
Surveillance nationale en laboratoire maladie invasive due au streptocoque au Canada, Rapport sommaire annuel de 2013

Tableau des matières
Figures et tableaux
- Figure 1. Incidence des PI au Canada, 2000-2013
- Figure 2, Tableau 1. Taux d'incidence des PI pour 100 000 au Canada, 2009-2013
- Tableau 2. Nombre d'isolats de S. pneumoniae causant une infection invasive, dans chaque province et territoire, 2013
- Figure 3. Sites d'isolement clinique de S. pneumoniae, 2013
- Figure 4 à 7. Distribution des sérotypes de S. pneumoniae causant une infection invasive par source clinique, 2013
- Figure 8 à 14. Sérotypes de S. pneumoniae causant une infection invasive selon l'âge, 2010 2013
- Figure 15 à 18. Sérotypes 3, 7F, 19A et 22F par année et par âge
- Figure 19 à 25. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive, selon l'âge, 2013
- Figure 26 à 29. Sérotypes 3, 7F, 19A et 22F par région et par groupe d'âge
- Tableau 3. Sérotypes vaccinaux, 2013
- Figure 30. Sérotypes vaccinaux par région, 2013
- Figure 31. Sérotypes vaccinaux, 2010-2013
- Figure 32, Tableau 4. Sérotypes (4, 6B, 9V, 14, 18C, 19F, 23F) du vaccin Pneu-C-7 par groupe d'âge, 2010-2013
- Figure 33, Tableau 5. Sérotypes (1, 5, 7F, 3, 6A, 19A) du vaccin Pneu-C-13 par groupe d'âge, 2010-2013
- Figure 34, Tableau 6. Sérotypes (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20, 22F, 33F) du vaccin Pneu-P-23 par groupe d'âge, 2010-2013
- Figure 35, Tableau 7. Sérotypes non inclus dans les vaccins par groupe d'âge, 2010-2013
- Figure 36. Résistance des isolats de S. pneumoniae aux antimicrobiens, 2011-2013
- Figure 37. Résistance de S. pneumoniae aux antimicrobiens par région, 2013
- Figure 38. Profils de multirésistance de S. pneumoniae, 2011-2013
- Figure 39. Profils de multirésistance des sérotypes de S. pneumoniae, 2013
- Tableau 8. Résistance des sérotypes de S. pneumoniae aux antimicrobiens, 2013
- Figure 40 à 47. Profils de résistance des sérotypes de S. pneumoniae par antibiotique, 2011-2013
- Tableau 9. Nombre d'isolats de S. pyogenes causant une infection invasive dans chaque province, 2013
- Figure 48 à 49. Sites d'isolement clinique de S. pyogenes chez les enfants et chez les adultes
- Figure 50 à 51. Types emm de S. pyogenes causant une infection invasive chez les enfants et chez les adultes
- Figure 52. Distribution régionale des types emm de S. pyogenes
- Figure 53abc. Types emm de S. pyogenes causant une infection invasive par source d'isolement clinique, 2013
- Figure 54, Tableau 10. Résistance de S. pyogenes causant une infection invasive aux antimicrobiens, 2010 - 2013
- Tableau 11. Types emm de S. pyogenes non sensibles au chloramphénicol, 2010-2013
- Figure 55. Types emm de S. pyogenes résistants au macrolide, 2013
- Tableau 12. Sérotypes de S. agalactiae causant une infection invasive, pour chaque groupe d'âge, 2013
- Figure 56 à 57, Tableau 13. Sites d'isolement clinique de S. agalactiae
- Figure 58. Distribution régionale des sérotypes de S. agalactiae causant une infection invasive, 2013
- Figure 59, Tableau 14. Résistance aux antimicrobiens des isolats de S. agalactiae causant une infection invasive, 2010-2013
- Tableau 15. Sérotypes de S. agalactiae non sensibles au chloramphénicol, 2010-2013
- Figure 60. Résistance des sérotypes de S. agalactiae au macrolide, 2013
- Annexe
- Tableau A. Proportion de cas sérotypés de Streptococcus pneumoniae causant une infection invasive au Canada, 2013
- Tableau B. Proportion de cas de Streptococcus pyogenes causant une infection invasive au Canada, 2013
Remerciements
Auteurs
Unité des streptocoques et des ITS
Programme de bactériologie et de maladies entériques
Laboratoire national de microbiologie
Agence de la santé publique du Canada
Walter H.B. Demczuk, Averil Griffith, Ravinder Singh, Irene Martin (chef de section)
Dr Michael Mulvey (chef, Résistance aux antimicrobiens et infections nosocomiales)
Autres collaborateurs :
Section des maladies évitables par les vaccins
Division de la surveillance et de l’intervention en cas d’épidémie
Centre de l’immunisation et des maladies respiratoires infectieuses (CIMRI)
Agence de la santé publique du Canada
Lindsey Sherrard et Susan Squires
Laboratoire de santé publique du Québec (LSPQ)
Dre Brigitte Lefebvre, Dr Marc-Christian Domingo et Dre Cécile Tremblay
Toronto Invasive Bacterial Diseases Network (TIBDN)
Département de microbiologie, Mount Sinai Hospital
Dre Allison McGeer, Agron Plevneshi, Sylvia Pong-Porter, Karen Green, Kenny Wong et enquêteurs et personnel sur place
The Alberta Provincial Laboratory for Public Health (ProvLab Alberta)
Dr Gregory J. Tyrrell et Dr Graham Tipples
Département de microbiologique médicale et des maladies infectieuses, faculté de médecine, Université du Manitoba
Dre Heather Adam, Dr Matthew Gilmour, Melanie Baxter, Kim Nichol, Barbara Weshnoweski, Ravi Vashisht et Dr George Zhanel
Le présent rapport a été soumis à l’examen des directeurs de laboratoires provinciaux et territoriaux du Réseau des laboratoires de santé publique du Canada (RLSPC).
Remerciements
Les résultats présentés dans ce rapport ont été obtenus grâce aux isolats de streptocoques ayant été aimablement acheminés au LNM par les hôpitaux ou les laboratoires provinciaux de santé publique suivants : Centre de contrôle des maladies de la Colombie-Britannique, Vancouver, Colombie-Britannique – Dre Judy Issac-Renton, Dre Linda Hoang, Ana Paccagnella, Loretta Janz, Sultana Mithan et Tazim Rahim; Saskatchewan Disease Control Laboratory, Regina, Saskatchewan – Dr Greg Horsman et Rosanne Kitzul; Laboratoire provincial de Cadham, Winnipeg, Manitoba – Dr Jared Bullard, Dr Paul Van Caeseele et Denise Sitter; Santé Publique Ontario, Toronto, Ontario – Dr. Frances Jamieson, Dr. Jonathan Gubbay, Deirdre Soares, Dr. Julianne Kus; Queen Elizabeth II Health Sciences Centre, Halifax, Nouvelle-Écosse – Dr David Haldane; Hôpitaux régionaux du Nouveau-Brunswick – Dr Alexander Doroshenko, Maryse Thibeault, Éric Brisson, Francine Plourde, Dr L. Thibault, Dr Lewis Abbott, Melanie Stace, Peter Delaney, Linda Turgeon et Tammy Mahaney; Hôpital Queen Elizabeth, Charlottetown, Île-du-Prince-Édouard – Dr G. German; Laboratoire de santé publique de Terre-Neuve, St. John’s, Terre-Neuve – Dr S. Ratnam; Stanton Territorial Hospital Laboratory, Yellowknife, Territoires du Nord-Ouest – Cheryl Case; Hôpital général de Whitehorse, Whitehorse, Yukon – Rosalyn Robertson, Becky Nash et Chris Cash.
Sommaire
- En 2013, 2 577 isolats de Streptococcus pneumoniae causant une pneumococcie invasive (PI) ont été caractérisés.
- L’incidence globale des PI, tous groupes d’âges combinés, est demeuré relativement stable, à environ 9,6 cas pour 100 000 personnes (variation : 9,0 à 9,8) de 2009 à 2013. Le taux d’incidence des PI a décliné chez les enfants de moins de 5 ans. Toutefois, les taux d’incidence chez les groupes plus âgés sont demeurés relativement inchangés.
- En 2013, les taux d’incidence les plus élevés ont été observés chez les adultes de 60 ans et plus (21,7 cas pour 100 000), les nourrissons d’un an et moins (17,8 cas pour 100 000) et les enfants de 1 à 4 ans (11,2 cas pour 100 000).
- La proportion des sérotypes contenus dans le vaccin Pneu-C-7 (4, 6B, 9V, 14, 18C, 19F, 23F) a décliné, passant de 9,5 % de tous les isolats de PI en 2010 à 5,0 % en 2013.
- La proportion des sérotypes contenus dans le vaccin Pneu-C-13 (1, 5, 3, 6A, 7F, 19A) a diminué pour tous les âges, présentant une baisse générale de 45,5 % en 2010 à 30,4 % en 2013.
- La proportion des sérotypes contenus dans le vaccin Pneu-P-23 (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20, 22F, 33F) a augmenté dans tous les groupes d’âge, soit une hausse générale de 24,7 % (n = 670) à 37,6 % (n = 970) entre 2010 et 2013.
- Le sérotype 22F était le sérotype le plus fréquemment observé en 2013; son taux est passé de 6,7 % en 2010 à 12,1 % en 2013, dans tous les groupes d’âge.
- Le taux du sérotype 19A a continué de décliner depuis 2010, passant de 19,0 % à 11,6 % en 2013. Par rapport à 2012, on a observé une réduction des taux chez les enfants de < 2 ans (de 20,7 % à 11,6 %) et chez les enfants de 2 à 4 ans (de 23,6 % à 18,7 %).
- Le taux du sérotype 7F a connu une baisse similaire, passant de 14,3 % en 2010 à 8,8 % en 2013, et aucun n’a pas été signalé chez les enfants de < 2 ans au cours de 2013.
- Le taux du sérotype 3 a connu une légère augmentation globale depuis 2010, passant de 8,3 % à 8,9 % des isolats de 2013; depuis 2012, il a diminué chez les enfants de < 2 ans, passant de 8,6 % à 5,4 %, et augmenté chez les enfants de 5 à 14 ans, pour passer de 2,9 % à 5,1 %.
- Selon un test de sensibilité aux antimicrobiens réalisé sur 1 061 isolats, 24,8 % étaient résistants à la clarithromycine, 11,9 % à la pénicilline, 9,9 % à la doxycycline, 5,9 % à la clindamycine, 7,4 % au triméthoprime-sulfaméthoxazole, 2,6 % au méropénème et 2,3 % à l’imipénème. Aucune résistance au linézolide, à la tigécycline ou à la vancomycine n'a été observée.
- Des augmentations de la résistance aux antimicrobiens ont été observées avec la clarithromycine, le triméthoprime-sulfaméthoxazole, l’imipénème et le méropénème.
- On a observé une multirésistance à 3 classes ou plus d’antimicrobiens dans 7,5 % des isolats testés, les taux les plus élevés étant associés aux sérotypes 15A (50,0 %) et 19A (28,9 %).
- Sur les 1 294 isolats analysés en 2013 de S. pyogenes (streptocoques du groupe A) causant une infection invasive, le type de gène emm1 est encore le plus prédominant, c’est-à-dire qu’on le retrouve dans 28,4 % des isolats chez les enfants de < 15 ans et dans 23,7 % des isolats chez les personnes de ≥ 15 ans. Le type emm89 était le deuxième en importance, avec 7,4 % des isolats chez les enfants et 9,9 % chez les adultes.
- Sur les 1 294 isolats de S. pyogenes analysés en 2013, 0,6 % étaient non sensibles au chloramphénicol, 8,5 % à l’érythromycine et 2,3 % à la clindamycine. Aucune résistance à la pénicilline ou à la vancomycine n’a été observée.
- 416 isolats de S. agalactiae (streptocoque du groupe B) causant une infection invasive ont été soumis au LNM durant l’année 2013. Les sérotypes III (23,8 %), V (19,7 %) et Ia (17,8 %) étaient les plus prédominants.
- Des augmentations de la résistance à l’érythromycine (48,8 %) et à la clarithromycine (28,8 %) ont été observées en 2013 pour les streptocoques du groupe B.
Introduction
Depuis le 1er avril 2010, le Laboratoire national de microbiologie (LNM), à Winnipeg, offre des services de surveillance, de diagnostic de référence et de soutien en cas d’éclosion d’infections invasives à Streptococcus pneumoniae (pneumocoque), à Streptococcus pyogenes (streptocoque du groupe A) et à Streptococcus agalactiae (streptocoque du groupe B). L'Unité des streptocoques et des ITS participe également à un certain nombre de programmes de surveillance internationaux, nationaux et régionaux.
Le présent rapport vise à tracer un tableau de la distribution actuelle des sérotypes de S. pneumoniae, des types emm de S. pyogenes et des sérotypes de S. agalactiae isolés de sites stériles, qui sont acheminés au LNM par les laboratoires de santé publique, les unités sanitaires régionales et les centres de référence des provinces et territoires du Canada. Pour rendre les données encore plus représentatives, les dénombrements regroupés incluent également les données soumises par le Laboratoire de santé publique du Québec (LSPQ), le Toronto Invasive Bacterial Diseases Network (TIBDN) et l’Alberta Provincial Laboratory for Public Health (ProvLab Alberta), des organismes qui ont leurs propres activités de sérotypage.
La pneumococcie invasive (PI, S. pneumoniae) cause des infections graves, comme la méningite et la bactériémie [Marchessault, 2002; Schuchat, 1997], les enfants et les personnes âgées étant les groupes les plus exposés à l’infection [Robinson, 2001; Scott, 1996]. Bien qu’il existe 92 sérotypes de pneumocoque différents actuellement reconnus, la majorité des cas observés sur la planète sont attribuables à quelques sérotypes seulement.
Un vaccin conjugué heptavalent contre le pneumocoque (Pneu-C-7) composé des sérotypes 4, 6B, 9V, 14, 18C, 19F et 23F a été adopté par tous les programmes de vaccination provinciaux et territoriaux entre 2002 et 2006 [Bettinger, 2010], ce qui a mené, chez les enfants, à une baisse radicale de l’incidence des infections invasives causées par les sérotypes contenus dans le vaccin [Bettinger, 2010; Bjornson, 2007; Bruce, 2008; Demczuk, 2012; Deng, 2013; DeWals, 2012; Kellner, 2008; Kellner, 2009; Lim, 2013; Lovgren, 1998; McIntosh, 2011; NACI, 2010; Shahidi, 2008; Tyrrell, 2009; Weinberger, 2011] (figure 1). Depuis le lancement des programmes de vaccination, on a observé, chez les enfants, une hausse de l’incidence des PI causées par une substitution des sérotypes à l’origine d’infections pneumococciques, avec pour conséquence une augmentation des infections par des sérotypes non contenus dans le vaccin Pneu-C-7, comme les sérotypes 7F et 19A [Kellner, 2009; Tyrrell, 2009]. En 2009, un vaccin décavalent contre le pneumocoque, Haemophilus influenzae et la diphtérie (vaccin PHiD10), constitué de tous les sérotypes du vaccin Pneu-C-7 et des sérotypes 1, 5 et 7F, a été utilisé au Québec, en Ontario et à Terre-Neuve-et-Labrador. En 2010, on a recommandé l’utilisation au Canada [Comité consultatif national de l’immunisation (CCNI), 2010] du vaccin conjugué 13-valent contre le pneumocoque (Pneu-C-13), qui comprend tous les sérotypes du vaccin PHiD10 et les sérotypes 3, 6A et 19A; toutes les provinces et territoires ont commencé à l’utiliser entre la mi-2010 et la mi-2011. Les calendriers d’immunisation varient selon la province et le territoire; toutefois, le CCNI et l’Agence de la santé publique du Canada (ASPC) ont publié des recommandations [CCNI, 2010; ASPC, 2013]. Il existe un vaccin pour les adultes, le vaccin polysaccharidique 23-valent contre le pneumocoque (Pneu-P-23), qui n’est pas efficace chez les enfants en raison de la faible réponse anticorps thymo-indépendante caractéristique des systèmes immunitaires non parvenus à maturité [Merck & Co. Inc.]. Il est important de surveiller la distribution des sérotypes de S. pneumoniae pour guider le choix de sérotypes devant composer le vaccin et pour surveiller un éventuel remplacement des sérotypes prédominants [Demczuk, 2010, Demczuk 2013].
Figure 1. Incidence des PI au Canada, 2000-2013
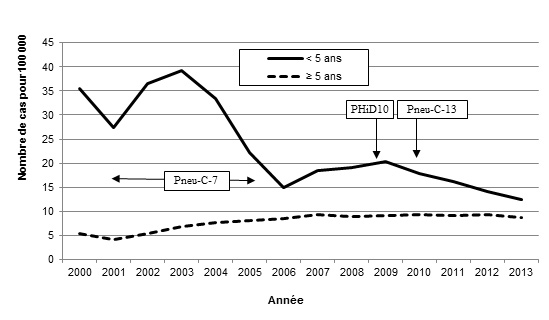
Incidence des PI au Canada, 2000-2013 - Description textuel
Ce figure représente le nombre de cas pour 100 000 des pneumococcies invasives distribués entre 2000 et 2013 pour les <5 ans et ≥5 ans.
| Année | Nombre de cas pour 100 000 | |
|---|---|---|
| <5 ans | ≥5 ans | |
| 2000 | 35.59 | 5.36 |
| 2001 | 27.47 | 4.25 |
| 2002 | 36.54 | 5.46 |
| 2003 | 39.22 | 6.85 |
| 2004 | 33.33 | 7.74 |
| 2005 | 22.28 | 8.11 |
| 2006 | 14.99 | 8.48 |
| 2007 | 18.41 | 9.36 |
| 2008 | 19.10 | 9.03 |
| 2009 | 20.33 | 9.14 |
| 2010 | 17.95 | 9.28 |
| 2011 | 16.27 | 9.11 |
| 2012 | 14.15 | 9.44 |
| 2013 | 12.44 | 8.76 |
L’infection invasive à streptocoque du groupe A (iSGA, S. pyogenes) est une maladie à déclaration obligatoire à l’échelle nationale. Cette infection est responsable d’un vaste éventail de maladies, dont la bactériémie, le syndrome de choc toxique et des infections de la peau et des tissus mous dont la fasciite nécrosante est la plus notoire [Cunningham, 2000]. Il est important de surveiller les souches pour repérer toute hausse des profils de virulence associés à ce microorganisme [Schwartz, 1990; Siljander, 2010]. La protéine M, codée par le gène emm, est un important facteur de virulence et un marqueur épidémiologique servant à caractériser les isolats de S. pyogenes.
Le streptocoque du groupe B Streptococcus (SGB, S. agalactiae) est généralement associé aux maladies néonatales pour lesquelles l’accouchement présente le risque d’infection le plus élevé, et qui sont souvent traitées par des antibiotiques à des fins prophylactiques. La maladie associée au nourrisson est une maladie à déclaration obligatoire. Toutefois, le SGB constitue également une préoccupation de santé croissante chez les adultes, car il cause la septicémie, la méningite, la pneumonie, ainsi que des infections des os, des articulations et des tissus. Les groupes d’adultes à risque englobent les personnes atteintes d’une affection sous-jacente, les femmes enceintes et les personnes qui résident dans des établissements de soins de longue durée [Lamangni, 2013].
Méthodes
Ce rapport porte sur un total de 2 577 isolats deS. pneumoniaecausant une infection invasive, de 1 294 isolats de S. pyogenes causant une infection invasive et de 420 isolats de S. agalactiae prélevés en 2013. Les données de 2013 englobent les résultats d’analyses des isolats soumis au LNM par les laboratoires de santé publique des provinces et des territoires et les données fournies par les provinces et les territoires notamment 529 isolats de PI sérotypés par le LSPQ, 342 isolats provenant du ProvLab Alberta et 310 du TIBDN.
Les données accompagnant les isolats bactériens comprenaient l’âge du patient, le sexe, la source clinique et la date du prélèvement. Les isolats multiples de même sérotype prélevés chez un même patient dans une période de 14 jours ont été comptés comme un seul, le site d'isolement le plus invasif étant consigné. Les isolats liés à la méningite ont été considérés comme les plus invasifs, suivis du sang, puis des autres sites stériles. Les données ont été réunies par groupe d'âge, soit les < 2 ans, les 2-14 ans, les 15-49 ans, les 50-64 ans et les ≥ 65 ans, et par région, soit l'Ouest (Colombie-Britannique, Alberta, Saskatchewan, Manitoba, Yukon, Territoires du Nord-Ouest et Nunavut), le Centre (Ontario et Québec) et l'Est (Nouveau-Brunswick, Nouvelle-Écosse, Île-du-Prince-Édouard et Terre-Neuve-et-Labrador). Il convient d’être prudent lors de l’interprétation des données présentées dans le rapport. En effet, l’interprétation globale des résultats est difficile en raison des limites associées aux isolats transmis à des fins d’analyse. Seul un sous-groupe d’isolats de laboratoire de chaque province peut être acheminé pour subir une analyse. Ce rapport ne constitue donc pas un véritable portrait de l’incidence ou du taux de la maladie au Canada. L’envoi d’isolats au LNM est volontaire et n’est pas systématique à l’échelle nationale. C’est pourquoi nous présentons ici des sommaires d’ensemble pour le Canada et les régions.
Les données de surveillance validées couvrant la période 2009 à 2012 sont tirées du Système de surveillance des maladies à déclaration obligatoire (SSMDO) [ASPC, 2014]. Les données sur la population correspondent aux estimations annuelles de la population au 1er juillet de Statistique Canada, de 2009 à 2013. La population des provinces et des territoires dont on n’a pas pu fournir de données sur les cas a été exclue du dénominateur.
Tous les isolats pneumococciques ont subi un test de solubilité biliaire et des analyses réalisées à l'aide d'optochine (Oxoid). Les infections invasives causées par le streptocoque du groupe A et le streptocoque du groupe B ont été confirmées à l'aide de la réaction PYR (pyrrolidone naphthylamide), de tests de sensibilité à la bacitracine (Oxoid) et de disques de sensibilité au triméthoprime-sulfaméthoxazole (BBL, 1,25/23,75 µg/ml) [Spellerberg, 2007]. Les sites d'isolement stériles cliniques sont les suivants : sang, liquide céphalorachidien (LCR) ou autre tissu nerveux, liquide pleural, liquide péritonéal, liquide péricardique, liquide articulaire et sites situés à l'intérieur du corps et des muscles, y compris les échantillons provenant d'une chirurgie ou d'une biopsie et les ponctions. En ce qui concerne S. pyogenes, les sites d'isolement ont été analysés lorsqu'un cas de syndrome de choc toxique ou de fasciite nécrosante était associé à l'infection [Relevé des maladies transmissibles au Canada, 2009; Minnesota Department of Health].
Au LNM, le sérotypage de l’agent responsable de la PI est réalisé en observant la réaction de Quellung à l’aide d’un pool commercial d’antisérums dirigés contre un groupe, un type ou un facteur (SSI Diagnostica, Statens Serum Institute, Copenhague, Danemark) [Austrian, 1976; Lovgren, 1998]. Les isolats associés à une absence de réaction de Quellung sont confirmés par un séquençage du gène rpoB [Drancourt, 2004; Clinical Laboratory Standards Institute (CLSI), 2008] de même qu’un sérotypage effectué par PCR, comme il est indiqué à l’adresse http://www.cdc.gov/ncidod/biotech/strep/pcr.htm.
En 2011, le LNM a entrepris un projet de collaboration avec le Health Sciences Centre – Canadian Antimicrobial Resistance Alliance (CARA) de l’Université du Manitoba en vue de la réalisation de tests de sensibilité aux antimicrobiens (TSA) sur les isolats de S. pneumoniae soumis au LNM; ce projet est désigné par l’acronyme SAVE (sérotypage de Streptococcus pneumoniae et sensibilité aux antimicrobiens : Évaluation de l’efficacité des vaccins au Canada après l’introduction du Pneu-C-13). Nous avons inclus dans l’étude tous les isolats ayant causé une PI, prélevés de sites stériles chez des sujets de n’importe quel groupe d’âge, qui ont été soumis par huit provinces participantes (Saskatchewan, Manitoba, Ontario, Québec, Nouvelle-Écosse, Île-du-Prince-Édouard, Nouveau-Brunswick et Terre-Neuve-et-Labrador). Pour les analyses, nous avons employé un panel composé de 18 antimicrobiens, dont les suivants : pénicilline, amoxicilline/clavulanate, céfuroxime, ceftriaxone, clarithromycine, ertapénème, méropénème, clindamycine, vancomycine, ciprofloxacine, lévofloxacine, moxifloxacine, linézolide, tigécycline, triméthoprime/sulfaméthoxazole et doxycycline. Les CMI de ces antimicrobiens sont déterminées par la technique de microdilution en milieu liquide du CLSI au moyen de plaques de microtitrage à 96 puits faites sur mesure [CLSI, 2011]. Les normes d’interprétation pour les CMI ont été définies en fonction des seuils publiés par le CLSI (M100-S21, 2011) pour tous les antibiotiques à l’exception de la ciprofloxacine et de la doxycycline, pour lesquelles on a plutôt employé les seuils d’interprétation approuvés par l’EUCAST [EUCAST, 2012].
On a réalisé le typage du gène emm de tous les isolats de streptocoque du groupe A causant une infection invasive acheminés au LNM. Les isolats ont été caractérisés à l’aide du protocole des CDC sur le séquençage du gène emm affiché à l’adresse http://www.cdc.gov/ncidod/biotech/strep/M-ProteinGene_typing.htm. Les séquences du gène emm obtenues ont été comparées à celles de la banque de données des CDC (Atlanta) (www.cdc.gov/ncidod/biotech/strep/strepblast.htm), et les résultats sont présentés par type, et non par sous-type (emm4.4 est classé comme emm4). Conformément aux lignes directrices du CLSI, la sensibilité aux antimicrobiens en ce qui concerne le chloramphénicol (CHL, 30 µg), l'érythromycine (ERY, 15 µg), la clindamycine (CLI, 2 µg), la pénicilline (PEN, 10 µg) et la vancomycine (VAN, 30, µg) été déterminée à l'aide de la méthode de diffusion en gélose utilisant des disques de Kirby-Bauer [Kellner, 2009].
Les sérotypes du streptocoque du groupe B ont été déterminés au moyen d’antisérums commerciaux d’agglutination au latex (SSI Diagnostica; Statens Serum Institute, Copenhague, Danemark).
Résultats et analyse
Streptococcus pneumoniae
Au Canada, le taux d'incidence global des PI est demeuré relativement stable au cours des cinq dernières années. En 2013, ce taux était de 9,0 cas pour 100 000, les taux les plus élevés étant toujours observés chez les nourrissons de < 1 an (17,8 cas pour 100 000), les enfants de 1-4 ans (11,2 cas pour 100 000) et les personnes âgées de 60 ans et plus (21,7 cas pour 100 000) (figure 2). Entre 2009 et 2013, on a observé une baisse régulière des taux de PI chez les nourrissons de < 1 an (de 27,6 à 17,8 cas pour 100 000) et chez les enfants de 1-4 ans (de 18,7 à 11,2 cas pour 100 000).
Figure 2. Taux d’incidence des PI pour 100 000 au Canada, 2009-2013
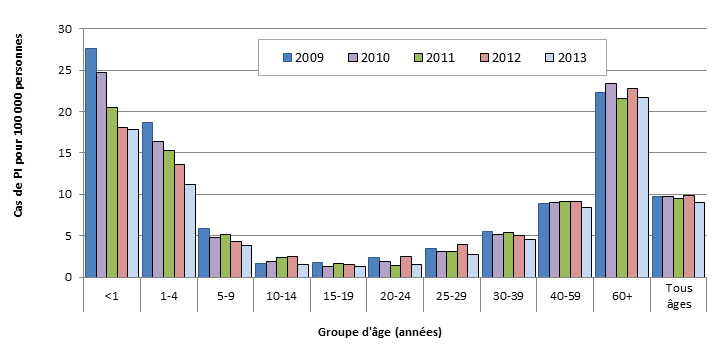
Taux d’incidence des PI pour 100 000 au Canada, 2009-2013 - Description textuel
Ce figure représente le nombre de cas par 100 000 des pneumococcies invasives distribués entre 2009 et 2013 par groupe d'âge.
| Groupe d'âge | Pourcentage par Année | ||||
|---|---|---|---|---|---|
| 2009 | 2010 | 2011 | 2012 | 2013 | |
| <1 | 27.6 | 24.7 | 20.5 | 18.1 | 17.8 |
| 1-4 | 18.7 | 16.4 | 15.3 | 13.6 | 11.2 |
| 5 to 9 | 5.8 | 4.8 | 5.2 | 4.4 | 3.8 |
| 10-14 | 1.6 | 1.8 | 2.4 | 2.4 | 1.5 |
| 15-19 | 1.8 | 1.3 | 1.6 | 1.6 | 1.2 |
| 20-24 | 2.4 | 1.9 | 1.4 | 2.5 | 1.5 |
| 25-29 | 3.5 | 3.1 | 3.0 | 4.0 | 2.8 |
| 30-39 | 5.5 | 5.1 | 5.4 | 5.1 | 4.5 |
| 40-59 | 8.9 | 9.0 | 9.1 | 9.1 | 8.4 |
| 60+ | 22.3 | 23.3 | 21.6 | 22.7 | 21.7 |
| All Ages | 9.8 | 9.8 | 9.6 | 9.8 | 9.0 |
| Année | Groupe d'âge (années) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| <1 | 1-4 | 5-9 | 10-14 | 15-19 | 20-24 | 25-29 | 30-39 | 40-59 | 60+ | Tout âge | |
| 2009 | 27.6 | 18.7 | 5.8 | 1.6 | 1.8 | 2.4 | 3.5 | 5.5 | 8.9 | 22.3 | 9.8 |
| 2010 | 24.7 | 16.4 | 4.8 | 1.8 | 1.3 | 1.9 | 3.1 | 5.1 | 9 | 23.3 | 9.8 |
| 2011 | 20.5 | 15.3 | 5.2 | 2.4 | 1.6 | 1.4 | 3 | 5.4 | 9.1 | 21.6 | 9.6 |
| 2012 | 18.1 | 13.6 | 4.4 | 2.4 | 1.6 | 2.5 | 4 | 5.1 | 9.1 | 22.7 | 9.8 |
| 2013 | 17.8 | 11.2 | 3.8 | 1.5 | 1.2 | 1.5 | 2.8 | 4.5 | 8.4 | 21.7 | 9.0 |
Les données sur la population correspondent aux estimations annuelles de la population au 1er juillet de Statistique Canada, de 2009 à 2013. La population des provinces et des territoires dont on n’a pas pu fournir de données sur les cas a été exclue du dénominateur.
Distribution des sérotypes de Streptococcus pneumoniae
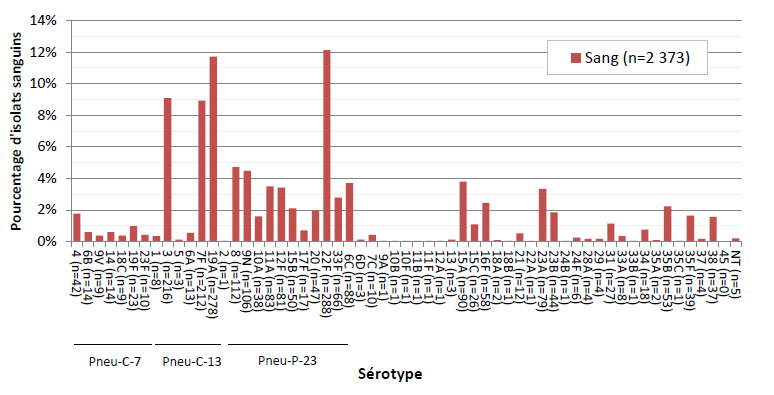
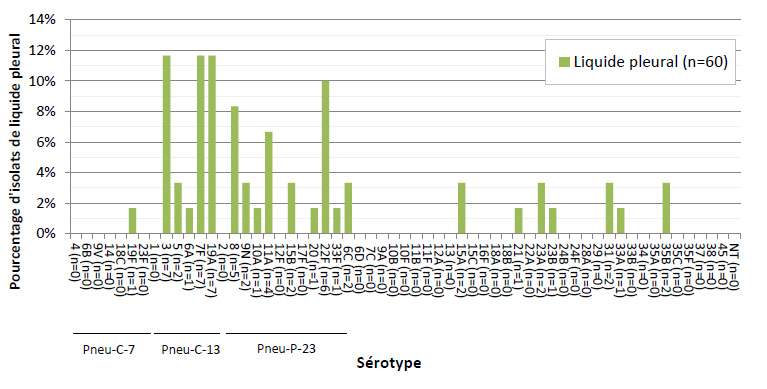
Sur les 2 577 isolats de PI sérotypés en 2013, 5,0 % (n = 129) ont été prélevés chez des enfants de < 2 ans, 2,9 % (n = 75) chez des enfants de 2-4 ans, 3,0 % (n = 78) chez des enfants de 5-14 ans, 20,6 % (n = 532) chez des adultes de 15-49 ans, 26,5 % (n = 682) chez des adultes de 50-64 ans et 40,6 % (n = 1 047) chez des personnes âgées de ≥ 65 ans (tableau 2). Le sang était le site d’isolement clinique le plus fréquent, représentant 92,1 % (n = 2 373) de l’ensemble des isolats (figures 3 et 4). Le sérotype 22F était uniformément présent dans toutes les sources, soit dans 10 à 12 % des isolats de chaque source d’isolement clinique, tandis que la proportion du sérotype 7F était plus forte dans le sang (8,9 %, n = 212) et dans les isolats de liquide pleural (11,7 %, n = 7) que dans le LCR (5,2 %, n = 4) ou d’autres sites d’isolement clinique stériles (4,5 %, n = 3). De même, le sérotype 3 était davantage présent dans le sang et le liquide pleural, représentant 9,1 % (n = 216) et 11,7 % (n = 7) des isolats, respectivement. Sur les 2 542 isolats pour lesquels le sexe était précisé, 55 % (n = 1 390) provenaient d’hommes.
En 2013, les sérotypes les plus répandus étaient 22F (12,1 %, n = 311), 19A (11,6 %, n = 299), 3 (8,9 %, n = 229) et 7F (8,8 %, n = 226), qui représentaient ensemble 41,3 % (n = 1 065) des cas de PI au Canada (figure 4). Le sérotype 22F (un composant du vaccin Pneu-C-23) a augmenté depuis 2010, devenant le sérotype le plus courant à l’échelle du Canada en 2013. Les sérotypes 19A, 7F et 6A poursuivent leur déclin constant depuis 2010 : le sérotype 19A est passé de 19,0 % (n = 517) en 2010 à 11,6 % (n = 299) en 2013, le sérotype 7F, de 14,3 % (n = 389) à 8,8 % (n = 226) et le sérotype 6A, de 2,7 % (n = 72) à 0,6 % (n = 15). De 2010 à 2013 le taux du sérotype 3 a légèrement augmenté, passant de 8,3 % (n = 225) à 8,9 % (n = 229), malgré une diminution parallèle des autres sérotypes associés au vaccin Pneu-C-13 (1, 5, 7F, 6A et 19A).
