
Truite arc-ciel anadrome (Oncorhynchus mykiss), population de la rivière Thompson et population de la rivière Chilcotin au Canada, 2018 : COSEPAC Résumés techniques et donnés d’appui pour les évaluations d’urgence
- Table des matières
- COSEPAC Sommaire de l’évaluation - Population de la rivière Thompson
- COSEPAC Sommaire de l’évaluation - Population de la rivière Chilcotin
- COSEPAC Résumé
- Introduction
- Structure taxonomique et unités désignables
- Renseignements accessibles pour l’évaluation
- Menaces
- Résultats de l’évaluation
- Immigration de source externe
- Remerciements
- Résumé technique 1- Truite arc-en-ciel anadrome (Population de la rivière Thompson)
- Résumé technique 2- Truite arc en ciel anadrome (Population de la rivière Chilcotin)
- Sources d’information
- Figure 1. Tendance du nombre d’individus matures de l’UD de la truite arc en ciel anadrome de la rivière Thompson, de 1978 à 2018, et régression log-linéaire ajustée au cours des 3 dernières générations (durée d’une génération : 5 ans).
- Figure 2.Tendance du nombre d’individus matures de l’UD de la truite arc en ciel anadrome de la rivière Chilcotin, de 1972 à 2018, et régression log-linéaire ajustée au cours des 3 dernières générations (durée d’une génération : 6 ans).
- Figure A1. Aire de répartition approximative de la truite arc en ciel anadrome en Colombie Britannique (à gauche de la ligne rouge, y compris toutes les îles côtières).
- Figure A2. Répartition des grands groupes phylogénétiques (GGP) de l’Oncorhynchus mykiss en Colombie-Britannique (d’après McCusker et al., 2000).
- Figure A3. Lieux où l’on rencontre la truite arc en ciel anadrome de la rivière Chilcotin (TAECA-RC) et la truite arc en ciel anadrome de la rivière Thompson (TAECA-RT; flèches noires) en Colombie Britannique et grands groupes phylogénétiques (voir la figure A2).
- Figure A4. Emplacement approximatif des populations de truites arc en ciel anadromes de la rivière Chilcotin (TAECA-RC) et de truites arc en ciel anadromes de la rivière Thompson (TAECA-RT) dans le contexte des zones climatiques de la Colombie Britannique, compte tenu de la température annuelle moyenne de l’air.
- Annexe 1
- Annexe 2
- Truites arc-en-ciel anadromes de la rivière Thompson et de la rivière Chilcotin - Structure des unités désignables (UD)
- (a) Distinction des truites arc-en-ciel anadromes des rivières Thompson et Chilcotin (TAECA-RTC) d’autres truites arc-en-ciel anadromes de la Colombie-Britannique
- (a) Distinction entre la TAECA-RT et la TAECA-RC
Truite arc-en-ciel anadrome Oncorhynchus mykiss, population de la rivière Thompson et population de la rivière Chilcotin au Canada, 2018 : COSEPAC Résumés techniques et donnés d’appui pour les évaluations d’urgence

COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
En Colombie-Britannique, deux lignées évolutives d’Oncorhynchus mykiss sont présentes, communément appelées O. mykiss « côtière » et « de l’intérieur ». Ces deux lignées d’O. mykiss se trouvent dans des populations ou des types de cycles vitaux dulcicoles et anadromes, connues respectivement comme étant la truite arc-en-ciel et la truite arc-en-ciel anadrome. L’O. mykiss de l’intérieur se trouve dans les rivières Thompson et Chilcotin (qui font partie du bassin versant du fleuve Fraser). Il existe un certain croisement entre les individus résidents d’eau douce et les individus anadromes, et les individus résidents d’eau douce pourraient produire une descendance anadrome et vice versa.
L’anadromie de la truite arc-en-ciel anadrome et son âge avancé (et sa plus grande taille) à maturité sont des aspects importants de son cycle vital qui la distinguent de la truite arc-en-ciel résidente d’eau douce. Il est probable que les truites arc-en-ciel anadromes des rivières Thompson et Chilcotin aient évolué à partir d’individus isolés dans le refuge du Columbia pendant la dernière glaciation, tandis que les autres truites arc-en-ciel anadromes du Canada pourraient provenir du refuge de Haida Gwaii. Selon des données génétiques, la truite arc-en-ciel anadrome de la rivière Thompson et celle de la rivière Chilcotin se distinguent des autres truites arc-en-ciel anadromes du Canada, et diffèrent l’une de l’autre. C’est pourquoi la truite arc-en-ciel anadrome de l’intérieur du fleuve Fraser satisfait au critère du COSEPAC permettant son évaluation en tant que deux unités désignables (UD) ou populations distinctes : la population de la rivière Thompson et la population de la rivière Chilcotin.
La truite arc-en-ciel anadrome des rivières Thompson et Chilcotin a récemment fait l’objet de préoccupations du public, notamment par la présentation d’une demande visant une évaluation d’urgence en janvier 2016. Ces commentaires du public sont venus accroître les préoccupations du COSEPAC concernant la truite arc-en-ciel anadrome des rivières Thompson et Chilcotin, et un rapport de situation devait être amorcé en 2018. Toutefois, les données les plus récentes sur les remontes des individus adultes indiquent que la situation s’aggrave et constitue une urgence. C’est ce qui a mené à la présentation d’une autre demande visant une évaluation d’urgence en novembre 2017.
Le nombre d’individus matures qui ont effectué une remonte vers l’eau douce à partir de la mer à l’automne 2017 et qui frayeront au printemps 2018 est de 177 pour la rivière Thompson et de 58 pour la rivière Chilcotin. Le nombre annuel moyen d’individus matures effectuant une remonte dans les rivières Thompson et Chilcotin au cours des trois dernières années (2016-2018) est le plus faible d’une série chronologique qui a commencé en 1978. Le déclin des individus matures dans la rivière Thompson au cours des trois dernières générations (15 ans) est de 79 %, et le déclin de la truite arc-en-ciel anadrome de la rivière Chilcotin au cours de trois générations (18 ans) est de 81 %. La mortalité attribuable aux prises accessoires de la pêche commerciale au saumon du Pacifique et le déclin de la qualité de l’habitat marin et dulcicole constituent les principaux facteurs de déclin des populations.
L’évaluation d’urgence a été effectuée le 10 janvier 2018. Les participants à l’évaluation d’urgence ont examiné ces données et conclu que le statut de la truite arc-en-ciel anadrome de l’UD de la rivière Thompson et de l’UD de la rivière Chilcotin est en voie de disparition et constitue un cas urgent. Le présent rapport regroupe les documents d’information utilisés pendant l’évaluation d’urgence et les conclusions formulées.
En réaction aux plus faibles remontes de la truite arc-en-ciel anadrome des rivières Thompson et Chilcotin observées à ce jour et aux préoccupations du public concernant le statut de conservation de l’espèce sauvage, le COSEPAC a effectué une évaluation d’urgence de l’espèce le 10 janvier 2018. Trente-et-une personnes ont participé à l’évaluation d’urgence, et leurs noms et associations sont énumérés à l’annexe 1. Le présent rapport fournit un résumé des renseignements examinés par des participants ainsi que les conclusions de l’évaluation d’urgence. En attendant la décision concernant la recommandation du COSEPAC voulant qu’un décret d’urgence soit pris (voir ci-dessous), un rapport de situation complet du COSEPAC sera produit en accéléré, et dans l’année suivant la prise d’un décret d’urgence en vertu de la Loi sur les espèces en péril (article 30.1).
