
Killer whales (Orcinus orca) recovery strategy: chapter 1
1. Background
The status report and assessment summary for resident killer whales is available from the Committee on the Status of Endangered Wildlife in Canada (COSEWIC) Secretariat.
COSEWIC Assessment Summary
Common Name:
killer whale, orca
Scientific Name:
Orcinus orca
Assessment Summary:
Assessed in 1999, reviewed and revised in 2001
COSEWIC Status:
‘Southern resident’ killer whales are designated as endangered, ‘northern resident’ killer whales as threatened
Species at Risk Act (SARA) Status:
‘Southern resident’ killer whales, endangered, on Schedule 1
‘Northern resident’ killer whales, threatened on Schedule 1
Reason for Designation:
The southern resident killer whale population is small, with recent declines of 17% between 1995 and 2001, and currently contains 85 members. The northern resident killer whale population is small at 205 members, with recent declines of 7% between 1997 and 2003. Seasonally, they are exposed to high levels of boat traffic. The availability of their prey is reduced relative to historic levels. High levels of persistent organic pollutants may be compromising their reproductive and immune systems, leading to reduced calving and/ or increased mortality rates.
Range in Canada:
Pacific Ocean
Status History:
In April 1999, the two North Pacific ‘resident’ killer whale populations were designated threatened. In November 2001, the southern resident population was designated endangered while the northern resident population remained threatened.
1.1.1 Species description
The killer whale is the largest member of the dolphin family, Delphinidae. Its size, striking black and white colouring and tall dorsal fin are the main identifying characteristics. Killer whales are mainly black above and white below, with a white oval eye patch, and a grey saddle patch below the dorsal fin. Each killer whale has a uniquely shaped dorsal fin and saddle patch, and most animals have naturally acquired nicks and scars. Individual killer whales are identified using photographs of the dorsal fin, saddle patch, and sometimes eye patches (Ford et al. 2000). They are sexually dimorphic. Maximum recorded lengths and weights for male killer whales are 9.0 m, and 5568 kg respectively, whereas females are smaller at 7.7 m and 4000 kg (Dahlheim and Heyning 1999). The tall triangular dorsal fin of adult males is often as high as 1.8 m, while in juveniles and adult females it reaches 0.9 m or less. In adult males, the paddle-shaped pectoral fins and tail flukes are longer and broader and the fluke tips curl downward (Bigg et al. 1987).
Currently, most authorities consider killer whales to be one species, Orcinus orca, having regional variations in diet, size, colouration, and vocal patterns (Heyning and Dahlheim 1988, Ford et al. 2000, Barrett-Lennard and Ellis 2001). Two and possibly three distinct species have recently been proposed for Antarctic populations (Mikhalev et al. 1981, Berzin and Vladimorov 1983, Pitman and Ensor 2003), but they are not currently widely accepted (Reeves et al. 2004). In addition, recent genetic studies report little global variation in mitochondrial DNA suggesting that the population segregation indicated by the morphological differences described above is relatively recent (Barrett-Lennard 2000, Hoelzel et al. 2002).
Three distinct forms, or ecotypes, of killer whale inhabit Canadian Pacific waters: transient, offshore and resident. These forms are sympatric but socially isolated and differ in their dietary preferences, genetics, morphology and behaviour (Ford et al. 1998, 2000, Barrett-Lennard and Ellis 2001). Transient killer whales feed on marine mammals; particularly harbour seals (Phoca vitulina), porpoises, and sea lions (Ford et al. 1998). They travel in small, acoustically quiet groups that rely on stealth to find their prey (Ford and Ellis 1999). To the experienced eye, the dorsal fins of transient whales tend to be pointed and their saddle patches are large and uniformly grey (Ford et al. 2000). Offshore killer whales are not as well understood as residents and transients, but they are thought to feed on fish (Ford et al. 2000, Heise et al. 2003). They travel in large acoustically active groups of 30 or more whales, using frequent echolocation and social calls (Ford et al. 2000). The dorsal fins of offshore killer whales are more rounded than those of transients, and their saddle patches may either be uniformly grey or may contain a black region.
Resident killer whales are the best understood of the three ecotypes. They feed exclusively on fish and cephalopods and travel in acoustically active groups of 10 to 25 or more whales (Ford et al. 2000). The tips of their dorsal fins tend to be rounded at the leading edge and have a fairly abrupt angle at the trailing edge. Their saddle patches may be uniformly grey or contain a black region. The social organization of resident killer whales is highly structured. Their fundamental unit is the matriline, comprising all surviving members of a female lineage. A typical matriline comprises an adult female, her offspring, and the offspring of her daughters. Both sexes remain within their natal matriline for life (Bigg et al. 1990). Social systems in which both sexes remain with their mother for life has only been described in one other mammalian species, the long-finned pilot whale (Globicephala melas) (Amos et al. 1993). Bigg et al. (1990) defined pods as groups of closely related matrilines that travel, forage, socialize and rest with each other at least 50% of the time, and predicted that pods, like matrilines, would be stable over many generations. However, Ford and Ellis (2002) showed that inter-matriline association patterns in the northern residents have evolved over the past decade such that some of the pods identified by Bigg et al. now fail to meet the 50% criterion. Their analysis suggests that pods are best defined as transitional groupings that reflect the relatedness of recently diverged matrilines.
