
Monarch (Danaus plexippus): COSEWIC assessment and status report 2016

Endangered
2016
Table of contents
- Table of contents
- Assessment summary
- Executive summary
- Technical summary
- Preface
- Wildlife species description and significance
- Distribution
- Habitat
- Biology
- Population sizes and trends
- Threats and limiting factors
- IUCN 1. Residential and commercial development (low impact)
- IUCN 2. Agriculture and aquaculture (medium impact)
- IUCN 3. Energy production and mining (negligible impact)
- IUCN 4. Transportation and service corridors (low)
- IUCN 5. Biological resource Use (high impact)
- IUCN 6. Human intrusions and disturbance (negligible impact)
- IUCN 7. Natural system modifications (negligible)
- IUCN 8. Invasive and other problematic species and genes (unknown impact)
- IUCN 9. Pollution (high impact)
- IUCN 11. Climate change and severe weather (low impact)
- Limiting factors
- Number of locations
- Protection, status and ranks
- Acknowledgements
- Authorities contacted
- Information sources
- Biographical summary of report writers
- Collections examined
List of figures
- Figure 1. Monarch (Danaus plexippus) adult. Photo by Jessica Linton.
- Figure 2. Monarch (Danaus plexippus) caterpillar. Photo by Jessica Linton.
- Figure 3. Monarchs gathering at overwintering site in Mexico. Photo Jessica Linton.
- Figure 4. North American (native) range of Monarch (Danaus plexippus) showing the approximate distribution of the six subspecies (adapted from Pierce et al. 2015).
- Figure 5. The Canadian range extent of the Monarch (Danaus plexippus). The dashed line is the approximate northern distribution limit for caterpillar food plants Swamp Milkweed, Common Milkweed, and Showy Milkweed. Dots north of the line and in coastal B.C. represent non-breeding vagrants and occurrences at isolated patches of milkweeds planted outside their native range. Overwintering areas of eastern and western Monarchs are shown (grey areas).
- Figure 6. Estimated abundance of Monarchs (Danaus plexippus) at overwintering sites in California 1997 to 2015 (Monroe et al. 2016).
- Figure 7. Total area (hectares) occupied by Monarch (Danaus plexippus) colonies at overwintering sites in Mexico 1994/95 to 2015/16 (Monarch Watch 2016).
- Figure 8. Total area (ha) occupied by Monarchs (Danaus plexippus) at the overwintering sites in Mexico, with fitted regression lines that show the decline over time. The dotted line shows the fit over the entire time series (1994/95 – 2015/16). The solid line shows the fitted regression over the last ten years (2006/07 – 2015/16).
List of tables
- Table 1. The International Union for the Conservation of Nature - Conservation Measures Partnership (IUCN- CMP) threats calculator (see Salafsky et al. 2008; Master et al. 2009) was used to classify and list threats to the Monarch. Both the eastern and western migratory routes were included in the assessment. The results of the threats calculator show an overall threat impact of Very High.
Document information
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
![]()
COSEPAC
Comité sur la situation
des espèces en péril
au Canada
COSEWIC status reports are working documents used in assigning the status of wildlife species suspected of being at risk. This report may be cited as follows:
COSEWIC. 2016. COSEWIC assessment and status report on the Monarch Danaus plexippus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xiii + 59 pp. (Species at Risk Public Registry website).
Previous report(s):
COSEWIC. 2010. COSEWIC assessment and status report on the Monarch Danaus plexippus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 43 pp. (Species at Risk Public Registry website)
COSEWIC. 2001. COSEWIC assessment and update status report on the Monarch Danaus plexippus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. v + 21 pp. (Species at Risk Public Registry website)
1997. Draft. COSEWIC status report on the Monarch Danaus plexippus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa.
Production note:
COSEWIC would like to acknowledge Allan Harris and Robert Foster for writing the status report on the Monarch, Danaus plexippus, in Canada, prepared under contract with Environment Canada. This report was overseen and edited by Jennifer Heron, Co-chair of the COSEWIC Arthropods Specialist Subcommittee.
For additional copies contact:
COSEWIC Secretariat
c/o Canadian Wildlife Service
Environment and Climate Change Canada
Ottawa, ON
K1A 0H3
Tel.: 819-938-4125
Fax: 819-938-3984
E-mail: COSEWIC E-mail
Website: COSEWIC
Également disponible en français sous le titre Ếvaluation et Rapport de situation du COSEPAC sur le Monarque (Danaus plexippus) au Canada.
Cover illustration/photo:
Monarch -- Photo provided by author.
COSEWIC assessment summary
Assessment summary – November 2016
- Common name
- Monarch
- Scientific name
- Danaus plexippus
- Status
- Endangered
- Reason for designation
- This large showy species is one of the most well-known butterflies in the world. The Canadian population is migratory with two distinct pathways and cumulative threats at both overwintering sites and along the long migratory routes. The migratory group west of the Rocky Mountains moves between coastal California and southern British Columbia. The group east of the Rocky Mountains represents the vast majority of the Canadian population and moves between the Oyamel Forest of central Mexico and southern Canada east of Alberta. The overwintering sites in central Mexico are extremely small, and threats to these areas include illegal logging and agricultural development, and increased frequency and severity of storms during key congregation times. Declines of greater than 50% have occurred over the past decade.
- Occurrence
- British Columbia, Alberta, Saskatchewan, Manitoba, Ontario, Quebec, New Brunswick, Prince Edward Island, Nova Scotia, Newfoundland and Labrador, Northwest Territories
- Status history
- Designated Special Concern in April 1997. Status re-examined and confirmed in November 2001 and in April 2010. Status re-examined and designated Endangered in November 2016.
COSEWIC executive summary
Monarch
Danaus plexippus
Wildlife species description and significance
The Monarch is a conservation icon and one of the most well-known and well-studied butterflies in the world. The species has four life stages. The adult Monarch is a large (wingspan 93 – 105 mm), showy butterfly with predominantly orange wings outlined by a broad black border and two rows of circular white spots. The caterpillar is distinctively white, yellow, and black-banded, with a pair of black filaments at its head and tail. The chrysalis is green and gold. The eggs are approximately 1 mm long, oval with a flat base and bluntly pointed apex.
The Monarch is one of a few butterflies that migrate and their migration from southern Canada to Mexico has been described as an endangered biological phenomenon. The Monarch is used in classrooms all over North America to teach children about biology, metamorphosis, conservation, and an appreciation for nature.
Distribution
The Monarch is a migratory butterfly. The overall native range of the Monarch occurs from Central America northward through the continental United States to southern Canada, and from the Atlantic Coast westward to the Pacific Coast. The Canadian range of occurrence includes portions of all ten provinces and the Northwest Territories. Monarchs are loosely divided into eastern and western subgroups based on their migratory routes and overwintering sites. Eastern Monarchs breed from Alberta east to Nova Scotia and migrate south to overwinter in the mountains of Central Mexico. Western Monarchs breed in southern British Columbia and migrate south to overwinter in coastal California. The breeding range in Canada is south of the 50° latitude in Ontario, Quebec and the Maritimes and extends north to the 54° latitude in Manitoba, Saskatchewan and Alberta.
The Monarch is being assessed as one designatable unit in Canada. There is some exchange of individuals between the eastern and western migratory routes and no genetic or morphological evidence to suggest two subspecies.
Monarchs have also colonized continental Europe, North Africa, Australia and many Pacific islands within the last 200 years but these colonized populations do not migrate.
Habitat
Milkweeds (numerous species) are the sole food plant for Monarch caterpillars. These plants grow predominantly in open and periodically disturbed habitats such as roadsides, fields, wetlands, prairies, and open forests. Milkweeds are often planted outside their native range, and sometimes wayward Monarchs are observed at these patches.
Overwintering Monarchs require a cool, humid microclimate that is protected from frost, excessive sunlight, wind, and heavy precipitation. These conditions are found along the Pacific coast of California and the high elevation forests of central Mexico. Eastern Monarchs overwinter at elevations of 2900 - 3300 m in the Oyamel Fir forests in Mexico. Western Monarchs overwinter within a few kilometres of the Pacific coast of California, mainly in stands of non-native eucalyptus trees that replaced native pines starting in the 1850s, which were planted to replace native tree species.
Monarchs require staging areas which are used to rest, feed, and avoid inclement weather during migration. In Canada, they are found along the north shores of the Great Lakes where Monarchs roost in trees before crossing large areas of open water.
Biology
Adults mate during the winter or early spring at the overwintering sites in Mexico or California and begin flying north in late February or early March. About 10% of eastern Monarchs arriving in Canada fly the entire journey but most females that leave the overwintering sites breed in the southern United States. Female Monarchs lay 300 - 400 eggs singly on the undersides of milkweed leaves. The eggs hatch in three to eight days and the caterpillars feed almost continuously as they increase their body weight 2000-fold. After 9 - 14 days of feeding, the caterpillar transforms into a chrysalis and the adult emerges 9 - 15 days later. Adults of the following generations continue the migration north, many of which breed, reproduce and complete another generation in the central United States. Most Monarchs that reach Canada are the great-grandchildren of those that left Mexico. Monarchs in southern Ontario and Quebec produce two to three generations between June and October each year. Monarchs in southern British Columbia produce at least one generation each summer. Summer adults live for two to five weeks, but overwintering adults live up to nine months. The late summer adults migrate south to Mexico or California, where they overwinter and the yearly migration begins again.
Monarch caterpillars sequester the chemicals present in milkweed plants, which make them, as well as adult butterflies, unpalatable to most birds and other vertebrates.
Population size and trends
Population size estimates are not available for Monarchs in Canada. Each fall hundreds of thousands of Monarchs migrate through Long Point in southern Ontario but it’s unknown what proportion of the Canadian population these individuals represent. Population estimates are available for the overwintering sites, which include Monarchs from both Canada and the United States. The total overwintering population size in Mexico (eastern Monarch) was estimated at 66 million individuals in 2014-2015 and 200 million in 2015-2016. A storm in March 2016 killed a large but unknown number of Monarchs at the eastern overwintering sites. Fewer than 500,000 Monarchs currently overwinter in California (western Monarch), and only a tiny percentage of these breed in Canada. In some years, the western Canadian breeding population (in British Columbia) is so small as to be undetectable.
The overwintering population in Mexico, as measured by area of occupied habitat (hectares), declined significantly over the period 1994-2015. A log-linear regression of the time series indicates an 83% decline. The 2012-2014 estimates were the lowest in the time series. The area of occupied habitat in 2015 (4 ha) was higher than the previous three years but below the time series average of 6 ha. The decline rate of the occupied habitat over a 10-year period was calculated using the slope of a log-linear regression of the entire time series applied to a period of 10 years. The estimated 10-year change was estimated to be -59%.
A long-term migration monitoring study at Long Point, Ontario showed modest declines when numbers were adjusted for weather effects but similar studies at Cape May (New Jersey) and the Peninsula Point (Michigan) showed no evidence of decline.
Threats and limiting factors
Most North American Monarchs are concentrated in a few hectares in the winter and are vulnerable to extreme weather events, fire, diseases, predation, and anthropogenic threats. Overwintering habitat in Oyamel Fir forest in Mexico has been fragmented and degraded by conversion to agriculture, fire, logging, and forest thinning. These practices increase the exposure of overwintering Monarchs to winter storms, cold temperatures and wet conditions, resulting in increased mortality. Climate change models predict that the area of suitable forest at the overwintering sites in Mexico will decline and the frequency of winter storms will increase resulting in catastrophic mortality of Monarchs. Degradation to the western Monarch overwintering habitat is caused mainly by real estate development along the California coast and by elimination of introduced eucalyptus upon which the butterflies overwinter.
The increased use of herbicides and subsequent decline in milkweeds is a significant threat facing Monarchs throughout their North American range. Increased herbicide use may also cause declines in nectar supplies needed by migrating Monarchs and reduce overwinter survival. Neonicotinoid pesticides are an emerging threat, the magnitude of which is poorly understood.
Protection, status, and ranks
The Monarch was assessed as Special Concern by COSEWIC in 1997. The status was re-examined and confirmed in 2001 and 2010 and designated Endangered in 2016. Monarchs are classified as Special Concern under the Ontario Endangered Species Act and the New Brunswick Species at Risk Act. The Monarch is listed as “under special protection” in Mexico and in the United States, Monarchs are under review for listing under the United States Endangered Species Act. Breeding populations of Monarchs are ranked as Apparently Secure (G4) globally and Secure for Canada and the United States. Migratory concentration areas are ranked as Vulnerable to Imperiled in the United States.
Technical summary
- Scientific name:
- Danaus plexippus
- English name:
- Monarch
- French name:
- Monarque
- Range of occurrence in Canada:
- British Columbia, Alberta, Saskatchewan, Manitoba, Ontario, Quebec, New Brunswick, Nova Scotia, Prince Edward Island, Northwest Territories and Newfoundland (Labrador).
Demographic information
| Summary items | Information |
|---|---|
| Generation time | 0.33 years Mean age at which a cohort of newborns produce offspring, assuming three generations per year. |
| Is there an [observed, inferred, or projected] continuing decline in number of mature individuals? | Yes, inferred. Assuming overwintering population declines are reflective of Canadian populations. |
| Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations]. | Unknown |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations]. | Yes, estimated and inferred 59% decline rate based on area (ha) of occupied overwintering habitat in Mexico (Figure 8). Assuming overwintering population declines are reflective of Canadian populations. |
| [Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations]. | Unknown |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future. | Unknown |
| Are the causes of the decline a. clearly reversible and b. understood and c. ceased? | a. No Decline probably caused by the cumulative effects of declining milkweed, forest loss and degradation at wintering sites, and climate change. The impacts of neonicotinoid insecticides, diseases, declining nectar sources, are lesser known threats. |
| Are there extreme fluctuations in number of mature individuals? | No. Although indices of overwintering numbers fluctuate, changes have been less than an order of magnitude between any two successive years. |
Extent and occupancy information
| Summary items | information |
|---|---|
| Estimated extent of occurrence | ~ 1.3 million km2 Refers to extent of occurrence in Canada only. |
| Index of area of occupancy (IAO) (Always report 2x2 grid value). |
Canadian breeding range: Total overwintering (western [California] and eastern [Mexico]) range: 588 km2 |
| Is the population “severely fragmented” i.e., is >50% of its total area of occupancy is in habitat patches that are (a) smaller than would be required to support a viable population, and (b) separated from other habitat patches by a distance larger than the species can be expected to disperse? | a. No b. No |
| Number of locations (use plausible range to reflect uncertainty if appropriate) (Note: See Definitions and abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) |
Not applicable. |
| Is there an [observed, inferred, or projected] decline in extent of occurrence? | No |
| Is there an [observed, inferred, or projected] decline in index of area of occupancy? | No |
| Is there an [observed, inferred, or projected] decline in number of subpopulations? | No |
| Is there an [observed, inferred, or projected] decline in number of “locations”? (Note: See Definitions and abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) |
No |
| Is there an [observed, inferred, or projected] decline in [area, extent and/or quality] of habitat? | Yes, inferred decline in quality of habitat. Assuming decreases in milkweed abundance observed in the United States also apply in Canada. Overwintering habitat quality has declined in Mexico and California. |
| Are there extreme fluctuations in number of subpopulations? | No |
| Are there extreme fluctuations in number of “locations”? (Note: See Definitions and abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.) |
No |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | No |
Number of mature individuals (in each subpopulation)
| Subpopulations (give plausible ranges) | N Mature Individuals |
|---|---|
| Western | Unknown |
| Eastern | Unknown |
| Total | - |
Quantitative analysis
| Summary items | Information |
|---|---|
| Probability of extinction in the wild is at least [20% within 20 years or 5 generations, or 10% within 100 years]. | Predict 11% to 57% chance of quasi-extinction of the eastern Monarch population over the next 20 years (Semmens et al. 2016). Because this model only includes the eastern Monarch, the E criteria was not invoked. |
Threats (direct, from highest impact to least, as per IUCN Threats Calculator)
Was a threats calculator completed for this species? Yes.
IUCN threats calculator – calculated impact Very High
- 9.3 Agricultural and forestry effluents (high impact)
- 2.1 Annual and perennial non-timber crops (medium impact)
- 5.3 Logging and wood harvesting (medium impact)
- 1.1 Housing and urban areas (low impact)
- 4.1 Roads and railroads (low impact)
- 11.1 Habitat shifting and alteration (low impact)
- 11.2 Droughts (low impact)
- 11.3 Temperature extremes (low impact)
- 11.4 Storms and flooding (low impact)
- 9.5 Air-borne pollutants (unknown impact)
What additional limiting factors are relevant? Threats include habitats outside Canada, including the migration route throughout the United States and overwintering sites in Mexico and California.
Rescue effect (immigration from outside Canada)
| Summary items | Information |
|---|---|
| Status of outside population(s) most likely to provide immigrants to Canada. | Declining. |
| Is immigration known or possible? | Yes, possible. |
| Would immigrants be adapted to survive in Canada? | In summer, yes. |
| Is there sufficient habitat for immigrants in Canada? | Yes. |
| Are conditions deteriorating in Canada? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
Yes Assuming decreases in milkweed abundance observed in the United States also apply in Canada. |
| Are conditions for the source population deteriorating? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
Yes, based on decreases in milkweed abundance in the United States and habitat declines at overwintering sites in both Mexico and California. |
| Is the Canadian population considered to be a sink? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
No |
| Is rescue from outside populations likely? | Not likely. Populations elsewhere in the world do not migrate, it is possible wayward individuals could recolonize North American populations. |
Data-sensitive species
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
Status history
| Summary items | Information |
|---|---|
| COSEWIC Status History: | Designated Special Concern in April 1997. Status re-examined and confirmed in November 2001 and in April 2010. Status in re-examined and designated Endangered in November 2016. |
Status and reasons for designation:
| Summary items | Information |
|---|---|
| Status: | Endangered |
| Alpha-numeric code: | A2bce |
| Reasons for designation: | This large showy species is one of the most well-known butterflies in the world. The Canadian population is migratory with two distinct pathways and cumulative threats at both overwintering sites and along the long migratory routes. The migratory group west of the Rocky Mountains moves between coastal California and southern British Columbia. The group east of the Rocky Mountains represents the vast majority of the Canadian population and moves between the Oyamel Forest of central Mexico and southern Canada east of Alberta. The overwintering sites in central Mexico are extremely small, and threats to these areas include illegal logging and agricultural development, and increased frequency and severity of storms during key congregation times. Declines of greater than 50% have occurred over the past decade. |
Applicability of criteria:
| Summary items | Information |
|---|---|
| Criterion A (Decline in total number of mature individuals): |
Meets Endangered, A2bce, based on a decline in population estimates at central Mexican overwintering sites (b), a decline in habitat quality at both overwintering sites and throughout the migration routes (c) and pollutants (e). |
| Criterion B (Small distribution range and decline or fluctuation): |
Not applicable. Almost meets the criteria with a small IAO and decline in habitat quality based on the overwintering site but the number of locations are numerous and the species is not severely fragmented. |
| Criterion C (Small and declining number of mature individuals): |
Not applicable. Insufficient data to estimate decline in total number of mature individuals and subpopulations. |
| Criterion D (Very small or restricted population): |
Not applicable. Insufficient data on number of mature individuals within population. |
| Criterion E (Quantitative analysis): |
Not applicable. Insufficient data for quantitative analysis. |
Preface
The Monarch was first assessed by COSEWIC as Special Concern in April 1997. The species status was re-examined and confirmed in November 2001 and in April 2010. Since the most recent status report (2010), evidence from population estimates at the overwintering sites in Mexico suggests Monarchs have continued to decline. Major advances in Monarch research have improved understanding of habitat trends, population dynamics at different stages in the migration cycle, and basic biology. The emergence of herbicide-resistant crops and the impacts of increased herbicide use on milkweeds and nectaring plants is an emerging threat, as is climate change. Population viability and quasi-extinction models are also available. A large volume of research and publication on the Monarch continues as of this writing. An updated status report was initiated prior to the ten-year review schedule based on this additional information.