Sérotype 22F : La plus grande augmentation de la proportion relative des isolats 22F a été observée chez les enfants de < 2 ans, soit une augmentation de 7,8 % (n = 9) en 2012 à 17,1 % (n = 22) en 2013. Des augmentations de 7,6 % (n = 8) à 10,3 % (n = 8) chez les enfants de 5-14 ans, et de 7,2 % (n = 44) à 10,7 % (n = 57) chez les adultes de 15-49 ans ont aussi été observées entre 2012 et 2013, respectivement. Le sérotype 22F a légèrement diminué chez les enfants de 2-4 ans, passant de 12,2 % (n = 15) des isolats en 2012 à 9,3 % (n = 7) en 2013, mais ces taux demeurent considérablement élevés par rapport au taux de 2010, qui s’établissait à 2,7 % (n = 4). Les taux du sérotype 22F chez les 50-64 ans et les ≥ 65 ans sont restés relativement constants depuis l’année dernière, à 11,4 % (n = 78) et à 12,9 % (n = 135), respectivement (figure 18).
Sérotype 19A : En 2013, on a observé une diminution continue du sérotype 19A dans tous les groupes d’âge, sauf chez les 5-14 ans, où les proportions ont augmenté à 28,2 % (n = 22) en 2013 par rapport à 27,6 % (n = 29) en 2012, et de façon plus importante par rapport à 12,7 % (n = 14) en 2010. Des baisses marquées par rapport aux taux de 2012 ont été observées chez les enfants de < 2 ans, de 20,7 % (n = 24) à 11,6 % (n = 15), et chez les enfants de 2-4 ans, de 23,6 % (n = 29) à 18,7 % (n = 14). On a observé des diminutions plus modestes chez les adultes de 15-49 ans, de 10,8 % (n = 66) en 2012 à 9,6 % (n = 51) en 2013; chez les adultes de 50-64 ans, de 12,8 % (n = 89) à 12,0 % (n = 82); et chez les personnes âgées de ≥ 65 ans, de 12,9 % (n = 135) à 10,9 % (n = 114) (figure 15).
Sérotype 7F : En 2013, aucun isolat du sérotype 7F n’a été relevé parmi les 129 isolats prélevés chez des enfants de < 2 ans, une diminution par rapport au taux de 11,4 % (n = 21) en 2010. De 2010 à 2013, les taux du sérotype 7F ont également diminué chez les enfants de 2-4 ans, passant de 9,5 % (n = 14) à 2,7 % (n = 2); chez les enfants de 5-14 ans, de 25,5 % (n = 28) à 9,0 % (n = 7); chez les adultes de 15-49 ans, de 24,3 % (n = 142) à 15,6 % (n = 83); chez les adultes de 50-64 ans, de 12,3 % (n = 85) à 8,7 % (n = 59); et chez les personnes âgées de ≥ 65 ans, de 9,9 % (n = 96) à 6,9 % (n = 72). Il est intéressant de noter que dans le groupe d’âge 5-14 ans, une diminution de 7F et une augmentation de 19A ont été observées en parallèle, même si les deux sérotypes sont des composants du vaccin Pneu-C-13 (figure 16).
Sérotype 3 : Après avoir augmenté chez les enfants de < 2 ans, passant de 6,5 % (n = 12) à 8,6 % (n = 10) des isolats de 2010 à 2012, le sérotype 3 a connu une diminution en 2013 pour passer à 5,4 % (n = 7). Inversement, chez les 5-14 ans, le sérotype 3 a diminué, passant de 7,3 % (n = 8) à 2,9 % (n = 3) de 2010 à 2012, puis augmenté au cours de 2013 pour atteindre 5,1 % (n = 4). Dans les autres groupes d’âge, le sérotype 3 est demeuré à des taux relativement similaires au cours des dernières années, soit à 9,3 % (n = 7) chez les 2-4 ans, à 7,5 % (n = 40) chez les 15-49 ans, à 9,5 % (n = 65) chez les 50-64 ans et à 9,7 % (n = 101) chez les ≥ 65 ans en 2013 (figure 17).
Autres sérotypes : Même s’il est actuellement présent en faibles quantités, le sérotype 15B du vaccin Pneu-P-23 a augmenté chez les enfants de < 2 ans, passant de 1,6 % (n = 3) à 7,8 % (n = 10), et chez les enfants de 2-4 ans, passant de 3,4 % (n = 5) à 10,7 % (n = 8), de 2012 à 2013. Les taux du sérotype 33F (Pneu-P-23) ont également augmenté depuis 2012 dans ces groupes d’âge, passant de 2,6 % (n = 3) à 8,5 % (n = 11) chez les < 2 ans, de 1,6 % (n = 2) à 4,0 % (n = 3) chez les 2-4 ans. Le sérotype 21 a vu sa prévalence augmenter chez les < 2 ans, passant de 1,7 % (n = 2) à 5,4 % (n = 7) de 2012 à 2013, et le taux de 23B continue d’augmenter chez les 2-4 ans, passant de 2,0 % (n = 3) à 9,0 % (n = 6).
| Province | < 2 | 2 – 4 | 5 – 14 | 15 – 49 | 50 – 64 | ≥65 | Non précisé | Total |
|---|---|---|---|---|---|---|---|---|
| Colombie-Britannique | 9 | 8 | 20 | 74 | 83 | 135 | - | 329 |
| Alberta | 14 | 15 | 9 | 93 | 101 | 105 | 5 | 342 |
| Saskatchewan | 9 | 3 | 3 | 42 | 30 | 38 | - | 125 |
| Manitoba | 11 | 3 | 5 | 40 | 23 | 41 | 8 | 131 |
| Ontario | 37 | 27 | 26 | 155 | 244 | 418 | 16 | 923 |
| Québec | 42 | 14 | 11 | 93 | 152 | 219 | - | 531 |
| Nouveau-Brunswick | 3 | 1 | 1 | 11 | 18 | 39 | 1 | 74 |
| Nouvelle-Écosse | 2 | 3 | 1 | 14 | 20 | 33 | - | 73 |
| Île-du-Prince-Édouard | - | - | - | 5 | 2 | 5 | 3 | 15 |
| Terre-Neuve-et-Labrador | - | 1 | 2 | 1 | 4 | 11 | 1 | 20 |
| Yukon | - | - | - | - | 2 | 1 | - | 3 |
| Territoires du Nord-Ouest | - | - | - | 3 | 3 | 2 | - | 8 |
| Nunavut | 2 | - | - | 1 | - | - | - | 3 |
| Canada | 129 | 75 | 78 | 532 | 682 | 1047 | 34 | 2577 |
Figure 3. Sites d’isolement clinique de S. pneumoniae, 2013
Figure 3a. Isolats, < 5 ans (N=204)Footnote *
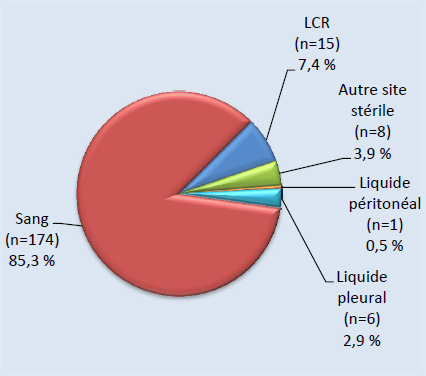
Sites d’isolement clinique de S. pneumoniae, 2013 - Description textuel
Ce figure représente les sites d'isolement cliniques de S. pneumoniae isolats soumis à des tests de particuliers <5 ans (N = 204).
| Source | Pourcentage d'isolats |
|---|---|
| Sang (n=174) | 85.3% |
| LCR (n=15) | 7.4% |
| Autre site stérile (n=8) | 3.9% |
| Liquide peritoneal (n=1) | 0.5% |
| Liquide pleural (n=6) | 2.9% |
Figure 3b. Isolats, ≥ 5 ans (N=2339)Footnote *
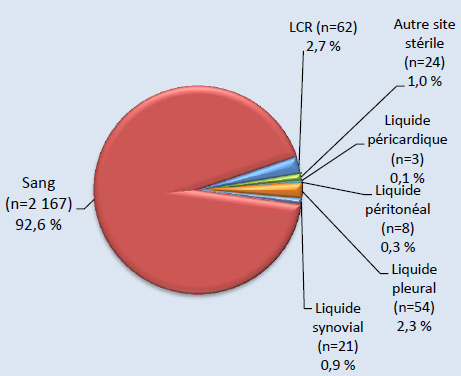
Sites d’isolement clinique de S. pneumoniae, 2013 - Description textuel
Ce figure représente les sites d'isolement cliniques de S. pneumoniae isolats soumis à des tests de particuliers ≥5 ans (N = 2339).
| Source | Pourcentage d'isolats |
|---|---|
| Sang (n=2167) | 92.6% |
| LCR (n=62) | 2.7% |
| Autre site stérile (n=24) | 1.0% |
| Liquide péricardique (n=3) | 0.1% |
| Liquide peritoneal (n=8) | 0.3% |
| Liquide pleural (n=54) | 2.3% |
| Liquide synovial (n=21) | 0.9% |
Figure 4. Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats sanguins, 2013
Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats sanguins, 2013 - Description textuel
Ce figure représente le pourcentage d'isolats distribués dans chaque sérotype pour tous les isolats de sang en 2013.
| Sérotype | Pourcentage d'isolats sanguins (n=2373) |
|---|---|
| 4(n=42) | 1.8% |
| 6B (n=14) | 0.6% |
| 9V (n=9) | 0.4% |
| 14 (n=14) | 0.6% |
| 18C (n=9) | 0.4% |
| 19F (n=23) | 1.0% |
| 23F (n=10) | 0.4% |
| 1 (n=8) | 0.3% |
| 3 (n=216) | 9.1% |
| 5 (n=3) | 0.1% |
| 6A (n=13) | 0.5% |
| 7F (n=212) | 8.9% |
| 19A (n=278) | 11.7% |
| 2 (n=1) | 0.0% |
| 8 (n=112) | 4.7% |
| 9N (n=106) | 4.5% |
| 10A (n=38) | 1.6% |
| 11A (n=83) | 3.5% |
| 12F (n=81) | 3.4% |
| 15B (n=50) | 2.1% |
| 17F (n=17) | 0.7% |
| 20 (n=47) | 2.0% |
| 22F (n=288) | 12.1% |
| 33F (n=66) | 2.8% |
| 6C (n=88) | 3.7% |
| 6D (n=3) | 0.1% |
| 7C (n=10) | 0.4% |
| 9A (n=1) | 0.0% |
| 10B (n=1) | 0.0% |
| 10F (n=1) | 0.0% |
| 11B (n=1) | 0.0% |
| 11F (n=1) | 0.0% |
| 12A (n=1) | 0.0% |
| 13 (n=3) | 0.1% |
| 15A (n=90) | 3.8% |
| 15C (n=26) | 1.1% |
| 16F (n=58) | 2.4% |
| 18A (n=2) | 0.1% |
| 18B (n=1) | 0.0% |
| 21 (n=12) | 0.5% |
| 22A (n=1) | 0.0% |
| 23A (n=79) | 3.3% |
| 23B (n=44) | 1.9% |
| 24B (n=1) | 0.0% |
| 24F (n=6) | 0.3% |
| 28A (n=4) | 0.2% |
| 29 (n=4) | 0.2% |
| 31 (n=27) | 1.1% |
| 33A (n=8) | 0.3% |
| 33B (n=1) | 0.0% |
| 34 (n=18) | 0.8% |
| 35A (n=2) | 0.1% |
| 35B (n=53) | 2.2% |
| 35C (n=1) | 0.0% |
| 35F (n=39) | 1.6% |
| 37 (n=4) | 0.2% |
| 38 (n=37) | 1.6% |
| 45 (n=0) | 0.0% |
| NT (n=5) | 0.2% |
Figure 5. Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats de LCR, 2013

Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats de LCR, 2013 - Description textuel
Ce figure représente le pourcentage d'isolats distribués dans chaque sérotype pour tous les CSF isole en 2013 (N = 77).
| Sérotype | Pourcentage d'isolats de LCR (n=77) |
|---|---|
| 4 (n=0) | 0.0% |
| 6B (n=1) | 1.3% |
| 9V (n=0) | 0.0% |
| 14 (n=1) | 1.3% |
| 18C (n=1) | 1.3% |
| 19F (n=0) | 0.0% |
| 23F (n=0) | 0.0% |
| 1(n=1) | 1.3% |
| 3 (n=3) | 3.9% |
| 5 (n=0) | 0.0% |
| 6A (n=0) | 0.0% |
| 7F (n=4) | 5.2% |
| 19A (n=4) | 5.2% |
| 2 (n=0) | 0.0% |
| 8 (n=1) | 1.3% |
| 9N (n=3) | 3.9% |
| 10A (n=6) | 7.8% |
| 11A (n=4) | 5.2% |
| 12F (n=5) | 6.5% |
| 15B (n=2) | 2.6% |
| 17F (n=0) | 0.0% |
| 20 (n=2) | 2.6% |
| 22F (n=9) | 11.7% |
| 33F (n=4) | 5.2% |
| 6C (n=0) | 0.0% |
| 6D (n=0) | 0.0% |
| 7C (n=0) | 0.0% |
| 9A (n=0) | 0.0% |
| 10B (n=0) | 0.0% |
| 10F (n=0) | 0.0% |
| 11B (n=0) | 0.0% |
| 11F (n=0) | 0.0% |
| 12A (n=0) | 0.0% |
| 13 (n=1) | 1.3% |
| 15A (n=3) | 3.9% |
| 15C (n=5) | 6.5% |
| 16F (n=1) | 1.3% |
| 18A (n=0) | 0.0% |
| 18B (n=0) | 0.0% |
| 21 (n=0) | 0.0% |
| 22A (n=0) | 0.0% |
| 23A (n=3) | 3.9% |
| 23B (n=4) | 5.2% |
| 24B (n=0) | 0.0% |
| 24F (n=0) | 0.0% |
| 28A (n=0) | 0.0% |
| 29 (n=0) | 0.0% |
| 31 (n=0) | 0.0% |
| 33A (n=0) | 0.0% |
| 33B (n=0) | 0.0% |
| 34 (n=1) | 1.3% |
| 35A (n=0) | 0.0% |
| 35B (n=3) | 3.9% |
| 35C (n=0) | 0.0% |
| 35F (n=5) | 6.5% |
| 37 (n=0) | 0.0% |
| 38 (n=0) | 0.0% |
| 45 (n=0) | 0.0% |
| NT (n=0) | 0.0% |
Figure 6. Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats de liquide pleural, 2013
Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats de liquide pleural, 2013 - Description textuel
Ce figure représente le pourcentage d'isolats distribués dans chaque sérotype pour tous liquide pleural isole en 2013 ( N = 60 ) .
| Sérotype | Pourcentage d'isolats de liquide pleural (n=60) |
|---|---|
| 4 (n=0) | 0.0% |
| 6B (n=0) | 0.0% |
| 9V (n=0) | 0.0% |
| 14 (n=0) | 0.0% |
| 18C (n=0) | 0.0% |
| 19F (n=1) | 1.7% |
| 23F (n=0) | 0.0% |
| 1 (n=0) | 0.0% |
| 3 (n=7) | 11.7% |
| 5 (n=2) | 3.3% |
| 6A (n=1) | 1.7% |
| 7F (n=7) | 11.7% |
| 19A (n=7) | 11.7% |
| 2 (n=0) | 0.0% |
| 8 (n=5) | 8.3% |
| 9N (n=2) | 3.3% |
| 10A (n=1) | 1.7% |
| 11A (n=4) | 6.7% |
| 12F (n=0) | 0.0% |
| 15B (n=2) | 3.3% |
| 17F (n=0) | 0.0% |
| 20 (n=1) | 1.7% |
| 22F (n=6) | 10.0% |
| 33F (n=1) | 1.7% |
| 6C (n=2) | 3.3% |
| 6D (n=0) | 0.0% |
| 7C (n=0) | 0.0% |
| 9A (n=0) | 0.0% |
| 10B (n=0) | 0.0% |
| 10F (n=0) | 0.0% |
| 11B (n=0) | 0.0% |
| 11F (n=0) | 0.0% |
| 12A (n=0) | 0.0% |
| 13 (n=0) | 0.0% |
| 15A (n=2) | 3.3% |
| 15C (n=0) | 0.0% |
| 16F (n=0) | 0.0% |
| 18A (n=0) | 0.0% |
| 18B (n=0) | 0.0% |
| 21 (n=1) | 1.7% |
| 22A (n=0) | 0.0% |
| 23A (n=2) | 3.3% |
| 23B (n=1) | 1.7% |
| 24B (n=0) | 0.0% |
| 24F (n=0) | 0.0% |
| 28A (n=0) | 0.0% |
| 29 (n=0) | 0.0% |
| 31 (n=2) | 3.3% |
| 33A (n=1) | 1.7% |
| 33B (n=0) | 0.0% |
| 34 (n=0) | 0.0% |
| 35A (n=0) | 0.0% |
| 35B (n=2) | 3.3% |
| 35C (n=0) | 0.0% |
| 35F (n=0) | 0.0% |
| 37 (n=0) | 0.0% |
| 38 (n=0) | 0.0% |
| 45 (n=0) | 0.0% |
| NT (n=0) | 0.0% |
Figure 7. Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats d'autres sites stériles, 2013
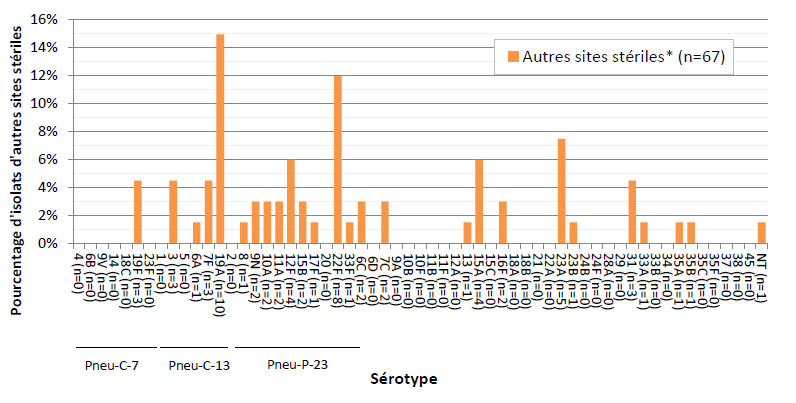
* « Autres sites stériles » comprend les isolats suivants : 4 liquides péricardiques, 10 liquides péritonéaux, 21 liquides synoviaux et 32 autres isolats issus d'échantillons de tissus profonds, de biopsies et de chirurgies.
Distribution des sérotypes de S. pneumoniae causant une infection invasive, isolats d'autres sites stériles, 2013 - Description textuel
Ce figure représente le pourcentage d'isolats distribués dans chaque sérotype pour tous les autres un site stérile isole en 2013 ( N = 67 ).
| Sérotype | Pourcentage d'isolats d'autres sites stériles* (n=67) |
|---|---|
| 4 (n=0) | 0.0% |
| 6B (n=0) | 0.0% |
| 9V (n=0) | 0.0% |
| 14 (n=0) | 0.0% |
| 18C (n=0) | 0.0% |
| 19F (n=3) | 4.5% |
| 23F (n=0) | 0.0% |
| 1 (n=0) | 0.0% |
| 3 (n=3) | 4.5% |
| 5 (n=0) | 0.0% |
| 6A (n=1) | 1.5% |
| 7F (n=3) | 4.5% |
| 19A (n=10) | 14.9% |
| 2 (n=0) | 0.0% |
| 8 (n=1) | 1.5% |
| 9N (n=2) | 3.0% |
| 10A (n=2) | 3.0% |
| 11A (n=2) | 3.0% |
| 12F (n=4) | 6.0% |
| 15B (n=2) | 3.0% |
| 17F (n=1) | 1.5% |
| 20 (n=0) | 0.0% |
| 22F (n=8) | 11.9% |
| 33F (n=1) | 1.5% |
| 6C (n=2) | 3.0% |
| 6D (n=0) | 0.0% |
| 7C (n=2) | 3.0% |
| 9A (n=0) | 0.0% |
| 10B (n=0) | 0.0% |
| 10F (n=0) | 0.0% |
| 11B (n=0) | 0.0% |
| 11F (n=0) | 0.0% |
| 12A (n=0) | 0.0% |
| 13 (n=1) | 1.5% |
| 15A (n=4) | 6.0% |
| 15C (n=0) | 0.0% |
| 16F (n=2) | 3.0% |
| 18A (n=0) | 0.0% |
| 18B (n=0) | 0.0% |
| 21 (n=0) | 0.0% |
| 22A (n=0) | 0.0% |
| 23A (n=5) | 7.5% |
| 23B (n=1) | 1.5% |
| 24B (n=0) | 0.0% |
| 24F (n=0) | 0.0% |
| 28A (n=0) | 0.0% |
| 29 (n=0) | 0.0% |
| 31 (n=3) | 4.5% |
| 33A (n=1) | 1.5% |
| 33B (n=0) | 0.0% |
| 34 (n=0) | 0.0% |
| 35A (n=1) | 1.5% |
| 35B (n=1) | 1.5% |
| 35C (n=0) | 0.0% |
| 35F (n=0) | 0.0% |
| 37 (n=0) | 0.0% |
| 38 (n=0) | 0.0% |
| 45 (n=0) | 0.0% |
| NT (n=1) | 1.5% |
* « Autres sites stériles » comprend les isolats suivants : 4 liquides péricardiques, 10 liquides péritonéaux, 21 liquides synoviaux et 32 autres isolats issus d'échantillons de tissus profonds, de biopsies et de chirurgies. |
|
Figure 8. Sérotypes de S. pneumoniae causant une infection invasive, tous les groupes d’âge combinés, 2010-2013
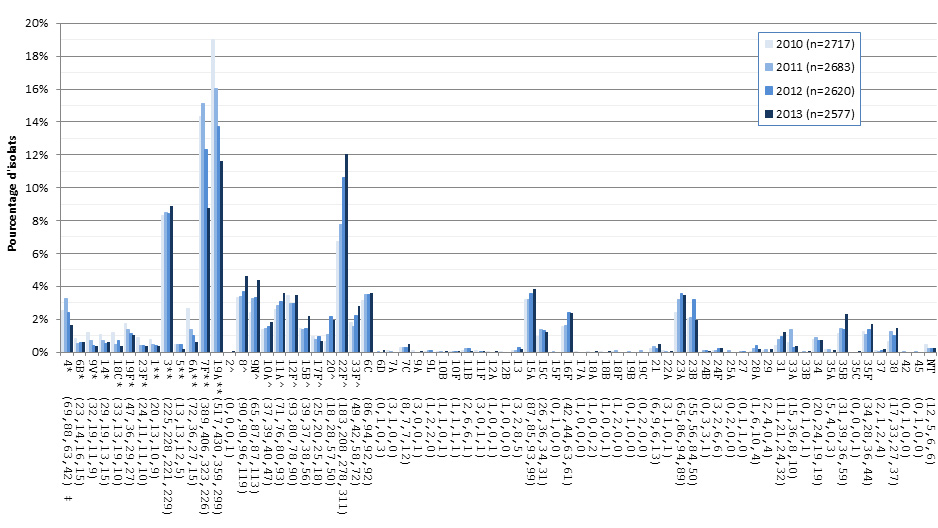
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive, tous les groupes d’âge combinés, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de tous les isolats prélevés entre 2010-2013.
| Sérotype | 2010 (n=2717) | 2011 (n=2683) | 2012 (n=2620) | 2013 (n=2577) |
|---|---|---|---|---|
| 4* (69,88,63,42) ‡ | 2.54% | 3.28% | 2.40% | 1.63% |
| 6B* (23,14,16,15) | 0.85% | 0.52% | 0.61% | 0.58% |
| 9V* (32,19,11,9) | 1.18% | 0.71% | 0.42% | 0.35% |
| 14* (29,19,13,15) | 1.07% | 0.71% | 0.50% | 0.58% |
| 18C* (33,13,19,10) | 1.21% | 0.48% | 0.73% | 0.39% |
| 19F* (47,36,29,27) | 1.73% | 1.34% | 1.11% | 1.05% |
| 23F* (24,11,11,10) | 0.88% | 0.41% | 0.42% | 0.39% |
| 1** (20,13,10,9) | 0.74% | 0.48% | 0.38% | 0.35% |
| 3** (225,228,221,229) | 8.28% | 8.50% | 8.44% | 8.89% |
| 5** (13,13,12,5) | 0.48% | 0.48% | 0.46% | 0.19% |
| 6A** (72,36,27,15) | 2.65% | 1.34% | 1.03% | 0.58% |
| 7F** (389,406,323,226) | 14.32% | 15.13% | 12.33% | 8.77% |
| 19A**(517,430,359,299) | 19.03% | 16.03% | 13.70% | 11.60% |
| 2^ (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.04% |
| 8^ (90,90,96,119) | 3.31% | 3.35% | 3.66% | 4.62% |
| 9N^ (65,87,87,113) | 2.39% | 3.24% | 3.32% | 4.38% |
| 10A^ (37,39,40,47) | 1.36% | 1.45% | 1.53% | 1.82% |
| 11A^ (71,76,80,93) | 2.61% | 2.83% | 3.05% | 3.61% |
| 12F^ (93,80,78,90) | 3.42% | 2.98% | 2.98% | 3.49% |
| 15B^ (39,37,38,56) | 1.44% | 1.38% | 1.45% | 2.17% |
| 17F^ (25,20,25,18) | 0.92% | 0.75% | 0.95% | 0.70% |
| 20^ (18,28,57,50) | 0.66% | 1.04% | 2.18% | 1.94% |
| 22F^ (183,208,278,311) | 6.74% | 7.75% | 10.61% | 12.07% |
| 33F^ (49,42,58,72) | 1.80% | 1.57% | 2.21% | 2.79% |
| 6C (86,94,92,92) | 3.17% | 3.50% | 3.51% | 3.57% |
| 6D (0,1,0,3) | 0.00% | 0.04% | 0.00% | 0.12% |
| 7A (3,1,0,0) | 0.11% | 0.04% | 0.00% | 0.00% |
| 7C (8,7,7,12) | 0.29% | 0.26% | 0.27% | 0.47% |
| 9A (3,0,0,1) | 0.11% | 0.00% | 0.00% | 0.04% |
| 9L (1,2,2,0) | 0.04% | 0.07% | 0.08% | 0.00% |
| 10B (1,1,0,1) | 0.04% | 0.04% | 0.00% | 0.04% |
| 10F (1,1,1,1) | 0.04% | 0.04% | 0.04% | 0.04% |
| 11B (2,6,6,1) | 0.07% | 0.22% | 0.23% | 0.04% |
| 11F (3,0,1,1) | 0.11% | 0.00% | 0.04% | 0.04% |
| 12A (1,0,0,1) | 0.04% | 0.00% | 0.00% | 0.04% |
| 12B (1,0,0,0) | 0.04% | 0.00% | 0.00% | 0.00% |
| 13 (3,2,8,5) | 0.11% | 0.07% | 0.31% | 0.19% |
| 15A (87,85,93,99) | 3.20% | 3.17% | 3.55% | 3.84% |
| 15C (26,36,34,31) | 0.96% | 1.34% | 1.30% | 1.20% |
| 15F (0,1,0,0) | 0.00% | 0.04% | 0.00% | 0.00% |
| 16F (42,44,63,61) | 1.55% | 1.64% | 2.40% | 2.37% |
| 17A (1,0,0,0) | 0.04% | 0.00% | 0.00% | 0.00% |
| 18A (1,0,0,2) | 0.04% | 0.00% | 0.00% | 0.08% |
| 18B (1,0,0,1) | 0.04% | 0.00% | 0.00% | 0.04% |
| 18F (1,2,0,0) | 0.04% | 0.07% | 0.00% | 0.00% |
| 19B (0,1,0,0) | 0.00% | 0.04% | 0.00% | 0.00% |
| 19C (0,2,0,0) | 0.00% | 0.07% | 0.00% | 0.00% |
| 21 (6,9,6,13) | 0.22% | 0.34% | 0.23% | 0.50% |
| 22A (3,1,0,1) | 0.11% | 0.04% | 0.00% | 0.04% |
| 23A (65,86,94,89) | 2.39% | 3.21% | 3.59% | 3.45% |
| 23B (55,56,84,50) | 2.02% | 2.09% | 3.21% | 1.94% |
| 24B (0,3,3,1) | 0.00% | 0.11% | 0.11% | 0.04% |
| 24F (3,2,6,6) | 0.11% | 0.07% | 0.23% | 0.23% |
| 25A (0,2,0,0) | 0.00% | 0.07% | 0.00% | 0.00% |
| 27 (0,1,1,0) | 0.00% | 0.04% | 0.04% | 0.00% |
| 28A (1,6,10,4) | 0.04% | 0.22% | 0.38% | 0.16% |
| 29 (2,4,0,4) | 0.07% | 0.15% | 0.00% | 0.16% |
| 31 (11,21,24,32) | 0.40% | 0.78% | 0.92% | 1.24% |
| 33A (15,36,8,10) | 0.55% | 1.34% | 0.31% | 0.39% |
| 33B (0,1,0,1) | 0.00% | 0.04% | 0.00% | 0.04% |
| 34 (20,24,19,19) | 0.74% | 0.89% | 0.73% | 0.74% |
| 35A (5,4,0,3) | 0.18% | 0.15% | 0.00% | 0.12% |
| 35B (31,39,36,59) | 1.14% | 1.45% | 1.37% | 2.29% |
| 35C (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.04% |
| 35F (34,28,36,44) | 1.25% | 1.04% | 1.37% | 1.71% |
| 37 (2,1,2,4) | 0.07% | 0.04% | 0.08% | 0.16% |
| 38 (17,33,27,37) | 0.63% | 1.23% | 1.03% | 1.44% |
| 42 (0,1,0,0) | 0.00% | 0.04% | 0.00% | 0.00% |
| 45 (0,1,0,0) | 0.00% | 0.04% | 0.00% | 0.00% |
| NT (12,5,6,6) | 0.44% | 0.19% | 0.23% | 0.23% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | ||||
Figure 9. Sérotypes de S. pneumoniae causant une infection invasive chez les enfants de < 2 ans, 2010-2013
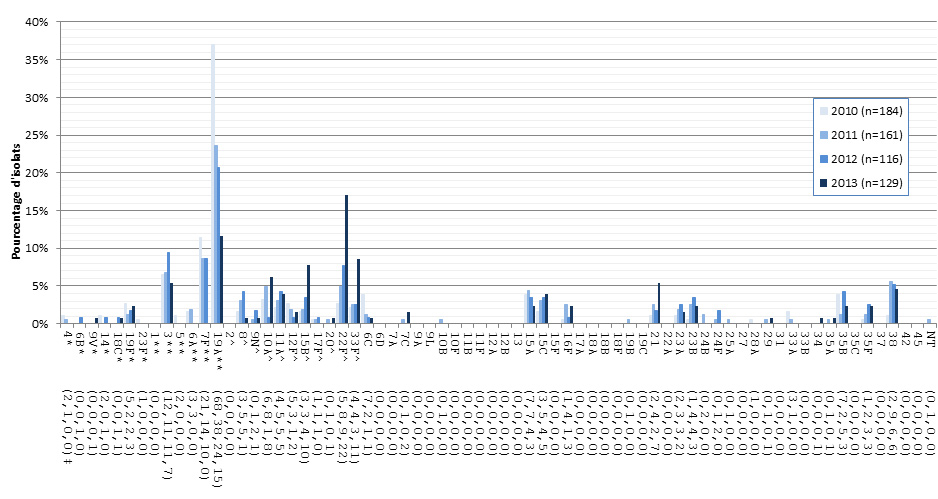
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive chez les enfants de < 2 ans, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de < 2 ans recueillies entre 2010-2013.
