
Fowler’s toad (Anaxyrus fowleri): COSEWIC assessment and status report 2010
Table of Contents
- COSEWIC Assessment Summary
- COSEWIC Executive Summary
- Species Information
- Name and classification
- Morphological description
- Genetic description
- Designatable units
- Distribution
- Global range
- Canadian range
- Habitat
- Habitat requirements
- Habitat variability
- Other habitat considerations
- Habitat trends
- Habitat protection/ownership
- Biology
- Life cycle and reproduction
- Feeding and predation
- Physiology
- Dispersal and migration
- Interspecific interactions
- Adaptability
- Population Sizes and Trends
- Search effort
- Abundance
- Fluctuations and trends
- Rescue effect
- Population viability analysis
- Limiting Factors and Threats
- Shoreline development and artificial stabilization
- Invasive species
- Agriculture and industrial contaminants
- Special Significance of the Species
- Existing Protection or Other Status Designations
- Technical Summary
- Acknowledgements and Authorities Consulted
- Information Sources
- Biographical Summary of Report Writers
- Collections Examined
List of Figures
- Figure 1. Fower’s Toad
- Figure 2. Neighbour–joining tree of sequence divergence (p–distance) from control region mtDNA of Fowler’s Toads of populations from the northern edge of their distribution
- Figure 3. North American range of Fowler’s Toad
- Figure 4. Canadian records of Fowler’s Toad
- Figure 5. Minimum convex polygon encompassing the current range of Fowler’s Toads in Canada
- Figure 6. Estimates of abundance in the three populations of Fowler’s Toad in Canada
- Figure 7. Fowler’s Toad habitat characterization along the Lake Erie shoreline, west of the Niagara shoreline
- Figure 8. Fowler’s Toad habitat characterization along the Niagara shoreline
- Figure 9. Diminution in numbers of calling male Fowler’s Toads and the extent of invasive reeds, Phragmites australis, in the Crown Marsh of Long Point, Ontario
- Figure 10. Survivorship curve for post–metamorphic Fowler’s Toads based on mark–recapture data from Long Point, Ontario
- Figure 11. Mean extinction probabilities for the Long Point population of Fowler’s Toad populations simulated over 100 years based on 1000 iterations of the population viability model showing the relative effects of different levels of dispersal survival
- Figure 12. Mean extinction probabilities for each of the three Canadian Fowler’s Toad populations simulated over 50 years based on 1000 iterations of the population viability model. Additionally, the effect of habitat loss due to expansion of invasive Phragmitesreeds is simulated for Long Point
- Figure 13. Mean extinction probabilities for Canadian Fowler’s Toads estimated over 50 years based on 1000 iterations of the population viability model under differing scenarios of dispersal between populations (basins) and habitat loss modelled as due to spread of Phragmites
List of Tables
- Table 1. Dispersal rates, as percentages, used between the 17 subpopulations of the population viability model
- Table 2. Estimates for area (ha), percent of total area, and percent annual change in area occupied by invasive Common Reed, Phragmites australis, at Long Point from 1999 to 2006
- Table 3. Summary of population viability analysis models and results
- Table 4. Current federal legislation that can directly or indirectly protect Fowler’s Toads and/or habitat
- Table 5. Current Ontario provincial legislation that can directly or indirectly protect Fowler’s Toads and/or their habitat (adapted from Green et al., 2008)
Fowler’s Toad Anaxyrus fowleri

Endangered – 2010
COSEWIC -- Committee on the Status of Endangered Wildlife in Canada

COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2010. COSEWIC assessment and status report on the Fowler’s Toad Anaxyrus fowleriin Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 58 pp.
Previous report(s):
COSEWIC. 2000. COSEWIC assessment and update status report on the Fowler’s Toad Bufo fowleri in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. v + 25 pp.
Green, D.M. 1999. Update COSEWIC status report on the Fowler’s Toad Bufo fowleri in Canada, in COSEWIC assessment and update status report on the Fowler’s Toad Bufo fowleri in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1–25 pp.
Green, D.M. 1986. COSEWIC status report on the Fowler’s Toad Bufo woodhousii fowleri in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 51 pp.
Production note:
COSEWIC would like to acknowledge David M. Green and M. Alexander Smith for writing the status report on Fowler’s Toad (Anaxyrus fowleri) in Canada, prepared under contract with Environment Canada. This report was overseen by Ronald J. Brooks, Chair of the COSEWIC Amphibians and Reptiles Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment Canada
Ottawa, ON
K1A 0H3
Tel.: 819–953–3215
Fax: 819–994–3684
E–mail
Website
Également disponible en français sous le titre Évaluation et Rapport de situation du COSEPAC sur le crapaud de Fowler (Anaxyrus fowleri) au Canada.
Cover illustration/photo:
Fowler’s Toad -- Andrée Jenks, Guelph, Ontario.
© Her Majesty the Queen in Right of Canada, 2010.
Catalogue CW69–14/124–2010E–PDF
ISBN978–1–100–15927–0

Assessment Summary – April 2010
Common name
Fowler’s Toad
Scientific name
Anaxyrus fowleri
Status
Endangered
Reason for designation
This species only occurs on sandy beaches in three disjunct areas along the north shore of Lake Erie. It has disappeared from numerous historic sites on the Lake Erie shore and continues to decline in abundance and number of populations with further habitat loss and degradation due to invasive species (Common Reed, Zebra Mussels) and anthropogenic activities including shoreline development, beach cleaning, construction of breakwalls, bulldozing of beaches, vehicle use on beaches and agricultural and industrial contaminants. In addition, a Population Viability Analysis (PVA) model suggests that over the last decade, the probability of extirpation within 20 years has increased substantially.
Occurrence
Ontario
Status history
Designated Special Concern in April 1986. Status re–examined and designated Threatened in April 1999. Status re–examined and confirmed in November 2000. Status re–examined and designated Endangered in April 2010.

Fowler’s Toad Anaxyrus fowleri
Species information
Adult Fowler’s Toads, Anaxyrus fowleri, are 50 to 80 mm in snout–vent length, with females growing larger than males. The dorsum is grey or buff, with small warts and tubercles. The belly is white with a single, dark, pectoral spot. The male’s call is a rather shrill scream. Tadpoles are up to 27 mm in total length and are mottled grey and black. Fowler’s Toads have a distinctive smell, reminiscent of unroasted peanuts. Until recently, the species was assigned to the genus Bufo.
Distribution
Although Fowler’s Toads inhabit much of North America east of the Great Plains and south of the Great Lakes, they occur in Canada only along the northern shore of Lake Erie in extreme southern Ontario. The species is now restricted to Rondeau Provincial Park, Long Point, and the shore from Low Point to Fort Erie (Niagara). It is considered extirpated from numerous historic sites. The Canadian Extent of Occurrence, exclusive of uninhabitable shoreline and open water of Lake Erie, is approximately 1,200 km². Area of inhabited shore line is estimated at no more than 65 km².
Habitat
Fowler’s Toads occur along the Lake Erie shoreline where there are well–drained sandy soils or sand dunes, sandy beaches, sandy deciduous woodland, and rocky, poorly vegetated areas. Eggs and tadpoles need sparsely vegetated ponds, sandy bottomed pools, shallow rocky shoals or rocky pools. These habitats are unstable and are subject to the lake’s naturally dynamic processes of erosion and sand deposition, storms and fluctuating water levels. The complex life cycle of the toad requires both aquatic and terrestrial habitats. Fowler’s Toad habitats are in general decline, principally due to shoreline development, beach stabilization, vehicular traffic and recreational use at Niagara, and the spread of the invasive Common Reed, Phragmites australis australis, at Long Point and Rondeau Provincial Park. Much of the range of Fowler’s Toad in Canada is Crown Land, including provincial parks and national wildlife areas at Rondeau Provincial Park and Long Point, although 91% of the shoreline in Niagara is privately owned.
Biology
Adult male Fowler’s Toads congregate to call at aquatic breeding sites in late spring, to attract females. The larval period takes 40 to 60 days and newly metamorphosed toadlets emerge in midsummer. First–year growth is rapid and most individuals attain adult size in one year. Fowler’s Toadsappear to have a potential life span of 3 to 5 years, but annual mortality is high at all life stages.
Fowler’s Toads are insectivores specializing on ants and beetles. Snakes, particularly gartersnakes, Thamnophis spp., are the toad’s main predators and are undeterred by the toads’ noxious skin secretions. Fowler’s Toads otherwise avoid predation chiefly by digging beneath the surface of the soil.
Fowler’s Toads can be found nightly sitting in shallow water or on wet sand along the beach. Generally sedentary, most individuals move no more than a few metres along the shoreline throughout the year. However, a few individuals have moved over 10 km in a year.
Fowler’s Toads are known to hybridize with American Toads, Anaxyrus americanus, at Long Point, yet introgression appears to be limited.
Population sizes and trends
The total abundance of adult Fowler’s Toads in Canada cannot be determined with precision, primarily due to stochastic fluctuations in population size. The best estimate is an effective population size of under 5,000 adult toads. Rescue effect between populations, or from U.S.populations on the south shore of Lake Erie, is highly unlikely.
A population viability analysis, using the program VORTEX, and based on current distribution and estimates of abundance and demographic characteristics of Fowler’s Toads in Canada, indicates a greater than 20% probability of extirpation of Fowler’s Toads from Canada within 20 years, and nearly 100% probability of extirpation within 100 years, under all scenarios.
Limiting factors and threats
Fowler’s Toad is threatened principally by loss and degradation of its shoreline beach and dune habitat. Threats include: shore development and artificial coastline stabilization including construction of break walls, roads, parking lots, piers and groynes; intensive mechanical beach maintenance activities such as grading, grooming and clearing of algae using beach grooming machines or bulldozers; intensive human recreational use of beaches and dunes, including vehicular traffic; loss and degradation of habitats due to invasive species, particularly Zebra Mussels (Dreissena polymorpha) and Common Reed, and agricultural and industrial contaminants.
Special significance of the species
Perhaps the greatest significance of this species is that it has been the subject of the longest (>30 years) population and demographic study of a Canadian amphibian. This research has elucidated much of the nature of demographic, intrinsic, and extrinsic factors on population fluctuations and abundance in an anuran species. In addition, the impacts of humans on the species is now becoming understood. Adult Fowler’s Toads are important small insectivores, specializing in ants and beetles, whereas their tadpoles are significant detritivores in small ponds, rocky pools and embayments.
Existing protection
The majority of Fowler’s Toad habitat in the Rondeau Provincial Park and Long Point areas is located within protected areas where there is no direct threat from industrial or urban development. However, there is little direct legal protection for the species along the Niagara shoreline outside of James N. Allan Provincial Park. Currently, Fowler’s Toad is listed as Threatened under Schedule 1 of the federal Species at Risk Act and under Ontario’s Endangered Species Act 2007. It is also listed as a “specially protected amphibian” under Ontario’s Fish and Wildlife Conservation Act. The species is not listed as a species of concern in the United States nor in any of the states adjacent to Ontario.

COSEWIC History
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal–Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC Mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC Membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non–government science members and the co–chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2009)
- Wildlife Species
-
A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
-
A wildlife species that no longer exists.
- Extirpated (XT)
-
A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
-
A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
-
A wildlife species likely to become endangered if limiting factors are not reversed.
- Special Concern (SC)*
-
A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
- Not at Risk (NAR)**
-
A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
- Data Deficient (DD)***
- A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
* Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.
** Formerly described as “Not In Any Category”, or “No Designation Required.”
*** Formerly described as “Indeterminate” from 1994 to 1999 or “ISIBD” (insufficient scientific information on which to base a designation) prior to 1994. Definition of the (DD) category revised in 2006.
The Canadian Wildlife Service, Environment Canada, provides full administrative and financial support to the COSEWIC Secretariat.
COSEWICStatus Report on the Fowler’s Toad Anaxyrus fowleri in Canada – 2010.
Fowler’s Toad was formally described and distinguished from the American Toad, Anaxyrus americanus, by Hinckley (1882) after the Massachusetts naturalist S.P. Fowler realized in the 1860s that two sorts of toads inhabited his vicinity (Myers 1931; Dexter 1973; Green 1989). Until fairly recently, Fowler’s Toad was considered to be a subspecies of Woodhouse’s Toad (A. woodhousii), a more western form, but recent evidence indicates that this classification cannot be justified (Sullivan et al. 1996; Crother et al. 2008). Until the recent study by Frost et al. (2007), Fowler’s Toad and related species were placed in the genus Bufo (but see Pauly et al. 2004).
Adult Fowler’s Toads, Anaxyrus fowleri (Figure 1A), are 50 to 80 mm in length from snout to vent; females are generally larger than males (Green 1989, 2005). The dorsum is grey or buff coloured with darker spots and numerous small reddish–brown warts on a granular, textured skin. There are usually three or more small warts per dark dorsal spot, although this is not always a reliable diagnostic character as the spots may be small in some specimens, particularly in Canada. The Canadian populations are subtly distinguishable from each other in that individuals from Niagara tend to have fairly bold dorsal spots whereas individuals from Long Point tend to be more uniform in colour and have smaller, less distinct dorsal spots. The under surface is finely granulated and white or cream coloured, and is either without spots or with only a single dark pectoral spot situated between the forelimbs. The throat is dark in adult males but white in females and juveniles. The snout is short and blunt and the bony cranial crests on the head are weak.
Figure 1. Fowler’s Toad. A) Male, Long Point, Ontario (D.M. Green, photo). B) Tadpoles, Gravelly Bay, Niagara (Rob Tervo, photo).
A

