
Steelhead Trout (Oncorhynchus mykiss), Thompson River and Chilcotin River populations in Canada, 2018: COSEWIC Technical summaries and supporting information for emergency assessments
- Table of contents
- COSEWIC assessment summary - Thompson River population
- COSEWIC assessment summary - Chilcotin River population
- COSEWIC executive summary
- Introduction
- Taxonomic structure and designatable units
- Available information for the assessment
- Threats
- Assessment results
- Rescue effect
- Acknowledgements
- Technical summary - Thompson River population
- Technical summary - Chilcotin River population
- Literature cited
- Figure 1. Trend in the number of mature individuals in the Thompson River Steelhead Trout DU, 1978-2018, and the fitted log-linear regression through the last 3 generations (5 year generation time).
- Figure 2. Trend in the number of mature individuals in the Chilcotin River Steelhead Trout DU, 1972-2018, and the fitted log-linear regression through the last three generations (6 year generation time).
- Figure A1. Approximate range of Steelhead Trout in BC (left of red line including all coastal islands).
- Figure A2. Distribution of major phylogenetic groups (MPG) of Oncorhynchus mykiss in BC (based on McCusker et al. 2000).
- Figure A3. Location of Chilcotin River (CRS) and Thompson River Steelhead Trout (TRS; black arrows) within BC and major phylogenetic groupings (see Figure A2).
- Figure A4. Approximate locations of populations of Chilcotin River Steelhead Trout (CRS) and Thompson River Steelhead Trout (TRS) within the context of climate zones of BC as measured by mean annual air temperature (climate map from ClimateBC 2017).
Steelhead Trout (Oncorhynchus mykiss), Thompson River and Chilcotin River populations in Canada, 2018: COSEWIC Technical summaries and supporting information for emergency assessments

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
In British Columbia, Oncorhynchus mykiss occurs as two evolutionary lineages, commonly referred to as “coastal” and “interior” O. mykiss. Both lineages of O. mykiss are found in freshwater-resident and anadromous (sea run) populations or life-history types, known as Rainbow Trout and Steelhead Trout, respectively. Interior O. mykiss are found in the Thompson-Chilcotin rivers (part of the Fraser River drainage). There is some interbreeding between freshwater-resident and anadromous individuals and freshwater-resident individuals may produce anadromous offspring and vice versa.
The anadromy of Steelhead Trout and their older age (and larger size) at maturity are significant aspects of their life history that set them apart from freshwater-resident Rainbow Trout. Thompson and Chilcotin Steelhead Trout are likely to have evolved from fish isolated in the Columbia refugium during the last glaciation while other Canadian Steelhead Trout may have arisen from the Haida Gwaii refugium. Based on genetic data, Steelhead Trout in the Thompson and Chilcotin rivers are discrete from all other Canadian Steelhead Trout, and also differ from each other. Thus, the interior Fraser River Steelhead Trout satisfy COSEWIC criteria to be assessed as two separate designatable units (DUs) or populations: Thompson River population and Chilcotin River population.
Thompson and Chilcotin Steelhead Trout have been the subject of considerable recent public concern, including the submission of an application for an Emergency Assessment in January 2016. Such input heightened concern for Thompson and Chilcotin Steelhead Trout already held by COSEWIC, and a status report was expected to be initiated in 2018. The most recent information on returns of adult fish, however, indicates that the situation is worsening and constituted an emergency. This resulted in an additional application for an Emergency Assessment submitted in November 2017.
The number of mature fish that have returned to fresh water from the sea in the fall of 2017, and that will spawn in the spring of 2018, are 177 and 58 for the Thompson and Chilcotin rivers, respectively. The average annual number of mature individuals returning to the Thompson and Chilcotin rivers in the last three years (2016-2018) is the lowest in a time series that began in 1978. The decline of mature individuals in the Thompson River over the last three generations (15 years) is 79%, and the decline of the Chilcotin River Steelhead Trout over three generations (18 years) is 81%. Bycatch mortality in commercial Pacific salmon fisheries and declines in marine and freshwater habitat quality are the key factors driving the declines.
The Emergency Assessment was conducted on January 10, 2018. The participants in the Emergency Assessment considered these data, and concluded that the status both of the Thompson River and Chilcotin River DUs of Steelhead Trout is Endangered and constitutes an emergency situation. This report documents the background material used during the Emergency Assessment and the conclusions reached.
In response to record low returns of Thompson/Chilcotin Steelhead Trout and public concerns over their conservation status, COSEWIC conducted an Emergency Assessment (EA) of those wildlife species on January 10, 2018. There were 31 participants in the EA and their names and affiliations are listed in Appendix One. This report provides a summary of the information reviewed by participants, and the conclusions of the EA. Pending the decision regarding COSEWIC’s recommendation that an Emergency Order be made (see below), a full COSEWIC status report will be produced on an expedited basis, and within one year of an Emergency Order being made as per the Species at Risk Act (s30.1).
The taxon Oncorhynchus mykiss exhibits two broad life-history types: a lake- and stream-resident form known as Rainbow Trout and an anadromous (sea run) form known as Steelhead Trout (McPhail 2007). Depending on the geographic context (e.g., distance from the sea, presence of migration barriers, presence of lakes within a watershed), one or both forms may occur in a single watershed and even in the same tributary within a watershed. Again, depending on geographic context, the forms may exist separately, co-exist at the same place and time as juveniles and spawning adults, or their ranges may be adjacent to one another (McPhail 2007). Predictably, there is also a variable degree of demographic and genetic interaction between the forms where they co-exist. In some instances, there is little detectable genetic differentiation between the forms and in other instances they may represent genetically-distinct populations (Docker and Heath 2003; McMillan et al. 2007; Pearse et al. 2009). Furthermore, there is evidence that in some systems, Steelhead Trout may be produced from Rainbow Trout mothers and some Steelhead Trout offspring may remain permanently in fresh water (termed “residuals”), especially when they experience faster growth as juveniles, e.g. as often occurs in hatchery-supplemented populations (Viola and Schuck 1995; Zimmerman and Reeves 2000; Thrower et al. 2004). These variable relationships between Steelhead and Rainbow Trout are also found in other salmonid species such as O. nerka where there are freshwater-resident (“Kokanee”) and anadromous forms (“Sockeye Salmon”) and Salmo salar (with freshwater “Ouananiche” and anadromous “Atlantic Salmon” forms). In the context of Thompson and Chilcotin rivers’ Steelhead Trout, there is no information on the genetic relationship between the two life-history forms. There is some evidence that Steelhead Trout in these systems may be produced from Rainbow Trout mothers (R. Bison, BC Ministry of Forests, Lands, Natural Resource Operations & Rural Development, Kamloops, BC, pers. comm.), but the spatial and temporal extent of this phenomenon is not well understood. Accordingly, and consistent with recent status assessments both for Atlantic Salmon (COSEWIC 2010) and Sockeye Salmon (COSEWIC 2018), this assessment of interior Fraser River O. mykiss concerns only Steelhead Trout. The COSEWIC approach is also consistent with that of USA fisheries management agencies; here, anadromous and freshwater-resident forms of O. mykiss are assessed separately (Hard et al. 2015).