Each resident pod has a unique dialect made up of approximately a dozen discrete calls (Ford 1989, 1991). These dialects can be distinguished, providing each pod with a unique acoustic signature. Dialects are probably learned from mothers and other associated kin and are highly stable over time (Ford et al. 2000). Their function is not entirely understood, although it appears that they play an important role in mate selection (Barrett-Lennard 2000, discussed below in Section 1.4.1. Culture). Despite having distinct dialects, some pods share certain calls and call variants. Pods that share one or more calls belong to a common clan.
Resident killer whales that share a common range and that associate at least occasionally are considered to be members of the same community or population. There are two communities of resident killer whales in British Columbia, the northern residents and the southern residents. They have not been observed interacting and genetic studies have revealed that the two populations rarely if ever interbreed (Barrett-Lennard and Ellis 2001). The northern resident community consists of three clans, and the southern resident community consists of one.
The existence of two distinct populations of resident killer whales using the waters of Washington and British Columbia has been recognized by both the Canadian and US governments. In 2001 COSEWIC assigned northern residents 'threatened' status, and southern residents 'endangered' status. In the United States, marine mammals are afforded federal protection under both the Marine Mammal Protection Act (MMPA) and, when listed, under the Endangered Species Act (ESA). The southern residents were listed as 'depleted' under the MMPA in 2003. In February 2006, southern resident killer whales were listed as endangered under the ESA. In June 2004, the Washington State Department of Fish and Wildlife added southern resident killer whales to their endangered species list.
1.2 Distribution
1.2.1 Global range
Killer whales are found in all oceans, and are most common in areas associated with high ocean productivity in mid to high latitudes (Forney and Wade in press). They are able to tolerate temperatures ranging from those found in polar waters to the tropics, and have been recorded in water ranging from shallow (several metres) to open ocean depths (Baird 2001).
1.2.2 Canadian Pacific range
Killer whales are found in all three of Canada's oceans, as well as occasionally in Hudson Bay and in the Gulf of St. Lawrence, but they appear to be uncommon in the Atlantic and the Arctic (COSEWIC 2003). In British Columbia (BC), they have been recorded throughout almost all salt-water areas, including many long inlets, narrow channels and deep embayments (Baird 2001). The three ecotypes of BC killer whales (offshore, transient, and resident) do not appear to interact socially despite their overlapping ranges (Ford et al. 2000). Offshore killer whales are most often sighted on the continental shelf off the outer coast, but they are occasionally found in protected inside waters (Ford et al. 2000). Transient killer whales range throughout the area, as do resident killer whales (Ford and Ellis 1999, Ford et al. 2000). Residents and transients have occasionally been seen in close proximity to each other, but rarely interact (Ford and Ellis 1999). Figure 1 shows many place names mentioned in the text, as well as the general ranges of northern and southern residents.
Figure 1: The coast of British Columbia and northwest Washington state showing the general ranges of northern and southern resident killer whales
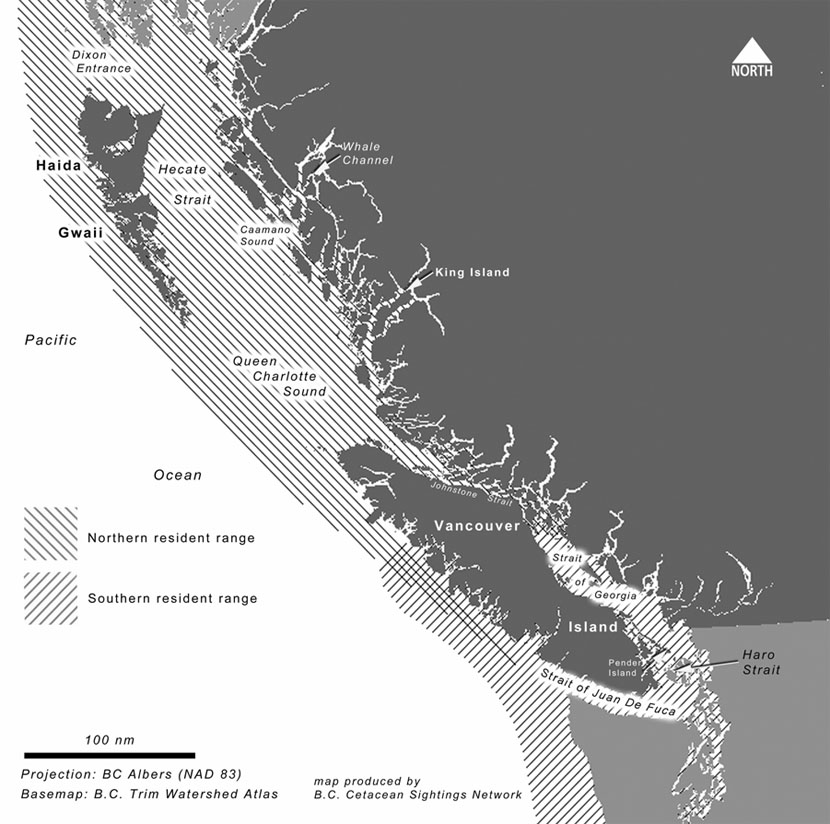
