
Caribou (Rangifer tarandus) barren-ground population COSEWIC assessment and status report 2016: appendices
Appendix A. Place names mentioned in this report. Active mines/ports and projects in advanced exploration phase are in black font, Projects in early design to small-scale exploration phases are in grey font. Active roads and railways are depicted in complete lines. The locations of proposed road projects mentioned in the report are approximate and are not meant to represent project specifications.
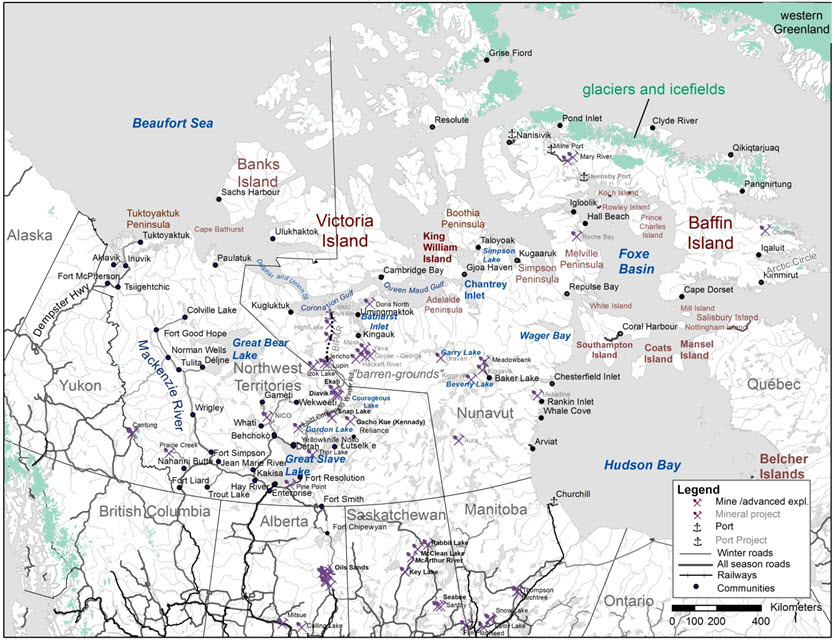
Long description for Appendix A
Map containing place names mentioned in this report.
Appendix B. Subpopulations on the Northeast Mainland.
By the mid-1990s on the northeast mainland, four additional subpopulations (Ahiak, Melville Peninsula, Lorillard and Wager Bay) were recognized based on pre-calving and calving aerial surveys and identifying the calving locations of cows fitted with VHF radio-collars (Heard et al. 1983; Heard and Stenhouse 1992; Buckland et al. 2000; Gunn et al. 2000a). Subsequently, using aerial surveys of calving distribution and satellite telemetry, Campbell (2005) combined the South Melville Peninsula into the Wager Bay subpopulation based on the movements of the satellite-collared cows.
The increase in the number of subpopulations continued when Gunn et al. (2000a) tentatively identified three subpopulations (Keith Bay, Simpson Peninsula, Boothia Peninsula) based on aerial surveys of calving areas (1974-1993) and satellite-collared cows (Gunn et al. 2000a; Gunn and Fournier 2000) (Figure A-1). Gunn and Fournier (2000) identified two other calving areas, but the aerial coverage was insufficient to define boundaries (Simpson Lake and Arrowsmith Lowlands) and whether the areas represented separate subpopulations.
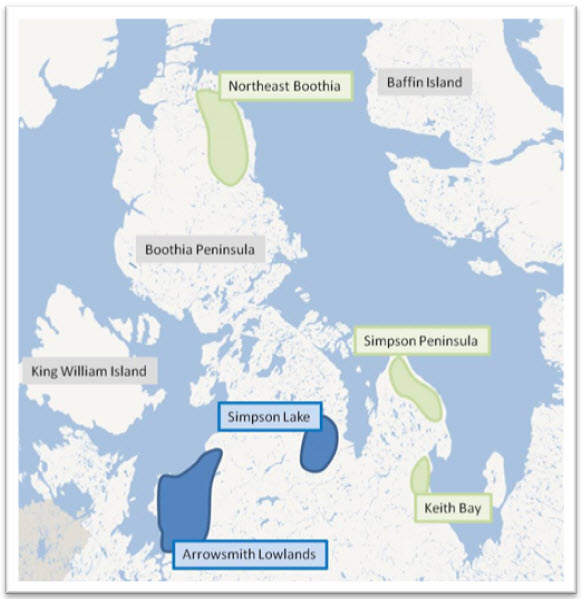
Long description for Figure A-1
Schematic showing subpopulation calving areas on the Boothia Isthmus area based on aerial surveys and limited satellite telemetry, from 1991 to 1993.
