
COSEWIC Assessment and Status Report on the Eastern Wolf Canis sp. cf. lycaon in Canada - 2015

Photo: © Michael Runtz, 2015
- Document Information
- Assessment Summary
- Executive Summary
- Technical Summary
- Preface
- Wildlife Species Description and Significance
- Distribution
- Habitat
- Biology
- Population Sizes and Trends
- Threats and Limiting Factors
- Protection, Status and Ranks
- Acknowledgements and Authorities Contacted
- Information Sources
- Biographical Summary of Report Writers
- Collections Examined
- Table 1. Sampling effort and number of Eastern Wolves found. Eastern Wolf occurrence is based on an assignment of Q≥0.8 in the program Structure where individuals were genotyped at 12 autosomal microsatellites. Source data are from literature noted below.
- Table 2. Abundance estimates of Eastern Wolves. Maximum # in Ontario (ON) sites is based on a density estimate of 3 wolves per 100 km2 documented in Algonquin Park (Rutledge et al. 2010e), estimated # is based on 69% of the individuals with Q≥0.8 in Structure (Rutledge et al. 2010c), and # of mature individuals is based on 45.9% of the population being adults (Pimlott et al. 1969). Sites in Québec (QC) use a density of 2.6 wolves per 100 km2 (Papineau-Labelle) or 1.9 per 100 km2 (remaining sites). The closest abundance estimate of Eastern Wolf is 236 mature individuals.
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Cananda
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2015. COSEWIC assessment and status report on the Eastern Wolf Canis sp. cf. lycaon in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xii + 67 pp. (Species at Risk Public Registry website).
COSEWIC 2001. COSEWIC assessment and update status report on the eastern wolf Canis lupus lycaon in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 19 pp.
Samson, C. 2001. Update COSEWIC status report on the eastern wolf Canis lupus lycaon in Canada, in COSEWIC assessment and update status report on the eastern wolf Canis lupus lycaon in Canada.
Van Zyll de Jong, C.G., and L.N. Carbyn. 1999. COSEWIC status report on the Grey Wolf Canis lupus in Canada. Committee on the Status of Endangered Wildlife in Canada. 61 pp.
COSEWIC would like to acknowledge Linda Rutledge for writing the status report on the Eastern Wolf (Canis sp. cf. lycaon), prepared under contract with Environment Canada. Modifications to the status report after acceptance of the provisional report were overseen by Graham Forbes, Co-chair of the COSEWIC Terrestrial Mammals Specialist Subcommittee (TM SSC), based on comments from jurisdictions, the TM SSC, COSEWIC members, and external experts.
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
E-mail: COSEWIC E-mail
Website: COSEWIC
Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur le Loup de l'Est (Canis sp. cf. lycaon) au Canada.
Eastern Wolf -- Photograph of Eastern Wolf from Algonquin Provincial Park. © Michael Runtz, used with permission.
The Eastern Wolf (putatively Canis lycaon, formerly Canis lupus lycaon) is an intermediate-sized canid weighing an average 24 kg for females and 29 kg for males. Pelage often is described as reddish-brown/tawny, but is highly variable. The Eastern Wolf is best defined by a combination of genetic distinctiveness, morphological characters, and an ecological role associated with a feeding preference for smaller prey than fed on by Gray Wolf (C. lupus). The Eastern Wolf population has a degree of hybridization with Coyote (C. latrans), and individuals are defined based on having a high level of genetic 'purity,' that is, distinctiveness from both Gray Wolf and Coyote as determined by molecular genetic analysis. It is important to note that the Eastern Wolf discussed in this report is not the same Eastern Wolf discussed in the Great Lakes region because those Canis are considered in this report as Great Lakes-Boreal Wolves, a hybrid between the Eastern Wolf and Gray Wolf. Although evidence is strong that the Eastern Wolf is a valid species, the taxonomy of Eastern Wolf is under debate; in this report the Eastern Wolf is considered to be Canis sp. cf. lycaon, a wildlife species as defined under SARA that is worthy of conservation because of its distinctiveness, persistence, and significance as a large carnivore, and likely part of the last remnant population of the large Canis from eastern North America. Aboriginal traditional knowledge also supports the existence of a medium-sized Canis in the region.
The current distribution of Eastern Wolves is thought to be restricted to the mixed coniferous-deciduous forests of central Ontario and southwestern Québec, namely the Great Lakes-St. Lawrence Forest Region. Eastern Wolves were extirpated from most of their original range in North America due to eradication of large Canis over much of the past 400 years. Genetic analyses suggest that the current distribution of Eastern Wolves mainly is in central Ontario and southern Québec (north of the St. Lawrence River), with concentrations in core areas, all of which are protected areas.
Eastern Wolves typically occur in deciduous and mixed forest landscapes with low human density, south of the Boreal Forest Region. Sandy soils are often preferred for den sites. Both den and rendezvous sites tend to be located in conifer/hardwood-dominated landscapes near a permanent water source. Territory size is often near 200 km2.
Eastern Wolves live in family-based packs composed of a breeding pair and offspring from the current and previous years. Females give birth to an average of five pups in late April - early May and they remain at the den site for 6 - 8 weeks. Dispersing juveniles leave the pack after 37 weeks. Eastern Wolves are primarily predators of White-tailed Deer (Odocoileus virginianus). Predator-prey and diet analyses indicate that Eastern Wolves can be effective predators of Moose (Alces americanus), although efficiency varies by pack, season, and year. Beaver (Castor canadensis) also constitutes a substantial portion of Eastern Wolf diet.
There have been 170 - 195 Eastern Wolf (all ages) identified in the last 10-15 years. The population size is unknown but likely less than 1000 mature individuals. The estimated minimum population size is 236 mature individuals, mainly located within protected areas. A best-possible-scenario maximum estimate of 1203 mature individuals within the extent of occurrence is based on there being an equally high density of Eastern Wolf outside protected areas. Most records though occur in protected areas and the population size of mature Eastern Wolf likely is closer to 236 individuals. There is no population trend information except for Algonquin Park, the site with the most Eastern Wolf records to date, which appears to be stable.
The main threat and limiting factor for Eastern Wolves outside the protected areas likely is human-caused mortality from hunting and trapping, which is facilitated by road networks. Based on research in Algonquin Park, excessive mortality likely limits dispersal, and alters pack breeding dynamics, leading to another main threat, gene introgression (hybridization) with Eastern Coyotes due to the lack of conspecific mates. Habitat loss and fragmentation associated with road networks and urbanization is expected to continue outside protected areas and likely will deter population expansion. Negative public attitudes towards wolves, and established packs of Eastern Coyote, may limit population expansion.
The Eastern Wolf is listed as Special Concern under the federal Species at Risk Act (SARA) and Ontario's Endangered Species Act, 2007. Both listings are as a subspecies of Gray Wolf (Canis lupus lycaon). No wolf species is listed under the Lois sur les espèces menacées ou vulnérables [Act respecting Threatened or Vulnerable Species in Québec]. Hunting and trapping of wolves is permitted in wildlife reserves, but not in national (federal or provincial) parks. In Ontario, wolves are protected from regulated hunting and trapping in Algonquin Park, in the townships surrounding Algonquin Park, and in all provincial Crown Game Preserves. Eastern Wolves are protected from hunting, but not from trapping, in French River Park. Wolves are protected from harvest in national parks. Aboriginal communities retain constitutional rights to harvest Wolves for sustenance and ceremonial purposes, including in protected areas. A small game licence is required to hunt Wolves in Ontario (limit of 2 per year) and Québec (no bag limit). NatureServe ranks Canis lupus lycaon as N4 (apparently secure). At the provincial scale, Eastern Wolf is ranked as S4 ('apparently secure') in Ontario, and is not ranked in Québec.
| Question | Information |
|---|---|
| Generation time Age of first breeding is > 2 years and adult lifespan highly variable depending on mortality rates. |
3.5 yrs |
| Is there an inferred continuing decline in number of mature individuals? The subpopulation in Algonquin Park appears stable but subpopulations elsewhere are not well surveyed; subpopulations in the larger area are small and relatively isolated and likely experience threats such as habitat alteration, road building, hunting/trapping, hybridization with Eastern Coyotes that decrease the population. |
Possibly |
| Estimated percent of continuing decline in total number of mature individuals within 2 generations (7 years). (see above) |
Unknown |
| Suspected percent reduction in total number of mature individuals over the last 3 generations (10.5 yrs). Unknown for most of range but in Algonquin Park area, the number of mature animals decreased 20% from 2002 - 2007 due to disease, but have since increased. |
Unknown |
| Suspected percent reduction in total number of mature individuals over the next 3 generations (10.5 yrs). (see above) |
Unknown |
| Observed/suspected percent reduction in total number of mature individuals over any 3 generation period (10.5 yrs), over a time period including both the past (observed) and the future (suspected). No evidence of recent decline. Historical decrease of approximately 50% of its Canadian range. Human-caused mortality between 1964 - 1971 (< 16% decline), and 1991 - 1999 in the Algonquin subpopulation (< 12% decline) (see Fluctuations and Trends). |
Unknown |
| Are the causes of the decline clearly reversible and understood and ceased? The cause of the historical decline was targeted eradication, which has ceased. Harvest outside protected zones has not ceased and continues to be a significant threat. Harvest could be a cause of potential future decline via gene introgression by Eastern Coyotes. |
No |
| Are there extreme fluctuations in number of mature individuals? | No |
| Question | Information |
|---|---|
| Estimated extent of occurrence Extent of occurrence (EOO) based on all records |
126,573 km2 |
| Index of area of occupancy (IAO) The area of occupancy, based on size of sites containing Eastern Wolf records, is 29,472 km2. |
10,000 km2 |
| Is the total population severely fragmented? The total population is fragmented by regions where there is no protection, high road networks, higher human density, and increased risk of hybridization with Eastern Coyotes. |
Yes |
| Number of locations (Note: See Definitions and Abbreviations on COSEWIC website and IUCN 2010 (not in information sources) for more information on this term.) Population exists mainly in 11 sites (plus numerous townships around Algonquin Park), and animals exist in numerous packs. If causes of mortality are independent among packs, then locations would be > 20. |
Unknown, likely >20 |
| Is there an inferred continuing decline in extent of occurrence? Canis mortality outside protected regions likely limits expansion of the range but the EOO is based on recent genetic tests and declines have not been noted over this short time period. |
Unknown |
| Is there an inferred continuing decline in index of area of occupancy? (see above) |
Unknown |
| Is there an inferred continuing decline in number of populations? (see above) |
Unknown |
| Is there an inferred continuing decline in number of locations? Wolf harvest outside protected regions may impact the locations. |
Unknown |
| Is there an inferred continuing decline in area, extent and/or quality of habitat? Hunting and trapping limit ability to establish relatively non-hybridized packs in other locations. Land conversion and road building in potential dispersal areas decreases habitat quality. |
Yes |
| Are there extreme fluctuations in number of populations? | Unknown, but unlikely |
| Are there extreme fluctuations in number of locations? | No |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | No |
| Population | N Mature Individuals |
|---|---|
| Ontario-Québec Estimated minimum number of mature individuals is 236, based on extrapolating results from several study areas to size of protected areas where Eastern Wolf have been recorded. A maximum estimate of 1203 mature individuals may occur within the extent of occurrence but only if it is assumed that areas outside protected areas have the same density of Eastern Wolf, which is very unlikely because few animals have been recorded outside protected areas. The population of mature individuals likely is closer to 236. |
- |
| Total | Unknown, but very likely < 1000 |
| Question | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. Several analyses were done > 10 years ago but only for the Algonquin Park area; methods varied, genetic issues were not included, and overall, their application is limited. One population viability analysis concluded cause for concern, while another concluded that the population was secure. |
Not available |
Hunting and Trapping, Gene introgression from Eastern Coyote, Road Networks facilitate mortality levels. Access to most of range limited by competition from established populations of Eastern Coyote.
| Question | Information |
|---|---|
| Status of outside population(s)? | No populations outside Canada are known. The Red Wolf is an unviable candidate for a translocation rescue effect in Canada because its genome has been impacted by founder effects and extensive hybridization. Natural colonization by Red Wolves is unlikely due to the high human density throughout the ≈1000 km geographic separation of the two populations. |
| Is immigration known or possible? | No |
| Would immigrants be adapted to survive in Canada? There is no immigrant source. |
N/A |
| Is there sufficient habitat for immigrants in Canada? | N/A |
| Is rescue from outside populations likely? | No |
In 1999, the Eastern Grey Wolf (Canis lupus lycaon) was considered a subspecies of the Grey Wolf and was placed in the Data Deficient category. Status was re-examined (as Eastern Wolf, Canis lupus lycaon) and designated Special Concern in May 2001. New genetic analyses indicate that the Eastern Wolf is not a subspecies of Grey Wolf. In May 2015, a new wildlife species, Eastern Wolf (Canis sp. cf. lycaon) was designated Threatened.
Throughout this report, the term "Eastern Wolf" will be used to identify the intermediate-sized canid that occurs in central Ontario and southwestern Québec. As per the 3-species hypothesis, "Eastern Coyote" will be used for the smaller Coyote - Eastern Wolf Canis hybrid (also known as Tweed Wolf, Brush Wolf, or Coywolf) that occurs from Ontario to Newfoundland, whereas "Great Lakes-Boreal Wolf" will be used for the putative Grey Wolf - Eastern Wolf Canis hybrid that occurs in more northern boreal forest landscapes of Manitoba, Ontario, and Québec. The term "Grey Wolf" will be used to refer to the larger non-hybridized wolf found in northwestern regions of Canada and "Coyote" (C. latrans) will be used to refer to the smaller non-hybridized Coyote from western North America. Only the status of Eastern Wolf is addressed in this COSEWIC report.
As part of COSEWIC status assessments, Aboriginal traditional knowledge (ATK) reports are prepared by the Aboriginal Traditional Knowledge Subcommittee (ATK SC). These initial reports compile and summarize ATK relevant to status assessment when ATK information is available and readily accessible. A Gathering Report may be undertaken if there are significant knowledge gaps, or if major contradictions exist between ATK and other forms of knowledge. The initial process used by the ATK SC did not identify ATK specific to the Eastern Wolf in Canada, but information was provided to the ATK SC after the 2-month review period, and that information is included in the report.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
The taxonomy of the Eastern Wolf is under debate and this report begins with a discussion of its eligibility for assessment by COSEWIC. COSEWIC criteria for species eligibility are based on: taxonomic validity (defined by peer-reviewed publications and communication with specialists); being native to Canada (naturally occurring or present for > 50 years); and occurring regularly in Canada (COSEWIC 2014).
At present, COSEWIC lists the Eastern Wolf as a subspecies of Gray Wolf (Canis lupus lycaon) (Samson 2001). However, based on genetic analyses in the last 10 years, there is now widespread agreement that the Eastern Wolf is not a subspecies of the Gray Wolf (see Appendix A). The taxonomic validity of Eastern Wolf is outlined below, and discussed in detail in Appendix A.
The large Canis east of the Great Lakes region has consistently been recognized as unique, and has been classified as a species, or subspecies for over 200 years. The type specimen for Eastern Wolf was collected (likely) near Québec City, Québec in 1761, and named Canis lycaon by Schreber in 1775 (Miller 1912; Manning and Sturtevant 1966). In the first major revision of Canis, Pocock (1935) used differences in morphology to retain the Eastern Wolf (re-labelled as Eastern Canadian Wolf) as a full species, and defined all remaining Canis in central and north North America as subspecies of Gray Wolf. Goldman (1937) labelled the Eastern Wolf as Canis lupus lycaon - Algonquin type. The inclusion of C. lycaon as a subspecies of C. lupus was due to a similarity in skull measurements to other Canis. Since then, the Eastern Wolf has been consistently recognized in all morphometric-based analyses as the smallest sized of all C. lupus subspecies (Young and Goldman 1944; Nowak 1995).
Aboriginal traditional knowledge from the Mohawk First Nation of Akwesasne notes that more than one type of Canid was recognized in the region before European contact, based on differences in body size, temperament, and size of prey consumed (Lickers pers. comm. 2015). The 'little wolf' was considered to be 'nice' (as compared to the more feared [Gray] Wolf) and are known to prey on White-tailed Deer. A smaller canid (Coyote) arrived later in the region (Lickers pers. comm. 2015).
Kyle et al. (2006), Chambers et al. (2012), and Mech et al. (2014) provide extensive reviews on the taxonomic history of the Eastern Wolf, and Mech (2011) documents the non-genetic evidence for the existence of the Eastern Wolf. Other reviews are found in Cronin and Mech (2009), Schwartz and Vucetich (2009), Mech (2010a), Benson et al. (2012), and Rutledge et al. (2012). Much of the debate on the taxonomy of Canis is associated with the arrival of Coyote (Canis latrans) into eastern North America. In a continental-scale invasion, Coyotes from the Prairie region of North America expanded northward and eastward; the first record in southeastern Ontario was in 1919 (Nowak 1979). These small Canis (e.g., adult male averages of 13 - 14 kg in different parts of the central Prairies [Parker 1995]) bred with a larger Canis in the Great Lakes region and produced an intermediate-sized animal (e.g., adult male averages of 14.6 - 21 kg in different parts of northeastern North America [Parker 1995; Villemure and Jolicoeur 2004]). The new animal, named the Eastern Coyote, then established itself across eastern Canada, reaching Québec in 1944, Nova Scotia in the 1970s, and Newfoundland in 1985 (Parker 1995; Naughton 2012).
In the other hypothesis (the 3-species model), the large Canis that bred with the 'western' Coyote was a separate species named Eastern Wolf (or Red Wolf) that existed mainly in eastern North America. The third species, Gray Wolf, immigrated from Eurasia 300,000 years ago (Wilson et al. 2003). In this hypothesis, the Eastern Wolf is a true species and the Eastern Coyote is a hybrid that resulted from mating between Coyote and Eastern Wolf, not between Coyote and Gray Wolf (Rutledge et al. 2012; Rutledge et al. in revision). Another large Canis, the Great Lakes-Boreal Wolf (see Search Effort) is a hybrid that resulted from Eastern Wolf breeding with Gray Wolf (e.g. Wheeldon and White 2009; Wheeldon et al. 2010; Fain et al. 2010; Rutledge et al. 2012, in review).
There have been publications (see Appendix A) where the Great Lakes-Boreal Wolf has been called Eastern Wolf (Canis lycaon) but, in this report, the Eastern Wolf refers to what is believed to be the most 'pure' and remnant population of the progenitor of the Great Lakes-Boreal Wolf and Eastern Coyote hybrids.
Notwithstanding the taxonomic debate, there is consensus that a unique Canis has been recognized to exist in the region since the mid-1700s and currently exists in parts of Ontario and Québec (Appendix A). The animal exploits a different ecological niche than Canis lupus-like canids and C. latrans-like canids (see Interspecific Interaction). It is largely found in minimally human-impacted mixed-woods plains region of Canada. Its contemporary distribution is influenced by genetic introgression from hybridization with Eastern Coyote in more anthropogenically altered regions in the south of Ontario and Québec, and hybridization with C. lupus in more boreal-dominated regions containing more Moose (Appendix A). Hence, there is a unique canid that exploits an ecological niche in, or proximate to, a few small protected regions of Ontario and Québec; protecting the largest canid within this narrowing/threatened ecological niche is a conservation concern.