Steelhead Trout in the Thompson River and Chilcotin River are discrete from other Canadian Steelhead Trout based on genetic data, and also differ from each other. Thompson and Chilcotin Steelhead Trout likely evolved from fish isolated in the Columbia refugium during the last glaciation while other Canadian Steelhead Trout may have arisen from the Haida Gwaii refugium. After reviewing available information on Designatable Units (DU) for Thompson/Chilcotin Steelhead Trout and applying the COSEWIC criteria of discreteness and significance, EA participants agreed that Thompson and Chilcotin Steelhead Trout should be assessed as two DUs separate from all other BC Steelhead Trout populations: Thompson River DU and Chilcotin River DU (see Appendix Two for additional details).
Annual monitoring information is available from a test fishery conducted in the Fraser River about 60 km upstream from the ocean (near Albion, BC). The fishing gear used is conventional salmon gillnets. A large mesh gillnet (8 inch) and a slightly smaller mesh gillnet (6.75 inch) are used on alternate days and on consistent tide stages during the early and peak time of Interior Fraser Steelhead Trout migration at that site. During the latter stage of the migration, only the smaller mesh gillnet is used on a daily basis. The catch statistic used in the forecasting of spawning (breeding) fish abundance is simply the number of Steelhead Trout caught per day.
For some of the tributaries of the Thompson and Chilcotin rivers where Steelhead Trout spawn, instream counts and estimates are also conducted. For the Thompson River tributaries, automated fish counters are used in the Deadman and Bonaparte rivers and periodic boat-based visual counts are used in a major tributary of the Nicola River watershed. These visual counts are combined in a maximum likelihood estimate model with observer efficiency, timing and spatial distribution estimates from external tagging and radio tagging to estimate abundance of Steelhead Trout in the Nicola River watershed, which includes estimates for the Coldwater River, Spius Creek and the lower Nicola River (Bison and Phelps 2017). For the Chilko River (a tributary of the Chilcotin River), periodic visual (helicopter based) counts are conducted over a distance of 24 km (i.e., from Brittany Creek confluence with the Chilko River up to Chilko Lake; Bison and Phelps 2017).
Catch (and release) in the sport fishery is estimated by random stratified on-the-ground angler surveys, where about one-third of the total effort is surveyed (Bison and Phelps 2017). A secondary estimate is based on an annual post-season angler questionnaire survey conducted province-wide by Fish & Wildlife Branch (Bison and Phelps 2017).
Bycatch in the commercial Pacific salmon fisheries is estimated indirectly, using trends and level of encounter rates and fishing mortality rates are estimated with the use of a simulation model (Bison 2016).
The trends in annual abundances of spawning fish for major tributaries of the Thompson and Chilcotin River DUs both show dramatic declines since the early 2000s (Figures 1 and 2). The rates of decline over three generations are 79% and 81% for the Thompson and Chilcotin DUs, respectively.
For the Thompson River DU, if the current rate of decline persists for another three generations, the number of spawning fish will decline to 37, which is 2% of the pre-2000 abundance. For the Chilcotin River DU, if the current rate of decline persists for another three generations, the number of spawning fish will decline to 11, which is about 0.9% of the pre-2000 abundance.
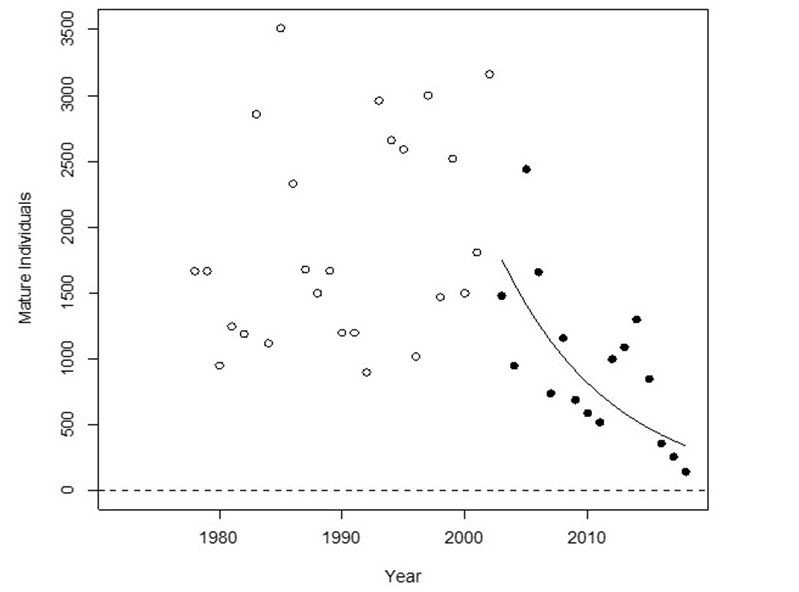
Long description for Figure 1
Chart illustrating the trend in the number of mature individuals in the Thompson River Steelhead Trout designatable unit for 1978 to 2018 and the fitted log-linear regression through the last three generations. The solid data points shown after the year 2000 were used in the decline estimate of 79 percent.
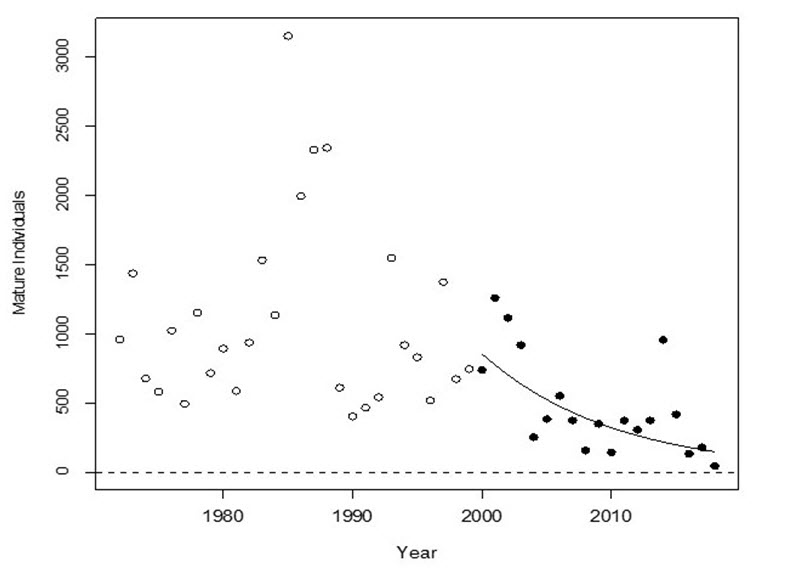
Long description for Figure 2
Chart illustrating the trend in the number of mature individuals in the Chilcotin River Steelhead designatable unit, from 1972 to 2018, and the fitted log-linear regression through the last three generations (six year generation time). Solid data points shown starting in the year 2000 were used in the decline estimate of 81 percent.