The community of southern residents comprises a single acoustic clan, J clan, which is composed of three pods (referred to as J, K, and L) containing a total of 20 matrilines (Ford et al. 2000). The known range of this community is from northern British Columbia to central California (Ford et al. 2000; unpublished data, Cetacean Research Program, Fisheries & Oceans Canada, Pacific Biological Station, Nanaimo, BC [Cetacean Research Program-Fisheries and Oceans Canada (CRP-DFO)]). During summer, its members are usually found in waters off southern Vancouver Island and northern Washington State, where they congregate to intercept migratory salmon. The main area of concentration for southern residents is Haro Strait and vicinity off southeastern Vancouver Island (Figure 1), but they are commonly seen in Juan de Fuca Strait, and the southern Strait of Georgia (Ford et al. 2000). Of the three southern resident pods, J pod is most commonly seen in inside waters throughout the year, and appears to seldom leave the Strait of Georgia-Puget Sound-Strait of Juan de Fuca region (Ford et al. 2000). K and L pods are more often found in western Juan de Fuca Strait and off the outer coasts of Washington State and Vancouver Island. Unlike J pod, K and L pods typically leave inshore waters in winter and return in May or June. Their range during this period is poorly known, but they have been sighted as far south as Monterey Bay, California and as far north as Langara Island, off Haida Gwaii (Ford et al. 2000, Black et al. 2001, unpublished data (CRP-DFO).
Northern residents
The northern resident killer whale community comprises three acoustic clans (A, G, and R) containing 34 matrilines, which range from Glacier Bay, Alaska to Grays Harbour, Washington (Ford et al. 2000, unpublished data CRP-DFO). From June to October, they frequent areas from mid Vancouver Island to southeastern Alaska, particularly Johnstone Strait and Queen Charlotte Strait (Figure 1), off northeastern Vancouver Island (Ford et al. 2000). Their range at other times of the year is poorly understood. Small groups of northern residents are sometimes seen in Johnstone Strait and other inshore waters along the BC coast in winter (Ford et al. 2000) but such sightings are rare even when seasonal changes in observer effort are taken into account.
There is no evidence that clans are restricted to specific regions within the range of their community, but some show an apparent preference for particular areas (Ford et al. 2000). For example, the most commonly sighted whales off northeastern Vancouver Island belong to A-clan, whereas most of the whales sighted off the west coast of Vancouver Island belong to G-clan, and R-clan seems to prefer the northern part of the community's range. The range of northern residents overlaps with southern residents and with a community referred to as the southern Alaskan residents. Northern residents have never been seen associating with members of the southern resident community, and while they were observed travelling in proximity to a southern Alaskan resident pod on one occasion (Dahlheim et al. 1997), it is not clear that social mixing took place. Genetic studies have not ruled out the possibility of occasional breeding between the northern resident and southern Alaskan resident communities (Barrett-Lennard and Ellis 2001).
1.3 Population size and trends
1.3.1 Global
Little is known of the historic abundance of killer whales, except that they were "not numerous" (Scammon 1874). Since the early 1970s, photo-identification studies have provided reasonable population estimates for killer whales in the near-shore waters of the northeastern Pacific (Washington, British Columbia, Alaska, and California), and similar work is now underway in several other coastal regions (e.g. the Gulf of California, the Russian Far East, New Zealand, Patagonia, Iceland and Norway). In other areas line transect surveys have been used to provide population estimates. These include the Antarctic (25 000 whales, Branch and Butterworth 2001) and the Eastern Tropical Pacific (85 500 whales, Wade and Gerodette 1993). As such, the worldwide abundance of killer whales is probably between 405 000 and 605 000 whales (Forney and Wade in press). Trend data for killer whales are generally not available, with the exception of resident populations of whales in British Columbia (discussed below) and southern Alaska (population increasing, Craig Matkin, North Gulf Oceanic Society personal communication, November 2005) and for a small population of transients in Prince William Sound (AT1s, currently in decline, not likely to recover, Saulitis et al. 2002).