The annual movements of satellite-collared females were aggregated as statistically defined clusters which were equated as subpopulations (Nagy et al. 2011). While the cluster analysis identified similar subpopulations as based on fidelity to calving grounds for most of the previously described subpopulations, it differed for the northeast mainland. The annual distribution of satellite-collared cows was used to designate three clusters for the northeast mainland which are Lorillard, Wager Bay and Queen Maud Gulf (Nagy et al. 2011; Campbell et al. 2014; Nagy and Campbell 2012).
The Wager Bay and Queen Maud Gulf clusters each included several previously described calving areas. Including several calving areas in two clusters may be an artefact of the technique to map calving areas (kernel analysis of small numbers of satellite-collared Caribou), the effect of overlapping winter distribution on subsequent subpopulation structure, and/or it may represent a change in calving distribution (see Abundance and Trends).
The Wager Bay cluster included the previously identified ranges used by Keith Bay and Simpson Peninsula subpopulations (Nagy and Campbell 2012). However, the within-cluster overlap of the range used by individual cows was lowest for the Wager Bay cluster compared to the other clusters and while this may reflect different calving dispersion behaviour (Nagy and Campbell 2012), it may also be that low sample size limited resolution of the cluster. Nagy and Campbell (Fig. 2.6 in 2012) mapped calving of the Wager Bay cluster and found similar areas of calving previously observed in the 1990s on eastern Boothia Peninsula, Melville Peninsula, Arrowsmith Lowlands and south of Boothia Isthmus. Both the Wager Bay and Queen Maud Gulf clusters included Boothia Peninsula (Figure A-2). However, in this COSEWIC report, Boothia Peninsula is included as a separate subpopulation as it is surveyed during calving and post-calving as a recognizable geographical unit (the peninsula).
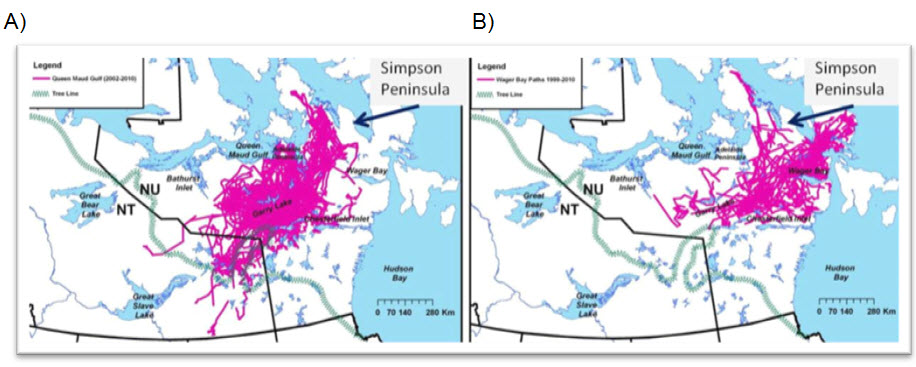
Long description for Figure A-2
Two map panels showing individual movement trajectories for the Queen Maud Gulf cluster of satellite-collared females tracked during 1999 to 2010 (left panel) and the Wager Bay cluster tracked during 2002 to 2011 (right panel).