| Sérotype | 2010 (n=184) | 2011 (n=161) | 2012 (n=116) | 2013 (n=129) |
|---|---|---|---|---|
| 4* (2,1,0,0)‡ | 1.09% | 0.62% | 0.00% | 0.00% |
| 6B* (0,0,1,0) | 0.00% | 0.00% | 0.86% | 0.00% |
| 9V* (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.78% |
| 14* (2,0,1,0) | 1.09% | 0.00% | 0.86% | 0.00% |
| 18C* (0,0,1,1) | 0.00% | 0.00% | 0.86% | 0.78% |
| 19F* (5,2,2,3) | 2.72% | 1.24% | 1.72% | 2.33% |
| 23F* (1,0,0,0) | 0.54% | 0.00% | 0.00% | 0.00% |
| 1** (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 3** (12,11,11,7) | 6.52% | 6.83% | 9.48% | 5.43% |
| 5** (2,0,0,0) | 1.09% | 0.00% | 0.00% | 0.00% |
| 6A** (3,3,0,0) | 1.63% | 1.86% | 0.00% | 0.00% |
| 7F** (21,14,10,0) | 11.41% | 8.70% | 8.62% | 0.00% |
| 19A** (68,38,24,15) | 36.96% | 23.60% | 20.69% | 11.63% |
| 2^ (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 8^ (3,5,5,1) | 1.63% | 3.11% | 4.31% | 0.78% |
| 9N^ (0,1,2,1) | 0.00% | 0.62% | 1.72% | 0.78% |
| 10A^ (6,8,1,8) | 3.26% | 4.97% | 0.86% | 6.20% |
| 11A^ (4,5,5,5) | 2.17% | 3.11% | 4.31% | 3.88% |
| 12F^ (5,3,1,2) | 2.72% | 1.86% | 0.86% | 1.55% |
| 15B^ (3,3,4,10) | 1.63% | 1.86% | 3.45% | 7.75% |
| 17F^ (1,1,1,0) | 0.54% | 0.62% | 0.86% | 0.00% |
| 20^ (0,1,0,1) | 0.00% | 0.62% | 0.00% | 0.78% |
| 22F^ (5,8,9,22) | 2.72% | 4.97% | 7.76% | 17.05% |
| 33F^ (4,4,3,11) | 2.17% | 2.48% | 2.59% | 8.53% |
| 6C (7,2,1,1) | 3.80% | 1.24% | 0.86% | 0.78% |
| 6D (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7C (0,1,0,2) | 0.00% | 0.62% | 0.00% | 1.55% |
| 9A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 9L (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 10B (0,1,0,0) | 0.00% | 0.62% | 0.00% | 0.00% |
| 10F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 13 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 15A (7,7,4,3) | 3.80% | 4.35% | 3.45% | 2.33% |
| 15C (3,5,4,5) | 1.63% | 3.11% | 3.45% | 3.88% |
| 15F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 16F (1,4,1,3) | 0.54% | 2.48% | 0.86% | 2.33% |
| 17A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19B (0,1,0,0) | 0.00% | 0.62% | 0.00% | 0.00% |
| 19C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 21 (2,4,2,7) | 1.09% | 2.48% | 1.72% | 5.43% |
| 22A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 23A (2,3,3,2) | 1.09% | 1.86% | 2.59% | 1.55% |
| 23B (1,4,4,3) | 0.54% | 2.48% | 3.45% | 2.33% |
| 24B (0,2,0,0) | 0.00% | 1.24% | 0.00% | 0.00% |
| 24F (0,1,2,0) | 0.00% | 0.62% | 1.72% | 0.00% |
| 25A (0,1,0,0) | 0.00% | 0.62% | 0.00% | 0.00% |
| 27 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 28A (1,0,0,0) | 0.54% | 0.00% | 0.00% | 0.00% |
| 29 (0,1,0,1) | 0.00% | 0.62% | 0.00% | 0.78% |
| 31 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 33A (3,1,0,0) | 1.63% | 0.62% | 0.00% | 0.00% |
| 33B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 34 (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.78% |
| 35A (0,1,0,1) | 0.00% | 0.62% | 0.00% | 0.78% |
| 35B (7,2,5,3) | 3.80% | 1.24% | 4.31% | 2.33% |
| 35C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 35F (1,2,3,3) | 0.54% | 1.24% | 2.59% | 2.33% |
| 37 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 38 (2,9,6,6) | 1.09% | 5.59% | 5.17% | 4.65% |
| 42 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 45 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| NT (0,1,0,0) | 0.00% | 0.62% | 0.00% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | ||||
Figure 10. Sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 2-4 ans, 2010-2013
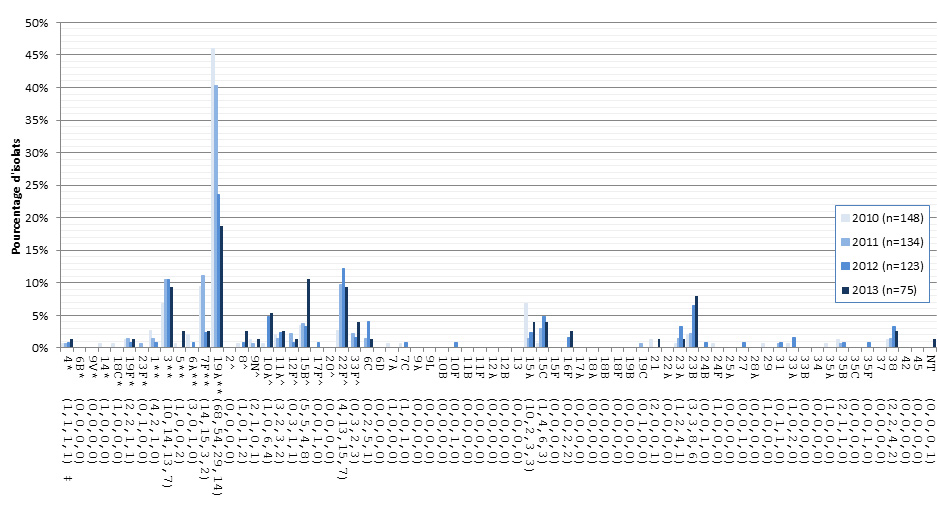
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 2-4 ans, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de olds 2-4 années recueillies entre 2010-2013.
| Sérotype | 2010 (n=148) | 2011 (n=134) | 2012 (n=123) | 2013 (n=75) |
|---|---|---|---|---|
| 4* (1,1,1,1) ‡ | 0.68% | 0.75% | 0.81% | 1.33% |
| 6B* (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 9V* (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 14* (1,0,0,0) | 0.68% | 0.00% | 0.00% | 0.00% |
| 18C* (1,0,0,0) | 0.68% | 0.00% | 0.00% | 0.00% |
| 19F* (2,2,1,1) | 1.35% | 1.49% | 0.81% | 1.33% |
| 23F* (0,1,0,0) | 0.00% | 0.75% | 0.00% | 0.00% |
| 1** (4,2,1,0) | 2.70% | 1.49% | 0.81% | 0.00% |
| 3** (10,14,13,7) | 6.76% | 10.45% | 10.57% | 9.33% |
| 5** (1,0,0,2) | 0.68% | 0.00% | 0.00% | 2.67% |
| 6A** (3,0,1,0) | 2.03% | 0.00% | 0.81% | 0.00% |
| 7F** (14,15,3,2) | 9.46% | 11.19% | 2.44% | 2.67% |
| 19A**(68,54,29,14) | 45.95% | 40.30% | 23.58% | 18.67% |
| 2^ (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 8^ (1,0,1,2) | 0.68% | 0.00% | 0.81% | 2.67% |
| 9N^ (2,1,0,1) | 1.35% | 0.75% | 0.00% | 1.33% |
| 10A^ (1,0,6,4) | 0.68% | 0.00% | 4.88% | 5.33% |
| 11A^ (3,2,3,2) | 2.03% | 1.49% | 2.44% | 2.67% |
| 12F^ (0,3,1,1) | 0.00% | 2.24% | 0.81% | 1.33% |
| 15B^ (5,5,4,8) | 3.38% | 3.73% | 3.25% | 10.67% |
| 17F^ (0,0,1,0) | 0.00% | 0.00% | 0.81% | 0.00% |
| 20^ (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 22F^ (4,13,15,7) | 2.70% | 9.70% | 12.20% | 9.33% |
| 33F^ (0,3,2,3) | 0.00% | 2.24% | 1.63% | 4.00% |
| 6C (0,2,5,1) | 0.00% | 1.49% | 4.07% | 1.33% |
| 6D (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7A (1,0,0,0) | 0.68% | 0.00% | 0.00% | 0.00% |
| 7C (1,0,1,0) | 0.68% | 0.00% | 0.81% | 0.00% |
| 9A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 9L (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 10F (0,0,1,0) | 0.00% | 0.00% | 0.81% | 0.00% |
| 11B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 13 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 15A (10,2,3,3) | 6.76% | 1.49% | 2.44% | 4.00% |
| 15C (1,4,6,3) | 0.68% | 2.99% | 4.88% | 4.00% |
| 15F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 16F (0,0,2,2) | 0.00% | 0.00% | 1.63% | 2.67% |
| 17A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19C (0,1,0,0) | 0.00% | 0.75% | 0.00% | 0.00% |
| 21 (2,0,0,1) | 1.35% | 0.00% | 0.00% | 1.33% |
| 22A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 23A (1,2,4,1) | 0.68% | 1.49% | 3.25% | 1.33% |
| 23B (3,3,8,6) | 2.03% | 2.24% | 6.50% | 8.00% |
| 24B (0,0,1,0) | 0.00% | 0.00% | 0.81% | 0.00% |
| 24F (1,0,0,0) | 0.68% | 0.00% | 0.00% | 0.00% |
| 25A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 27 (0,0,1,0) | 0.00% | 0.00% | 0.81% | 0.00% |
| 28A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 29 (1,0,0,0) | 0.68% | 0.00% | 0.00% | 0.00% |
| 31 (0,1,1,0) | 0.00% | 0.75% | 0.81% | 0.00% |
| 33A (1,0,2,0) | 0.68% | 0.00% | 1.63% | 0.00% |
| 33B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 34 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 35A (1,0,0,0) | 0.68% | 0.00% | 0.00% | 0.00% |
| 35B (2,1,1,0) | 1.35% | 0.75% | 0.81% | 0.00% |
| 35C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 35F (0,0,1,0) | 0.00% | 0.00% | 0.81% | 0.00% |
| 37 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 38 (2,2,4,2) | 1.35% | 1.49% | 3.25% | 2.67% |
| 42 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 45 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| NT (0,0,0,1) | 0.00% | 0.00% | 0.00% | 1.33% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010, 2011, 2012 et 2013, respectivement. | ||||
Figure 11. Sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 5-14 ans, 2010-2013
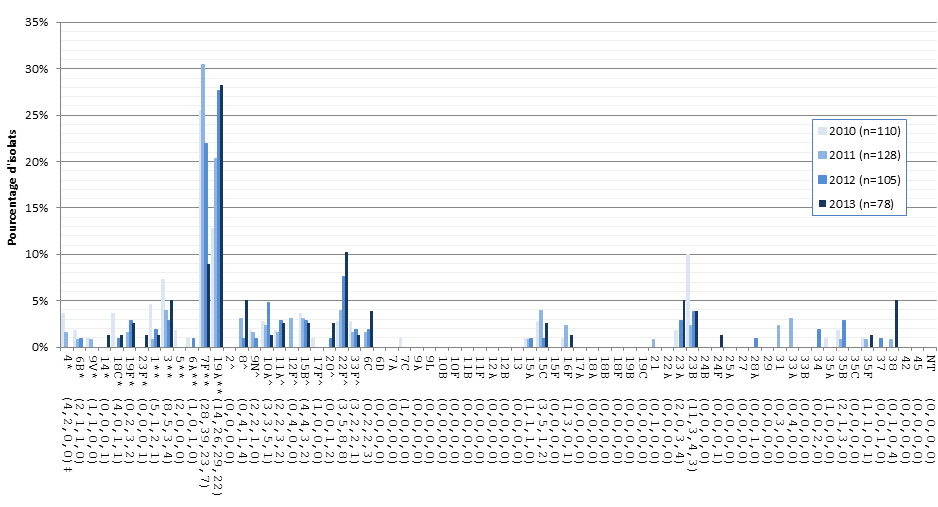
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 5-14 ans, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype 5-14 ans recueillies entre 2010-2013.
| Sérotype | 2010 (n=110) | 2011 (n=128) | 2012 (n=105) | 2013 (n=78) |
|---|---|---|---|---|
| 4* (4,2,0,0)‡ | 3.64% | 1.56% | 0.00% | 0.00% |
| 6B* (2,1,1,0) | 1.82% | 0.78% | 0.95% | 0.00% |
| 9V* (1,1,0,0) | 0.91% | 0.78% | 0.00% | 0.00% |
| 14* (0,0,0,1) | 0.00% | 0.00% | 0.00% | 1.28% |
| 18C* (4,0,1,1) | 3.64% | 0.00% | 0.95% | 1.28% |
| 19F* (0,2,3,2) | 0.00% | 1.56% | 2.86% | 2.56% |
| 23F* (0,0,0,1) | 0.00% | 0.00% | 0.00% | 1.28% |
| 1** (5,1,2,1) | 4.55% | 0.78% | 1.90% | 1.28% |
| 3** (8,5,3,4) | 7.27% | 3.91% | 2.86% | 5.13% |
| 5** (2,0,0,0) | 1.82% | 0.00% | 0.00% | 0.00% |
| 6A** (1,0,1,0) | 0.91% | 0.00% | 0.95% | 0.00% |
| 7F** (28,39,23,7) | 25.45% | 30.47% | 21.90% | 8.97% |
| 19A**(14,26,29,22) | 12.73% | 20.31% | 27.62% | 28.21% |
| 2^ (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 8^ (0,4,1,4) | 0.00% | 3.13% | 0.95% | 5.13% |
| 9N^ (2,2,1,0) | 1.82% | 1.56% | 0.95% | 0.00% |
| 10A^ (3,3,5,1) | 2.73% | 2.34% | 4.76% | 1.28% |
| 11A^ (2,2,3,2) | 1.82% | 1.56% | 2.86% | 2.56% |
| 12F^ (0,4,0,0) | 0.00% | 3.13% | 0.00% | 0.00% |
| 15B^ (4,4,3,2) | 3.64% | 3.13% | 2.86% | 2.56% |
| 17F^ (1,0,0,0) | 0.91% | 0.00% | 0.00% | 0.00% |
| 20^ (0,0,1,2) | 0.00% | 0.00% | 0.95% | 2.56% |
| 22F^ (3,5,8,8) | 2.73% | 3.91% | 7.62% | 10.26% |
| 33F^ (3,2,2,1) | 2.73% | 1.56% | 1.90% | 1.28% |
| 6C (0,2,2,3) | 0.00% | 1.56% | 1.90% | 3.85% |
| 6D (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7C (1,0,0,0) | 0.91% | 0.00% | 0.00% | 0.00% |
| 9A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 9L (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 10F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 13 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 15A (1,1,1,0) | 0.91% | 0.78% | 0.95% | 0.00% |
| 15C (3,5,1,2) | 2.73% | 3.91% | 0.95% | 2.56% |
| 15F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 16F (1,3,0,1) | 0.91% | 2.34% | 0.00% | 1.28% |
| 17A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 21 (0,1,0,0) | 0.00% | 0.78% | 0.00% | 0.00% |
| 22A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 23A (2,0,3,4) | 1.82% | 0.00% | 2.86% | 5.13% |
| 23B (11,3,4,3) | 10.00% | 2.34% | 3.81% | 3.85% |
| 24B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 24F (0,0,0,1) | 0.00% | 0.00% | 0.00% | 1.28% |
| 25A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 27 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 28A (0,0,1,0) | 0.00% | 0.00% | 0.95% | 0.00% |
| 29 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 31 (0,3,0,0) | 0.00% | 2.34% | 0.00% | 0.00% |
| 33A (0,4,0,0) | 0.00% | 3.13% | 0.00% | 0.00% |
| 33B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 34 (0,0,2,0) | 0.00% | 0.00% | 1.90% | 0.00% |
| 35A (1,0,0,0) | 0.91% | 0.00% | 0.00% | 0.00% |
| 35B (2,1,3,0) | 1.82% | 0.78% | 2.86% | 0.00% |
| 35C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 35F (1,1,0,1) | 0.91% | 0.78% | 0.00% | 1.28% |
| 37 (0,0,1,0) | 0.00% | 0.00% | 0.95% | 0.00% |
| 38 (0,1,0,4) | 0.00% | 0.78% | 0.00% | 5.13% |
| 42 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 45 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| NT (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | ||||
Figure 12. Sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 15-49 ans, 2010-2013
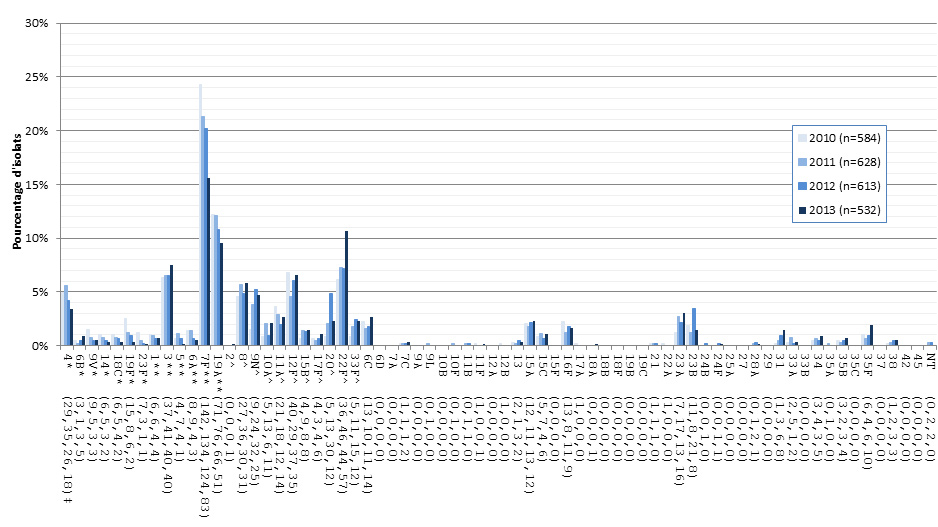
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 15-49 ans, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de 15-49 ans recueillies entre 2010-2013.
| Sérotype | 2010 (n=584) | 2011 (n=628) | 2012 (n=613) | 2013 (n=532) |
|---|---|---|---|---|
| 4* (29,35,26,18) | 4.97% | 5.57% | 4.24% | 3.38% |
| 6B* (3,1,3,5) | 0.51% | 0.16% | 0.49% | 0.94% |
| 9V* (9,5,3,3) | 1.54% | 0.80% | 0.49% | 0.56% |
| 14* (6,5,3,2) | 1.03% | 0.80% | 0.49% | 0.38% |
| 18C* (6,5,4,2) | 1.03% | 0.80% | 0.65% | 0.38% |
| 19F* (15,8,6,2) | 2.57% | 1.27% | 0.98% | 0.38% |
| 23F* (7,3,1,1) | 1.20% | 0.48% | 0.16% | 0.19% |
| 1** (6,6,4,4) | 1.03% | 0.96% | 0.65% | 0.75% |
| 3** (37,41,40,40) | 6.34% | 6.53% | 6.53% | 7.52% |
| 5** (4,7,4,1) | 0.68% | 1.11% | 0.65% | 0.19% |
| 6A** (8,9,4,3) | 1.37% | 1.43% | 0.65% | 0.56% |
| 7F** (142,134,124,83) | 24.32% | 21.34% | 20.23% | 15.60% |
| 19A**(71,76,66,51) | 12.16% | 12.10% | 10.77% | 9.59% |
| 2^ (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.19% |
| 8^ (27,36,30,31) | 4.62% | 5.73% | 4.89% | 5.83% |
| 9N^ (9,24,32,25) | 1.54% | 3.82% | 5.22% | 4.70% |
| 10A^ (5,13,6,11) | 0.86% | 2.07% | 0.98% | 2.07% |
| 11A^ (21,18,12,14) | 3.60% | 2.87% | 1.96% | 2.63% |
| 12F^ (40,29,37,35) | 6.85% | 4.62% | 6.04% | 6.58% |
| 15B^ (4,9,8,8) | 0.68% | 1.43% | 1.31% | 1.50% |
| 17F^ (4,3,4,6) | 0.68% | 0.48% | 0.65% | 1.13% |
| 20^ (5,13,30,12) | 0.86% | 2.07% | 4.89% | 2.26% |
| 22F^ (36,46,44,57) | 6.16% | 7.32% | 7.18% | 10.71% |
| 33F^ (5,11,15,12) | 0.86% | 1.75% | 2.45% | 2.26% |
| 6C (13,10,11,14) | 2.23% | 1.59% | 1.79% | 2.63% |
| 6D (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7C (1,1,1,2) | 0.17% | 0.16% | 0.16% | 0.38% |
| 9A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 9L (0,1,0,0) | 0.00% | 0.16% | 0.00% | 0.00% |
| 10B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 10F (0,1,0,0) | 0.00% | 0.16% | 0.00% | 0.00% |
| 11B (0,1,1,0) | 0.00% | 0.16% | 0.16% | 0.00% |
| 11F (1,0,0,1) | 0.17% | 0.00% | 0.00% | 0.19% |
| 12A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 12B (1,0,0,0) | 0.17% | 0.00% | 0.00% | 0.00% |
| 13 (2,1,3,2) | 0.34% | 0.16% | 0.49% | 0.38% |
| 15A (12,11,13,12) | 2.05% | 1.75% | 2.12% | 2.26% |
| 15C (5,7,4,6) | 0.86% | 1.11% | 0.65% | 1.13% |
| 15F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 16F (13,8,11,9) | 2.23% | 1.27% | 1.79% | 1.69% |
| 17A (1,0,0,0) | 0.17% | 0.00% | 0.00% | 0.00% |
| 18A (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.19% |
| 18B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 21 (1,1,1,0) | 0.17% | 0.16% | 0.16% | 0.00% |
| 22A (1,0,0,0) | 0.17% | 0.00% | 0.00% | 0.00% |
| 23A (7,17,13,16) | 1.20% | 2.71% | 2.12% | 3.01% |
| 23B (11,8,21,8) | 1.88% | 1.27% | 3.43% | 1.50% |
| 24B (0,0,1,0) | 0.00% | 0.00% | 0.16% | 0.00% |
| 24F (0,0,1,1) | 0.00% | 0.00% | 0.16% | 0.19% |
| 25A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 27 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 28A (0,1,2,1) | 0.00% | 0.16% | 0.33% | 0.19% |
| 29 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 31 (1,3,6,8) | 0.17% | 0.48% | 0.98% | 1.50% |
| 33A (2,5,1,2) | 0.34% | 0.80% | 0.16% | 0.38% |
| 33B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 34 (3,4,3,5) | 0.51% | 0.64% | 0.49% | 0.94% |
| 35A (0,1,0,0) | 0.00% | 0.16% | 0.00% | 0.00% |
| 35B (3,2,3,4) | 0.51% | 0.32% | 0.49% | 0.75% |
| 35C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 35F (6,4,6,10) | 1.03% | 0.64% | 0.98% | 1.88% |
| 37 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 38 (1,2,3,3) | 0.17% | 0.32% | 0.49% | 0.56% |
| 42 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 45 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| NT (0,2,2,0) | 0.00% | 0.32% | 0.33% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | ||||
Figure 13. Sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 50-64 ans, 2010-2013
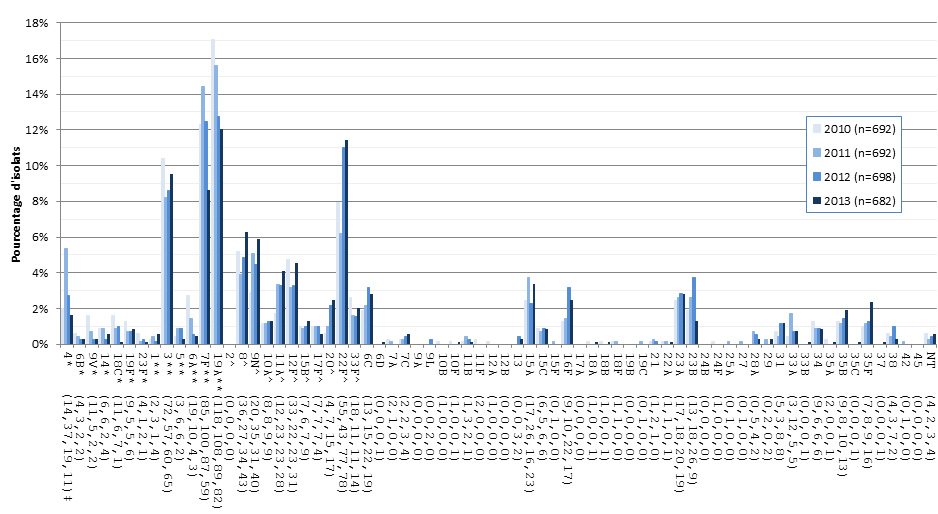
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 50-64 ans, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de jeunes 50-64 ans recueillies entre 2010-2013.
| Sérotype | 2010 (n=692) | 2011 (n=692) | 2012 (n=698) | 2013 (n=682) |
|---|---|---|---|---|
| 4* (14,37,19,11) | 2.02% | 5.35% | 2.72% | 1.61% |
| 6B* (4,3,2,2) | 0.58% | 0.43% | 0.29% | 0.29% |
| 9V* (11,5,2,2) | 1.59% | 0.72% | 0.29% | 0.29% |
| 14* (6,6,2,4) | 0.87% | 0.87% | 0.29% | 0.59% |
| 18C* (11,6,7,1) | 1.59% | 0.87% | 1.00% | 0.15% |
| 19F* (9,5,5,6) | 1.30% | 0.72% | 0.72% | 0.88% |
| 23F* (4,1,2,1) | 0.58% | 0.14% | 0.29% | 0.15% |
| 1** (2,3,1,4) | 0.29% | 0.43% | 0.14% | 0.59% |
| 3** (72,57,60,65) | 10.40% | 8.24% | 8.60% | 9.53% |
| 5** (3,6,6,2) | 0.43% | 0.87% | 0.86% | 0.29% |
| 6A** (19,10,4,3) | 2.75% | 1.45% | 0.57% | 0.44% |
| 7F** (85,100,87,59) | 12.28% | 14.45% | 12.46% | 8.65% |
| 19A**(118,108,89,82) | 17.05% | 15.61% | 12.75% | 12.02% |
| 2^ (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 8^ (36,27,34,43) | 5.20% | 3.90% | 4.87% | 6.30% |
| 9N^ (20,35,31,40) | 2.89% | 5.06% | 4.44% | 5.87% |
| 10A^ (8,8,9,9) | 1.16% | 1.16% | 1.29% | 1.32% |
| 11A^ (12,23,23,28) | 1.73% | 3.32% | 3.30% | 4.11% |
| 12F^ (33,22,23,31) | 4.77% | 3.18% | 3.30% | 4.55% |
| 15B^ (7,6,7,9) | 1.01% | 0.87% | 1.00% | 1.32% |
| 17F^ (7,7,7,4) | 1.01% | 1.01% | 1.00% | 0.59% |
| 20^ (4,7,15,17) | 0.58% | 1.01% | 2.15% | 2.49% |
| 22F^ (55,43,77,78) | 7.95% | 6.21% | 11.03% | 11.44% |
| 33F^ (18,11,11,14) | 2.60% | 1.59% | 1.58% | 2.05% |
| 6C (13,15,22,19) | 1.88% | 2.17% | 3.15% | 2.79% |
| 6D (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.15% |
| 7A (2,1,0,0) | 0.29% | 0.14% | 0.00% | 0.00% |
| 7C (2,2,3,4) | 0.29% | 0.29% | 0.43% | 0.59% |
| 9A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 9L (0,0,2,0) | 0.00% | 0.00% | 0.29% | 0.00% |
| 10B (1,0,0,0) | 0.14% | 0.00% | 0.00% | 0.00% |
| 10F (1,0,0,1) | 0.14% | 0.00% | 0.00% | 0.15% |
| 11B (1,3,2,1) | 0.14% | 0.43% | 0.29% | 0.15% |
| 11F (2,0,0,0) | 0.29% | 0.00% | 0.00% | 0.00% |
| 12A (1,0,0,0) | 0.14% | 0.00% | 0.00% | 0.00% |
| 12B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 13 (0,0,3,2) | 0.00% | 0.00% | 0.43% | 0.29% |
| 15A (17,26,16,23) | 2.46% | 3.76% | 2.29% | 3.37% |
| 15C (6,5,6,6) | 0.87% | 0.72% | 0.86% | 0.88% |
| 15F (0,1,0,0) | 0.00% | 0.14% | 0.00% | 0.00% |
| 16F (9,10,22,17) | 1.30% | 1.45% | 3.15% | 2.49% |
| 17A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18A (1,0,0,1) | 0.14% | 0.00% | 0.00% | 0.15% |
| 18B (1,0,0,1) | 0.14% | 0.00% | 0.00% | 0.15% |
| 18F (1,1,0,0) | 0.14% | 0.14% | 0.00% | 0.00% |
| 19B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19C (0,1,0,0) | 0.00% | 0.14% | 0.00% | 0.00% |
| 21 (1,2,1,0) | 0.14% | 0.29% | 0.14% | 0.00% |
| 22A (1,1,0,1) | 0.14% | 0.14% | 0.00% | 0.15% |
| 23A (17,18,20,19) | 2.46% | 2.60% | 2.87% | 2.79% |
| 23B (13,18,26,9) | 1.88% | 2.60% | 3.72% | 1.32% |
| 24B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 24F (1,0,0,0) | 0.14% | 0.00% | 0.00% | 0.00% |
| 25A (0,1,0,0) | 0.00% | 0.14% | 0.00% | 0.00% |
| 27 (0,1,0,0) | 0.00% | 0.14% | 0.00% | 0.00% |
| 28A (0,5,4,2) | 0.00% | 0.72% | 0.57% | 0.29% |
| 29 (0,2,0,2) | 0.00% | 0.29% | 0.00% | 0.29% |
| 31 (5,3,8,8) | 0.72% | 0.43% | 1.15% | 1.17% |
| 33A (3,12,5,5) | 0.43% | 1.73% | 0.72% | 0.73% |
| 33B (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.15% |
| 34 (9,6,6,6) | 1.30% | 0.87% | 0.86% | 0.88% |
| 35A (2,0,0,1) | 0.29% | 0.00% | 0.00% | 0.15% |
| 35B (9,8,10,13) | 1.30% | 1.16% | 1.43% | 1.91% |
| 35C (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.15% |
| 35F (7,8,9,16) | 1.01% | 1.16% | 1.29% | 2.35% |
| 37 (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.15% |
| 38 (4,3,7,2) | 0.58% | 0.43% | 1.00% | 0.29% |
| 42 (0,1,0,0) | 0.00% | 0.14% | 0.00% | 0.00% |
| 45 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| NT (4,2,3,4) | 0.58% | 0.29% | 0.43% | 0.59% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | ||||
Figure 14. Sérotypes de S. pneumoniae causant une infection invasive chez les personnes âgées de ≥ 65 ans, 2010-2013
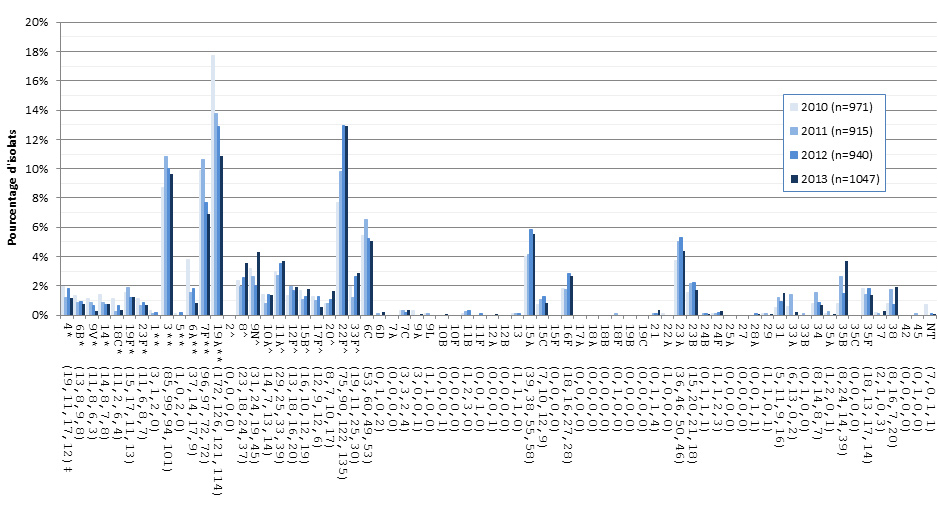
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Sérotypes de S. pneumoniae causant une infection invasive chez les personnes âgées de ≥ 65 ans, 2010-2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de ≥ jeunes de 65 ans recueillies entre 2010-2013.
| Sérotype | 2010 (n=971) | 2011 (n=915) | 2012 (n=940) | 2013 (n=1047) |
|---|---|---|---|---|
| 4* (19,11,17,12) | 1.96% | 1.20% | 1.81% | 1.15% |
| 6B* (13,8,9,8) | 1.34% | 0.87% | 0.96% | 0.76% |
| 9V* (11,8,6,3) | 1.13% | 0.87% | 0.64% | 0.29% |
| 14* (14,8,7,8) | 1.44% | 0.87% | 0.74% | 0.76% |
| 18C* (11,2,6,4) | 1.13% | 0.22% | 0.64% | 0.38% |
| 19F* (15,17,11,13) | 1.54% | 1.86% | 1.17% | 1.24% |
| 23F* (11,6,8,7) | 1.13% | 0.66% | 0.85% | 0.67% |
| 1** (3,1,2,0) | 0.31% | 0.11% | 0.21% | 0.00% |
| 3** (85,99,94,101) | 8.75% | 10.82% | 10.00% | 9.65% |
| 5** (1,0,2,0) | 0.10% | 0.00% | 0.21% | 0.00% |
| 6A** (37,14,17,9) | 3.81% | 1.53% | 1.81% | 0.86% |
| 7F** (96,97,72,72) | 9.89% | 10.60% | 7.66% | 6.88% |
| 19A**(172,126,121,114) | 17.71% | 13.77% | 12.87% | 10.89% |
| 2^ (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 8^ (23,18,24,37) | 2.37% | 1.97% | 2.55% | 3.53% |
| 9N^ (31,24,19,45) | 3.19% | 2.62% | 2.02% | 4.30% |
| 10A^ (14,7,13,14) | 1.44% | 0.77% | 1.38% | 1.34% |
| 11A^ (29,25,33,39) | 2.99% | 2.73% | 3.51% | 3.72% |
| 12F^ (13,18,16,20) | 1.34% | 1.97% | 1.70% | 1.91% |
| 15B^ (16,10,12,19) | 1.65% | 1.09% | 1.28% | 1.81% |
| 17F^ (12,9,12,6) | 1.24% | 0.98% | 1.28% | 0.57% |
| 20^ (8,7,10,17) | 0.82% | 0.77% | 1.06% | 1.62% |
| 22F^ (75,90,122,135) | 7.72% | 9.84% | 12.98% | 12.89% |
| 33F^ (19,11,25,30) | 1.96% | 1.20% | 2.66% | 2.87% |
| 6C (53,60,49,53) | 5.46% | 6.56% | 5.21% | 5.06% |
| 6D (0,1,0,2) | 0.00% | 0.11% | 0.00% | 0.19% |
| 7A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 7C (3,3,2,4) | 0.31% | 0.33% | 0.21% | 0.38% |
| 9A (3,0,0,1) | 0.31% | 0.00% | 0.00% | 0.10% |
| 9L (1,1,0,0) | 0.10% | 0.11% | 0.00% | 0.00% |
| 10B (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.10% |
| 10F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 11B (1,2,3,0) | 0.10% | 0.22% | 0.32% | 0.00% |
| 11F (0,0,1,0) | 0.00% | 0.00% | 0.11% | 0.00% |
| 12A (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.10% |
| 12B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 13 (1,1,1,0) | 0.10% | 0.11% | 0.11% | 0.00% |
| 15A (39,38,55,58) | 4.02% | 4.15% | 5.85% | 5.54% |
| 15C (7,10,12,9) | 0.72% | 1.09% | 1.28% | 0.86% |
| 15F (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 16F (18,16,27,28) | 1.85% | 1.75% | 2.87% | 2.67% |
| 17A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 18F (0,1,0,0) | 0.00% | 0.11% | 0.00% | 0.00% |
| 19B (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 19C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 21 (0,1,1,4) | 0.00% | 0.11% | 0.11% | 0.38% |
| 22A (1,0,0,0) | 0.10% | 0.00% | 0.00% | 0.00% |
| 23A (36,46,50,46) | 3.71% | 5.03% | 5.32% | 4.39% |
| 23B (15,20,21,18) | 1.54% | 2.19% | 2.23% | 1.72% |
| 24B (0,1,1,1) | 0.00% | 0.11% | 0.11% | 0.10% |
| 24F (1,1,2,3) | 0.10% | 0.11% | 0.21% | 0.29% |
| 25A (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 27 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 28A (0,0,1,1) | 0.00% | 0.00% | 0.11% | 0.10% |
| 29 (1,1,0,1) | 0.10% | 0.11% | 0.00% | 0.10% |
| 31 (5,11,9,16) | 0.51% | 1.20% | 0.96% | 1.53% |
| 33A (6,13,0,2) | 0.62% | 1.42% | 0.00% | 0.19% |
| 33B (0,1,0,0) | 0.00% | 0.11% | 0.00% | 0.00% |
| 34 (8,14,8,7) | 0.82% | 1.53% | 0.85% | 0.67% |
| 35A (1,2,0,1) | 0.10% | 0.22% | 0.00% | 0.10% |
| 35B (8,24,14,39) | 0.82% | 2.62% | 1.49% | 3.72% |
| 35C (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 35F (18,13,17,14) | 1.85% | 1.42% | 1.81% | 1.34% |
| 37 (2,1,0,3) | 0.21% | 0.11% | 0.00% | 0.29% |
| 38 (8,16,7,20) | 0.82% | 1.75% | 0.74% | 1.91% |
| 42 (0,0,0,0) | 0.00% | 0.00% | 0.00% | 0.00% |
| 45 (0,1,0,0) | 0.00% | 0.11% | 0.00% | 0.00% |
| NT (7,0,1,1) | 0.72% | 0.00% | 0.11% | 0.10% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | ||||
Figure 15. Sérotype 19A par âge
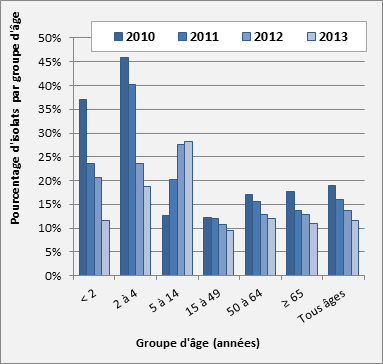
Sérotype 19A par âge - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 19A pour chaque groupe d'âge entre 2010 à 2013.