B

The call of the male Fowler’s Toad has been described variously as a “prolonged and rather shrill scream”, an “almost agonized wail”, a “penetrating droning scream”, a “weird, wailing scream” or “a muffled scream from a small and distressed sheep” (Green 2005). Call characteristics vary with temperature; as the temperature goes up, call pulse rate increases and call duration declines (Zweifel 1968). The dominant frequency is negatively correlated with body size. Males also issue a grumbling, vibrating release call when handled, whether by humans or by other toads (Brown and Littlejohn 1972), serving to announce their sex if clasped by another male. Females are silent.
Eggs are 3 to 4 mm in diameter and are laid in long twin strands. Counts of ovarian eggs range from 2,000 to 10,000 per female, but the average clutch size for toads at Long Point, based on direct counts of eggs laid, is around 4,200 (Green 2005). Larger females tend to have more eggs. Eggs hatch no more than one week after oviposition. The tadpoles are mottled grey and black (Figure 1B), with an emarginate oral disc and dorsally placed eyes. They do not exceed 27 mm in total length. The ratio of tail height to tail musculature height is more than 2.00, distinguishing them from the tadpoles of the American Toad (Hinckley 1882).
Fowler’s Toads have a distinctive smell, reminiscent of unroasted peanuts (Green 2005) or, according to Miller and Chapin (1910), Ailanthus (Tree–of–Heaven) wood.
There is significant phylogeographic structure to the genetic variability of Fowler’s Toad populations in Canada (Figure 2). There are two distinct mitochondrial DNA (mtDNA) lineages, or phylogroups, among Canadian Fowler’s Toads, based on mtDNA sequence variation in the control region (d–loop) and cytochrome c oxidase 1 (CO1), among 161 individual toads from 21 localities (Smith and Green 2004). The two control region sequences diverge by 6%. Phylogroup 2 includes Fowler’s Toads at Long Point as well as central and southern U.S. Fowler’s Toad populations whereas Phylogroup 1 includes the rest of the Fowler’s Toad populations surrounding Lake Erie. The mtDNA of Phylogroup 1 Fowler’s Toads is almost identical to the mtDNA found in American Toads in the same region. There is evidence of further geographic variation in mtDNAs within the Fowler’s Toad over large geographic scales (Smith and Green 2004).
Of the several possible explanations for this unusual mtDNAdiversity in the Lake Erie region, introgression of American Toad mtDNA into Fowler’s Toad populations via past hybridization is the most plausible (Smith and Green 2004). The variation in the mtDNA control region and CO1 sequences among Phylogroup 1 Fowler’s Toads (Smith and Green 2004) indicates that this introgression is not recent. A further three subdivisions among Phylogroup 1 Fowler’s Toads are recognizable at below the level of 0.5% sequence divergence (Figure 2), corresponding roughly to the basin structure of Lake Erie and the three areas of Rondeau, Long Point, and Niagara where the toads occur and within each of which there is little genetic variation (Smith and Green 2004). This is indicative that Rondeau Provincial Park, Long Point and Niagara function as three separate populations because both genetic and demographic estimates of neighborhood size are larger than the areas occupied by the many, seemingly discrete breeding localities.
Figure 2. Neighbour–joining tree of sequence divergence (p–distance) from control region mtDNA of Fowler’s Toads of populations from the northern edge of their distribution. Bootstrap values > 80% are shown. From Smith and Green (2004).

The retention of what appears to be an ancestral Fowler’s Toad mtDNAhaplotype among Long Point Fowler’s Toads (Phylogroup 2) is probably the result of multiple invasions of Fowler’s Toads into the Lake Erie basin following the Pleistocene glaciation (Smith and Green 2004). Some corroboration for this interpretation comes from nuclear DNA sequence information, although it is not as variable as the mtDNA sequences and no sequence variation has been found between Fowler’s Toads from populations of Long Point, Rondeau Provincial Park and Niagara at either of the nuclear genetic loci, Rag–1 (Recombination–activating gene) and 18S rRNA (Smith and Green in prep.). However, there is a single nucleotide difference in 18S between these Fowler’s Toads and American Toads, which is an indication of a diagnostic single nucleotide polymorphism (SNP) that differentiates the two species. Thus this nuclear gene region shows a different picture from the mtDNA sequences in that they unite the Fowler’s Toads of two mtDNA phylogroups to the exclusion of American Toads. This can only be reconciled with the mtDNA phylogroup structure of Canadian Fowler’s Toad populations if there had been past hybridization of Fowler’s Toads with American Toads that resulted in the introgression of American Toad mtDNA into the Fowler’s Toads that gave rise to Phylogroup1.
The three extant populations of Fowler’s Toads in Canada (Rondeau, Long Point and Niagara) are genetically distinct and, to the practised eye, morphologically distinguishable. There is no evidence of any significant, ongoing genetic exchange between the three populations (Smith and Green, 2004), and each population shows little intrapopulation variation. There is genetic distinction between the Long Point toads and those from the other two areas, arising from a past hybridization with American Toads that occurred before the Long Point toads invaded Lake Erie. There is disjunction among the three populations, but it isn’t possible to decide if this is from natural or anthropogenic causes. Similarly, the genotypes of the extirpated populations (e.g., Pelee Island, Point Pelee, Niagara area sub–populations) are unknown so the original phylogeographic pattern along Lake Erie is also uncertain. The three populations occupy the same ecozone. In terms of significance, again the picture is murky. There is mtDNA sequence variation, but not nuclear DNA sequence variation. The importance of the mtDNA variation is unclear, so it is also unclear whether the loss of one of the populations would constitute a significant loss to the species in Canada. The populations do not exist in unique ecological settings. The threats to continued existence at Niagara differ somewhat from those affecting populations at Rondeau Provincial Park and Long Point; however, again there is overlap in threats among the three areas (See Limiting Factors and Threats). Therefore, the three populations could qualify as separate designatable units, or one could also dispute this decision. Regardless of what decision is made, it is clear that each population would qualify separately or as part of the whole species as Endangered, so there would appear to be negligible assessment or conservation value added in assessing two or three separate DUs, and given the conflicting evidence the most parsimonious decision is to treat the three populations as a single DU.
Fowler’s Toad inhabits much of North America east of the Great Plains and south of the Great Lakes (Green 2005), though it is absent from the southern Atlantic coastal plain from the Carolinas to northern Florida (Figure 3). On the western edge of the range, the considerably larger, more western Woodhouse’s Toad intergrades with Fowler’s Toad along a line south from Lake Michigan to the coast of the Gulf of Mexico at the Texas/ Louisiana border (Meacham 1962). The species’ northern range limit encompasses the west side of Michigan’s Lower Peninsula, reaching almost to the Mackinac Strait, the shores of Lake Erie in Ohio, Ontario and northern Pennsylvania and extends though New Jersey, southeast New York and southern Vermont and New Hampshire to reach the Atlantic coast almost to Maine (Green 2005). Populations along the southern shore of Lake Erie in Pennsylvania and Ohio are not continuous with other populations of the species further south. Recent evidence indicates that these populations are related to, and possibly derived from, Canadian populations on the north shore of the lake and are subject to the same threats (Smith 2004).
Figure 3. North American range of Fowler’s Toad (after Conant and Collins, 1998).

Fowler’s Toadis documented in Canada only from sandy or rocky points and sandy beaches along the northern shore and islands in the western basin of Lake Erie in extreme southern Ontario (Figure 4). It is now restricted to Rondeau Provincial Park, Long Point and the eastern (Niagara) Lake Erie shore from Low Point (James N. Allan Provincial Park) to Fort Erie. It has disappeared, and is considered extirpated, from numerous historic sites. For example, Fowler’s Toad was last recorded at Fisher’s Glen in 1927, Point Pelee in 1949, Turkey Point in 1956, Pelee Island in 1960, and Rock Point (and adjacent Port Maitland) in 1986 (Green 2000, Green et al. 2008, Yagi and Tervo 2006a).
Figure 4. Canadian records of Fowler’s Toad. Based on georeferenced data from the Ontario NHIC (historical records) and from the work of Sandy Dobbyn (Rondeau), Alex Smith (Long Point), David M. Green (Long Point) and Anne Yagi (Niagara). Map by Sara Lourie.

To estimate the extent of occurrence (EO) of Fowler’s Toads in Canada, a minimum convex polygon encompassing the three populations of Rondeau Provincial Park, Long Point and Niagara can be constructed (Figure 5). This EO is 4670 km². However, the shoreline between the three populations consists largely of bluffs or other unsuitable habitat for the toads and the polygon extends over a considerable area of open water of the lake. Excluding these areas from the calculation, the EO is approximately 1,200 km².
Figure 5. Minimum convex polygon encompassing the current range of Fowler’s Toads in Canada, distinguishing the habitable area within the polygon usable for calculation of extent of occurrence (EO) from uninhabitable area including open lake.

No verified records of Fowler’s Toad in Canada are from more than half a kilometre from the Lake Erie coast. Accordingly, the species can be considered to occupy a virtually linear range, with the length of inhabited coastline (Figures 6, 7), consisting of Type 1 and Type 2 habitats as defined by Yagi and Tervo (2006a,b), as the logical basis for estimating a biologically meaningful area of occupancy (AO). These habitats occupy a total of about 129 km of the Lake Erie shoreline (64 km from Low Point to Fort Erie, 42 km from Hahn Beach to the Long Point tip and 23 km at Rondeau Provincial Park from Erie Beach to the coast south of Morpeth). Assuming a rather generous occupied zone about 500 m wide, the AO is no more than 65 km². A superimposed 1 x 1 km grid to estimate the index of area of occupancy (IAO) provides a maximum estimate of 129 x 0.5 = 120 km². An IAO based on a 2x2 km² provides an estimate of 480 km². Again, this is a maximum estimate as it assumes that there is no overlap among grid squares and that the entire coastline is occupied by toads. Neither assumption is correct.
Figure 6. Estimates of abundance in the three populations of Fowler’s Toad in Canada. A) Annual spring survey of males along ca. 8 km of shoreline at the base of Long Point. Estimates are based on intensive mark– recapture data using nightly surveys over 6 weeks in May through early June and calculated using the M(th) estimator in the program CAPTURE. B) Summer surveys of all toads > 48 mm at Nickel Beach, Niagara, using a Lincoln–Peterson estimate based on weekly surveys. C) Summer surveys of all toads > 48 mm at Rondeau, covering the entire shoreline inhabited by toads, using a Lincoln–Peterson estimate based on three surveys during the summer. No suvey was conducted at Rondeau Provincial Park in 2006.

Figure 7. Fowler’s Toad habitat characterization along the Lake Erie shoreline, west of the Niagara shoreline. Habitats are evaluated from Type 1 (toads present in suitable habitat) to Type 4 (wholly unsuitable habitat). Adapted from Yagi and Tervo (2006).

Figure 8. Fowler’s Toad habitat characterization along the Niagara shoreline. Habitats are evaluated from Type 1 (toads present in suitable habitat) to Type 4 (wholly unsuitable habitat). Adapted from Yagi and Tervo (2006a).