Reduced marine survival of Steelhead Trout is considered to be a key factor driving population declines since the early 1990s (Kendall et al. 2017). Similar poor ocean survival-based declines have been reported in recent COSEWIC reports for Sockeye Salmon and Coho Salmon (O. kisutch). Although relationships between marine temperature and survival have been identified, the underlying causal mechanisms driving the relationships are poorly understood for Steelhead Trout. Ocean temperatures have warmed an average of 0.5°C over the past two decades and have likely contributed to declining survival of Steelhead Trout as has been suggested for Sockeye Salmon (Hinch and Martins 2011). Ocean temperatures in the Gulf of Alaska where Steelhead Trout spend much of their marine life are predicted to increase 1-2°C by the 2040s (Abdul-Aziz et al. 2011). Berejikian et al. (2016) suggested that predation by Harbour Seals (Phoca vitulina) contributed to mortality of migrating juvenile Steelhead Trout off Washington State, and they hypothesized that documented changes in the Puget Sound ecosystem may currently put Steelhead Trout at greater risk of predation by Harbour Seals and possibly other predators.
Bycatch of returning mature fish in purse seine and gillnet fisheries directed at Pacific salmon is a better-quantified threat compared with marine survival. There are no directed commercial fisheries for Steelhead Trout in BC and the sport fishery operates on a catch-and-release basis with closures if in-season abundance estimates are below pre-determined limits. The estimated mortality rate from all bycatch in commercial fisheries is in the range of 15-25% annually (Bison 2016). This alone could explain a large proportion of the observed decline in mature individuals.
While it is generally considered that the quality of freshwater habitat is declining, the severity of the freshwater habitat-based threats in the Thompson and Chilcotin rivers is not well understood.
The EA concluded that for the Thompson River DU, a designation of Endangered applies (Endangered A2bd+4bd; C2a(i); D1 - see Technical Summary 1). For the Chilcotin River DU, the EA also concluded that Endangered applies (Endangered A2bd+4bd; C2a(i,ii); D1 - see Technical Summary 2).
The EA also used RAMAS Red List V3.0 (RAMAS Software by Applied Biomathematics) software for a rapid assessment using International Union for the Conservation of Nature (IUCN) criteria. For both DUs, the RAMAS procedure using the most recent spawning adult abundance estimates from the Province of BC and the decline rates used in this report indicated a “Critically Endangered” IUCN status.
As noted earlier, freshwater-resident Rainbow Trout may produce offspring that become anadromous (e.g., Zimmerman and Reeves 2000). The same literature, however, also indicates that the phenomenon is a watershed-specific characteristic, and the extent to which this occurs within the Thompson and Chilcotin watersheds is not well known. Regardless, given the observed declines (figure 1 and figure 2), there is no evidence that any potential contribution of resident Rainbow Trout is mitigating the recent precipitous decline in Steelhead Trout or that it might do so in the future.
The authors thank the many COSEWIC members and members of the Marine Fishes Species Specialist subcommittee who contributed to this report. Alan Sinclair, John Reynolds, John Post, Greg Wilson, and Robert Bison (BC Ministry of Forests, Lands, Natural Resource Operations and Rural Development) led the preparation of material supporting the Emergency Assessment. They also thank all in the COSEWIC secretariat who have contributed to this urgent matter.
| Summary items | Information |
|---|---|
| Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines (2011) is being used). | 5 yrs |
| Is there an [observed, inferred, or projected] continuing decline in number of mature individuals? | Yes |
| Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations] | 71% decline in last 2 generations |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations]. | 79% decline in last 3 generations |
| [Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations]. | If the current rate of decline persists for another 3 generations, the number of spawning fish will decline to 37, which is 2.0% of the pre-2000 abundance. |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future. | 79% decline inferred over this time period assuming the same decline rate as in the last 3 generations |
| Are the causes of the decline a. clearly reversible and b. understood and c. ceased? | a. Partially if bycatch fishing mortality is reduced. b. Bycatch mortality well understood but declines in marine and freshwater environments less so. c. No |
| Are there extreme fluctuations in number of mature individuals? | No |
| Summary items | Information |
|---|---|
| Estimated extent of occurrence (EOO) | > 20,000 km2 |
| Index of area of occupancy (IAO) (Always report 2x2 grid value.) |
< 500 km2 |
| Is the population “severely fragmented” i.e., is >50% of its total area of occupancy in habitat patches that are (a) smaller than would be required to support a viable population, and (b) separated from other habitat patches by a distance larger than the species can be expected to disperse? | a. No b. No |
| Number of “locations” See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term. (use plausible range to reflect uncertainty if appropriate) |
NA |
| Is there an [observed, inferred, or projected] decline in extent of occurrence? | No |
| Is there an [observed, inferred, or projected] decline in index of area of occupancy? | No |
| Is there an [observed, inferred, or projected] decline in number of subpopulations? | No |
| Is there an [observed, inferred, or projected] decline in number of “locations” See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.? |
NA |
| Is there an [observed, inferred, or projected] decline in [area, extent and/or quality] of habitat? | Yes, there is an inferred decline in habitat quality. |
| Are there extreme fluctuations in number of subpopulations? | No |
| Are there extreme fluctuations in Number of “locations” See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.? |
No |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | No |
| Subpopulations (give plausible ranges) | N mature individuals |
|---|---|