COSEWIC history
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) was created in 1977 as a result of a recommendation at the Federal-Provincial Wildlife Conference held in 1976. It arose from the need for a single, official, scientifically sound, national listing of wildlife species at risk. In 1978, COSEWIC designated its first species and produced its first list of Canadian species at risk. Species designated at meetings of the full committee are added to the list. On June 5, 2003, the Species at Risk Act (SARA) was proclaimed. SARA establishes COSEWIC as an advisory body ensuring that species will continue to be assessed under a rigorous and independent scientific process.
COSEWIC mandate
The Committee on the Status of Endangered Wildlife in Canada (COSEWIC) assesses the national status of wild species, subspecies, varieties, or other designatable units that are considered to be at risk in Canada. Designations are made on native species for the following taxonomic groups: mammals, birds, reptiles, amphibians, fishes, arthropods, molluscs, vascular plants, mosses, and lichens.
COSEWIC membership
COSEWIC comprises members from each provincial and territorial government wildlife agency, four federal entities (Canadian Wildlife Service, Parks Canada Agency, Department of Fisheries and Oceans, and the Federal Biodiversity Information Partnership, chaired by the Canadian Museum of Nature), three non-government science members and the co-chairs of the species specialist subcommittees and the Aboriginal Traditional Knowledge subcommittee. The Committee meets to consider status reports on candidate species.
Definitions (2016)
- Wildlife species
- A species, subspecies, variety, or geographically or genetically distinct population of animal, plant or other organism, other than a bacterium or virus, that is wild by nature and is either native to Canada or has extended its range into Canada without human intervention and has been present in Canada for at least 50 years.
- Extinct (X)
- A wildlife species that no longer exists.
- Extirpated (XT)
- A wildlife species no longer existing in the wild in Canada, but occurring elsewhere.
- Endangered (E)
- A wildlife species facing imminent extirpation or extinction.
- Threatened (T)
- A wildlife species likely to become endangered if limiting factors are not reversed.
-
Special concern (SC)
(Note: Formerly described as “Vulnerable” from 1990 to 1999, or “Rare” prior to 1990.) - A wildlife species that may become a threatened or an endangered species because of a combination of biological characteristics and identified threats.
-
Not at risk (NAR)
(Note: Formerly described as “Not In Any Category”, or “No Designation Required.”) - A wildlife species that has been evaluated and found to be not at risk of extinction given the current circumstances.
-
Data deficient (DD)
(Note: Formerly described as “Indeterminate” from 1994 to 1999 or “ISIBD” [insufficient scientific information on which to base a designation] prior to 1994. Definition of the [DD] category revised in 2006.) - A category that applies when the available information is insufficient (a) to resolve a species’ eligibility for assessment or (b) to permit an assessment of the species’ risk of extinction.
The Canadian Wildlife Service, Environment and Climate Change Canada, provides full administrative and financial support to the COSEWIC Secretariat.
Wildlife species description and significance
Name and classification
Kingdom : Animalia – animals
Phylum : Arthropoda –arthropods
Subphylum : Hexapoda – hexapods
Class : Insecta – insects
Subclass : Pterygota – winged insects
Order : Lepidoptera – butterflies and moths
Family : Nymphalidae Rafinesque 1815 – admirals, anglewings, brush-footed butterflies, checker-spots, crescent-spots, fritillaries, mourning clocks, purples
Subfamily : Danainae Boisduval, 1833 – Monarchs
Genus : Danaus Kluk 1780
Subgenus : Danaus (Danaus) Kluk 1780
Species : Danaus plexippus (Linnaeus 1758)
English Common Name: Monarch
French Common Name: Monarque
The Monarch (Danaus plexippus (L.)) was previously in the family Danaidae, but is now in subfamily Danainae: a diverse group that includes nymphs, satyrs, arctics, morphos, fritillaries, owl butterflies, and snouts (Opler and Warren 2002). It is the only representative of the milkweed butterflies in Canada (Marshall 2006).
Six subspecies of Monarch have been named. Danaus plexippus plexippus occurs in North America while the other subspecies occur in the Caribbean, Central America, and South America (Opler and Warren 2002). Two other members of the genus Danaus, the Queen (Danaus gilippus) and Soldier (D. eresimus) breed in the southern United States (US) (Scott 1986).
The discussion in this report often refers separately to the Monarchs east of the Rocky Mountains (eastern Monarchs) and west of the Rocky Mountains (western Monarchs). Eastern and western Monarchs have separate breeding and wintering ranges (see Global and Canadian range) but genetic evidence confirms both subgroups intermix and are subspecies plexippus (Lyons et al. 2012) (see Population Spatial Structure and Variability).
Morphological description
The Monarch butterfly has four separate and distinct life stages: egg, caterpillar, chrysalis and adult.
Adults:
The Monarch is a large (93-105 mm wing span), showy, orange and black butterfly (Figure 1). The wings are bright orange with heavy black veins and a thick black border containing two rows of circular white spots (Layberry et al. 1998). The males have a distinct black spot (scent gland) on the cubital vein of each hind wing; females do not have this black spot (Layberry et al. 1998).
Eggs:
Monarch eggs (approximately 1 mm long) are oval with a flat base and bluntly pointed apex. A series of approximately 18 ridges run from base to apex of the egg, which is whitish when first laid but darkens with age. Eggs are laid singly.
Caterpillar:
The caterpillar (larva) (Figure 2) are striped with alternating white, yellow, and black transverse bands. There is one pair of black filaments near the head and another near the end of the abdomen. Late instar caterpillars are up to 5 cm in length (Scott 1986; Schappert 2004).
Chrysalis:
The chrysalis (pupa) is green with a black and metallic gold band at the junction of the abdomen and thorax and scattered gold spots elsewhere (Scott 1986; Schappert 2004). The chrysalis hangs from a branch or other substrate, attached by a pad of silk spun by the caterpillar before pupation (Schappert 2004).
The Viceroy (Limenitis archippus) is similar in appearance to the Monarch and overlaps most of the Monarch’s Canadian range (Layberry et al. 1998). The Viceroy is smaller and has a black line crossing the veins on the hind wing that is lacking in the Monarch (Scott 1986; Layberry et al. 1998).

Long description for Figure 1
Photo of an adult Monarch, lateral view, among sunflowers. This large, showy butterfly has bright orange wings with heavy black veins and a thick black border containing two rows of circular white spots.

Long description for Figure 2
Photo of a Monarch caterpillar on a leaf. The caterpillar is striped with alternating white, yellow, and black transverse bands. There is one pair of black filaments near the head and another near the end of the abdomen.

Long description for Figure 3
Photo of clouds of Monarchs gathering at an overwintering site in Mexico.
Population spatial structure and variability
Monarch butterflies are migratory insects, and their broad distribution, migratory routes and overwintering sites have led to many questions and much research about their genetic lineages and evolutionary history (recently summarized in Pierce et al. 2015). It was initially proposed, based on the tagging methods of Urquhart and Urquhart (1977), that the eastern and western Monarchs use different migration routes and wintering grounds, although no genetic evidence was collected. However, subsequent genetic analysis, additional specimen tagging evidence, and field observations since this initial model (i.e., Urquhart and Urquhart 1977) indicate that these migratory pathways are not as well defined, are more complex, and that there is interchange of eastern and western Monarchs during migration (reviewed in Pierce et al. 2015 and Pyle 2015). A new and more complex model is suggested (Brower and Pyle 2004; Dingle et al. 2005; Pyle 2015) although further research is needed to explain the unknown degree of interchange between individuals at the California and Mexican overwintering sites.
The field verification available to support the interchange of eastern and western Monarch individuals, including extensive tagging studies, is summarized in Pyle (2015). One study, the Southwest Monarch Study, tagged wild-caught and captive bred Monarchs, released them in Arizona and then recovered some of those individuals at California and Mexican overwintering sites. When only wild-monarch data were considered, there is strong evidence that wild-caught Monarchs from west of the Continental Divide migrated both to Mexican and Californian overwintering sites (Southwest Monarch Study 2013 as read in Pyle 2015).
No separate genetic lineages have been revealed between the eastern or western Monarchs, probably because interbreeding occurs during migration when Monarchs from a broad geographical area intermix. The earliest genetic study used allozyme markers (enzymatic variants of proteins that represent genetic variation) (Eanes and Koehn 1978). The study showed that any genetic differentiation detected across breeding sites was erased during annual migration because Monarchs from different sites of pre-migration origin mix and mate randomly during migration; resulting in low genetic differentiation between eastern and western Monarchs (Eanes and Koehn 1978).
Mitochondrial DNA (DNA located in the mitochondria and inherited only from the mother) from 28 specimens was used to compare eastern and western migratory Monarchs with those in the neotropics (Brower and Boyce 1991). Specimens used in this study showed no differences in mitochondrial genome size among or within individuals, and only a single polymorphism was found (revealed by digestion with the restriction enzyme Ase1), although it was observed in Monarchs collected in both overwintering sites (Brower and Boyce 1991), and thus did not support differences among individuals migrating to eastern and western overwintering sites. Almost identical genetic patterns were shown in all populations, as all other restriction fragments were monomorphic among all individuals tested (Brower and Boyce 1991).
The most recent study involved microsatellite markers (short sequences of DNA that are repeated in tandem) and compared five samples: one from Florida in eastern North America, two from California in western North America, and samples from non-migratory populations in Hawai’i and New Zealand (Lyons et al. 2012). The value of FST (the proportion of the total genetic variation attributable to differences between populations) between the North American samples was very low, 0.0012, and not statistically different from 0 (P = 0.2). Lyons et al. (2012) concluded that North American Monarchs constitute a single, genetically admixed population.
Designatable units
The Monarch is being assessed as one designatable unit in Canada. Genetic and field evidence indicates that eastern and western Monarchs intermix (see Population Spatial Structure and Variability). Further, there is no evidence of morphological, behavioural, or molecular genetic differences between eastern and western Monarchs.
Monarchs range in the southern portions of the Atlantic, Boreal, Great Lakes Plains, Pacific, Prairie and Southern Mountain COSEWIC (2017) ecological areas.
Special significance
The Monarch is a conservation icon and one of the most well-known and well-studied butterflies in the world (Gustafsson et al. 2015). Monarch migration from southern Canada through the continental United States to Mexico has been described as an endangered biological phenomenon (Brower 1996). The Monarch is used in classrooms all over North America to teach children about biology and natural history, life cycles and metamorphosis, conservation, and an appreciation for nature.
Distribution
Global range
The global range of the Monarch extends from southern Canada, through the United States and Central America to northern South America; and from the Atlantic to the Pacific (Figure 5). The Monarch is often reported outside this native range, often due to the release of captive bred and wayward individuals. These sightings are not considered part of its natural range.
Six subspecies of Monarch have been named: D. p. plexippus (North America and all areas where the species occurs outside the Americas [the subspecies being assessed in this status report]), D. p. leucogyne (Virgin Islands), D. p. portoricensis (Puerto Rico), D. p. tobagi (Tobago), D.p. megalippe (Caribbean and Central America, Smith et al. 1994), and D. p. nigrippus (South America) (Opler and Warren 2002; Pelham 2008; NatureServe 2015) (Figure 4). Some genetic studies suggest that some or all of the populations inhabiting the Caribbean Islands and South America constitute a single subspecies (e.g. Smith et al. 2005). Unlike the more northern populations, the Central American and Caribbean subspecies are relatively sedentary and do not contribute to the migratory population of Monarchs found in Canada and the United States (Crolla and Lafontaine 1996). Genetic research shows that migratory Monarchs dispersed out of North America and are ancestral to the non-migratory Caribbean, Central American and South American subspecies (Zhan et al. 2014). There is also some evidence that migratory populations are genetically distinct from the non-migratory population (Zhan et al. 2014).
Vagrant Monarchs apparently crossed the Atlantic in the mid- to late 1800s to islands of the eastern Atlantic and continental Europe (Fernandez-Haeger et al. 2015). Populations are now established in North Africa, Spain, Portugal, the Azores, Canary Islands, and other islands (Fernandez-Haeger et al. 2015). How Monarchs reached Australia is unknown, but they were first reported around 1870 and spread through eastern Australia and parts of Western Australia (Zalucki and Rochester 2004). Monarchs apparently colonized Hawaii, New Zealand, and the larger islands in the Pacific Ocean (Galapagos, Solomons, Norfolk, Philippines, and Taiwan) after 1940 (Vane-Wright 1993).
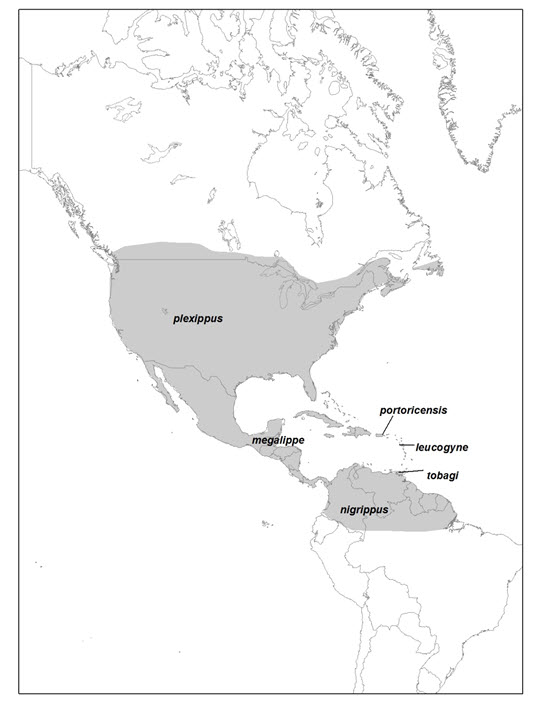
Long description for Figure 4
Map of the American (native) range of the Monarch, showing the approximate distribution of the six subspecies. The Monarch occurs from southern Canada, through the United States and Central America to northern South America, and from the Atlantic to the Pacific.
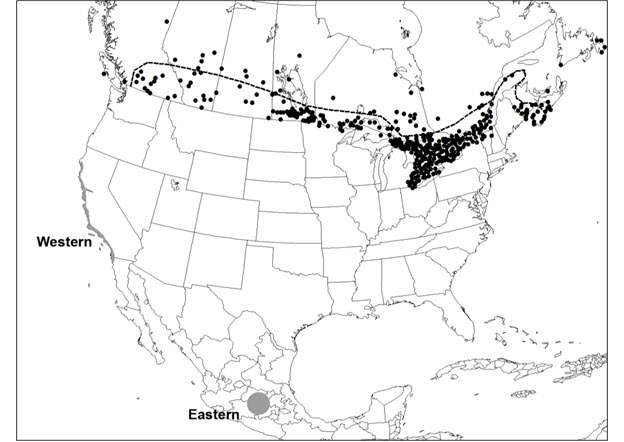
Long description for Figure 5
Map of the range extent of the Monarch in Canada, where it has been recorded from all 10 provinces and the Northwest Territories. The approximate northern distribution limit for caterpillar food plants (Swamp Milkweed, Common Milkweed, and Showy Milkweed) is indicated.
Eastern Monarchs breed east of the Rockies from the Gulf Coast to southern Canada and migrate to central Mexico to overwinter (Urquhart 1987). Western Monarchs breed from the southwestern United States (Arizona and New Mexico) northwards to southern Canada (British Columbia [BC]) and from the Rocky Mountains westwards to the Pacific Coast. They overwinter along the coast of California with small numbers extending south into northern Mexico (Lane 1984).
South Florida and Cuban populations are non-migratory, although some migrants from northern populations migrate through Florida and become assimilated with resident breeding populations (Dockx et al. 2004; Knight and Brower 2009). The offspring of these individuals apparently do not migrate north in the following spring. In contrast to overwintering Monarchs in Mexico and California, resident Monarchs in south Florida do not form overwintering clusters or enter diapause. They are active year round and breed continuously (Knight and Brower 2009).
Canadian range
Monarchs have been recorded in Canada from all ten provinces and the Northwest Territories (NT) (Layberry et al. 1998; Environment and Climate Change Canada 2016) (Figure 5). The breeding range in Canada is south of the 50° latitude in Ontario (ON), Quebec (QC) and the Maritime Provinces (New Brunswick [NB], Newfoundland [NF], Nova Scotia [NS] and Prince Edward Island [PE]) and extends north to the 54° latitude through Manitoba (MB), Saskatchewan (SK) and Alberta (AB). The continuous range corresponds with the range of the Monarch food plants, milkweeds (Asclepias spp.) (Brower 1996; Crolla and Lafontaine 1996). In NS breeding is confined mainly to the Annapolis Valley (Crolla and Lafontaine 1996) (Figure 5). Monarch records north and east of the native range of milkweeds represent non-breeding vagrants and breeding occurrences, at planted milkweeds. Vagrant butterflies have been observed in NF and NT, as well as in northern areas of other provinces (Environment and Climate Change Canada 2016).