1.3.2 British Columbia
There are no population estimates for killer whales in British Columbia prior to 1960. Population censuses for killer whales are now conducted annually using photo-identification of individuals. Population trends vary by community and clan. For the purposes of the recovery strategy, data held by the Centre for Whale Research (Centre for Whale Research (CWR)), Friday Harbor, Washington, were used to describe the population status and trends of southern resident killer whales. Data held by the Cetacean Research Program, Fisheries and Oceans Canada (DFO) Nanaimo, BC (CRP-DFO), were used to describe the northern resident killer whale population. Whales are censused slightly differently by each research group.Footnote 3
The southern resident count includes all whales that are seen during a calendar year, and mortalities are included in the count depending on when they take place. For example, a whale that is not seen from March onwards is assumed to be dead. There is less certainty that a whale that is not seen in November or December is dead, and it may be included in the count. In recent years, observer effort has been high and members of the southern resident community are photographed annually, so the count is reasonably precise.
The northern resident count includes all whales that are known to be alive on July 1 of each year. However, not all members of the resident community are seen each year, so the count data are generally less precise than for the southern residents.
In 2003, there were a total of 290 northern and southern resident killer whales (unpublished data, CWR, and CRP-DFO). By comparison there are approximately 220 transient and 200 offshore killer whales, although these numbers are less precise than the resident counts, because not all individuals are encountered each year (Ford et al. 2000).
Southern residents
The size of the southern resident community has been known since the first complete photo-identification census in 1976, and was estimated for the years prior to that (Olesiuk et al. 1990, unpublished data CWR). Figure 2 shows the size of each pod as well as the fluctuation in the total population of the southern resident community from 1974-2003.
Figure 2: Population size and trends for southern resident killer whales from 1974-2003
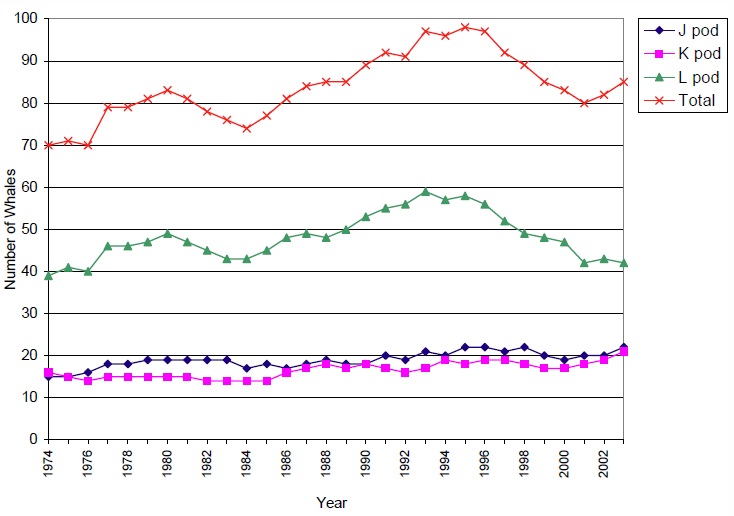
Source: Unpublished data from the Centre for Whale Research
Although the southern resident community was likely increasing in size in the early 1960s, the number of whales in the community dropped dramatically in the late 1960s and early 1970s due to live capture for aquariums (Bigg and Wolman 1975). A total of 47 individuals that are known or likely to have been southern residents were captured and removed from the population (Bigg et al. 1990). The population increased 19% (3.1% per year) from a low of 70 after the live-captures ended in 1973 to 83 whales in 1980, although the growth rate varied by pod (Figure 2). From 1981-1984 the population declined 11% (-2.7% per year) to 74 whales as a result of lower birth rates, higher mortality for adult females and juveniles (Taylor and Plater 2001), and lower numbers of mature animals, especially males, which was caused by selective cropping in previous years (Olesiuk et al. 1990). From 1985 to 1995, the number of southern residents increased by 34% (2.9% per year) to 99 animals. A surge in the number of mature individuals, an increase in births, and a decrease in deaths contributed to the population growth. The latest decline began in 1996, with an extended period of poor survival (Taylor and Plater 2001, Krahn et al. 2002) and low fecundity (Krahn et al. 2004) resulting in a decline of 17% (-2.9% per year) to 81 whales in 2001. Since 2001, the number of southern residents has increased slightly to 85 in 2003Footnote 4 (unpublished data CWR). The growth has been in J and K pods, whereas L pod has continued to decline.
Population viability analyses (PVA) have been used to estimate the extinction risk of southern resident killer whales (Taylor and Plater 2001, and Krahn et al. 2002, 2004). As would be expected, extinction risk increases when the frequency and magnitude of catastrophes such as oil spills and disease epidemics is elevated. The models predict that if the mortality and reproductive rates of the 1990s persist, there is a 6-100 % probability that the population will be extinct within 100 years, and a 68-100% risk that the population will be extinct within 300 years. Extinction of the southern resident population can be regarded as inevitable in these scenarios under the assumptions of the analyses, and catastrophic events simply hasten its demise. However, when the mortality and reproductive rates of the entire 1974-2000 period are used, the risk of the population going extinct declines to 0-55% over 100 years and 2-100% over 300 years.
