
Ambystoma unisexué (Ambystoma laterale) et 3 sous-espèces : évaluation et rapport de situation du COSEPAC – 2016

Population dépendante de la salamandre à petite bouche - En voie de disparition
population dépendante de la salamandre de Jefferson - En voie de disparition
population dépendante de la salamandre à points bleus - Non en péril
2016
Table des matières
- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique (UD 1)
- Résumé technique (UD 2)
- Résumé technique (UD 3)
- Préface
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Facteurs limitatifs
- Menaces
- UD 1 : Salamandres unisexuées dépendantes de la salamandre à petite bouche (impact global des menaces calculé = élevé)
- UD 2 : Salamandres unisexuées dépendantes de la salamandre de jefferson (impact global des menaces calculé = très élevé)
- UD 3 : Salamandres unisexuées dépendantes de la salamandre à points bleus (impact global des menaces calculé = faible)
- Nombre de localités
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique du rédacteur du rapport
- Collections examinées
Liste des figures
- Figure 1. Salamandre unisexuée dépendante de la salamandre à petite bouche (Ambystoma laterale - (2) texanum ou LTT) provenant de l'île Pelée (Ontario). Spécimen recueilli sous forme de larve par L. Licht et élevé en laboratoire.
- Figure 2. Salamandre unisexuée dépendante de la salamandre de Jefferson (Ambystoma laterale - (2 )jeffersonianum ou LJJ) provenant de Hilton Falls (Ontario). Spécimen recueilli par K. Bériault et utilisé dans le cadre d'expériences de radiodétection (Bériault, 2005).
- Figure 3. Salamandre unisexuée dépendante de la salamandre à points bleus (Ambystoma(2)laterale - jeffersonianum ou LLJ) provenant du Bois de Saraguay, sur l'île de Montréal (Québec). Spécimen recueilli par Jean-François Desroches.
- Figure 4. Aire de répartition des Ambystoma unisexués qui ont été identifiés au moyen de marqueurs génétiques de 1979 à 2015. Les différents symboles représentent les populations unisexuées qui dépendent de différentes espèces donneuses de sperme qui sont partitionnées en trois unités désignables (figures 5 à 7).
- Figure 5. Aire de répartition mondiale des Ambystoma unisexués dépendants de la salamandre à petite bouche, qui ont besoin de la salamandre à petite bouche (A. texanum) comme espèce donneuse de sperme. Ces populations contiennent des individus qui possèdent au moins une garniture chromosomique de la salamandre à points bleus et au moins une garniture chromosomique de la salamandre à petite bouche (LT et LTT).
- Figure 6. Aire de répartition mondiale des Ambystoma unisexués dépendants de la salamandre de Jefferson, qui ont besoin de la salamandre de Jefferson (A. jeffersonianum) comme espèce donneuse de sperme. Ces populations contiennent des individus qui possèdent normalement une garniture chromosomique de la salamandre à points bleus et deux garnitures chromosomiques de la salamandre de Jefferson (LJJ).
- Figure 7. Aire de répartition mondiale des Ambystoma unisexués dépendants de la salamandre à points bleus, qui ont besoin de la salamandre à points bleus (A. laterale) comme espèce donneuse de sperme. Ces populations contiennent des individus qui possèdent normalement deux garnitures chromosomiques de la salamandre à points bleus et une garniture chromosomique de la salamandre de Jefferson (LLJ).
- Figure 8. Des salamandres sont recueillies dans des pièges à ménés dans l'aire de conservation de Hilton Falls à la fin mars, alors que la glace libère les bords de l'étang. Ce piège contient des salamandres de Jefferson, des salamandres unisexuées dépendantes de la salamandre de Jefferson et des salamandres maculées (A. maculatum).
- Figure 9. Événements hypothétiques pouvant expliquer la baisse et le remplacement génomiques chez les salamandres unisexuées de l'île Pelée. Des salamandres à points bleus (Ambystoma laterale ou LL) et des salamandres à petite bouche (A. texanum ou TT) sont présentes sur l'île. Un individu triploïde unisexué LLT (niveau A) pond des œufs LLT à ploïdie non réduite qui peuvent produire des larves LLT par gynogenèse (le sperme est rejeté). Si le sperme d'un mâle A. laterale est incorporé, cependant, l'individu produit sera un LLLT tétraploïde; si un spermatozoïde d'A. texanum est incorporé, l'individu produit sera un LLTT tétraploïde symétrique (niveau B). Les œufs d'un individu tétraploïde symétrique pourraient être non réduits sur le plan de la ploïdie et se développer par gynogenèse, ou subir une méiose réductionnelle produisant des œufs LT (niveau C). Les œufs LT diploïdes peuvent se développer par gynogenèse ou être fertilisés par du sperme d'A. laterale pour produire des individus LLT, ou par du sperme d'A. texanum pour produire des individus LTT (niveau D). Tous ces biotypes sont présents sur l'île Pelée (tableau 1), et un tel système pourrait expliquer la formation d'individus unisexués diploïdes LT. L'incorporation de sperme hausse la ploïdie au niveau triploïde et explique le remplacement génomique. Un génome L (chez un individu unisexué de niveau A) est remplacé par un nouveau génome L ou T chez les unisexués de niveau D.
Liste des tableaux
- Tableau 1. Fréquences des génomotypes trouvés au sein des sous-populations d'Ambystoma unisexués en Ontario où des échantillons de taille adéquate étaient disponibles pour estimer les distributions des fréquences d'individus bisexués et d'individus unisexués diploïdes ou polyploïdes. Les fréquences sont présentées selon le nombre d'individus de chaque génomotype et le pourcentage dans la sous-population (en parenthèses). Tous les génomotypes unisexués ont au moins une garniture chromosomique de l'A. laterale (L), et au moins une garniture ou un génome de l'A. jeffersonianum (J) ou de l'A. texanum (T). Les individus diploïdes ont 2 garnitures chromosomiques, les triploïdes en ont 3, les tétraploïdes en ont 4 et les pentaploïdes en ont 5.
- Tableau 1a. Salamandres unisexuées dépendantes de la salamandre à petite bouche.
- Tableau 1b. Salamandres unisexuées dépendantes de la salamandre de Jefferson.
- Tableau 1c. Salamandres unisexuées dépendantes de la salamandre à points bleus.
- Tableau 2. Sommaire de l'impact des menaces pesant sur les trois unités désignables de l'Ambystomaunisexué, d'après l'évaluation effectuée le 2 février 2015. Voir les tableaux d'évaluation complets aux annexes 2 à 4.
Liste des annexes
- Annexe 1. Hausse et baisse de la ploïdie
- Annexe 2. Tableau d'évaluation des menaces, y compris les commentaires, pour l'Ambystoma unisexué dépendant de la salamandre à petite bouche (UD 1).
- Annexe 3. Tableau d'évaluation des menaces, y compris les commentaires, pour l'Ambystoma unisexué dépendant de la salamandre de jefferson (UD 2).
- Annexe 4. Tableau d'évaluation des menaces, y compris les commentaires, pour l'Ambystoma unisexué dépendant de la salamandre à points bleus (UD 3).
Information sur le document
COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur l'Ambystoma unisexué (Ambystoma laterale), population dépendante de la salamandre à petite bouche, population dépendante de la salamandre de Jefferson et la population dépendante de la salamandre à points bleus, au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xxiii + 70 p.
Note de production :
Le COSEPAC remercie Jim Bogart d'avoir rédigé le rapport de situation sur l'Ambystoma unisexué au Canada. Le présent rapport a été préparé aux termes d'un marché conclu avec Environnement Canada. La supervision a été assurée par Kristiina Ovaska, coprésidente du Sous-comité de spécialistes des amphibiens et des reptiles du COSEPAC.
Pour obtenir des exemplaires supplémentaires, s'adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Unisexual Ambystoma, Ambystoma laterale, Small-mouthed Salamander dependent population, Jefferson Salamander dependent population and the Blue-spotted Salamander dependent population, in Canada.
Illustration/photo de la couverture :
Ambystoma unisexué - Individu unisexué triploïde dépendant de la salamandre de Jefferson (LJJ) recueilli dans la région d'Halton, en Ontario (10 avril 2013; longueur totale = 150 mm; longueur museau-cloaque = 80 mm). Photo de J.P. Bogart.
COSEPAC sommaire de l'évaluation
Sommaire de l'évaluation - mai 2016
- Nom commun
- Ambystoma unisexué - population dépendante de la salamandre à petite bouche
- Nom scientifique
- Ambystoma laterale - texanum
- Statut
- En voie de disparition
- Justification de la désignation
- Ces salamandres unisexuées inhabituelles ne sont présentes que sur une seule île isolée au Canada (île Pelée dans le lac Érié) et dépendent pour le recrutement d'une espèce donneuse de sperme en voie de disparition, la salamandre à nez court (Ambystoma texanum). De nombreuses menaces pèsent sur la salamandre, ce qui rend son existence précaire. Ces menaces comprennent la prédation et la modification de l'habitat par les dindons sauvages introduits, les activités de drainage qui peuvent causer l'assèchement précoce des étangs de reproduction, la mortalité attribuable à la circulation routière pendant les migrations saisonnières, le développement urbain et les activités récréatives.
