodoq : ENV/2005/0099, Québec, Canada.
- MDDEP (2012). Personal communication with C. Robert.
- Meneely, J.P., Chevallier, O.P., Graham, S., Greer, B., Green, B.D. and Elliott, C.T. (2016). Beta-methylamino-L-alanine (BMAA) is not found in the brains of patients with confirmed Alzheimer's disease. Sci. Rep., 6(Article number: 36363): 1-9.
- Merel, S., Walker, D., Chicana, R., Snyder, S., Baurès, E. and Thomas, O. (2013). State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environment International, 59: 303-327.
- Meriluoto, J., Blaha, L., Bojadzija, G., Bormans, M., Brient, L., Codd, G.A., Drobac, D., Faassen, E.J., Fastner, J., Hiskia, A., Ibelings, B.W., Kaloudis, T., Kokocinski, M., Kurmayer, R., Pantelić, D., Quesada, A., Salmaso, N., Tokodi, N., Triantis, T.M., Visser, P.M., and Svirčev, Z. (2017). Toxic cyanobacteria and cyanotoxins in European waters – recent progress achieved through the CYANOCOST Action and challenges for further research. Advances in Oceanography and Limnology, 8(1): 161-178.
- Metcalf, J.S., Banack, S.A., Lindsay, J., Morrison, L.F., Cox, P.A. and Codd, G.A. (2008). Co-occurrence of beta-N-methylamino-L-alanine, a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies. Environmental Microbiology, 10(3): 702-708.
- Millie, D.F., Fahnenstiel, G.L., Dyble Bressie, J., Pigg, R.J., Rediske, R.R., Klarer, D.M., Tester, P.A. and Litaker, R.W. (2009). Late-summer phytoplankton in Western Lake Erie (Laurentian Great Lakes): Bloom distributions, toxicity, and environmental influences. Aquat. Ecol., 43(4): 915-934.
- Millie, D.F., Pigg, R.J., Fahnenstiel, G.L., and Carrick, H.J. (2010). Algal Chlorophylls: A synopsis of analytical methodologies. In: Algae Source to Treatment. AWWA Manual M57. American Water Works Association, Denver, CO., pp. 93-122.Murch, S.J., Cox, P.A., Banack, S.A., Steele, J.C. and Sacks, O.W. (2004). Occurrence of beta-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol. Scand., 110(4): 267-269.
- Negri, A.P., Jones, G.J. and Hindmarsh, M. (1995). Sheep mortality associated with paralytic shellfish poisons from the cyanobacterium Anabaena circinalis. Toxicon, 33(10): 1321-1329.
- New Zealand Ministry for the Environment and Ministry of Health, (2009). New Zealand guidelines for cyanobacteria in recreational fresh waters – interim guidelines. Prepared for the New Zealand Ministry for the Environment and the Ministry of Health by Wood, S.A., Hamilton, D.P., Paul, W. J., Safi, K. A., and Williamson, W. M., Wellington: Ministry for the Environment.
- Newcombe, G., (ed.) (2009). International guidance manual for the management of toxic cyanobacteria. Global Water Research Coalition, London, United Kingdom.
- Newcombe, G., House, J., Ho, L., Baker, P. and Burch, M. (2010). Management strategies for cyanobacteria (blue-green algae): A guide for water utilities. Water Quality Research Australia, Report Number 74, Adelaide, Australia.
- Ngwa, F.F., Madramootoo, C.A. and Jabaji, S. (2014). Comparison of cyanobacterial microcystin synthetase (mcy) E gene transcript levels, mcy E gene copies, and biomass as indicators of microcystin risk under laboratory and field conditions. Microbiologyopen, 3(4): 411-425.
- NHMRC (2008). Guidelines for managing risks in recreational water. National Health and Medical Research Council, Australian Government, Canberra.
- NHMRC (2011). Australian drinking water guidelines 6 - national water quality management strategy. National Health and Medical Research Council, National Resource Management Ministerial Council, Commonwealth of Australia, Canberra.
- Oh, H.M., Lee, S.J., Kim, J.H., Kim, H.S. and Yoon, B.D. (2001). Seasonal variation and indirect monitoring of microcystin concentrations in Daechung Reservoir, Korea. Appl. Environ. Microbiol., 67(4): 1484-1489.
- Ohkouchi, Y., Tajima, S., Nomura, M., and Itoh, S. (2015). Inflammatory responses and potencies of various lipopolysaccharides from bacteria and cyanobacteria in aquatic environments and water supply systems. Toxicon, 97: 23-31.
