
Évaluation et Rapport de situation du COSEPAC sur le Loup de l'Est Canis sp. cf. lycaon au Canada - 2015

Photo : © Michael Runtz, 2015
- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique
- Préface
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances de la population
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique de la rédactrices du rapport
- Collections examinées
- Figure 1. Photographie d'un loup de l'Est du parc provincial Algonquin. © Michael Runtz, reproduit avec sa permission
- Figure 2. Répartitions des formes des espèces de Canis en Ontario : a) proposées par Kolenosky et Standfield (1975) d'après la morphologie du crâne et le poids corporel; b) proposées par Rutledge (2010) d'après des analyses génétiques et le poids corporel. Kolenosky et Standfield (1975) ont mal défini les limites de la répartition du C. l. lycaon forme Tweed, mais ils en ont défini une zone centrale indiquée par les triangles, le plus grand des triangles une fréquence d'occurrence plus élevée. Ils ont également estimé que les zones agricoles étaient habitées par le coyote (C. latrans), présumément le coyote de l'ouest. Très peu de C. l. hudsonicus étaient disponibles pour les deux études, mais dans les deux cas il semblait s'agir d'une sous espèce du loup gris vivant dans les régions côtières de la baie d'Hudson et de la baie James. La région pointillée chevauchant la partie centrale de l'aire de répartition du coyote de l'Est en b) indique la zone approximative où l'on a documenté la présence de loups de l'Est du parc Algonquin, bien que certains se dispersent vers l'est au Québec. La limite plus au nord de l'aire de répartition du coyote de l'Est indiquée en b) est fondée sur la présence confirmée de coyotes de l'Est à Nakina (Ontario). L'aire de répartition du loup boréal/des Grands Lacs s'étend au Manitoba, au Québec et dans les États du Minnesota, du Wisconsin et du Michigan, dans la région ouest des Grands Lacs (Rutledge, 2010a,b)
- Figure 3. Aire de répartition mondiale historique du loup de l'Est (zone en gris) selon a) le scénario 1 d'après Nowak (1995) et Leonard et al. (2005) et b) le scénario 2 d'après Wilson et al. (2000) et Rutledge et al. (2010d). La délimitation des forêts tempérées de l'est est tirée de CEC (1997)
- Figure 4. Zone d'occurrence du loup de l'Est et localités des individus identifiés par une méthode commune à diverses études publiées [Rutledge et al., 2010a, 2010b, 2010c; Rutledge et White, 2013, 2014]) et indiqués par les étoiles rouges et des individus identifiés par Rogic et al. (2014), Hénault (données inédites) et Tessier (données inédites) et indiqués par des cercles. Le nombre dans chaque cercle indique le nombre d'individus, et la position du cercle indique leur emplacement approximatif. Les aires protégées et les réserves fauniques sont indiquées par l'ombrage foncé, et les zones d'exploitation contrôlée (ZEC) par l'ombrage plus pâle
- Figure 5. Localisation des échantillons utilisés pour déterminer la répartition du loup de l'Est au Canada (voir Activités de recherche pour les détails concernant les échantillons)
- Figure 6. Localisation des aires protégées et des principales routes asphaltées (indiquées par les lignes noires) dans l'aire de répartition du loup de l'Est. Les aires protégées varient selon leur degré de protection contre la chasse et le piégeage. La récolte de loups est interdite dans les parcs Algonquin, Queen Elizabeth II, du Mont Tremblant et de la Mauricie et permise dans les réserves fauniques et les zones d'exploitation contrôlées (ZEC)
- Tableau 1. Effort d'échantillonnage et nombre de loups de l'Est identifiés. L'occurrence de l'espèce est fondée sur l'assignation d'un Q ≥ 0,8 dans le programme structure pour le génotypage de 12 microsatellites chez les individus. Les sources des données sont présentées sous le tableau
- Tableau 2. Estimations de l'abondance du loup de l'Est. Le nombre maximal d'individus aux sites de l'Ontario (Ontario) est fondé sur une estimation de la densité égale à 3 loups par 100 km2 observée dans le parc Algonquin (Rutledge et al., 2010e); le nombre estimé d'individus est fondé sur le fait que 69 % des individus a un Q ≥ 0,8 dans le programme structure (Rutledge et al., 2010c); le nombre d'individus matures est fondé sur le fait que 45,9 % de la population se compose d'adultes (Pimlott et al., 1969). Pour les sites au Québec (Québec), on a utilisé une densité de 2,6 loups par 100 km2 (Papineau-Labelle) ou de 1,9 loup par 100 km2 (autres sites). L'estimation de l'abondance du loup de l'Est la plus proche est de 236 individus matures.
COSEPAC
Comité sur la situation
des espèces en péril
au Cananda

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2015. Évaluation et Rapport de situation du COSEPAC sur leloup de l'Est (Canis sp. cf. lycaon) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xii + 73 p. (Registre public des espèces en péril ).
COSEPAC. 2001. Évaluation et Rapport de situation du COSEPAC sur le loup de l'Est (Canis lupus lycaon au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa, vii + 20 p.
Samson, C. 2001. Update COSEWIC status report on the eastern wolf Canis lupus lycaon in Canada in COSEWIC assessment and update status report the eastern wolf Canis lupus lycaon in Canada, Committee on the Status of Endangered Wildlife in Canada, Ottawa, 1-17 p.
Van Zyll de Jong, C.G., et L.N. Carbyn. 1999. COSEWIC status report on the Grey Wolf Canis lupus in Canada, Committee on the Status of Endangered Wildlife in Canada, 61 p.
Le COSEPAC remercie Linda Rutledge d'avoir rédigé le rapport de situation sur le loup de l'Est (Canis sp. cf. lycaon) en vertu d'un contrat avec Environnement Canada. Graham Forbes, coprésident du Sous-comité de spécialistes des mammifères terrestres (SCSMT) du COSEPAC, a supervisé les modifications qui ont été apportées au rapport de situation (après l'acceptation de la version provisoire du rapport) selon les commentaires formulés par les provinces concernées, le SCSMT, des membres du COSEPAC et des spécialistes externes.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Eastern Wolf Canis sp. cf. lycaon in Canada.
Illustration/photo de la couverture :
Loup de l'Est dans le parc provincial Algonquin. © Michael Runtz, photo reproduite avec sa permission.
Le loup de l'est (espèce présumée Canis lycaon, anciennement lupus lycaon) est un canidé de taille intermédiaire dont les femelles pèsent en moyenne 24 kg et les mâles 29 kg. La couleur de son pelage est souvent décrite comme étant brun-roux ou fauve, mais elle est très variable. Le loup de l'Est se définit par une combinaison de différences génétiques, de traits morphologiques et du rôle écologique qu'il joue en raison de sa préférence pour de plus petites proies que le loup gris (C. lupus). Comme le loup de l'Est s'hybride à divers degrés avec le coyote (C. latrans), on en définit les individus comme ceux présentant une grande « pureté » génétique distincte du loup gris et du coyote dans les analyses de génétique moléculaire. Il est important de remarquer que le loup de l'Est abordé dans le présent rapport n'est pas le même que le loup de l'Est de la région des Grands Lacs qui est abordé dans des rapports antérieurs et que le présent rapport désigne loup boréal/des Grands Lacs (Great Lakes/Boreal Wolf), un hybride entre le loup de l'Est et le loup gris. Bien que beaucoup de données appuient la validité du loup de l'Est comme espèce, sa taxinomie reste controversée. Le présent rapport considère le loup de l'Est comme Canis sp. cf. lycaon, une espèce sauvage qui doit être protégée en raison de son caractère distinct, de sa persistance et de son importance comme grand carnivore, et parce qu'il s'agit sans doute d'une partie de la dernière population vestige de l'espèce de Canis de grande taille de l'est de l'Amérique du Nord. Les connaissances traditionnelles autochtones mentionnent l'existence d'un Canis de taille moyenne dans la région.
On croit que la répartition du loup de l'Est est actuellement restreinte aux forêts mixtes (conifères et feuillus) du centre de l'Ontario et du sud-ouest du Québec, soit dans la région des Grands Lacs et du Saint-Laurent. Le loup de l'Est est disparu de la majeure partie de son aire de répartition d'origine en Amérique du Nord en raison de l'éradication des grands Canis durant quatre siècles. Selon les analyses génétiques, le loup de l'Est est présent dans le centre de l'Ontario et le sud du Québec (au nord du fleuve Saint-Laurent), formant des concentrations dans des aires protégées.
Le loup de l'Est vit habituellement dans des paysages de forêt feuillue ou mixte à faible densité humaine, au sud de la région de la forêt boréale. Il préfère souvent aménager sa tanière dans un sol sableux. Ses tanières et ses sites de rendez-vous se trouvent habituellement près d'une source d'eau permanente dans des paysages dominés par des conifères ou des feuillus. Son territoire couvre souvent une superficie de près de 200 km2.
Le loup de l'Est vit en meute familiale composée d'un couple de géniteurs et leur progéniture de l'année en cours et de l'année précédente. La femelle donne naissance à cinq louveteaux à la fin d'avril ou au début de mai. Les louveteaux restent à la tanière durant leur six à huit premières semaines; ils quittent la meute en se dispersant au bout de 37 semaines. La principale proie de ce prédateur est le cerf de Virginie (Odocoileus virginianus). Des analyses prédateur-proie et des analyses de son régime alimentaire indiquent que le loup de l'Est peut aussi chasser l'orignal (Alces americanus), mais avec une efficacité variable selon la meute, la saison ou l'année. Le castor (Castor canadensis) constitue également une bonne part de l'alimentation du loup de l'Est.
Entre 170 et 195 loups de l'Est (de tous âges) ont été identifiés depuis 10 ou 15 ans. On ignore la taille de la population, mais elle compte vraisemblablement moins de 1000 individus matures. La population minimale estimée est de 236 individus matures, se trouvant pour la plupart dans des aires protégées. L'estimation maximale est de 1203 individus matures selon le meilleur scénario possible, c'est-à-dire si la densité de l'espèce est la même à l'extérieur des aires protégées. Or, comme la plupart des mentions de l'espèce ont été faites dans des aires protégées, la taille de la population mature du loup de l'Est se rapproche sans doute plutôt de 236 individus. Le parc Algonquin, où la plupart des mentions de l'espèce ont été faites, est le seul endroit pour lequel il existe des données sur la tendance de population : la tendance semble stable.
La principale menace, ou facteur limitatif, hors des aires protégées est sans doute la mortalité d'origine humaine par chasse et piégeage, qui est facilitée par le réseau routier. D'après la recherche menée dans le parc Algonquin, une mortalité excessive limiterait la dispersion et modifierait la dynamique de reproduction de la meute, entraînant une autre menace importante, l'introgression de gènes (hybridation) avec le coyote de l'Est en raison du manque de partenaire de la même espèce. La perte et la fragmentation d'habitat liées à l'expansion du réseau routier et l'urbanisation devraient se poursuivre hors des aires protégées et empêcheront sans doute l'expansion de la population.
Le loup de l'Est, sous-espèce du loup gris (Canis lupus lycaon), est désigné espèce préoccupante en vertu de la Loi sur les espèces en péril (LEP) du gouvernement fédéral et la Loi de 2007 sur les espèces en voie de disparition de l'Ontario. Aucune espèce de loup n'est visée par la Loi sur les espèces menacées ou vulnérables du Québec. Dans cette province, la chasse et le piégeage de loups sont permis dans les réserves fauniques, mais pas dans les parcs nationaux (fédéraux ou provinciaux). En Ontario, les loups sont protégés contre la chasse et le piégeage dans le parc Algonquin, dans les cantons entourant le parc, ainsi que dans les réserves fauniques de la Couronne provinciale (Crown Game Preserves). L'espèce est protégée contre la chasse, mais pas contre le piégeage dans le parc provincial de la Rivière des Français. Les loups ne peuvent être ni chassés ni piégés dans les parcs nationaux. Les collectivités autochtones conservent le droit constitutionnel de récolter des loups pour leur subsistance et à des fins cérémoniales, y compris dans les aires protégées. Un permis de chasse au petit gibier est requis pour chasser le loup en Ontario (limite de deux par année) et au Québec (aucune limite). NatureServe cote le Canis lupus lycaon N4 (apparemment non en péril). À l'échelle provinciale, le loup de l'Est est coté S4 (apparemment non en péril) en Ontario et n'est pas coté au Québec.
| Question | Information |
|---|---|
| Durée d'une génération L'âge à la première reproduction est de plus de 2 ans, et la durée de vie est très variable selon le taux de mortalité. |
3,5 ans |
| Y a-t-il un déclin continu inféré du nombre total d'individus matures? La sous-population du parc Algonquin semble stable, tandis que les autres sous-populations sont mal recensées; celles-ci sont petites et relativement isolées et subissent des menaces, comme l'altération de l'habitat, la construction de routes, la chasse et le piégeage ainsi que l'hybridation avec le coyote de l'Est, qui diminuent la population. |
Possiblement |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur deux générations (7 ans). (voir ci-dessus) |
Inconnu |
| Pourcentage présumé de réduction du nombre total d'individus matures au cours des trois dernières générations (10,5 ans). Ce pourcentage est inconnu dans la majeure partie de l'aire de répartition; dans le secteur du parc Algonquin, le nombre d'individus matures a diminué de 20 % de 2002 à 2007 en raison de la maladie, mais a augmenté depuis. |
Inconnu |
| Pourcentage présumé de réduction du nombre total d'individus matures au cours des trois prochaines générations (10,5 ans). (voir ci-dessus) |
Inconnu |
| Pourcentage observé et présumé de réduction du nombre total d'individus matures au cours de toute période de trois générations (10,5 ans) commençant dans le passé (observé) et se terminant dans le futur (présumé). Aucun signe d'un déclin récent. Déclin historique d'environ 50 % de l'aire de répartition au Canada. Mortalité anthropique de 1964 à 1971 (déclin de moins de 16 %) et de 1991 à 1999 (déclin de moins de 12 %) de la sous-population du parc Algonquin (voir Fluctuations et tendances). |
Inconnu |
| Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé? La cause du déclin historique était l'éradication ciblée, qui a cessé. La chasse et le piégeage hors des aires protégées n'ont pas cessé et continuent d'être une menace importante; elles pourraient causer un déclin futur en causant l'introgression de gènes du coyote de l'Est. |
Non |
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Non |
| Question | Information |
|---|---|
| Superficie estimée de la zone d'occurrence Superficie de la zone d'occurrence estimée d'après toutes les mentions |
126 573 km² |
| Indice de zone d'occupation (IZO) La zone d'occupation estimée d'après la superficie de tous les sites abritant le loup de l'Est est de 29 472 km2. |
>10 000 km² |
| La population totale est-elle gravement fragmentée? La population totale est fragmentée par des régions où l'espèce n'est pas protégée et qui présentent un dense réseau routier, une densité humaine relativement forte et un risque accru d'hybridation avec le coyote de l'Est. |
Oui |
| Nombre de localités (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) La population est principalement présente dans 11 sites (et dans de nombreux cantons autour du parc Algonquin) et forme de nombreuses meutes. Si les causes de mortalité étaient indépendantes entre les meutes, il y aurait plus de 20 localités. |
Inconnu, probablement plus de 20 |
| Y a-t-il un déclin continu inféré de la zone d'occurrence? La mortalité hors des aires protégées limite probablement l'expansion de l'aire de répartition, mais la superficie de la zone d'occurrence est estimée d'après des analyses génétiques récentes, et aucun déclin n'a été observé durant cette courte période. |
Inconnu |
| Y a-t-il un déclin continu inféré de l'indice de zone d'occupation? (voir ci-dessus) |
Inconnu |
| Y a-t-il un déclin continu inféré du nombre de populations? (voir ci-dessus) |
Inconnu |
| Y a-t-il un déclin continu inféré du nombre de localités? La chasse et le piégeage hors des aires protégées pourraient influer sur ce nombre. |
Inconnu |
| Y a-t-il un déclin continu inféré du nombre de la superficie ou de la qualité de l'habitat? La chasse et le piégeage limitent la possibilité d'établir des meutes non hybrides dans d'autres localités. La conversion de terres et la construction de routes dans des zones de dispersion possible y réduisent la qualité de l'habitat. |
Oui |
| Y a-t-il des fluctuations extrêmes du nombre de populations? | Inconnu, mais peu probable |
| Y a-t-il des fluctuations extrêmes du nombre de localités? | Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
| Population | Nombre d'individus matures |
|---|---|
| Ontario-Québec Le nombre minimal d'individus matures est estimé à 236 d'après l'extrapolation des résultats pour plusieurs aires d'étude à la superficie des aires protégées où l'espèce a été observée. La zone d'occurrence pourrait abriter au maximum 1203 individus matures si l'on présume que la densité du loup de l'Est est la même hors des aires protégées, ce qui est très improbable parce que peu d'individus ont été observés hors des aires protégées. La population d'individus matures se rapproche sans doute plutôt de 236 individus. |
- |
| Total | Inconnu, mais très probablement moins de 1000 |
| Question | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. Plusieurs analyses ont été effectuées il y a plus de 10 ans, mais seulement pour le secteur du parc Algonquin : les méthodes ont varié, les questions d'ordre génétiques n'ont pas été prises en considération, et les analyses sont généralement peu applicables. Une analyse de la viabilité de la population a conclu qu'il y avait matière à préoccupation, tandis qu'une autre a conclu que la population n'était pas en péril. |
Non disponible |
Chasse et piégeage, introgression de gènes du coyote de l'Est, réseaux routiers qui favorisent la mortalité. L'accès à la majeure partie de l'aire de répartition est limité par la concurrence des populations de coyotes de l'Est.
| Question | Information |
|---|---|
| Situation des populations de l'extérieur? | On ne connaît aucune population de loups de l'Est à l'extérieur du Canada. Le loup roux n'est pas un candidat viable à la translocation au Canada parce que son génome a été altéré par des effets fondateurs et une forte hybridation. La colonisation naturelle par le loup roux au Canada est improbable en raison de la forte densité humaine dans le territoire qui sépare les deux populations sur une distance d'environ 1000 km. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Non |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? Il n'y a aucune source d'individus immigrants. | S.O. |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | S.O. |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Non |
En 1999, le loup gris de l'Est (Canis lupus lycaon) a été considéré comme une sous espèce du loup gris et classé dans la catégorie « données insuffisantes ». En mai 2001, sa situation (à titre de loup de l'Est, Canis lupus lycaon) a été réexaminée et il a été désigné espèce préoccupante. De nouvelles analyses génétiques indiquent que le loup de l'Est n'est pas une sous espèce du loup gris. En mai 2015, la nouvelle espèce loup de l'Est (Canis sp. cf. lycaon) a été désignée espèce menacée.
En 1999, le loup de l'Est a été désigné loup gris de l'Est (Canis lupus lycaon), sous-espèce du loup gris, dans le rapport du COSEPAC sur le loup gris (van Zyll de Jong et Carbyn, 1999) et classé dans la catégorie « données insuffisantes ». En 2001, le loup de l'Est a été désigné espèce préoccupante (Samson, 2001). Le statut taxinomique du loup gris est controversé, mais il y a maintenant un consensus à savoir que le loup de l'Est n'est pas une sous-espèce du loup gris (Kyle et al., 2006; vonHoldt et al., 2011; Rutledge et al., 2012). Dans le présent rapport du COSEPAC, le loup de l'Est est désigné Canis sp. cf. lycaon, et on en identifie les individus comme ceux qui ont un degré élevé de « pureté » (c.-à-d. un Q supérieur ou égal à 80 d'un groupement génétique unique au C. lupus et au C. lycaon). Les individus au Q inférieur à 80 sont considérés trop hybridés pour être des loups de l'Est et ne sont pas inclus dans les estimations de l'abondance et de la répartition.
Dans le présent rapport, le terme « loup de l'Est » désigne le canidé de taille intermédiaire qui vit dans le centre de l'Ontario et le sud-ouest du Québec. Conformément à l'hypothèse des trois espèces, le terme « coyote de l'Est » désigne l'hybride coyote-loup de l'Est de petite taille qui est présent dans le territoire s'étendant de l'Ontario jusqu'à Terre-Neuve, tandis que le terme « loup boréal/des Grands Lacs » désigne l'hybride présumé entre le loup gris et le loup de l'Est qui vit plus au nord dans la forêt boréale du Manitoba, de l'Ontario et du Québec. Le terme « loup gris » désigne le loup non hybride de plus grande taille qui vit dans le nord-ouest du Canada, et « coyote » (C. latrans) désigne le coyote non hybride de plus petite taille de l'ouest de l'Amérique du Nord. Le présent rapport ne porte que sur la situation du loup de l'Est.
Dans le cadre des évaluations des situations des espèces du COSEPAC, le sous-comité des connaissances traditionnelles autochtones (SCCTA) rédige des rapports sur les connaissances traditionnelles autochtones (CTA). Ces rapports préliminaires compilent et résument les CTA facilement accessibles qui s'appliquent aux évaluations. Un rapport de collecte de CTA peut être rédigé s'il existe d'importantes lacunes dans les connaissances ou des contradictions majeures entre les CTA et d'autres types de connaissances. Dans son processus initial, le SCCTA n'a pas relevé de CTA propres au loup de l'Est au Canada, mais on lui en a fourni après la période d'examen de deux mois, et cette information est incluse dans le rapport.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Le Service canadien de la faune d'Environnement Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
La taxinomie du loup de l'Est est controversée, et le présent rapport commence par une discussion de son admissibilité à l'évaluation par le COSEPAC. Voici les critères d'admissibilité des espèces du COSEPAC : validité taxinomique (définie par des études publiées revues par des pairs et des communications avec des spécialistes), origine indigène au Canada (espèce naturellement présente ou présente depuis plus de 50 ans) et présence régulière au Canada (COSEPAC, 2014).
À l'heure actuelle, le COSEPAC considère le loup de l'Est comme une sous-espèce du loup gris (Canis lupus lycaon) (Samson, 2001). Toutefois, à la suite d'analyses génétiques effectuées depuis 10 ans, il y a maintenant un large consensus à savoir que le loup de l'Est n'est pas une sous-espèce du loup gris (voir l'annexe A). La présente section donne un aperçu de la validité taxinomique du loup de l'Est, laquelle est présentée plus en détail à l'annexe A.
Le Canis de grande taille à l'est de la région des Grands Lacs a toujours été reconnu comme étant distinct et est classé comme une espèce ou une sous-espèce depuis plus de deux siècles. Le spécimen type du loup de l'Est a (probablement) été capturé près de la ville de Québec (Québec) en 1761, et Schreber lui a donné le nom de Canis lycaon en 1775 (Miller, 1912; Manning et Sturtevant, 1966). Dans la première grande révision du genre Canis, Pocock (1935) s'est appuyé sur des différences morphologiques pour maintenir le statut du loup de l'Est (renommé loup de l'Est du Canada) comme espèce à part entière et a défini tous les autres Canis dans le centre et l'ouest du nord de l'Amérique du Nord comme des sous-espèces du loup gris. Goldman (1937) a désigné le loup de l'Est Canis lupus lycaon - type algonquin. Le C. lycaon a été inclus comme sous-espèce du C. lupus parce que les mesures de son crâne étaient semblables à celles des autres Canis. Depuis, toutes les analyses morphométriques ont montré que le loup de l'Est est le plus petit de toutes les sous-espèces de C. lupus (Young et Goldman, 1944; Nowak, 1995).
Selon les CTA de la Première Nation mohawk d'Akwesasne, plus d'un type de canidés étaient reconnus dans la région avant l'arrivée des Européens sur la base de différences dans leur taille corporelle, leur tempérament et la taille de leurs proies (Lickers, comm. pers., 2015). Le « petit loup » était considéré comme « gentil » (comparativement au loup gris plus redoutable), et on savait qu'il chassait le cerf de Virginie. Un plus petit canidé (le coyote) est arrivé plus tard dans la région (Lickers, comm. pers., 2015).
Kyle et al. (2006), Chambers et al. (2012) et Mech et al. (2014) ont publié des synthèses détaillées sur l'histoire taxinomique du loup de l'Est. Mech (2011) présente les preuves non génétiques de l'existence du loup de l'Est. Cronin et Mech (2009), Schwartz et Vucetich (2009), Mech (2010a), Benson et al. (2012) et Rutledge et al. (2012) passent également en revue la taxinomie du loup dans l'est de l'Amérique du Nord. Une bonne partie de la controverse entourant la taxinomie des Canis est liée à l'arrivée du coyote (Canis latrans) dans l'est de l'Amérique du Nord. En effet, dans une invasion à l'échelle continentale, le coyote de la région des Prairies de l'Amérique du Nord s'est répandu vers le nord et l'est; la première mention de sa présence dans le sud-est de l'Ontario remonte à 1919 (Nowak, 1979). Ce Canis de petite taille (adultes pesant en moyenne 13 ou 14 kg dans diverses régions du centre des Prairies [Parker, 1995]) s'est accouplé avec un Canis plus grand dans la région des Grands Lacs pour donner un Canis de taille intermédiaire (adultes pesant en moyenne de 14,6 à 21 kg dans diverses régions du nord-est de l'Amérique du Nord [Parker, 1995; Villemure et Jolicoeur, 2004]). Ce nouvel animal, nommé le coyote de l'Est, s'est ensuite établi dans l'est du Canada, atteignant Québec en 1944, la Nouvelle-Écosse dans les années 1970 et Terre-Neuve en 1985 (Parker, 1995; Naughton, 2012).
Les chercheurs s'entendent pour dire que le coyote de l'Est est un hybride (Wilson et al., 2009; vonHoldt et al., 2011). Le débat vise à savoir si le Canis de grande taille qui s'est hybridé avec le coyote était le loup gris ou le loup de l'Est. Dans l'hypothèse des deux espèces, ce Canis de grande taille était le loup gris, et son hybridation avec le coyote a produit divers Canis hybrides, soit le loup de l'Est, le loup roux (C. rufus, désigné espèce en péril aux États-Unis), le loup boréal/des Grands Lacs et le coyote de l'Est (Lehman et al., 1991; Kays et al., 2010; vonHoldt et al., 2011). Le coyote de l'Est, le loup de l'Est et le loup roux ont tous une taille intermédiaire à celles du loup gris et du coyote; cette taille intermédiaire correspond au résultat attendu d'une hybridation et sous-tend en partie l'argument voulant que le loup de l'est ne soit pas une espèce.