Southern ON and QC hold the most significant Monarch breeding areas in Canada (White 1996). Long Point (both the provincial park [PP] and surrounding habitats), Rondeau PP, and Point Pelee National Park (NP) on the north shore of Lake Erie and Presqu’ile PP on Lake Ontario are important staging areas where Monarchs cluster before crossing the Great Lakes. Monarchs are common in southern QC but are rarely observed north of 50° latitude. In the fall, large concentrations of migrating Monarchs aggregate in specific areas in southern QC such as along Valleyfield and Missiquoi bays (Handfield et al. 1999).
In AB, SK and MB, the Monarch primarily ranges in the south and abundance decreases north and west from southern MB to AB (Klassen et al. 1989, Layberry et al. 1998). In some years, Monarchs breed as far north as Edmonton (AB). Monarchs are fairly common in southern MB, except in the southeast which is extensively wooded and supports less Monarch habitat (Klassen et al. 1989).
In BC, breeding is reported from scattered sites in the southern portions of the province, particularly in the Okanagan and Similkameen valleys and the Kootenays (southeast) (Guppy and Shepard 2001; St. John et al. 2014; B.C. Conservation Data Centre 2016). Wayward individuals are reported from the lower Fraser Valley and Vancouver Island, but these areas are not within the native distribution of milkweeds and these geographic areas are not considered part of the Monarch’s native range.
Approximately 10% of the global breeding range of the Monarch is in Canada. This is based on records of caterpillar observations along the international border from the Maritimes west to southern BC.
At the time of writing this status report, there was no Aboriginal Traditional Knowledge available on Monarch distribution or abundance in Canada.
Extent of occurrence and area of occupancy
The Monarch’s extent of occurrence in Canada (excluding records of vagrants and those breeding at milkweeds in gardens outside their historical range) corresponds with the distribution of the three primary caterpillar food plants; Swamp Milkweed (Asclepias incarnata), Common Milkweed (Asclepias syriaca), and Showy Milkweed (Asclepias speciosa) (White 1996). A convex polygon encompassing the native distribution of milkweed species in Canada (Figure 5) covers over 3.5 million km2 and approximates the extent of occurrence for eastern and western Monarchs. Given the migratory nature of this species and the weather-induced variance in the extent to which it spreads north each year, the extent of occurrence in Canada fluctuates considerably between years.
The index of area of occupancy (IAO) in Canada, based on a 2 km x 2 km grid intersecting breeding occurrences, was not calculated due to a lack of detailed information on the distribution of breeding sites throughout the country. Regardless, the IAO undoubtedly exceeds 2,000 km2.
The IAO for the overwintering range of the eastern Monarch was estimated using the number of overwintering colonies in Mexico. In 2015, the overwintering area consisted of nine colonies ranging in size from 0.13 ha to 1.09 ha (Anonymous 2016). If each colony occupies a unique 2 km x 2 km grid square, the maximum IAO in 2015 was 36 km2.
The IAO for the overwintering range of western Monarchs in California spanned 138 sites in 2015 (Monroe et al. 2016) for a maximum IAO of 552 km2.
The total IAO for the western and eastern Monarch overwintering sites in 2015 was 588 km2.
Search effort
The distribution of the Monarch in Canada was determined by the efforts of collectors, researchers, and citizen scientists. Monarch distribution and abundance have been summarized at the national level in Layberry et al. (1998) and regionally for BC (Guppy and Shepard 2001; B.C. Conservation Data Centre 2016), AB (Bird et al. 1995), Manitoba (Klassen et al. 1989), ON (Holmes et al. 1991, Macnaughton et al. 2016), QC (Handfield et al. 1999), NB (Thomas 1996), and the Maritime Provinces (Maritimes Butterfly Atlas 2016) and within the species national recovery plan (Environment and Climate Change Canada 2016).
The Toronto Entomologists’ Association has compiled annual summaries of butterfly sightings from across ON since 1968 (Macnaughton 2010) including regular status reports on [eastern] Monarchs (e.g. Hess 1992). The Ontario Butterfly Atlas Online (Macnaughton et al. 2016) includes 280,000 records of butterfly species from the Toronto Entomologists’ Association annual summaries and eButterfly.
Monarch observations in BC [western Monarchs] are primarily incidental sightings or reports from naturalists or biologists with few, if any, surveys specifically targeting Monarchs (St. John et al. 2014). A targeted survey in the Okanagan and Similkameen river valleys in 2014 searched 758 Showy Milkweed sites over 37 days in mid- to late summer (St. John et al. 2014). During 2014, eggs from a gravid female Monarch were raised in captivity and the adults tagged and released in an effort to gain natural history knowledge on Monarch populations in the south Okanagan (Antonijevic et al. 2014).
The Monarch’s widespread appeal along with its ease of observation and identification make it suitable for monitoring by the public (Commission for Environmental Cooperation 2009). Ongoing citizen science projects relevant to Canadian Monarchs include Monarch Watch (tagging, fall migration), Mission Monarch (egg and caterpillar counts), Journey North (sightings, migration monitoring), Monarch Caterpillar Monitoring Project (egg and caterpillar surveys), North American Butterfly Association (annual censuses), eButterfly (photos and sightings), the Maritime Butterfly Atlas, and Ontario Butterfly Atlas (Howard and Davis 2015; Oberhauser et al. 2015a; Macnaughton et al. 2016; Maritimes Butterfly Atlas 2016).
Habitat
Habitat requirements
Monarch butterflies require different habitats depending on their life stage. The breeding and migration habitats described below apply to habitats in Canada. The overwintering habitats refer to the sites in Mexico (eastern Monarchs) and California (western Monarchs).
Breeding habitat:
In Canada, Monarch caterpillars feed exclusively on milkweed plants (Asclepias spp.) and the breeding habitat is confined to places where milkweeds grow. Monarchs are known to use 11 of the 14 milkweed species that occur in Canada (White 1996; Borders and Lee-Mader 2014). However, in Canada the most common milkweeds used by Monarchs are Showy Milkweed in BC, AB, SK and MB; and Common Milkweed and Swamp Milkweed in ON and eastern Canada. Milkweeds occur in a wide range of habitats including roadsides, fields, wetlands, meadows, prairies, and open forests (Borders and Lee-Mader 2014).
Milkweeds are often planted in gardens beyond the natural range of the plant and potentially extending the breeding range of Monarchs. Milkweeds in soybean and corn fields tend to have higher density of Monarch eggs and higher survival of eggs and early instars than milkweeds in non-agricultural habitats, possibly due to less predation and higher palatability of the leaves (Pleasants 2015). An estimated 80% of Monarchs in the Midwest United States fed on milkweed in agricultural fields (Pleasants 2015).
Adult Monarchs feed at milkweed flowers but require other wildflowers for nectar, especially when milkweeds are not in bloom (Crolla and Lafontaine 1996). In Canada, the most commonly used alternate nectar sources are goldenrods (Solidago spp.), asters (Doellingeria spp., Eurybia spp., Oclemena spp., Symphyotrichum spp. and Virgulus spp.), the non-native Purple Loosestrife (Lythrum salicaria), and various clovers (Trifolium spp. and Melilotus spp.) (Crolla and Lafontaine 1996). Nectar sources are especially important during the fall migration when sugars from the nectar are converted to fat reserves required for winter survival (Urquhart 1987; Gibo and McCurdy 1993).
Migration or staging habitat:
Monarchs require areas to rest, feed, and avoid inclement weather during migration. In Canada, staging areas occur along the north shores of the Great Lakes where Monarchs roost in trees before crossing these large areas of open water. Notable sites include Presqu’ile PP, Point Pelee NP, Rondeau and Long Point. Monarchs are highly adaptable in terms of roost attributes but apparently choose staging areas based on proximity to large lakes, tree size, and wind direction (Schappert 2004; Davis et al. 2012). In the span of a few hours, tens to hundreds of thousands of Monarchs have been observed leaving the tip of Point Pelee, apparently after staging on the point overnight (Wormington 1997).
Overwintering habitat:
Overwintering Monarchs require a cool microclimate, but also protection from seasonal frost. High humidity and protection from excessive sunlight, wind, and precipitation are also crucial (Williams and Brower 2015). These conditions are found in the Pacific coast of California and the high elevation forests of central Mexico.
Eastern Monarchs overwinter in the high elevation Oyamel Fir (Abies religiosa) forests of central Mexico (Williams and Brower 2015). Oyamel forest is found at 2900m above sea level (asl) to 3300 m asl and has a spatial area of approximately 80 km x 80 km. Suitable forest patches within that area total about 562 km2 (Slayback et al. 2007). Overwintering colonies are found in areas of the forest with greater tree cover than randomly selected non-colony areas (Williams et al. 2007). The forest canopy protects against precipitation and winds, freezing temperatures as well as warm temperatures so Monarchs can maintain a low metabolic rate (Williams and Brower 2015).
Western Monarchs overwinter along a 1000 km (approximate) stretch of the Pacific coast of California south into northern Baja California. Approximately 450 overwintering sites have been recorded along this stretch (Jepsen and Black 2015), most of them associated with stands of non-native Australian eucalyptus trees (Eucalyptus globalus and E. camaldulensis) (Frey and Schaffner 2004; Jepsen and Black 2015). Eucalyptus trees were introduced in the 1850s for landscaping, as windbreaks, and for use as fuel, resulting in an increase in suitable overwintering habitat for Monarchs (Crolla and Lafontaine 1996). This growth of eucalyptus coincided with cutting of the dense coastal stands of native tree species, such as Monterey Pine (Pinus radiata) and Monterey Cypress (Cupressus macrocarpa), which are also used by Monarchs as overwintering sites where they persist (Lane 1993; Jepsen and Black 2015).
Overwintering sites in California are associated with maritime climate usually within 2.4 km of the coast where temperatures rarely drop below freezing (mean January temperature of 10° to 12° C) but remain cool enough to allow Monarchs to conserve energy. These sites also have high humidity and morning dew to prevent dehydration. Most sites are on slopes oriented to the south or west which offer solar radiation and wind protection (Leong et al. 2004).
Habitat trends
Overwintering habitat:
The historical extent of eastern Monarch overwintering habitat in the Oyamel Fir forest (Mexico) is not well documented; however, prior to 1971 much of the area was continuous high-density forest (Brower et al. 2002). Between 1971 and 1999, 44% of the dense forest (i.e. >80% cover) was clearcut, thinned, or burned. The largest patch of high-quality forest was reduced from 271 km2 to 58 km2 (Brower et al. 2002).
The Monarch Butterfly Biosphere Reserve was designated in 1980 and its boundaries defined in 1986. It has a core area of 130 km2 and buffer area of more than 420 km2 (Jepsen et al. 2015). Between 1986 and 2012, a total of 4,300 ha of Oyamel Fir forest was lost or disturbed (8% of the reserve) due to illegal logging, fires, domestic timber extraction and agricultural clearing (Ramirez et al. 2015). The rate of illegal logging increased immediately after the reserve was established but then decreased from 2005-2007 (731 ha) to 2014 (5.2 ha) (Vidal and Rendón-Salinas 2014). Illegal logging of 19 ha occurred in the state of Michoacan in 2014 – 2015 (WWF 2015).
Overwintering habitat in California has also declined. At least 38 Monarch wintering sites were destroyed by housing developments before 1991. An additional 11 sites were destroyed by housing developments in the 1990s and two others are presently slated for development (Jepsen and Black 2015). Degradation of some sites has occurred due to tree trimming, eucalyptus removal, and die-off of older trees (Frey and Schaffner 2004; Jepsen and Black 2015).
Climate models predict that suitable climatic habitat for Oyamel Fir will decline by 50% by 2030 and be completely eliminated by 2090 as the impacts of pests and disease increase with increasing drought stress (Ramirez et al. 2015) (see Threats section 11.1).
Breeding Habitat:
Monarch breeding habitat in many parts of Canada has undergone significant changes over the last 200 years. Milkweeds are adapted to open habitat and disturbed soil and until the 1880s, the prairies of central North America were the main breeding area of eastern Monarchs (Brower 1995). Extensive prairie and savannah were also present in southern ON and QC, and maintained by First Nations peoples’ burning practices (Riley 2013), which ceased following the European settlement of these areas. Deciduous forest expanded as First Nations’ burning practices declined (Riley 2013) and Monarch habitat declined with the increasing forest cover. In the late1800s, clearing of the deciduous forest for agriculture in northeastern North America including southern ON and QC, and portions of the Maritime Provinces, undoubtedly allowed the rapid spread of milkweeds and eventually Monarchs (Brower 1995; Pleasants 2015).
Habitat trends can be estimated from remote sensing data, assuming that Monarch habitat consists of cropland and grassland cover types. More recently, Monarch habitat in southern Canada has declined with the conversion of grassland to more intensive agricultural cropland and other land uses. Between 2000 and 2011 (the most recent data available), about 2,600 km2 of cropland and grassland were converted to urban, rural and industrial land use, 10,500 km2 succeeded to forest and shrub, and 5,300 km2 were lost to other causes (Agriculture and Agri-Food Canada 2012). During the same time span, approximately 6,400 km2 of forest was converted to cropland. In total, approximately 12,000 km2 of Monarch habitat is considered lost during this period. About 7,000 km2 of grassland was converted to cropland during this period but the impacts on Monarchs are unknown.
In the United States, since the mid-1990s, approximately 700,000 km2 of Monarch breeding habitat east of the Rockies (eastern Monarchs) has been lost due to agricultural changes and development (The Center for Biological Diversity et al. 2014). This area is approximately one third of the Monarch’s summer breeding range. Much of the habitat loss was caused by increased use of glyphosate on herbicide resistant crops (see Threats). Other habitat loss was caused by conversion of unused lands to farmland and by commercial and residential development. Increased demand for biofuels has caused the loss of about 45,000 km2 of Monarch breeding habitat. Fallow farmlands (i.e. lands enrolled in the Conservation Reserve Program) were converted to corn and soybeans between 2007 and 2014 (The Center for Biological Diversity et al. 2014). The increased use of herbicides on these lands reduced the habitat value for Monarchs by killing milkweeds and nectaring plants (see Threats). Between 1982 and 2010, 174,000 km2 of land in the United States were converted to residential and commercial development (The Center for Biological Diversity et al. 2014).
West of the Rocky Mountains (western Monarchs), breeding habitat trends are less well studied and the most important breeding areas have not been identified (Jepsen and Black 2015), particularly within BC (Guppy pers. comm. 2016; Heron pers. comm. 2016; St. John pers. comm. 2016). Showy Milkweed is abundant along roadsides in southern BC and does not appear to be a limiting factor for breeding Monarchs (Dyer pers. comm. 2015; Guppy pers. comm. 2015; St John pers. comm. 2015).
Biology
The Monarch is one of the most well studied insects in North America. Researchers in Canada and the United States have completed extensive studies on migration, genetics, overwintering physiology, feeding habits, predators and parasites, and diseases. Tagging and isotope studies have filled information gaps about migration biology, while modelling has provided insights into population dynamics. Compilations by Oberhauser and Solensky (2004) and Oberhauser et al. (2015a) include recent research and comprehensive reviews of many aspects of Monarch biology. Summaries of Monarch biology are also found in Schappert (2004), Commission for Environmental Cooperation (2008), and Jepsen et al. (2015).
Life cycle and reproduction
The Monarch life cycle, like all butterflies, has four life stages: egg, caterpillar (caterpillar), chrysalis (chrysalis), and adult. Summer Monarchs (i.e., not overwintering adults) mate when adults are three to eight days old and females begin to lay eggs immediately after mating (Oberhauser 2004). Females mate up to 10 times, receiving nutrients and salts in the form of a spermatophore (Suzuki and Zalucki 1986). Monarchs lay 300 to 400 eggs on the underside of milkweed leaves (Oberhauser 2004). Eggs are laid singly, but several eggs are often laid on different leaves of the same plant (Crolla and Lafontaine 1996). The eggs hatch in three to eight days (Schappert 2004).
The caterpillar stage (Figure 2) lasts nine to 14 days under normal summer temperatures but longer under cooler conditions (Oberhauser 2004). Caterpillar feed on milkweed leaves almost continuously as they grow to a total length of 5 cm and increase their body weight 2000-fold (Oberhauser 2004). In preparation for the chrysalis stage, the caterpillar spins a pad of silk and hangs upside down as it transforms into a chrysalis (Urquhart 1987). Emergence of the adult occurs 9 to 15 days after pupation and about 10% of eggs survive to adulthood (Oberhauser 2004).
Monarchs in southern ON and QC produce two to three generations between June and October each year (Holmes et al. 1991; Crolla and Lafontaine 1996). Monarchs in southern BC produce at least one generation each summer (Guppy and Shepard 2001; St. John pers. comm. 2015; B.C. Conservation Data Centre 2015). Summer adults live for two to five weeks, but overwintering adults live up to nine months (Oberhauser 2004).
The generation of Monarchs that emerges in late summer or fall is in reproductive diapause and migrates from Canada (and elsewhere in the United States) to the overwintering grounds (Solensky 2004). Diapause is triggered by decreasing day length, decreasing temperatures, and senescing food plants in autumn, but a few individuals break diapause during migration and reproduce in the southern United States before reaching the wintering grounds (Solensky 2004).
Overwintering Monarchs mate during the winter or early spring but do not oviposit until they find milkweeds on their northward migration in the southern or central United States. These adults emerge and continue the migration north. About 90% of Monarchs reaching the breeding range in the Great Lakes region originate from the southern (south of 35°N latitude) or central (35°N to 41°N latitude) United States. The remaining 10% complete the migration in a single sweep (Miller et al. 2012). Monarchs from the central United States originate from both overwintering parents and first generation young from the southern United States indicating that most Canadian Monarchs require at least one generation to complete the northward migration (Miller et al. 2012; Flockhart et al. 2013). Western Monarchs are less well studied but apparently go through several generations between leaving California and arriving in southern BC (Jepsen et al. 2015).
Physiology and adaptability
Habitat adaptability:
The Monarch has been able to adapt to some changes in the environment and colonize new habitats. Within the last 200 years, Monarchs have successfully colonized Australia, Europe, North Africa, and numerous oceanic islands around the world, demonstrating that they are able to adapt to new physical environments and food plants. Most or all of these new populations are non-migratory.
Western Monarchs have successfully switched from overwintering in native coastal forests to non-native eucalyptus trees in California (summarized in Jepsen and Black 2015). Eastern Monarchs were able to colonize new habitats when the eastern deciduous forests were cleared and converted to farmland, roadsides, and railways (Crolla and Lafontaine 1996). More recently Monarchs have been recorded from patches of cultivated milkweed planted outside their native range (e.g., within homesteads and gardens).
