
Monarque (Danaus plexippus) : évaluation et rapport de situation du COSEPAC 2016

En voie de disparition
2016
Table des matières
- Table des matières
- Sommaire de l’évaluation
- Résumé
- Résumé technique
- Préface
- Description et importance de l’espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances de la population
- Menaces et facteurs limitatifs
- Développement résidentiel et commercial (UICN 1, impact faible)
- Agriculture et aquaculture (UICN 2, impact moyen)
- Production d’énergie et exploitation minière (UICN 3, impact négligeable)
- Corridors de transport et de service (UICN 4, impact faible)
- Utilisation des ressources biologiques (IUCN 5, impact élevé)
- Intrusions et perturbations humaines (UICN 6, impact négligeable)
- Modifications des systèmes naturels (UICN 7, impact négligeable)
- Espèces et gènes envahissants ou problématiques (IUCN 8, impact inconnu)
- Pollution (UICN 9, impact élevé)
- Changements climatiques et phénomènes météorologiques violents (UICN 11, impact faible)
- Facteurs limitatifs
- Nombre de localités
- Protection, statuts et classements
- Remerciements
- Experts contactés
- Sources d’information
- Sommaire biographique des rédactrices du rapport
- Collections examinées
Liste des figures
- Figure 1. Adulte du monarque (Danaus plexippus). Photo : Jessica Linton.
- Figure 2. Chenille du monarque (Danaus plexippus). Photo : Jessica Linton.
- Figure 3. Monarques se rassemblant dans un site d’hivernage, au Mexique. Photo : Jessica Linton.
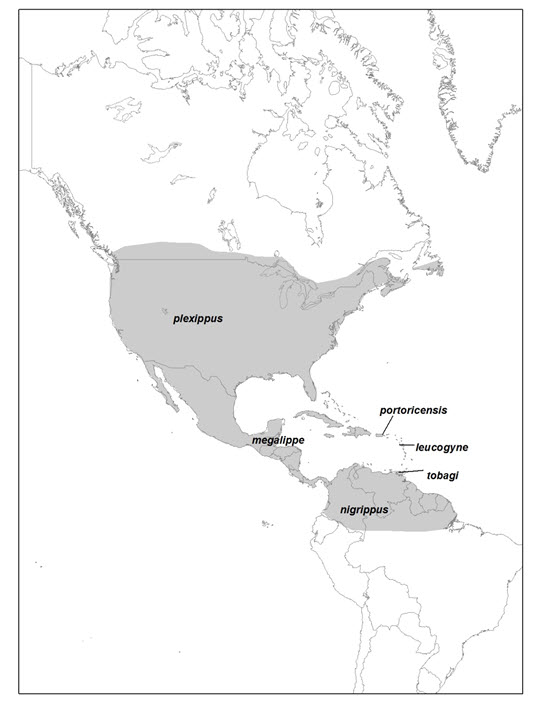
- Figure 4. Aire de répartition nord-américaine (indigène) du monarque (Danaus plexippus), avec indication de la répartition approximative des six sous-espèces (adapté de Pierce et al., 2015).
- Figure 5. Aire de répartition canadienne du monarque (Danaus plexippus). La ligne tiretée représente la limite nord approximative des plantes hôtes de la chenille de l’espèce, soit l’asclépiade incarnate, l’asclépiade commune et la belle asclépiade. Les points situés au nord de cette ligne et dans la région côtière de la Colombie-Britannique représentent des individus errants non reproducteurs et les occurrences observées dans les parcelles isolées où des asclépiades ont été plantées à l’extérieur de leur aire de répartition indigène. Les zones d’hivernage des populations de l’est et de l’ouest sont indiquées en gris.
- Figure 6. Estimation de l’abondance du monarque (Danaus plexippus) dans les sites d’hivernage de la Californie, de 1997 à 2015 (Monroe et al., 2016).
- Figure 7. Superficie totale (hectares) occupée par les colonies de monarques (Danaus plexippus) dans les sites d’hivernage du Mexique, de 1994-1995 à 2015-2016 (Monarch Watch, 2016).
- Figure 8. Superficie totale (ha) occupée par le monarque (Danaus plexippus) dans les sites d’hivernage du Mexique, avec droites de régression d’ajustement montrant le déclin au fil du temps. La ligne pointillée représente la droite d’ajustement pour la totalité de la série chronologique (1994-1995 à 2015-2016), alors que la ligne pleine représente celle pour les dix dernières années (2006-2007 à 2015-2016).
Liste des tableaux
- Tableau 1. Classification des menaces pesant sur le monarque (voir Salafsky et al., 2008; Master et al., 2009), réalisée à l’aide du système de classification des menaces proposé par l’Union internationale pour la conservation de la nature (UICN) et le Partenariat pour les mesures de conservation (Conservation Measures Partnership, ou CMP) (IUCN-CMP). Les voies migratoires de l’est et de l’ouest ont été prises en compte pour l’évaluation. L’impact global des menaces calculé pour le monarque est très élevé.
Information sur le document
COSEPAC
Comité sur la situation
des espèces en péril
au Canada
![]()
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l’on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur le monarque (Danaus plexippus) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xiv + 65 p. (Registre public des espèces en péril site Web).
Rapport(s) précédent(s) :
COSEPAC. 2010. Évaluation et Rapport de situation du COSEPAC sur le monarque (Danaus plexippus) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. vii + 51 p. (Registre public des espèces en péril site Web).
COSEPAC. 2001. Évaluation et Rapport de situation du COSEPAC sur le monarque (Danaus plexippus) au Canada – Mise à jour. Comité sur la situation des espèces en péril au Canada. Ottawa. v + 23 p. (Registre public des espèces en péril site Web).
1997. Draft. COSEWIC status report on the Monarch Danaus plexippus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa.
Note de production :
Le COSEPAC remercie Allan Harris et Robert Foster d’avoir rédigé le rapport de situation sur le monarque (Danaus plexippus) au Canada, aux termes d’un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Jennifer Heron, coprésidente du Sous-comité de spécialistes des arthropodes du COSEPAC.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Monarch Danaus plexippus in Canada.
Illustration/photo de la couverture :
Monarque -- Photo fournie par les rédacteurs.
COSEPAC Sommaire de l’évaluation
Sommaire de l’évaluation – novembre 2016
- Nom commun
- Monarque
- Nom scientifique
- Danaus plexippus
- Statut
- En voie de disparition
- Justification de la désignation
- Cette grande espèce remarquable est un des papillons les plus connus dans le monde. La population canadienne est migratrice et elle emprunte deux voies distinctes. Des menaces cumulatives pèsent sur les sites d’hivernage et le long des voies de migration. Le groupe migrateur à l’ouest des montagnes Rocheuses se déplace entre la côte de la Californie et le sud de la Colombie-Britannique. Le groupe à l’est des montagnes Rocheuses représente la vaste majorité de la population canadienne et se déplace entre la forêt de sapin oyamel du centre du Mexique et le sud du Canada à l’est de l’Alberta. Les sites d’hivernage dans le centre du Mexique sont extrêmement petits, et les menaces qui pèsent sur ces aires incluent l’exploitation forestière illégale et le développement agricole, ainsi qu’une augmentation de la fréquence et de la gravité des tempêtes pendant les périodes clés de rassemblement. Des déclins de plus de 50 % se sont produits au cours de la dernière décennie.
- Répartition
- Territoires-du-Nord-Ouest, Colombie-Britannique, Alberta, Saskatchewan, Manitoba, Ontario, Québec, Nouveau-Brunswick, Île-du-Prince-Édouard , Nouvelle-Écosse , Terre-Neuve-et-Labrador
- Historique du statut
- Espèce désignée « préoccupante » en avril 1997. Réexamen et confirmation du statut en novembre 2001 et en avril 2010. Réexamen du statut : l'espèce a été désignée « en voie de disparition » en novembre 2016.
COSEPAC Résumé
Monarque
Danaus plexippus
Description et importance de l’espèce sauvage
Le monarque est une espèce emblématique de la conservation et compte parmi les espèces de papillons les plus connues et les plus étudiées dans le monde. Le cycle vital du monarque comprend quatre stades. Le monarque adulte est un superbe et grand papillon (envergure de 93 à 105 mm) dont les ailes sont principalement orange et comportent une large bordure noire piquée de deux rangées de points circulaires blancs. La chenille, facilement reconnaissable, est annelée de blanc, de jaune et de noir et porte deux paires de filaments noirs, une au niveau de la tête et l’autre à l’extrémité de l’abdomen. La chrysalide est vert et doré. L’œuf du monarque mesure environ 1 mm de longueur et est légèrement ovale, avec la base aplatie et le sommet formant une pointe arrondie.
Le monarque est l’une des rares espèces de papillons qui migrent, et sa migration depuis le sud du Canada jusqu’au Mexique est considérée comme un phénomène biologique en péril. Partout en Amérique du Nord, le monarque est utilisé dans les écoles pour enseigner aux enfants la biologie, la métamorphose et la conservation et leur faire apprécier la nature.
Répartition
Le monarque est une espèce migratrice. L’aire de répartition indigène globale du monarque s’étend depuis l’Amérique centrale jusqu’au Canada, en passant par la partie continentale des États-Unis, et de la côte de l’Atlantique jusqu’à la côte du Pacifique. Au Canada, l’espèce se rencontre dans certaines portions des dix provinces et des Territoires du Nord-Ouest. L’espèce peut être divisée en deux sous-groupes, la population de l’est et la population de l’ouest, en fonction des voies migratoires et des sites d’hivernage des individus. L’aire de reproduction de la population de l’est s’étend depuis l’Alberta jusqu’à la Nouvelle-Écosse, et les individus de cette population migrent vers le sud pour aller hiverner dans les montagnes du centre du Mexique. Les individus de la population de l’ouest se reproduisent dans le sud de la Colombie-Britannique et migrent vers le sud jusqu’à leur aire d’hivernage, située sud les côtes de la Californie. L’aire de reproduction de l’espèce au Canada se situe au sud du 50e parallèle en Ontario, au Québec et dans les Maritimes et au sud du 54e parallèle au Manitoba, en Saskatchewan et en Alberta.
Le monarque est considéré comme formant une seule unité désignable au Canada aux fins de la présente évaluation. Des échanges d’individus ont été observés entre les populations de l’est et de l’ouest durant la migration, et aucune observation génétique ou morphologique n’indique qu’il pourrait y avoir deux sous-espèces.
Le monarque a colonisé l’Europe continentale, l’Afrique du Nord, l’Australie et de nombreuses îles du Pacifique au cours des 200 dernières années, mais les populations ainsi établies ne migrent pas.
Habitat
Les asclépiades (nombreuses espèces) sont les seules plantes dont se nourrit la chenille du monarque. Ces plantes poussent principalement dans les milieux dégagés et subissant périodiquement des perturbations, comme les bords de route, les champs, les milieux humides, les prairies et les forêts claires. Les asclépiades sont souvent plantées à l’extérieur de leur aire de répartition indigène, et des monarques errants sont parfois observés dans ces parcelles.
Pour l’hivernage, le monarque a besoin d’un microclimat frais et humide et d’être protégé du gel, d’une exposition excessive au soleil, du vent et des précipitations fortes. Il trouve ces conditions en Californie, dans la région de la côte du Pacifique, et dans le centre du Mexique, dans les forêts de haute altitude. Les individus de la population de l’est hivernent à des altitudes de 2 900 à 3 300 m, dans les forêts de sapin oyamel du Mexique. Ceux de la population de l’ouest hivernent à quelques kilomètres de la côte du Pacifique, en Californie, principalement dans des peuplements d’eucalyptus, arbres exotiques qui ont été plantés à partir des années 1850 pour remplacer les essences indigènes, dont les pins.
Le monarque a besoin d’aires de rassemblement où les individus peuvent se reposer, s’alimenter et s’abriter des intempéries durant la migration. Au Canada, on trouve ce type d’aires le long des rives nord des Grands Lacs, où les monarques se reposent sur les arbres avant de traverser ces vastes étendues d’eau libre.
Biologie
Les adultes se reproduisent durant l’hiver et au début du printemps dans les sites d’hivernage du Mexique ou de la Californie, puis entreprennent leur migration vers le nord à la fin février ou au début mars. Environ 10 % des individus de la population de l’est font le voyage en entier jusqu’au Canada, mais la plupart des femelles qui quittent les sites d’hivernage se reproduisent dans le sud des États-Unis. La femelle du monarque pond 300 à 400 œufs, chaque œuf étant déposé individuellement sur la face inférieure des feuilles d’asclépiade. L’éclosion survient trois à huit jours après la ponte, puis la chenille se nourrit des feuilles d’asclépiade de façon presque continue, multipliant son poids corporel par 2 000. Après s’être alimentée durant 9 à 14 jours, la chenille se transforme en chrysalide, et l’adulte émerge 9 à 15 jours plus tard. Les adultes de la génération suivante poursuivent la migration vers le nord, et plusieurs d’entre eux se reproduisent dans le centre des États-Unis, donnant naissance à une nouvelle génération. La plupart des individus qui atteignent le Canada sont en fait les arrière-petits-enfants de ceux qui ont quitté le Mexique. Dans le sud de l’Ontario et du Québec, l’espèce produit chaque année deux ou trois générations, de juin à octobre. Dans le sud de la Colombie-Britannique, l’espèce produit au moins une génération chaque été. Les adultes estivaux ont une durée de vie de deux à cinq semaines, mais les adultes hivernants peuvent vivre jusqu’à neuf mois. Les adultes qui émergent à la fin de l’été migrent vers le Mexique ou la Californie, où ils hivernent avant d’entreprendre une nouvelle migration en sens inverse.
La chenille du monarque stocke les composés chimiques présents chez les asclépiades, ce qui rend l’espèce, également au stade adulte, non comestible pour la plupart des oiseaux et des autres vertébrés.
Taille et tendances de la population
On ne dispose d’aucune estimation de la taille de la population de monarque au Canada. Chaque automne, des centaines de milliers de monarques migrent à partir de la région de Long Point, dans le sud de l’Ontario, mais on ignore quelle proportion de la population canadienne ces individus représentent. Une estimation de la population a été faite dans les sites d’hivernage, où se rassemblent les individus provenant du Canada et des États-Unis. L’effectif total de la population hivernant au Mexique (population de l’est) a été évalué à 66 millions d’individus en 2014-2015, et à 200 millions d’individus en 2015-2016. Une tempête survenue en mars 2016 a tué un grand nombre d’individus dans les sites d’hivernage de la population de l’est, mais on ignore l’ampleur exacte des pertes. Moins de 500 000 individus hivernent actuellement en Californie (population de l’ouest), et seulement un très faible pourcentage de ceux-ci se reproduisent au Canada. Certaines années, la population reproductrice de l’ouest (Colombie-Britannique) est si petite qu’elle n’est pas détectée.
La population hivernante du Mexique, dont la taille est mesurée d’après la superficie d’habitat occupée (hectares), a considérablement diminué au cours de la période de 1994 à 2015. Selon une régression linéaire logarithmique de cette série chronologique, la population a subi un déclin de 83 %. Les estimations pour la période 2012-2014 ont été les plus basses de cette série. La superficie d’habitat occupée en 2015 (4 ha) a été plus élevée que celle enregistrée les trois années précédentes, mais tout de même inférieure à la moyenne pour la série chronologique, qui est de 6 ha. Le taux de déclin de l’habitat occupé au cours d’une période de 10 ans a été calculé au moyen la pente de la régression linéaire logarithmique établie d’après la série chronologique entière, appliquée à une période de 10 ans. Le changement a ainsi été évalué à - 59 % sur une période de 10 ans.
Une étude à long terme sur la migration a été réalisée dans la région de Long Point, en Ontario; selon les données ajustées en fonction des conditions météorologiques, la population a connu un faible déclin. Toutefois, aucun signe de déclin n’a été observé dans le cadre d’études semblables menées à Cape May (New Jersey) et à Peninsula Point (Michigan).
Menaces et facteurs limitatifs
Durant l’hiver, la majeure partie des monarques d’Amérique du Nord se concentrent sur seulement quelques hectares et sont donc vulnérables aux phénomènes météorologiques extrêmes, aux incendies, aux maladies, à la prédation et aux menaces anthropiques. L’habitat d’hivernage situé dans les forêts de sapin oyamel du Mexique a été fragmenté et dégradé par l’aménagement de terres agricoles, les incendies, l’exploitation forestière et les coupes d’éclaircie. Ces pratiques augmentent le risque d’exposition des individus hivernants aux tempêtes hivernales ainsi qu’aux conditions froides et humides, ce qui entraîne une hausse du taux de mortalité. Selon les modèles de changements climatiques, dans l’aire d’hivernage du Mexique, il y aura une diminution de la superficie de forêts convenant à l’espèce et une augmentation de la fréquence des tempêtes hivernales, ce qui entraînera une mortalité catastrophique. Dans le cas de la population de l’ouest, la dégradation de l’habitat d’hivernage résulte principalement du développement immobilier qui se poursuit le long de la côte de la Californie et de l’élimination des eucalyptus introduits sur lesquels les monarques se rassemblent pour hiverner.
L’utilisation accrue d’herbicides et les déclins connexes des asclépiades représentent une menace considérable pour le monarque dans l’ensemble de son aire de répartition nord-américaine. L’utilisation accrue d’herbicides peut également causer un déclin des plantes nectarifères dont les individus migrateurs ont besoin et ainsi réduire les taux de survie hivernale. Les pesticides de la catégorie des néonicotinoïdes sont une menace nouvelle, mais l’ampleur de leur impact est mal comprise.
Protection, statuts et classements
Le monarque a été désigné espèce préoccupante par le COSEPAC en 1997. Son statut a été réexaminé et confirmé en 2001 et en 2010, et l’espèce a été désignée espèce en voie de disparition en 2016. Le monarque est considéré comme une espèce préoccupante aux termes la Loi de sur les espèces en voie de disparition de l’Ontario et de la Loi sur les espèces en péril du Nouveau-Brunswick. Il est « sous protection particulière » au Mexique, et il fait actuellement l’objet d’une évaluation en vue de son inscription à l’Endangered Species Act aux États-Unis. Les populations reproductrices du monarque sont classées apparemment non en péril (G4) à l’échelle mondiale et non en péril au Canada et aux États-Unis. Les zones de concentration d’individus migrateurs sont classées comme vulnérables à en péril aux États-Unis.
Résumé technique
- Nom scientifique :
- Danaus plexippus
- Nom français :
- Monarque
- Nom anglais :
- Monarch
- Répartition au Canada :
- Colombie-Britannique, Alberta, Saskatchewan, Manitoba, Ontario, Québec, Nouveau-Brunswick, Nouvelle-Écosse, Île-du-Prince-Édouard, Territoires du Nord-Ouest et Terre-Neuve-et-Labrador.
Données démographiques
| Éléments du résumé technique | information |
|---|---|
| Durée d’une génération | -0,33 an Âge moyen auquel une nouvelle cohorte peut produire une descendance, si on suppose qu’il y a trois générations par année. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d’individus matures? | Oui, inféré. Si on infère que les déclins de la population hivernante reflètent ceux des populations canadiennes. |
| Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations]. | Oui, réduction estimée et inférée de 59 % d’après la diminution de la superficie (ha) d’habitat d’hivernage occupée au Mexique (figure 8). Si on infère que les déclins de la population hivernante reflètent ceux des populations canadiennes. |
| Pourcentage [prévu ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
| Est-ce que les causes du déclin sont a. clairement réversibles et b. comprises et c. ont effectivement cessé? | a. Non Le déclin est probablement causé par les effets cumulatifs de la raréfaction des asclépiades, de la déforestation, de la dégradation des sites d’hivernage et des changements climatiques. Les répercussions des néonicotinoïdes, les maladies et le déclin des sources de nectar sont des menaces mal comprises. |
| Y a-t-il des fluctuations extrêmes du nombre d’individus matures? | Non. Les indices d’abondance de la population hivernante fluctuent, mais les changements ont été inférieurs à un ordre de grandeur sur deux années successives. |
Information sur la répartition
| Éléments du résumé technique | information |
|---|---|
| Superficie estimée de la zone d’occurrence | ~ 1,3 million de km2 Superficie de la zone d’occurrence au Canada uniquement. |
Indice de zone d’occupation (IZO) (Fournissez toujours une valeur établie à partir d’une grille à carrés de 2 km de côté) |
Aire de reproduction au Canada : Aire d’hivernage totale (ouest [Californie] et est [Mexique]) : 588 km2 |
| La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d’occupation totale se trouvent dans des parcelles d’habitat qui sont a) plus petites que la superficie nécessaire au maintien d’une population viable et b) séparées d’autres parcelles d’habitat par une distance supérieure à la distance de dispersion maximale présumée pour l’espèce? | a. Non b. Non |
| Nombre de localités (utilisez une fourchette plausible pour refléter l’incertitude, le cas échéant) (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Sans objet. |
| Y a-t-il un déclin [observé, inféré ou prévu] de la zone d’occurrence? | Non |
| Y a-t-il un déclin [observé, inféré ou prévu] de l’indice de zone d’occupation? | Non |
| Y a-t-il un déclin [observé, inféré ou prévu] du nombre de sous-populations? | Non |
| Y a-t-il un déclin [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non |
| Y a-t-il un déclin [observé, inféré ou prévu] de [la superficie, l’étendue ou la qualité] de l’habitat? | Oui, déclin inféré de la qualité de l’habitat. Si on suppose que les diminutions de l’abondance des asclépiades observées aux États-Unis s’appliquent également au Canada. La qualité de l’habitat d’hivernage a diminué au Mexique et en Californie. |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? | Non |
| Y a-t-il des fluctuations extrêmes de la zone d’occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation? | Non |
Nombre d’individus matures dans chaque sous-population
| Sous-population (utilisez une fourchette plausible) : | Nombre d’individus matures |
|---|---|
| Ouest | Inconnu |
| Est | Inconnu |
| Total | - |
Analyse quantitative
| Éléments du résumé technique | information |
|---|---|
| La probabilité de disparition de l’espèce à l’état sauvage est d’au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans] | Selon les prédictions, les probabilités de quasi-disparition de la population de l’est au cours des 20 prochaines années sont de 11 % à 57 % (Semmens et al., 2016). Puisque le modèle comprend uniquement la population de l’est, le critère E n’a pas été invoqué. |
Menaces (directes, de l’impact le plus élevé à l’impact le plus faible, selon le calculateur des menaces de l’UICN)
Un calculateur des menaces a-t-il été rempli pour l’espèce? Oui.
Calculateur des menaces de l’UICN – impact calculé très élevé
- 9.3 Effluents agricoles et sylvicoles (impact élevé)
- 2.1 Cultures annuelles et pérennes de produits autres que le bois (impact moyen)
- 5.3 Exploitation forestière et récolte du bois (impact moyen)
- 1.1 Zones résidentielles et urbaines (impact faible)
- 4.1 Routes et voies ferrées (impact faible)
- 11.1 Déplacement et altération de l’habitat (impact faible)
- 11.2 Sécheresses (impact faible)
- 11.3 Températures extrêmes (impact faible)
- 11.4 Tempêtes et inondations (impact faible)
- 9.5 Polluants atmosphériques (impact inconnu)
Quels autres facteurs limitatifs sont pertinents? L’espèce est exposée à des menaces dans ses habitats à l’extérieur du Canada, notamment sur sa route migratoire qui traverse les États-Unis et dans les sites d’hivernage au Mexique et en Californie.