Fowler’s Toads occur in areas with loose, well–drained gravelly or sandy soils, including sand dunes, sandy deciduous woodland, and rocky, poorly vegetated areas (Hubbs 1918; Smith 1961; Minton 1972; Brown 1974; Green 1989; Klemens 1993). Wright and Wright (1949) noted that in the eastern United States, Fowler’s Toads were common along roadsides, near homes, and in fields, pastures, gardens, and sand dunes. They are a typical species of the New Jersey Pine Barrens (Zampella and Bunnell 2000). In comparison to American Toads, which occur in wet shady woodlands and uplands, Fowler’s Toads prefer dry scrub, sand dunes and open country (Lazell 1976; Klemens 1993).
The complex life cycle of Fowler’s Toad involves the use of both aquatic and terrestrial habitats (Tupper and Cook 2008). Eggs and tadpoles need sparsely vegetated, stillwater ponds, sandy bottom pools, shallow rocky shoals or rocky pools that endure at least until midsummer in order to complete development. Juvenile and adult toads inhabit sandy riverside and lakeshore habitats with bare to sparse vegetation cover and need sand dunes for hibernation, open beach for feeding and re–hydration and sandy beaches and foredunes for hiding during the day during the active season. Thus, Fowler’s Toad require five habitat types: a) hibernation habitat, b) breeding, egg–laying and tadpole development habitat, c) feeding and hydration habitat, d) daytime retreat and aestivation habitat and e) dispersal corridor habitat. All of these must be in close proximity to sustain a population.
Fowler’s Toads must dig into sandy areas to avoid extreme temperatures and harsh environmental conditions. These areas include sand bars, beaches and dunes along the Lake Erie shoreline. The occupied burrow is a subterranean space in open to moderately open sandy areas which are sufficiently deep and well drained that the toad can dig below the frost line to just above the water table and remain over winter (7 to 8 months from late September to mid–May) (Green et al. 2008). They are not freeze–tolerant like some frogs, and therefore to hibernate must dig deep enough to be below the frost. In Niagara, in sandy soils along the Lake Erie shoreline, frost depth was greater than 2 m during more severe winters (Yagi 2008). The burrow is moist and facilitates aerobic metabolism, but toads can not sit in water because they will continue to absorb through their skin and bloat. Therefore fluctuations in the groundwater table must not reach the burrow, or the toads will drown (Green et al. 2008). Given the proximity of these burrows to the shoreline and the occurrence of high water levels in winter, there is a thin zone between frost and the water table which may be of major importance to survivorship over winter (S. Hecnar pers. comm. Jan 2010).
Fowler’s Toads breed in the shallow, still water of semi–permanent ponds, flooded low ground, temporary pools, rocky headland pools and lakeshores in shallow bays (Wright and Wright 1949; Smith 1961; Mount 1975; Collins and Wilbur 1979; Green and Pauley 1987; Breden 1988; Dundee and Rossman 1989; Tupper and Cook 2008). Breden (1988) and Smith and Green (2004) described breeding ponds used by the toads as shallow with sandy bottoms and gradually sloping banks, vegetated primarily with sedges and bulrushes. Breeding sites are open to moderately vegetated oligotrophic sandy bottom ponds, wetlands or pools or bedrock pool areas located within the full range of Lake Erie Water levels (Green et al. 2008).
During the active season in sand–dune areas such as at Long Point and Rondeau, Fowler’s Toads use the lower beach near the water for night–time foraging and hydration and the upper beach and foredune as retreats during the day (Green 2008). Animals may use particular hiding places during the day, such as driftwood logs, or else bury themselves in sand down to the level of the damp sand layer. However, toads may also rest in the dry sand at the surface with just the head or head and back exposed.
At Rondeau Provincial Park and Long Point, the human impact on beaches and foredunes is not so profound as at Niagara (Green et al. 2008; Yagi and Tervo 2008). At Niagara, beaches are hardened by vehicular traffic and dunes are reinforced and stabilized. Thus, daytime retreats usable by toads are much fewer, and toads should be expected to be more philopatric to their hiding places. Toads along the Niagara shoreline are likely to hide in crevices and fissures in rock walls and breakwaters.
Fowler’s Toad is adapted to early stages of ecological succession in sand dune and lake shore habitats (Breden 1988; Green 2005). These habitats are unstable and subject to the lake’s naturally dynamic processes of erosion and sand deposition, storms and fluctuating water levels. Severe, destructive storms that can drastically alter the shoreline are common on Lake Erie (Hazen 2000). When water levels are high, beaches and dunes erode and the sand gets moved and re–deposited by alongshore drift. Dune blowouts caused by storms are an important source of the open, sandy habitat preferred by Fowler’s Toads. When water is calm and levels are low, the same dune and beach areas accumulate sand. Fowler’s Toad is adapted to this changeable environment, and these natural processes are not a threat to the toad.
As an example of the dynamism of the Lake Erie shoreline, the evidence of Long Point’s history of constant change is readily apparent (Stone 1988, Hazen 2000). In April and again in December 1985, high water and storm winds inundated and destroyed large sections of dune and foreshore at Long Point, including some 56 cottages (Hazen 2000). The dune blow–outs caused by storms created gaps in the beach face and covered portions of the marsh with tracts of new sand. In this way, although the storm decimated the hibernating population of Fowler’s Toads, it also created new, open habitat and sandy–bottomed ponds which were used by the surviving adults for breeding. Yet these alterations were only temporary and much of the open habitat has since become overgrown. A small clean–bottomed pond at the CWS Thoroughfare Point Unit, formed by the 1985 storm and favoured by the toads, had by 1996 become completely choked with bulrushes, cattails and, now, reeds. Toads did not sing from this pond after 1994 (D.M. Green pers. obs.).
The sand dune and beach habitats required by Fowler’s Toads are affected and maintained by the movements of sand along the Lake Erie shoreline. Rondeau Provincial Park and Long Point are sand spit formations that are dynamically maintained by Lake Erie storms, fluctuating water levels, and littoral drift processes. Beaches and dune systems on the Niagara shoreline form within bays located between rocky headlands, which affect beach erosion and sand deposition rates. Longshore transport deposits sand in the bay and then waves and wind action push the sand onto the beach. Wind deposits loose, dry sand onto the dunes. Therefore, events and landscapes at some distance from the immediate areas where the toads are found have tangible effects upon toad habitats. The movement of large amounts of sand regenerates, reforms and shifts beaches and dunes. Beach sand is moved along the shoreline by wave action, wind and currents and dunes are formed from wind–blown sand. Many of the large dunes inland of Lake Erie formed 4,000 to 6,000 years ago during the post–glacial Nipissing period and are important sources of sand. In the Point Pelee area, where Fowler’s Toads are now extirpated, historic sand dredging operations off the tip as well as increasing shoreline protection measures on either side of Point Pelee National Park resulted in reduced sediment delivery to the shoreline and may partly explain their extirpation.
Numerous anthropogenic habitats, particularly along the Niagara shoreline, are periodically or incidentally used by Fowler’s Toads. These include drain outlets, ponds, wetlands, culverts, roads, buildings, cracks within shore wall structures, gardens, boats, docks, decks, debris, loose materials and litter, sand and algae piles. Many of these features may be “behavioural sinks” in that they are incidentally used by toads because they are located between needed habitat features but interfere with normal dispersal and movement patterns. Toads appear to be present at these sites only because they lie within the species’ activity range, not because they are favoured habitat. In other cases, anthropogenic habitat alterations may appear to attract toads. For example, a portion of the Casey Drain at Morgan’s Point and the artificial waterfowl feeding ponds in Long Point Provincial Park are known to attract breeding choruses of toads (Yagi et al. 2006). How successful they are as breeding habitat in terms of the recruitment of young toads is unknown. If they attract breeding adults, but are unsuitable for the successful growth and metamorphosis of tadpoles, they are detrimental population sinks.
The habitats required by Fowler’s Toads are in general decline, principally because of shoreline development, beach stabilization, vehicular traffic and recreational use at Niagara and the spread of the invasive Common Reed Grass, Phragmites australis australis, at Long Point and Rondeau Provincial Park.
The Crown Marsh at Long Point was previously a highly productive and well–populated breeding site for Fowler’s Toads (Green 1981, 1989) but has not been used by Fowler’s Toads as a breeding site for many years (Green and Sanderson 2006, 2007a; Green and Summerfield 2008). The site used by the Fowler’s Toads is immediately north of Erie Boulevard (Hwy 59), between the cross–streets of Winston Pkwy and Pines Pkwy at Long Point (Figure 9). Toads have bred at the site both during times of low water, as during the early 1990s and high water, as in the late 1990s. The area is a sandy–bottomed former dune blowout and was dominated by cattails and low sedge. Phragmites initially invaded during the 1990s and since about 2003, it has completely dominated the site (D.M. Green pers. obs.; Meyer 2003; Badzinski et al. 2008).
Figure 9. Diminution in numbers of calling male Fowler’s Toads and the extent of invasive reeds, Phragmites australis, in the Crown Marsh of Long Point, Ontario. A) Observed calling males in the Crown Marsh, 1990–2009 (D.M. Green, unpublished). B) Extent of Phragmites reeds and other vegetation types in 2006 in the Crown Marsh area formerly used by the toads (box) (after Badzinski et al. 2007). No toads called from the Crown Marsh from 2005 on.
A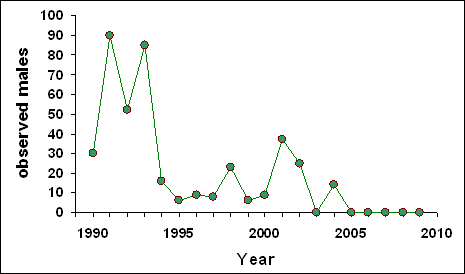
B 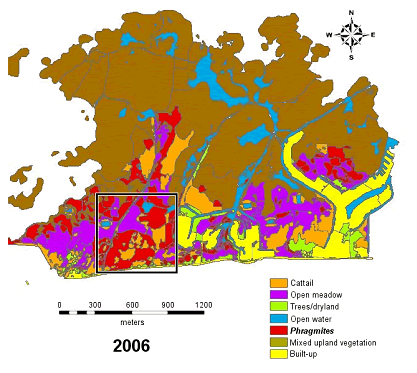
B
Figure 10. Survivorship curve for post–metamorphic Fowler’s Toads based on mark–recapture data from Long Point, Ontario.