A wildlife species is defined in the Species at Risk Act (SARA) and COSEWIC (2014) as a species, subspecies, variety or geographically or genetically distinct population that is native to Canada and wild by nature. The Eastern Wolf is not a subspecies of Gray Wolf (see Taxonomic Validity) and therefore could not be a Designatable Unit of Gray Wolf. In summary, this report considers the Eastern Wolf as a unique, persistent, and evolutionarily significant wildlife species in Canada. The remaining Eastern Wolves are naturally occurring residents in Canada that have been present for > 50 years. A plausible mechanism exists for the maintenance of the Eastern Wolf in regional sympatry with the Eastern Coyote (see Appendix A). The proposed name for this report is Eastern Wolf (Canis sp. cf. lycaon) (Appendix E4 in COSEWIC 2014).
The high plasticity in morphological features, especially in body size ranges (Rutledge et al. 2010c), makes the Eastern Wolf difficult to distinguish from Great Lakes-Boreal Wolves and Eastern Coyotes without assignment tests based on genetic markers. However, Eastern Wolves (Figure 1) have been differentiated from both Great Lakes-Boreal Wolves and Eastern Coyotes in skull morphology, body size, habitat, prey base, and genetic signature. Both skull morphology and body size of Eastern Wolves are intermediate to Great Lakes-Boreal Wolves and Eastern Coyotes (Kolenosky and Standfield 1975; Schmitz and Kolenosky 1985; Sears et al. 2003; Theberge and Theberge 2004;Rutledge et al. 2010c, 2010d; Benson et al. 2012). Long before genetics was used to identify species, Kolenosky and Standfield (1975) categorized wolves in Algonquin Provincial Park (hereafter. 'Algonquin Park') separately from surrounding Canis types based on body size and skull morphology. Their distribution map of Canis in Ontario has since been supported by the patterns identified with molecular genetic techniques (Rutledge 2010a; Figure 2).
Photo: © Michael Runtz, 2015
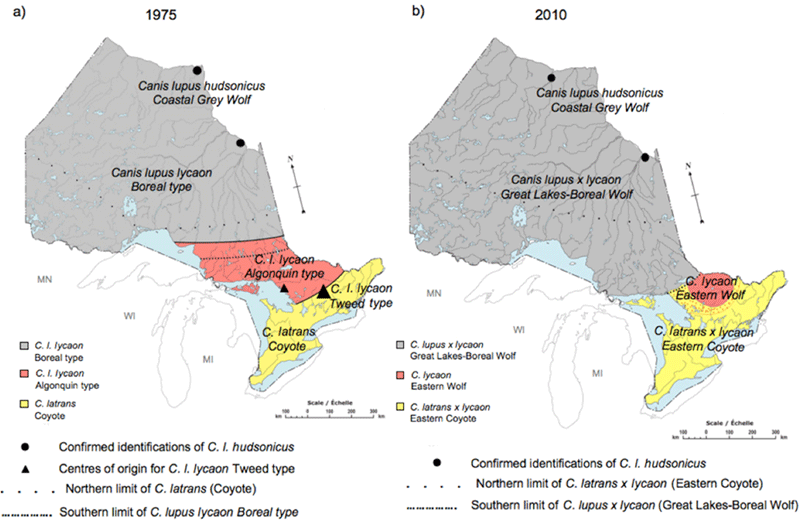
Long description for Figure 2
Two maps illustrating the distributions of types of Canis species in Ontario. Map A shows the distributions of proposed by Kolenosky and Standfield (1975) based on skull morphology and body mass. Map B shows the distributions proposed by Rutledge (2010). For details, refer to Figure 2 caption and report text.
Eastern Wolves are typically < 30 kg (Theberge and Theberge 2004). Based on data collected in Algonquin Park from 2002 - 2007, female average yearling weight is 18.1 kg and female average adult weight is 24.2 kg, whereas male average yearling weight is 23.5 kg and average adult weight is 29.3 kg. Jolicoeur and Hénault (2010) reported the mean weight of Canis found in Réserve faunique La Papineau-Labelle (hereafter, 'Papineau-Labelle Reserve') as 24.6 kg (±0.7 SE) for males and 22.2 kg (±0.9 SE) for females. In Parc national de la Mauricie (hereafter, 'La Mauricie Park'), adult female Canis have an average weight of about 28.7 kg, with shoulder height of 71.8 cm and an average male adult weight of approximately 44.5 kg, with shoulder height of 81.2 cm (Villemure and Festa-Bianchet 2002). Although sample sizes were low (3 females, 2 males), these sizes are larger than those estimated for Eastern Wolves and likely include Great Lakes-Boreal Wolves in the sample. Average adult shoulder height for Eastern Wolf from the Algonquin Park region is 63.8 cm for females and 70.0 cm for males (Patterson 2011b, pers. comm.). Eastern Wolves are longer (from tip of nose to base of tail) than Eastern Coyotes, with average Eastern Wolf female length of 109.3 cm (± 1.3 SE) and average male length of 113.0 (± 1.8 SE) (Benson et al. 2012). Body size is important in predators because morphological characteristics affect energetic requirements and influence the size of prey that wolves can effectively hunt (Carbone et al. 1999;MacNulty et al. 2009).
Coat colour is not a reliable descriptor of the different Canis species because of the variability in pelage among Canis. However, Eastern Wolves (Figure 1) typically have coats with more reddish-brown/tawny colouration and reddish forelegs, when compared to Gray Wolves and Great Lakes-Boreal Wolves.
Population (genetic) structure of Canis in the region varies due to the introgression of Canis latrans, and C. lupus genes into the remaining Eastern Wolf population. The Canid types are separated based on use of Q values, which represent the probability of belonging to a cluster, as identified in the genetic analysis program STRUCTURE 2.3 (Hubisz et al. 2009; Appendix B).
The amount of variation within the Eastern Wolf population is unknown.
Existing Eastern Wolves likely represent a relict population that once inhabited the eastern temperate forests of North America, including the southern regions of Ontario and Québec, prior to the arrival of European settlers (Rutledge et al. 2010d; Kyle et al. 2006; Figure 3). Clearing of land, invasion of non-endemic Coyotes, and targeted persecution of large canids may have pushed Eastern Wolves north into Algonquin Park and south-central Quebec, which probably represents a refugium at the northern limit of their historical distribution.
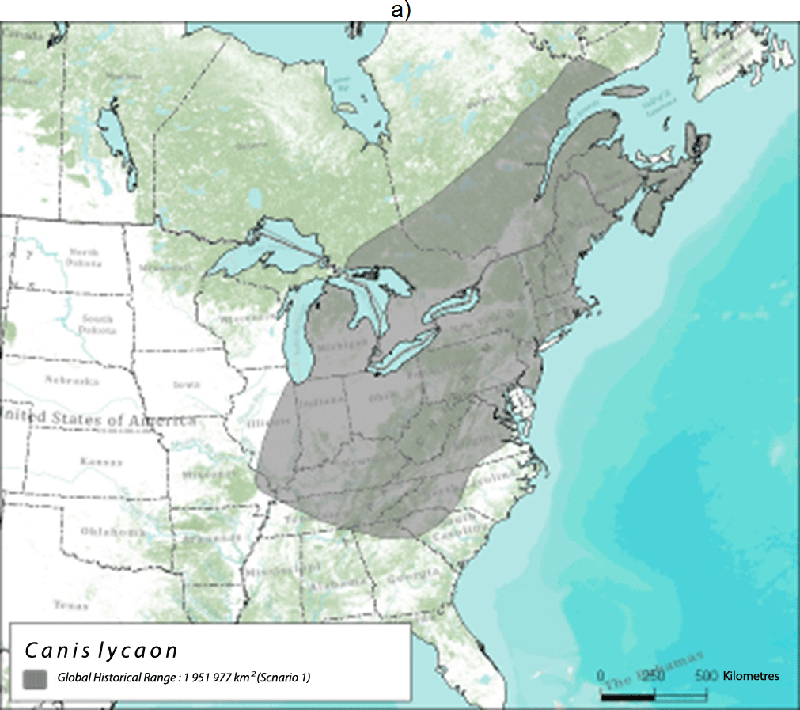
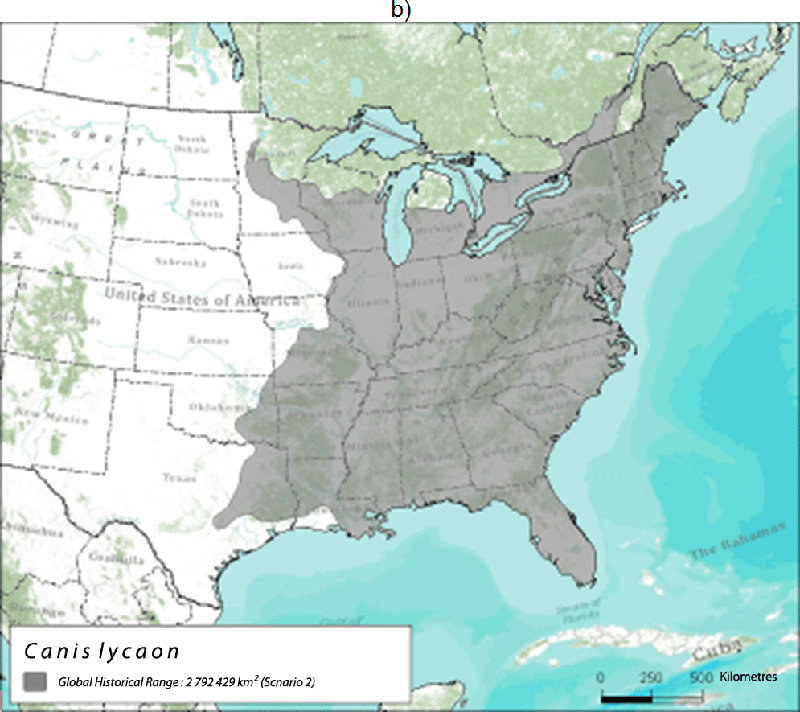
Long description for Figure 3
Two maps illustrating the global historical range of the Eastern Wolf. Map A illustrates the range based Nowak (1995) and Leonard et al. (2005). Map B illustrates the range based on Wilson et al. (2000) and Rutledge et al. (2010d). For discussion, refer to report text.
The current socio-economic importance of Eastern Wolves is difficult to quantify, but, of note, an estimated > 162,000 people have ventured to Algonquin Park to participate in the educational Wolf Howl Program since the 1960s (Steinberg pers. comm. 2013). This program provides a unique educational and recreational component of tourism in Ontario.
The value of Eastern Wolf to trappers is unknown. Large canids are trapped and hunted within the range of the Eastern Wolf (see Legal Protection and Status) but the number of pelts from Eastern Wolves is unknown because pelts of mixed/unknown genotypes typically are grouped together for auction.
The Eastern Wolf also has special significance because it may be the last significant wild population of the Red Wolf, a critically endangered species in the United States (see Taxonomic Validity; Appendix A). In addition, larger carnivores influence ecosystems (Chapron et al. 2008; Beschta and Ripple 2009, 2010), and the role of the Eastern Wolf in the region likely differs from that of the smaller Eastern Coyote, and the larger Gray Wolf.
The Eastern Wolf, as described here, is not currently found in the United States, although it probably did historically range across the eastern temperate forest region of North America (Figure 3) (Wilson et al. 2000; Kyle et al. 2006; Rutledge et al. 2010d).
The historical distribution for this species is difficult to assess because of the limited availability of fossil information and the varying interpretations of Eastern Wolf taxonomy. If one accepts the interpretation that the Eastern Wolf and the Red Wolf were historically the same species, then both genetic data and the fossil record suggest that the Eastern Wolf/Red Wolf historically occupied all of the eastern temperate forests of North America (Wilson et al. 2000; Rutledge et al. 2010d), an area nearly 3 million km2 (Figure 3). This range extended west just beyond the Mississippi River, south to Florida, and east to the Atlantic coast. Although Red Wolves historically occupied regions west of the Mississippi River in Louisiana and Texas (where the captive breeding population was collected), the introgression of Coyote, Gray Wolf, and possibly Dog genes in the Red Wolf population suggest these regions were historical hybrid zones. Thus, the Mississippi River may have been a natural geographic barrier that presumably limited hybridization eastward. Nowak (1995) suggested a smaller, more northern historical range for Eastern Wolves (Figure 3a), independent of Red Wolves.
Presently, Eastern Wolf, as defined in this report (Q≥0.8; see Appendix B), are not believed to be present outside Canada. Under the 3-species hypothesis, the Red Wolf and Eastern Wolf likely were the same species but have since differentiated enough due to introgression from Dog and Coyote genes during the breeding program (see Appendix A) that they no longer fit the Q≥0.8 threshold used in this report (see Appendix B). The Eastern Wolf mtDNA found in samples in Minnesota and area are considered hybrids of Eastern Wolf and Gray Wolf (= Great Lakes-Boreal Wolf, or Great Lakes Wolf) (see Appendix A).
The historical Canadian range of Eastern Wolves likely included the Deciduous Forest region of southern Ontario and Québec, but may have overlapped into the southern part of the Great Lakes-St. Lawrence Forest region. Nowak (1995) suggested a northern range that included the Great Lakes-St. Lawrence Forest region and possibly the Acadian Forest (Figure 3a). The historical Canadian range is therefore estimated at between 112,610 - 500,533 km2.
The present Canadian range of the Eastern Wolf (Figure 4) is based on genetic analysis of Canis specimens sampled across much of the Great Lakes region and from Manitoba to the Maritime Provinces (see Search Effort and Taxonomic Validity). The distribution of the Eastern Wolf is based on the available literature that relies on genetic analyses (see Appendix B). Most Canis individuals recorded outside the core range are identified primarily as Great Lakes-Boreal Wolves or Eastern Coyotes (Stronen et al. 2012; Rutledge et al. in prep.)

Long description for Figure 4
Map delineating the present extent of occurrence (126,573 sq. km) of the Eastern Wolf based on genetic analysis of Canis specimens sampled across much of the Great Lakes region and from Manitoba to the Maritime Provinces. Localities where the Eastern Wolf was found are indicated by symbols. Protected areas, reserves, and controlled exploitation zones are indicated by shading.
Although an Eastern Wolf mtDNA control region haplotype and ATPase haplotype (C3/Ccr13/GL2; Catp13) have been found in Manitoba Wolves (Stronen et al. 2010; Rutledge et al. 2010b), most wolves in Manitoba have Gray Wolf mtDNA (Stronen 2009; Stronen et al. 2010). Stronen et al. (2012) examined nuclear microsatellites but only included eight individuals from Algonquin Park, which is considered to be likely insufficient for cluster identification in the presence of gene flow (Fogelqvist et al. 2010), and therefore would have likely been unable to identify Eastern Wolf by the same definition used for the Algonquin Park subpopulation (see papers by Rutledge, Benson; Appendices A, B). However, evidence of Eastern Wolf genes (termed 'new world mtDNA', separate from old world Gray Wolf mtDNA) were found across the Québec to Saskatchewan region, which is interpreted under the 3-species hypothesis as evidence of Great Lakes-Boreal Wolf (Great Lakes Wolf)) being hybrids from Eastern Wolf x Gray Wolf. In general, Manitoba Wolves cluster more closely with Great Lakes-Boreal Wolves of the Great Lakes states, northern Ontario, and northern Québec, although with greater influence from Gray Wolves from the Northwest Territories (Wheeldon 2009). Wolves from Manitoba are considered different from the Eastern Wolves of Algonquin Park (Wheeldon 2009; Wilson et al. 2000).
The distribution of Eastern Wolf in Ontario is from west of Algonquin Park to the Ottawa Valley, and south to Frontenac Park. Distribution is discontinuous though, with records mainly confined to various protected areas (Figure 4).
Twelve Eastern Wolves have been identified in Kawartha Highlands Park (376 km2) south of Algonquin Park (Benson et al. 2012). Although there is some evidence that reproducing Eastern Wolves have established themselves in WMU49 (Benson et al. 2012), Eastern Coyotes and admixed individuals are more prevalent there, likely due to higher mortality of Eastern Wolves outside protected areas (Benson et al. 2012).
Eastern Wolf in Québec were identified by two methods. The methods are very similar except that the analyses by Stronen et al. (2012), Rogic et al. (2014), Henault (unpub. data), and Tessier (unpub. data) used fewer samples from the Algonquin Park area, resulting in a broader definition of Eastern Wolf and thus includes more samples that would be labelled as admixed animals than if the method in Rutledge and White (2013, 2014) was used. Results from both methods are presented (Figure 4) but the method used by Rutledge and White (2013, 2014) was used in both Ontario and Québec and is given priority in establishing the boundaries of the extent of occurrence (EO). Samples from the other method located beyond the EO are not included here. More work on the distribution of Eastern Wolf would help define the distribution of Eastern Wolf in Québec.
In Québec, 16Eastern Wolves were identified from 457 Canis samples using the same techniques used to identify Canis individuals from Ontario (Rutledge and White 2013, 2014; Table 1; Figure 4). For example, five Eastern Wolves were located in Papineau-Labelle Reserve (1628 km2), two were located north of Mattawa, one was found in the Réserve faunique Rouge-Matawin (Rouge-Matawin Reserve) and another was sampled northwest of the Reserve. Single Eastern Wolves were found in the Réserve faunique des Laurentides (Laurentides Wildlife Reserve), and near Saint-Basile, Québec (Figure 4). A sample of a single male trapped south of the St. Lawrence River in Québec near the village of Sainte-Marguerite-de-Lingwick had previously been identified as an Eastern Wolf because its microsatellite profile was consistent with an Eastern Wolf from Algonquin Park (Villemure and Jolicoeur 2004), but recent work has established it as admixed, and not an Eastern Wolf (Rutledge and White 2014).
| Location | Sample Size | # Eastern Wolves (Q≥0.8) | % Eastern Wolves |
|---|---|---|---|
| ManitobaNotebof Table 1 | 36 | 0 | 0 |
| OntarioNotecof Table 1 Notedof Table 1 Noteeof Table 1 Notefof Table 1 | 558 | 154 | 28 |
| QuébecNotegof Table 1 Notehof Table 1 | 457 | 16 (+25Noteaof Table 1) | 3 |
| Total | 1051 | 170 (+25Noteaof Table 1) | 16 |
The single occurrences are possibly vagrants because it is unknown if these animals represent an undersampled subpopulation of resident animals, or whether they are dispersers. They are included as part of the range of Eastern Wolf until further sampling confirms other Eastern Wolves are not in the area.
The method used by Rogic et al. 2014, Tessier (unpub. data) identified 25 additional records during approximately the last 10 years (Figure 4). Eight Eastern Wolf were identified in La Mauricie Park or within 10 km of the boundary (Tessier unpub. data). Wolves in La Mauricie Park had previously been classified as Eastern Wolves, based largely on morphology, the distribution proposed by Nowak (1995), and the presence of an Eastern Wolf mtDNA haplotype (Villemure 2003; Villemure and Festa-Bianchet 2002; Villemure 2011, pers. comm.). Adult wolves in La Mauricie Park are similar in size to Great Lakes-Boreal Wolves and it is likely that several Canis types reside there (Villemure 2003).
Similarly, Rogic et al. (2014) and Tessier (unpub. data) identified 12 Eastern Wolves in Parc national du Mont-Tremblant ('Mont-Tremblant Park') based on recently collected samples analyzed with genetic analysis in Stronen et al. (2012). There also are four recent records from the area north of Mont-Laurier, and one record near Québec City (Hénault unpub. data).