| Année | Pourcentage d'isolats par groupe d'âge | ||||||
|---|---|---|---|---|---|---|---|
| < 2 | 2 to 4 | 5 to 14 | 15 to 49 | 50 to 64 | ≥ 65 | Tous âges | |
| 2010 | 37.0% | 45.9% | 12.7% | 12.2% | 17.1% | 17.7% | 19.0% |
| 2011 | 23.6% | 40.3% | 20.3% | 12.1% | 15.6% | 13.8% | 16.0% |
| 2012 | 20.7% | 23.6% | 27.6% | 10.8% | 12.8% | 12.9% | 13.7% |
| 2013 | 11.6% | 18.7% | 28.2% | 9.6% | 12.0% | 10.9% | 11.6% |
Figure 16. Sérotype 7F par âge
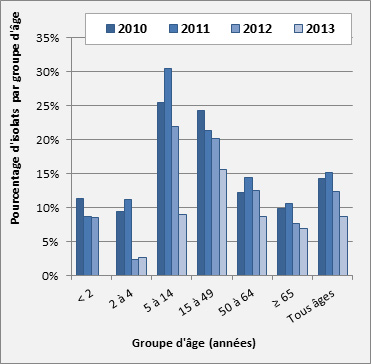
Sérotype 7F par âge - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 7F pour chaque groupe d'âge entre 2010 à 2013.
| Année | Pourcentage d'isolats par groupe d'âge | ||||||
|---|---|---|---|---|---|---|---|
| < 2 | 2 to 4 | 5 to 14 | 15 to 49 | 50 to 64 | ≥ 65 | Tous âges | |
| 2010 | 11.4% | 9.5% | 25.5% | 24.3% | 12.3% | 9.9% | 14.3% |
| 2011 | 8.7% | 11.2% | 30.5% | 21.3% | 14.5% | 10.6% | 15.1% |
| 2012 | 8.6% | 2.4% | 21.9% | 20.2% | 12.5% | 7.7% | 12.3% |
| 2013 | 0.0% | 2.7% | 9.0% | 15.6% | 8.7% | 6.9% | 8.8% |
Figure 17. Sérotype 3 par âge
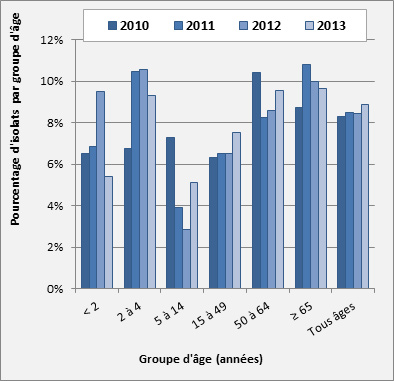
Sérotype 3 par âge - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 3 pour chaque groupe d'âge entre 2010 à 2013.
| Année | Pourcentage d'isolats par groupe d'âge | ||||||
|---|---|---|---|---|---|---|---|
| < 2 | 2 to 4 | 5 to 14 | 15 to 49 | 50 to 64 | ≥ 65 | Tous âges | |
| 2010 | 6.5% | 6.8% | 7.3% | 6.3% | 10.4% | 8.8% | 8.3% |
| 2011 | 6.8% | 10.4% | 3.9% | 6.5% | 8.2% | 10.8% | 8.5% |
| 2012 | 9.5% | 10.6% | 2.9% | 6.5% | 8.6% | 10.0% | 8.4% |
| 2013 | 5.4% | 9.3% | 5.1% | 7.5% | 9.5% | 9.6% | 8.9% |
Figure 18. Sérotype 22F par âge
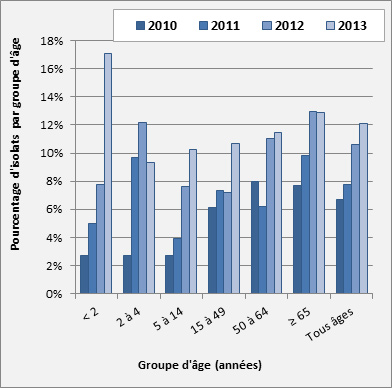
Sérotype 22F par âge - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 22F pour chaque groupe d'âge entre 2010 à 2013.
| Année | Pourcentage d'isolats par groupe d'âge | ||||||
|---|---|---|---|---|---|---|---|
| < 2 | 2 to 4 | 5 to 14 | 15 to 49 | 50 to 64 | ≥ 65 | Tous âges | |
| 2010 | 2.7% | 2.7% | 2.7% | 6.2% | 7.9% | 7.7% | 6.7% |
| 2011 | 5.0% | 9.7% | 3.9% | 7.3% | 6.2% | 9.8% | 7.8% |
| 2012 | 7.8% | 12.2% | 7.6% | 7.2% | 11.0% | 13.0% | 10.6% |
| 2013 | 17.1% | 9.3% | 10.3% | 10.7% | 11.4% | 12.9% | 12.1% |
Distribution régionale des sérotypes de S. pneumoniae
En 2013, le sérotype 19A était le plus fréquent dans les régions du Centre et de l’Ouest, représentant 8,2 % (n = 77) et 13,6 % (n = 197) des isolats, respectivement, alors que dans les régions de l’Est, le 7F était le plus fréquent avec 15,4 % (n = 28). La présence du sérotype 7F a aussi été importante dans le Centre du Canada, représentant 8,4 % (n = 122) des isolats, mais ce sérotype ne correspondait qu’à 8,1 % (n = 76) des isolats de l’Ouest du pays. Le sérotype 22F était réparti uniformément entre les régions, soit à 11,5 % (n = 108), à 12,8 % (n = 186) et à 9,3 % (n = 17) des totaux régionaux dans l’Ouest, dans le Centre et dans l’Est, respectivement. Le sérotype 3 était aussi réparti uniformément entre les régions canadiennes, représentant 8,7 % (n = 82) des isolats dans l’Ouest, 9,0 % (n = 131) dans le Centre et 8,8 % (n = 16) dans l’Est (figure 19).
Dans les isolats prélevés chez les enfants de < 2 ans (figure 20), la proportion du sérotype 19A était plus élevée dans les régions du Centre (15,2 %, n = 12) que dans les régions de l’Ouest (4,4 %, n = 2). Dans l’Ouest du Canada, la proportion du sérotype 33F était relativement élevée dans les isolats de ce groupe d’âge (13,3 %, n = 6).
Dans les isolats prélevés chez les enfants de 2-4 ans (figure 21), le sérotype 19A est prédominant dans les régions du Centre, représentant 22,0 % (n = 9) des isolats. Quant aux sérotypes 23B (13,8 %, n = 4), 33F (10,3 %, n = 3) et 15A (6,9 %, n = 2), ils sont plus fréquents dans l’Ouest que dans les autres régions.
Les sérotypes des isolats prélevés chez les enfants de 5-14 ans (figure 22) et chez les adultes de 15-49 ans (figure 23) étaient répartis de façon assez uniforme entre les régions, la proportion des sérotypes 3, 19A et 22F étant légèrement plus élevée dans le Centre du Canada et celle des sérotypes 8, 9N, 12F et 20 étant légèrement plus élevée dans l’Ouest.
Chez les adultes de 50-64 ans (figure 24), les sérotypes 19A et 7F étaient présents en plus faible proportion dans l’Ouest du Canada (7,4 %, n = 18 et 6,2 %, n = 15, respectivement) que dans les régions du Centre (14,9 %, n = 59 et 9,3 %, n = 37, respectivement) et de l’Est (11,4 %, n = 5 et 15,9 %, n = 7, respectivement). Le sérotype 3 était présent en proportions relativement similaires dans l’Ouest (10,7 %, n = 26) et le Centre du Canada (9,3 %, n = 37), et des concentrations moindres ont été observées dans l’Est (4,6 %, n = 2). De même, le sérotype 22F était réparti uniformément dans les régions de l’Ouest (12,0 %, n = 29) et les régions du Centre (11,9 %, n = 47) et se trouvait en plus faibles proportions dans les régions de l’Est (4,6 %, n = 2). D’autres sérotypes présentent un taux de prévalence relativement plus élevé, notamment les sérotypes 12F, 20 et 23B dans les régions de l’Ouest (7,4 %, n = 18; 6,2 % n = 15; et 2,5 %, n = 6, respectivement), les sérotypes 15A et 23A dans les régions du Centre (4,8 %, n = 19 et 3,5 %, n = 14, respectivement) et les sérotypes 8, 33F et 6C dans les régions de l’Est (13,6 %, n = 6; 6,8 %, n = 3; et 6,8 %, n = 3, respectivement).
Dans le groupe des personnes âgées de ≥ 65 ans (figure 25), la distribution régionale correspond à celle des adultes de 50-64 ans. Les taux des sérotypes 19A et 7F sont plus faibles dans l’Ouest (8,4 %, n = 27 et 5,9 %, n = 19, respectivement) que dans l’Est (14,8 %, n = 13 et 11,6 %, n = 74, respectivement) et le Centre du Canada (11,6 %, n = 74 et 7,1 %, n = 45, respectivement). Dans ce groupe d’âge, les sérotypes 3 et 22F étaient répartis uniformément dans l’ensemble du Canada à environ 10 % et 13 %, respectivement. Comme dans le cas de la distribution régionale des sérotypes chez les 50-64 ans, la proportion des sérotypes 12F, 20 et 23B était plus élevée dans l’Ouest (4,7 %, n = 15; 4,0%, n = 13; et 3,1 %, n = 10, respectivement), et dans le Centre du Canada, la proportion des sérotypes 15A et 23A (6,9 %, n = 44 et 5,2 %, n = 33, respectivement) était élevée par rapport à d’autres régions.
Figure 19. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive, tous les groupes d'âge combinés, 2013
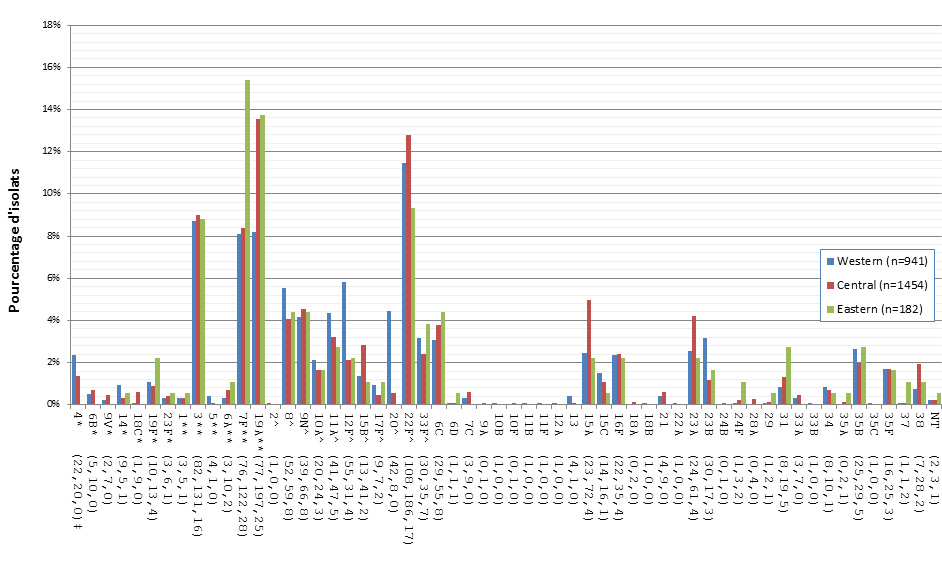
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive, tous les groupes d'âge combinés, 2013 - Description textuel
Ce figure représente le nombre d'isolats de tous les sérotypes répartis régionalement, recueillies en 2013.
| Sérotype | Ouest (n=941) | Centre (n=1454) | Est (n=182) |
|---|---|---|---|
| 4* (22,20,0)‡ | 2.30% | 1.40% | 0.00% |
| 6B* (5,10,0) | 0.50% | 0.70% | 0.00% |
| 9V* (2,7,0) | 0.20% | 0.50% | 0.00% |
| 14* (9,5,1) | 1.00% | 0.30% | 0.50% |
| 18C* (1,9,0) | 0.10% | 0.60% | 0.00% |
| 19F* (10,13,4) | 1.10% | 0.90% | 2.20% |
| 23F* (3,6,1) | 0.30% | 0.40% | 0.50% |
| 1** (3,5,1) | 0.30% | 0.30% | 0.50% |
| 3** (82,131,16) | 8.70% | 9.00% | 8.80% |
| 5** (4,1,0) | 0.40% | 0.10% | 0.00% |
| 6A** (3,10,2) | 0.30% | 0.70% | 1.10% |
| 7F** (76,122,28) | 8.10% | 8.40% | 15.40% |
| 19A**(77,197,25) | 8.20% | 13.50% | 13.70% |
| 2^ (1,0,0) | 0.10% | 0.00% | 0.00% |
| 8^ (52,59,8) | 5.50% | 4.10% | 4.40% |
| 9N^ (39,66,8) | 4.10% | 4.50% | 4.40% |
| 10A^ (20,24,3) | 2.10% | 1.70% | 1.60% |
| 11A^ (41,47,5) | 4.40% | 3.20% | 2.70% |
| 12F^ (55,31,4) | 5.80% | 2.10% | 2.20% |
| 15B^ (13,41,2) | 1.40% | 2.80% | 1.10% |
| 17F^ (9,7,2) | 1.00% | 0.50% | 1.10% |
| 20^ (42,8,0) | 4.50% | 0.60% | 0.00% |
| 22F^ (108,186,17) | 11.50% | 12.80% | 9.30% |
| 33F^ (30,35,7) | 3.20% | 2.40% | 3.80% |
| 6C (29,55,8) | 3.10% | 3.80% | 4.40% |
| 6D (1,1,1) | 0.10% | 0.10% | 0.50% |
| 7C (3,9,0) | 0.30% | 0.60% | 0.00% |
| 9A (0,1,0) | 0.00% | 0.10% | 0.00% |
| 10B (1,0,0) | 0.10% | 0.00% | 0.00% |
| 10F (0,1,0) | 0.00% | 0.10% | 0.00% |
| 11B (1,0,0) | 0.10% | 0.00% | 0.00% |
| 11F (1,0,0) | 0.10% | 0.00% | 0.00% |
| 12A (1,0,0) | 0.10% | 0.00% | 0.00% |
| 13 (4,1,0) | 0.40% | 0.10% | 0.00% |
| 15A (23,72,4) | 2.40% | 5.00% | 2.20% |
| 15C (14,16,1) | 1.50% | 1.10% | 0.50% |
| 16F (22,35,4) | 2.30% | 2.40% | 2.20% |
| 18A (0,2,0) | 0.00% | 0.10% | 0.00% |
| 18B (1,0,0) | 0.10% | 0.00% | 0.00% |
| 21 (4,9,0) | 0.40% | 0.60% | 0.00% |
| 22A (1,0,0) | 0.10% | 0.00% | 0.00% |
| 23A (24,61,4) | 2.60% | 4.20% | 2.20% |
| 23B (30,17,3) | 3.20% | 1.20% | 1.60% |
| 24B (0,1,0) | 0.00% | 0.10% | 0.00% |
| 24F (1,3,2) | 0.10% | 0.20% | 1.10% |
| 28A (0,4,0) | 0.00% | 0.30% | 0.00% |
| 29 (1,2,1) | 0.10% | 0.10% | 0.50% |
| 31 (8,19,5) | 0.90% | 1.30% | 2.70% |
| 33A (3,7,0) | 0.30% | 0.50% | 0.00% |
| 33B (1,0,0) | 0.10% | 0.00% | 0.00% |
| 34 (8,10,1) | 0.90% | 0.70% | 0.50% |
| 35A (0,2,1) | 0.00% | 0.10% | 0.50% |
| 35B (25,29,5) | 2.70% | 2.00% | 2.70% |
| 35C (1,0,0) | 0.10% | 0.00% | 0.00% |
| 35F (16,25,3) | 1.70% | 1.70% | 1.60% |
| 37 (1,1,2) | 0.10% | 0.10% | 1.10% |
| 38 (7,28,2) | 0.70% | 1.90% | 1.10% |
| NT (2,3,1) | 0.20% | 0.20% | 0.50% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 20. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les enfants de < 2 ans, 2013
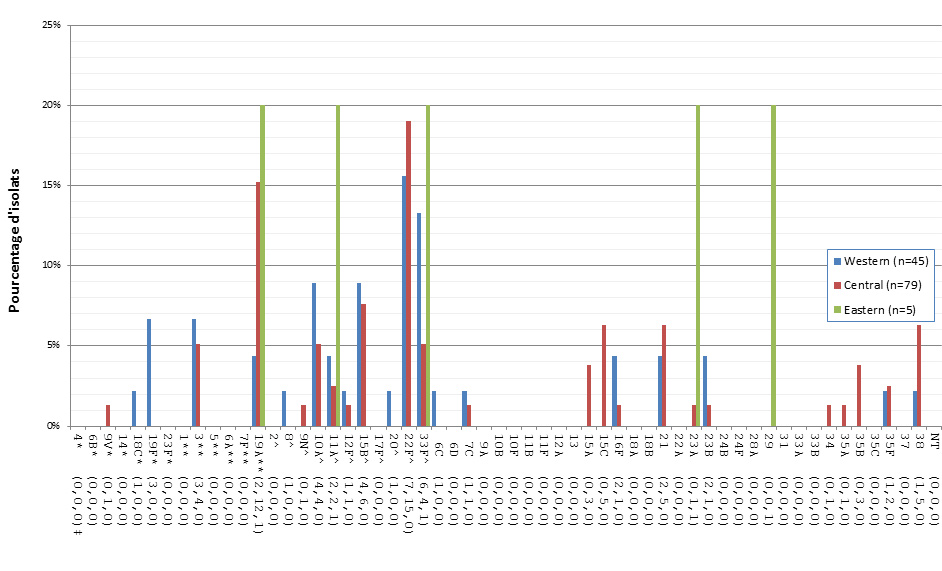
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les enfants de < 2 ans, 2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de < 2 ans distribués régionalement, recueillies en 2013.
| Sérotype | Ouest (n=45) | Centre (n=79) | Est (n=5) |
|---|---|---|---|
| 4* (0,0,0)‡ | 0.00% | 0.00% | 0.00% |
| 6B* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 9V* (0,1,0) | 0.00% | 1.30% | 0.00% |
| 14* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 18C* (1,0,0) | 2.20% | 0.00% | 0.00% |
| 19F* (3,0,0) | 6.70% | 0.00% | 0.00% |
| 23F* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 1** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 3** (3,4,0) | 6.70% | 5.10% | 0.00% |
| 5** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 6A** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7F** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 19A**(2,12,1) | 4.40% | 15.20% | 20.00% |
| 2^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 8^ (1,0,0) | 2.20% | 0.00% | 0.00% |
| 9N^ (0,1,0) | 0.00% | 1.30% | 0.00% |
| 10A^ (4,4,0) | 8.90% | 5.10% | 0.00% |
| 11A^ (2,2,1) | 4.40% | 2.50% | 20.00% |
| 12F^ (1,1,0) | 2.20% | 1.30% | 0.00% |
| 15B^ (4,6,0) | 8.90% | 7.60% | 0.00% |
| 17F^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 20^ (1,0,0) | 2.20% | 0.00% | 0.00% |
| 22F^ (7,15,0) | 15.60% | 19.00% | 0.00% |
| 33F^ (6,4,1) | 13.30% | 5.10% | 20.00% |
| 6C (1,0,0) | 2.20% | 0.00% | 0.00% |
| 6D (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7C (1,1,0) | 2.20% | 1.30% | 0.00% |
| 9A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 13 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 15A (0,3,0) | 0.00% | 3.80% | 0.00% |
| 15C (0,5,0) | 0.00% | 6.30% | 0.00% |
| 16F (2,1,0) | 4.40% | 1.30% | 0.00% |
| 18A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 21 (2,5,0) | 4.40% | 6.30% | 0.00% |
| 22A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 23A (0,1,1) | 0.00% | 1.30% | 20.00% |
| 23B (2,1,0) | 4.40% | 1.30% | 0.00% |
| 24B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 24F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 28A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 29 (0,0,1) | 0.00% | 0.00% | 20.00% |
| 31 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 33A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 33B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 34 (0,1,0) | 0.00% | 1.30% | 0.00% |
| 35A (0,1,0) | 0.00% | 1.30% | 0.00% |
| 35B (0,3,0) | 0.00% | 3.80% | 0.00% |
| 35C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35F (1,2,0) | 2.20% | 2.50% | 0.00% |
| 37 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 38 (1,5,0) | 2.20% | 6.30% | 0.00% |
| NT (0,0,0) | 0.00% | 0.00% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 21. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 2-4 ans, 2013
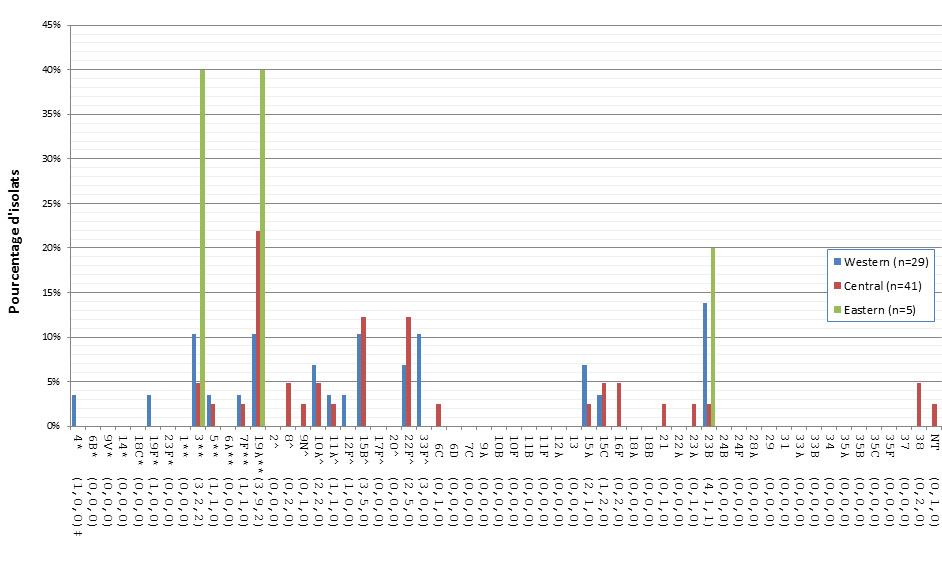
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 2-4 ans, 2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de 2 - enfants de 4 ans répartis régionalement, recueillies en 2013.
| Sérotype | Ouest (n=29) | Centre (n=41) | Est (n=5) |
|---|---|---|---|
| 4* (1,0,0)‡ | 3.40% | 0.00% | 0.00% |
| 6B* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 9V* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 14* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 18C* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 19F* (1,0,0) | 3.40% | 0.00% | 0.00% |
| 23F* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 1** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 3** (3,2,2) | 10.30% | 4.90% | 40.00% |
| 5** (1,1,0) | 3.40% | 2.40% | 0.00% |
| 6A** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7F** (1,1,0) | 3.40% | 2.40% | 0.00% |
| 19A**(3,9,2) | 10.30% | 22.00% | 40.00% |
| 2^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 8^ (0,2,0) | 0.00% | 4.90% | 0.00% |
| 9N^ (0,1,0) | 0.00% | 2.40% | 0.00% |
| 10A^ (2,2,0) | 6.90% | 4.90% | 0.00% |
| 11A^ (1,1,0) | 3.40% | 2.40% | 0.00% |
| 12F^ (1,0,0) | 3.40% | 0.00% | 0.00% |
| 15B^ (3,5,0) | 10.30% | 12.20% | 0.00% |
| 17F^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 20^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 22F^ (2,5,0) | 6.90% | 12.20% | 0.00% |
| 33F^ (3,0,0) | 10.30% | 0.00% | 0.00% |
| 6C (0,1,0) | 0.00% | 2.40% | 0.00% |
| 6D (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 9A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 13 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 15A (2,1,0) | 6.90% | 2.40% | 0.00% |
| 15C (1,2,0) | 3.40% | 4.90% | 0.00% |
| 16F (0,2,0) | 0.00% | 4.90% | 0.00% |
| 18A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 21 (0,1,0) | 0.00% | 2.40% | 0.00% |
| 22A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 23A (0,1,0) | 0.00% | 2.40% | 0.00% |
| 23B (4,1,1) | 13.80% | 2.40% | 20.00% |
| 24B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 24F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 28A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 29 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 31 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 33A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 33B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 34 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 37 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 38 (0,2,0) | 0.00% | 4.90% | 0.00% |
| NT (0,1,0) | 0.00% | 2.40% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 22. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 5-14 ans, 2013
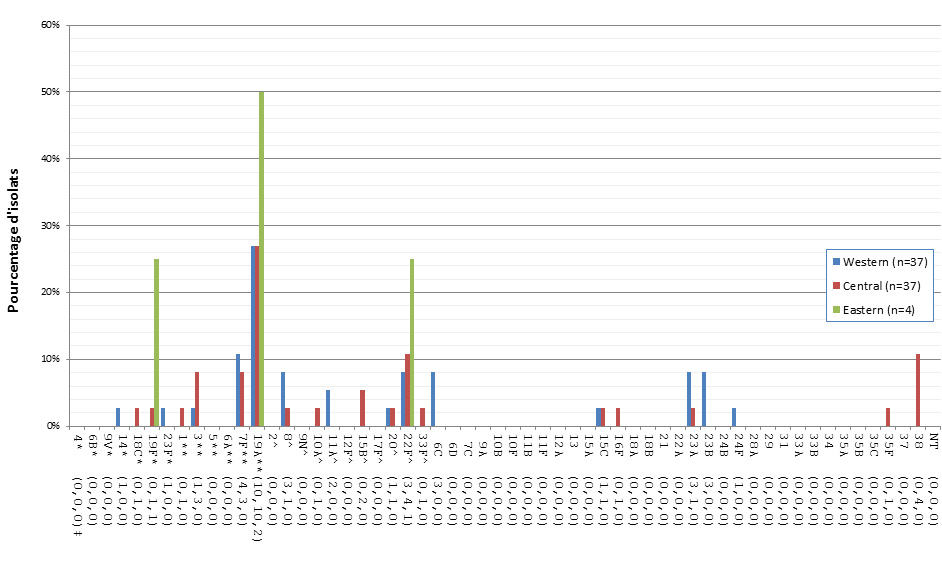
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les enfants de 5-14 ans, 2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de 5-14 ans distribués régionalement, recueillies en 2013.
| Sérotype | Ouest (n=37) | Centre (n=37) | Est (n=4) |
|---|---|---|---|
| 4* (0,0,0)‡ | 0.00% | 0.00% | 0.00% |
| 6B* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 9V* (0,0,0) | 0.00% | 0.00% | 0.00% |
| 14* (1,0,0) | 2.70% | 0.00% | 0.00% |
| 18C* (0,1,0) | 0.00% | 2.70% | 0.00% |
| 19F* (0,1,1) | 0.00% | 2.70% | 25.00% |
| 23F* (1,0,0) | 2.70% | 0.00% | 0.00% |
| 1** (0,1,0) | 0.00% | 2.70% | 0.00% |
| 3** (1,3,0) | 2.70% | 8.10% | 0.00% |
| 5** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 6A** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7F** (4,3,0) | 10.80% | 8.10% | 0.00% |
| 19A**(10,10,2) | 27.00% | 27.00% | 50.00% |
| 2^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 8^ (3,1,0) | 8.10% | 2.70% | 0.00% |
| 9N^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10A^ (0,1,0) | 0.00% | 2.70% | 0.00% |
| 11A^ (2,0,0) | 5.40% | 0.00% | 0.00% |
| 12F^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 15B^ (0,2,0) | 0.00% | 5.40% | 0.00% |
| 17F^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 20^ (1,1,0) | 2.70% | 2.70% | 0.00% |
| 22F^ (3,4,1) | 8.10% | 10.80% | 25.00% |
| 33F^ (0,1,0) | 0.00% | 2.70% | 0.00% |
| 6C (3,0,0) | 8.10% | 0.00% | 0.00% |
| 6D (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 9A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 13 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 15A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 15C (1,1,0) | 2.70% | 2.70% | 0.00% |
| 16F (0,1,0) | 0.00% | 2.70% | 0.00% |
| 18A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 21 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 22A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 23A (3,1,0) | 8.10% | 2.70% | 0.00% |
| 23B (3,0,0) | 8.10% | 0.00% | 0.00% |
| 24B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 24F (1,0,0) | 2.70% | 0.00% | 0.00% |
| 28A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 29 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 31 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 33A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 33B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 34 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35F (0,1,0) | 0.00% | 2.70% | 0.00% |
| 37 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 38 (0,4,0) | 0.00% | 10.80% | 0.00% |
| NT (0,0,0) | 0.00% | 0.00% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 23. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 15-49 ans, 2013
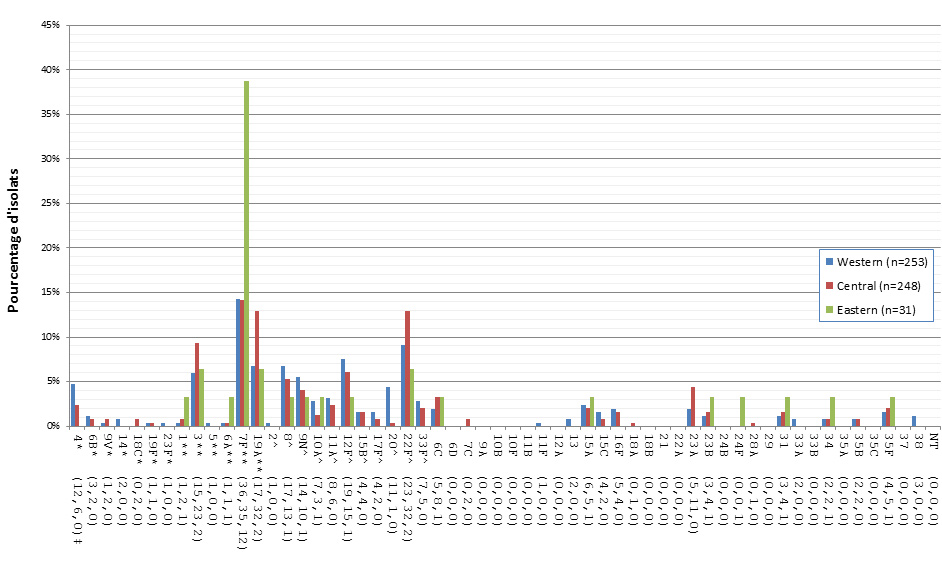
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 15-49 ans, 2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de 15-49 ans distribués régionalement, recueillies en 2013.
| Sérotype | Ouest (n=253) | Centre (n=248) | Est (n=31) |
|---|---|---|---|
| 4* (12,6,0)‡ | 4.70% | 2.40% | 0.00% |
| 6B* (3,2,0) | 1.20% | 0.80% | 0.00% |
| 9V* (1,2,0) | 0.40% | 0.80% | 0.00% |
| 14* (2,0,0) | 0.80% | 0.00% | 0.00% |
| 18C* (0,2,0) | 0.00% | 0.80% | 0.00% |
| 19F* (1,1,0) | 0.40% | 0.40% | 0.00% |
| 23F* (1,0,0) | 0.40% | 0.00% | 0.00% |
| 1** (1,2,1) | 0.40% | 0.80% | 3.20% |
| 3** (15,23,2) | 5.90% | 9.30% | 6.50% |
| 5** (1,0,0) | 0.40% | 0.00% | 0.00% |
| 6A** (1,1,1) | 0.40% | 0.40% | 3.20% |
| 7F** (36,35,12) | 14.20% | 14.10% | 38.70% |
| 19A**(17,32,2) | 6.70% | 12.90% | 6.50% |
| 2^ (1,0,0) | 0.40% | 0.00% | 0.00% |
| 8^ (17,13,1) | 6.70% | 5.20% | 3.20% |
| 9N^ (14,10,1) | 5.50% | 4.00% | 3.20% |
| 10A^ (7,3,1) | 2.80% | 1.20% | 3.20% |
| 11A^ (8,6,0) | 3.20% | 2.40% | 0.00% |
| 12F^ (19,15,1) | 7.50% | 6.00% | 3.20% |
| 15B^ (4,4,0) | 1.60% | 1.60% | 0.00% |
| 17F^ (4,2,0) | 1.60% | 0.80% | 0.00% |
| 20^ (11,1,0) | 4.30% | 0.40% | 0.00% |
| 22F^ (23,32,2) | 9.10% | 12.90% | 6.50% |
| 33F^ (7,5,0) | 2.80% | 2.00% | 0.00% |
| 6C (5,8,1) | 2.00% | 3.20% | 3.20% |
| 6D (0,0,0) | 0.00% | 0.00% | 0.00% |
| 7C (0,2,0) | 0.00% | 0.80% | 0.00% |
| 9A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11F (1,0,0) | 0.40% | 0.00% | 0.00% |
| 12A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 13 (2,0,0) | 0.80% | 0.00% | 0.00% |
| 15A (6,5,1) | 2.40% | 2.00% | 3.20% |
| 15C (4,2,0) | 1.60% | 0.80% | 0.00% |
| 16F (5,4,0) | 2.00% | 1.60% | 0.00% |
| 18A (0,1,0) | 0.00% | 0.40% | 0.00% |
| 18B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 21 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 22A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 23A (5,11,0) | 2.00% | 4.40% | 0.00% |
| 23B (3,4,1) | 1.20% | 1.60% | 3.20% |
| 24B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 24F (0,0,1) | 0.00% | 0.00% | 3.20% |
| 28A (0,1,0) | 0.00% | 0.40% | 0.00% |
| 29 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 31 (3,4,1) | 1.20% | 1.60% | 3.20% |
| 33A (2,0,0) | 0.80% | 0.00% | 0.00% |
| 33B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 34 (2,2,1) | 0.80% | 0.80% | 3.20% |
| 35A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35B (2,2,0) | 0.80% | 0.80% | 0.00% |
| 35C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35F (4,5,1) | 1.60% | 2.00% | 3.20% |
| 37 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 38 (3,0,0) | 1.20% | 0.00% | 0.00% |
| NT (0,0,0) | 0.00% | 0.00% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 24. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 50-64 ans, 2013
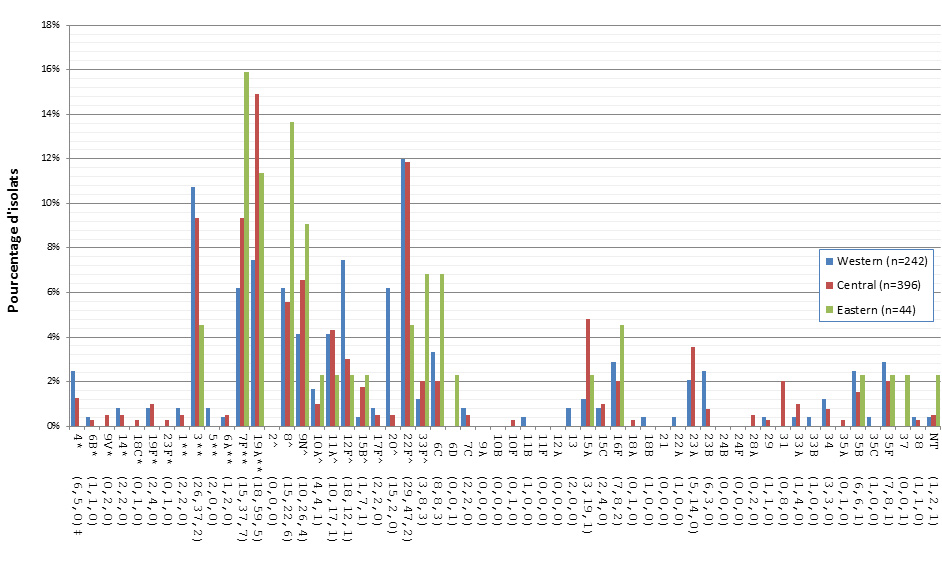
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les adultes de 50-64 ans, 2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de jeunes 50-64 ans distribués régionalement, recueillies en 2013.
