
Beluga whale (Delphinapterus leucas) COSEWIC assessment and status report: chapter 8
Population Identification
- St. Lawrence Estuary Population
- Ungava Bay Population
- Eastern Hudson Bay Population
- Western Hudson Bay Population
- Eastern High Arctic - Baffin Bay Population
- Cumberland Sound Population
- Eastern Beaufort Sea Population
- Problems with the Identification of Beluga Populations in Canada
In the past, for management purposes, seven separate populations of belugas were recognized to be present in Canadian waters during at least part of the year (Fig. 2). These consist of (1) the St. Lawrence Estuary population (Pippard 1985) (2) the Ungava Bay population; (3) the Eastern Hudson Bay population (Reeves and Mitchell 1989, DFO 2001); (4) the Western Hudson Bay population (Richard 1993); (5) the Eastern High Arctic - Baffin Bay population (Doidge and Finley 1993); (6) the Cumberland Sound population (Richard and Orr 1991, DFO 2002a); (7) the Eastern Beaufort Sea population (DFO 2000).
Figure 2: Location of the of Canadian Beluga Populations: (1) St. Lawrence Estuary Population (2) Ungava Bay Population (3) Eastern Hudson Bay Population (4) Western Hudson Bay Population (5) Eastern High Arctic – Baffin Bay Population (6) Cumberland Sound Population (7) Eastern Beaufort Sea Population
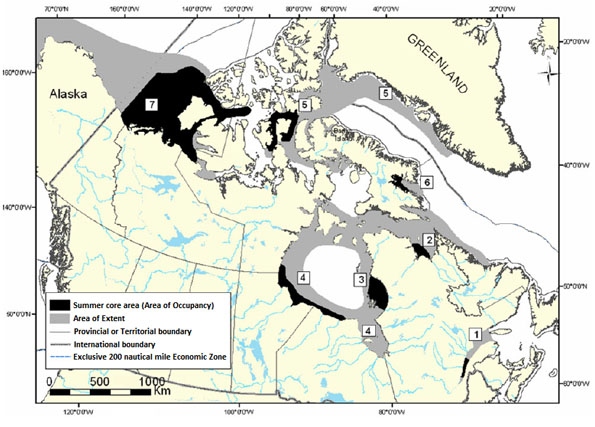
Modified from DFO 2002b.
Using this scheme of population division, the Canadian populations of belugas all have estuarine centres of aggregation during the summer open-water season. In most cases their summer coastal and offshore distribution is separate from other populations. Autumn, winter, and spring ranges are often contiguous or overlapping with other populations.
The first evidence for discriminating between beluga populations was based on apparent size differences between belugas collected in different parts of the Arctic (Sergeant and Brodie 1969). Reexamination of the data and methods of analysis confirm that minor differences do occur, but not to the degree that they might be practically used as criteria to differentiate closely adjacent beluga populations (Doidge 1990). Stewart (1994) found significant size differences between several populations, which were well separated geographically, but notes that body size differences are of waning importance to population identity issues, especially in the light of new genetic population discrimination techniques.
More generally accepted was the delineation of geographical populations based on their summer distributions, which were thought to be mainly coastal and centred on sites such as the estuaries where significant numbers of individuals gathered (Sergeant and Brodie 1975, Smith et al. 1985). This was supported by behavioural studies in estuaries, which showed that belugas are philopatric and strongly site tenacious (Caron and Smith 1990, Smith et al. 1994). In one of those estuaries, individuals known by distinct scars were seen to return frequently, and in a short time, to the estuary after hunting had taken place (Caron and Smith 1990).
Recently, emphasis has been placed on the use of genetic analyses, both mitochondrial DNA (mtDNA) and microsatellites to differentiate populations of belugas (de March et al. 2001, 2002) (Fig. 3). There has been a gradual realization that there are a number of complex problems associated with the representative sampling of animals, the scope and breath of the genetic analyses themselves, and their standardization between studies, as well as in the types of statistical analyses which are appropriate in analysing the complex genetic data sets (de March et al. 2002, Palsbøll et al. 2002, de March and Postma 2003). These genetic studies confirm the existence of some previously defined populations. They also indicate the existence of new populations. Other types of studies, aimed at defining the timing of migration and routes, used satellite-tagged belugas (Martin et al. 1993), detaÎled aboriginal traditional knowledge, and other markers such as contaminant signatures (Innes et al. 2002a).