- Ontario Ministry of the Environment (2012). Personal communication with S. Deshpande.
- Orihel, D.M., Bird, D.F., Brylinsky, M., Chen, H., Donald, D.B., Huang, D.Y., Giani, A., Kinniburgh, D., Kling, H., Kotak, B.G., Leavitt, P.R., Nielsen, C.C., Reedyk, S., Rooney, R.C., Watson, S.B., Zurawell, R.W., and Vinebrooke, R.D. (2012). High microcystin concentrations occur only at low nitrogen-to-phosphorus ratios in nutrient-rich Canadian lakes. Can. J. Fish. Aquat. Sci., 69: 1457-1462.
- Osborne, N.J. and Shaw, G.R. (2008). Dermatitis associated with exposure to a marine cyanobacterium during recreational water exposure. BMC Dermatol., 8(5).
- Osborne, N.J., Webb, P.M. and Shaw, G.R. (2001). The toxins of Lyngbya majuscula and their human and ecological health effects. Environment International, 27(5): 381-392.
- Otten, T.G., and Paerl, H.W. (2015). Health effects of toxic cyanobacteria in U.S. drinking and recreational waters: our current understanding and proposed direction. Curr. Envir. Health Rpt, 2:75-84.
- Pablo, J., Banack, S.A., Cox, P.A., Johnson, T.E., Papapetropoulos, S., Bradley, W.G., Buck, A. and Mash, D.C. (2009). Cyanobacterial neurotoxin BMAA in ALS and Alzheimer's disease. Acta Neurol. Scand., 120(4): 216-225.
- Pace, M.L., Batt, R.D., Buelo, C.D., Carpenter, S.R., Cole, J.J., Kurtzweil, J.T. and Wilkinson, G.M. (2017). Reversal of a cyanobacterial bloom in response to early warnings. Proc. Natl. Acad. Sci. USA, 114(2): 352-357.
- Paerl, H.W., Xu, H., McCarthy, M.J., Zhu, G., Qin, B., Li, Y. and Gardner, W.S. (2011a). Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Research, 45(5): 1973-1983.
- Paerl, H.W., Hall, N.S. and Calandrino, E.S. (2011b). Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Science of the Total Environment, 409(10): 1739-1745.
- Paerl, H.W. (2017). Controlling cyanobacterial harmful blooms in freshwater ecosystems. Microbial Biotechnology, 10(5): 1106-1110.
- Pearson, L., Mihali, T., Moffitt, M., Kellmann, R. and Neilan, B. (2010). On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs, 8(5): 1650-1680.
- Phillips, G., Pietiläinen, O.P., Carvalho, L., Solimini, A., Lyche Solheim, A., and Cardoso, A.C. (2008). Chlorophyll-nutrient relationships of different lake types using a large European dataset. Aquat. Ecol., 42(2):213–226.
- Pilotto, L., Hobson, P., Burch, M.D., Ranmuthugala, G., Attewell, R. and Weightman, W. (2004). Acute skin irritant effects of cyanobacteria (blue-green algae) in healthy volunteers. Aust. New Zealand J. Public Health, 28(3): 220-224.
- Pilotto, L.S., Douglas, R.M., Burch, M.D., Cameron, S., Beers, M., Rouch, G.J., Robinson, P., Kirk, M., Cowie, C.T., Hardiman, S., Moore, C. and Attewell, R.G. (1997). Health effects of exposure to cyanobacteria (blue-green algae) during recreational water-related activities. Aust. N. Z. J. Public Health, 21(6): 562-566.
- Puschner, B., Hoff, B. and Tor, E. (2008). Diagnosis of anatoxin-a poisoning in dogs from North America. Journal of Veterinary Diagnostic Investigation, 20: 89-92.
- Quiblier, C., Wood, S., Echenique-Subiabre, I., Heath, M., Villeneuve, A. and Humbert, J.F. (2013). A review of current knowledge on toxic benthic freshwater cyanobacteria – ecology, toxin production and risk management. Water Research, 47(15): 5464-5479.
- Rinta-Kanto, J.M., Ouellette, A.J., Boyer, G.L., Twiss, M.R., Bridgeman, T.B. and Wilhelm, S.W. (2005). Quantification of toxic Microcystis spp. during the 2003 and 2004 blooms in Western Lake Erie using quantitative real-time PCR. Environ. Sci. Technol., 39(11): 4198-4205.