Based on morphological features that included cranial measurements, Jolicoeur and Hénault (2010) suggested that two types of wolves are found in Québec, a smaller wolf (average weight 23 kg) consistent with the distribution of White-tailed Deer (Odocoileus virginianus; hereafter 'Deer'), and a larger wolf within the distribution of Moose and Caribou (Rangifer tarandus) (average weight 30 kg).
Grewal et al. (2004) suggested that wolves in Réserve faunique La Vérendrye (La Verendrye Reserve) were more similar to samples from Abitibi-Temiscamingue, than samples from Algonquin Park, but still thought they were more genetically similar to those from Algonquin Park than areas in southeastern Ontario. However, this conclusion was not based on the more sensitive approach of Bayesian clustering in STRUCTURE, but rather on loosely interpreted RSTand FSTvalues determined on data from eight microsatellites.
Nowak (1995) suggested a historical presence of Eastern Wolves in the Acadian Forest Region of New Brunswick and Nova Scotia and a northern limit was proposed by Nowak (2009) that includes the Great Lakes-St. Lawrence Forest Region from the southeast corner of Lake Superior, through Ontario and Québec to the Gaspé Peninsula (Figure 3a). A Canis type existed in the Gaspé Peninsula and Maritime provinces but was extirpated in the 19th century; the taxonomy is unknown because of a lack of any specimens (Lohr and Ballard 1996). Eastern Coyote are now common in the region but no microsatellite profiles specific to the Eastern Wolf have been found in the population (Way et al. 2010). Therefore, the current Canadian range of the Eastern Wolf does not include the Maritime Provinces.
The current extent of occurrence, based on the minimum convex polygon connecting the points of observed Eastern Wolf occurrence, is estimated to be 126,573 km2 (Figure 4). The extent of occurrence (EOO) is based on records identified from the method used for both Ontario and Québec (Rutledge et al. 2010c; Benson et al. 2012; Rutledge and White 2013, 2014). This area also contains most of the records identified by a similar method used by Rogic et al. 2014, and Tessier and Henault, unpub. data for Québec. Records identified by Stronen et al. (2012) and Rogic et al. (2014) outside the EOO are not included (see 'Quebec' section of Canadian Range). Single records are difficult to assign as resident or vagrant animals but are included in the EOO because the animal may be evidence of a resident population.
The area of occupancy is 29,472 km2 and is based on the size of the sites where the Eastern Wolf has been recorded. The size of each site is used because these sites are protected areas and Canis mortality is less (see Mortality Rates and Threats), or likely less in the case of Québec Wildlife Reserves, where hunting and trapping are allowed, but development and human density typically is lower when compared to non-protected areas.
Many studies have used Canis samples from Eastern Wolf range and compared them to samples from the Great Lakes region, the southern US, and parts of Canada. An unknown number of the same samples are reused in different studies but sampling appears to be intensive and extensive (Figure 5). Wheeldon (2009) analyzed 627 canid samples (at 12 nuclear microsatellites) from Northwestern Ontario (n=87); Northeastern Ontario (n=93); Michigan (n=90); Wisconsin (n=48); Minnesota (n=53); Northwest Territories (n=56); Manitoba, n=36 (north of Duck Mountain National Park, n=11; Duck Mountain National Park, n=13, Riding Mountain National Park, n=12); Québec, n=34 (western Québec, n=24; eastern Québec, n=10); Algonquin Park, n=54; Frontenac Axis, n=52; Texas, n=24. Way et al. (2010) analyzed 583 Canis samples from Northwest Territories, Ontario (including Algonquin Park), Québec, New Brunswick, and the eastern United States. In Manitoba, Stronen (2009) conducted analyses of 221 wolf faecal and hair samples identified as different individuals from Riding Mountain National Park and the Duck Mountains collected from 2003 - 2005.
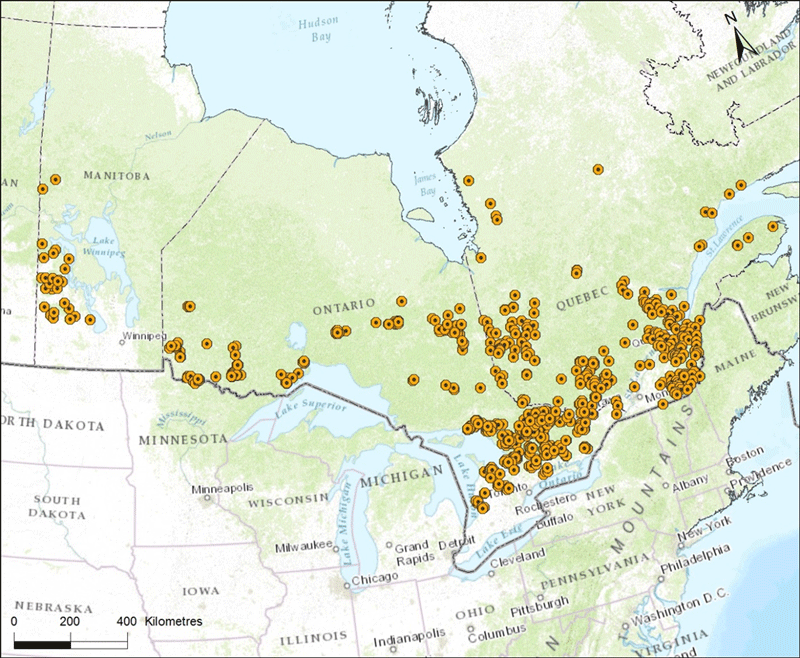
Long description for Figure 5
Map showing the locations in Manitoba, Ontario, and Quebec of Canis samples used by many studies in identifying the distribution of the Eastern Wolf in Canada.
Most sampling of Eastern Wolves in Ontario has been part of a larger study of Canis species conducted by the Ontario Ministry of Natural Resources since 2002. Algonquin Park has been sampled extensively as a result of ongoing wolf research in the Park. Between 1909 - 1958, Park policy promoted a targeted wolf eradication program that ceased to allow a government research project on the ecology of wolves. As part of the project, Pimlott et al. (1969) lethally sampled 106 wolves from the Park, and 19 of those were analyzed by Wilson et al. (2000) at the mtDNA control region. Seventeen of the samples described in Pimlott et al. (1969) have been genetically analyzed with autosomal microsatellites (Rutledge et al. 2011). Grewal et al. (2004) analyzed 102 blood and tissue samples from 27 of the 35 known wolf packs in Algonquin Park sampled between 1987 - 1999. Rutledge et al. (2010c) analyzed 128 samples from Algonquin Park in combination with 51 from northeastern Ontario and 38 from the Frontenac Axis in southern Ontario. Wilson et al. (2009) analyzed 269 samples from 6 geographic regions including southern Ontario, Magnetewan west of Algonquin Park, and northern Ontario.
Stronen et al. (2012) used 109 samples from the Canadian Prairies and 154 from Québec, plus 8 from Algonquin Park, to determine the extent of Eastern Wolf genes from Saskatchewan to Québec. vonHoldt et al. (2011) used a sample of 208 wolves from a worldwide sample, plus 57 Coyote from North America, and other Canis, including domestic dog, to assess issues of hybridization. Only three samples were from the extant range of Eastern Wolf; 2 samples were from Algonquin Park, and 1 from southern Québec.
A total of 457 samples, collected from across Québec (Figure 4), were analyzed with Ontario samples, using the same technique, and interpretation as had been used in Ontario (Rutledge and White 2013, 2014). Rogic et al. (2014) analyzed 15 samples (14 tissue, 1 fecal) collected between 2007 - 2013 in and around Mont-Tremblant Park, and compared them with samples from Stronen et al. (2012).
Numerous research projects on the ecology (e.g. food habits, survival, movement, etc.) of Eastern Wolf have been conducted in Algonquin Park and surrounding areas over the last 50 years (Pimlott et al. 1969; Forbes and Theberge 1995; Forbes and Theberge 1996b; Wydeven et al. 1998; Mills et al. 2008; Loveless 2010; Rutledge et al. 2010e; Benson et al. 2013b, 2014). In Québec, ecological studies also have been conducted in areas containing Eastern Wolf, such as Papineau-Labelle Reserve (Potvin 1988; Potvin et al. 1988, 1992a, b). The degree to which data are specific to Eastern Wolf in these areas is unknown.
Research on habitat use by Canis in Algonquin Park, and Papineau-Labelle Reserve suggests that the Eastern Wolf is not restricted to any particular forest type (Pimlott et al. 1969; Potvin et al. 1988; Theberge and Theberge 2004). The definition of habitat (realized niche) includes competition, predation, prey availability, and den sites. The availability of prey, and mortality, appear to be important habitat features, with evidence that Eastern Wolf are most abundant in areas with abundant prey, such as Deer, and low levels of human-related mortality (see Interspecific Interactions and Threats). The ecological environment and management practices in Algonquin Park represent the most suitable habitat for Eastern Wolves because prey is available and the large size of the Park (7,571 km2), plus the surrounding zone where Canis are free from harvest (6,340 km2), creates the largest number of Eastern Wolf packs exposed to low mortality levels and the least amount of Coyote introgression (Benson et al. 2012). In Québec, sites containing Eastern Wolf also are mainly protected areas where prey populations likely are abundant, although mortality rates may be higher because wolf harvest is permitted in some sites (e.g., Papineau-Labelle Reserve (see Legal Protection and Status).
Pack size is likely associated with territory size and both may be linked to prey density. Eastern Wolves have an average territory size of approximately 190 km2 (±SD 88, n=12; Loveless 2010) in Algonquin Park and 199 km2 (± SE 16, n=19) in Papineau-Labelle Reserve (Potvin 1988). The upper limit of Eastern Wolf pack size ranged from 9 - 14 in Algonquin (Loveless 2010; Theberge and Theberge 2004) and 10 in Papineau-Labelle Reserve (Potvin 1988). Due to their energetic requirements, Eastern Wolves require large prey such as Deer, which are abundant in moderate to high density in the southern part of Ontario (Cervid Ecological Zone E3; OMNR 2009), and Beaver, which are found across Ontario primarily in forested regions but whose abundance fluctuates with habitat suitability (Fryxell 2001). Eastern Wolves may shift from Deer to Moose through the winter via predation (Loveless 2010) or scavenging on Moose killed in association with Winter Tick (Dermacentor albipictus) infestations (Forbes and Theberge 1992).
Den sites are an important habitat feature but dens have been located in a wide range of soil and forest types and are not expected to be limiting factors. In the eastern part of Algonquin Park, den and rendezvous sites are located primarily in pine (Pinus spp.) forests (Norris et al. 2002) possibly due to the sandy, easily excavated soil. Pups are relocated by adults after human disturbance but disturbed dens are often used in subsequent years (Argue et al. 2008). Various rendezvous sites are used into the fall (Mills et al. 2008). Both den and rendezvous sites tend to be located in conifer-dominated landscapes near a permanent water source (Pimlott et al. 1969; Norris et al. 2002).
The amount and extent of vegetative habitat supporting Deer populations likely is increasing in parts of Eastern Wolf range; previously agricultural-dominated landscapes in southeastern Ontario are succeeding into forest at an estimated rate of 1.9% per decade (Lancaster et al. 2008) and may represent improved habitat for Eastern Wolves. Forested landscapes with minimal road networks are still available directly south, west, and east of Algonquin Park, as well as large parts of the range in Québec (Figure 6). The smaller areas in the southern reaches of Ontario may contain only one pack, and likely are too small to sustain viable wolf populations (Benson et al. 2014). A forested corridor with lower road density connects Bon Echo Provincial Park to Frontenac Provincial Park and the Charleston Lake area west of Algonquin Park. The Queen Elizabeth II Wildlands Park probably represents prime Eastern Wolf habitat, and recent work suggests Eastern Wolves are present (Patterson pers. comm. 2014). In Algonquin Park, fire protection and relatively low intensity forest harvest could negatively impact the future amount of early-succession associated species such as Deer and Beaver (Quinn 2004, 2005).
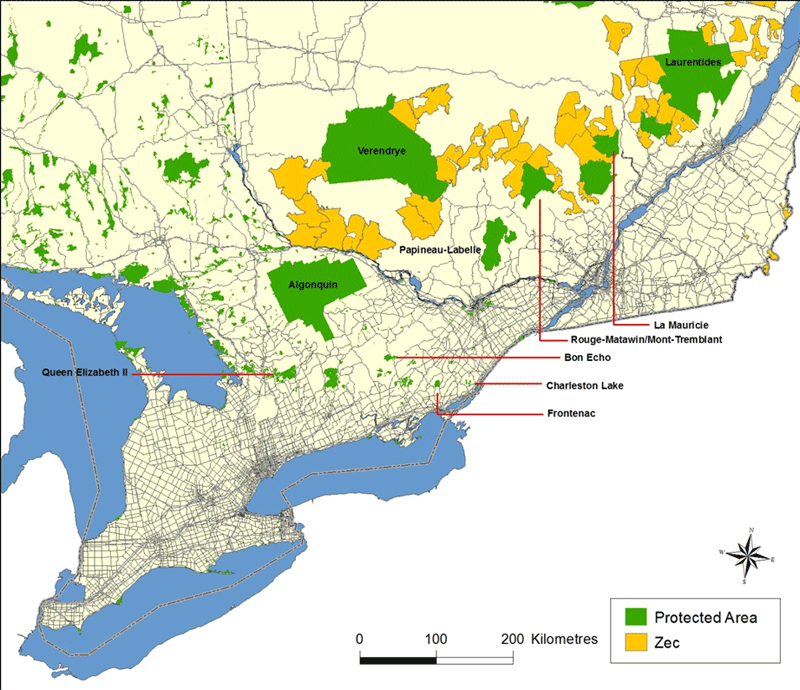
Long description for Figure 1
Map showing the locations of protected areas, controlled exploitation zones, and major paved roads within the range of the Eastern Wolf. Protected areas vary in degree of protection from hunting and trapping, with wolf harvest not permitted in Algonquin, Queen Elizabeth II, Mont-Tremblant, and La Mauricie parks. Reserves and controlled exploitation zones areas are subject to hunting and trapping.
Potential expansion habitats include forested regions extending from Algonquin Park that have limited human-altered landscapes, few road networks/low traffic volume, and protection from hunting and trapping. These areas include regions north to Killarney Park, west to Georgian Bay, south to Hwy 7 and perhaps along the Frontenac Axis through the Algonquin to Adirondack region (although increasing road networks and urbanization may present difficulties in that area), and east across southern Québec's parks and faunal reserves to Saguenay Fjord Park.
Expansion of Eastern Wolves will not occur without protection from hunting and trapping throughout its range because juvenile dispersers are more susceptible to harvest (Theberge and Theberge 2004). Four of five radio-tagged juveniles died from trapping < 1 year after dispersal to areas outside Algonquin Park, and Eastern Wolf were more vulnerable to harvest than other Canis types outside Algonquin Park (survival rates of 33% for Eastern Wolf, compared to 66% for Eastern Coyote, and 85% for Canis within Algonquin Park (Benson et al. 2014)). As well, high harvest rates increase hybridization with Eastern Coyotes (Rutledge et al. 2011; Benson et al. 2014).
Wolves, in general, may benefit from roads because of increased hunting efficiency (Whittington et al. 2011; Lesmerises et al. 2012) but populations can decline if increased access for hunters and trappers facilitates unsustainable harvest rates (Mech et al. 1988; Person and Russel 2008). A road density of < 0.4 - 0.7 km of road per km2 and human density of < 4 per km2 have been suggested as necessary to support large Canids, if mortality from humans is a concern (Wydeven et al. 1998). In most of southeastern Ontario, road density is > 0.6 km per km2 (Buss and deAlmeida 1997; Figure 6) and in southern Ontario road length increased from 7,133 km in 1935 to 23,806 km in 1965, and to 35,637 km in 1995 (Fenech et al. 2000). This trend may limit Eastern Wolf dispersal southward.
Much of our knowledge of wolves derives from studies on Gray Wolves or Great Lakes-Boreal Wolves (e.g. Mech and Boitani 2003). Substantially less information is available on Eastern Wolves. The information presented here is primarily a synthesis of research conducted on Eastern Wolves in Algonquin Park, as well as research from Québec (e.g., Papineau-Labelle Reserve) where Eastern Wolf has been recorded.
Early life stages are divided into: 1) denning (age < 6 weeks); 2) early rendezvous (age 6 - 12 weeks); 3) late rendezvous (age 12 - 17.9 weeks); and 4) rendezvous abandonment (age ≥18 weeks) (Mills 2006). Pups are born in late April to early May. At 6 - 8 weeks they are moved from the natal den to an initial rendezvous site. Throughout the summer, pups are moved to subsequent rendezvous sites that occur at increasing distance from the den (Mills et al. 2008) until the sites are abandoned in fall. Eastern Wolves hunt in packs but pups do not hunt until they are about 18 weeks old and when final rendezvous sites are abandoned.
The average life span of wolves has been reported as 4 or 5 years (Fuller et al. 2003) with some animals reaching at least 15 years (Theberge and Theberge 2004). Survival rates for adults were 63-70% in the late 1980s - early 1990s (Forbes and Theberge 1995). Average annual survival rate (ŝ) of pups is estimated at 0.749 (±0.061 SE) in eastern Algonquin Park and 0.246 (±0.073 SE) in western Algonquin Park (Benson et al. 2013b). The lower survival in western Algonquin Park is influenced by low Beaver density and unavailability of wintering Deer. Survival of yearlings and adults in Algonquin Park was higher (ŝ = 0.852 (±0.05 SE)) than pup survival (Benson et al. 2014). Using these data, life expectancy (LE) (calculated as LE = -1/(ln(ŝ)), where ŝ is the average annual survival rate), of pups is between 0.7 and 3.5 years, whereas yearlings and adults have a life expectancy of 6.2 years. Females do not typically breed before 2 years and therefore the generation time is likely to be approximately 3.5 years.
Eastern Wolves typically have a single, unrelated breeding pair within each pack. Pimlott et al. (1969) documented that 59% of all female adults (≥ 2 years) sampled in Algonquin Park had produced at least one litter. Litters are produced annually and range between 2 - 7 pups, with an average of 4.9 (Mills et al. 2008). Sex-ratio of 5.5 week old pups is 1:1 and mortality is not different between sexes (Mills et al. 2008). Vucetich and Paquet (2000) suggested the average age of Eastern Wolves in Algonquin Park prior to the harvest ban was 2 - 3 years old, which is lower than the post-ban average age of 5 years. Based on capture statistics in Algonquin Park from 2002 - 2007, the proportion of pups in the population is estimated at 39.1%; yearlings and adults comprise 15.0%, and 45.9%, respectively (Patterson, unpub. data).
Most Eastern Wolves exist in packs that are highly social and territorial. Average pack size in Algonquin Park and Papineau-Labelle Reserve is around six animals (Potvin 1988; Forbes and Theberge 1996b). Wolf social structure is important for effective resource use (Sand et al. 2006; Stahler et al. 2006; Loveless 2010), pup survival (Brainerd et al. 2008; Schmidt et al. 2008), avoidance of incestuous mating (vonHoldt et al. 2008), and precluding hybridization with Coyotes (Rutledge et al. 2011).
When naturally regulated, Eastern Wolves live in family-based social packs with little evidence of inbreeding. Rutledge et al. (2010e) documented unrelated males joining packs and becoming the breeder (after the known breeding male could no longer be located with telemetry equipment), and daughters of breeding females were identified as subsequent breeders with unrelated males within their natal pack. When influenced by high human-caused mortality, Eastern Wolf packs become fractured with higher adoption of unrelated individuals (Grewal et al. 2004) and more prevalent hybridization with Eastern Coyotes (Rutledge et al. 2011).