- Répartition
- Ontario
- Historique du statut
- Espèce désignée « en voie de disparition » en avril 2016.
Sommaire de l'évaluation - mai 2016
- Nom commun
- Ambystoma unisexué - population dépendante de la salamandre de Jefferson
- Nom scientifique
- Ambystoma laterale - (2) jeffersonianum
- Statut
- En voie de disparition
- Justification de la désignation
- Ces salamandres unisexuées inhabituelles occupent des zones restreintes dans des aires peuplées et fortement modifiées de l'Ontario et dépendent pour le recrutement d'une espèce donneuse de sperme en voie de disparition, la salamandre de Jefferson (Ambystoma jeffersonianum). La salamandre fait face à de nombreuses menaces résultant d'activités humaines, menant à la perte et à la fragmentation de l'habitat, ce qui rend son existence précaire.
- Répartition
- Ontario
- Historique du statut
- Espèce désignée « en voie de disparition » en avril 2016.
Sommaire de l'évaluation - mai 2016
- Nom commun
- Ambystoma unisexué - population dépendante de la salamandre à points bleus
- Nom scientifique
- Ambystoma (2) laterale - jeffersonianum
- Statut
- Non en péril
- Justification de la désignation
- Ces salamandres unisexuées inhabituelles dépendent d'une espèce donneuse de sperme, la salamandre à points bleus (Ambystoma laterale), laquelle a une vaste répartition au Canada. Elle est présente de la Nouvelle-Écosse au Manitoba et des Grands Lacs à la baie James et le nord du Québec. Les salamandres unisexuées qui dépendent de cette espèce ont été identifiées dans des sites dans l'ensemble de l'aire de répartition et existent probablement dans de nombreux autres sites qui n'ont pas été soumis à des analyses génétiques. Bien que des déclins dans le sud-ouest de l'Ontario aient été observés et sont attendus pour la salamandre à points bleus ainsi que les salamandres unisexuées qui en dépendent, les menaces sont localisées et auraient peu d'impact sur l'ensemble de la population canadienne.
- Répartition
- Ontario, Québec, Nouveau-Brunswick, Nouvelle-Écosse
- Historique du statut
- Espèce désignée « non en péril » en avril 2016.
COSEPAC résumé
Ambystoma unisexué
Ambystoma laterale
population dépendante de la salamandre à petite bouche
(Ambystoma laterale - texanum)
population dépendante de la salamandre de Jefferson
(Ambystoma laterale - (2) jeffersonianum)
population dépendante de la salamandre à points bleus
(Ambystoma (2) laterale - jeffersonianum)
Description et importance de l'espèce sauvage
Les populations d'Ambystoma formées uniquement de femelles (individus unisexués) sont membres de la famille des salamandres fouisseuses, soit les Ambystomatidés. Leur morphologie, variable, est déterminée par leurs génomes nucléaires. Les individus unisexués qui possèdent deux garnitures chromosomiques ou plus de la salamandre à points bleus (A. laterale) sont noirs et présentent des mouchetures bleues plus ou moins nombreuses, des membres relativement courts et une tête étroite. Les individus unisexués qui possèdent deux garnitures chromosomiques ou plus de la salamandre de Jefferson (A. jeffersonianum) sont quant à eux plus gros, leur peau est de couleur grise ou brune avec quelques mouchetures bleues, leurs membres sont relativement plus longs et leur tête est plus large. Enfin, les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre à petite bouche (A. texanum) sont gris, leur corps est plus mince et leur tête est étroite.
Les Ambystoma unisexués partagent tous un ADN mitochondrial très similaire, différent de celui des espèces bisexuées. Ces individus ont un système génétique unique et représentent une lignée monophylétique distincte qui est apparue il y a de 3 à 5 millions d'années, ce qui en fait la plus ancienne lignée connue de vertébrés unisexués. Les œufs se forment normalement par gynogenèse. Ce processus exige du sperme, qui provient d'espèces sympatriques bisexuées. Le sperme ne sert qu'à déclencher la formation des œufs et n'est habituellement pas incorporé au développement de l'embryon. Dans de rares cas, le sperme est incorporé et, lorsque l'ADN du sperme est incorporé, la ploïdie des embryons augmente (de triploïde à tétraploïde).
Répartition
Les salamandres unisexuées se trouvent en association avec les espèces bisexuées appropriées, dont les mâles servent de donneurs de sperme. L'aire de répartition géographique des salamandres unisexuées du genre Ambystoma coïncide à peu près avec les forêts décidues et mixtes du nord-est de l'Amérique du Nord, soit de la Nouvelle-Écosse et des États de la Nouvelle-Angleterre à l'Indiana. La limite nord de l'aire de répartition se trouve au Minnesota, dans le centre-nord de l'Ontario et dans le sud du Québec, et la limite sud se trouve au Kentucky. Trois unités désignables sont prises en compte dans le présent rapport, selon les espèces dont les mâles servent de donneurs de sperme. Au Canada, les salamandres unisexuées se trouvent en association avec la salamandre à points bleus en Nouvelle-Écosse, au Nouveau-Brunswick, au Québec et en Ontario, avec la salamandre de Jefferson en Ontario et avec la salamandre à petite bouche sur l'île Pelée, dans le lac Érié, en Ontario. Au Canada, des populations unisexuées de salamandres sont présentes au sein de toutes les populations connues de salamandres de Jefferson et de salamandres à petite bouche, ainsi que dans la majorité des populations de salamandres à points bleus qui ont été examinées. Les salamandres unisexuées peuvent être beaucoup plus nombreuses que les individus des espèces sympatriques bisexuées qui servent de donneurs de sperme.
Habitat
Les salamandres unisexuées ont les mêmes besoins en matière d'habitat que leurs espèces donneuses de sperme respectives. Elles se trouvent normalement dans des forêts décidues ou mixtes qui contiennent des étangs de reproduction convenables ou qui y sont adjacentes. Ces étangs sont habituellement des mares éphémères ou printanières qui s'assèchent à la fin de l'été. L'habitat terrestre se situe dans des boisés humides où les salamandres s'abritent des prédateurs et du dessèchement sous des arbres tombés ou des pierres, ainsi que dans des terriers de mammifères. Les adultes s'alimentent lorsque les conditions sont humides la nuit, sur le tapis forestier, à moins de 1 km de l'étang de reproduction. Ces salamandres ont aussi besoin de sites d'hivernage sous la ligne de gel.
Biologie
En conjonction avec les individus de leurs espèces donneuses de sperme respectives, les adultes unisexués migrent en direction et en provenance des étangs de reproduction la nuit, très tôt au printemps. La plupart de ces épisodes de migration coïncident avec la pluie ou des conditions très humides. Des mâles sympatriques bisexués prennent part à la parade nuptiale et, un jour ou deux après l'accouplement, les salamandres unisexuées pondent plusieurs masses d'œufs sur des brindilles ou de la végétation émergente à diverses profondeurs dans l'étang de reproduction. La ponte peut aussi avoir lieu sous la glace. La durée du développement des œufs et des larves est variable et dépend de la température. Les larves sont carnivores et se nourrissent d'une variété d'invertébrés; elles sont aussi cannibales. Au Canada, les larves se métamorphosent normalement en juillet ou au début août, et quittent l'étang. Les jeunes et les adultes sont entièrement terrestres, sauf durant la période de reproduction annuelle.
Taille et tendances de la population
Il est difficile d'estimer la taille des populations de salamandres unisexuées, car ces dernières sont semblables sur le plan morphologique aux femelles de leurs espèces donneuses de sperme respectives. La plupart des sites historiques qui ont fait l'objet de recherches visant la salamandre de Jefferson en 1990 et 1991 n'accueillaient plus ni de salamandres de Jefferson ni de salamandres unisexuées en 2003 et 2004. En outre, à certains sites où des salamandres de Jefferson et des salamandres unisexuées étaient toujours présentes en 2003-2004, on a constaté une réduction notable du nombre de masses d'œufs par rapport aux quantités observées au cours de relevés précédents. La taille des populations de salamandres unisexuées varie en fonction de l'espèce donneuse de sperme et de la région géographique. Toutes les sous-populations de salamandres de Jefferson et de salamandres à petite bouche contiennent aussi des individus unisexués qui peuvent représenter quelque 85 % des individus à un site donné. Le pourcentage d'individus unisexués trouvés dans les étangs de reproduction de la salamandre à points bleus varie davantage, et certains de ces étangs ne contiennent aucun individu unisexué.
Menaces et facteurs limitatifs
La perte de donneurs de sperme sexués constitue un facteur limitatif unique aux salamandres unisexuées du genre Ambystoma, car celles-ci ont besoin de mâles diploïdes sexués pour se reproduire. Les menaces qui pèsent sur l'espèce sont :l'élimination partielle ou absolue de l'habitat convenable causée par l'aménagement des terres, y compris la perte d'étangs de reproduction, d'arbres et de couverture végétale;les obstacles (routes, clôtures anti-érosion) qui bloquent les voies migratoires reliées aux étangs de migration;l'assèchement prématuré des étangs durant l'été.