Selon l'autre hypothèse (le modèle des trois espèces), le Canis de grande taille qui s'est croisé avec le coyote « de l'ouest » serait une espèce distincte nommée loup de l'Est (ou loup roux) qui était présente surtout dans l'est de l'Amérique du Nord. La troisième espèce, le loup gris, aurait immigré depuis l'Eurasie il y a 300 000 ans (Wilson et al., 2003). Selon cette hypothèse, le loup de l'Est est une véritable espèce, et le coyote de l'Est est un hybride issu du croisement entre le coyote et le loup de l'Est, non pas entre le coyote et le loup gris (Rutledge et al., 2012; Rutledge et al., en révision). Un autre grand Canis, le loup boréal/des Grands Lacs (voir Activités de recherche) serait un hybride issu du croisement entre le loup de l'Est et le loup gris (p. ex. Wheeldon et White, 2009; Wheeldon et al., 2010; Fain et al., 2010; Rutledge et al., 2012, en révision).
Dans certaines études publiées (voir l'annexe A), le loup boréal/des Grands Lacs a été appelé loup de l'Est (Canis lycaon), mais dans le présent rapport le loup de l'Est désigne ce qu'on croit être la population vestige la plus « pure » de l'ancêtre des hybrides que sont le loup boréal/des Grands Lacs et le coyote de l'Est.
Il y a de fortes preuves que le loup de l'Est devrait être considéré comme une espèce distincte (annexe A), mais le COSEPAC reconnaît que la controverse se poursuit (NCEAS, 2014) et que le problème taxinomique n'est pas entièrement résolu. De plus, la population actuelle de l'espèce présumée loup de l'Est contient plus ou moins de gènes du coyote, et on en identifie donc les individus en fonction de la probabilité de les assigner à un groupement génétiquement semblable, donc identifiable (annexe B).
Peu importe la controverse taxinomique, il y a consensus à savoir qu'un Canis distinct est reconnu dans la région depuis le milieu du XVIIIe siècle et qu'il est encore présent dans certaines parties de l'Ontario et du Québec (annexe A). Ce loup exploite une niche écologique différente de celles des canidés semblables à Canis lupus et des canidés semblables à C. latrans (voir Relations interspécifiques). On le trouve surtout dans des zones peu touchées par les activités humaines dans la région des plaines à forêts mixtes du Canada. Il est davantage hybridé (introgression génétique) avec le coyote de l'Est dans les régions plus modifiées par les activités humaines du sud de l'Ontario et du Québec, et avec le C. lupus dans les régions plus boréales qui abritent plus d'orignaux (annexe A). Il s'agit donc d'un canidé distinct qui exploite une niche écologique dans quelques petites aires protégées et les alentours en Ontario et au Québec; il est important de protéger le plus grand canidé dans cette niche écologique qui est menacée et qui s'amenuise.
La Loi sur les espèces en péril (LEP) et le COSEPAC (2014) définissent espèce sauvage comme une espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte qui est indigène au Canada et à l'état sauvage. Le loup de l'Est n'est pas une sous-espèce du loup gris (voir Validité taxinomique) et ne peut donc pas être considéré comme une unité désignable du loup gris. En résumé, le présent rapport considère le loup de l'Est comme une espèce sauvage distincte, persistante et importante sur le plan de l'évolution au Canada. Les loups de l'Est qui restent sont des résidants naturellement présents au Canada depuis plus de 50 ans. Il existe un mécanisme plausible qui maintiendrait le loup de l'Est en sympatrie régionale avec le coyote de l'Est (voir l'annexe A). Le nom proposé pour le présent rapport est loup de l'Est (Canis sp. c.f. lycaon) (annexe E4 dans COSEPAC, 2014).
En raison de la forte plasticité des caractères morphologiques, particulièrement la taille corporelle (Rutledge et al. 2010c), le loup de l'Est est difficile à distinguer du loup boréal/des Grands Lacs et du coyote de l'Est sans test d'assignation fondé sur des marqueurs génétiques. Des loups de l'Est (figure 1) ont cependant été différenciés des loups boréaux/des Grands Lacs et des coyotes de l'Est par la morphologie de leur crâne, leur taille corporelle, leur habitat, leurs proies et leur signature génétique. La morphologie du crâne et la taille corporelle du loup de l'Est sont intermédiaires entre celles du loup boréal/des Grands Lacs et du coyote de l'Est (Kolenosky et Standfield, 1975; Schmitz et Kolenosky, 1985; Sears et al., 2003; Théberge et Théberge, 2004;Rutledge et al., 2010c, 2010d; Benson et al., 2012). Bien avant l'utilisation de la génétique pour identifier des espèces, Kolenosky et Standfield (1975) ont distingué les loups du parc provincial Algonquin (ci-après désigné « parc Algonquin ») des autres types de Canis dans la région d'après leur taille corporelle et la morphologie de leur crâne. Leur carte de la répartition des Canis en Ontario a depuis été corroborée par des techniques de génétique moléculaire (Rutledge 2010a; figure 2).
Photo: © Michael Runtz, 2015
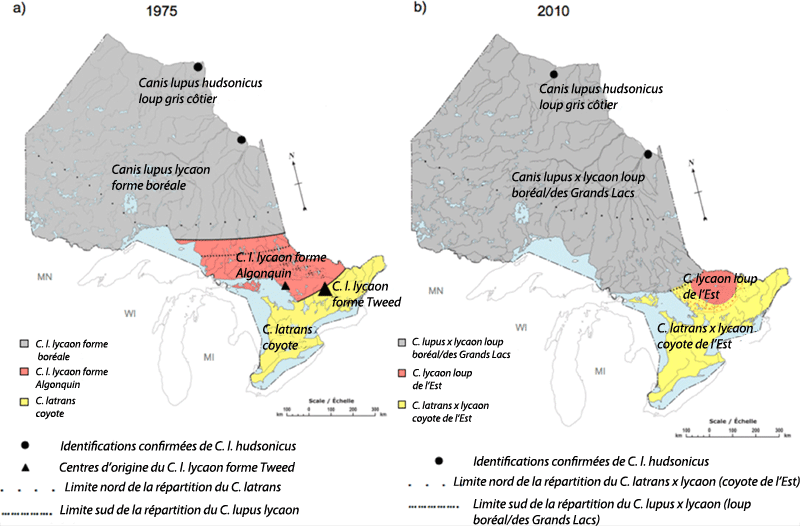
Description longue pour la figure 2
Deux cartes montrant les répartitions proposées des formes des espèces de Canis en Ontario. La carte A montre les répartitions proposées par Kolenosky et Standfield (1975) d'après la morphologie du crâne et le poids corporel. La carte B montre les répartitions proposées par Rutledge (2010). Pour de plus amples détails, veuillez consulter la légende de la figure 2 et le texte du rapport.
Le loup de l'Est pèse habituellement moins de 30 kg (Théberge et Théberge, 2004). D'après les données recueillies dans le parc Algonquin de 2002 à 2007, le poids moyen est de 18,1 kg pour les femelles d'un an, de 24,2 kg pour les femelles adultes, de 23,5 kg pour les mâles d'un an et de 29,3 kg pour les mâles adultes. Selon Jolicoeur et Hénault (2010), le poids moyen des Canis de la réserve faunique Papineau-Labelle (ci-après, réserve Papineau-Labelle) est de 24,6 kg (± erreur type de 0,7) pour les mâles et de 22,2 kg (± erreur type de 0,9) pour les femelles. Dans le parc national de la Mauricie (ci-après, parc de la Mauricie), les Canis femelles adultes ont un poids moyen d'environ 28,7 kg et une hauteur au garrot de 71,8 cm, tandis que les mâles adultes ont un poids moyen d'environ 44,5 kg et une hauteur au garrot de 81,2 cm (Villemure et Festa-Bianchet, 2002). Bien qu'il s'agissait d'un petit échantillon (trois femelles et deux mâles), ces tailles sont supérieures à celles estimées pour le loup de l'Est, et l'échantillon comprenait donc sans doute des loups boréaux/des Grands Lacs. Dans la région du parc Algonquin, la hauteur moyenne au garrot du loup de l'Est est de 63,8 cm chez les femelles et de 70,0 cm chez les mâles (Patterson 2011b, comm, pers.). Le loup de l'Est est plus long (longueur du bout du museau à la base de la queue) que le coyote de l'Est; chez le loup de l'Est, la longueur moyenne est de 109,3 cm (± erreur type de 1,3) pour les femelles et de 113,0 (± erreur type de 1,8) pour les mâles (Benson et al., 2012). La taille corporelle est importante chez les prédateurs, car elle influe sur leurs besoins en énergie et la taille des proies qu'ils peuvent chasser efficacement (Carbone et al., 1999;MacNulty et al., 2009).
La couleur du pelage n'est pas un descripteur fiable des différentes espèces de Canis en raison de sa variabilité. Toutefois, le pelage du loup de l'Est (figure 1) est habituellement plus fauve ou brun roux (roux sur les pattes avant) que ceux du loup gris et du loup boréal/des Grands Lacs.
La structure de population (génétique) des Canis dans la région varie en raison de l'introgression de gènes du Canis latrans et du C. lupus dans la population de loups de l'Est qui restent. Les différentes formes de canidés sont distinguées par leur valeur Q, qui représente la probabilité que l'individu appartienne à un groupement selon le programme d'analyse génétique structure 2.3 (Hubisz et al., 2009; annexe B).
On ignore la quantité de variations au sein de la population de loups de l'Est.
Une seule unité désignable est recommandée pour le loup de l'Est. La définition du loup de l'Est se fonde en partie sur sa différenciation des autres Canis dans la région (c.-à-d. Q ≥ 0,8; annexe B); les individus dont la valeur Q dépasse ce seuil sont considérés comme semblables.
Les loups de l'Est existants constituent sans doute une population relique qui, avant l'arrivée des colons européens, habitait dans les forêts tempérées de l'est de l'Amérique du Nord, y compris les régions méridionales de l'Ontario et du Québec (Rutledge et al., 2010d;Kyle et al., 2006; figure 3). Le défrichage des terres, l'invasion du coyote, qui n'est pas endémique à la région, et la persécution visant les grands canidés auraient poussé le loup de l'Est vers le nord dans le parc Algonquin et le centre-sud du Québec, qui constituent vraisemblablement un refuge à la limite nord de son aire de répartition historique.
Il est difficile de quantifier l'importance socio-économique actuelle du loup de l'Est, mais on estime que plus de 162 000 personnes ont visité le parc Algonquin pour participer au programme éducatif Wolf Howl (appel des loups) depuis les années 1960 (Steinberg, comm. pers., 2013). Ce programme est une composante éducative et récréative particulière du tourisme en Ontario.
La valeur du loup de l'Est pour les trappeurs est inconnue. Des canidés de grande taille sont chassés et piégés dans l'aire de répartition du loup de l'Est (voir Statuts et protection juridiques), mais on ignore le nombre de peaux de loups de l'Est récoltées parce que les peaux de génotype mixte ou inconnu sont habituellement regroupées pour les ventes aux enchères.
Le loup de l'Est a également une importance particulière parce qu'il constitue peut-être la dernière population sauvage importante du loup roux, espèce gravement menacée d'extinction aux États-Unis (voir Validité taxinomique; annexe A). De plus, les grands carnivores influent sur les écosystèmes (Chapron et al., 2008; Beschta et Ripple, 2009, 2010), et le rôle du loup de l'Est dans la région diffère sans doute de celui du coyote de l'Est plus petit et de celui du loup gris plus grand.
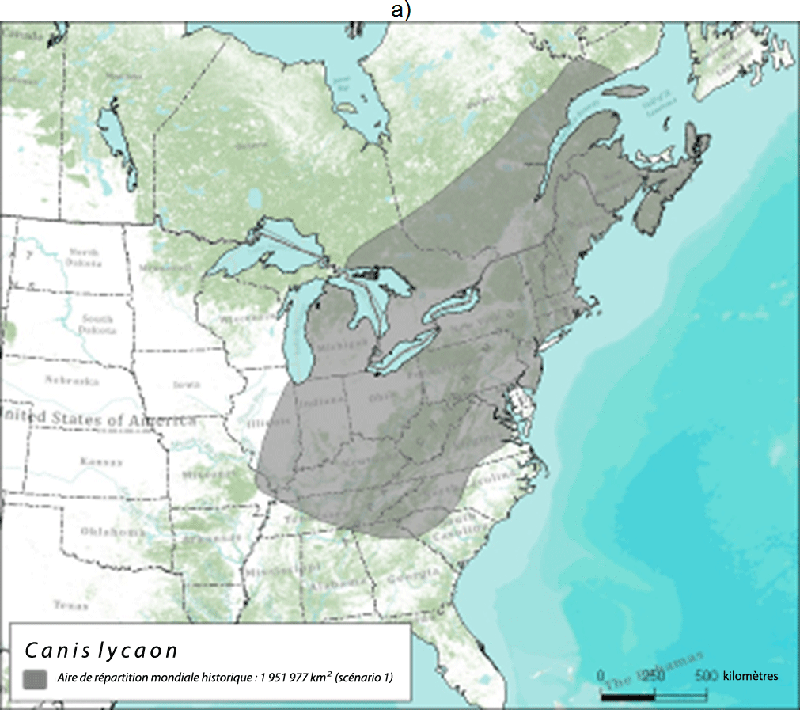
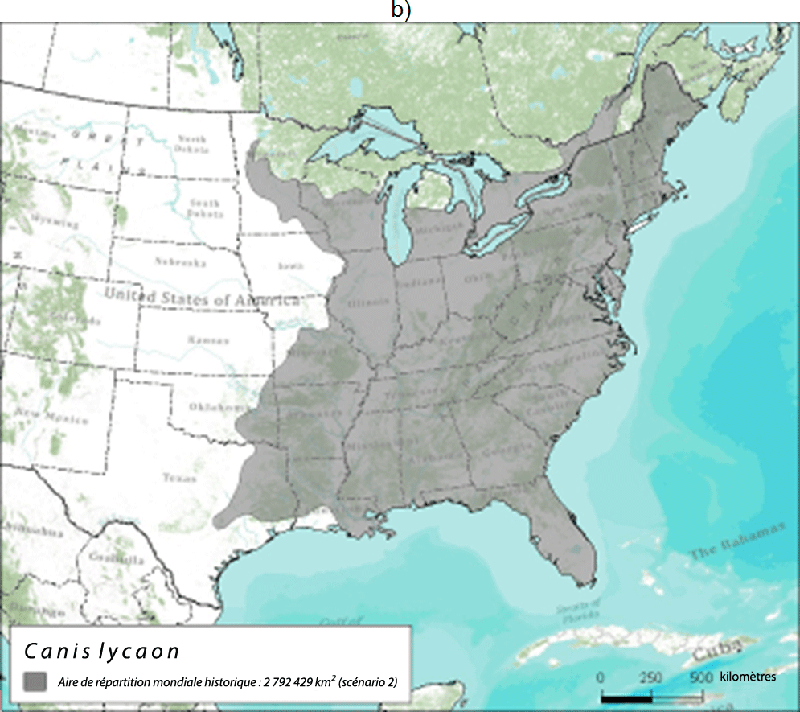
Description longue pour la figure 3
Deux cartes montrant l'aire de répartition mondiale historique du loup de l'Est. La carte A montre l'aire de répartition selon Nowak (1995) et Leonard et al. (2005). La carte B montre l'aire de répartition selon Wilson et al. (2000) et Rutledge et al. (2010d). Pour de plus amples détails, veuillez consulter le texte du rapport.
Le loup de l'Est, tel qu'il est décrit dans le présent rapport, est actuellement absent des États-Unis, bien que son aire de répartition historique couvrait probablement toute la région de la forêt tempérée de l'est de l'Amérique du Nord (figure 3) (Wilson et al., 2000; Kyle et al., 2006; Rutledge et al., 2010d).
Il est difficile de délimiter l'aire de répartition historique du loup de l'Est parce qu'il en existe peu de fossiles et qu'il y a diverses interprétations de sa taxinomie. Si l'on accepte l'interprétation selon laquelle le loup de l'Est et le loup roux étaient historiquement de la même espèce, alors les données génétiques et les fossiles portent à croire que le loup de l'Est/loup roux occupait toutes les forêts tempérées de l'est de l'Amérique du Nord (Wilson et al., 2000; Rutledge et al., 2010d), soit une superficie de presque 3 millions de km2 (figure 3). Cette aire de répartition s'étendait vers l'ouest juste au-delà du fleuve Mississippi, vers le sud jusqu'en Floride et vers l'est jusqu'à la côte atlantique. Bien que par le passé le loup roux occupait des régions à l'ouest du Mississippi en Louisiane et au Texas (d'où provient la population reproductrice en captivité), l'introgression de gènes du coyote, du loup gris et peut-être aussi du chien dans la population de loups roux laisse croire que ces régions étaient des zones d'hybridation historique. Ainsi, le fleuve Mississippi a peut-être constitué une barrière géographique naturelle qui aurait limité l'hybridation à l'est. Selon Nowak (1995), l'aire de répartition historique du loup de l'Est, indépendant du loup roux, aurait été plus petite et plus au nord (figure 3a).
On ne croit pas que le loup de l'Est, tel qu'il est défini dans le présent rapport (Q ≥ 0,8; voir l'annexe B), soit actuellement présent hors du Canada. Selon l'hypothèse des trois espèces, le loup roux et le loup de l'Est étaient sans doute de la même espèce, mais le loup roux se serait par la suite suffisamment différencié, en raison de l'introgression de gènes du chien et du coyote durant le programme d'élevage en captivité (voir l'annexe A), pour ne plus atteindre le seuil de Q de 0,8 utilisé dans le présent rapport (voir l'annexe B). D'après l'ADN mitochondrial (ADNmt) du loup de l'Est trouvé chez des loups échantillonnés dans la région du Minnesota, on considère que ces loups sont des hybrides du loup de l'Est et du loup gris (= loup boréal/des Grands Lacs, ou loup des Grands Lacs) (voir l'Annexe A).
L'aire de répartition canadienne historique du loup de l'Est comprenait probablement la région forestière des feuillus du sud de l'Ontario et du Québec, mais s'étendait peut-être à la partie sud de la région forestière des Grands Lacs et du Saint-Laurent. Selon Nowak (1995), cette aire de répartition s'étendait plus au nord et comprenait la région forestière des Grands Lacs et du Saint-Laurent et peut-être la forêt acadienne (figure 3a). On estime donc que l'aire de répartition canadienne historique du loup de l'Est couvrait entre 112 610 et 500 533 km2.
L'aire de répartition canadienne actuelle du loup de l'Est (figure 4) est délimitée d'après les analyses génétiques publiées (voir l'annexe B) de spécimens de Canis échantillonnés depuis le Manitoba jusqu'aux Provinces maritimes, notamment dans une bonne partie de la région des Grands Lacs (voir Activités de recherche et Validité taxinomique). La plupart des Canis observés hors de la partie centrale de l'aire de répartition sont identifiés comme étant des loups boréaux/des Grands Lacs ou des coyotes de l'Est (Stronen et al., 2012; Rutledge et al., en préparation).
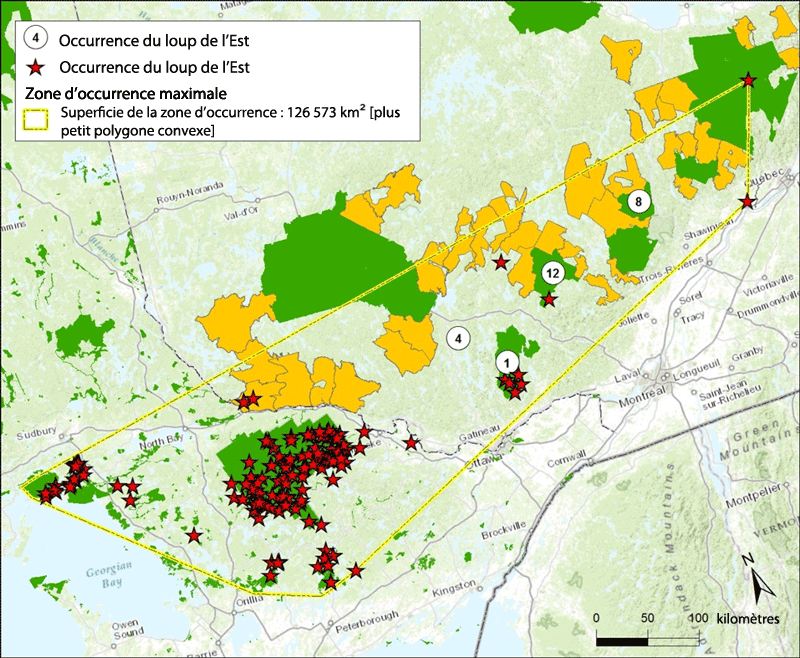
Description longue pour la figure 4
Carte délimitant la zone d'occurrence actuelle (126 573 km2) du loup de l'Est d'après l'analyse génétique de Canis échantillonnés depuis le Manitoba jusqu'aux provinces maritimes, notamment dans une bonne partie de la région des Grands Lacs. Les sites où le loup de l'Est a été trouvé sont indiqués par des symboles. Les aires protégées, les réserves fauniques et les zones d'exploitation contrôlée sont indiquées par des zones ombragées.
Bien qu'on ait trouvé chez des loups du Manitoba un haplotype de la région de contrôle de l'ADNmt et un haplotype de l'ATPase (C3/Ccr13/GL2; Catp13) caractéristiques du loup de l'Est (Stronen et al., 2010; Rutledge et al., 2010b), la plupart des loups du Manitoba présentent de l'ADNmt du loup gris (Stronen, 2009; Stronen et al., 2010). Stronen et al. (2012) ont analysé des microsatellites nucléaires, mais n'ont inclus que huit individus du parc Algonquin, ce qui est sans doute insuffisant pour l'identification de groupements en présence de flux de gènes (Fogelqvist et al., 2010) et n'aurait donc pas permis d'identifier des loups de l'Est selon la définition utilisée pour la sous-population du parc Algonquin (voir les articles de Rutledge et de Benson; annexes A et B). Toutefois, on a trouvé à travers la région s'étendant du Québec à la Saskatchewan des gènes du loup de l'Est (désignés « ADNmt du Nouveau Monde », distinct de l'ADNmt du loup gris de l'Ancien Monde), ce qui, selon l'hypothèse des trois espèces, est interprété comme indiquant que le loup boréal/des Grands Lacs (loups des Grands Lacs) est un hybride entre le loup de l'Est et le loup gris. En général, les analyses de groupement montrent que les loups du Manitoba sont plus apparentés aux loups boréaux/des Grands Lacs des États américains des Grands Lacs et du nord de l'Ontario et du Québec, avec une plus forte influence du loup gris des Territoires du Nord-Ouest (Wheeldon, 2009). Les loups du Manitoba sont considérés comme différents du loup de l'Est du parc Algonquin (Wheeldon, 2009; Wilson et al., 2000).
L'aire de répartition du loup de l'Est en Ontario s'étend de la région à l'ouest du parc Algonquin jusqu'à la vallée de l'Outaouais à l'est et au parc Frontenac au sud. Il s'agit cependant d'une répartition discontinue, les mentions de l'espèce étant restreintes pour la plupart à diverses aires protégées (figure 4).
Le parc Killarney et la région au sud de Sudbury entre ce parc et la route 69 semblent constituer la limite nord de la répartition du loup de l'Est en Ontario (Rutledge et al., en préparation). D'après la taille et la morphologie des individus, ainsi que des données de télémétrie), le loup de l'Est serait résidant dans le parc provincial Queen Elizabeth II Wildlands, ce qui reste à être confirmé par analyse génétique (Patterson, comm. pers., 2014). Dans la région de Magnetawan, à l'ouest du parc Algonquin, la plupart des loups étaient d'ascendance mixte, quoiqu'un petit nombre (<10) s'apparentaient au Canis du parc Algonquin (Wilson et al., 2009). Plus récemment, Benson et al. (2012) ont analysé 342 Canis dans le parc Algonquin et les environs : ils ont identifié trois loups de l'Est dans l'unité de gestion de la faune (UGF) 47 et un dans l'UGF 49, mais ont constaté que les individus hors des parcs Algonquin et Kawartha Highlands (soit les UGF 47 et 49) étaient surtout des coyotes de l'Est ou des individus d'ascendance mixte.
Douze loups de l'Est ont été identifiés dans le parc Kawartha Highlands (376 km2), situé au sud du parc Algonquin (Benson et al., 2012). Bien qu'une population reproductrice de loups de l'Est semblerait s'être établie dans l'UGF 49 (Benson et al. 2012), le coyote de l'Est et les individus d'ascendance mixte y sont plus abondants, sans doute en raison du plus fort taux de mortalité du loup de l'Est hors des aires protégées (Benson et al., 2012).
Au Québec, le loup de l'Est a été identifié par deux méthodes. Celles-ci sont très semblables, sauf que les analyses de Stronen et al. (2012), de Rogic et al. (2014), de Hénault (données inédites) et de Tessier (données inédites) ont utilisé moins d'échantillons de la région du parc Algonquin, ce qui a donné une plus large définition du loup de l'Est et a donc inclus davantage d'échantillons identifiés comme d'ascendance mixte que si la méthode de Rutledge et White (2013, 2014) avait été utilisée. Les résultats des deux méthodes sont présentés à la figure 4), mais celle de Rutledge et White (2013, 2014) a été utilisée en Ontario et au Québec, et on lui donne priorité pour délimiter la zone d'occurrence. Le présent rapport ne tient pas compte des échantillons de l'autre méthode qui se trouvaient hors de la zone d'occurrence. Il faudrait mener d'autres études pour préciser la répartition du loup de l'Est au Québec.
Au Québec, 16 loups de l'Est ont été identifiés parmi 457 échantillons deCanis selon les mêmes techniques que celles utilisées en Ontario (Rutledge et White 2013, 2014; tableau 1; figure 4). Cinq loups de l'Est ont été trouvés dans la réserve faunique Papineau-Labelle (1628 km2), deux au nord de Mattawa, un dans la réserve faunique Rouge-Matawin et un au nord-ouest de cette réserve. Un seul individu isolé a été identifié dans la réserve faunique des Laurentides et un autre près de Saint-Basile (figure 4). Un échantillon d'un mâle capturé au sud du fleuve Saint-Laurent, près de Sainte-Marguerite-de-Lingwick, avait été identifié comme un loup de l'Est parce que son profil de microsatellites correspondait à celui d'un loup de l'Est du parc Algonquin (Villemure et Jolicoeur, 2004), mais Rutledge et White (2014) ont récemment établi qu'il s'agissait plutôt d'un individu d'ascendance mixte.
| Province | Taille de l'échantillon | Nbre de loups de l'Est (Q ≥ 0,8) | % de loups de l'Est |
|---|---|---|---|
| ManitobaNotebdu tableau 1 | 36 | 0 | 0 |
| OntarioNotecdu tableau 1 Noteddu tableau 1 Noteedu tableau 1 Notefdu tableau 1 | 558 | 154 | 28 |
| QuébecNotegdu tableau 1 Notehdu tableau 1 | 457 | 16 (+25Noteadu tableau 1) | 3 |
| Total | 1051 | 170 (+25Noteadu tableau 1) | 16 |
Les occurrences d'un seul individu isolé pourraient être des individus errants, car on ignore s'ils représentent une sous-population sous-échantillonnée de loups résidants ou des loups se dispersant. Ces occurrences sont incluses dans la délimitation de l'aire de répartition du loup de l'Est jusqu'à ce que d'autres études confirment l'absence de l'espèce dans la région.