Based on research in the eastern part of Algonquin Park, Theberge et al. (2006) estimated an average Eastern Wolf adult mortality rate of 0.30 between 1989 - 1999, although there was high variability in the estimates (Patterson and Murray 2008). Pimlott et al. (1969) noted high mortality in Eastern Wolves during the first year of life. Theberge and Theberge (2004) noted that yearlings and subadults were more susceptible than adults to human-caused mortality. Pup survival is estimated at 81% up to about 37 weeks of age with no observable mortality in the first 11 weeks (Mills et al. 2008). Within Algonquin Park, natural causes account for the majority of deaths (conspecific strife, drowning, Black Bear [Ursus americanus] predation, injuries caused by ungulates) followed by human-causes (poisoning and shooting after movement/dispersal outside the protected area).
Canine parvovirus (CPV-2) and canine distemper (CD) are not associated with pup deaths although exposure is evident (Mills et al. 2008). Pup mortality seems to vary among packs, for unknown reasons. Benson et al. (2013b) documented a total mortality rate in Algonquin Park of 44.3%, with natural causes accounting for 33.9% of deaths, 4.6% caused by human factors, and 5.8% unknown causes; pup mortality risk is associated with Beaver density and Deer availability. Outside Algonquin Park, high adult mortality is linked to Eastern Wolf ancestry (Benson et al. 2014) and poor pup survival is linked to Eastern Wolf x Eastern Coyote admixed ancestry (Benson et al. 2013b), making dispersal and expansion difficult for Eastern Wolves.
Theberge et al. (1994) reported that 6 of 18 radio-collared wolves died of rabies during an outbreak in 1990 - 91. Rabies has not been recorded in the population since then (Patterson 2011a, pers. comm.). Although human-caused mortality accounted for approximately 67% of wolf deaths in the eastern part of the Park prior to 2001, a ban on hunting and trapping in the three townships at the south end of the Park and surrounding townships, was implemented in December 2001 and has been successful in reducing human-caused mortality to 16% of all deaths (Rutledge et al. 2010e). The number of natural deaths, however, increased and generally offset those from human causes. Inter-pack strife is currently the main natural cause of death for adult Eastern Wolves in eastern Algonquin Park (Rutledge et al. 2010e) and starvation is a significant mortality risk for pups in the western part of the Park (Benson et al. 2013b).
In 2002/2003, directly after the implementation of the hunting and trapping ban in townships surrounding Algonquin Park, Eastern Wolf survival in the Park was 95.2% (±0.03 SE), but levelled off between 2003 - 2006 with survival rates between 81.6% and 84.2% (Patterson, unpub. data). Survival decreased significantly in 2006/2007 to 69.7% (±0.06 SE) when mange (caused by Sarcoptes scabiei) killed a large number of Eastern Wolves in the Park (see Fluctuations and Trends). Mange was rare and not debilitating for wolves in Algonquin Park from 1987 - 1999 (Theberge and Theberge 2004) suggesting that mortality (and thus population size) can fluctuate significantly from year to year due to unexpected events such as disease outbreaks.
Wolf hunting and trapping in Québec is prohibited in national and provincial parks but occurs elsewhere, including in wildlife reserves (St-Louis pers. comm. 2012). Human-caused mortality from trapping, vehicles, and hunting removed about 25-30% of a tagged population in the 1980s - 1990s (Jolicoeur and Hénault 2010). Harvest rates in Québec range regionally from 2.8 to 29.5%, with a provincial average of 5.9%. Villemure and Festa-Bianchet (2002) documented mortality of 53.3% in radio-collared wolves from La Mauricie Park, mostly due to trapping outside Park boundaries, which accounted for 87.5% of all mortalities, with pups most affected. A study conducted in Papineau-Labelle Reserve in the early 1980s documented a 34% mortality rate, with hunting and trapping accounting for 66% and roadkill 33% of the 33 Canis dying (Potvin 1988). Overall harvest rate in the Papineau-Labelle region was 9.0 - 17.3% in the late 1990s (Hénault and Jolicoeur 2003).
Eastern Wolves also suffered some mortality as a result of capture associated with research activities in Ontario. Between 2002 - 2011, 8 of 328 (2.4%) captured wolves died from capture-related mortality caught in traps, live-snares, or by heli-net-gunning (Patterson 2011c, pers. comm.). Two pups drowned post-capture because they had been relocated to a den site that later flooded; a third pup died 8 days post-capture of unknown causes (Argue et al. 2008).
Gray Wolves are typically defined as highly adaptable to climatic variations given that they historically occurred from Mexico to the Arctic. There is considerably less information available on Eastern Wolves but it is likely that they have similar adaptability as other large Canis species.
Individual wolves can become acclimated when fed by people and when scavenging at landfill sites, and young dispersers sometimes frequent landfills (Mills et al. 2008).
Some adult Eastern Wolves in Algonquin Park migrated up to 42 km annually between their non-winter territory boundary and Deer concentration areas in response to Deer leaving the area during winter (Forbes and Theberge 1995; Forbes and Theberge 1996b; Loveless 2010; Rutledge et al. 2010e).
Annual dispersal probabilities were 0.22 (95% CI=0.108 - 0.318), 0.508 (0.341 - 0.633), and 0.144 (0.099 - 0.187) for pup, yearling, and adult Canis in Algonquin Park, respectively (B. Patterson unpub. data). Although dispersers often die or sometimes wander indefinitely, they typically form new packs or join existing packs as early as December. Once rendezvous sites are abandoned, juveniles start hunting and moving along with the pack.
Eastern Wolves have dispersed up to 555 km, and can cross mixed landscapes of forest, farmland, and 4-lane highways (Wydeven et al. 1998). Wolves from Algonquin Park dispersed south into the Kawartha Highlands Park, west toward Georgian Bay, and east into Québec. Radio-tagged juvenile Eastern Wolves typically did not disperse north, although one disperser from Algonquin Park was located in Nakina, northwestern Ontario, a direct distance of approximately 800 km (Patterson pers. comm.).
The main impediment to successful dispersal is high human-caused mortality, primarily hunting and trapping, outside protected areas (Forbes and Theberge 1996a; Wydeven et al. 1998; Benson et al. 2013b, 2014). In Ontario, when human-caused mortality was high, hybridization with Eastern Coyote increased (Rutledge et al. 2011) probably in response to a lack of conspecifc mates (Rutledge et al. 2010c) and disruption of pack social structure (Rutledge et al. 2010e). Expansion into historical range is also limited by lack of sufficiently connected forested areas containing low road and human densities (Harrison and Chapin 1998). Expansion northward appears to be limited, possibly due to the presence of the larger Great Lakes-Boreal Wolf and/or ecological requirements.
In Papineau-Labelle Reserve, Canis (of which some proportion likely were Eastern Wolf) mainly consumed Deer during winter, and Beaver and Moose during summer (Potvin et al. 1988). Beaver populations increased by 30% after 60% of Canis were removed (Potvin et al. 1992b). Similarly, Forbes and Theberge (1996b)found a high correlation between Eastern Wolf density and Deer numbers in winter. Prey selection by Eastern Wolves varies across months, seasons, and years (Forbes and Theberge 1996b; Loveless 2010). For example, in the eastern part of Algonquin Park, Theberge and Theberge (2004) noted that use of Beaver decreased from May to June and that consumption of adult Deer was highest in July. In some years, Moose were consumed more than Deer, although a high proportion was likely from scavenging. Eastern Wolves were documented as mostly scavengers of Moose in the 1980s (Forbes and Theberge 1992). More recent work suggests that they are an effective Moose predator; in 2006, Loveless (2010) noted that the proportion of Moose in the diet of Eastern Wolves was higher (64%) than in the preceding year (47%), and that overall biomass consumption increased from fall through mid-winter and decreased in late winter. Most Moose carcasses fed on were adults and the overall proportion of Moose vs. Deer in the diet tended to increase throughout the winter. Most Moose were depredated by packs that appeared to specialize on Moose. In the winter, pack size was correlated with Moose predation. Higher kill rates on Deer occurred in the fall, presumably in response to the fall migration of Deer. Older moose (>11 yrs) and calves were taken in higher proportion than their presence in the overall population.
The only known predator of adult Eastern Wolves are humans, but Black Bear occasionally prey on wolf pups (Mills et al. 2008). There have been no documented human deaths as a result of Eastern Wolf-human interactions, although injuries to people have occurred in Algonquin Park, likely as a result of wolves being fed within the campgrounds (Linnell et al. 2002).
Interactions with Eastern Coyote are discussed in the Threats section.Diseases are discussed in the Threats section because most diseases associated with Eastern Wolves are related to invasive disease and domestic animals.
The population size is unknown, but likely is < 1000 mature animals. Eastern Wolf are identified by genetic analyses of samples that have been collected in haphazard manner with different effort and during different time periods; an accurate population cannot be derived from such data. There have been170 individuals identified in the common analysis for Ontario and Québec samples (Rutledge et al. 2010c, Benson et al. 2012; Rutledge and White 2013, 2014) and another 25 identified by Rogic et al. (2014), Henault (unpub. data), Tessier (unpub. data) within the extent of occurrence, for a total of 195 Eastern Wolf specimens from various sampling efforts over the last 10 - 15 years.
| Location | Area (km2) | Maximum # of Eastern Wolves in these Sites |
Estimated # of Eastern Wolves (Q≥0.8) |
Estimated # Mature Individuals |
|---|---|---|---|---|
| Algonquin Park, ON | 7,571 | 227 | 157 | 72 |
| Algonquin Townships, ON | 6,340 | 190 | 131 | 60 |
| Kawartha Highlands Park, ON | 376 | 11 | 8 | 4 |
| Killarney Park, ON | 645 | 19 | 13 | 6 |
| Burwash/Bird Lake Area, ON | 360 | 11 | 7 | 3 |
| French River Park, ON | 735 | 22 | 15 | 7 |
| Queen Elizabeth II Park, ON | 255 | 8 | 5 | 2 |
| Papineau-Labelle Reserve, QC | 1,628 | 42 | 29 | 13 |
| Rouge-Matawin/Mont Tremblant, QC | 3,165 | 60 | 41 | 19 |
| La Mauricie Park, QC | 536 | 10 | 7 | 3 |
| Laurentide Reserve, QC | 7,861 | 149 | 103 | 47 |
| Total | 29,472 | 749 | 516 | 236 |
| Total (Extent of Occurrence)Noteiof Table 2 | 126,573 | 3797 | 2620 | 1203 |
In Québec, Canis densities are available for two sites containing Eastern Wolf; Papineau-Labelle Reserve has an estimated 2.6 Wolves/100 km2 and Rouge-Matawin Reserve has 1.6 - 1.9 Wolves/100 km2 (Jolicoeur and Hénault 2010). It is assumed that Mont Tremblant Park, which is adjacent to Rouge-Matawin Reserve (Figure 6), has the same Canis density as Rouge-Matawin Reserve, as do the La Maurice and Laurentide sites, which are to the east. The Québec population size is estimated to be 82 mature animals.
The population of "Algonquin ecotype" wolves in Québec had been roughly estimated at 585 Wolves, based on a deciduous forest habitat and distribution of Deer (Jolicoeur and Henault 2002). The genetic analysis of Québec Canis, however, suggests few of these animals likely are Eastern Wolf with Q≥0.8 (Rutledge and White 2014; Figure 4).
In the previous COSEWIC status assessment (Samson 2001), the number of adult and juvenile Eastern Wolves in Canada was estimated at 1,311 - 2,684 over 210,000 km2. The current minimum estimate of Eastern Wolves is 344 (including individuals of all ages) and the maximum is 1203 over a total extent of occurrence of 126,573 km2 (Table 2). Lower numbers between COSEWIC reports reflect a reassessment of the extent of occurrence and changes to how Eastern Wolves are identified, rather than a measurable/comparable change in population size and range.
The large Canis species was extirpated from the Maritimes, eastern United States, and southern parts of Ontario and Québec. If the Eastern Wolf inhabited this entire region (Figure 3b), or a more northerly part only (Figure 3a), then the historical loss of the Eastern Wolf population has been at least 50% of its range.
Recent population trends in Québec are not known. In Ontario, research efforts since the late 1950s have documented several atypical mortality events that caused short-term declines. Killing of wolves for research in 1964 - 1965 (Pimlott et al. 1969) decreased wolf density in the 1,700 km2 study area from 4.7 wolves per 100 km2 (n=80) in 1964 (prior to the study) to 2.4 per 100 km2 in 1972 - 1973 (n=41). Between 1988 - 1999, Theberge and Theberge (2004) recorded a trend of decreasing wolf abundance in the eastern part of Algonquin Park due to human-caused mortality. Wolf density averaged 2.37 wolves per 100 km2 (±0.57 SD) with the lowest recorded density at 1.4 per 100 km2 in 1999. Estimated abundance in the study area was highest in 1991 - 1992 at 93 adults, and lowest in 1998 - 1999 at 38, suggesting a decrease in adults from 43 to 17, a 60% decrease in the study area, and a 5.0 - 16.6% decrease in the overall population of mature individuals. Survival was high immediately after the hunting ban was implemented in December 2001 (s = 95.2%), which presumably accounted for an increase in density. Survival stabilized between 2003 - 2006 at 81.6 - 84.2%, and was 85% between 2007 and 2010 (Benson et al. 2014). In 2006/2007, the population decreased significantly (p< 0.05) to 69.7% due to a mange outbreak (Patterson 2011b, pers. comm.) but increased afterwards.
A population viability analysis (PVA) by Vucetich and Paquet (2000) suggested that the population of Eastern Wolves in Algonquin Park was in decline between 1988 - 1999 and that annual mortality (33%) exceeded annual recruitment (21%). Subsequently, Theberge et al. (2006) suggested it would be difficult to identify a positive response in wolf abundance to the harvest ban within 30 months of its implementation due to high stochasticity. Patterson and Murray (2008) suggested these analyses were flawed and in their re-analysis they concluded that a response in abundance would be detectable and that the population of wolves was unlikely to decrease significantly within the next 20 years. In Algonquin Park, the low occurrence of incestuous mating since the implementation of the harvest ban suggests inbreeding is not currently an issue within the Park; levels of inbreeding outside the Park are unknown but are believed to be higher, based on genetic analyses (Rutledge et al. 2010e).
The Eastern Wolf, as presently described, is not present in the United States and thus rescue effect is not possible. Although Red Wolves in the southeastern United States and Eastern Wolves in Canada probably evolved from the same historical population, there are high levels of hybridization in the Red Wolf population, including genetic contributions from Coyote, Gray Wolf, and Dog (see Appendix A). This mixed genome, combined with the limited evolutionary potential due to a genetic bottleneck associated with a founder effect of the captive breeding program, make the Red Wolf population an unviable option for an Eastern Wolf rescue effect in Canada.
The Threats Calculator results are summarized in Appendix C. The overall threat score was Very High to High, based mainly on the likely continuing threat of mortality, and its impact on population expansion outside protected areas.
Territorial behaviour by Eastern Coyote may limit the expansion of Eastern Wolf from protected areas. There is evidence from research conducted in and near Algonquin Park that both Eastern Coyote and Eastern Wolf packs are territorial (Benson and Patterson 2013a). Eastern Wolf individuals travelling by themselves are unlikely to establish in areas already containing a pack of Eastern Coyote. Within the extent of occurrence, most areas outside the cores areas of Eastern Wolf contain Eastern Coyote.
In general, wolves are capable of living in agricultural areas and low density residential and commercial areas (Musiani et al. 2010). Deer populations may be abundant enough in rural areas, and food could be supplemented by scavenging garbage. However, wolves generally are not tolerated by society and may be killed; wolves typically are more common in areas without human activity or in areas with a lower density of agricultural, residential, or commercial development. The Threats Calculator exercise considers proximate factors, and therefore direct threats, such as habitat loss due to the expansion of housing, are presented in the Urban and Residential Development category, while a mortality event, regardless of it occurring in residential or forest areas, is discussed in the Hunting and Trapping category.
Wolves are a regulated furbearer species in the range of Eastern Wolf (see Legal Protection and Status). Eastern Wolves are protected from harvest within their core range of Algonquin Park, surrounding townships, and some other protected areas (see legal Protection and Status). In the remainder of their range, Eastern Wolf is subject to harvest. The number harvested is unknown because the canids appear similar enough that they are combined in auction summary data. In the 2011/2012 season, 571 'wolf' pelts from trappers in Québec were sold (MFFP, 2012c). In the Great Lakes - Ontario region, 98 "Timber Wolves" including 'Arctic' and 'Eastern' were offered in the auction and 5,545 'Coyote' pelts, including those from western and eastern geographies (Fur Harvesters Auction Inc. 2011).
Although most of the known population exists in protected areas, it is likely that mortality from humans limits the population elsewhere, and restricts expansion. Prior to the implementation of a hunting ban around Algonquin Park, human-caused mortality accounted for approximately 67% of wolf mortality in the eastern part of the park (Forbes and Theberge 1996a); post-ban (see Legal Protection and Status) it has decreased to 16% (Rutledge et al. 2010e). Outside the protected regions, hunting and trapping results in high mortality rates (Jolicoeur and Hénault 2010; Benson et al. 2014).
The degree to which pelt records indicate mortality rates is unknown. In Ontario, some people shoot wolves and Coyotes while hunting for Deer or Moose, in the belief that this increases ungulate populations (Patterson 2012b, pers. comm.). The extent of mortality from this behaviour is unknown in Eastern Wolf range.
Road density has been identified as a primary limiting factor to Eastern Wolf dispersal because roads are a proxy for increased hunting and trapping harvest. Hybridization with Eastern Coyotes is higher in regions of high road density (Benson et al. 2012), likely because high mortality rates associated with harvest access appear to facilitate Coyote introgression (see Taxonomic Validity and Habitat Trends). High road density in southern Ontario and Québec (Figure 6) likely would be a potential for roadkill and may severely impede natural expansion of Eastern Wolves into their historical range. Between 1985 and 1995 the number of single and multi-lane paved roads in southern Ontario increased by 3,025 km (Fenech et al. 2000), an average increase of 302.5 km per year, and suggestive that road density will continue to increase outside protected areas.
The presence of agricultural areas may mean increased Deer availability but these sites may be problematic for Eastern Wolf because of mortality by people in response to depredation of livestock in potential expansion areas. This threat is difficult to quantify because of the high variability in response from farmers (e.g. Stronen et al. 2007) and the divisive attitudes towards wolf management in agricultural areas where livestock predation occurs (Mech 2010b; Way and Bruskotter 2012).
Although positive attitudes towards wolves have increased over the past century (and in particular since the 1970s), attitudes can still be characterized as hostile in certain regions (Bruskotter et al. 2007, 2011; Williams et al. 2002). In the 1980s and 1990s, attitudes towards wolves in communities near Algonquin Park were generally unfavourable (Theberge and Theberge 2004). In Québec, the attitudes of trappers to wolves is variable, with about 42% opposing management to increase wolf numbers and the remainder either supporting the increase (29%), or undecided (29%), especially as it pertains to La Mauricie Park and surrounding regions (Bath 2006). A survey of residents near La Mauricie Park suggests most are sympathetic to the cause of the Eastern Wolf but most overestimated the number of wolves in the region and therefore were not necessarily supportive of management that aimed at increasing the population size (Parcs Canada 2007). The results of the survey suggest that better education of the public regarding wolves is an important component of wolf conservation in Québec. Societal shifts in values have been proposed as the most important factor in sustaining wolf populations worldwide (Musiani and Paquet 2004).