- Rogers, E.H., Hunter, E.S., Moser, V.C., Phillips, P.M., Herkovits, J., Muñoz, L., Hall, L.L. and Chernoff, N. (2005). Potential developmental toxicity of anatoxin-a, a cyanobacterial toxin. Journal of Applied Toxicology, 25(6): 527-534.
- Rosen, B.H., and St. Amand, A. (2015). Field and laboratory guide to freshwater cyanobacteria harmful algal blooms for Native American and Alaska Native Communities. U.S. Geological Survey Open-File Report 2015-1164.
- Roy-Lachapelle, A., Solliec, M. and Sauve, S. (2015). Determination of BMAA and three alkaloid cyanotoxins in lake water using dansyl chloride derivatization and high-resolution mass spectrometry. Anal. Bioanal Chem., 407(18): 5487-5501.
- Sabart, M., Pobel, D., Briand, E., Combourieu, B., Salencon, M.J., Humbert, J.F. and Latour, D. (2010). Spatiotemporal variations in microcystin concentrations and in the proportions of microcystin-producing cells in several Microcystis aeruginosa populations. Appl. Environ. Microbiol., 76(14): 4750-4759.
- Saccà, A. (2016). A simple yet accurate method for the estimation of the biovolume of planktonic microorganisms. Plos One, 11(5): e0151955.
- Schets, F.M., Schijven, J.F. and de Roda Husman, A.M. (2011). Exposure assessment for swimmers in bathing waters and swimming pools. Water Res., 45(7): 2392-2400.
- Schwimmer, M. and Schwimmer, D. (1968). Medical aspects of phycology. In: Algae, man, and the environment. Jackson, D.F. (ed.). Syracuse University Press, New York, N.Y., pp. 279-358.
- Sinha, R., Pearson, L.A., Davis, T.W., Burford, M.A., Orr, P.T. and Neilan, B.A. (2012). Increased incidence of Cylindrospermopsis raciborskii in temperate zones – is climate change responsible? Water Research, 46(5): 1408-1419.
- Smith, M.J., Shaw, G.R., Eaglesham, G.K., Ho, L., and Brookes, J.D. (2008). Elucidating the factors influencing the biodegradation of cylindrospermopsin in drinking water sources. Environmental Toxicology, DOI 10.1002/tox
- Søndergaard, M., Larsen, S.E., Jørgensen, T.B. and Jeppesen, E. (2011). Using chlorophyll a and cyanobacteria in the ecological classification of lakes. Ecological Indicators, 11(5): 1403-1412.
- Spoof, L., and Arnaud, C. (2017). Appendix 3. Tables of microcystins and nodularins. In: Meriluoto, J., Spoof, L., and Codd, G.A. (Eds). Handbook of cyanobacterial monitoring and cyanotoxin analysis. Wiley, European Cooperation in Science and Technology, Cyanocost, United Kingdom.
- Srivastava, A., Choi, G., Ahn, C., Oh, H., Ravi, A.K. and Asthana, R.K. (2012). Dynamics of microcystin production and quantification of potentially toxigenic Microcystis sp. using real-time PCR. Water Research, 46(3): 817-827.
- Stevens, D.K. and Krieger, R.I. (1991). Stability studies on the cyanobacterial nicotinic alkaloid anatoxin-a. Toxicon, 29: 167–179.
- Stewart, I. (2004). Recreational exposure to freshwater cyanobacteria: Epidemiology, dermal toxicity and biological activity of cyanobacterial lipopolysaccharides. Doctor of Philosophy. University of Queensland, Australia.
- Stewart, I., Seawright, A.A. and Shaw, G.R. (2008). Cyanobacterial poisoning in livestock, wild mammals and birds – an overview. In: Cyanobacterial harmful algal blooms: State of the science and research needs. . Hudnell, H.K. (ed.). Springer, New York, NY, pp. 613-637.
- Stewart, I., Webb, P.M., Schluter, P.J., Fleming, L.E., Burns, J.W., Gantar, M., Backer, L.C. and Shaw, G.R. (2006). Epidemiology of recreational exposure to freshwater cyanobacteria - an international prospective cohort study. BMC Public Health, 6(Article Number: 93): 1-11.
- Suppes, L.M., Abrell, L., Dufour, A.P. and Reynolds, K.A. (2014). Assessment of swimmer behaviors on pool water ingestion. J. Water. Health., 12(2): 269-279.