La méthode utilisée par Rogic et al. (2014) et Tessier (données inédites) leur a permis d'identifier 25 autres loups de l'Est depuis une décennie (figure 4). Tessier (données inédites) a identifié huit loups de l'Est dans le parc de la Mauricie et à moins de 10 km des limites du parc. Les loups du parc de la Mauricie avaient auparavant été classés comme des loups de l'Est d'après leur morphologie, la répartition proposée par Nowak (1995) et la présence d'un haplotype d'ADNmt caractéristique du loup de l'Est (Villemure, 2003; Villemure et Festa-Bianchet, 2002; Villemure, 2011, comm. pers.). Les loups adultes dans le parc ont une taille semblable à celle des loups boréaux/des Grands Lacs, et plusieurs formes de Canis sont probablement résidantes dans le parc (Villemure, 2003).
De même, Rogic et al. (2014) et Tessier (données inédites) ont identifié 12 loups de l'Est dans le parc national du Mont-Tremblant (parc du Mont-Tremblant) par analyse génétique d'échantillons qui avaient récemment été recueillis et par comparaison avec les résultats de l'analyse génétique de Stronen et al. (2012). Il y a aussi quatre mentions récentes de l'espèce dans la région au nord de Mont-Laurier et une près de la ville de Québec (Hénault, données inédites).
D'après des caractères morphologiques incluant des mesures du crâne, Jolicoeur et Hénault (2010) estiment qu'on trouve deux formes de loups au Québec : un loup de petite taille (poids moyen de 23 kg) dont la répartition suit celle du cerf de Virginie (Odocoileus virginianus) et un loup de grande taille (poids moyen de 30 kg) dans les aires de répartition de l'orignal et du caribou (Rangifer tarandus).
Selon Grewal et al. (2004), les loups dans la réserve faunique La Vérendrye ressemblent davantage à ceux de l'Abitibi-Témiscamingue qu'à ceux du parc Algonquin, mais ils seraient plus génétiquement apparentés à ceux du parc Algonquin qu'à ceux du sud-est de l'Ontario. Toutefois, cette conclusion était fondée sur une méthode moins sensible que l'analyse de groupement bayésien du programme structure, c.-à-d. sur une libre interprétation des valeurs RST et FST calculées à partir des données sur huit microsatellites.
Selon Nowak (1995), le loup de l'Est était présent par le passé dans la région forestière acadienne du Nouveau-Brunswick et de la Nouvelle-Écosse, et Nowak (2009) estime que la limite nord de l'aire de répartition de l'espèce s'étend dans la région forestière des Grands Lacs et du Saint-Laurent, en Ontario et au Québec, depuis l'extrémité sud-est du lac Supérieur jusqu'à la péninsule gaspésienne (figure 3a). Dans la péninsule gaspésienne et les provinces maritimes, il existait une forme de Canis qui a disparu au XIXe siècle; on en ignore la taxinomie en raison de l'absence de spécimen (Lohr et Ballard, 1996). Le coyote de l'Est est maintenant commun dans la région, mais aucun profil de microsatellites caractéristique du loup de l'Est n'a été trouvé dans cette population (Way et al., 2010). Par conséquent, l'aire de répartition canadienne actuelle du loup de l'Est ne comprend pas les provinces maritimes.
On estime à 126 573 km2 la superficie de la zone d'occurrence actuelle du loup de l'Est calculée selon la méthode du plus petit polygone convexe englobant toutes les occurrences du loup de l'Est (figure 4). La zone d'occurrence est fondée sur les mentions de l'espèce identifiée par la méthode utilisée en Ontario et au Québec (Rutledge et al., 2010c; Benson et al., 2012; Rutledge et White, 2013, 2014.). Cette zone englobe également la plupart des mentions de l'espèce identifiée par une méthode semblable utilisée par Rogic et al. (2014) et Tessier et Henault (données inédites) au Québec. Les mentions d'individus identifiés par Stronen et al. (2012) et Rogic et al. (2014) hors de la zone d'occurrence ne sont pas incluses (voir section « Québec » dans Aire de répartition canadienne). Il est difficile de déterminer si les mentions d'un seul individu isolé correspondent à des loups résidants ou errants, mais elles sont incluses dans la zone d'occurrence parce que chacun de ces individus pourrait appartenir à une population résidante.
La superficie de la zone d'occupation est estimée à 29 472 km2 d'après la taille des sites où le loup de l'Est a été observé. On s'est servi de la taille de chaque site parce que ces sites sont des aires protégées où la mortalité des Canis est moindre (voir Taux de mortalité et Menaces), ou probablement moindre dans le cas des réserves fauniques du Québec (où la chasse et le piégeage sont permis, mais la densité humaine est faible) que dans les zones non protégées.
De nombreuses études ont comparé des échantillons de Canis recueillis dans l'aire de répartition du loup de l'Est avec des échantillons provenant de la région des Grands Lacs, du sud des États-Unis et d'autres parties du Canada. Un nombre inconnu des mêmes échantillons sont utilisés dans différentes études, mais l'échantillonnage semble avoir été intensif et extensif (figure 5). Wheeldon (2009) a analysé 12 microsatellites nucléaires sur 627 échantillons de canidés du nord-ouest de l'Ontario (n = 87), du nord-est de l'Ontario (n = 93), du Michigan (n = 90), du Wisconsin (n = 48), du Minnesota (n = 53), des Territoires du Nord-Ouest (n = 56), du Manitoba (n = 36 : au nord du parc national du mont Duck, n = 11; parc national du mont Duck, n = 13, parc national du mont-Riding, n = 12), du Québec (n = 34; ouest du Québec, n = 24; est du Québec, n = 10), du parc Algonquin (n = 54), de l'axe de Frontenac (n = 52) et du Texas (n = 24). Way et al. (2010) ont analysé 583 échantillons de Canis provenant des Territoires du Nord-Ouest, de l'Ontario (y compris le parc Algonquin), du Québec, du Nouveau-Brunswick et de l'est des États-Unis. Au Manitoba, Stronen (2009) a analysé 221 échantillons d'excréments et de poils de loups recueillis de 2003 à 2005 dans le parc national du mont-Riding et les monts Duck.
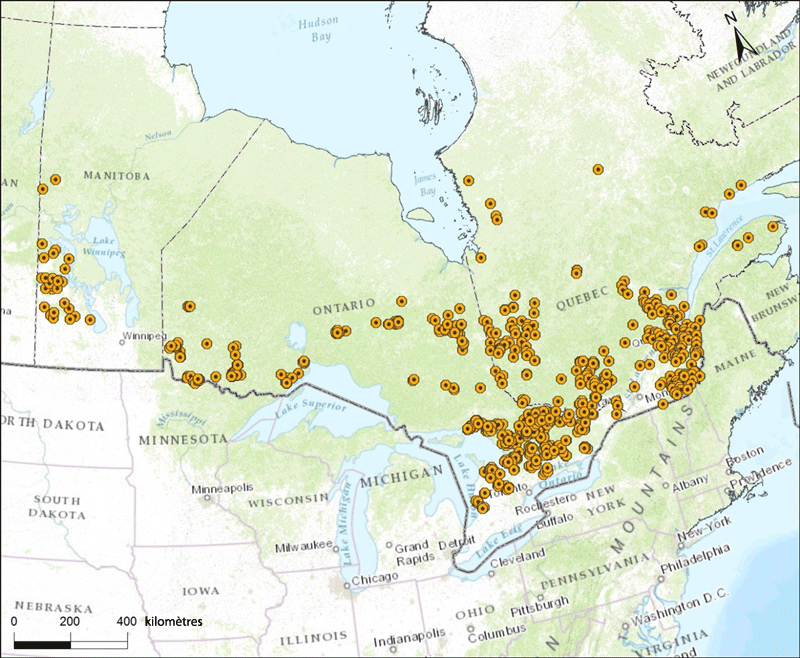
Description longue pour la figure 5
Carte montrant les sites au Manitoba, en Ontario et au Québec d'où proviennent les échantillons de Canis utilisés par de nombreuses études pour déterminer la répartition du loup de l'Est au Canada.
L'échantillonnage de loups de l'Est en Ontario a principalement été effectué dans le cadre d'une vaste étude des espèces de Canis menée par le ministère des Richesses naturelles de l'Ontario depuis 2002. Le parc Algonquin est échantillonné extensivement dans le cadre de la recherche constante sur le loup dans le parc. De 1909 à 1958, le parc avait un programme d'éradication ciblée du loup, auquel on a mis fin pour réaliser un projet de recherche gouvernemental sur l'écologie du loup. Dans ce projet, Pimlott et al. (1969) ont échantillonné 106 loups en les abattant dans le parc; Wilson et al. (2000) ont analysé la région de contrôle de l'ADNmt chez 19 de ces loups. Dix-sept des échantillons de Pimlott et al. (1969) ont fait l'objet de l'analyse génétique de microsatellites autosomiques (Rutledge et al., 2011). Grewal et al. (2004) ont analysé 102 échantillons de sang et de tissus prélevés chez 27 des 35 meutes de loups connues du parc Algonquin entre 1987 et 1999. Rutledge et al. (2010c) ont analysé 128 échantillons du parc Algonquin ainsi que 51 provenant du nord-est de l'Ontario et 38 de l'axe de Frontenac, dans le sud de l'Ontario. Wilson et al. (2009) ont analysé 269 échantillons recueillis dans six régions, notamment le sud de l'Ontario, la région de Magnetewan, à l'ouest du parc Algonquin, et le nord de l'Ontario.
Stronen et al. (2012) ont analysé 109 échantillons des Prairies canadiennes, 154 du Québec et 8 du parc Algonquin pour déterminer la répartition des gènes du loup de l'Est de la Saskatchewan au Québec. vonHoldt et al. (2011) ont analysé 208 loups de partout au monde, 57 coyotes de l'Amérique du Nord et d'autres Canis, y compris le chien domestique, pour étudier l'hybridisation. Seuls trois de ces échantillons provenaient de l'aire de répartition actuelle du loup de l'Est : deux du parc Algonquin et un du sud du Québec.
Au total, 457 échantillons prélevés au Québec (figure 4) ont été analysés en même temps et par la même technique et la même interprétation que des échantillons de l'Ontario (Rutledge et White, 2013, 2014). Rogic et al. (2014) ont analysé 15 échantillons (14 échantillons de tissu, 1 échantillon fécal) recueillis de 2007 à 2013 dans le parc du Mont-Tremblant et sa périphérie et les ont comparés aux échantillons de Stronen et al. (2012).
De nombreux projets de recherche sur l'écologie (p. ex. habitudes alimentaires, survie, déplacements, etc.) du loup de l'Est ont été menés dans le parc Algonquin depuis 50 ans (Pimlott et al., 1969; Forbes et Théberge, 1995; Forbes et Théberge, 1996b; Wydeven et al., 1998; Mills et al., 2008; Loveless, 2010; Rutledge et al., 2010e; Benson et al., 2013b, 2014). Au Québec, des études écologiques sur le loup ont été menées dans des régions abritant le loup de l'Est, comme la réserve Papineau-Labelle (Potvin, 1988; Potvin et al., 1988, 1992a, b), mais on ignore dans quelle mesure les données concernent le loup de l'Est dans ces régions.
La recherche sur l'utilisation de l'habitat par les Canis dans le parc Algonquin et la réserve Papineau-Labelle laisse croire que le loup de l'Est n'est pas restreint à un type de forêt particulier (Pimlott et al., 1969; Potvin et al., 1988; Théberge et Théberge, 2004). La définition d'habitat (niche réalisée) comprend la compétition, la prédation, la disponibilité de proies et les sites de tanières. La disponibilité de proies et la mortalité semblent être d'importantes caractéristiques de l'habitat, le loup de l'Est étant plus abondant dans les milieux où les proies, comme le cerf de Virginie, sont abondantes et où la mortalité anthropique est faible (voir Relations interspécifiques et Menaces). Le cadre écologique et les pratiques de gestion du parc Algonquin offrent le milieu qui convient le mieux au loup de loup de l'Est en raison de la disponibilité de proies; le vaste territoire du parc (7571 km2) et de sa périphérie où les Canis ne sont ni chassés ni piégés (6340 km2) abrite le plus grand nombre de meutes de loups de l'Est exposées à une faible mortalité et à la plus faible introgression de gènes de coyote (Benson et al., 2012). Au Québec, les zones qui abritent le loup de l'Est sont aussi principalement des aires protégées où les populations de proies sont sans doute abondantes, bien que la mortalité pourrait être plus élevée parce que la récolte de loups est permise dans certaines zones (p. ex. la réserve Papineau-Labelle (voir Statuts et protection juridiques).
La taille d'une meute est sans doute liée à la taille du territoire, et les deux pourraient être liées à la densité des proies. La superficie moyenne du territoire du loup de l'Est est de 190 km2 (± écart type de 88, n = 12; Loveless, 2010) dans le parc Algonquin et de 199 km2 (± erreur type de 16, n = 19) dans la réserve Papineau-Labelle (Potvin, 1988). La limite supérieure de la taille des meutes de loups de l'Est était de 9 à 14 individus dans le parc Algonquin (Loveless, 2010; Théberge et Théberge, 2004) et de 10 individus dans la réserve Papineau-Labelle (Potvin, 1988). Étant donné ses besoins énergétiques, le loup de l'Est a besoin de proies de grande taille comme le cerf de Virginie, qui est présent en densité moyenne à élevée dans le sud de l'Ontario (zone écologique des cervidés E3; OMNR, 2009), et le castor, qu'on trouve partout en Ontario surtout dans les régions forestières, mais dont l'abondance fluctue selon la qualité de l'habitat (Fryxell, 2001). L'hiver, le loup de l'Est peut remplacer le cerf par l'orignal dans son alimentation, soit en chassant l'orignal (Loveless, 2010), soit en se nourrissant de carcasses d'orignaux tués par des infestations de tiques d'hiver (Dermacentor albipictus) (Forbes et Théberge, 1992).
Les sites de tanières constituent un important élément de l'habitat, mais des tanières ont été trouvées dans de nombreux types de sols et de forêts et ne devraient pas être un facteur limitatif. Dans l'est du parc Algonquin, les tanières et les sites de rendez-vous se trouvent pour la plupart dans des forêts de pins (Pinus spp.) (Norris et al., 2002) peut-être parce que les sols y sont sableux et faciles à creuser. Après une perturbation humaine de leur tanière, les adultes déplacent leurs louveteaux, mais les tanières perturbées sont souvent utilisées les années suivantes (Argue et al., 2008). Divers sites de rendez-vous sont utilisés jusqu'à l'automne (Mills et al., 2008). Les tanières et les sites de rendez-vous se trouvent souvent dans des paysages à prédominance de conifères près d'une source d'eau permanente (Pimlott et al., 1969; Norris et al., 2002).
La quantité et l'étendue d'habitat végétal abritant des populations de cerfs de Virginie augmentent sans doute dans certaines parties de l'aire de répartition du loup de l'Est, car des paysages autrefois dominés par l'agriculture retournent à la forêt à un rythme estimé de 1,9 % par décennie (Lancaster et al., 2008), ce qui pourrait améliorer l'habitat du loup de l'Est. Il existe encore des paysages forestiers à faible densité de routes immédiatement au sud, à l'ouest et à l'est du parc Algonquin, ainsi que dans une bonne partie de l'aire de répartition de l'espèce au Québec (figure 6). Les plus petites zones d'habitat dans le sud de l'Ontario n'abritent peut-être qu'une meute et sont sans doute trop petites pour soutenir des populations viables de loups (Benson et al., 2014). Un corridor forestier à faible densité de routes relie le parc provincial Bon Echo, le parc provincial Frontenac et la région du lac Charleston, à l'ouest du parc Algonquin. Le parc Queen Elizabeth II Wildlands Park constitue probablement un habitat optimal pour le loup de l'Est, et de récents travaux portent à croire que l'espèce y est présente (Patterson, comm. pers., 2014). Dans le parc Algonquin, la protection contre les feux et l'exploitation forestière d'intensité relativement faible pourraient réduire l'abondance future d'espèces associées au début de la succession forestière comme le cerf de Virginie et le castor (Quinn, 2004, 2005).
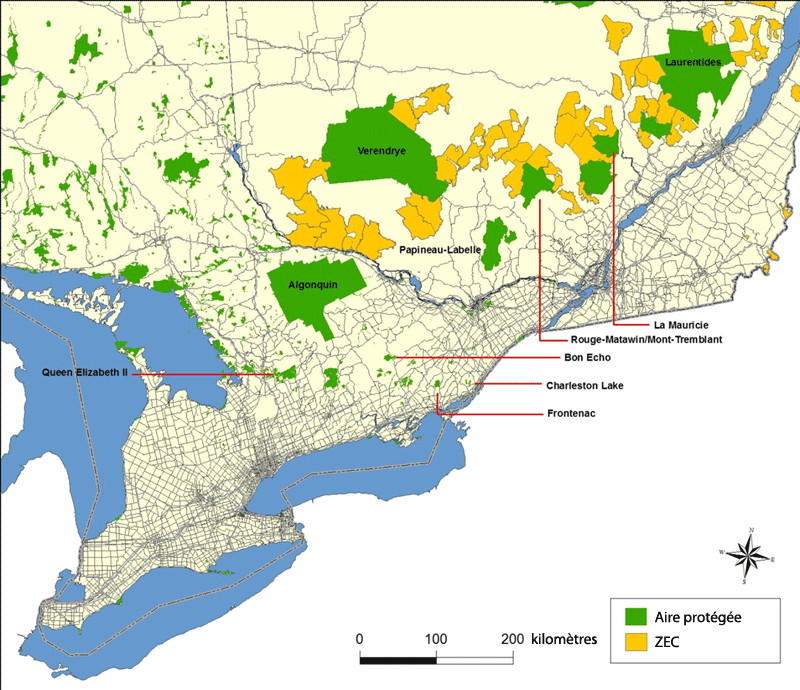
Description longue pour la figure 6
Carte montrant les aires protégées, les zones d'exploitation contrôlée et les principales routes asphaltées dans l'aire de répartition du loup de l'Est. Les aires protégées varient selon leur degré de protection contre la chasse et le piégeage. La récolte de loups est interdite dans les parcs Algonquin, Queen Elizabeth II, du Mont-Tremblant et de la Mauricie alors qu'elle est permise dans les réserves fauniques et les zones d'exploitation contrôlée.
Les habitats d'expansion potentiels comprennent les régions forestières autour du parc Algonquin qui présentent des paysages peu modifiés par les activités humaines, peu de routes et une faible circulation routière et où la chasse et le piégeage sont interdits. Il s'agit des régions autour du parc Algonquin qui s'étendent vers le nord jusqu'au parc Killarney, vers l'ouest jusqu'à la baie Georgienne, vers le sud jusqu'à la route 7 et peut-être aussi sur l'axe de Frontenac jusqu'à la région des Adirondacks (bien que le développement du réseau routier et l'urbanisation pourraient être problématiques dans cette région), ainsi que vers l'Est jusqu'au parc du Fjord-du-Saguenay, en passant par les parcs et réserves fauniques du sud du Québec.
L'expansion de la répartition du loup de l'Est ne se produira pas sans protection contre la chasse et le piégeage dans toute son aire de répartition parce que les jeunes qui se dispersent sont plus susceptibles d'être chassés ou piégés (Théberge et Théberge, 2004). Quatre de cinq juvéniles munis d'un émetteur radio sont morts dans des pièges moins d'un an après s'être dispersés hors du parc Algonquin, et le loup de l'Est était plus vulnérable à la récolte que les autres formes de Canis; les taux de survie étaient de 33 % pour le loup de l'Est et de 66 % pour le coyote de l'Est hors du parc et de 85 % pour tous Canis dans le parc (Benson et al., 2014). De plus, les forts taux de récolte accroissent l'hybridation avec le coyote de l'Est (Rutledge et al., 2011; Benson et al., 2014).
En général, les loups peuvent profiter des routes, car elles accroissent leur efficacité de chasse (Whittington et al., 2011; Lesmerises et al., 2012), mais leurs populations peuvent diminuer si l'accès accru des chasseurs et des trappeurs se traduit par des taux de récolte non viables (Mech et al., 1988; Person et Russel, 2008). Selon Wydeven et al. (1998), une densité de routes de moins 0,4 - 0,7 km par km2 et une densité humaine de moins de 4 personnes par km2 sont nécessaires pour soutenir les populations de canidés de grande taille, si la mortalité anthropique est préoccupante. Dans la majeure partie du sud-est de l'Ontario, la densité des routes est supérieure à 0,6 km par km2 (Buss et deAlmeida, 1997; figure 6), et la longueur totale des routes dans le sud de l'Ontario est passée de 7 133 km en 1935 à 23 806 km en 1965, puis à 35 637 km en 1995 (Fenech et al. 2000). Cette tendance pourrait restreindre la dispersion du loup de l'Est vers le sud.
Une bonne partie des connaissances sur les loups proviennent d'études sur le loup gris ou le loup boréal/des Grands Lacs (p. ex. Mech et Boitani, 2003). On en connaît beaucoup moins sur le loup de l'Est. L'information présentée ici constitue une synthèse des recherches menées sur le loup de l'Est dans le parc Algonquin et au Québec (p. ex. dans la réserve Papineau-Labelle).
Voici les premières étapes de la vie d'un louveteau selon Mills (2006) : 1) il reste à la tanière (jusqu'à l'âge de 6 semaines), 2) il reste au(x) site(s) de rendez-vous (de l'âge de 6 à 12 semaines), 3) il s'éloigne des sites de rendez-vous mais y revient (de l'âge de 6 à 12 semaines) et 4) il abandonne les sites de rendez-vous (à partir de 18 semaines). Les louveteaux naissent à la fin d'avril ou au début de mai. Lorsqu'ils atteignent l'âge de 6 à 8 semaines, la meute les déplace de leur tanière natale au premier site de rendez-vous. Au cours de l'été, la meute les emmène à d'autres sites de rendez-vous de plus en plus éloignés de la tanière (Mills et al., 2008) jusqu'à ce que les sites soient abandonnés à l'automne. Le loup de l'Est chasse en meute, mais les louveteaux ne participent pas à la chasse avant l'âge d'environ 18 semaines lorsque les derniers sites de rendez-vous sont abandonnés.
Les loups vivent en moyenne 4 ou 5 ans (Fuller et al., 2003), mais certains individus atteignent l'âge de 15 ans ou plus (Théberge et Théberge, 2004). Le taux de survie des adultes variait de 63 à 70 % à la fin des années 1980 et au début des années 1990 (Forbes et Théberge, 1995). Benson et al. (2013b) ont estimé le taux de survie annuel moyen (ŝ) des louveteaux à 0,749 (± erreur type de 0,061) dans l'est du parc Algonquin et à 0,246 (± erreur type de 0,073) dans l'ouest du parc. La plus faible survie dans l'ouest du parc est liée à la faible densité de castors et au manque de cerfs l'hiver. La survie des jeunes d'un an et des adultes dans le parc Algonquin était plus élevée (ŝ = 0,852 ± erreur type de 0,05) que celle des louveteaux (Benson et al., 2014). On calcule à partir de ces données que l'espérance de vie [EV = -1/(ln(ŝ), où ŝ est le taux de survie annuel moyen] se situe entre 0,7 et 3,5 ans pour les louveteaux et à 6,2 ans pour les jeunes d'un an et les adultes. La femelle ne se reproduit habituellement pas avant l'âge de 2 ans, de sorte que la durée d'une génération serait d'environ 3,5 ans.
Chaque meute de loups de l'Est comprend habituellement un seul couple de géniteurs non apparentés. Pimlott et al. (1969) ont observé que 59 % des femelles adultes (âgées d'au moins 2 ans) échantillonnées dans le parc Algonquin avaient eu au moins une portée. Les femelles ont une portée de 2 à 7 louveteaux (moyenne de 4,9) chaque année (Mills et al., 2008). Le rapport des sexes des louveteaux de 5,5 semaines est de 1/1, et la mortalité ne varie pas selon le sexe (Mills et al., 2008). Vucetich et Paquet (2000) ont estimé que l'âge moyen des loups de l'Est dans le parc Algonquin et du piégeage était de 2 à 3 ans avant l'interdiction de la chasse et du piégeage, contre 5 ans après l'interdiction. D'après les données de capture obtenues de 2002 à 2007, on estime que la population est constituée à 39,1 % de louveteaux, à 15,0 % de jeunes d'un an et de 45,9 % d'adultes (Patterson, données inédites).
La plupart des loups de l'Est vivent en meutes très sociales et territoriales. La taille moyenne d'une meute dans le parc Algonquin et la réserve Papineau-Labelle est d'environ six individus (Potvin, 1988; Forbes et Théberge, 1996b). La structure sociale de la meute est importante pour assurer son utilisation efficace des ressources (Sand et al., 2006; Stahler et al., 2006; Loveless, 2010) et la survie des louveteaux (Brainerd et al., 2008; Schmidt et al., 2008), ainsi que pour éviter les accouplements incestueux (vonHoldt et al., 2008) et empêcher l'hybridation avec le coyote (Rutledge et al,. 2011).
Lorsqu'ils sont naturellement régulés, les loups de l'Est vivent en meutes familiales et présentent peu de signes de consanguinité. Rutledge et al. (2010e) ont observé des cas où un mâle non apparenté s'est joint à une meute pour en devenir le géniteur (après qu'on a perdu la trace du géniteur connu muni d'un émetteur radio) et où les filles des femelles génitrices se sont par la suite accouplées avec ces mâles au sein de leur meute natale. Lorsqu'ils subissent une forte mortalité anthropique, les meutes se fragmentent, adoptent plus souvent des individus non apparentés (Grewal et al., 2004) et s'hybrident davantage avec le coyote de l'Est (Rutledge et al., 2011).