This report attempts to determine Designatable Units based on range disjunction and genetic divergence (COSEWIC Appendix F5 2003). In recent years, new genetic studies have indicated that there are more beluga populations than was once thought, but because of sampling inadequacies, have not yet been able to define these precisely or practically for management purposes. Since none of the new genetic data contradict the existence of the previously recognized populations, which have all received status designations, these will form the basis and departure point of the present review. Where there exist strong genetic, range, migration or other ecological evidence for designating other Evolutionarily Significant Units (Waples 1991), these are noted.
Figure 3: Beluga mtDNA Haplotype Frequencies at 13 Locations in North America and Greenland
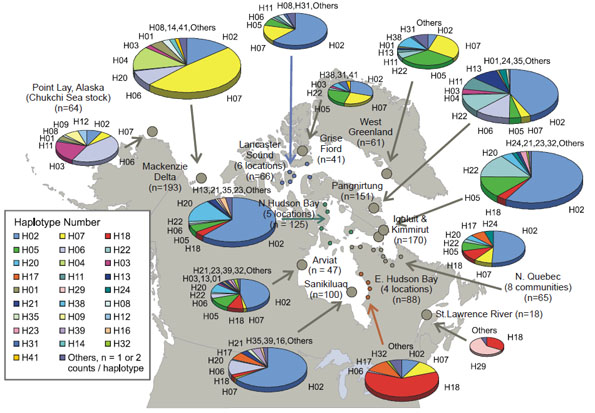
Areas of the pies are proportional to sample size (from de March et al. 2002, p 25, figure 2).
The St. Lawrence Estuary population has long been presumed to be a relict Arctic population. Recognition of the discreteness of this small population of belugas, mainly centred near the mouth of the Saguenay River, relied on the absence of any significant numbers of belugas in the areas contiguous to their location. Few belugas are found along the north shore of the St. Lawrence or the south Labrador coast, confirming the lack of an extant connection between the belugas of the St. Lawrence and those populations occupying the sub-Arctic coastline of Quebec (Reeves and Mitchell 1984, Michaud et al. 1990, Curren and Lien 1998).
The St. Lawrence belugas are more genetically distinct than all other Canadian beluga populations. This population has one haplotype which has not yet been found anywhere else. It contains another haplotype which is common only in belugas of eastern Hudson Bay, particularly those sampled in the Nastapoka River Estuary. The geographically isolated St. Lawrence population and the Eastern Hudson Bay population show the lowest diversities of mtDNA haplotypes and microsatellite alleles within Canadian populations (de March et al. 2002). It has been postulated that both the St. Lawrence and Eastern Hudson Bay beluga might have originated from the inland sea of the Lakes Agassiz and Ojibway, a refugium extant during the Wisconsin period of glaciation. More recently, de March et al. (2002) postulate that the Eastern Hudson Bay belugas might have been the first arrivals from an Atlantic refugium after the first ice barrier disappeared some 8,000 B.P. (Fulton 1989). It is postulated that all the other Canadian populations originated from a western refugium (Brown 1996, Brown-Gladden et al. 1997, Brown-Gladden et al. 1999). This would have included the other eastern Arctic populations and that of Western Hudson Bay. More studies are needed, especially of belugas in western Russian waters, in order to elucidate the post-glacial dispersion of this species.
The belugas of the postulated Ungava Bay population were defined by their summer distribution, primarily centred near the Mucalic River estuary, although other small estuarine concentrations, such as the George and Whale Rivers, were also known. Never very large (Reeves and Mitchell 1989), this population was reduced significantly by both commercial and subsistence hunting. The Mucalic River no longer is a significant gathering place for belugas (T. Smith unpublished data, personal observations 1985). Because of the low numbers of animals taken in this area in recent years, there have been few samples obtained to study the genetic structure. Mancuso (1995), from a small sample of belugas (n=11) taken in the Ungava Bay area, found the diversity of haplotypes to be higher than expected given the small population size (Smith and Hammill 1986). This has since been corroborated by further sampling and analyses (de March and Maiers2001) in which high haplotype diversity was demonstrated in 33 belugas from several locations in the Ungava Bay area, although there are concerns that the locations of some of these kills were not within the core range of the Ungava Bay population, and thus that they may be animals from other populations. The high genetic diversity leads to the hypothesis that animals in Ungava Bay area now are part of one or more other populations. Since the main centres of aggregation, such as the Mucalic, are now almost unoccupied, we may never know if Ungava belugas had different allele frequencies than belugas from other areas of the Arctic.