Eastern Monarchs have recently been recorded overwintering in small numbers in Texas and on the Gulf Coast of the United States, suggesting that new wintering areas can be established. These Monarchs were generally in groups of less than 10 individuals rather than forming the large overwintering clusters observed in Mexico and California. It is unknown if these individuals go on to breed and contribute to the next generation (Howard et al. 2010).
Compound sequestration:
Many predators avoid eating Monarchs because the caterpillars sequester cardenolides (steroids produced for protection against herbivores) from milkweeds, which deter predation by most birds and other vertebrates (Malcolm and Brower 1989). The adults also have these sequestered compounds.
Energy reserves:
Prior to migration, adult Monarchs store lipid nutrients (from nectar sources) as energy reserves that can metabolize quickly (Gibo and McCurdy 1993).
Dispersal and migration
Monarchs famously migrate from southern Canada and the northern United States to Mexico and California each fall, and return back to these areas again each spring. The northward migration from Mexico typically starts from March to early April. Several successive generations advance northward and typically reach Canada in late May or early June (Wormington 2008). About 10% of individuals that wintered in Mexico make the entire journey to Ontario (Miller et al. 2012). Such individuals have been recorded at Point Pelee NP in late April and early May albeit in a worn condition (Wormington pers. comm. 2007).
The southward migration typically starts in early August and may extend through to early November (Wormington pers. comm. 2007). Staging Monarchs cluster together on trees along the north shores of Lake Ontario and Lake Erie to form overnight roosts containing hundreds to thousands of individuals. The most significant staging areas are at Presqu’ile PP, Long Point, Long Point National Wildlife Area, Rondeau PP, and Point Pelee NP (Crolla and Lafontaine 1996). The departure of hundreds of thousands of Monarchs from Point Pelee NP has been observed numerous times (Wormington 1994; 1997; 2008).
Eastern Monarchs follow a central flyway through Kansas, Oklahoma, and Texas to Mexico, or an eastern flyway along the Atlantic seaboard Gulf Coast (Brindza et al. 2008; Howard and Davis 2009). Western Monarchs follow a shorter migration route south along the coast of California as far as northern Mexico (Crolla and Lafontaine 1996).
Monarchs can reduce their energy expenditure during the fall migration by soaring, gliding, and riding columns of rising warm air to reach altitudes where strong prevailing winds speed their flight (Gibo and Pallett 1979; Gibo 1981). Monarchs orient using the sun (Perez et al.1997) and magnetic compasses contained in light-sensitive magneto-sensors in the antennae (Guerra et al. 2014).
Interspecific interactions
The obligate protozoan parasite, Ophryocystis elektroscirrha, is present in North American, South American, and Australian Monarch populations (Altizer and Oberhauser 1999). The parasite has little effect on Monarch survival or reproduction, except at high levels of infection (Altizer and Oberhauser 1999). Monarchs inoculated with high levels of parasites had decreased survival to hatching, smaller wingspans, lower body mass, shorter flight distances, and slower flight speeds than non-inoculated adults (Altizer and Oberhauser 1999; Bradley and Altizer 2005). Migratory Monarch populations have a lower infection rate than non-migratory populations. Infected Monarchs fly less efficiently and migration may weed out the infected animals and reduce parasite prevalence and transmission between generations (Bradley and Altizer 2005; Satterfield et al. 2015).
Arthropod predators on Monarchs include spiders, lacewings, mantids, dragonflies, ants, and wasps. Some predators can cope with the cardenolide toxins by overcoming taste aversion, selecting Monarchs with low levels of toxins, selectively feeding on parts of the body with low toxin levels, or other mechanisms (Fink and Brower 1981, Oberhauser et al. 2015b). Parasitoids include 12 species of tachinid flies (especially Lespesia archippivora) and a brachonid wasp (Oberhauser et al. 2015b). Two species of birds, the Black-headed Grosbeak (Pheucticus melanocephalus) and the Black-backed Oriole (Icterus galbula abeillei), feed extensively on Monarchs at the overwintering sites in Mexico (see Threats and Limiting Factors).
Population sizes and trends
Sampling effort and methods
There is little population size or trend data available for Monarchs breeding in Canada. Migration monitoring at Long Point provides a trend index for Monarchs originating north of Lake Erie. Trends can be inferred from overwintering numbers in California and Mexico where Canadian and United States Monarchs share roosts.
The most systematic monitoring data are collected at wintering sites in California (western Monarchs) and Mexico (eastern Monarchs) where large numbers of Monarchs are concentrated in small areas. Since 1993, population trends in Mexico have been monitored at about 20 sites by measuring the spatial area (hectares [ha]) occupied by clusters of overwintering Monarchs (Brower et al. 2012; Garcia-Serrano et al. 2004; Vidal and Rendon-Salinas 2014). Two methods have been used to estimate total overwintering population size. One method extrapolates from mark-release-recapture techniques. Another measures the density of Monarchs on a subsample of branches and extrapolates for the size of the colony. Data are insufficient to assess which method is most accurate (Calvert 2004; Solensky 2004).
Monitoring at overwintering sites in California has taken place since 1997 (Jepsen and Black 2015). The Western Monarch Thanksgiving Count engages volunteers to count roosting Monarchs during a two week window in the fall at about 100 coastal sites (Monroe et al. 2015).
The North American Butterfly Association Seasonal Count Program, eButterfly, the Illinois Butterfly Monitoring Network, Monarch Caterpillar Monitoring Program, and the Ontario and Maritime Butterfly atlases are among the volunteer-based programs applied in the breeding range (Macnaughton et al. 2016; Maritimes Butterfly Atlas 2016; Reis et al. 2015a,b; Inamine et al. 2016).
Migrating Monarchs at Long Point on the north shore of Lake Erie (ON) have been monitored since 1995 (Crewe and McCracken 2015). Standardized daily counts of adults are made at two stations. Similar counts are conducted at Cape May, New Jersey and Peninsula Point, Michigan in the United States (Crewe and McCracken 2015).
Abundance
No estimates are available for the Monarch population size in Canada. “Hundreds of thousands” of Monarchs migrate through Long Point each fall (Crewe and McCracken 2015) but it’s unknown what proportion of the Canadian population this estimate represents. In the Maritime Provinces, Monarchs are widespread but generally uncommon and abundance varies significantly from year to year (Maritimes Butterfly Atlas 2016). In the Maritime Provinces, high numbers were seen in 2012, when there was a massive migration (Klymko pers. comm. 2016). In NB, Monarchs are recorded annually, usually in small numbers and breeding occurs mainly along the banks of the Saint John River where Common Milkweed is locally abundant (Klymko pers. comm. 2016; Maritimes Butterfly Atlas 2016). Monarchs breed on PE on patches of Swamp Milkweed, which is native, and on Common Milkweed, which has been introduced in this province.
Canadian and United States Monarchs share overwintering areas but the relative proportions from each country are unknown. Fewer than 500,000 Monarchs currently overwinter in California (NatureServe 2015), and only a tiny percentage of these breed in Canada. In some years, the western Canadian breeding population (in British Columbia) is so small as to be undetectable (St. John pers. comm. 2016).
The total overwintering population size in Mexico was estimated at 66 million in 2014 – 2015 and 200 million in 2015 – 2016 based on extrapolation from the combined area of overwintering colonies and an average density of 50 million Monarchs per hectare. A storm in March 2016 killed a large but unknown number of Monarchs in Mexico after the population survey had been completed (Monarch Watch 2016).
Fluctuations and trends
The most rigorous population monitoring data are from overwintering sites in California and Mexico. These data show that since the late 1990s, Monarch overwintering populations have declined in both these areas (Figures 7 and 8).
Numbers of western Monarchs overwintering in California trended downward from the late 1990s until 2002 but showed no clear trend between 2002 and 2015, fluctuating from a low of 58,468 in 2009 to a maximum of 271,924 in 2015 (NatureServe 2015; Monroe et al. 2016). Between 1997 and 2015, the population declined by over 80% (Figure 6) (NatureServe 2015; Semmens et al. 2016). In the most recent ten-year period (2006 and 2015) western Monarchs increased by 2.1%/year. Because of the very small portion of Canadian individuals that overwinter in California relative to those that overwinter in Mexico, the relevant trend in this assessment is solely that of the Mexican overwintering population.
The overwintering population in Mexico, as measured by area (ha) of occupied habitat, declined significantly over the period 1994-2015 (Figure 7, Monarch Watch 2016). Using this data, a log-linear regression of the time series indicates an 83% decline. The 2012-2014 estimates were the lowest in the time series. The area of occupied habitat in 2015 (4 ha) was higher than the previous three years but below the time series average of 6 ha. The impacts of a severe storm on Monarch populations in March 2016 (after the 2015-2016 count was completed) have yet to be determined (Cave 2016).
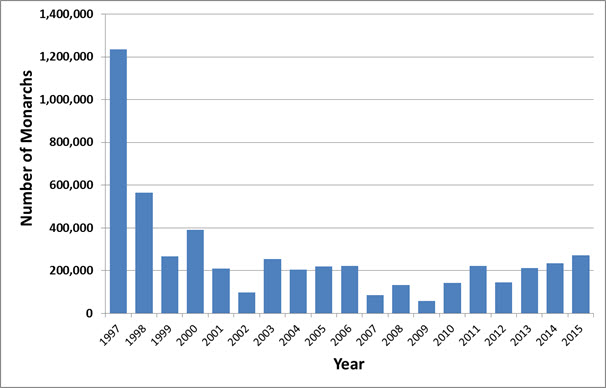
Long description for Figure 6
Chart illustrating estimated abundance of Monarchs at overwintering sites in California from 1997 to 2015.
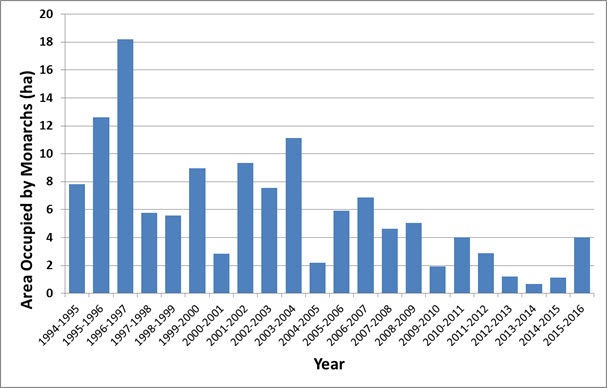
Long description for Figure 7
Chart illustrating total area occupied by Monarch colonies at overwintering sites in Mexico from 1994-1995 to 2015-2016.
The decline rate of the occupied habitat at the overwintering sites in Mexico, over a 10-year period (Figure 7, Monarch Watch 2016) was calculated using the slope of a log-linear regression of the entire time series applied to a period of 10 years. This is consistent with IUCN guidelines on applying the A criterion for time series with wide fluctuations. The estimated slope was -0.089 (p>0.0001) and the 10-year change was estimated to be -59% (Figure 8).
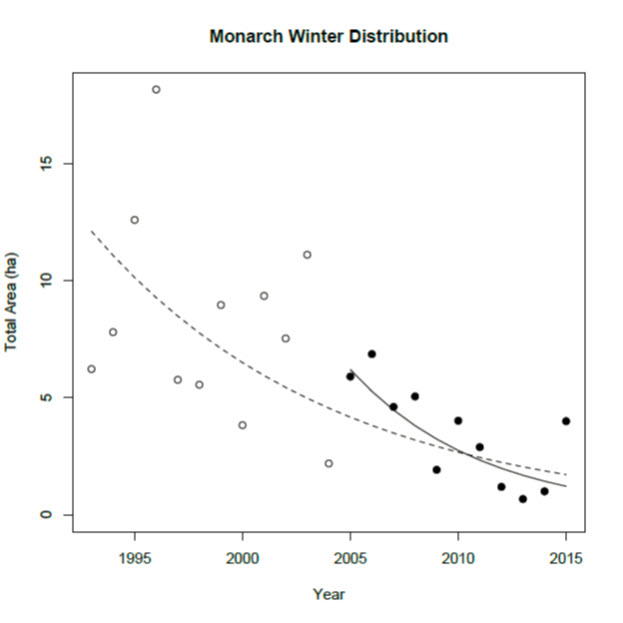
Long description for Figure 8
Chart illustrating total area occupied by Monarchs at the overwintering sites in Mexico, with fitted regression lines that show the decline over time.
Although overwintering data in Mexico indicate a significant downward trend, population trend data as a whole for eastern Monarchs do not align when different migratory stages are compared (see Ries et al. 2015a,b; Inamine et al. 2016; Pleasants et al. 2016). Indices of breeding population size, as estimated through the North American Butterfly Association Seasonal Count Program and the Illinois Butterfly Monitoring Network, showed high year-to-year variability but no statistically detectable trend between 1993 and 2013, during a period when overwintering numbers in Mexico were declining (Reis et al. 2015a,b). The drivers behind population declines are difficult to differentiate and whether this occurs during the summer breeding (milkweed limitation hypothesis) or other cumulative threats during seasonal migration. Possible reasons for the differing results include biased sampling that missed declines in agricultural regions (Pleasants et al. 2016), significant year-to-year variability making it difficult to detect trends, or mortality during fall migration.
Migration monitoring does not consistently show population declines. A long-term (1995 – 2014) migration monitoring study at Long Point, ON, is one of the few available trend studies from Canada (Crewe and McCracken 2015). Monarch abundance counts declined an average of 5.11% per year, when numbers were adjusted for weather effects. Numbers were highly variable between years and showed different trajectories between two monitoring sites. Lower and less variable counts were seen between 2010 and 2014 (Crewe and McCracken 2015). Similar studies at Cape May, New Jersey and Peninsula Point, Michigan station showed no evidence of decline (Davis 2012, Badgett and Davis 2015).
There are two recent publications that outline Monarch population models and estimate extinction risk and population declines. The first by Flockhart et al. (2014) predicts a >5% risk of quasi-extinction over 100 years. The study used a spatially structured stochastic and density-dependent periodic projection matrix model to estimate the probability of quasi-extinction, for which the threshold for quasi-extinction was set at <1000 individuals. Along with the quasi-extinction probability, the paper projects a 14% population decline over 100 years. The use of a quasi-extinction threshold of 1000 individuals has been criticized as being too optimistic (Semmens et al. 2016). Based on an assumed density of 40 million monarchs/ha in overwintering habitat, 1000 monarchs would occupy just 0.25 square metres of forest, an area insufficient for population functions to remain intact at such a reduced abundance for a species that clusters in winter for thermal regulation, that needs to find mates during northward migration across the entire eastern part of North America, and is susceptible to extreme weather (Semmens et al. 2016).
The second quasi-extinction model is detailed in a recent paper by Semmens et al. (2016) and is applicable to the eastern Monarchs only. The model predicts the probability of quasi-extinction to be 11-57% over 20 years using a starting population that corresponds to the winter 2014/2015 estimate of 1.13ha, and a range of quasi-extinction thresholds from 0.01-0.25 ha. The model was fit using occupied overwintering hectares of forest across all years there was data (1993–2014) and total annual egg production in the Midwest (1999–2014), a population growth rate (estimated at 0.94), as well as a yearly deviation. The model assumes a normal distribution and a standard deviation termed process noise (or population stochasticity due to biological and environmental variability) and that the noise in the time series is greater than the measurement error associated with the Mexican overwintering data.
The assumptions made for the Midwestern egg production estimates are similar to those for the overwintering portion of the model. The egg production per year was based on the average estimated eggs per milkweed stem for that year multiplied by the number of available milkweed stems on the landscape in that year. The data was indexed from citizen scientist observations of eggs, and the density of milkweeds in different habitats throughout the landscape based on survey data from the USDA (Semmens et al. 2016).
Rescue effect
Rescue effect may possible. However, Monarchs that dispersed naturally to Europe and elsewhere within the last 200 years did not retain the migratory behaviour required for rescue of Canadian populations (Fernandez-Haeger et al. 2015). Given the strong flight capability of Monarchs, it is possible wayward individuals find their way back and forth across the oceans and recolonize North America.
Threats and limiting factors
The International Union for the Conservation of Nature - Conservation Measures Partnership (IUCN- CMP) threats calculator (see Salafsky et al. 2008; Master et al. 2009) was used to classify and list threats to the Monarch (Table 1). Threats are listed sequentially according to Table 1. The threats discussed below apply to the Monarch throughout its entire range, including the overwintering sites and migratory pathway through the United States. Threats are discussed together for both the western and eastern Monarchs; however, in some instances threats are different for these geographic areas and are explained accordingly. Threats applicable to the Canadian range are explicitly stated. The overall threat impact for the Monarch was assessed at Very High.
Table 1. The International Union for the Conservation of Nature - Conservation Measures Partnership (IUCN- CMP) threats calculator (see Salafsky et al. 2008; Master et al. 2009) was used to classify and list threats to the Monarch. Both the eastern and western migratory routes were included in the assessment. The results of the threats calculator show an overall threat impact of Very High.
- Species English Name
- Monarch
- Species or ecosystem scientific name:
- Danaus plexippus
- Element ID
- -
- Elcode
- -
- Date:
- 2016-01-12
- Assessor(s):
- Jenny Heron (Arthropods SSC Co-chair and moderator), Al Harris (co-author), Paul Grant (Arthropods SSC Co-chair), Sheila Colla (Arthropods SSC member), Jessica Linton (Arthropods SSC member), John Klymko (Arthropods SSC member), Dave McCorquodale (Arthropods SSC Member), Remi Hebert (Arthropods SSC members), Leah Deforest (Parks Canada Agency), Medea Curteanu (Environment and Climate Change Canada), Nathalie Desrosiers (COSEWIC member for Quebec) and Angèle Cyr (COSEWIC Secretariat and recorder). Further input was provided by Ruben Boles (Environment and Climate Change Canada), Dave Fraser (BC Ministry of Environment), Jon McCraken (Birds SSC Co-chair) and Justina Ray (Terrestrial Mammals SSC Co-chair).