Loss of habitat appears to be the underlying cause of the disappearance of Fowler’s Toads from Rock Point Provincial Park (RPPP) as well (A. Yagi, pers. comm.). In the mid–1980s the dunes at Rock Point were severely eroded by Lake Erie storms. The park managers began planting the remaining dunes to protect the shoreline without engineered revetment. Fowler’s Toads have not been seen since the mid–1980s at RPPP. In 2003, a careful removal of some of the vegetation was begun on the dunes in an attempt to open up the dunes to attract toads. In addition, several truck loads of Zebra Mussel (Dreissena polymorpha), and Quagga Mussel (D. rostriformis bugensis) shells were removed from the rocky pool areas fronting the dunes to improve breeding site quality. The re–colonization of the RPPP area is important for the James N. Allan site, the closest western site.
Much of the range of Fowler’s Toad in Canada is Crown Land, including provincial parks and national wildlife areas (NWAs). The species is extirpated from Turkey Point Provincial Park and Point Pelee National Park, and may now also be absent from Rock Point Provincial Park.
Within Rondeau Provincial Park, there are some 290 privately owned family cottages, two churches, a yacht club and a community centre on land leased from the park. Cottages increase roadkill, invasive plants on the dunes, and loss of oak–savannah habitat to crushed rock imported for parking, driving and recreational areas. The leases expire in 2017, at which time all must be demolished (Welsman 2009).
Long Point is contained within the Long Point Biosphere Reserve. Most of the point is protected area, consisting of Big Creek and Long Point National Wildlife Areas controlled by the Canadian Wildlife Service, Long Point Provincial Park in two sections, and the tip of Long Point controlled by the Federal Ministry of Transport. The Long Point Company maintains and preserves a large central section of the point as a private hunting and fishing reserve. The Anderson Property occupies a much smaller portion of the north side of the outer point. The Crown Marsh, administered by CWS lies on the Inner Bay side of the point near the old section of Long Point Provincial Park. At the base of the point from the Big Creek Marsh on the west to the new section of Long Point Provincial Park are many private cottages, a motel and restaurants. Several privately owned marinas occupy the north side of the point on the Inner Bay.
Unlike Rondeau Provincial Park and Long Point, 91% of the eastern Lake Erie shoreline near Niagara is privately owned (Yagi and Tervo 2006a). Many seasonal summer cottages line the shore and it is popular for summer recreation. Although there are also small provincial parks and conservation areas, most of the Point Abino peninsula is privately owned by American seasonal cottage owners. Adjacent Sherkston Shores is a privately owned seasonal campground although it is becoming a more permanent trailer park. Crystal Beach, east of Point Abino, was a popular amusement park that closed in 1989 (Jackson 1967). It is now the site of the Crystal Beach Tennis and Yacht Club gated community. The town of Fort Erie occupies the eastern end of the Niagara shoreline and contains Fowler’s Toad habitats at Windmill Point. The town of Port Colborne straddles the Lake Erie entrance to the Welland Canal and includes Nickel Beach, which is owned by INCO. James N. Allan Provincial Park at Low Point, Rock Point Provincial Park, Long Beach Conservation Area and Morgan’s Point Conservation Area all receive heavy recreational use during summer (Yagi et al. 2006).
Fowler’s Toads congregate at breeding sites in late spring, depending upon conditions of temperature and rainfall (Green 2005a). The male toads sing at a minimum body temperature of about 13°C. On Lake Erie, Fowler’s Toads begin to sing any time from the last days of April to the last week of May, continuing, depending on the weather, for three to four weeks. Calling Fowler’s Toads usually can be heard at Long Point until early June. In the Niagara area, chorusing typically begins a few weeks later in the year, which corresponds with prolonged cooler water temperatures associated with the eastern basin of Lake Erie (Yagi 2008). Breeding congregations comprise chiefly chorusing males, who respond to each other’s vocalizations. Females orient to particular males’ calls based on persistence and intensity (Given 1996). In general, female toads are attracted to males that call more frequently regardless of temperature (Sullivan 1982, 1983). Not all males in a breeding aggregation will call, however, thus the intensity of a chorus is not a good indicator of the actual number of animals present (Shirose et al. 1997).
Tadpoles may be observed in large aggregations in stream and pond habitats, typically in standing water and resting on a muddy or sandy substrate. The larval period takes 40 to 60 days and tadpoles metamorphose at about 8 to 12 mm snout to vent length (SVL). Logier (1931) reported recently metamorphosed individuals present at Long Point on 13 July 1929. At Nickel beach near Port Colborne, metamorphs were observed as early as 25 June 2007 (SVL 10.7 to 28.8 mm) and as late as 25 July 2006 (Yagi 2008).
First–year growth is rapid. Clarke (1974b) reported an average 6.58–fold increase in size during the first year in Connecticut. Females grow significantly faster than do males and thus reach a larger size (Clarke 1974b). Most males begin to show secondary sexual characteristics about one year after metamorphosis. Females attain adult size about one month later, but also generally at two years of age (Breden 1987). A variable proportion of the animals within a year’s cohort achieve sexual maturity in their first year (Kellner and Green 1995). In other years, however, maturity may be delayed such that some individuals do not become sexually mature until age 3.
Fowler’s Toadappears to have a potential life span of 3 to 5 years (Clarke 1977; Kellner and Green 1995), but annual mortality is high at all life stages (Green 1999a). Survivorship of tadpoles to one year old was estimated to be only 0.0013 (Green 1999a). It is not known how much that is affected by survival of the tadpoles until metamorphosis or by subsequent survival of the toadlets through their first winter. Clarke (1977) found 30% annual survival among post–metamorphic individuals at a site in Connecticut; highest mortality was among young–of–the–year juveniles prior to their first winter. Among 17,000 electrophoretically identified tadpoles introduced into an Indiana population, Breden (1987) recovered 8,539 postmetamorphic toadlets and, of these, recaptured 37 as breeding adults. Green (1999) calculated 27% annual survivorship among adults.
Fowler’s Toad is an insectivore, specializing in ants and beetles (Judd 1957; Bush and Melnick 1962). During the active season, they forage on the beach and dunes at night. Tadpoles are significant detritivores in small ponds, rocky pools and embayments.
Snakes, particularly gartersnakes, Thamnophis spp., are the main predators of toads, including tadpoles and toadlets. Common Gartersnakes (Thamnophis sirtalis) are quite abundant on Long Point and Eastern Hog–nosed Snakes (Heterodon platirhinos), now threatened in Ontario, are specialized toad eaters. Northern Watersnakes (Nerodia sipedon) are also predators of toads. Raccoons (Procyon lotor), Largemouth Bass (Micropterus salmoides), Bullfrogs (Lithobates catesbeiana) and some birds, such as bitterns and owls, have also been observed to take Fowler’s Toads. Ring–billed Gulls (Larus delawarensis) at James N. Allan Provincial Park were observed eating adult male and juvenile toads respectively from the Niagara population (Smith and Green 2002, 2005a). Evidence of predation by gartersnakes and Raccoons comes from recent telemetry studies of the Niagara toad population (Limnoterra 2006). A gartersnake consumed a telemetered toad and was tracked for several days at Nickel Beach in Port Colborne. Another transmitter was recovered from an arboreal site several metres above the ground at Morgan’s Point Conservation Area. The transmitter and skin remains of the toad were recovered from a crotch in the tree suggesting Raccoon predation. Raccoons, instead of owls, were suspected because they are not tolerant to the skin toxins but are known to eat the belly of the toad and leave the skin (Green pers. obs.). Potential predators may often injure toads. Nematode worms parasitize Fowler’s Toads (Green pers. obs.), but there are no data on mortality rates caused by such infections.
Fowler’s Toads will often remain immobile when disturbed, especially when threatened by snakes. The usual fright reaction of a toad is to inflate its lungs, elevate its body and lower its head towards the potential threat. The noxious and toxic skin secretions render toads generally unpalatable to most potential predators. The snakes, Thamnophis, Nerodia and Heterodon,though, all have substantial tolerance to toad skin toxins.
Fowler’s Toads avoid predation chiefly by digging out of sight or by camouflage. The light grey, mottled dorsum blends in well with sandy substrates. This behaviour was confirmed on several occasions during telemetry studies in the Niagara Area (Limnoterra 2006; Yagi 2008) and Long Point (Green 2008). During the day, toads were found in terrestrial sites just under the sand surface, in burrows at the crest of dunes, under shoreline debris (logs, washed up vegetation), in bedrock fissures and under rocks.
Postmetamorphic Fowler’s Toads, like all terrestrial amphibians, are continuously challenged with maintaining an optimal water balance (Hillman et al. 2009). During the active season, they accomplish this by moving from daytime hiding places, which are under cover in cool dry sandy areas where they lose moisture, to the Lake Erie Shoreline at night where they reabsorb water through their skin. They are found nightly often in large numbers, sitting in shallow water or wet sand along the shoreline during the active season. They may stay there for only a few minutes or longer depending on the weather conditions (Yagi et al., 2008).
Most Fowler’s Toads move small distances along the shorelines of lakes or large ponds but they are not territorial in any way (Green 2008). On Lake Michigan and Lake Erie, they emerge in the evening from under the sand up to 400 m from the water’s edge and move to the beach to rehydrate and forage over about 8 m of shorefront. On rainy evenings, they may be observed actively foraging over the beach and foreshore dunes. Generally sedentary, about 70% of individuals, both juveniles and adults of both sexes, move no more than a few metres laterally along the shoreline from day to day or even month to month (Smith and Green 2004). However, both adults and juvenile toads are capable of long–distance dispersal, as about 2% of individuals move over 10 km a year and even up to 35 km at Long Point (Smith 2004; Smith and Green 2006). At Nickel beach near Port Colborne, adult telemetered toads (n = 356) seasonally moved over 1 km along the shoreline from breeding sites to overwintering sites with average seasonal movements of 70 m per year and mean daily movements of 12 m/day (Yagi 2008). After several years of mark recapture data there is evidence that a small percentage of toads marked outside Nickel Beach have been recaptured at Nickel beach (Planck and Blott, 2006; Yagi 2008). This is consistent with DNA evidence that demonstrates the genetic similarity within the Niagara population region (Smith and Green 2004).
Both juveniles and adults can disperse widely and can recolonize habitats after local extinctions provided there are no barriers placed in the way of their movements along the beach.But there are such barriers (Figures 6 and 7). The presence of steep bluffs along much of the Lake Erie coastline limits areas where there may be suitable breeding sites or foraging habitats available. Lack of such avenues for dispersal may explain, in part, the continuing absence of the species from Pelee Island and Point Pelee, which are isolated from the rest of the Canadian range.
Hybrids between the American Toad and Fowler’s Toad have been noted in various places in eastern North American, including Long Point, Ontario, since early this century (Green 1984; Green and Parent 2003). The American Toad is generally largerand commonly has a profusely spotted belly and comparatively large warts. The call of the American Toadis a long pulsed trill easily distinguished from the bleat of a Fowler’s Toad (Zweifel 1968; Green 1982). The differences between the two species are summarized by Wright and Wright (1949) and Zweifel (1968). Hybrids are viable and fertile yet introgression at Long Point appears currently to be limited (Green 1984; Green and Parent 2003). It appears that the extent of hybridization may vary year–to–year and locality–to–locality depending upon local conditions.
Fowler’s Toad is a lakeshore and sand dune specialist at the northern limit of its range in Canada where it is restricted to a limited number of suitable sites along the shoreline of Lake Erie. Because it breeds preferentially in oligotrophic, sandy–bottomed ponds in early successional habitats, it depends upon breeding sites that are continually created or maintained by disturbance lest they be overgrown and eutrophic due to ecological succession. Both adults and juveniles disperse widely (Clarke 1974a; Breden 1987, 1988; Smith and Green 2005b, 2006) and can recolonize habitats after local extinctions provided there are no barriers placed in the way of their movements along the beach.
Fowler’s Toad is perhaps the most thoroughly studied and surveyed amphibian in Canada. The extent of this work provides a comprehensive estimation of changes in the approximate size and constitution of Fowler’s Toad populations.
Green (1981) undertook an extensive search for Fowler’s Toads along the whole north shore of Lake Erie from 1978 to 1981. During 2001–2003, Alex Smith also carried out an extensive search for Fowler’s Toads (Smith 2004), and Anne Yagi and her staff (OMNR) surveyed the Lake Erie coast to characterize suitable habitat (Yagi and Tervo 2006a,b).
Intensive surveys at Long Point, Rondeau Provincial Park and Niagara have also been done. Since 1988, the ecology and abundance of Fowler’s Toads at the base of Long Point has been annually surveyed (Laurin and Green 1989; Green and Cantin 1990; Green and Porebski 1991; Green et al. 1991, 1993, 1995; Green and Chan Tang 1994; Green and Rabinowitz 1996; Green 1997a, 1999; Green and Hensley 1998; Green and Bol 1999; Green and Smith 2000, 2003; Green and McTavish 2005; Green and Sanderson 2006, 2007b,c; Green and Summerfield 2008). The Long Point surveys have largely taken place during the emergence and breeding seasons (May–June), to estimate and describe the total population each year and thereby come to an understanding of demographic parameters. Population surveys using mark–recapture have been done at Rondeau Provincial Park in 2004, 2005, 2007 and 2008 (Dobbyn 2005, 2008) and at Morgan’s Point, Gravelly Bay and Nickel Beach on the Niagara shoreline (Yagi and Mills 2003; Yagi et al. 2004; Yagi and Tervo 2006a; Yagi 2008). There has also been radio–tracking of individual toads at Gravelly Bay (Yagi 2008) and on the Thoroughfare and Hahn beaches at Long Point (Sanderson and Green 2007; Green 2008).
Despite the intensive surveys, the total abundance of adult Fowler’s Toads in Canada cannot be determined with any precision. This is primarily due to stochastic fluctuations in population size in all three populations (Figure 6). Stochastic episodes of rise and decline within populations reduce the effective size of those populations relative to the actual number of animals that may be present. This is more important to estimating long–term survivorship than are population maxima or a long–term mean population size (Meffe and Carroll 1994). In such highly variable populations, the harmonic mean abundance, rather than the arithmetic mean, is the better approximation of effective population size.
In 4 years of study at Rondeau Provincial Park, estimated numbers of adult toads ranged from 195 ± 64 (S.E.) to 1401 ± 107 (Dobbyn unpublished data), for a harmonic mean of 386. At the study site at the base of Long Point between Big Creek Marsh and the CWS Thoroughfare Point Unit, estimated numbers of males ranged from 12 ± 0 to 515 ± 62 per year over 21 years (Green and Summerfield 2008), for a harmonic mean of 63. Doubling that to account for females (at a 1:1 sex ratio) and assuming that this is one of about eight areas of productivity (breeding localities) for the toads at Long Point results in a harmonic mean estimate of about 1,090 adult toads at Long Point. At Nickel Beach, Niagara, the estimated numbers of adult toads ranged from 257 ± 21 to 861 ± 126 over 6 years (Yagi 2008); the harmonic mean is 420. Assuming that Nickel Beach is one of about eight areas of productivity at Niagara results in a harmonic mean estimate of about 3,360 adult toads at Niagara. The total effective population size estimated this way is thus under 5,000 adult toads.
Fowler’s Toad populations exhibit large fluctuations in population size even from one year to the next (Figure 6). Such fluctuations have been observed in many other species of pond–breeding anurans, especially those with short life expectancies and high early mortality rates (Berven 1990, 1995; Green 1997b; Pechmann et al. 1991; Green 1997b). Fowler’s Toads rarely live more than 3 years post–metamorphosis. Only one 5–year old toad has ever been determined in the Long Point population by skeletochronology (Green unpublished data); most breeding adults are 2 years of age (Kellner and Green 1995). Based on calculated survival rates at Long Point (Green unpublished data) and elsewhere (Clarke 1977), survivorship from egg to adult is only about 0.0007 (= 1 out of 1,430 eggs). These demographic characteristics lend themselves to expectations of inherently high stochasticity in population size and of local extinctions (Blaustein et al. 1995; Pechmann and Wilbur 1996). Depopulated habitat patches normally can be rescued mainly by dispersal of adults and juveniles from neighbouring sites – as is suggested by observations of long–distance dispersal events.
Because of the highly variable population sizes, trends in abundance are difficult to discern. The overall trend in abundance at the study site at the base of Long Point is downward. Over a 21–year span, peak abundance occurred in the early 1990s, and recent years have witnessed a steady diminution in numbers. This may be misleading as clearly the habitat conditions have changed over the same period and animals are probably breeding in other areas. It is probable that any storm of sufficient strength to create new, high productivity breeding sites will also have a transient, negative effect on overall abundance.
There is little or no chance of a rescue effect between Rondeau Provincial Park to the west, Long Point in the centre and Niagara to the east, or from U.S. populations on the south shore of Lake Erie. The distances between these populations are too great for the toads to traverse and there is no suitable intervening habitat (Yagi and Tervo 2006a,b). Information from mitochondrial DNA–sequencing shows that Long Point toads are genetically distinct from other populations, indicating that there has been no exchange of individuals between these populations for thousands of years. Furthermore, any toad that was swept into open water would tend to be transported by the general surface circulation patterns in Lake Erie from the north shore to the south shore. Thus the U.S. populations are more likely to benefit from a rescue effect from Canada than vice–versa (Smith and Green 2004).
A population viability analysis (PVA) was performed using the program VORTEX, version 9.4. The model was based on the current distribution, estimated abundance and known or estimated demographic characteristics of Fowler’s Toads in Canada.
For use with VORTEX, the model was spatially structured to represent the three Canadian Fowler’s Toad populations by postulating the existence of multiple subpopulations linked by varying degrees of dispersal: three subpopulations were used to represent the Rondeau Provincial Park population, six to represent Long Point and eight to represent Niagara. These 17 subpopulations were based on areas of known or probable critical habitat, characterized by the proximity of beaches, dunes and breeding sites.
VORTEX does not deal with populations and subpopulations in a hierarchical manner, therefore the existence of a higher level population structure was simulated in the model by setting varying levels of dispersal potential between the subpopulations. The dispersal potential values were calculated based on the power law approximation of the dispersal kernel derived by Smith and Green (2006) using mark/recapture methods. This procedure produced moderate to high estimated levels of dispersal within populations and low levels between them (Table 1). The minimum age at dispersal was set at 1 year of age, the maximum age at dispersal was set at 5 years of age and both females and males could disperse. Because VORTEXregards dispersal as the percentage of animals moving annually between subpopulations, survival rate during dispersal could be varied in the model by applying a percentage multiplier. In the basic model, survival rate was set at 50%, but different scenarios were also investigated at both higher and lower levels.
The model was further parameterized using the basic population data derived from studies at the western base of Long Point (Green 2000 and unpublished data), augmented by data from Rondeau Provincial Park (Dobbyn 2008) and the Niagara region (Yagi 2008). The larval stage of the life cycle, from egg through to metamorphosis, was ignored for the purposes of parameterizing the demographic variables of the model because inclusion of the thousands of eggs produced even by a single female yielded numbers that VORTEX was unable to process. Green and Summerfield (2008) estimated average clutch size, based on counts of eggs laid, to be 4618 ± 1317 eggs (n = 30). In the model, therefore, yearling juvenile toads represented the age 0 class, as though females gave birth to litters of juveniles. Mean “litter” size (yearlings per female) was calculated to be 10.5 ± 16 based on the total number of females present. However, this number was adjusted in the model in consideration of a breeding pool defined to be 90% of adult females present, as well as taking into account inter–individual variance in clutch size. Fifty percent of available females were set to breed in years of low density, while 100% of available females could breed in years at carrying capacity. In the age–structured model, survival of yearling juveniles (Year 0) to 2–year–old adults (Year 1) was parameterized to 17.2 ± 19.8 (S.D.)% of the starting cohort. Survival to older year classes was 4.2 ± 3.8% to Year 2, 1.6 ± 3.7% to Year 3, and 0.01 ± 0.02% to Year 4 (Figure 10). Age of first reproduction was modelled as Year 1 as toads commonly breed at 2 years of age (Kellner and Green 1995). The maximum breeding age was set at 5 years and the sex ratio at birth was set to 1:1. Mating was defined as polygynous, with 20% of adult males allowed in the breeding pool but 90% ± 45 (std. dev.) of adult females producing offspring.
Initial abundances of animals within the modelled subpopulations were set at 500 post–metamorphic individuals in all Niagara populations, one Long Point subpopulation and two of three Rondeau Provincial Park subpopulations. The remainder of the subpopulations were initially set to 1000 individuals. Carrying capacity (K) was set, optimistically, to 5000 ± 1500 (S.D.) individuals. Extinction was defined as the absence of animals of either sex. No inbreeding depression or density dependence was modelled as both of these effects are likely to be minor.
To account for random events of weather and climate, limited environmental stochasticity (e.g. major storms) was designed into the model. Such storms may extirpate local subpopulations, but at the same time they may create new dune habitat and lead to temporary subpopulation increases, if toads are able to reinvade these areas. Increasing development pressure makes it more difficult for toads to disperse successfully among subpopulations (See Limiting Factors and Threats). Additionally, one stochastically occurring storm of the century (“environmental catastrophe”, as termed by VORTEX), was included in the model to simulate the effects of particularly severe storms, though this low frequency is probably an underestimate of the actual frequency of devastating storms on Lake Erie (see Hazen 2000). Modelling additional “catastrophes” would increase the simulated rate of extirpations. Average probabilities of extirpation for each simulation were calculated from the results of 1000 iterations of the model, each run for the model equivalent of 100 years.
Using the basic model, the likely effects of particular threats or population characteristics were simulated by altering the appropriate parameters in different scenarios. Thus the effect of fragmentation was simulated by altering the estimated dispersal survival value, and thereby, the connectivity between subpopulations. Scenarios with dispersal survival values of 25%, 50%, 75% and 100% were run for the Long Point subpopulations. Similarly, the effect of habitat loss, for example the expansion of the invasive strain of Common Reed Grass (Phragmites a. australis) at Long Point, was modelled as a deterministic reduction in the estimated carrying capacities of the subpopulations. In the case of Phragmites, the extent of the reduction was based on the estimate by (Badzinski et al. 2008) of an average rate of expansion of 25.3% of area per year for Phragmitesat Long Point from 1999 to 2006 (Table 2).
Finally, the effect of isolation of the three populations was investigated by constructing two scenarios with differing estimates of dispersal structure. One scenario held that low yet positive percentages of individuals could disperse from any one population to any other whereas the other scenario held that individual toads were only able to successfully disperse among subpopulations within each population. This latter, more structured scenario is almost certainly the more accurate.
All scenarios under the model successfully mimicked the fluctuations in population size documented at Long Point, Niagara (Nickel Beach) and Rondeau Provincial Park (Figure 6). The model was virtually insensitive to alterations in age structure, whether 50% juveniles or 90% juveniles (Table 2), but like the older analysis performed by Green (2000), it did react to the number of populations employed, the postulated levels of immigration and emigration and the levels of variance in demographic parameters.
Altering survival during dispersal greatly affected population viability (Figure 11; Table 3). Using Long Point as the exemplar, under the most optimistic scenario of 100% survival during dispersal, the mean time to first extinction was 29.56 years ± 1.12 (S.E.), and the mean growth rate (r) was −0.1241 ± 0.0119 (S.E.). The scenario of 25% survival, however, yielded a mean time to first extinction of 22.14 years ± 0.81 (S.E.) and a mean r of −0.2647 ± 0.0154 (S.E.).
Figure 11. Mean extinction probabilities for the Long Point population of Fowler’s Toad populations simulated over 100 years based on 1000 iterations of the Population Viability model showing the relative effects of different levels of dispersal survival.