While the Ahiak subpopulation’s calving overlapped the eastern portion of the previously described coastal calving ground (Gunn et al. 2013a), the cluster extended east of Chantrey Inlet and included the Arrowsmith Lowlands and Simpson Lake calving areas. Pathways of satellite-collared Caribou tracked in 1999-2010 (the cluster analysis did not use the earlier 1990s satellite telemetry data) did not include the Keith Bay or Simpson Peninsula areas (Figure A-2), suggesting a change in calving distribution, loss of the previously identified subpopulations or an effect of low sample size (aerial surveys and collared Caribou). The coalescence of the calving areas may also be an artefact of the technique to map calving areas (kernel analysis of small numbers of satellite-collared Caribou), or it may represent a change in calving distribution.
Table A-1 summarizes survey information for northeast mainland calving areas.
| Subpopulation | Calving area | Aerial surveys calving | Incidental aerial sightings calving | Satellite collars (n, mean ± SE) |
|---|---|---|---|---|
| Ahiak cluster (Campbell et al. 2012) | Tuktoyaktuk Peninsula, | 2007, 2008, 2009 | blank | 2006-2012 (12 ± 2.05) |
| Ahiak cluster (Campbell et al. 2012) | Arrowsmith Lowlands, | 1975, 1985, 1986, 2010, 2011 | 1989,91 | blank |
| Ahiak cluster (Campbell et al. 2012) | Keith Bay, | 2010; 2011 | 1991 | 1991-92 (1) |
| Ahiak cluster (Campbell et al. 2012) | Boothia Peninsula East, | 1975, 1985, 2006, 2010; 2011 | blank | 1991-93 (3) |
| Ahiak cluster (Campbell et al. 2012) | Simpson Lake, | 1975, 1985 1989, 2010; 2011 | blank | 1991-93 (1) |
| Ahiak cluster (Campbell et al. 2012) | Simpson Peninsula | 1989; 2010; 2011 | blank | 1991-93 (1) |
| Wager Bay cluster (Nagy et al. 2011) | Wager Bay | 1974, 1976; 1977, 2000, 2002, 2004 | 1986, 1987,1988 | 2000-06 (5.5 ± 1.28) |
| Wager Bay cluster (Nagy et al. 2011) | Lorillard | 1976, 1977; 1999, 2001, 2003 | 1986, 1987,1988 | 2000-2006 (8.6 ± 1.23) |
| Wager Bay cluster (Nagy et al. 2011) | South Melville Peninsula | 1974, 1976 | blank | 1991 (1) 2000-2006(5.5 ± 1.28) |
| Subpopulation | Year | Month | Estimate | Precision | Precision type | Survey design a | Estimate type | Age class. | Report type | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Porcupine | 1987 | July | 165,000 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Incl. calves | Summary | Caikoski 2009 |
| Porcupine | 1989 | July | 178,000 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Incl. calves | Summary | Caikoski 2009 |
| Porcupine | 1992 | July | 160,000 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Incl. calves | Summary | Caikoski 2009 |
| Porcupine | 1994 | July | 152,000 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Incl. calves | Summary | Caikoski 2009 |
| Porcupine | 1998 | July | 129,000 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Incl. calves | Summary | Caikoski 2009 |
| Porcupine | 2001 | July | 123,000 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Incl. calves | Summary | Caikoski 2009 |
| Porcupine | 2010 | July | 169,000 | 153,493–184,403 | 95% CI | Post-calving photo | Rivest | Incl. calves | Detailed | Caikoski 2013 |
| Porcupine | 2013 | July | 197,228 | 13,772; 168,667–225,789 | SE; 95% CI | Post-calving photo | Rivest | Incl. calves | Detailed | Caikoski 2015 |
| Tuktoyaktuk | 2005 | Sept | 2,700b | blank | blank | Strip transect | Extrapolation from density & sex age composition | Incl. calves | Summary | Branigan 2005 |
| Tuktoyaktuk | 2006 | July | 3,078 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Nagy and Johnson 2006 |
| Tuktoyaktuk | 2009 | July | 2,753 | 276 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Davison et al. 2014 |
| Tuktoyaktuk | 2012 | July | 2,192 | 178 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | None | Davison 2016 |
| Tuktoyaktuk | 2015 | July | 1,701 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Non-calf | None | Davison 2016 |
| blank | blank | blank | blank | blank | blank | blank | blank | Non-calf | blank | blank |