- Teneva, I., Asparuhova, D., Dzhambazov, B., Mladenov, R. and Schirmer, K. (2003). The freshwater cyanobacterium Lyngbya aerugineo-coerulea produces compounds toxic to mice and to mammalian and fish cells. Environ. Toxicol., 18(1): 9-20.
- Tillett, D., Dittmann, E., Erhard, M., von Döhren, H., Börner, T. and Neilan, B.A. (2000). Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: an integrated peptide–polyketide synthetase system. Chemistry & Biology, 7(10): 753-764.
- U.S. EPA (2011) Exposure Factors Handbook: 2011 Edition (Final). Washington, DC; EPA/600/R-09/052F. Available at https://www.epa.gov/expobox/about-exposure-factors-handbook.
- U.S. EPA (2015). Drinking water health advisory for the cyanobacterial toxin cylindrospermopsin. United States Environmental Protection Agency, Office of Water, Document Number: EPA-820-R-15-101
- U.S. EPA (2017). Recommendations for Cyanobacteria and cyanotoxin monitoring in recreational waters. United States Environmental Protection Agency, Office of Water, Document Number: EPA 820-R-17-001.
- U.S. EPA (2019) Recommended Human Health Recreational Ambient Water Quality Criteria or Swimming Advisories for Microcystins and Cylindrospermopsin. Washington, DC. EPA/822/R-19/001. May 2019.
- Vidal, F., Sedan, D., D'Agostino, D., Cavalieri, M.L., Mullen, E., Parot Varela, M.M., Flores, C., Caixach, J. and Andrinolo, D. (2017). Recreational exposure during algal bloom in Carrasco Beach, Uruguay: A liver failure case report. Toxins (Basel), 9(Article Number: 267): 1-12.
- Vijayavel, K., Sadowsky, M.J., Ferguson, J.A. and Kashian, D.R. (2013). The establishment of the nuisance cyanobacteria Lyngbya wollei in Lake St. Clair and its potential to harbor fecal indicator bacteria. Journal of Great Lakes Research, 39(4): 560-568.
- Vis, C., Cattaneo, A. and Hudon, C. (2008). Shift from chlorophytes to cyanobacteria in benthic macroalgae along a gradient of nitrate depletion. J. Phycol., 44(1): 38-44.
- WHO (2003). Guidelines for safe recreational water environments, Volume 1: Coastal and fresh waters. World Health Organization, Geneva, Switzerland.
- Wiese, M., D'Agostino, P.,M., Mihali, T.K., Moffitt, M.C. and Neilan, B.A. (2010). Neurotoxic alkaloids: Saxitoxin and its analogs. Marine Drugs, 8(7): 2185-2211.
- Williams, C.D., Aubel, M.T., Chapman, A.D. and D'Aiuto, P.E. (2007). Identification of cyanobacterial toxins in Florida's freshwater systems. Lake Reserv. Manage., 23(2): 144-152.
- Winter, J.G., DeSellas, A.M., Fletcher, R., Heintsch, L., Morley, A., Nakamoto, L. and Utsumi, K. (2011). Algal blooms in Ontario, Canada: Increases in reports since 1994. Lake Reserv. Manage., 27(2): 107-114.
- Wood, S.A., Paul, W.J. and Hamilton, D.P. (2008). Cyanobacterial biovolumes for the Rotorua Lakes. Cawthorn Institute, Prepared for Environment Bay of Plenty, 1504, New Zealand.
- Wood, S.A., Kuhajek, J.M., de Winton, M. and Phillips, N.R. (2012). Species composition and cyanotoxin production in periphyton mats from three lakes of varying trophic status. FEMS Microbiol. Ecol., 79(2): 312-326.
- Wood, S.A., Hawes, I., McBride, G., Truman, P., and Dietrich, D. (2015). Advice to inform the development of a benthic cyanobacteria attribute. Prepared for New Zealand Ministry for the Environment. Cawthron Report No. 2752. 91 p.
- Wood, S.A., and Puddick J. (2017). The abundance of toxic genotypes is a key contributor to anatoxin variability in Phormidium-dominated benthic mats. Marine Drugs, 15 (307): 1-11.
- Wörmer, L. et al. (2008). Cylindrospermopsin is not degraded by co-occurring natural bacterial communities during a 40-day study. Harmful Algae, 7: 206–213.