| Sérotype | Ouest (n=242) | Centre (n=396) | Est (n=44) |
|---|---|---|---|
| 4* (6,5,0)‡ | 2.50% | 1.30% | 0.00% |
| 6B* (1,1,0) | 0.40% | 0.30% | 0.00% |
| 9V* (0,2,0) | 0.00% | 0.50% | 0.00% |
| 14* (2,2,0) | 0.80% | 0.50% | 0.00% |
| 18C* (0,1,0) | 0.00% | 0.30% | 0.00% |
| 19F* (2,4,0) | 0.80% | 1.00% | 0.00% |
| 23F* (0,1,0) | 0.00% | 0.30% | 0.00% |
| 1** (2,2,0) | 0.80% | 0.50% | 0.00% |
| 3** (26,37,2) | 10.70% | 9.30% | 4.50% |
| 5** (2,0,0) | 0.80% | 0.00% | 0.00% |
| 6A** (1,2,0) | 0.40% | 0.50% | 0.00% |
| 7F** (15,37,7) | 6.20% | 9.30% | 15.90% |
| 19A**(18,59,5) | 7.40% | 14.90% | 11.40% |
| 2^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 8^ (15,22,6) | 6.20% | 5.60% | 13.60% |
| 9N^ (10,26,4) | 4.10% | 6.60% | 9.10% |
| 10A^ (4,4,1) | 1.70% | 1.00% | 2.30% |
| 11A^ (10,17,1) | 4.10% | 4.30% | 2.30% |
| 12F^ (18,12,1) | 7.40% | 3.00% | 2.30% |
| 15B^ (1,7,1) | 0.40% | 1.80% | 2.30% |
| 17F^ (2,2,0) | 0.80% | 0.50% | 0.00% |
| 20^ (15,2,0) | 6.20% | 0.50% | 0.00% |
| 22F^ (29,47,2) | 12.00% | 11.90% | 4.50% |
| 33F^ (3,8,3) | 1.20% | 2.00% | 6.80% |
| 6C (8,8,3) | 3.30% | 2.00% | 6.80% |
| 6D (0,0,1) | 0.00% | 0.00% | 2.30% |
| 7C (2,2,0) | 0.80% | 0.50% | 0.00% |
| 9A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 10F (0,1,0) | 0.00% | 0.30% | 0.00% |
| 11B (1,0,0) | 0.40% | 0.00% | 0.00% |
| 11F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 12A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 13 (2,0,0) | 0.80% | 0.00% | 0.00% |
| 15A (3,19,1) | 1.20% | 4.80% | 2.30% |
| 15C (2,4,0) | 0.80% | 1.00% | 0.00% |
| 16F (7,8,2) | 2.90% | 2.00% | 4.50% |
| 18A (0,1,0) | 0.00% | 0.30% | 0.00% |
| 18B (1,0,0) | 0.40% | 0.00% | 0.00% |
| 21 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 22A (1,0,0) | 0.40% | 0.00% | 0.00% |
| 23A (5,14,0) | 2.10% | 3.50% | 0.00% |
| 23B (6,3,0) | 2.50% | 0.80% | 0.00% |
| 24B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 24F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 28A (0,2,0) | 0.00% | 0.50% | 0.00% |
| 29 (1,1,0) | 0.40% | 0.30% | 0.00% |
| 31 (0,8,0) | 0.00% | 2.00% | 0.00% |
| 33A (1,4,0) | 0.40% | 1.00% | 0.00% |
| 33B (1,0,0) | 0.40% | 0.00% | 0.00% |
| 34 (3,3,0) | 1.20% | 0.80% | 0.00% |
| 35A (0,1,0) | 0.00% | 0.30% | 0.00% |
| 35B (6,6,1) | 2.50% | 1.50% | 2.30% |
| 35C (1,0,0) | 0.40% | 0.00% | 0.00% |
| 35F (7,8,1) | 2.90% | 2.00% | 2.30% |
| 37 (0,0,1) | 0.00% | 0.00% | 2.30% |
| 38 (1,1,0) | 0.40% | 0.30% | 0.00% |
| NT (1,2,1) | 0.40% | 0.50% | 2.30% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 25. Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les personnes âgées de ≥ 65 ans, 2013
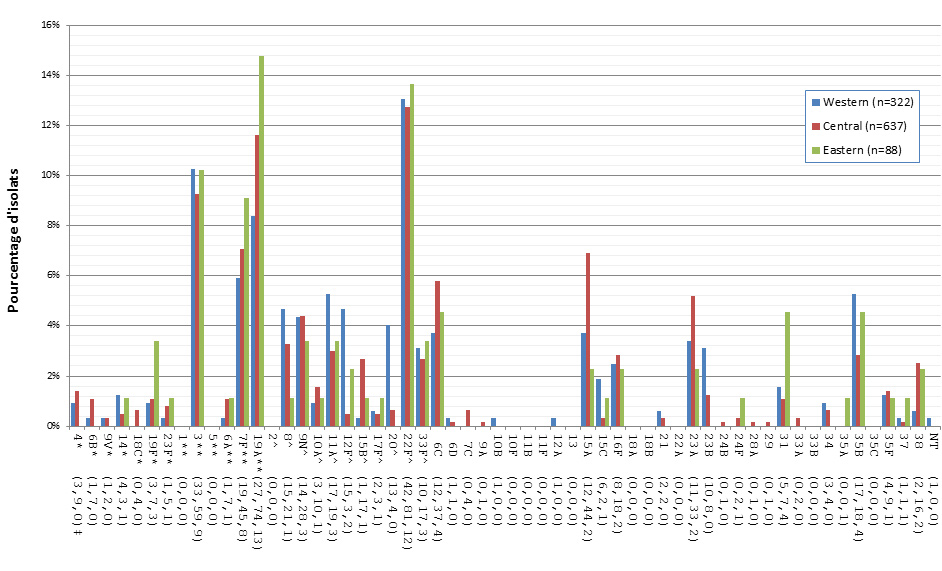
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement.
Distribution régionale des sérotypes de S. pneumoniae causant une infection invasive chez les personnes âgées de ≥ 65 ans, 2013 - Description textuel
Ce figure représente le nombre d'isolats de chaque sérotype de Olds ≥65 ans distribués régionalement, recueillies entre 2010-2013.
| Sérotype | Ouest (n=322) | Centre (n=637) | Est (n=88) |
|---|---|---|---|
| 4* (3,9,0)‡ | 0.90% | 1.40% | 0.00% |
| 6B* (1,7,0) | 0.30% | 1.10% | 0.00% |
| 9V* (1,2,0) | 0.30% | 0.30% | 0.00% |
| 14* (4,3,1) | 1.20% | 0.50% | 1.10% |
| 18C* (0,4,0) | 0.00% | 0.60% | 0.00% |
| 19F* (3,7,3) | 0.90% | 1.10% | 3.40% |
| 23F* (1,5,1) | 0.30% | 0.80% | 1.10% |
| 1** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 3** (33,59,9) | 10.20% | 9.30% | 10.20% |
| 5** (0,0,0) | 0.00% | 0.00% | 0.00% |
| 6A** (1,7,1) | 0.30% | 1.10% | 1.10% |
| 7F** (19,45,8) | 5.90% | 7.10% | 9.10% |
| 19A**(27,74,13) | 8.40% | 11.60% | 14.80% |
| 2^ (0,0,0) | 0.00% | 0.00% | 0.00% |
| 8^ (15,21,1) | 4.70% | 3.30% | 1.10% |
| 9N^ (14,28,3) | 4.30% | 4.40% | 3.40% |
| 10A^ (3,10,1) | 0.90% | 1.60% | 1.10% |
| 11A^ (17,19,3) | 5.30% | 3.00% | 3.40% |
| 12F^ (15,3,2) | 4.70% | 0.50% | 2.30% |
| 15B^ (1,17,1) | 0.30% | 2.70% | 1.10% |
| 17F^ (2,3,1) | 0.60% | 0.50% | 1.10% |
| 20^ (13,4,0) | 4.00% | 0.60% | 0.00% |
| 22F^ (42,81,12) | 13.00% | 12.70% | 13.60% |
| 33F^ (10,17,3) | 3.10% | 2.70% | 3.40% |
| 6C (12,37,4) | 3.70% | 5.80% | 4.50% |
| 6D (1,1,0) | 0.30% | 0.20% | 0.00% |
| 7C (0,4,0) | 0.00% | 0.60% | 0.00% |
| 9A (0,1,0) | 0.00% | 0.20% | 0.00% |
| 10B (1,0,0) | 0.30% | 0.00% | 0.00% |
| 10F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 11F (0,0,0) | 0.00% | 0.00% | 0.00% |
| 12A (1,0,0) | 0.30% | 0.00% | 0.00% |
| 13 (0,0,0) | 0.00% | 0.00% | 0.00% |
| 15A (12,44,2) | 3.70% | 6.90% | 2.30% |
| 15C (6,2,1) | 1.90% | 0.30% | 1.10% |
| 16F (8,18,2) | 2.50% | 2.80% | 2.30% |
| 18A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 18B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 21 (2,2,0) | 0.60% | 0.30% | 0.00% |
| 22A (0,0,0) | 0.00% | 0.00% | 0.00% |
| 23A (11,33,2) | 3.40% | 5.20% | 2.30% |
| 23B (10,8,0) | 3.10% | 1.30% | 0.00% |
| 24B (0,1,0) | 0.00% | 0.20% | 0.00% |
| 24F (0,2,1) | 0.00% | 0.30% | 1.10% |
| 28A (0,1,0) | 0.00% | 0.20% | 0.00% |
| 29 (0,1,0) | 0.00% | 0.20% | 0.00% |
| 31 (5,7,4) | 1.60% | 1.10% | 4.50% |
| 33A (0,2,0) | 0.00% | 0.30% | 0.00% |
| 33B (0,0,0) | 0.00% | 0.00% | 0.00% |
| 34 (3,4,0) | 0.90% | 0.60% | 0.00% |
| 35A (0,0,1) | 0.00% | 0.00% | 1.10% |
| 35B (17,18,4) | 5.30% | 2.80% | 4.50% |
| 35C (0,0,0) | 0.00% | 0.00% | 0.00% |
| 35F (4,9,1) | 1.20% | 1.40% | 1.10% |
| 37 (1,1,1) | 0.30% | 0.20% | 1.10% |
| 38 (2,16,2) | 0.60% | 2.50% | 2.30% |
| NT (1,0,0) | 0.30% | 0.00% | 0.00% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante de PPV23 ; ‡ Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. | |||
Figure 26. Sérotype 19A par région
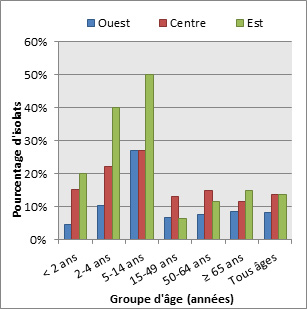
Sérotype 19A par région - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 19A pour tous les groupes d'âge répartis par région en 2013.
| Groupe d'âge | Ouest | Centre | Est |
|---|---|---|---|
| <2 yrs | 4.4% | 15.2% | 20.0% |
| 2 - 4 yrs | 10.3% | 22.0% | 40.0% |
| 5 - 14 yrs | 27.0% | 27.0% | 50.0% |
| 15 - 49 yrs | 6.7% | 12.9% | 6.5% |
| 50 - 64 yrs | 7.4% | 14.9% | 11.4% |
| ≥65 yrs | 8.4% | 11.6% | 14.8% |
| Tous âges | 8.2% | 13.5% | 13.7% |
Figure 27. Sérotype 7F par région

Sérotype 7F par région - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 7F pour tous les groupes d'âge répartis par région en 2013.
| Groupe d'âge | Ouest | Centre | Est |
|---|---|---|---|
| <2 yrs | 0.0% | 0.0% | 0.0% |
| 2 - 4 yrs | 3.4% | 2.4% | 0.0% |
| 5 - 14 yrs | 10.8% | 8.1% | 0.0% |
| 15 - 49 yrs | 14.2% | 14.1% | 38.7% |
| 50 - 64 yrs | 6.2% | 9.3% | 15.9% |
| ≥65 yrs | 5.9% | 7.1% | 9.1% |
| Tous âges | 8.1% | 8.4% | 15.4% |
Figure 28. Sérotype 3 par région
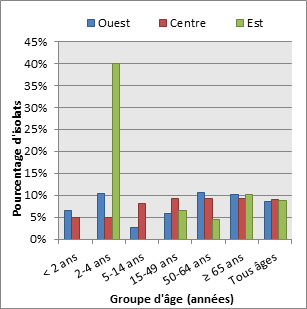
Sérotype 3 par région - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 3, pour tous les groupes d'âge répartis par région en 2013.
| Groupe d'âge | Ouest | Centre | Est |
|---|---|---|---|
| <2 yrs | 6.7% | 5.1% | 0.0% |
| 2 - 4 yrs | 10.3% | 4.9% | 40.0% |
| 5 - 14 yrs | 2.7% | 8.1% | 0.0% |
| 15 - 49 yrs | 5.9% | 9.3% | 6.5% |
| 50 - 64 yrs | 10.7% | 9.3% | 4.5% |
| ≥65 yrs | 10.2% | 9.3% | 10.2% |
| Tous âges | 8.7% | 9.0% | 8.8% |
Figure 29. Sérotype 22F par région
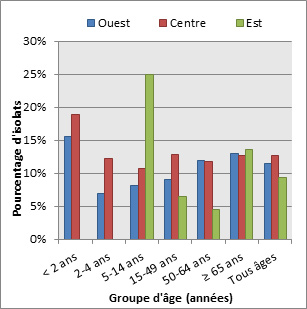
Sérotype 22F par région - Description textuel
Ce figure représente le pourcentage d'isolats au sein de sérotype 22F pour tous les groupes d'âge répartis par région en 2013.
| Groupe d'âge | Ouest | Centre | Est |
|---|---|---|---|
| <2 yrs | 15.6% | 19.0% | 0.0% |
| 2 - 4 yrs | 6.9% | 12.2% | 0.0% |
| 5 - 14 yrs | 8.1% | 10.8% | 25.0% |
| 15 - 49 yrs | 9.1% | 12.9% | 6.5% |
| 50 - 64 yrs | 12.0% | 11.9% | 4.5% |
| ≥65 yrs | 13.0% | 12.7% | 13.6% |
| Tous âges | 11.5% | 12.8% | 9.3% |
Sérotypes vaccinaux
Le nombre total de sérotypes contenus dans le vaccin Pneu-C-7 (4, 6B, 9V 14, 18C 19F, 23F) est très faible dans les groupes d’âge de < 15 ans; toutefois, les proportions ont continué de diminuer dans la plupart des groupes d’âge en 2013, sauf chez les enfants de 2-4 ans, pour qui les proportions sont passées de 1,6 % (n = 2) en 2012 à 2,7 % (n = 2) en 2013, et chez les enfants de 5-14 ans, pour qui les proportions ont passé de 4,8 % (n = 5) à 6,4 % (n = 5) (figure 32).
Au Canada, la proportion des sérotypes contenus dans le vaccin Pneu-C-13 (1, 3, 5, 6A, 7F et 19A) a continué de diminuer dans tous les groupes d’âge combinés, passant de 36,3 % (n = 952) en 2012 à 30,4 % (n = 783) en 2013 (figure 31). Cette même année, la proportion la plus élevée des sérotypes du vaccin Pneu-C-13 a été observée dans les régions de l’Est, soit 39,6 % (n = 72), une légère baisse par rapport à 2012, où cette proportion était de 42,1 % (n = 75) (figure 30). On a observé une diminution des sérotypes contenus dans le vaccin Pneu-C-13 dans tous les groupes d’âge entre 2012 et 2013, ce qui est généralement attribuable à la réduction de la proportion des sérotypes 7F et 19A. En 2013, les sérotypes contenus dans le vaccin Pneu-C-13 représentaient 17,1 % (n = 22) des isolats chez les enfants de < 2 ans, 33,3 % (n = 27) chez les enfants de 2-4 ans, 43,6 % (n = 34) chez les enfants de 5-14 ans, 34,2 % (n = 182) chez les adultes de 15-49 ans, 31,5 % (n = 215) chez les adultes de 50-64 ans et 28,3 % (n = 296) chez les personnes âgées de ≥ 65 ans (figure 33).
La proportion globale des sérotypes contenus dans les vaccins Pneu-C-7 et Pneu-C-13 a connu une diminution, passant de 42,5 % (n = 1 114) en 2012 à 35,4 % (n = 911) en 2013 (figure 31). En 2013, le grand total des sérotypes contenus dans les vaccins Pneu-C-7 et Pneu-C-13 représentait 42,9 % (n = 78) des isolats prélevés dans l’Est, 36,9 % (n = 536) des isolats du Centre et 31,6 % (n = 297) des isolats provenant de l’Ouest. Dans les régions du Centre et de l’Ouest, la proportion des sérotypes contenus dans les vaccins Pneu-C-7 et Pneu-C-13 a décliné, passant de 40,5 % (n = 586) à 32,1 % (n = 466) et de 29,3 % (n = 291) à 26,0 % (n = 245), respectivement (figure 30).
La proportion d’isolats représentant les sérotypes contenus dans le vaccin Pneu-P-23 (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20, 22F, 33F) a augmenté dans les isolats pour tous les groupes d’âge combinés, passant de 24,7 % (n = 670) en 2010 à 37,6 % (n = 970) en 2013 (figure 31). La hausse la plus marquée en 2013 a été observée chez les enfants de < 2 ans, pour lesquels la proportion est passée de 26,7 % (n = 31) en 2012 à 47,3 % (n = 61) en 2013. De 2012 à 2013, la proportion des sérotypes du vaccin Pneu-P-23 dans les isolats prélevés chez les 2-4 ans a connu une augmentation de 26,8 % (n = 33) à 37,3 % (n=28); chez les 5-14 ans, elle a diminué, passant de 2,9 % (n = 24) à 2,1 % (n = 24); chez les 15-49 ans, elle est passée de 35,6 % (n = 218) à 39,8 % (n = 212); chez les 50-64 ans, de 34,0 % (n = 237) à 40,0 % (n = 273); et chez les personnes âgées de ≥ 65 ans, de 30,4 % (n = 286) à 36,6 % (n = 362), respectivement (figure 34).
| VaccinFootnote * | Groupe d'âge (N) | ||||||
|---|---|---|---|---|---|---|---|
| <2 | 2-4 | 5-14 | 15-19 | 50-64 | ≥65 | Tous âgesFootnote ** | |
| Pneu-C-7 | 3,9% (5) | 2,7% (2) | 6,4% (5) | 6,2% (33) | 4% (27) | 5,3% (55) | 5% (128) |
| Pneu-C-13 | 17,1% (22) | 33,3% (25) | 43,6% (34) | 34,2% (182) | 31,5% (215) | 28,3% (296) | 30,4% (783) |
| Pneu-C-13 Tous | 20,9% (27) | 36% (27) | 50% (39) | 40,4% (215) | 35,5% (242) | 33,5% (351) | 35,4% (911) |
| Pneu-P-23 | 47,3% (61) | 37,3% (28) | 25,6% (20) | 39,8% (212) | 40% (273) | 34,6% (362) | 37,6% (970) |
| Pneu-P-23 Tous | 68,2% (88) | 73,3% (55) | 75,6% (59) | 79,7% (424) | 75,1% (512) | 67,2% (704) | 70,6% (24) |
| SNIV Total | 31,8% (41) | 26,7% (20) | 24,4% (19) | 19,7% (105) | 24,5% (167) | 31,9% (334) | 27% (696) |
| Tous | (129) | (75) | (78) | (532) | (682) | (1047) | (2577) |
Figure 30. Sérotypes vaccinaux par région, 2013
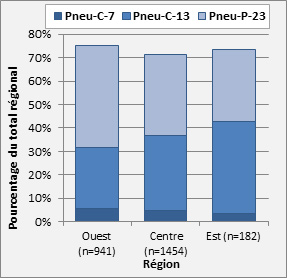
Sérotypes vaccinaux par région, 2013 - Description textuel
Ce figure représente les sérotypes vaccinaux de pourcentage trouvés dans chaque région en 2013.
| Sérotypes | Ouest (n=941) | Centre (n=1454) | Est (n=182) |
|---|---|---|---|
| Pneu-C-7 | 5.5% | 4.8% | 3.3% |
| Pneu-C-13 | 26.0% | 32.0% | 39.6% |
| Pneu-P-23 | 43.6% | 34.7% | 30.8% |
Figure 31. Sérotypes vaccinaux 2010-2013
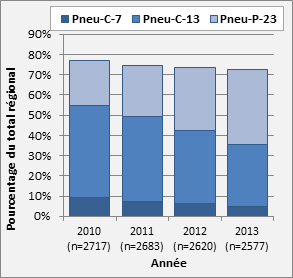
Sérotypes vaccinaux 2010-2013 - Description textuel
Ce figure représente le pourcentage des sérotypes vaccinaux par année , de 2010 à 2013.
| Sérotypes | 2010 (n=2717) | 2011 (n=2683) | 2012 (n=2620) | 2013 (n=2577) |
|---|---|---|---|---|
| Pneu-C-7 | 9.5% | 7.5% | 6.2% | 5.0% |
| Pneu-C-13 | 45.5% | 42.0% | 36.3% | 30.4% |
| Pneu-P-23 | 22.0% | 25.0% | 30.9% | 37.1% |
Figure 32. Sérotypes (4, 6B, 9V, 14, 18C, 19F, 23F) du vaccin Pneu-C-7 par groupe d'âge, 2010-2013

Sérotypes (4, 6B, 9V, 14, 18C, 19F, 23F) du vaccin Pneu-C-7 par groupe d'âge, 2010-2013 - Description textuel
Ce figure représente le pourcentage des sérotypes PCV7 par groupes d'âge et par année de 2010 à 2013.
| Année | < 2 ans | 2 à 4 ans | 5 à 14 ans | 15 à 49 ans | 50 à 64 ans | ≥65 ans |
|---|---|---|---|---|---|---|
| 2010 | 5.4% | 3.4% | 10.0% | 12.8% | 8.5% | 9.7% |
| 2011 | 1.9% | 3.0% | 4.7% | 9.9% | 9.1% | 6.6% |
| 2012 | 4.3% | 1.6% | 4.8% | 7.5% | 5.6% | 6.8% |
| 2013 | 3.9% | 2.7% | 6.4% | 6.2% | 4.0% | 5.3% |
| Groupe d'âge (Années) | Année | |||
|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | |
| <2 | 10 | 3 | 5 | 5 |
| 2 - 4 | 5 | 4 | 2 | 2 |
| 5 - 14 | 11 | 6 | 5 | 5 |
| 15 - 49 | 75 | 62 | 46 | 33 |
| 50 - 64 | 59 | 63 | 39 | 27 |
| ≥65 | 94 | 60 | 64 | 55 |
Figure 33. Sérotypes (1, 5, 7F, 3, 6A, 19A) du vaccin Pneu-C-13 par groupe d'âge, 2010-2013

Sérotypes (1, 5, 7F, 3, 6A, 19A) du vaccin Pneu-C-13 par groupe d'âge, 2010-2013 - Description textuel
Ce figure représente le pourcentage de sérotypes de PCV13 par groupe d'âge et par année de 2010 à 2013.
| Année | < 2 ans | 2 à 4 ans | 5 à 14 ans | 15 à 49 ans | 50 à 64 ans | ≥65 ans |
|---|---|---|---|---|---|---|
| 2010 | 57.6% | 67.6% | 52.7% | 45.9% | 43.2% | 40.6% |
| 2011 | 41.0% | 63.4% | 55.5% | 43.5% | 41.0% | 36.8% |
| 2012 | 38.8% | 38.2% | 55.2% | 39.5% | 35.4% | 32.8% |
| 2013 | 17.1% | 33.3% | 43.6% | 34.2% | 31.5% | 28.3% |
| Groupe d'âge (Années) | Année | |||
|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | |
| <2 | 106 | 66 | 45 | 22 |
| 2 - 4 | 100 | 85 | 47 | 25 |
| 5 - 14 | 58 | 71 | 58 | 34 |
| 15 - 49 | 268 | 273 | 242 | 182 |
| 50 - 64 | 299 | 284 | 247 | 215 |
| ≥65 | 394 | 337 | 308 | 296 |
Figure 34. Sérotypes (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20, 22F, 33F) du vaccin Pneu P-23 par groupe d'âge, 2010-2013
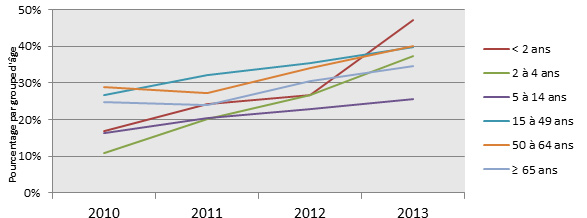
Sérotypes (2, 8, 9N, 10A, 11A, 12F, 15B, 17F, 20, 22F, 33F) du vaccin Pneu P-23 par groupe d'âge, 2010-2013 - Description textuel
Ce figure représente le pourcentage de sérotypes de PCV23 par groupe d'âge et par année de 2010 à 2013.
| Année | < 2 ans | 2 à 4 ans | 5 à 14 ans | 15 à 49 ans | 50 à 64 ans | ≥65 ans |
|---|---|---|---|---|---|---|
| 2010 | 16.8% | 10.8% | 16.4% | 26.7% | 28.9% | 24.7% |
| 2011 | 24.2% | 20.1% | 20.3% | 32.2% | 27.3% | 23.9% |
| 2012 | 26.7% | 26.8% | 22.9% | 35.6% | 34.0% | 30.4% |
| 2013 | 47.3% | 37.3% | 25.6% | 39.8% | 40.0% | 34.6% |
| Groupe d'âge (Années) | Année | |||
|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | |
| <2 | 31 | 39 | 31 | 61 |
| 2 - 4 | 16 | 27 | 33 | 28 |
| 5 - 14 | 18 | 26 | 24 | 20 |
| 15 - 49 | 156 | 202 | 218 | 212 |
| 50 - 64 | 200 | 189 | 237 | 273 |
| ≥65 | 240 | 219 | 286 | 362 |
Figure 35. Sérotypes non inclus dans les vaccins par groupe d'âge, 2010-2013
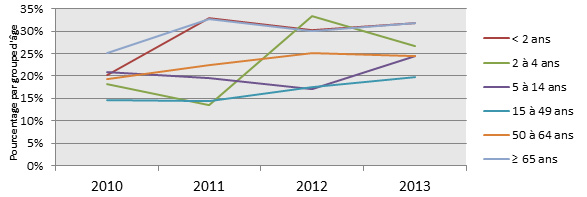
Sérotypes non inclus dans les vaccins par groupe d'âge, 2010-2013 - Description textuel
Ce figure représente le pourcentage des sérotypes non vaccinaux par groupe d'âge et par année de 2010 à 2013.
| Année | < 2 ans | 2 à 4 ans | 5 à 14 ans | 15 à 49 ans | 50 à 64 ans | ≥65 ans |
|---|---|---|---|---|---|---|
| 2010 | 20.1% | 18.2% | 20.9% | 14.6% | 19.4% | 25.0% |
| 2011 | 32.9% | 13.4% | 19.5% | 14.5% | 22.5% | 32.7% |
| 2012 | 30.2% | 33.3% | 17.1% | 17.5% | 25.1% | 30.0% |
| 2013 | 31.8% | 26.7% | 24.4% | 19.7% | 24.5% | 31.9% |
| Groupe d'âge (Années) | Année | |||
|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | |
| <2 | 37 | 53 | 35 | 41 |
| 2 - 4 | 27 | 18 | 41 | 20 |
| 5 - 14 | 23 | 25 | 18 | 19 |
| 15 - 49 | 85 | 91 | 107 | 105 |
| 50 - 64 | 134 | 156 | 175 | 167 |
| ≥65 | 243 | 299 | 282 | 334 |
Résistance de S. pneumoniae aux antimicrobiens
En 2013, dans le cadre d’un programme conjoint entre le LNM et la CARA appelé SAVE, 1 061 isolats de S. pneumoniae ayant causé une infection invasive ont fait l’objet de tests de sensibilité aux antimicrobiens. Ces isolats, prélevés chez des sujets de tous les groupes d’âge, ont été soumis par huit provinces participantes (Saskatchewan, Manitoba, Ontario, Québec, Nouvelle-Écosse, Île-du-Prince-Édouard, Nouveau-Brunswick, Terre-Neuve-et-Labrador).
Les taux élevés de résistance aux antimicrobiens concernaient la clarithromycine en 2013 (24,8 %, n = 263), une légère augmentation par rapport aux taux de 2011 (21,4 %, n = 242). Les taux les moins élevés touchaient la pénicilline en fonction des doses seuils applicables à la méningite, qui sont passés de 11,9 % (n = 135) en 2011 à 10,0 % (n = 106) en 2013. Le taux de résistance à la clindamycine a également décliné depuis 2011, passant de 7,1 % (n = 80) à 5,9 % (n = 62) en 2013. La résistance à la doxycycline est demeurée relativement stable depuis 2010, à un taux de 9,9 % (n = 105) en 2013, tandis que le taux de résistance de S. pneumoniae au triméthoprime-sulfaméthoxazole a augmenté, passant de 5,1 % (n = 58) à 7,4 % (n = 78) au cours de la même période. On a également observé une légère augmentation des taux de résistance à l’imipénème et au méropénème entre 2011 et 2013, c’est-à-dire de 1,0 % (n = 11) à 2,3 % (n = 24) et de 2,0 % (n = 23) à 2,6 % (n = 28), respectivement. Aucun isolat n’était non sensible au linézolide, à la tigécycline ou à la vancomycine (figure 36). Selon une comparaison régionale des taux de résistance de S. pneumoniae aux antimicrobiens au cours de l’année 2013, l’Ouest du Canada présentait des taux de résistance légèrement plus élevés à la clarithromycine (29,8 %, n = 73), à la pénicilline en fonction des doses seuils applicables à la méningite (11,0 %, n = 27) et au triméthoprime-sulfaméthoxazole (9,4 %, n=23), tandis que la résistance à la clindamycine et à la doxycycline était plus fréquente dans les régions du Centre (6,5 %, n = 44 et 11,5 %, n = 78, respectivement) (figure 37).
En 2013, les isolats du sérotype 15A présentaient généralement les taux de résistance les plus élevés, entre autres à la pénicilline en fonction des doses seuils applicables à la méningite (40,9 %, n = 9), à la clarithromycine (77,3 %, n = 17), à la clindamycine (31,8 %, n = 7) et à la doxycycline (72,7 %, n = 16) (tableau 8).
Au cours de cette même année, la résistance à la clarithromycine était associée aux sérotypes 19A (59,6 %, n = 68), 10A (22,6 %, n = 7), 12F (81,1 %, n = 43), 15A (77,3 %, n = 17), 22F (28,0 %, n = 33) et 33F (81,6 %, n = 40) (figure 40). Les sérotypes de S. pneumoniae résistants au céfuroxime administré par voie parentérale étaient principalement 19A (26,3 %, n = 30) et 35B (21,4 %, n = 6), et des taux plus élevés de résistance à la clindamycine ont été relevés pour les sérotypes 19A (26,3 %, n = 30), 33F (16,3 %, n = 8) et 15A (31,8 %, n = 7). Parmi les sérotypes résistants à la doxycycline figuraient 19A (33,3 %, n = 38) et 15A (72,7 %, n = 16). La résistance à l’imipénème et au méropénème était principalement associée aux isolats du sérotype 19A (20,2 %, n = 23 et 21,1 %, n = 24, respectivement). Des taux relativement élevés de résistance à la pénicilline (méningite) étaient notables pour les sérotypes 6A (60,0 %, n = 3), 6C (33,3 %, n = 11), 15A (40,9 %, n = 9), 19A (28,9 %, n = 33), 23A (30,0 %, n = 9), 23B (38,1 %, n = 8) et 35B (25,0 %, n = 7). La résistance au triméthoprime-sulfaméthoxazole était associée aux sérotypes 19A (27,2 %, n = 31) et 6C (21,2 %, n = 7) (figure 47).