All available genetic, distribution, and abundance data indicates that this population is at undetectably low levels or has been extirpated.
Belugas were fairly common along the northern Labrador coast in summer until the 1950s, but have been very scarce since (Brice-Bennett 1978). The Labrador Inuit Association receives reports of approximately a dozen summer sightings, and 2-3 summer catches, of belugas per year (J. Rowell, Labrador Inuit Association, pers. comm., 2004). Based upon geographic proximity, these may have had affinities with the Ungava Bay population.
This population was originally defined by its centres of estuarine concentration at the Nastapoka and Little Whale Rivers, with the main area of summer coastal occupation being from Kujjuarapik to Inukjuak. Aerial surveys, which began in 1983, showed that there was an offshore distribution of animals throughout this area as far offshore as the Belcher Islands (Smith and Hammill 1986, Kingsley 2000, Gosselin et al. 2002).
Mitochondrial DNA analyses have shown that there is a genetic difference between eastern and western Hudson Bay belugas (Mancuso 1995, Brennin et al. 1997, Brown-Gladden et al. 1997, de March and Postma 2003). Further analyses of mitochrondrial DNA and microsatellites showed that Belcher Island belugas are consistently different from both Eastern Hudson Bay and Western Hudson Bay populations. These belugas may be a separate population or a consistent mixture of more than one population (de March and Postma 2003).
The southern boundary of the Eastern Hudson Bay population has not been clearly delineated. Sanikiluaq, in the Belcher Islands, close to the summer range of the Eastern Hudson Bay belugas, appears to harvest whales either from a separate population or from a consistent mixture of other populations. A large group of belugas spends the summer months in James Bay, an area contiguous to the south with both the Eastern Hudson Bay population and Sanikiluaq. Belugas also occupy the northern Ontario coastline from James Bay west to Churchill, Manitoba. The few genetic samples from these neighboring animals that have been analysed indicate relatedness to the Western Hudson Bay population (B. de March, pers. com.). Recent tagging of belugas in the Nelson River, Manitoba, also shows that Western Hudson Bay belugas frequent the northern Ontario coastline and travel into James Bay (P. Richard pers. com.).
Belugas taken during the spring and autumn hunts, along the south coast of the Hudson Strait, contain a mixture of genotypes as do the small number of belugas actually harvested in Ungava Bay. These areas receive belugas from Eastern Hudson Bay (Bourdages et al. 2002), Western Hudson Bay and southeastern Baffin Island because of the favorable winter ice regime immediately to the north of Ungava Bay and in the offshore areas of Hudson Strait (de March and Postma 2003).
There appears to be a strong genetic basis for designating belugas of Eastern Hudson Bay as a separate population and increasingly good evidence that they contribute to the harvests in other Nunavik communities as far distant as Ungava Bay.
The belugas of this population contain many of the haplotypes common to all other Canadian populations. However, the haplotypes most common in Eastern Hudson Bay, the St. Lawrence Estuary, and some from the Beaufort Sea population are relatively rare in Western Hudson Bay (de March and Postma 2003). Western Hudson Bay genetic samples were obtained from communities as far distant from one another as Churchill, Manitoba, Sanikiluaq, Repulse Bay, and the Northwest Territories, and have a high haplotype and microsatellite diversity. This group, as a whole, is thus genetically less homogeneous than the Eastern Hudson Bay belugas. For example, in recent studies only 75/103 (73%) of Western Hudson Bay samples were correctly reassigned to their population of origin as compared to 79/96 (83%) in Eastern Hudson Bay (de March and Postma 2003).
It is possible that the Churchill area belugas from the Western Hudson Bay represent a distinct population centred on the Churchill River since a large percentage of these animals all contain a common haplotype (de March and Postma 2003). Because Western Hudson Bay has been considered one large population and exploitation is assumed to be sustainable, little genetic sampling has been done to cover the whole range of this presumed, but little studied, single population. The genetic identities of Western Hudson Bay belugas are, however, of interest to managers since they travel into Hudson Strait and form part of the annual harvest of the southeast Baffin and the Hudson Strait communities, which also harvest belugas from depleted populations in northern Quebec. Animals summering off southeast Baffin Island, but outside Cumberland Sound, are (in the scheme used here) also tentatively assigned to the Western Hudson Bay population (see below).