Immigration de source externe (immigration de l’extérieur du Canada)
| Éléments du résumé technique | information |
|---|---|
| Situation des populations de l’extérieur les plus susceptibles de fournir des individus immigrants au Canada | En déclin. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Oui, possible. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui, durant l’été. |
| Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants? | Oui. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
Oui Si on suppose que les diminutions de l’abondance des asclépiades observées aux États-Unis s’appliquent également au Canada. |
| Les conditions de la population source se détériorent-elles? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
Oui, d’après les diminutions de l’abondance des asclépiades observées aux États-Unis et les déclins de l’habitat dans les sites d’hivernage au Mexique et en Californie. |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
Non |
| La possibilité d’une immigration depuis des populations externes existe-t-elle? | Probablement pas. Les populations ne migrent pas ailleurs dans le monde, mais il est possible que des individus errants recolonisent les populations nord-américaines. |
Nature délicate de l’information sur l’espèce
| Éléments du résumé technique | information |
|---|---|
| L’information concernant l’espèce est-elle de nature délicate? | Non |
Historique du statut
Statut et justification de la désignation :
| Éléments du résumé technique | information |
|---|---|
| Statut : | En voie de disparition |
| Code alphanumérique : | A2bce |
| Justification de la désignation : | Cette grande espèce remarquable est un des papillons les plus connus dans le monde. La population canadienne est migratrice et elle emprunte deux voies distinctes. Des menaces cumulatives pèsent sur les sites d’hivernage et le long des voies de migration. Le groupe migrateur à l’ouest des montagnes Rocheuses se déplace entre la côte de la Californie et le sud de la Colombie-Britannique. Le groupe à l’est des montagnes Rocheuses représente la vaste majorité de la population canadienne et se déplace entre la forêt de sapin oyamel du centre du Mexique et le sud du Canada à l’est de l’Alberta. Les sites d’hivernage dans le centre du Mexique sont extrêmement petits, et les menaces qui pèsent sur ces aires incluent l’exploitation forestière illégale et le développement agricole, ainsi qu’une augmentation de la fréquence et de la gravité des tempêtes pendant les périodes clés de rassemblement. Des déclins de plus de 50 % se sont produits au cours de la dernière décennie. |
Applicabilité des critères :
| Éléments du résumé technique | information |
|---|---|
| Critère A (déclin du nombre total d’individus matures) : |
Correspond au critère de la catégorie « en voie de disparition », A2bce, car un déclin de l’effectif estimatif a été observé dans les sites d’hivernage du centre du Mexique (b) et un déclin de la qualité de l’habitat a été observé dans les sites d’hivernage et tout le long des voies migratoires (c) et à cause des polluants (e). |
| Critère B (petite aire de répartition, et déclin ou fluctuation) : |
Sans objet. Correspond presque au critère relatif au faible IZO et au déclin de la qualité de l’habitat si l’on se fie au site d’hivernage, mais le nombre de localités est élevé et la population de l’espèce n’est pas gravement fragmentée. |
| Critère C (nombre d’individus matures peu élevé et en déclin) : |
Sans objet. On ne dispose pas de suffisamment de données pour estimer le déclin du nombre total d’individus matures et du nombre de sous-populations. |
| Critère D (très petite population totale ou répartition restreinte) : |
Sans objet. On ne dispose pas de suffisamment de données sur le nombre d’individus matures que compte la population. |
| Critère E (analyse quantitative) : |
Sans objet. On ne dispose pas de suffisamment de données pour faire une analyse quantitative. |
Préface
Le monarque a été évalué pour la première fois par le COSEPAC en avril 1997 et a alors été désigné « espèce préoccupante ». Son statut a été réexaminé et confirmé en novembre 2001 et en avril 2010. Depuis la publication du rapport de situation précédent (2010), les estimations de la population effectuées dans les sites d’hivernage du Mexique donnent à penser que le déclin du monarque s’est poursuivi. Les recherches sur le monarque ont permis des avancées majeures des connaissances relatives aux tendances en matière d’habitat, à la dynamique de la population à différents stades du cycle de migration et à la biologie générale de l’espèce. La mise au point de plantes cultivées résistantes aux herbicides et les effets de l’utilisation accrue des herbicides sur les asclépiades et autres plantes nectarifères représentent une menace nouvelle, tout comme le changement climatique. Des modèles de viabilité de la population et de quasi-extinction ont également été créés. Un grand volume de recherches et de publications portant sur le monarque était encore en cours au moment de la publication du rapport. Une mise à jour du rapport de situation a donc été entreprise avant la fin de la période de dix ans prévue pour la réévaluation, d’après cette information additionnelle.
Historique du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Mandat du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Composition du COSEPAC
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Définitions (2016)
- Espèce sauvage
- Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d’animal, de plante ou d’un autre organisme d’origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s’est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
- Disparue (D)
- Espèce sauvage qui n’existe plus.
- Disparue du pays (DP)
- Espèce sauvage qui n’existe plus à l’état sauvage au Canada, mais qui est présente ailleurs.
-
En voie de disparition (VD)
(Remarque : Appelée « espèce disparue du Canada » jusqu’en 2003.) - Espèce sauvage exposée à une disparition de la planète ou à une disparition du pays imminente.
- Menacée (M)
- Espèce sauvage susceptible de devenir en voie de disparition si les facteurs limitants ne sont pas renversés.
-
Préoccupante (P)
(Remarque : Appelée « espèce en danger de disparition » jusqu’en 2000.) - Espèce sauvage qui peut devenir une espèce menacée ou en voie de disparition en raison de l'effet cumulatif de ses caractéristiques biologiques et des menaces reconnues qui pèsent sur elle.
-
Non en péril (NEP)
(Remarque : Appelée « espèce rare » jusqu’en 1990, puis « espèce vulnérable » de 1990 à 1999.) - Espèce sauvage qui a été évaluée et jugée comme ne risquant pas de disparaître étant donné les circonstances actuelles.
-
Données insuffisantes (DI)
(Remarque :Autrefois « aucune catégorie » ou « aucune désignation nécessaire ».) - Une catégorie qui s’applique lorsque l’information disponible est insuffisante (a) pour déterminer l’admissibilité d’une espèce à l’évaluation ou (b) pour permettre une évaluation du risque de disparition de l’espèce.
Le Service canadien de la faune d’Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Description et importance de l’espèce sauvage
Nom et classification
Règne : Animalia – Animaux
Embranchement : Arthropoda – Arthropodes
Sous-embranchement : Hexapoda – Hexapodes
Classe : Insecta – Insectes
Sous-classe : Pterygota – Ptérygotes, ou insectes ailés
Ordre : Lepidoptera – Lépidoptères ou papillons (diurnes et nocturnes)
Famille : Nymphalidae Rafinesque 1815 – Nymphalidés
Sous-famille : Danainae Boisduval, 1833 – Danaïnés
Genre : Danaus Kluk 1780
Sous-genre : Danaus (Danaus) Kluk 1780
Espèce : Danaus plexippus(Linnaeus 1758)
Nom français : Monarque
Nom anglais : Monarch
Le monarque (Danaus plexippus (L.)) était autrefois classé dans la famille des Danaïdés, mais il fait maintenant partie de la sous-famille des Danaïnés, groupe diversifié auquel appartiennent également les satyres, les nordiques, les morphos, les papillons-hiboux et les papillons longs-palpes (Opler et Warren, 2002). Le monarque est le seul représentant canadien de la sous-famille des Danaïnés (Marshall, 2006).
Le Danaus plexippus compte six sous-espèces : le Danaus plexippus plexippus est présent en Amérique du Nord, alors que les autres sous-espèces se rencontrent dans les Caraïbes, en Amérique centrale et en Amérique du Sud (Opler et Warren, 2002). Deux autres membres du genre Danaus, le D. gilippus et le D. eresimus se reproduisent dans le sud des États-Unis (Scott, 1986).
Dans le présent rapport, les individus présents à l’est des Rocheuses (population de l’est) et ceux présents à l’ouest des Rocheuses (population de l’ouest) font souvent l’objet d’analyses distinctes. En effet, les populations de l’est et de l’ouest ont des aires de reproduction et des aires d’hivernage distinctes (voit les sections Aire de répartition mondiale et Aire de répartition canadienne), mais des preuves génétiques confirment que les deux sous-groupes se mélangent et appartiennent à la sous-espèce plexippus (Lyons et al., 2012) (voir la section Structure spatiale et variabilité de la population).
Description morphologique
Le cycle vital du monarque comprend quatre stades distincts : l’œuf, la chenille, la chrysalide et l’adulte.
Adulte
Le monarque est un superbe papillon diurne orange et noir de grande taille (figure 1). Les ailes, d’une envergure de 93 à 105 mm, sont orange vif et comportent des nervures noires marquées et une large bordure noire piquée de deux rangées de points circulaires blancs (Layberry et al., 1998). Chez les mâles, chacune des ailes postérieures porte sur la nervure cubitale une tache noire distincte constituée d’écailles androconiales; cette tache fait défaut chez les femelles (Layberry et al., 1998).
Œuf
L’œuf du monarque mesure environ 1 mm de longueur et est ovale, avec la base aplatie et le sommet formant une pointe arrondie. Environ 18 crêtes le parcourent de la base au sommet. L’œuf est blanchâtre juste après la ponte, mais devient plus foncé avec le temps. Les œufs sont pondus individuellement.
Chenille
La chenille (larve) (figure 2) est annelée de blanc, de jaune et de noir. Elle est pourvue d’une paire de filaments noirs près de sa tête et d’une autre paire à l’extrémité de son abdomen. Au dernier stade larvaire, la chenille peut mesurer jusqu’à 5 cm de longueur (Scott, 1986; Schappert, 2004).
Chrysalide
La chrysalide (nymphe) est verte avec une bande noire et une bande dorée à la jonction de l’abdomen et du thorax et avec des points dorés épars sur le reste de sa surface (Scott, 1986; Schappert, 2004). Elle pend d’une branche ou d’un autre substrat auquel elle est fixée par un coussinet de soie tissé par la chenille avant la nymphose (Schappert, 2004).
L’aire de répartition du vice-roi (Limenitis archippus) chevauche la majeure partie de celle du monarque au Canada, et les deux espèces se ressemblent (Layberry et al., 1998). Le vice-roi est toutefois plus petit que le monarque et s’en distingue par une ligne noire traversant les nervures des ailes postérieures (Scott, 1986; Layberry et al., 1998).

Description longue pour la figure 1
Photo d’un monarque adulte, en vue latérale, parmi des fleurs. Ce superbe et grand papillon possède des ailes principalement orange ornées de fortes nervures noires et d’une large bordure noire piquée de deux rangées de points circulaires blancs.

Description longue pour la figure 2
Photo d’une chenille de monarque sur une feuille. La chenille est annelée de blanc, de jaune et de noir. Elle est pourvue d’une paire de filaments noirs près de sa tête et d’une autre paire près de l’extrémité de son abdomen.

Description longue pour la figure 3
Photo d’une nuée de monarques se rassemblant dans un site d’hivernage, au Mexique.
Structure spatiale et variabilité des populations
Le monarque est une espèce migratrice, et sa vaste répartition, ses voies migratoires et ses sites d’hivernage ont soulevé beaucoup de questions, de sorte que de nombreuses recherches ont été menées sur les lignées génétiques de l’espèce et son histoire évolutionnaire (résumé récent dans Pierce et al., 2015). Il avait initialement été avancé, selon les données obtenues à l’aide des méthodes de marquage de Urquhart et Urquhart (1977), que les populations de l’est et de l’ouest utilisaient des voies migratoires et des aires d’hivernage différentes, mais aucune donnée génétique n’avait été recueillie. Toutefois, depuis ce modèle initial (Urquhart et Urquhart, 1977), des analyses génétiques, des données provenant du marquage de spécimens et des observations faites sur le terrain indiquent que ces voies migratoires ne sont pas bien définies et sont plus complexes qu’on le croyait, et que des échanges se produisent entre les populations de l’est et de l’ouest durant la migration (analyse de Pierce et al., 2015, et de Pyle 2015). Un nouveau modèle, plus complexe, est maintenant proposé (Brower et Pyle, 2004; Dingle et al., 2005; Pyle, 2015), mais il faut encore mener des recherches pour expliquer les échanges, dont l’ampleur est inconnue, entre les individus des sites d’hivernage de la Californie et du Mexique.
Un résumé des vérifications sur le terrain appuyant l’existence d’échanges d’individus entre les populations de monarque de l’est et de l’ouest, notamment de vastes études de marquage, est présenté dans Pyle (2015). Dans le cadre d’une étude, intitulée « Southwest Monarch Study », des individus sauvages capturés et des individus élevés en captivité ont été marqués puis relâchés en Arizona; certains de ces individus ont été retrouvés dans les sites d’hivernage de la Californie, et d’autres dans ceux du Mexique. Les données obtenues pour les monarques sauvages uniquement montrent clairement que les monarques capturés à l’ouest de la ligne continentale de partage des eaux migraient aussi bien vers les sites d’hivernage du Mexique que vers ceux de la Californie (Southwest Monarch Study, 2013, dans Pyle, 2015).
Il a été impossible de distinguer des lignées génétiques entre les populations de l’est et de l’ouest, probablement parce qu’il y a des croisements durant la migration, les individus provenant d’une vaste région géographique se mélangeant à cette occasion. La première étude génétique portant sur l’espèce était fondée sur des allozymes marqueurs (variantes enzymatiques de certaines protéines qui représentent une variation génétique) (Eanes et Koehn, 1978). Cette étude a révélé que les différences génétiques détectées entre les sites de reproduction étaient effacées durant la migration annuelle, car les monarques des différents sites prémigratoires se mélangeaient et se reproduisaient de manière aléatoire durant la migration; ce phénomène explique donc les faibles différences génétiques observées entre les populations de l’est et de l’ouest (Eanes et Koehn, 1978).
L’ADN mitochondrial (ADN qui se trouve dans les mitochondries et est transmis par le parent femelle) de 28 spécimens a été utilisé pour comparer les individus des populations migratrices de l’est et de l’ouest avec les individus des néotropiques (Brower et Boyce, 1991). Aucune différence n’a été observée quant à la taille du génome mitochondrial entre les individus ou chez un même individu, et un seul polymorphisme (révélé par digestion par l’enzyme de restriction Ase1) a été trouvé; toutefois, ce polymorphisme a été détecté chez des spécimens provenant des deux sites d’hivernage (Brower et Boyce, 1991), et il n’indique donc pas de différences entre les individus qui migrent vers les sites d’hivernage de l’est et de l’ouest. Des profils génétiques presque identiques ont été observés dans toutes les populations, les autres fragments de restriction étant tous monomorphes chez tous les spécimens analysés (Brower et Boyce, 1991).
Dans le cadre de la plus récente étude génétique, des marqueurs microsatellites (courtes séquences d’ADN répétées en tandem) ont été utilisés pour comparer cinq échantillons : un de Floride, dans l’est de l’Amérique du Nord, deux de Californie, dans l’ouest de l’Amérique du Nord, ainsi qu’un des populations non migratrices d’Hawaii et de Nouvelle-Zélande (Lyons et al., 2012). La valeur FST (proportion de la variation génétique totale attribuable à des différences entre les populations) obtenue par comparaison des échantillons nord-américains était de 0,0012, ce qui est très faible et non statistiquement différent de 0 (P = 0,2). Lyons et al. (2012) en ont donc conclu que les monarques d’Amérique du Nord constituent une seule population présentant un mélange des gènes.
Unités désignables
Le monarque est considéré comme formant une seule unité désignable au Canada aux fins de la présente évaluation. Des données génétiques et des informations recueillies sur le terrain indiquent que les individus des populations de l’est et de l’ouest se mélangent (voir la section Structure spatiale et variabilité de la population). En outre, aucune différence morphologique, comportementale ou génétique moléculaire n’a été constatée entre les individus des populations de l’est et de l’ouest.
Le monarque est présent dans la portion méridionale des aires écologiques du COSEPAC (COSEWIC, 2017) de l’Atlantique, boréale, des plaines des Grands Lacs, du Pacifique, des Prairies et des montagnes du Sud.
Importance de l’espèce
Le monarque est une espèce emblématique de la conservation et compte parmi les espèces de papillons les plus connues et les plus étudiées dans le monde (Gustafsson et al., 2015). La migration du monarque depuis le sud du Canada jusqu’au Mexique, en passant par la partie continentale des États-Unis, a été décrite comme un phénomène biologique en péril (Brower, 1996). Partout en Amérique du Nord, le monarque est utilisé dans les écoles pour enseigner aux enfants la biologie et l’histoire naturelle, les cycles de vie et la métamorphose et la conservation et pour leur faire apprécier la nature.
Répartition
Aire de répartition mondiale
L’aire de répartition mondiale du monarque s’étend depuis le sud du Canada jusque dans le nord de l’Amérique du Sud, en passant par les États-Unis et l’Amérique centrale, et de l’Atlantique au Pacifique. Le monarque est fréquemment signalé à l’extérieur de son aire de répartition indigène, phénomène souvent associé à des individus élevés en captivité puis relâchés ainsi qu’à des individus errants. On ne considère pas que ces observations font partie de l’aire de répartition naturelle de l’espèce.
Six sous-espèces de monarques ont été identifiées : le D. p. plexippus (Amérique du Nord et toutes les autres régions où l’espèce a été signalée à l’extérieur des Amériques [sous-espèce évaluée dans le cadre du présent rapport de situation]), le D. p. leucogyne (îles Vierges), le D. p. portoricensis (Porto Rico), le D. p. tobagi (Tobago), le D. p. megalippe (Caraïbes et Amérique centrale; Smith et al., 1994) et le D. p. nigrippus (Amérique du Sud) (Opler et Warren, 2002; Pelham, 2008; NatureServe, 2015) (figure 4). Selon certaines études génétiques, une partie des populations, voire toutes les populations, des îles des Caraïbes et d’Amérique du Sud appartiendraient en fait à la même sous-espèce (p. ex. Smith et al., 2005). Contrairement à la sous-espèce des populations plus septentrionales, la sous-espèce d’Amérique centrale et des Caraïbes est relativement sédentaire et ne contribue pas à la population migratrice du Canada et des États-Unis (Crolla et Lafontaine, 1996). Les recherches génétiques montrent que des individus des populations migratrices se sont dispersés à l’extérieur de l’Amérique du Nord et sont les ancêtres des sous-espèces non migratrices des Caraïbes, d’Amérique centrale et d’Amérique du Sud (Zhan et al., 2014). En outre, certains éléments semblent indiquer que les populations migratrices sont distinctes sur le plan génétique des populations non migratrices (Zhan et al., 2014).
Des individus errants ont apparemment traversé l’Atlantique au milieu ou à la fin du 19e siècle et ont atteint des îles de l’est de l’Atlantique et l’Europe continentale (Fernandez-Haeger et al., 2015). Des populations sont maintenant établies en Afrique du Nord, en Espagne, au Portugal, dans les Açores, dans les îles Canaries ainsi que dans d’autres îles (Fernandez-Haeger et al., 2015). La façon dont le monarque a atteint l’Autralie est inconnue, mais l’espèce y a été signalée pour la première fois vers 1870 et s’est dispersée dans tout l’est et dans certaines portions de l’est du pays (Zalucki et Rochester, 2004). Le monarque a apparemment colonisé Hawaii, la Nouvelle-Zélande et les grandes îles du Pacifique (Galapagos, îles Salomon, Norfolk, Philippines et Taïwan) après 1940 (Vane-Wright, 1993).
Description longue pour la figure 4
Carte de l’aire de répartition américaine (indigène) du monarque, montrant la répartition approximative des six sous-espèces. Le monarque est présent depuis le sud du Canada jusque dans le nord de l’Amérique du Sud, en passant par les États-Unis et l’Amérique centrale, et le l’Atlantique au Pacifique.
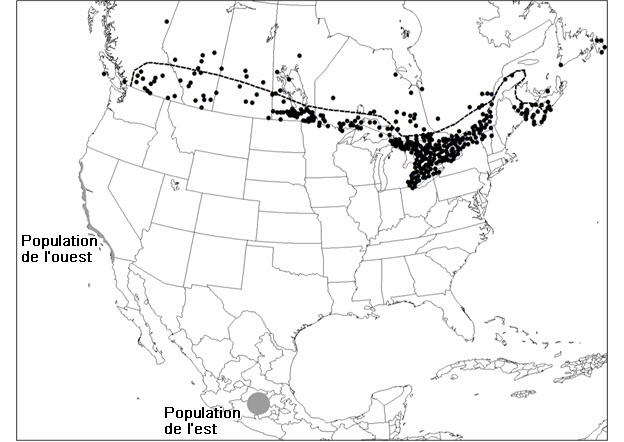
Description longue pour la figure 5
Carte de la répartition du monarque au Canada, où l’espèce a été signalée dans les 10 provinces et dans les Territoires du Nord-Ouest. La limite nord approximative des plantes hôtes de la chenille de l’espèce (asclépiade incarnate, asclépiade commune et belle asclépiade) est indiquée.
L’aire de reproduction de la population de l’est s’étend à l’est des Rocheuses, depuis la côte du golfe du Mexique jusque dans le sud du Canada, et les individus migrent vers le centre du Mexique pour y hiverner (Urquhart, 1987). L’aire de reproduction de la population de l’ouest s’étend depuis le sud-ouest des États-Unis (Arizona et Nouveau-Mexique) jusque dans le sud du Canada (Colombie-Britannique) et depuis les Rocheuses jusqu’à la côte du Pacifique. Les individus de cette population hivernent le long de la côte de la Californie, et un petit nombre d’individus poussent leur voyage plus au sud, jusque dans le nord du Mexique (Lane, 1984).
Les populations du sud de la Floride et de Cuba sont non migratrices, mais certains individus migrants des populations du nord passent par la Floride durant leur migration et s’assimilent aux populations reproductrices résidentes (Dockx et al., 2004; Knight et Brower, 2009). La descendance de ces individus ne migre apparemment pas vers le nord au printemps suivant. Contrairement aux individus qui hivernent au Mexique et en Californie, les individus résidents du sud de la Floride ne se regroupent pas pour hiverner et n’entrent pas en diapause. Ils sont actifs durant toute l’année et se reproduisent de façon continue (Knight et Brower, 2009).
Aire de répartition canadienne
Au Canada, le monarque a été signalé dans les dix provinces ainsi que dans les Territoires du Nord-Ouest (Layberry et al., 1998; Environment and Climate Change Canada, 2016) (figure 5). L’aire de reproduction de l’espèce au Canada se situe au sud du 50e parallèle en Ontario, au Québec et dans les Maritime (Nouveau-Brunswick, Terre-Neuve-et-Labrador, Nouvelle-Écosse et Île-du-Prince-Édouard) et au sud du 54e parallèle au Manitoba, en Saskatchewan et en Alberta. L’aire de répartition continue de l’espèce correspond à celle de ses plantes hôtes, les asclépiades (Asclepias spp.) (Brower, 1996; Crolla et Lafontaine, 1996). En Nouvelle-Écosse, l’aire de reproduction de l’espèce se limite presque uniquement à la vallée de l’Annapolis (Crolla et Lafontaine, 1996) (figure 5). Les individus signalés au nord et à l’est de l’aire de répartition indigène des asclépiades représentent des individus non reproducteurs errants et des individus se reproduisant dans des parcelles où des asclépiades ont été plantées. Des individus errants ont été observés à Terre-Neuve-et-Labrador et dans les Territoires du Nord-Ouest, ainsi que dans le nord d’autres provinces (Environment and Climate Change Canada, 2016).