- References:
- -
| Threat impact | Threat impact (descriptions) | Level 1 Threat impact counts: high range |
Level 1 Threat impact counts: low range |
|---|---|---|---|
| A | Very high | 0 | 0 |
| B | High | 1 | 1 |
| C | Medium | 2 | 2 |
| D | Low | 3 | 3 |
| - | Calculated overall threat impact: | Very High | Very High |
| # | Threat | Impact (calculated) |
Scope (next 10 Yrs) |
Severity (10 Yrs or 3 Gen.) |
Timing | Comments |
|---|---|---|---|---|---|---|
| 1 | Residential and commercial development | D - Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | - |
| 1.1 | Housing and urban areas | D - Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Applicable to small areas of habitat in both the eastern and western Monarch migratory habitats. |
| 1.2 | Commercial and industrial areas | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Applicable to small areas of habitat in both the eastern and western Monarch migratory habitats. |
| 1.3 | Tourism and recreation areas | Negligible | Negligible (<1%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | Considered negligible at the overwintering sites in California (western Monarch); higher potential for infrastructure development at overwintering sites in Mexico (eastern Monarch). |
| 2 | Agriculture and aquaculture | C - Medium | Restricted (11-30%) | Serious (31-70%) | High (Continuing) | - |
| 2.1 | Annual and perennial non-timber crops | D - Medium | Restricted (11-30%) | Serious (31-70%) | High (Continuing) | Applicable to both eastern and western Monarchs. |
| 2.2 | Wood and pulp plantations | - | - | - | - | Not applicable. |
| 2.3 | Livestock farming and ranching | Not a Threat | Negligible (<1%) | Neutral or Potential Benefit | High (Continuing) | Applicable to both eastern and western Monarchs, not as consumption of food plants but as an impact to habitat. Considered a potential benefit. |
| 2.4 | Marine and freshwater aquaculture | - | - | - | - | Not applicable. |
| 3 | Energy production and mining | Negligible | Negligible (<1%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | - |
| 3.1 | Oil and gas drilling | Negligible | Negligible (<1%) | Negligible (<1%) | Moderate (Possibly in the short term, < 10 yrs) | Not considered a high threat. There is oil and gas development, but this development would probably open up habitats and milkweeds would grow in these areas. |
| 3.2 | Mining and quarrying | - | - | - | - | Not applicable. |
| 3.3 | Renewable energy | Negligible | Negligible (<1%) | Unknown | High (Continuing) | Applicable but considered negligible. More applicable to eastern Monarchs. |
| 4 | Transportation and service corridors | D - Low | Small (1-10%) | Extreme (71-100%) | High (Continuing) | - |
| 4.1 | Roads and railroads | D - Low | Small (1-10%) | Extreme (71-100%) | High (Continuing) | Applicable. Road mortality is the threat, as well as roadside maintenance activities that included pesticide application. |
| 4.2 | Utility and service lines | Not a Threat | Small (1-10%) | Neutral or Potential Benefit | Moderate (Possibly in the short term, < 10 yrs) | Not considered a threat. The creation of utility and service lines, or their maintenance, would keep habitats open and milkweeds would grow in these areas. |
| 4.3 | Shipping lanes | - | - | - | - | Not applicable. |
| 4.4 | Flight paths | - | - | - | - | Not applicable. |
| 5 | Biological resource use | C - Medium | Large (31-70%) | Moderate (11-30%) | High (Continuing) | - |
| 5.1 | Hunting and collecting terrestrial animals | Negligible | Negligible (<1%) | Unknown | High (Continuing) | Applicable. |
| 5.2 | Gathering terrestrial plants | Not a Threat | Negligible (<1%) | Neutral or Potential Benefit | High (Continuing) | Not considered a threat. |
| 5.3 | Logging and wood harvesting | C - Medium | Large (31-70%) | Moderate (11-30%) | High (Continuing) | Applicable to overwintering sites in Mexico (eastern Monarchs). |
| 5.4 | Fishing and harvesting aquatic resources | - | - | - | - | Not applicable. |
| 6 | Human intrusions and disturbance | Negligible | Negligible (<1%) | Unknown | High (Continuing) | - |
| 6.1 | Recreational activities | Negligible | Negligible (<1%) | Unknown | High (Continuing) | Applicable. |
| 6.2 | War, civil unrest and military exercises | - | - | - | - | Not applicable. |
| 6.3 | Work and other activities | - | - | - | - | Not applicable. |
| 7 | Natural system modifications | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | - |
| 7.1 | Fire and fire suppression | Negligible | Negligible (<1%) | Negligible (<1%) | High (Continuing) | Applicable. The threat of wildfire at the overwintering sites (both eastern and western) is possible, although the overwintering sites are spread over a large area and the possibility of fire affecting all sites at once is considered negligible. |
| 7.2 | Dams and water management/use | - | - | - | - | Not applicable. |
| 7.3 | Other ecosystem modifications | - | - | - | - | Not applicable. The threat of conversion of agriculture to soy or corn (more resistant) to be accounted for under pollution. |
| 8 | Invasive and other problematic species and genes | Unknown | Large (31-70%) | Unknown | High (Continuing) | - |
| 8.1 | Invasive non-native/alien species | Unknown | Large (31-70%) | Unknown | High (Continuing) | Applicable. |
| 8.2 | Problematic native species | Unknown | Unknown | Unknown | High (Continuing) | Applicable. |
| 8.3 | Introduced genetic material | Unknown | Unknown | Unknown | High (Continuing) | Applicable. Diseases introduced from captive rearing. |
| 9 | Pollution | B - High | Large (31-70%) | Extreme - Serious (31-100%) | High (Continuing) | - |
| 9.1 | Household sewage and urban waste water | - | - | - | - | Not applicable. |
| 9.2 | Industrial and military effluents | - | - | - | - | Not applicable. |
| 9.3 | Agricultural and forestry effluents | B - High | Large (31-70%) | Extreme - Serious (31-100%) | High (Continuing) | Applicable. |
| 9.4 | Garbage and solid waste | - | - | - | - | Not applicable. |
| 9.5 | Air-borne pollutants | Unknown | Large (31-70%) | Unknown | High (Continuing) | Applicable throughout the species migratory range and overwintering sites (both eastern and western Monarchs). |
| 9.6 | Excess energy | - | - | - | - | Not applicable. |
| 10 | Geological events | - | - | - | - | - |
| 10.1 | Volcanoes | - | - | - | - | Not applicable in Canadian range. |
| 10.2 | Earthquakes/ tsunamis | - | - | - | - | Not applicable in Canadian range. |
| 10.3 | Avalanches/landslides | - | - | - | - | Not applicable in Canadian range. |
| 11 | Climate change and severe weather | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | - |
| 11.1 | Habitat shifting and alteration | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Applicable. Difficult to assign an impact, but habitat shifting and alteration will likely affect 1% of the habitat at the overwintering sites. |
| 11.2 | Droughts | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Not applicable to milkweeds. Applicable to forest habitats at overwintering sites. Difficult to assign an impact, but droughts that this will likely affect 1% of the habitat at the overwintering sites. |
| 11.3 | Temperature extremes | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Applicable at overwintering sites. Difficult to assign an impact, but frost and temperature extremes will likely affect 1% of the population at the overwintering sites. |
| 11.4 | Storms and flooding | Low | Small (1-10%) | Slight (1-10%) | High (Continuing) | Applicable. Difficult to assign an impact, but storms and flooding will likely affect 1% of the habitat at the overwintering sites. |
IUCN 1. Residential and commercial development (low impact)
1.1 Housing and urban areas (low impact) and 1.2 commercial and industrial areas (negligible impact)
Development within Canada:
Residential and commercial development is applicable in central Canada, predominantly within southern ON and applicable to eastern Monarchs. Abandoned farmlands that currently provide suitable breeding and nectaring habitat for the eastern Monarchs are at risk of residential or commercial development, (Environment Canada 2014). About 2,500 km2 of cropland and grassland in southern Canada were converted to built-up areas between 2000 and 2011 (see Habitat Trends) (Agriculture and Agri-Food Canada 2012). Much of the development pressure is near population centres in southern ON and QC, where the highest numbers of breeding Monarchs also occur.
Eastward and westward this threat is considered negligible. In BC (western Monarchs), human population growth and increasing land development threaten Monarch habitat in the Okanagan Valley. The human population in this valley has tripled every 30 to 40 years since 1940 (Jensen and Epp 2002) and is projected to experience significant growth over the next two decades (Hobson and Associates 2006). Yet despite this development, milkweed plants are abundant, widespread and common throughout the area (Dyer pers. comm. 2016; St. John pers. comm. 2016).
Development within the overwintering sites:
Real estate development along the California coast is a threat to the western Monarch overwintering sites. Since the 1980s, at least 30 overwintering sites in California have been lost to housing and commercial developments (Jepsen et al. 2015). Activities associated with residential use, such as tree trimming and removal, can alter the microclimate of an overwintering site and make it less suitable for Monarchs (Jepsen et al. 2015).
Within Mexico (eastern Monarchs), the overwintering sites are mostly clustered in a biosphere reserve and are protected. This habitat and the adjacent non-protected habitat are not considered at high risk from residential and commercial development.
Overall, the threat impact from 1.1 Housing and urban areas and 1.2 Commercial and industrial areas is considered low.
1.3 Tourism and recreational development (negligible impact)
The threat of recreational and tourism development to Monarch habitat in Canada is considered negligible across the species’ range.
The western Monarch overwintering sites in California are not known to have new infrastructure development; most sites are interspersed between residential areas and are in public parks. These areas are still visited by people; however, the development of new trails or infrastructure is low. The eastern Monarch overwintering habitats may be at risk of tourism and recreational development. Hundreds of thousands of people visit these overwintering sites each year, and the need for facilities may increase in the future.
IUCN 2. Agriculture and aquaculture (medium impact)
2.1 Annual and perennial non-timber crops (medium impact)
The abundance of milkweed throughout both Canada and the United States is declining from the conversion of farmland or agricultural areas into larger and more intensive industrial agricultural use. Conversely, within some parts of Canada, abandoned farmland is slowly converting to forest through natural succession. This overall change in agricultural habitats is leading to a decline in the abundance and distribution of milkweeds throughout the species’ breeding and migratory range.
Abandoned farmlands that currently provide suitable breeding and nectaring habitat for the eastern Monarchs are at risk of conversion to more intensive agriculture (Environment Canada 2014). In addition, the conversion of crops with milkweed growing throughout the crop (e.g., as a weed that is not controlled) within, for example, corn and soybean crops, are being converted to more intensive agriculture that is not conducive to the growth of these weedy milkweed patches (Pleasants 2017).
Data on milkweed population trends in Canada are unavailable. However in the United States, loss of milkweed is one of the most important factors in the decline of eastern Monarchs. Since the late 1990s, Common Milkweed has undergone a massive decline in the Midwest United States (Jepsen et al. 2015; Pleasants 2017).
A study in Iowa (within the eastern Monarch range) found a 64% decline in milkweed in agricultural areas between 1999 and 2010 (data presented in Pleasants and Oberhauser 2012 were corrected by Pleasants 2015). Eastern Monarch populations declined by 88% in this study area during this same period (Pleasants 2015).
Population modelling by Flockhart et al. (2014) for eastern Monarchs predicts further declines of 14% within 100 years and concludes that recent Monarch declines were caused by reduction in milkweed plants in the United States rather than from climate change or degradation of forest habitats in Mexico. The impact on western Monarchs may be less since fewer breed in soybean, corn and canola fields. Conversely, a recent paper by Inamine et al. (2016) tested this milkweed limitation hypothesis by analyzing citizen science records from four monitoring programs across North America. Results suggest no statistically significant temporal trends and that lack of milkweed availability is unlikely to be driving Monarch population declines.
Further analysis by Pleasants (2017) estimates that from 1999 – 2014 850 million milkweed stems were lost in the Midwest due to herbicide use in corn and soybean fields; and since 2008 an additional 11 million stems lost due to the conversion of grasslands to cropland. The result is an estimated 71% loss in support capacity for migrating Monarchs, based on milkweed stems in agricultural fields averaging 3.9 times more Monarch eggs than those in non-agricultural habitats (Pleasants 2017). Overall, there has been an estimated 40% decline in milkweed stems between 1999 – 2014 (Pleasants 2017).
Canadian croplands, particularly in the prairies and southern ON, have undergone similar changes in agricultural practices although there is no current data or analysis available.
Common Milkweed is considered to be a noxious weed in the Weed Control Acts of Manitoba (Province of Manitoba 2010), Quebec (Schappert 2004) and Nova Scotia (Province of Nova Scotia 2016) and until recently (2014) was listed as a noxious weed in Ontario but has since been removed. In Manitoba, Showy and Swamp milkweeds are also listed as noxious weeds.
IUCN 3. Energy production and mining (negligible impact)
3.3 Renewable energy (negligible impact)
Energy production and mining are unlikely to influence Monarch populations. Collisions with wind turbines have been identified as a potential threat for migrating Monarchs in the previous COSEWIC status report, but have not been documented in Canada and the magnitude of this threat is unknown. A review of all publically available monitoring reports for ON wind farms along the Lake Ontario and Lake Erie shorelines in 2013 found no records of Monarch or other butterfly mortalities (Linton pers. data. 2016).
Some solar farms can cause Monarch mortality. Some solar flux facilities use mirrors to reflect sunlight to a boiler situated on a tower. Large numbers of dead Monarchs were observed at such a solar flux facility in California (Kagan et al. 2014). Monarchs were apparently killed as they flew into the concentrated light near the boiler. Solar flux technology is not presently used in Canada (Natural Resources Canada 2016).
IUCN 4. Transportation and service corridors (low)
4.1 Roads and railroads (low impact)
Roadsides as breeding sites:
Roadsides are important habitat for breeding Monarchs, as roadside verges are periodically disturbed thus allowing milkweeds to grow. These areas are increasingly important as milkweeds in agricultural fields are declining throughout the United States. Road rights-of-way support about 10% of milkweeds in North America (Flockhart et al. 2014) and are estimated to produce 35% of Monarchs in the Midwest United States (unpublished data cited by The Center for Biological Diversity et al. 2014). Threats in roadside habitats include adult mortality from vehicle collisions, loss of food plants from excessive mowing and herbicides, and exposure to road salt.
Road mortality to Monarchs:
Monarchs are vulnerable to mortality from vehicle collisions, particularly in the summer range. A study in central Illinois estimated 500,000 Monarchs were killed annually in Illinois (McKenna et al. 2001). At least 7% of butterflies (all species combined) inhabiting roadsides in a study in Poland were killed by vehicle collisions. Monarch road kills increased with increased traffic volume, road width, and mowing (Skórkaa et al. 2013).
Road maintenance activities:
Excessive mowing and use of herbicides on roadsides could kill caterpillars and remove milkweeds and nectaring plants (Jepsen et al. 2015). However, some mowing may be important to maintain milkweed on roadsides. Green Milkweed (Asclepias viridis) in Oklahoma regrew after mowing of roadsides but in the absence of mowing, died back in late summer and was unavailable for Monarchs (Baum and Mueller 2015).
Milkweeds exposed to road salt runoff have higher sodium content than milkweeds grown away from roads (Snell-Rooda et al. 2014). Monarch caterpillars raised on roadside milkweeds had lower survival rates. Among the survivors, males had increased muscle mass and females had greater neural investment (Snell-Rooda et al. 2014). The implications for Monarch fitness are unknown.
IUCN 5. Biological resource use (high impact)
5.1 Hunting and collecting terrestrial animals (low impact)
Monarchs are collected at winter roosts in California to augment breeding stock for the commercial Monarch production industry, but the scale of the practice is unknown (Jepsen and Black 2015).
In Canada, there are incidental collection of eggs and caterpillars for rearing in southern ON and the Maritimes, although the number of specimens collected is unknown. There are few restrictions to the importation or movement of Monarchs from one geographic location to another in Canada. See Threat 6.1 for threats associated with parasites and protozoans; and Threat 8.3 for threats associated with the introduction of genetic material.
5.3 Logging and wood harvesting (high impact)
Forest loss and degradation at the overwintering sites in Mexico was considered a higher threat in the past than at present; although this threat continues. A total of 4300 ha (8%) of Oyamel Fir forest within the Monarch Butterfly Biosphere Reserve was lost or disturbed between 1986 and 2012, largely to logging (Ramirez et al. 2015). Even small-scale logging is a potential threat because openings and thinned areas in the forest expose overwintering Monarchs to winter storms, cold temperatures and wet conditions, potentially causing mortality (Jepsen et al. 2015; Williams and Brower 2015). Logging has taken place in this area since the late nineteenth century and included both commercial harvest and domestic use for building material, firewood, and charcoal (Ramirez et al. 2015). Although logging is prohibited in the core of the reserve, illegal logging is ongoing (Jepsen et al. 2015). In 2015, loggers illegally cut 10 ha of forest on Sierra Chincua, only 600 m from where Monarchs were roosting (Davis and Linton 2016).
IUCN 6. Human intrusions and disturbance (negligible impact)
6.1 Recreational activities (negligible impact)
Unsustainable ecotourism is a potential threat to overwintering Monarchs in Mexico (Commission for Environmental Cooperation 2008, The Center for Biological Diversity et al. 2014). The effects have not been quantified but some researchers suspect that disturbance and habitat degradation may be harming Monarchs. The overwintering sites in Mexico have an estimated 100,000 to 150,000 annual visitors (Vidal et al. 2014). Disturbance of overwintering Monarchs in California was not listed as a threat in a review of the conservation status of Monarchs in the United States (Jepsen et al. 2015).
IUCN 7. Natural system modifications (negligible)
7.1 Fire and fire suppression (negligible)
Fire in the overwintering range of Monarchs in both California and Mexico is a potential threat. At the overwintering sites in Mexico, 45 fires occurred in the Monarch Butterfly Biosphere Reserve in 2012, covering 176.6 ha or 0.31% of the reserve (Martinez-Torres et al. 2015). Most were caused by agricultural activities, campfires, or forestry activities, with only one originating from a lightning strike (Martinez-Torres et al. 2015). The historical forest regime of the Oyamel Fir, the impacts of fire suppression, and effectiveness of reforestation efforts remain poorly understood.
At the overwintering sites in California, non-native eucalyptus trees provide the majority of Monarch winter roost trees (Frey and Schaffner 2004). Programs to remove eucalyptus and restore native tree cover and reduce fire risk may inadvertently threaten Monarchs. Eucalyptus provides wind cover and their leaf and branch structure are suitable for clustering, but native tree species such as Monterey Pine and Monterey Cypress are preferred where available (Jepsen et al. 2015). Habitat degradation also results from aging and die-back of Monterey Pines and other trees in the overwintering range of western Monarchs (Jepsen et al. 2015).
IUCN 8. Invasive and other problematic species and genes (unknown impact)
8.1 Invasive / non-native alien species (unknown impact)
Dog-strangling Vine (Cynanchum vincetoxicum) and Black Swallowwort (C. louiseae or V. nigrum) are non-native plants in the milkweed family, spreading throughout central and southern ON and southern QC (Ontario’s Invading Species Awareness Program 2015). Monarchs are known to oviposit on these non-native plants, but caterpillars have significantly lower survival than those on native milkweeds and often do not develop past the first instar (DiTommaso and Losey 2003; Mattila and Otis 2003; Casagrande and Dacey 2007). However, there is a strong preference for native milkweeds over these non-native plants.