| Thompson River, includes spawning in the following tributaries: Deadman, Bonaparte, Coldwater rivers and Spius Creek and Nicola River in most recent survey year (2017). | 177 (the average of last 3 years is 255) |
| Total | 177 |
| Summary items | Information |
|---|---|
| Is the probability of extinction in the wild at least [20% within 20 years or 5 generations, or 10% within 100 years]? | Not calculated |
| Summary items | Information |
|---|---|
| Was a threats calculator completed for this species? No. This population faces a number of threats in the marine and freshwater environments, many of which are similar to Sockeye Salmon in the Fraser River drainage and Coho Salmon in the Interior Fraser River. Fishery removals for the interior Fraser River Steelhead Trout vary from 15-25% per year, depending on the abundance of Pacific salmon targeted in commercial fisheries. What additional limiting factors are relevant? |
|
| Summary items | Information |
|---|---|
| Status of outside population(s) most likely to provide immigrants to Canada. | Thompson Steelhead Trout are endemic to this watershed and rescue is not possible from other Steelhead Trout populations. Rescue from non-anadromous trout within this watershed is unlikely. |
| Is immigration known or possible? | NA |
| Would immigrants be adapted to survive in Canada? | NA |
| Is there sufficient habitat for immigrants in Canada? | NA |
| Are conditions deteriorating in Canada? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
NA |
| Are conditions for the source (i.e., outside) population deteriorating? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
NA |
| Is the Canadian population considered to be a sink? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
NA |
| Is rescue from outside populations likely? | NA |
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
| Summary items | Information |
|---|---|
| COSEWIC | Not previously assessed |
| Summary items | Information |
|---|---|
| Recommended Status | Endangered |
| Alpha-numeric codes | A2bd+4bd; C2a(i); D1 |
| Reasons for designation | This wildlife species faces a number of threats, including declining habitat quality both in marine and freshwater environments, and bycatch mortality from Pacific salmon fisheries. The number of spawning fish was variable with little trend prior to 2000. Since then, the population has declined dramatically (79%) over the last three generations and it is now the lowest on record. The 177 mature fish observed in the most recent survey are only about 9.5% of the pre-2000 mean. If the current rate of decline persists for another three generations, the number of spawning fish will decline to 37, which is 2.0% of the pre-2000 abundance. |
| Summary items | Information |
|---|---|
| Criterion A (Decline in Total Number of Mature Individuals) | Meets Endangered A2bd+4bd. The number of mature individuals has declined by 79% over the past 3 generations and it is inferred that this decline will continue into the future. |
| Criterion B (Small Distribution Range and Decline or Fluctuation) | Does not meet criterion. IAO meets criterion for Endangered and the quality of the freshwater and marine habitats is declining, but the population is not severely fragmented, the criterion for restricted number of locations does not apply and there are no extreme fluctuations. |
| Criterion C (Small and Declining Number of Mature Individuals) | Meets Endangered C2a(i). The number of mature individuals for all subpopulations is 177 in the most recent survey year, and no subpopulation is estimated to have more than 250 individuals. |
| Criterion D (Very Small or Restricted Population) | Meets Endangered D1 because the number of mature individuals in the last survey year is 177 (most recent 3 year average is 255). |
| Criterion E (Quantitative Analysis) | Not done. |
| Generation time (usually average age of parents in the population; indicate if another method of estimating generation time indicated in the IUCN guidelines (2011) is being used). | 6 yrs |
| Is there an [observed, inferred, or projected] continuing decline in number of mature individuals? | Yes |
| Estimated percent of continuing decline in total number of mature individuals within [5 years or 2 generations] | 59% decline in last 2 generations |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over the last [10 years, or 3 generations]. | 81% decline in last 3 generations |
| [Projected or suspected] percent [reduction or increase] in total number of mature individuals over the next [10 years, or 3 generations]. | If the current rate of decline persists for another 3 generations, the number of spawning fish would decline to 11, or about 0.9% of the pre-2000 abundance. |
| [Observed, estimated, inferred, or suspected] percent [reduction or increase] in total number of mature individuals over any [10 years, or 3 generations] period, over a time period including both the past and the future. | 81% decline inferred over three generations assuming the same decline rate as in the last 3 generations |
| Are the causes of the decline a. clearly reversible and b. understood and c. ceased? | a. Partially if bycatch fishing mortality is reduced. b. Bycatch mortality well understood but declines in marine and freshwater environments less so. c. No |
| Are there extreme fluctuations in number of mature individuals? | No |
| Summary items | Information |
|---|---|
| Estimated extent of occurrence (EOO) | > 20,000 km2 |
| Index of area of occupancy (IAO) (Always report 2x2 grid value). |
< 500 km2 |
| Is the population “severely fragmented” i.e., is >50% of its total area of occupancy in habitat patches that are (a) smaller than would be required to support a viable population, and (b) separated from other habitat patches by a distance larger than the species can be expected to disperse? | a. No b. No |
| Number of “locations” See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term. (use plausible range to reflect uncertainty if appropriate) |
NA |
| Is there an [observed, inferred, or projected] decline in extent of occurrence? | No |
| Is there an [observed, inferred, or projected] decline in index of area of occupancy? | No |
| Is there an [observed, inferred, or projected] decline in number of subpopulations? | No |
| Is there an [observed, inferred, or projected] decline in Number of “locations” See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term. |
NA |