When the three Canadian populations of Fowler’s Toads were analyzed individually, the Rondeau Provincial Park population, comprised of only three subpopulations, fared the poorest and emerged as the most likely to be extirpated (r= −0.123 ± 0.686 S.D.), whereas Long Point (r = −0.251 ± 0.607 S.D.) and Niagara (r = −0.271 ± 0.590 S.D.) were very similar in outcome (Figure 12). The effect of habitat loss at Long Point, modelled as due to encroaching Phragmites, though, was dramatic as in this scenario r was reduced to −0.267 ± 0.709 (S.D.).
Figure 12. Mean extinction probabilities for each of the three Canadian Fowler’s Toad populations simulated over 50 years based on 1000 iterations of the Population Viability model. Additionally, the effect of habitat loss due to expansion of invasive Phragmites reeds is simulated for Long Point, only.

Figure 13. Mean extinction probabilities for Canadian Fowler’s Toads estimated over 50 years based on 1000 iterations of the Population Viability model under differing scenarios of dispersal between populations (basins) and habitat loss modelled as due to spread of Phragmites.

Considering all Canadian Fowler’s Toad populations collectively in the basic scenario of high population connectivity and no habitat loss, the mean time to extirpation was 23.28 years ± 0.26 (S.E.) and r was −0.2140 ± 0.0044 (S.E.). The probability of extirpation was 37.6 % ± 1.53 in 20 years (Table 3). Introducing habitat loss, such as due to the expansion of Phragmites, reduced the mean time to first extinction to 19.94 years ± 0.22 (S.E.) and mean r to −0.2085 ± 0.0052 (S.E.) while raising the probability of extirpations to 58% in 20 years. Including the high likelihood that the three populations are wholly isolated from each other, making rescue improbable, further reduced the mean time to extirpation to 19.10 years ± 0.23 (S.E.) and the mean r to −0.1793 ± 0.0051 (S.E.). The probability of extirpation for all populations after 20 years in this most pessimistic yet most plausible scenario was 69% (Table 3).
This PVA indicates a greater than 20% probability of extirpation of Fowler’s Toads from Canada within 20 years, and nearly 100% probability of extirpation within 100 years, under all scenarios of prevailing conditions, provided the parameters of the model are valid. Habitat loss, such as due to encroaching Common Reed (Phragmites), emerges as the greatest cause of increased extirpation probability, followed by habitat fragmentation that causes loss of inter–population connectivity (Table 3).
The simulated fluctuations in abundance of Fowler’s Toads so evident in all scenarios of the PVA model are largely due to high variances in the demographic parameters of the populations themselves, especially the survival and recruitment rates. Population fluctuations over several orders of magnitude are not uncommon for amphibian species (Green 2003). For example, in South Carolina, Pechmann et al. (1991) recorded fluctuations in female breeding population sizes and juvenile recruitment of over three orders of magnitude for three species of salamanders and one species of frog. Thus extirpations of subpopulations in the PVA model can largely be attributed to stochastic variation in population size, making the model sensitive to certain parameters in particular, such as starting population size, alterations in carrying capacity, the number of subpopulations, levels of immigration and emigration, mortality rates of both juveniles and adults and demographic stochasticity. Reduction in carrying capacity due to habitat loss, and accompanying reduction in juvenile recruitment, is particularly detrimental. In reality, these translate into threats that result in reduced dispersal potential, reduced survivorship, and reduced carrying capacity, all of which negatively affect the viability of Fowler’s Toad populations in Canada.
This PVA model, like all such estimations, has its limitations. Many parameters remain only educated approximations and many more years of population data would be required to accurately estimate the variances associated with them. It is not robust to changes in its variables; populations can be made to live forever with appropriately increased recruitment rates and lowered variances. But many more years of data may be a luxury not available to the ongoing study of Canadian Fowler’s Toads. Fowler’s Toad population sizes, like those of Natterjack Toads (Anaxyrus calamita) and many other amphibians (Berven and Grudzien 1990; Berven 1995; Beebee et al. 1996; Scribner et al. 1997) appear to be substantially recruitment–driven, subject to random extinction, and recoverable by immigration. As this analysis indicates, if the Canadian populations of Fowler’s Toads become increasingly isolated in diminished habitats where recruitment is reduced, the species will not survive.
Fowler’s Toad is extremely habitat–limited. As it is confined to only the shoreline of Lake Erie in areas where there is the right combination of natural beach, accessible dunes and oligotrophic, stillwater pools and marshes, the species is vulnerable to a variety of events or threats along the lake shore. The major threats, therefore, to the continued survival of Fowler’s Toad in Canada are related to habitat loss and habitat degradation, including outright loss of physical habitat, lost connectivity between areas of habitat, and declines in habitat quality.
Fowler’s Toad is adapted to and not threatened by the natural processes of shoreline change. Such changes are necessary for this and other species of amphibians that specialize in early succession habitats (Beebee et al.1996). Artificial interventions attempting to stabilize the shoreline by curtailment and control of the natural agents of change constitute serious threats to the species.
The development and stabilization of the shoreline are of major consequence to toad habitats. Breakwalls may eliminate hibernation sites outright or else obstruct access to them from the beach. Furthermore, backwash from waves hitting breakwalls increases removal of sand and disrupts the natural maintenance of beaches. This loss of beach is readily apparent in front of breakwalls all along the lakefront. Beach loss deprives the toads of foraging habitats and dispersal corridors as it turns the lakeshore into artificial bluffs where the toads do not survive.
Until recently, the seasonal cottages that line the shore of Lake Erie in many areas occupied by toads, including the base of Long Point, the east side of Rondeau Provincial Park and much of the Niagara shore, have not been appreciably detrimental to the toads. However, the small, seasonal cottages are yielding increasingly to high intensity development of larger homes intended for year–round occupation. This pattern of urbanizing the shoreline is particularly evident at Long Point and in the townships of Fort Erie, Port Colborne and Wainfleet in the Niagara Peninsula. Existing cottages have been re–developed or renovated into larger estates with greatly increased property values. A direct consequence of this development activity is the construction of shoreline protection in front of the dunes in aid of protecting valuable beach front properties from wind–blown sand, high water and storms off the lake. The intense storms of 1985 and high water of the late 1990s particularly precipitated this shoreline construction.
The construction of piers and groynes has occurred along the Lake Erie shoreline since early settlement. Some, such as the piers at Port Maitland and the Welland Shipping Canal at Port Colborne, were constructed to keep shipping lanes open. Other, smaller structures were constructed to protect shorelines from erosion or wave damage. The piers at Port Maitland have evidently affected the longshore drift process of sand movements in the lake; sand deposition on the west side and erosion of sand on the east side are both increasing. Because such structures lie perpendicular to the shoreline, they may also interfere with abilities of the toads to disperse, which may help to explain why toads have not re–colonized certain habitable areas.
Dunes are also stabilized by the colonization of natives and non–native plants naturally during times of low lake levels and few storm events. However, the rate of colonization is much greater when natural erosion process have been interfered with by the construction of roads, breakwalls, groynes and compaction for parking areas. When the dune becomes over–vegetated the population of toads declines. This has been observed in the last 3 years at Morgan’s Point Conservation Area and in the last 20 years at Rock Point Provincial Park where toads are no longer found (Yagi et al. 2006). At Nickel Beach in Port Colborne, parts of the dunes have been colonized by several exotic species including fescue (Festuca sp.), Scots Pine (Pinus sylvestris), Common Reed (Phragmites a. australis) and White Poplar (Populus alba) (Limnoterra 2006; A. Yagi, pers. comm.).
Beach cleaning activities, such as grading, grooming and clearing of algae are frequent along the shoreline, especially at public beaches. These result in the direct mortality of toads that are resting on or just under the sand. There are currently no regulations preventing these activities on beaches and dunes that are above the seasonal high water line. Beach cleaning with machines at Long Point Provincial Park was stopped by the former park supervisor, John Marchington, in 2007.
At Niagara, some beaches, including Gravelly Bay Beach, have a seasonally high motor vehicle use and/or are used as parking lots. Compacted sand beaches are inhospitable to toads and the vehicular traffic represents a source of direct mortality for toads especially when vehicles are driven on the beach at night. Although no specific surveys have been conducted to quantify the mortality rates, there have been observations of toads run over and killed on such beaches (Yagi 2008).
Located on the Erie lakefront just west of Morgan’s Point in Wainfleet, the Lakewood Easter Seals Camp for Children with Disabilities closed in 2005. It had been in operation since 1953 and is a known locality for Fowler’s Toads. In April 2007, a large amount of shoreline beach and dune sand was unlawfully removed with heavy equipment from a Fowler’s Toad habitat enhancement site on the property and deposited on private property to fill in a wetland (A. Yagi pers. comm.). In September, 2007, the Wainfleet Township council voted to amend a zoning bylaw to allow the new owner of the property to build 35 residential condominium units (Speck 2007). The full plan is to build 56 units in two phases (Bott 2007). The potential of this development to have a profoundly negative effect on the Fowler’s Toads that inhabit the site has been raised (Watts 2009) and considered by the Ontario Municipal Board under the Planning Act (Case No. PL070864, May 5, 2008). The Welland Township council has now absolved itself of responsibility for maintenance or operation of water and sewage systems for the housing development (Welland Tribune, May 2009).
With increasing development along the Lake Erie shoreline, sand dunes are often threatened. Existing, well–established sand dunes have been removed during the shoreline housing development projects at several sites on the Niagara Peninsula. Wind–blown sand will accumulate around any windbreak or object on the shore, including buildings. Such naturally forming, nascent dunes that occur on properties and access roads are in most cases removed as part of regular maintenance of roads and properties. At Long Point, accumulated sand is routinely bulldozed from lake–facing properties down to the water line. During evening toad beach surveys at Long Point, toads are noticeably more prevalent along the beach in front of the two sections of Long Point Provincial Park, where the beach is not ploughed, than along intervening sections of disturbed beach in front of lakeside cottages.
The extirpation of the toads from Point Pelee, a high–use national park that attracts considerable numbers of people, is a troubling precedent (Green et al. 2008). Beginning between 1894 and 1895 approximately 60% of the former wetland area between Point Pelee and Hillman Marsh was drained, which likely reduced the amount of available breeding habitat. In 1953, the Marentette Drainage Scheme drained the last section of marsh immediately north of the Park. Cottage and housing development along the shoreline west of the Park began in the mid–1930s and east of the Park in 1953. This likely also reduced the amount of habitat available for Fowler’s Toads as it increased human use of the area.
The invasive strain of Common Reed, Phragmites a. australis, appears to pose a growing and considerable threat to breeding areas of Fowler’s Toad at Rondeau Provincial Park and Long Point (Table 1), as well as many wetland–dependant species (Badzinski et al. 2008; S. Dobbyn pers. comm. 2008). On Long Point, some historic breeding sites such as the Crown Marsh are now overgrown with Phragmites and no longer used (D. Green, unpublished data). Phragmites is emerging as an aggressive invasive species throughout the lower Great Lakes (Wilcox et al. 2003; T’ulbure et al. 2007).