Protection, statuts et classements
Les salamandres unisexuées coexistent avec certaines espèces qui ont reçu une cote de conservation, et sont impossibles à distinguer de ces espèces sur le plan morphologique. L'État du Connecticut désigne le « complexe » A. jeffersonianum et le « complexe » A. laterale comme des espèces préoccupantes. En Ontario, les salamandres polyploïdes dominées par la salamandre de Jefferson sont des individus unisexués qui ont besoin de salamandres de Jefferson mâles. Depuis 2010, ces individus reçoivent la même protection sur le plan de l'habitat que la salamandre de Jefferson en vertu de la Loi de 2007 sur les espèces en voie de disparition (LEVD) (voir Règl. de l'Ont. 242/08, art. 28). Jusqu'ici, aucun règlement semblable n'a été adopté pour les salamandres unisexuées du Canada qui vivent avec la salamandre à petite bouche (A. texanum; en voie de disparition) sur l'île Pelée (Ontario) ni pour les salamandres unisexuées qui dépendent de la salamandre à points bleus (A. laterale).
Résumé technique (UD 1)
- Nom scientifique :
- Ambystoma laterale - texanum
- Nom français :
- Ambystoma unisexué (population dépendante de la salamandre à petite bouche)
- Nom anglais :
- Unisexual Ambystoma(Small-mouthed Salamander-dependent population)
- Répartition au Canada (province/territoire/océan ) :
- Ontario (île Pelée, comté d'Essex)
Données démographiques
| Sujet | Information |
|---|---|
| Durée d’une génération | Environ 8 ans (voir Cycle vital et reproduction) |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures? | Oui, il y a un déclin continu inféré et prévu d'après le déclin prévu de la salamandre à petite bouche (voir COSEWIC, 2004, 2014) et d'autres menaces répertoriées. |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. | Inconnu |
| Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Une réduction est prévue et présumée d'après l'évaluation des menaces, qui a révélé que l'impact global des menaces était « élevé » (déclin prévu de 10 à 70 %). |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
Est-ce que les causes du déclin sont
|
|
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Des fluctuations ont lieu parce que le recrutement varie considérablement d'une année à l'autre, mais ces fluctuations ne sont probablement pas extrêmes. |
Information sur la répartition
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 20 km2 |
| Indice de zone d'occupation (IZO) (Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté). | 20 km2, d'après 5 grilles de 2 km de côté superposées sur 4 étangs de reproduction récemment occupés. |
| La population totale est-elle gravement fragmentée c.-à-d. que plus de 50 % de sa zone d'occupation totale se trouvent dans des parcelles d'habitat qui sont a) plus petites que la superficie nécessaire au maintien d'une population viable et b) séparées d'autres parcelles d'habitat par une distance supérieure à la distance de dispersion maximale présumée pour l'espèce? | a) Non; plus de la moitié des sites occupés semblent soutenir des populations viables; b) oui, les étangs de reproduction sont séparés par plus de 1 km, et il est peu probable que les salamandres se dispersent entre eux. |
| Nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) (utilisez une fourchette plausible pour refléter l'incertitude, le cas échéant) |
Quatre localités sont situées sur l'île Pelée. Une perte de deux étangs de reproduction (localités) historiques s'est produite entre 1991 et 2000 (COSEWIC, 2004). |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | Oui, la perte de deux étangs de reproduction historiques a été observée. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | Oui, il y a un déclin inféré de l'IZO fondé sur la perte d'étangs de reproduction entre 1991 et 2000 (COSEWIC, 2004). |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations? | Oui, il y a un déclin inféré puisque chaque étang de reproduction est considéré comme une sous-population. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Oui, il y a une perte inférée d'étangs de reproduction, qui correspondent à des localités, d'après les données dont on dispose sur la salamandre à petite bouche (COSEWIC, 2004, 2014). |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? | Oui, il y a une perte observée et inférée d'étangs de reproduction fondée sur les données dont on dispose sur la salamandre à petite bouche (COSEWIC, 2004, 2014). |
| Y a-t-il des fluctuations extrêmes du nombre de populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
Nombre d'individus matures dans chaque sous-population
| Sous-population (indiquez des fourchettes plausibles) | Nombre d'individus matures |
|---|---|
| Pointe Fish (extrémité sud de l'île Pelée) | Inconnu |
| Étang (milieu de l'île Pelée) | Inconnu |
| Pointe Sheridan (extrémité nord de l'île Pelée) | Inconnu |
| Chemin Stone | Inconnu |
| Total | Inconnu, mais possiblement moins de 1 000 |
Analyse quantitative
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Non effectuée en raison du manque de données |
Menaces (réelles ou imminentes pour les populations ou leur habitat, de l'impact le plus élevé à l'impact le plus faible)
| Sujet | Information |
|---|---|
|
Un calculateur des menaces a-t-il été rempli pour l'espèce, et dans l'affirmative, par qui? Oui, par Leslie Anthony, Jim Bogart, Joe Crowley, Yohann Dubois, Isabelle Gauthier, Bev McBride (Secrétariat du COSEPAC), Kristiina Ovaska, Mary Sabine. |
Immigration de source externe (immigration de l'extérieur du Canada)
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur les plus susceptibles de fournir des individus immigrants au Canada | L'Ambystoma unisexué n'a pas de statut en Ohio ni au Michigan. La salamandre à petite bouche est une espèce menacée au Michigan, principalement en raison de la perte d'habitat convenable. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Non |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Non. Ces salamandres unisexuées dépendent des mâles de la salamandre à petite bouche qui, au Canada, n'existe que sur l'île Pelée, qui est isolée des populations des États-Unis. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Oui |
| Les conditions de la population source se détériorent-elles? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Inconnu |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Non |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Non |
Nature délicate de l'information sur l'espèce
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Oui, car ces salamandres unisexuées vivent avec la salamandre à petite bouche, dont elles dépendent. Il s'agit d'une espèce en voie de disparition, et l'information qui la concerne est de nature délicate. |
Historique du statut
| Sujet | Information |
|---|---|
| COSEPAC | L'espèce n'a pas été antérieurement évaluée. |
Statut et justification de la désignation
| Sujet | Information |
|---|---|
| Statut | En voie de disparition |
| Codes alphanumériques | B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv) |
| Justification de la désignation | Ces salamandres unisexuées inhabituelles ne sont présentes que sur une seule île isolée au Canada (île Pelée dans le lac Érié) et dépendent pour le recrutement d'une espèce donneuse de sperme en voie de disparition, la salamandre à nez court (Ambystoma texanum). De nombreuses menaces pèsent sur la salamandre, ce qui rend son existence précaire. Ces menaces comprennent la prédation et la modification de l'habitat par les dindons sauvages introduits, les activités de drainage qui peuvent causer l'assèchement précoce des étangs de reproduction, la mortalité attribuable à la circulation routière pendant les migrations saisonnières, le développement urbain et les activités récréatives. |
Applicabilité des critères
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Ne correspond pas au critère. L'ampleur des déclins est inconnue. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Correspond aux critères de la catégorie « en voie de disparition », B1ab(i,ii,iii,iv)+2ab(i,ii,iii,iv). La zone d'occurrence et l'IZO sont inférieurs aux seuils de la catégorie « en voie de disparition ». On compte moins de 5 localités (sous-critère a), la zone d'occurrence a été réduite par la perte de 2 sites historiques (b i); la quantité et la qualité de l'habitat sont en baisse à cause des dindons sauvages et d'autres menaces (b iii); un déclin continu des sous-populations (localités) est prévu (b iv), ce qui mènera à une réduction de l'IZO (b ii). |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Ne s'applique pas. Les seuils de la catégorie « en voie de disparition » ne sont pas atteints. |
| Critère D (très petite population totale ou répartition restreinte) | Ne s'applique pas. Les seuils de la catégorie « en voie de disparition » ne sont pas atteints. |
| Critère E (analyse quantitative) | Non effectuée en raison du manque de données. |
Résumé technique (UD 2)
- Nom scientifique :
- Ambystoma laterale - (2) jeffersonianum
- Nom français :
- Ambystoma unisexué (population dépendante de la salamandre de Jefferson)
- Nom anglais :
- Unisexual Ambystoma (Jefferson Salamander-dependent population)
- Répartition au Canada (province/territoire/océan) :
- Ontario
Données démographiques
| Sujet | Information |
|---|---|
| Durée d’une génération | Environ 11 ans (voir Cycle vital et reproduction) |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures? | Oui, un déclin continu est observé, inféré et prévu d'après le déclin de la salamandre de Jefferson et les menaces qui pèsent sur l'habitat |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. | Déclin présumé de plus de 90 % au cours des 3 dernières générations (33 ans) fondé sur les ensembles de données à long terme pour la salamandre de Jefferson. Les dénombrements des masses d'œufs (indice d'abondance) de 1976 à 2006 montrent un déclin de plus de 90 %. |
| Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Réduction prévue d'après l'évaluation des menaces, qui a révélé un impact global des menaces « très élevé » (réduction de 50 à 100 %). |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Déclin présumé de plus de 90 % fondé sur les déclins passés et les déclins prévus dans l'avenir. |
Est-ce que les causes du déclin sont
|
|
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Des fluctuations ont lieu parce que le recrutement varie considérablement d'une année à l'autre, mais ces fluctuations ne sont probablement pas extrêmes. |
Information sur la répartition
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 9 457 km2. En tenant compte des localités historiques et récentes, on a calculé une zone d'occurrence de 24 624 km2. Cependant, ces salamandres dépendent de la salamandre de Jefferson, et la zone d'occurrence la plus récente calculée pour cette espèce est de 9 457 km2. |
| Indice de zone d'occupation (IZO) (Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté). | 188 km2. En tenant compte des localités historiques et récentes et d'une grille à carrés de 2 km de côté, on a calculé un IZO de 728 km2. Le calcul le plus récent pour la salamandre de Jefferson a produit un IZO de 188 km2. |
La population totale est-elle gravement fragmentée c.-à-d. que plus de 50 % de sa zone d'occupation totale se trouvent dans des parcelles d'habitat qui sont
|
|
| Nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) (utilisez une fourchette plausible pour refléter l'incertitude, le cas échéant) |
On estime qu'il y a quelque 30 localités distinctes sur le plan géographique ou écologique pour la salamandre de Jefferson. À chaque localité, un seul événement menaçant pour l'espèce, comme un changement des propriétés hydrologiques ou de l'hydropériode ou diverses activités humaines, peut rapidement toucher tous les individus. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | Oui. Les sous-populations actuelles sont davantage associées à l'escarpement du Niagara. Plusieurs sous-populations historiques à l'est et à l'ouest de l'escarpement ont été perdues, et une réduction de 62 % de la zone d'occurrence a été constatée. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | Oui. Il y a un déclin observé et prévu pour la salamandre de Jefferson, qui aura des répercussions sur les individus unisexués qui dépendent de cette espèce. En comparant les données de 1979 à 2003 et de 2004 à 2015 sur la salamandre de Jefferson, on obtient une baisse du nombre d'étangs de reproduction convenables de 74 % pour la génération la plus récente (2004 à 2015). |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations? | Oui. Il y a des déclins observés et prévus du nombre de sous-populations (qui correspondent aux étangs de reproduction distincts). |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Oui. Il y a un déclin observé et prévu du nombre de localités (qui correspondent aux étangs de reproduction distincts). |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? | Oui. Il y a un déclin observé et prévu de la superficie, de l'étendue et de la qualité de l'habitat. |
| Y a-t-il des fluctuations extrêmes du nombre de populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
Nombre d'individus matures dans chaque sous-population
| Sous-population (indiquez des fourchettes plausibles) | Nombre d'individus matures |
|---|---|
| On estime qu'il y a environ 30 étangs de rétablissement distincts, et chacun représente une sous-population; on s'attend à ce que l'échange génétique entre les sous-populations soit très faible ou nul. | Inconnu, mais probablement < 10 000. Il pourrait y avoir < 2 500 salamandres de Jefferson adultes (COSEWIC, 2010). Les individus unisexués qui dépendent de la salamandre de Jefferson sont plus nombreux que ceux de leur espèce donneuse de sperme et, dans l'ensemble de l'aire de répartition de la salamandre de Jefferson, constituent de 60 à 95 % des sous-populations, avec une moyenne de 80 %, soit < 10 000 individus unisexués adultes. |
| Total | < 10 000 |
Analyse quantitative
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Non effectuée en raison du manque de données |
Menaces (réelles ou imminentes pour les populations ou leur habitat, de l'impact le plus élevé à l'impact le plus faible)
| Sujet | Information |
|---|---|
|
Un calculateur des menaces a-t-il été rempli pour l'espèce, et dans l'affirmative, par qui? Oui, par Leslie Anthony, Jim Bogart, Joe Crowley, Yohann Dubois, Isabelle Gauthier, Bev McBride (secrétariat du COSEPAC), Kristiina Ovaska, Mary Sabine. |
Immigration de source externe (immigration de l'extérieur du Canada)
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur les plus susceptibles de fournir des individus immigrants au Canada | Les salamandres unisexuées n'ont pas de statut dans les États adjacents (Vermont, New York, Michigan et Ohio). L'espèce donneuse de sperme, la salamandre de Jefferson, est une espèce menacée au Vermont, et n'est pas présente au Québec ni au Michigan. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Peu probable, mais possible depuis le Vermont. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Non. L'habitat se limite à des régions du sud-ouest de l'Ontario où une réduction a été constatée. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Oui. La perte et la dégradation de l'habitat ont été constatées dans l'ensemble de l'aire de répartition de la salamandre de Jefferson au Canada. |
| Les conditions de la population source se détériorent-elles? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Inconnu |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Non |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Non |
Nature délicate de l'information sur l'espèce
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Oui, car ces salamandres unisexuées vivent avec la salamandre de Jefferson. Il s'agit d'une espèce en voie de disparition, et l'information qui la concerne est de nature délicate. |
Historique du statut
| Sujet | Information |
|---|---|
| COSEPAC | L'espèce n'a pas été antérieurement évaluée. |
Statut et justification de la désignation
| Sujet | Information |
|---|---|
| Statut | En voie de disparition |
| Codes alphanumériques | A2bc+3c+4bc; B2ab(i,ii,iii,iv,v) |
| Justification de la désignation | Ces salamandres unisexuées inhabituelles occupent des zones restreintes dans des aires peuplées et fortement modifiées de l'Ontario et dépendent pour le recrutement d'une espèce donneuse de sperme en voie de disparition, la salamandre de Jefferson (Ambystoma jeffersonianum). La salamandre fait face à de nombreuses menaces résultant d'activités humaines, menant à la perte et à la fragmentation de l'habitat, ce qui rend son existence précaire. |
Applicabilité des critères
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Correspond au critère de la catégorie « en voie de disparition », A2, car il y a un déclin observé, inféré et présumé de > 50 % du nombre d'individus matures au cours des 3 dernières générations (depuis 1982) fondé sur une baisse de l'indice d'abondance (sous-critère b) et de l'IZO et de la qualité de l'habitat (sous-critère c); correspond aussi au critère A3 en raison de déclins futurs présumés et au critère A4 en raison de déclins semblables présumés, tant dans le futur que dans le passé. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Correspond au critère de la catégorie « en voie de disparition », B2, car l'IZO est inférieur au seuil; correspond au sous-critère a parce que la population est gravement fragmentée; correspond aussi au sous-critère b, car il y a une réduction continue inférée et prévue de la zone d'occurrence (i), de l'indice de zone d'occupation (ii), de l'étendue et/ou de la qualité de l'habitat (iii), du nombre de localités et de sous-populations (iv) et du nombre d'individus matures (v). |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Ne s'applique pas. Les seuils de la catégorie « en voie de disparition » ne sont pas atteints. |
| Critère D (très petite population totale ou répartition restreinte) | Ne s'applique pas. La population n'est ni très petite ni restreinte. |
| Critère E (analyse quantitative) | Non effectuée en raison du manque de données. |
Résumé technique (UD 3)
- Nom scientifique :
- Ambystoma (2) laterale - jeffersonianum
- Nom français :
- Ambystoma unisexué (population dépendante de la salamandre à points bleus)
- Nom anglais :
- Unisexual Ambystoma (Blue-spotted Salamander-dependent population)
- Répartition au Canada (province/territoire/océan) :
- Ontario, Québec, Nouvelle-Écosse, Nouveau-Brunswick
Données démographiques
| Sujet | Information |
|---|---|
| Durée d’une génération Voir Cycle vital et reproduction |
Environ 8 ans (voir Cycle vital et reproduction) |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures? | Oui. Il y a un déclin inféré et prévu. Même si les individus unisexués qui dépendent de la salamandre à points bleus ont une grande aire de répartition et sont abondants au Canada, la perte d'habitat et de milieux humides dans certaines parties de l'aire de répartition de leur espèce donneuse de sperme a donné lieu à un déclin. |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations] | Inconnu |
| Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu. Le calcul des menaces a révélé que l'impact global des menaces était faible (réduction de 0 à 10 %). |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
Est-ce que les causes du déclin sont
|
|