Les plus importantes aires de reproduction du monarque au Canada se trouvent dans le sud de l’Ontario et du Québec (White, 1996). Le parc provincial Long Point et les milieux environnants, le parc provincial Rondeau et le parc national de la Pointe-Pelée, sur la rive nord du lac Érié, et le parc provincial Presqu’ile, en bordure du lac Ontario, sont d’importants lieux où se rassemblent les monarques avant leur traversée des Grands Lacs. Le monarque est commun dans le sud du Québec, mais il est rarement observé au-delà du 50e parallèle. En automne, les monarques en migration se rassemblent en grand nombre dans des régions précises du sud de la province, en particulier le long de la baie de Valleyfield et de la baie Missisquoi (Handfield et al., 1999).
En Alberta, en Saskatchewan et au Manitoba, le monarque est principalement présent dans le sud, et son abondance diminue vers le nord et vers l’est, du sud du Manitoba vers l’Alberta (Klassen et al., 1989, Layberry et al., 1998). Certaines années, la reproduction du monarque a été observée au nord jusqu’à Edmonton, en Alberta. L’espèce est plutôt commune dans le sud du Manitoba, sauf dans sa portion sud-est, en large partie boisée et comportant donc moins d’habitat propice à l’espèce (Klassen et al., 1989).
En Colombie-Britannique, la reproduction de l’espèce a été observée dans des sites épars dans la portion sud de la province, particulièrement dans les vallées de l’Okanagan et de la Similkameen et dans les Kootenays (sud-est) (Guppy et Shepard, 2001; St. John et al., 2014; B.C. Conservation Data Centre, 2016). Des individus errants ont été signalés dans la vallée du bas Fraser et dans l’île de Vancouver, mais ces régions ne font pas partie de l’aire de répartition indigène des asclépiades et ne sont donc pas considérées comme faisant partie de l’aire de répartition indigène du monarque.
Environ 10 % de l’aire de reproduction mondiale du monarque se trouvent au Canada. Cette évaluation est fondée sur les observations des chenilles de l’espèce le long de la frontière internationale, depuis les Maritimes jusque dans le sud de la Colombie-Britannique.
Aucune connaissance traditionnelle autochtone liée à la répartition ou à l’abondance du monarque au Canada n’était connue au moment de la rédaction du présent rapport de situation.
Zone d’occurrence et zone d’occupation
Au Canada, la zone d’occurrence du monarque (excluant les mentions d’individus errants et des mentions d’individus se reproduisant dans des colonies d’asclépiades situées dans des jardins, à l’extérieur de leur aire de répartition historique) correspond à la répartition des trois principales plantes hôtes de la chenille de l’espèce : l’asclépiade incarnate (Asclepias incarnata), l’asclépiade commune (Asclepias syriaca) et la belle asclépiade (Asclepias speciosa) (White, 1996). La superficie délimitée par un polygone convexe comprenant l’aire de répartition indigène de ces trois espèces d’asclépiades au Canada (figure 5) s’élève à plus de 3,5 millions de km2, et ce polygone représente approximativement la zone d’occurrence des populations de l’est et de l’ouest du monarque. Compte tenu du comportement migratoire du monarque et de l’étendue de sa dispersion annuelle vers le nord, qui diffère selon les conditions météorologiques, la zone d’occurrence au Canada fluctue considérablement d’une année à l’autre.
L’indice de zone d’occupation (IZO) au Canada, généralement établi à partir d’une grille à carrés de 2 km de côté recoupant les occurrences reproductrices, n’a pas été calculé à cause du manque de renseignements détaillés concernant la répartition des sites de reproduction dans l’ensemble du pays. Néanmoins, il est certain que l’IZO est supérieur à 2 000 km2.
L’IZO de l’aire d’hivernage de la population de l’est a été estimé d’après le nombre de colonies qui hivernent au Mexique. En 2015, la zone d’hivernage comprenait neuf colonies, occupant chacune une superficie de 0,13 ha à 1,09 ha (anonyme, 2016). Si on suppose que chaque colonie occupait un seul carré de 2 km de côté, l’IZO maximal en 2015 était de 36 km2.
L’IZO de l’aire d’hivernage de la population de l’ouest, en Californie, comprenait 138 sites en 2015 (Monroe et al., 2016) et s’élevait au maximum à 552 km2.
L’IZO total pour les sites d’hivernage des populations de l’ouest et de l’est en 2015 était donc de 588 km2.
Activités de recherche
La répartition du monarque au Canada a pu être déterminée grâce aux efforts de collectionneurs, de chercheurs et de citoyens-chercheurs. Un sommaire de la répartition et de l’abondance du monarque à l’échelle nationale a été présenté dans Layberry et al. (1998), et ce type de sommaire a également été préparé à l’échelle régionale pour la Colombie-Britannique (Guppy et Shepard, 2001; B.C. Conservation Data Centre, 2016), l’Alberta (Bird et al., 1995), le Manitoba (Klassen et al., 1989), l’Ontario (Holmes et al., 1991, Macnaughton et al., 2016), le Québec (Handfield et al., 1999), le Nouveau-Brunswick (Thomas, 1996) et les Maritimes (Maritimes Butterfly Atlas, 2016), et dans le plan national de rétablissement de l’espèce (Environment and Climate Change Canada, 2016).
La Toronto Entomologists’ Association a compilé les observations annuelles de papillons effectuées depuis 1968 pour tout l’Ontario (Macnaughton, 2010), y compris celles provenant des rapports fréquents sur la situation du monarque (population de l’est) (p. ex. Hess, 1992). La version en ligne de l’Ontario Butterfly Atlas (Macnaughton et al., 2016) comprend 280 000 mentions de papillons provenant des sommaires annuels de la Toronto Entomologists’ Association et de la plateforme eButterfly.
Les mentions du monarque en Colombie-Britannique (population de l’ouest) correspondent principalement à des observations fortuites ou à des observations faites par des naturalistes ou des biologistes, et peu de relevés ciblant l’espèce ont été réalisés dans la province, voire aucun (St. John et al., 2014). Un relevé ciblé a été effectué dans les vallées de l’Okanagan et de la Similkameen en 2014, dans le cadre duquel 758 sites où pousse la belle asclépiade ont été visités sur 37 jours, du milieu à la fin de l’été (St. John et al., 2014). En 2014, les œufs d’une femelle gravide ont été élevés en captivité, et les adultes issus de ces œufs ont été marqués et relâchés en vue de produire des données sur l’histoire naturelle des populations de monarques dans le sud de la vallée de l’Okanagan (Antonijevic et al., 2014).
Le monarque bénéficie d’une popularité générale et est facile à observer et à identifier, ce qui le rend propice à un suivi par le public (Commission for Environmental Cooperation, 2009). Il existe plusieurs projets continus de science citoyenne liés aux populations canadiennes de monarque, dont Monarch Watch (marquage, migration automnale), Mission monarque (dénombrement des œufs et des chenilles), Journey North (observations, suivi de la migration), le Monarch Caterpillar Monitoring Project (recensement des œufs et des chenilles), la North American Butterfly Association (recensements annuels), eButterfly (photos et observations), le Maritime Butterfly Atlas et l’Ontario Butterfly Atlas (Howard et Davis, 2015; Oberhauser et al., 2015a; Macnaughton et al., 2016; Maritimes Butterfly Atlas, 2016).
Habitat
Besoins en matière d’habitat
Le monarque a besoin de différents types d’habitat selon le stade de son cycle vital. Les habitats de reproduction et de migration décrits ci-dessous sont ceux de l’espèce au Canada. Les habitats d’hivernage se situent au Mexique (population de l’est) et en Californie (population de l’ouest).
Habitat de reproduction
Au Canada, la chenille du monarque se nourrit exclusivement sur les asclépiades (Asclepias spp.), et l’habitat de reproduction est donc limité aux endroits où les plantes de ce genre poussent. Le monarque utilise 11 des 14 espèces d’asclépiades présentes au Canada (White, 1996; Borders et Lee-Mader, 2014). Toutefois, au Canada, les espèces les plus couramment utilisées par le monarque sont la belle asclépiade en Colombie-Britannique, en Alberta, en Saskatchewan et au Manitoba ainsi que l’asclépiade commune et l’asclépiade incarnate en Ontario et dans l’est du Canada. Les asclépiades poussent dans une grande variété de milieux, notamment en bordure des routes ainsi dans les champs, les milieux humides, les prés, les prairies et les forêts dégagées (Borders et Lee-Mader, 2014).
Des asclépiades sont souvent plantées dans des jardins à l’extérieur de leur aire de répartition naturelle, ce qui peut étendre l’aire de reproduction du monarque. On observe généralement chez les asclépiades qui poussent dans les champs de soja et de maïs des densités d’œufs et des taux de survie des œufs et des chenilles des premiers stades larvaires plus élevés que chez les asclépiades qui poussent dans les milieux non agricoles, peut-être parce que la prédation est moindre et que les feuilles présentent une meilleure appétibilité en milieu agricole (Pleasants, 2015). Selon les estimations, 80 % des individus présents dans les États du Midwest se sont nourri sur des asclépiades poussant en milieu agricole (Pleasants, 2015).
Les adultes se nourrissent du nectar des fleurs d’asclépiade, mais ils ont besoin d’autres espèces de plantes nectarifères sauvages, en particulier durant les périodes de l’année où les asclépiades ne sont pas en fleurs (Crolla et Lafontaine, 1996). Au Canada, les verges d’or (Solidago spp.), les asters (Doellingeria spp., Eurybia spp., Oclemena spp., Symphyotrichum spp. et Virgulus spp.), la salicaire commune (Lythrum salicaria), espèce introduite, et diverses espèces de trèfles (Trifolium spp.) et de mélilot (Melilotus spp.) sont les autres sources de nectar les plus utilisées par le monarque (Crolla et Lafontaine, 1996). Les sources de nectar sont particulièrement importantes durant la migration automnale, l’espèce convertissant les sucres du nectar en réserves de graisses nécessaires à sa survie en hiver (Urquhart, 1987; Gibo et McCurdy, 1993).
Habitat de migration ou de rassemblement
Le monarque a besoin de milieux où se reposer, s’alimenter et s’abriter des intempéries durant sa migration. Au Canada, on trouve des aires de rassemblement le long des rives nord des Grands Lacs, où les monarques se reposent sur les arbres avant de traverser ces vastes étendues d’eau libre. Les principales aires se trouvent dans le parc provincial Presqu’ile, le parc provincial Point Pelee ainsi que les régions de Rondeau et de Long Point. Le monarque possède une bonne capacité d’adaptation quant aux caractéristiques des aires de repos, mais il choisit apparemment ses aires de rassemblement en fonction de leur proximité de grands lacs, de la taille des arbres et de la direction du vent (Schappert, 2004; Davis et al., 2012). En quelques heures, des dizaines à des centaines de milliers d’individus ont été observés en trait de quitter l’extrémité de la pointe Pelée, apparemment après s’y être reposés pendant la nuit (Wormington, 1997).
Habitat d’hivernage
Durant son hivernage, le monarque a besoin d’un microclimat frais, mais il doit également être protégé du gel saisonnier. Il a également besoin d’un taux d’humidité élevé et d’une protection contre l’exposition excessive au soleil, le vent et les précipitations (Williams et Brower, 2015). Il trouve ces conditions en Californie, dans la région de la côte du Pacifique, et dans le centre du Mexique, dans les forêts de haute altitude.
Les individus de la population de l’est passent l’hiver dans les forêts de sapin oyamel (Abies religiosa) de haute altitude du centre du Mexique (Williams et Brower, 2015). Ces forêts se situent à 2 900 à 3 300 m au-dessus du niveau de la mer et ont une étendue d’environ 80 km sur 80 km. Les parcelles de forêt convenant à l’espèce dans cette étendue occupent au total environ 562 km2(Slayback et al., 2007). Les colonies hivernantes ont été observées dans les zones de la forêt qui présentaient un couvert plus dense que d’autres zones non occupées choisies au hasard (Williams et al., 2007). Le couvert forestier protège les individus des précipitations et des vents, des températures inférieures au point de congélation ainsi que des températures chaudes, ce qui permet aux individus de se maintenir dans un état de métabolisme ralenti (Williams et Brower, 2015).
Les individus de la population de l’ouest hivernent dans un secteur s’étendant sur environ 1 000 km le long de la côte de la Californie, jusque dans le nord de la Basse-Californie. Environ 450 sites d’hivernage ont été recensés le long de cette portion de côte (Jepsen et Black, 2015), la plupart étant associés à des peuplements d’eucalyptus (Eucalyptus globalus et E. camaldulensis), arbres exotiques provenant d’Australie (Frey et Schaffner, 2004; Jepsen et Black, 2015). L’eucalyptus a été introduit dans les années 1850 à des fins ornementales ou comme brise-vents ou source de combustibles, ce qui a conduit à une augmentation des sites d’hivernage propices (Crolla et Lafontaine, 1996). La croissance des populations d’eucalyptus a coïncidé avec l’élimination des peuplements côtiers denses d’essences indigènes comme le pin de Monterey (Pinus radiata) et le cyprès de Monterey (Cupressus macrocarpa). Les quelques peuplements qui n’ont pas été détruits sont encore utilisés comme sites d’hivernage par le monarque (Lane, 1993; Jepsen et Black, 2015).
En Californie, les sites d’hivernage sont associés à un climat maritime et se trouvent généralement à moins de 2,4 km de la côte, secteur où les températures descendent rarement sous le point de congélation (températures moyennes en janvier de 10 à 12 °C) mais demeurent suffisamment fraîches pour permettre aux monarques de conserver leur énergie. En outre, le taux d’humidité est élevé dans ces sites et on y observe une rosée matinale, ce qui protège les individus de la déshydratation. La plupart des sites se trouvent sur des pentes orientées vers le sud ou l’ouest, qui reçoivent donc un bon rayonnement solaire et sont protégées du vent (Leong et al., 2004).
Tendances en matière d’habitat
Habitat d’hivernage
L’étendue historique de l’habitat d’hivernage de la population de l’est dans les forêts de sapin oyamel, au Mexique, n’est pas bien documentée, mais on sait qu’avant 1971 la majeure partie de la région était occupée par une forêt dense et continue (Brower et al., 2002). De 1971 à 1999, 44 % de la forêt dense (couvert d’une densité de > 80 %) a subi une coupe à blanc, a été éclaircie ou a été brûlée. La plus grande parcelle de forêt de haute qualité est passée de 271 km2 à 58 km2 (Brower et al., 2002).
La Réserve de la biosphère du monarque a été créée en 1980, et ses limites ont été établies en 1986. Elle comprend une zone centrale de 130 km2 ainsi qu’une zone tampon de plus de 420 km2 (Jepsen et al., 2015). De 1986 à 2012, un total de 4 300 ha de forêt de sapin oyamel ont été détruits ou ont subi des perturbations (8 % de la réserve) à cause de l’exploitation forestière illégale, des incendies, de la récolte de bois à usage domestique et du défrichage agricole (Ramirez et al., 2015). Le taux d’exploitation forestière illégale a augmenté tout juste après la création de la réserve, mais il a ensuite diminué de 2005-2007 (731 ha) à 2014 (5,2 ha) (Vidal et Rendón-Salinas, 2014). Une exploitation illégale a été effectuée sur 19 ha dans l’État du Michoacán en 2014-2015 (WWF, 2015).
L’habitat d’hivernage a également subi un déclin en Californie. Au moins 38 sites d’hivernage du monarque ont été détruits par la construction d’ensembles résidentiels avant 1991. Onze autres sites ont été détruits par cette même activité dans les années 1990, et deux sites supplémentaires sont actuellement destinés à la construction résidentielle (Jepsen et Black, 2015). De plus, certains sites ont été dégradés par l’élagage des arbres, l’élimination des eucalyptus et la mort de vieux arbres (Frey et Schaffner, 2004; Jepsen et Black, 2015).
Des modèles climatiques prédisent que les milieux qui présentent des conditions climatiques propices au sapin oyamel auront diminué de 50 % d’ici 2030 et que les forêts de sapin oyamel auront complètement disparu d’ici 2090, car les effets des ravageurs et des maladies s’accentueront avec la hausse du stress hydrique (Ramirez et al., 2015) (voir la section Menaces, point 11.1).
Habitat de reproduction
Dans de nombreuses parties du Canada, l’habitat de reproduction du monarque a subi de considérables changements au cours des 200 dernières années. Les asclépiades sont adaptées aux milieux dégagés et aux sols perturbés, et, jusque dans les années 1880, les prairies du centre de l’Amérique du Nord représentaient la principale aire de reproduction de la population de l’est (Brower, 1995). Il y avait également de vastes étendues de prairie et de savane dans le sud de l’Ontario et du Québec, qui étaient maintenues par les pratiques de brûlage des Premières Nations (Riley, 2013), celles-ci ayant toutefois cessé après l’établissement européen dans ces régions. Les forêts de feuillus ont donc empiété sur ces milieux à mesure que les pratiques de brûlage des Premières Nations ont décliné (Riley, 2013), et l’habitat du monarque a diminué avec l’augmentation de la couverture forestière. À la fin du 19e siècle, la coupe des forêts de feuillus à des fins agricoles réalisée dans le nord-est de l’Amérique du Nord, notamment dans le sud de l’Ontario et du Québec et certaines portions des Maritimes, a certainement permis la dispersion rapide des asclépiades et par la suite celle du monarque (Brower, 1995; Pleasants, 2015).
Les tendances en matière d’habitat peuvent être estimées d’après des données de télédétection, si on suppose que les types de couverture constituant l’habitat du monarque sont les terres cultivées et les prairies. Récemment, l’habitat du monarque dans le sud du Canada a connu un déclin à cause de la conversion de prairies en terres cultivées de manière intensive et en terres utilisées à d’autres fins. De 2000 à 2011 (données les plus récentes accessibles), les terres agricoles et les prairies ont été converties en zones urbaines, rurales et industrielles sur environ 2 600 km2, ont été remplacées par la forêt ou des arbustaies sur 10 500 km2, et ont été détruites par d’autres causes sur 5 300 km2 (Agriculture and Agri-Food Canada, 2012). Durant la même période, environ 6 400 km2 de forêt ont été convertis en terres cultivées. Au total, la perte d’habitat du monarque s’élève donc à environ 12 000 km2 durant cette période. Approximativement 7 000 km2 de prairie ont été convertis en terres cultivées durant le même laps de temps, mais les effets de cette conversion sont inconnus.
Aux États-Unis, depuis le milieu des années 1990, environ 700 000 km2 de l’habitat de reproduction du monarque se trouvant à l’est des Rocheuses (population de l’est) ont été détruits par la modification des pratiques agricoles et le développement (The Center for Biological Diversity et al., 2014). Cette superficie représente environ le tiers de l’aire de reproduction estivale du monarque. Cette perte d’habitat a en grande partie été causée par l’utilisation accrue du glyphosate dans les cultures résistantes à cet herbicide (voir la section Menaces). Elle a également été causée par la conversion de terres inutilisées en terres agricoles et par le développement commercial et résidentiel. La demande accrue de biocombustibles a entraîné la destruction d’environ 45 000 km2d’habitat de reproduction du monarque. Des terres agricoles en jachère (terres faisant partie du Conservation Reserve Program) ont été converties en cultures de maïs et de soja de 2007 à 2014 (The Center for Biological Diversity et al., 2014). L’utilisation accrue d’herbicides dans ces terres a entraîné une diminution de leur valeur comme habitat pour le monarque, en tuant les asclépiades et autres plantes nectarifères (voir la section Menaces). De 1982 à 2010, aux États-Unis, 174 000 km2 de terres ont été convertis en zones résidentielles et commerciales (The Center for Biological Diversity et al., 2014).
À l’ouest des Rocheuses (population de l’ouest), les tendances relatives à l’habitat de reproduction ont été étudiées moins en détail, et les principales aires de reproduction n’ont pas été déterminées (Jepsen et Black, 2015), particulièrement en Colombie-Britannique (Guppy, comm. pers., 2016; Heron, comm. pers., 2016; St. John, comm. pers., 2016). La belle asclépiade est abondante en bordure des routes dans le sud de la Colombie-Britannique et ne semble pas représenter un facteur limitatif pour la reproduction du monarque (Dyer, comm. pers., 2015; Guppy, comm. pers., 2015; St John, comm. pers., 2015).
Biologie
Le monarque est un des insectes ayant été les plus étudiés en Amérique du Nord. Au Canada et aux États-Unis, les chercheurs ont mené de vastes études sur la migration, la génétique, la physiologie de l’hivernage et les habitudes alimentaires de l’espèce ainsi que sur ses prédateurs, ses parasites et ses maladies. Des études de marquage et des études réalisées à l’aide d’isotopes ont permis de combler les lacunes dans les connaissances concernant la biologie de la migration de l’espèce, et la modélisation a permis de mieux comprendre la dynamique des populations. Oberhauser et Solensky (2004) et Oberhauser et al. (2015a) ont compilé les recherches récentes et ont fait un examen approfondi de plusieurs aspects de la biologie du monarque. Un sommaire des connaissances sur la biologie du monarque est également présenté dans les sources suivantes : Schappert (2004), Commission for Environmental Cooperation (2008) et Jepsen et al. (2015).
Cycle vital et reproduction
Le cycle vital du monarque, comme celui de tous les papillons, comprend quatre stades : l’œuf, la chenille, la chrysalide et l’adulte. Les adultes estivaux (qui ne sont pas les mêmes adultes qui hivernent) se reproduisent lorsqu’ils sont âgés de trois à huit jours, et les femelles commencent à pondre immédiatement après l’accouplement (Oberhauser, 2004). Les femelles s’accouplent à jusqu’à 10 reprises, recevant chaque fois un spermatophore contenant des éléments nutritifs et des sels (Suzuki et Zalucki, 1986). Elles pondent 300 à 400 œufs sur la face inférieure des feuilles d’asclépiade (Oberhauser, 2004). Chaque œuf est déposé individuellement, mais plusieurs œufs sont souvent pondus sur différentes feuilles d’une même plante (Crolla et Lafontaine, 1996). L’éclosion survient trois à huit jours après la ponte (Schappert, 2004).
Le stade larvaire (figure 2) dure 9 à 14 jours en présence de températures estivales normales, mais plus longtemps lorsque les conditions sont fraîches (Oberhauser, 2004). Durant leur développement, les chenilles se nourrissent des feuilles des asclépiades de façon presque continue et atteignent une longueur totale de 5 cm, et leur poids corporel est multiplié par 2 000 (Oberhauser, 2004). Les chenilles tissent un coussinet de soie puis s’y suspendent la tête en bas en préparation de la nymphose, puis se transforment en chrysalide (Urquhart, 1987). Les adultes émergent 9 à 15 jours après le début de la nymphose; le taux de survie de l’œuf au stade adulte est d’environ 10 % (Oberhauser, 2004).
Dans le sud de l’Ontario et du Québec, le monarque produit chaque année deux ou trois générations de juin à octobre (Holmes et al., 1991; Crolla et Lafontaine, 1996). Dans le sud de la Colombie-Britannique, l’espèce produit au moins une génération chaque été (Guppy et Shepard, 2001; St. John, comm. pers., 2015; B.C. Conservation Data Centre, 2015). Les adultes estivaux ont une durée de vie de deux à cinq semaines, mais les adultes hivernants peuvent vivre jusqu’à neuf mois (Oberhauser, 2004).