The impact of vehicular collisions on the population of Eastern Wolf is unknown. Roads with high traffic volume exist throughout the extent of occurrence but are much less common in the protected sites. It is unlikely that there will be a large increase in new high volume roads, but traffic volume will likely increase on existing roads. A third of radio-collared Canis in Papineau-Labelle Reserve died from vehicles (Potvin 1988), and 2.7 ± 1% (SE) of adults, and 3.7 ± 2% (SE) of pups died in a sample of radio-collared Eastern Wolf in the Algonquin Park area from 2002 - 2007 (B. Patterson, unpub. data). From 2004 - 2010, 4.9% of Eastern Wolf were killed by vehicular collision in the same study area (Benson et al. 2014). The indirect threat of roads is discussed under biological resource use (IUCN 5.1).
Coyotes were first documented in Ontario in Lambton County in 1919 (Nowak 1979). Since that time, they have hybridized with Eastern Wolves, resulting in a thriving population of Eastern Coyotes that are common across southern Ontario and from the eastern United States to Newfoundland (Way et al. 2010). The adaptability of these animals to human-modified landscapes has resulted in rapid colonization of areas in eastern North America (Kays et al. 2010) historically occupied by Eastern Wolves. High human-caused mortality is probably the primary cause of current hybridization of Eastern Wolves with Eastern Coyotes, except in Algonquin Park, where protection has curtailed ongoing hybridization (Rutledge et al. 2011). Outside the protected regions, however, almost all Canis are identified either as smaller Eastern Coyotes or highly admixed individuals (Eastern Wolf x Eastern Coyote hybrids). Very few are identified as Eastern Wolves despite absence of physical/geographic barriers. Thus, outside protected areas, gene introgression by Eastern Coyotes threatens the persistence of Eastern Wolves due to initial hybridization between the two species and then back-crossing to Eastern Coyotes (see Habitat Requirements and Habitat Trends).
An increase in modified habitat, including land clearing for housing, can represent a decrease in habitat quality if prey species are not present. An area with high housing density will generally not contain prey for Eastern Wolf. However, much of Eastern Wolf range is composed of rural farms, cottages, and small towns, and Deer would occur in the rural areas. The main threat likely relates to increased contact between people and wolves, which could lead to higher wolf mortality; this indirect threat is discussed under biological resource use (IUCN 5.1). Urbanization and ongoing land conversion will continue to indirectly threaten Eastern Wolves' potential expansion. Residential and commercial development presently is a minor threat to Eastern Wolf because most of the population is located in protected areas that limit development (Figure 4). The projected rate of development of residential and commercial activity outside protected areas is unknown, but it is not likely to decrease.
An increase in modified habitat, including land clearing for agriculture, can be a direct problem for Eastern Wolf if prey species are not present. Although prey base in areas of Eastern Wolf range is unknown, Deer often are present in agricultural areas and agriculture is decreasing in some areas and reverting to forest (see Habitat Trends section).
Human intrusions, such as visitation to dens or rendezvous sites, could have an impact on Eastern Wolves because wolves tend to avoid humans and relocate pups after den disturbance (Frame et al. 2007). The effect likely would be limited because the core of the population occurs in protected areas where human activities are regulated. The extent of disturbance is unknown, but likely is negligible.
Domestic dogs have exposed Eastern Wolves to domestic diseases, including canine parvovirus-2 (CPV-2), canine distemper (CD), and canine hepatitis (CH). Antibodies for these diseases are found in most Eastern Wolf pups at an early age, but exposure does not typically result in death (Theberge and Theberge 2004; Mills et al. 2008). Theberge and Theberge (2004) noted antibodies in Eastern Wolves for CPV (82%), CH (76%) CD (46%), and rabies (20%). Although domestic disease-related deaths have been rare in wolves from Algonquin Park, the Isle Royale National Park, Minnesota, wolf population suffered a dramatic decline in numbers between 1980 - 1982 due to a CPV outbreak that left the population with only 14 wolves (Peterson et al. 1998). Disease can be an important extinction risk in small relict wild canid populations (Woodroffe et al. 2004).
The threat exists because there is a potential for mortality from domestic dogs associated with tourism in protected areas, and a decrease in adult animals could increase levels of Coyote gene introgression in Eastern Wolf (see Taxonomic Validity). The threat level is considered to be low because a level of immunity to parvovirus appears to exist in the population.
Rabies in wolves and coyotes is rare, although 15 cases were documented in Ontario between 1960 - 1994 (Theberge et al. 1994). During winter of 1990 - 1991, 4 wolves (perhaps 6) died of rabies (Theberge et al. 1994), but no rabies-related deaths have since been documented (Patterson 2011, pers. comm.). The rabies outbreak in Algonquin Park wolves was thought to be linked to an outbreak of fox strain rabies during 1990 - 1991 (Theberge and Theberge 2004). The survival of those Eastern Wolves that had antibodies for rabies is thought to be due to ingestion of oral rabies vaccine bait that was distributed in eastern Algonquin Park in 1991 - 1992.
There currently are no direct estimates of how climate change could affect Eastern Wolves. However, effects of climate change on Moose populations predict a decline in Moose density in the southern limits (south of Hwy 101; ≈48th parallel) of Ontario, particularly in the region of Algonquin Park (Rempel 2011). In general, models predict a northward movement of Moose with populations in the southern regions receding due to marginal climate and habitat (Murray et al. 2006) and expansion of parasitic disease associated with increased Deer density (Lankester 2010). Although Murray et al. (2006) suggested that climate change should benefit Deer populations, they also noted that it is difficult to predict how warmer temperatures will impact deer parasites. In general, Deer density is expected to increase across Ontario and Québec (Thompson et al. 1998), which could benefit Eastern Wolves. Lankester (2010) also suggested winters will get shorter and milder with longer snow-free periods, which will reduce the hunting success of Eastern Wolves since they have an advantage compared to ungulates in deep snow conditions (DelGiudice et al. 2002; Crête and Larivière 2003).
Most studies on climate change have focused on plant-herbivore actions and there are relatively few studies that examine the impact of climate change on high-level trophic interactions. Wilmers et al. (2007) demonstrated that in the absence of wolves, Moose populations are predicted to be more susceptible to climatic variations. Wilmers et al. (2006) provided evidence that wolves mediate the impact of climate change on ungulate populations. Thus a healthy Eastern Wolf population may be important for mediating fluctuating ungulate populations predicted in response to climate change. The direct and indirect effects of climate change on the Eastern Wolf population are unknown.
The Eastern Wolf is listed as Special Concern on Schedule 1 of the federal Species at Risk Act (SARA), under the scientific name Canis lupus lycaon. An Eastern Wolf Management Plan, required under SARA for special concern species, is under development by Environment Canada.
Gray Wolves are listed under Appendix II of the Convention on International Trade in Endangered Species (CITES), which is administered in Canada by Environment Canada. CITES permits are required to export wolves from Canada, including Eastern Wolf specimens (recognized as Canis lupus lycaon). Other than CITES, there are currently no international agreements for protection that would include Eastern Wolf.
The Eastern Wolf is considered to be a subspecies of Canis lupus.
The Eastern Wolf is listed under Ontario's Endangered Species Act, 2007 (Ontario Government 2007) as a Gray Wolf subspecies (C. lupus lycaon) with a status of Special Concern. The NatureServe S-rank is S4 for Ontario.
Under the Fish and Wildlife Conservation Act, 1997 (Ontario Government 1997), all animals of the species Canis lupus are considered furbearing mammals, and are provided protection. Hunting and trapping of wolves are regulated under regulations of the Fish and Wildlife Conservation Act, 1997, and harvest is controlled and monitored. In addition to a small game licence, a wolf/coyote game seal (maximum of 2 per year) is required to hunt wolves in core wolf range (Wildlife Management Units 1A, 1C, 1D, 2-42, 46-50 and 53-58). Wolves can also be hunted or trapped under a trapping licence. The Act provides authority to limit harvest by trappers if necessary.
Where regulated hunting and trapping are permitted, the wolf and Coyote season is closed from April 1 to September 14 across core wolf range identified above. Wolves (along with Coyotes) can be hunted and trapped year round south of core wolf range, and there are no limits on harvest. However, this area is considered outside the current extent of occurrence of Eastern Wolves, but within their historical range.
There is no open hunting/trapping season for wolves in 40 townships around Algonquin Park: Airy, Alice, Ballantyne, Boulter, Boyd, Burns, Butt, Calvin, Cameron, Chisholm, Clancy, Clara, Dickens, Dudley, Eyre, Finlayson, Franklin, Fraser, Hagarty, Harburn, Harcourt, Havelock, Head, Herschel, Lauder, Livingstone, Maria, McClintock, McClure, McCraney, McKay, Murchison, Papineau, Paxton, Petawawa, Richards, Rolph, Sabine, Sinclair, and Wylie.
Wolves may not be hunted or trapped in the following protected areas: Pukaskwa Park, Algonquin Park, and all Crown Game Preserves. They are protected from regulated hunting in all other Provincial Parks (except Point Farms Provincial Park in southern Ontario, which is presumed to be outside core wolf range), and are protected from trapping in over half of the Provincial Parks. Wolves can be trapped in Kawartha Highlands and French River Park. Trapping will be eliminated from one-third of the remaining parks where it currently occurs under a phase out policy.
Mandatory reporting of activity and harvests is required by hunters and trappers. A person who kills a wolf in protection of property must either register the acquisition of the carcass online, if keeping the carcass, by submitting a Notice of Possession, or report the killing immediately to the Ministry if not keeping the carcass.
Residents and non-residents must have an Ontario export licence for furbearing mammals and their pelts to export a wolf or Coyote, as well as its pelt, from Ontario whether it originated from Ontario or not, and including carcasses and live wolves and coyotes. Royalties are payable on Ontario wolves and Coyotes prior to their export from Ontario.
The Strategy for Wolf Conservation in Ontario (OMNR 2005) provides the strategic policy direction for the management of wolves in Ontario.
Den and rendezvous sites for Eastern Wolf on Crown Land are protected as part of provincial forest management guidelines (OMNR 2010).
In Québec, wolves are considered a furbearer and are protected under An Act Respecting the Conservation and Development of Wildlife but not under Loi sur les espèces menacées ou vulnérables [Act respecting Threatened or Vulnerable Species in Québec]. The Eastern Wolf is not officially recognized because the province does not recognize wolf subspecies (MFFP 2011); the Act only acknowledges taxonomic nomenclature as outlined by the Smithsonian Museum of Natural History and the Integrated Taxonomic Information System (MFFP 2012a). Currently, wolf hunting and trapping is prohibited in all federal and national (provincial) parks but permitted elsewhere, including wildlife reserves (St. Louis pers. comm.). In most areas, the harvest season is late October - late March, and there is no bag limit.
NatureServe (http://www.natureserve.org/explorer/) lists the Eastern Wolf as a subspecies of Gray Wolf (Canis lupus lycaon) and ranks it as G4G5TNR (apparently secure/secure) and N4 nationally. At the provincial scale, Québec has not assessed the Eastern Wolf, and Ontario has assigned a rank of S4 - apparently secure.
Mortality from human activity is the most important feature of habitat for Eastern Wolves. Within the current occupied areas in Ontario, the total habitat without harvest of Eastern Wolf includes Algonquin Park (7,571 km2) and surrounding townships (6,340 km2), Queen Elizabeth II Wildlands (255 km2), and Killarney Park (645 km2), for a total of 14,811 km2. Wolf harvest is permitted in Kawartha Highlands Park (376 km2) and French River Park (735 km2), but they would represent areas where human activity (e.g., buildings, roads, etc.) would be relatively limited.
In Québec, there are wildlife reserves and Zecs (zones d'expoitation contrôlées [controlled exploitation zones]) (e.g. Papineau-Labelle [1628 km2], Rouge-Matawin, Mastigouche, Saint Maurice, Port-Neuf, and Laurentides) that would contain less development than elsewhere and likely more prey, but these reserves generally are oriented towards extractive activities such as hunting and trapping. As such, their role as sites that would minimize Eastern Coyote gene introgression is uncertain. Harvest of Canids is not permitted in Mont Tremblant (1510 km2) and La Mauricie (536 km2) Parks.
Thank you to Jenny Wu at Environment Canada for creating the map figures for this report and to Dr. Graham Forbes for overseeing and providing editorial comments on the draft versions. Also, thank you to the following people and agencies that were contacted for information, input, and/or feedback for the status report.
Maria de Almeida, Wildlife Policy Section, Ontario Ministry of Natural Resources and Forestry, Peterborough ON.
Jeff Bowman. Ontario Ministry Natural Resources and Forestry, Peterborough, ON.
Robert Craig, Natural Heritage Information Centre Information Analyst, Ontario Ministry of Natural Resources, ON.
Marie-France Dalcourt, Acting Manager Population Conservation Section, Canadian Wildlife Service - Québec Region.
Dave Duncan, Regional Director, Canadian Wildlife Service, Prairie and Northern Regions, Environment Canada, MB.
Isabelle Gauthier, Ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec, Biologiste. Coordonnatrice provinciale, espèces fauniques menacées et vulnérables, QC.
Hélène Jolicoeur, Retired from Ministère des Ressources Naturelles et de la Faune (MFFP) du Québec.
Christopher Kyle, Assistant Professor, Trent University, Peterborough, ON.
Michel Henault, Ministère des Forêts, de la Faune et des Parcs, Direction de la gestion de la faune de Lanaudière et des Laurentides.
Briar Howes, Critical Habitat Biologist, Species at Risk, Parks Canada.
Hank Hristienko, Manitoba Conservation Wildlife and Ecosystem Protection Branch, MB.
Henry Lickers, Director of Environment, Mohawk Council of Akwesasne, Ontario/Quebec.
Angela McConnell, Canadian Wildlife Service, Ottawa, ON.
Patrick Nantel, Parks Canada, Conservation Biologist, Species at Risk Program, QC.
Dean Nernberg, Species at Risk Officer, National Defence.
Ronald Nowak, retired US Fish and Wildlife Service.
Michael Oldham, Natural Heritage Information Centre, Ontario Ministry of Natural Resources, ON.
Sylvain Paradis, Coordinator, Species at Risk, Parks Canada, QC.
Brent Patterson, Wildlife Research and Development Section, Ontario Ministry of Natural Resources and Forestry, ON.
Annie Paquet, Centre de données sur le patrimoine naturel du Québec. Ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec. Direction de l'expertise sur la faune et ses habitats.
Jason Pitre, ATK Coordinator, COSEWIC Secretariat, Canadian Wildlife Service.
Chris Risley, Bird and Mammal Species at Risk Specialist, Species at Risk Branch, Ontario Ministry of Natural Resources, ON.
Brad Steinberg, Management Biologist, Algonquin Park, ON.
Antoine St.-Louis, Ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec, Biologiste des espèces menacées.
Astrid vik Stronen, BIOCONSUS Fellow, Mammal Research Institute, Poland
Rick Stronks, Chief Park Naturalist, Algonquin Park, ON.
Richard Thiel, Retired Wildlife Biologist, Wisconsin Department of Natural Resources.
Mario Villemure, Resource Conservation Technician, La Mauricie National Park, QC.
Bradley White, Chair of Biology, Trent University, Peterborough, ON.
Paul Wilson, Canada Research Chair in DNA Profiling, Trent University, Peterborough, ON.
Adrian Wydeven, Mammalian Ecologist/Conservation Biologist, Wisconsin Department of Natural Resources.
Argue, A.M., Mills, K.J., and B.R. Patterson. 2008. Behavioural response of eastern wolves (Canis lycaon) to disturbance at homesites and its effects on pup survival. Canadian Journal of Zoology 86:400-406.
Bath, A.J. 2006. Human dimensions in wolf conservation: understanding trapper's perceptions toward wolves in and around La Mauricie National Park of Canada, Rapport présenté au Centre de services de Parcs Canada au Québec, 96 pp.
Benson, J.F., Patterson, B.R., and T.J. Wheeldon. 2012. Spatial genetic and morphologic structure of wolves and coyotes in relation to environmental heterogeneity in a Canis hybrid zone. Molecular Ecology 21:5934-5954.
Benson, J.F. and B. R. Patterson. 2013a. Inter-specific territoriality in a Canis hybrid zone: spatial segregation between wolves, coyotes, and hybrids. Oecologia. 173:1539-1550.
Benson, J. F., Mills, K.J., Loveless, K.M., and B.R. Patterson. 2013b. Genetic and environmental influences on pup mortality risk for wolves and coyotes within a Canis hybrid zone. Biological Conservation. 166:133-141.
Benson, J.F., Patterson, B.R., and P.J. Mahoney. 2014. A protected area influences genotype-specific survival and the structure of a Canis hybrid zone. Ecology 95:254-264.
Bertorelle, G., and L. Excoffier 1998. Inferring admixture proportions from molecular data. Molecular Biology and Evolution 15:1298-1311.
Beschta, R.L., and W.J. Ripple. 2009. Large predators and trophic cascades in terrestrial ecosystems of the western United States. Biological Conservation. 142:2401-2414.
Beschta, R.L., and W.J. Ripple. 2010. Recovering riparian plant communities with wolves in northern Yellowstone, USA. Restoration Ecology 18:380-389.
Brainerd, S.M., (and 6 co-authors). 2008. The effects of breeder loss on wolves. Journal of Wildlife Management 72:89-98.
Bruskotter, J.T., Schmidt, R.H., and T.L. Teel. 2007. Are attitudes toward wolves changing? A case study in Utah. Biological Conservation 139:211-218.
Bruskotter, J.T., Toman, E., Enzler, S.A., and R.H. Schmidt. 2011. Gray wolves not out of the woods yet. Science 327:30-31.
Buss, M., and M. deAlmeida. 1997. A review of wolf and coyote status and policy in Ontario. Ontario Ministry of Natural Resources. 88 pp.
Carbone, C., Mace, G.M., Roberts, S.C., and D.W. Macdonald. 1999. Energetic constraints on the diet of terrestrial carnivores. Nature 402:286-288.
CEC (Commission for Environmental Cooperation) 1997. Ecological regions of North America: a common perspective. Montreal, QC.
Chambers, S., S. Fain, B. Fazio, and M. Amaral. 2012. An account of the taxonomy of North American wolves from morphological and genetic analyses. North American Fauna 77:1-67. USFWS. Doi:10.3996/nafa.77.0001.
COSEWIC (Committee on the Status of Endangered Wildlife in Canada). 2014. Operations and Procedures Manual, April 2014. COSEWIC Secretariat. Website: COSEWIC Guidelines on Manipulated Populations [accessed February 2015].
Cronin, M.A, and L.D. Mech. 2009. Problems with the claim of ecotype and taxon status of the wolf in the Great Lakes region. Molecular Ecology 18:4991-4993.
Chapron, G., Andren, H., and O. Liberg. 2008. Conserving top predators in ecosystems. Science. 320: 47.
Crête, M., and S. Larivière. 2003. Estimating the costs of locomotion in snow for coyotes. Canadian Journal of Zoology 81:1808-1814.
DelGiudice, G.D., Riggs, M.R., Joly, P., and W. Pan. 2002. Winter severity, survival, and cause-specific mortality of female white-tailed deer in north-central Minnesota. Journal of Wildlife Management. 66:698-717.
Dowling, T., (and 4 co-authors). 1992. Response to Wayne, Nowak, and Phillips and Henry: Use of molecular characteristics in conservation biology. Conservation Biology 6:600-603.