On a observé une multirésistance à 3 classes ou plus d’antimicrobiens dans 7,5 % (n = 80) des isolats de S. pneumoniae analysés en 2013, une légère diminution par rapport au taux de 8,0 % (n = 91) observé en 2011 (figure 38). Les sérotypes affichant les taux les plus élevés de multirésistance au cours de l’année 2013 étaient les suivants : 15A (n = 11), dont 50,0 % des isolats étaient résistants à 3 classes ou plus d’antimicrobiens (figure 39), ce qui représente une diminution par rapport à 2011 (74,1 %, n = 20); 19A, à 28,9 % (n = 33), un taux similaire à celui de 2011 (29,1 %, n = 43); 6C, à 24,2 % (n = 8), une hausse par rapport à 2011 (8,7 %, n = 4); et 19F, à 23,1 % (n = 3), le même pourcentage qu’en 2011. Le principal profil de multirésistance dans le cas des isolats du sérotype 15A était β-lactamine-macrolide-clindamycine-tétracycline, pour les sérotypes 19A et 19F, il s’agissait de β-lactamine-macrolide-clindamycine-tétracycline- triméthoprime-sulfaméthoxazole et pour le sérotype 6C, β-lactamine-macrolide-triméthoprime-sulfaméthoxazole. On a observé une résistance aux 6 classes d’antimicrobiens dans un isolat du sérotype 23F.
Figure 36. Résistance des isolats de S. pneumoniae aux antimicrobiens, 2011-2013
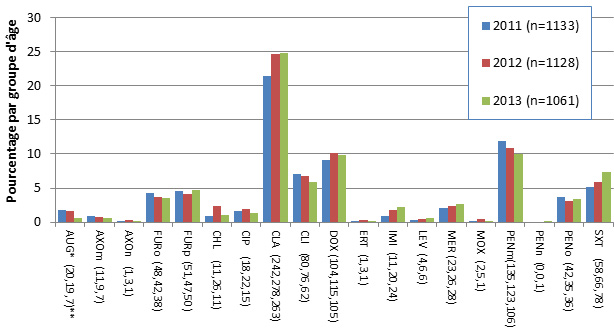
*AUG = amoxicilline/acide clavulanique; PENm = pénicilline en fonction de la norme d'interprétation du CLSI pour la dose parentérale en cas de méningite; PENn = pénicilline en fonction de la norme d'interprétation pour la dose parentérale dans les cas autres que la méningite; PENo = pénicilline en fonction de la norme d'interprétation pour la dose orale de la pénicilline V; LEV = lévofloxacine; MOX = moxifloxacine; AXOm = ceftriaxone en fonction de la norme d'interprétation pour la dose parentérale en cas de méningite; AXOn = ceftriaxone en fonction de la norme d'interprétation pour la dose parentérale dans les cas autres que la méningite; FURo = céfuroxime en fonction de la norme d'interprétation pour la dose orale; FURp = céfuroxime en fonction de la norme d'interprétation pour la dose parentérale; ETP = ertapénème; IMI = imipénème; MER = méropénème; CLA = clarithromycine; CLI = clindamycine; CHL = chloramphénicol; DOX = doxycycline; SXT = triméthoprime sulfaméthoxazole. On n'a observé aucune non-sensibilité dans les cas suivants : daptomycine (aucune norme d'interprétation), linézolide, tigécycline (aucune norme d'interprétation) et vancomycine. On s'est servi des seuils d'interprétation EUCAST [EUCAST, 2012] pour la DOX; dans tous les autres cas, on a employé les seuls CLSI [CLSI, 2011].**Nombre d'isolats en 2011, 2012 et 2013, respectivement.
Résistance des isolats de S. pneumoniae aux antimicrobiens, 2011-2013 - Description textuel
Ce figure représente le pourcentage d'isolats résistants aux antibiotiques par année et en 2011-2013.
| Région | AUGFootnote * (20,19,7)Footnote ** |
AXOm (11,9,7) |
AXOn (1,3,1) |
FURo (48,42,38) |
FURp (51,47,50) |
CHL (11,26,11) |
CIP (18,22,15) |
CLA (242,278,263) |
CLI (80,76,62) |
DOX (104,115,105) |
ERT (1,3,1) |
IMI (11,20,24) |
LEV (4,6,6) |
MER (23,26,28) |
MOX (2,5,1) |
PENm (135,123,106) |
PENn (0,0,1) |
PENo (42,35,36) |
SXT (58,66,78) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2011 (n=1133) | 1.7% | 1.0% | 0.1% | 4.2% | 4.5% | 1.0% | 1.6% | 21.4% | 7.1% | 9.2% | 0.1% | 0.1% | 0.4% | 2.0% | 0.2% | 11.9% | 0.0% | 3.7% | 5.4% |
| 2012 (n=1128) | 1.7% | 0.8% | 0.3% | 3.7% | 4.2% | 2.3% | 2.0% | 24.6% | 6.7% | 10.2% | 0.3% | 1.8% | 0.5% | 2.3% | 0.4% | 10.9% | 0.0% | 3.1% | 5.9% |
| 2013 (n=1061) | 0.7% | 0.7% | 0.1% | 3.6% | 4.7% | 1.0% | 1.4% | 24.8% | 5.8% | 9.9% | 0.1% | 2.3% | 0.6% | 2.6% | 0.1% | 10.0% | 0.1% | 3.4% | 7.4% |
Figure 37. Résistance de S. pneumoniae aux antimicrobiens par région, 2013

*AUG = amoxicilline/acide clavulanique; PENm = pénicilline en fonction de la norme d'interprétation du CLSI pour la dose parentérale en cas de méningite; PENn = pénicilline en fonction de la norme d'interprétation pour la dose parentérale dans les cas autres que la méningite; PENo = pénicilline en fonction de la norme d'interprétation pour la dose orale de la pénicilline V; LEV = lévofloxacine; MOX = moxifloxacine; AXOm = ceftriaxone en fonction de la norme d'interprétation pour la dose parentérale en cas de méningite; AXOn = ceftriaxone en fonction de la norme d'interprétation pour la dose parentérale dans les cas autres que la méningite; FURo = céfuroxime en fonction de la norme d'interprétation pour la dose orale; FURp = céfuroxime en fonction de la norme d'interprétation pour la dose parentérale; ETP = ertapénème; IMI = imipénème; MER = méropénème; CLA = clarithromycine; CLI = clindamycine; CHL = chloramphénicol; DOX = doxycycline; SXT = triméthoprime sulfaméthoxazole. On n'a observé aucune non-sensibilité dans les cas suivants : daptomycine (aucune norme d'interprétation), linézolide, tigécycline (aucune norme d'interprétation) et vancomycine. On s'est servi des seuils d'interprétation EUCAST [EUCAST, 2012] pour la DOX; dans tous les autres cas, on a employé les seuls CLSI [CLSI, 2011].**Nombre d'isolats en 2011, 2012 et 2013, respectivement.
Résistance de S. pneumoniae aux antimicrobiens par région, 2013 - Description textuel
Ce figure représente le pourcentage d'isolats résistants par antibiotiques et par région pour 2013.
| Région | AUGFootnote * (2,2,3)Footnote ** |
AXOm (2,3,2) |
AXOn (0,0,1) |
FURo (9,25,4) |
FURp (13,31,6) |
CHL (0,10,1) |
CIP (1,14,0) |
CLA (73,156,34) |
CLI (13,44,5) |
DOX (14,78,13) |
ERT (0,0,1) |
IMI (6,14,4) |
LEV (1,5,0) |
MER (6,18,4) |
MOX (0,1,0) |
PENm (27,69,10) |
PENn (0,1,0) |
PENo (7,25,4) |
SXT (23,47,8) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ouest (n=245) | 0.8% | 0.8% | 0.0% | 3.7% | 5.3% | 0.0% | 0.4% | 29.8% | 5.3% | 5.7% | 0.0% | 2.4 | 0.4% | 2.4% | 0.0% | 11.0% | 0.0% | 2.9% | 9.4% |
| Centre (n=676) | 0.3% | 0.4% | 0.0% | 3.7% | 4.6% | 1.5% | 2.1% | 23.0% | 6.5% | 11.5% | 0.0% | 2.1% | 0.7% | 2.7% | 0.1% | 10.2% | 0.1% | 3.7% | 7.0% |
| Est (n=140) | 2.1% | 1.4% | 0.7% | 2.9% | 4.3% | 0.7% | 0.0% | 24.3% | 3.6% | 9.3% | 0.7% | 2.9% | 0.0% | 2.9% | 0.0% | 7.1% | 0.0% | 2.9% | 5.7% |
Figure 38. Profils de multirésistance de S. pneumoniae, 2011-2013
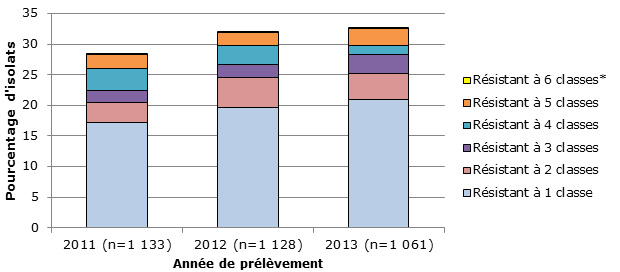
*Les classes d'antimicrobiens sont les suivantes: β-lactamines (amoxicilline/acide clavulanique, pénicilline en fonction des doses seuils pour la méningite, ceftriaxone en fonction des doses seuils pour la méningite, céfuroxime en fonction de la dose seuil parentérale, ertapénème, imipénème et méropénème); macrolides (clarithromycine); fluoroquinolones (lévofloxacine et moxifloxacine); tétracyclines (doxycycline); inhibiteurs de la voie folate (triméthoprime-sulfaméthoxazole); phénicols (chloramphénicol); lincosamides (clindamycine); oxazolidinones (linézolide).
Profils de multirésistance de S. pneumoniae, 2011-2013 - Description textuel
Ce figure représente le pourcentage d'isolats qui sont résistants à une ou plusieurs classes de médicaments par année de 2011 à 2013.
| Année | Résistant à 1 classe | Résistant à 2 classes | Résistant à 3 classes | Résistant à 4 classes | Résistant à 5 classes | Résistant à 6 classesFootnote * |
|---|---|---|---|---|---|---|
| 2011 (n=1133) | 17.1% | 3.3% | 2.0% | 3.6% | 2.3% | 0.1% |
| 2012 (n=1128) | 19.6% | 4.9% | 2.1% | 3.1% | 2.2% | 0.2% |
| 2013 (n=1061) | 20.9% | 4.2% | 3.2% | 1.4% | 2.8% | 0.1% |
Figure 39. Profils de multirésistance des sérotypes de S. pneumoniae, 2013
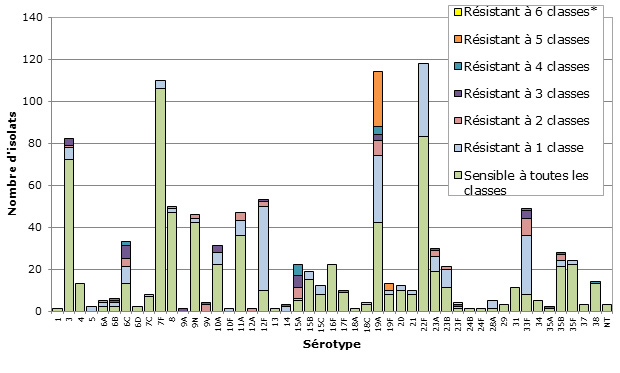
* Les classes d'antimicrobiens sont les suivantes: β-lactamines (amoxicilline/acide clavulanique, pénicilline en fonction des doses seuils pour la méningite, ceftriaxone en fonction des doses seuils pour la méningite, céfuroxime en fonction de la dose seuil parentérale, ertapénème, imipénème et méropénème); macrolides (clarithromycine); fluoroquinolones (lévofloxacine et moxifloxacine); tétracyclines (doxycycline); inhibiteurs de la voie folate (triméthoprime-sulfaméthoxazole); phénicols (chloramphénicol); lincosamides (clindamycine); oxazolidinones (linézolide).
Profils de multirésistance des sérotypes de S. pneumoniae, 2013 - Description textuel
Ce figure représente le nombre d'isolats résistants à une ou plusieurs classes de médicaments par sérotype.
| Sérotype | Sensible à toutes les classes | Résistant à 1 classe | Résistant à 2 classes | Résistant à 3 classes | Résistant à 4 classes | Résistant à 5 classes | Résistant à 6 classesFootnote * |
|---|---|---|---|---|---|---|---|
| 1 | 1 | ||||||
| 3 | 72 | 6 | 1 | 3 | |||
| 4 | 13 | ||||||
| 5 | 2 | ||||||
| 6A | 2 | 2 | 1 | ||||
| 6B | 2 | 2 | 1 | 1 | |||
| 6C | 13 | 8 | 4 | 6 | 2 | ||
| 6D | 2 | ||||||
| 7C | 7 | 1 | |||||
| 7F | 106 | 4 | |||||
| 8 | 47 | 2 | 1 | ||||
| 9A | 1 | ||||||
| 9N | 42 | 2 | 2 | ||||
| 9V | 3 | 1 | |||||
| 10A | 22 | 6 | 3 | ||||
| 10F | 1 | ||||||
| 11A | 36 | 7 | 4 | ||||
| 12A | 1 | ||||||
| 12F | 10 | 40 | 2 | 1 | |||
| 13 | 1 | ||||||
| 14 | 2 | 1 | |||||
| 15A | 5 | 1 | 5 | 6 | 5 | ||
| 15B | 15 | 4 | |||||
| 15C | 8 | 4 | |||||
| 16F | 22 | ||||||
| 17F | 9 | 1 | |||||
| 18A | 1 | ||||||
| 18C | 3 | 1 | |||||
| 19A | 42 | 32 | 7 | 3 | 4 | 26 | |
| 19F | 8 | 2 | 3 | ||||
| 20 | 10 | 2 | |||||
| 21 | 8 | 2 | |||||
| 22F | 83 | 35 | |||||
| 23A | 19 | 7 | 3 | 1 | |||
| 23B | 11 | 9 | 1 | ||||
| 23F | 1 | 1 | 1 | 1 | |||
| 24B | 1 | ||||||
| 24F | 1 | ||||||
| 28A | 1 | 4 | |||||
| 29 | 3 | ||||||
| 31 | 11 | ||||||
| 33F | 8 | 28 | 8 | 4 | 1 | ||
| 34 | 5 | ||||||
| 35A | 1 | 1 | |||||
| 35B | 21 | 3 | 3 | 1 | |||
| 35F | 22 | 2 | |||||
| 37 | 3 | ||||||
| 38 | 13 | 1 | |||||
| NT | 3 | ||||||
| Classe | Pénicillines | Céphèmes | Carbapénèmes | Fluoroquinolones | Autres | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sérotype | AUGFootnote a | PENm | PENn | PENo | AXOm | AXOn | FURo | FURp | ERT | IMI | MER | LEV | MOX | CLA | CLI | CHL | DOX | SXT |
| 1 (n=1) | -Footnote b | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 3 (n=82) | - | 1,2Footnote c | - | - | - | - | - | - | - | - | - | - | - | 3,7 | 2,4 | 7,3 | 11,0 | 1,2 |
| 4 (n=13) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 5 (n=2) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 |
| 6A (n=5) | - | 60,0 | - | - | - | - | - | - | - | - | - | 20,0 | - | - | - | - | - | - |
| 6B (n=6) | - | 16,7 | - | - | - | - | - | - | - | - | - | - | - | 33,3 | 16,7 | - | 33,3 | 50,0 |
| 6C (n=33) | - | 33,3 | - | 3,0 | - | - | - | 6,1 | - | - | - | - | - | 45,5 | 12,1 | - | 12,1 | 21,2 |
| 6D (n=2) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 7C (n=8) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 12,5 |
| 7F (n=110) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3,6 | - |
| 8 (n=50) | - | 2,0 | - | 2,0 | - | - | 2,0 | 2,0 | - | - | - | - | - | 2,0 | 2,0 | - | 6,0 | 2,0 |
| 9A (n=1) | - | 100 | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | - | 100 |
| 9N (n=46) | - | 2,2 | - | - | - | - | - | - | - | - | - | - | - | 4,3 | - | - | 4,3 | - |
| 9V (n=4) | - | 100 | - | 25,0 | - | - | 50,0 | 75,0 | - | - | - | - | - | 25,0 | - | - | - | 100 |
| 10A (n=31) | - | 6,5 | - | - | - | - | - | - | - | - | - | - | - | 22,6 | 6,5 | - | 3,2 | 9,7 |
| 10F (n=1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 |
| 11A (n=47) | - | 2,1 | - | - | - | - | - | - | - | - | - | 2,1 | - | 12,8 | - | - | - | 10,6 |
| 12A (n=1) | - | - | - | - | - | - | - | - | - | - | - | - | - | 100 | - | - | 100 | - |
| 12F (n=53) | - | - | - | - | - | - | - | - | - | - | - | - | - | 81,1 | 1,9 | - | 5,7 | - |
| 13 (n=1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 14 (n=3) | - | 33,3 | - | 33,3 | - | - | 33,3 | 33,3 | - | - | 33,3 | 33,3 | 33,3 | 66,7 | - | - | 33,3 | - |
| 15A (n=22) | - | 40,9 | - | - | - | - | - | 9,1 | - | - | - | - | - | 77,3 | 31,8 | - | 72,7 | - |
| 15B (n=19) | - | - | - | - | - | - | - | - | - | - | - | - | - | 10,5 | - | - | 5,3 | 5,3 |
| 15C (n=12) | - | 16,7 | - | - | - | - | - | - | - | - | - | - | - | 16,7 | - | - | - | - |
| 16F (n=22) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 17F (n=10) | - | 10,0 | - | - | - | - | - | - | - | - | - | - | - | 10,0 | 10,0 | - | 10,0 | - |
| 18A (n=1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 18C (n=4) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 25,0 |
| 19A(n=114) | 6,1 | 28,9 | - | 22,8 | 4,4 | 0,9 | 24,6 | 26,3 | 0,9 | 20,2 | 21,1 | - | - | 59,6 | 26,3 | - | 33,3 | 27,2 |
| 19F (n=13) | - | 38,5 | - | 23,1 | 7,7 | - | 15,4 | 23,1 | - | 7,7 | 15,4 | - | - | 23,1 | 23,1 | - | 23,1 | 23,1 |
| 20 (n=12) | - | - | - | - | - | - | - | - | - | - | - | - | - | 8,3 | - | - | - | - |
| 21 (n=10) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 20,0 |
| 22F(n=118) | - | 0,8 | 0,8 | 0,8 | 0,8 | - | 0,8 | 0,8 | - | - | - | - | - | 28,0 | - | 0,8 | - | - |
| 23A (n=30) | - | 30,0 | - | - | - | - | - | - | - | - | - | - | - | 3,3 | - | 3,3 | 16,7 | 3,3 |
| 23B (n=21) | - | 38,1 | - | - | - | - | - | - | - | - | - | - | - | 4,8 | - | - | 4,8 | 4,8 |
| 23F (n=4) | - | 50,0 | - | 25,0 | - | - | 25,0 | 25,0 | - | - | - | 75,0 | - | 50,0 | 25,0 | 25,0 | 25,0 | 25,0 |
| 24B (n=1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 24F (n=1) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 28A (n=5) | - | - | - | - | - | - | - | - | - | - | - | - | - | 20,0 | - | 40,0 | 40,0 | 20,0 |
| 29 (n=3) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 31 (n=11) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 33F (n=49) | - | 2,0 | - | - | - | - | - | - | - | - | - | - | - | 81,6 | 16,3 | - | 10,2 | 10,2 |
| 34 (n=5) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 35A (n=2) | - | - | - | - | - | - | - | - | - | - | - | - | - | 50,0 | - | - | 50,0 | 50,0 |
| 35B (n=28) | - | 25,0 | - | 3,6 | - | - | 7,1 | 21,4 | - | - | 3,6 | - | - | 14,3 | - | - | - | 3,6 |
| 35F (n=24) | - | - | - | - | - | - | - | - | - | - | - | - | - | 4,2 | - | - | - | - |
| 37 (n=3) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 38 (n=14) | - | 7,1 | - | - | - | - | - | - | - | - | - | - | - | 6,7 | 6,7 | - | 6,7 | - |
| NT (n=3) | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Tous (n=1061) | 0,7 | 10,0 | 0,1 | 3,4 | 0,7 | 0,1 | 3,6 | 4,7 | 0,1 | 2,3 | 2,6 | 0,6 | 0,1 | 24,8 | 5,8 | 1,0 | 9,9 | 7,4 |
Figure 40. Résistance des sérotypes de S. pneumoniae à la clarithromycine, 2011-2013
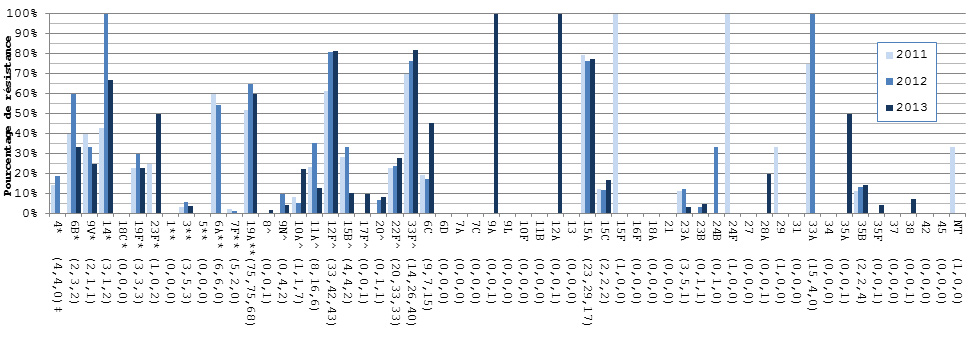
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae à la clarithromycine, 2011-2013 - Description textuel
Ce figure représente le pourcentage d'isolats résistants à la clarithromycine par type de vaccin, le sérotype et l'année 2011-2013
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4* (4,4,0)‡ | 14.3% | 19.0% | 0.0% |
| 6B* (2,3,2) | 40.0% | 60.0% | 33.3% |
| 9V* (2,1,1) | 40.0% | 33.3% | 25.0% |
| 14* (3,1,2) | 42.9% | 100.0% | 66.7% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (3,3,3) | 23.1% | 30.0% | 23.1% |
| 23F* (1,0,2) | 25.0% | 0.0% | 50.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (3,5,3) | 3.3% | 6.0% | 3.7% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (6,6,0) | 60.0% | 54.5% | 0.0% |
| 7F** (5,2,0) | 2.2% | 1.1% | 0.0% |
| 19A**(75,75,68) | 51.7% | 64.7% | 59.6% |
| 8^ (0,0,1) | 0.0% | 0.0% | 2.0% |
| 9N^ (0,4,2) | 0.0% | 9.8% | 4.3% |
| 10A^ (1,1,7) | 8.3% | 5.3% | 22.6% |
| 11A^ (8,16,6) | 23.5% | 35.6% | 12.8% |
| 12F^ (33,42,43) | 61.1% | 80.8% | 81.1% |
| 15B^ (4,4,2) | 28.6% | 33.3% | 10.5% |
| 17F^ (0,0,1) | 0.0% | 0.0% | 10.0% |
| 20^ (0,1,1) | 0.0% | 6.7% | 8.3% |
| 22F^ (20,33,33) | 23.0% | 23.7% | 28.0% |
| 33F^ (14,26,40) | 70.0% | 76.5% | 81.6% |
| 6C (9,7,15) | 19.1% | 17.5% | 45.5% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9A (0,0,1) | 0.0% | 0.0% | 100.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,1) | 0.0% | 0.0% | 100.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (23,29,17) | 79.3% | 76.3% | 77.3% |
| 15C (2,2,2) | 12.5% | 11.8% | 16.7% |
| 15F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (3,5,1) | 11.1% | 12.5% | 3.3% |
| 23B (0,1,1) | 0.0% | 3.1% | 4.8% |
| 24B (0,1,0) | 0.0% | 33.3% | 0.0% |
| 24F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,1) | 0.0% | 0.0% | 20.0% |
| 29 (1,0,0) | 33.3% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (15,4,0) | 75.0% | 100.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,1) | 0.0% | 0.0% | 50.0% |
| 35B (2,2,4) | 11.1% | 13.3% | 14.3% |
| 35F (0,0,1) | 0.0% | 0.0% | 4.2% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,1) | 0.0% | 0.0% | 7.1% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (1,0,0) | 33.3% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 41. Résistance des sérotypes de S. pneumoniae au céfuroxime (parentéral), 2011-2013
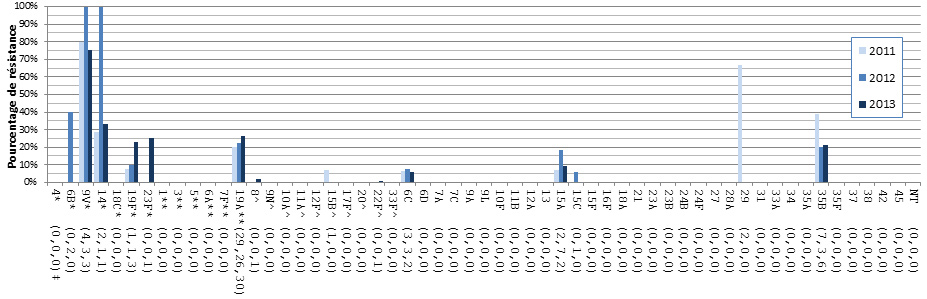
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae au céfuroxime (parentéral), 2011-2013 - Description textuel
Ce figure représente le pourcentage d'isolats résistants au céfuroxime (parentérale) par type de vaccin, par sérotype et par année.
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4* (0,0,0)‡ | 0.0% | 0.0% | 0.0% |
| 6B* (0,2,0) | 0.0% | 40.0% | 0.0% |
| 9V* (4,3,3) | 80.0% | 100.0% | 75.0% |
| 14* (2,1,1) | 28.6% | 100.0% | 33.3% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (1,1,3) | 7.7% | 10.0% | 23.1% |
| 23F* (0,0,1) | 0.0% | 0.0% | 25.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7F** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19A**(29,26,30) | 20.0% | 22.4% | 26.3% |
| 8^ (0,0,1) | 0.0% | 0.0% | 2.0% |
| 9N^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15B^ (1,0,0) | 7.1% | 0.0% | 0.0% |
| 17F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (0,0,1) | 0.0% | 0.0% | 0.8% |
| 33F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6C (3,3,2) | 6.4% | 7.5% | 6.1% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (2,7,2) | 6.9% | 18.4% | 9.1% |
| 15C (0,1,0) | 0.0% | 5.9% | 0.0% |
| 15F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 29 (2,0,0) | 66.7% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35B (7,3,6) | 38.9% | 20.0% | 21.4% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 42. Résistance des sérotypes de S. pneumoniae à la clindamycine, 2011-2013

* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae à la clindamycine, 2011-2013 - Description textuel
Ce figure représente le pourcentage de clindamycine S. pneumoniae résistant par le vaccin, le sérotype et l'année 2011-2013.
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4* (0,1,0)‡ | 0.0% | 4.8% | 0.0% |
| 6B* (0,1,1) | 0.0% | 20.0% | 16.7% |
| 9V* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 14* (2,1,0) | 28.6% | 100.0% | 0.0% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (3,1,3) | 23.1% | 10.0% | 23.1% |
| 23F* (0,0,1) | 0.0% | 0.0% | 25.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (1,2,2) | 1.1% | 2.4% | 2.4% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (1,0,0) | 10.0% | 0.0% | 0.0% |
| 7F** (0,1,0) | 0.0% | 0.6% | 0.0% |
| 19A**(35,28,30) | 24.1% | 24.1% | 26.3% |
| 8^ (0,0,1) | 0.0% | 0.0% | 2.0% |
| 9N^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10A^ (0,1,2) | 0.0% | 5.3% | 6.5% |
| 11A^ (1,0,0) | 2.9% | 0.0% | 0.0% |
| 12F^ (1,1,1) | 1.9% | 1.9% | 1.9% |
| 15B^ (2,0,0) | 14.3% | 0.0% | 0.0% |
| 17F^ (1,1,1) | 16.7% | 9.1% | 10.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (1,3,0) | 1.1% | 2.2% | 0.0% |
| 33F^ (2,6,8) | 10.0% | 17.6% | 16.3% |
| 6C (4,2,4) | 8.5% | 5.0% | 12.1% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (22,22,7) | 75.9% | 57.9% | 31.8% |
| 15C (1,1,0) | 6.3% | 5.9% | 0.0% |
| 15F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (1,3,0) | 3.7% | 7.5% | 0.0% |
| 23B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 29 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,1,0) | 0.0% | 25.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,1) | 0.0% | 0.0% | 7.1% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 43. Résistance des sérotypes de S. pneumoniae à la doxycycline, 2011-2013
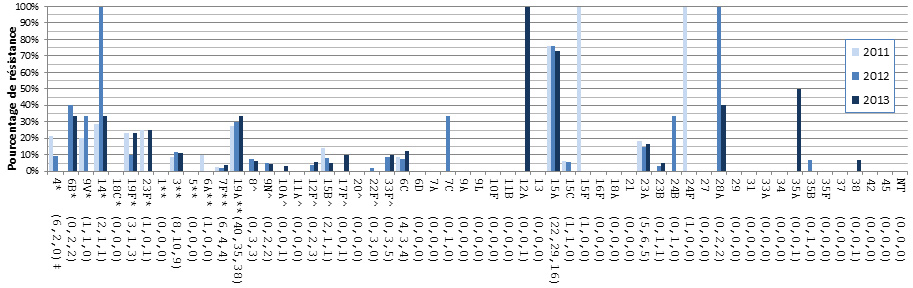
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae à la doxycycline, 2011-2013 - Description textuel
Ce figure représente le pourcentage de la doxycycline S. pneumoniae résistant par le vaccin, le sérotype et l'année 2011-2013.
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4*(6,2,0)‡ | 21.4% | 9.5% | 0.0% |
| 6B* (0,2,2) | 0.0% | 40.0% | 33.3% |
| 9V* (1,1,0) | 20.0% | 33.3% | 0.0% |
| 14* (2,1,1) | 28.6% | 100.0% | 33.3% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (3,1,3) | 23.1% | 10.0% | 23.1% |
| 23F* (1,0,1) | 25.0% | 0.0% | 25.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (8,10,9) | 8.7% | 11.9% | 11.0% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (1,0,0) | 10.0% | 0.0% | 0.0% |
| 7F** (6,4,4) | 2.6% | 2.3% | 3.6% |
| 19A**(40,35,38) | 27.6% | 30.2% | 33.3% |
| 8^ (0,3,3) | 0.0% | 7.7% | 6.0% |
| 9N^ (0,2,2) | 0.0% | 4.9% | 4.3% |
| 10A^ (0,0,1) | 0.0% | 0.0% | 3.2% |
| 11A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12F^ (0,2,3) | 0.0% | 3.8% | 5.7% |
| 15B^ (2,1,1) | 14.3% | 8.3% | 5.3% |
| 17F^ (0,0,1) | 0.0% | 0.0% | 10.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (0,3,0) | 0.0% | 2.2% | 0.0% |
| 33F^ (0,3,5) | 0.0% | 8.8% | 10.2% |
| 6C (4,3,4) | 8.5% | 7.5% | 12.1% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,1,0) | 0.0% | 33.3% | 0.0% |
| 9A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,1) | 0.0% | 0.0% | 100.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (22,29,16) | 75.9% | 76.3% | 72.7% |
| 15C (1,1,0) | 6.3% | 5.9% | 0.0% |
| 15F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (5,6,5) | 18.5% | 15.0% | 16.7% |
| 23B (0,1,1) | 0.0% | 3.1% | 4.8% |
| 24B (0,1,0) | 0.0% | 33.3% | 0.0% |
| 24F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,2,2) | 0.0% | 100.0% | 40.0% |
| 29 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,1) | 0.0% | 0.0% | 50.0% |
| 35B (0,1,0) | 0.0% | 6.7% | 0.0% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,1) | 0.0% | 0.0% | 7.1% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 44. Résistance des sérotypes de S. pneumoniae à l'imipénème, 2011-2013

* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae à l'imipénème, 2011-2013 - Description textuel
Ce figure représente le pourcentage de S. pneumoniae résistant à l'imipénème par le vaccin, le sérotype et l'année 2011-2013.
| Sérotype | 2010 | 2011 | 2012 |
|---|---|---|---|
| 4* (0,0,0)‡ | 0.0% | 0.0% | 0.0% |
| 6B* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9V* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 14* (0,0,0) | 7.7% | 10.0% | 7.7% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (1,1,1) | 0.0% | 0.0% | 0.0% |
| 23F* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (0,0,0) | 6.9% | 16.4% | 20.2% |
| 7F** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19A**(10,19,23) | 0.0% | 0.0% | 0.0% |
| 8^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9N^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15B^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 17F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 29 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 45. Résistance des sérotypes de S. pneumoniae au méropénème, 2011-2013
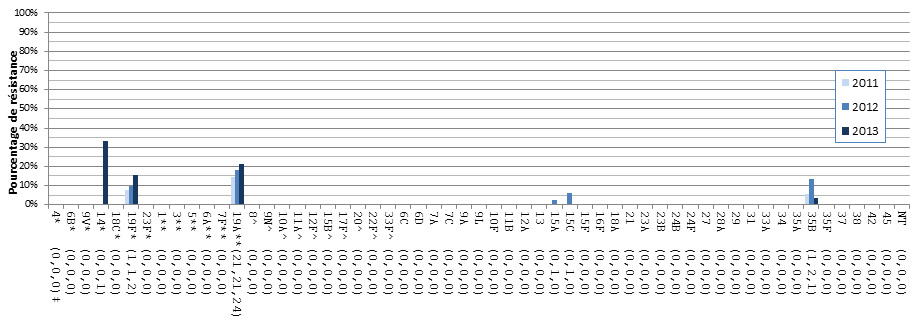
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae au méropénème, 2011-2013 - Description textuel
Ce figure représente le pourcentage de méropénème S. pneumoniae résistant par le vaccin, le sérotype et l'année 2011-2013.
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4* (0,0,0)‡ | 0.0% | 0.0% | 0.0% |
| 6B* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9V* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 14* (0,0,1) | 0.0% | 0.0% | 33.3% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (1,1,2) | 7.7% | 10.0% | 15.4% |
| 23F* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7F** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19A**(21,21,24) | 14.5% | 18.1% | 21.1% |
| 8^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9N^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11A^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15B^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 17F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (0,1,0) | 0.0% | 2.6% | 0.0% |
| 15C (0,1,0) | 0.0% | 5.9% | 0.0% |
| 15F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 29 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35B (1,2,1) | 5.6% | 13.3% | 3.6% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 46. Résistance des sérotypes de S. pneumoniae à la pénicilline (doses seuils pour la méningite), 2011-2013
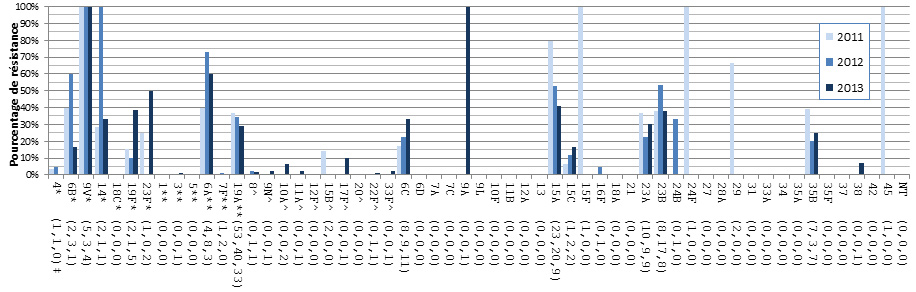
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae à la pénicilline (doses seuils pour la méningite), 2011-2013 - Description textuel
Ce figure représente le pourcentage de S. pneumoniae résistant à la pénicilline par le vaccin, le sérotype et l'année 2011-2013.