La génération qui émerge à la fin de l’été ou à l’automne est dans un état de diapause reproductive et migre du Canada (et d’autres endroits aux États-Unis) jusqu’aux aires d’hivernage (Solensky, 2004). La diapause est déclenchée en automne par les jours décroissants, la diminution des températures et la sénescence des plantes servant de source de nourriture, mais quelques individus sortent de diapause durant la migration et se reproduisent dans le sud des États-Unis avant d’atteindre les aires d’hivernage (Solensky, 2004).
Les individus hivernants se reproduisent en hiver ou au début du printemps, mais ils ne pondent que lorsqu’ils trouvent des asclépiades sur le chemin de leur migration vers le nord, dans le sud ou le centre des États-Unis. Les adultes issus de ces œufs poursuivent la migration vers le nord. Environ 90 % des monarques qui atteignent l’aire de reproduction située dans la région des Grands Lacs proviennent du sud (sud du 35e parallèle) ou du centre (35e au 41e parallèles) des États-Unis. Les 10 % restants ont effectué la migration complète en une seule fois (Miller et al., 2012). Les individus qui proviennent du centre des États-Unis sont issus soit de parents hivernants, soit de jeunes individus de la première génération produite dans le sud des États-Unis, ce qui indique qu’au moins une génération est nécessaire pour réaliser la migration vers le nord jusqu’au Canada (Miller et al. 2012; Flockhart et al. 2013). La population de l’ouest a été moins bien étudiée, mais le voyage entre la Californie et le sud de la Colombie-Britannique nécessite apparemment plusieurs générations (Jepsen et al., 2015).
Physiologie et adaptabilité
Adaptabilité en matière d’habitat
Le monarque a été en mesure de s’adapter à certains changements environnementaux et de coloniser de nouveaux habitats. Au cours des 200 dernières années, l’espèce est parvenue à coloniser l’Australie, l’Europe, l’Afrique du Nord et de nombreuses îles océaniques dans le monde, ce qui montre qu’elle est capable de s’adapter à de nouveaux environnements physiques et à de nouvelles plantes hôtes. Ces nouvelles populations sont pour la plupart ou toutes non migratrices.
La population de l’ouest est parvenue à s’adapter à la perte des peuplements côtiers d’essences indigènes qui lui servaient de sites hivernage en Californie, en adoptant les peuplements d’eucalyptus non indigènes qui les ont remplacés (résumé dans Jepsen et Black, 2015). La population de l’est a été capable de coloniser les terres agricoles ainsi que les bords de route et de voies ferrées qui ont remplacé les forêts de feuillus dans l’est (Crolla et Lafontaine, 1996). Plus récemment, le monarque a été observé dans des parcelles où des asclépiades étaient cultivées à l’extérieur de leur aire de répartition indigène (p. ex. dans des fermes ou des jardins).
Un petit nombre d’individus de la population de l’est a été observé en train d’hiverner au Texas et sur la côte du golfe du Mexique, aux États-Unis, ce qui donne à penser que de nouveaux sites d’hivernage peuvent être établis. Ils formaient généralement des groupes de moins de 10 individus, plutôt que de denses groupes comme ce qui est observé dans les aires d’hivernage du Mexique et de la Californie. On ignore si ces individus se reproduisent après l’hivernage et contribuent à la génération suivante (Howard et al., 2010).
Stockage des composés
La chenille du monarque stocke les cardénolides (stéroïdes produits à des fins de protection contre les herbivores) présentes chez les asclépiades, ce qui dissuade la plupart des oiseaux et autres vertébrés prédateurs (Malcolm et Brower, 1989). Ces composés stockés sont également présents chez l’adulte.
Réserves d’énergie
Avant la migration, les adultes de l’espèce emmagasinent des lipides (à partir du nectar) qui leur servent de réserve d’énergie à métabolisation rapide (Gibo et McCurdy, 1993).
Déplacements et dispersion
Le monarque effectue chaque automne sa célèbre migration depuis le sud du Canada et le nord des États-Unis jusqu’au Mexique et à la Californie, puis retourne dans au nord chaque printemps. Il entame généralement sa migration vers le nord depuis le Mexique de mars au début d’avril. Plusieurs générations se succèdent dans cette migration vers le nord, et l’espèce atteint généralement le Canada à la fin mai ou au début juin (Wormington, 2008). Environ 10 % des individus qui ont hiverné au Mexique font le voyage en entier jusqu’en Ontario (Miller et al., 2012). Des individus dans cette situation ont été signalés dans le parc national de la Pointe-Pelée à la fin avril et au début mai, mais ils étaient usés (Wormington, comm. pers., 2007).
La migration vers le sud débute généralement au début août, mais pourrait se prolonger jusqu’au début novembre (Wormington, comm. pers., 2007). Les individus peuvent se rassembler par centaines ou par milliers pour passer la nuit sur des arbres le long des rives nord des lacs Ontario et Érié. Parmi les plus importantes aires de rassemblement, on compte le parc provincial Presqu’ile, la région de Long Point, la réserve nationale de faune de Long Point, le parc provincial Rondeau et le parc national de la Pointe-Pelée (Crolla et Lafontaine, 1996). L’envolée de centaines de milliers de monarques a été observée de nombreuses fois au parc national de la Pointe-Pelée (Wormington, 1994, 1997, 2008).
La population de l’est suit une voie centrale qui traverse le Kansas, l’Oklahoma et le Texas jusqu’au Mexique, ou alors une voie orientale qui longe la côte atlantique puis celle du golfe du Mexique (Brindza et al., 2008; Howard et Davis, 2009). La voie migratoire de la population de l’ouest est plus courte et longe la côte jusqu’en Californie et même jusque dans le nord du Mexique (Crolla et Lafontaine, 1996).
Durant la migration automnale, les monarques peuvent réduire leurs dépenses énergétiques en planant et en se laissant glisser et porter par des colonnes ascendantes d’air chaud jusqu’à des altitudes où leur vol sera accéléré par les forts vents dominants (Gibo et Pallett, 1979; Gibo, 1981). Le monarque s’oriente grâce au soleil (Perez et al.,1997) et à des capteurs magnétiques photosensibles qui sont logés dans ses antennes et jouent le rôle d’une boussole (Guerra et al., 2014).
Relations interspécifiques
Le protozoaire parasite obligatoire Ophryocystis elektroscirrha est présent dans les populations de monarques d’Amérique du Nord, d’Amérique du Sud et d’Australie (Altizer et Oberhauser, 1999). Ce parasite a peu d’effet sur la survie ou la reproduction du monarque, sauf lorsqu’il est présent à fortes concentrations (Altizer et Oberhauser, 1999). Une diminution du taux de survie des œufs jusqu’à l’éclosion, de l’envergure des ailes, de la masse corporelle, des distances de vol et des vitesses de vol a été observée dans le cas des individus auxquels on avait inoculé de fortes concentrations du parasite comparativement aux individus n’ayant reçu aucune inoculation (Altizer et Oberhauser, 1999; Bradley et Altizer, 2005). Le taux d’infection est moins élevé dans les populations migratrices que dans les populations non migratrices. Il est possible que l’efficacité de vol réduite des individus infectés entraîne l’élimination de ceux-ci durant la migration, ce qui réduirait la présence du parasite et sa transmission d’une génération à l’autre (Bradley et Altizer, 2005; Satterfield et al., 2015).
Parmi les arthropodes prédateurs du monarque, on compte les araignées, les chrysopes, les mantes, les libellules, les fourmis et les guêpes. Certains prédateurs parviennent à composer avec les cardénolides, en passant outre leur mauvais goût, en choisissant les individus qui renferment de faibles concentrations de ces toxines, en consomment uniquement les parties qui renferment de faibles concentrations de ces toxines ou en utilisant d’autres mécanismes (Fink et Brower, 1981; Oberhauser et al., 2015b). Les parasitoïdes du monarque comprennent 12 espèces de mouches tachinaires (en particulier le Lespesia archippivora) et une guêpe de la famille des Braconidés (Oberhauser et al., 2015b). Deux espèces d’oiseaux, le Cardinal à tête noire (Pheucticus melanocephalus) et l’Oriole d’Abeillé (Icterus galbula abeillei), consomment une grande quantité de monarques dans les sites d’hivernage au Mexique (voir la section Menaces et facteurs limitatifs).
Taille et tendances de la population
Activités et méthodes d’échantillonnage
On dispose de peu de données sur la taille ou les tendances de la population reproductrice au Canada. Le suivi de la migration effectué dans la région de Long Point fournit un indice de tendance pour les individus provenant du nord du lac Érié. En outre, des tendances peuvent être inférées d’après le nombre d’individus provenant du Canada et des États-Unis qui se rassemblent pour hiverner en Californie et au Mexique.
Des données de suivi sont recueillies de manière systématique dans les sites d’hivernage en Californie (population de l’ouest) et au Mexique (population de l’est), où un grand nombre d’individus sont concentrés sur de petites superficies. Depuis 1993, dans environ 20 sites au Mexique, la superficie (en hectares [ha]) occupée par les groupes de monarques hivernants est mesurée à des fins de suivi des tendances de la population (Brower et al., 2012; Garcia-Serrano et al., 2004; Vidal et Rendon-Salinas, 2014). Deux méthodes ont été utilisées pour estimer la taille totale de la population hivernante. La première méthode consiste à extrapoler les données obtenues grâce à des techniques de marquage-lâcher-recapture. L’autre méthode consiste à mesurer les individus sur un sous-échantillon de branches puis à extrapoler les données en fonction de la taille de la colonie. On ne dispose pas de suffisamment de données pour évaluer laquelle de ces méthodes donne les résultats les plus précis (Calvert, 2004; Solensky, 2004).
En Californie, un suivi est réalisé dans les sites d’hivernage depuis 1997 (Jepsen et Black, 2015). Une activité de recensement, nommée « Western Monarch Thanksgiving Count », fait appel à des bénévoles sur une période de deux semaines en automne pour le dénombrement des individus qui se rassemblent dans environ 100 sites côtiers (Monroe et al., 2015).
Plusieurs programmes reposant sur des bénévoles sont menés dans l’aire de reproduction de l’espèce, dont le Seasonal Count Program de la North American Butterfly Association, eButterfly, l’Illinois Butterfly Monitoring Network, le Monarch Caterpillar Monitoring Program ainsi que les atlas des papillons de l’Ontario et des Maritimes (Macnaughton et al., 2016; Maritimes Butterfly Atlas, 2016; Reis et al., 2015a,b; Inamine et al., 2016).
Un suivi des individus migrateurs est réalisé depuis 1995 dans la région de Long Point, sur la rive nord du lac Érié, en Ontario (Crewe et McCracken, 2015). Des dénombrements journaliers normalisés des individus adultes sont effectués dans deux stations. Des dénombrements semblables sont réalisés à Cape May, dans le New Jersey, et à Peninsula Point, dans le Michigan, aux États-Unis (Crewe et McCracken, 2015).
Abondance
Il n’existe aucune estimation de la taille de la population de monarque au Canada. Des « centaines de milliers » d’individus passent par la région de Long Point chaque automne pour la migration (Crewe et McCracken, 2015), mais on ignore quelle proportion de la population canadienne représentent ces individus. Dans les Maritimes, le monarque est répandu mais généralement peu commun, et son abondance varie considérablement d’une année à l’autre (Maritimes Butterfly Atlas, 2016). Une migration massive de monarques y a été observée en 2012 (Klymko, comm. pers., 2016). Au Nouveau-Brunswick, le monarque est signalé chaque année, généralement en petit nombre, et se reproduit principalement le long des berges du fleuve Saint-Jean, où l’asclépiade commune est localement abondante (Klymko, comm. pers., 2016; Maritimes Butterfly Atlas, 2016). Dans l’Île-du-Prince-Édouard, le monarque se reproduit dans des parcelles d’asclépiade incarnate, espèce indigène de la province, et d’asclépiade commune, espèce introduite dans la province.
Les individus du Canada et des États-Unis partagent les mêmes aires d’hivernage, mais les proportions relatives d’individus de chacun des pays sont inconnues. Moins de 500 000 monarques hivernent actuellement en Californie (NatureServe, 2015), et seulement un très faible pourcentage de ceux-ci se reproduisent au Canada. Certaines années, la population reproductrice de l’ouest (Colombie-Britannique) est si petite qu’elle n’est pas détectée (St. John, comm. pers., 2016).
Au Mexique, la taille totale de la population hivernante a été évaluée à 66 millions d’individus en 2014-2015, et à 200 millions en 2015-2016, d’après une extrapolation réalisée à partir de la superficie combinée des colonies hivernantes et d’une densité moyenne de 50 millions d’individus par hectare. Une tempête survenue au Mexique en mars 2016 a tué un grand nombre d’individus après le recensement de la population, mais le nombre exact de morts est inconnu (Monarch Watch, 2016).
Fluctuations et tendances
Les données de suivi de la population les plus rigoureuses ont été recueillies dans les sites d’hivernage de la Californie et du Mexique. Ces données montrent que, depuis la fin des années 1990, les populations hivernantes ont connu un déclin dans ces deux régions (figures 7 et 8).
Le nombre d’individus qui hivernent en Californie (population de l’ouest) a connu une tendance à la baisse de la fin des années 1990 à 2002; toutefois, aucune tendance claire n’a pu être dégagée de 2002 à 2015, le nombre d’individus ayant fluctué au cours de cette période entre 58 468 en 2009 et 271 924 en 2015 (NatureServe, 2015; Monroe et al., 2016). De 1997 à 2015, la population a connu un déclin de plus de 80 % (figure 6) (NatureServe, 2015; Semmens et al., 2016). Au cours de la période de dix ans la plus récente (2006 à 2015) la population de l’ouest a augmenté de 2,1 % par année. La proportion des individus du Canada qui hiverne en Californie est très faible par rapport à celle qui hiverne au Mexique, de sorte que la tendance de la population hivernante du Mexique est la seule considérée comme pertinente aux fins de la présente évaluation.
La population hivernante du Mexique, mesurée d’après la superficie (ha) d’habitat occupée, a connu un déclin considérable au cours de la période de 1994-2015 (figure 7; Monarch Watch, 2016). La régression linéaire logarithmique de la série chronologique a été calculée d’après les données recueillies durant cette période et indique un déclin de 83 %. Les estimations pour la période 2012-2014 ont été les plus basses de cette série. La superficie d’habitat occupée en 2015 (4 ha) a été plus élevée que les trois années précédentes, mais tout de même inférieure à la moyenne de la série chronologique (6 ha). Les effets sur les populations de monarque d’une tempête grave survenue en mars 2016 (après la réalisation du recensement de 2015-2016) restent à déterminer (Cave, 2016).
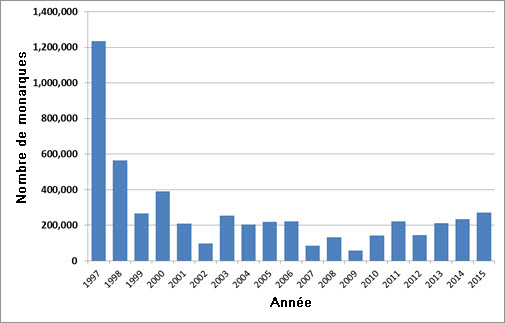
Description longue pour la figure 6
Graphique illustrant l’abondance estimative du monarque dans les sites d’hivernage en Californie, de 1997 à 2015.
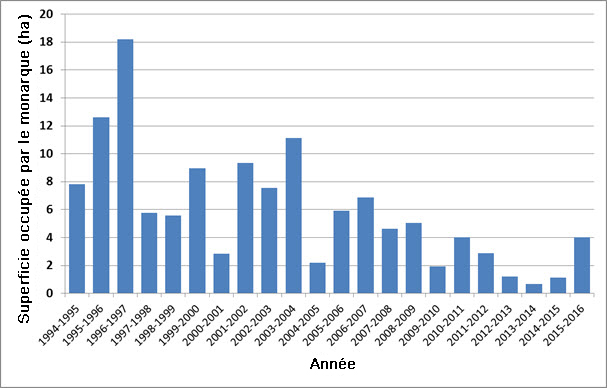
Description longue pour la figure 7
Graphique illustrant la superficie totale occupée par les colonies de monarques dans les sites d’hivernage du Mexique, de 1994-1995 à 2015-2016.
Le taux de déclin de l’habitat occupé dans les sites d’hivernage du Mexique au cours d’une période de 10 ans (figure 7; Monarch Watch, 2016) a été calculé au moyen la pente de la régression linéaire logarithmique établie d’après la série chronologique entière, appliquée à une période de 10 ans. Cette méthode est conforme aux lignes directrices de l’UICN relatives à l’application du critère A dans le cas des séries chronologiques comportant d’importantes fluctuations. La valeur de la pente a été estimée à -0,089 (p < 0,0001), et le changement sur 10 ans a été estimé à -59 % (figure 8).
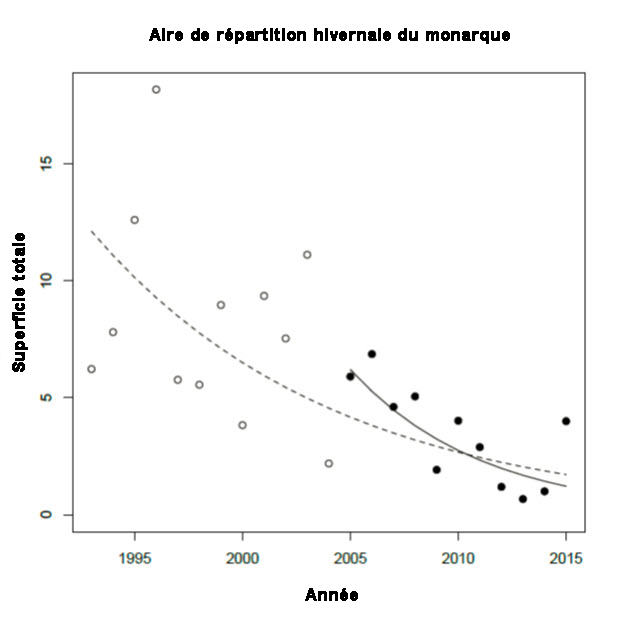
Description longue pour la figure 8
Graphique illustrant la superficie totale occupée par les colonies de monarques dans les sites d’hivernage du Mexique, avec droites de régression d’ajustement montrant le déclin au fil du temps.
Même si les données sur la population hivernante recueillies au Mexique indiquent une tendance marquée à la baisse, les données sur les tendances de la population de l’est dans son ensemble ne sont pas cohérentes lorsque différents stades de la migration sont comparés (voir Ries et al., 2015a, b; Inamine et al., 2016; Pleasants et al., 2016). Les indices de la taille de la population reproductrice, estimée d’après les données du Seasonal Count Program de la North American Butterfly Association et de l’Illinois Butterfly Monitoring Network, ont montré qu’il y avait une grande variabilité interannuelle mais qu’il n’y avait aucune tendance statistiquement détectable entre 1993 et 2013, période où le nombre d’individus hivernants au Mexique a connu un déclin (Reis et al., 2015a, b). Il est difficile de distinguer les facteurs responsables du déclin et de déterminer si le déclin se produit durant la reproduction estivale (hypothèse selon laquelle les asclépiades sont un facteur limitatif) ou à cause d’autres menaces cumulatives durant la migration saisonnière. La différence entre les résultats pourrait s’expliquer par un échantillonnage biaisé n’ayant pas détecté les déclins en régions agricoles (Pleasants et al., 2016), par une considérable variabilité interannuelle rendant la détection des tendances difficile ou par la mortalité durant la migration automnale.
Les activités de suivi de la migration n’indiquent pas toutes que la population connaît un déclin. Quelques études des tendances ont été réalisées au Canada, dont une étude à long terme (1995-2014) sur la migration menée dans la région de Long Point, en Ontario (Crewe et McCracken, 2015). Les individus dénombrés ont diminué en moyenne de 5,11 % par année, une fois les données ajustées en fonction des effets des conditions météorologiques. Le nombre d’individus variait considérablement d’une année à l’autre, et des orientations différentes ont été observées entre les deux sites de suivi. De 2010 à 2014, une diminution du nombre d’individus et de la variabilité de celui-ci a été observée (Crewe et McCracken, 2015). Dans le cadre d’études semblables réalisées à Cape May, dans le New Jersey, et à Peninsula Point, dans le Michigan, aucun signe de déclin n’a été observé (Davis, 2012; Badgett et Davis, 2015).
Dans deux publications récentes, des modèles démographiques ont été utilisés pour estimer le risque de disparition de l’espèce et les déclins de la population. Selon les prédictions de la publication de Flockhart et al. (2014), le risque de quasi-disparition sur 100 ans serait de plus de 5 %. Dans le cadre de cette étude, un modèle matriciel de projection périodique stochastique et dépendant de la densité structuré spatialement a été utilisé pour estimer les probabilités de quasi-disparition, le seuil de quasi-disparition ayant été fixé à < 1 000 individus. En plus de la probabilité de quasi-extinction, la publication prévoit un déclin de la population de 14 % sur 100 ans. Le seuil de quasi-disparition fixé à 1 000 individus a été critiqué, étant jugé trop optimiste (Semmens et al., 2016). Supposant une densité de 40 millions d’individus par hectare dans l’habitat d’hivernage, 1 000 individus occuperaient seulement 0,25 mètre carré de forêt; cette superficie est insuffisante pour que les fonctions de la population demeurent indemnes, puisque les individus de l’espèce se regroupent en hiver à des fins de thermorégulation, doivent trouver des partenaires dans toute la portion est de l’Amérique du Nord durant leur migration vers le nord et sont vulnérables aux conditions météorologiques extrêmes (Semmens et al., 2016).
Le deuxième modèle de quasi-disparition, qui s’applique uniquement à la population de l’est, est présenté dans un récent article de Semmens et al. (2016). Selon ce modèle, la probabilité de quasi-disparition est de 11 à 57 % sur 20 ans, si on utilise comme population de départ celle estimée à l’hiver 2014-2015, qui occupait 1,13 ha, et comme seuil de quasi-disparition une superficie de 0,01 à 0,25 ha. Le modèle a été ajusté en fonction du nombre d’hectares de forêt occupés durant l’hivernage, d’après l’ensemble des années pour lesquelles on dispose de données (1993-2014), et de la production annuelle d’œufs dans le Midwest américain (1999-2014), ainsi que du taux de croissance de la population (estimé à 0,94) et d’une déviation annuelle. Le modèle suppose une répartition normale et un écart-type correspondant à un bruit de processus (ou une stochasticité démographique attribuable à une variabilité biologique et environnementale) et suppose que le bruit dans la série chronologique est supérieur à l’erreur de mesure associée aux données recueillies dans les sites d’hivernage au Mexique.
La méthode utilisée pour l’estimation de la production annuelle d’œufs dans le Midwest américain est semblable à celle utilisée pour la portion du modèle concernant l’hivernage. La production annuelle d’œufs a été estimée d’après le nombre moyen d’œufs par tige d’asclépiade pour une année donnée, multiplié par le nombre de tiges d’asclépiade présentes dans le paysage cette même année. Les données ont été indexées à partir des observations d’œufs réalisées dans le cadre des projets de science citoyenne, et la densité d’asclépiades dans les différents types d’habitats dans le paysage, à partir des données des relevés de l’USDA (Semmens et al., 2016).