- Wörmer, L. et al. (2009). Advances in solid phase extraction of the cyanobacterial toxin cylindrospermopsin. Limnol. Oceanogr. Methods, 7: 568–575.
- Xie, L., Hagar, J., Rediske, R.R., O'Keefe, J., Dyble, J., Hong, Y. and Steinman, A.D. (2011). The influence of environmental conditions and hydrologic connectivity on cyanobacteria assemblages in two drowned river mouth lakes. Journal of Great Lakes Research, 37(3): 470-479.
- Yilmaz, M. and Phlips, E.J. (2011). Diversity of and selection acting on cylindrospermopsin cyrB gene adenylation domain sequences in Florida. Appl. Environ. Microbiol., 77(7): 2502-2507.
- Yoshida, M., Yoshida, T., Satomi, M., Takashima, Y., Hosoda, N., and Hiroishi, S. (2008). Intra-specific phenotypic and genotypic variation in toxic cyanobacterial Microcystis strains. Journal of Applied Microbiology, 105:407-415.
- Yuan, L.A., Pollard, A.I., Pather, S., Oliver, J.L. and D'Anglada, L. (2014). Managing microcystin: Identifying national-scale thresholds for total nitrogen and chlorophyll a. Freshwater Biology, 59: 1970-1981.
- Zamyadi, A., MacLeod, S.L., Fan, Y., McQuaid, N., Dorner, S., Sauvé, S., and Prévost, M. (2012a). Toxic cyanobacterial breakthrough and accumulation in a drinking water plant: a monitoring and treatment challenge. Water Research, 46: 1511-1523.
- Zamyadi, A., McQuaid, N., Prévost, M., and Dorner, S. (2012b). Monitoring of potentially toxic cyanobacteria using an online multi-probe in drinking water sources. J. Environ Monit, 14:579-588.
- Zastepa, A., Watson, S.B., Kling, H. and Kotak, B. (2017). Spatial and temporal patterns in microcystin toxins in Lake of the Woods surface waters. Lake Reserv. Manage., 33(4): 433-443.
- Zohary, T., Shneor, M. and Hambright, K.D. (2016). PlanktoMetrix – a computerized system to support microscope counts and measurements of plankton. Inland Waters, 6(2): 131-135.
Appendix A: List of acronyms
- ALS
- amyotrophic lateral sclerosis
- ALS-PDC
- amyotrophic lateral sclerosis/parkinsonism–dementia complex
- BMAA
- β-methylamino-L-alanine
- bw
- body weight
- CI
- confidence interval
- DNA
- deoxyribonucleic acid
- ELISA
- enzyme-linked immunosorbent assay
- HABs
- hazardous algal blooms
- HBV
- health-based value
- HPLC
- high-performance liquid chromatography
- LC
- liquid chromatography
- LOAEL
- lowest-observed-adverse-effect level
- MC
- microcystin variant (e.g., MC-LR, MC-LA, MC-YA, MC-RR, MC-YR)