Tropical Milkweed (Asclepias curassavica), is rapidly spreading in the Monarch’s range from Texas to Florida (Batalden and Oberhauser 2015). During the fall migration south, Monarchs will preferentially oviposit on Tropical Milkweed over native milkweeds, possibly because native species are beginning to senesce and Tropical Milkweed is typically in good condition. Most females are in reproductive diapause during the fall migration although the option to break reproductive diapause when these plants are available could have adverse consequences to the natural life cycle of the species (Batalden and Oberhauser 2015). These effects have not yet been demonstrated but are considered potential threats to the species (Batalden and Oberhauser 2015). In addition, higher levels of the parasite Ophyrocystis elektroscirrha are found in non-migratory Monarchs feeding on Tropical Milkweed (Satterfield et al. 2015).
The release of commercially bred Monarchs (see Threat 8.3) has the potential to transmit disease and parasites (The Center for Biological Diversity et al. 2014, North American Butterfly Association 2014). Captive-raised Monarchs can be heavily infested with the protozoan parasite Ophyrocystis elektroscirrha, the bacteria Serratia, the fungus Nosema, and cytoplasmic viruses (The Center for Biological Diversity et al. 2014). Transmission of these diseases from captive-raised to wild Monarchs is possible but has not been demonstrated.
8.2 Problematic native species (unknown impact)
Abandoned farmlands that currently provide suitable breeding and nectaring habitat for the eastern Monarchs are at risk of being lost, as they either regenerate into forest or are developed further to more intensive agricultural farmland (see Habitat trends).
8.3 Introduced genetic material (unknown)
The release of commercially bred Monarchs at weddings and other events has been identified as a potential threat to wild Monarch populations through the introduction of undesirable genetic traits (The Center for Biological Diversity et al. 2014; North American Butterfly Association 2014). Millions of Monarchs are released each year in the United States (The Center for Biological Diversity et al. 2014), but the scale of the practice is unknown in Canada. The level of genetic diversity in captive Monarchs is unknown but mixing with wild populations could cause inbreeding depression and introduction of deleterious adaptations and potentially reduced survivorship of wild Monarchs (The Center for Biological Diversity et al. 2014) and impede the scientific study of Monarch movement (Pyle 2015).
IUCN 9. Pollution (high impact)
9.3 Agricultural and forestry effluents (high impact)
Milkweed decline is strongly correlated with the use of herbicide-resistant crops. Although glyphosate has been in use since the 1970s, herbicide-resistant crops have enabled farmers to apply glyphosate more frequently, at higher rates, and later in the growing season than with conventional crops (Center for Food Safety 2015). Common Milkweed is tolerant of the use of glyphosate as a pre-emergent treatment on corn and soybean fields. Milkweeds grow from perennial roots and are not significantly affected by pre-emergent herbicides but are killed or defoliated by glyphosate applied later in the season (Hartzler 2010).
Herbicide-resistant crop use in Canada has increased substantially over the past twenty years. Herbicide-resistant canola was first used in Canada in 1995, followed by soybeans in 1997, corn in 1998, and wheat in 2004 (Beckie et al. 2006). By 2005, about 95% of canola crop area and 60% of soybean crop area in Canada were glyphosate-resistant (Beckie et al. 2006). Adoption of herbicide-resistant corn was slower than soybeans and totalled only 17% to 18% of the total crop area by 2005. More recent Canadian data are unavailable but herbicide-resistant crop use probably followed trends similar to those in the United States.
Almost all corn and soybeans (91%) in the United States are sprayed with glyphosate. The area of these crops treated with glyphosate increased nine-fold between 1995 and 2013 (Center for Food Safety 2015). In these 18 years, the application rate of glyphosate increased by 58% on soybeans and 43% on corn; and application frequency increased by 64% on soybeans and 16% on corn. In total, glyphosate use on corn and soybeans increased more than 20-fold, from 4.5 million to 93 million kg/year (Center for Food Safety 2015).
Increased herbicide use may also reduce populations of flowers that provide nectar throughout Monarch migration routes. Monarchs increase their feeding rates while migrating south, particularly as they reach Texas and northern Mexico. Nectar is converted to lipids and used as an energy source by overwintering Monarchs (Brower et al. 2006). Declining nectar supplies have been proposed as a possible contributing factor in overwinter declines (e.g. Brower et al. 2006, Inamine et al. 2016).
Neonicotinoid insecticides were introduced in the 1990s and include imidacloprid, clothianidin, thiamethoxam, acetamiprid, thiacloprid, and dinotefuran. Neonicotinoids are approved for use as seed treatments, soil applications, and foliar sprays on oilseeds, grains, pulse crops, fruits, vegetables, greenhouse crops, ornamental plants, and Christmas trees in Canada (Health Canada 2016). In Ontario, nearly 100% of corn seed and roughly 60% of soybean seed are treated with neonicotinoids (Ontario Ministry of Agriculture, Food and Rural Affairs 2014). The use of neonicotinoid insecticides has increased substantially in recent years and has been implicated in declines of non-target species (Douglas et al. 2014; Jepsen et al. 2015).
Neonicotinoids persist in soil and are translocated to plant tissues to levels potentially lethal to non-target insects (Goulson 2013). Monarch caterpillars were fed milkweed leaves with levels of clothianidin similar to milkweeds exposed to spray from a treated field. Exposed caterpillars grew more slowly than unexposed caterpillars (Pecenka and Lundgren 2015). A neonicotinoid insecticide (imidacloprid) applied to soil was translocated to milkweed tissues, causing reduced survival of Monarch caterpillars feeding on the leaves, but not nectaring adults (Krischik et al. 2015). Sub-lethal impacts of neonicotinoids on bees and other invertebrates include altered foraging behaviour, reduced reproduction, and greater susceptibility to pathogens (Goulson 2013, van der Sluijs et al. 2013).
Monarchs in western Canada have probably been less affected by herbicides than those in eastern Canada although insecticides are used in the vineyards and fruit orchards in the Okanagan Valley (BC). Both of these agricultural systems use large amounts of pesticides but the effects on Monarchs are unknown.
9.5 Airborne pollutants (unknown impact)
During agricultural crop management practices, the soil is tilled before sowing a field and significant dust can form and dissipate into the airspace. Depending on the weather (e.g., dry or windy conditions), this dust can blow long distances and/or linger in the area for a few days. The residual pesticides mixed in the soil become airborne with the dust, and eventually settle on plants, including milkweeds within adjacent areas. For example, Bt corn is a genetically modified crop that is an indirect threat to Monarchs. Bt refers to the bacterium Bacillus thuringiensis, from which a gene that produces a protein toxic to Lepidoptera caterpillars. This gene has been genetically incorporated into corn making it less palatable to Lepidoptera caterpillar, and reducing the need for pesticide application on the crop. Laboratory studies have showed that Bt corn pollen blown onto milkweed leaves which are then consumed by Monarch caterpillar caused reduced growth and survival (Losey et al. 1999). Conversely, subsequent field studies found that concentrations of pollen were unlikely to significantly harm caterpillar (Pleasants 2015).
IUCN 11. Climate change and severe weather (low impact)
The impacts of climate change on Monarch breeding dynamics are complex and require additional study (Zipkin et al. 2012). Climate change could act on Monarchs through a number of mechanisms including severe weather (e.g., early or late season frost at overwintering sites and throughout migratory range), heat stress and loss of milkweeds in the breeding range, altered timing of migration, and loss of overwintering habitat due to habitat shifting. Modelling of future climate scenarios suggests that climate change will have an effect on overwintering Monarchs in Mexico. It is difficult to assign an impact; however, climate change will likely affect 1% of the population at the overwintering sites.
11.1 Habitat shifting and alteration (low impact)
Asynchrony between milkweed development and Monarch migration is a potential threat (Robinson et al. 2009). Butterflies exhibited a stronger response to climate shifts than their food plants, which could result in Monarchs starting northward migration before milkweeds are available for caterpillar food (Parmesan 2007). One climate change model predicts that Monarch-preferred breeding temperature and precipitation will move northward within the next 50 years (Batalden et al. 2007). Monarchs and milkweeds will need to expand their range northward under this scenario (Batalden et al. 2007). In contrast, another model suggests that climate change during the breeding season is unlikely to cause eastern Monarch population declines (Zalucki et al. 2015).
Changing climate could also alter habitat in the overwintering habitats. As the climate becomes warmer, the area of suitable habitat for Oyamel Fir is predicted to decline by 50% by 2030 and completely disappear by 2090 as tree mortality increases due to pests and disease (Ramirez et al. 2015). Climate change may also have an impact on the overwintering habitat of western Monarchs through increased incidence of wildfires (Commission for Environmental Cooperation 2008).
11.2 Droughts (low impact)
Climate models predict decreasing soil moisture in large parts of the breeding range, potentially reducing breeding success (Stevens and Frey 2010). Yet milkweed plants are considered drought hardy and grow in areas where moisture is low.
Increased drought stress associated with climate change may make Oyamel Fir more vulnerable to diseases and insect pest including bark beetles (Scolytus spp.) (Ramirez et al. 2015).
11.3 Temperature extremes (low impact)
Caterpillar exposure to continuous high temperatures (36°C) resulted in higher mortality, longer development time, and lighter adult mass (York and Oberhauser 2002). However, temperatures fluctuating between 27°C and 36°C resulted in very little mortality and shorter development times than controls.
11.4 Storms and flooding (low impact)
Storms at the overwintering sites in both Mexico and California can cause catastrophic mortality to Monarchs during the months when butterflies are roosted at these sites. A storm in January 2002 killed an estimated 500 million overwintering Monarchs or about 75% of the population at the time. Heavy rain and snow caused extreme wetting of butterflies and was followed by temperatures dropping as low as -4°C resulting in hypothermia and mortality (Brower et al. 2004). A rain and sleet storm accompanied by strong winds on March 8 - 9 2016 buried large numbers of Monarchs in sleet (Monarch Watch 2016). Mortality was estimated at 3 - 50% of the overwintering population, although many Monarchs had already left on their northward migration and avoided the storm. One climate model predicts that cool weather precipitation will increase and cause more frequent large scale Monarch mortality events (Oberhauser and Peterson 2003). The scope and severity of winter storms at the overwintering sites is substantial, but the impact on the longer ten-year time frame is hard to estimate.
Limiting factors
Overwintering habitat:
Most of the adult population from eastern North America is concentrated in a small number of hectares in the overwintering grounds in Mexico, and is vulnerable to extreme weather events, fire, diseases and parasites, predation, and anthropogenic threats.
Migration:
Long distance migration exposes Monarchs to extreme weather events and requires a continuous supply of breeding and feeding habitat from Mexico to Canada to complete the migratory cycle. The eastern Canadian breeding population is strongly influenced by breeding success of returning migrants in the southern United States, especially Texas and the central United States (Miller et al. 2012; Ries et al. 2015b). Reduced breeding success caused by drought and other threats in Texas and the central United States can greatly reduce the numbers of breeding Monarchs reaching Canada.
Predation:
The Black-headed Grosbeak and the Black-backed Oriole feed extensively on Monarchs at the overwintering sites in Mexico. Predation has occurred at a rate of up to 34,000 butterflies killed per day (Snook 1993). About 2 million Monarchs (10% of the total colony) were eaten by birds over a single winter (Arellano et al. 1993). Birds feed on the perimeter of the colony and smaller colonies, with a proportionally greater circumference, suffer greater rates of predation, as high as 44% (Calvert et al. 1979). The Black-eared Mouse (Peromyscus melanotis) also feeds extensively on both live and dead Monarchs (Glendinning 1993).
Number of locations
The location concept does not apply to the Monarch. The species’ range is vast, and the threats are complex, varying with the migratory cycle stage, the type of breeding habitat used (agricultural verses natural), and in the case of California overwintering habitat, local land management may also be a factor.
Protection, status and ranks
Legal protection and status
The Monarch was designated as Special Concern by COSEWIC in 1997 and listed as such under the federal Species at Risk Act in 2004. The status was re-examined and confirmed in 2001 and 2010.
Monarchs are listed as Special Concern under the Ontario Endangered Species Act (2007) and the New Brunswick Species at Risk Act (2013).
In Ontario, Monarchs are protected under the Fish and Wildlife Conservation Act.
In Quebec, the Monarch is not currently listed under the: Loi sur les espèces menacées ou vulnérables” (RLRQ, c E-12.01) (LEMV) (Act respecting threatened or vulnerable species) (CQLR, c E-12.01) (MRNF 2011) and is not afforded protection under the “Loi sur la conservation et la mise en valeur de la faune” (RLRQ, c. C- 61.1) (LCMVF) (Act respecting the conservation and development of wildlife) (CQLR, c. C-61.1).
Non-legal status and ranks
Breeding populations of Monarchs (Natureserve 2015):
Global conservation status rank: Apparently Secure (G4).
Canadian conservation status rank: Secure (N5B).
United States conservation status rank: Secure (N5B).
Provincial conservation status ranks:
Manitoba: Secure (S5B)
Ontario: Apparently Secure (S4B)
B.C., AB, NB, SK: Vulnerable (S3B)
Nova Scotia and Newfoundland: Imperiled (S2B)
PEI: Critically Imperiled (S1B).
Quebec: not ranked
In Ontario, the conservation status rank of non-breeding populations is Imperiled (S2N) to reflect the scarcity of significant migratory concentration areas (NHIC 2015).
US state conservation status ranks: Secure (S5) in 27 states, Apparently Secure (S4 or S4S5) in 10 states, Vulnerable (S3) in California, and not ranked in 11 states (NatureServe 2015).
Conservation status rank of US migratory concentration areas: Vulnerable to Imperiled (N2N3N).
The International Union for Conservation of Nature (IUCN) designated Monarch migration as an endangered phenomenon, recognizing that migration is imperiled, although the species is not in danger of extinction (Commission for Environmental Cooperation 2008). This was the first designation of a biological phenomenon, as opposed to a species.
The Monarch Butterfly Biosphere Reserve in Mexico was inscribed as a World Heritage Site in 2008 (UNESCO 2015). Fifteen of the overwintering colonies are in the reserve and seven are outside the reserve (Garcia-Serrano et al. 2004).
In Mexico, Monarchs are listed as “under special protection” in the Species at Risk standard (NOM-059-Semarnat-2001) (Commission for Environmental Cooperation 2008).
In the United States, Monarchs are presently under review for listing under the United States Endangered Species Act in response to a petition from environmental groups (The Center for Biological Diversity et al. 2014).
Habitat protection and ownership
Most Monarch breeding habitat in Canada is on private land, rights-of-way and other unprotected areas.
In Canada, there are migration staging areas in federal protected areas on the shores of lakes Ontario and Erie in southern Ontario including Point Pelee National Park, Long Point National Wildlife Area, and Prince Edward Point National Wildlife Area. These areas were designated as part of a network of protected areas for Monarchs through an international agreement with Mexico and the United States (Commission for Environmental Cooperation 2008). Other staging areas in Ontario are in municipal and provincial parks and private land (Hess 1992).
In Mexico, most overwintering colonies are within the federally protected Monarch Butterfly Biosphere Reserve (MBBR), administered by the National Commission of Natural Protected Areas (Commission for Environmental Cooperation 2008). Smaller colonies outside the MBBR have varying degrees of federal protection (Commission for Environmental Cooperation 2008). The Monarch Butterfly Biosphere Reserve in Mexico was inscribed as a World Heritage Site in 2008, but Illegal logging, growth in human population, tourism pressures, agricultural advances, and forest fires are continuing threats (UNESCO 2015).
In the United States, Monarch overwintering habitat in California is protected under a variety of federal, state, and local laws and policies. However, collecting or disturbing Monarchs and tree removal and trimming in roosts is typically not restricted except on federal and state land (International Environmental Law Project 2012).
Acknowledgements
Thank you to all the people listed in the Authorities Contacted, who provided valuable information, advice, and knowledge. Thank you for information from Phil Schappert, Mar Elderkin, Pail Johanson, Pail and Victoria Leck. Specific thank you to Ruben Boles (Canadian Wildlife Service, Environment and Climate Change Canada), Alan Sinclair (Marine Fish SSC Co-chair), Jessica Linton (Arthropods SSC member), Paul Grant (Arthropods SSC Co-chair), Justina Ray (Terrestrial Mammals SSC Co-chair), Patrick Nantel (Parks Canada), Cory Sheffield (Royal Saskatchewan Museum) and Dave Fraser (BC Ministry of Environment) for valuable insight and careful review of the document. Thank you to additional Arthropod SSC members Colin Jones, Syd Cannings, Sheila Colla, Vincent Bereczky and Cory Sheffield. Thank you to Jennifer Heron (Arthropods SSC Co-chair) for editorial and technical support, Angèle Cyr (COSEWIC Secretariat) for technical support and Jenny Wu (COSEWIC Secretariat) for mapping support. Karine Bériault wrote the previous (2010) status report. The cover photograph is by Jessica Linton.
Authorities contacted
Boles, Ruben. Canadian Wildlife Service, Environment and Climate Change Canada, Gatineau, QC.
Brownell, Vivian. Ontario Ministry of Natural Resources and Forestry Policy Division, Peterborough ON.
Cannings, Syd. Environment and Climate Change Canada, Canadian Wildlife Service, Pacific and Yukon Region, Whitehorse, YT.
Curteanu, Medea. Canadian Wildlife Service, Environment and Climate Change Canada, Edmonton, AB.
Cyr, Angèle. COSEWIC Secretariat, Canadian Wildlife Service, Environment and Climate Change Canada, Gatineau, QC.
Desrosiers, Nathalie. Direction de la biodiversité et des maladies de la faune
Direction générale de l’expertise sur la faune et ses habitats, Ministère des Forêts, de la Faune et des Parcs du Québec, Québec.
Fraser, David. British Columbia Ministry of Environment, Species Conservation Science Unit, Victoria, BC.
Gauthier, Isabelle. Coordonnatrice provinciale des espèces fauniques menacées et vulnérables, Ministère des Forêts, de la Faune et des Parcs, QC.
Gelling, Lea. British Columbia Conservation Data Centre, Victoria, BC.
Heron, Jennifer. British Columbia Ministry of Environment, Species Conservation Science Unit, Vancouver, BC.
Huntley, Matt. Environment and Climate Change Canada, Canadian Wildlife Service, Pacific and Yukon Region, Delta, BC.