Immigration de source externe
Une immigration de source externe pourrait être possible. Toutefois, les individus qui se sont dispersés naturellement jusqu’en Europe et ailleurs dans le monde au cours des 200 dernières années n’ont pas conservé le comportement migratoire qui serait nécessaire à une telle immigration vers le Canada (Fernandez-Haeger et al., 2015). Vu la grande capacité de vol du monarque, il est possible que des individus errants puissent traverser les océans et recoloniser l’Amérique du Nord.
Menaces et facteurs limitatifs
La classification des menaces pesant sur le monarque (voir Salafsky et al., 2008; Master et al., 2009) été réalisée à l’aide du système de classification des menaces proposé par l’Union internationale pour la conservation de la nature (UICN) et le Partenariat pour les mesures de conservation (Conservation Measures Partnership, ou CMP) (IUCN-CMP) (tableau 1). Les menaces sont présentées selon leur ordre d’apparition dans le tableau 1. Les menaces analysées ci-dessous s’appliquent à l’ensemble de l’aire de répartition du monarque, y compris les sites d’hivernage et les voies migratoires qui traversent les États-Unis. Les menaces sont analysées pour les populations de l’ouest et de l’est combinées, mais dans certains cas les menaces diffèrent entre ces aires géographiques, et des explications sont alors fournies. Les menaces qui s’appliquent à l’aire de répartition canadienne sont explicitement énoncées. L’impact global des menaces calculé pour le monarque est très élevé.
Tableau 1. Classification des menaces pesant sur le monarque (voir Salafsky et al., 2008; Master et al., 2009), réalisée à l’aide du système de classification des menaces proposé par l’Union internationale pour la conservation de la nature (UICN) et le Partenariat pour les mesures de conservation (Conservation Measures Partnership, ou CMP) (IUCN-CMP). Les voies migratoires de l’est et de l’ouest ont été prises en compte pour l’évaluation. L’impact global des menaces calculé pour le monarque est très élevé.
- Nom français
- Monarque
- Nom scientifique
- Danaus plexippus
- Code de l’élément
- -
- Date
- 2016-01-12
- Évaluateur(s) :
- Jenny Heron (coprésidente du Sous-comité de spécialistes des arthropodes et animatrice), Al Harris (corédacteur), Paul Grant (coprésident du Sous-comité de spécialistes des arthropodes), Sheila Colla (membre du Sous-comité de spécialistes des arthropodes), Jessica Linton (membre du Sous-comité de spécialistes des arthropodes), John Klymko (membre du Sous-comité de spécialistes des arthropodes), Dave McCorquodale, (membre du Sous-comité de spécialistes des arthropodes), Remi Hebert (membre du Sous-comité de spécialistes des arthropodes), Leah Deforest (Agence Parcs Canada), Medea Curteanu (Environnement et Changement climatique Canada), Nathalie Desrosiers (membre du COSEPAC pour le Québec) et Angele Cyr (Secrétariat du COSEPAC et secrétaire de séance). Les personnes suivantes ont également formulé des commentaires : Ruben Boles (Environnement et Changement climatique Canada), Dave Fraser (Ministry of Environment de la C.-B.), Jon McCraken (coprésident du Sous-comité de spécialistes des oiseaux) et Justina Ray (coprésidente du Sous-comité de spécialistes des mammifères terrestres).
- Références :
- -
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l’intensité de leur impact : Maximum de la plage d’intensité |
Comptes des menaces de niveau 1 selon l’intensité de leur impact : Minimum de la plage d’intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 1 | 1 |
| C | Moyen | 2 | 2 |
| D | Faible | 3 | 3 |
| - | Impact global des menaces calculé : | Très élevé | Très élevé |
| # | Menace | Impact (calculé) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | D - Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | - |
| 1.1 | Habitations et zones urbaines | D - Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | S’applique à de petites superficies de l’habitat migratoire des populations de l’est et de l’ouest. |
| 1.2 | Zones commerciales et industrielles | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | S’applique à de petites superficies de l’habitat migratoire des populations de l’est et de l’ouest. |
| 1.3 | Tourisme et espaces récréatifs | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Modérée (possiblement à court terme, < 10 ans) | Considérée comme négligeable dans les sites d’hivernage de Californie (population de l’ouest); le risque de développement d’infrastructures est plus élevé dans le cas des sites d’hivernage du Mexique (population de l’est). |
| 2 | Agriculture et aquaculture (en anglais seulement) | C - Moyen | Restreinte (11-30 %) | Grave (31-70 %) | Élevée (constante) | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | D - Moyen | Restreinte (11-30 %) | Grave (31-70 %) | Élevée (continue) | S’applique aux populations de l’est et de l’ouest. |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | Sans objet. |
| 2.3 | Élevage et élevage à grande échelle | Pas une menace | Négligeable (< 1 %) | Neutre ou avantage possible | Élevée (continue) | S’applique aux populations de l’est et de l’ouest. La consommation des plantes sources de nourriture de l’espèce n’est pas prise en compte ici; on évalue plutôt l’effet de cette activité sur l’habitat. Avantage possible. |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | Sans objet. |
| 3 | Production d’énergie et exploitation minière (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Modérée (possiblement à court terme, < 10 ans) | - |
| 3.1 | Forage pétrolier et gazier | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Modérée (possiblement à court terme, < 10 ans) | Activité non considérée comme une menace grave. Il est probable que l’asclépiade pourrait pousser dans les milieux dégagés pour l’exploitation pétrolière et gazière. |
| 3.2 | Exploitation de mines et de carrières | - | - | - | - | Sans objet. |
| 3.3 | Énergie renouvelable | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (continue) | Menace s’appliquant à l’espèce mais considérée comme négligeable. S’applique davantage à la population de l’est. |
| 4 | Corridors de transport et de service (en anglais seulement) | D - Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 4.1 | Routes et voies ferrées | D - Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | Menace s’appliquant à l’espèce. Menace associée à la mortalité routière ainsi qu’aux activités d’entretien des routes, notamment l’application de pesticides. |
| 4.2 | Lignes de services publics | Pas une menace | Petite (1-10 %) | Neutre ou avantage possible | Modérée (possiblement à court terme, < 10 ans) | Pas considérées comme une menace. La création de lignes de services publics et l’entretien de ces lignes permettraient le maintien de milieux dégagés où l’asclépiade peut pousser. |
| 4.3 | Transport par eau | - | - | - | - | Sans objet. |
| 4.4 | Trajectoires de vol | - | - | - | - | Sans objet. |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | C - Moyen | Grande (31-70 %) | Modérée (11-30 %) | Élevée (constante) | - |
| 5.1 | Chasse et prélèvement d’animaux terrestres | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (constante) | Menace s’appliquant à l’espèce. |
| 5.2 | Cueillette de plantes terrestres | Pas une menace | Négligeable (< 1 %) | Neutre ou avantage possible | Élevée (continue) | Pas considérée comme une menace. |
| 5.3 | Exploitation forestière et récolte du bois | C - Moyen | Grande (31-70 %) | Modérée (11-30 %) | Élevée (continue) | Menace s’appliquant aux sites d’hivernage du Mexique (population de l’est). |
| 5.4 | Pêche et récolte des ressources aquatiques | - | - | - | - | Sans objet. |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (constante) | - |
| 6.1 | Activités récréatives | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (constante) | Menace s’appliquant à l’espèce. |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | Sans objet. |
| 6.3 | Travaux et autres activités | - | - | - | - | Sans objet. |
| 7 | Modification du système naturel (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | Menace s’appliquant à l’espèce. Les feux de friche représentent une menace possible dans les sites d’hivernage (populations de l’est et de l’ouest), mais ces sites sont répartis dans une vaste région, et le risque que le feu touche tous les sites en même temps est considéré comme négligeable. |
| 7.2 | Barrages, gestion et utilisation de l’eau | - | - | - | - | Sans objet. |
| 7.3 | Autres modifications de l’écosystème | - | - | - | - | Sans objet. La menace que représente la conversion des terres agricoles en cultures de soja ou de maïs (plantes résistantes aux herbicides) est traitée dans la section sur la pollution. |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | Inconnu | Grande (31-70 %) | Inconnue | Élevée (continue) | - |
| 8.1 | Espèces exotiques/non indigènes envahissantes | Inconnu | Grande (31-70 %) | Inconnue | Élevée (constante) | Menace s’appliquant à l’espèce. |
| 8.2 | Espèces indigènes problématiques | Inconnu | Inconnue | Inconnue | Élevée (continue) | Menace s’appliquant à l’espèce. |
| 8.3 | Introduction de matériel génétique | Inconnu | Inconnue | Inconnue | Élevée (continue) | Menace s’appliquant à l’espèce. Maladies introduites par les individus élevés en captivité. |
| 9 | Pollution (en anglais seulement) | B - Élevé | Grande (31-70 %) | Extrême - Élevée (31-100 %) | Élevée (continue) | - |
| 9.1 | Eaux usées domestiques et urbaines | - | - | - | - | Sans objet. |
| 9.2 | Effluents industriels et militaires | - | - | - | - | Sans objet. |
| 9.3 | Effluents agricoles et forestiers | B - Élevé | Grande (31-70 %) | Extrême - Élevée (31-100 %) | Élevée (continue) | Applicable. |
| 9.4 | Détritus et déchets solides | - | - | - | - | Sans objet. |
| 9.5 | Polluants atmosphériques | Inconnu | Grande (31-70 %) | Inconnue | Élevée (continue) | Menace s’appliquant dans l’ensemble de l’aire de migration et des sites d’hivernage de l’espèce (populations de l’est et de l’ouest). |
| 9.6 | Énergie excessive | - | - | - | - | Sans objet. |
| 10 | Phénomènes géologiques (en anglais seulement) | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | Menace ne s’appliquant pas dans l’aire de répartition canadienne. |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | Menace ne s’appliquant pas dans l’aire de répartition canadienne. |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | Menace ne s’appliquant pas dans l’aire de répartition canadienne. |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | - |
| 11.1 | Déplacement et altération de l’habitat | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | Menace s’appliquant à l’espèce. Il est difficile de déterminer son impact, mais il est probable que 1 % de l’habitat d’hivernage sera touché par cette menace. |
| 11.2 | Sécheresses | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | Ne s’applique pas aux asclépiades. Menace pour les forêts des sites d’hivernage. Il est difficile de déterminer son impact, mais il est probable que 1 % de l’habitat d’hivernage sera touché par cette menace. |
| 11.3 | Températures extrêmes | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | Menace pour les sites d’hivernage. Il est difficile de déterminer son impact, mais il est probable que 1 % de l’habitat d’hivernage sera touché par cette menace. |
| 11.4 | Tempêtes et inondations | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | Menace s’appliquant à l’espèce. Il est difficile de déterminer son impact, mais il est probable que 1 % de l’habitat d’hivernage sera touché par cette menace. |
Développement résidentiel et commercial (UICN 1, impact faible)
1.1 Zones résidentielles et urbaines (impact faible) et 1.2 Zones commerciales et industrielles (impact négligeable)
Développement au Canada
Le développement résidentiel et commercial représente une menace dans le centre du Canada, principalement dans le sud de l’Ontario, et donc pour la population de l’est. Les terres agricoles abandonnées qui fournissent actuellement des habitats de reproduction et d’alimentation des adultes aux individus de la population de l’est risquent d’être converties en zones résidentielles ou commerciales (Environment Canada, 2014). Dans le sud du Canada, environ 2 500 km2 de terres agricoles et de prairies ont été convertis en zones urbanisées de 2000 à 2011 (voir la section Tendances en matière d’habitat) (Agriculture and Agri-Food Canada, 2012). Le développement représente principalement une menace à proximité des agglomérations du sud de l’Ontario et du Québec, régions où on trouve le plus grand nombre d’individus reproducteurs.
Cette menace est considérée comme négligeable dans l’est et dans l’ouest. En Colombie-Britannique (population de l’ouest), la croissance de la population humaine et l’intensification de l’aménagement du territoire menacent l’habitat du monarque dans la vallée de l’Okanagan. La population humaine dans cette vallée a triplé tous les 30 à 40 ans depuis 1940 (Jensen et Epp, 2002), et l’on s’attend à ce qu’elle continue de croître au même rythme au cours des deux prochaines décennies (Hobson et Associates, 2006). Malgré ce développement, les asclépiades demeurent abondantes, répandues et communes dans la région (Dyer, comm. pers., 2016; St. John, comm. pers., 2016).
Développement dans les sites d’hivernage
Le développement immobilier le long de la côte californienne représente une menace pour les sites d’hivernage de la population de l’ouest. Depuis las années 1980, au moins 30 sites d’hivernage ont été détruits par le développement commercial et résidentiel en Californie (Jepsen et al., 2015). Les activités associées aux zones résidentielles, comme la taille ou l’élimination des arbres, peuvent modifier le microclimat d’un site d’hivernage et le rendre moins propice pour le monarque (Jepsen et al., 2015).
Au Mexique (population de l’est), les sites d’hivernage sont principalement regroupés dans une réserve de la biosphère et sont protégés. On considère qu’il y a un risque élevé que cet habitat et l’habitat adjacent non protégé soient touchés par le développement résidentiel et commercial.
Dans l’ensemble, l’impact des zones résidentielles et urbaines (1.1) et des zones commerciales et industrielles (1.2) est considéré comme faible.
1.3 Zones touristiques et récréatives (impact négligeable)
Le développement associé aux activités récréatives et touristiques représente une menace négligeable pour l’habitat du monarque dans l’ensemble de l’aire de répartition canadienne de l’espèce.
En Californie, il ne semble y avoir aucun développement de nouvelles infrastructures dans les sites d’hivernage de la population de l’ouest; la plupart des sites se trouvent dans des parcs publics, dispersés entre des zones résidentielles. Ces lieux sont encore visités par les humains, mais les probabilités sont faibles que de nouveaux sentiers ou de nouvelles infrastructures y soient aménagés. L’habitat d’hivernage de la population de l’est pourrait être menacé par le développement associé aux activités récréatives et touristiques. Ces sites reçoivent des centaines de milliers de visiteurs chaque année, et le besoin de créer de nouvelles installations pourrait augmenter dans le futur.
Agriculture et aquaculture (UICN 2, impact moyen)
2.1 Cultures annuelles et pérennes de produits autres que le bois (impact moyen)
Un déclin de l’abondance des asclépiades est observé au Canada et aux États-Unis, parce que les terres et les zones agricoles sont de plus en plus utilisées pour de vastes cultures industrielles intensives. À l’inverse, dans certaines parties du Canada, la succession naturelle fait en sorte que la forêt empiète lentement sur des terres agricoles abandonnées. Globalement, les changements touchant les milieux agricoles entraînent un déclin de l’abondance et de la répartition des asclépiades dans l’ensemble des aires de reproduction et de migration de l’espèce.
Les terres agricoles abandonnées qui fournissent actuellement un habitat de reproduction et d’alimentation des adultes à la population de l’est risquent de faire l’objet d’une exploitation agricole intensive (Environment Canada, 2014). De plus, les cultures dans lesquelles les asclépiades poussaient à l’état adventice (par exemple en l’absence de lutte contre les mauvaises herbes) sont remplacées par des cultures intensives comme le maïs et le soja, qui ne sont pas propices à la présence de colonies d’asclépiades (Pleasants, 2017).
Il n’existe pas de données sur les tendances des populations d’asclépiades au Canada. Cependant, on sait qu’aux États-Unis la perte des asclépiades est l’un des principaux facteurs contribuant au déclin de la population de l’est. Depuis la fin des années 1990, l’asclépiade commune a subi un déclin très important dans le Midwest américain (Jepsen et al., 2015; Pleasants, 2017).
Selon une étude menée en Iowa (dans l’aire de répartition de la population de l’est), les asclépiades ont connu un déclin de 64 % dans les zones agricoles de 1999 à 2010 (les données présentées dans Pleasants et Oberhauser, 2012, ont été corrigées par Pleasants, 2015). Durant cette même période, la population de l’est a subi un déclin de 88 % dans la région où été menée l’étude (Pleasants, 2015).
Le modèle démographique de Flockhart et al. (2014) pour la population de l’est prédit un déclin supplémentaire de 14 % d’ici 100 ans et conclu que les récents déclins de l’espèce ont été causés par une diminution de la présence des asclépiades aux États-Unis plutôt que par les changements climatiques ou la dégradation des habitats forestiers au Mexique. Les répercussions pourraient être moindres dans le cas de la population de l’ouest, puisqu’elle se reproduit dans une plus faible mesure dans les champs de soja, de maïs et de canola. Toutefois, Inamine et al. (2016) ont récemment évalué l’hypothèse selon laquelle l’espèce est limitée par les asclépiades, en analysant les données de science citoyenne de quatre programmes de suivi menés en Amérique du Nord. Selon leurs résultats, il n’y a aucune tendance temporelle significative sur le plan statistique, et il est peu probable que la faible présence d’asclépiades soit responsable des déclins de la population de monarques.
Selon une analyse plus approfondie menée par Pleasants (2017), de 1999 à 2014, dans le Midwest américain, 850 millions d’asclépiades ont été détruites par les herbicides utilisés dans les champs de maïs et de soja, et depuis 2008, 11 millions d’asclépiades supplémentaires sont disparues à cause de la conversion de prairies en terres agricoles. On estime que cela représente une perte de 71 % des supports dont disposent les individus en migration pour la ponte, puisque les asclépiades poussant dans les champs agricoles portent 3,9 fois plus d’œufs que celles poussant en milieu non agricole (Pleasants, 2017). Dans l’ensemble, on estime qu’il y a eu un déclin de 40 % du nombre d’asclépiades de 1999 à 2014 (Pleasants, 2017).
Les pratiques agricoles ont changé de façon semblable au Canada, particulièrement dans les Prairies et dans le sud de l’Ontario, mais on ne dispose actuellement d’aucune donnée ni d’aucune analyse à ce sujet.
L’asclépiade commune est considérée comme une mauvaise herbe nuisible en vertu de lois provinciales régissant la lutte contre les mauvaises herbes au Manitoba (Province of Manitoba, 2010), au Québec (Schappert, 2004), en Nouvelle-Écosse (Province of Nova Scotia, 2016) et, jusqu’à récemment (2014), en Ontario, mais elle n’est maintenant plus considérée comme nuisible dans cette province. Au Manitoba, la belle asclépiade et l’asclépiade incarnate sont elles aussi considérées comme des mauvaises herbes nuisibles.
Production d’énergie et exploitation minière (UICN 3, impact négligeable)
3.3 Énergie renouvelable (impact négligeable)
Il est peu probable que la production d’énergie et l’exploitation minière aient une incidence sur les populations de monarques. Dans le rapport de situation du COSEPAC précédent, les collisions avec les éoliennes ont été jugées comme une menace potentielle pour les individus migrants, mais ce phénomène n’a pas été signalé au Canada, et l’ampleur de cette menace est inconnue. En 2013, tous les rapports de suivi accessibles au public pour les parcs éoliens de l’Ontario situés le long des rives des lacs Ontario et Érié ont été examinés, et aucun cas de mortalité de monarques ou d’autres papillons n’y était mentionné (Linton, données pers., 2016).
Certaines centrales solaires peuvent causer la mort d’individus de l’espèce, notamment celles qui utilisent des miroirs pour réfléchir le rayonnement vers une chaudière située sur une tour. Un grand nombre de monarques morts a été observé dans ce type de centrale à concentration en Californie (Kagan et al., 2014). Les individus ont apparemment été tués lorsqu’ils ont traversé le faisceau concentré à proximité de la chaudière. Les technologies de concentration du rayonnement ne sont pas actuellement utilisées au Canada (Natural Resources Canada, 2016).
Corridors de transport et de service (UICN 4, impact faible)
4.1 Routes et voies ferrées (impact faible)
Sites de reproduction situés en bordure des routes
Les bords des routes sont un important habitat de reproduction pour le monarque, car ils sont périodiquement perturbés, ce qui permet aux asclépiades d’y pousser. Ces zones sont de plus en plus importantes, puisqu’il y a un déclin des asclépiades dans les champs agricoles partout aux États-Unis. Les emprises routières hébergent environ 10 % des asclépiades en Amérique du Nord (Flockhart et al., 2014), et on estime que 35 % des monarques y naîtraient dans le Midwest américain (données inédites, citées dans The Center for Biological Diversity et al., 2014). Les menaces qui pèsent sur l’espèce dans ce type d’habitat sont la mortalité des adultes attribuable aux collisions avec les véhicules, les fauchages excessifs et l’application d’herbicides qui détruisent les plantes servant de nourriture ainsi que l’exposition au sel de voirie.
Mortalité routière
Le monarque est vulnérable aux collisions avec les véhicules, particulièrement dans son aire de répartition estivale. Selon une étude menée dans le centre de l’Illinois, 500 000 monarques sont tués chaque année dans cet État (McKenna et al., 2001). Dans le cadre d’une étude menée en Pologne, les collisions avec des véhicules ont causé la mort d’au moins 7 % des papillons (toutes espèces combinées) vivant en bordure des routes. Le taux de mortalité routière des monarques augmentait avec l’augmentation de la densité de la circulation, de la largeur des routes et de la fréquence des fauchages (Skórkaa et al., 2013).
Entretien des routes
Les fauchages excessifs et l’utilisation d’herbicides sur le bord des routes pourraient tuer les chenilles de l’espèce et éliminer les asclépiades et les plantes nectarifères (Jepsen et al., 2015). Toutefois, un certain degré de fauchage pourrait être important pour le maintien des asclépiades en bordure des routes. En Oklahoma, l’Asclepias viridis repoussait en bordure des routes après les fauchages, mais en l’absence de fauchage il mourrait à la fin de l’été et n’était plus disponible pour le monarque (Baum et Mueller, 2015).
Les asclépiades exposées au ruissellement des sels de voirie présentent des taux de sodium plus élevés que les asclépiades qui poussent à l’écart des routes (Snell-Rooda et al., 2014). Le taux de survie chenilles du monarque qui grandissent sur des asclépiades poussant en bordure des routes est inférieur; chez les individus survivants, une masse musculaire accrue a été observée dans le cas des mâles, et un développement accru des tissus nerveux a été observé dans le cas des femelles (Snell-Rooda et al., 2014). On ignore toutefois l’effet de ce phénomène sur la valeur adaptative du monarque.
Utilisation des ressources biologiques (IUCN 5, impact élevé)
5.1 Chasse et capture d’animaux terrestres (impact faible)
Des individus sont capturés dans les aires de repos hivernales en Californie, pour servir de reproducteurs pour la production commerciale de monarques, mais l’ampleur de cette pratique est inconnue (Jepsen et Black, 2015).
Au Canada, des œufs et des chenilles sont prélevés en vue de leur élevage dans le sud de l’Ontario et dans les Maritimes, mais le nombre de spécimens ainsi récoltés est inconnu. Il y a peu de restrictions relatives à l’importation ou au déplacement de monarques d’un emplacement géographique à un autre au Canada. Voir la menace 6.1 pour des renseignements sur l’impact des parasites et protozoaires et la menace 8.3 pour des renseignements sur l’impact de l’introduction de matériel génétique.