Selon les recherches menées dans l'est du parc Algonquin, Theberge et al. (2006) ont estimé que le taux de mortalité moyen des loups de l'Est adultes avait été de 0,30 entre 1989 et 1999, bien que les estimations ont été très variables (Patterson et Murray, 2008). Pimlott et al. (1969) ont observé un taux de mortalité élevé chez le loup de l'Est au cours de leur première année de vie. Theberge et Theberge (2004) ont mentionné que les individus d'un an et les sous-adultes étaient plus sensibles à la mortalité causée par l'humain que les adultes. Le taux de survie des petits est estimé à 81 % jusqu'à environ l'âge de 37 semaines, et aucun cas de mortalité n'a été observé au cours des 11 premières semaines de vie (Mills et al., 2008). Dans le parc Algonquin, les causes naturelles sont responsables de la majorité des cas de mortalité (bataille entre congénères, noyade, prédation par l'ours noir [Ursus americanus], blessures causées par les ongulés); les causes liées à des activités humaines arrivent au second rang (empoisonnement et chasse lorsque les loups de l'Est se déplacent ou se dispersent à l'extérieur de l'aire protégée).
Le parvovirus canin et la maladie de Carré ne sont pas associés à la mortalité des petits, bien qu'il soit évident que ces derniers y sont exposés (Mills et al., 2008). Le taux de mortalité des petits semble varier d'une meute à l'autre, mais on ne sait pas pourquoi. Benson et al. (2013b) ont documenté un taux de mortalité totale de 44,3 % dans le parc Algonquin, et des causes naturelles responsables de 33,9 % des cas de mortalité, des causes liées aux activités humaines responsables de 4,6 % des cas de mortalité, et des causes inconnues responsables de 5,8 % des cas de mortalité; le risque de mortalité des petits est associé à la densité des castors et à la disponibilité des cerfs. À l'extérieur du parc Algonquin, le taux élevé de mortalité des adultes est lié à l'ascendance du loup de l'Est (Benson et al., 2014), et le faible taux de survie des petits est lié à l'ascendance mixte du loup de l'Est et du coyote de l'Est (Benson et al., 2013b), ce qui rend difficiles la dispersion et l'expansion des loups de l'Est.
Theberge et al. (1994) ont mentionné que 6 des 18 individus munis de colliers radio étaient morts durant une épidémie de rage en 1990-1991. Aucun cas de rage n'a été signalé dans la population depuis (Patterson, 2011a, comm. pers.). Bien que la mortalité causée par l'activité humaine représente environ 67 % des cas de mortalité de loups dans l'est du parc Algonquin avant 2001, une réglementation interdisant à quiconque de chasser ou de piéger le loup dans les trois comtés situés à l'extrémité sud du parc et dans les comtés environnants, adoptée en décembre 2001, a permis de réduire le taux de mortalité causée par l'activité humaine à 16 % du nombre total de cas de mortalité (Rutledge et al.,2010e). Cependant, le nombre de cas de morts naturelles a augmenté et compense généralement la réduction du taux de mortalité causée par l'activité humaine. Les batailles entre individus d'une même meute constituent actuellement la principale cause naturelle de mortalité des loups de l'Est adultes dans l'est du parc Algonquin (Rutledge et al., 2010e), et le manque de nourriture représente un risque de mortalité important pour les petits dans l'ouest du parc (Benson et al.,2013b).
En 2002-2003, immédiatement après l'adoption de la réglementation interdisant à quiconque de chasser ou de piéger le loup dans les comtés avoisinants du parc Algonquin, le taux de survie du loup de l'Est dans le parc était de 95,2 % (erreur-type = ±0,03), mais ce taux s'est stabilisé entre 2003 et 2006 pour atteindre 81,6 % et 84,2 % (Patterson, données inédites). En 2006-2007, le taux de survie a diminué considérablement jusqu'à 69,7 % (erreur-type = ±0,06) lorsque la gale animale (causée par le Sarcoptes scabiei) a tué un grand nombre de loups de l'Est dans le parc (voir la section « Fluctuations et tendances »). De 1987 à 1999, la gale animale a été rare et n'a pas été débilitante pour les loups du parc Algonquin (Theberge et Theberge, 2004), ce qui porte à croire que le taux de mortalité (et donc la taille de la population) peut fluctuer considérablement d'année en année en raison d'événements imprévus comme les épidémies de maladies.
Il est interdit de chasser ou de piéger le loup dans les parcs nationaux et provinciaux au Québec, mais la chasse au loup et le piégeage du loup sont permis ailleurs, notamment dans les réserves fauniques (St-Louis, comm. pers., 2012). La mortalité causée par l'activité humaine comme le piégeage, les collisions avec les véhicules et la chasse, a éliminé environ 25 à 30 % d'une population dont les individus avaient été marqués dans les années 1980-1990 (Jolicoeur et Hénault, 2010). Le taux de récolte des loups au Québec varie d'une région à l'autre de 2,8 à 29,5 %, et le taux de récolte moyen à l'échelle de la province est de 5,9 %. Villemure et Festa-Bianchet (2002) ont documenté un taux de mortalité de 53,3 % chez les loups munis d'un collier radio provenant du parc de la Mauricie, et ce taux de mortalité est principalement attribuable au piégeage effectué à l'extérieur des limites du parc, qui représente 87,5 % de tous les cas de mortalité; et ce sont les petits qui sont les plus touchés. Une étude menée au début des années 1980 dans la réserve Papineau-Labelle a révélé un taux de mortalité de 34 %, la chasse et le piégeage comptant pour 66 %, et la mortalité routière comptant pour 33 % des 33 Canis qui sont morts (Potvin, 1988). Le taux de récolte général dans la région de Papineau-Labelle était de 9,0 à 17,3 % à la fin des années 1990 (Hénault et Jolicoeur, 2003).
Certains cas de mortalité du loup de l'Est résultent aussi de la capture d'individus dans le cadre d'activités de recherche menées en Ontario. Entre 2002 et 2011, 8 des 328 (2,4 %) loups capturés sont morts après leur capture dans des pièges, des collets ou dans des filets lancés à partir d'hélicoptères (Patterson 2011c, comm. pers.). Deux petits se sont noyés après avoir été capturés, parce qu'ils avaient été déplacés dans une tanière qui a été inondée par la suite; un troisième petit est mort 8 jours après sa capture de causes inconnues (Argue et al., 2008).
Le loup gris est généralement considéré comme une espèce qui s'adapte facilement aux variations climatiques, parce qu'il était autrefois présent du Mexique jusqu'à l'Arctique. Il existe considérablement moins d'information sur le loup de l'Est, mais il est probable que sa capacité d'adaptation soit semblable à celle d'autres Canis de grande taille.
Les individus peuvent s'acclimater lorsqu'ils sont nourris par les humains et lorsqu'ils cherchent des restes de nourriture dans les décharges, et les jeunes qui se dispersent fréquentent parfois les décharges (Mills et al., 2008).
Certains loups de l'Est adultes du parc Algonquin se sont déplacés chaque année sur une distance pouvant atteindre 42 km entre les limites de leur territoire non hivernal et les zones de concentrations de cerfs, parce que les cerfs avaient quitté la zone du parc durant l'hiver (Forbes et Theberge, 1995; Forbes et Theberge, 1996b; Loveless, 2010; Rutledge et al., 2010e).
Les probabilités de dispersion annuelle étaient de 0,22 (IC à 95 % = 0,108 - 0,318), 0,508 (0,341 - 0,633) et 0,144 (0,099 - 0,187) pour les petits, les petits d'un an et les adultes du genre Canis dans le parc Algonquin, respectivement (B. Patterson, données inédites). Bien qu'il arrive souvent que les jeunes qui se dispersent meurent ou errent indéfiniment, ils forment généralement de nouvelles meutes ou se joignent à des meutes existantes dès décembre. Après avoir abandonné les lieux de rendez-vous, les jeunes commencent à chasser et se déplacent avec la meute.
Certains loups de l'Est se sont dispersés sur une distance allant jusqu'à 555 km et peuvent traverser des paysages mixtes composés de forêts, de terres agricoles et de routes à 4 voies (Wydeven et al., 1998). Les loups du parc Algonquin se sont dispersés vers le sud jusque dans le parc Kawartha Highlands, vers l'ouest en direction de la baie Georgienne, et vers l'est jusqu'au Québec. Les jeunes loups de l'Est munis d'un collier radio ne se sont généralement pas dispersés vers le nord, bien qu'un des jeunes en dispersion qui provenait du parc Algonquin ait été repéré à Nakina, dans le nord-ouest de l'Ontario, à une distance directe d'environ 800 km (Patterson, comm. pers.).
Le principal obstacle à une dispersion fructueuse est la forte mortalité causée par l'humain, principalement par la chasse et le piégeage, à l'extérieur des aires protégées (Forbes et Theberge, 1996a; Wydeven et al., 1998; Benson et al., 2013b, 2014). En Ontario, lorsque la mortalité causée par l'humain était élevée, l'hybridation avec le coyote de l'Est a augmenté (Rutledge et al., 2011), probablement en raison du manque de partenaires de la même espèce (Rutledge et al., 2010c) et d'une perturbation de la structure sociale des meutes (Rutledge et al., 2010e). L'expansion de l'espèce dans l'aire de répartition historique est limitée aussi par le manque d'étendues de forêts suffisamment reliées et comportant de faibles densités de routes et d'humains (Harrison et Chapin, 1998). L'expansion vers le nord semble limitée, peut-être en raison de la présence du loup boréal/des Grands Lacs, de plus grande taille, ou d'exigences écologiques de l'espèce.
Dans la réserve Papineau-Labelle, les Canis (dont une certaine proportion était probablement des loups de l'Est) ont consommé principalement des cerfs durant l'hiver, et des castors et des orignaux durant l'été (Potvin et al., 1988). Les populations de castors ont augmenté de 30 % après qu'on ait éliminé 60 % des Canis (Potvin et al., 1992b). De la même façon, Forbes et Theberge (1996b) ont observé une forte corrélation entre la densité du loup de l'Est et le nombre de cerfs en hiver. La sélection de proies par le loup de l'Est varie d'un mois à l'autre, d'une saison à l'autre et d'année en année (Forbes et Theberge, 1996b; Loveless, 2010). Ainsi, dans l'est du parc Algonquin, Theberge et Theberge (2004) ont constaté que la consommation de castors diminuait de mai à juin et que la consommation de cerfs adultes était maximale en juillet. Certaines années, la consommation d'orignaux a été plus importante que la consommation de cerfs, bien que les carcasses ont constitué probablement une proportion élevée des orignaux consommés. Il a été mentionné que le loup de l'Est se nourrissait principalement de carcasses d'orignaux dans les années 1980 (Forbes et Theberge, 1992). Selon des travaux récents, le loup de l'Est serait un efficace prédateur de l'orignal; en 2006, Loveless (2010) a observé que la proportion d'orignaux dans le régime alimentaire du loup de l'Est était plus élevée (64 %) que celle de l'année précédente (47 %), et que la consommation générale de biomasse avait augmenté de l'automne jusqu'au milieu de l'hiver, et qu'elle avait diminué ensuite jusqu'à la fin de l'hiver. La plupart des carcasses d'orignaux consommées par le loup de l'Est étaient des carcasses d'individus adultes, et la proportion globale d'orignaux par rapport à celle des cerfs dans le régime alimentaire a eu tendance à augmenter durant l'hiver. La plupart des orignaux victimes des prédateurs ont été attaqués par des meutes de loups qui semblaient spécialistes de la chasse à l'orignal. En hiver, la taille des meutes était corrélée avec le taux de prédation des orignaux. Le taux de prédation maximal des cerfs a été observé en automne, probablement en raison de la migration qu'effectuent les cerfs durant cette saison. Les orignaux âgés (> 11 ans) et les jeunes orignaux ont été attaqués de manière proportionnellement plus importante que ne l'est leur présence dans la population générale.
Bien que les humains soient les seuls prédateurs connus du loup de l'Est adulte, l'ours noir attaque de manière occasionnelle les petits du loup de l'Est (Mills et al., 2008). Aucun cas de mortalité humaine liée aux interactions avec le loup de l'Est n'a été documenté, mais certaines personnes ont été blessées par des loups dans le parc Algonquin, probablement parce que les loups avaient été nourris dans les campings (Linnell et al., 2002).
Les interactions avec le coyote de l'Est sont traitées dans la section « Menaces ». Les maladies sont abordées dans la section « Menaces », parce que la plupart des maladies associées au loup de l'Est sont liées à des maladies envahissantes et à des animaux de compagnie.
La taille de la population est déterminée à partir des estimations de densité, des cotes d'assignation génétique nucléaire (c.-à-d. le Q), et des proportions des classes d'âge de loups dans le parc Algonquin. La méthode est appliquée ensuite aux secteurs dans lesquels les analyses génétiques ont permis d'établir une localité du loup de l'Est (voir les sections « Aire de répartition mondiale » et « Aire de répartition canadienne »).
La taille de la population n'est pas connue, mais elle est probablement de moins de 1 000 individus matures. Le loup de l'Est a été identifié au moyen des analyses génétiques d'échantillons recueillis de manière désordonnée, à savoir avec différents efforts d'échantillonnage et durant différentes périodes; une taille de la population précise ne peut être obtenue à partir de pareilles données. L'analyse commune des échantillons de l'Ontario et du Québec a permis d'identifier 170 individus (Rutledge et al., 2010c; Benson et al., 2012; Rutledge et White, 2013, 2014) et 25 autres individus ont été identifiés par Rogic et al. (2014), Henault (données inédites) et Tessier (données inédites) dans la zone d'occurrence, ce qui représente un total de 195 spécimens de loups de l'Est provenant de différents efforts d'échantillonnage au cours des 10 à 15 dernières années.
Il est possible d'estimer grossièrement l'abondance si l'on présume que le pourcentage de loups de l'Est (Q ≥ 0,8) et la proportion d'individus matures échantillonnés dans une aire protégée (parc Algonquin), et la densité de Canis à plusieurs sites sont semblables à ceux d'autres aires protégées où le loup de l'Est a été signalé. Il est présumé aussi que ces valeurs sont demeurées constantes au fil du temps. Au cours des 3 dernières générations (10,5 ans), la densité de loups dans le parc Algonquin est demeurée à environ 3 loups par 100 km2 (Rutledge et al., 2010e), ce qui donne à penser à un nombre total extrapolé de 227 loups à l'intérieur des limites du parc (tableau 2). Sur les 128 échantillons de Canis provenant du parc Algonquin qui ont été analysés par Rutledge et al. (2010c), 88 (69 %) avait une cote d'assignation de Q ≥ 0,80. Pour estimer le nombre d'individus matures (c.-à-d. le nombre d'individus capables de se reproduire), seuls les individus considérés comme étant âgés de > 2 ans sont inclus, parce c'est l'âge de la première reproduction (Pimlott et al., 1969). Par conséquent, on estime que le nombre de loups de l'Est matures dans le parc Algonquin (si l'on considère que 45,9 % de la population se compose d'adultes) est de 72 (tableau 2). L'extrapolation de cette méthode aux 6 autres sites en Ontario a permis de dénombrer 82 autres individus, pour un total de 154 loups de l'Est matures en Ontario (tableau 2).
| Localité | Superficie(km2) | Nombre maximal de loups de l'Est à ces sites | Nombre estimé de loups de l'Est (Q ≥ 0,8) | Nombre estimé d'individus matures |
|---|---|---|---|---|
| Parc Algonquin (Ontario) | 7 571 | 227 | 157 | 72 |
| Canton d'Algonquin (Ontario) | 6 340 | 190 | 131 | 60 |
| Parc Kawartha Highlands (Ontario) | 376 | 11 | 8 | 4 |
| Parc Killarney (Ontario) | 645 | 19 | 13 | 6 |
| Secteur des lacs Burwash/Bird (Ontario) | 360 | 11 | 7 | 3 |
| Parc French River (Ontario) | 735 | 22 | 15 | 7 |
| Parc Queen Elizabeth II (Ontario) | 255 | 8 | 5 | 2 |
| Réserve Papineau-Labelle (Québec) | 1 628 | 42 | 29 | 13 |
| Rouge-Matawin/Mont-Tremblant (Québec) | 3 165 | 60 | 41 | 19 |
| Parc de la Mauricie (Québec) | 536 | 10 | 7 | 3 |
| Réserve des Laurentides (Québec) | 7 861 | 149 | 103 | 47 |
| Total | 29 472 | 749 | 516 | 236 |
| Total (zone d'occurrence)Noteidu tableau 2 | 126 573 | 3 797 | 2 620 | 1 203 |
Au Québec, les densités des Canis sont disponibles pour deux sites occupés par le loup de l'Est; la densité estimée dans la réserve Papineau-Labelle est de 2,6 loups/100 km2, alors qu'elle est de 1,6 - 1,9 loup/100 km2 dans la réserve Rouge-Matawin (Jolicoeur et Hénault, 2010). Il est présumé que la densité des Canis dans le parc du Mont-Tremblant, qui est adjacent à la réserve Rouge-Matawin (figure 6), est la même que celle de la réserve, tout comme le sont les densités des sites de La Maurice et des Laurentides, situés plus à l'est. La taille de la population au Québec est estimée à 82 individus matures.
La population de loups de l'« écotype Algonquin » au Québec a été estimée grossièrement à 585 loups, compte tenu de l'habitat de forêt feuillue et de la répartition des cerfs (Jolicoeur et Henault, 2002). Cependant, l'analyse génétique des Canis auQuébec laisse croire qu'un petit nombre de ces individus sont probablement des loups de l'Est avec un Q ≥ 0,8 (Rutledge et White 2014; figures 4).
Il existe quelques mentions du loup de l'Est provenant de l'extérieur des aires protégées mais de l'intérieur de la zone d'occurrence. Ces mentions sont généralement associées à des individus seuls (figure 3). Une estimation de la population maximale a été obtenue à partir de l'hypothèse selon laquelle la densité des loups de l'Est à l'extérieur des aires protégées était égale à la densité des loups de l'Est dans le parc Algonquin. Au Canada, la population maximale serait estimée à 1 203 individus matures dans 126 573 km2. Cependant, l'hypothèse ne cadre pas avec les tendances observées de l'ascendance mixte, et le nombre d'individus est considéré comme étant une surestimation (c.-à-d. un scénario optimiste) de la population des adultes.
Dans le précédent rapport de situation du COSEPAC (Samson, 2001), le nombre de loups de l'Est adultes et jeunes au Canada a été estimé à 1 311 - 2 684 individus dans 210 000 km2. L'estimation minimale actuelle de loups de l'Est est de 344 individus (y compris les individus de tout âge), et l'estimation maximale est de 1 203 individus dans une zone d'occurrence d'une superficie totale de 126 573 km2 (tableau 2). La baisse de la population entre les rapports du COSEPAC résulte de la réévaluation de la zone d'occurrence et des changements dans la façon dont le loup de l'Est est identifié, plutôt que d'un changement mesurable ou comparable de la taille et de la répartition de la population.
Le Canis de grande taille a disparu des Maritimes, de l'est des États-Unis et du sud de l'Ontario et du Québec. Si le loup de l'Est a occupé l'ensemble de cette région (figure 3b), ou seulement une partie du nord de cette région (figure 3a), la perte de la population historique de loups de l'Est a touché au moins 50 % de son aire de répartition.
Les récentes tendances de la population au Québec ne sont pas connues. En Ontario, les activités de recherche menées depuis la fin des années 1950 ont révélé plusieurs cas de mortalité atypique ayant causé des déclins à court terme. Le fait d'avoir tué des loups à des fins de recherche en 1964-1965 (Pimlott et al., 1969) a fait diminuer la densité des loups dans la zone d'étude de 1 700 km2; cette densité est passée de 4,7 loups par 100 km2 (n = 80) en 1964 (avant l'étude) à 2,4 loups par 100 km2 en 1972-1973 (n = 41). Entre 1988 et 1999, Theberge et Theberge (2004) ont observé une tendance à la baisse pour ce qui est de l'abondance des loups dans l'est du parc Algonquin en raison de la mortalité causée par l'humain. La densité de loups était en moyenne de 2,37 individus par 100 km2 (écart-type = ±0,57), et la plus faible densité enregistrée a été de 1,4 individu par 100 km2 en 1999. L'abondance estimée dans la zone d'étude a été la plus élevée en 1991-1992 avec 93 adultes, et la plus faible en 1998-1999 avec 38 adultes, ce qui laisse croire que le nombre d'adultes a baissé de 43 à 17 individus, soit une diminution de 60 % dans la zone d'étude, et une diminution de 5,0 à 16,6 % dans la population générale d'individus matures. Le taux de survie était élevé immédiatement après l'adoption de la réglementation interdisant la chasse en décembre 2001 (s = 95,2 %), ce qui aurait favorisé une augmentation de la densité de loups. Le taux de survie s'est stabilisé entre 2003 et 2006 à 81,6 - 84,2 %, et a atteint 85 % entre 2007 et 2010 (Benson et al., 2014). En 2006-2007, la population a diminué considérablement (p < 0,05) à 69,7 % en raison d'une épidémie de gale animale (Patterson 2011b, comm. pers.), et a augmenté par la suite.
La tendance de la population canadienne au cours des trois dernières générations (c.-à-d. 2005 - 2015) n'est pas connue.
Une analyse de viabilité de la population (AVP) menée par Vucetich et Paquet (2000) indique que la population de loups de l'Est dans le parc Algonquin a connu un déclin entre 1988 et 1999 et que le taux de mortalité annuel (33 %) a dépassé le taux de recrutement annuel (21 %). Par la suite, Theberge et al. (2006) ont suggéré qu'il serait difficile d'établir s'il y avait eu une réponse positive de l'abondance des loups à la réglementation interdisant la chasse dans les 30 mois suivant son adoption en raison d'une forte stochasticité. Patterson et Murray (2008) ont indiqué que ces analyses étaient erronées et ont conclu, à la suite de leur propre analyse, qu'une réponse de l'abondance serait détectable et qu'il était peu probable que la population de loups diminue considérablement au cours des 20 prochaines années. Dans le parc Algonquin, la faible occurrence d'accouplements incestueux depuis l'adoption de la réglementation interdisant la chasse laisse croire que la consanguinité ne constitue pas un problème pour le moment dans le parc; le taux de consanguinité à l'extérieur du parc n'est pas connu, mais il serait plus élevé, selon les analyses génétiques (Rutledge et al., 2010e).
Le loup de l'Est, tel qu'il est décrit présentement, est absent des États-Unis, et une immigration de source externe n'est donc pas possible. Bien que le loup roux dans le sud-est des États-Unis et le loup de l'Est au Canada aient probablement évolué à partir de la même population historique, il existe un taux élevé d'hybridation dans la population de loups roux, notamment des contributions génétiques provenant du coyote, du loup gris et du chien (voir l'annexe A). Ce génome mixte, combiné au potentiel d'évolution limité attribuable à un étranglement génétique lié à un effet fondateur du programme d'élevage en captivité, fait de la population de loups roux une option non viable pour une immigration de source externe au Canada.
Les résultats du calculateur des menaces sont résumés à l'annexe C. L'impact global des menaces est de très élevé à élevé, et repose principalement sur la menace continue et probable de mortalité et de son impact sur l'expansion de la population à l'extérieur des aires protégées.
Le comportement territorial du coyote de l'Est limite peut-être l'expansion du loup de l'Est à partir des aires protégées. Des données issues de recherches menées dans le parc Algonquin et ses environs indiquent que les meutes de coyotes de l'Est et les meutes de loups de l'Est sont territoriales (Benson et Patterson, 2013a). Les loups de l'Est qui se déplacent individuellement sont peu susceptibles de s'établir dans des secteurs déjà occupés par une meute de coyotes de l'Est. Dans la zone d'occurrence. la plupart des zones situées à l'extérieur des zones principales occupées par le loup de l'Est contiennent des coyotes de l'Est.
En général, les loups sont capables de vivre dans les zones agricoles ainsi que dans les zones résidentielles et commerciales de faible densité (Musiani et al., 2010). Les populations de cerfs peuvent être assez abondantes dans les zones rurales, et les loups peuvent aussi trouver de la nourriture dans les déchets. Cependant, les loups ne sont généralement pas tolérés par la société, et il arrive qu'ils soient tués; ils sont habituellement plus communs dans les secteurs sans activité humaine ou dans les secteurs où le développement agricole, résidentiel ou commercial se fait à faible densité. Étant donné que le calculateur de menaces tient compte des facteurs proximaux, les menaces directes, comme la perte d'habitat causée par l'expansion résidentielle, sont présentées dans la catégorie « Développement urbain et résidentiel », alors que tout cas de mortalité, peu importe qu'il survienne dans un secteur résidentiel ou forestier, est traité dans la catégorie « chasse et piégeage ».
Dans l'aire de répartition du loup de l'Est, les loups font partie des espèces d'animaux à fourrure réglementées (voir la section « Statuts et protection juridiques »). Le loup de l'Est est protégé de la récolte dans son aire de répartition principale dans le parc Algonquin, les comtés environnants, et certaines autres aires protégées (voir la section « Statuts et protection juridiques »). Dans le reste de son aire de répartition, le loup de l'Est est visé par la récolte. Le nombre d'individus récoltés n'est pas connu, parce que les canidés sont assez semblables et qu'ils sont regroupés dans les données sommaires des ventes aux enchères. En 2011-2012, 571 peaux de « loups » provenant des piégeurs au Québec ont été vendues (MFFP, 2012c). Dans la région des Grands Lacs et de l'Ontario, 98 loups des bois, y compris ceux provenant des sections « Arctique » et « de l'Est » ont été vendus aux enchères ainsi que 5 545 peaux de « coyotes », notamment celles qui provenaient de l'ouest et de l'est (Fur Harvesters Auction Inc., 2011).
Bien que la majeure partie de la population connue se trouve dans des aires protégées, il est probable que la mortalité causée par l'humain limite la population ailleurs et qu'elle en limite l'expansion. Avant l'adoption de la réglementation interdisant la chasse autour du parc Algonquin, la mortalité causée par l'humain représentait approximativement 67 % des cas de mortalité de loups dans l'est du parc (Forbes et Theberge, 1996a); après l'adoption de la réglementation (voir la section « Statuts et protection juridiques »), les cas de mortalité de loups ont diminué jusqu'à 16 % (Rutledge et al., 2010e). À l'extérieur des aires protégées, la chasse et le piégeage ont entraîné des taux de mortalité élevés (Jolicoeur et Hénault, 2010; Benson et al., 2014).
La mesure dans laquelle les données sur les peaux constituent un indicateur du taux de mortalité n'est pas connue. En Ontario, certaines personnes abattent les loups et les coyotes durant la chasse au cerf ou à l'orignal, car elles pensent que le fait de tuer les loups et les coyotes fera augmenter les populations d'ongulés (Patterson, 2012b, comm. pers.). L'étendue des cas de mortalité associés à ce comportement n'est pas connue dans l'aire de répartition du loup de l'Est.