Le taxon Oncorhynchus mykiss présente deux grands types de cycles vitaux : une forme résidente des lacs et des cours d’eau connue sous le nom de « truite arc en ciel » et une forme anadrome (allant à la mer), soit la « truite arc en ciel anadrome » (McPhail, 2007). Tout dépendant du contexte géographique (p. ex. la distance de la mer, la présence d’obstacles à la migration, la présence de lacs à l’intérieur d’un bassin versant), une forme, ou les deux, pourraient se trouver dans un seul bassin versant et même dans le même affluent au sein d’un bassin versant. Encore une fois, tout dépendant du contexte géographique, les formes pourraient exister séparément, coexister au même endroit, au même moment, en tant que juvéniles et adultes reproducteurs, ou leurs aires de répartition pourraient être adjacentes les unes des autres (McPhail, 2007). Comme on peut s’y attendre, il y a également un degré variable d’interaction démographique et génétique entre les formes aux endroits où elles coexistent. Dans certains cas, il y a peu de différence génétique décelable entre les formes et, dans d’autres cas, les formes pourraient représenter des populations distinctes sur le plan génétique (Docker et Heath, 2003; McMillan et al., 2007; Pearse et al., 2009). De plus, des éléments prouvent que, dans certains réseaux, des truites arc en ciel anadromes pourraient être produites par des truites arc en ciel résidentes d’eau douce, et que des descendants de truites arc-en ciel anadromes pourraient demeurer en permanence en eaux douces (nommés « résidus »), particulièrement lorsque les juvéniles connaissent une croissance plus rapide, notamment comme c’est souvent le cas dans les populations enrichies de poissons d’écloseries (Viola et Schuck, 1995; Zimmerman et Reeves, 2000; Thrower et al., 2004). Ces liens variables entre la truite arc-en-ciel d’eau douce et la truite arc-en-ciel anadrome se trouvent également chez d’autres espèces de salmonidés, comme l’O. nerka, aux endroits où il y a des formes résidentes d’eau douce (Kokanee) et des formes anadromes (saumon sockeye), et le Salmo salar (avec le ouananiche dulcicole et les formes anadromes du saumon de l’Atlantique). Dans le cas de la truite arc-en-ciel anadrome des rivières Thompson et Chilcotin, il n’y a aucun renseignement sur le lien génétique entre les deux types de cycles vitaux. Il semble que les truites-arc-en-ciel anadromes dans ces réseaux pourraient être produites par des truites arc-en-ciel résidentes d’eau douce (R. Bison, Ministry of Forests, Lands, Natural Resource Operations and Rural Development de la Colombie-Britannique, Kamloops [Colombie-Britannique], comm. pers.), mais l’étendue spatiale et temporelle de ce phénomène n’est pas bien comprise. Par conséquent, et conformément aux récentes évaluations de la situation du saumon de l’Atlantique (COSEWIC, 2010) et du saumon sockeye (COSEWIC, 2018), cette évaluation de l’O. mykiss du Fraser intérieur concerne seulement la truite arc-en-ciel anadrome. L’approche du COSEPAC est également conforme à celle des organismes de gestion des pêches des États Unis; ici, les formes anadrome et résidente d’eau douce d’O. mykiss sont évaluées séparément (Hard et al., 2015).
Selon des données génétiques, la truite arc-en-ciel anadrome des rivières Thompson et Chilcotin se distingue des autres truites arc-en-ciel anadromes du Canada, et les populations des deux rivières se distinguent l’une de l’autre. La truite arc-en-ciel anadrome des rivières Thompson et Chilcotin a probablement évolué à partir de poissons isolés dans le refuge du Columbia pendant la dernière glaciation, tandis que les autres truites arc-en-ciel anadromes du Canada pourraient provenir du refuge de Haida Gwaii. Après avoir examiné les renseignements accessibles sur les unités désignables (UD) de la truite arc-en-ciel anadrome des rivières Thompson et Chilcotin et appliqué les critères du COSEPAC concernant le caractère distinct et l’importance, les participants à l’évaluation d’urgence ont convenu que la truite arc-en-ciel anadrome des rivières Thompson et Chilcotin devait être évaluée en tant que deux UD distinctes de toutes les autres populations de truites arc-en-ciel anadromes de la Colombie-Britannique : l’UD de la rivière Thompson et l’UD de la rivière Chilcotin (voir l’annexe 2 pour de plus amples détails).
Des renseignements sur la surveillance annuelle sont accessibles grâce à une pêche expérimentale effectuée dans le fleuve Fraser, environ 60 km en amont de l’océan (près d’Albion, en Colombie-Britannique). Les engins de pêche utilisés sont des filets maillants conventionnels de pêche au saumon. Un filet à grandes mailles (8 pouces) et un filet à mailles légèrement plus petites (6,75 pouces) sont utilisés en alternance et à des hauteurs de marée constantes pendant le début et la période de pointe de la migration de la truite arc-en-ciel anadrome du Fraser intérieur à cet endroit. Pendant la dernière étape de la migration, seul le filet à mailles plus petites est utilisé chaque jour. Les statistiques sur les prises utilisées dans la prévision de l’abondance des individus reproducteurs sont simplement le nombre de truites arc-en-ciel anadromes capturées par jour.
Pour certains affluents des rivières Thompson et Chilcotin où la truite arc-en-ciel anadrome fraye, des dénombrements et des estimations sont également effectués dans les cours d’eau. En ce qui concerne les affluents de la rivière Thompson, des compteurs automatisés de poissons sont utilisés dans les rivières Deadman et Bonaparte, et des dénombrements visuels périodiques à partir d’embarcations sont effectués dans un affluent majeur du bassin versant de la rivière Nicola. Ces dénombrements visuels sont combinés dans un modèle d’estimation de vraisemblance maximale avec l’efficacité des observateurs, le moment des observations et les estimations de la répartition spatiale fondées sur le marquage externe et le radiopistage pour estimer l’abondance de truites arc-en-ciel anadromes dans le bassin versant de la rivière Nicola, qui comprend des estimations pour la rivière Coldwater, le ruisseau Spius et le cours inférieur de la rivière Nicola (Bison et Phelps, 2017). Pour ce qui est de la rivière Chilko (un affluent de la rivière Chilcotin), des dénombrements visuels périodiques (à partir d’un hélicoptère) sont effectués sur une distance de 24 km (c.-à-d. à partir de la confluence du ruisseau Brittany avec la rivière Chilko jusqu’au lac Chilko; Bison et Phelps, 2017).
Les prises (et la remise à l’eau) de la pêche sportive sont estimées par des sondages aléatoires et stratifiés menés auprès des pêcheurs sur le terrain, où environ le tiers de l’effort total fait l’objet de sondages (Bison et Phelps, 2017). Une estimation secondaire est fondée sur un questionnaire de sondage annuel mené auprès des pêcheurs de l’ensemble de la province après la saison de la pêche par la Fish & Wildlife Branch (Bison et Phelps, 2017).
Les prises accessoires de la pêche commerciale au saumon du Pacifique sont estimées indirectement, d’après les tendances et le niveau des taux de rencontre, tandis que le taux de mortalité des poissons est estimé au moyen d’un modèle de simulation (Bison, 2016).
Les tendances de l’abondance annuelle des individus reproducteurs dans les affluents majeurs des UD des rivières Thompson et Chilcotin montrent toutes deux des déclins dramatiques depuis le début des années 2000 (figures 1 et 2). Les taux de déclin pendant trois générations sont de 79 % pour l’UD de la rivière Thompson et de 81 % pour l’UD de la rivière Chilcotin.
Pour ce qui est de l’UD de la rivière Thompson, si le taux de déclin actuel persiste pendant trois autres générations, le nombre d’individus reproducteurs chutera pour atteindre 37 individus, ce qui représente 2 % de l’abondance précédant 2000. Pour ce qui est de l’UD de la rivière Chilcotin, si le taux de déclin actuel persiste pendant trois autres générations, le nombre d’individus reproducteurs chutera pour atteindre 11 individus, ce qui représente environ 0,9 % de l’abondance précédant 2000.
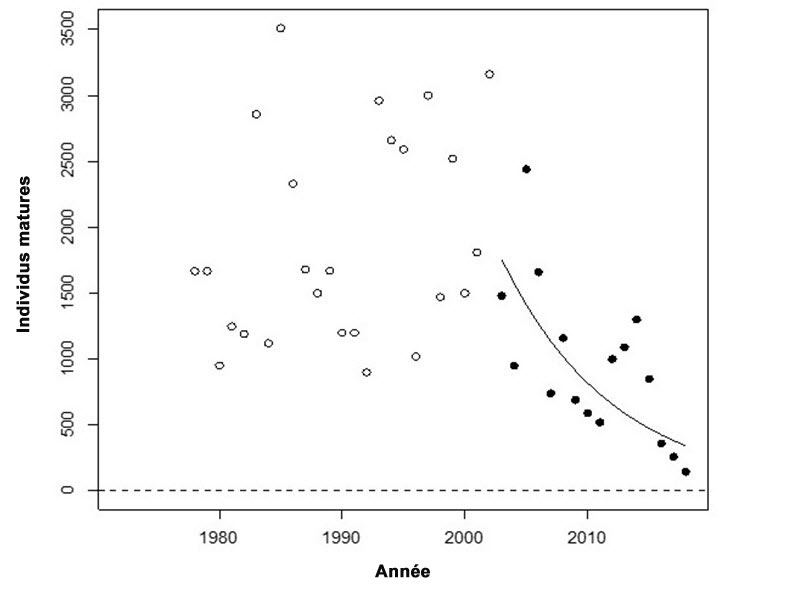
Description longue de la figure 1
Diagramme illustrant la tendance du nombre d’individus matures dans l’unité désignable de la truite arc-en-ciel anadrome de la rivière Thompson, de 1978 à 2018, et la régression log-linéaire ajustée au cours des trois dernières générations. Les points de données pleins montrés après 2000 ont été utilisés dans l’estimation du déclin de 79 %.