5.3 Exploitation forestière et récolte du bois (impact élevé)
La destruction et la dégradation de la forêt dans les sites d’hivernage au Mexique étaient considérées comme une menace plus importante dans le passé qu’aujourd’hui, mais cette menace existe toujours à l’heure actuelle. Au total, dans la Réserve de la biosphère du monarque, 4 300 ha (8 %) de forêt de sapin oyamel ont été détruits ou ont subi des perturbations de 1986 à 2012, en grande partie à cause de l’exploitation forestière (Ramirez et al., 2015). Même l’exploitation à petite échelle représente une menace, car les trouées et les éclaircies dans le couvert forestier pourraient entraîner une mortalité parmi les colonies, celles-ci étant plus exposées aux effets des tempêtes hivernales, des basses températures et du temps humide (Jepsen et al., 2015; Williams et Brower, 2015). Cette région fait l’objet d’une exploitation forestière depuis la fin du 19e siècle, à des fins commerciales et domestiques pour la production de matériaux de construction, de bois de chauffage et de charbon (Ramirez et al., 2015). Bien que l’exploitation forestière soit interdite dans le cœur de la réserve, cette activité y est actuellement pratiquée de manière illégale (Jepsen et al., 2015). En 2015, 10 ha de forêt ont été coupés illégalement à Sierra Chincua, à seulement 600 m d’une aire de repos du monarque (Davis et Linton, 2016).
Intrusions et perturbations humaines (UICN 6, impact négligeable)
6.1 Activités récréatives (impact négligeable)
L’écotourisme réalisé de manière non durable représente une menace potentielle pour le monarque dans son habitat d’hivernage du Mexique (Commission for Environmental Cooperation, 2008; The Center for Biological Diversity et al., 2014). Les effets de cette activité n’ont pas été quantifiés, mais certains chercheurs soupçonnent que les perturbations et la dégradation de l’habitat qui en résultent pourraient nuire au monarque. On estime que les sites d’hivernage au Mexique recevraient 100 000 à 150 000 visiteurs chaque année (Vidal et al., 2014). Ce type de perturbation n’a pas été considéré comme une menace pour les individus hivernant en Californie dans le cadre d’un examen du statut de conservation du monarque aux États-Unis (Jepsen et al., 2015).
Modifications des systèmes naturels (UICN 7, impact négligeable)
7.1 Incendies et suppression des incendies (impact négligeable)
Les incendies représentent une menace potentielle pour le monarque dans ses aires d’hivernage en Californie et au Mexique. Dans les sites d’hivernage du Mexique, 45 incendies se sont produits dans la Réserve de la biosphère du monarque en 2012, touchant une superficie de 176,6 ha, soit 0,31 % du territoire de la réserve (Martinez-Torres et al., 2015). La plupart de ces incendies ont été causés par les activités agricoles, les feux de camp ou les activités forestières, et un seul incendie a été déclenché par la foudre (Martinez-Torres et al., 2015). Le régime forestier historique de la forêt de sapin oyamel, les répercussions de la suppression des incendies et l’efficacité des efforts de reboisement demeurent mal compris.
Dans les sites d’hivernage de Californie, les monarques se reposent principalement dans les peuplements d’eucalyptus, plante non indigène de la région (Frey et Schaffner, 2004). Les programmes destinés à éliminer les eucalyptus pour rétablir un couvert arboré indigène et à réduire les risques d’incendie pourraient par inadvertance constituer une menace pour le monarque. Les eucalyptus offrent une protection contre le vent, et leurs feuilles et la structure de leurs branches est propice au regroupement des monarques, mais l’espèce privilégie les essences indigènes comme le pin de Monterey et le cyprès de Monterey lorsque celles-ci sont présentes (Jepsen et al., 2015). En outre, une dégradation de l’habitat est associée au vieillissement et à la mort des pins de Monterey et des arbres d’autres espèces dans l’aire d’hivernage de la population de monarque de l’ouest (Jepsen et al., 2015).
Espèces et gènes envahissants ou problématiques (IUCN 8, impact inconnu)
8.1 Espèces exotiques (non indigènes) envahissantes (impact inconnu)
Le dompte-venin officinal (Cynanchum vincetoxicum) et le dompte-venin noir (C. louiseae ou Vincetoxicum nigrum) sont des plantes exotiques qui appartiennent à la même famille que les asclépiades et se propagent dans le centre et le sud de l’Ontario et le sud du Québec (Ontario’s Invading Species Awareness Program, 2015). Le monarque pond sur ces plantes exotiques, mais les chenilles nées sur ces espèces présentent un taux de survie significativement plus faible que celles nées sur les asclépiades, et elles ne dépassent souvent pas le premier stade larvaire (DiTommaso et Losey, 2003; Mattila et Otis, 2003; Casagrande et Dacey, 2007). Toutefois, le monarque préfère fortement les asclépiades indigènes à ces espèces non indigènes.
L’Asclepias curassavica se répand rapidement dans l’aire de répartition du monarque du Texas à la Floride (Batalden et Oberhauser, 2015). Durant la migration automnale vers le sud, le monarque préfère pondre sur l’Asclepias curassavica que sur les asclépiades indigènes, peut-être parce que les espèces indigènes sont en début de sénescence alors que l’Asclepias curassavica est généralement en bonne condition. La plupart des femelles sont en état de diapause reproductive durant la migration automnale, et la possibilité qu’elles sortent de cet état en présence de l’Asclepias curassavica pourrait avoir des conséquences négatives sur le cycle vital naturel du monarque (Batalden et Oberhauser, 2015). Ces effets n’ont pas encore été démontrés, mais ils sont considérés comme une menace potentielle pour l’espèce (Batalden et Oberhauser, 2015). De plus, des taux élevés du parasite Ophyrocystis elektroscirrha ont été observés chez les individus non migrateurs qui se nourrissent sur l’Asclepias curassavica (Satterfield et al., 2015).
Les monarques qui sont élevés à des fins commerciales et libérés dans la nature (voir la menace 8.3) pourraient être des vecteurs de maladies et de parasites (The Center for Biological Diversity et al., 2014; North American Butterfly Association, 2014). En effet, les monarques élevés en captivité peuvent être fortement infestés par le protozoaire parasite Ophyrocystis elektroscirrha, des bactéries du genre Serratia, des champignons du genre Nosema et des virus cytoplasmiques (The Center for Biological Diversity et al., 2014). Il est possible que les individus élevés en captivité puissent transmettre ces organismes nuisibles aux individus sauvages, mais le phénomène n’a pas été démontré.
8.2 Espèces indigènes problématiques (impact inconnu)
Les terres agricoles abandonnées qui fournissent actuellement à la population de l’est des milieux propices à la reproduction et à l’alimentation des adultes risquent de ne plus convenir à l’espèce, car elles sont remplacées par la forêt ou sont utilisées à des fins de culture intensive (voir la section Tendances en matière d’habitat).
8.3 Matériel génétique introduit (impact inconnu)
Les monarques élevés à des fins commerciales qui sont relâchés à l’occasion de mariages et d’autres événements sont considérés comme une menace potentielle pour les populations sauvages de l’espèce, parce qu’ils peuvent mener à l’introduction de caractères génétiques indésirables (The Center for Biological Diversity et al., 2014; North American Butterfly Association, 2014). Des millions de monarques sont relâchés chaque année aux États-Unis (The Center for Biological Diversity et al., 2014), mais l’ampleur de cette pratique au Canada est inconnue. Le degré de diversité génétique des individus élevés en captivité est inconnu, mais le mélange de ceux-ci avec les populations sauvages pourrait entraîner une dépression de consanguinité, l’introduction de caractères nuisibles et potentiellement une diminution du taux de survie des individus sauvages (The Center for Biological Diversity et al., 2014), en plus de nuire à l’étude scientifique des déplacements du monarque (Pyle, 2015).
Pollution (UICN 9, impact élevé)
9.3 Effluents agricoles et sylvicoles (impact élevé)
Il y a une forte corrélation entre le déclin des asclépiades et la culture plantes résistantes aux herbicides. Le glyphosate est utilisé depuis les années 1970, mais les plantes résistantes aux herbicides ont permis aux producteurs d’augmenter la fréquence d’utilisation de ce produit et les doses appliquées ainsi que de réaliser des traitements plus tard dans la saison que dans les cultures classiques (Center for Food Safety, 2015). L’asclépiade commune est tolérante au glyphosate lorsque celui-ci est utilisé en traitement de prélevée dans les champs de maïs et de soja. Les asclépiades sont vivaces et repoussent à partir de leur système racinaire déjà établi, de sorte les traitements herbicides de prélevée n’ont pas beaucoup d’effet sur elles; toutefois, le glyphosate les tue ou cause la chute de leurs feuilles lorsqu’il est appliqué plus tard dans la saison (Hartzler, 2010).
Au Canada, la culture de plantes résistante aux herbicides a considérablement augmenté au cours des vingt dernières années. Les plantes résistantes aux herbicides ont été utilisées pour la première fois au Canada en 1995 dans le cas du canola, en 1997 dans le cas du soja, en 1998 dans le cas du maïs et en 2004 dans le cas du blé (Beckie et al., 2006). En 2005, environ 95 % des superficies consacrées au canola et 60 % des superficies consacrées au soja au Canada étaient occupés par des plantes résistantes au glyphosate (Beckie et al., 2006). Le maïs résistant aux herbicides a été adopté moins rapidement que le soja résistant et représentait seulement 17 % à 18 % des superficies totales consacrées à cette plante en 2005. On ne dispose pas de données plus récentes pour le Canada, mais l’utilisation de plantes résistantes aux herbicides a probablement suivi des tendances semblables à celles observées aux États-Unis.
Aux États-Unis, presque toutes (91 %) les cultures de maïs et de soja sont traitées au glyphosate. Les superficies traitées avec ce produit ont été multipliées par neuf de 1995 à 2013 (Center for Food Safety, 2015). Durant cette période de 18 ans, les doses de glyphosate appliquées ont augmenté de 58 % dans les cultures de soja et de 43 % dans les cultures de maïs; la fréquence d’application a quant à elle augmenté de 64 % dans les cultures de soja et de 16 % dans les cultures de maïs. Au total, l’utilisation du glyphosate dans les champs de maïs et de soja a été multipliée par 20 et est passée de 4,5 millions kg/année à 93 millions kg/année (Center for Food Safety, 2015).
En outre, l’utilisation accrue des herbicides pourrait réduire les populations de plantes à fleurs qui fournissent du nectar au monarque durant sa migration. Le taux d’alimentation du monarque augmente durant sa migration vers le sud, particulièrement lorsqu’il atteint le Texas et le nord du Mexique. Le nectar est converti en lipides et utilisé comme source d’énergie par les individus hivernants (Brower et al., 2006). Le déclin des sources de nectar a été avancé comme facteur pouvant contribuer au déclin de la population hivernante (p. ex. Brower et al., 2006; Inamine et al., 2016).
Les insecticides de la catégorie des néonicotinoïdes, introduits dans les années 1990, comprennent l’imidaclopride, la clothianidine, le thiaméthoxame, l’acétamipride, le thiaclopride et le dinotéfurane. Au Canada, l’utilisation des néonicotinoïdes est approuvée pour le traitement des semences, l’épandage sur les sols et les traitements foliaires des cultures d’oléagineux, de grains, de légumineuses, de fruits, de légumes, de plantes en serre, de plantes ornementales et d’arbres de Noël (Health Canada, 2016). En Ontario, près de 100 % des semences de maïs et environ 60 % des semences de soja sont traitées aux néonicotinoïdes (Ontario Ministry of Agriculture, Food and Rural Affairs, 2014). L’utilisation des néonicotinoïdes a augmenté considérablement au cours des dernières années et a participé au déclin d’espèces non ciblées (Douglas et al., 2014; Jepsen et al., 2015).
Les néonicotinoïdes persistent dans le sol et circulent jusqu’aux tissus des plantes, pouvant atteindre des concentrations mortelles pour des insectes non ciblés (Goulson, 2013). On a nourri des chenilles de monarque avec des feuilles d’asclépiades renfermant des concentrations de clothianidine semblables à celles observées chez les asclépiades poussant dans des champs traités. La croissance de ces chenilles a été plus lente que celle des chenilles qui n’ont pas été exposées à l’insecticide (Pecenka et Lundgren, 2015). Un insecticide de la catégorie des néonicotinoïdes (imidaclopride) appliqué au sol a été absorbé par les asclépiades et transporté jusqu’à leurs tissus; une diminution du taux de survie des chenilles du monarque s’étant nourries des feuilles a été observée, mais cet effet n’a pas été observé chez les adultes ayant consommé le nectar (Krischik et al., 2015). Les néonicotinoïdes peuvent avoir des effets sublétaux chez les abeilles et autre invertébrés, dont une modification du comportement de quête alimentaire, une diminution de la reproduction et une augmentation de la vulnérabilité aux agents pathogènes (Goulson, 2013; van der Sluijs et al., 2013).
L’espèce a probablement été moins touchée par les herbicides dans l’ouest du Canada que dans l’est du pays, mais des insecticides sont utilisés dans les vignobles et les vergers de la vallée de l’Okanagan (Colombie-Britannique). De grandes quantités de pesticides sont utilisées dans ces deux types de systèmes agricoles, mais les effets de cette utilisation sur le monarque sont inconnus.
9.5 Polluants atmosphériques (impact inconnu)
Durant les activités de conduite des cultures, le sol est labouré avant l’ensemencement, ce qui soulève de la poussière qui est ensuite dispersée dans l’atmosphère. Selon les conditions météorologiques (p. ex., conditions sèches ou venteuses), cette poussière peut être transportée sur de longues distances ou demeurer dans la même zone durant quelques jours. Les résidus de pesticides mélangés au sol sont transportés dans l’atmosphère avec les autres particules de poussière, et finissent par se poser sur des plantes, dont des asclépiades, dans les zones adjacentes. Par exemple, le maïs Bt est une plante génétiquement modifiée qui représente une menace indirecte pour le monarque. L’acronyme Bt fait référence à la bactérie Bacillus thuringiensis, de laquelle ont a tiré un gène permettant la production d’une protéine toxique pour les chenilles de Lépidoptères. Ce gène a été incorporé dans le matériel génétique du maïs, rendant le goût de celui-ci moins agréable pour les chenilles et réduisant ainsi les besoins de traitements pesticides. Des études en laboratoire ont montré que la présence de pollen de maïs Bt déposé sur les feuilles d’asclépiades consommées par les chenilles du monarque causait une diminution des taux de croissance et de survie de celles-ci (Losey et al., 1999). Par contre, selon des études subséquentes réalisées au champ, il est peu probable que les concentrations de pollen soient suffisantes pour qu’il y ait des effets négatifs considérables sur les chenilles (Pleasants, 2015).
Changements climatiques et phénomènes météorologiques violents (UICN 11, impact faible)
Les effets des changements climatiques sur la dynamique de la reproduction du monarque sont complexes et doivent être étudiés plus en détail (Zipkin et al., 2012). Un certain nombre de mécanismes associés aux changements climatiques pourraient avoir des répercussions sur le monarque, notamment les phénomènes météorologiques violents (p. ex., gelées hâtives ou tardives dans les sites d’hivernage et dans l’aire de migration), le stress thermique, la perte d’asclépiades dans l’aire de reproduction, la modification des périodes de migration et la perte de l’habitat d’hivernage causée par le déplacement de celui-ci. La modélisation des scénarios climatiques futurs semble indiquer que les changements climatiques auront un impact sur les individus qui hivernent au Mexique. Il est difficile d’attribuer un niveau de gravité d’impact à cette menace, mais il est probable que les changements climatiques toucheront environ 1 % de la population dans les sites d’hivernage.
11.1 Déplacement et altération de l’habitat (impact faible)
Une asynchronie entre le développement des asclépiades et la migration du monarque représente une menace potentielle pour l’espèce (Robinson et al., 2009). L’évolution du climat a eu un effet plus marqué sur le monarque que sur ses plantes hôtes, de sorte que le monarque pourrait entreprendre sa migration vers le nord avant que les asclépiades servant de nourriture aux chenilles soient disponibles (Parmesan, 2007). Selon un modèle de changements climatiques, le régime de températures et de précipitations préféré par le monarque pour sa reproduction connaîtra un déplacement vers le nord au cours des 50 prochaines années (Batalden et al., 2007). Le monarque et les asclépiades devront donc étendre leurs aires de répartition vers le nord selon ce scénario (Batalden et al., 2007). En revanche, un autre modèle semble indiquer qu’il est peu probable que les changements du climat durant la saison de reproduction causent un déclin de la population de l’est (Zalucki et al., 2015).
En outre, les changements climatiques pourraient modifier l’habitat dans les aires d’hivernage. Avec le réchauffement climatique, la superficie des milieux convenant au sapin oyamel pourrait diminuer de 50 % d’ici 2030, et les forêts de cette espèce pourraient disparaître complètement d’ici 2090, en raison de la hausse de la mortalité due aux ravageurs et aux maladies (Ramirez et al., 2015). Les changements climatiques pourraient aussi avoir des répercussions sur l’habitat d’hivernage de la population de l’ouest, en causant une augmentation de la fréquence des feux de friche (Commission for Environmental Cooperation, 2008).
11.2 Sécheresses (impact faible)
Les modèles climatiques prédisent que la teneur en eau du sol diminuera dans de grandes portions de l’aire de reproduction du monarque, ce qui pourrait entraîner une diminution du succès de reproduction (Stevens et Frey, 2010). Toutefois, les asclépiades sont considérées comme des plantes résistantes à la sécheresse, et elles se rencontrent dans des régions où l’humidité est faible.
Le stress hydrique associé aux changements climatiques pourrait rendre le sapin oyamel plus vulnérable aux maladies et aux insectes ravageurs, notamment aux scolytes (Scolytus spp.) (Ramirez et al., 2015).
11.3 Températures extrêmes (impact faible)
L’exposition des chenilles à des températures élevées continues (36 °C) a entraîné une hausse du taux de mortalité et du temps de développement ainsi qu’une diminution de la masse des adultes (York et Oberhauser, 2002). Cependant, leur exposition à des températures fluctuant entre 27 °C et 36 °C a entraîné une très faible mortalité et une diminution du temps de développement par rapport au témoin.
11.4 Tempêtes et inondations (impact faible)
Dans les sites d’hivernage du Mexique et de la Californie, les tempêtes peuvent entraîner une mortalité catastrophique chez le monarque durant les mois où les individus de l’espèce se reposent dans ces sites. En janvier 2002, une tempête a tué environ 500 millions d’individus hivernants, soit environ 75 % de la population à ce moment. De fortes précipitations de pluie et de neige ont fait en sorte que les individus sont devenus extrêmement mouillés, puis les températures ont chuté jusqu’à -4 °C, causant l’hypothermie et la mort des individus (Brower et al., 2004). Une tempête de pluie et de grésil accompagnée de forts vents est survenue les 8 et 9 mars 2016, et un grand nombre d’individus ont été ensevelis sous le grésil (Monarch Watch, 2016). Cette tempête aurait causé la mort de 3 à 50 % de la population hivernante, mais de nombreux individus avaient déjà entamé leur migration vers le nord et ont ainsi évité la tempête. Un modèle climatique prédit une augmentation des précipitations associées à du temps frais, ce qui causera une hausse de la fréquence des épisodes de mortalité massive (Oberhauser et Peterson, 2003). La portée et la gravité des tempêtes hivernales sont considérables dans les sites d’hivernage, mais il est difficile d’évaluer l’impact de cette menace à long terme, sur une période de dix ans.
Facteurs limitatifs
Habitat d’hivernage
La majeure partie de la population adulte de l’est de l’Amérique du Nord se concentre sur un petit nombre d’hectares dans l’aire d’hivernage du Mexique, ce qui la rend vulnérable aux phénomènes météorologiques extrêmes, aux incendies, aux maladies et aux parasites, à la prédation et aux menaces anthropiques.
Migration
Puisqu’il migre sur de longues distances, le monarque est exposé à des phénomènes météorologiques extrêmes et a besoin d’habitats continus de reproduction et d’alimentation des adultes entre le Mexique et le Canada pour compléter son cycle migratoire. Au Canada, la population reproductrice de l’est dépend fortement du succès de la reproduction des individus migrateurs dans le sud des États-Unis, particulièrement au Texas et dans les États du centre (Miller et al., 2012; Ries et al., 2015b). Une diminution du succès de la reproduction causée par la sécheresse et d’autres menaces au Texas et dans le centre des États-Unis peut entraîner une importante réduction du nombre d’individus reproducteurs qui atteint le Canada.
Prédation
Le Cardinal à tête noire et l’Oriole d’Abeillé consomment une grande quantité de monarques dans les sites d’hivernage au Mexique. Les pertes dues à la prédation peuvent atteindre 34 000 individus par jour (Snook, 1993). Environ 2 millions d’individus (10 % de l’effectif total de la colonie) ont été consommés par des oiseaux au cours d’un seul hiver (Arellano et al., 1993). Les oiseaux se nourrissent à la limite externe des colonies, de sorte que les colonies de petite taille, qui ont une circonférence proportionnellement plus grande, subissent donc des taux de prédation plus élevés, pouvant atteindre 44 % (Calvert et al., 1979). En outre, une espèce de souris mexicaine, le Peromyscus melanotis, se nourrit abondamment de monarques morts ou vivants (Glendinning, 1993).
Nombre de localités
Le concept de localité ne s’applique pas dans le cas du monarque. L’espèce a une vaste aire de répartition, et les menaces qui pèsent sur elle sont complexes, varient selon le stade du cycle migratoire et le type d’habitat de reproduction (agricole ou naturel), et, dans le cas de l’habitat d’hivernage de Californie, la gestion locale des terres peut également être un facteur.
Protection, statuts et classements
Statuts et protection juridiques
Le monarque a été désigné espèce préoccupante par le COSEPAC en 1997 et a été inscrit à ce titre à la Loi sur les espèces en péril en 2004. Son statut a été réexaminé et confirmé en 2001 et en 2010.
Le monarque est considéré comme une espèce préoccupante aux termes la Loi de 2007 sur les espèces en voie de disparition de l’Ontario et de la Loi sur les espèces en péril du Nouveau-Brunswick (2013).
En Ontario, le monarque est protégé en vertu de la Loi sur la protection du poisson et de la faune.
Au Québec, le monarque n’est pas protégé par la Loi sur les espèces menacées ou vulnérables (RLRQ, c E-12.01) (LEMV) (MRNF, 2011) ni par la Loi sur la conservation et la mise en valeur de la faune (RLRQ, c. C-61,1) (LCMVF).
Statuts et classements non juridiques
Populations reproductrices du monarque (Natureserve, 2015)
Cote de conservation mondiale : apparemment non en péril (G4).
Cote de conservation au Canada : non en péril (N5B).
Cote de conservation aux États-Unis : non en péril (N5B).
Cotes de conservation à l’échelle provinciale :
Manitoba : non en péril (S5B).
Ontario : apparemment non en péril (S4B).
Colombie-Britannique, Alberta, Nouveau-Brunswick, Saskatchewan : vulnérable (S3B).
Nouvelle-Écosse et Terre-Neuve-et-Labrador : en péril (S2B).
Île-du-Prince-Édouard : gravement en péril (S1B).
Québec : non cotée.
En Ontario, une cote de conservation de S2N (en péril) a été attribuée aux populations non reproductrices, pour refléter la rareté des zones de concentration d’individus migrateurs (NHIC, 2015).
Cotes de conservation assignées à l’échelle subnationale aux États-Unis : non en péril (S5) dans 27 États, apparemment non en péril (S4 ou S4S5) dans 10 États, vulnérable (S3) en Californie et non cotée dans 11 États (NatureServe, 2015).
Cote de conservation attribuée aux zones de concentration d’individus migrateurs aux États-Unis : vulnérable à en péril (N2N3N).
L’Union internationale pour la conservation de la nature (UICN) a désigné la migration du monarque comme un phénomène en péril et reconnaît ainsi que la migration de l’espèce est en péril même si l’espèce elle-même n’est pas menacée d’extinction (Commission for Environmental Cooperation, 2008). Il s’agit du premier cas où un phénomène biologique est désigné plutôt qu’une espèce.