In addition to analyses focused solely on the southern residents, Krahn et al. (2002) ran simulations assuming that the southern resident population was part of a larger breeding population including northern and southern Alaskan resident killer whales, which greatly decreased its extinction risk. However this scenario does not reflect present evidence that suggests that southern residents are genetically isolated from other populations (Barrett-Lennard 2000; Barrett-Lennard and Ellis 2001).
Northern residents
The northern resident community was likely increasing in size during the early 1960s, but was cropped by the live capture fishery of 1964-1973, during which at least 14 individuals were removed. Twelve of those are known to have been from one pod (A5, Bigg et al. 1990). When first censused in 1974, the northern resident community was estimated to contain approximately 120 whales. Although abundance estimates for northern residents are less precise than those for southern residents, because not all matrilines are seen each year, it appears that the northern community grew steadily during the period 1974 to 1991 (approximately 3.4% per year, Figure 3). The census method used for northern residents is to estimate the population size based on the number of animals that are known to be alive on July 1 of each year. The population increased to a peak of 220 animals in 1997 (growth of 3.0% per year, unpublished data CRP-DFO). Several reasons have been postulated for the northern residents' success relative to southern residents during this period: the population's larger size may have buffered changes in birth and death rates, fewer animals were captured during the live-capture fishery (Olesiuk et al. 1990), and in general they are exposed to less disturbance and environmental contamination. Between 1997 and 2003, the northern resident community declined 7% to 205 whales in 2003 (unpublished data CRP-DFO, Figure 3). As with southern resident killer whales, the cause(s) of the decline are not known. No population viability analysis has yet been conducted for the northern resident killer whales exclusively.
Figure 3: Population Size and Trends for Northern Resident Killer Whales from 1974 to 2003
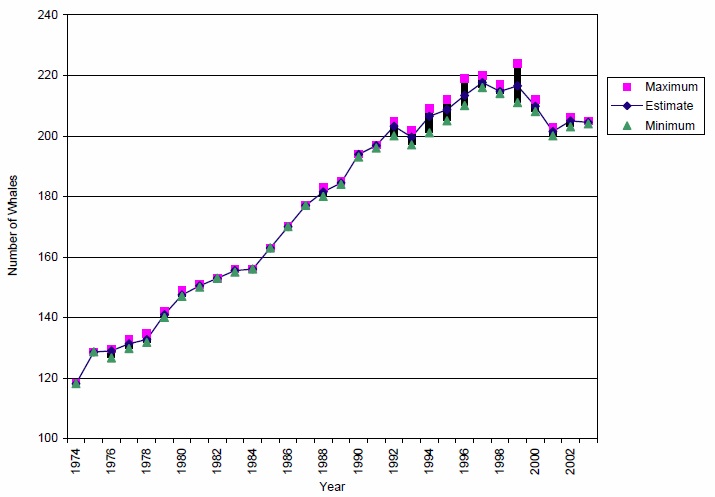
Values reflect the minimum, maximum and estimated number of animals alive as of July 1 in each year. Source: Unpublished data, CRP-DFO, Nanaimo.
1.4 Natural factors affecting population viability and recovery
It is important to appreciate that northern and southern resident killer whales have been studied primarily in protected waters during the months of May to October (Ford et al. 1998, 2000). Their behaviour and ecology in other areas and seasons is poorly known.
1.4.1 Biological limiting factors
The following description of the biology of killer whales is based on data from both the northern and southern resident populations. Essentially, resident killer whales feed on fish and do not switch to marine mammals when their principal prey species are not abundant. They are long-lived animals with no natural predators. On average, females produce a single calf every five to six years during a 25-year reproductive period, and as a result the population has an inherently slow rate of growth. Resident killer whales have strong cultural traditions that influence their association and mating behaviours, which also limits the capacity for the population to grow. More detailed information on the factors that may limit the ability of resident killer whale populations to grow is provided below.
Diet
Although killer whales feed on a wide range of prey species globally, northern and southern resident killer whales are dietary specialists, feeding primarily on fish (Ford et al. 1998). Unlike transient killer whales, resident killer whales do not feed on marine mammals and the breadth of their diet appears to be quite limited. Extensive surface observations and collection of prey fragments from sites of kills by resident whales have shown that these whales forage selectively for certain salmonids regardless of their abundance (Ford and Ellis 2005). Chinook salmon (Oncorhynchus tshawytscha) is the predominant prey species taken by both northern and southern resident communities during May-August, but chum salmon (O. keta) is more prevalent in September-October. Coho salmon (O. kisutch) are taken in low numbers in June-October, but sockeye (O. nerka) and pink (O. gorbuscha) salmon are not significant prey species despite their high seasonal abundance. Non-salmonid fishes do not appear to represent an important component of resident whale diet during May-October.