In Niagara during times of low water levels, Fowler’s Toad breeding sites consist of flooded rocky pools near headlands and drain outlets near beaches. Recently observed large accumulations of Zebra Mussel (Dreissena polymorpha) shells in rocky pool areas have eliminated breeding habitat (A. Yagi pers. comm.). At Rock Point Provincial Park, as part of a habitat enhancement project, several truckloads of Zebra Mussel shells were removed in an effort to improve habitat (A. Yagi pers. comm.). The abundance of Zebra Mussels may be producing a threat of botulism poisoning among Great Lakes fish and birds (Riley et al. 2008), and possibly also Fowler’s Toads. The botulism toxin is produced by the bacterium, Clostridium botulinum, which proliferates in anaerobic conditions (Reed and Rocke 1992). Zebra Mussels may enhance the growth of nearshore benthic algae (Hecky et al. 2004) and when both dead Zebra Mussels and decomposing algal mats are cast up on shore, their decay creates the anoxic conditions required by C. botulinum (Riley et al. 2008). Invertebrates that consume the bacteria may pass the toxin on to their predators (Jensen and Allen 1960).
Most Fowler’s Toad habitat is too sandy and too close to Lake Erie to be suitable for farming. However, the use of chemicals in agriculture may have contributed to the declines in toad populations in southwestern Ontario. Point Pelee and Pelee Island, especially, have been subject to intensive agricultural activity since World War II. Sanders (1970) found tadpoles of A. fowleri to be highly susceptible to the herbicide Trifluralin and the insecticide Endrin in particular. Tadpoles became more susceptible as they grew older. Adults are equally susceptible to poisoning by these chemicals (Ferguson and Gilbert 1968). Campbell (1975) reported significant, sublethal levels of toxic chemicals accumulated in specimens of A. fowleri from Ontario. The last observation of Fowler’s Toad at Point Pelee National Park in 1949 closely coincides with the beginning of DDT use for mosquito control in the Park (Russell et al. 1999).
There is considerable potential for metal contamination at Nickel Beach and vicinity. Vale INCO owns the Nickel Beach property and the Port Colborne Refinery, which produces platinum–group metals and packages nickel for distribution. The refinery was commissioned in 1918. A 2001 class action suit launched against INCO by the residents of Port Colborne cites the area’s soil contamination with as much as 14,000 parts per million of nickel, including carcinogenic nickel oxide, a level 70 times that considered safe for residential areas in Ontario (Globe and Mail, March 27, 2001). Modified in 2005 (Ontario Court of Appeal Docket C42414), the suit has yet to go to trial. Nevertheless, the population effects on Fowler’s Toads of contaminating heavy metals in the environment are effectively unknown. Laboratory tests of metal toxicity (Birge et al. 2000; Linder and Grillitsch 2000) showed that Fowler’s Toads have higher tolerances for some metals, such as Cesium (LC50=1076 mg/L) and Magnesium (LC50 = 807 mg/L), than others, including Zinc (LC50= 87 mg/L ), Copper (LC50 = 25 mg/L), Chromium (LC50= 0.11 mg/L), Gallium (LC50= 0.13 mg/L), Titanium (LC50= 0.24 mg/L), or Aluminum (LC50= 0.28 mg/L).
As noted here and elsewhere in this report, (see sections on: Genetic description; Designatable units; Population viability analysis; Limiting factors and Threats), Fowler’s Toads occur in three populations which are fairly distinct and genetically isolated from each other. Furthermore, subpopulations, especially in the Niagara area, are increasingly isolated from one another by anthropogenic pressures and habitat destruction and degradation. For example, Figures 7, 8 show uninhabitable areas of shoreline and these are growing. Thus local extinctions, from storms or from loss of habitat to invasive plants, become less likely to be “rescued” by movements among subpopulations. Recent meta–analyses (Reed et al. 2003, Traill et al. 2007) and research cited within these reports suggest that for long–term viability, populations should have 4000 to 5000 individuals. As pointed out in the section on Abundance, the total effective population size for this species in Canada is likely only about 5000 individuals. Therefore, the viability of the three isolated populations, or even more so of the isolated smaller subpopulations, is probably not sustainable. This conclusion is amply borne out by the PVA. Thus, the species qualifies as severely fragmented.
Fowler’s Toads play the role of small insectivores, specializing in ants and beetles (Judd 1957; Bush and Melnick 1962). Tadpoles are significant detritivores in small ponds, rocky pools and embayments. No other anurans in the Great Lakes region habitually and primarily forage along lakeshores and dunes. In turn, higher–level carnivores including snakes, birds, fish, mammals and other frogs consume them.
The majority of Fowler’s Toad habitat in the Rondeau Provincial Park and Long Point areas is located within a Provincial Park or National Wildlife Area where there is no direct threat from industrial or urban development. However, human activities within and surrounding the parks affect the quality and quantity of available habitat within these protected areas. In the Niagara Area, the vast majority of Fowler’s Toad habitat is on private land. The only Crown Land that supports a population of Fowler’s Toads is James N. Allan Provincial Park. They were historically found at Rock Point Provincial Park. Fowler’s Toads have been found in Long Beach and Morgan’s Point Conservation Areas.
Fowler’s Toad was assessed by COSEWIC as “Rare” in 1986 and then “Vulnerable” and “Special Concern” as COSEWIC changed its terminology. It was assessed as “Threatened” in 1999 (Green, 1999b) and that assessment was confirmed in 2000 (Green 2000). In 2003, the species was legally listed under Schedule 1 of the Species at Risk Act (SARA). Because SARA applies only to federal Crown Lands, it affords protection to the toads only in Big Creek and Long Point National Wildlife Areas and the Ministry of Transport controlled tip of Long Point. The federal Fisheries Actand Environmental Assessment Act (Table 4) is also indirectly applicable to Fowler’s Toad habitats.
The Ontario Endangered Species Act 2007 lists Fowler’s Toad as a Threatened species, thus invoking prohibitions on killing, harming, harassing, transporting, selling etc. or damage or destruction to its habitat. A recovery strategy is required under the Act. Permits are available. The Ontario Fish and Wildlife Conservation Act considers Fowler’s Toad to be a “specially protected amphibian” and thus prohibits hunting, trapping or keeping in captivity. Licences are available. Other provincial laws (Table 5) are also indirectly applicable to Fowler’s Toad habitats.
The species is not listed as a species of concern in the United States or in any of the states adjacent to Ontario. It has a NatureServe status of G5, or Secure.
Anaxyrus fowleri
Fowler’s Toad – crapaud de Fowler
Demographic Information
population. The toads mature at 2 years of age. Of an
average clutch of ~ 4500 eggs, about 10 survive to be
yearlings. Survival of these yearlings to 2–year–old adults
was estimated at 17 % of the starting cohort. Survival to
older year classes was 4 % to age 3, 1% to age 4, and
0.01 % to age 5 years. (See section on Population Viability
Analysis and Figure 10).
decline in number of mature individuals?
mature individuals within [5 years or 2 generations]
[reduction or increase] in total number of mature
individuals over the last [10 years, or 3 generations].
in total number of mature individuals over the next
[10 years, or 3 generations].
[reduction or increase] in total number of mature individuals
over any [10 years, or 3 generations] period, over a time
period including both the past and the future.
Extent and Occupancy Information
Based on minimum convex polygon of all extant localities (= 4,690 km²) minus uninhabitable reaches of coastline lake surface.
Based on the length of shoreline occupied by extant populations the AO is ~ 65 km². This is appropriate as the species is restricted to the linear habitat of the lakeshore and is nowhere found as much as 500 m inland. An IAO using a 1x1 km² grid
gives an area of 120 km², and a 2x2 km² grid adds up to an IAO of 480 km².
IAO (1x1 km²) = 120 km²
IAO (2x2 km²) = 480 km²
The three populations are isolated. Within the Niagara
population, development of the shoreline has further isolated
subpopulations from each other. (See section at end of
Limiting Factors and Threats)
be considered three locations.
in extent of occurrence? Subsequent to the loss of populations
at Pelee Island, Point Pelee and Turkey Point, population losses
over the past 30 years have been within the EO as calculated.
Yes, due to losses of breeding sites at Rock Point and Port Maitland and encroachment by invasive Common Reed at Long Point and Rondeau Provincial Park.
in number of populations?
in number of locations?
in [area, extent and/or quality] of habitat?
Habitat decline mainly due to development and disturbance of shoreline at Niagara and invasive Common Reed at Long Point
and Rondeau Provincial Park.
* See definition of location.
Number of Mature Individuals (in each population)
estimates of adults, 2004–2008)
Quantitative Analysis
[20% within 20 years or 5 generations, or 10%
within 100 years]. Population viability analysis. The most realistic scenario is based on a model allowing for limited environmental stochasticity, no connectivity between populations, structured age–specific estimates of abundance and loss of habitat from invasive Phragmites reeds. A best case scenario yields a 22% probability of extirpation in 20 years.
Threats (actual or imminent, to populations or habitats)
- Loss and deterioration of habitats due to shoreline development and artificial coastline stabilization, including construction of breakwalls, roads, parking lots, piers and groynes.
- Intensive mechanical beach maintenance, such as grading, grooming and clearing of algae using beach grooming machines or bulldozers.
- Intensive human recreational use of beaches and dunes, including vehicular traffic.
- Loss and degradation of habitats due to invasive species, particularly Zebra Mussels and Common Reed.
- Agricultural and industrial contaminants.
Rescue Effect (immigration from outside Canada)
U.S. populations are separated from Canadian populations by Lake Erie.
Current Status
Status and Reasons for Designation
Reasons for designation:
This species only occurs on sandy beaches in three disjunct areas along the north shore of Lake Erie. It has disappeared from numerous historic sites on the Lake Erie shore and continues to decline in abundance and number of populations with further habitat loss and degradation due to invasive species (Common Reed, Zebra Mussels) and anthropogenic activities including shoreline development, beach cleaning, construction of breakwalls, bulldozing of beaches, vehicle use on beaches and agricultural and industrial contaminants. In addition, a population viability analysis (PVA) model suggests that over the last decade, the probability of extirpation within 20 years has increased substantially.
Applicability of Criteria
Criterion A (Decline in Total Number of Mature Individuals): Does not meet criterion A as the size of decline can not be determined.
Criterion B (Small Distribution Range and Decline or Fluctuation): Meets Endangered B1ab(ii,iii,v)c(iv)+2ab(ii,iii,v)c(iv) as its distribution range is severely fragmented, EO (1,200 km²) and IAO (480 km²) are below thresholds, and area of occupancy, habitat area and quality, and the number of mature individuals are declining. Meets c(iv) because population fluctuates more than an order of magnitude.
Criterion C (Small and Declining Number of Mature Individuals): Not applicable. Although the minimum number of mature individuals from Rondeau, Long Point and Niagara is below the Endangered threshold (2347), the mean number (4740) is above the Endangered threshold, the decline rates are not calculable, some populations exceed 250 mature individuals, and no population contains more than 95% of all mature individuals.
Criterion D (Very Small or Restricted Total Population): Does not meet criterion D as the total number of mature individuals exceeds threshold.
Criterion E (Quantitative Analysis): Meets Endangered under criterion E as a quantitative analysis based on a PVA model allowing for limited environmental stochasticity, no connectivity between populations, structured age–specific estimates of abundance and loss of habitat from invasive Phragmites reeds estimates a 69% probability of extirpation in 20 years. This is based on the most realistic scenario. A best case scenario yields a 22% probability of extirpation in 20 years.