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4* (1,1,0)‡ | 3.6% | 4.8% | 0.0% |
| 6B* (2,3,1) | 40.0% | 60.0% | 16.7% |
| 9V* (5,3,4) | 100.0% | 100.0% | 100.0% |
| 14* (2,1,1) | 28.6% | 100.0% | 33.3% |
| 18C* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 19F* (2,1,5) | 15.4% | 10.0% | 38.5% |
| 23F* (1,0,2) | 25.0% | 0.0% | 50.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (0,0,1) | 0.0% | 0.0% | 1.2% |
| 5** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 6A** (4,8,3) | 40.0% | 72.7% | 60.0% |
| 7F** (1,2,0) | 0.4% | 1.1% | 0.0% |
| 19A**(53,40,33) | 36.6% | 34.5% | 28.9% |
| 8^ (0,1,1) | 0.0% | 2.6% | 2.0% |
| 9N^ (0,0,1) | 0.0% | 0.0% | 2.2% |
| 10A^ (0,0,2) | 0.0% | 0.0% | 6.5% |
| 11A^ (0,0,1) | 0.0% | 0.0% | 2.1% |
| 12F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15B^ (2,0,0) | 14.3% | 0.0% | 0.0% |
| 17F^ (0,0,1) | 0.0% | 0.0% | 10.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (0,1,1) | 0.0% | 0.7% | 0.8% |
| 33F^ (0,0,1) | 0.0% | 0.0% | 2.0% |
| 6C (8,9,11) | 17.0% | 22.5% | 33.3% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,0) | 0.0% | 0.0% | 0.0% |
| 9A (0,0,1) | 0.0% | 0.0% | 100.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (23,20,9) | 79.3% | 52.6% | 40.9% |
| 15C (1,2,2) | 6.3% | 11.8% | 16.7% |
| 15F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 16F (0,1,0) | 0.0% | 4.5% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 23A (10,9,9) | 37.0% | 22.5% | 30.0% |
| 23B (8,17,8) | 38.1% | 53.1% | 38.1% |
| 24B (0,1,0) | 0.0% | 33.3% | 0.0% |
| 24F (1,0,0) | 100.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 29 (2,0,0) | 66.7% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35B (7,3,7) | 38.9% | 20.0% | 25.0% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,1) | 0.0% | 0.0% | 7.1% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (1,0,0) | 100.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Figure 47. Résistance des sérotypes de S. pneumoniae au triméthoprime-sulfaméthoxazole, 2011-2013
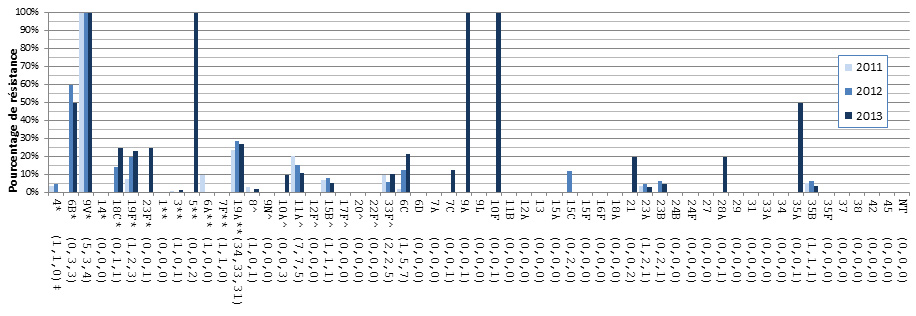
* Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement.
Résistance des sérotypes de S. pneumoniae au triméthoprime-sulfaméthoxazole, 2011-2013 - Description textuel
Ce figure représente le pourcentage de triméthoprime / sulfaméthoxazole S. pneumoniae résistant par le vaccin, le sérotype et l'année 2011-2013.
| Sérotype | 2011 | 2012 | 2013 |
|---|---|---|---|
| 4* (1,1,0)‡ | 3.6% | 4.8% | 0.0% |
| 6B* (0,3,3) | 0.0% | 60.0% | 50.0% |
| 9V* (5,3,4) | 100.0% | 100.0% | 100.0% |
| 14* (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18C* (0,1,1) | 0.0% | 14.3% | 25.0% |
| 19F* (1,2,3) | 7.7% | 20.0% | 23.1% |
| 23F* (0,0,1) | 0.0% | 0.0% | 25.0% |
| 1** (0,0,0) | 0.0% | 0.0% | 0.0% |
| 3** (1,0,1) | 1.1% | 0.0% | 1.2% |
| 5** (0,0,2) | 0.0% | 0.0% | 100.0% |
| 6A** (1,0,0) | 10.0% | 0.0% | 0.0% |
| 7F** (1,1,0) | 0.4% | 0.6% | 0.0% |
| 19A**(34,33,31) | 23.4% | 28.4% | 27.2% |
| 8^ (1,0,1) | 3.2% | 0.0% | 2.0% |
| 9N^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10A^ (0,0,3) | 0.0% | 0.0% | 9.7% |
| 11A^ (7,7,5) | 20.6% | 15.6% | 10.6% |
| 12F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15B^ (1,1,1) | 7.1% | 8.3% | 5.3% |
| 17F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 20^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 22F^ (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33F^ (2,2,5) | 10.0% | 5.9% | 10.2% |
| 6C (1,5,7) | 2.1% | 12.5% | 21.2% |
| 6D (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 7C (0,0,1) | 0.0% | 0.0% | 12.5% |
| 9A (0,0,1) | 0.0% | 0.0% | 100.0% |
| 9L (0,0,0) | 0.0% | 0.0% | 0.0% |
| 10F (0,0,1) | 0.0% | 0.0% | 100.0% |
| 11B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 12A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 13 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 15C (0,2,0) | 0.0% | 11.8% | 0.0% |
| 15F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 16F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 18A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 21 (0,0,2) | 0.0% | 0.0% | 20.0% |
| 23A (1,2,1) | 3.7% | 5.0% | 3.3% |
| 23B (0,2,1) | 0.0% | 6.3% | 4.8% |
| 24B (0,0,0) | 0.0% | 0.0% | 0.0% |
| 24F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 27 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 28A (0,0,1) | 0.0% | 0.0% | 20.0% |
| 29 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 31 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 33A (0,0,0) | 0.0% | 0.0% | 0.0% |
| 34 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 35A (0,0,1) | 0.0% | 0.0% | 50.0% |
| 35B (1,1,1) | 5.6% | 6.7% | 3.6% |
| 35F (0,0,0) | 0.0% | 0.0% | 0.0% |
| 37 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 38 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 42 (0,0,0) | 0.0% | 0.0% | 0.0% |
| 45 (0,0,0) | 0.0% | 0.0% | 0.0% |
| NT (0,0,0) | 0.0% | 0.0% | 0.0% |
| * Composante du PCV7 ; ** Composante du PCV13 ; ^ Composante du PPV23 ; ‡ Nombre d'isolats pour 2010 , 2011, 2012 et 2013 , respectivement. | |||
Streptococcus pyogenes (streptocoque du groupe A)
Sur les 1 294 isolats de Streptococcus pyogenes analysés au LNM par typage du gène emm, 155 (12,0 %) avaient été prélevés chez des enfants de < 15 ans, 1 096 (84,7 %) chez des adultes de ≥ 15 ans, ainsi que chez 43 personnes dont on ne connaissait pas l’âge. Les renseignements sur le sexe étaient disponibles pour 1 269 isolats, dont 54,2 % (n = 688) provenaient d’hommes.
Aucune différence majeure n’a été décelée entre les adultes et les enfants dans les proportions relatives selon les sites d’isolement clinique, sinon que davantage de cas liés au LCR et au liquide pleural ont été observés dans les isolats provenant d’enfants (2,6 %, n = 4 et 6,5 %, n = 10, respectivement) que dans les isolats provenant d’adultes (0,2 %, n = 2 et 1,5 %, n = 16, respectivement) (figures 48 et 49).
Le type emm le plus fréquent au Canada en 2013 est encore emm1, qui représente 28,4 % (n = 44) des isolats prélevés chez les enfants de < 15 ans et 23,7 % (n = 260) des isolats prélevés chez les adultes de ≥ 15 ans (figures 50 et 51). Le deuxième en importance est emm89, qui correspond à 7,1 % (n = 11) des isolats chez les enfants et à 9.9 % (n = 108) des isolats chez les adultes. La majorité des isolats de types emm68 (n = 46), emm53 (n = 32), emm80 (n = 28), emm77 (n = 14), emm22 (n = 12) et emm41 (n = 6) provenaient de l’Ouest du Canada, tandis que la majorité des isolats de types emm89 (n = 90), emm4 (n = 54), emm28 (n = 54), emm12 (n = 49), emm59 (n = 33) et emm2 (n = 277) provenaient des régions du Centre (figure 52).
Parmi les isolats prélevés chez les enfants, les adultes et les sujets d’âge inconnu (n = 887), les proportions des types emm1 et emm89 dans les isolats sanguins étaient beaucoup plus élevées (26,6 %, n = 236 et 9,1 %, n = 81, respectivement) que celles des isolats de liquide synovial (10,2 %, n = 10 et 6,1 %, n = 6, respectivement). Par contre, les proportions des types emm68 et emm82 étaient plus élevées dans les isolats de liquide synovial (7,1 %, n = 7 et 8,2 %, n = 8, respectivement) que dans les isolats sanguins (3,0 %, n = 27 et 1,2 %, n = 11, respectivement) (figure 53).
| Province | < 2 | 2 – 4 | 5 – 14 | 15 – 49 | 50 – 64 | ≥ 65 | Non précisé | Total |
|---|---|---|---|---|---|---|---|---|
| Colombie-Britannique | 3 | 2 | 9 | 61 | 23 | 43 | 1 | 142 |
| Alberta | - | - | - | - | - | 1 | - | 1 |
| Saskatchewan | 5 | 1 | 5 | 43 | 22 | 17 | 2 | 95 |
| Manitoba | 11 | 2 | 7 | 75 | 47 | 16 | 12 | 170 |
| Ontario | 27 | 6 | 26 | 167 | 115 | 158 | 19 | 518 |
| Québec | 10 | 8 | 30 | 121 | 51 | 91 | 8 | 319 |
| Nouveau-Brunswick | - | - | 1 | 7 | 1 | 8 | - | 17 |
| Nouvelle-Écosse | - | - | - | 5 | 3 | 4 | - | 12 |
| Île-du-Prince-Édouard | - | - | - | - | 1 | - | 1 | 2 |
| Terre-Neuve-et-Labrador | - | - | 2 | 3 | 6 | 1 | - | 12 |
| Yukon Territories | - | - | - | 1 | 1 | - | - | 2 |
| Territoires du Nord-Ouest | - | - | - | 2 | 1 | 1 | - | 4 |
| Canada | 56 | 19 | 80 | 485 | 271 | 340 | 43 | 1294 |
Figure 48. Sites d'isolement clinique de S. pyogenes chez les enfants de < 15 ans (N = 155)
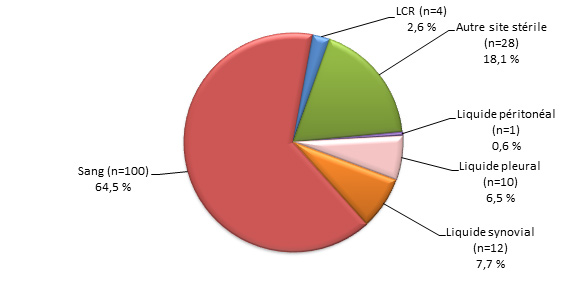
Les autres sites stériles sont notamment les suivants : tissus profonds, échantillons prélevés au cours d'une biopsie ou d'une chirurgie, os, mastoïde et sources cliniques associées à la fasciite nécrosante. L'âge du patient n'était pas précisé pour 43 isolats.
Sites d'isolement clinique de S. pyogenes chez les enfants de < 15 ans (N = 155) - Description textuel
Ce figure représente le pourcentage de S. pyogenes isole de divers sites d'isolement cliniques chez les enfants de moins de 15 ans.
| Le site d'isolement clinique | Pourcentage |
|---|---|
| Sang (n=100) | 64.5% |
| LCR (n=4) | 2.6% |
| Autre site stérile (n=28) | 18.1% |
| Liquide peritoneal (n=1) | 0.6% |
| Liquide pleural (n=10) | 6.5% |
| Liquide synovial (n=12) | 7.7% |
Les autres sites stériles sont notamment les suivants : tissus profonds, échantillons prélevés au cours d'une biopsie ou d'une chirurgie, os, mastoïde et sources cliniques associées à la fasciite nécrosante. L'âge du patient n'était pas précisé pour 43 isolats.
Figure 49. Sites d'isolement clinique de S. pyogenes chez les adultes ≥ 15 ans (N = 1 096)
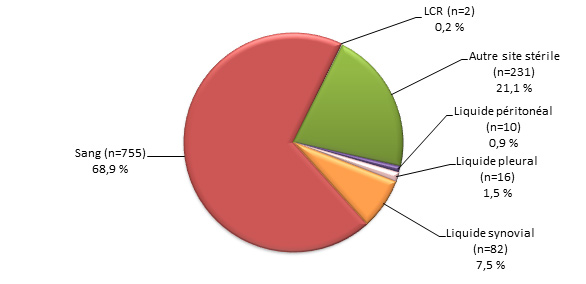
Les autres sites stériles sont notamment les suivants : tissus profonds, échantillons prélevés au cours d'une biopsie ou d'une chirurgie, os, mastoïde et sources cliniques associées à la fasciite nécrosante. L'âge du patient n'était pas précisé pour 43 isolats.
Sites d'isolement clinique de S. pyogenes chez les adultes ≥ 15 ans (N = 1 096 - Description textuel
Ce figure représente le pourcentage de S. pyogenes de divers sites d'isolement cliniques des adultes (âgés de plus de 14 années).
| Le site d'isolement clinique | Pourcentage |
|---|---|
| Sang (n=755) | 68.9% |
| LCR (n=2) | 0.2% |
| Autre site stérile (n=231) | 21.1% |
| Liquide peritoneal (n=10) | 0.9% |
| Liquide pleural (n=16) | 1.5% |
| Liquide synovial (n=82) | 7.5% |
Les autres sites stériles sont notamment les suivants : tissus profonds, échantillons prélevés au cours d'une biopsie ou d'une chirurgie, os, mastoïde et sources cliniques associées à la fasciite nécrosante. L'âge du patient n'était pas précisé pour 43 isolats.
Figure 50. Types emm de S. pyogenes causant une infection invasive chez les enfants de < 15 ans, 2010-2013
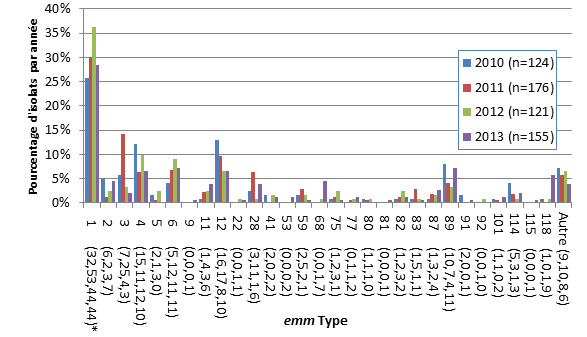
*Nombre d'isolats pour 2010, 2011, 2012 et 2013, respectivement
Types emm de S. pyogenes causant une infection invasive chez les enfants de < 15 ans, 2010-2013 - Description textuel
Ce chiffre représente le nombre de types emm S. pyogenes envahissantes trouvées chez les enfants de moins de 15 ans par année de 2010 à 2013.
| emm | 2010 (n=124) | 2011 (n=176) | 2012 (n=121) | 2013 (n=155) |
|---|---|---|---|---|
| 1 (32,53,44,44)Footnote * | 25.80% | 30.10% | 36.40% | 28.40% |
| 2 (6,2,3,7) | 4.80% | 1.10% | 2.50% | 4.50% |
| 3 (7,25,4,3) | 5.60% | 14.20% | 3.30% | 1.90% |
| 4 (15,11,12,10) | 12.10% | 6.30% | 9.90% | 6.50% |
| 5 (2,1,3,0) | 1.60% | 0.60% | 2.50% | 0.00% |
| 6 (5,12,11,11) | 4.00% | 6.80% | 9.10% | 7.10% |
| 9 (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.60% |
| 11 (1,4,3,6) | 0.80% | 2.30% | 2.50% | 3.90% |
| 12 (16,17,8,10) | 12.90% | 9.70% | 6.60% | 6.50% |
| 22 (0,0,1,1) | 0.00% | 0.00% | 0.80% | 0.60% |
| 28 (3,11,1,6) | 2.40% | 6.30% | 0.80% | 3.90% |
| 41 (2,0,2,2) | 1.60% | 0.00% | 1.70% | 1.30% |
| 53 (0,0,0,2) | 0.00% | 0.00% | 0.00% | 1.30% |
| 59 (2,5,2,1) | 1.60% | 2.80% | 1.70% | 0.60% |
| 68 (0,0,1,7) | 0.00% | 0.00% | 0.80% | 4.50% |
| 75 (1,2,3,1) | 0.80% | 1.10% | 2.50% | 0.60% |
| 77 (0,1,1,2) | 0.00% | 0.60% | 0.80% | 1.30% |
| 80 (1,1,1,0) | 0.80% | 0.60% | 0.80% | 0.00% |
| 81 (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.60% |
| 82 (1,2,3,2) | 0.80% | 1.10% | 2.50% | 1.30% |
| 83 (1,5,1,1) | 0.80% | 2.80% | 0.80% | 0.60% |
| 87 (1,3,2,4) | 0.80% | 1.70% | 1.70% | 2.60% |
| 89 (10,7,4,11) | 8.10% | 4.00% | 3.30% | 7.10% |
| 91 (2,0,0,1) | 1.60% | 0.00% | 0.00% | 0.60% |
| 92 (0,0,1,0) | 0.00% | 0.00% | 0.80% | 0.00% |
| 101 (1,1,0,2) | 0.80% | 0.60% | 0.00% | 1.30% |
| 114 (5,3,1,3) | 4.00% | 1.70% | 0.80% | 1.90% |
| 115 (0,0,0,1) | 0.00% | 0.00% | 0.00% | 0.60% |
| 118 (1,0,1,9) | 0.80% | 0.00% | 0.80% | 5.80% |
| Autre (9,10,8,6) | 7.30% | 5.70% | 6.60% | 3.90% |
Figure 51. Types emm de S. pyogenes causant une infection invasive chez les adultes de ≥ 15 ans, 2010-2013
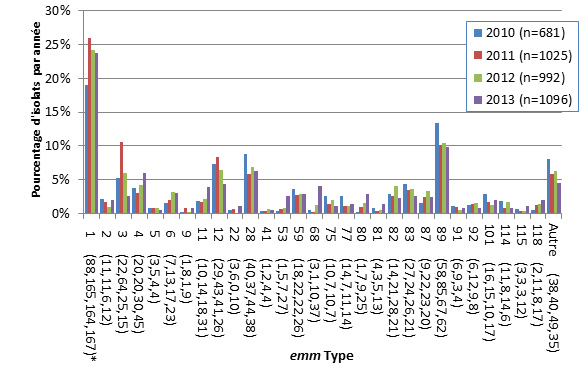
*Nombre d'isolats pour 2010 , 2011, 2012 et 2013, respectivement
Types emm de S. pyogenes causant une infection invasive chez les adultes de ≥ 15 ans, 2010-2013 - Description textuel
Ce figure représente le nombre de types emm S. pyogenes invasives chez les adultes de plus de 14 ans par année de 2010 à 2013.
| emm | 2010 (n=681) | 2011 (n=1025) | 2012 (n=992) | 2013 (n=1096) |
|---|---|---|---|---|
| 1 (88,165,164,167)Footnote * | 18.90% | 26.00% | 24.20% | 23.70% |
| 2 (11,11,6,12) | 2.20% | 1.70% | 1.00% | 2.00% |
| 3 (22,64,25,15) | 5.30% | 10.50% | 5.90% | 2.60% |
| 4 (20,20,30,45) | 3.80% | 3.00% | 4.20% | 5.90% |
| 5 (3,5,4,4) | 0.70% | 0.80% | 0.80% | 0.50% |
| 6 (7,13,17,23) | 1.50% | 2.00% | 3.20% | 3.00% |
| 9 (1,8,1,9) | 0.10% | 0.80% | 0.10% | 0.80% |
| 11 (10,14,18,31) | 1.80% | 1.80% | 2.10% | 3.90% |
| 12 (29,43,41,26) | 7.30% | 8.30% | 6.50% | 4.30% |
| 22 (3,6,0,10) | 0.40% | 0.70% | 0.00% | 1.10% |
| 28 (40,37,44,38) | 8.80% | 5.90% | 6.90% | 6.30% |
| 41 (1,2,4,4) | 0.30% | 0.40% | 0.70% | 0.50% |
| 53 (1,5,7,27) | 0.30% | 0.60% | 0.80% | 2.60% |
| 59 (18,22,22,26) | 3.70% | 2.70% | 2.80% | 2.80% |
| 68 (3,1,10,37) | 0.40% | 0.20% | 1.20% | 4.00% |
| 75 (10,7,10,7) | 2.60% | 1.50% | 2.00% | 1.10% |
| 77 (14,7,11,14) | 2.60% | 1.10% | 1.10% | 1.50% |
| 80 (1,7,9,25) | 0.10% | 1.00% | 1.50% | 2.80% |
| 81 (4,3,5,13) | 0.70% | 0.40% | 0.50% | 1.40% |
| 82 (14,21,28,21) | 2.90% | 2.50% | 4.00% | 2.30% |
| 83 (27,24,26,21) | 4.40% | 3.50% | 3.60% | 2.60% |
| 87 (9,22,23,20) | 1.50% | 2.40% | 3.30% | 2.50% |
| 89 (58,85,67,62) | 13.40% | 10.10% | 10.50% | 9.90% |
| 91 (6,9,3,4) | 1.00% | 0.90% | 0.50% | 0.70% |
| 92 (6,12,9,8) | 1.20% | 1.40% | 1.50% | 0.70% |
| 101 (16,15,10,17) | 2.90% | 1.70% | 1.30% | 2.00% |
| 114 (11,8,14,6) | 1.80% | 0.80% | 1.60% | 0.80% |
| 115 (3,3,3,12) | 0.60% | 0.40% | 0.30% | 1.10% |
| 118 (2,11,8,17) | 0.40% | 1.30% | 1.40% | 1.90% |
| Autre (38,40,49,35) | 8.10% | 5.90% | 6.30% | 4.60% |
Figure 52. Distribution régionale des types emm de S. pyogenes

Distribution régionale des types emm de S. pyogenes - Description textuel
Ce figure représente la distribution des types emm par région (Europe occidentale, centrale et orientale).
| Type emm | Ouest (n=414) | Centre (n=837) | Est (n=43) |
|---|---|---|---|
| 1 | 64 | 240 | 7 |
| 89 | 24 | 90 | 7 |
| 4 | 16 | 54 | 8 |
| 28 | 18 | 54 | 4 |
| 12 | 12 | 49 | 2 |
| 68 | 46 | 9 | |
| 11 | 26 | 22 | 2 |
| 6 | 12 | 31 | 1 |
| 53 | 32 | 2 | |
| 59 | 33 | ||
| 118 | 10 | 22 | 1 |
| 2 | 5 | 27 | |
| 3 | 3 | 24 | 5 |
| 80 | 28 | 4 | |
| 87 | 16 | 15 | |
| 83 | 11 | 19 | |
| 82 | 20 | 9 | |
| 101 | 7 | 17 | |
| 77 | 14 | 4 | 1 |
| 81 | 2 | 14 | |
| 22 | 12 | 2 | |
| 115 | 9 | 4 | |
| 75 | 2 | 10 | 1 |
| 114 | 5 | 7 | |
| 9 | 11 | ||
| 91 | 2 | 7 | |
| 92 | 8 | ||
| 41 | 6 | 1 | |
| 5 | 6 | ||
| 58 | 1 | 3 | |
| 49 | 4 | ||
| 76 | 1 | 3 | |
| 44 | 1 | 2 | |
| 94 | 3 | ||
| 102 | 2 | 1 | |
| 169 | 2 | ||
| 73 | 2 | ||
| 174 | 2 | ||
| 88 | 2 | ||
| 74 | 1 | 1 | |
| 100 | 2 | ||
| 170 | 2 | ||
| 183 | 2 | ||
| 90 | 1 | 1 | |
| 85 | 2 | ||
| 103 | 1 | 1 | |
| 238 | 1 | ||
| 230 | 1 | ||
| 194 | 1 | ||
| 8 | 1 | ||
| 64 | 1 | ||
| 33 | 1 | ||
| 56 | 1 | ||
| 29 | 1 | ||
| 223 | 1 | ||
| 119 | 1 | ||
| 110 | 1 | ||
| 239 | 1 | ||
| 109 | 1 | ||
| 71 | 1 | ||
| NT | 1 |
Figure 53a. Distribution des types emm de S. pyogenes causant une infection invasive isoler du sangFootnote *, 2013
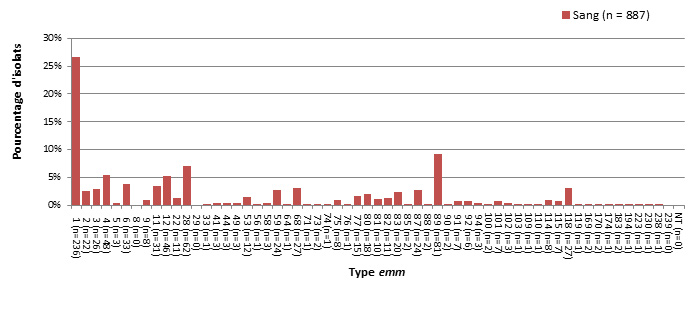
Types emm de S. pyogenes causant une infection invasive par source d'isolement clinique - Description textuel
Ce figure représente le pourcentage de types emm dans diverses sources d'isolement cliniques en 2013.
| Sérotype | Sang (n=887) |
|---|---|
| 1 (n=236) | 26.61% |
| 2 (n=22) | 2.48% |
| 3 (n=26) | 2.93% |
| 4 (n=48) | 5.41% |
| 5 (n=3) | 0.34% |
| 6 (n=33) | 3.72% |
| 8 (n=0) | 0.00% |
| 9 (n=8) | 0.90% |
| 11 (n=31) | 3.49% |
| 12 (n=46) | 5.19% |
| 22 (n=11) | 1.24% |
| 28 (n=62) | 6.99% |
| 29 (n=0) | 0.00% |
| 33 (n=1) | 0.11% |
| 41 (n=3) | 0.34% |
| 44 (n=3) | 0.34% |
| 49 (n=3) | 0.34% |
| 53 (n=12) | 1.35% |
| 56 (n=1) | 0.11% |
| 58 (n=3) | 0.34% |
| 59 (n=24) | 2.71% |
| 64 (n=1) | 0.11% |
| 68 (n=27) | 3.04% |
| 71 (n=1) | 0.11% |
| 73 (n=2) | 0.23% |
| 74 (n=1) | 0.11% |
| 75 (n=8) | 0.90% |
| 76 (n=1) | 0.11% |
| 77 (n=15) | 1.69% |
| 80 (n=18) | 2.03% |
| 81 (n=10) | 1.13% |
| 82 (n=11) | 1.24% |
| 83 (n=20) | 2.25% |
| 85 (n=2) | 0.23% |
| 87 (n=24) | 2.71% |
| 88 (n=2) | 0.23% |
| 89 (n=81) | 9.13% |
| 90 (n=2) | 0.23% |
| 91 (n=7) | 0.79% |
| 92 (n=6) | 0.68% |
| 94 (n=3) | 0.34% |
| 100 (n=2) | 0.23% |
| 101 (n=7) | 0.79% |
| 102 (n=3) | 0.34% |
| 103 (n=1) | 0.11% |
| 109 (n=1) | 0.11% |
| 110 (n=1) | 0.11% |
| 114 (n=8) | 0.90% |
| 115 (n=7) | 0.79% |
| 118 (n=27) | 3.04% |
| 119 (n=1) | 0.11% |
| 169 (n=2) | 0.23% |
| 170 (n=2) | 0.23% |
| 174 (n=1) | 0.11% |
| 183 (n=2) | 0.23% |
| 194 (n=1) | 0.11% |
| 223 (n=1) | 0.11% |
| 230 (n=1) | 0.11% |
| 238 (n=1) | 0.11% |
| 239 (n=0) | 0.00% |
| NT (n=0) | 0.00% |
Figure 53b. Distribution des types emm de S. pyogenes causant une infection invasive isoler de la liquide synovialFootnote *, 2013

Distribution des types emm de S. pyogenes causant une infection invasive isoler de la liquide synovial - Description textuel
Ce figure représente le pourcentage de types emm isoler de la liquide synovial en 2013.
| Sérotype | Liquide Synoviale (n=98) |
|---|---|
| 1 (n=10) | 10.20% |
| 2 (n=3) | 3.06% |
| 3 (n=3) | 3.06% |
| 4 (n=7) | 7.14% |
| 5 (n=0) | 0.00% |
| 6 (n=3) | 3.06% |
| 8 (n=0) | 0.00% |
| 9 (n=0) | 0.00% |
| 11 (n=6) | 6.12% |
| 12 (n=4) | 4.08% |
| 22 (n=0) | 0.00% |
| 28 (n=4) | 4.08% |
| 29 (n=1) | 1.02% |
| 33 (n=0) | 0.00% |
| 41 (n=0) | 0.00% |
| 44 (n=0) | 0.00% |
| 49 (n=1) | 1.02% |
| 53 (n=5) | 5.10% |
| 56 (n=0) | 0.00% |
| 58 (n=1) | 1.02% |
| 59 (n=3) | 3.06% |
| 64 (n=0) | 0.00% |
| 68 (n=7) | 7.14% |
| 71 (n=0) | 0.00% |
| 73 (n=0) | 0.00% |
| 74 (n=1) | 1.02% |
| 75 (n=2) | 2.04% |
| 76 (n=1) | 1.02% |
| 77 (n=3) | 3.06% |
| 80 (n=2) | 2.04% |
| 81 (n=4) | 4.08% |
| 82 (n=8) | 8.16% |
| 83 (n=2) | 2.04% |
| 85 (n=0) | 0.00% |
| 87 (n=2) | 2.04% |
| 88 (n=0) | 0.00% |
| 89 (n=6) | 6.12% |
| 90 (n=0) | 0.00% |
| 91 (n=0) | 0.00% |
| 92 (n=1) | 1.02% |
| 94 (n=0) | 0.00% |
| 100 (n=0) | 0.00% |
| 101 (n=3) | 3.06% |
| 102 (n=0) | 0.00% |
| 103 (n=0) | 0.00% |
| 109 (n=0) | 0.00% |
| 110 (n=0) | 0.00% |
| 114 (n=1) | 1.02% |
| 115 (n=2) | 2.04% |
| 118 (n=2) | 2.04% |
| 119 (n=0) | 0.00% |
| 169 (n=0) | 0.00% |
| 170 (n=0) | 0.00% |
| 174 (n=0) | 0.00% |
| 183 (n=0) | 0.00% |
| 194 (n=0) | 0.00% |
| 223 (n=0) | 0.00% |
| 230 (n=0) | 0.00% |
| 238 (n=0) | 0.00% |
| 239 (n=0) | 0.00% |
| NT (n=0) | 0.00% |
Figure 53c. Distribution des types emm de S. pyogenes causant une infection invasive isoler d'un autre site stérileFootnote *, 2013
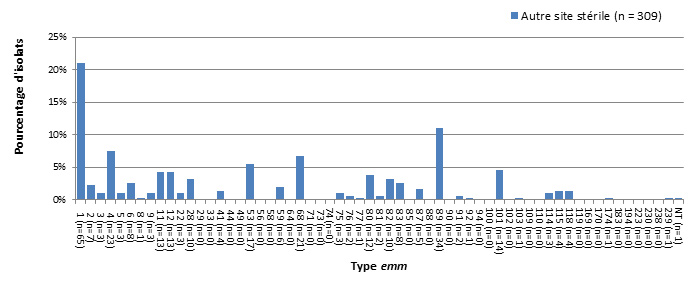
Distribution des types emm de S. pyogenes causant une infection invasive isoler d'un autre site stérile - Description textuel
Ce figure représente le pourcentage de types emm isoler d'un autre site stérile en 2013.
| Sérotype | Autre site stérile (n=309) |
|---|---|
| 1 (n=65) | 21.04% |
| 2 (n=7) | 2.27% |
| 3 (n=3) | 0.97% |
| 4 (n=23) | 7.44% |
| 5 (n=3) | 0.97% |
| 6 (n=8) | 2.59% |
| 8 (n=1) | 0.32% |
| 9 (n=3) | 0.97% |
| 11 (n=13) | 4.21% |
| 12 (n=13) | 4.21% |
| 22 (n=3) | 0.97% |
| 28 (n=10) | 3.24% |
| 29 (n=0) | 0.00% |
| 33 (n=0) | 0.00% |
| 41 (n=4) | 1.29% |
| 44 (n=0) | 0.00% |
| 49 (n=0) | 0.00% |
| 53 (n=17) | 5.50% |
| 56 (n=0) | 0.00% |
| 58 (n=0) | 0.00% |
| 59 (n=6) | 1.94% |
| 64 (n=0) | 0.00% |
| 68 (n=21) | 6.80% |
| 71 (n=0) | 0.00% |
| 73 (n=0) | 0.00% |
| 74 (n=0) | 0.00% |
| 75 (n=3) | 0.97% |
| 76 (n=2) | 0.65% |
| 77 (n=1) | 0.32% |
| 80 (n=12) | 3.88% |
| 81 (n=2) | 0.65% |
| 82 (n=10) | 3.24% |
| 83 (n=8) | 2.59% |
| 85 (n=0) | 0.00% |
| 87 (n=5) | 1.62% |
| 88 (n=0) | 0.00% |
| 89 (n=34) | 11.00% |
| 90 (n=0) | 0.00% |
| 91 (n=2) | 0.65% |
| 92 (n=1) | 0.32% |
| 94 (n=0) | 0.00% |
| 100 (n=0) | 0.00% |
| 101 (n=14) | 4.53% |
| 102 (n=0) | 0.00% |
| 103 (n=1) | 0.32% |
| 109 (n=0) | 0.00% |
| 110 (n=0) | 0.00% |
| 114 (n=3) | 0.97% |
| 115 (n=4) | 1.29% |
| 118 (n=4) | 1.29% |
| 119 (n=0) | 0.00% |
| 169 (n=0) | 0.00% |
| 170 (n=0) | 0.00% |
| 174 (n=1) | 0.32% |
| 183 (n=0) | 0.00% |
| 194 (n=0) | 0.00% |
| 223 (n=0) | 0.00% |
| 230 (n=0) | 0.00% |
| 238 (n=0) | 0.00% |
| 239 (n=1) | 0.32% |
| NT (n=1) | 0.32% |
Résistance de S. pyogenes aux antimicrobiens
Sur les 1 290 isolats de S. pyogenes causant une infection invasive analysés par la méthode de diffusion en gélose en 2013, 0,6 % (n = 8) étaient non sensibles au chloramphénicol (4 résistants et 4 intermédiaires), une baisse par rapport au taux de 9,4 % (n = 36) observé en 2010. La résistance à l’érythromycine a également diminué, passant de 14,4 % (n = 55) à 8,5 % (n = 110) de 2010 à 2013, tandis que la résistance à la clindamycine est demeurée relativement la même à 2.2 % (n = 29) (figure 54). Une résistance induite à la clindamycine a également été observée dans 6,9 % (n = 89) des isolats. Aucune résistance à la pénicilline et à la vancomycine n’a été observée.