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Des fluctuations ont lieu parce que le recrutement varie considérablement d'une année à l'autre, mais ces fluctuations ne sont probablement pas extrêmes. |
Information sur la répartition
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 671 668 km2 |
| Indice de zone d'occupation (IZO) (Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté). | 1 932 km2 (calculé d'après les localités documentées; la valeur réelle est probablement beaucoup plus élevée) |
| La population totale est-elle gravement fragmentée c.-à-d. que plus de 50 % de sa zone d'occupation totale se trouvent dans des parcelles d'habitat qui sont a) plus petites que la superficie nécessaire au maintien d'une population viable et b) séparées d'autres parcelles d'habitat par une distance supérieure à la distance de dispersion maximale présumée pour l'espèce? | a. Non b. Non |
| Nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) (utilisez une fourchette plausible pour refléter l'incertitude, le cas échéant) |
Très nombreuses; >> 100 localités |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | Oui. Un déclin est inféré dans certaines parties de l'aire de répartition (sud de l'Ontario) d'après la perte d'habitat, mais peu de données sont disponibles. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations? | Oui. Il y a un déclin observé et inféré dans certaines parties de l'aire de répartition de l'espèce dans le sud de l'Ontario et du Québec à proximité des zones urbaines ou dans les endroits où des milieux humides ont été drainés. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Oui. Il y a un déclin observé et inféré dans certaines parties de l'aire de répartition de l'espèce dans le sud-ouest de l'Ontario et le sud du Québec à proximité des zones urbaines ou dans les endroits où des milieux humides ont été drainés. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? | Oui. Il y a un déclin inféré de la superficie, de l'étendue et de la qualité de l'habitat dans certaines parties de l'aire de répartition. |
| Y a-t-il des fluctuations extrêmes du nombre de populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
Nombre d'individus matures dans chaque sous-population
| Sous-population (indiquez des fourchettes plausibles) | Nombre d'individus matures |
|---|---|
| Individus unisexués dépendants de la salamandre à points bleus (Nouvelle-Écosse à Ontario) | Probablement > 1 000 000, mais de grandes parties de l'aire de répartition n'ont pas été examinées. |
| Total | > 1 000 000 |
Analyse quantitative
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Non effectuée |
Menaces (réelles ou imminentes pour les populations ou leur habitat, de l'impact le plus élevé à l'impact le plus faible)
| Sujet | Information |
|---|---|
|
Un calculateur des menaces a-t-il été rempli pour l'espèce, et dans l'affirmative, par qui? Oui, par Leslie Anthony, Jim Bogart, Joe Crowley, Yohann Dubois, Isabelle Gauthier, Bev McBride (secrétariat du COSEPAC), Kristiina Ovaska, Mary Sabine. |
Immigration de source externe (immigration de l'extérieur du Canada)
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur les plus susceptibles de fournir des individus immigrants au Canada | L'Ambystoma unisexué n'a pas de statut dans les États adjacents (Maine, Vermont, New York, Pennsylvanie, Ohio, Michigan, Wisconsin et Minnesota), mais la salamandre à points bleus est une espèce menacée en Ohio. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Non constatée, mais possible |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Oui |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Oui, dans certaines parties de l'aire de répartition, comme dans le sud-ouest de l'Ontario |
| Les conditions de la population source se détériorent-elles? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Probablement. La salamandre à points bleus est menacée en Ontario par la perte de son habitat. |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Non |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Peu probable à une échelle significative |
Nature délicate de l'information sur l'espèce
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Non |
Historique du statut
| Sujet | Information |
|---|---|
| COSEPAC | L'espèce n'a pas été antérieurement évaluée. |
Statut et justification de la désignation
| Sujet | Information |
|---|---|
| Statut | Non en péril |
| Codes alphanumériques | Sans objet |
| Justification de la désignation | Ces salamandres unisexuées inhabituelles dépendent d'une espèce donneuse de sperme, la salamandre à points bleus (Ambystoma laterale), laquelle a une vaste répartition au Canada. Elle est présente de la Nouvelle-Écosse au Manitoba et des Grands Lacs à la baie James et le nord du Québec. Les salamandres unisexuées qui dépendent de cette espèce ont été identifiées dans des sites dans l'ensemble de l'aire de répartition et existent probablement dans de nombreux autres sites qui n'ont pas été soumis à des analyses génétiques. Bien que des déclins dans le sud-ouest de l'Ontario aient été observés et sont attendus pour la salamandre à points bleus ainsi que les salamandres unisexuées qui en dépendent, les menaces sont localisées et auraient peu d'impact sur l'ensemble de la population canadienne. |
Applicabilité des critères
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Ne correspond pas au critère. Bien qu'il y ait un déclin inféré et prévu du nombre d'adultes matures, l'ampleur de ce déclin est inconnue. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Ne correspond pas au critère. La zone d'occurrence et l'IZO se situent vraisemblablement au-delà des seuils. |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Ne correspond pas au critère. La population compte probablement beaucoup plus que 10 000 individus matures. |
| Critère D (très petite population totale ou répartition restreinte) | Ne correspond pas au critère. La population n'est ni très petite ni restreinte. |
| Critère E (analyse quantitative) | Non effectuée en raison du manque de données. |
Préface
L'Ambystoma unisexué est apparu il y a 3 à 5 millions d'années, et partage un ancêtre maternel éloigné avec une population de salamandres des ruisseaux (A. barbouri) du Kentucky (Bi et Bogart, 2010). Dans leur aire de répartition, les Ambystoma unisexués partagent des génomes nucléaires (chromosomes) avec trois différentes espèces bisexuées d'Ambystoma au Canada (la salamandre de Jefferson; la salamandre à points bleus; la salamandre à petite bouche) ainsi qu'avec la salamandre des ruisseaux et la salamandre tigrée de l'Est (A. tigrinum) aux États-Unis. Les populations unisexuées d'Ambystoma comprennent plus de 20 combinaisons génomiques nucléaires (chromosomiques) diploïdes, triploïdes, tétraploïdes et pentaploïdes de deux ou trois de ces cinq espèces (Bogart, 2003; Bogart et al., 2009). Tous les individus unisexués ont au moins un génome nucléaire (garniture chromosomique) de la salamandre à points bleus ainsi qu'un ADN mitochondrial (ADNmt) très similaire qui diffère clairement des séquences mitochondriales des cinq espèces dont les génomes nucléaires peuvent résider dans divers individus unisexués (Hedges et al., 1992; Bogart, 2003). Cela élimine la possibilité que les populations unisexuées d'Ambystoma puissent être apparues à la suite d'épisodes d'hybridation contemporains ou historiques impliquant des femelles des salamandres à points bleus, de Jefferson, à petite bouche ou tigrée de l'Est.
Pour se reproduire, tous les individus unisexués ont besoin de sperme provenant d'une espèce donneuse cooccurrente; dans l'éventualité d'une disparition de cette espèce, les individus unisexués disparaîtraient eux aussi. Dans le cas des taxons qui dépendent obligatoirement d'autres taxons pour accomplir une partie ou l'ensemble de leur cycle vital, des valeurs biologiquement appropriées pour le taxon « hôte » devraient être utilisées à des fins d'évaluation (voir la définition de « population » dans COSEWIC, 2015a). La salamandre à petite bouche (Ambystoma texanum) a été désignée espèce en voie de disparition au Canada par le COSEPAC en 2004 (statut confirmé en 2014), et la salamandre de Jefferson (A. jeffersonianum) a été désignée espèce en voie de disparition au Canada en 2010. La salamandre à points bleus (A. laterale) n'a pas été évaluée par le COSEPAC.
Historique du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Mandat du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Composition du COSEPAC
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Définitions (2016)
- Espèce sauvage
- Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d'animal, de plante ou d'un autre organisme d'origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s'est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
- Disparue (D)
- Espèce sauvage qui n'existe plus.
- Disparue du pays (DP)
- Espèce sauvage qui n'existe plus à l'état sauvage au Canada, mais qui est présente ailleurs.
-
En voie de disparition (VD)
(Remarque : Appelée « espèce disparue du Canada » jusqu'en 2003.) - Espèce sauvage exposée à une disparition de la planète ou à une disparition du pays imminente.
- Menacée (M)
- Espèce sauvage susceptible de devenir en voie de disparition si les facteurs limitants ne sont pas renversés.
-
Préoccupante (P)
(Remarque : Appelée « espèce en danger de disparition » jusqu'en 2000.) - Espèce sauvage qui peut devenir une espèce menacée ou en voie de disparition en raison de l'effet cumulatif de ses caractéristiques biologiques et des menaces reconnues qui pèsent sur elle.
-
Non en péril (NEP)
(Remarque : Appelée « espèce rare » jusqu'en 1990, puis « espèce vulnérable » de 1990 à 1999.) - Espèce sauvage qui a été évaluée et jugée comme ne risquant pas de disparaître étant donné les circonstances actuelles.