| Cape Bathurst | 1986 | July | 13,476c | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Nagy 2009a |
| Cape Bathurst | 1987 | July | 12,516c | 3,504 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Cape Bathurst | 1992 | July | 19,278 | 5,397 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Cape Bathurst | 2000 | July | 11,089 | 1,756 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Cape Bathurst | 2005 | July | 2,434 | 257 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Cape Bathurst | 2006 | July | 1,821 | 149 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Nagy and Johnson 2006 |
| Cape Bathurst | 2009 | July | 1,934 | 350 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Davison et al. 2014 |
| Cape Bathurst | 2012 | July | 2,427 | blank | blank | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Cape Bathurst | 2015 | July | 2,259 | 84 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2016 |
| blank | blank | blank | blank | blank | blank | blank | blank | Non-calf | blank | blank |
| Bluenose-West | 1986 | July | 88,369[4] | 6,899 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Nagy 2009a |
| Bluenose-West | 1987 | July | 106,887 | 4,655 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Bluenose-West | 1992 | July | 112,360 | 25,566 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Bluenose-West | 2000 | July | 76,376 | 14,347 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Bluenose-West | 2005 | July | 20,800 | 2,040 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Bluenose-West | 2006 | July | 18,050 | 527 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Nagy and Johnson 2006 |
| Bluenose-West | 2009 | July | 17,897 | 1,310 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Davison et al. 2014 |
| Bluenose-West | 2012 | July | 20,465 | 3,490 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Bluenose-West | 2015 | July | 15,268 | 1,369 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2016 |
| Bluenose-East | 2000 | July | 104,000 | 84 412 - 126 100 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Patterson et al. 2004 |
| Bluenose-East | 2005 | July | 70,081 | 8,120 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Summary | Davison 2015 |
| Bluenose-East | 2006 | July | 65,119 | 3,504 | 95% CI | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Nagy and Tracz 2006 |
| Bluenose-East | 2010 | July | 98,481 | 7,125 | SE | Post-calving photo | Lincoln-Peterson estimate | Non-calf | Detailed | Adamczewski et al. 2013 |
| Bluenose-East | 2010 | June | 114,472 | 6,908 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Adamczewski et al. 2013 |
| Bluenose-East | 2013 | June | 68,295 | 7,610 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Boulanger et al. 2014 |
| Bluenose-East | 2015 | June | 38,592 | 33,859 - 43,325 | 95% CI | Calving photo/ double counting | Extrapolation from density & sex age composition | Non-calf | Detailed | Boulanger 2015 |
| Bathurst | 1986 | June | 472,000 | 72,900 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Heard and Williams 1991 |
| Bathurst | 1990 | June | 351,683 | 77,800 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Heard and Williams 1991 |
| Bathurst | 1996 | June | 349,046 | 94,900 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Gunn et al. 1997 |
| Bathurst | 2003 | June | 186 005 | 40 146 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Gunn et al. 2005 |
| Bathurst | 2006 | June | 128,047 | 27,300 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Nishi et al. 2007 |
| Bathurst | 2009 | June | 31,982 | 5,306 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Nishi et al. 2014 |
| Bathurst | 2012 | June | 34,690 | 4,691 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Boulanger et al. 2014 |
| Bathurst | 2015 | June | 19,769 | 12,349-27,189 | 95% CI | Calving photo/ double counting | Extrapolation from density & sex age composition | Non-calf | Detailed | Boulanger et al. 2015 |