Une non-sensibilité au chloramphénicol a été relevée dans un très petit nombre d’isolats appartenant notamment aux types emm11 (n = 1), emm89 (n = 2), emm92 (n = 1), emm110 (n = 1), emm115 (n = 1) et emm180 (n = 2) (tableau 11). Une résistance relativement élevée à l’érythromycine (macrolide) a été observée en ce qui concerne les isolats appartement aux types emm9 (72,7 %, n = 11), emm11 (61,7 %, n = 47), emm92 (87,5 %, n = 8) et emm114 (75,1 %, n = 12) (figure 55).
Figure 54. Résistance de S. pyogenes causant une infection invasive aux antimicrobiens, 2010-2013

CHL-NS = Non-sensibilité au chloramphénicol (résistant ou intermédiaire); ERY-R = Résistance à l'érythromycine; CLI R = Résistance constitutive à la clindamycine.
Résistance de S. pyogenes causant une infection invasive aux antimicrobiens, 2010-2013 - Description textuel
Ce figure représente le pourcentage de S. pyogenes qui sont non-sensible au chloramphénicol, résistantes à l'érythromycine et résistants à la clindamycine par année de 2010 à 2013.
| Année | CHL-NS | ERY-R | CLI-R |
|---|---|---|---|
| 2010 (n=383) | 9.4% | 14.4% | 2.3% |
| 2011 (n=1230) | 4.5% | 9.5% | 1.8% |
| 2012 (n=1122) | 2.0% | 10.1% | 2.6% |
| 2013 (n=1290) | 0.6% | 8.5% | 2.2% |
CHL-NS = Chloramphenicol non susceptible (resistant or intermediate); ERY-R = Erythromycin resistant; CLI-R = constitutively clindamycin resistant.
| Antimicrobien | Année | |||
|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | |
| CHL-NS* | 36 | 55 | 23 | 8 |
| ERY-R | 55 | 117 | 113 | 110 |
| CLI-R | 9 | 22 | 29 | 29 |
| CHL-NS = Non-sensibilité au chloramphénicol (résistant ou intermédiaire); ERY-R = Résistance à l’érythromycine; CLI-R = Résistance constitutive à la clindamycine. | ||||
| Type emm | Année de prélèvement (nbre d'échantillons testés) | |||
|---|---|---|---|---|
| 2010 (n=383) | 2011 (n=1230) | 2012 (n=1122) | 2013 (n=1290) | |
| 1 | 8,8% (n=7) | 4,8% (n=16) | 1,7% (n=5) | - |
| 2 | 22,2% (n=2) | 10,5% (n=2) | - | - |
| 3 | 9,1% (n=1) | 1,4% (n=2) | - | - |
| 4 | - | - | 1,8% (n=1) | - |
| 6 | - | - | 2,3% (n=1) | - |
| 9 | - | 12,5% (n=1) | - | - |
| 11 | 14,3% (n=1) | - | - | 2,1% (n=1) |
| 12 | 4,9% (n=2) | 4,9% (n=5) | - | - |
| 22 | - | 14,3% (n=1) | - | - |
| 28 | 5,6% (n=2) | 1,4% (n=1) | 2,8% (n=2) | - |
| 29 | - | - | 7,1% (n=1) | - |
| 41 | 50% (n=1) | - | 11,1% (n=1) | - |
| 48 | - | 25% (n=1) | - | - |
| 58 | - | 42,9% (n=3) | - | - |
| 59 | 13,3% (n=2) | - | - | - |
| 63 | - | 50% (n=1) | - | - |
| 68 | 100% (n=1) | - | 15,4% (n=2) | - |
| 73 | - | - | 11,1% (n=1) | - |
| 75 | - | 5,9% (n=1) | - | - |
| 80 | - | 9,1% (n=1) | - | - |
| 82 | - | 6,9% (n=2) | - | - |
| 83 | 20% (n=3) | - | 2,6% (n=1) | - |
| 87 | - | - | 2,9% (n=1) | - |
| 89 | 17,4% (n=8) | 12,3% (n=14) | 4,5% (n=5) | 1,7% (n=2) |
| 90 | - | 100% (n=1) | - | - |
| 91 | 33,3% (n=1) | - | - | - |
| 92 | - | - | 6,3% (n=1) | 12,5% (n=1) |
| 101 | 28,6% (n=2) | 11,1% (n=2) | - | - |
| 110 | - | - | - | 100% (n=1) |
| 114 | 37,5% (n=3) | 7,7% (n=1) | 5,9% (n=1) | - |
| 115 | - | - | - | 7,7% (n=1) |
| 183 | - | - | - | 100% (n=2) |
| Tous | 9,4% (n=36) | 4,5% (n=55) | 2% (n=23) | 0,6% (n=8) |
Figure 55. Types emm de S. pyogenes résistants au macrolide, 2013
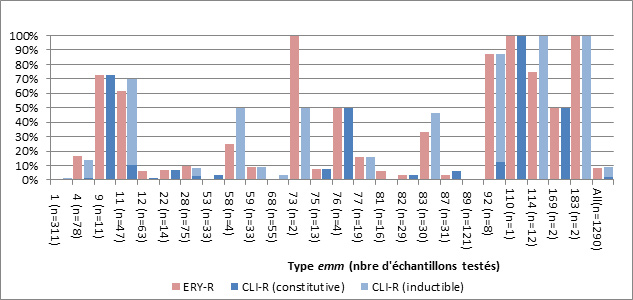
Types emm de S. pyogenes résistants au macrolide, 2013 - Description textuel
Ce figure représente la résistance de macrolides à différent types emm en 2013.
| Type emm | ERY-R | CLI-R (Constitutive) | CLI-R (Inductible) |
|---|---|---|---|
| 1 (n=311) | 0.6% | 0.3% | 0.3% |
| 4 (n=78) | 16.7% | 1.3% | 12.8% |
| 9 (n=11) | 72.7% | 72.7% | 0.0% |
| 11 (n=47) | 61.7% | 10.6% | 59.6% |
| 12 (n=63) | 6.3% | 1.6% | 0.0% |
| 22 (n=14) | 7.1% | 7.1% | 0.0% |
| 28 (n=75) | 9.3% | 2.7% | 5.3% |
| 53 (n=33) | 0.0% | 3.0% | 0.0% |
| 58 (n=4) | 25.0% | 0.0% | 50.0% |
| 59 (n=33) | 9.1% | 0.0% | 9.1% |
| 68 (n=55) | 0.0% | 0.0% | 3.6% |
| 73 (n=2) | 100.0% | 0.0% | 50.0% |
| 75 (n=13) | 7.7% | 7.7% | 0.0% |
| 76 (n=4) | 50.0% | 50.0% | 0.0% |
| 77 (n=19) | 15.8% | 0.0% | 15.8% |
| 81 (n=16) | 6.3% | 0.0% | 0.0% |
| 82 (n=29) | 3.4% | 3.4% | 0.0% |
| 83 (n=30) | 33.3% | 0.0% | 46.7% |
| 87 (n=31) | 3.2% | 6.5% | 0.0% |
| 89 (n=121) | 0.8% | 0.0% | 0.8% |
| 92 (n=8) | 87.5% | 12.5% | 75.0% |
| 110 (n=1) | 100.0% | 100.0% | 0.0% |
| 114 (n=12) | 75.0% | 0.0% | 100.0% |
| 169 (n=2) | 50.0% | 50.0% | 0.0% |
| 183 (n=2) | 100.0% | 0.0% | 100.0% |
| Tous (n=1290) | 8.5% | 2.2% | 6.9% |
Infection invasive à Streptococcus agalactiae (streptocoque du groupe B)
Sur les 416 isolats de Streptococcus agalactiae analysés par le LNM, 16 cultures (3,8 %) provenaient de cas d’infection précoce, soit de nourrissons de < 8 jours; 7 cultures (1,7 %) provenaient de cas d’infection tardive, soit de nourrissons de 8 à 31 jours; 4 cultures (1,0 %) provenaient d’enfants âgés de 1 mois à 14 ans, 177 (42,5 %), d’adultes de 15 à 64 ans et 165 (39,7 %), de personnes âgées de ≥ 65 ans. Quarante-sept isolats provenaient de personnes d’âge inconnu (tableau 12). Les isolats prélevés chez les hommes représentaient 52,8 % (n = 84) des 159 isolats pour lesquels on disposait de renseignements sur le sexe.
Le sérotype III était le plus fréquent, représentant 23,8 % (n = 99) de l’ensemble des isolats testés en 2013, et 62,5 % (n = 10) des isolats de cas d’infection précoce, 57,1 % (n = 4) des isolats de cas d’infection tardive et 50,0 % (n = 2) des isolats provenant d’enfants. Le sérotype V était le plus prédominant chez les adultes et les personnes âgées, correspondant à 24,3 % (n = 43) et à 20,6 % (n = 34) des isolats dans ces groupes d’âge, respectivement (tableau 12).
La majorité des streptocoques du groupe B provenait des isolats sanguins, à savoir 75,7 % (n = 315) de l’ensemble des isolats, suivi des isolats de liquide synovial, soit 4,8 % (n = 20) des isolats (figure 56). Le sérotype III représentait 25,7 % (n = 81) des isolats sanguins et 60,0 % (n = 6) des isolats de LCR, tandis que le sérotype II représentait 30,0 % (n = 3) des isolats de liquide péritonéal, et que le sérotype V représentait 50,0 % (n = 10) des isolats de liquide synovial et 25 % (n = 14) des isolats prélevés sur d’autres sites stériles (figure 57).
| Sérotype | Nourrisson (infection précoce) | Nourris-son (infection tardive) | Enfant | Adulte | Personne âgée | Âge non précisé | Total |
|---|---|---|---|---|---|---|---|
| Ia | - | 28,6%(2) | 50,0%(2) | 15,3%(27) | 18,8%(31) | 25,5%(12) | 17,8%(74) |
| Ib | - | 14,3%(1) | - | 15,3%(27) | 17,6%(29) | 10,6%(5) | 14,9%(62) |
| II | 6,3%(1)Footnote b | - | - | 15,3%(27) | 9,7%(16) | 8,5%(4) | 11,5%(48) |
| III | 62,5%(10) | 57,1%(4) | 50,0%(2) | 19,2%(34) | 18,2%(30) | 40,4%(19) | 23,8%(99) |
| IV | 25,0%(4) | - | - | 7,9%(14) | 9,7%(16) | 6,4%(3) | 8,9%(37) |
| V | 6,3%(1) | - | - | 24,3%(43) | 20,6%(34) | 8,5%(4) | 19,7%(82) |
| VI | - | - | - | - | 2,4%(4) | - | 1,0%(4) |
| VII | - | - | - | 0,6%(1) | 0,6%(1) | - | 0,5%(2) |
| VIII | - | - | - | 0,6%(1) | 1,2%(2) | - | 0,7%(3) |
| IX | - | - | - | 0,6%(1) | 0,6%(1) | - | 0,5%(2) |
| NT | - | - | - | 1,1%(2) | 0,6%(1) | - | 0,7%(3) |
| Total | (16) | (7) | (4) | (177) | (165) | (47) | (416) |
Figure 56. Sites d'isolement clinique de S. agalactiae
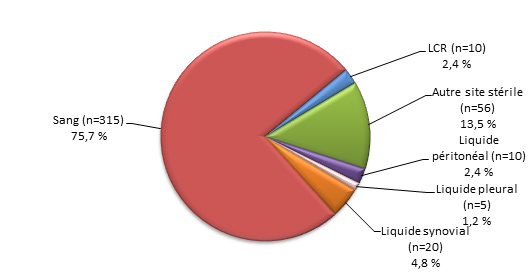
Sites d'isolement clinique de S. agalactiae - Description textuel
Ce figure représente le pourcentage de S. agalactiae constaté dans les différents sites d'isolement cliniques.
| Le site d'isolement clinique | Pourcentage |
|---|---|
| Sang (n=315) | 75.7% |
| LCR (n=10) | 2.4% |
| Autre site stérile (n=56) | 13.5% |
| Liquide péritonéal (n=10) | 2.4% |
| Liquide pleural (n=5) | 1.2% |
| Liquide synovial (n=20) | 4.8% |
Figure 57. Sérotypes de S. agalactiae causant une infection invasive, par site d'isolement clinique
Sérotypes de S. agalactiae causant une infection invasive, par site d'isolement clinique - Description textuel
Ce figure représente le pourcentage de S. agalactiae constaté dans les différents sites d'isolement cliniques par sérotype.
| Sérotype | Sang (n=315) | LCR (n=10) | Autre site stérile (n=56) | Liquide péritonéal (n=10) | Liquide pleural (n=5) | Liquide synovial (n=20) |
|---|---|---|---|---|---|---|
| Ia | 18.4% | 20.0% | 10.7% | 20.0% | 40.0% | 20.0% |
| Ib | 15.9% | 10.0% | 12.5% | 10.0% | 40.0% | 5.0% |
| II | 11.1% | 0.0% | 14.3% | 30.0% | 0.0% | 10.0% |
| III | 25.7% | 60.0% | 16.1% | 20.0% | 0.0% | 5.0% |
| IV | 7.3% | 10.0% | 17.9% | 10.0% | 20.0% | 5.0% |
| IX | 0.3% | 0.0% | 0.0% | 0.0% | 0.0% | 5.0% |
| NT | 1.0% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| V | 18.1% | 0.0% | 25.0% | 10.0% | 0.0% | 50.0% |
| VI | 1.0% | 0.0% | 1.8% | 0.0% | 0.0% | 0.0% |
| VII | 0.6% | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| VIII | 0.6% | 0.0% | 1.8% | 0.0% | 0.0% | 0.0% |
| Sérotype | Sang | LCR | Autre Site Stérile | Liquide Péritonéal | Liquide Pleural | Liquide Synovial |
|---|---|---|---|---|---|---|
| NT | 3 | |||||
| IX | 1 | 1 | ||||
| VIII | 2 | 1 | ||||
| VII | 2 | |||||
| VI | 3 | 1 | ||||
| V | 57 | 14 | 1 | 10 | ||
| IV | 23 | 1 | 10 | 1 | 1 | 1 |
| III | 81 | 6 | 9 | 2 | 1 | |
| II | 35 | 8 | 3 | 2 | ||
| Ib | 50 | 1 | 7 | 1 | 2 | 1 |
| Ia | 58 | 2 | 6 | 2 | 2 | 4 |
Figure 58. Distribution régionale des sérotypes de S. agalactiae causant une infection invasive, 2013
* Nombre d'isolats pour les régions de l'Ouest, Centrale et de l’Est, respectivement. Pour améliorer l’échelle, 2 isolats contenant le sérotype IV et représentant 100 % des isolats de la région de l’Est ont été exclus de la figure.
Distribution régionale des sérotypes de S. agalactiae causant une infection invasive, 2013 - Description textuel
Ce figure représente la distribution des sérotypes de S. agalactiae à travers les régions de l'ouest et du centre du Canada en 2013.
| Sérotype | Ouest (n=163) | Centre (n=251) |
|---|---|---|
| Ia (23,51,0)* | 14.1% | 20.3% |
| Ib (18,44,0) | 11.0% | 17.5% |
| II (26,22,0) | 16.0% | 8.8% |
| III (36,63,0) | 22.1% | 25.1% |
| IV (21,14,2) | 12.9% | 5.6% |
| V (33,49,0) | 20.2% | 19.5% |
| VI (2,2,0) | 1.2% | 0.8% |
| VII (0,2,0) | 0.0% | 0.8% |
| VIII (3,0,0) | 1.8% | 0.0% |
| IX (0,2,0) | 0.0% | 0.8% |
| NT (1,2,0) | 0.6% | 0.8% |
Note: Pour améliorer l'échelle, 2 isolats contenant le sérotype IV et représentant 100 % des isolats de la région de l'Est ont été exclus de la figure.
Résistance de S. agalactiae aux antimicrobiens
Sur les 420 isolats de S. agalactiae causant une infection invasive analysés en 2013 par la méthode de diffusion en gélose, 1,9 % (n = 8) étaient non sensibles au chloramphénicol (2 intermédiaires et 6 résistants), un taux demeuré relativement constant depuis 2010 (2,1 %, n = 5). La résistance au macrolide a augmenté de même que la résistance à la clindamycine, passant de 25,5 % (n = 84) à 28,8 % (n = 121) des isolats, et la résistance à l’érythromycine est passée de 40,7 % (n = 134) à 48,8 % (n = 205) entre 2012 et 2013 (figure 59).
Comme pour l’autre streptocoque bêta-hémolytique du groupe A, une non-sensibilité au chloramphénicol chez les streptocoques du groupe B a été observée pour un très petit nombre d’isolats, notamment les sérotypes Ia (n = 1), III (n = 2), IV (n = 1) et V (n = 4) (tableau 15). Une résistance relativement élevée à l’érythromycine (macrolide) a été observée pour le sérotype II (59,6 %, n = 28), IV (60,5 %, n = 23) et V (61,2 %, n = 52) (figure 60).
Figure 59. Résistance aux antimicrobiens des isolats de S. agalactiae causant une infection invasive, 2010 - 2013
CHL-NS = Non-sensibilité au chloramphénicol (résistant ou intermédiaire); ERY-R = Résistance à l'érythromycine; CLI R = Résistance constitutive à la clindamycine
Résistance aux antimicrobiens des isolats de S. agalactiae causant une infection invasive, 2010 - 2013 - Description textuel
Ce figure représente le pourcentage de antimicrobiens résistants S. agalactiae (IGBS) isolé par année de 2010 à 2013.
| Année | CHL-NS | ERY-R | CLI-R |
|---|---|---|---|
| 2010 (n=235) | 2.1% | 39.1% | 24.3% |
| 2011 (n=392) | 2.6% | 41.6% | 25.5% |
| 2012 (n=329) | 2.7% | 40.7% | 25.5% |
| 2013 (n=420) | 1.9% | 48.8% | 28.8% |
CHL-NS = Non-sensibilité au chloramphénicol (résistant ou intermédiaire); ERY-R = Résistance à l’érythromycine; CLI R = Résistance constitutive à la clindamycine
| Antimicrobiens | Année | |||
|---|---|---|---|---|
| 2010 | 2011 | 2012 | 2013 | |
| CHL-NS | 5 | 10 | 9 | 8 |
| ERY-R | 92 | 163 | 134 | 205 |
| CLI-R | 57 | 100 | 84 | 121 |
| CHL-NS = Non-sensibilité au chloramphénicol (résistant ou intermédiaire); ERY-R = Résistance à l’érythromycine; CLI R = Résistance constitutive à la clindamycine | ||||
| Sérotype | Année de prélèvement (nbre d'échantillons testés) | |||
|---|---|---|---|---|
| 2010 (n=235) | 2011 (n=392) | 2012 (n=329) | 2013 (n=420) | |
| Ia | 4,8% (n=2) | - | 1,8% (n=1) | 1,4% (n=1) |
| Ib | - | - | - | - |
| II | - | - | 3,4% (n=1) | - |
| III | 1,7% (n=1) | 2,1% (n=2) | 3,5% (n=3) | 2% (n=2) |
| IV | 5,3% (n=1) | 9,8% (n=4) | 11,8% (n=4) | 2,6% (n=1) |
| V | 1,8% (n=1) | 3,8% (n=3) | - | 4,7% (n=4) |
| VI | - | - | - | - |
| VII | - | - | - | |
| VIII | - | 16,7% (n=1) | - | |
| IX | - | - | - | - |
| NT | - | - | - | - |
| Tous | 2,1% (n=5) | 2,6% (n=10) | 2,7% (n=9) | 1,9% (n=8) |
Figure 60. Résistance des sérotypes de S. agalactiae au macrolide, 2013
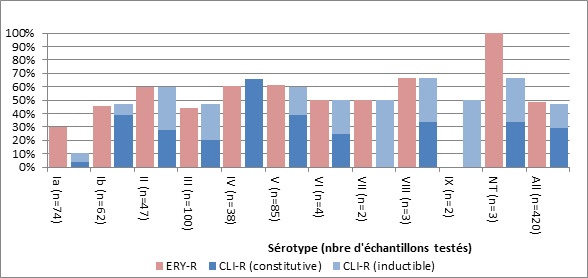
Résistance des sérotypes de S. agalactiae au macrolide, 2013 - Description textuel
Ce figure représente le pourcentage de résistants macrolide (résistant à l'érythromycine, la clindamycine résistant constitutive et inductible à la clindamycine résistant) S. agalactiae par le sérotype en 2013.
| Sérotype | ERY-R | CLI-R (Constitutive) | CLI-R (Inductible) |
|---|---|---|---|
| Ia (n=74) | 29.7% | 4.1% | 6.8% |
| Ib (n=62) | 45.2% | 38.7% | 8.1% |
| II (n=47) | 59.6% | 27.7% | 31.9% |
| III (n=100) | 44.0% | 20.0% | 27.0% |
| IV (n=38) | 60.5% | 65.8% | 0.0% |
| V (n=85) | 61.2% | 38.8% | 21.2% |
| VI (n=4) | 50.0% | 25.0% | 25.0% |
| VII (n=2) | 50.0% | 0.0% | 50.0% |
| VIII (n=3) | 66.7% | 33.3% | 33.3% |
| IX (n=2) | 0.0% | 0.0% | 50.0% |
| NT (n=3) | 100.0% | 33.3% | 33.3% |
| Tous (n=420) | 48.8% | 28.8% | 17.9% |
Conclusion
Dans le cas de S. pneumoniae, la proportion des sérotypes contenus dans le vaccin Pneu-C-7 demeure faible et elle continue de diminuer dans les sérotypes prédominants (7F et 19A) du vaccin Pneu-C-13. Cette diminution, associée à la diminution constante de l’incidence des infections invasives chez les enfants, donne une preuve des effets des programmes d’immunisation au vaccin Pneu-C-13 au Canada. Toutefois, il faut continuer à être vigilant dans le but de reconnaître les hausses potentielles touchant des sérotypes non contenus dans le vaccin Pneu-C-13 en circulation au Canada, comme les sérotypes 22F, 23A et 23B. Le sérotype 3, bien qu’il se retrouve dans le vaccin Pneu-C-13, n’a pas suivi le même déclin que les autres sérotypes constitutifs, ce qui soulève des préoccupations quant à la virulence, à l’immunogénicité et à l’efficacité de ce sérotype. Surveillance continue de la fréquence relative des sérotypes présents au Canada nous guidera dans la mise au point et le choix des composants des nouveaux vaccins, de manière à réduire le fardeau global de la maladie.
De manière générale, la résistance des isolats de S. pneumoniae aux antimicrobiens diminue, principalement à cause du déclin du sérotype 19A qui est très résistant; toutefois, la résistance aux pénicillines, aux macrolides, aux tétracyclines, aux sulfonamides et aux fluoroquinolones, ainsi que la multirésistance sont encore fréquentes. À l’heure actuelle, les taux de résistance aux antimicrobiens sont relativement faibles au Canada, mais il est essentiel de surveiller les profils de sensibilité aux antimicrobiens des sérotypes communs de S. pneumoniae si l’on veut orienter les traitements empiriques et les traitements ciblés.
Les types emm1 et emm89 de S. pyogenescontinuent d’être les souches dominantes au Canada, tandis que la variabilité annuelle des autres types emm évolue. La résistance du streptocoque du groupe A aux antimicrobiens a diminué depuis 2010. Toutefois, étant donné la gravité des infections à streptocoque du groupe A, leur risque élevé de survenue et la sensibilisation croissante du public, il importe de maintenir un suivi et une surveillance des sérotypes en circulation et des taux de résistance aux antimicrobiens pour être en mesure de repérer les éclosions et pour guider les interventions en santé publique.
Les sérotypes III et IV de S. agalactiaesont les souches prédominantes au Canada, et la résistance aux antimicrobiens est à la hausse, surtout la résistance aux macrolides. Le SGB entraîne des complications graves chez les nouveau-nés; on observe toutefois une augmentation du fardeau de la maladie chez les adultes. Il est important de surveiller les changements dans la répartition des sérotypes et les taux de résistance aux antimicrobiens, ainsi que de recueillir des données épidémiologiques supplémentaires améliorées pour pouvoir déterminer les facteurs de risque potentiels, la propagation des souches invasives et pour mieux faire connaître les options futures de prévention et de traitement.
Annexe
| Groupe d'âge | Nombre total d'isolats sérotypés | Nombre total de cas de maladie déclarés au SSMDOFootnote ** | Pourcentage de cas sérotypés |
|---|---|---|---|
| <1 an | 51 | 68 | 75,0% |
| 1 à 4 ans | 153 | 172 | 89,0% |
| 5 à 39 ans | 345 | 444 | 77,7% |
| 40 à 59 ans | 678 | 851 | 79,7% |
| ≥60 ans | 1316 | 1627 | 80,9% |
| Tous âges | 2577Footnote * | 3162 | 81,5% |
| Groupe d'âge | Nombre total d'isolats | Nombre total de cas de maladie déclarés au SSMDCFootnote ** | Pourcentage de cas |
|---|---|---|---|
| <1 an | 43 | 37 | 116,2% |
| 1 à 4 ans | 34 | 71 | 47,9% |
| 5 à 39 ans | 366 | 497 | 73,6% |
| 40 à 59 ans | 389 | 517 | 75,2% |
| ≥60 ans | 421 | 536 | 78,5% |
| Tous âges | 1294Footnote * | 1658 | 78,0% |
Références
- Agence de la santé publique du Canada. 2014. Maladies à déclaration obligatoire en direct. http://dsol-smed.phac-aspc.gc.ca/dsol-smed/ndis/graphiques.php?c=pl [consultation le 4 septembre 2004).
- Agence de la santé publique du Canada. 2014. Programmes d’immunisation subventionnés par l’État au Canada – Calendrier d’immunisation systématique des nourrissons et des enfants incluant les programmes de rappel (en date de septembre 2014) [en ligne]. Accessible à l’adresse : http://www.phac-aspc.gc.ca/im/ptimprog-progimpt/table-1-fra.php [consultation le 17 octobre 2013].
- Austrian R. The Quellung reaction, a neglected microbiological technique. 1976. Mt. Sinai J. Med. 43:699–709.
- Bettinger JA, Scheifele DA, Kellner JD, et al. 2010. The effect of routine vaccination on invasive pneumococcal infections in Canadian children, Immunization Monitoring Program, Active 2000–2007. Vaccine 28:2130–2136.
- Bjornson G, Scheifele DW, Bettinger J, et al. 2007. Effectiveness of Pneumococcal Conjugate Vaccine in Greater Vancouver, Canada: 2004-2005. Ped.Inf.Dis.J. 26(6):540-542.
- Bruce, MG, Deeks SL, Zulz T, et al. 2008. International Circumpolar Surveillance System for Invasive Pneumococcal Disease, 1999-2005. Emerging.Infect.Dis. 14(1):25-33.
- Clinical Laboratory Standards Institute (CLSI). 2008. Interpretative Criteria for Identification of Bacteria and Fungi by DNA Target Sequencing, Approved Guideline. MM18-A; 28(12):21-24.
- Clinical Laboratory Standards Institute (CLSI). January 2011. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-First Informational Supplement. M100-S21; 31(1).
- Comité consultatif national de l’immunisation (CCNI). 2010 (novembre). Une déclaration d’un comité consultatif (DCC), Mise à jour sur l’utilisation des vaccins conjugués contre le pneumocoque chez les enfants. RMTC 36(ACS-12):1-23.
- Cunningham MW. 2000 (July). Pathogenesis of Group A Streptococcal Infections. Clin. Micro. Rev. 470-511.
- Définitions nosologiques des maladies transmissibles faisant l’objet d’une surveillance nationale – 2009. Novembre 2009. RMTC 35S2. Accessible à l’adresse : http://www.phac-aspc.gc.ca/publicat/ccdr-rmtc/09vol35/35s2/Pneumoco-fra.php.
- Demczuk WHB, Martin I, Griffith A, et al. 2012. Serotype distribution of invasive Streptococcus pneumoniae in Canada during the introduction of the 13-valent pneumococcal conjugate vaccine, 2010. Can.J.Microbiol. = Rev. ca. microbiol. 58:1008-1017.
- Demczuk WHB, Martin I, Griffith A, et al. 2013. Serotype distribution of invasive Streptococcus pneumoniae in Canada after the introduction of the 13-valent pneumococcal conjugate vaccine, 2010-2012. Can. J. Microbiol. = Rev. ca. microbiol. 59:778-788.
- Deng X, Church D, Vanderkooi OG, et al. 2013. Streptococcus pneumoniae infection: a Canadian perspective. Expert Rev. Anti Infect. Ther. 11(8):781-791.
- De Wals P, Lefebvre B, Defay F, et al. 2012. Invasive pneumococcal diseases in birth cohorts vaccinated with PCV-7 and/or PHiD-CV in the province of Quebec, Canada. Vaccine 30:6416-6420.
- Drancourt M, Roux V, Fournier PE, Raoult D. 2004 (February). rpoB gene sequence-based identification of aerobic gram-positive cocci of the genera Streptococcus, Enterococcus, Gemella, Abiotrophia and Granulicatella. J. Clin. Micro. 42(2):497-504.
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). 2012. Clinical Breakpoint Tableau. Version 2.0. Accessible à l’adresse : http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/Breakpoint_table_v_2.0_120221.pdf
- Kellner, JD, Sheifele, D, Vanderkooi, OG, et al. 2008. Effects of Routine Infant Vaccination with the 7-Valent Pneumococcal Conjugate Vaccine on Nasopharyngeal Colonization with Streptococcus pneumoniae in Children in Calgary, Canada. The Ped. Infect. Dis. Journal. 27(6):526-532.
- Kellner JD, Vanderkooi OG, MacDonald J, Church DL, Tyrrell GJ, Scheifele DW. 2009 (July). Changing epidemiology of invasive pneumococcal disease in Canada, 1998-2007: update from the Calgary-area Streptococcus pneumoniae research (CASPER) study. Clin Infect Dis. 49(2):205-12.
- Lamangni, TL, Keshishian C, Efstratiou A, et al. 2013. Emerging Trends in the Epidemiology of Invasive Group B Streptococcal Disease in England and Wales, 1991 – 2010. Clin Inf Dis 2013;57(5):682-8.
- Lim, GH, Wormsbecker, AE, McGeer A, et al. 2013. Have changing pneumococcal vaccination programmes impacted disease in Ontario? Vaccine 31:2680-2685.
- Lovgren M, Spika JS, Talbot JA. 1998. Invasive Streptococcus pneumoniae infections: serotype distribution and antimicrobial resistance in Canada, 1992-1995. Can.Med.Assoc.J. 158(3):327-331.
- Marchessault V, éditeur. 2002. Guide canadien d’immunisation. 6e éd. Ottawa, Association médicale canadienne.
- Merck & Co. Inc., Whitehouse Station, NJ 08889, USA. Pneumovax® 23 (vaccin polyvalent antipneumococcique).
- McIntosh ED, Reinert RR. 2011 (Jan). Global prevailing and emerging pediatric pneumococcal serotypes. Expert Rev Vaccines. 10(1):109-29.
- Minnesota Department of Health, Infectious Disease Epidemiology, Prevention and Control Division. Accessible à l’adresse : http://www.health.state.mn.us/divs/idepc/dtopics/invbacterial/sterile.html.
- Robinson KA, Baughman W, Rothrock G, Barrett NL, Pass M, Lexau C, et al. 2001 (April).
- Epidemiology of invasive Streptococcus pneumoniae infections in the United States, 1995–1998: opportunities for prevention in the conjugate vaccine era. JAMA. 285(13):1729–35.
- Schuchat A, Robinson K, Wenger JD, Harrison LH, Farley M, Reingold AL, et al. 1997.
- Bacterial meningitis in the United States in 1995. Active Surveillance Team. N Engl J Med;337(October (14)):970–6.
- Schwartz B, Facklam RR, Breiman RF. 1990. Changing epidemiology of group A streptococcal infection in the USA. Lancet 336:1167-1171.
- Scott JA, Hall AJ, Dagan R, Dixon JM, Eykyn SJ, Fenoll A, et al. 1996 (June). Serogroup-specific epidemiology of Streptococcus pneumoniae: associations with age, sex, and geography in 7,000 episodes of invasive disease. Clin Infect Dis. 22(6):973–81.
- Shahidi N, Dhaliwa J, Tyrrell G, et al. 2008. Trends in incidence of invasive pneumococcal disease following introduction of the universal infant immunization program in British Columbia, 2001 – 2006. BC Medical Journal 50(1):18-21.
- Siljander T, Lyytikäinen O, Vähäkuopus S, et al. 2010. Epidemiology, outcome and emm types of invasive group A streptococcal infections in Finland. Eur J Clin Microbiol Infect Dis; 29:1229-1235.
- Spellerberg B, Brandt C. Streptococcus. In: Murray PR, Baron EL, Jorgensen JH, Landry ML, Pfaller MA. editors. 2007. Manual of Clinical Microbiology. 9th ed. Washington: American Society for Microbiology; p. 412-429.
- Tyrrell GJ, Lovgren M, Chui N, et al. 2009. Serotypes and antimicrobial susceptibilities of invasive Streptococcus pneumoniae pre- and post-seven valent pneumococcal conjugate vaccine introduction in Alberta, Canada 2000-2006. Vaccine; 27:3553-3560.
- Weinberger DM, Malley R, Lipsitch M. 2011 (April). Serotype replacement in disease after pneumococcal vaccination. The Lancet, Accessible en ligne : ISSN 0140-6736, DOI: 10.1016/S0140-6736(10)62225-8. (http://www.sciencedirect.com/science/article/B6T1B-52M217X-4/2/f3141605bd8e55b78bbc1df8f2dd8677)