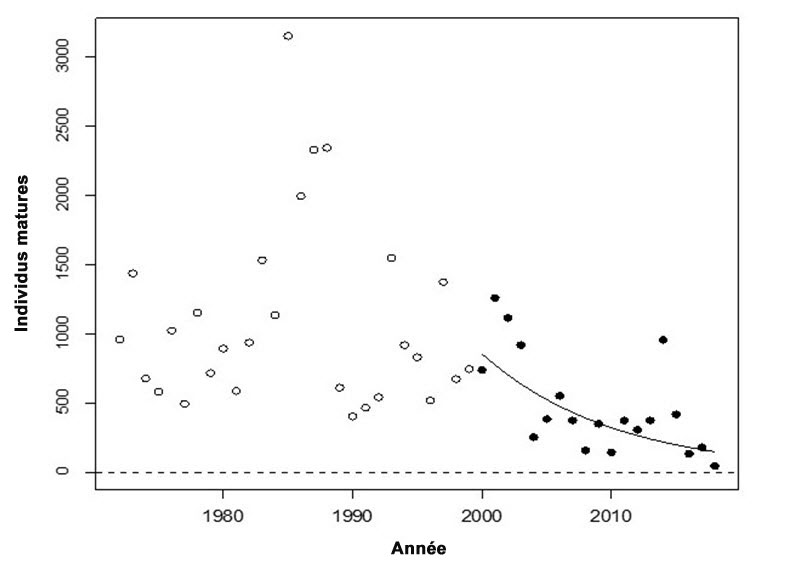
Description longue de la figure 2
Diagramme illustrant la tendance du nombre d’individus matures dans l’unité désignable de la truite arc-en-ciel anadrome de la rivière Chilcotin, de 1972 à 2018, et la régression log-linéaire ajustée au cours des trois dernières générations (durée d’une génération : six ans). Les points de données pleins montrés après 2000 ont été utilisés dans l’estimation du déclin de 81 %.
La diminution de la survie de la truite arc-en-ciel anadrome en milieu marin est considérée comme étant un facteur clé des déclins des populations depuis le début des années 1990 (Kendall et al., 2017). Des déclins semblables causés par de faibles taux de survie en océan ont été signalés dans les récents rapports du COSEPAC sur le saumon sockeye et le saumon coho (O. kisutch). Bien que des liens entre la température de l’eau et la survie aient été cernés, les mécanismes causaux sous-jacents expliquant ces liens ne sont pas bien compris pour la truite arc-en-ciel anadrome. Les températures de l’océan ont augmenté en moyenne de 0,5 °C au cours des deux dernières décennies et ont probablement contribué au déclin du taux de survie de la truite arc-en-ciel anadrome, comme on l’a laissé entendre pour le saumon sockeye (Hinch et Martins, 2011). Il est prévu que la température de l’eau du golfe de l’Alaska, où la truite arc-en-ciel anadrome passe la majorité de sa vie en milieu marin, augmente de 1 à 2 °C d’ici les années 2040 (Abdul-Aziz et al., 2011). Berejikian et al. (2016) ont avancé que la prédation par les phoques communs (Phoca vitulina) a contribué à la mortalité de truites arc-en-ciel anadromes migrant à partir de l’État de Washington, et ils ont formulé l’hypothèse selon laquelle les changements documentés dans l’écosystème de Puget Sound pourraient actuellement entraîner un plus grand risque de prédation par les phoques communs et possiblement d’autres prédateurs pour la truite arc-en-ciel anadrome.
La prise accessoire d’individus matures en remonte dans le cadre de la pêche à senne coulissante et à filets maillants visant le saumon du Pacifique constitue une menace mieux quantifiée par rapport à la survie en milieu marin. Aucune pêche commerciale ne vise la truite arc-en-ciel anadrome en Colombie-Britannique, et la pêche sportive se limite à la capture et remise à l’eau; cette activité prend fin si les estimations de l’abondance en saison sont inférieures aux limites prédéterminées. L’estimation du taux de mortalité attribuable aux prises accessoires de la pêche commerciale se situe entre 15 et 25 % par année (Bison, 2016). Cette menace à elle seule pourrait expliquer en grande partie le déclin observé chez les individus matures.
Bien qu’on considère généralement que la qualité de l’habitat dulcicole connaît un déclin, la gravité des menaces qui pèsent sur l’habitat dulcicole des rivières Thompson et Chilcotin n’est pas bien comprise.
L’évaluation d’urgence a permis de conclure que, pour l’UD de la rivière Thompson, une désignation d’espèce en voie de disparition s’applique (en voie de disparition, A2bd+4bd; C2a(i); D1 - voir le résumé technique 1). Pour l’UD de la rivière Chilcotin, l’évaluation d’urgence a également permis de conclure qu’une désignation d’espèce en voie de disparition s’applique (en voie de disparition, A2bd+4bd; C2a(i,ii); D1 - voir le résumé technique 2).
Dans l’évaluation d’urgence, on a également utilisé le logiciel RAMAS Red List V3.0 (RAMAS Software by Applied Biomathematics) pour une évaluation rapide au moyen des critères de l’Union internationale pour la conservation de la nature (UICN). Pour les deux UD, la procédure RAMAS utilisant les plus récentes estimations de l’abondance des adultes reproducteurs de la province de la Colombie Britannique et les taux de déclin utilisés dans le présent rapport indiquaient un statut de l’UICN « gravement en péril ».
Comme indiqué précédemment, la truite arc-en-ciel résidente d’eau douce pourrait produire une descendance qui deviendrait anadrome (p. ex. Zimmerman et Reeves, 2000). Toutefois, la même documentation indique également que le phénomène est une caractéristique propre au bassin versant et que l’ampleur dans laquelle il se produit dans les bassins versants des rivières Thompson et Chilcotin n’est pas bien connue. Néanmoins, étant donné les déclins observés (figure 1 et figure 2), aucun élément ne prouve que toute contribution potentielle de la truite arc-en-ciel résidente d’eau douce atténue le récent déclin précipité de la truite arc-en-ciel anadrome, ou que ce pourrait être le cas à l’avenir.
Les auteurs tiennent à remercier les nombreux membres du COSEPAC et les membres du Sous-comité de spécialistes des poissons marins qui ont contribué au présent rapport. Alan Sinclair, John Reynolds, John Post, Greg Wilson et Robert Bison (Ministry of Forests, Lands, Natural Resource Operations and Rural Development de la Colombie-Britannique) ont dirigé la préparation de la documentation à l’appui de l’évaluation de l’urgence. Ils remercient également tous les membres de l’équipe du Secrétariat du COSEPAC qui ont offert leur aide dans le cadre de cette situation urgente.