Anne R. Yagi (Ontario Ministry of Natural Resources) and Sandy Dobbyn (Ontario Parks) were especially valuable sources of information and commentary for the report. The following additional members of the Fowler’s Toad recovery team were also greatly helpful (alphabetical order): James Duncan (Nature Conservancy Canada), Kim Frohlich (Niagara Peninsula Conservation Authority), Ron Gould (Ontario Ministry of Natural Resources), Kate Hayes (Environment Canada), Deb Jacobs (Ontario Ministry of Natural Resources), Bob Johnson (Toronto Zoo), Kate Macintyre (Ontario Ministry of Natural Resources), John Marchington (Ontario Parks), Vicki McKay (Parks Canada Agency), Devin Mills (Ontario Ministry of Natural Resources), Michael Oldham (Ontario Ministry of Natural Resources), Sherry Pineo (Ontario Parks), Jeff Robinson (Canadian Wildlife Service), Tim Seburn–Bert Miller Nature Club, Fort Erie, Rob Tervo (Ontario Ministry of Natural Resources), Wayne Weller (Ontario Power Group), Allen Woodliffe (Ontario Ministry of Natural Resources). Sara Lourie (McGill University) produced the Canadian range map (Figure 4).
Badzinski, S.S., S. Proracki, S.A. Petrie and D. Richards. 2007. Changes in the distribution & abundance of common reed (Phragmites australis) between 1999 & 2006 in marsh complexes at Long Point – Lake Erie. Unpublished report to Ontario Ministry of Natural Resources.
Badzinski, S.S., S. Proracki, S.A. Petrie, and D. Richards. 2008. Changes in the distribution & abundance of common reed (Phragmites australis) between 1999 & 2006 in marsh complexes at Long Point – Lake Erie. Ontario Ministry of Natural Resources, Peterborough.
Beebee, T.J.C., J.S. Denton, and J. Buckley. 1996. Factors affecting population densities of adult natterjack toads, Bufo calamita, in Britain. Journal of Applied Ecology 33:263–268.
Berven, K.A. 1990. Factors affecting population fluctuations in larval and adult stages of the wood frog, (Rana sylvatica). Ecology 71:1599–1608
Berven, K.A. 1995. Population regulation in the wood frog, Rana sylvatica, from three diverse localities. Australian Journal of Ecology 20:385–392.
Berven, K.A., and T.A. Grudzien. 1990. Dispersal in the wood frog (Rana sylvatica): implications for population structure. Evolution 44:2047–2056.
Birge, W.J., A.G. Westerman, and J.A. Spromberg. 2000. Comparative toxicology and risk assessment of amphibians. pp. 727–791. In D.W. Sparling, G. Linder and C.A. Bishop (eds.) Ecotoxicology of amphibians and reptiles. SETAC Press, Pensacola, Florida.
Blaustein, A.R., L.K. Belden, D.H. Olson, D.M. Green, T.L. Root, and J.M. Kiesecker. 2001. Amphibian breeding and climate change. Conservation Biology. 15:1804–1809.
Bossert, M., M. Draud and T. Draud. 2003. Bufo fowleri (Fowler’s Toad) and Malaclemys terrapin terrapin (Northern Diamondback Terrapin). Refugia and nesting. Herpetological Review 34(2):135.
Bott, L. 2007. Wainfleeters must fight urbanization. Welland Tribune. October, 2007
Breden, F. 1987. The effects of post–metamorphic dispersal on the population genetic structure of Fowler’s Toad Bufo fowleri. Copeia 1987:386–395.
Breden, F. 1988. Natural history and ecology of Fowler’s Toad, Bufo woodhousei fowleri (Amphibia: Bufonidae), in the Indiana Dunes National Lakeshore. Fieldiana: Zoology 49:1–16.
Brown, L.E., and M.J. Littlejohn. 1972. Male release call in the Bufo americanus group. Pages 310–323 in W. F. Blair, editor. Evolution in the Genus Bufo. University of Texas Press, Austin, Texas.
Brown, R.L. 1974. Diets and habitat preferences of selected anurans in southeast Arkansas. American Midland Naturalist 91:468–473.
Bush, F.M., and E.F. Melnick. 1962. The food of Bufo woodhousei fowleri. Herpetologica 18:110–114.
Campbell, C.A. 1975. Reproduction and ecology of turtles and other reptiles and amphibians of lakes Erie and St. Clair in relation to toxic chemicals. Unpublished report to Canadian Wildlife Service.
Clarke, R.D. 1974a. Postmetamorphic growth rates in a natural population of Fowler’s Toad, Bufo woodhousei fowleri. Canadian Journal of Zoology 52:1489–1497.
Clarke, R.D. 1974b. Activity and movement patterns in a population of Fowler’s Toad, Bufo fowleri. American Midland Naturalist 92:257–74
Clarke, R.D. 1977. Postmetamorphic survivorship of Fowler’s Toad, Bufo woodhousei fowleri. Copeia 1977:594–597.
Collins, J.P., and H.M. Wilbur. 1979. Breeding habits and habitats of the amphibians of the Edwin S. George Reserve Michigan and notes on the local distribution of fishes. Occasional Paper of the Museum of Zoology, University of Michigan 686:1–34.
Conant, R., and J. T.Collins. 1998). A Field Guide to Reptiles and Amphibians of Eastern and Central North America. 3rd Edition. Houghton Mifflin Company, Boston.
Crother, B.I. (ed.). 2008. Scientific and standard English names of amphibians and reptiles of North America north of Mexico. SSAR Herpetological Circular 37:1–84.
Dexter, R.W. 1973. Nomenclatural history and status of Fowler’s toad (Bufo fowleri). HISS News Journal 1:155–157.
Dobbyn, S. 2005. A Preliminary Assessment of the Fowler’s Toad Population in Rondeau Provincial Park. 2004 Field Season Report. Ontario Parks, Morpeth, Ontario. 20 pp.
Dobbyn, S. 2008. Rondeau Provincial ParkFowlers Toad Mark/Recapture Study 2004–2008. Draft report, Ontario Parks, Morpeth, Ontario.
Dundee, H.A., and D.A. Rossman. 1989. The Amphibians and Reptiles of Louisiana. Louisiana State University Press, Baton Rouge, Louisiana.
Ferguson, D.E., and C.C. Gilbert. 1968. Tolerances of three species of anuran amphibians to five chlorinated hydrocarbon insecticides. Journal of the Mississippi Academy of Sciences 13:135–138.
Frost, D.R., T. Grant, J. Faivovich, R. Bain, A. Haas, C.F.B. Haddad, R. de Sá, A. Channing, M. Wilkinson, S.C. Donnellan, C. Raxworthy, J.A. Campbell, B.L. Blotto, P. Moler, R.C. Drewes, R.A. Nussbaum, J.D. Lynch, D.M. Green, and W. Wheeler. 2007. The amphibian tree of life. Bulletin of the American Museum of Natural History 297:1–370.
Gerhardt, H.C. 1975. Sound pressure levels and radiation patterns of the vocalizations of some North American frogs and toads. Journal of Comparative Physiology A 102:1–12.
Given, M.F. 1996. Intensity modulation of advertisement calls in Bufo woodhousii fowleri. Copeia 1996:970–977.
Given, M.F. 2002. Interrelationships among calling effort, growth rate, and chorus tenure in Bufo fowleri. Copeia. 2002:979–987.
Green, D.M. 1982. Mating call characteristics of hybrid toads (Bufo americanus x B. fowleri) at Long Point, Ontario. Canadian Journal of Zoology 60:3293–3297.
Green, D.M. 1981. Theoretical analysis of hybrid zones derived from an examination of two dissimilar zones of hybridization in toads (genus Bufo). Ph.D. Thesis, University of Guelph.
Green, D.M. 1984. Sympatric hybridization and allozyme variation in the toads Bufo americanus and B. fowleri in southern Ontario. Copeia 1984:18–26.
Green, D.M. 1989. Fowler’s Toads, (Bufo woodhousii fowleri) in Canada: Biology and population status. Canadian Field–Naturalist 103:486–496.
Green, D.M. 1997a. Temporal variation in abundance and age structure in Fowler’s Toads, Bufo fowleri, at Long Point, Ontario. Pages 45–56 in D. M. Green, editor. Amphibians in decline: Canadian studies of a global problem. Society for the Study of Amphibians and Reptiles, St. Louis.
Green, D.M. 1997b. Population Ecology of Fowler’s Toads, Bufo fowleri, at Long Point, Ontario. World Wildlife Fund, Endangered Species Recovery Fund.
Green, D.M. 1999a. Status report update on Fowler’s Toad; Bufo fowleri, in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa Ontario.
Green, D.M. 1999b. Population ecology of Fowler’s Toads, Bufo fowleri, at Long Point, Ontario, in 1999. World Wildlife Fund Canada Endangered Species Recovery Fund.
Green, D.M. 2000. Status report update on Fowler’s Toad; Bufo fowleri, in Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa Ontario.
Green, D.M. 2003. The ecology of extinction: population fluctuation and decline in amphibians. Biological Conservation 111:331–343.
Green, D.M. 2005. Bufo fowleri, Fowler’s toad. pp. 766–778. In Amphibian Declines: the conservation status of United States species. M.J. Lannoo (ed.). University of California Press, Berkeley.
Green, D.M. 2008. Movements and habitat use by Fowler’s Toads, Bufo (Anaxyrus) fowleri, at Hahn Beach, Big Creek NWA, Long Point, Ontario. Report to Canadian Wildlife Service, Environment Canada. 30 pp.
Green, D.M., and L. Bol. 1999. Population ecology of Fowler’s Toads, Bufo fowleri, at Long Point, Ontario, in 1998.Ontario Ministry of Natural Resources, Canadian Wildlife Service, and World Wildlife Fund Canada Endangered Species Recovery Fund.
Green, D.M., and D. Cantin. 1990. Spring emergence and breeding behaviour of Fowler’s Toads, Bufo woodhousii fowleri at Long Point, Ontario, in 1989. Report to Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., and H.S. Chan Tang. 1994. Breeding activity and growth rate in Fowler’s Toads (Bufo woodhousii fowleri) at Long Point, Ontario in 1993. Ontario Ministry of Natural Resources, Canadian Wildlife Service, and World Wildlife Fund Canada.
Green, D.M., and J. Hensley. 1998. Age structure, and abundance of Fowler’s Toads, Bufo fowleri, at Long Point, Ontario, in 1997.Ontario Ministry of Natural Resources, Canadian Wildlife Service, and World Wildlife Fund Canada Endangered Species Recovery Fund.
Green, D.M., and E.J.B. McTavish. 2005. Population ecology of Fowler’s Toad (Bufo fowleri) at Long Point, Ontario. Field Report for 2004. Ontario Ministry of Natural Resources.
Green, D.M., and C. Parent. 2003. Variable and asymmetric introgression in a hybrid zone in the toads, Bufo americanus, and Bufo fowleri. Copeia 2003:34–43.
Green, N.B., and T.K. Pauley. 1987. Amphibians and Reptiles in West Virginia. University of Pittsburgh Press, Pittsburgh, Pennsylvania.
Green, D.M., and Z.S. Porebski. 1991. Breeding activity of Fowler’s Toads, Bufo woodhousii fowleri at Long Point, Ontario, in 1990. Report to Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., and J. Rabinowitz. 1996. Spring emergence, age structure, and abundance of Fowler’s Toads, Bufo fowleri, at Long Point, Ontario, in 1995. Ontario Ministry of Natural Resources, Canadian Wildlife Service, and World Wildlife Fund Canada.
Green, D.M. and N.K. Sanderson. 2006. Population Ecology of Fowler’s Toad (Bufo fowleri) at Long Point, Ontario. Field Report for 2005. Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., and N.K. Sanderson. 2007. Population ecology of Fowler’s Toad (Bufo fowleri) at Long Point, Ontario. Field Report for 2006. Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., and N.K. Sanderson. 2007. Population ecology of Fowler’s Toad (Bufo fowleri) at Long Point, Ontario. Field Report for 2007. Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., and N.K. Sanderson. 2007. Radio–tracking Fowler’s Toads in Long Point, Ontario. Field Report for 2007, Canadian Wildlife Service, Ontario Region,
Green, D.M., and M.A. Smith. 2000 Ecology and viability of Fowler’s Toads (Bufo fowleri) Populations. World Wildlife Fund Canada Endangered Species Recovery Fund.
Green, D.M., and M.A. Smith. 2003. Spatial ecology of Fowler’s toad (Bufo fowleri) populations in Canada. World Wildlife Fund Canada Endangered Species Recovery Fund.
Green, D.M., and A. Summerfield. 2008. Population ecology of Fowler’s Toad (Bufo fowleri) at Long Point, Ontario. Field Report for 2008. Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., C.W. Zeyl and A. El–Yassir. 1991. Spring emergence and abundance of Fowler’s Toads, Bufowoodhousii fowleri at Long Point, Ontario, in 1991: implications for conservation. Report to Ontario Ministry of Natural Resources, Canadian Wildlife Service, and World Wildlife Fund Canada.
Green, D.M., A. Valachovic, G. Tambasco, and A. Kelner. 1993. Spring emergence, age structure, and abundance of Fowler’s Toads, Bufo woodhousii fowleri, at Long Point, Ontario, in 1992. Report to Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Green, D.M., M. Ouellet, and C. Saumure. 1995. Breeding activity and growth rate in Fowler’s Toads (Bufo woodhousii fowleri) at Long Point, Ontario in 1994. Ontario Ministry of Natural Resources, Canadian Wildlife Service, and World Wildlife Fund Canada.
Green, D.M., A.R. Yagi, M.A. Smith, S. Dobbyn, J. Duncan, K. Frohlich, R. Gould, K. Hayes, D. Jacobs, B. Johnson, K. Macintyre, J. Marchington, V. McKay, D. Mills, M.Oldham, S. Pineo, J. Robinson, T. Seburn, R. Tervo, W. Weller, and A. Woodliffe. 2008. Recovery Strategy for the Fowler’s Toad (Bufo[Anaxyrus] fowleri) in Ontario. Species at Risk Act Recovery Strategy Series. Ontario Ministry of Natural Resources, Peterborough. 68 pp.(draft)
Hazen, S. (ed.) 2000. Down by the Bay: a history of Long Point and Port Rowan 1799–1999. Boston Mills Press, Erin, Ont.
Hecky, R.E., R.E.H. Smith, D.R. Barton, S.J. Guilford, and W.D. Taylor. 2004. The nearshore phosphorus shunt: a consequence of ecosystem engineering by dreissenids in the Laurentian Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 61:1285–1293.
Hillman, S.S., P.C. Withers, R.C. Drewes, and S.D. Hillyard 2009. Ecological and Environmental Physiology of Amphibians. Oxford University Press, Oxford.