Belugas occupying the Canadian High Arctic regions of Lancaster Sound, Barrow Strait, Peel Sound and Baffin Bay during the summer open water season were commonly assumed to occupy the ice-free or pack–ice area along the West Greenland coast during the winter months. High harvests of belugas have been common in West Greenland during the last century (Reeves and Mitchell 1987).
In the 1980s, prior to genetic sampling and satellite telemetry studies, aerial surveys documented the presence of belugas in the pack-ice areas as far south as 66°N along the Greenland side of Baffin Bay (Koski and Davis 1979). The western side of Baffin Bay, along the Canadian Baffin Island coast, has always had much more extreme land-fast ice and heavy concentrations of pack ice. Late winter aerial surveys found that an unknown number of belugas apparently remained to over-winter in the north Baffin Bay area of the North Water polynya (Finley and Renaud 1980, Stirling 1980, Richard et al. 1998a).
From 1987 to 2000 a significant number of belugas were tagged with satellite transmitters in the Canadian High Arctic, but only one of 39 animals was observed to move into the West Greenland area (Smith and Martin 1994, Richard et al. 1998b, Reeves and St. Aubin 2001). In 2001, 3 of 5 belugas tagged in Creswell Bay in the Canadian High Arctic, ended up in West Greenland (Heide-Jørgensen et al. 2003). This was the same location from which originated the only previous tagged beluga to end up in West Greenland (Smith and Martin 1994). A reanalysis of all belugas that retained their tags after October 1 (n=26), reveals that 15% of these animals ended up in West Greenland. This is still based on a small sample size and there are problems with the apparent non-random distribution of West Greenland belugas whÎle they are in their summer Canadian High Arctic habitat. This makes it difficult to obtain a clear picture on the proportion of belugas from the Canadian High Arctic which might over-winter in either the North Water / Lancaster Sound area, or along the West Greenland coast (Heide-Jørgensen et al. 2003).
Genetic evidence also suggests that at least two populations inhabit this large geographic area (de March et al. 2002). A high percentage of the belugas sampled from West Barrow Strait and Somerset Island locations have the most common Western Hudson Bay haplotype, which is less common in samples from Peel Sound and West Greenland (de March et al. 2002). One of the common haplotypes from Peel Sound is also common in West Greenland and the Beaufort Sea, but less common in West Barrow Strait and Somerset Island. Microsatellite allele frequencies also differ significantly in these two areas.
Genetic results to date point to overall differences between Lancaster Sound and Greenland belugas (de March et al. 2002). However, individual lots of samples from the same location, taken at different times, show different affiliations confirming that a mixture of populations coexist during the summer and also during the fall and spring movements. A clear picture of the number and geographical distribution of these populations will not emerge until much more systematic sampling is done.
Inuit from several West Greenland communities report the presence of distinctive belugas in their catches, which they assume come from Canadian waters (Thomsen 1993). These are generally described as longer animals that have an elongate caudal peduncle and are thought to be adapted to swimming in areas with strong currents. Some hunters also mention belugas with long teeth, which they term kigutikaat. Another type, infrequently seen, is nujalik, which refers to a hairy head (see Smith and Sleno 1986 for a possible explanation).
In summary, there exists considerable distributional (Richard et al. 1998a), movement, (Heide-Jørgensen et al. 2003), and genetic evidence (de March and Postma 2003) to indicate that the High Arctic – Baffin Bay population occupies two distinct winter habitats, one in the North Water and the other along the West Greenland coast. It is not yet clear how or if those populations also separate in their summering areas in the Canadian High Arctic. Indications are that the belugas in West Greenland have been reduced significantly in numbers in the last decade (Heide-Jǿrgensen et al. 1993, Heide-Jǿrgensen and Aquarone 2002) and that current harvest levels cannot be sustained (Innes and Stewart 2002).
The belugas found during the summer in the Cumberland Sound area have long been thought of as a distinct population (Sergeant and Brodie 1975), possibly because of their summer aggregation, in significant numbers, in the inner reaches of Cumberland Sound, at Clearwater Fiord. There the Ranger River forms an estuary, which is utilized by belugas in much the same way as many other estuaries throughout the Canadian Arctic (Brodie 1971, Sergeant and Brodie 1975, Fraker et al. 1979, Smith et al 1994). There is no other such estuarine or other centre of aggregation for belugas in the southeastern Baffin area. The Cumberland Sound belugas are genetically diverse, with a notable number of uncommon haplotypes and microsatellite loci that are not found elsewhere.