| Sujet | Information |
|---|---|
| Durée d’une génération (généralement, âge moyen des parents dans la population; indiquez si une méthode d’estimation de la durée d’une génération autre que celle qui est présentée dans les lignes directrices de l’UICN [2011] est utilisée). | 5 ans |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d’individus matures? | Oui |
| Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations]. | Déclin de 71 % au cours des deux dernières générations |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations]. | Déclin de 79 % au cours des trois dernières générations |
| [Pourcentage [prévu ou présumé] [de changement, de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Si le pourcentage de déclin actuel persiste pendant 3 autres générations, le nombre d’individus reproducteurs diminuera pour atteindre 37 individus matures, ce qui constitue 2,0 % de l’abondance précédant 2000. |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Déclin inféré de 79 % pendant cette période si l’on présume que le taux de déclin sera le même qu’au cours des trois dernières générations |
| Est-ce que les causes du déclin sont a) clairement réversibles et b) comprises et c) ont effectivement cessé? | a. Partiellement si la mortalité causée par les prises accessoires est réduite. b. La mortalité causée par les prises accessoires est bien comprise, mais les déclins dans les milieux dulcicoles et marins le sont moins. c. Non |
| Y a-t-il des fluctuations extrêmes du nombre d’individus matures? | Non |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d’occurrence | > 20 000 km2 |
| Indice de zone d’occupation (IZO) (Fournissez toujours une valeur établie à partir d’une grille à carrés de 2 km de côté.) | < 500 km2 |
| La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d’occupation totale se trouvent dans des parcelles d’habitat qui sont a) plus petites que la superficie nécessaire au maintien d’une population viable et b) séparées d’autres parcelles d’habitat par une distance supérieure à la distance de dispersion maximale présumée pour l’espèce? | a. Non b. Non |
| Nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme. (utilisez une fourchette plausible pour refléter l’incertitude, le cas échéant) |
S.O. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d’occurrence? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l’indice de zone d’occupation? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de sous-populations? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
S.O. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l’étendue ou la qualité] de l’habitat? | Oui, il y a un déclin inféré de la qualité de l’habitat. |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d’occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation? | Non |
| Sous-populations (utilisez une fourchette plausible) | Nombre d’individus matures |
|---|---|
| Rivière Thompson, y compris la fraye dans les affluents suivants : rivières Deadman, Bonaparte et Coldwater, ruisseau Spius et rivière Nicola pendant l’année du relevé le plus récent (2017). | 177 (la moyenne des 3 dernières années est de 255) |
| Total | 177 |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l’espèce à l’état sauvage est d’au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Non calculée |
| Sujet | Information |
|---|---|
| Un calculateur des menaces a-t-il été rempli pour l’espèce? | Non. Cette population fait face à certaines menaces dans les milieux dulcicoles et marins, et bon nombre de ces menaces sont semblables à celles qui pèsent sur le saumon sockeye dans le bassin versant du fleuve Fraser et le saumon coho dans le fleuve Fraser intérieur. Les captures de truites arc-en-ciel anadromes dans le fleuve Fraser intérieur varient de 15 à 25 % par année, tout dépendant de l’abondance du saumon du Pacifique visé par la pêche commerciale. Quels sont les autres facteurs limitatifs pertinents? |
| Sujet | Information |
|---|---|
| Situation des populations de l’extérieur les plus susceptibles de fournir des individus immigrants au Canada. | La truite arc en ciel anadrome de la rivière Thompson est endémique à ce bassin versant, et l’immigration est impossible à partir d’autres populations de truites arc en ciel anadromes. L’immigration de truites non anadromes au sein de ce bassin versant est improbable. |
| Une immigration a-t-elle été constatée ou est-elle possible? | S.O. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | S.O. |
| Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants? | S.O. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
S.O. |
| Les conditions de la population source (c.-à-d. de l’extérieur) se détériorent elles? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
S.O. |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
S.O. |
| La possibilité d’une immigration depuis des populations externes existe-t-elle? | S.O. |
| Sujet | Information |
|---|---|
| L’information concernant l’espèce est-elle de nature délicate? | Non |
| Sujet | Information |
|---|---|
| COSEPAC | L’espèce n’a pas été évaluée antérieurement. |
| Sujet | Information |
|---|---|
| Statut recommandé | En voie de disparition |
| Codes alphanumériques | A2bd+4bd; C2a(i); D1 |
| Justification de la désignation | Certaines menaces pèsent sur cette espèce sauvage, notamment le déclin de la qualité de l’habitat dans les milieux dulcicoles et marins ainsi que la mortalité attribuable aux prises accessoires de la pêche au saumon du Pacifique. Le nombre d’individus reproducteurs était variable, malgré peu de tendances, avant 2000. Depuis, la population a connu un déclin considérable (79 %) au cours des trois dernières générations et a maintenant atteint son plus faible effectif reconnu à ce jour. Les 177 individus matures observés dans le cadre du plus récent relevé représentent 9,5 % de la moyenne précédant 2000. Si le pourcentage de déclin actuel persiste pendant trois autres générations, le nombre d’individus matures déclinera pour atteindre 37 individus, ce qui représente 2,0 % de l’abondance précédant 2000. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d’individus matures) | Correspond au critère de la catégorie « espèce en voie de disparition » A2bd+4bd. Le nombre d’individus matures a chuté de 79 % au cours des 3 dernières générations et il est inféré que ce déclin se poursuivra à l’avenir. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Ne correspond pas au critère. L’IZO correspond au critère de la catégorie « espèce en voie de disparition » et la qualité de l’habitat dulcicole et marin connaît un déclin, mais la population n’est pas gravement fragmentée, le critère de nombre restreint de localités ne s’applique pas et il n’y a pas de fluctuations extrêmes. |
| Critère C (nombre d’individus matures peu élevé et en déclin) | Correspond au critère de la catégorie « espèce en voie de disparition » C2a(i). Selon le relevé le plus récent, le nombre d’individus matures de l’ensemble des sous-populations est de 177, et on estime qu’aucune sous-population ne compterait plus de 250 individus. |
| Critère D (très petite population totale ou répartition restreinte) | Correspond au critère de la catégorie « espèce en voie de disparition » D1, car le nombre d’individus matures observés pendant le dernier relevé est de 177 (la moyenne des trois dernières années est de 255). |
| Critère E (analyse quantitative) | Aucune analyse quantitative n’a été effectuée. |
| Sujet | Information |
|---|---|
| Durée d’une génération (généralement, âge moyen des parents dans la population; indiquez si une méthode d’estimation de la durée d’une génération autre que celle qui est présentée dans les lignes directrices de l’UICN [2011] est utilisée). | 6 ans |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d’individus matures? | Oui |
| Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations]. | Déclin de 59 % au cours des 2 dernières générations |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations]. | Déclin de 81 % au cours des 3 dernières générations |
| [Pourcentage [prévu ou présumé] [de changement, de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Si le pourcentage de déclin persiste pendant les 3 prochaines générations, le nombre d’individus reproducteurs chutera pour atteindre 11 individus, ou environ 0,9 % de l’abondance précédant 2000. |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Déclin inféré de 81 % au cours de 3 générations si l’on présume que le pourcentage de déclin demeurera le même qu’au cours des 3 dernières générations. |
| Est-ce que les causes du déclin sont a) clairement réversibles et b) comprises et c) ont effectivement cessé? | a. Partiellement si la mortalité causée par les prises accessoires est réduite. b. La mortalité causée par les prises accessoires est bien comprise, mais les déclins dans les milieux marins et dulcicoles le sont moins. c. Non |
| Y a-t-il des fluctuations extrêmes du nombre d’individus matures? | Non |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d’occurrence | > 20 000 km2 |
| Indice de zone d’occupation (IZO) (Fournissez toujours une valeur établie à partir d’une grille à carrés de 2 km de côté). | < 500 km2 |
| La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d’occupation totale se trouvent dans des parcelles d’habitat qui sont a) plus petites que la superficie nécessaire au maintien d’une population viable et b) séparées d’autres parcelles d’habitat par une distance supérieure à la distance de dispersion maximale présumée pour l’espèce? | a. Non b. Non |
| nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme. (utilisez une fourchette plausible pour refléter l’incertitude, le cas échéant) |
S.O. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d’occurrence? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l’indice de zone d’occupation? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de sous-populations? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
S.O. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l’étendue ou la qualité] de l’habitat? | Oui, il y a un déclin inféré de la qualité de l’habitat. |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d’occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation? | Non |
| Sous-populations (utilisez une fourchette plausible) | Nombre d’individus matures |
|---|---|
| Rivière Chilcotin, y compris les affluents de fraye suivants : rivières Taseko, Chilko et Little Chilcotin pendant l’année du relevé le plus récent (2017) | 58 (la moyenne des 3 dernières années est de 120) |
| Total | 58 |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l’espèce à l’état sauvage est d’au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans] | Non calculée |
| Sujet | Information |
|---|---|
| Un calculateur des menaces a-t-il été rempli pour l’espèce? | Non. Cette population fait face à certaines menaces dans les milieux marins et dulcicoles, et bon nombre de ces menaces sont semblables à celles qui pèsent sur le saumon sockeye dans le bassin versant du fleuve Fraser et le saumon coho dans le fleuve Fraser intérieur. Les captures de truites arc-en-ciel anadromes dans le fleuve Fraser intérieur varient de 15 à 25 % par année, tout dépendant de l’abondance du saumon visé par la pêche commerciale.