- MS
- mass spectrometry
- MS/MS
- tandem mass spectrometry
- NOAEL
- no-observed-adverse-effect level
- PCR
- polymerase chain reaction
- PDA
- photodiode array
- PPIA
- protein phosphatase inhibition assay
- PSP
- paralytic shellfish poisoning
- qPCR
- quantitative polymerase chain reaction
- rRNA
- ribosomal ribonucleic acid
- TDI
- tolerable daily intake
- TEQ
- toxic equivalency
- UF
- uncertainty factor
- UHPLC
- ultra-high-performance liquid chromatography
- U.S. EPA
- United States Environmental Protection Agency
- UV
- ultraviolet
Appendix B: Flow chart for monitoring cyanobacteria and their toxins
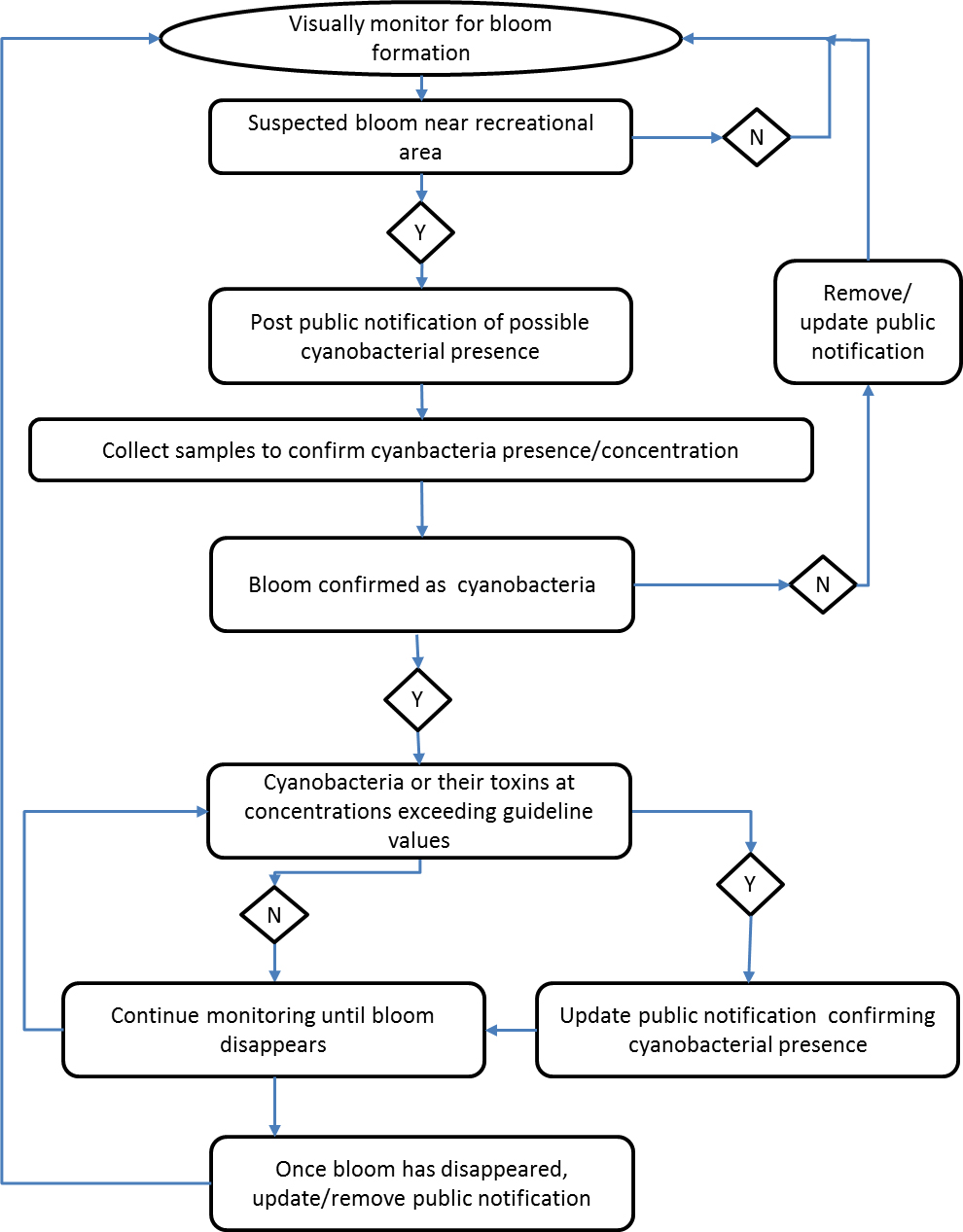
Text description
The process for monitoring for planktonic cyanobacteria and their toxins is presented in a flow chart. The flow chart is made up of a series of boxes connected by unidirectional arrows that flow to subsequent boxes. Some boxes contain questions, while other boxes contain recommendations or sampling information. The boxes that contain questions connect to two boxes, and the path taken depends on whether the answer to the question is yes or no.
The flow chart begins by advising the user to visually monitor for bloom formation. If no bloom is suspected, the flowchart loops back and visual monitoring for blooms continues. If a suspected bloom is detected near a recreational area, the “yes” path is taken and a public notification is posted of the possible presence of cyanobacteria. The next step in the flow chart is to collect samples to confirm cyanobacteria presence/concentration. If the bloom is not cyanobacteria, the public notification should be removed or updated and visual monitoring for bloom formation should continue. If the bloom is confirmed as cyanobacteria, the “yes” path is followed. This paths requires the determination of whether cyanobacteria or their toxins are at concentrations exceeding the guideline values. If the answer is no, then monitoring should continue until the bloom disappears. If the answer is yes, the public notification should be updated to confirm the presence of cyanobacteria. The next step is to then continue monitoring until the bloom disappears. Once the bloom has disappeared, the last step in the flow chart is to update or remove the public notification and then to start back at the beginning of the flow chart with visual monitoring.