La Réserve de la biosphère du monarque au Mexique a été inscrite sur la Liste du patrimoine mondial en 2008 (UNESCO, 2015). Quinze des colonies hivernantes se trouvent dans cette réserve, et sept autres à l’extérieur de la réserve (Garcia-Serrano et al., 2004).
Au Mexique, le monarque est « sous protection particulière » aux termes du règlement régissant les espèces en péril (NOM-059-Semarnat-2001) (Commission for Environmental Cooperation, 2008).
Aux États-Unis, le monarque fait actuellement l’objet d’une évaluation en vue de son inscription à l’Endangered Species Act, grâce à une pétition lancée par des groupes environnementaux (The Center for Biological Diversity et al., 2014).
Protection et propriété de l’habitat
Au Canada, la majeure partie de l’habitat de reproduction du monarque se trouve sur des terrains privés, dans des emprises routières et dans d’autres types de milieux non protégés.
Au Canada, on trouve des aires de rassemblement migratoires dans des aires protégées fédérales, sur les rives des lacs Ontario et Érié, dans le sud de l’Ontario, notamment dans le parc national de la Pointe-Pelée, la réserve nationale de faune de Long Point et la réserve nationale de faune de Prince Edward. Ces aires font partie d’un réseau d’aires créé pour la protection du monarque dans le cadre d’une entente internationale conclue avec le Mexique et les États-Unis (Commission for Environmental Cooperation, 2008). On trouve d’autres aires de rassemblement en Ontario, dans des parcs municipaux et provinciaux ainsi que sur des terrains privés (Hess, 1992).
Au Mexique, la plupart des colonies hivernent dans la Réserve de la biosphère du monarque, protégée par le gouvernement fédéral et administré par la Commission nationale des aires naturelles protégées (Comisión Nacional de Áreas Naturales Protegidas) (Commission for Environmental Cooperation, 2008). De petites colonies hivernent à l’extérieur de la réserve et jouissent de divers degrés de protection fédérale (Commission for Environmental Cooperation, 2008). La Réserve de la biosphère du monarque, située au Mexique, a été inscrite sur la Liste du patrimoine mondial en 2008, mais l’exploitation forestière illégale, la croissance de la population humaine, les pressions associées au tourisme, les progrès agricoles et les feux de friche y représentent des menaces continues pour l’espèce (UNESCO, 2015).
Aux États-Unis, l’habitat d’hivernage du monarque en Californie est protégé par diverses lois et politiques fédérales, étatiques et locales. Toutefois, aucune contrainte n’est généralement imposée pour la collecte ou le dérangement des individus de l’espèce et pour l’élimination et la taille des arbres sur lesquels les individus se reposent, sauf sur les terres appartenant au gouvernement fédéral ou à l’État (International Environmental Law Project, 2012).
Remerciements
Merci à toutes les personnes énumérées dans la section Experts contactés, qui ont fourni des renseignements, des conseils et des connaissances précieux. Merci aussi à Phil Schappert, Mar Elderkin, Pail Johanson, Pail et Victoria Leck pour les renseignements qu’ils ont fournis. Des remerciements particuliers sont adressés à Ruben Boles (Service canadien de la faune, Environnement et Changement climatique Canada), Alan Sinclair (coprésident du Sous-comité de spécialistes des poissons marins), Jessica Linton (membre du Sous-comité de spécialistes des arthropodes), Paul Grant (coprésident du Sous-comité de spécialistes des arthropodes), Justina Ray (coprésidente du Sous-comité de spécialistes des mammifères terrestres), Patrick Nantel (Parcs Canada), Cory Sheffield (Royal Saskatchewan Museum) et Dave Fraser (Ministry of Environment de la Colombie-Britannique), qui ont fait une précieuse analyse et une révision minutieuse du document. Merci aussi à Colin Jones, Syd Cannings, Sheila Colla, Vincent Bereczky et Cory Sheffield, également membres du Sous-comité de spécialistes des arthropodes. Enfin, merci à Jennifer Heron (coprésidente du Sous-comité de spécialistes des arthropodes) pour son soutien technique et en matière de rédaction, Angele Cyr (Secrétariat du COSEPAC) pour son soutien technique et Jenny Wu (Secrétariat du COSEPAC) pour son soutien en matière de cartographie. Karine Bériault a rédigé le rapport de situation précédent (2010). La photographie de la couverture a été prise par Jessica Linton.
Experts contactés
Boles, Ruben. Service canadien de la Faune, Environnement et Changement climatique Canada, Gatineau (Québec).
Brownell, Vivian. Ministère des Richesses naturelles et des Forêts de l’Ontario, Division des politiques, Peterborough (Ontario).
Cannings, Syd. Environnement et Changement climatique Canada, Service canadien de la faune, Région du Pacifique et du Yukon, Whitehorse (Yukon).
Curteanu, Medea. Environnement et Changement climatique Canada, Service canadien de la Faune, Edmonton (Alberta).
Cyr, Angele. Secrétariat du COSEPAC, Service canadien de la faune, Environnement et Changement climatique Canada, Gatineau (Québec).
Desrosiers, Nathalie. Direction de la biodiversité et des maladies de la faune, Direction générale de l’expertise sur la faune et ses habitats, ministère des Forêts, de la Faune et des Parcs du Québec (Québec).
Fraser, David. British Columbia Ministry of Environment, Species Conservation Science Unit, Victoria (Colombie-Britannique).
Gauthier, Isabelle. Coordonnatrice provinciale des espèces fauniques menacées et vulnérables, ministère des Forêts, de la Faune et des Parcs (Québec).
Gelling, Lea. Conservation Data Centre de la Colombie-Britannique, Victoria (Colombie-Britannique).
Heron, Jennifer. British Columbia Ministry of Environment, Species Conservation Science Unit, Vancouver (Colombie-Britannique).
Huntley, Matt. Environnement et Changement climatique Canada, Service canadien de la faune, Région du Pacifique et du Yukon, Delta (Colombie-Britannique).
Jones, Colin. Centre d’information sur le patrimoine naturel de l’Ontario, Peterborough (Ontario).
Jones, Neil. Secrétariat du COSEPAC, Service canadien de la faune, Environnement et Changement climatique Canada, Gatineau (Québec).
Klymko, John. Centre de données sur la conservation du Canada atlantique, Sackville (Nouveau-Brunswick).
Nantel, Patrick. Agence Parcs Canada, Gatineau (Québec).
Linton, Jessica. Natural Resources Solutions, Waterloo (Ontario).
Pardy, Shelley. Department of Environment and Conservation, Endangered Species and Biodiversity, St. John (Terre-Neuve-et-Labrador).
Perrault, Julie. Secrétariat du COSEPAC Service canadien de la faune, Environnement et Changement climatique Canada, Gatineau (Québec).
Schnobb, Sonia. Secrétariat du COSEPAC Service canadien de la faune, Environnement et Changement climatique Canada, Gatineau (Québec).
Sheffield, Cory. Royal Saskatchewan Museum, Regina (Saskatchewan).
Stipec, Katrina. Conservation Data Centre de la Colombie-Britannique, Victoria (Colombie-Britannique).
Wu, Jenny. Secrétariat du COSEPAC, Service canadien de la faune, Environnement et Changement climatique Canada, Gatineau (Québec).
Sources d’information
Agriculture and Agri-Food Canada. 2012. 2011 AAFC crop type map of Canada. Site Web : ftp://ftp.agr.gc.ca/pub/outgoing/aesb-eos-gg/Crop_Inventory/2011/ (consulté le 9 octobre 2012). Statistics Canada, Environment Accounts and Statistics Division, 2013, special tabulation. [Consulté en août 2016]. (Également disponible en français : Agriculture et Agroalimentaire Canada. 2012. 2011 AAC Cartographie des cultures au Canada. Site Web : ftp://ftp.agr.gc.ca/pub/outgoing/aesb-eos-gg/Crop_Inventory/2011/. Statistique Canada, Division des comptes et de la statistique de l’environnement, 2013, tabulation spéciale.)
Altizer, S.M. et K.S. Oberhauser. 1999. Effects of the protozoan parasite Ophryocystis elektroscirrha on the fitness of Monarch butterflies (Danaus plexippus). Journal of Invertebrate Pathology 74(1): 76-83.
Anonymous. 2016. Press release: Monarch butterflies triple the area occupied for their Mexican wintering sites [PDF, 144 Mo]. Site Web : [Consulté en août 2016]. [en anglais seulement]
Antonijevic, E. D. St. John et J. Heron. 2014. Citizen science observations from rearing, tagging and releasing Monarch butterflies (Danaus plexippus) in the South Okanagan, British Columbia, in 2014. Rapport inédit. B.C. Ministry of Environment, Vancouver, B.C. 31pp.
Arellano, G., J.I. Glendinning, J.B. Anderson et L.P. Brower. 1993. Interspecific comparisons of the foraging dynamics of black-backed orioles and black-headed grosbeaks on overwintering monarch butterflies in Mexico. pp. 315-322. In Malcolm, S.B. and M.P. Zalucki (eds.), Biology and conservation of the Monarch butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Badgett, G. et A.K. Davis. 2015. Population trends of Monarchs at a northern monitoring site: analyses of 19 years of fall migration counts at Peninsula Point, MI. Annals of the Entomological Society of America: 1–7.
Batalden, R.V., K. Oberhauser et A.T. Peterson. 2007. Ecological niches in sequential generations of eastern North American Monarch butterflies (Lepidoptera: Danaidae): The ecology of migration and likely climate change implications. Environmental Entomology. 36:1365–1373.
Batalden, R.V. et K.S. Oberhauser. 2015. Potential changes in eastern North American Monarch migration in response to an introduced milkweed, Asclepias curassvica. Pp. 215-224. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Baum, K.A. et E.K. Mueller 2015. Grassland and roadside management practices affect milkweed abundance and opportunities for Monarch recruitment. Pp. 197-202. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Beckie, H. J., Harker, K. N., Hall, L. M., Warwick, S. I., Légère, A., Sikkema, P. H., Clayton, G. W., Thomas, A. G., Leeson, J. Y., Séguin-Swartz, G. et Simard, M.-J. 2006. A decade of herbicide-resistant crops in Canada. Canadian Journal of Plant Science 86: 1243–1264.
Bird, C.D., Hilchie, G. J., Kondla, N. G., Pike, E. M., & Sperling, F. A. H. 1995. Alberta Butterflies. The Provincial Museum of Alberta, Edmonton, Alberta. 349 pp.
Borders, B. et E. Lee-Mader. 2014. Milkweeds: A Conservation Practitioners Guide. 146 pp. Portland OR. The Xerces Society for Invertebrate Conservation.
Bradley, C.A. et S. Altizer. 2005. Parasites hinder Monarch butterfly flight: implications for disease spread in migratory hosts. Ecology Letters 8(3): 290-300.
Brindza, L.J., L.P. Brower, A.K. Davis et T. van Hook. 2008. Comparative success of Monarch Butterfly migration to overwintering sites in Mexico from inland and coastal sites in Virginia. Journal of the Lepidopterist’s Society 62:189-200.
Brower, L.P. 1995.Understanding and misunderstanding the migration of the monarch butterfly (Nymphalidae) in North America: 1857-1995 [PDF, 35.1 Mo]. Journal of the Lepidopterists Society 49:304–385. Site Web : [Consulté le 14 avril 2016]. [en anglais seulement]
Brower, L.P. 1996. Monarch butterfly orientation: missing pieces of a magnificent puzzle. The Journal of Experimental Biology 199:93-103.
Brower, A.V.Z. et T.M. Boyce. 1991. Mitochondrial DNA variation in monarch butterflies. Evolution 45: 1281 – 1286.
Brower, L.P. et R. M. Pyle. 2004. The interchange of migratory monarchs between Mexico and the western United States, and the importance of floral corridors to the fall and spring migrations. In G.P. Nabhan, ed., Conserving migratory pollinators and nectar corridors in western North America, pp. 144-166. Tuscan University of Arizona Press.
Brower, L.P., G. Castilleja, A.Peralta, J. Lopez-Garcia, L. Bojorquez-Tapia, S. Diaz, D. Melgarejo et M. Missrie. 2002. Quantitative changes in forest quality in a principal overwintering area of the Monarch Butterfly in Mexico, 1971-1999. Conservation Biology 16(2): 346-359.
Brower L.P., D.R. Kust, E. Rendon-Salinas, E.G. Serrano, K.R. Kust, J. Miller, C. Fernandez del Rey et K. Pape. 2004. Catastrophic winter storm mortality of Monarch butterflies in Mexico during January 2002. Pp. 151-166 in K.S. Oberhauser et M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Brower, L. P., O. R. Taylor, E. H. Williams, D. A. Slayback, R. R. Zubieta et M. I. Ramirez. 2012. Decline of monarch butterflies overwintering in Mexico: is the migratory phenomenon at risk? Insect Conservation and Diversity 5: 95–100.
Calvert, W. 2004. Two methods of estimating overwintering monarch population size in Mexico. Pp. 121-128. in K.S. Oberhauser et M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Calvert, W.H., L.E. Hedrick et L.P. Brower. 1979. Mortality of the Monarch butterfly (Danaus plexippus L.): avian predation at five overwintering sites in Mexico. Science 204: 847-851.
Casagrande, R.A. et J.E. Dacey, 2007. Monarch oviposition on swallow-worts (Vincetoxicum spp.). Environmental Entomology 36(3):631-636.
Cave, R. 2016. Monarch butterfly sanctuary in Mexico hit by cold weather. Site Web : [Consulté le 11 avril 2016].
Center for Food Safety (CFS). 2015. Monarchs in Peril Herbicide-Resistant Crops and the Decline Of Monarch Butterflies In North America. Site Web : http://www.centerforfoodsafety.org/files/cfs-monarch-report_2-4-15_design_05341.pdf. [Consulté en novembre 2015].
Commission for Environmental Cooperation. 2008. North American Monarch Conservation Plan. Site Web : [Consulté en novembre 2015]. (Également disponible en français : Commission de coopération environnementale. 2008. Plan nord-américain de conservation du monarque.
Commission for Environmental Cooperation. 2009. Monarch Butterfly Monitoring in North America: Overview of Initiatives and Protocols. Site Web : http://www.mlmp.org/Resources/pdf/Monarch-Monitoring_en.pdf [Consulté le 13 avril 2016]. (Également disponible en français : Commission de coopération environnementale. 2009. Surveillance du monarque en Amérique du Nord : aperçu des initiatives et des protocoles [PDF, 4.49 Mo].
COSEWIC. 2017. National ecological areas. Site Web : [consulté en février 2017]. (Également disponible en français : COSEPAC. 2017. Aires écologiques nationales du COSEPAC.
{kind=link}
{kind=link}
Crewe, T.L. et J. D. McCracken. 2015. Long-term trends in the number of Monarch Butterflies (Lepidoptera:Nymphalidae) counted on fall migration at Long Point, Ontario, Canada (1995–2014). Annals of the Entomological Society of America: 1–11
Crolla, J.P. et J.D. Lafontaine.1996. COSEWIC Status Report on the Monarch Butterfly. 23 pp.
Davis, A. K. 2012. Are migratory monarchs really declining in eastern North America? Examining evidence from two fall census programs. Insect Conservation and Diversity 5: 101–105.
Davis, A.K., N.P. Nibbelink et E. Howard. 2012. Identifying large- and small-scale habitat characteristics of Monarch butterfly migratory roost sites with citizen science observations. International Journal of Zoology. Volume 2012 (2012), Article ID 149026, 9 pages.
Davis, D. et J. Linton. 2016. Monarch wintering population increases by 255 percent however their fate remains uncertain after a devastating winter storm. Ontario Insects 21 (3):13-14.
DeMarco, E. 2015. Monarchs pose a puzzle. Science 349: 570-571.
Dingle, H., M.P. Zalucki, W.A. Rochester et T.Armijo-Prewitt. 2005. Distribution of the monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Nymphalidae), in western North America. Biological Journal of the Linnaean Society 85: 491-500.
DiTommaso, A. et J.E. Losey. 2003. Oviposition preference and larval performance of Monarch Butterflies (Danaus plexippus) on two invasive swallow-wort species. Entomologia Experimentalis et Applicata 108(3): 205-209.
Dockx, X., L.P Brower, L.I. Wasserman et K.A. Hobson. 2004. Do North American Monarch butterflies travel to Cuba? Stable isotope and chemical tracer techniques. Ecological Applications 14 :1106-1114.
Douglas, M.R., J.R. Rohr et J.F. Tooker. 2014. Neonicotinoid insecticide travels through a soil food chain, disrupting biological control of non-target pests and decreasing soya bean yield. Journal of Applied Ecology. 52 (1): 250-260.
Eanes, W.F. et R.K. Koehn. 1978. Analysis of genetic structure in the monarch butterfly, Danaus plexippus L. Evolution 32: 784 – 797.
Environment and Climate Change Canada. 2016. Management Plan for the Monarch (Danaus plexippus) in Canada. Species at Risk Act Management Plan Series. Environment Canada, Ottawa. iv + 45 pp. (Également disponible en français : Plan de gestion du monarque (Danaus plexippus) au Canada. 2016. Série de Plans de gestion de la Loi sur les espèces en péril, Environnement et Changement climatique Canada, Ottawa, v + 47 p.)
Fernandez-Haeger, J., D. Jordano et M.P. Zalucki. 2015. Monarchs across the Atlantic Ocean. Pp. 247-256. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Fink, L.S. et L.P. Brower. 1981. Birds can overcome the cardelonide defense of Monarch butterflies in Mexico. Nature 291: 67-70.
Flockhart, D.T.T., J.P. Pichancourt, D.R. Norris et T.G. Martin. 2014. Unravelling the annual cycle in a migratory animal: breeding-season habitat loss drives population declines of monarch butterflies. Journal of Animal Ecology. 84(1): 155-165.
Flockhart, D.T.T., L.I. Wassenaar, T.G. Martin, K.A. Hobson, M.B. Wunder et D.R. Norris. 2013. Tracking multi-generational colonization of the breeding grounds by monarch butterflies in eastern North America. Proceedings of the Royal Society B 280: 20131087.
rey, D. et A. Schaffner. 2004. Spatial and temporal pattern of Monarch overwintering abundance in western North America. Pp. 167-176 In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Garcia-Serrano, E., J.L. Reyes et B.X. Alvarez. 2004. Locations and areas occupied by Monarch butterflies overwintering in Mexico from 1993 to 2002. pp. 129-133. In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Gauthier, I. comm. pers. 2016. Communication par courriel adressée à A. Harris, avril 2016, biologiste et coordonnatrice provinciale des espèces fauniques menacées ou vulnérables. Direction générale de la gestion de la faune et des habitats. Ministère des Forêts, de la Faune et des Parcs. Québec.
Gibo, D.L. 1981. Altitudes attained by migrating Monarch Butterflies, Danaus plexippus (Lepidoptera: Danaidae) as reported by glider pilots. Canadian Journal of Zoology 59(3): 571-572
Gibo, D.L. et J.A. McCurdy. 1993. Lipid accumulation by migrating Monarch butterflies, Danaus plexippus, (Nymphalidae). Canadian Journal of Zoology 71: 76-82.
Gibo, D.L. et M.J. Pallett. 1979. Soaring flight of monarch butterflies, Danaus plexippus (Lepidoptera: Danaidae), during the late summer migration in southern Ontario. Canadian Journal of Zoology. 57:1393-1401.
Glendinning, J.I. 1993. Comparative feeding responses of the mice Peromyscus melanotis, P. aztecus, Reithrodontomys sumichrasti, and Microtus mexicanus to overwintering Monarch butterflies in Mexico. pp. 323-333. In Malcom, S.B. and M.P. Zalucki (eds.), Biology and conservation of the Monarch butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Goulson, D. 2013. An overview of the environmental risks posed by neonicotinoid insecticides. Journal of Applied Ecology. 50: 977–987.
Guerra, P.A., R.J. Gegear et S.M. Reppert. 2014. A magnetic compass aids monarch butterfly migration. Nature Communications 5: 4164.
Guppy, C.S. et J.H. Shepard. 2001. Butterflies of British Columbia: Including Western Alberta, Southern Yukon, the Alaskan Panhandle, Washington, Northern Oregon, Northern Idaho and Northwestern Montana. UBC Press, Vancouver, B.C . 414 pp.
Gustafsson, K.M., A.A. Agrawal, B.V. Lewenstein et S.A. Wolf. 2015. The Monarch butterfly through time and space: the social construction of an icon. Bioscience 65 (6): 612-622.
Handfield, L., J.D. Lafontaine, B. Landry, F. Lessard, D. Rancourt, E. Rassart et P. Thibault. 1999. Le Guide des Papillons du Québec. Volume 1. Broquet inc., Ottawa. 123 pp.
Hartzler, R.G. 2010. Reduction in common milkweed (Asclepias syriaca) occurrence in Iowa cropland from 1999 to 2009. Crop Protection 29: 1542-1544.
Health Canada. 2016. Update on Neonicotinoid Pesticides and Bee Health. Site Web : [Consulté en août 2016]. (Également disponible en français : Santé Canada. 2016. Mise à jour sur les pesticides de la classe des néonicotinoïdes et sur la santé des abeilles.
Hess, Q.F. 1992. The Monarch in Ontario. pp. 8-10. In: Hanks, A.J. et Q.F Hess (eds.), Butterflies of Ontario and Summaries of Lepidoptera Encountered in Ontario in 1991. Occasional Publication # 24-92. Toronto Entomologists’ Association.
Hobson and Associates. 2006. Environmental Issues and Options for the South Okanagan Regional Growth Strategy. Vol 1: Background. 119 pp. Disponible à l’adresse : http://www.rdosmaps.bc.ca/min_bylaws/planning/rgs/ReportsAndStudies/RGSVol1EnvIssuesOptionFinal06.pdf [consulté en décembre 2015]
Holmes, A.M., Q.F. Hess, R.R. Tasker et A.J. Hanks, 1991. The Ontario Butterfly Atlas. Toronto Entomologists’ Association, Toronto, 167 pp.
Howard, E., H. Aschen et A.K. Davis. 2010. Citizen science observations of monarch butterfly overwintering in the southern United States. Psyche 2010: 1-6.
Howard, E. et A.K. Davis. 2009. The fall migration flyways of monarch butterflies in eastern North America revealed by citizen scientists. Journal of Insect Conservation 13: 279-286.
Howard, E. et A.K. Davis. 2015. Investigating long-term changes in the spring migration of Monarch Butterflies (Lepidoptera: Nymphalidae) using 18 years of data from Journey North, a citizen science program. Annals of the Entomological Society of America: 1–6.
Inamine, H., S. P. Ellner, J.P. Springer et A.A. Agrawal. 2016. Linking the continental migratory cycle of the monarch butterfly to understand its population decline. Oikos doi: [10.1111/oik.03196].
International Environmental Law Project. 2012. The legal status of Monarch butterflies in California. Monarch Joint Venture and Xerces Society for Invertebrate Conservation. Site Web : http://www.xerces.org/wp-content/uploads/2008/09/legal-status-of-california-monarchs.pdf . [Consulté en novembre 2015].
IUCN Species Survival Commission. 2010. Guidelines for Application of IUCN Red List Criteria at Regional and National Levels Version 4.0. Site Web : http://www.iucnredlist.org/documents/reg_guidelines_en.pdf [Consulté en décembre 2015]. (Également disponible en français : Santé Canada. 2016. Mise à jour sur les pesticides de la classe des néonicotinoïdes et sur la santé des abeilles.