Fain, S.R., Straughan, D.J., and B.F. Taylor. 2010. Genetic outcomes of wolf recovery in the western Great Lakes states. Conservation Genetics 11:1747-1765.
Fenech, A., Taylor, B., Hansell, R., and G. Whitelaw. 2000. Major road changes in southern Ontario 1935-1995: Implications for protected areas. Integrated Mapping Assessment Project. University of Toronto. Website: http://www.utoronto.ca/imap/papers/road_changes.htm [accessed January 9, 2011]
Fogelqvist J., Niittyvuopio, A., Ågren J., Savolainen O., and M. Lascoux. 2010. Cryptic population genetic structure: the number of inferred clusters depends on samples size. Molecular Ecology Resources 10:314-323.
Forbes, G.J., and J.B. Theberge. 1992. Importance of scavenging on moose by wolves in Algonquin Park, Ontario. Alces 28:235-241.
Forbes, G.J., and J.B. Theberge. 1995. Influences of a migratory deer herd on wolf movements and mortality in and around Algonquin Park, Ontario. Pp. 303-314 in Ecology and Conservation of Wolves in a Changing World. L. Carbyn, S. Fritts, and D. Seip (eds). Canadian Circumpolar Institute, University of Alberta, Edmonton, Alberta. 619 pp.
Forbes, G.J., and J.B. Theberge. 1996a. Cross-boundary management of Algonquin Park wolves. Conservation Biology 10:1091-1097.
Forbes, G.J., and J.B. Theberge. 1996b. Response of wolves to prey variation in central Ontario. Canadian Journal of Zoology 74:1511-1520.
Frame. P., D. Cluff and D. Hik. 2007. Response of wolves to experimental disturbance at homesites. Journal of Wildlife Management 71:313-320.
Fryxell, J.M. 2001. Habitat suitability and source-sink dynamics of beavers. Journal of Animal Ecology 70:310-316.
Fuller, T.K., Mech, D.L., and J.F. Cochrane. 2003. Wolf population dynamics. Pp. 161-191 in Wolves: Behavior, Ecology, and Conservation. D.L. Mech and L. Boitani (eds). University of Chicago Press, Chicago.
Fur Harvesters Auction Inc. 2011. Sale Results for January 7, 2011. (PDF; 43.7 KB).
Garcia-Moreno J., M. Matocq, M. Roy, E. Geffen E, and R. Wayne. 1996. Relationships and genetic purity of the endangered Mexican wolf based on analysis of microsatellite loci. Conservation Biology 10:376-389.
Goldman, E. 1937. The wolves of North America. Journal of Mammalogy 18:37-45.
Grewal, S.K., Wilson, P.J., Kung, T.K., Shami, K., Theberge, M.T., Theberge, J.B., and B.N. White. 2004. A genetic assessment of the eastern wolf (Canis lycaon) in Algonquin Provincial Park. Journal of Mammalogy 85:625-632.
Hailer F., and J.A. Leonard. 2008. Hybridisation among three native North American Canis species in a region of natural sympatry. PLoS ONE 3(10): e3333. doi:10.1371/journal.pone.0003333.
Harrison, D.J., and T.G. Chapin 1998. Extent and connectivity of habitat for wolves in eastern North America. Wildlife Society Bulletin 26:767-775.
Hedrick P., P. Miller, E. Geffen, and R. Wayne. 1997. Genetic evaluation of three captive Mexican wolf lineages. Zoological Biology 16:47-69.
Hénault, M. Unpublished data. Information contained in Québec jurisdictional review of Eastern Wolf Report. February 2015.
Hénault, M., and H. Jolicouer. 2003. Les loups au Québec: Meutes et mystères. Société de la faune et des Parcs du Québec, Direction de l'aménagement de la région des Laurentides, Direction du développement de la faune, 129 p.
Hubisz, M.J., Falush, D., Stephens, M., Pritchard, J.K. 2009. Inferring weak population structure with the assistance of sample group information. Molecular Ecology Resources 9:1322-1332.
Jolicoeur, H., and Hénault, M. 2002. Répartition géographique du loup et du coyote au sud du 52e parallèle et estimation de la population de loups au Québec. Société de la faune et des parcs du Québec. ISBN: 2-550-39485-2.
Jolicoeur, H., and M. Hénault. 2010. Current status and management of Wolves in Québec. Wolf Stewards Meeting, Dorset, Ontario.
Jost, L. 2008. Gst and its relatives do not measure differentiation. Molecular Ecology 17:4015-4026.
Kays, R., Curtis, A., and J.J. Kirchman. 2010. Rapid adaptive evolution of northeastern coyotes via hybridisation with wolves. Biology Letters 6:89-93.
Kennedy, L.J. 2012. pers. comm. Email correspondence to L Rutledge January 9, 2012. Senior Scientist, University of Manchester, Centre for Integrated Genomic Medical Research, Manchester, United Kingdom.
Koblmüller, S., Nord, M., Wayne, R.K., Leonard, J.A. 2009. Origin and status of the Great Lakes wolf. Molecular Ecology. 11:2313-2326.
Kolenosky, G., and R. Standfield. 1975. Morphological and ecological variation among Gray wolves (Canis lupus) of Ontario, Canada. Pp 62-72. in M. Fox (ed.). The Wild Canids. Van Nostrand Reinhold: New York.
Kurten, B., and E. Anderson, 1980. Pleistocene mammals of North America. Columbia Press, NY.
Kyle, C.J., Johnson, A.R., Patterson, B.R., Wilson, P.J., Shami, K., Grewal, S.K., and B.N. White. 2006. Genetic nature of eastern wolves: past, present and future. Conservation Genetics 7:273-287.
Kyle, C.J., Johnson, A.R., Patterson, B.R., Wilson, P.J., and B.N. White. 2008. The conspecific nature of eastern and red wolves: conservation and management implications. Conservation Genetics. 9:699-701.
Lancaster, P.A., Bowman, J., and B.A. Pond. 2008. Fishers, farms, and forests in eastern North America. Environmental Management 42:93-101.
Lankester, M.W. 2010. Understanding the impact of meningeal worm, Parelaphostrongylus tenuis, on moose populations. Alces 46:53-70.
Lehman, N., (and 6 co-authors). 1991. Introgression of coyote mitochondrial DNA genotype frequencies: estimation of the effective number of alleles. Evolution 45:104-119.
Leonard, J.A., and R.K. Wayne. 2005. Legacy lost: genetic variability and population size of extirpated US Gray wolves (Canis lupus). Molecular Ecology 14:9-17.
Leonard, J.A., and R.K. Wayne. 2008. Native Great Lakes wolves were not restored. Biology Letters 4:95-98.
Lesmarises, F., C. Dussault, and M-H. St-Laurent. 2012. Wolf habitat selection is shaped by human activities in a highly managed boreal forest. Forest Ecology and Management 276:125-131.
Lickers, H. Email correspondence with G. Goulet, TMSSC Member, COSEWIC. April 2015.
Linnell, J.D.C., Andersen, R., Andersone, Z. et al. 2002. The fear of wolves: A review of wolf attacks on humans. NINA Oppdragsmelding: 731: 1-65. PDF
Lohr, C., and W.B. Ballard. 1996. Historical occurrence of wolves, Canis lupus in the Maritime Provinces. Canadian Field-Naturalist 110:607-610.
Loveless, K. 2010. Foraging strategies of eastern wolves in relation to migratory prey and hybridisation. Masters thesis. Trent University, Peterborough, ON. 73 pp.
MacNulty, D.R., Smith, D.W., Mech, L.D., and L.E. Eberly. 2009. Body size and predatory performance in wolves: is bigger better? Journal of Animal Ecology 78:532-539.
Manning, R., and W. Sturtevant 1966. Early specimens of the eastern wolf, Canis lupus lycaon. Chesapeake Science 7:218-220.
McCarley, H., and J. Carley. 1979. Recent changes in distribution and status of red wolves (Canis rufus). Endangered Species Report no.4. U.S. Fish and Wildlife Service, Albuquerque, NM.
Mech, L.D., and L. Boitani. 2003. Wolf social ecology. Pp. 1-34. in D.L. Mech, and L. Boitani (eds.). Wolves: Behavior, Ecology and Conservation. University of Chicago Press, Chicago.
Mech, D., S. Fritts, and G. Radde. 1988. Wolf distribution and road density in Minnesota. Wildlife Society Bulletin 16:85-87.
Mech, L.D. 2010a. What is the taxonomic identity of Minnesota wolves? Canadian Journal of Zoology 88:129-138.
Mech, L.D. 2010b. Considerations for developing wolf harvest regulations in the contiguous United States. Journal of Wildlife Management 74:1421-1424.
Mech, L.D. 2011. Non-genetic data supporting genetic evidence for the eastern wolf. Northeastern Naturalist 18:521-526.
Mech, L.D., (and 4 co-authors). 2014. Production of hybrids between western gray wolves and western coyotes. PloSONE 9(2):e88861.doi:10.1371.
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2011.
Email correspondence to L Rutledge from Annie Paquet. May 13, 2011. Technicienne de la faune. Service de la biodiversité et des maladies de la faune, Québec.
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2012a.
Comments on the COSEWIC status report for the eastern wolf (Canis lycaon). E-mail correspondence to Graham Forbes, July 26, 2012.
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2012b. Web site: Portrait of the Territory [accessed December 8, 2011].
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2012c. Web site: Trapping statistics [accessed August 20, 2012].
Miller, G. 1912. The names of two North American wolves. Proceedings Biological Society of Washington. 25:93.
Mills, K.J. 2006. Wolf (Canis lycaon) pup survival, dispersal, and movements in Algonquin Provincial Park, Ontario. Masters thesis. Trent University, Peterborough, ON, 67 pp.
Mills, K.J., Patterson, B.R., and D.L. Murray. 2008. Direct estimation of early survival and movements in eastern wolf pups. Journal of Wildlife Management 72:949-954.
Murray, D.L., and L.P. Waits. 2008. Taxonomic status and conservation strategy of the endangered red wolf: a response to Kyle et al. (2006). Conservation Genetics 8:1483-1485.
Murray, D.L., (and 7 co-authors). 2006. Pathogens, nutritional deficiency, and climate influences on a declining moose population. Wildlife Monographs 166:1-20.
Musiani, M., L. Boitani, and P. Paquet. 2010. The World of Wolves: New Perspectives on Ecology, Behaviour and Management. University of Calgary Press. 398 pp.
Musiani, M., and P.C. Paquet. 2004. The practices of wolf persecution, protection, and restoration in Canada and the United States. BioScience 54:50-60.
Naughton, D. 2012. The Natural History of Canadian Mammals. University of Toronto Press, Toronto, ON. 784 pp.
NCEAS (National Center for Ecological Analysis and Synthesis). 2014. Review of the proposed rule regarding status of the wolf under the ESA. Santa Barbara, CA. 68 pp.
Norris, D.R., Theberge, M.T., and J.B. Theberge. 2002. Forest composition around wolf (Canis lupus) dens in eastern Algonquin Provincial Park, Ontario. Canadian Journal of Zoology 80:866-872.
Nowak, R.M. 1979. North American Quaternary Canis. Museum of Natural History, University of Kansas (Monograph No. 6), Lawrence, Kansas.
Nowak, R.M. 1992. The red wolf is not a hybrid. Conservation Biology 6:593-595.
Nowak, R.M. 1995. Another look at wolf taxonomy. Pp. 375-398, in L.N. Carbyn, S.H. Fritts and D.R. Seip (eds.). Ecology and Conservation of Wolves in a Changing World. Canadian Circumpolar Institute, Occasional Publication No. 35, 642 pp.
Nowak, R.M. 2002. The original status of wolves in eastern North America. Southeastern Naturalist 1(2):95-130.
Nowak, R.M. 2009. Taxonomy, morphology, and genetics of wolves in the Great Lakes region. in A.P. Wydeven, T.R. vanDeelen, and E. Heske (eds.), Recovery of Gray Wolves in the Great Lakes Region of the United States. Springer Publishing, 350 pp.
Nowak, R.M., pers. comm. 2011. Email correspondence to L. Rutledge. August, 2011. Wolf taxonomist, retired from US Fish and Wildlife Service.
OMNR (Ontario Ministry of Natural Resources). 2005. Strategy for Wolf Conservation in Ontario, Ontario Ministry of Natural Resources, report. http://www.mnr.gov.on.ca/stdprodconsume/groups/lr/@mnr/@fw/documents/document/263811.pdf. [accessed February, 2015].
OMNR (Ontario Ministry of Natural Resources). 2009. Cervid Ecological Framework. http://www.ontario.ca/environment-and-energy/cervid-ecological framework. [accessed February, 2015].
OMNR (Ontario Ministry of Natural Resources). 2010. Forest Management Guide for Conserving Biodiversity at the Stand and Site Scales. Queen's Printer, Toronto, ON.
OMNR (Ontario Ministry of Natural Resources). 2012. Wildlife Management Unit Maps. Find a wildlife management unit (WMU) map. [accessed February, 2015].
Ontario Government, Fish and Wildlife Conservation Act, 1997, S.O. 1997, c. 41.
Ontario Government, Endangered Species Act, 2007, S.O. 2007, c. 6,
Parcs Canada. 2007. Étude sur les attitudes, perceptions et connaissances des résidents de la région limitrophe du parc national du Canada de la Mauricie à l'égard du loup de l'Est. Rapport final, Mars 2007. Numéro de contrat: 5P201-060514 - POR-273-06.
Parker, G. 1995. Eastern Coyote: The Story of its Success. Nimbus Publishing, Halifax, NS. 254 pp.
Patterson, B.R., 2011a, pers. comm. Email correspondence to L Rutledge. October 14, 2011. Research Scientist, Ontario Ministry of Natural Resources, Trent University, Peterborough, Ontario.
Patterson, B.R., 2011b, pers. comm. Email correspondence to L Rutledge November 21, 2011. Research Scientist, Ontario Ministry of Natural Resources, Trent University, Peterborough, Ontario.
Patterson, B.R., 2011c, pers. comm. Email correspondence to L Rutledge December 18, 2011. Research Scientist, Ontario Ministry of Natural Resources, Trent University, Peterborough, Ontario.
Patterson, B.R., 2012, pers. comm. Email correspondence to L Rutledge in comment on draft. January 12, 2012. Research Scientist, Ontario Ministry of Natural Resources, Trent University, Peterborough, Ontario.
Patterson, B.R., 2012b, pers. comm. Email correspondence to L Rutledge. February 1, 2012. Research Scientist, Ontario Ministry of Natural Resources, Trent University, Peterborough, Ontario.
Patterson, B.R., 2014, pers. comm. Email correspondence to L Rutledge. July 2, 2014. Research Scientist, Ontario Ministry of Natural Resources, Trent University, Peterborough, Ontario.
Patterson, B.R., and D.L. Murray. 2008. Flawed population viability analysis can result in misleading population assessment: a case study for wolves in Algonquin Park, Canada. Biological Conservation 141:669-680.
Person, D., and A. Russell. Correlates of wolf mortality in an exploited wolf population. Journal of Wildlife Management 72:1540-1549.
Peterson, R.O., Thomas, N.J., Thurber, J.M., Vucetich, J.A., and T.A. Waite. 1998. Population limitation and the wolves of Isle Royale. Journal of Mammalogy 79:828-841.
Pilgrim KL, Boyd DK, Forbes SH (1998) Testing for wolf-coyote hybridization in the Rocky Mountains using mitochondrial DNA. Journal of Wildlife Management 62:683-689.
Pimlott, D.H., Shanon, J.A., and G.B. Kolenosky. 1969. The ecology of the timber wolf in Algonquin Provincial Park. Report No. 87. Ontario Ministry of Natural Resources, Toronto, Ontario, Canada.
Phillips, M., and V. Henry. 1992. Comments on red wolf taxonomy. Conservation Biology 6:596-599.
Pocock, R. 1935. The races of Canis lupus. Proceedings Zoological Society of London. Part 3; 647-686.
Potvin, F. 1988. Wolf movements and population dynamics in Papineau-Labelle Reserve, Québec. Canadian Journal of Zoology 66:1266-1273.
Potvin, F., H. Jolicoeur, L. Breton and R. Lemiuex. 1992a. Evaluation of an experimental wolf reduction and its impact on deer in Papineau-Labelle Reserve, Québec. Canadian Journal of Zoology 70:1595-1603.
Potvin, F., H. Jolicoeur, and J. Huot. 1988. Wolf diet and prey selectivity during two periods for deer in Québec: decline versus expansion. Canadian Journal of Zoology 66:1274-1279.
Potvin, F., L. Breton, C. Pilon, and M. Mactuart. 1992b. Impact of an experimental wolf reduction on beaver in Papineau-Labelle. Reserve, Québec. Canadian Journal of Zoology 70:180-183.
Quinn, N. 2004. The presettlement hardwood forests and wildlife of Algonquin Provincial Park: A synthesis of historic evidence and recent research. Forestry Chronicle 80:705-717.
Quinn, N. 2005. Reconstructing changes in abundance of White-tailed Deer, Odocoileus virgnianus, Moose, Alces alces, and Beaver, Castor Canadensis, in Algonquin Park, Ontario, 1860-2004. Canadian Field-Naturalist 119:330-342.
Reich, R. Wayne, and D. Goldstein. 1999. Genetic evidence for a recent origin by hybridization of red wolves. Molecular Ecology 8:139-144.
Rempel, R.S. 2011. Effects of climate change on moose populations: exploring the response horizon through biometric and systems models. Ecological Modelling 222:3355-3365.
Rogic, A., N. Tessier, and F.-J. Lapointe. 2014. Identification of canids found within Parc national du Mont-Tremblant and its surroundings using microsatellite markers. University of Montreal, Ministère des Forêts, de la Faune et des Parcs. 30 p.
Roy M., E. Geffen, D. Smith, E. Ostrander, and R. Wayne. 1994. Patterns of differentiation and hybridization in North American Wolf-like Canids, revealed by analysis of microsatellite loci. Molecular Biology and Evolution 11:553-570.
Roy M., E. Geffen, D. Smith, and R. Wayne. 1996. Molecular genetics of pre-1940 red wolves. Conservation Biology 10:1413-1424.
Rutledge, L.Y. 2010a. Evolutionary origins, social structure, and hybridisation of the eastern wolf (Canis lycaon). Ph.D. dissertation, Trent University, Peterborough, Ontario, Canada. 187 pp.
Rutledge, L.Y., Patterson B.R., and B.N. White. 2010b. Analysis of Canis mitochondrial DNA demonstrates high concordance between the control region and ATPase genes. BMC Evolutionary Biology 10: 215.
Rutledge, L.Y., Garroway, C.J., Loveless, K.M., and B.R. Patterson. 2010c. Genetic differentiation of eastern wolves in Algonquin Park despite bridging gene flow between coyotes and Gray wolves. Heredity 105:520-531.
Rutledge, L.Y., Bos, K.I., Pearce, R.J., White, B.N. 2010d. Genetic and morphometric analysis of sixteenth century Canis skull fragments: implications for historical eastern and Gray wolf distribution in North America. Conservation Genetics. 11:1273-1281.
Rutledge, L.Y., Patterson, B.R., Mills, K.J., Loveless, K.M., Murray, D.L., and B.N. White. 2010e. Protection from harvesting restores the natural social structure of eastern wolf packs. Biological Conservation 143:332-339.
Rutledge L. and B. White. 2013. Genetic characterization of Canis in Québec. Contrat de services abrégé 111720000. Final Report. 10 p.