In a reconsideration of the populations for management purposes, Richard and Orr (1986) argued that the belugas of Cumberland Sound belonged to a more dispersed group of belugas frequenting the Iqaluit, Kimmirut and Cumberland Sound region. They described this population collectively as the Southeast Baffin population (Richard and Orr 1991). However, recent evidence from satellite tagging, genetics, organochlorine contaminant signatures, and traditional knowledge (Kilabuk 1998), seem to point to Cumberland Sound as a separate population. Aerial surveys (Richard and Baratin 2002) and studies of belugas tagged with satellite linked VHF transmitters (Richard 2002) have shown that the Cumberland Sound whales remain within their immediate area, possibly throughout the winter (P. Richard pers. com., DFO, Winnipeg). DNA samples obtained over the years since the late 1980s also differentiate between Cumberland Sound belugas and others sampled elsewhere in the southeast Baffin region and the High Arctic (Brown-Gladden et al. 1997, de March et al. 2002).
The delimitation of the beluga populations occupying the rest of the southeastern Baffin region is not at all clear (see below). Genetic samples collected there show that many resemble the Western Hudson Bay haplotypes. They appear to be a mixed stock dominated by Western Hudson Bay belugas.
It is possible that more than one population summers in Cumberland Sound. Genetic and organochlorine contaminant signatures, even though showing significant differences among belugas hunted in Pangnirtung, Iqaluit, and Kimmirut, are not inconsistent with this possibility (B. de March, pers. com. DFO Winnipeg). Hunters in Cumberland Sound recognize three types of belugas (Kilabuk 1998, DFO 2002a). Those hunted at the floe edge in spring are smaller, thinner and very white. Those taken during July-August, in Clearwater fiord, are larger and have a yellow colouration showing signs of epidermal moult. Those taken outside of Clearwater Fiord, on the west side of Cumberland Sound, are smaller and thinner. Their epidermis (maayak or muktuk) is thicker and stronger tasting.
Overall, recent genetic, distributional and abundance data reinforce the historical view that the belugas of Cumberland Sound form a discrete population.
A large population of belugas spends the early part of the summer in the southeastern Beaufort Sea, frequenting the Mackenzie River estuary (Fraker et al. 1979, Harwood et al. 1996, DFO 2000). Fraker (1980), without direct evidence, considered this a separate population because of its known migratory movements and the lack of a contiguous summer concentration of belugas along the Alaskan coast. More recent aerial surveys (Harwood et al. 1996) and tagging studies (Richard et al. 2001b) have strongly reinforced this view.
Genetic analyses have confirmed that Beaufort Sea belugas are clearly separated from other Canadian populations (Brown-Gladden et al. 1997) and also from the more closely adjacent Alaskan populations with which they may share a common wintering habitat (Brown-Gladden et al. 1997, O’Corry-Crowe et al. 1997, Brown-Gladden et al. 1999). This separation is primarily on the basis of the maternally inherited mitochondrial DNA haplotypes and weakly on the basis of microsatellite data. Thus these populations may interbreed in winter, but separate in their summering areas
There are three principal areas of uncertainty with the beluga population identification scheme outlined above:
- Southeast Baffin Island. The status of animals found off Southeast Baffin Island, but outside Cumberland Sound, is uncertain. Current information indicates that they are not part of the Cumberland Sound population. Inuit hunters from Iqaluit and Kimmirut, on the southeast Baffin coast, mention different body sizes and shapes in their landed catches and believe that they are harvesting from several different populations (Kilabuk 1998). These two villages are located near the assumed wintering areas of the Hudson Bay populations, and appear to harvest animals from these populations. Some of the genetic and organochlorine contaminant (de March et al. In press) sampling of Southeast Baffin belugas indicates that this might be the case. Since individual, or grouped samples, taken during spring or autumn migratory periods are expected to contain mixed populations of belugas, areas such as Southeast Baffin which are contiguous with wintering grounds shared by several populations pose a particularly difficult problem in population identification and management. Animals using these regions in the summer may share closest affinities with the Hudson Bay populations.
- Southern Hudson Bay. There are few data on the animals that summer in James Bay and the waters off the neighbouring Ontario coastline. However, the data that are available (see above) suggest that they are more closely related to the Western Hudson Bay population than the Eastern Hudson Bay population.
- High Arctic. There is increasing evidence that the Eastern High Arctic - Baffin Bay population has important substructure (see above). However, with the data currently available splitting this population is probably not justified under COSEWIC guidelines.