Jones, Colin. Ontario Natural Heritage Information Centre, Peterborough, ON.
Jones, Neil. COSEWIC Secretariat, Canadian Wildlife Service, Environment and Climate Change Canada, Gatineau, QC.
Klymko, John. Atlantic Conservation Data Centre, Sackville, NB.
Nantel, Patrick. Parks Canada Agency, Gatineau, QC.
Linton, Jessica. Natural Resources Solutions, Waterloo, ON.
Pardy, Shelley. Department of Environment and Conservation, Endangered Species and Biodiversity, St. John, NF.
Perrault, Julie. COSEWIC Secretariat, Canadian Wildlife Service, Environment and Climate Change Canada, Gatineau, QC.
Schnobb, Sonia. COSEWIC Secretariat, Canadian Wildlife Service, Environment and Climate Change Canada, Gatineau, QC.
Sheffield, Cory. Royal Saskatchewan Museum, Regina, SK.
Stipec, Katrina. British Columbia Conservation Data Centre, Victoria, BC.
Wu, Jenny. COSEWIC Secretariat, Canadian Wildlife Service, Environment and Climate Change Canada, Gatineau, QC.
Information sources
Agriculture and Agri-Food Canada. 2012. 2011 AAFC crop type map of Canada. Web site: ftp://ftp.agr.gc.ca/pub/outgoing/aesb-eos-gg/Crop_Inventory/2011/ (accessed October 9, 2012). Statistics Canada, Environment Accounts and Statistics Division, 2013, special tabulation. [Accessed August 2016].
Altizer, S.M. and K.S. Oberhauser. 1999. Effects of the protozoan parasite Ophryocystis elektroscirrha on the fitness of Monarch butterflies (Danaus plexippus). Journal of Invertebrate Pathology 74(1): 76-83.
Anonymous. 2016. Press release: Monarch butterflies triple the area occupied for their Mexican wintering sites [PDF, 144 Mb]. Web site: [Accessed August 2016].
Antonijevic, E. D. St. John and J. Heron. 2014. Citizen science observations from rearing, tagging and releasing Monarch butterflies (Danaus plexippus) in the South Okanagan, British Columbia, in 2014. Unpublished report. B.C. Ministry of Environment, Vancouver, B.C. 31pp.
Arellano, G., J.I. Glendinning, J.B. Anderson, and L.P. Brower. 1993. Interspecific comparisons of the foraging dynamics of black-backed orioles and black-headed grosbeaks on overwintering monarch butterflies in Mexico. pp. 315-322. In Malcolm, S.B. and M.P. Zalucki (eds.), Biology and conservation of the Monarch butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Badgett, G. and A.K. Davis. 2015. Population trends of Monarchs at a northern monitoring site: analyses of 19 years of fall migration counts at Peninsula Point, MI. Annals of the Entomological Society of America: 1–7.
Batalden, R.V., K. Oberhauser, and A.T. Peterson. 2007. Ecological niches in sequential generations of eastern North American Monarch butterflies (Lepidoptera: Danaidae): The ecology of migration and likely climate change implications. Environmental Entomology. 36:1365–1373.
Batalden, R.V. and K.S. Oberhauser. 2015. Potential changes in eastern North American Monarch migration in response to an introduced milkweed, Asclepias curassvica. Pp. 215-224. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Baum, K.A. and E.K. Mueller 2015. Grassland and roadside management practices affect milkweed abundance and opportunities for Monarch recruitment. Pp. 197-202. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Beckie, H. J., Harker, K. N., Hall, L. M., Warwick, S. I., Légère, A., Sikkema, P. H., Clayton, G. W., Thomas, A. G., Leeson, J. Y., Séguin-Swartz, G. and Simard, M.-J. 2006. A decade of herbicide-resistant crops in Canada. Canadian Journal of Plant Science 86: 1243–1264.
Bird, C.D., Hilchie, G. J., Kondla, N. G., Pike, E. M., & Sperling, F. A. H. 1995. Alberta Butterflies. The Provincial Museum of Alberta, Edmonton, AB. 349 pp.
Borders, B. and E. Lee-Mader. 2014. Milkweeds: A Conservation Practitioners Guide. 146 pp. Portland OR. The Xerces Society for Invertebrate Conservation.
Bradley, C.A. and S. Altizer. 2005. Parasites hinder Monarch butterfly flight: implications for disease spread in migratory hosts. Ecology Letters 8(3): 290-300.
Brindza, L.J., L.P. Brower, A.K. Davis and T. van Hook. 2008. Comparative success of Monarch Butterfly migration to overwintering sites in Mexico from inland and coastal sites in Virginia. Journal of the Lepidopterist’s Society 62:189-200.
Brower, L.P. 1995. Understanding and misunderstanding the migration of the monarch butterfly (Nymphalidae) in North America: 1857-1995 [PDF, 35.1 Mb]. Journal of the Lepidopterists Society 49:304–385. Web site: [Accessed April 14, 2016].
Brower, L.P. 1996. Monarch butterfly orientation: missing pieces of a magnificent puzzle. The Journal of Experimental Biology 199:93-103.
Brower, A.V.Z. and T.M. Boyce. 1991. Mitochondrial DNA variation in monarch butterflies. Evolution 45: 1281 – 1286.
Brower, L.P. and R. M. Pyle. 2004. The interchange of migratory monarchs between Mexico and the western United States, and the importance of floral corridors to the fall and spring migrations. In G.P. Nabhan, ed., Conserving migratory pollinators and nectar corridors in western North America, pp. 144-166. Tuscan University of Arizona Press.
Brower, L.P., G. Castilleja, A.Peralta, J. Lopez-Garcia, L. Bojorquez-Tapia, S. Diaz, D. Melgarejo, and M. Missrie. 2002. Quantitative changes in forest quality in a principal overwintering area of the Monarch Butterfly in Mexico, 1971-1999. Conservation Biology 16(2): 346-359.
Brower L.P., D.R. Kust, E. Rendon-Salinas, E.G. Serrano, K.R. Kust, J. Miller, C. Fernandez del Rey, and K. Pape. 2004. Catastrophic winter storm mortality of Monarch butterflies in Mexico during January 2002. Pp. 151-166 in K.S. Oberhauser and M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Brower, L. P., O. R. Taylor, E. H. Williams, D. A. Slayback, R. R. Zubieta, and M. I. Ramirez. 2012. Decline of monarch butterflies overwintering in Mexico: is the migratory phenomenon at risk? Insect Conservation and Diversity 5: 95–100.
Calvert, W. 2004. Two methods of estimating overwintering monarch population size in Mexico. Pp. 121-128. in K.S. Oberhauser and M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Calvert, W.H., L.E. Hedrick and L.P. Brower. 1979. Mortality of the Monarch butterfly (Danaus plexippus L.): avian predation at five overwintering sites in Mexico. Science 204: 847-851.
Casagrande, R.A. and J.E. Dacey, 2007. Monarch oviposition on swallow-worts (Vincetoxicum spp.). Environmental Entomology 36(3):631-636.
Cave, R. 2016. Monarch butterfly sanctuary in Mexico hit by cold weather. Web site: [Accessed April 11 2016].
Center for Food Safety (CFS). 2015. Monarchs in Peril Herbicide-Resistant Crops and the Decline Of Monarch Butterflies In North America. Web site: http://www.centerforfoodsafety.org/files/cfs-monarch-report_2-4-15_design_05341.pdf. [Accessed November 2015].
Commission for Environmental Cooperation. 2008. North American Monarch Conservation Plan. Web site: [Accessed November 2015].
Commission for Environmental Cooperation. 2009. Monarch Butterfly Monitoring in North America: Overview of Initiatives and Protocols. Web site: http://www.mlmp.org/Resources/pdf/Monarch-Monitoring_en.pdf [Accessed April 13 2016].
COSEWIC. 2017. National ecological areas. Web site: [accessed February 2017].
{kind=link}
Crewe, T.L. and J. D. McCracken. 2015. Long-term trends in the number of Monarch Butterflies (Lepidoptera:Nymphalidae) counted on fall migration at Long Point, Ontario, Canada (1995–2014). Annals of the Entomological Society of America: 1–11
Crolla, J.P. and J.D. Lafontaine.1996. COSEWIC Status Report on the Monarch Butterfly. 23 pp.
Davis, A. K. 2012. Are migratory monarchs really declining in eastern North America? Examining evidence from two fall census programs. Insect Conservation and Diversity 5: 101–105.
Davis, A.K., N.P. Nibbelink, and E. Howard. 2012. Identifying large- and small-scale habitat characteristics of Monarch butterfly migratory roost sites with citizen science observations. International Journal of Zoology. Volume 2012 (2012), Article ID 149026, 9 pages.
Davis, D. and J. Linton. 2016. Monarch wintering population increases by 255 percent however their fate remains uncertain after a devastating winter storm. Ontario Insects 21 (3):13-14.
DeMarco, E. 2015. Monarchs pose a puzzle. Science 349: 570-571.
Dingle, H., M.P. Zalucki, W.A. Rochester and T.Armijo-Prewitt. 2005. Distribution of the monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Nymphalidae), in western North America. Biological Journal of the Linnaean Society 85: 491-500.
DiTommaso, A. and J.E. Losey. 2003. Oviposition preference and larval performance of Monarch Butterflies (Danaus plexippus) on two invasive swallow-wort species. Entomologia Experimentalis et Applicata 108(3): 205-209.
Dockx, X., L.P Brower, L.I. Wasserman and K.A. Hobson. 2004. Do North American Monarch butterflies travel to Cuba? Stable isotope and chemical tracer techniques. Ecological Applications 14 :1106-1114.
Douglas, M.R., J.R. Rohr and J.F. Tooker. 2014. Neonicotinoid insecticide travels through a soil food chain, disrupting biological control of non-target pests and decreasing soya bean yield. Journal of Applied Ecology. 52 (1): 250-260.
Eanes, W.F. and R.K. Koehn. 1978. Analysis of genetic structure in the monarch butterfly, Danaus plexippus L. Evolution 32: 784 – 797.
Environment and Climate Change Canada. 2016. Management Plan for the Monarch (Danaus plexippus) in Canada. Species at Risk Act Management Plan Series. Environment Canada, Ottawa. iv + 45 pp.
Fernandez-Haeger, J., D. Jordano, and M.P. Zalucki. 2015. Monarchs across the Atlantic Ocean. Pp. 247-256. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Fink, L.S. and L.P. Brower. 1981. Birds can overcome the cardelonide defense of Monarch butterflies in Mexico. Nature 291: 67-70.
Flockhart, D.T.T., J.P. Pichancourt, D.R. Norris and T.G. Martin. 2014. Unravelling the annual cycle in a migratory animal: breeding-season habitat loss drives population declines of monarch butterflies. Journal of Animal Ecology. 84(1): 155-165.
Flockhart, D.T.T., L.I. Wassenaar, T.G. Martin, K.A. Hobson, M.B. Wunder and D.R. Norris. 2013. Tracking multi-generational colonization of the breeding grounds by monarch butterflies in eastern North America. Proceedings of the Royal Society B 280: 20131087.
Frey, D. and A. Schaffner. 2004. Spatial and temporal pattern of Monarch overwintering abundance in western North America. Pp. 167-176 In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Garcia-Serrano, E., J.L. Reyes, and B.X. Alvarez. 2004. Locations and areas occupied by Monarch butterflies overwintering in Mexico from 1993 to 2002. pp. 129-133. In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Gauthier, I. pers. comm. 2016. Email communication to A. Harris. April 2016. Biologist Coordonnatrice provinciale des espèces espèces fauniques menacées ou vulnérables. Direction générale de la gestion de la faune et des habitats. Ministère des Forêts, de la Faune et des Parcs. Québec.
Gibo, D.L. 1981. Altitudes attained by migrating Monarch Butterflies, Danaus plexippus (Lepidoptera: Danaidae) as reported by glider pilots. Canadian Journal of Zoology 59(3): 571-572
Gibo, D.L. and J.A. McCurdy. 1993. Lipid accumulation by migrating Monarch butterflies, Danaus plexippus, (Nymphalidae). Canadian Journal of Zoology 71: 76-82.
Gibo, D.L. and M.J. Pallett. 1979. Soaring flight of monarch butterflies, Danaus plexippus (Lepidoptera: Danaidae), during the late summer migration in southern Ontario. Canadian Journal of Zoology. 57:1393-1401.
Glendinning, J.I. 1993. Comparative feeding responses of the mice Peromyscus melanotis, P. aztecus, Reithrodontomys sumichrasti, and Microtus mexicanus to overwintering Monarch butterflies in Mexico. pp. 323-333. In Malcom, S.B. and M.P. Zalucki (eds.), Biology and conservation of the Monarch butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Goulson, D. 2013. An overview of the environmental risks posed by neonicotinoid insecticides. Journal of Applied Ecology. 50: 977–987.
Guerra, P.A., R.J. Gegear, and S.M. Reppert. 2014. A magnetic compass aids monarch butterfly migration. Nature Communications 5: 4164.
Guppy, C.S. and J.H. Shepard. 2001. Butterflies of British Columbia: Including Western Alberta, Southern Yukon, the Alaskan Panhandle, Washington, Northern Oregon, Northern Idaho and Northwestern Montana. UBC Press, Vancouver, B.C . 414 pp.
Gustafsson, K.M., A.A. Agrawal, B.V. Lewenstein and S.A. Wolf. 2015. The Monarch butterfly through time and space: the social construction of an icon. Bioscience 65 (6): 612-622.
Handfield, L., J.D. Lafontaine, B. Landry, F. Lessard, D. Rancourt, E. Rassart and P. Thibault. 1999. Le Guide des Papillons du Québec. Volume 1. Broquet inc., Ottawa. 123 pp.
Hartzler, R.G. 2010. Reduction in common milkweed (Asclepias syriaca) occurrence in Iowa cropland from 1999 to 2009. Crop Protection 29: 1542-1544.
Health Canada. 2016. Update on Neonicotinoid Pesticides and Bee Health. Web site: [Accessed August 2016].
Hess, Q.F. 1992. The Monarch in Ontario. pp. 8-10. In: Hanks, A.J. and Q.F Hess (eds.), Butterflies of Ontario and Summaries of Lepidoptera Encountered in Ontario in 1991. Occasional Publication # 24-92. Toronto Entomologists' Association.
Hobson and Associates. 2006. Environmental Issues and Options for the South Okanagan Regional Growth Strategy. Vol 1: Background. 119 pp. Available at: http://www.rdosmaps.bc.ca/min_bylaws/planning/rgs/ReportsAndStudies/RGSVol1EnvIssuesOptionFinal06.pdf [accessed December 2015]
Holmes, A.M., Q.F. Hess, R.R. Tasker, and A.J. Hanks, 1991. The Ontario Butterfly Atlas. Toronto Entomologists' Association, Toronto, 167 pp.
Howard, E., H. Aschen, and A.K. Davis. 2010. Citizen science observations of monarch butterfly overwintering in the southern United States. Psyche 2010: 1-6.
Howard, E. and A.K. Davis. 2009. The fall migration flyways of monarch butterflies in eastern North America revealed by citizen scientists. Journal of Insect Conservation 13: 279-286.
Howard, E. and A.K. Davis. 2015. Investigating long-term changes in the spring migration of Monarch Butterflies (Lepidoptera: Nymphalidae) using 18 years of data from Journey North, a citizen science program. Annals of the Entomological Society of America: 1–6.
Inamine, H., S. P. Ellner, J.P. Springer, and A.A. Agrawal. 2016. Linking the continental migratory cycle of the monarch butterfly to understand its population decline. Oikos doi: [10.1111/oik.03196].
International Environmental Law Project. 2012. The legal status of Monarch butterflies in California. Monarch Joint Venture and Xerces Society for Invertebrate Conservation. Web site: http://www.xerces.org/wp-content/uploads/2008/09/legal-status-of-california-monarchs.pdf . [Accessed November 2015].
IUCN Species Survival Commission. 2010. Guidelines for Application of IUCN Red List Criteria at Regional and National Levels Version 4.0. Web site: http://www.iucnredlist.org/documents/reg_guidelines_en.pdf [Accessed December 2015].
Jensen, E.V., and P.F. Epp. 2002. Water quality trends in Okanagan, Skaha and Osoyoos Lakes in response to nutrient reductions and hydrologic variation. Ministry of Water, Land and Air Protection. Penticton, B.C.
Jepsen, S. and S.H. Black. 2015. Understanding and conserving the western North American Monarch population. Pp. 147-156. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Jepsen, S., D.F. Schweitzer, B. Young, N. Sears, M. Ormes, and S.H. Black. 2015. Conservation Status and Ecology of the Monarch Butterfly in the United States. NatureServe, Arlington, Virginia, and the Xerces Society for Invertebrate Conservation, Portland, Oregon. 39 pp.
Kagan, R.A., T.C. Viner, P.W. Trail, and E.O. Espinoza. 2014. Avian Mortality at Solar Energy Facilities in Southern California: A Preliminary Analysis. National Fish and Wildlife Forensics Laboratory. Unpublished report.
Klassen, P., R. Westwood, W.B. Preston, and W.B. McKillop.1989. The Butterflies of Manitoba. Manitoba Museum of Man and Nature, Winnipeg, Manitoba.
Klymko, J. pers. comm. 2016. Email communication to A. Harris. April 2016. Zoologist Atlantic Canada Conservation Data Centre, Sackville, NB.
Knight, A. and L.P. Brower. 2009. The influence of eastern North American autumnal migrant Monarch butterflies (Danaus plexippus L.) on continuously breeding resident Monarch populations in Southern Florida. Journal of Chemical Ecology online: doi 10.1007/s10886-009-9655-z.
Krischik V., M. Rogers, G. Gupta, and A. Varshney. 2015. Soil-applied imidacloprid translocates to ornamental flowers and reduces survival of adult Coleomegilla maculata, Harmonia axyridis, and Hippodamia convergens lady beetles, and larval Danaus plexippus and Vanessa cardui butterflies. PLoS ONE 10(3): e0119133. doi:10.1371/journal.pone.0119133
Lane, J. 1984. The status of monarch butterfly overwintering sites in Alta California. Atala 9:17-20.
Lane, J. 1993. Overwintering Monarch Butterflies in California: Past and Present. pp. 335-344. In Malcolm, S.B., and M.P. Zaluchi (eds), Biology and Conservation of the Monarch Butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Layberry, R.A., P.W. Hall, and J.D. Lafontaine. 1998. Butterflies of Canada. University of Toronto Press. 280 pp.