Jensen, E.V. et P.F. Epp. 2002. Water quality trends in Okanagan, Skaha and Osoyoos Lakes in response to nutrient reductions and hydrologic variation. Ministry of Water, Land and Air Protection. Penticton, B.C.
Jepsen, S. et S.H. Black. 2015. Understanding and conserving the western North American Monarch population. Pp. 147-156. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Jepsen, S., D.F. Schweitzer, B. Young, N. Sears, M. Ormes et S.H. Black. 2015. Conservation Status and Ecology of the Monarch Butterfly in the United States. NatureServe, Arlington, Virginia et the Xerces Society for Invertebrate Conservation, Portland, Oregon. 39 pp.
Kagan, R.A., T.C. Viner, P.W. Trail et E.O. Espinoza. 2014. Avian Mortality at Solar Energy Facilities in Southern California: A Preliminary Analysis. National Fish and Wildlife Forensics Laboratory. Unpublished report.
Klassen, P., R. Westwood, W.B. Preston et W.B. McKillop.1989. The Butterflies of Manitoba. Manitoba Museum of Man and Nature, Winnipeg, Manitoba.
Klymko, J. comm. pers. 2016. Communication par courriel adressée à A. Harris, avril 2016, zoologiste Centre de données sur la conservation du Canada Atlantique, Sackville, Nouveau-Brunswick.
Knight, A. et L.P. Brower. 2009. The influence of eastern North American autumnal migrant Monarch butterflies (Danaus plexippus L.) on continuously breeding resident Monarch populations in Southern Florida. Journal of Chemical Ecology online: doi 10.1007/s10886-009-9655-z.
Krischik V., M. Rogers, G. Gupta et A. Varshney. 2015. Soil-applied imidacloprid translocates to ornamental flowers and reduces survival of adult Coleomegilla maculata, Harmonia axyridis, and Hippodamia convergens lady beetles, and larval Danaus plexippus and Vanessa carduibutterflies. PLoS ONE 10(3): e0119133. doi:10.1371/journal.pone.0119133
Lane, J. 1984. The status of monarch butterfly overwintering sites in Alta California. Atala 9:17-20.
Lane, J. 1993. Overwintering Monarch Butterflies in California: Past and Present. pp. 335-344. In Malcolm, S.B. et M.P. Zaluchi (eds), Biology and Conservation of the Monarch Butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Layberry, R.A., P.W. Hall et J.D. Lafontaine. 1998. Butterflies of Canada. University of Toronto Press. 280 pp.
Leong, K., D., W.H. Sakai, W. Bremer, D. Feuerstein et G. Yoshimura. 2004. Analysis of the pattern of distribution and abundance of monarch overwintering sites along the California coastline. Pp. 177-186 in K.S. Oberhauser et M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Linton, J. comm. pers. 2016. Communication par courriel adressée à A. Harris, avril 2016. Natural Resources Solutions Inc. Waterloo, Ontario.
Linton, J. données pers. Communication par courriel adressée à J.Heron et à A. Harris, novembre 2016. Natural Resources Solutions Inc. Waterloo, Ontario.
Losey, J.E., L.S. Rayor et M.E.Carter. 1999. Transgenic pollen harms monarch larvae. Nature 399: 214.
Lyons, J.I., A.A. Pierce, S.M. Barribeau, E.D. Sternberg, A.J. Mongue et J.C. De Roode. 2012. Lack of genetic differentiation between monarch butterflies with divergent migration destinations. Molecular Ecology 21: 3433-3444.
Malcolm, S.B. et L.P. Brower. 1989. Evolutionary and ecological implications of cardenolide sequestration in the Monarch butterfly. Experientia 45:284-295.
Marshall, S.A. 2006. Insects: Their Natural History and Diversity. Firefly Books Ltd., Richmond Hill, Ont.
Mattila, H.R. et G.W. Otis. 2003. A comparison of the host preference of Monarch butterflies (Danaus plexippus) for milkweed (Asclepias syriaca) over dog-strangler vine (Vincetoxinum rossicum). Entomologia Experimentalis et Applicata 107(3): 193-199.
MacNaughton, A., R. Layberry, C. Jones et B. Edwards. 2016. Ontario Butterfly Atlas Online. Site Web : [Consulté le 11 avril 2016].
MacNaughton, A. 2010. The birth of Ontario Insects 1995. Ontario Insects 15 (3): 40-41.
Maritimes Butterfly Atlas. 2016. Atlantic Canada Conservation Data Centre, Sackville, New Brunswick. Site Web : [Consulté le 1er avril 2016].
Martinez-Torres, H., M. Cantu-Fernandez, M.I. Ramirez et D.R. Perez-Salicrup. 2015. Fires and fire management in the Monarch Butterfly Biosphere Reserve. Pp. 179-189. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Master, L., D. Faber-Langendoen, R. Bittman, G. A. Hammerson, B. Heidel, J. Nichols, L. Ramsay et A. Tomaino. 2009. NatureServe Conservation Status Assessments: Factors for Assessing Extinction Risk. NatureServe, Arlington, VA.
McKenna D.D., K.M. McKenna, S.B. Malcom et M.R. Berenbaum. 2001. Mortality of Lepidoptera along roadways in central Illinois. Journal of the Lepidopterists’ Society 55(2): 63-68.
Miller, N.G., L.I. Wassenaar, K.A. Hobson et D.R. Norris. 2012. Migratory connectivity of the Monarch butterfly (Danaus plexippus): patterns of spring re-colonization in eastern North America. PLoS ONE 7(3): e31891. doi:10.1371/journal.pone.0031891
Monarch Watch. 2016. Monarch population status. Site Web : [Consulté le 29 août 2016].
Monroe, M., Pelton, E., C. Fallon, D. Frey et S. Stevens. 2016. Western Monarch Thanksgiving Count Data from 1997–2015. Site Web : http://www.westernmonarchcount.org/data/. [Consulté en avril 2016].
Natural Heritage Information Centre (NHIC) 2015. Site Web : https://www.ontario.ca/page/get-natural-heritage-information. [Consulté en novembre 2015]. (Également disponible en français : Centre d’information sur le patrimoine naturel. 2015.
Natural Resources Canada. 2016. Renewables. Site Web : [Consulté le 7 avril 2016]. (Également disponible en français : Ressources naturelles Canada. 2016. Énergies renouvelables.
Nature Serve. 2015. NatureServe Explorer: An online encyclopedia of life [application Web]. Version 7.1. NatureServe, Arlington, Virginia. [consulté en octobre 2015]
North American Butterfly Association. 2014. There’s no need to release butterflies -- they’re already free. Site Web : [Consulté en novembre 2015]. [en anglais seulement]
Oberhauser K.S. 2004. Overview of Monarch breeding biology. Pp. 3-8. In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser and M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Oberhauser K.S. and M.J. Solensky (eds). 2004. The Monarch Butterfly: Biology and Conservation. Oberhauser and Solensky Cornell University Press, Ithaca, New York.
Oberhauser, K.S., L, Ries, S. Altizer, R.V. Batalden, J. Kudell-Ekstrum, M. Garland, E. Howard, S. Jepsen, J. Lovett, M. Monroe, G. Morris, E. Rendón-Salinas, R.G. RuBino, A, Ryan, O.R. Taylor, R. Trevino, F.X. Villablanco et D. Walton. 2015a. Contributions to Monarch biology and conservation through citizen science. Pp. 13-30. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Oberhauser, K.S., M. Anderson, S. Anderson, W. Caldwell, A. De Anda, M. Hunter, M.C. Kaiser et M.J. Solensky. 2015b. Lacewings, wasps, and flies – oh my. Pp. 71-82. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Oberhauser, K. et A.T. Peterson. 2003. Modeling current and future potential wintering distributions of eastern North American Monarch butterflies. Proceedings of the National Academy of Sciences of the United States of America 100:14063-14068.
Ontario’s Invading Species Awareness Program. 2015. Dog-strangling Vine. Site Web : http://www.invadingspecies.com/invaders/plants-terrestrial/dog-strangling-vine/. [Consulté en novembre 2015]
Ontario Ministry of Agriculture, Food and Rural Affairs. 2014 (mise à jour le 3 août 2016). Pollinator Health. Site Web : [Consulté en août 2016]. (Également disponible en français : Ontario. Ministère de l’Agriculture, de l’Alimentation et des Affaires rurales. 2014 (mis à jour le 3 août 2016). La santé des pollinisateurs.
Opler, P. A. et A. D. Warren. 2002. Butterflies of North America. 2. Scientific Names List for Butterfly Species of North America, north of Mexico. C.P Gillette Museum of Arthropod Diversity, Department of Bioagricultural Sciences and Pest Management, Colorado State University, Fort Collins, Colorado. 79 pp.
Parmesan, C. 2007. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology (13): 1860-1872.
Pecenka, J.R. et J.G. Lundgren. 2015. Non-target effects of clothianidin on monarch butterflies. The Science of Nature 102: 19.
Perez, S.M., O.R. Taylor et R. Jander. 1997. A sun compass in monarch butterflies. Nature 387: 29.
Pierce, A.A.,S. Altizer, N.L. Chamberlain, M.R. Kronforst et J.C. de Roode. 2015. Unraveling the mysteries of Monarch migration and global dispersal through molecular genetic techniques. Pp. 257-267. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Pleasants, J.M. et K.S. Oberhauser. 2012. Milkweed loss in agricultural fields because of herbicide use: effect on the monarch butterfly population. Insect Conservation and Diversity (2012) doi: 10.1111: 1-10.
Pleasants, J.M. 2015. Monarch butterflies and agriculture. Pp. 169-178. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Pleasants, J.M., E.H. Williams, L.P. Brower, K.S. Oberhauser et O.R. Taylor. 2016. Conclusion of no decline in summer monarch population not supported. Annals of the Entomological Society of America. 2016: 1–3 doi: 10.1093/aesa/sav115
Pleasants, J.M. 2017. Milkweed restoration in the Midwest for monarch butterfly recovery: estimates of milkweeds lost, milkweeds remaining and milkweeds that must be added to increase the monarch population. Insect Conservation and Diversity 10(1): 42 – 53.
Province of Manitoba. 2010. The Noxious Weeds Act. Manitoba Laws. Site Web : [consulté le 14 avril 2016]. (Également disponible en français : Province du Manitoba. 2010. Loi sur la destruction des mauvaises herbes.
Province of Nova Scotia. 2016. Weed control regulations. Site Web : [Consulté le 14 avril 2016]
Pyle, R.M. 2015. Monarchs in the mist. Pp. 236-246. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Ramirez, M.I., C. Saenz-Romero, G. Rehfeld et L. Salas-Canela. 2015. Threats to the availability of overwintering habitat in the Monarch Butterfly Biosphere Reserve. Pp. 157-168. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Rendón-Salinas, E., A. Fajardo-Arroyo et G. Tavera-Alonso. 2014. Forest surface occupied by Monarch butterfly hibernation colonies in December 2014 [PDF, 556 Ko]. Site Web : [Consulté le 12 avril 2016]. [en anglais seulement]
Ries, L., D.J. Taron et E. Rendon-Salinas. 2015a. The disconnect between summer and winter Monarch trends for the eastern migratory population: possible links to differing drivers. Annals of the Entomological Society of America 1–9.
Ries, L., D.J. Taron, E. Rendon-Salinas et K.S. Oberhauser. 2015b. Connecting eastern Monarch population dynamics across their migratory cycle. Pp. 268-281. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Riley, J.L. 2013. The Once and Future Great Lakes Country. An Ecological History. McGill – Queen’s University Press.
Robinson, R.A., H.Q.P. Crick, J.A. Learmonth, I.M.D. Maclean, C.D. Thomas, F. Bairlein, M.C. Forchhammer, C.M. Francis, J.A. Gill, B.J. Godley, J. Harwood, G. C. Hays, B. Huntley, A.M. Hutson, G.J. Pierce, M.M. Rehfisch, D.W. Sims, M.B. Santos, T H. Sparks, D.A. Stroud et M.E. Visser. 2009. Travelling through a warming world: climate change and migratory species. Endangered Species Research (7): 87-99.
Salafsky, N., D. Salzer, A. J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O’Connor et D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conservation Biology 22:897-911. Site Web : http://conservationmeasures.org/CMP/IUCN/browse.cfm?TaxID=DirectThreats [Consulté le 10 avril 2016].
Satterfield D.A., J.C. Maerz et S. Altizer. 2015. Loss of migratory behaviour increases infection risk for a butterfly host. Proc. R. Soc. B 282: 20141734. [en anglais seulement]
Schappert, P. 2004. Monarch Butterflies: Saving the King of the New World. Key Porter Books, Toronto, ON. 128 pp.
Scott, J.A. 1986. The Butterflies of North America: A Natural History and Field Guide. Stanford University Press, Stanford, California. 583 pp.
Semmens, B.X., D.J. Semmens, W.E. Thogmartin, R. Wiederholt, L. López-Hoffman, J.E. Diffendorfer, J.M. Pleasants, K.S. Oberhauser et O.R. Taylor. 2016. Quasi-extinction risk and population targets for the Eastern, migratory population of monarch butterflies (Danaus plexippus). Scientific Reports 6, Article number: 23265 doi:10.1038/srep23265
Skórkaa, P., M. Lendab, D. Morońc, K. Kalarusd, P. Tryjanowskia. 2013. Factors affecting road mortality and the suitability of road verges for butterflies. Biological Conservation 159: 148–157.
Slayback, D.A., L.P. Brower, M.I. Ramírez et L.S. Fink. 2007. Establishing the presence and absence of overwintering colonies of the Monarch Butterfly in Mexico by the use of small aircraft. American Entomologist: 27-40.
Smith, D.A.S., G. Lushai et J.A. Allen. 2005. A classification of Danaus butterflies (Lepidoptera: Nymphalidae) based upon data from morphology and DNA. Zoological Journal of the Linnean Society 144(2):191-212.
Snell-Rooda, E.C., A. Espeseta, C.J. Bosera, W.A. White et R. Smykalskia. 2014. Anthropogenic changes in sodium affect neural and muscle development in butterflies. PNAS 111(28) 10221–10226, doi: 10.1073/pnas.1323607111
Snook, L.C. 1993. Conservation of the Monarch butterfly reserves in Mexico: Focus on the Forest. Pages 363-375 In Malcom, S. B. et M. P. Zalucki (eds.), Biology and conservation of the Monarch butterfly. Publications of the Natural History Museum of Los Angeles County, Los Angeles, California. Science series no.38.
Solensky, M.J. 2004. Overview of monarch migration. Pp. 79-83. in K.S. Oberhauser et M.J. Solensky (eds).The Monarch Butterfly: Biology and Conservation. Cornell University Press, Ithaca New York.
Southwest Monarch Study. 2013. Consulté en février 2017.
St. John, D, E. Antonijevic et J. Heron. 2014. Surveys for the Monarch Butterfly (Danaus plexippus) and Showy Milkweed (Asclepias speciosa) in the Okanagan and Similkameen river valleys of British Columbia. Rapport inédit. B.C. Ministry of Environment, Vancouver, B.C. 53pp.
Stevens, S.R. et D.F. Frey. 2010. Host plant pattern and variation in climate predict the location of natal grounds for migratory monarch butterflies in western North America. Insect Conserv. 14:731–744.
Suzuki, Y. et M.P. Zalucki. 1986. The influence of sex ratio on female dispersal in Danaus plexippus (L.)(Lepidopetra: Danaidae). Journal of the Australian Entomological Society 25: 31-35.
The Center for Biological Diversity, Center for Food Safety, The Xerces Society & L. Brower 2014. Petition to protect the Monarch Butterfly (Danaus plexippus plexippus) under the Endangered Species Act. Site Web : http://xerces.org/wp-content/up loads/2014/08/monarch-esa-petition.pdf. [Consulté le 10 avril 2016].
Thomas, A.W. 1996. A Preliminary Atlas of the Butterflies of New Brunswick. Natural Resources Canada, Canadian Forest Service, Atlantic Forestry Centre, Fredericton, New Brunswick. New Brunswick Museum Publications in Natural Science 11. 41 pp.
United Nations Educational, Scientific and Cultural Organization [PDF, 1.73 Mb] (UNESCO). 2015. State of conservation of properties inscribed on the World Heritage List. Site Web : http://whc.unesco.org/archive/2015/whc15-39com-7BAdd-en.pdf [Consulté le 13 avril 2016]. (Également disponible en français : Organisation des Nations Unies pour l’éducation, la science et la culture [PDF, 1,72 Mo] (UNESCO). 2015. Etat de conservation de biens inscrits sur la Liste du patrimoine mondial.
Urquhart, F. A.1987. The Monarch Butterfly: International Traveller. Nelson-Hall, Chicago. 42 pp.
van der Sluijs, J.P., N. Simon-Delso, D. Goulson, L. Maxim, J.M. Bonmatin et L.P. Belzunces. 2013. Neonicotinoids, bee disorders and the sustainability of pollinator services. Current Opinion in Environmental Sustainability. 5:293–305.
Vane-Wright, R.I. 1993. The Columbus hypothesis: an explanation for the dramatic 19th century range expansion of the monarch butterfly. In S.B. Malcolm & M. Zalucki (eds), Biology and conservation of the monarch butterfly, pp. 179–187. Science Series (38), Natural History Museum of Los Angeles County, Los Angeles.
Vidal, O., J. López-García et E. Rendón-Salinas. 2014. Trends in deforestation and forest degradation after a decade of monitoring in the Monarch Butterfly Biosphere Reserve in Mexico. Conserv Biol. 28(1): 177–186.
Vidal, O. et E. Rendón-Salinas. 2014. Dynamics and trends of overwintering colonies of the monarch butterfly in Mexico. Biological Conservation 180: 165 – 175.
White, D.J. 1996. The milkweeds of Canada (Asclepias), status, distribution, and potential impacts from noxious weeds legislation. Report prepared for the CWS, consulté le from Monarch Watch, June 14, 2008.
Williams, E.H. et L.P. Brower. 2015. Microclimatic protection of overwintering Monarchs provided by Mexico’s high elevation Oyamel Fir forests. Pp. 109-116. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Williams, J., D. Stow et L. Brower, 2007. the influence of forest fragmentation on the location of overwintering Monarch butterflies in central Mexico, Journal of the Lepidopterist’s Society 61: 90-104.
Wormington, A. comm. pers. 2007. Communication par courriel adressée à K. Bériault, janvier 2007.
Wormington, A. 1994. A mass migration of Monarchs at Point Pelee, Ontario. Toronto Entomologists Association, Occasional Publication #26-94, pp. 26-27.
Wormington, A. 1997. Another mass migration of Monarchs at Point Pelee, Ontario. Toronto Entomologists Association, Occasional Publication #29-97, p. 11.
Wormington, A. 2008. The Butterflies of Point Pelee National Park, Ontario. Unpublished Manuscript.
World Wildlife Fund (WWF). 2015. 96% of all deforestation within Mexico’s Monarch butterfly sanctuaries occur in a single community. Site Web : [Consulté le 11 avril 2016]. [en anglais seulement]
York, H.A. et K.S. Oberhauser. 2002. Effects of duration and timing of heat stress on monarch butterfly (Danaus plexippus) (Lepidoptera: Nymphalidae) development. Journal of the Kansas Entomological Society, Vol. 75 (4): 290-298.
Zalucki, M.P.et W.A. Rochester. 2004. Spatial and temporal population dynamics of Monarchs Down-Under: lessons for North America. Pp. 219-228. In The Monarch Butterfly: Biology and Conservation. K.S. Oberhauser et M.J. Solensky (eds). Cornell University Press, Ithaca, New York.
Zalucki, M.P., L.P. Brower, S.B. Malcolm et B.H Slager. 2015. Estimating the climate signal in Monarch population decline. Pp. 130-141. in K.S. Oberhauser, K.R. Nail et S. Altizer (eds.). Monarchs in a Changing World: Biology and Conservation of an Iconic Butterfly. Cornell University Press, Ithaca New York.
Zhan, S., W. Zhang, K. Niitepold, J. Hsu, J. Fernandez Haeger, M.P. Zalucki, S. Altizer, J.C. de Roode, S.M. Reppert and M.R. Kronforst. 2014. The genetics of monarch butterfly migration and warning colouration. Nature 514: 317-321.
Zipkin, E. F., L. Ries, R. Reeves, J. Regetz et K. Oberhauser. 2012. Tracking climate impacts on the migratory monarch butterfly. Global Change Biology 18: 3039 – 3049.
Sommaire biographique des rédactrices du rapport
Allan Harris compte plus de 25 années d’expérience comme biologiste dans le nord de l’Ontario. Il détient un diplôme de baccalauréat (B.Sc.) en biologie de la faune de l’Université de Guelph (University of Guelph) et un diplôme de maîtrise (M.Sc.) en biologie de l’Université Lakehead. Après avoir occupé un poste de biologiste au ministère des Richesses naturelles de l’Ontario pendant sept ans, il a cofondé Northern Bioscience, une entreprise de consultation écologique établie à Thunder Bay, en Ontario. Allan Harris est auteur ou coauteur de douzaines d’articles scientifiques, de rapports techniques et d’articles de vulgarisation, y compris des rapports de situation du COSEPAC sur le gomphe riverain, le gomphe de Laura, le gomphe des rapides, la cicindèle à grandes taches de Gibson, la cicindèle verte des pinèdes, l’hespérie de Powesheik, le mormon, l’amiral de Weidemeyer, l’hémileucin du ményanthe, le perceur du ptéléa, la gnaphose de Snohomish, l’escargot-forestier à larges bandes, l’aster de la Nahanni, l’aster fausse-prenanthe, la buchnéra d’Amérique, le trille à pédoncule incliné et le lipocarphe à petites fleurs. Il est également auteur d’un rapport provincial sur la situation du caribou des bois en Ontario et auteur ou coauteur de programmes de rétablissement nationaux et provinciaux visant des espèces de plantes vasculaires et d’oiseaux en péril.
Robert Foster est cofondateur et partenaire principal de Northern Bioscience, entreprise qui offre des services professionnels de consultation écologique en appui aux travaux de gestion, de planification et de recherche sur les écosystèmes. Il est titulaire d’un baccalauréat en biologie (B.Sc) de l’Université Lakehead et d’un doctorat en zoologie (D.Phil.) de l’Université d’Oxford (Oxford University). Robert Forster a travaillé pendant plus de 20 ans comme écologiste en Ontario. Il est auteur ou coauteur de rapports de situation du COSEPAC sur le gomphe riverain, le gomphe de Laura, le gomphe des rapides, la cicindèle à grandes taches de Gibson, la cicindèle verte des pinèdes, l’hespérie de Powesheik, le mormon, l’amiral de Weidemeyer, l’hémileucin du ményanthe, le perceur du ptéléa, la gnaphose de Snohomish, l’escargot-forestier à larges bandes, l’aster de la Nahanni, l’aster fausse-prenanthe, la buchnéra d’Amérique et le trille à pédoncule incliné, de même que des programmes de rétablissement ciblant des espèces de plantes, de lichens et d’odonates rares. Robert Forster a réalisé de nombreux relevés d’odonates et d’autres insectes à des fins de planification et d’évaluation environnementale d’aires protégées en Ontario, ainsi qu’au Manitoba, au Minnesota, au Québec et en Colombie-Britannique.
Collections examinées
Aucune.