Il a été établi que la densité des routes constituait l'un des principaux facteurs limitatifs à la dispersion du loup de l'Est, parce que les routes sont un indicateur de l'augmentation de la récolte issue de la chasse et du piégeage. Les cas d'hybridation avec le coyote de l'Est sont plus fréquents dans les régions à forte densité routière (Benson et al., 2012), probablement parce que les taux de mortalité élevés associés à l'accès à la récolte semblent faciliter l'introgression par le coyote (voir les sections « Validité taxinomique » et « Tendances en matière d'habitat »). La forte densité routière dans le sud de l'Ontario et du Québec (figure 6) constitue probablement un risque de mortalité routière et peut nuire considérablement à l'expansion naturelle du loup de l'Est dans son aire de répartition historique. Entre 1985 et 1995, la longueur des routes asphaltées à voie simple ou à voies multiples a augmenté de 3 025 km dans le sud de l'Ontario (Fenech et al., 2000), soit une augmentation moyenne de 302,5 km par année, ce qui laisse croire que la densité des routes continuera à augmenter à l'extérieur des aires protégées.
La présence de zones agricoles peut signifier que la disponibilité des cerfs est accrue, mais ces sites peuvent poser problème au loup de l'Est en raison de la mortalité causée par l'humain en réponse à la prédation du bétail par les loups dans des zones d'expansion possible. Il est difficile de quantifier cette menace, parce que la réaction des agriculteurs est très variable (p. ex. Stronen et al. [2007]) et que les divers points de vue sont source de division en ce qui concerne la gestion des loups dans les zones agricoles où ces derniers attaquent le bétail (Mech, 2010b; Way et Bruskotter, 2012).
Bien que les attitudes positives envers les loups aient augmenté au cours du siècle dernier (en particulier depuis les années 1970), d'autres attitudes peuvent encore être considérées comme hostiles dans certaines régions (Bruskotter et al., 2007, 2011; Williams et al., 2002). Dans les années 1980 et 1990, les attitudes envers les loups dans les collectivités situées à proximité du parc Algonquin étaient généralement défavorables (Theberge et Theberge, 2004). Au Québec, l'attitude des piégeurs envers les loups est variable; environ 42 % d'entre eux s'opposent à la gestion visant à faire augmenter le nombre de loups, et le reste des piégeurs appuient leur augmentation (29 %) ou sont indécis (29 %), en particulier en ce qui a trait au parc de la Mauricie et aux régions environnantes (Bath, 2006). Une enquête menée auprès de résidents à proximité du parc de la Mauricie révèle que la plupart d'entre eux sont sympathiques à la cause du loup de l'Est, mais que la plupart surestiment le nombre de loups dans la région et qu'ils ne soutiennent donc pas la gestion visant à faire augmenter la taille de la population (Parcs Canada, 2007). Les résultats de cette enquête donnent à penser qu'une meilleure éducation du public à l'égard des loups constitue un élément clé de la conservation du loup au Québec. Il a été proposé que la modification des valeurs sociales constitue le facteur le plus important pour le soutien des populations de loups dans le monde entier (Musiani et Paquet, 2004).
L'impact des collisions avec les véhicules sur la population de loups de l'Est n'est pas connu. Dans l'ensemble de la zone d'occurrence, il existe des routes où circulent beaucoup de véhicules, mais ce genre de route est moins commun dans les sites protégés. Il est peu probable qu'on assiste à une forte augmentation des routes où circulent beaucoup de véhicules, mais le volume de circulation augmentera sans doute sur les routes existantes. Un troisième Canis, muni d'un collier radio, est mort après avoir été frappé par un véhicule dans la réserve Papineau-Labelle (Potvin, 1988), et 2,7 ± 1 % (écart-type) des adultes, et 3,7 ± 2 % (écart-type) des petits sont morts dans un échantillon de loups de l'Est munis de colliers radio dans le secteur du parc Algonquin de 2002 à 2007 (B. Patterson, données inédites). De 2004 à 2010, 4,9 % des loups de l'Est ont été tués à la suite de collisions avec des véhicules dans la même zone d'étude (Benson et al., 2014). La menace indirecte que posent les routes est abordée dans la section « Utilisation des ressources biologiques » (UICN 5.1).
La présence du coyote a été documentée pour la première fois en Ontario dans le comté de Lambton en 1919 (Nowak, 1979). Depuis ce temps, le coyote s'est hybridé avec le loup de l'Est, ce qui a donné une population en pleine croissance de coyotes de l'Est, qui sont communs dans le sud de l'Ontario ainsi que depuis l'est des États-Unis jusqu'à Terre-Neuve (Way et al., 2010). La capacité d'adaptation du coyote aux paysages modifiés par l'humain a entraîné une colonisation rapide de régions de l'est de l'Amérique du Nord (Kays et al., 2010) qu'occupait par le passé le loup de l'Est. Le fort taux de mortalité associé à l'humain constitue probablement la principale cause de l'hybridation actuelle du loup de l'Est avec le coyote de l'Est, à l'exception du parc Algonquin où la protection du loup de l'Est a freiné l'hybridation continue (Rutledge et al., 2011). Cependant, à l'extérieur des régions protégées, la plupart des Canis ont été identifiés comme étant soit des coyotes de l'Est de petite taille, soit des individus d'ascendance très mixte (hybrides du loup de l'Est x coyote de l'Est). Très peu d'individus ont été identifiés comme étant des loups de l'Est et ce, malgré l'absence d'obstacles physiques ou géographiques. Par conséquent, à l'extérieur des aires protégées, l'introgression de gènes par le coyote de l'Est menace la persistance du loup de l'Est, parce qu'il y a déjà eu une première hybridation entre les deux espèces, puis un rétrocroisement du coyote de l'Est (voir les sections « Besoins en matière d'habitat » et « Tendances en matière d'habitat »).
Une augmentation des superficies d'habitat modifié, notamment par le déboisement de terres à des fins résidentielles, peut entraîner une diminution de la qualité de l'habitat si les espèces proies ne sont pas présentes. Une zone à forte densité d'habitations ne contient généralement pas de proies pour le loup de l'Est. Cependant, une grande partie de l'aire de répartition du loup de l'Est comporte des fermes, des chalets et de petites villes, et le cerf est présent dans les zones rurales. La principale menace est probablement liée à l'augmentation des contacts entre les humains et les loups, qui peut faire accroître la mortalité des loups; cette menace indirecte est abordée dans la section « Utilisation des ressources biologiques » (UICN 5.1). L'urbanisation et la conversion des terres actuelle vont continuer à menacer indirectement l'expansion possible du loup de l'Est, qui est menacée. Le développement résidentiel et commercial constitue pour le moment une menace mineure pour le loup de l'Est, parce que la majeure partie de la population se trouve dans des aires protégées où le développement est limité (figure 4). Le rythme prévu de développement des activités résidentielles et commerciales à l'extérieur des aires protégées n'est pas connu, mais il est peu probable qu'il diminue.
Une augmentation des superficies d'habitat modifié, notamment par le déboisement de terres à des fins agricoles, peut poser une menace directe pour le loup de l'Est si les espèces proies ne sont pas présentes. Bien que les proies dans certaines zones de l'aire de répartition du loup de l'Est ne soient pas connues, on sait que le cerf est souvent présent dans les zones agricoles et que l'agriculture diminue dans certaines zones qui sont converties en forêt (voir la section « Tendances en matière d'habitat »).
Les intrusions par les humains, comme les visites des tanières ou des lieux de rendez-vous, pourraient avoir un impact sur le loup de l'Est, parce que ce dernier a tendance à éviter les humains et à déplacer ses petits lorsque la tanière a été dérangée (Frame et al., 2007). L'effet serait probablement limité, parce que la majeure partie de la population se trouve dans des aires protégées où les activités sont réglementées. On ne connaît pas le degré de perturbation, mais il est probablement négligeable.
Les chiens domestiques ont exposé le loup de l'Est à des maladies indigènes, notamment le parvovirus-2, la maladie de Carré et l'hépatite canine. On trouve des anticorps contre ces maladies chez la plupart des petits du loup de l'Est en bas âge, mais le fait d'être exposé à ces maladies n'entraîne généralement pas la mort (Theberge et Theberge, 2004; Mills et al., 2008). Theberge et Theberge (2004) ont observé chez le loup de l'Est des anticorps contre le parvovirus (82 %), l'hépatite canine (76 %), la maladie de Carré (46 %) et la rage (20 %). Bien que les cas de mortalité liés aux maladies indigènes aient été rares chez les loups du parc Algonquin, la population du parc national Isle Royale, au Minnesota, a connu un déclin spectaculaire entre 1980 et 1982 en raison d'une épidémie de parvovirus qui a fait chuter la population à seulement 14 individus (Peterson et al., 1998). Les maladies peuvent constituer un important risque de disparition dans les petites populations reliques de canidés sauvages (Woodroffe et al., 2004).
La menace existe en raison d'une possibilité de mortalité associée aux chiens domestiques des touristes dans les aires protégées, et la diminution du nombre de loups adultes pourrait faire augmenter le degré d'introgression des gènes par le coyote chez le loup de l'Est (voir la section « Validité taxinomique »). L'ampleur de la menace est considérée comme étant faible, parce qu'il semble exister un certain degré d'immunité au parvovirus dans la population.
Bien qu'il soit rare que les loups et les coyotes contractent la rage, 15 cas ont été documentés en Ontario entre 1960 et 1994 (Theberge et al., 1994). Durant l'hiver de 1990 - 1991, 4 loups (peut-être 6) sont morts de la rage (Theberge et al., 1994), mais aucune mort liée à la rage n'a été signalée depuis (Patterson, 2011, comm. pers.). L'épidémie de rage chez les loups du parc Algonquin aurait été liée à une épidémie de rage du renard en 1990-1991 (Theberge et Theberge, 2004). La survie des loups de l'Est qui présentaient des anticorps contre la rage serait due à l'ingestion d'appâts vaccinaux antirabiques qui ont été distribués dans l'est du parc Algonquin en 1991-1992.
Il n'existe actuellement aucune estimation directe de la manière dont les changements climatiques pourraient toucher le loup de l'Est. Cependant, Il est prévu que les effets des changements climatiques sur les populations d'orignaux mèneront à une réduction de la densité des orignaux aux limites sud (au sud de la route 101; environ au 48e parallèle) de l'Ontario, particulièrement dans la région du parc Algonquin (Rempel, 2011). En général, les modèles prédisent un déplacement vers le nord de l'orignal et une baisse des populations dans les régions du sud en raison des conditions climatiques limites et de l'habitat marginal (Murray et al., 2006) ainsi que de l'expansion des maladies parasitaires associées à l'augmentation de la densité des cerfs (Lankester, 2010). Bien que Murray et al. (2006) aient suggéré que les changements climatiques seraient profitables pour les populations de cerfs, ils ont constaté aussi qu'il est difficile de prédire dans quelle mesure des températures plus élevées auraient un impact sur les parasites des cerfs. En général, la densité des cerfs devrait augmenter en Ontario et au Québec (Thompson et al., 1998), ce dont pourrait profiter le loup de l'Est. Lankester (2010) a suggéré aussi que les hivers deviendront plus courts et plus doux et que les périodes libres de neige seront plus longues, ce qui aura comme conséquence de réduire le succès de la chasse pour le loup de l'Est, car ce dernier est avantagé par rapport aux ongulés lorsque la neige est profonde (DelGiudice et al., 2002; Crête et Larivière, 2003).
La plupart des études sur les changements climatiques ont mis l'accent sur les interactions entre les plantes et les herbivores, et il existe assez peu d'études qui examinent l'impact des changements climatiques sur les interactions aux niveaux supérieurs de la chaîne trophique. Wilmers et al. (2007) ont montré que, en absence de loups, les populations d'orignaux devraient être plus sensibles aux variations climatiques. Wilmers et al. (2006) ont fourni des données selon lesquelles les loups modifient l'impact des changements climatiques sur les populations d'ongulés. Par conséquent, une population de loups de l'Est en santé pourra jouer un grand rôle sur la fluctuation prévue des populations d'ongulés en réponse aux changements climatiques. Les effets directs et indirects des changements climatiques sur la population de loups de l'Est ne sont pas connus.
Le loup de l'Est figure parmi les espèces préoccupantes à l'annexe 1 de la Loi sur les espèces en péril (LEP), sous le nom scientifique Canis lupus lycaon. Un plan de gestion du loup de l'Est, exigé par la Loi dans le cas des espèces préoccupantes, est en voie d'être élaboré par Environnement Canada.
Le loup gris figure à l'annexe II de la Convention sur le commerce international des espèces de faune et de flore sauvages menacées d'extinction (CITES), qui est mise en application au Canada par Environnement Canada. Une licence d'exportation CITES est requise pour toute exportation de loups du Canada, notamment les spécimens de loups de l'Est (identifiés comme étant des Canis lupus lycaon). Outre la CITES, il n'existe actuellement aucune autre entente internationale visant la protection d'espèces qui inclurait le loup de l'Est.
Le loup de l'Est est considéré comme une sous-espèce du Canis lupus.
Le loup de l'Est est visé par la Loi de 2007 sur les espèces en voie de disparition de l'Ontario (Ontario Government, 2007) en tant que sous-espèce du loup gris (C. lupus lycaon) et en tant qu'espèce préoccupante. La cote infranationale (S) de NatureServe est S4 pour l'Ontario.
En vertu de la Loi de 1997 sur la protection du poisson et de la faune (Ontario Government, 1997), tout individu de l'espèce Canis lupus est considéré comme un mammifère à fourrure et est protégé par la Loi. La chasse au loup et le piégeage du loup sont réglementés aux termes de la Loi, et la récolte est contrôlée et surveillée. En plus d'un permis de chasse au petit gibier, un sceau de gibier pour le loup ou le coyote (maximum de 2 par année) est nécessaire pour chasser le loup dans la partie principale de son aire de répartition (unités de gestion de la faune 1A, 1C, 1D, 2-42, 46-50 et 53-58). Les loups peuvent aussi être chassés ou piégés en vertu de permis de piégeage. La Loi prescrit les pouvoirs nécessaires pour limiter la récolte par piégeage, le cas échéant.
Lorsque la chasse et le piégeage réglementés sont permis, la saison de chasse et de piégeage du loup et du coyote est suspendue du 1er avril au 14 septembre dans la zone principale de l'aire de répartition définie plus haut. Les loups (ainsi que les coyotes) peuvent être chassés et piégés toute l'année au sud de la zone principale de l'aire de répartition du loup, et aucune limite de récolte n'a été fixée. Cette zone est considérée comme étant à l'extérieur de la zone d'occurrence actuelle du loup de l'Est, mais à l'intérieur de son aire de répartition historique.
Il n'y a pas de saison de chasse au loup ou de piégeage du loup dans les 40 comtés suivants, qui sont voisins du parc Algonquin : Airy, Alice, Ballantyne, Boulter, Boyd, Burns, Butt, Calvin, Cameron, Chisholm, Clancy, Clara, Dickens, Dudley, Eyre, Finlayson, Franklin, Fraser, Hagarty, Harburn, Harcourt, Havelock, Head, Herschel, Lauder, Livingstone, Maria, McClintock, McClure, McCraney, McKay, Murchison, Papineau, Paxton, Petawawa, Richards, Rolph, Sabine, Sinclair et Wylie.
Il est interdit de chasser ou de piéger le loup dans les aires protégées suivantes : parc Pukaskwa, parc Algonquin et réserves de chasse de la Couronne. Le loup est protégé contre la chasse réglementée dans tous les autres parcs provinciaux (à l'exception du parc provincial Point Farms, dans le sud de l'Ontario, qui se trouverait à l'extérieur de la zone principale de l'aire de répartition), et il est protégé contre le piégeage dans plus de la moitié des parcs provinciaux. Le piégeage du loup est permis dans les parcs Kawartha Highlands et French River. Dans le cadre d'une politique d'abandon graduel du piégeage, il sera interdit de piéger le loup dans un tiers des autres parcs où la pratique du piégeage est permise actuellement.
Les chasseurs et les piégeurs sont tenus de remplir une déclaration obligatoire des activités et des récoltes. Une personne qui tue un loup en voulant protéger sa propriété doit enregistrer la carcasse en ligne si elle a l'intention de la conserver, en soumettant un avis de possession, ou signaler immédiatement la mort de l'animal au Ministère si elle n'a pas l'intention de conserver la carcasse.
Les résidents et les non-résidents doivent détenir une licence d'exportation de l'Ontario pour les mammifères à fourrure et leurs peaux s'ils veulent exporter un loup ou un coyote, ou leur peau, depuis l'Ontario, que l'animal provienne ou non de l'Ontario, et incluant les carcasses ainsi que les loups et coyotes vivants. Des redevances sur les loups et coyotes de l'Ontario sont payables avant leur exportation de l'Ontario.
La Stratégie pour la conservation des loups en Ontario (OMNR, 2005) présente l'orientation politique stratégique en matière de gestion des loups dans la province.
Les tanières et les lieux de rendez-vous du loup de l'Est sur les terres de la Couronne sont protégés, car ils sont visés par les lignes directrices en matière de gestion des forêts de la province (OMNR, 2010).
Au Québec, les loups sont considérés comme des animaux à fourrure et sont protégés en vertu de la Loi sur la conservation et la mise en valeur de la faune, mais ne sont pas protégés en vertu de la Loi sur les espèces menacées ou vulnérables. Le loup de l'Est n'est pas reconnu officiellement, parce que la province ne reconnaît pas les sous-espèces de loups (MFFP, 2011); la Loi sur la conservation et la mise en valeur de la faune reconnaît seulement la nomenclaturetaxinomique du Smithsonian Museum of Natural History et du Système d'information taxinomique intégré [en anglais seulement] (MFFP, 2012a). Actuellement, la chasse au loup et le piégeage du loup sont interdits dans tous les parcs fédéraux et nationaux (provinciaux), mais sont permis ailleurs dans la province, notamment dans les réserves fauniques (St. Louis, comm. pers.). Dans la plupart des régions, la saison de récolte va de la fin d'octobre à la fin de mars, et il n'existe aucune limite de prises.
NatureServe (http://www.natureserve.org/explorer/) estime que le loup de l'Est est une sous-espèce du loup gris (Canis lupus lycaon) et lui a attribué la cote G4G5TNR (apparemment non en péril/non en péril) et la cote N4 à l'échelle nationale. À l'échelle provinciale, le gouvernement du Québec n'a pas évalué le loup de l'Est, et le gouvernement de l'Ontario lui a attribué la cote S4 - apparemment non en péril.
La mortalité causée par l'activité humaine constitue la caractéristique la plus importante de l'habitat du loup de l'Est. Dans les zones occupées actuellement en Ontario, les superficies d'habitat où il n'y a pas de récolte de loup de l'Est comprennent le parc Algonquin (7 571 km2) et les comtés environnants (6 340 km2), les Queen Elizabeth II Wildlands (255 km2 et le parc Killarney (645 km2), ce qui représente une superficie totale de 14 811 km2. La récolte du loup est permise dans le parc Kawartha Highlands (376 km2) et le parc French River (735 km2), mais ces parcs se trouvent dans des secteurs où l'activité humaine (immeubles, routes, etc.) est relativement limitée.
Au Québec, bien qu'il existe des réserves fauniques et des ZEC (zones d'exploitation contrôlées) (p. ex. Papineau-Labelle [1 628 km2], Rouge-Matawin, Mastigouche, Saint-Maurice, Portneuf et des Laurentides), qui sont moins aménagées que d'autres régions de la province et qui contiennent probablement un plus grand nombre de proies, ces réserves fauniques visent généralement les activités de prélèvement, comme la chasse et le piégeage. On ignore donc le rôle que ces réserves pourraient jouer pour réduire au minimum l'introgression des gènes par le coyote de l'Est. La récolte de canidés est interdite dans le parc du Mont-Tremblant (1 510 km2) et dans le parc de la Mauricie (536 km2).
Nous remercions Jenny Wu, d'Environnement Canada, d'avoir créé les figures cartographiques du rapport et Graham Forbes (Ph.D.) d'avoir supervisé la rédaction du rapport et fourni des commentaires sur ses versions préliminaires. Nous remercions également les personnes et organismes suivants qui ont contribué au rapport en fournissant de l'information ou des commentaires :
Maria de Almeida, Section des politiques de la faune, ministère des Richesses naturelles et des Forêts de l'Ontario, Peterborough (Ontario).
Jeff Bowman, ministère des Richesses naturelles et des Forêts de l'Ontario, Peterborough (Ontario).
Robert Craig, analyste de l'information, Centre d'information sur le patrimoine naturel, ministère des Richesses naturelles de l'Ontario (Ontario).
Marie-France Dalcourt, gestionnaire par intérim, section Conservation des populations, Service canadien de la faune - Région du Québec.
Dave Duncan, directeur régional, Service canadien de la faune, région des Prairies et du Nord, Environnement Canada (Manitoba).
Isabelle Gauthier, biologiste et coordonnatrice provinciale, espèces fauniques menacées et vulnérables, ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec (Québec).
Hélène Jolicoeur, retraitée du ministère des Ressources naturelles et de la Faune (MFFP) du Québec.
Christopher Kyle, professeur adjoint, Trent University, Peterborough (Ontario).
Michel Henault, ministère des Forêts, de la Faune et des Parcs, Direction de la gestion de la faune de Lanaudière et des Laurentides.
Briar Howes, biologiste de l'habitat essentiel, Espèces en péril, Parcs Canada.
Hank Hristienko, Conservation Manitoba, Direction de la protection de la faune et des écosystèmes (Manitoba).
Henry Lickers, directeur de l'environnement, Conseil des Mohawks d'Akwesasne (Ontario/Québec).
Angela McConnell, Service canadien de la faune, Ottawa (Ontario).
Patrick Nantel, biologiste de la conservation, Programme des espèces en péril, Parcs Canada (Québec).
Dean Nernberg, agent des espèces en péril, Defense nationale.
Ronald Nowak, retraité du US Fish and Wildlife Service.
Michael Oldham, Centre d'information sur le patrimoine naturel, ministère des Richesses naturelles de l'Ontario (Ontario).
Sylvain Paradis, coordonnateur, Espèces en péril, Parcs Canada (Québec).
Brent Patterson, Section de recherche-développement en matière de faune, ministère des Richesses naturelles et des Forêts de l'Ontario (Ontario).
Annie Paquet, Centre de données sur le patrimoine naturel du Québec, Direction de l'expertise sur la faune et ses habitats, ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec.
Jason Pitre, coordonnateur des CTA, Secrétariat du COSEPAC, Service canadien de la faune.
Chris Risley, biologiste des espèces d'oiseaux et de mammifères en péril, Direction des espèces en péril, ministère des Richesses naturelles de l'Ontario (Ontario).
Brad Steinberg, biologiste de la gestion des ressources, parc Algonquin (Ontario).
Antoine St.-Louis, biologiste des espèces menacées, ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec.
Astrid vik Stronen, BIOCONSUS Fellow, Mammal Research Institute, Pologne.
Rick Stronks, naturaliste en chef, parc Algonquin (Ontario).
Richard Thiel, Retired Wildlife Biologist, Wisconsin Department of Natural Resources.
Mario Villemure, technicien en conservation des ressources, parc national de la Mauricie (Québec).
Bradley White, Chaire de biologie, Trent University, Peterborough (Ontario).
Paul Wilson, Chaire de recherche du Canada en analyse des empreintes génétiques, Trent University, Peterborough (Ontario).
Adrian Wydeven, Mammalian Ecologist/Conservation Biologist, Wisconsin Department of Natural Resources.
Argue, A.M., Mills, K.J. et B.R. Patterson. 2008. Behavioural response of eastern wolves (Canis lycaon) to disturbance at homesites and its effects on pup survival. Canadian Journal of Zoology 86:400-406.
Bath, A.J. 2006. Human dimensions in wolf conservation: understanding trapper's perceptions toward wolves in and around La Mauricie National Park of Canada, Rapport présenté au Centre de services de Parcs Canada au Québec, 96 pp.
Benson, J. F., Mills, K.J., Loveless, K.M. et B.R. Patterson. 2013b. Genetic and environmental influences on pup mortality risk for wolves and coyotes within a Canis hybrid zone. Biological Conservation. 166:133-141.
Benson, J.F. et B. R. Patterson. 2013a. Inter-specific territoriality in a Canis hybrid zone: spatial segregation between wolves, coyotes, and hybrids. Oecologia. 173:1539-1550.
Benson, J.F., Patterson, B.R. et P.J. Mahoney. 2014. A protected area influences genotype-specific survival and the structure of a Canis hybrid zone. Ecology 95:254-264.
Benson, J.F., Patterson, B.R. et T.J. Wheeldon. 2012. Spatial genetic and morphologic structure of wolves and coyotes in relation to environmental heterogeneity in a Canis hybrid zone. Molecular Ecology 21:5934-5954.
Bertorelle, G. et L. Excoffier 1998. Inferring admixture proportions from molecular data. Molecular Biology and Evolution 15:1298-1311.
Beschta, R.L. et W.J. Ripple. 2009. Large predators and trophic cascades in terrestrial ecosystems of the western United States. Biological Conservation. 142:2401-2414.
Beschta, R.L. et W.J. Ripple. 2010. Recovering riparian plant communities with wolves in northern Yellowstone, USA. Restoration Ecology 18:380-389.
Brainerd, S.M. (et 6 co-auteurs). 2008. The effects of breeder loss on wolves. Journal of Wildlife Management 72:89-98.
Bruskotter, J.T., Schmidt, R.H. et T.L. Teel. 2007. Are attitudes toward wolves changing? A case study in Utah. Biological Conservation 139:211-218.
Bruskotter, J.T., Toman, E., Enzler, S.A. et R.H. Schmidt. 2011. Gray wolves not out of the woods yet. Science 327:30-31.
Buss, M. et M. deAlmeida. 1997. A review of wolf and coyote status and policy in Ontario. Ontario Ministry of Natural Resources. 88 pp.
Carbone, C., Mace, G.M., Roberts, S.C. et D.W. Macdonald. 1999. Energetic constraints on the diet of terrestrial carnivores. Nature 402:286-288.
CEC (Commission for Environmental Cooperation) 1997. Ecological regions of North America: a common perspective. Montreal, QC. (Également disponible en français : CCE (Commission de coopération environnementale). 1997. Les régions écologiques de l'Amérique du Nord : vers une perspective commune. Montréal, QC).
Chambers, S., S. Fain, B. Fazio et M. Amaral. 2012. An account of the taxonomy of North American wolves from morphological and genetic analyses. North American Fauna 77:1-67. USFWS. Doi:10.3996/nafa.77.0001.
Chapron, G., Andren, H. et O. Liberg. 2008. Conserving top predators in ecosystems. Science. 320: 47.
Coordonnateur de l'équipe de rétablissement des oiseaux de proie du Québec (ÉROP). MFFP, Québec, QC.