Resident whales likely forage selectively for chinook salmon over other available salmonids because of the large size, high fat content, and year-round availability of this species in coastal waters (Ford et al. 1998, Ford and Ellis 2005). Killer whales feeding at Langara Island in Haida Gwaii (Queen Charlotte Islands) are known to feed on chinook from stocks returning to rivers as far north as the Skeena River near Prince Rupert and as far south as the Columbia River in Oregon (unpublished data CRP-DFO).
Despite over 30 years of study in British Columbia, only 14 stomachs from resident killer whales have been recovered and examined (Ford et al. 1998, unpublished data CRP-DFO). The extent to which stranded individuals provide accurate insight into the dietary preferences of healthy, free-ranging killer whales is not certain. However, salmon was identified in all seven stomachs that contained prey, including four in which chinook was positively identified. Two contained squid and one also contained bottom fish. It is possible that bottom fish (including ling cod, kelp greenling and sablefish), as well as squid, comprises a significant component of killer whale diet in some areas or during certain times of the year, but more research is needed to determine the year-round diet of killer whales.
It is not known whether resident killer whales depend on specific salmon runs, but their occurrence has been correlated with the abundance of various salmonid species in several past studies (Heimlich-Boran 1986, Nichol and Shackleton 1996, Osborne 1999). The role of these geographical correlations with regard to prey selection is uncertain, since some of these species (sockeye and pink salmon) are not taken in significant numbers compared to chinook salmon (Ford et al. 1998, Ford and Ellis 2005). It is likely that whale occurrence in such areas is driven primarily by the availability of migrating chinook salmon, especially in summer months, and correlations with pink and sockeye salmon are an incidental result of their great abundance during the same period. In fall, the presence of chum salmon appears to influence the movements of resident whales. In Johnstone Strait, chum salmon is the primary prey species taken by northern residents from late September through October (Ford and Ellis 2005). Fall movements of southern resident pods into Puget Sound were roughly correlated with runs of chum salmon, as well as chinook (Osborne 1999). Recent winter sightings of southern resident killer whales in central California were coincident with high local densities of chinook salmon (N. Black, Monterey Bay Whale Watch, unpublished. data).
Social organization
The social structure of killer whales in British Columbia appears to be complex and differs among the three ecotypes (Ford and Ellis 1999, Ford et al. 2000). The social structure of resident killer whales is the best understood, and one of its unique features is that there is no permanent dispersal of either sex from the natal group. The basic social unit of resident killer whales is the matriline, composed of an older female (or matriarch) her male and female offspring, and the offspring of her daughters (Ford et al. 2000). Because matriarchs have long life spans, some matrilines may contain up to four generations. In over three decades of study, immigration and emigration have rarely been observed (Bigg et al. 1990, Ford et al. 2000). Two recent cases of juvenile whales leaving their matrilines and traveling alone are considered to be exceptional, isolated incidents. One, a female calf referred to as A73, or Springer, was separated from her pod shortly after her mother died and was observed alone after a brief period of association with a pod from another clan. She was subsequently reunited with her pod and joined another matriline. The second incident involved a male calf L98, or Luna, who became isolated from his pod and all other killer whales for unknown reasons in 2001. Although individuals do not disperse from their natal group, sisters often begin to spend more and more time apart after their mother dies, and their own matrilines may eventually become socially independent (Bigg et al. 1990, Ford et al. 2000, Ford and Ellis 2002).
Reproductive parameters
Females reach sexual maturity, defined as the age of first successful pregnancy, at 14.9 years on average (range 12-18 years,Olesiuk et al. 1990). Males reach sexual maturity, defined as when the dorsal fin shape changes sufficiently to distinguish males from females, at 15 years on average (range, 10 -17.4 years). Males reach physical maturity (when the dorsal fin reaches its full height) at about 20 years. Genetic paternity testing indicates that males rarely reproduce before 25 years of age (Barrett-Lennard 2000). The gestation period of killer whales is typically 16 to 17 months, one of the longest of all whales (Walker et al. 1988, Duffield et al. 1995). Only single calves are normally born. Only one possible case of twins has been reported (Olesiuk et al. 1990.
Approximately equal number of males and females are born (Dahlheim and Heyning 1999) and newborn calves are between 218 and 257 cm long (Olesiuk et al. 1990). Haenel (1986) estimated that calves are weaned at 1.0-1.5 to two years of age. The interval between calving is usually about 5.2 years for northern residents and 6.2 years for southern residents (unpublished data CRP-DFO). However the interval is highly variable, and ranges from two to 12 years, and increases with age until menopause (Olesiuk et al. 1990). Overall, females have an average of 5.25 viable calves in a 25.2 year reproductive lifespan (Olesiuk et al. 1990). Calving occurs year-round in the northern resident community, but appears to peak between fall and spring. Southern residents do not appear to calve in the summer (unpublished data CWR).