| Ahiak (< 2011) | 1986 | June | 30,000 | blank | blank | Systematic strip survey | Extrapolation from density | Non-calf | Detailed | Gunn et al. 2000 |
| Ahiak (< 2011) | 1996 | June | 200,000 | blank | blank | Systematic strip survey | Extrapolation from density | Non-calf | Detailed | Gunn et al. 2000 |
| Ahiak (< 2011) | 2007 | June | 235,000 | blank | blank | Systematic strip survey | Extrapolation from density | Non-calf | Summary | Johnson et al. 2008 |
| Beverly (< 2011) | 1984 | June | 263,691 | 80,652 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Summary | Heard and Jackson 1990 |
| Beverly (< 2011) | 1988 | June | 189,561 | 70,961 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Heard and Jackson 1990 |
| Beverly (< 2011) | 1994 | June | 276,000 | 106,600 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Williams 1995 |
| Beverly (< 2011) | 2002 | June | Too low to estimate | blank | blank | Systematic strip survey | blank | Non-calf | Detailed | Johnson and Mulders 2009 |
| Beverly (< 2011) | 2007 | June | Too low to estimate | blank | blank | Systematic strip survey | blank | Non-calf | Detailed | Johnson et al. 2009 |
| Qamanirjuaq | 1985 | June | 272,032 | 142,199 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Summary | Heard and Jackson 1990 |
| Qamanirjuaq | 1988 | June | 220,999 | 72,459 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Heard and Jackson 1990 |
| Qamanirjuaq | 1994 | June | 495,665 | 105,426 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Summary | Campbell et al. 2010 |
| Qamanirjuaq | 2008 | June | 348,661 | 44,861 | SE | Calving photo | Extrapolation from density & sex age composition | Non-calf | Detailed | Campbell et al. 2010 |
| Qamanirjuaq | 2014 | June | 264,718 | 44,084 | 95% CI | Calving photo/ double counting | Extrapolation from density & sex age composition | Non-calf | Detailed | Campbell et al. 2015 |
| Southampton | 1987 | June | 4,033 | 659 | SE | Stratified random bloc (helicopter) | Extrapolation from density | Non-calf | Summary | Heard and Ouellet 1994 |
| Southampton | 1990 | March | 9,319 | 3,325 | SE | Stratified transects | Extrapolation from density | Incl. calves | Summary | Heard and Ouellet 1994 |
| Southampton | 1991 | June | 13,676 | 1,584 | SE | Stratified random bloc | Extrapolation from density | Non-calf | Summary | Ouellet et al. 1996 |
| Southampton | 1995 | June | 18,275 | 1,390 | SE | blank | blank | blank | Summary | Campbell 2006 |
| Southampton | 1997 | June | 30,381 | 3,982 | SE | Stratified random transect | Extrapolation from density | Non-calf | Detailed | Campbell 2006 |
| Southampton | 2003 | June | 17,981 | 2,127 CI=3,982 | SE | Stratified random transect | Extrapolation from density | Non-calf | Detailed | Campbell 2006 |
| Southampton | 2005 | June | 20,582 | 3, 065 CI=3,065 | SE | Stratified random transect | Extrapolation from density | Non-calf | Detailed | Campbell 2006 |
| Southampton | 2007 | June | 15,452 | 1,858 CI=1,858 | SE | Stratified random transect | Extrapolation from density | Non-calf | Detailed | Campbell and Boulanger 2016 |
| Southampton | 2009 | June | 13,956 | 1,790 | 95% CI | Stratified random transect | Extrapolation from density | Non-calf | Detailed | Campbell and Boulanger 2016 |
| Southampton | 2011 | June | 7,903 | 1,261 | 95% CI | Stratified random transect/ double counting | Extrapolation from density | Non-calf | Detailed | Campbell and Boulanger 2016 |
| Southampton | 2013 | May | 7,287 | 1,045 | 95% CI | Stratified random transect/ double counting | Extrapolation from density | Non-calf | Detailed | Campbell and Boulanger 2016 |
| Southampton | 2015 | May | 12,297 | 1,844 | 95% CI | Stratified random transect/ double counting | Extrapolation from density | Non-calf | Detailed | Campbell and Boulanger 2016 |
a Fixed-wing unless otherwise specified.
b Total number of Rangifer in the area estimated at 3,890; 30% assumed to be reindeer, as assessed in independent helicopter survey.
c Recalculated retroactively by Nagy (2009a)