COSEWIC (Committee on the Status of Endangered Wildlife in Canada). 2014. Operations and Procedures Manual, April 2014. COSEWIC Secretariat. COSEWIC Guidelines on Manipulated Populations. [consulté en février 2015]. (Également disponible en français : COSEPAC (Comité sur la situtation des espèces en péril au Canada). 2014. Manuel des opérations et des procédures, avril 2014. Secrétariat du COSEPAC. Lignes directrices du COSEPAC concernant les populations manipulées).
Crête, M. et S. Larivière. 2003. Estimating the costs of locomotion in snow for coyotes. Canadian Journal of Zoology 81:1808-1814.
Cronin, M.A. et L.D. Mech. 2009. Problems with the claim of ecotype and taxon status of the wolf in the Great Lakes region. Molecular Ecology 18:4991-4993.
DelGiudice, G.D., Riggs, M.R., Joly, P. et W. Pan. 2002. Winter severity, survival, and cause-specific mortality of female white-tailed deer in north-central Minnesota. Journal of Wildlife Management. 66:698-717.
Dowling, T. (et 4 co-auteurs). 1992. Response to Wayne, Nowak, and Phillips and Henry: Use of molecular characteristics in conservation biology. Conservation Biology 6:600-603.
Fain, S.R., Straughan, D.J. et B.F. Taylor. 2010. Genetic outcomes of wolf recovery in the western Great Lakes states. Conservation Genetics 11:1747-1765.
Fenech, A., Taylor, B., Hansell, R. et G. Whitelaw. 2000. Major road changes in southern Ontario 1935-1995: Implications for protected areas. Integrated Mapping Assessment Project. University of Toronto. Website: http://www.utoronto.ca/imap/papers/road_changes.htm [consulté le 9 janvier 2011]
Fogelqvist J., Niittyvuopio, A., Ågren J., Savolainen O. et M. Lascoux. 2010. Cryptic population genetic structure: the number of inferred clusters depends on samples size. Molecular Ecology Resources 10:314-323.
Forbes, G.J. et J.B. Theberge. 1992. Importance of scavenging on moose by wolves in Algonquin Park, Ontario. Alces 28:235-241.
Forbes, G.J. et J.B. Theberge. 1995. Influences of a migratory deer herd on wolf movements and mortality in and around Algonquin Park, Ontario. Pp. 303-314 in Ecology and Conservation of Wolves in a Changing World. L. Carbyn, S. Fritts et D. Seip (eds). Canadian Circumpolar Institute, University of Alberta, Edmonton, Alberta. 619 pp.
Forbes, G.J. et J.B. Theberge. 1996a. Cross-boundary management of Algonquin Park wolves. Conservation Biology 10:1091-1097.
Forbes, G.J. et J.B. Theberge. 1996b. Response of wolves to prey variation in central Ontario. Canadian Journal of Zoology 74:1511-1520.
Forbes, Graham, comm. Pers. 26 juillet 2012. Correspondance par courriel : commentaires sur le rapport de situation du COSEPAC sur le loup de l'Est (Canis lycaon).
Frame. P., D. Cluff et D. Hik. 2007. Response of wolves to experimental disturbance at homesites. Journal of Wildlife Management 71:313-320.
Fryxell, J.M. 2001. Habitat suitability and source-sink dynamics of beavers. Journal of Animal Ecology 70:310-316.
Fuller, T.K., Mech, D.L. et J.F. Cochrane. 2003. Wolf population dynamics. Pp. 161-191 in Wolves: Behavior, Ecology, and Conservation. D.L. Mech and L. Boitani (eds). University of Chicago Press, Chicago.
Fur Harvesters Auction Inc. 2011. Sale Results for January 7, 2011. (PDF; 43.7 KB) .
Garcia-Moreno J., M. Matocq, M. Roy, E. Geffen et R. Wayne. 1996. Relationships and genetic purity of the endangered Mexican wolf based on analysis of microsatellite loci. Conservation Biology 10:376-389.
Goldman, E. 1937. The wolves of North America. Journal of Mammalogy 18:37-45.
Grewal, S.K., Wilson, P.J., Kung, T.K., Shami, K., Theberge, M.T., Theberge, J.B. et B.N. White. 2004. A genetic assessment of the eastern wolf (Canis lycaon) in Algonquin Provincial Park. Journal of Mammalogy 85:625-632.
Hailer F. et J.A. Leonard. 2008. Hybridisation among three native North American Canis species in a region of natural sympatry. PLoS ONE 3(10): e3333. doi:10.1371/journal.pone.0003333.
Harrison, D.J. et T.G. Chapin 1998. Extent and connectivity of habitat for wolves in eastern North America. Wildlife Society Bulletin 26:767-775.
Hedrick P., P. Miller, E. Geffen et R. Wayne. 1997. Genetic evaluation of three captive Mexican wolf lineages. Zoological Biology 16:47-69.
Hénault, M. Données inédites. Information contained in Québec jurisdictional review of Eastern Wolf Report. February 2015.
Hénault, M. et H. Jolicouer. 2003. Les loups au Québec: Meutes et mystères. Société de la faune et des Parcs du Québec, Direction de l'aménagement de la région des Laurentides, Direction du développement de la faune, 129 p.
Hubisz, M.J., Falush, D., Stephens, M., Pritchard, J.K. 2009. Inferring weak population structure with the assistance of sample group information. Molecular Ecology Resources 9:1322-1332.
Jolicoeur, H. et Hénault, M. 2002. Répartition géographique du loup et du coyote au sud du 52e parallèle et estimation de la population de loups au Québec. Société de la faune et des parcs du Québec. ISBN: 2-550-39485-2.
Jolicoeur, H. et M. Hénault. 2010. Current status and management of Wolves in Québec. Wolf Stewards Meeting, Dorset, Ontario.
Jost, L. 2008. Gst and its relatives do not measure differentiation. Molecular Ecology 17:4015-4026.
Kays, R., Curtis, A. et J.J. Kirchman. 2010. Rapid adaptive evolution of northeastern coyotes via hybridisation with wolves. Biology Letters 6:89-93.
Kennedy, L.J. 2012. pers. comm. Email correspondence to L Rutledge January 9, 2012. Senior Scientist, University of Manchester, Centre for Integrated Genomic Medical Research, Manchester, United Kingdom.
Koblmüller, S., Nord, M., Wayne, R.K., Leonard, J.A. 2009. Origin and status of the Great Lakes wolf. Molecular Ecology. 11:2313-2326.
Kolenosky, G. et R. Standfield. 1975. Morphological and ecological variation among Gray wolves (Canis lupus) of Ontario, Canada. Pp 62-72. in M. Fox (ed.). The Wild Canids. Van Nostrand Reinhold: New York.
Kurten, B. et E. Anderson, 1980. Pleistocene mammals of North America. Columbia Press, NY.
Kyle, C.J., Johnson, A.R., Patterson, B.R., Wilson, P.J. et B.N. White. 2008. The conspecific nature of eastern and red wolves: conservation and management implications. Conservation Genetics. 9:699-701.
Kyle, C.J., Johnson, A.R., Patterson, B.R., Wilson, P.J., Shami, K., Grewal, S.K. et B.N. White. 2006. Genetic nature of eastern wolves: past, present and future. Conservation Genetics 7:273-287.
Lancaster, P.A., Bowman, J. et B.A. Pond. 2008. Fishers, farms, and forests in eastern North America. Environmental Management 42:93-101.
Lankester, M.W. 2010. Understanding the impact of meningeal worm, Parelaphostrongylus tenuis, on moose populations. Alces 46:53-70.
Lehman, N. (et 6 co-auteurs). 1991. Introgression of coyote mitochondrial DNA genotype frequencies: estimation of the effective number of alleles. Evolution 45:104-119.
Leonard, J.A. et R.K. Wayne. 2005. Legacy lost: genetic variability and population size of extirpated US Gray wolves (Canis lupus). Molecular Ecology 14:9-17.
Leonard, J.A. et R.K. Wayne. 2008. Native Great Lakes wolves were not restored. Biology Letters 4:95-98.
Lesmarises, F., C. Dussault et M-H. St-Laurent. 2012. Wolf habitat selection is shaped by human activities in a highly managed boreal forest. Forest Ecology and Management 276:125-131.
Lickers, H. Email correspondence with G. Goulet, TMSSC Member, COSEWIC. April 2015.
Linnell, J.D.C., Andersen, R., Andersone, Z. et al. 2002. The fear of wolves: A review of wolf attacks on humans. NINA Oppdragsmelding: 731: 1-65. PDF
Lohr, C. et W.B. Ballard. 1996. Historical occurrence of wolves, Canis lupus in the Maritime Provinces. Canadian Field-Naturalist 110:607-610.
Loveless, K. 2010. Foraging strategies of eastern wolves in relation to migratory prey and hybridisation. Masters thesis. Trent University, Peterborough, ON. 73 pp.
MacNulty, D.R., Smith, D.W., Mech, L.D. et L.E. Eberly. 2009. Body size and predatory performance in wolves: is bigger better? Journal of Animal Ecology 78:532-539.
Manning, R. et W. Sturtevant 1966. Early specimens of the eastern wolf, Canis lupus lycaon. Chesapeake Science 7:218-220.
McCarley, H. et J. Carley. 1979. Recent changes in distribution and status of red wolves (Canis rufus). Endangered Species Report no.4. U.S. Fish and Wildlife Service, Albuquerque, NM.
Mech, D., S. Fritts et G. Radde. 1988. Wolf distribution and road density in Minnesota. Wildlife Society Bulletin 16:85-87.
Mech, L.D. (et 4 co-auteurs). 2014. Production of hybrids between western gray wolves and western coyotes. PloSONE 9(2):e88861.doi:10.1371.
Mech, L.D. 2010a. What is the taxonomic identity of Minnesota wolves? Canadian Journal of Zoology 88:129-138.
Mech, L.D. 2010b. Considerations for developing wolf harvest regulations in the contiguous United States. Journal of Wildlife Management 74:1421-1424.
Mech, L.D. 2011. Non-genetic data supporting genetic evidence for the eastern wolf. Northeastern Naturalist 18:521-526.
Mech, L.D. et L. Boitani. 2003. Wolf social ecology. Pp. 1-34. in D.L. Mech et L. Boitani (eds.). Wolves: Behavior, Ecology and Conservation. University of Chicago Press, Chicago.
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2011.
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2012a.
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2012b. Site Web : Énergie et Ressources naturelles [consulté le 8 décembre 2011].
MFFP (Ministère des Forêts, de la Faune et des Parcs). 2012c. Web site: Trapping statistics (en anglais seulement) [consulté le 20 août 2012].
Miller, G. 1912. The names of two North American wolves. Proceedings Biological Society of Washington. 25:93.
Mills, K.J. 2006. Wolf (Canis lycaon) pup survival, dispersal, and movements in Algonquin Provincial Park, Ontario. Mémoire de maîtrise. Trent University, Peterborough, ON, 67 pp.
Mills, K.J., Patterson, B.R. et D.L. Murray. 2008. Direct estimation of early survival and movements in eastern wolf pups. Journal of Wildlife Management 72:949-954.
Murray, D.L. (et 7 co-auteurs). 2006. Pathogens, nutritional deficiency, and climate influences on a declining moose population. Wildlife Monographs 166:1-20.
Murray, D.L. et L.P. Waits. 2008. Taxonomic status and conservation strategy of the endangered red wolf: a response to Kyle et al. (2006). Conservation Genetics 8:1483-1485.
Musiani, M. et P.C. Paquet. 2004. The practices of wolf persecution, protection, and restoration in Canada and the United States. BioScience 54:50-60.
Musiani, M., L. Boitani et P. Paquet. 2010. The World of Wolves: New Perspectives on Ecology, Behaviour and Management. University of Calgary Press. 398 pp.
Naughton, D. 2012. The Natural History of Canadian Mammals. University of Toronto Press, Toronto, ON. 784 pp.
NCEAS (National Center for Ecological Analysis and Synthesis). 2014. Review of the proposed rule regarding status of the wolf under the ESA. Santa Barbara, CA. 68 pp.
Norris, D.R., Theberge, M.T. et J.B. Theberge. 2002. Forest composition around wolf (Canis lupus) dens in eastern Algonquin Provincial Park, Ontario. Canadian Journal of Zoology 80:866-872.
Nowak, R.M. 1979. North American Quaternary Canis. Museum of Natural History, University of Kansas (Monograph No. 6), Lawrence, Kansas.
Nowak, R.M. 1992. The red wolf is not a hybrid. Conservation Biology 6:593-595.
Nowak, R.M. 1995. Another look at wolf taxonomy. Pp. 375-398, in L.N. Carbyn, S.H. Fritts and D.R. Seip (eds.). Ecology and Conservation of Wolves in a Changing World. Canadian Circumpolar Institute, Occasional Publication No. 35, 642 pp.
Nowak, R.M. 2002. The original status of wolves in eastern North America. Southeastern Naturalist 1(2):95-130.
Nowak, R.M. 2009. Taxonomy, morphology, and genetics of wolves in the Great Lakes region. in A.P. Wydeven, T.R. vanDeelen, and E. Heske (eds.), Recovery of Gray Wolves in the Great Lakes Region of the United States. Springer Publishing, 350 pp.
Nowak, R.M., comm. pers. Correspondance adressée à L. Rutledge en août 2011. Taxonomiste des loups, retraité de l'US Fish and Wildlife Service.
OMNR (Ontario Ministry of Natural Resources). 2005. Strategy for Wolf Conservation in Ontario, Ontario Ministry of Natural Resources, report. http://www.ontario.ca/environment-and-energy/wolf-conservation-strategy. [consulté en février 2015]. (Également disponible en français : MRNO (Ministère des Richesses naturelles de l'Ontario). 2005. Stratégie pour la conservation des loups en Ontario. http://www.ontario.ca/fr/document/strategie-pour-la-conservation-des-loups.
OMNR (Ontario Ministry of Natural Resources). 2009. Cervid Ecological Framework. http://www.ontario.ca/environment-and-energy/cervid-ecological framework. [consulté en février 2015]. (Également disponible en français : MRNO (Ministère des Richesses naturelles de l'Ontario). 2009. Cadre de gestion écologique des cervidés. http://www.ontario.ca/fr/document/cadre-de-gestion-ecologique-des-cervides).
OMNR (Ontario Ministry of Natural Resources). 2010. Forest Management Guide for Conserving Biodiversity at the Stand and Site Scales. Queen's Printer, Toronto, ON.
OMNR (Ontario Ministry of Natural Resources). 2012. Wildlife Management Unit Maps. Find a wildlife management unit (WMU) map [consulté en février 2015]. (Également disponible en français : MRNO (Ministère des Richesses naturelles de l'Ontario). 2012. Cartes des unités de gestion de la faune. Trouver une carte des unités de gestion de la faune (UGF)).
Ontario Government, Endangered Species Act, 2007, S.O. 2007, c. 6,. (Également disponible en français : Gouvernement de l'Ontario, Loi de 2007 sur les espèces en voie de disparition, L.O. 1997, ch. 6, ).
Ontario Government, Fish and Wildlife Conservation Act, 1997, S.O. 1997, c. 41. (Également disponible en français : Gouvernement de l'Ontario, Loi de 1997 sur la protection du poisson et de la faune, L.O. 1997, ch. 41,).
Paquet, Annie, comm. pers.Correspondance par courriel adressée à L. Rutledge le13 mai 2011. Technicienne de la faune. Service de la biodiversité et des maladies de la faune, Québec.
Parcs Canada. 2007. Étude sur les attitudes, perceptions et connaissances des résidents de la région limitrophe du parc national du Canada de la Mauricie à l'égard du loup de l'Est. Rapport final, Mars 2007. Numéro de contrat: 5P201-060514 - POR-273-06.
Parker, G. 1995. Eastern Coyote: The Story of its Success. Nimbus Publishing, Halifax, NS. 254 pp.
Patterson, B.R. et D.L. Murray. 2008. Flawed population viability analysis can result in misleading population assessment: a case study for wolves in Algonquin Park, Canada. Biological Conservation 141:669-680.
Patterson, B.R., comm. pers. 2011a. Correspondance par courriel adressée à L. Rutledge le 14 octobre 2011. Chercheur scientifique, ministère des Richesses naturelles de l'Ontario, Trent University, Peterborough (Ontario).
Patterson, B.R., comm. pers. 2011b. Correspondance par courriel adressée à L. Rutledge le21 novembre 2011. Chercheur scientifique, ministère des Richesses naturelles de l'Ontario, Trent University, Peterborough (Ontario).
Patterson, B.R., comm. pers. 2011c. Correspondance par courriel adressée à L. Rutledge le 18 décembre 2011. Chercheur scientifique, ministère des Richesses naturelles de l'Ontario, Trent University, Peterborough (Ontario)..
Patterson, B.R., comm. pers. 2012. Correspondance par courriel (commentaire sur l'ébauche du rapport) adressée à L. Rutledge le 12 janvier 2012. Chercheur scientifique, ministère des Richesses naturelles de l'Ontario, Trent University, Peterborough (Ontario).
Patterson, B.R., comm. pers. 2012b. Correspondance par courriel adressée à L. Rutledge le 1er février 2012. Chercheur scientifique, ministère des Richesses naturelles de l'Ontario, Trent University, Peterborough (Ontario).
Patterson, B.R., comm. pers. 2014. Correspondance par courriel adressée à L. Rutledge le 2 juillet 2014. Chercheur scientifique, ministère des Richesses naturelles de l'Ontario, Trent University, Peterborough (Ontario).
Person, D. et A. Russell. Correlates of wolf mortality in an exploited wolf population. Journal of Wildlife Management 72:1540-1549.
Peterson, R.O., Thomas, N.J., Thurber, J.M., Vucetich, J.A. et T.A. Waite. 1998. Population limitation and the wolves of Isle Royale. Journal of Mammalogy 79:828-841.
Phillips, M. et V. Henry. 1992. Comments on red wolf taxonomy. Conservation Biology 6:596-599.
Pilgrim KL, Boyd DK et Forbes SH (1998) Testing for wolf-coyote hybridization in the Rocky Mountains using mitochondrial DNA. Journal of Wildlife Management 62:683-689.
Pimlott, D.H., Shanon, J.A. et G.B. Kolenosky. 1969. The ecology of the timber wolf in Algonquin Provincial Park. Report No. 87. Ontario Ministry of Natural Resources, Toronto, Ontario, Canada.
Pocock, R. 1935. The races of Canis lupus. Proceedings Zoological Society of London. Part 3; 647-686.
Potvin, F. 1988. Wolf movements and population dynamics in Papineau-Labelle Reserve, Québec. Canadian Journal of Zoology 66:1266-1273.
Potvin, F., H. Jolicoeur et J. Huot. 1988. Wolf diet and prey selectivity during two periods for deer in Québec: decline versus expansion. Canadian Journal of Zoology 66:1274-1279.
Potvin, F., H. Jolicoeur, L. Breton et R. Lemieux. 1992a. Evaluation of an experimental wolf reduction and its impact on deer in Papineau-Labelle Reserve, Québec. Canadian Journal of Zoology 70:1595-1603.
Potvin, F., L. Breton, C. Pilon et M. Mactuart. 1992b. Impact of an experimental wolf reduction on beaver in Papineau-Labelle. Reserve, Québec. Canadian Journal of Zoology 70:180-183.
Quinn, N. 2004. The presettlement hardwood forests and wildlife of Algonquin Provincial Park: A synthesis of historic evidence and recent research. Forestry Chronicle 80:705-717.
Quinn, N. 2005. Reconstructing changes in abundance of White-tailed Deer, Odocoileus virgnianus, Moose, Alces alces, and Beaver, Castor Canadensis, in Algonquin Park, Ontario, 1860-2004. Canadian Field-Naturalist 119:330-342.
Reich, R. Wayne et D. Goldstein. 1999. Genetic evidence for a recent origin by hybridization of red wolves. Molecular Ecology 8:139-144.
Rempel, R.S. 2011. Effects of climate change on moose populations: exploring the response horizon through biometric and systems models. Ecological Modelling 222:3355-3365.
Rogic, A., N. Tessier et F.-J. Lapointe. 2014. Identification of canids found within Parc national du Mont-Tremblant and its surroundings using microsatellite markers. University of Montreal, Ministère des Forêts, de la Faune et des Parcs. 30 p. (Également disponible en français : Rogic, A., N. Tessier et F.-J. Lapointe. 2014. Identification des canidés du Parc national du Mont-Tremblant et de sa périphérie à l'aide de marqueurs microsatellites. Université de Montréal, Ministère des Forêts, de la Faune et des Parcs. 30 p.)
Roy M., E. Geffen, D. Smith et R. Wayne. 1996. Molecular genetics of pre-1940 red wolves. Conservation Biology 10:1413-1424.
Roy M., E. Geffen, D. Smith, E. Ostrander et R. Wayne. 1994. Patterns of differentiation and hybridization in North American Wolf-like Canids, revealed by analysis of microsatellite loci. Molecular Biology and Evolution 11:553-570.
Rutledge L. et B. White. 2013. Genetic characterization of Canis in Québec. Contrat de services abrégé 111720000. Final Report. 10 p.
Rutledge L. et B. White. 2014. Genetic characterization of Canis in Québec. Contrat de services abrégé 111720000. Final Report Addendum.14 p.
Rutledge, L.Y. 2010a. Evolutionary origins, social structure, and hybridisation of the eastern wolf (Canis lycaon). Thèse de doctorat, Trent University, Peterborough, Ontario, Canada. 187 pp.
Rutledge, L.Y., Bos, K.I., Pearce, R.J., White, B.N. 2010d. Genetic and morphometric analysis of sixteenth century Canis skull fragments: implications for historical eastern and Gray wolf distribution in North America. Conservation Genetics. 11:1273-1281.
Rutledge, L.Y., Desy, G., Fryxell J., Patterson, B.R. et B.N. White. En préparation. Distribution of eastern wolves refined with non-invasive sampling. Canadian Journal of Zoology.
Rutledge, L.Y., Devillard S., Boone, J.Q., Hohenlohe, P.A. et B.N. White. In Review. RAD sequencing and genomic simulations resolve hybrid origins within North America Canis. Biology Letters.
Rutledge, L.Y., Garroway, C.J., Loveless, K.M. et B.R. Patterson. 2010c. Genetic differentiation of eastern wolves in Algonquin Park despite bridging gene flow between coyotes and Gray wolves. Heredity 105:520-531.
Rutledge, L.Y., Patterson B.R. et B.N. White. 2010b. Analysis of Canis mitochondrial DNA demonstrates high concordance between the control region and ATPase genes. BMC Evolutionary Biology 10: 215.
Rutledge, L.Y., Patterson, B.R., Mills, K.J., Loveless, K.M., Murray, D.L. et B.N. White. 2010e. Protection from harvesting restores the natural social structure of eastern wolf packs. Biological Conservation 143:332-339.
Rutledge, L.Y., White, B.N., Row, J.R. et B.R. Patterson. 2011. Intense harvesting of eastern wolves facilitated hybridisation with coyotes. Ecology and Evolution DOI: 10.1002/ece3.61.
Rutledge, L.Y., Wilson, P.J., Klütsch, C.F.C., Patterson, B.R. et B.N. White. 2012. Conservation genomics in perspective: a holistic approach to understanding Canis evolution in North America. Biological Conservation 155:186-192.
Samson, C. 2001. COSEWIC Assessment Update Status Report of the Eastern Wolf (Canis lupus lycaon) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 17 pp.
Sand, H., Wikenros, C., Wabakken, P. et O. Liberg. 2006. Effects of hunting group size, snow depth and age on the success of wolves hunting moose. Animal Behaviour 72:781-789.
Schmidt, K., Jędrzejewski, W., Theuerkauf, J., Kowalczyk, R., Okarma, H. et B. Jędrzejewski. 2008. Reproductive behaviour of wild-living wolves in Bialowieza Primeval Forest (Poland). Journal of Ethology 26:69-78.
Schmitz, O.J. et G.B. Kolenosky. 1985. Wolves and coyotes in Ontario - morphological relationships and origins. Canadian Journal of Zoology 63:1130-1137.
Schreber, J. 1775. Die Säughthiere in Abbildungen nach der Natur mit Beschreibungen. Sweeter Theil. Tafeln. Erlangen. (Walther). Vol. 3, 353, plate 89. Erlangen.
Schwartz, M.K. et J.A. Vucetich. 2009. Molecules and beyond: assessing the distinctness of the Great Lakes wolf. Molecular Biology 18:2307-2309.
Sears, H.J., Theberge, J.B., Theberge, M.T., Thornton, I. et G.D. Campbell. 2003. Landscape influence on Canis morphological and ecological variation in a coyote-wolf C. lupus x latrans hybrid zone, southeastern Ontario. Canadian Field-Naturalist 117:589-600.
St. Louis, Antoine, comm. pers. 2012. Correspondance par courriel adressée à L. Rutledge le 6 août 2012. Biologiste des espèces menacées.
Stahler, D.R., Smith, D.W. et D.S. Guernsey. 2006. Foraging and feeding ecology of the Gray wolf (Canis lupus): lessons from Yellowstone National Park, Wyoming, USA. Journal of Nutrition 26: 1923S.
Steinberg, B. comm. pers. 2011. Correspondance par courriel adressée à L. Rutledge en mai 2011. Management Biologist, parc Algonquin (Ontario).
Stronen, A.V. (et 9 co-auteurs). 2012. Canid hybridization: contemporary evolution in human-modified landscapes. Ecology and Evolution 2:2128-2140.
Stronen, A.V. 2009. Dispersal in a Plain Landscape: Wolves in Southwestern Manitoba, Canada. Thèese de doctorat. University of New Brunswick, Fredericton, New Brunswick, Canada.
Stronen, A.V., Brook, R.K., Paquet, P.C., Mclachlan, S. 2007. Farmer attitudes toward wolves: implications for the role of predators in managing disease. Biological Conservation 135:1-10.
Stronen, A.V., Forbes, G.J., Sallows, T., Goulet, G., Musiani, M. et P.C. Paquet. 2010. Wolf body mass, skull morphology, and mitochondrial DNA haplotypes in the Riding Mountain National Park region of Manitoba, Canada. Canadian Journal of Zoology 88:496-507.
Tessier, N. 2015. Données inédites. Ministère des Forêts, de la Faune et des Parcs (MFFP) du Québec.
Theberge, J. et M. Theberge. 2004. The Wolves of Algonquin Park: A 12-Year Ecological Study. Publication Series Number 56, Department of Geogrpahy, University of Waterloo, Waterloo, Canada.
Theberge, J.B. 1991. Ecological classification, status and management of the Gray Wolf, Canis lupus, in Canada. The Canadian Field-Naturalist 105:459-463.