| Is there an [observed, inferred, or projected] decline in [area, extent and/or quality] of habitat? | Yes, there is an inferred decline in habitat quality. |
| Are there extreme fluctuations in number of subpopulations? | No |
| Are there extreme fluctuations in Number of “locations” See Definitions and Abbreviations on COSEWIC website and IUCN (Feb 2014) for more information on this term.? |
No |
| Are there extreme fluctuations in extent of occurrence? | No |
| Are there extreme fluctuations in index of area of occupancy? | No |
| Subpopulations (give plausible ranges) | N mature individuals |
|---|---|
| Chilcotin River, including the following spawning tributaries: Taseko, Chilko and Little Chilcotin rivers in the most recent survey year (2017) | 58 (average of last 3 years is 120) |
| Total | 58 |
| Summary items | Information |
|---|---|
| Is the probability of extinction in the wild at least [20% within 20 years or 5 generations, or 10% within 100 years]? | Not calculated |
| Summary items | Information |
|---|---|
| Was a threats calculator completed for this species? No. This population faces a number of threats in the marine and freshwater environments, many of which are similar to Sockeye Salmon in the Fraser River drainage and Coho Salmon in the Interior Fraser River. Fishery removals for the interior Fraser Steelhead Trout vary from 15-25% per year, depending on the abundance of salmon targeted in commercial fisheries.What additional limiting factors are relevant? |
|
| Summary items | Information |
|---|---|
| Status of outside population(s) most likely to provide immigrants to Canada. | Chilcotin Steelhead Trout are endemic to this watershed and rescue is not possible from other Steelhead Trout populations. Rescue from non-anadromous trout within this watershed is unlikely. |
| Is immigration known or possible? | NA |
| Would immigrants be adapted to survive in Canada? | NA |
| Is there sufficient habitat for immigrants in Canada? | NA |
| Are conditions deteriorating in Canada? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
NA |
| Are conditions for the source (i.e., outside) population deteriorating? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
NA |
| Is the Canadian population considered to be a sink? See Table 3 (Guidelines for modifying status assessment based on rescue effect). |
NA |
| Is rescue from outside populations likely? | NA |
| Summary items | Information |
|---|---|
| Is this a data sensitive species? | No |
| Summary items | Information |
|---|---|
| COSEWIC | Not previously assessed |
| Summary items | Information |
|---|---|
| Recommended Status | Endangered |
| Alpha-numeric codes | A2bd+4bd; C2a(i,ii); D1 |
| Reasons for designation | This wildlife species faces a number of threats, including declining habitat quality both in marine and freshwater environments, and bycatch mortality from Pacific salmon fisheries. The population has declined dramatically (81%) over the last three generations and it is now the lowest on record. The number of spawning fish was high and variable with little trend prior to 2000. The 58 mature fish observed in the most recent survey are only 5% of the pre-2000 mean. If the current rate of decline persists for another three generations, the number of spawning fish will decline to 11, which is 0.9% of the pre-2000 abundance. |
| Summary items | Information |
|---|---|
| Criterion A (Decline in Total Number of Mature Individuals) | Meets Endangered A2bd+4bd. The number of mature individuals has declined by 81% over the past 3 generations and it is inferred that this decline will continue into the near future. |
| Criterion B (Small Distribution Range and Decline or Fluctuation) | Does not meet criterion. IAO meets criterion for Endangered and the quality of the freshwater and marine habitats is declining, but the population is not severely fragmented, the criterion of restricted number of locations does not apply and there are no extreme fluctuations. |
| Criterion C (Small and Declining Number of Mature Individuals) | Meets Endangered C2a(i,ii). The number of mature individuals is 58 in the most recent survey year, there has been a 59% decline in the number of mature individuals over the last 2 generations, no subpopulation has more than 250 individuals, and one subpopulation has more than 95% of all mature individuals. |
| Criterion D (Very Small or Restricted Population) | Meets Endangered D1 because the number of mature individuals is less than 250 (most recent year is 58, with an average of 120 over the most recent three years). |
| Criterion E (Quantitative Analysis) | Not done |
| Participant | Role |
|---|---|
| Eric Taylor | COSEWIC Chair |
| John Neilson | Co-chair Marine Fishes |
| Dwayne Lepitzki | Co-chair Molluscs |
| Dave Fraser | Jurisdiction - BC |
| Greg Wilson | Jurisdiction - BC |
| Syd Cannings | Jurisdiction - CWS |
| Jennifer Shaw | Jurisdiction - DFO |
| Simon Nadeau | Jurisdiction - DFO |
| Robert Bison | British Columbia government area fishery specialist |
| John Post | Co-chair Freshwater Fishes |
| John Reynolds | Non-government Science member |
| Alan Sinclair | EA SSC special member or observer |
| Arne Mooers | Non-government Science member |
| Donna Hurlburt | Co-chair ATK |
| Roger Gallant | Co-chair ATK |
| Paul Grant | Co-chair Arthropods |
| Ross Claytor | Co-chair Marine Fishes |
| Aaron McNeil | Marine Fishes SSC member |
| Bruce Atkinson | Marine Fishes SSC member |
| Craig Purchase | Marine Fishes SSC member |
| David Hardie | Marine Fishes SSC member |
| Ian Fleming | Marine Fishes SSC member |
| Laura Weir | Marine Fishes SSC member |
| Margaret Treble | Marine Fishes SSC member |
| Nancy Shackell | Marine Fishes SSC member |
| Peter Westley | Marine Fishes SSC member |
| Carrie Holt | Marine Fishes SSC member |
| Marc Trudel | Marine Fishes SSC member |
| Bev McBride | Secretariat |
| Karen Timm | Secretariat |
| Lisa Twolan | Secretariat |
British Columbia contains myriad populations of Steelhead Trout (anadromous Oncorhynchus mykiss) from south coastal areas to northwestern British Columbia with perhaps 1,200 or more watersheds potentially supporting Steelhead Trout populations (Fig. A1). The Thompson and Chilcotin rivers’ Steelhead Trout (TCS) constitute two designatable units (DUs) within this assemblage as they satisfy both the discrete and significance criteria for recognizing DUs (COSEWIC 2016).
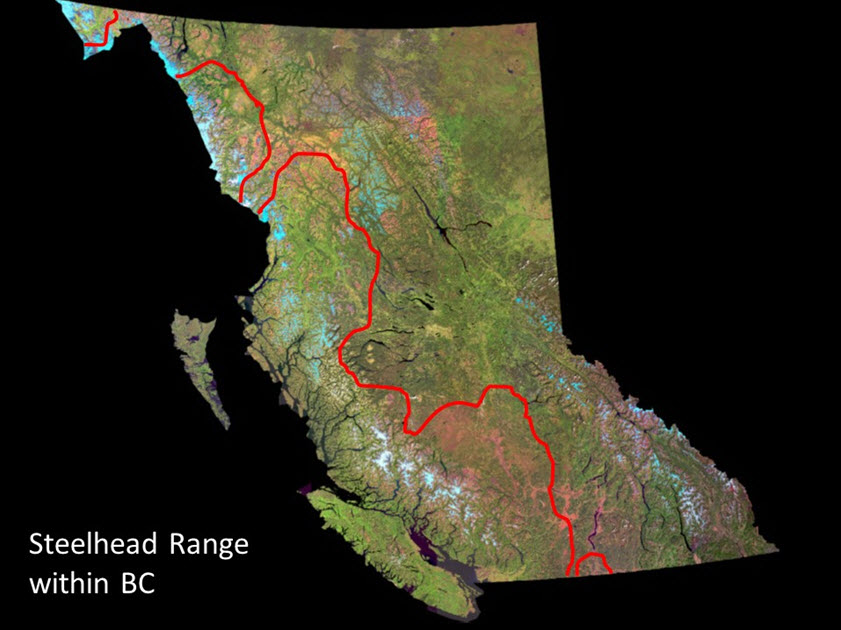
Long description for A1
Long descriptions available in the accompanying text.