-
Données insuffisantes (DI)
(Remarque :Autrefois « aucune catégorie » ou « aucune désignation nécessaire ».) - Une catégorie qui s'applique lorsque l'information disponible est insuffisante (a) pour déterminer l'admissibilité d'une espèce à l'évaluation ou (b) pour permettre une évaluation du risque de disparition de l'espèce.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Description et importance de l'espèce sauvage
Nom et classification
Le statut taxinomique des lignées unisexuées d'Ambystoma fait l'objet de nombreux débats, car ces salamandres ne correspondent à aucun concept d'espèce si ce n'est qu'elles représentent une lignée mitochondriale monophylétique. Premier chercheur à reconnaître le caractère particulier de ces salamandres, Clanton (1934) a discerné, chez des populations du sud du Michigan considérées comme des salamandres de Jefferson (A. jeffersonianum), des individus « sombres » des deux sexes (rapport des sexes de 1:1) et des individus « pâles », qui étaient tous des femelles. D'après les observations de Clanton et les variations morphologiques tenues pour présentes à l'époque chez les salamandres à points bleus (A. laterale) et de Jefferson, Bishop (1947) a considéré que toutes ces salamandres représentaient une seule et même espèce variable - A. jeffersonianum. Au Canada, Logier et Toner (1961) ont par conséquent combiné tous les sites connus de salamandres de Jefferson et de salamandres à points bleus. Minton (1954) a postulé que les formes intermédiaires étaient des hybrides entre l'A. laterale et l'A. jeffersonianum, et Uzzell (1964) a décrit les hybrides commet deux espèces triploïdes qui seraient en fait des clones indépendants capables de se reproduire. D'après les divers synonymes disponibles pour l'A. jeffersonianum, Uzzell a introduit les noms d'espèces suivants : les salamandres unisexuées possédant une série de chromosomes de l'A. laterale et deux séries de chromosomes de l'A. jeffersonianum (un génomotype des individus LJJ) représentaient la salamandre argentée (A. platineum Cope 1867), et celles possédant deux séries de chromosomes de l'A. laterale et une série de chromosomes de l'A. jeffersonianum (LLJ) représentaient la salamandre de Tremblay (A. tremblayi Comeau 1943). Lowcock et al. (1987) ont toutefois démontré que ni l'A. platineum ni l'A. tremblayi ne pouvaient être considérées comme des espèces valides puisque de nombreuses autres variantes chromosomiques existent en plus de ces deux triploïdes réciproques. Une troisième « espèce » unisexuée, la salamandre d'île Kelleys, a été décrite par Kraus (1985a) comme étant l'A. nothagenes de l'île Kelleys, en Ohio. Cette espèce a été décrite comme une Ambystoma laterale - texanum - tigrinum trihybride (LTTi), et l'on pensait aussi qu'il s'agissait d'une lignée évolutive distincte (un autre clone). Bogart et al. (1987) ont cependant montré que l'A. nothagenus n'était pas monophylétique, et qu'il pouvait être recréé par des individus unisexués diploïdes de l'A. laterale - texanum (LT) dans une même période de reproduction par l'incorporation de sperme de l'A. tigrinum provenant de mâles cooccurrents de la salamandre tigrée de l'Est sur l'île Kelleys. Par conséquent, les salamandres unisexuées sont distinctes de toute autre espèce d'Ambystoma, mais elles demeurent à nommer. Lowcock et al. (1987) a donné des noms non officiels aux salamandres unisexuées connues en accord avec le système utilisé par Shultz (1969) pour les poissons unisexués du genre Poeciliopsis. Par exemple, un individu unisexué triploïde possédant un génome de la salamandre à points bleus et deux génomes de la salamandre de Jefferson serait nommé Ambystoma laterale - (2) jeffersonianum et désigné par l'acronyme LJJ.
Les salamandres unisexuées du genre Ambystoma sont admissibles à l'évaluation par le COSEPAC parce qu'elles forment des populations génétiquement distinctes d'origine sauvage et sont présentes au Canada depuis plus de 50 ans. Le COSEPAC exige des noms communs pour toutes les espèces évaluées. Il n'y a aucun nom scientifique ou commun officiel pour les salamandres unisexuées du genre Ambystoma selon Crother (2012), l'autorité scientifique approuvée par le COSEPAC en matière de noms communs d'amphibiens. Les salamandres unisexuées sont mentionnées en conjonction avec l'Ambystoma jeffersonianum, car : [traduction] « La reconnaissance taxinomique soulève des questions complexes concernant la discordance entre les gènes cytoplasmiques et nucléaires, l'évolution réticulée et les permutations du génome » (Crother, 2012, p. 23). Si aucun nom commun n'existe, ces noms doivent être inventés au moyen de protocoles propres au groupe taxinomique examiné (COSEWIC, 2015b). Bien que de nombreuses publications incluent les salamandres unisexuées dans le complexe de la salamandre de Jefferson, certaines populations ne possèdent aucun chromosome de la salamandre de Jefferson. Toutes les salamandres unisexuées ainsi que leurs espèces donneuses de sperme sont incluses dans le genre Ambystoma de la famille des salamandres fouisseuses, soit les Ambystomatidés. Au Canada, le complexe peut être divisé génétiquement et géographiquement comme suit : population dépendante de la salamandre à petite bouche; population dépendante de la salamandre de Jefferson; population dépendante de la salamandre à points bleus.
Description morphologique
Avant de pouvoir recourir à des marqueurs génétiques qui établissent une distinction entre toutes les combinaisons génétiques des individus unisexués (génomotypes), les taxinomistes étaient déconcertés par le « complexe de la salamandre de Jefferson » qui, par le passé, ne comprenait que les salamandres à points bleus, les salamandres de Jefferson et les individus unisexués qui vivaient avec ces deux espèces. Il n'existe pas de morphologie typique des individus unisexués. En fait, leur morphologie reflète le contenu génomique de leur noyau (chromosomes de deux espèces ou plus), et varie tant au sein des populations qu'entre celles-ci. Les salamandres unisexuées tendent à être robustes, de couleur grise à bleu-noir, avec une longueur du museau au cloaque d'environ 80 mm et une queue presque aussi longue que le corps, comprimée latéralement. Dans les étangs de reproduction, les individus unisexués sont plus gros que les femelles de la salamandre à points bleus, mais environ de la même taille que les femelles de la salamandre de Jefferson (Lowcock et al., 1992) et de la salamandre à petite bouche (Licht, 1989). Leurs caractéristiques morphologiques précises sont intermédiaires à celles des espèces dont elles possèdent les chromosomes. Comme toutes les salamandres unisexuées ont au moins une (haploïde) série de chromosomes de la salamandre à points bleus, leurs flancs montrent presque toujours des mouchetures bleues plus ou moins distinctes. Les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre à points bleus sont noirs et présentent divers degrés de mouchetures bleues; ils ont aussi des membres relativement courts et la tête étroite. Les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre de Jefferson sont quant à eux plus gros, leur peau est de couleur grise ou brune avec quelques mouchetures bleues, leurs membres sont relativement plus longs et leur tête est plus large. Enfin, les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre à petite bouche (A. texanum) sont gris, leur corps est plus mince et leur tête est étroite.
Identification génétique
L'Ambystoma « platineum » et l'A. « tremblayi » ont été décrits comme triploïdes par Uzzell (1964) d'après la découverte de 42 chromosomes chez certains individus unisexués du Michigan. Le nombre diploïde de chromosomes (2n) des espèces bisexuées d'Ambystoma est de 28. La détermination de la ploïdie des individus a été la première méthode utilisée pour distinguer les individus bisexués des individus unisexués. Cette détermination peut être accomplie par le dénombrement des chromosomes et/ou l'estimation de la ploïdie par la comparaison de la taille des érythrocytes (Wilbur, 1976; Austin et Bogart, 1982). Plus récemment, la taille du génome (et la ploïdie) a été estimée par cytométrie en flux (Lowcock et al., 1991; Lowcock et Murphy, 1991; Ramsden et al., 2006). Le fait de connaître la ploïdie, ne permet cependant pas de distinguer les différents génomotypes. En outre, les individus unisexués diploïdes, qui ont été les premiers dont l'existence a été reconnue dans le complexe par Bogart et Klemens (1997), seraient « identifiés » incorrectement comme individus appartenant à une espèce bisexuée [voir Réduction de la ploïdie à l'annexe 1]. Les premières tentatives d'identification des chromosomes des salamandres unisexuées comparaient les caryotypes des individus unisexués à ceux de leurs espèces donneuses de sperme mais, avec les techniques classiques, les caryotypes sont trop semblables et ne peuvent pas être utilisés pour l'identification des espèces ou des génomes (Taylor et Bogart, 1990). Grâce aux nouvelles techniques de cytogénétique, les chromosomes de chaque génome d'un individu unisexué peuvent désormais être identifiés par l'application de molécules d'ADN fluorescentes ciblant des chromosomes propres à chaque espèce dans les préparations de chromosomes d'individus unisexués (Bi et Bogart, 2006; Bogart et al., 2009).
Tous les individus unisexués possèdent le même ADN mitochondrial (ADNmt) transmis par la mère, qui est différent des séquences d'ADNmt des autres espèces. Par conséquent, les salamandres unisexuées peuvent être facilement distinguées d'espèces bisexuées sympatriques par le séquençage de gènes mitochondriaux (Hedges et al., 1992; Bogart, 2003; Nöel et al., 2008; Bi et Bogart, 2010) ou par la comparaison des polymorphismes de la longueur des fragments de restriction (RFLP) du génome mitochondrial (Spolski et al., 1992). Des marqueurs génétiques nucléaires doivent ensuite être utilisés pour déterminer la ploïdie et les génomes nucléaires que possèdent les individus unisexués par rapport aux génomes des espèces bisexuées. Par exemple, les salamandres de Jefferson peuvent être facilement distinguées des salamandres à points bleus d'après la présence de différents allèles révélés par électrophorèse (allozymes) à plusieurs loci d'isozymes qui sont homozygotes pour la plupart (p > 0,90), mais qui présentent des mobilités électrophorétiques différentes chez les deux espèces. Les salamandres unisexuées qui vivent avec les salamandres de Jefferson ou les salamandres à points bleus ont des allozymes des deux espèces, et l'intensité de la coloration, ou le dosage, des allozymes renseigne sur le nombre de chromosomes de salamandre de Jefferson ou de salamandre à points bleus qui sont présents (Bogart, 1982; Bogart et Klemens, 1997, 2008). Les isozymes ont aussi été utilisées pour identifier les génomotypes des individus unisexués qui possèdent des jeux de chromosomes de la salamandre à petite bouche et/ou de la salamandre tigrée de l'Est (Bogart et al., 1985, 1987).