Theberge, J.B., Forbes, G.J., Barker, I.K. et T. Bollinger. 1994. Rabies in wolves of the Great-Lakes region. Journal of Wildlife Diseases 30:563-566.
Theberge, J.B., Theberge, M.T., Vucetich, J.A. et P.C. Paquet. 2006. Pitfalls of applying adaptive management to a wolf population in Algonquin Provincial Park, Ontario. Environmental Management 37:451-460.
Thompson, I.D., Fannigan, M.D., Wotton, B.M. et R. Suffling. 1998. The effects of climate change on landscape diversity: and example in Ontario Forests. Environmental Monitoring and Assessment 49:213-233.
Toews, D.P. et A. Brelsford. 2012. The biogeography of mitochondrial and nuclear discordance in animals. Molecular Ecology 16:3907-3930.
USFWS (United States Fish and Wildlife Service). 2015a. Endangered Species Act web page [en anglais seulement]. [consulté en février 2015].
USFWS (United States Fish and Wildlife Service). 2015b. Red Wolf recovery web page [en anglais seulement]. [consulté en février 2015].
USFWS, comm. pers. 2012. Correspondance par courriel adressée à L. Rutledge le 21 mai 2012.
Vähä J-P et C.R. Primmer. 2006. Efficiency of model-based Bayesian methods for detecting hybrid individuals under different hybridization scenarios and with different numbers of loci. Molecular Ecology 15:63-72.
Van Zyll de Jong, C.G. et L. Carbyn. 1999. COSEWIC Status Report on the Gray Wolf (Canis lupus) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 61 pp.
Veradri, A., V. Lucchini et E. Randi. 2006. Detecting introgressive hybridization between free-ranging domestic dogs and wild wolves (Canis lupus) by admixture linkage disequilibrium analysis. Molecular Ecology 15:2845-2855.
Villemure, M. 2003. Écologie et conservation du loup dans la région du parc national de la Mauricie. Mémoire de maîtrise. Université de Sherbrooke, Sherbrooke, Québec, Canada. 92 p.
Villemure, M. et H. Jolicoeur. 2004. First confirmed occurrence of a wolf, Canis lupus, south of the St. Lawrence River in over 100 years. Canadian Field-Naturalist 118:608-610.
Villemure, M. et M. Festa-Bianchet. 2002. Écologie du loup au parc national de la Mauricie. Rapport rédigé pour Parcs Canada. Université de Sherbrooke. Département de Biologie, Sherbrooke, Québec. 130 p.
Villemure, M., comm. pers. 2011. Correspondance par courriel adressée à L. Rutledge en août 2011. Technicien de la conservation des ressources, parc national de la Mauricie, Parcs Canada, St-Mathieu-du Parc (Québec).
vonHoldt, B.M. (et 18 co-auteurs). 2011. A genome-wide perspective on the evolutionary history of enigmatic wolf-like canids. Genome Research 21:1294-1305.
vonHoldt, B.M., Stahler, D.R., Smith, D.W., et al. 2008. The genealogy and genetic viability of reintroduced Yellowstone Gray wolves. Molecular Ecology 17:252-274.
Vucetich, J. et P. Paquet. 2000. The demographic population viability of Algonquin wolves. Report prepared for The Algonquin Wolf Advisory Committee.
Way, J.G. et J. T. Bruskotter. 2012. Additional considerations for gray wolf management after their removal from Endangered Species Act protections. Journal of Wildlife Management 76:457-461.
Way, J.G., Rutledge, L., Wheeldon, T. et B.N. White. 2010. Genetic characterization of eastern "coyotes" in eastern Massachusetts. Northeastern Naturalist.17:189-204.
Wayne, R. et N. Lehman. 1992. Mitochondrial DNA analysis of the Eastern Coyote: Origins and hybridization. Pp 9-22 in Ecology and Management of the Eastern Coyote. A. Boer (ed.). Wildlife Research Unit. Fredericton, NB. 194 pp.
Wayne, R. et S. Jenks. 1991. Mitochondrial DNA analysis supports extensive hybridization of the endangered red wolf (Canis rufus). Nature 351:565-568.
Wheeldon, T. 2009. Genetic characterization of Canis populations in the western Great Lakes region. Masters thesis, Trent University, Peterborough, Ontario, Canada. 120 pp.
Wheeldon, T. et B.N. White. 2009. Genetic analysis of historical western Great Lakes region wolf samples reveals early Canis lupus/lycaon hybridisation. Biology Letters 5:101-104.
Wheeldon, T.J., Patterson, B.R. et White, B.N. 2010. Sympatric wolf and coyote populations of the western Great Lakes region are reproductively isolated. Molecular Ecology. 19:4428-4440.
Wheeldon, T.J., Rutledge, L.Y., Patterson, B.R., White, B.N. et Wilson, P.J. 2013. Y-chromosome evidence supports asymmetric dog introgression into eastern coyotes. Ecology and Evolution. 3:3005-3020.
Whittington, J. (et 6 co-auteurs). 2011. Caribou encounters with wolves increase near roads and trails: a time-to-event approach. Journal of Applied Ecology 48:1535-1542.
Williams, C.K., Ericsson, G. et T.A. Heberlein. 2002. A quantitative summary of attitudes toward wolves and their reintroduction (1972-2000). Wildlife Society Bulletin 30:575-584.
Wilmers, C.C., Post, E. et A. Hastings. 2007. The anatomy of predator-prey dynamics in a changing climate. Journal of Animal Ecology 76:1037-1044.
Wilmers, C.C., Post, E., Peterson, R.O. et J.A. Vucetich. 2006. Predator disease out-break modulates top-down, bottom-up and climatic effects on herbivore population dynamics. Ecology Letters 9:383-389.
Wilson, P.J. (et 14 co-auteurs). 2000. DNA profiles of the eastern Canadian wolf and the red wolf provide evidence for a common evolutionary history independent of the Gray wolf. Canadian Journal of Zoology 78:2156-2166.
Wilson, P.J., Grewal, S., McFadden, T., Chambers, R.C. et B.N. White. 2003. Mitochondrial DNA extracted from eastern North American wolves killed in the 1800s is not of Gray Wolf origin. Canadian Journal of Zoology 81:936-940.
Wilson, P.J., Grewal, S.K., Mallory, F.F. et B.N. White. 2009. Genetic characterization of hybrid wolves across Ontario. Journal of Heredity. 100: S80-89.
Wilson, P.J., Rutledge, L.Y., Wheeldon, T.J., Patterson, B.R. et B.N. White. 2012. Y-chromosome evidence supports widespread signatures of three-species Canis hybridisation in eastern North America. Ecology and Evolution. DOI: 10.1002/ece3.301.
Woodroffe R., Cleaveland, S., Courtenay, O., Laurenson, M.K. et M. Artois. 2004. Infectious Disease: Infectious Disease in the management and Conservation of Wild Canids. Pp. 123-142 in Biology and Conservation of Wild Canids (D.W. MadDonald and C. Sillero-Zubiri (eds.). Oxford University Press, Oxford, UK.
Wydeven, A.P. (et 6 co-auteurs). 2009. History, Population Growth, and Management of Wolves in Wisconsin. Pp. 87 - 105 in Recovery of Gray Wolves in the Great Lakes Region of the United States. A.P. Wydeven et al. (eds), Springer, DOI: 10.1007/978-0-387-85952-1.
Wydeven, A.P., Fuller, T.K., Weber, W. et K. MacDonald. 1998. The potential for wolf recovery in the northeastern United States via dispersal from southeastern Canada. Wildlife Society Bulletin 26:776-784.
Young S. et E. Goldman. 1944. The Wolves of North America. Dover Publications, New York, NY. 636 pp.
Wydeven, A.P., (and 6 co-authors). 2009. History, Population Growth, and Management of Wolves in Wisconsin. Pp. 87 - 105 in Recovery of Gray Wolves in the Great Lakes Region of the United States. A.P. Wydeven et al. (eds), Springer, DOI: 10.1007/978-0-387-85952-1.
Young S., and E. Goldman. 1944. The Wolves of North America. Dover Publications, New York, NY. 636 pp.
Linda Rutledge est chercheure au niveau post-doctoral et chargée de cours au département de biologie, à l'Université Trent, à Peterborough (Ontario). Ses travaux de doctorat ont porté sur l'histoire de l'évolution, la structure des meutes et l'hybridation du loup de l'Est dans le parc provincial Algonquin. Les champs d'intérêt pour la recherche de madame Rutledge sont la détermination des changements sur le plan de l'évolution dans les génomes des espèces, ces changements étant des indicateurs des réactions comportementales, de l'adaptation, de l'hybridation et de la spéciation, en particulier en ce qui concerne les espèces en péril. La chercheure s'intéresse aussi à la manière dont l'être humain influe sur la trajectoire évolutive des espèces sauvages. Dans le cadre de ses travaux actuels, elle utilise la génomique comparative d'échantillons anciens et contemporains pour identifier les gènes qui sont responsables des adaptations écologiques dans l'espace et dans le temps.
Les collections examinées sont décrites dans le présent rapport. Les données brutes utilisées aux fins d'analyse proviennent de la banque de données sur l'ADN du loup et du coyote, à l'Université Trent [en anglais seulement].
L'identification d'une entité distincte (espèce, sous-espèce, unité désignable) par des techniques génétiques est compliquée par l'apparition de nouvelles techniques et l'interprétation des résultats. La présence d'haplotypes particuliers propres à une région géographique et leur persistance au fil du temps peuvent indiquer une adaptation locale et être utilisées pour la validation d'une espèce. Certains logiciels de simulation (p. ex. structure, Geneland) classent les spécimens en groupements d'animaux présentant des génotypes similaires. Cette dernière méthode repose sur une probabilité d'assignation et est sensible à la taille de l'échantillon, car quelques échantillons pourraient ne pas suffire à la différenciation comme groupement distinct au sein d'un échantillonnage plus grand (Fogelqvist et al., 2010). Aussi, la capacité de différencier dépend des différences relatives parmi l'échantillonnage; un sous-ensemble limité des divers génotypes signifie que certains groupements distincts ne seront pas identifiés en raison de l'absence de spécimens. Un autre aspect créant de la confusion dans la taxinomie du genre Canis tient au fait que plusieurs auteurs (vonHoldt et al., 2011, Koblmüller et al., 2009) ont mélangé des échantillons de loups de l'Est avec des échantillons de loups des Grands Lacs et les ont tous deux appelés Canis lycaon, puis ils ont conclu que le loup de l'Est, ou C. lycaon, était un hybride. Toutefois, selon l'hypothèse des trois espèces (voir plus bas), le loup de l'Est est une espèce valide, distincte du loup des Grands Lacs, lequel est un hybride entre le loup de l'Est et le loup gris.
La plupart des travaux de recherche sur l'hybridation des Canis visaient au départ à déterminer si le loup roux était un hybride, car les loups roux étaient protégés aux termes de l'Endangered Species Act (ESA) des États-Unis, alors que les hybrides ne l'étaient pas (USFWS, 2015a). Wayne et Jenks (1991) ainsi que Roy et al. (1994) ont utilisé l'absence de sites de restriction uniques de l'ADNmt (qui auraient réuni les loups roux en un seul clade) pour démontrer que le loup roux était un hybride. Par contre, Dowling et al. (1992) ont répliqué que la méthode et l'interprétation déficientes utilisées par Wayne et Jenks (1991) ne permettaient pas de tirer une telle conclusion. Selon des travaux ultérieurs, la période d'hybridation estimée ayant mené à la création du loup roux a eu lieu au cours des 2500 dernières années, et peut-être au cours des 250 dernières années (Reich et al., 1999). vonHoldt et al. (2011) ont conclu que les loups roux étaient des hybrides, mais Rutledge et al. (2012) ainsi que Wilson et al. (2012) ont noté que von Holdt et al. (2012) avaient utilisé des échantillons (n = 12) d'animaux qui étaient présents après la récente introgression de gènes du coyote, ce qui constitue un biais par rapport à l'identification des loups roux auparavant « purs ». Même si le débat se poursuit, le loup roux demeure inscrit sur la liste des espèces en voie de disparition des États-Unis.
Les études génétiques sur le loup de l'Est ont été faites dans le cadre d'une évaluation des Canis de la région des Grands Lacs qui a débuté dans les années 1990 (Lehman et al., 1991; Roy et al., 1994; Koblmüller et al., 2009). Par exemple, Wayne et Lehman (1992) ont eu recours à l'analyse des polymorphismes des sites de restriction de l'ADNmt pour 226 échantillons prélevés du Manitoba au Maine, combinés à 538 échantillons de Canis provenant d'ailleurs en Amérique du Nord, et ont conclu que le coyote de l'Est était un hybride, mais aussi que tous les Canis de la région des Grands Lacs étaient des hybrides. En 2009, Koblmüller et al. ont utilisé des marqueurs maternels, paternels et des marqueurs hérités des deux parents pour en arriver à la conclusion que les Canis de grande taille dans la région des Grands Lacs (loup des Grands Lacs) formaient un écotype ou une population de loups gris unique ayant subi un degré élevé d'introgression de gènes du coyote et du loup gris de l'Ouest dans l'Antiquité et récemment. vonHoldt et al. (2011) ont analysé 48 000 polymorphismes mononucléotidiques (SNP pour single-nucleotid-polymorphism) dans des échantillons de Canis prélevés dans le monde entier et ont conclu que le loup roux et le loup des Grands Lacs étaient différents, et que le loup des Grands Lacs était un hybride comportant une part importante du génome du loup gris.
L'une des critiques des travaux de recherche taxinomique antérieurs sur le genre Canis était liée au recours (par tous les chercheurs) à un seul paramètre (ADNmt). L'utilisation de plusieurs paramètres, comme le chromosome Y, est maintenant courante (p. ex. Wilson et al., 2012). Parmi les autres problèmes, citons le faible nombre de marqueurs généralement utilisés (p. ex. 12; Wilson et al., 2000; Rutledge et al., 2010b). Le grand nombre (48 000) de SNP utilisé par vonHoldt et al. (2011) est cité comme une amélioration par rapport aux travaux antérieurs (R. Wayne, dans NCEAS, 2014).
En résumé, selon l'hypothèse des deux espèces, il n'y aurait que deux espèces (le coyote et le loup gris), et les hybridations survenues récemment et historiquement auraient donné lieu à l'ensemble des hybrides de Canis.
L'affirmation selon laquelle le loup de l'Est et le loup roux sont des espèces valides provient d'une hypothèse soutenant que trois Canis de grande taille étaient présents en Amérique du Nord. Compte tenu de la divergence des séquences des haplotypes, Wilson et al. (2000) ont affirmé que le progéniteur ancestral du loup gris, du loup de l'Est/loup roux et du coyote avait résidé en Amérique du Nord. Il y a un ou deux millions d'années, le loup gris a divergé de l'ancêtre et émigré en Eurasie. Le coyote et le loup de l'Est/loup roux ont ensuite divergé l'un de l'autre il y a 150 000 à 300 000 ans (Wilson et al., 2000), et le loup gris a quitté l'Eurasie et est revenu en Amérique du Nord il y a quelque 300 000 ans (Kurten et Anderson, 1980). Les coyotes étaient limités aux zones situées à l'ouest de la rivière Mississippi, puis ils se sont déplacés vers le nord et vers l'est avec la colonisation et les pratiques d'utilisation des terres des Européens, et avec la disparition des loups dans l'est de l'Amérique du Nord. S'il y a un hybride du coyote de l'Est entre le loup de l'Est et le coyote « de l'Ouest », c'est qu'il s'agit d'espèces étroitement apparentées. Le loup des Grands Lacs (ou loup boréal/des Grands Lacs) est considéré comme un hybride entre le loup de l'Est et le loup gris (Wilson et al., 2000, 2009; Wheeldon, 2009).
Les résultats des travaux ci-dessus ont constitué la base de l'identification du loup de l'Est par le COSEPAC en 2001 (Samson, 2001). À l'heure actuelle, le loup de l'Est figure parmi les sous-espèces du loup gris (Canis lupus lycaon). Cependant, les analyses génétiques effectuées dans le cadre de nombreux projets ont permis de constater qu'il n'y avait pas suffisamment d'haplotypes et d'allèles de C. lupus chez le loup de l'Est/loup roux pour que le loup de l'Est soit considéré comme une sous-espèce du loup gris (Lehman et al., 1991; Roy et al., 1994, 1996; Wilson et al., 2000, 2003; Grewal et al., 2004; vonHoldt et al., 2011). En outre, certains haplotypes d'ADN mitochondrial non trouvés chez les populations de loups gris existantes ont été trouvés dans des échantillons de loup de l'Est datant des années 1800, c'est-à-dire avant l'arrivée des coyotes (Wilson et al., 2003).
Les premiers fondements de l'hypothèse des trois espèces résident dans les travaux sur les séquences de 8 locus microsatellites et de la région de contrôle de l'ADNmt de 68 échantillons prélevés dans le parc Algonquin, comparativement à des loups roux (n = 67), à des coyotes du Texas (n = 24) et à des loups gris des Territoires du Nord-Ouest (n = 67) [Wilson et al., 2000]. Les Canis du parc Algonquin ont été différenciés du loup gris, et le loup roux était semblable aux échantillons du parc Algonquin. Contrairement à ce qui a été observé pour le loup des Grands Lacs, l'ADNmt du loup gris n'a pas été trouvé dans les échantillons du centre de l'Ontario, comme on s'y serait attendu si ces animaux avaient été des hybrides du loup gris et du coyote. Wilson et al. (2009) ont plus tard analysé l'ADNmt de 269 échantillons de l'Ontario (109 du parc Algonquin et 186 en tout de l'aire de répartition du loup de l'Est), ainsi que de 393 échantillons de Canis d'ailleurs en Amérique du Nord, et ont conclu à l'existence d'une espèce de Canis distincte dans le centre de l'Ontario. Seulement 4 échantillons du parc Algonquin sur 92 contenaient des haplotypes du loup gris. Des travaux ultérieurs sur les haplotypes du chromosome Y chez le loup de l'Est ont permis de constater que ces haplotypes étaient absents chez les coyotes « de l'Ouest », ce qui constitue un élément de preuve de l'évolution du loup dans l'est de l'Amérique du Nord, et qui concorde avec l'hypothèse des trois espèces (Wilson et al., 2012).
D'autres études ont également montré qu'une espèce de Canis autre que le loup gris a déjà été présente dans l'est de l'Amérique du Nord (Bertorelle et Excoffier, 1998). L'ADNmt du loup de l'Est a été trouvé dans des échantillons du Wisconsin et du Michigan datant de 100 ans (Leonard et Wayne, 2008), et Wheeldon et White (2009) ont trouvé de l'ADNmt du loup de l'Est de même que la signature nucléaire de loups gris-loups de l'Est d'ascendance mixte dans des échantillons de loups du Minnesota et du Wisconsin datant de 100 ans. Les données génétiques semblent indiquer qu'il y aurait eu des loups de l'Est dans le Maine et l'État de New York il y a 100 ans, avant l'expansion vers l'est des coyotes (Wilson et al., 2003). Les analyses tant génétiques que morphométriques d'échantillons prélevés par excavation dans un village iroquois préhistorique (v. 1530) à London, en Ontario, portent à croire que les loups de l'Est étaient présents dans les forêts caroliniennes du sud de l'Ontario avant l'arrivée des colons européens (Rutledge et al., 2010d). Un Canis de petite taille a été trouvé dans les fossiles de l'est de l'Amérique du Nord datant du Pleistocène (Nowak, 1995).
La conclusion tirée par vonHoldt et al. dans son article de 2011 et dans des articles antérieurs, selon laquelle le loup de l'Est est un hybride du loup gris et du coyote, a été remise en question, ou n'a pas été appuyée, dans de récentes publications (Rutledge et al., 2010c, 2012; Wilson et al., 2012; Benson et al., 2012, 2014). Cette remise en question a pour origine le faible nombre d'échantillons de loups de l'Est connus. L'argument des partisans de l'hypothèse des trois espèces tient au fait que le loup de l'Est est confiné à une zone relativement petite et que les échantillons prélevés dans la région des Grands Lacs (nord de l'Ontario, Minnesota) et réunis en un échantillon de loup des Grands Lacs (p. ex. Koblmüller et al., 2009; vonHoldt et al., 2011) ne constituent pas une mesure valide de l'existence du loup de l'Est. La taille de l'échantillon influera sur la probabilité de différencier un groupement d'animaux particuliers parce que le processus fait appel aux génotypes individuels assignés à un taxon en fonction de la taille de l'échantillon et de la probabilité (Fogelqvist et al., 2010). Dans l'étude de Wayne et Lehman (1992), seulement 3 échantillons sur 764 provenaient de la région du parc Algonquin (dont deux de la même meute; G. Forbes, données inédites), et 16 échantillons en tout provenaient de l'aire de répartition existante du loup de l'Est (figure 4). De plus, on ne sait pas avec certitude si ces spécimens étaient des loups de l'Est ou des Canis d'ascendance mixte (coyote de l'Est) et, en conséquence, on ignore dans quelle mesure les loups de l'Est ont été vraiment bien échantillonnés. Dans l'étude de Koblmüller et al. (2009), quelques échantillons représentaient le génotype Canis avant l'introgression des gènes du coyote; seulement 4 échantillons sur 401 provenaient de spécimens historiques (1910-1916) dans l'aire de répartition actuelle du loup de l'Est.
En réponse à l'étude de vonHoldt et al. (2011), Rutledge et al. (2012) ont fait remarquer que les deux échantillons du parc Algonquin ne suffisaient pas à différencier le loup de l'Est. Toutefois, lorsqu'une nouvelle analyse phylogénétique des 48 000 SNP de l'étude de von Holdt et al. (2011) a été faite avec les deux échantillons, et que ces derniers ont ensuite été combinés à d'autres échantillons locaux, les deux loups de l'Est du parc Algonquin formaient alors un groupement indépendant des autres groupes de Canis et étaient situés en position basale par rapport aux loups roux et aux coyotes au sein d'une lignée nord-américaine (Rutledge et al., 2012), ce qui indique une ascendance avec le coyote, comme on pouvait s'y attendre selon l'hypothèse des trois espèces. Aussi, la structure basale était comparable à celle observée tant avec l'ADNmt (Rutledge et al., 2010b) qu'avec le chromosome Y (Wilson et al., 2012). Ces résultats fournissent des données génétiques sur la contribution maternelle, paternelle et biparentale démontrant que le loup de l'Est est une espèce. Plus récemment, une analyse de 127 000 SNP à l'échelle du génome a démontré au moyen de simulations d'hybridation que le loup de l'Est est une espèce distincte, et son intégration comme troisième espèce résout la question des origines hybrides des loups boréaux/des Grands Lacs et des coyotes de l'Est (Rutledge et al., en révision).
Rutledge et al. (2012) ont aussi remarqué que vonHoldt et al. (2011) s'étaient servi d'une méthode reposant sur un modèle à deux espèces, lequel ne permet le partage des origines qu'entre les loups gris et les coyotes, un protocole qui ne pouvait différencier le loup de l'Est. Rutledge et al. (2012) soutiennent également qu'il y a une hypothèse biaisée selon laquelle l'ensemble des SNP établis issus du génome du chien domestique (Canis lupus familiaris) rendent bien compte de la variabilité réelle des espèces sauvages de Canis.
Même si le débat entourant la taxinomie des Canis se poursuit, les éléments ci-dessous sont jugés suffisamment convaincants pour que l'on considère le loup de l'Est comme une entité taxinomique valide.
- Deux haplotypes d'ADN du loup de l'Est (C1 et C3) forment des groupements monophylétiques et sont en position basale par rapport à tous les autres haplotypes nord-américains évolués (coyote/loup roux) [Rutledge et al., 2010b].
- Il existe trois haplotypes du chromosome Y du loup de l'Est (4AA, 4BB, 4BR) qui sont plus étroitement associés aux coyotes, mais qui sont aussi différents des coyotes que les coyotes le sont des loups gris (Wilson et al., 2012).
- Les loups du parc Algonquin forment un groupement monophylétique primitif distinct dans l'analyse phylogénétique de 48 000 SNP (Rutledge et al., 2012).
- Il existe un groupement indépendant des microsatellites autosomaux analysés chez les loups du parc Algonquin (Rutledge et al., 2010c; Rutledge et al., 2012; Benson et al., 2012).
- Il existe un groupement indépendant et une absence de chevauchement avec les hybrides simulés de loups gris et de coyotes d'après les 127 000 SNP à l'échelle du génome (Rutledge et al., en révision).
- Parmi un échantillon comportant des canidés du parc Algonquin (n = 37), des loups gris d'Amérique du Nord (n = 175), des loups gris d'Europe (n = 222) et des coyotes de partout en Amérique du Nord (n = 70), l'identification de trois haplotypes uniques du complexe majeur d'histocompatibilité, un groupe de gènes participant à la réponse immunitaire, chez les loups du parc Algonquin porte à croire que ces derniers sont une espèce bien distincte des autres canidés (Kennedy, comm. pers., 2012).
- Même si l'ADNmt apparenté au coyote est introgressé chez les loups du parc Algonquin (Rutledge et al., 2010c), il existe peu de données sur l'introgression de l'ADN nucléaire (Rutledge et al., 2011). Cette discordance entre l'ADNmt et l'ADN nucléaire est courante dans les systèmes animaux (voir le compte rendu de Toews et Brelsord, 2012) et on ignore toujours si la présence d'ADNmt apparenté au coyote chez le loup de l'Est est due à un tri des lignées incomplet attribuable à leur divergence récente (il y a 150 000 à 300 000 ans [Wilson et al., 2000]).
- Il existe des données sur la morphologie (Kolenosky et Standfield, 1975; Schmitz et Kolenosky, 1985; Rutledge et al., 2010e; Benson et al., 2012) qui plaident en faveur de l'existence du loup de l'Est (voir Description morphologique).
- Un canidé distinct est reconnu dans la région des Grands Lacs - Est de l'Amérique du Nord depuis le milieu des années 1700 (voir la section Validité taxinomique), et des animaux possédant les mêmes caractéristiques génétiques que le loup de l'Est d'aujourd'hui ont été trouvés dans des échantillons datant de 100 ans (Wilson et al., 2003) et d'avant l'arrivée des coyotes. Au Québec, la présence du coyote n'a été signalée qu'après les années 1940 (Naughton, 2012), mais le spécimen type pour le Canis lycaon a été recueilli au Québec en 1761 (voir Validité taxinomique). Selon l'hypothèse des deux espèces, le loup de l'Est est le produit du croisement entre le coyote et le loup gris; cependant, d'après les données historiques, le loup de l'Est était déjà présent avant les coyotes, ce qui porte à croire que le loup de l'Est est une véritable espèce et non le produit de l'hybridation du coyote et du loup gris (Rutledge et al., 2010d).