The TCS spawn within tributaries of the Thompson and Chilcotin rivers of the Fraser River drainage and thus are spatially discrete from other Steelhead Trout populations in BC. Given the well-documented homing to natal streams for spawning of most anadromous salmonids like Steelhead Trout, there is a high degree of spatial genetic population structure in interior Fraser River Steelhead Trout. For instance, Beacham et al. (2004) used 14 microsatellite DNA loci and demonstrated that TCS, and a group of Steelhead Trout from the mid-Fraser River (MFS; Stein, Nahatlatch, and Bridge rivers), formed a well-defined cluster of populations (75% bootstrap support, Fig. 2 in Beacham et al. 2004) distinct from 46 other populations from northwestern BC to US portions of the upper Columbia River. In fact, the TCS (and MFS) were more similar genetically to Steelhead Trout from the upper Columbia River than they were to Steelhead Trout from the lower Fraser River (e.g., Chilliwack and Coquihalla rivers, Fig. 2 of Beacham et al. 2004). Furthermore, the TCS are part of the admixed south coast/interior phylogenetic group as inferred from mtDNA that is unique in BC (Fig. 9, 10 in McCusker et al. 2000; Fig. A2, A3). In addition, the Thompson River component of the TCS are discrete from the MFS as well as from the Chilcotin River Steelhead Trout (CRS) when assayed using these same microsatellite loci (98% bootstrap support). Allele frequency tests based on the microsatellite DNA data of Beacham et al. (2004) and four polymorphic allozyme loci studied by Parkinson (1984, Table 1), however, both indicate that the Chilcotin River Steelhead Trout are also significantly distinct from all three MFS samples (all P < 0.001, E. Taylor, University of British Columbia, Vancouver, unpublished results). In the case of the allozyme loci, most of the differentiation is attributable to differences between the Chilcotin River and the Stein/ Nahatlatch River samples, but one locus (AGP) also distinguished Chilcotin River fish from Bridge River fish (randomization test P = 0.0003). These two samples were also significantly distinct when combining probabilities across all four loci (Fisher’s combined probability test, P < 0.001, E. Taylor, University of British Columbia, Vancouver, unpublished analysis). The Bridge River fish are also the most similar of the MFS samples to CRS in terms of microsatellite loci. The proportion of total variation in microsatellite allele frequencies attributable to differences between Bridge River fish and CRS (FST) is 2.5% (P < 0.002) whereas it is between 5.6% and 8.8% between Nahatlatch and Stein rivers and CRS, respectively (both P < 0.002, T.D. Beacham, DFO, Nanaimo, BC, pers. comm. Jan. 12, 2017 based on data in Beacham et al. 2004). Although the degree of differentiation is variable, the TCS are, demonstrably discrete from all other BC Steelhead Trout, and the Thompson and Chilcotin rivers’ populations are discrete from each other (see below).
The genetic data cited above also point to the significance of the TCS as a discrete assemblage of Steelhead Trout. The microsatellite and mtDNA data both suggest that the TCS have had a unique glacial and postglacial history in BC in that they share a close affinity with Steelhead Trout from the south coast (mtDNA) as well as from the upper Columbia River (microsatellites). This suggests that the TCS may result from a double invasion of the current waterscape from two glacial refugia - again, a situation that appears to be unique within the evolutionary legacy of BC Steelhead Trout (Fig. 9 in McCusker et al. 2000; Fig. A2, A3).
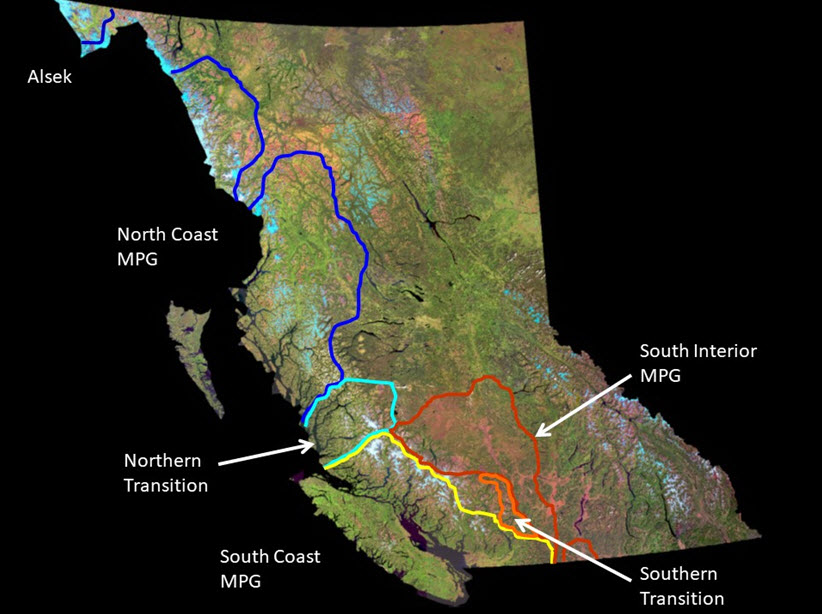
Long description for A2
Long descriptions available in the accompanying text.
Further evidence for the evolutionary significance of the discreteness of TCS from other Steelhead Trout comes from the studies of allozyme differentiation and its apparent association with swimming stamina. The TCS, represented by samples from the Thompson River, have higher frequencies of lactate dehydrogenase phenotypes that are associated with substantially greater prolonged swimming performance compared to fish from the lower Fraser River (Tsuyuki and Williscroft 1977). Such physiological differences are also apparent between coastal and interior populations of Coho Salmon (Taylor and McPhail 1985) and point to the actual and potential adaptive characteristics of salmonid fishes with long upstream migrations in the Fraser River. Other differences between TCS and south coast Steelhead Trout include their fall-season run timing and the immature state of gonads during migration - a phenomenon known as “premature migration”. By contrast, south coast Steelhead Trout typically migrate through the lower Fraser River after TCS and with gonads in more advanced states of maturity. The premature migration phenotype appears to have a relatively simple genetic basis, to be under strong positive selection, and is considered critical for the persistence of Steelhead Trout biodiversity in other portions of its range (Prince et al. 2017). The TCS also differ in several aspects of migration timing, speed, age at maturation, and smolt age from fish in the lower and middle Fraser River. These differences are especially evident between the Chilcotin River fish and all others and likely reflect adaptations to the longer and more arduous migrations of these fish and the distinct climates that they live in (see below).
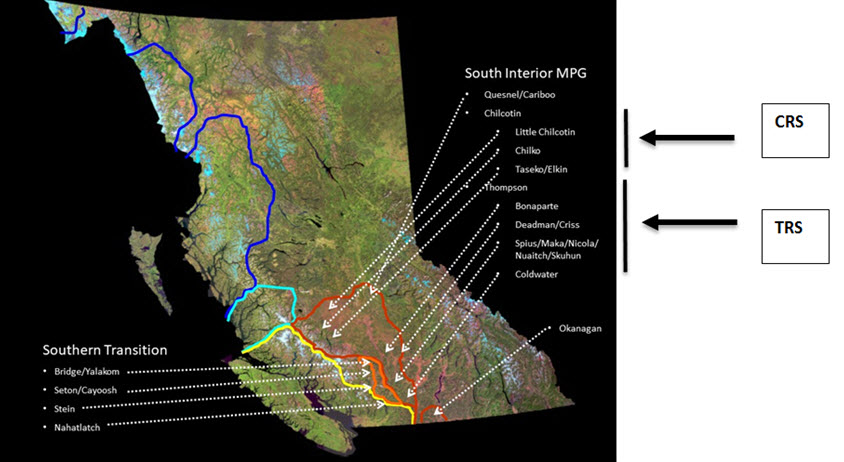
Long description for A3
Long descriptions available in the accompanying text.