Leong, K., D., W.H. Sakai, W. Bremer, D. Feuerstein, and G. Yoshimura. 2004. Analysis of the pattern of distribution and abundance of monarch overwintering sites along the California coastline. Pp. 177-186 in K.S. Oberhauser and M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Linton, J. pers. comm. 2016. Email communication to A. Harris. April 2016. Natural Resources Solutions Inc. Waterloo, Ontario.
Linton, J. pers. data. Email communication to J.Heron and A. Harris. November 2016. Natural Resources Solutions Inc. Waterloo, Ontario.
Losey, J.E., L.S. Rayor, and M.E.Carter. 1999. Transgenic pollen harms monarch larvae. Nature 399: 214.
Lyons, J.I., A.A. Pierce, S.M. Barribeau, E.D. Sternberg, A.J. Mongue, and J.C. De Roode. 2012. Lack of genetic differentiation between monarch butterflies with divergent migration destinations. Molecular Ecology 21: 3433-3444.
Malcolm, S.B., and L.P. Brower. 1989. Evolutionary and ecological implications of cardenolide sequestration in the Monarch butterfly. Experientia 45:284-295.
Marshall, S.A. 2006. Insects: Their Natural History and Diversity. Firefly Books Ltd., Richmond Hill, Ont.
Mattila, H.R. and G.W. Otis. 2003. A comparison of the host preference of Monarch butterflies (Danaus plexippus) for milkweed (Asclepias syriaca) over dog-strangler vine (Vincetoxinum rossicum). Entomologia Experimentalis et Applicata 107(3): 193-199.
MacNaughton, A., R. Layberry, C. Jones and B. Edwards. 2016. Ontario Butterfly Atlas Online. Web site: [Accessed April 11 2016].
MacNaughton, A. 2010. The birth of Ontario Insects 1995. Ontario Insects 15 (3): 40-41.
Maritimes Butterfly Atlas. 2016. Atlantic Canada Conservation Data Centre, Sackville, New Brunswick. Web site: [Accessed April 1 2016].
Martinez-Torres, H., M. Cantu-Fernandez, M.I. Ramirez, and D.R. Perez-Salicrup. 2015. Fires and fire management in the Monarch Butterfly Biosphere Reserve. Pp. 179-189. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Master, L., D. Faber-Langendoen, R. Bittman, G. A. Hammerson, B. Heidel, J. Nichols, L. Ramsay and A. Tomaino. 2009. NatureServe Conservation Status Assessments: Factors for Assessing Extinction Risk. NatureServe, Arlington, VA.
McKenna D.D., K.M. McKenna, S.B. Malcom, and M.R. Berenbaum. 2001. Mortality of Lepidoptera along roadways in central Illinois. Journal of the Lepidopterists' Society 55(2): 63-68.
Miller, N.G., L.I. Wassenaar, K.A. Hobson, and D.R. Norris. 2012. Migratory connectivity of the Monarch butterfly (Danaus plexippus): patterns of spring re-colonization in eastern North America. PLoS ONE 7(3): e31891. doi:10.1371/journal.pone.0031891
Monarch Watch. 2016. Monarch population status. Web site: [Accessed August 29 2016].
Monroe, M., Pelton, E., C. Fallon, D. Frey, and S. Stevens. 2016. Western Monarch Thanksgiving Count Data from 1997–2015. Web site: http://www.westernmonarchcount.org/data/. [Accessed April 2016].
Natural Heritage Information Centre (NHIC) 2015. Web site: [Accessed November 2015].
Natural Resources Canada. 2016. Renewables. Web site: [Accessed April 7 2016].
Nature Serve. 2015. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. [accessed October 2015]
North American Butterfly Association. 2014. There's no need to release butterflies -- they're already free. Web site: [Accessed November 2015].
Oberhauser K.S. 2004. Overview of Monarch breeding biology. Pp. 3-8. In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Oberhauser K.S. and M.J. Solensky (eds). 2004. The Monarch Butterfly: Biology and Conservation. Oberhauser and Solensky Cornell University Press, Ithaca, New York.
Oberhauser, K.S., L, Ries, S. Altizer, R.V. Batalden, J. Kudell-Ekstrum, M. Garland, E. Howard, S. Jepsen, J. Lovett, M. Monroe, G. Morris, E. Rendón-Salinas, R.G. RuBino, A, Ryan, O.R. Taylor, R. Trevino, F.X. Villablanco, and D. Walton. 2015a. Contributions to Monarch biology and conservation through citizen science. Pp. 13-30. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Oberhauser, K.S., M. Anderson, S. Anderson, W. Caldwell, A. De Anda, M. Hunter, M.C. Kaiser, and M.J. Solensky. 2015b. Lacewings, wasps, and flies – oh my. Pp. 71-82. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Oberhauser, K., and A.T. Peterson. 2003. Modeling current and future potential wintering distributions of eastern North American Monarch butterflies. Proceedings of the National Academy of Sciences of the United States of America 100:14063-14068.
Ontario’s Invading Species Awareness Program. 2015. Dog-strangling Vine. Web site: http://www.invadingspecies.com/invaders/plants-terrestrial/dog-strangling-vine/. [Accessed November 2015]
Ontario Ministry of Agriculture, Food and Rural Affairs. 2014 (updated August 3 2016). Pollinator Health. Web site: [Accessed August 2016].
Opler, P. A., and A. D. Warren. 2002. Butterflies of North America. 2. Scientific Names List for Butterfly Species of North America, north of Mexico. C.P Gillette Museum of Arthropod Diversity, Department of Bioagricultural Sciences and Pest Management, Colorado State University, Fort Collins, Colorado. 79 pp.
Parmesan, C. 2007. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology (13): 1860-1872.
Pecenka, J.R. and J.G. Lundgren. 2015. Non-target effects of clothianidin on monarch butterflies. The Science of Nature 102: 19.
Perez, S.M., O.R. Taylor, and R. Jander. 1997. A sun compass in monarch butterflies. Nature 387: 29.
Pierce, A.A.,S. Altizer, N.L. Chamberlain, M.R. Kronforst, and J.C. de Roode. 2015. Unraveling the mysteries of Monarch migration and global dispersal through molecular genetic techniques. Pp. 257-267. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Pleasants, J.M. and K.S. Oberhauser. 2012. Milkweed loss in agricultural fields because of herbicide use: effect on the monarch butterfly population. Insect Conservation and Diversity (2012) doi: 10.1111: 1-10.
Pleasants, J.M. 2015. Monarch butterflies and agriculture. Pp. 169-178. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Pleasants, J.M., E.H. Williams, L.P. Brower, K.S. Oberhauser, and O.R. Taylor. 2016. Conclusion of no decline in summer monarch population not supported. Annals of the Entomological Society of America. 2016: 1–3 doi: 10.1093/aesa/sav115
Pleasants, J.M. 2017. Milkweed restoration in the Midwest for monarch butterfly recovery: estimates of milkweeds lost, milkweeds remaining and milkweeds that must be added to increase the monarch population. Insect Conservation and Diversity 10(1): 42 – 53.
Province of Manitoba. 2010. The Noxious Weeds Act. Manitoba Laws. Web site: [accessed April 14 2016].
Province of Nova Scotia. 2016. Weed control regulations. Web site: [Accessed April 14 2016]
Pyle, R.M. 2015. Monarchs in the mist. Pp. 236-246. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Ramirez, M.I., C. Saenz-Romero, G. Rehfeld, and L. Salas-Canela. 2015. Threats to the availability of overwintering habitat in the Monarch Butterfly Biosphere Reserve. Pp. 157-168. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Rendón-Salinas, E., A. Fajardo-Arroyo, and G. Tavera-Alonso. 2014. Forest surface occupied by Monarch butterfly hibernation colonies in December 2014 [PDF, 556 Kb]. Web site: [Accessed April 12 2016].
Ries, L., D.J. Taron, and E. Rendon-Salinas. 2015a. The disconnect between summer and winter Monarch trends for the eastern migratory population: possible links to differing drivers. Annals of the Entomological Society of America 1–9.
Ries, L., D.J. Taron, E. Rendon-Salinas and K.S. Oberhauser. 2015b. Connecting eastern Monarch population dynamics across their migratory cycle. Pp. 268-281. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Riley, J.L. 2013. The Once and Future Great Lakes Country. An Ecological History. McGill – Queen’s University Press.
Robinson, R.A., H.Q.P. Crick, J.A. Learmonth, I.M.D. Maclean, C.D. Thomas, F. Bairlein, M.C. Forchhammer, C.M. Francis, J.A. Gill, B.J. Godley, J. Harwood, G. C. Hays, B. Huntley, A.M. Hutson, G.J. Pierce, M.M. Rehfisch, D.W. Sims, M.B. Santos, T H. Sparks, D.A. Stroud and M.E. Visser. 2009. Travelling through a warming world: climate change and migratory species. Endangered Species Research (7): 87-99.
Salafsky, N., D. Salzer, A. J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O’Connor, and D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conservation Biology 22:897-911. Web site: http://conservationmeasures.org/CMP/IUCN/browse.cfm?TaxID=DirectThreats [Accessed April 10 2016].
Satterfield D.A., J.C. Maerz, and S. Altizer. 2015. Loss of migratory behaviour increases infection risk for a butterfly host. Proc. R. Soc. B 282: 20141734.
Schappert, P. 2004. Monarch Butterflies: Saving the King of the New World. Key Porter Books, Toronto, ON. 128 pp.
Scott, J.A. 1986. The Butterflies of North America: A Natural History and Field Guide. Stanford University Press, Stanford, California. 583 pp.
Semmens, B.X., D.J. Semmens, W.E. Thogmartin, R. Wiederholt, L. López-Hoffman, J.E. Diffendorfer, J.M. Pleasants, K.S. Oberhauser and O.R. Taylor. 2016. Quasi-extinction risk and population targets for the Eastern, migratory population of monarch butterflies (Danaus plexippus). Scientific Reports 6, Article number: 23265 doi:10.1038/srep23265
Skórkaa, P., M. Lendab, D. Morońc, K. Kalarusd, P. Tryjanowskia. 2013. Factors affecting road mortality and the suitability of road verges for butterflies. Biological Conservation 159: 148–157.
Slayback, D.A., L.P. Brower, M.I. Ramírez, and L.S. Fink. 2007. Establishing the presence and absence of overwintering colonies of the Monarch Butterfly in Mexico by the use of small aircraft. American Entomologist: 27-40.
Smith, D.A.S., G. Lushai, and J.A. Allen. 2005. A classification of Danaus butterflies (Lepidoptera: Nymphalidae) based upon data from morphology and DNA. Zoological Journal of the Linnean Society 144(2):191-212.
Snell-Rooda, E.C., A. Espeseta, C.J. Bosera, W.A. White, and R. Smykalskia. 2014. Anthropogenic changes in sodium affect neural and muscle development in butterflies. PNAS 111(28) 10221–10226, doi: 10.1073/pnas.1323607111
Snook, L.C. 1993. Conservation of the Monarch butterfly reserves in Mexico: Focus on the Forest. Pages 363-375 In Malcom, S. B. and M. P. Zalucki (eds.), Biology and conservation of the Monarch butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Solensky, M.J. 2004. Overview of monarch migration. Pp. 79-83. in K.S. Oberhauser and M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Southwest Monarch Study. 2013. Accessed February 2017.
St. John, D, E. Antonijevic and J. Heron. 2014. Surveys for the Monarch Butterfly (Danaus plexippus) and Showy Milkweed (Asclepias speciosa) in the Okanagan and Similkameen river valleys of British Columbia. Unpublished report. B.C. Ministry of Environment, Vancouver, B.C. 53pp.
Stevens, S.R. and D.F. Frey. 2010. Host plant pattern and variation in climate predict the location of natal grounds for migratory monarch butterflies in western North America. Insect Conserv. 14:731–744.
Suzuki, Y. and M.P. Zalucki. 1986. The influence of sex ratio on female dispersal in Danaus plexippus (L.)(Lepidopetra: Danaidae). Journal of the Australian Entomological Society 25: 31-35.
The Center for Biological Diversity, Center for Food Safety, The Xerces Society & L. Brower 2014. Petition to protect the Monarch Butterfly (Danaus plexippus plexippus) under the Endangered Species Act. Web site: http://xerces.org/wp-content/up loads/2014/08/monarch-esa-petition.pdf. [Accessed April 10 2016].
Thomas, A.W.. 1996. A Preliminary Atlas of the Butterflies of New Brunswick. Natural Resources Canada, Canadian Forest Service, Atlantic Forestry Centre, Fredericton, New Brunswick. New Brunswick Museum Publications in Natural Science 11. 41 pp.
United Nations Educational, Scientific and Cultural Organization [PDF, 1.73 Mb] (UNESCO). 2015. State of conservation of properties inscribed on the World Heritage List. Web site: [Accessed April 13 2016].
Urquhart, F. A.1987. The Monarch Butterfly: International Traveller. Nelson-Hall, Chicago. 42 pp.
van der Sluijs, J.P., N. Simon-Delso, D. Goulson, L. Maxim, J.M. Bonmatin and L.P. Belzunces. 2013. Neonicotinoids, bee disorders and the sustainability of pollinator services. Current Opinion in Environmental Sustainability. 5:293–305.
Vane-Wright, R.I. 1993. The Columbus hypothesis: an explanation for the dramatic 19th century range expansion of the monarch butterfly. In S.B. Malcolm & M. Zalucki (eds), Biology and conservation of the monarch butterfly, pp. 179–187. Science Series (38), Natural History Museum of Los Angeles County, Los Angeles.
Vidal, O., J. López-García, and E. Rendón-Salinas. 2014. Trends in deforestation and forest degradation after a decade of monitoring in the Monarch Butterfly Biosphere Reserve in Mexico. Conserv Biol. 28(1): 177–186.
Vidal, O. and E. Rendón-Salinas. 2014. Dynamics and trends of overwintering colonies of the monarch butterfly in Mexico. Biological Conservation 180: 165 – 175.
White, D.J. 1996. The milkweeds of Canada (Asclepias), status, distribution, and potential impacts from noxious weeds legislation. Report prepared for the CWS, accessed from Monarch Watch, June 14, 2008.
Williams, E.H. and L.P. Brower. 2015. Microclimatic protection of overwintering Monarchs provided by Mexico’s high elevation Oyamel Fir forests. Pp. 109-116. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Williams, J., D. Stow, and L. Brower, 2007. the influence of forest fragmentation on the location of overwintering Monarch butterflies in central Mexico, Journal of the Lepidopterist’s Society 61: 90-104.
Wormington, A. pers. comm. 2007. Email communication to K. Bériault. January 2007.
Wormington, A. 1994. A mass migration of Monarchs at Point Pelee, Ontario. Toronto Entomologists Association, Occasional Publication #26-94, pp. 26-27.
Wormington, A. 1997. Another mass migration of Monarchs at Point Pelee, Ontario. Toronto Entomologists Association, Occasional Publication #29-97, p. 11.
Wormington, A. 2008. The Butterflies of Point Pelee National Park, Ontario. Unpublished Manuscript.
World Wildlife Fund (WWF). 2015. 96% of all deforestation within Mexico’s Monarch butterfly sanctuaries occur in a single community. Web site: [Accessed April 11 2016].
York, H.A. and K.S. Oberhauser. 2002. Effects of duration and timing of heat stress on monarch butterfly (Danaus plexippus) (Lepidoptera: Nymphalidae) development. Journal of the Kansas Entomological Society, Vol. 75 (4): 290-298.
Zalucki, M.P.and W.A. Rochester. 2004. Spatial and temporal population dynamics of Monarchs Down-Under: lessons for North America. Pp. 219-228. In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Zalucki, M.P., L.P. Brower, S.B. Malcolm, and B.H Slager. 2015. Estimating the climate signal in Monarch population decline. Pp. 130-141. in K.S. Oberhauser, K.R. Nail, and S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Zhan, S., W. Zhang, K. Niitepold, J. Hsu, J. Fernandez Haeger, M.P. Zalucki, S. Altizer, J.C. de Roode, S.M. Reppert and M.R. Kronforst. 2014. The genetics of monarch butterfly migration and warning colouration. Nature 514: 317-321.
Zipkin, E. F., L. Ries, R. Reeves, J. Regetz and K. Oberhauser. 2012. Tracking climate impacts on the migratory monarch butterfly. Global Change Biology 18: 3039 – 3049.
Biographical summary of report writers
Allan Harris is a biologist with over 25 years’ experience in northern Ontario. He has a B.Sc. in Wildlife Biology from the University of Guelph and an M.Sc. in Biology from Lakehead University. After spending seven years as a biologist with Ontario Ministry of Natural Resources, he co-founded Northern Bioscience, an ecological consulting company based in Thunder Bay, Ontario. Al has authored or coauthored dozens of scientific papers, technical reports, and popular articles, including COSEWIC status reports for Riverine Clubtail, Laura’s Clubtail, Rapids Clubtail, Gibson’s Big Sand Tiger Beetle, Northern Barrens Tiger Beetle, Powesheik Skipperling, Mormon Metalmark, Weidemeyer’s Admiral, Bogbean Buckmoth, Hop-tree Borer, Georgia Basin Bog Spider, Broad-banded Forestsnail, Nahanni Aster, Crooked-stem Aster, Bluehearts, Drooping Trillium and Small-flowered Lipocarpha. Al also authored the Ontario provincial status report for woodland caribou, and has authored or coauthored national and provincial recovery strategies for vascular plants and birds.
Robert Foster is co-founder and principal of Northern Bioscience, an ecological consulting firm offering professional consulting services supporting ecosystem management, planning, and research. Dr. Foster has a B.Sc. in Biology from Lakehead University and a D. Phil in Zoology from the University of Oxford. Rob has worked as an ecologist in Ontario for over 20 years, and has authored or coauthored COSEWIC status reports on the Riverine Clubtail, Laura’s Clubtail, Rapids Clubtail, Gibson’s Big Sand Tiger Beetle, Northern Barrens Tiger Beetle, Powesheik Skipperling, Mormon Metalmark, Weidemeyer’s Admiral, Bogbean Buckmoth, Hop-tree Borer, Georgia Basin Bog Spider, Broad-banded Forestsnail, Nahanni Aster, Crooked-stem Aster, Bluehearts, and Drooping Trillium, as well as recovery plans for rare plants, lichens, and odonates. Rob has conducted numerous odonate and other insect surveys for protected areas planning and environmental assessments in Ontario, as well as Manitoba, Minnesota, Quebec, and British Columbia.
Collections examined
None.