Quels sont les autres facteurs limitatifs pertinents? |
| Sujet | Information |
|---|---|
| Situation des populations de l’extérieur les plus susceptibles de fournir des individus immigrants au Canada. | La truite arc-en-ciel anadrome de la rivière Chilcotin est endémique à ce bassin versant et l’immigration d’autres populations de truites arc-en-ciel anadromes est impossible. L’immigration de truites non anadromes au sein de ce bassin versant est improbable. |
| Une immigration a-t-elle été constatée ou est-elle possible? | S.O. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | S.O. |
| Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants? | S.O. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
S.O. |
| Les conditions de la population source (c.-à-d. de l’extérieur) se détériorent-elles? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
S.O. |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
S.O. |
| La possibilité d’une immigration depuis des populations externes existe-t-elle? | S.O. |
| Sujet | Information |
|---|---|
| L’information concernant l’espèce est-elle de nature délicate? | Non |
| Sujet | Information |
|---|---|
| COSEPAC | L’espèce n’a pas été évaluée antérieurement. |
| Sujet | Information |
|---|---|
| Statut recommandé | En voie de disparition |
| Codes alphanumériques | A2bd+4bd; C2a(i,ii); D1 |
| Justification de la désignation | Certaines menaces pèsent sur cette espèce sauvage, notamment le déclin de la qualité de l’habitat dans les milieux marins et dulcicoles ainsi que la mortalité attribuable aux prises accessoires de la pêche au saumon du Pacifique. La population a connu un déclin considérable (81 %) au cours des trois dernières générations et a maintenant atteint son plus faible effectif reconnu à ce jour. Le nombre d’individus reproducteurs était élevé et variable, malgré peu de tendances, avant 2000. Les 58 individus matures observés pendant le relevé le plus récent représentent seulement 5 % de la moyenne précédant 2000. Si le pourcentage de déclin actuel persiste pendant trois autres générations, le nombre d’individus reproducteurs déclinera pour atteindre 11 individus, ce qui représente 0,9 % de l’abondance précédant 2000. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d’individus matures) | Correspond au critère de la catégorie « espèce en voie de disparition » A2bd+4bd. Le nombre d’individus matures a chuté de 81 % au cours des trois dernières générations et il est inféré que ce déclin se poursuivra dans un avenir rapproché. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Ne correspond pas au critère. L’IZO correspond au critère de la catégorie « espèce en voie de disparition » et la qualité de l’habitat dulcicole et marin connaît un déclin, mais la population n’est pas gravement fragmentée, le critère de nombre restreint de localités ne s’applique pas et il n’y a pas de fluctuations extrêmes. |
| Critère C (nombre d’individus matures peu élevé et en déclin) | Correspond au critère de la catégorie « espèce en voie de disparition » C2a(i,ii). Le nombre d’individus matures était de 58 dans le relevé le plus récent, le nombre d’individus matures a connu un déclin de 59 % au cours des deux dernières générations, aucune sous-population ne compte plus de 250 individus et une seule sous-population compte plus de 95 % de tous les individus matures. |
| Critère D (très petite population totale ou répartition restreinte) | Correspond au critère de la catégorie « espèce en voie de disparition » D1, car le nombre d’individus matures est inférieur à 250 (58 pendant l’année la plus récente, et une moyenne de 120 au cours des trois dernières années). |
| Critère E (analyse quantitative) | Aucune analyse quantitative n’a été effectuée. |
| Nom | Rôle |
|---|---|
| Eric Taylor | Président du COSEPAC |
| John Neilson | Coprésident, SCS des poissons marins |
| Dwayne Lepitzki | Coprésident, SCS des mollusques |
| Dave Fraser | Responsable - Colombie-Britannique |
| Greg Wilson | Responsable - Colombie-Britannique |
| Syd Cannings | Responsable - SCF |
| Jennifer Shaw | Responsable - MPO |
| Simon Nadeau | Responsable - MPO |
| Robert Bison | Spécialiste des pêches du secteur, gouvernement de la Colombie-Britannique |
| John Post | Coprésident, SCS des poissons d’eau douce |
| John Reynolds | Membre scientifique non gouvernemental |
| Alan Sinclair | SCS, évaluations d’urgence, membre spécial ou observateur |
| Arne Mooers | Membre scientifique non gouvernemental |
| Donna Hurlburt | Coprésidente, Sous-comité des CTA |
| Roger Gallant | Coprésident, Sous-comité des CTA |
| Paul Grant | Coprésident, SCS des arthropodes |
| Ross Claytor | Coprésident, SCS des poissons marins |
| Aaron McNeil | Membre, SCS des poissons marins |
| Bruce Atkinson | Membre, SCS des poissons marins |
| Craig Purchase | Membre, SCS des poissons marins |
| David Hardie | Membre, SCS des poissons marins |
| Ian Fleming | Membre, SCS des poissons marins |
| Laura Weir | Membre, SCS des poissons marins |
| Margaret Treble | Membre, SCS des poissons marins |
| Nancy Shackell | Membre, SCS des poissons marins |
| Peter Westley | Membre, SCS des poissons marins |
| Carrie Holt | Membre, SCS des poissons marins |
| Marc Trudel | Membre, SCS des poissons marins |
| Bev McBride | Secrétariat |
| Karen Timm | Secrétariat |
| Lisa Twolan | Secrétariat |
La Colombie-Britannique abrite une myriade de populations de truites arc-en-ciel anadromes (Oncorhynchus mykiss anadrome) réparties depuis les zones côtières méridionales jusqu’au nord-ouest de la province, ces populations étant potentiellement présentes dans environ 1 200 bassins versants ou plus (figure A1). Les truites arc-en-ciel anadromes des rivières Thompson et Chilcotin (TAECA-RTC) constituent deux unités désignables (UD) dans cet assemblage, car elles satisfont aux critères de caractère distinct et d’importance nécessaires à la reconnaissance des UD (COSEWIC, 2016).
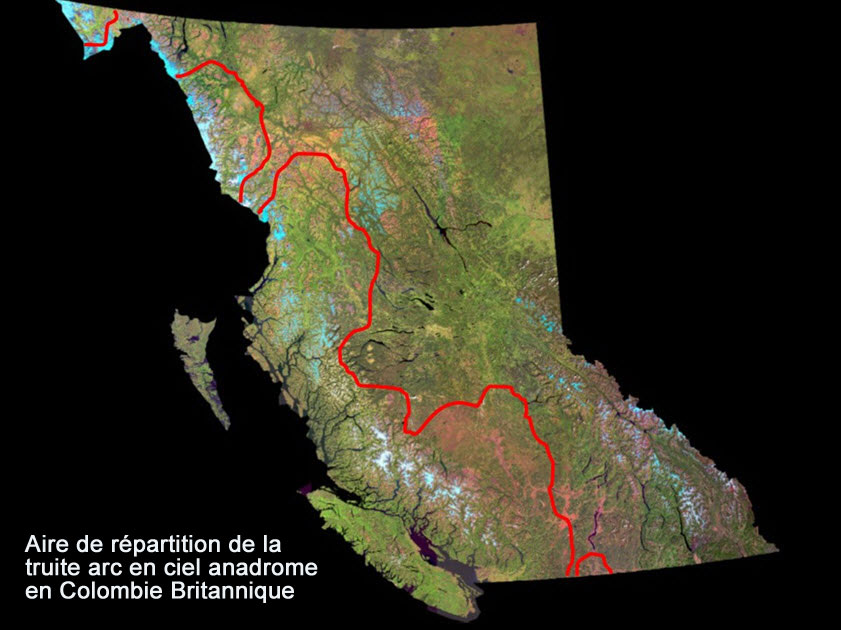
Description longue de la figure A1
Les descriptions longues sont fournies dans le texte d’accompagnement.