Given all the above, TCS satisfy both the discrete and significance criteria for their recognition as at least one distinct DU within O. mykiss. Further, it is proposed that TCS be subdivided into two DUs distinct from one another: Thompson River Steelhead Trout (TRS) and Chilcotin River Steelhead Trout (CRS).
As discussed above the genetic data of Beacham et al. (2004) clearly (i.e., with 98% bootstrap support) identified TRS as a genetic cluster distinct from other Steelhead Trout including the CRS. Genetic distance (FST) between TRS and CRS at microsatellite loci accounted for between 6.2% and 8.3% of the total variation when assaying those two samples (all P < 0.001, pers. comm. from T.D. Beacham, DFO, Nanaimo, BC Jan. 12, 2017 based on data in Beacham et al. 2004). Parkinson’s (1984) data also showed that CRS had multilocus genotypes across four allozyme loci (SOD, LDH, MDH, and AGP) that were distinct from samples of Thompson, and MFS rivers’ Steelhead Trout (see above). The TRS and CRS are also spatially discrete (see below) and phenotypically discrete from each other, most notably in terms of adult age at maturation, migration timing and behaviour, and smolt age (discussed below under significance).
The TRS and CRS differ in several aspects of migration timing, speed, and behaviour that can be plausibly interpreted as adaptations to the different locations of their spawning areas. In general, genetic mixture and telemetry studies indicate that the CRS enter the Fraser River earlier, migrate upriver faster, and exhibit less “milling” behaviour than TRS (i.e., “milling” is a behaviour where fish remain relatively stationary in certain areas en route to the spawning or overwintering areas). For instance, Bison (unpublished data) reported a mean difference in the date of migration past river km 235 (near the Nahatlatch River) of 13.8 days (i.e., these fish arrived on average almost 14 days earlier than the date averaged across all populations) for CRS compared to 0.2 to -4.3 days for the TRS (and -1.6 to -8.3 days for the later-arriving MFS fish, respectively, N = 49 fish radio-tagged from all areas). These differences likely result from selection for earlier and more direct migration in CRS because they have to surpass three major migration hurdles prior to the onset of winter (two in the lower Fraser River canyon at river kms 185 and 210, and one at Bridge River rapids at river km 340). By contrast, TRS need only surpass two hurdles in the lower Fraser River canyon. Further, CRS have further to travel to their overwintering sites which are at least as far upstream as river km 522 in the Chilcotin River and river km 510 in the Fraser River (~100 km upstream of the Chilcotin-Fraser confluence; Renn et al. 2001). By contrast, TRS overwinter only as far upstream as the outlet of Kamloops Lake at river km 375 from where the Fraser River enters the Strait of Georgia (note: the latest-arriving Nahatlatch River fish travel only 238 km from the mouth of the Fraser River). Finally, TRS and CRS differ from each other both in smolt age and adult age of return to fresh water; the majority of TRS smolts are age two years when they migrate to sea (93%), while the majority of CRS smolts are age three years (83%, Bison 2012). The older age at smolting of CRS drives their older adult age at return. The age at first spawning is typically five years (more rarely six or seven) for TRS, but age six years (rarely seven or eight) for CRS (N = 14 - 215 fish annually over 40 years of monitoring, Bison 2012). The older age of smolting and adult maturation in CRS are likely adaptations to the longer and/or more arduous migrations experienced by these fish (see above). Finally, the TRS and CRS are found in different biogeoclimatic zones of BC; the TRS are found primarily in the Interior Douglas Fir Zone whereas the CRS are found across a mix of several smaller zones. The former is considered more of a semi-desert region with generally higher mean annual air temperature (see Fig. A4). The higher temperature may result in greater growth opportunity for TRS smolts and, in part, explain their younger average age at smolting relative to CRS. Several studies have provided evidence of divergence in thermal tolerance physiology in O. mykiss from non-BC populations along a similar desert-montane environmental gradient (Rodnick et al. 2004; Narum et al. 2010, 2013) and it is plausible that similar differences exist between TRS and CRS.
The TRS and CRS should be assessed as two distinct DUs.
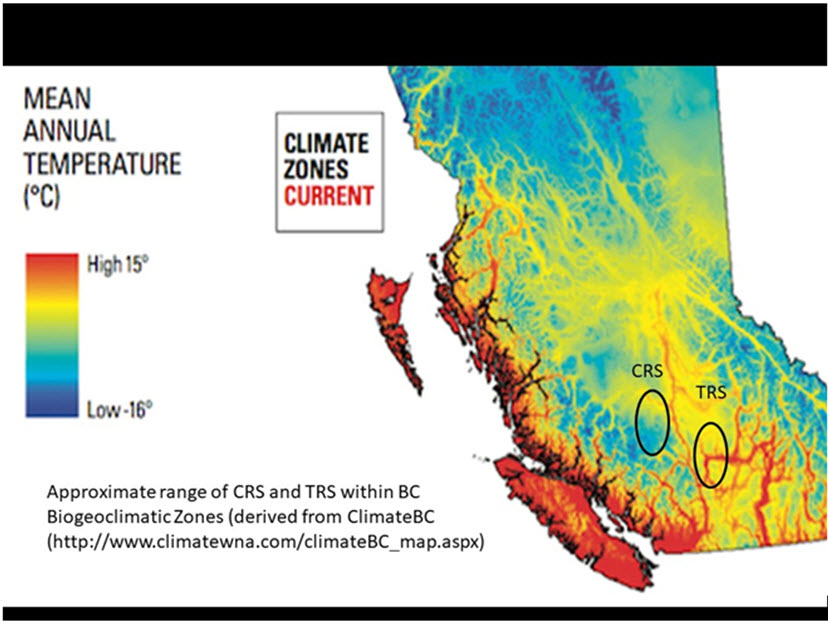
Long description for Figure A4
Long descriptions available in the accompanying text.
Abdul-Aziz O., Mantua N., and Myers K.W. 2011. Potential climate change impacts on thermal habitats of Pacific salmon (Oncorhynchus spp.) in the North Pacific Ocean and adjacent seas. Canadian Journal of Fisheries and Aquatic Sciences 68:1660-1680.
Berejikian, B.A., Moore, M.E. and Jeffries, S.J. 2016. Predator-prey interactions between harbor seals and migrating steelhead trout smolts revealed. Marine Ecology Progress Series 543: 21-35.
Bison, R. 2012. Population attributes for Fraser River late-run summer steelhead. Unpublished MS. BC Ministry of Natural Resource Operations, Fish & Wildlife Branch, Kamloops, BC. 55 p.