Hinckley, M.H. 1882. On some differences in the mouth–structure of tadpoles of the anurous batrachians found in Milton, Massachusetts. Proceedings of the Boston Society of Natural History 21:307–314.
Hubbs, C.L. 1918. Bufo fowleri in Michigan, Indiana and Illinois. Copeia 55:40–43.
Jackson, J.N. 1967. Recreational development and the Lake Erie shore. Niagara Regional Development Council, St. Catharines, Ontario.
Jensen, W.I., and J.P. Allen. 1960. A possible relationship between aquatic invertebrates and avian botulism. Transactions of the North American Wildlife Natural Resources Conference 25:171–180.
Judd, W.W. 1957. Fowler’s Toads on the Lake Erie shore. Bulletin of the Federation of Ontario Naturalists 78:13–15.
Kellner, A., and D.M. Green. 1995. Age structure and age at maturity in Fowler’s Toads, Bufo woodhousii fowerli, at their northern range limit. Journal of Herpetology 29:485–489.
Klemens, M.W. 1993. Amphibians and reptiles of Connecticut and adjacent regions. Bulletin of the State Geological and Natural History Survey of Connecticut 112:1–318.
Laurin, G., and D.M. Green 1989. Spring emergence and male breeding behaviour of Fowler’s Toads, Bufowoodhousii fowleri, at Long Point, Ontario. Report to Ontario Ministry of Natural Resources and Canadian Wildlife Service.
Laurin, G., and D.M. Green. 1990. Spring emergence and male chorus behaviour in Fowler’s Toads, Bufo woodhousii fowleri, at Long Point, Ontario. Canadian Field–Naturalist 104:429–434.
Lazell, J.D., Jr. 1976. This broken archipelago. Cape Cod and the islands, amphibians and reptiles. Demeter Press, New York.
Limnoterra Ltd. 2006. Nickel Beach, Port Colborne, Fowler’s Toad (Bufo fowleri) Data Analysis and Management Implications. (Draft) Prepared for Fowler’s Toad Recovery Team. 69 pp.
Linder, G., and B. Grillitsch. 2000. Ecotoxicology of metals. pp.325–459. InD.W. Sparling, G. Linder and C.A. Bishop (eds.) Ecotoxicology of amphibians and reptiles. SETAC Press, Pensacola, Florida.
Logier, E.B.S. 1931. The amphibians and reptiles of Long Point. Transactions of the Royal Canadian Institute 18:229–236.
Meacham, W.R. 1962. Factors affecting secondary intergradation between two allopatric populations in the Bufo woodhouseicomplex. American Midland Naturalist 67:282–304.
Meffe, G.K., and Carroll, C.R. 1994. Principles of Conservation Biology. Sinauer Associates, Inc. Sunderland, Massachusetts. 600 pp.
Meyer, S.M. 2003. Comparative use of Phragmites australis and other habitats by birds, amphibians, and small mammals at Long Point, Ontario. M.Sc. thesis, Faculty of Graduate Studies University of Western Ontario London, Ontario, Canada. September 2003.
Miller, W.deW., and J. Chapin. 1910. The toads of the northeastern United States. Science 32:315–317.
Minton, S.A. 1972. Amphibians and Reptiles of Indiana. Monograph Number 3, Indiana Academy of Science, Indianapolis, Indiana.
Mount, R.M. 1975. The Reptiles and Amphibians of Alabama. Auburn University Press, Auburn, Alabama
Myers, G.S. 1931. The original descriptions of Bufo fowleri and Bufo americanus. Copeia 1931:94–96.
Pauly, G.B., D.M. Hillis, and D.C. Cannatella. 2004. The history of a Nearctic colonization: Molecular phylogenetics and biogeography of the Nearctic toads (Bufo). Evolution 58(11): 2517–2535.
Pechmann, J.H.K., D.E. Scott, R.D. Semlitsch, J.P. Caldwell, L.J. Vitt, and J.W. Gibbons. 1991. Declining amphibian populations: the problem of separating human impacts from natural fluctuations. Science 253:892–895.
Pechmann, J.H.K., Wilbur, H.M., 1996. Putting declining amphibian populations in perspective, natural fluctuations and human impacts. Herpetologica 50, 65–84
Planck, R.J. and C. Blott. 2006. DRAFT Nickel Beach Port Colborne, Fowler’s Toad (Bufo fowleri) Data Analysis and Management Implications prepared for the National Fowler’s Toad Recovery Team.
Reed, T.M., and T.E. Rocke. 1992. The role of avian carcasses in botulism epizootics. Wildlife Society Bulletin 20:175–182.
Riley, S.C., K.R. Munkittrick, , A.N. Evans, and C.C. Krueger. 2008. Understanding the ecology of disease in Great Lakes fish populations. Aquatic Ecosystem Health and Management 11:321–334.
Russell, R.W., S.J. Hecnar, G. Mouland, and G.D. Haffner. 1999. Pesticide accumulation in Point Pelee amphibians. Proceedings of the Parks Research Forum of Ontario, 1999 Annual Meeting. Guelph, ON. April, 1999.
Sanders, H.D. 1970. Pesticide toxicities to tadpoles of the western chorus frog, Pseudacris triseriata, and Fowler’s Toad, Bufo woodhousei fowleri. Copeia 1970:246–251.
Scribner, K.T., J.W. Arntzen and T. Burke. 1997. Effective number of breeding adults in Bufo bufo estimated from age–specific variation at mini–satellite loci. Molecular Ecology 6:701–712.
Shirose, L.J., C.A. Bishop, D.M. Green, C.J. Macdonald, R.J. Brooks, and N.J. Helferty. 1997. Validation tests of an amphibian call count survey technique in Ontario, Canada. Herpetologica 53:312–320.
Smith, M.A. 2004. Spatial ecology of Bufo fowleri. Ph.D. Thesis. Department of Biology, McGill University, Montreal.
Smith, M.A. and D.M. Green. 2002. Bufo fowleri (Fowler’s Toad). Predation. Herpetological Review 33(2):125.
Smith, M.A. and D.M. Green. 2004. Phylogeography of Bufo fowleri at its northern range limit. Molecular Ecology 13:3723–3733.
Smith, M.A., and D.M. Green. 2005a. Bufo fowleri (Fowler’s Toad) Predation. Herpetological Reveiw 36(2):159–160.
Smith, M.A, and D.M. Green. 2005b. Are all amphibian populations metapopulations? Dispersal and the metapopulation paradigm in amphibian ecology. Ecography 28:110–128.
Smith, M.A, and D.M. Green. 2006. Sex, isolation and fidelity: unbiased long distance dispersal in a terrestrial amphibian. Ecography 29:649–658.
Smith, P.W. 1961. The Amphibians and Reptiles of Illinois. Illinois Natural History Survey Bulletin 28:1–298.
Speck, M. 2007. Development at Lakewood Beach camp approved. Niagara This Week. September 14, 2007.
Stone, D. 1988. Long Point: Last Port of Call. Boston Mills Press, Erin, Ontario.
Sullivan, B.K. 1982. Significance of site, temperature and call attributes to sexual selection in Bufo woodhousei australis. Journal of Herpetology 16:103–106.
Sullivan, B.K. 1983. Sexual selection in Woodhouse’s toad (Bufo woodhousei). II. Female choice. Animal Behaviour 31:1011–1017.
Sullivan, B.K., K.B. Malmos and M.F. Given. 1996. Systematics of the Bufo woodhousii Complex (Anura: Bufonidae): advertisement call variation. Copeia 1996:274–280.
T’ulbure, M.G., C.A. Johnston and D.L. Auger. 2007. Rapid invasion of a Great Lakes coastal wetland by non–native Phragmites australis and Typha. Journal of Great Lakes Research 33:269–279.
Tupper, T.A., and R.P. Cook. 2008. Habitat variables influencing breeding effort in northern clade Bufo fowleri: Implications for conservation. Applied Herpetology 5: 101–119.
Vences, M., M.W. Penuel–Matthews, D.R. Vietites and R. Altig. 2003. Rana temporaria (Common Frog) and Bufo fowleri (Fowler’s Toad) tadpoles. Protozoan infestation. Herpetological Review 34(3):237–238.
Watts A. 2009. Fowler conflict. Welland Tribune, April, 2009.
Welsman, P. 2009. Cottager Asks Important Questions… Letter to the Editor. The Chatham Daily News, Saturday February 28th.
Wilcox, K.L., S. A. Petrie, L. A. Maynard, and S. W. Meyer. 2003. Historical distribution and abundance of Phragmites australis at Long Point, Lake Erie, Ontario. Journal of Great Lakes Research 29:664–680.
Wright, A.H., and A.A. Wright. 1949. Handbook of Frogs and Toads, 3rd edition. Comstock, Ithaca, New York.
Yagi A.R. 2008. Fowler’s Toad (Bufo fowleri) distribution and abundance in the Niagara Population Region 2001 to 2006. Report in preparation. Ontario Ministry of Natural Resources.
Yagi, A.R., and D. Mills. 2003. Fowler’s Toad (Bufo fowleri) abundance and habitat use at Morgan’s Point Conservation Area with habitat enhancement recommendations, Summer 2003. Ontario Ministry of Natural Resources Report.
Yagi. A.R., D. Mills and R. Tervo. 2004. Aylmer District Fowler’s toad (Bufo fowleri) Field Trip 2004 – Historic Elemental Occurrence Verification, Current Presence/Absence information with notes on Preliminary Habitat Characterization for Recovery Planning Purposes. Ontario Ministry of Natural Resources Report.
Yagi. A.R.,and R. Tervo. 2006. Distribution of Fowler’s Toad (Bufo fowleri) in Aylmer District based upon field surveys conducted in 2004 and 2005 with notes on Habitat for Recovery Planning Purposes. Report prepared for Ontario Ministry of Natural Resources Aylmer District and OMNR SAR. 21 pp.
Yagi A.R., and R. Tervo. 2008. Species at Risk Habitat Mapping for the Fowler’s toad (Bufo fowleri) – a Test of Draft Habitat Mapping Guidelines. Ontario Ministry of Natural Resources. 19 pp.
Yagi A.R., A. Brant, S.Meyer, D.M. Green, S.Dobbyn, K.Frohlich, K. Hayes, B. Johnson, M. Oldham, and R. Tervo. 2008. The Fowler’s Toad Stewardship Guide, prepared for Environment Canada Habitat Stewardship Program. 60 pp.
Yagi. A.R., D. Mills, and R. Tervo. 2004. Aylmer District Fowler’s Toad (Bufo fowleri) field trip 2004 – Historic elemental occurrence verification, current presence/absence information with notes on preliminary habitat characterization for recovery planning purposes. Ontario Ministry of Natural Resources Report.
Yagi.A.R., R. Tervo and K. Frohlich. 2006. Fowler’s Toad Habitat Enhancement Projects in the Vicinity of Morgan’s Point and the Easter Seal Camp Lakewood in Wainfleet. Interim Unpublished Report. Environment Canada Habitat Stewardship Program. 31 pp.
Zampella, R.A., and J.F. Bunnell. 2000. The distribution of anurans in two river systems of a coastal plain watershed. Journal of Herpetology 34:210–221
Zweifel, R.G. 1968. Effects of temperature, body size and hybridization on mating call of toads, Bufo a. americanusand Bufo woodhousei fowleri. Copeia 1968:269–285.
Professor David M. Green was born in Vancouver, British Columbia. He obtained a B.Sc. (hons) in Zoology from the University of British Columbia (1976) and an M.Sc. (1979) and Ph.D. (1982), both also in Zoology, from the University of Guelph. He was an NSERC Postdoctoral Research Fellow at the Museum of Vertebrate Zoology, University of California, Berkeley, and came to the Redpath Museum of McGill University in 1986 as an NSERC University Research Fellow and Assistant Professor. He became a Full Professor in 2004 and has been Director of the Redpath Museum since 2005.
Prof. Green’s research concerns the ecology, genetics, and evolution of amphibians. He has particular interest in species at risk, including the determinants of species’ ranges and population declines, population dynamics, dispersal and recruitment in amphibians, and declining amphibian populations. He has authored over 120 refereed publications and more than 100 miscellaneous other publications and reports. His books include The Amphibians and Reptiles of British Columbia (2007), Amphibians in Decline. Canadian Studies of a Global Problem (1997), and Amphibian Cytogenetics and Evolution (1991). Few of his publications fail to mention frogs or toads in some manner.
Prof. Green is a Fellow of the Linnean Society and a member of the American Society of Ichthyologists and Herpetologists, the Society for the Study of Amphibians and Reptiles, the Herpetologists’ League and the Society for Conservation Biology. He is, or has been, Associate Editor for Diversity and Distributions, (2007– ), Ecography (2007– ), the Zoological Journal of the Linnean Society (2006– ) and Herpetologica (1995–2002). He is currently on the editorial boards of the Canadian Journal of Zoology (2005 – ) and Biochemical Systematics and Ecology (1996– ). He was President of the Society for the Study of Amphibians and Reptiles from 2001–2002 and founding President of the Canadian Association of Herpetologists from 1987–1990. He was first National Co–ordinator of the Canadian Declining Amphibian Populations Task Force (DAPCAN) from 1991–1994, the fore–runner to the Canadian Amphibians and Reptiles Conservation Network (CARCNET). He was Chair of the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) from 1998–2002, co–chair of COSEWIC’s Amphibians and Reptiles Subcommittee from 1995–2009 and served on the Science Advisory Council of Fisheries and Oceans Canada from 2002–2005.
Professor M. Alexander (Alex) Smith grew up in the Ottawa Valley, Ontario. He obtained a B.Sc. (hons) and a M.Sc. (1998) in Biology from Trent University, and a Ph.D. (2004) from McGill University. He was an FQAR Postdoctoral Research Fellow at the University of Guelph, and in 2008 was hired as an assistant professor in Molecular Ecology at the University of Guelph. As a biologist, he works to integrate field and molecular biology into questions of ecological and evolutionary importance to conservation. He currently works on multiple projects that include molecular estimates of biodiversity and the distribution of phylogenetic diversity, the co–evolutionary ecology of host/parasitoid/symbiont relationships and the phylogeography of both ants and amphibians.
No collections needed to be re–examined for the preparation of this updated report.