Les TAECA-RTC frayent dans les affluents des rivières Thompson et Chilcotin du bassin versant du fleuve Fraser, et elles sont donc séparées sur le plan spatial des autres populations de truites arc-en-ciel anadromes présentes en Colombie Britannique. Compte tenu de la fidélité bien documentée de la plupart des salmonidés anadromes, comme la truite arc en ciel anadrome, à leur frayère natale, les populations de truites arc en ciel anadromes du Fraser intérieur sont très structurées sur le plan spatial. Par exemple, Beacham et al. (2004) ont utilisé 14 locus de microsatellites d’ADN pour montrer que les TAECA-RTC et un groupe de truites arc en ciel anadromes du mi Fraser (TAECA-MF; rivières Stein, Nahatlatch et Bridge) constituent un regroupement bien défini de populations (valeur de bootstrap de 75 %, figure 2 dans Beacham et al., 2004), distinct de 46 autres populations réparties depuis le nord ouest de la Colombie Britannique jusque dans des portions américaines du haut Columbia. En fait, les TAECA-RTC (et les TAECA-MF) ressemblent plus, sur le plan génétique, aux truites arc en ciel anadromes du haut Columbia qu’elles ne ressemblent à celles du bas Fraser (présentes notamment dans les rivières Chilliwack et Coquihalla; figure 2 dans Beacham et al., 2004). Les TAECA-RTC font également partie du groupe phylogénétique d’ascendance mixte de la partie sud de la zone côtière et de l’intérieur, tel qu’inféré à partir de leur ADNmt qui est unique en Colombie Britannique (figures 9 et 10 dans McCusker et al., 2000; figures A2 et A3). De plus, la composante de la rivière Thompson (TAECA-RT) des TAECA-RTC se distingue des TAECA-MF ainsi que de la truite arc en ciel anadrome de la rivière Chilcotin (TAECA-RC) dans les analyses réalisées au moyen des mêmes locus microsatellites (valeur de bootstrap de 98 %). Des tests relatifs aux fréquences alléliques, fondés sur des données de microsatellites d’ADN de Beacham et al. (2004) et sur quatre locus d’alloenzymes polymorphes examinés par Parkinson (1984, tableau 1), indiquent toutefois que les TAECA-RC sont aussi significativement distinctes des trois échantillons de TAECA-MF (toujours P < 0,001, E. Taylor, Université de la Colombie Britannique, Vancouver, résultats inédits). Dans le cas des alloenzymes, la majeure partie de la différentiation est attribuable à des différences entre les échantillons de la rivière Chilcotin et des rivières Stein/Nahatlatch, mais un locus (AGP) permet aussi de distinguer les TAECA-RC des individus de la rivière Bridge (test de randomisation, P = 0,0003). Ces deux échantillons se sont aussi avérés distincts de façon significative si on combine les probabilités pour les quatre locus (test de probabilité combinée de Fisher, P < 0,001, E. Taylor, Université de la Colombie Britannique, Vancouver, analyse inédite). Parmi les échantillons de TAECA-MF, ce sont les échantillons de truites arc en ciel anadromes de la rivière Bridge qui ressemblent le plus aux TAECA-RC sur le plan des locus de microsatellites d’ADN. La proportion de la variation totale des fréquences alléliques des microsatellites attribuable à des différences entre les individus de la rivière Bridge et les TAECA-RC (FST) est de 2,5 % (P < 0,002), tandis qu’elle se situe entre 5,6 % et 8,8 % entre les individus des rivières Nahatlatch et Stein et les TAECA-RC, respectivement (les deux ayant une valeur P < 0,002, T.D. Beacham, MPO, Nanaimo, Colombie Britannique, comm. pers., 12 janvier 2017, d’après les données de Beacham et al., 2004). Malgré que le degré de différenciation soit variable, les TAECA-RTC sont manifestement distinctes de toutes les autres truites arc en ciel anadromes de la Colombie Britannique, et les populations de la rivière Thompson et de la rivière Chilcotin sont distinctes entre elles (voir ci dessous).
Les données génétiques mentionnées ci-dessus font aussi ressortir l’importance des TAECA-RTC en tant qu’assemblage distinct de truites arc-en-ciel anadromes. Les données de microsatellites et d’ADNmt semblent indiquer que les TAECA-RTC ont connu une évolution glaciaire et postglaciaire unique en Colombie-Britannique; elles ont une étroite affinité avec les truites arc-en-ciel anadromes de la partie sud de la zone côtière (ADNmt) ainsi qu’avec celles du haut Columbia (microsatellites). Cela porte à croire que les TAECA-RTC pourraient être le résultat d’une double invasion du paysage marin actuel en provenance de deux refuges glaciaires - encore là, une situation qui semble être unique au patrimoine évolutionnaire de la truite arc-en-ciel anadrome de la Colombie-Britannique (figure 9 dans McCusker et al. 2000; figures A2 et A3).
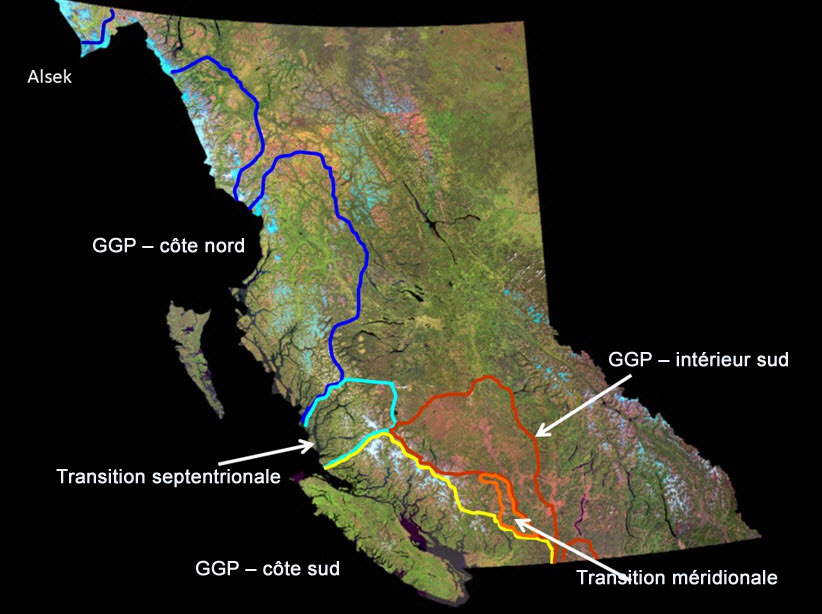
Description longue de la figure A2
Les descriptions longues sont fournies dans le texte d’accompagnement.
D’autres données appuyant l’importance sur le plan évolutif du caractère distinct des TAECA-RTC par rapport aux autres truites arc en ciel anadromes proviennent d’études de la différenciation des alloenzymes et de son lien apparent avec la résistance natatoire. Les TAECA-RTC, représentées par des échantillons provenant de la rivière Thompson, présentent des fréquences plus élevées de phénotypes du lactate-déshydrogénase qui sont associés à une performance en nage de longue durée nettement supérieure, comparativement aux truites arc en ciel anadrome du bas Fraser (Tsuyuki et Williscroft, 1977). De telles différences physiologiques sont aussi apparentes entre les populations côtières et de l’intérieur du saumon coho (Taylor et McPhail, 1985), et elles mettent en évidence les caractéristiques adaptatives réelles et potentielles des salmonidés qui migrent vers l’amont sur de longues distances dans le fleuve Fraser. Parmi les autres différences entre les TAECA-RTC et les truites arc en ciel anadrome de la partie sud de la zone côtière, on compte le moment de la remonte en automne et l’état immature des gonades durant la migration - un phénomène que l’on pourrait appeler « migration prématurée ». En revanche, les truites arc-en-ciel anadrome de la partie sud de la zone côtière migrent habituellement par le bas Fraser après les TAECA-RTC, et leurs gonades sont dans un état de maturité plus avancé. Le phénotype associé à la migration prématurée semble avoir une base génétique relativement simple et ferait l’objet d’une forte sélection positive, et il est considéré comme essentiel à la persistance de la biodiversité de la truite arc en ciel anadrome dans d’autres portions de son aire de répartition (Prince et al., 2017). Les TAECA-RTC diffèrent aussi sur plusieurs aspects, notamment le moment et la vitesse de la migration, l’âge à la maturation et l’âge à la smoltification, des truites arc en ciel anadrome du bas Fraser et du mi-Fraser. Ces différences sont particulièrement évidentes entre les individus de la rivière Chilcotin et tous les autres individus, et elles reflètent probablement des adaptations à la migration plus longue et plus ardue dans la rivière Chilcotin et les climats distincts dans lesquels elles vivent (voir ci dessous).