Mating Behaviour
Mating behaviour between male and female killer whales has rarely been observed in the wild. However, genetic evidence has revealed that resident killer whales have a propensity to mate outside their matriline (and clan, in the case of northern residents) but inside their community (Barrett-Lennard 2000, Barrett-Lennard and Ellis 2001). This minimizes the possibility of inbreeding very effectively, but restricts the options for mating if the population becomes very small. For example, in the southern resident community there may be an extreme shortage of sexually mature males, particularly for L pod females, assuming females select mates outside their pod.
Survival and Longevity
Survival of resident killer whales varies with age. Neonate mortality (from birth to six months of age) is high, reported at approximately 43% for all residents (Olesiuk et al. 1990), and 42% for northern residents (Bain 1990). Accordingly, average life expectancy is reported for an animal that survives the first six months, and is estimated to be 50.2 years for females and 29.2 years for males (Olesiuk et al. 1990). Maximum longevity for females is an estimated 80-90 years and for males is 50-60 years (Olesiuk et al. 1990). Although a typical trait in most mammals, the shorter lifespan of males could be related to sexual selection (Baird 2000) or to higher levels of persistent chemicals, such as Polychlorinated Biphenyls (PCBs) (Ross et al. 2000). The bioaccumulation of toxins is discussed in greater detail in Section 2.2.1. Atypical, however, is the prolonged post reproductive period of females, discussed in the following section. Recent evidence suggests that declines in both the northern and southern resident populations (all age and sex classes) can be attributed to an increase in mortality rates (Ford et al. 2005) as well as a decrease in fecundity for southern residents (Krahn et al. 2004). The potential causes of the population declines are discussed in Section 2.
Reproductive Senescence
The average life span of female resident killer whales is approximately 50 years, but on average they produce their last calf at 39, and a significant number live to 70 years or more (Olesiuk et al. 1990). The 'grandmother hypothesis' suggests that the presence of older females in a group can increase the survival of offspring, and this may indeed be true for killer whales (see discussion under Culture below). In any case, when evaluating the status of killer whale populations, it is important to consider the age structure of the population and to note that post-reproductive adult females are no longer able to contribute directly to population growth. In an endangered population of transients in southern Alaska (AT1s), no calves have been born since 1984. Since the remaining females are near or beyond their reproductive years, the population is on the verge of extinction (Saulitis et al. 2002), with virtually no prospect for recovery, even though it may persist for many more years.
Culture
Culture refers to a body of information and behavioural traits that are transmitted within and between generations by social learning. Until recently, culture was generally considered a distinguishing feature of human societies. Of late, the concept of culture has been broadened to include non-human mammals and birds (reviewed in Rendell and Whitehead 2001) and there is strong evidence for it in both northern and southern resident killer whales, and southern Alaskan resident killer whales (Ford 1991, Ford et al. 1998, Barrett-Lennard et al. 2001, Yurk et al. 2002). There is also evidence for culture in other cetaceans, such as sperm whales (Whitehead and Rendell 2004), although not to the same extent as for resident killer whales (Rendell and Whitehead 2001).
Dialects are the best studied form of culture in killer whales. A calf learns its dialect from its mother and other closely related adults, retains it for life, and passes it on to the next generation with few modifications (Ford 1991, Deecke et al. 2000, Miller and Bain 2000). These culturally-transmitted dialects may play an important role in inbreeding avoidance, since females apparently prefer males from dialect groups other than their own (Barrett-Lennard 2000, Yurk et al. 2002). Culture also appears to play an important role in feeding, with dietary preferences and probably foraging techniques and areas passed on culturally (Ford et al. 1998). Culture may also select for longevity in killer whales, as it provides a mechanism for older individuals to increase the fitness of their offspring and relatives by transferring knowledge to them (Barrett-Lennard et al. 2001). In African elephants, older matriarchs are better able to discriminate between threatening and non-threatening disturbances than younger animals, and pass this knowledge on to other members of their group (McComb et al. 2001).
Culture may help animals to learn to adapt to changing environments by allowing them to learn from each other in addition to learning from experience. For example, based on differences in foraging success by sympatric clans of sperm whales under different climatic regimes, Whitehead et al. (2004) suggest that cultural diversity may be even more significant than genetic diversity in helping sperm whales to deal with a changing ocean climate. While we do not know if this is true for resident killer whales, we do know that they respond culturally to anthropogenic changes in their environment. In Alaska, resident killer whales responded to longline fishing in areas of Alaska by learning to raid the gear and take fish, and this behaviour spread rapidly throughout the population (Matkin and Saulitis 1994).