Rutledge L. and B. White. 2014. Genetic characterization of Canis in Québec. Contrat de services abrégé 111720000. Final Report Addendum.14 p.
Rutledge, L.Y., White, B.N., Row, J.R., and B.R. Patterson. 2011. Intense harvesting of eastern wolves facilitated hybridisation with coyotes. Ecology and Evolution DOI: 10.1002/ece3.61.
Rutledge, L.Y., Wilson, P.J., Klütsch, C.F.C., Patterson, B.R., and B.N. White. 2012. Conservation genomics in perspective: a holistic approach to understanding Canis evolution in North America. Biological Conservation 155:186-192.
Rutledge, L.Y., Devillard S., Boone, J.Q., Hohenlohe, P.A., and B.N. White. In Review. RAD sequencing and genomic simulations resolve hybrid origins within North America Canis. Biology Letters.
Rutledge, L.Y., Desy, G., Fryxell J., Patterson, B.R., and B.N. White. Distribution of eastern wolves refined with non-invasive sampling. in prep. Canadian Journal of Zoology.
Samson, C. 2001. COSEWIC Assessment Update Status Report of the Eastern Wolf (Canis lupus lycaon) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 17 pp.
Sand, H., Wikenros, C., Wabakken, P., and O. Liberg. 2006. Effects of hunting group size, snow depth and age on the success of wolves hunting moose. Animal Behaviour 72:781-789.
Schmidt, K., Jędrzejewski, W., Theuerkauf, J., Kowalczyk, R., Okarma, H., and B. Jędrzejewski. 2008. Reproductive behaviour of wild-living wolves in Bialowieza Primeval Forest (Poland). Journal of Ethology 26:69-78.
Schmitz, O.J., and G.B. Kolenosky. 1985. Wolves and coyotes in Ontario - morphological relationships and origins. Canadian Journal of Zoology 63:1130-1137.
Schreber, J. 1775. Die Säughthiere in Abbildungen nach der Natur mit Beschreibungen. Sweeter Theil. Tafeln. Erlangen. (Walther). Vol. 3, 353, plate 89. Erlangen.
Schwartz, M.K., and J.A. Vucetich. 2009. Molecules and beyond: assessing the distinctness of the Great Lakes wolf. Molecular Biology 18:2307-2309.
Sears, H.J., Theberge, J.B., Theberge, M.T., Thornton, I., and G.D. Campbell. 2003. Landscape influence on Canis morphological and ecological variation in a coyote-wolf C. lupus x latrans hybrid zone, southeastern Ontario. Canadian Field-Naturalist 117:589-600.
St. Louis, Antoine, 2012, pers. comm. Email correspondence to L. Rutledge. August 6, 2012. Biologiste des espèces menacees.
Coordonnateur de l'équipe de rétablissement des oiseaux de proie du Québec (ÉROP). MFFP, Québec, QC.
Steinberg, B. 2011. E-mail correspondence to L. Rutledge. May, 2011. Management Biologist, Algonquin Park, Ontario.
Stahler, D.R., Smith, D.w., and D.S. Guernsey. 2006. Foraging and feeding ecology of the Gray wolf (Canis lupus): lessons from Yellowstone National Park, Wyoming, USA. Journal of Nutrition 26: 1923S.
Stronen, A.V., Brook, R.K., Paquet, P.C., Mclachlan, S. 2007. Farmer attitudes toward wolves: implications for the role of predators in managing disease. Biological Conservation 135:1-10.
Stronen, A.V. 2009. Dispersal in a Plain Landscape: Wolves in Southwestern Manitoba, Canada, PhD Dissertation. University of New Brunswick, Fredericton, New Brunswick, Canada.
Stronen, A.V., Forbes, G.J., Sallows, T., Goulet, G., Musiani, M., and P.C. Paquet. 2010. Wolf body mass, skull morphology, and mitochondrial DNA haplotypes in the Riding Mountain National Park region of Manitoba, Canada. Canadian Journal of Zoology 88:496-507.
Stronen, A.V., (and 9 co-authors). 2012. Canid hybridization: contemporary evolution in human-modified landscapes. Ecology and Evolution 2:2128-2140.
Tessier, N. 2015. Unpublished data. Ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec.
Theberge, J.B. 1991. Ecological classification, status and management of the Gray Wolf, Canis lupus, in Canada. The Canadian Field-Naturalist 105:459-463.
Theberge, J.B., Forbes, G.J., Barker, I.K., and T. Bollinger. 1994. Rabies in wolves of the Great-Lakes region. Journal of Wildlife Diseases 30:563-566.
Theberge, J., and M. Theberge. 2004. The Wolves of Algonquin Park: A 12-Year Ecological Study. Publication Series Number 56, Department of Geogrpahy, University of Waterloo, Waterloo, Canada.
Theberge, J.B., Theberge, M.T., Vucetich, J.A., and P.C. Paquet. 2006. Pitfalls of applying adaptive management to a wolf population in Algonquin Provincial Park, Ontario. Environmental Management 37:451-460.
Thompson, I.D., Fannigan, M.D., Wotton, B.M., and R. Suffling. 1998. The effects of climate change on landscape diversity: and example in Ontario Forests. Environmental Monitoring and Assessment 49:213-233.
Toews, D.P., and A. Brelsford. 2012. The biogeography of mitochondrial and nuclear discordance in animals. Molecular Ecology 16:3907-3930.
USFWS (United States Fish and Wildlife Service). 2015a. Endangered Species Act web page. [Accessed February 2015].
USFWS (United States Fish and Wildlife Service). 2015b. Red Wolf recovery web page. [Accessed February 2015].
USFWS, pers. comm. 2012. Email correspondence to L Rutledge May 21, 2012.
Vähä J-P, and C.R. Primmer. 2006. Efficiency of model-based Bayesian methods for detecting hybrid individuals under different hybridization scenarios and with different numbers of loci. Molecular Ecology 15:63-72.
Van Zyll de Jong, C.G., and L. Carbyn. 1999. COSEWIC Status Report on the Gray Wolf (Canis lupus) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 61 pp.
Veradri, A., V. Lucchini, and E. Randi. 2006. Detecting introgressive hybridization between free-ranging domestic dogs and wild wolves (Canis lupus) by admixture linkage disequilibrium analysis. Molecular Ecology 15:2845-2855.
Villemure, M. 2003. Écologie et conservation du loup dans la région du parc national de la Mauricie. Maître és sciences. Université de Sherbrooke, Sherbrooke, Québec, Canada. 92 pp.
Villemure, M., pers. comm. 2011. Email correspondence to L. Rutledge. August, 2011. Resource Conservation Technician, La Mauricie National Park, Parks Canada, St-Mathieu-du Parc, QC.
Villemure, M., and M. Festa-Bianchet. 2002. Écologie du loup au parc national de La Mauricie. Report prepared for Parks Canada. Université de Sherbrooke. Département de Biologie, Sherbrooke, Québec. 130p.
Villemure, M., and H. Jolicoeur. 2004. First confirmed occurrence of a wolf, Canis lupus, south of the St. Lawrence River in over 100 years. Canadian Field-Naturalist 118:608-610.
vonHoldt, B.M., Stahler, D.R., Smith, D.W., et al. 2008. The genealogy and genetic viability of reintroduced Yellowstone Gray wolves. Molecular Ecology 17:252-274.
vonHoldt, B.M., (and 18 co-authors). 2011. A genome-wide perspective on the evolutionary history of enigmatic wolf-like canids. Genome Research 21:1294-1305.
Vucetich, J., and P. Paquet. 2000. The demographic population viability of Algonquin wolves. Report prepared for The Algonquin Wolf Advisory Committee.
Way, J.G., Rutledge, L., Wheeldon, T., and B.N. White. 2010. Genetic characterization of eastern "coyotes" in eastern Massachusetts. Northeastern Naturalist.17:189-204.
Way, J.G., and J. T. Bruskotter. 2012. Additional considerations for gray wolf management after their removal from Endangered Species Act protections. Journal of Wildlife Management 76:457-461.
Wayne, R., and S. Jenks. 1991. Mitochondrial DNA analysis supports extensive hybridization of the endangered red wolf (Canis rufus). Nature 351:565-568.
Wayne, R., and N. Lehman. 1992. Mitochondrial DNA analysis of the Eastern Coyote: Origins and hybridization. Pp 9-22 in Ecology and Management of the Eastern Coyote. A. Boer (ed.). Wildlife Research Unit. Fredericton, NB. 194 pp.
Wheeldon, T. 2009. Genetic characterization of Canis populations in the western Great Lakes region. Masters thesis, Trent University, Peterborough, Ontario, Canada. 120 pp.
Wheeldon, T., and B.N. White. 2009. Genetic analysis of historical western Great Lakes region wolf samples reveals early Canis lupus/lycaon hybridisation. Biology Letters 5:101-104.
Wheeldon, T.J., Patterson, B.R., and White, B.N. 2010. Sympatric wolf and coyote populations of the western Great Lakes region are reproductively isolated. Molecular Ecology. 19:4428-4440.
Wheeldon, T.J., Rutledge, L.Y., Patterson, B.R., White, B.N., and Wilson, P.J. 2013. Y-chromosome evidence supports asymmetric dog introgression into eastern coyotes. Ecology and Evolution. 3:3005-3020.
Whittington, J., (and 6 co-authors). 2011. Caribou encounters with wolves increase near roads and trails: a time-to-event approach. Journal of Applied Ecology 48:1535-1542.
Williams, C.K., Ericsson, G., and T.A. Heberlein. 2002. A quantitative summary of attitudes toward wolves and their reintroduction (1972-2000). Wildlife Society Bulletin 30:575-584.
Wilmers, C.C., Post, E., Peterson, R.O., and J.A. Vucetich. 2006. Predator disease out-break modulates top-down, bottom-up and climatic effects on herbivore population dynamics. Ecology Letters 9:383-389.
Wilmers, C.C., Post, E., and A. Hastings. 2007. The anatomy of predator-prey dynamics in a changing climate. Journal of Animal Ecology 76:1037-1044.
Wilson, P.J., (and 14 co-authors). 2000. DNA profiles of the eastern Canadian wolf and the red wolf provide evidence for a common evolutionary history independent of the Gray wolf. Canadian Journal of Zoology 78:2156-2166.
Wilson, P.J., Grewal, S., McFadden, T., Chambers, R.C., and B.N. White. 2003. Mitochondrial DNA extracted from eastern North American wolves killed in the 1800s is not of Gray Wolf origin. Canadian Journal of Zoology 81:936-940.
Wilson, P.J., Grewal, S.K., Mallory, F.F., and B.N. White. 2009. Genetic characterization of hybrid wolves across Ontario. Journal of Heredity. 100: S80-89.
Wilson, P.J., Rutledge, L.Y., Wheeldon, T.J., Patterson, B.R., and B.N. White. 2012. Y-chromosome evidence supports widespread signatures of three-species Canis hybridisation in eastern North America. Ecology and Evolution. DOI: 10.1002/ece3.301.
Woodroffe R., Cleaveland, S., Courtenay, O., Laurenson, M.K., and M. Artois. 2004. Infectious Disease: Infectious Disease in the management and Conservation of Wild Canids. Pp. 123-142 in Biology and Conservation of Wild Canids (D.W. MadDonald and C. Sillero-Zubiri (eds.). Oxford University Press, Oxford, UK.
Wydeven, A.P., Fuller, T.K., Weber, W., and K. MacDonald. 1998. The potential for wolf recovery in the northeastern United States via dispersal from southeastern Canada. Wildlife Society Bulletin 26:776-784.
Wydeven, A.P., (and 6 co-authors). 2009. History, Population Growth, and Management of Wolves in Wisconsin. Pp. 87 - 105 in Recovery of Gray Wolves in the Great Lakes Region of the United States. A.P. Wydeven et al. (eds), Springer, DOI: 10.1007/978-0-387-85952-1.
Young S., and E. Goldman. 1944. The Wolves of North America. Dover Publications, New York, NY. 636 pp.
Linda Rutledge is a post-doctoral researcher and instructor in the Biology department at Trent University in Peterborough, Ontario. Her doctoral research focused on the evolutionary history, pack structure, and hybridization of Eastern Wolves in Algonquin Provincial Park. Dr. Rutledge's research interests focus on identifying evolutionary changes in species genomes that are indicative of behavioural responses, adaptation, hybridization, and speciation, especially as it applies to species at risk. She is also interested in how human influences impact the evolutionary trajectory of wildlife. Her current work involves utilizing comparative genomics of ancient and contemporary samples to identify genes responsible for ecological adaptations over space and time.
The collections examined are documented throughout the report. Raw data used for analysis was from the Wolf and Coyote DNA Bank at Trent University.
Much of the research on Canis hybridization was initially related to whether the Red Wolf was a hybrid because Red Wolves were being protected under the US Endangered Species Act (ESA), but hybrids were not eligible for protection (USFWS 2015a). Wayne and Jenks (1991) and Roy et al. (1994) used the absence of unique mtDNA restriction sites (that would have united Red Wolf into a single clade) as evidence that the Red Wolf was a hybrid. In a rebuttal, Dowling et al. (1992) noted that flawed methodology and interpretation by Wayne and Jenks (1991) limited any such conclusion. In later work, the estimated period of hybridization that created Red Wolf occurred in the last 2500 years, and possibly in the last 250 years (Reich et al. 1999). vonHoldt et al. (2011) concluded that Red Wolf were hybrids, but it was noted by Rutledge et al. (2012) and Wilson et al. (2012) that von Holdt et al. (2012) used samples (n=12) from animals that existed after the recent Coyote introgression, which would be biased against identifying previously 'pure' Red Wolf. Although the debate continues, the Red Wolf remains on the US Endangered Species list.
The genetic studies on the Eastern Wolf were part of a Great Lakes region assessment of Canis, beginning in the 1990s (Lehman et al. 1991; Roy et al. 1994; Koblmüller et al. 2009). For example, using mtDNA restriction site polymorphisms on 226 samples collected from Manitoba to Maine, combined with 538 Canis samples from elsewhere in North America, Wayne and Lehman (1992)concluded that the Eastern Coyote was a hybrid, but also that all Canis in the Great Lakes region were hybrids. In 2009, Koblmüller et al. used maternal, paternal, and biparentally inherited markers to conclude that the large Canis in the Great Lakes region (Great Lakes Wolf) was a unique population or ecotype of Gray Wolf that experienced a high degree of introgression from Coyote and western Gray Wolf in ancient, and recent, times. vonHoldt et al. (2011) analyzed 48,000 single-nucleotide-polymorphisms (SNP) in Canis samples collected worldwide and concluded that Red Wolf and Great Lakes Wolf were not similar, and that Great Lakes Wolf was a hybrid with large amounts of Gray Wolf genome.
A criticism of earlier Canis taxonomic research was a reliance (by all researchers) on a single metric (mtDNA). Multiple metrics, such as the Y-chromosome, are now commonly used (e.g., Wilson et al. 2012). Another issue has been the low number of markers typically used (e.g., 12; Wilson et al. 2000, Rutledge et al. 2010b). The large number (48,000) used by vonHoldt et al. (2011) is cited as an improvement over earlier work (R. Wayne, in NCEAS 2014).
In summary, the 2-species hypothesis proposes that only two species (Coyote and Gray Wolf) exist and that hybrid events in recent and historical times have produced the array of hybrid Canis.
The contention that Eastern Wolf and Red Wolf are valid species is derived from a hypothesis that three large Canis were present in North America. Based on sequence divergence of haplotypes Wilson et al. (2000) proposed that the ancestral progenitor of the Gray Wolf, Eastern Wolf/Red Wolf, and Coyote resided in North America. Approximately 1 - 2 million years ago, the Gray Wolf diverged from the ancestor and emigrated to Eurasia. The Coyote and Eastern Wolf/Red Wolf then diverged from each other between 150,000 to 300,000 years ago (Wilson et al. 2000), and the Gray Wolf returned to North America from Eurasia around 300,000 years ago (Kurten and Anderson 1980). Coyote were restricted to areas west of the Mississippi River, then moved northward and eastward in association with settlement and land-use practices by Europeans, and the extirpation of wolves in eastern North America. The reason there is an Eastern Coyote hybrid between the Eastern Wolf and 'western' Coyote is because they are closely related. The Great Lakes Wolf (or Great Lakes Boreal Wolf) is considered a hybrid between Eastern Wolf and Gray Wolf (Wilson et al. 2000, 2009; Wheeldon 2009).
The results of the above work was the basis for identifying the Eastern Wolf by COSEWIC in 2001 (Samson 2001). At present, the Eastern Wolf is listed as a subspecies of Gray Wolf (Canis lupus lycaon). However, genetic analyses in numerous projects noted the absence of enough C. lupus alleles and haplotypes in the Eastern Wolf/Red Wolf to consider Eastern Wolf as a subspecies of Gray Wolf (Lehman et al. 1991; Roy et al. 1994, 1996; Wilson et al. 2000, 2003; Grewal et al. 2004; vonHoldt et al. 2011). As well, mitochondrial DNA haplotypes not found in extant Gray Wolf populations were found in Eastern Wolf samples from the 1800s, a period before Coyote arrived (Wilson et al. 2003).
The initial basis for the 3-species hypothesis was work on 8 microsatellite loci and mtDNA control-region sequences from 68 samples collected in Algonquin Park, compared to Red Wolf (n=67), Texas Coyotes (n=24), and Northwest Territories Gray Wolf (n=67) (Wilson et al. 2000).Canis from Algonquin were differentiated from Gray Wolf, and Red Wolf was similar to Algonquin samples.Unlike the Great Lakes Wolf, mtDNA from Gray Wolf was not found in the central Ontario samples, as would be expected if these animals were hybrids of Gray Wolf x Coyote. Wilson et al. (2009) later analyzed mtDNA of 269 samples from Ontario (109 from Algonquin Park and 186 total from Eastern Wolf range), plus 393 Canis samples from elsewhere in North America, to conclude that a distinct Canis exists in central Ontario. Only 4 of 92 samples from Algonquin Park contained Gray Wolf haplotypes. Further work using Y-chromosome haplotypes in Eastern Wolf noted that these haplotypes are absent from western Coyote, which is evidence of a wolf evolving in eastern North America, consistent with the 3-species hypothesis (Wilson et al. 2012).
Other studies similarly found evidence that a Canis other than Gray Wolf was historically present in eastern North America (Bertorelle and Excoffier 1998). Eastern Wolf mtDNA has been found in 100-year-old samples from Wisconsin and Michigan (Leonard and Wayne 2008), and Wheeldon and White (2009) found Eastern Wolf mtDNA and admixed Gray Wolf - Eastern Wolf nuclear signature in 100-year-old wolf samples from Minnesota and Wisconsin. Genetic evidence suggests that Eastern Wolves may have been present in Maine and New York 100 years ago, prior to the eastward expansion of Coyotes (Wilson et al. 2003). Both genetic and morphometric analyses of samples excavated from a prehistoric (c.1530) Iroquois village in London, Ontario suggest that Eastern Wolves occurred in the Carolinian forests of southern Ontario prior to the arrival of European settlers (Rutledge et al. 2010d). A small Canis was present in the Pleistocene fossil record of eastern North America (Nowak 1995).