- D'après les connaissances traditionnelles autochtones, différents types de canidés étaient présents dans la région avant l'arrivée des colons européens (Lickers, comm. pers.).
- L'analyse génétique de fragments d'os de crâne donne à penser que le loup de l'Est (et non le loup gris) est présent dans le sud-ouest de l'Ontario depuis les années 1500 (Rutledge et al., 2010d).
- On croit que le Canis de grande taille qui s'est reproduit avec le coyote « de l'Ouest » est le loup de l'Est parce que le loup gris et le coyote ne s'accoupleraient pas nécessairement entre eux dans la nature (Kyle et al., 2007; Mech, 2011). Un tel accouplement serait requis pour la création d'un hybride du loup de l'Est. Mech et al. (2014) ont inséminé artificiellement des femelles coyotes « de l'Ouest » avec du sperme de loups de l'Ouest et les femelles ont eu des petits, ce qui prouve que le croisement est possible. Les résultats étaient cependant mitigés, car 8 ou 9 femelles ont présenté un comportement anormal ou ont mangé les fœtus.
- Le loup gris et le coyote sont sympatriques dans le sud-ouest de l'Amérique du Nord (c.-à-d. le loup du Mexique, C. lupus baileyi) et dans le nord-ouest de l'Amérique du Nord, mais l'hybridation n'a pas été observée (Hedrick et al., 1997; Garcia-Moreno et al., 1996; Pilgrim et al., 1998), ce qui va à l'encontre de l'hypothèse des deux espèces selon laquelle il y a hybridation lorsque le loup gris et le coyote sont sympatriques. L'hybridation est plus probable pour les espèces étroitement apparentées, ce qui tend à confirmer la conclusion de l'hypothèse des trois espèces selon laquelle le loup de l'Est/loup roux et le coyote s'hybrident dans l'est de l'Amérique du Nord parce qu'ils sont plus étroitement apparentés qu'ils ne le sont avec le loup gris (Wilson et al., 2000).
Les loups de l'Est identifiés par les analyses génétiques se trouvent presque tous dans des zones protégées, même si l'échantillonnage a été fait sur une vaste gamme de zones du parc et hors du parc (voir la section Activités de recherche; figures 4, 6). On ne connaît pas le mécanisme qui fait en sorte qu'une espèce distincte est créée et maintenue parmi des formes de Canis étroitement apparentées, mais des données probantes montrent que l'introgression de gènes du coyote/coyote de l'Est est facilitée par un changement dans la dynamique des meutes. On suppose que la mortalité excessive des individus reproducteurs modifie la structure sociale reproductive de la meute (Rutledge et al., 2010c, 2011; Benson et al., 2013a, 2014). En règle générale, seul le couple dominant (alpha) se reproduit, et la perte du reproducteur peut détruire la cohésion de la meute et changer les habitudes d'accouplement (Brainerd et al., 2008). Malgré l'introgression chez certains individus (Rutledge et al., 2010c; Grewal et al., 2004), les loups de l'Est du parc Algonquin ont conservé une signature génétique distincte pendant au moins 50 ans (Rutledge et al., 2011). Les loups dans ce parc de plus de 7 000 km2 ont été protégés contre la chasse et le piégeage durant la majeure partie de cette période. Dans les années 1980 et 1990, de nombreux loups du parc ont été tués lorsqu'ils ont suivi les cerfs à l'extérieur de la limite est du parc (Forbes et Theberge, 1996b). Ces mortalités élevées ont joué un rôle dans l'introgression de l'ADNmt du coyote et dans l'apparence de l'ADN nucléaire du coyote de l'Est durant cette période, car la signature nucléaire du loup de l'Est a été rétablie après qu'une interdiction visant la chasse et le piégeage a été mise en œuvre dans les zones adjacentes au parc en 2001 (Rutledge et al., 2011). De même, les loups de l'Est dispersés à partir des secteurs ouest du parc sont très vulnérables à la mortalité due à la chasse (Benson et al., 2014), et ce, malgré la présence de quelques lieux de résidence observés dans les zones non protégées du secteur de gestion de la faune (SGF) 47 (n = 3) et du SGF 49 (n = 1) [Benson et al., 2012]. L'hybridation continue avec les coyotes de l'Est a été réduite par une protection qui rend vraisemblablement les congénères partenaires plus faciles à trouver (Rutledge et al.,2010c, 2011).
Malgré les preuves de flux génique entre les formes de Canis en général, on observe une nette structure génétique et morphologique chez les loups de l'Est. Les différences écologiques dans l'habitat et le bassin de proies, de même que le choix des congénères partenaires pourraient susciter la divergence (Rutledge et al., 2010c). Les niveaux d'hétérozygotie, d'après les marqueurs microsatellites autosomaux, sont généralement élevés (Ho = 0,645; erreur type = 0,04; Rutledge et al., 2010c) et même si les mesures traditionnelles de la différenciation génétique comme l'indice FST sont relativement basses (indice FST entre les loups de l'Est et les loups boréaux/des Grands Lacs = 0,105; et entre les loups de l'Est et les coyotes de l'Est = 0,052), les différences sont statistiquement significatives (p < 0,001). Aussi, de nouvelles mesures reposant sur les allèles partagés (Jost, 2008) donnent à penser que les niveaux de différenciation sont plus élevés entre les loups de l'Est et les loups boréaux/des Grands Lacs (Jost, indice D = 0,207) qu'entre les loups de l'Est et les coyotes de l'Est (Jost, indice D = 0,090) [Rutledge et al., 2010c]. Les analyses bayésienne et multivariable des groupements fondées sur les microsatellites autosomaux ont révélé une différenciation marquée des loups de l'Est dans le parc Algonquin par rapport aux formes de Canis des régions avoisinantes, et ce, malgré l'absence de barrière physique (Rutledge et al.,2010c; Benson et al., 2012). Les meutes de loups de l'Est sont séparées les unes des autres ainsi que des autres formes de Canis dans l'espace (Benson et Patterson, 2013a).
La signature génétique unique pourrait aussi être reliée à l'homogamie et/ou à la sélection défavorable à l'introgression nucléaire par les coyotes de l'Est (Rutledge et al., 2010c) parce que les loups de l'Est se distinguent sur le plan écologique par l'habitat et les besoins énergétiques associés à la grande taille des proies (Benson et al., 2012; Rutledge et al., 2010c). Loveless (2010) a laissé entendre que le rétablissement de la structure sociale naturelle dans le parc Algonquin (Rutledge et al., 2010e) pourrait être à l'origine de l'efficacité accrue de la prédation par les orignaux grâce à l'apprentissage transmis entre les membres d'une famille. La signature génétique unique n'est probablement pas due à une dérive génétique, car cette dernière est le plus souvent associée à des populations isolées, et le parc Algonquin n'est pas isolé. Le flux génique peut se produire dans l'ensemble de la région, mais le génotype des Canis dans le parc Algonquin se maintient au moins depuis les années 1960, probablement à cause de l'homogamie (Rutledge et al., 2011). En outre, la diversité génétique est relativement élevée (hétérozygotie observée de 0,645; Rutledge et al., 2010c) et il n'y a aucun signe de consanguinité (Rutledge et al., 2010e).
D'après l'hypothèse des trois espèces, il est possible que le loup de l'Est et le loup roux soient la même espèce. Les données issues des microsatellites autosomaux semblent indiquer un chevauchement dans leur signature génétique (Wilson et al., 2000; Kyle et al., 2006, 2008; Murray et Waits, 2008; Rutledge et al., 2012), et l'analyse phylogénétique des 48 000 SNP porte à croire que les loups du parc Algonquin sont en position basale par rapport aux loups roux au sein d'une lignée nord-américaine (Rutledge et al., 2012). L'aire de répartition aurait probablement compris les régions de l'est des États-Unis et les régions du sud de l'Ontario et du Québec, une aire de répartition correspondant en grande partie aux forêts tempérées de l'est et à la répartition historique du cerf (Nowak, 1995, 2002, figure 3).
La population de loups roux existante a été produite dans le cadre d'un programme de reproduction en captivité qui a commencé avec 14 animaux capturés dans la région Texas-Louisiane durant les années 1970. Ces animaux avaient été identifiés comme des loups roux en fonction de critères morphométriques et de leurs vocalisations (McCarley et Carley, 1979). Certains de leurs descendants ont été relâchés en Caroline du Nord et maintenant, ils se reproduisent (USFWS, 2015b). Les premiers reproducteurs renfermaient cependant du matériel génétique du coyote (vonHoldt et al., 2011), du chien et/ou du loup gris (Hailer et Leonard, 2008; Wilson et al.,2012) et, malgré la gestion contrôlée de la reproduction, on craint que l'échantillon de loup roux existant ne soit pas une représentation complète du génome du loup roux. Ainsi, l'utilisation d'un échantillon génétique actuel pour déterminer si les animaux étaient de la même espèce est compromise.
Le problème de taxinomie a eu des incidences sur la liste des espèces en voie de disparition aux États-Unis. En mai 2011, le Fish and Wildlife Service des États-Unis (USFWS) a officiellement reconnu le loup de l'Est comme une espèce distincte (C. lycaon), mais il est revenu sur sa déclaration en décembre 2011. Une analyse de la taxinomie des hybrides, des sous-espèces et des espèces de Canis en Amérique du Nord a donc été faite (Chambers et al. 2012), et a constitué la base d'une proposition de décision sur l'inscription par l'USFWS. D'après les données recueillies par vonHoldt et al. (2011), l'USFWS a entrepris un exercice de consultation auprès d'un groupe d'experts (NCEAS, 2014) pour examiner l'analyse de Chambers et al. (2012). Il importe de souligner que l'analyse de Chambers et al. (2012) ne portait pas sur le loup de l'Est (tel qu'il est défini dans le présent rapport), parce que ces chercheurs ont combiné tous les Canis dans la région Québec-Grands Lacs en un seul « loup de l'Est (Canis lycaon) » alors que dans le présent rapport, on soutient que le loup de l'Est ne fait référence au Canis que dans le centre de l'Ontario et le sud du Québec (voir l'annexe B; la figure 4). Dans son rapport, le NCEAS (2014) conclut qu'il n'y a pas de consensus sur la taxinomie des Canis : les partisans de l'hypothèse des deux espèces et ceux de l'hypothèse des trois espèces conviennent qu'ils sont en désaccord (NCEAS, 2014). Dans son rapport, le NCEAS ne conclut pas que le loup de l'Est est une espèce non valide, mais uniquement qu'il n'y a pas de consensus sur la taxinomie des Canis, et que Chambers et al. (2012) n'ont pas suffisamment bien mis en évidence ce point de vue; par conséquent, la proposition de décision n'est pas fondée sur les « meilleures données scientifiques disponibles ».
En dépit du débat, il existe un consensus sur le fait que les populations actuelles des loups roux et des loups de l'Est possèdent une certaine partie du génome du coyote. Aussi, on s'entend sur le fait que le coyote de l'Est est un hybride, qui est maintenant présent dans tout l'est de l'Amérique du Nord. Ces Canis se sont établis grâce à l'expansion naturelle de l'aire de répartition et à l'interaction. Le problème des hybridations, en particulier dans les milieux modifiés par l'humain, soulève des questions sur les limites de la notion d'espèce et de la nomenclature taxinomique comme base pour la gestion des espèces qui ne sont pas « pures » (Wilson et al., 2012). La controverse sur l'intégration d'hybrides possibles comme le loup roux parmi les espèces en voie de disparition protégées a donné lieu à de nombreuses discussions sur l'identification des espèces d'après leurs caractéristiques écologiques et comportementales et non uniquement leurs caractéristiques génétiques (p. ex. Theberge, 1991; Nowak, 1992; Phillips et Henry, 1992). Dowling et al. (1992) ont fait observer que l'ESA visait à protéger les formes distinctives sans égard au fait qu'elles trouvent leur place dans la notion d'espèces. L'Endangered Species Act des États-Unis reconnaît les unités évolutionnaires significatives et, dans le cadre de son application, la Loi sur les espèces en péril du Canada reconnaît une définition élargie d'espèce sauvage; ces deux lois facilitent donc la protection d'entités distinctes et importantes d'un rang inférieur à celui d'espèce.
Certains individus doivent être assignés à la population des loups de l'Est au Canada, car l'aire de répartition du loup de l'Est comprend des Canis de génotypes mixtes. L'assignation d'individus comme les loups de l'Est est fondée sur l'analyse de marqueurs génétiques microsatellites autosomaux nucléaires hérités des deux parents en raison de : a) l'introgression d'haplotypes de l'ADNmt et du chromosome Y du loup de l'Est chez le loup boréal/des Grands Lacs (Rutledge et al., 2010c) et le coyote de l'Est (Way et al., 2010) à l'échelle de l'aire de répartition de ces espèces; b) la divergence ADNmt-ADN nucléaire observée dans l'ensemble des systèmes animaux, dont le genre Canis (Toews et Brelsford, 2012); et c) la possibilité d'un tri des lignées incomplet de l'ADNmt entre le loup de l'Est et les coyotes (Wheeldon et White, 2009).
Les méthodes d'identification des formes de Canis font habituellement appel à 12 marqueurs microsatellites autosomaux pour la réalisation des tests d'assignation dans un cadre analytique bayésien intégré au logiciel structure 2.3 (Hubisz et al., 2009). D'après les tests de simulation (Rutledge et al., 2010c), un score d'assignation Q ≥ 0,8 (où Q est la probabilité d'appartenance à un groupement) obtenu avec le logiciel structure 2.3 (Hubisz et al., 2009) est établi comme critère pour une identification fiable du loup de l'Est. Les individus présentant une valeur Q inférieure à 0,8 sont considérés comme étant d'ascendance mixte (c.-à-d. possédant un mélange d'allèles nucléaires issus d'au moins deux des trois formes de Canis). Avec cette méthode, les échantillons peuvent être assignés à l'une des trois formes de Canis, soit le loup de l'Est, le loup boréal/des Grands Lacs ou le coyote de l'Est, ou identifiés comme étant « d'ascendance mixte », ce qui signifie que l'hybridation était manifeste entre deux ou trois formes de Canis.
L'utilisation de la valeur seuil Q < 0,8 repose sur les modes de transmission héréditaire de base, la littérature, des analyses et certaines hypothèses. Le mode de transmission héréditaire de base désigne tout hybride F1 possédant une valeur Q égale à 0,5 et qui devient égale à 0,75 s'il y a recroisement avec un parent. L'utilisation d'une valeur Q supérieure à 0,8 permet d'éviter que de tels cas soient inclus. La notion de valeur seuil Q a été validée ailleurs (p. ex. Vähä et Primmer, 2006) de même qu'avec l'analyse de simulations d'hybrides qui a révélé une forte démarcation du loup de l'Est à l'intérieur et à l'extérieur du parc Algonquin, une conséquence que l'on croit reliée aux effets de la récolte de loups de l'Est et aux taux d'introgression des gènes du coyote (voir l'annexe A - « Mécanisme »). Benson et al. (2012) ont analysé la validité des assignations selon la valeur Q < 0,8 à l'aide de méthodes d'ACP et ont découvert que le taux de conformité entre les assignations du logiciel structure et celles de la méthode d'ACP était de 90 %. La méthode de la valeur Q est une méthode d'identification courante des formes de Canis (p. ex. Veradri et al,. 2006; Benson et al., 2012; Wheeldon et al., 2013).
Avec la méthode de la valeur seuil Q, une hypothèse sur ce que serait un génotype « pur » de loup de l'Est doit être formulée. Cependant, pour caractériser un loup de l'Est pur, nous n'avons pas suffisamment de spécimens recueillis avant l'arrivée des coyotes. L'utilisation de la valeur seuil Q ≥ 0,8 pour l'identification du loup de l'Est détermine le nombre d'animaux et de sites présentant ce qui est considéré comme un loup de l'Est. Même si Q ≥ 0,8 est un seuil valable, une carte des animaux dont la valeur Q est égale à 0,75 permettrait d'identifier plus d'animaux et, par conséquent, de déterminer la mesure dans laquelle le loup de l'Est est limité aux zones protégées, comme cela semble le cas. Il a été impossible de produire une telle carte parce que les sites correspondant aux 457 échantillons analysés au Québec (Rutledge et White, 2014) n'étaient connus que pour les 11 spécimens présentant une valeur Q ≥ 0,8. Dans la région du parc Algonquin, si la valeur Q était modifiée pour 0,75 et/ou 0,85, le nombre d'animaux assignés à une catégorie de Canis ou à une autre varierait de plus de 10 % pour certaines zones échantillonnées (C. Kyle, données inédites). De plus, dans certaines régions échantillonnées où on ne trouve pas de loups de l'Est dont la valeur Q est supérieure à 0,8, il y a des animaux dont la valeur Q est supérieure à 0,75. Il est alors possible que la taille de la population de loups de l'Est varie, mais uniquement d'un petit nombre par rapport à l'estimation fondée sur la valeur seuil Q ≥ 0,8 (tableau 2). L'aire de répartition (zone d'occurrence) ne changerait probablement pas, car elle englobe déjà une vaste zone où il y a eu quelques mentions. En résumé, l'utilisation du seuil Q ≥ 0,8 est la méthode qui a le plus d'appui et qui semble donner une représentation relativement bonne de la répartition et de la taille de la population.
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de nniveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de nniveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 1 | 0 |
| C | Moyen | 2 | 2 |
| D | Faible | 1 | 2 |
| - | Impact global des menaces calculé : | Très élevé | Élevé |
| # | Menace | Impact (calculé) Critère |
Impact (calculé) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 1.1 | Zones résidentielles et urbaines | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | D'après la zone d'occurrence de l'espèce indiquée à la figure 4 du rapport. Portée : tous sont d'accord que les nouveaux aménagements seraient inférieurs à 10 %. Gravité : d'après la population qui sera touchée par les nouvelles zones résidentielles et urbaines (et non uniquement le nombre de mortalités). * L'impact est extrêmement élevé si l'on tient compte des nouveaux aménagements urbains, mais les loups peuvent utiliser les zones de villégiature/résidentielles qui restent relativement rurales. * En Ontario, le loup de l'Est est absent des zones urbaines. * Au Québec, on s'attend à une hausse de la construction de chalets. Remarque : peut-être à revoir compte tenu de l'incertitude au Québec; le Québec tient compte des mortalités uniquement en fonction des nouveaux aménagements (ainsi négligeables), car les loups quitteraient la zone. |
| 1.2 | Zones commerciales et industrielles | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | Portée * Le Québec suggère une petite (1-10 %) portée, mais pourrait être d'accord avec une portée négligeable. * Il se pourrait que de nouveaux centres commerciaux soient construits en Ontario, mais cela ne devrait pas faire augmenter la portée à un niveau supérieur à 1 %. |
| 1.3 | Zones touristiques et récréatives | D | Faible | Petite (1-10 %) | Élevée - Modérée (11-70 %) | Élevée (continue) | Portée * Qc - petite (1-10 %) * L'Ontario est saturé de terrains de golf et n'appuiera probablement pas l'aménagement d'autres terrains de golf dans les 10 prochaines années; petite (1-10 %) en fonction d'autres installations récréatives (p. ex. camps d'été, chalets, etc.). |
| 2 | Agriculture et aquaculture (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) | Élevée - Légère (1-70 %) | Modérée (peut-être à court terme, < 10 ans) | - |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | - | Négligeable | Négligeable (< 1 %) | Élevée - Légère (1-70 %) | Modérée (peut-être à court terme, < 10 ans) | Portée : négligeable selon une entente Ont./Qc Gravité : * La plupart des loups sont dans des zones protégées; cependant, les données de répartition sont limitées lorsque les loups se déplacent entre les parcs; une plage est utilisée pour indiquer l'incertitude. * En Ontario, le couvert forestier s'accroît et l'étendue des terres agricoles diminue; encore une fois, si on ne sait pas où les loups vont lorsqu'ils sont à l'extérieur du parc, beaucoup d'incertitude demeure. |
| 2.2 | Plantations pour la production de bois et de pâte | - | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | * Ont. : ne représentent pas un enjeu. * Qc : représenteront une certaine menace. |
| 2.3 | Élevage de bétail | - | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (continue) | Gravité * Ont. : l'élevage pourrait s'accroître. * Qc : entre le parc Algonquin et les parcs dans l'est de l'aire de répartition, il y a beaucoup d'incertitude. |
| 3 | Production d'énergie et exploitation minière (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 3.1 | Forage pétrolier et gazier | - | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Négligeable (passée ou aucun effet direct) | * Ont. : pas d'enjeu connu. * Qc : on croit qu'il y en aura un peu. |
| 3.2 | Exploitation de mines et de carrières | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | Portée * Ont. : l'impact pourrait être plus important, mais il serait près de la limite inférieure d'une portée jugée petite (1-10 %); cependant, d'après la figure 4a, la plus grande partie touche le Québec et on s'entendra pour une portée négligeable (< 1 %). |
| 3.3 | Énergie renouvelable | - | - | - | - | - | - |
| 4 | Corridors de transport et de service (en anglais seulement) | CD | Moyen - Faible | Généralisée (71-100 %) | Modérée - Légère (1-30 %) | Élevée (continue) | - |
| 4.1 | Routes et voies ferrées | CD | Moyen - Faible | Généralisée (71-100 %) | Modérée - Légère (1-30 %) | Élevée (continue) | Portée : généralisée (71-100 %) parce que cela comprend toutes les routes (dont celles utilisées pour l'exploitation forestière/minière) Gravité : plage utilisée pour signaler l'incertitude. * Ont. : des animaux meurent certainement sur les routes, mais il ne s'agit pas d'une cause majeure du déclin de la population. Il y a eu quelques cas de mortalité sur les routes récemment (p. ex. route 116), mais il s'agit d'un petit nombre. * Qc : il existe peu de données, mais les chiffres pourraient être plus élevés. Selon une récente étude, 6 loups sur 60 ont été tués; par contre, on ignore s'il s'agissait de loups de l'Est ou d'hybrides. |
| 4.2 | Lignes de services publics | - | Négligeable | Petite (1-10 %) | Négligeable (< 1 %) | Élevée (continue) | Portée : ne tient compte que des nouvelles lignes de services publics. * Ont. : aucun nouvel aménagement qui pourrait toucher la population; < 1 %. * Qc : près de 1 %, mais on ignore si d'autres zones seront touchées dans les années à venir. |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | BC | Élevé - Moyen | Grande (31-70 %) | Élevée - Modérée (11-70 %) | Élevée (continue) | - |
| 5.1 | Chasse et capture d'animaux terrestres | BC | Élevé - Moyen | Grande (31-70 %) | Élevée - Modérée (11-70 %) | Élevée (continue) | Portée : tient compte de la chasse et légale et illégale et des prises accidentelles. Une grande proportion de la population se trouve dans le parc Algonquin; une portée généralisée (71-100 %) serait trop élevée. * Ont. : une fois qu'ils ont quitté la zone protégée, les loups sont beaucoup plus menacés. À l'extrémité est, le braconnage/la récolte ne sont pas permis. Gravité : plage utilisée pour signaler l'incertitude. * Qc : dépend des divers groupes et de la proportion d'animaux qui vont à l'extérieur de la zone protégée; n'est peut-être pas élevée à certains endroits. |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | Ne représente pas un enjeu. |
| 5.3 | Exploitation forestière et récolte du bois | - | - | - | - | - | Ne représentent pas un enjeu. * Ont. : pourraient être un avantage sur une période de 10 ans. |
| 5.4 | Pêche et récolte de ressources aquatiques | - | - | - | - | - | Ne représentent pas un enjeu. |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | - | Négligeable | Généralisée (71-100 %) | Négligeable (< 1 %) | Élevée (continue) | - |
| 6.1 | Activités récréatives | - | Négligeable | Généralisée (71-100 %) | Négligeable (< 1 %) | Élevée (continue) | Portée * Qc : la pratique de la motoneige est très importante. * Ont. : élevée si les activités comme le canotage et d'autres activités pratiquées au chalet sont prises en considération. |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | Ne représentent pas un enjeu. |
| 6.3 | Travail et autres activités | - | - | - | - | - | Ne représentent pas un enjeu. |
| 7 | Modification du système naturel (en anglais seulement) | - | Négligeable | Restreinte - Petite (1-30 %) | Négligeable (< 1 %) | Élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | - | Négligeable | Restreinte - Petite (1-30 %) | Négligeable (< 1 %) | Élevée (continue) | - |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | C | Moyen | Généralisée (71-100 %) | Modérée (11-30 %) | Élevée (continue) | - |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | - | Inconnu | Généralisée (71-100 %) | Inconnue | Élevée (continue) | Portée * Ont. : serait généralisée (71-100 %). Remarque : chiffres fondés sur des études menées en 2007 (une mauvaise année); probablement 11 % des animaux sont morts. * Qc : aucune donnée. Gravité : inconnue. * L'Ontario peut fournir des chiffres précis sur la mortalité, mais ne peut faire une meilleure estimation de la gravité. Remarque : la question du matériel génétique du coyote est traitée en 8.3 |
| 8.2 | Espèces indigènes problématiques | - | Inconnu | Grande (31-70 %) | Inconnue | Élevée (continue) | Portée : quel pourcentage des loups de l'Est est touché par des espèces indigènes problématiques? * Ont. : tous les animaux qui sont à l'extérieur du parc Algonquin. Remarque : il s'agit d'un facteur limitatif plutôt que d'une menace; traitées comme un facteur limitatif dans le rapport. |
| 8.3 | Matériel génétique introduit | C | Moyen | Généralisée (71-100 %) | Modérée (11-30 %) | Élevée (continue) | Gravité * Ont. : n'importe quel loup quittant le parc Algonquin pourrait avoir des gènes de chien/coyote dans une proportion de plus de 30 %, mais on peut convenir d'une plage de 11 à 30 % pour toutes les populations. |
| 9 | Pollution (en anglais seulement) | - | - | - | - | - | - |
| 9.4 | Déchets solides et ordures | - | Négligeable | Petite (1-10 %) | Négligeable (< 1 %) | Élevée (continue) | Ne comptent pas dans le calcul de l'impact global. * Score fondé sur le Québec, mais cela pourrait en fait avoir un effet positif. * Ont. : les sites d'enfouissement/ déchets ont un impact (positif et négatif) sur les loups. |
| 9.5 | Polluants atmosphériques | - | - | - | - | - | - |
| 9.6 | Apports excessifs d'énergie | - | - | - | - | - | - |
| 10 | Phénomènes géologiques (en anglais seulement) | - | - | - | - | - | Ne représentent pas un enjeu. |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | - | - | - | - | - | Inconnus. Les changements climatiques ne datent pas d'assez longtemps pour que l'on observe des modifications. À long terme, la densité des orignaux (et des cerfs) pourrait changer et toucher la population des loups. |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | Inconnus |