Julian et al. (2003) ont élaboré des amorces pour plusieurs loci d'ADN microsatellite de la salamandre de Jefferson et fourni des fourchettes de tailles de fragments pour les loci qui pourraient aussi être amplifiés chez la salamandre à points bleus. Les amorces utilisées pour amplifier les microsatellites génomiques nucléaires à plusieurs loci amplifient des fragments de tailles distinctes (non chevauchantes) chez les deux espèces, et les fragments sont assez variables pour déterminer le nombre de chromosomes chez un individu polyploïde (Ramsden et al., 2006). Avec ces amorces mises au point pour la salamandre de Jefferson et d'autres élaborées pour les microsatellites de la salamandre à petite bouche (Williams et DeWoody, 2004), il est possible d'identifier les salamandres de Jefferson, à points bleus, à petite bouche et tigrée de l'Est, ainsi que tous les génomotypes unisexués en examinant l'ADN extrait d'un petit bout de queue ou d'un doigt; il n'est donc pas nécessaire de sacrifier des individus pour l'analyse des isozymes (Bogart et al., 2007, 2009). Plus récemment, Greenwald et Gibbs (2012) ont montré que les polymorphismes mononucléotidiques (SNP) pouvaient aussi servir à identifier les génomotypes bisexués et unisexués d'après l'ADN extrait.
Structure spatiale et variabilité de la population
Pour se reproduire, toutes les salamandres unisexuées au Canada ont besoin de sperme provenant de mâles cooccurrents de la salamandre à points bleus, de Jefferson ou à petite bouche. Certaines salamandres unisexuées aux États-Unis peuvent aussi utiliser le sperme de salamandres tigrées de l'Est (Bogart et al., 1987) ou de la salamandre des ruisseaux (Bogart et al. 2009). Comme les Ambystomatidés (toutes les espèces de la famille Ambystomatidae) sont difficiles à trouver dans les milieux terrestres, les relevés ciblent des étangs de reproduction où les adultes se concentrent pendant une courte période au printemps. Les salamandres unisexuées ont été définies comme des parasites sexuels (Uzzell, 1964; Bogart, 2003), parce qu'elles s'accouplent avec et obtiennent le sperme de leurs hôtes bisexués. Dans les étangs de reproduction, les salamandres unisexuées sont habituellement plus nombreuses que leurs hôtes sexuels. Comme les individus unisexués se reproduisent généralement par gynogénétique, tous leurs descendants auraient le même génotype que leur mère et seraient membres du même clone (voir l'annexe 1). La variabilité intrapopulation des individus unisexués a été documentée, toutefois, au moyen d'analyses des microsatellites de génotypes multilocus (MLG, pour Multi-Locus Genotype), qui permet d'identifier un même clone génétique au sein des populations (Bogart et al., 2007; Ramsden, 2008; Nöel et al., 2011). De nouveaux clones unisexués dans un étang de reproduction peuvent provenir de l'immigration, d'une mutation ou d'un rare cas de hausse ou de remplacement génomique (voir l'annexe 1). Par conséquent, le nombre de MLG chez les salamandres unisexuées varie au sein des étangs de reproduction et entre eux, mais il est toujours beaucoup plus faible que le nombre de MLG chez les individus bisexués, qui ont chacun leur propre MLG.
Unités désignables
Dans l'ensemble de leur aire de répartition, les salamandres unisexuées ont un ADNmt très similaire qui diffère nettement des séquences d'ADNmt de tous les autres Ambystomatidés (Hedges et al., 1992; Bogart, 2003; Bogart et al., 2007; Bi et Bogart, 2010). Par conséquent, les salamandres unisexuées ne peuvent pas être considérées comme des hybrides ayant des liens avec les femelles d'une autre espèce contemporaine. Elles ont cependant des affinités chromosomiques avec leurs espèces donneuses de sperme (Bi et Bogart, 2006; Bogart et al., 2009), qui peuvent être utilisées pour distinguer trois unités désignables (UD) pour l'Ambystoma unisexué au Canada. Ces unités correspondent génétiquement et géographiquement aux populations de salamandres à petite bouche (Ambystoma texanum), de salamandres de Jefferson (A. jeffersonianum) et de salamandres à points bleus (A. laterale) [voir Nom et classification]. Au Canada, l'Ambystoma unisexué est réparti comme suit : une population dépendante de la salamandre à petite bouche (UD 1, figure 1); une population dépendante de la salamandre de Jefferson (UD 2, figure 2); une population dépendante de la salamandre à points bleus (UD 3, figure 3). Toutes les salamandres unisexuées possèdent au moins une garniture chromosomique de la salamandre à points bleus, mais les autres garnitures chromosomiques proviennent de l'une des deux autres espèces.

Description longue de la figure 1
Photo d'une salamandre unisexuée dépendante de la salamandre à petite bouche (Ambystoma laterale - (2) texanum ou LTT) provenant de l'île Pelée (Ontario). Par rapport aux autres salamandres unisexuées, les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre à petite bouche (A. texanum) sont plus gris, leur corps est plus mince et leur tête est plus étroite.

Description longue de la figure 2
Photo d'une salamandre unisexuée dépendante de la salamandre de Jefferson (Ambystoma laterale - (2) jeffersonianum ou LJJ) provenant de Hilton Falls (Ontario). Les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre de Jefferson sont de couleur grise ou brune avec quelques mouchetures bleues. Ils sont plus gros que les autres salamandres unisexuées, leurs membres sont relativement plus longs et leur tête est plus large.

Description longue de la figure 3
Photo d'une salamandre unisexuée dépendante de la salamandre à points bleus (Ambystoma (2) laterale - jeffersonianum ou LLJ) provenant du Bois de Saraguay, sur l'île de Montréal (Québec). Les individus unisexués qui possèdent au moins deux garnitures chromosomiques de la salamandre à points bleus sont noirs et présentent divers degrés de mouchetures bleues; ils ont aussi des membres relativement courts et la tête étroite.
La reconnaissance des UD (voir la définition d'une « unité désignable » dans COSEWIC, 2015a [version française]) exige que les UD satisfassent aux critères de caractère distinct et d'importance. Les trois UD pour les salamandres unisexuées sont distinctes et importantes d'après les caractéristiques génétiques qui reflètent la profonde divergence phylogénétique intraspécifique et les différentes garnitures chromosomiques. Chaque UD présente des adaptations et une écologie qui sont semblables à celles de son espèce donneuse de sperme cooccurrente. Chaque UD est importante parce qu'elle présente des différences quantitatives sur le plan des allèles communs, et serait amenée à disparaître de concert avec la disparition de son espèce donneuse de sperme sympatrique.
Bien que les individus triploïdes soient dominants, la plupart des populations contiennent aussi des salamandres unisexuées tétraploïdes [voir Hausse de la ploïdie à l'annexe 1]. Tant les individus triploïdes que tétraploïdes sont inclus dans leurs UD respectives, selon l'espèce donneuse de sperme. Les individus unisexués diploïdes sont rares et, lorsqu'on les trouve, ils vivent habituellement dans les mêmes populations que les individus triploïdes (Lowcock, 1991; Bogart et Klemens, 1997, 2008; Bogart, 2003; Bogart et al., 2007). Les salamandres unisexuées diploïdes peuvent être incluses dans une UD par leur association avec une espèce donneuse de sperme ou si elles sont trouvées avec leurs congénères triploïdes respectives. Les 36 individus échantillonnés dans une population à Mont-Saint-Hilaire, dans le sud du Québec, étaient tous des unisexués LJ diploïdes (Nöel et al., 2011). Aucun individu donneur de sperme ou triploïde n'a été trouvé dans la population. On s'attend à ce que la salamandre à points bleus soit l'espèce donneuse de sperme au Québec et, en attendant d'autres activités de recherche dans les populations voisines, cette population LJ diploïde serait incluse dans l'UD dépendante de la salamandre à points bleus.