In a rebuttal to vonHoldt et al. (2011), Rutledge et al. (2012) noted that the two samples from Algonquin Park were insufficient to differentiate Eastern Wolf. However, when a phylogenetic re-analysis of the 48K SNP data in von Holdt et al. (2011) was constructed with the two samples, but then combined with additional local samples, the two Eastern Wolves from Algonquin Park now clustered independently of other Canis groups and were located basal to Red Wolves and Coyotes within a North American lineage (Rutledge et al. 2012), indicating ancestry with Coyote, and as expected in a 3-species hypothesis. As well, the basal pattern was similar to that seen with both mtDNA (Rutledge et al. 2010b) and the Y-chromosome (Wilson et al. 2012). These results provide maternal, paternal, and bi-parental genetic evidence for the Eastern Wolf as a species. Most recently, an analysis of 127K genome-wide SNPs demonstrated through hybrid simulations that the Eastern Wolf is a distinct species, and inclusion of it as a third species resolves the hybrid origins of Great Lakes-Boreal Wolves and Eastern Coyotes (Rutledge et al. in revision).
As well, Rutledge et al. (2012) noted that vonHoldt et al. (2011) used a method that assumes a 2-species model which only allows ancestry to be divided between Gray Wolves and Coyotes, a design which could not delineate Eastern Wolf. As well, Rutledge et al. 2012) contends that there is a biased assumption that the SNP panel ascertained from the domestic Dog (Canis lupus familiaris) genome captures the true variability in wild Canis species.
Although the debate on Canis taxonomy continues, the evidence below is considered compelling enough to consider the Eastern Wolf as a valid taxonomic entity:
The Eastern Wolves identified by genetic analyses are almost all found in protected areas, even though sampling was conducted over a wide range of park, and non-park, areas (see Search Effort; Figure 4 and Figure 6). The mechanism to create and maintain unique species among closely related Canis types is unknown but there is evidence that introgression of Coyote/Eastern Coyote genes is facilitated by a change in pack dynamics. It is hypothesized that excessive mortality of breeding animals alters the socially based breeding structure of the pack (Rutledge et al. 2010c, 2011; Benson et al. 2013a, 2014). Typically, only the dominant (alpha) pair breed, and breeder loss can lead to dissolution of pack cohesion and changes in mating patterns (Brainerd et al. 2008). Despite introgression in some individuals (Rutledge et al. 2010c, Grewal et al. 2004), the Eastern Wolves in Algonquin Park have retained a genetically distinct signature for at least 50 years (Rutledge et al. 2011). Wolves in this >7000km2 park have been protected from hunting and trapping for much of this time. During the 1980s and 1990s, many Park wolves were killed when they followed Deer out of the east side of the Park (Forbes and Theberge 1996b). Those high mortality rates have been implicated in the introgression of Coyote mtDNA, and appearance of Eastern Coyote nuclear DNA during this period, because the Eastern Wolf nuclear signature was restored after a hunting and trapping ban in areas adjacent to the Park was implemented in 2001 (Rutledge et al. 2011). Similarly, Eastern Wolves dispersing from western regions of the Park are highly vulnerable to harvest mortality (Benson et al. 2014), despite some residency noted in the unprotected areas of Wildlife Management Unit (WMU) 47 (n=3) and WMU49 (n=1) (Benson et al. 2012). Ongoing hybridization with Eastern Coyotes has been curtailed by protection that presumably allows for conspecific mates to be more readily found (Rutledge et al. 2010c, 2011).
Despite evidence of gene flow between Canis types in general, there is substantial genetic and morphological structuring within Eastern Wolves. Ecological differences in habitat and prey base, as well as conspecific mate choice, may drive the divergence (Rutledge et al. 2010c). The levels of heterozygosity, based on autosomal microsatellite markers, are generally high (Ho = 0.645; SE = 0.04, Rutledge et al. 2010c) and although traditional measures of genetic differentiation such as Fst are relatively low (Fst between Eastern Wolves and Great Lakes-Boreal Wolves = 0.105; and between Eastern Wolves and Eastern Coyotes = 0.052), the differences are statistically significant (p<0.001). Also, new measures based on allele sharing (Jost 2008) suggest higher levels of differentiation exist between Eastern Wolves and Great Lakes-Boreal Wolves (Jost D = 0.207) than between Eastern Wolves and Eastern Coyotes (Jost D = 0.090) (Rutledge et al. 2010c). Bayesian and multivariate clustering analyses based on autosomal microsatellites reveal substantial differentiation of Eastern Wolves in Algonquin Park from Canis types in neighbouring regions, despite the lack of any physical barrier (Rutledge et al. 2010c; Benson et al. 2012). Eastern Wolf packs are spatially segregated from each other and from other Canis types (Benson and Patterson 2013a).
The unique genetic signature may also be related to assortative mating and/or selection against nuclear introgression from Eastern Coyotes (Rutledge et al. 2010c) because Eastern Wolves are ecologically differentiated based on habitat and the energetic requirements associated with larger size of prey (Benson et al. 2012; Rutledge et al. 2010c). Loveless (2010) suggested that the restoration of a natural social structure in Algonquin Park (Rutledge et al. 2010e) may be responsible for the improved efficiency at Moose predation via learning passed down through family members. The unique genetic signature is unlikely to be due to genetic drift because genetic drift is most commonly associated with isolated populations, but Algonquin Park is not isolated. Gene flow could occur across the region but the Canis genotype in Algonquin Park has been maintained at least since the 1960s, likely due to assortative mating (Rutledge et al. 2011). Also, genetic diversity is relatively high (observed heterozygosity = 0.645; Rutledge et al. 2010c) and there is no evidence of inbreeding Rutledge et al. 2010e)
Under the 3-species hypothesis, it is possible that the Eastern Wolf and the Red Wolf are the same species. Evidence from autosomal microsatellites suggests overlap in their genetic signature (Wilson et al. 2000; Kyle et al. 2006, 2008; Murray and Waits 2008; Rutledge et al. 2012), and phylogenetic analysis of 48K SNP data suggests Algonquin Wolves are basal to Red Wolves within a North American lineage (Rutledge et al. 2012). The range likely would have included eastern regions of the United States and southern regions of Ontario and Québec, a range largely consistent with the eastern temperate forests and the historical distribution of Deer (Nowak 1995, 2002, Figure 3).
The extant Red Wolf population has been produced by a captive breeding program that originated with 14 animals captured in the Texas-Louisiana region during the 1970s. They were identified as Red Wolf based on morphometrics and vocalizations (McCarley and Carley 1979), and some of their descendants were released into North Carolina, and are now reproducing (USFWS 2015b). However, the original breeders contained Coyote (vonHoldt et al. 2011), Dog and/or Gray Wolf genetic material (Hailer and Leonard 2008; Wilson et al. 2012) and despite controlled breeding management, there is concern that the existing Red Wolf sample is not a full representation of the Red Wolf genome. As such, the ability to use extant genetic samples to assess whether they were the same species is compromised.
The taxonomic issue has affected Endangered Species listing in the United States. In May 2011, the United States Fish and Wildlife Service (USFWS) formally recognized the Eastern Wolf as a distinct species (C. lycaon), but retracted the statement in December, 2011. A review of Canis species, subspecies, and hybrid taxonomy in North America was then produced (Chambers et al. 2012), which formed the basis for a proposed ruling on listing by the USFWS. Based on the findings of vonHoldt et al. (2011), the USFWS undertook an expert panel exercise (NCEAS 2014) to discuss the Chambers et al. (2012) review. Of note, the Chambers et al. (2012) review was not a review of Eastern Wolf (as defined in this COSEWIC report), because Chambers et al. (2012) combined all Canis in the Québec-Great Lakes region into one 'Eastern Wolf (Canis lycaon)', whereas it is the contention of this report that Eastern Wolf only refers to the Canis in central Ontario, and southern Québec (see Appendix B; Figure 4). The conclusion of the NCEAS (2014) report was that there was no consensus on Canis taxonomy, with proponents of the '2-species' or '3-species' hypotheses 'agreeing to disagree' (NCEAS 2014). The report does not conclude that Eastern Wolf is an invalid species but only that there is a lack of consensus on Canis taxonomy, and that Chambers et al. (2012) did not reflect this perspective well enough, and therefore the proposed ruling is not based on the 'best available science'.
Notwithstanding the debate, there is consensus that extant populations of both the Red Wolf and Eastern Wolf have some degree of Coyote genome. As well, it is agreed that the Eastern Coyote is a hybrid, which now exists across eastern North America. These Canis have established through natural range expansion and interaction. The issue of hybridization events, especially in human-modified environments, raises questions on the limitations of the species concept and taxonomic nomenclature as a basis for managing species that are not 'pure' (Wilson et al. 2012). The controversy over including possible hybrids such as the Red Wolf in Endangered Species protection resulted in numerous arguments for identifying species based on ecological and behavioural characteristics, and not simply genetic ones (e.g., Theberge 1991; Nowak 1992; Phillips and Henry 1992). Dowling et al. (1992) noted that the intent of the ESA was to protect distinctive forms, regardless of how well they fit into the species concept. The US Endangered Species Act recognizes evolutionary significant units, and implementation of the Canadian Species at Risk Act recognizes a broader definition of Wildlife Species, both of which facilitate protection of distinct and significant entities below the species level.
Individual animals need to be assigned to the population of Eastern Wolf in Canada because the range of Eastern Wolf contains Canis of mixed genotype. The assignment of individuals as Eastern Wolves is based on an analysis of bi-parentally inherited nuclear autosomal microsatellite genetic markers because of: a) the introgression of Eastern Wolf mtDNA and Y-chromosome haplotypes into Great Lakes-Boreal Wolves (Rutledge et al. 2010c) and Eastern Coyotes (Way et al. 2010) across their ranges; b) the mtDNA-nuclear DNA discordance known to occur throughout animal systems, including the Canis genus (Toews and Brelsford 2012); and c) the potential for incomplete lineage sorting of mtDNA between Eastern Wolves and Coyotes (Wheeldon and White 2009).
The use of Q<0.8 as a threshold is based on basic patterns of inheritance, support from literature, analyses, and some assumptions. The basic patterns of inheritance refers to any F1 hybrid possessing a Q=0.5, and, if backcrossed with a parent, becomes Q=0.75. A Q>0.8 avoids capturing such events. The Q value threshold concept has been validated elsewhere (e.g., Vähä and Primmer 2006) as well as with hybrid simulation analysis that indicated a strong demarcation of Eastern Wolf within, and outside, Algonquin Park, a result believed to be related to effects of Eastern Wolf harvest and rates of Coyote introgression (see Appendix A - 'Mechanism'). Benson et al. (2012) tested the validity of Q<0.8 assignments using PCA methods and found a 90% compliance between assignments from STRUCTURE, and PCA. The Q value approach is a common approach for identifying Canis types (e.g., Veradri et al. 2006; Benson et al. 2012; Wheeldon et al. 2013).
The Q value threshold approach requires an assumption of what a 'pure' Eastern Wolf genotype would be. However, we lack enough specimens that have been collected before Coyotes were present to characterize a pure Eastern Wolf. The use of Q≥0.8 as a threshold for identifying Eastern Wolf determines the number of animals and sites containing what is considered an Eastern Wolf. Although Q≥0.8 has value as a threshold, a map of animals with Q=0.75 could identify more animals, and consequently, the extent to which Eastern Wolf are restricted to protected areas, as appears to be the case. Production of a map was not possible because sites on the 457 samples analyzed in Québec (Rutledge and White 2014) were only available for the 11 specimens with Q≥0.8. In the Algonquin Park area, if the Q value was changed to either 0.75 and/or 0.85, some sampled regions vary by > 10% in the number of animals assigned to one category of Canis, or another (C. Kyle, unpubl. data). Further, in some regions sampled that have no Q>0.8 Eastern Wolf, there are Q>0.75 animals. It is possible then that the population size of Eastern Wolf varies, but only a small amount from the estimate based on Q≥0.8 (Table 2). The range (extent of occurrence) likely would not change because it already encompasses a large area with few records. In summary, the use of Q≥0.8 has the most support, and appears to capture the distribution and population size relatively well.
| Threat Impact | Threat Impact (descriptions) | Level 1 Threat Impact Counts: high range |
Level 1 Threat Impact Counts: low range |
|---|---|---|---|
| A | Very High | 0 | 0 |
| B | High | 1 | 0 |
| C | Medium | 2 | 2 |
| D | Low | 1 | 2 |
| - | Calculated Overall Threat Impact: | Very High | High |
| # | Threat | Impact (calculated) Criterion |
Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|---|
| 1 | Residential and commercial development | D | Low | Small (1-10%) | Extreme (71-100%) | High (Continuing) | - |
| 1.1 | Housing & urban areas | D | Low | Small (1-10%) | Extreme (71-100%) | High (Continuing) | Based on the species extent of occurrence from Figure 4 of the report. Scope: all in agreement that new development would be less than 10% Severity: based on the population to be affected by the new housing and urban areas (not just counting mortality) * impact is extremely high when we consider new urban development, however, wolves can use cottages/housing area that remain relatively rural * ON has no Eastern Wolf present in urban areas * QC is expecting more cottage development Note: May need to revisit based on QC uncertainty; QC are considering only mortality based on new development (thus negligible) as wolves would leave the area. |
| 1.2 | Commercial & industrial areas | - | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | Scope * QC suggest small (1-10%) but can agree with negligible * there is a possibilty of new shopping centres in ON, but doesn't believe it will push scope above 1% |
| 1.3 | Tourism & recreation areas | D | Low | Small (1-10%) | Serious - Moderate (11-70%) | High (Continuing) | Scope * QC - small (1-10%) * ON is saturated with golf courses and probably won't support anymore in the next 10 years; small (1-10%) based on other recreation (ex. Summer camps, cottage country recreation etc.) |
| 2 | Agriculture and aquaculture | - | Negligible | Negligible (<1%) | Serious - Slight (1-70%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 2.1 | Annual & perennial non-timber crops | - | Negligible | Negligible (<1%) | Serious - Slight (1-70%) | Moderate (Possibly in the short term, < 10 yrs) | Scope: ON/QC agreement of negligible Severity: * most of the wolves are within protected area, however, with limited distribution data when wolves move between the parks, a range is used to indicate uncertainty * ON forest covers are increasing and agriculture extent is going down and again, without knowing where wolves move outside the park, there is a lot of uncertainty |
| 2.2 | Wood & pulp plantations | - | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | * ON: not an issue * QC: there will be some threat |
| 2.3 | Livestock farming & ranching | - | Negligible | Negligible (<1%) | Unknown | High (Continuing) | Severity * ON: livestock might be increasing * QC: between Algonquin and parks in the eastern range, there is a lot of uncertainty |
| 3 | Energy production and mining | - | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | - |
| 3.1 | Oil & gas drilling | - | Negligible | Negligible (<1%) | Negligible (<1%) | Insignificant/Negligible (Past or no direct effect) | * ON: not aware of any issue * QC: believes there will be a little bit |
| 3.2 | Mining & quarrying | - | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | Scope * ON: could be a bigger iimpact but on the low end of small (1-10%) but based on Figure 4a, most occur in QC and will agree with negligible (<1%) |
| 3.3 | Renewable energy | - | - | - | - | - | - |
| 4 | Transportation and service corridors | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | - |
| 4.1 | Roads & railroads | CD | Medium - Low | Pervasive (71-100%) | Moderate - Slight (1-30%) | High (Continuing) | Scope: pervasive (71-100%) because this includes all roads (including logging/mining) Severity: range used to denote uncertainty * ON: road kill certainly happens but not a major cause of decline in population level. There have been a few kills recently (ex. Hwy 116) but it's a small number * QC: there are few data but numbers could be higher. One recent study found 6 wolves killed out of 60; although uncertain if wolves killed were eastern or a hybrid. |
| 4.2 | Utility & service lines | - | Negligible | Small (1-10%) | Negligible (<1%) | High (Continuing) | Scope: only looking at new service and utility lines * ON: no new development that would affect population; < 1% * QC: close to 1% but not aware in the coming years if more area would be impacted |
| 5 | Biological resource use | BC | High - Medium | Large (31-70%) | Serious - Moderate (11-70%) | High (Continuing) | - |
| 5.1 | Hunting & collecting terrestrial animals | BC | High - Medium | Large (31-70%) | Serious - Moderate (11-70%) | High (Continuing) | Scope: takes into account both legal and illegal hunting and bycatch. A large proportion of the population is found within Algonquin Park, pervasive (71-100%) would be too high * ON: once wolves leaves the protected area, they fair very poorly. On the eastern end, there is no poaching/harvesting allowed Severity: range used to denote uncertainty * QC: depends on the different group and the proportion that move outside the protected area; may not be high in some places |
| 5.2 | Gathering terrestrial plants | - | - | - | - | - | Not an issue |
| 5.3 | Logging & wood harvesting | - | - | - | - | - | Not an issue. * ON: could be a benefit over the 10 year period |
| 5.4 | Fishing & harvesting aquatic resources | - | - | - | - | - | Not an issue |
| 6 | Human intrusions and disturbance | - | Negligible | Pervasive (71-100%) | Negligible (<1%) | High (Continuing) | - |
| 6.1 | Recreational activities | - | Negligible | Pervasive (71-100%) | Negligible (<1%) | High (Continuing) | Scope * QC: snowmobiling activity is very high * ON: high if we include activities like canoeing and other cottage recreation |
| 6.2 | War, civil unrest & military exercises | - | - | - | - | - | Not an issue |
| 6.3 | Work & other activities | - | - | - | - | - | Not an issue |
| 7 | Natural system modifications | - | Negligible | Restricted - Small (1-30%) | Negligible (<1%) | High (Continuing) | - |
| 7.1 | Fire & fire suppression | - | Negligible | Restricted - Small (1-30%) | Negligible (<1%) | High (Continuing) | - |
| 7.2 | Dams & water management/use | - | Negligible | Negligible (<1%) | Extreme (71-100%) | High (Continuing) | - |
| 8 | Invasive and other problematic species and genes | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | - |
| 8.1 | Invasive non-native/alien species | - | Unknown | Pervasive (71-100%) | Unknown | High (Continuing) | Scope * ON would be pervasive (71-100%). Note: numbers based on studies in 2007 (which was a bad year); probably 11% died. * QC: has no data Severity: Unknown * ON can provide precise numbers on mortality rate but don't have a better guess for severity Note: coyote genetic issue dealt with in 8.3 |
| 8.2 | Problematic native species | - | Unknown | Large (31-70%) | Unknown | High (Continuing) | Scope: what % of Eastern Wolf experience problematic native species? * ON: all the ones outside of Algonquin Park Note: This is a limiting factor rather than a threat; discussed in report as limiting factor |
| 8.3 | Introduced genetic material | C | Medium | Pervasive (71-100%) | Moderate (11-30%) | High (Continuing) | Severity * ON: potentially any wolves that leave Agonquin Park would have higher than 30% of dog/coyote gene but can agree with the 11-30% range for all population |
| 9 | Pollution | - | - | - | - | - | - |
| 9.4 | Garbage & solid waste | - | Negligible | Small (1-10%) | Negligible (<1%) | High (Continuing) | Not counting towards overall calculator * score based QC, but this could actually have a positive effect * ON: landfill/garbage does have an impact on wolves (both postively and negatively) |
| 9.5 | Air-borne pollutants | - | - | - | - | - | - |
| 9.6 | Excess energy | - | - | - | - | - | - |
| 10 | Geological events | - | - | - | - | - | Not an issue |
| 11 | Climate change and severe weather | - | - | - | - | - | Unknown. Changes to climate haven't been long enough to see changes. Long term moose density (and deer) could change and affect the wolf population. |
| 11.1 | Habitat shifting & alteration | - | - | - | - | - | Unknown |