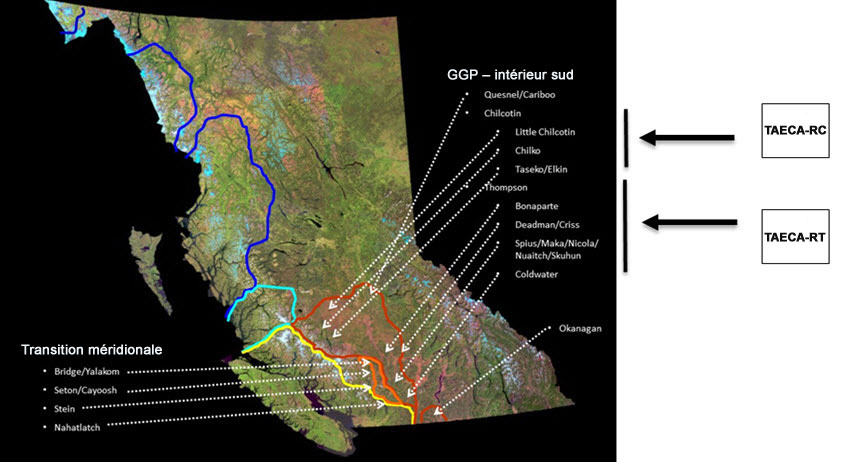
Description longue de la figure A3
Les descriptions longues sont fournies dans le texte d’accompagnement.
Compte tenu de ce qui précède, les truites arc-en-ciel anadromes des rivières Thompson et Chilcotin (TAECA-RTC) satisfont à la fois aux critères de caractère distinct et d’importance nécessaires à leur reconnaissance comme constituant au moins une UD distincte au sein de l’espèce O. mykiss. De plus, il est proposé que les TAECA-RTC soient subdivisées en deux UD distinctes l’une de l’autre : la truite arc-en-ciel anadrome de la rivière Thompson (TAECA-RT) et la truite arc-en-ciel anadrome de la rivière Chilcotin (TAECA-RC).
Comme il a été mentionné ci-dessus, les données génétiques de Beacham et al. (2004) identifient clairement (c.-à-d. selon une valeur de bootstrap de 98 %) la TAECA-RT comme un groupe génétique distinct d’autres truites arc-en-ciel anadromes, y compris la TAECA-RC. La distance génétique (FST) entre la TAECA-RT et la TAECA-RC aux locus de microsatellites représente entre 6,2 % et 8,3 % de la variation totale lorsqu’on analyse ces deux échantillons (toujours P < 0,001, comm. pers de T.D. Beacham, MPO, Nanaimo, Colombie-Britannique, 12 janvier 2017, basé sur des données dans Beacham et al., 2004). Les données de Parkinson (1984) ont aussi montré que la TAECA-RC présentait des génotypes multilocus sur quatre locus d’alloenzymes (SOD, LDH, MDH, et AGP) qui étaient distincts par rapport à des échantillons de truites arc-en-ciel anadromes de la rivière Thompson et du mi-Fraser (voir ci-dessus). La TAECA-RT et la TAECA-RC sont aussi distinctes l’une de l’autre sur le plan spatial (voir ci-dessous) et en termes de phénotypes, plus particulièrement en ce qui concerne l’âge à la maturation, le moment et le comportement de migration et l’âge à la smoltification (abordé ci-dessous à la section sur l’importance).
La TAECA-RT et la TAECA-RC se distinguent par plusieurs aspects relatifs à la migration, soit le moment, la vitesse et le comportement, que l’on peut interpréter comme étant des adaptations aux différents emplacements de leurs frayères. En général, les études du mélange génétique et de télémesure indiquent que la TAECA-RC se retrouve plus tôt dans le fleuve Fraser, migre vers l’amont plus rapidement et fait moins souvent halte (en restant relativement stationnaire à certains endroits) en route vers les frayères et les aires d’hivernage que la TAECA-RT. Par exemple, Bison (données inédites) rapporte une différence moyenne dans la date de passage au kilomètre 235 (près de la rivière Nahatlatch) lors de la migration de 13,8 jours pour la TAECA-RC (c.-à-d. que les individus sont arrivés, en moyenne, presque 14 jours avant la date moyenne pour toutes les populations), comparativement à 0,2 à -4,3 jours pour la TAECA-RT (et à -1,6 à -8,3 jours pour les TAECA-MF qui arrivent plus tard, respectivement; N = 49 individus munis de radioémetteurs provenant de toutes les zones). Ces différences découlent probablement de la migration plus hâtive et plus directe de la TAECA-RC, parce que les individus doivent surmonter trois obstacles majeurs à la migration avant le début de l’hiver (deux dans le canyon du bas Fraser aux kilomètres 185 et 210, et un aux rapides de la rivière Bridge au kilomètre 340). En revanche, la TAECA-RT n’a que deux obstacles à surmonter dans le canyon du bas Fraser. De plus, les TAECA-RC ont une plus grande distance à parcourir pour se rendre à leurs sites d’hivernage, qui se trouvent au moins aussi loin en amont que le kilomètre 522 de la rivière Chilcotin et que le kilomètre 510 du Fraser (~100 km en amont de la confluence du Fraser et de la rivière Chilcotin; Renn et al., 2001). Pour leur part, les TAECA-RT n’hivernent pas plus loin en amont que la décharge du lac Kamloops au kilomètre 375 depuis le point où le Fraser se déverse dans le détroit de Georgia (note : les truites de la rivière Nahatlatch qui arrivent le plus tard ne parcourent que 238 km à partir de l’embouchure du Fraser). Finalement, la TAECA-RT et la TAECA-RC se distinguent l’une de l’autre à la fois par l’âge à la smoltification et l’âge auquel les adultes reviennent en eau douce; la majorité des smolts de la TAECA-RT migrent vers la mer à deux ans (93 %), tandis que la majorité des smolts de la TAECA-RC le font à trois ans (83 %; Bison, 2012). Le fait que la smoltification a lieu à un âge plus tardif chez la TAECA-RC fait en sorte que les adultes reviennent à un âge plus avancé. Habituellement, les TAECA-RT frayent pour la première fois à cinq ans (plus rarement à six ou sept ans), tandis que les TAECA-RC frayent pour la première fois à six ans (rarement à sept ou huit ans) (N = 14 à 215 individus annuellement sur 40 ans de suivi; Bison, 2012). L’âge plus tardif de smoltification et de maturation chez les TAECA-RC résulte probablement d’adaptations à leur migration plus longue et/ou plus ardue (voir ci-dessus). Finalement, la TAECA-RT et la TAECA-RC sont présentes dans des zones biogéoclimatiques différentes de la Colombie-Britannique; la TAECA-RT est principalement présente dans la zone intérieure à douglas, alors que la TAECA-RC est présente dans un mélange de plusieurs zones plus petites. La zone intérieure à douglas est considérée davantage comme une région semi-désertique où la température annuelle moyenne de l’air est généralement plus élevée (voir la figure A4). La température plus élevée pourrait offrir de meilleures possibilités de croissance aux smolts de la TAECA-RT et pourrait, en partie, expliquer pourquoi la smoltification se produit à un plus jeune âge moyen chez celle-ci comparativement à la TAECA-RC. Plusieurs études ont fourni des données indiquant une divergence en ce qui concerne la physiologie de la tolérance thermique chez l’O. mykiss au sein de populations de l’extérieur de la Colombie-Britannique, le long d’un gradient environnemental désertique-montagnard similaire (Rodnick et al., 2004; Narum et al., 2010, 2013); il se peut que des différences semblables existent entre la TAECA-RT et la TAECA-RC.
La TAECA-RT et la TAECA-RC devraient être évaluées comme étant deux UD distinctes.
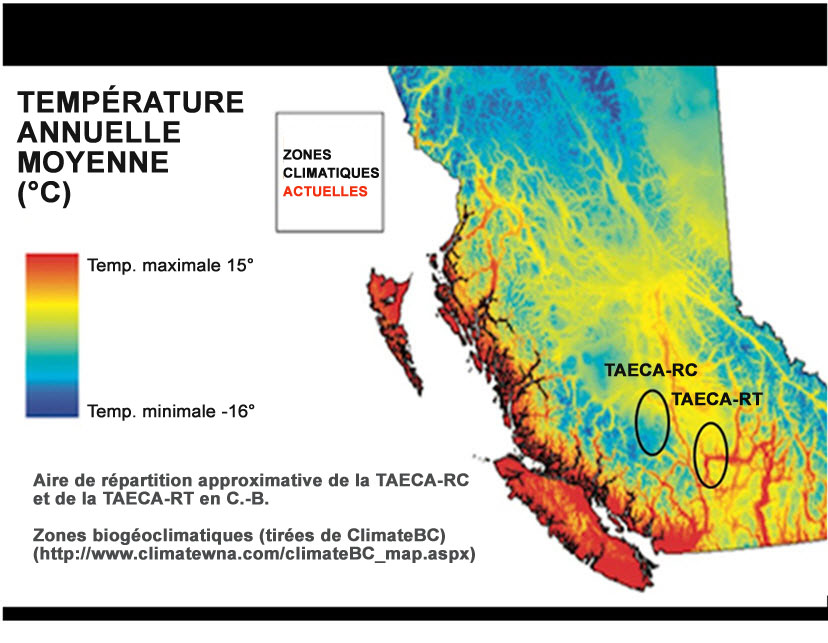
Description longue de la figure A4
Les descriptions longues sont fournies dans le texte d’accompagnement.