Bison, R. 2016. Fishing Mortality Trends for Thompson River Steelhead from 1991 to 2015. BC Ministry of Natural Resource Operations, Fish & Wildlife Branch, Kamloops, BC. 30 p.
Bison, R. and Phelps, M. 2017. Statistics for the 2016 Thompson River Steelhead Sport Fishery and Relationships between the Fishery and Steelhead Abundance. BC Ministry of Natural Resource Operations, Fish & Wildlife Branch, Kamloops, BC. 37 p.
Beacham, T.D., Le, K.D. and Candy, J.R., 2004. Population structure and stock identification of Steelhead Trout (Oncorhynchus mykiss) in British Columbia and the Columbia River based on microsatellite variation. Environmental Biology of Fishes 69:95-109.
ClimateBC. 2017. An interactive platform for visualization and data access. Centre for Forest Conservation Genetics, University of British Columbia, Vancouver. ClimateBC_Map. Accessed Dec 23, 2017.
COSEWIC. 2010. COSEWIC assessment and status report on the Atlantic Salmon Salmo salar (Nunavik population, Labrador population, Northeast Newfoundland population, South Newfoundland population, Southwest Newfoundland population, Northwest Newfoundland population, Quebec Eastern North Shore population, Quebec Western North Shore population, Anticosti Island population, Inner St. Lawrence population, Lake Ontario population, Gaspé-Southern Gulf of St. Lawrence population, Eastern Cape Breton population, Nova Scotia Southern Upland population, Inner Bay of Fundy population, Outer Bay of Fundy population) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xlvii + 136 pp.
COSEWIC. 2016. Operations and procedures manual. Appendix F5. Committee on the Status of Endangered Wildlife in Canada. Ottawa. In press.
COSEWIC. 2018. COSEWIC assessment and status report on the Sockeye Salmon (Oncorhynchus nerka) in the Fraser River drainage. Committee on the Status of Endangered Wildlife in Canada. Ottawa. In press.
Docker, M.F. and Heath, D.D. 2003. Genetic comparison between sympatric anadromous steelhead and freshwater resident rainbow trout in British Columbia, Canada. Conservation Genetics 4:227-231.
Hard, J.J., J.M. Myers, E.J. Connor, R.A. Hayman, R.G. Kope, G. Lucchetti, A.R. Marshall, G.R. Pess, and B.E. Thompson. 2015. Viability criteria for steelhead within the Puget Sound distinct population segment. U.S. Dept. Commerce, NOAA Tech. Memo. NMFS-NWFSC-129, 367 p.
Hinch, S.G. and E.G. Martins. 2011. A review of potential climate change effects on survival of Fraser River sockeye salmon and an analysis of interannual trends in en route loss and pre-spawn mortality. Cohen Commission Tech. Rept. 9: 134p. Vancouver, B.C.
Kendall, N.W., Marston, G.W. and Klungle, M.M. 2017. Declining patterns of Pacific Northwest steelhead trout (Oncorhynchus mykiss) adult abundance and smolt survival in the ocean. Canadian Journal of Fisheries and Aquatic Sciences 74:1275-1290.
McCusker, M.R., Parkinson, E.A., and Taylor, E.B. 2000. Phylogenetic Conservation Units for Rainbow Trout in British Columbia. Province of British Columbia, Fisheries Management Report 112. University of British Columbia, Vancouver, B.C. 35 p.
McMillan, J.R., Katz, S.L. and Pess, G.R., 2007. Observational evidence of spatial and temporal structure in a sympatric anadromous (winter steelhead) and resident rainbow trout mating system on the Olympic Peninsula, Washington. Transactions of the American Fisheries Society 136: 736-748.
McPhail, J.D. 2007. Freshwater fishes of British Columbia. University of Alberta Press, Edmonton.
Narum, S.R., Campbell, N.R., Kozfkay, C.C. and Meyer, K.A., 2010. Adaptation of redband trout in desert and montane environments. Molecular Ecology 19(21):4622-4637.
Narum, S.R., Campbell, N.R., Meyer, K.A., Miller, M.R., Hardy, R.W. 2013. Thermal adaptation and acclimation of ectotherms from differing aquatic climates. Molecular Ecology 22(11):3090-3097.
Parkinson, E.A. 1984. Genetic variation in populations of Steelhead Trout (Salmo gairdneri) in British Columbia. Canadian Journal of Fisheries and Aquatic Sciences 41:1412-1420.
Pearse, D.E., Hayes, S.A., Bond, M.H., Hanson, C.V., Anderson, E.C., Macfarlane, R.B. and Garza, J.C., 2009. Over the falls? Rapid evolution of ecotypic differentiation in steelhead/rainbow trout (Oncorhynchus mykiss). J. Heredity 100: 515-525.
Prince, D.J., O’Rourke, S.M., Thompson, T.Q., Ali, O.A., Lyman, H.S., Saglam, I.K., Hotaling, T.J., Spidle, A.P. and Miller, M.R., 2017. The evolutionary basis of premature migration in Pacific salmon highlights the utility of genomics for informing conservation. Science Advances 3(8), p.e1603198.
Renn, J.R., Bison R. Hagen, J. and Nelson, T.C. 2001. Migration characteristics and stock composition of interior Fraser steelhead as determined by radio telemetry, 1996-1999. BC Ministry of Water, Land and Air Protection, Fisheries Branch. 150p.
Rodnick, K.J., Gamperl, A.K., Lizars, K.R., Bennett, M.T., Rausch, R.N. and Keeley, E.R., 2004. Thermal tolerance and metabolic physiology among redband trout populations in south-eastern Oregon. J. Fish Biology 64(2):310-335.
Taylor, E.B. and McPhail, J.D., 1985. Variation in burst and prolonged swimming performance among British Columbia populations of coho salmon, Oncorhynchus kisutch. Canadian Journal of Fisheries and Aquatic Sciences 42(12):2029-2033.
Thrower F. P., Hard J. J., Joyce J. E. 2004-Genetic architecture of growth and early life-history transitions in anadromous and derived freshwater populations of steelhead. J. Fish Biology 65: 286-307.
Tsuyuki, H. and Williscroft, S.N., 1977. Swimming stamina differences between genotypically distinct forms of rainbow (Salmo gairdneri) and Steelhead Trout. Journal of the Fisheries Board of Canada 34:996-100.
Viola, A.E. and Schuck, M.L., 1995. A method to reduce the abundance of residual hatchery steelhead in rivers. North American Journal of Fisheries Management 15:488-493.
Zimmerman, C.E. and Reeves. G.H. 2000. Population structure of sympatric anadromous and nonanadromous Oncorhynchus mykiss: evidence from spawning surveys and otolith microchemistry. Canadian Journal of Fisheries and Aquatic Sciences 57:2152-2162.