Depensation
Resident killer whale populations are at risk simply by virtue of their low population size. In general, small populations generally have an increased likelihood of inbreeding and lower reproductive rates, which can lead to low genetic variability, reduced resilience against disease and pollution, reduced population fitness, and elevated extinction risks due to catastrophic events. Pacific resident killer whale populations are considered small, at 85 southern residents in 2003Footnote 5 (unpublished data, CWR), and 205 northern residents in 2003 (unpublished data, CRP-DFO). If either resident population continues to decline, they may be faced with a shortage of suitable mates. Among the southern residents, L pod females may be particularly vulnerable to this scenario because of the small number of reproductive males in J and K pod. Even under ideal conditions, the population will recover slowly because killer whales calve relatively infrequently.
Inbreeding appears to be less of a risk for resident killer whales than might be expected based on the small size of their populations. They may avoid inbreeding and its inherent risks through non-random mate selection. Resident killer whales select mates from outside their natal pod, which may make small populations of killer whales more genetically viable than would be expected from population size alone (Barrett-Lennard and Ellis 2001).
Natural Mortality
Killer whales have no recorded predators, other than humans. There are several potential sources of natural mortality that may impact killer whales: entrapment in coastal lagoons or constricted bays, accidental beaching, disease, parasitism, biotoxins, and starvation (Baird 2001). However, it cannot be ruled out that anthropogenic factors may make killer whales more vulnerable to natural sources of mortality. For example, disturbance from intense noise may cause animals to strand (Perrin and Geraci 2002). The proximate cause of death, stranding, is a natural source of mortality, but the death is ultimately human-caused.
1.4.2 Other natural limiting factors
Entrapment and/or accidental beaching
Accidental beaching and entrapment are sometimes a source of mortality for killer whales. At least four mass strandings involving more than 36 individuals occurred in BC in the 1940s (Carl 1946, Pike and MacAskie 1969, Mitchell and Reeves 1988, Cameron 1941). Although the causes of mass strandings in toothed whales are uncertain, disease, parasitism, and disturbance from intense underwater noise have been suggested as possible causes (Perrin and Geraci 2002). Two possible cases of temporary entrapment have been reported for southern resident killer whales (Shore 1995, 1998). In 1991, J-pod spent 11 days in Sechelt Inlet, apparently reluctant to exit through a constricted entrance with tidal rapids. In 1997, nineteen killer whales spent 30 days in Dyes Inlet, Puget Sound, possibly because they were reluctant to pass under a noisy bridge (Shore 1998).
Disease and Parasitism
Diseases in captive killer whales have been well studied, but little is known of diseases in wild killer whales (Gaydos et al. 2004). Causes of mortality for captive killer whales include pneumonia, systemic mycosis, other bacterial infections, and mediastinal abscesses (Greenwood and Taylor 1985). Of 16 pathogens identified in killer whales, three have been detected in wild individuals: marine Brucella, Edwardsiella tarda, and cetacean poxvirus (Gaydos et al. 2004). A severe infection of E. tarda resulted in the death of a southern resident male in 2000 (Ford et al. 2000). Marine Brucella may cause abortions and reduced fecundity in killer whales (Gaydos et al. 2004). Cetacean poxvirus can cause mortality in calves and causes skin lesions (Van Bressem et al. 1999). Twenty-seven additional pathogens have been identified in sympatric odontocetes that may be transmittable to killer whales (Gaydos et al. 2004).
External parasites of killer whales have been reported in Mexico (Black et al. 1997), but none have been observed on killer whales in BC (Baird 2001). Internal parasites of killer whales include various trematodes, cestodes, and nematodes (Heyning and Dahlheim 1988, Raverty and Gaydos 2004). These endoparasites are usually acquired through infected food, but the amount of infection and their contribution to killer whale mortality are not known at this time.
Algal Blooms
Harmful algal blooms (HABs) are blooms of algae that produce biotoxins such as paralytic shellfish poison, domoic acid, saxitoxin and brevitoxin. Such toxins can accumulate in the tissues of species that ingest them and are magnified up the food chain. Mortality of humpback whales (Megaptera novaeangliae) off Massachusetts in 1987 and California sea lions (Zalophus californianus) in California in 1998 have been linked to biotoxin exposure (Geraci et al. 1989, Scholin et al. 2000). Several species of marine mammals have been shown to have a potential susceptibility to the neurotoxic effects of biotoxins (Trainer and Baden 1999). Given the apparent increase in HAB event frequency, and the potential for toxic effects in killer whales, there may be some risk to resident killer whales exposed to biotoxins through HABs, although the risk is thought to be low (Krahn et al. 2002).
Regime Shifts
In the North Pacific, there are widespread changes that occur in the circulation and physical properties of the ocean. These changes take place on decadal time scales and are referred to as 'regime shifts' (see reviews in Francis et al. 1998, Benson and Trites 2002). Such shifts may happen quite quickly, and result in dramatic changes in the distribution and/ or abundance of many species, ranging from zooplankton to fish and possibly marine mammals and seabirds. If the distribution or abundance of resident killer whale prey changed significantly following a regime shift, it is possible that killer whales could be affected.