Abdul-Aziz, O., Mantua, N. et Myers, K.W. 2011. Potential climate change impacts on thermal habitats of Pacific salmon (Oncorhynchus spp.) in the North Pacific Ocean and adjacent seas. Canadian Journal of Fisheries and Aquatic Sciences 68:1660-1680.
Berejikian, B.A., Moore, M.E. et Jeffries, S.J. 2016. Predator-prey interactions between harbor seals and migrating steelhead trout smolts revealed. Marine Ecology Progress Series 543: 21-35.
Bison, R. 2012. Population attributes for Fraser River late-run summer steelhead. Unpublished MS. BC Ministry of Natural Resource Operations, Fish & Wildlife Branch, Kamloops, BC. 55 p.
Bison, R. 2016. Fishing Mortality Trends for Thompson River Steelhead from 1991 to 2015. BC Ministry of Natural Resource Operations, Fish & Wildlife Branch, Kamloops, BC. 30 p.
Bison, R. et Phelps, M. 2017. Statistics for the 2016 Thompson River Steelhead Sport Fishery and Relationships between the Fishery and Steelhead Abundance. BC Ministry of Natural Resource Operations, Fish & Wildlife Branch, Kamloops, BC. 37 p.
Beacham, T.D., Le, K.D. et Candy, J.R., 2004. Population structure and stock identification of Steelhead Trout (Oncorhynchus mykiss) in British Columbia and the Columbia River based on microsatellite variation. Environmental Biology of Fishes 69:95-109.
ClimateBC. 2017. An interactive platform for visualization and data access. Centre for Forest Conservation Genetics, University of British Columbia, Vancouver. ClimateBC_Map (consulté le 23 décembre 2017).
COSEPAC. 2010. Évaluation et rapport de situation du COSEPAC sur le Saumon atlantique, Salmo salar, population du Nunavik, population du Labrador, population du nord-est de Terre-Neuve, population du sud de Terre-Neuve, population du sud-ouest de Terre-Neuve, population du nord-ouest de Terre-Neuve, population de l’est de la Côte-Nord du Québec, population de l’ouest de la Côte-Nord du Québec, population de l’île d’Anticosti, population de l’intérieur du Saint-Laurent, population du lac Ontario, population de la Gaspésie-sud du golfe Saint-Laurent, population de l'est du Cap-Breton, population des hautes terres du sud de la Nouvelle-Écosse, population de l'intérieur de la baie de Fundy, population de l’extérieur de la baie de Fundy, au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xlix + 162 p.
COSEPAC. 2016. Manuel des opérations et des procédures. Annexe F5. Comité sur la situation des espèces en péril au Canada. Ottawa. Sous presse.)
COSEWIC. 2018. COSEWIC assessment and status report on the Sockeye Salmon (Oncorhynchus nerka) in the Fraser River drainage. Committee on the Status of Endangered Wildlife in Canada. Ottawa. Sous presse.
Docker, M.F. et Heath, D.D. 2003. Genetic comparison between sympatric anadromous steelhead and freshwater resident rainbow trout in British Columbia, Canada. Conservation Genetics 4:227-231.
Hard, J.J., J.M. Myers, E.J. Connor, R.A. Hayman, R.G. Kope, G. Lucchetti, A.R. Marshall, G.R. Pess et B.E. Thompson. 2015. Viability criteria for steelhead within the Puget Sound distinct population segment. U.S. Dept. Commerce, NOAA Tech. Memo. NMFS-NWFSC-129, 367 p.
Hinch, S.G. et E.G. Martins. 2011. A review of potential climate change effects on survival of Fraser River sockeye salmon and an analysis of interannual trends in en route loss and pre-spawn mortality. Cohen Commission Tech. Rept. 9: 134p. Vancouver, B.C.
Kendall, N.W., Marston, G.W. et Klungle, M.M. 2017. Declining patterns of Pacific Northwest steelhead trout (Oncorhynchus mykiss) adult abundance and smolt survival in the ocean. Canadian Journal of Fisheries and Aquatic Sciences 74:1275-1290.
McCusker, M.R., Parkinson, E.A. et Taylor, E.B. 2000. Phylogenetic Conservation Units for Rainbow Trout in British Columbia. Province of British Columbia, Fisheries Management Report 112. University of British Columbia, Vancouver, B.C. 35 p.
McMillan, J.R., Katz, S.L. et Pess, G.R., 2007. Observational evidence of spatial and temporal structure in a sympatric anadromous (winter steelhead) and resident rainbow trout mating system on the Olympic Peninsula, Washington. Transactions of the American Fisheries Society 136: 736-748.
McPhail, J.D. 2007. Freshwater fishes of British Columbia. University of Alberta Press, Edmonton.
Narum, S.R., Campbell, N.R., Kozfkay, C.C. et Meyer, K.A., 2010. Adaptation of redband trout in desert and montane environments. Molecular Ecology 19(21):4622-4637.
Narum, S.R., Campbell, N.R., Meyer, K.A., Miller, M.R. et Hardy, R.W. 2013. Thermal adaptation and acclimation of ectotherms from differing aquatic climates. Molecular Ecology 22(11):3090-3097.
Parkinson, E.A. 1984. Genetic variation in populations of Steelhead Trout (Salmo gairdneri) in British Columbia. Canadian Journal of Fisheries and Aquatic Sciences 41:1412-1420.
Pearse, D.E., Hayes, S.A., Bond, M.H., Hanson, C.V., Anderson, E.C., Macfarlane, R.B. et Garza, J.C., 2009. Over the falls? Rapid evolution of ecotypic differentiation in steelhead/rainbow trout (Oncorhynchus mykiss). J. Heredity 100: 515-525.
Prince, D.J., O’Rourke, S.M., Thompson, T.Q., Ali, O.A., Lyman, H.S., Saglam, I.K., Hotaling, T.J., Spidle, A.P. et Miller, M.R. 2017. The evolutionary basis of premature migration in Pacific salmon highlights the utility of genomics for informing conservation. Science Advances 3(8), p.e1603198.
Renn, J.R., Bison R. Hagen, J. et Nelson, T.C. 2001. Migration characteristics and stock composition of interior Fraser steelhead as determined by radio telemetry, 1996-1999. BC Ministry of Water, Land and Air Protection, Fisheries Branch. 150p.
Rodnick, K.J., Gamperl, A.K., Lizars, K.R., Bennett, M.T., Rausch, R.N. et Keeley, E.R. 2004. Thermal tolerance and metabolic physiology among redband trout populations in south-eastern Oregon. J. Fish Biology 64(2):310-335.
Taylor, E.B. et McPhail, J.D., 1985. Variation in burst and prolonged swimming performance among British Columbia populations of coho salmon, Oncorhynchus kisutch. Canadian Journal of Fisheries and Aquatic Sciences 42(12):2029-2033.
Thrower, F. P., Hard, J.J. et Joyce, J.E. 2004. Genetic architecture of growth and early life-history transitions in anadromous and derived freshwater populations of steelhead. J. Fish Biology 65: 286-307.
Tsuyuki, H. et Williscroft, S.N., 1977. Swimming stamina differences between genotypically distinct forms of rainbow (Salmo gairdneri) and Steelhead Trout. Journal of the Fisheries Board of Canada 34:996-100.
Viola, A.E. et Schuck, M.L., 1995. A method to reduce the abundance of residual hatchery steelhead in rivers. North American Journal of Fisheries Management 15:488-493.
Zimmerman, C.E. et Reeves, G.H. 2000. Population structure of sympatric anadromous and nonanadromous Oncorhynchus mykiss: evidence from spawning surveys and otolith microchemistry. Canadian Journal of Fisheries and Aquatic Sciences 57:2152-2162.