L'île Pelée présente une situation unique parce qu'elle accueille deux espèces donneuses de sperme (les salamandres à petite bouche et à points bleus) ainsi que des salamandres unisexuées diploïdes, triploïdes et tétraploïdes (Bogart et al., 1985) (tableau 1). Il s'agit de la seule localité canadienne pour la population dépendante de la salamandre à petite bouche qui comprend des individus unisexués LT, LTT et LTTT vivant en sympatrie avec la salamandre à petite bouche (A. texanum). D'autres salamandres unisexuées (LT, LLT et LLLT) se trouvent dans le même étang que la salamandre à points bleus sur l'île Pelée mais, contrairement à l'UD dépendante de la salamandre à points bleus du continent, qui possède un chromosome de la salamandre de Jefferson, toutes les salamandres unisexuées de l'île Pelée possèdent au moins une garniture chromosomique de la salamandre à petite bouche, et aucun chromosome de la salamandre de Jefferson. L'île Pelée compte aussi une fréquence relativement élevée d'individus unisexués diploïdes (LT) et tétraploïdes symétriques (LLLT) par rapport à ce que l'on trouve ailleurs dans l'aire de répartition géographique des salamandres unisexuées (voir le tableau 1). Bogart et Bi (2013) ont formulé l'hypothèse que les individus unisexués interagissaient avec les salamandres à petite bouche et à points bleus sur l'île [voir Réduction de la ploïdie et remplacement génomique à l'annexe 1] et qu'aucune distinction ni aucun partitionnement génétiques ne pouvaient s'appliquer aux salamandres unisexuées de l'île Pelée. L'inclusion de l'ensemble des salamandres unisexuées de l'île Pelée dans une seule et même UD dépendante de la salamandre à petite bouche concorde avec les données disponibles sur les Ambystoma des îles du lac Érié (Downs, 1978; Bogart et al., 1985, 1987; Bogart et Licht, 1986). Bien que des individus unisexués dépendants de la salamandre à petite bouche existent aussi dans les régions continentales de l'Ohio et dans l'extrême sud du Michigan (Downs, 1978; Kraus, 1985b), l'UD dépendante de la salamandre à petite bouche au Canada est distincte parce qu'elle est depuis longtemps isolée sur l'île Pelée.
Importance de l'espèce
La première espèce de vertébré unisexuée, le molly amazone (Poecilia formosa) du Mexique et du sud du Texas, a été décrite par Hubbs et Hubbs en 1932 (ils ont nommé ce poisson en l'honneur de la tribu légendaire de femmes guerrières, et non en référence à l'emplacement géographique). Depuis, des taxons unisexués ont été découverts chez divers genres de poissons, d'amphibiens et de reptiles de cinq continents (Dawley et Bogart, 1989; Vrijenhoek et al., 1989) mais, collectivement, les individus unisexués ne constituent qu'environ 0,1 % de toutes les espèces existantes de vertébrés (Avise, 2008). Même parmi les vertébrés unisexués, les Ambystoma unisexués sont importants sur le plan de l'évolution. Ils représentent la plus ancienne lignée connue de vertébrés unisexués (Hedges et al., 1992; Bi et Bogart, 2010) et ont un système de reproduction unique qui est décrit comme cleptogénétique (Bogart et al., 2007); les Ambystoma unisexués peuvent aussi se reproduire gynogénétiquement, ce qui semble être la norme. D'autres vertébrés unisexués ont une histoire évolutive plus récente, et ils sont : parthénogénétiques, c'est-à-dire qu'ils n'ont pas besoin de sperme pour se reproduire (p. ex. plusieurs espèces de lézards; Vrijenhoek et al., 1989); gynogénétiques, auquel cas du sperme est nécessaire, mais seulement pour déclencher le développement des œufs (p. ex. le molly amazone); hybridogénétiques, ce qui signifie que le génome paternel est éliminé durant la méiose et remplacé par un nouveau génome mâle à chaque génération (p. ex. les poissons du genre Poeciliopsis; Schultz, 1977).
Répartition
Aire de répartition mondiale
L'aire de répartition de l'Ambystoma unisexué s'étend vers le nord jusque dans le centre du Québec, le centre de l'Ontario et le nord du Minnesota, et vers le sud, jusque dans le New Jersey et le nord du Kentucky. D'est en ouest, ces salamandres unisexuées sont présentes depuis la Nouvelle-Écosse jusqu'en Indiana et au Minnesota (figure 4). Il est très probable que la répartition des salamandres unisexuées à la figure 4 soit sous-estimée, parce que des analyses génétiques sont nécessaires pour établir une distinction entre les individus unisexués et les espèces donneuses de sperme, et de nombreuses populations n'ont pas encore été soumises à de telles analyses. La concentration des populations qui ont été examinées dans le sud de l'Ontario et dans le sud des États de New York et du New Jersey (figure 4) est un artéfact d'une série d'activités de collecte visant la salamandre de Jefferson (en voie de disparition) en Ontario et deux espèces donneuses de sperme préoccupantes sur le plan de la conservation au New Jersey (A. laterale [S1] et A. jeffersonianum [S3]). Les populations qui semblent isolées dans la figure 4 résultent de la collecte fortuite d'un nombre relativement faible d'individus qui ont fait l'objet d'une analyse génétique (voir Activités de recherche).
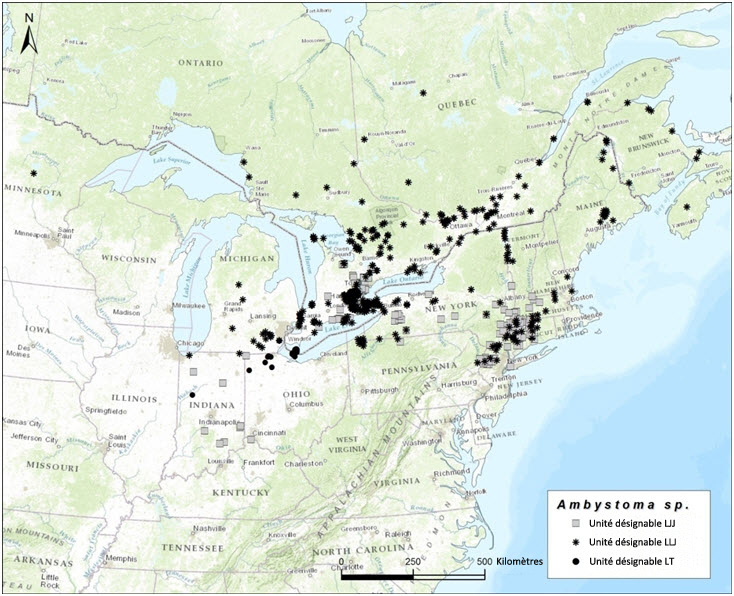
Description longue de la figure 4
Carte de l'aire de répartition mondiale des Ambystoma unisexués identifiés au moyen de marqueurs génétiques de 1979 à 2015. Différents symboles indiquent les trois unités désignables (LJJ, LLJ et LT). L'aire de répartition s'étend vers le nord jusque dans le centre du Québec, le centre de l'Ontario et le nord du Minnesota, et vers le sud, jusque dans le New Jersey et le nord du Kentucky. D'est en ouest, ces salamandres unisexuées sont présentes depuis la Nouvelle-Écosse jusqu'en Indiana et au Minnesota.
Aire de répartition canadienne
Des Ambystoma unisexués ont été trouvés de la Nouvelle-Écosse au lac Supérieur, en Ontario (figure 4). Les aires de répartition des salamandres unisexuées qui dépendent de la salamandre à petite bouche (figure 5) ou de la salamandre de Jefferson (figure 6) se limitent au sud de l'Ontario et sont relativement bien documentées (voir Activités de recherche). La salamandre à points bleus est largement répartie dans l'est du Canada (Cook, 1984), et comparativement peu de populations ont fait d'objet d'analyses visant à déceler la présence d'individus unisexués; la répartition des salamandres unisexuées dépendantes de la salamandre à points bleus (figure 7) n'est donc pas bien comprise.
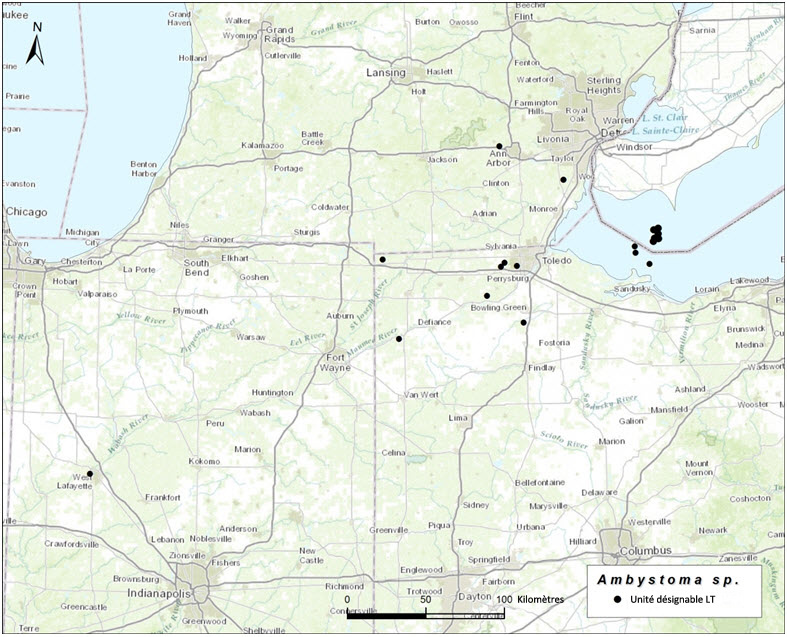
Description longue de la figure 5
Carte de l'aire de répartition mondiale de l'Ambystoma unisexué dépendant de la salamandre à petite bouche, qui a besoin de la salamandre à petite bouche (A. texanum) comme espèce donneuse de sperme. Ces populations contiennent des individus qui possèdent au moins une garniture chromosomique de la salamandre à points bleus et au moins une garniture chromosomique de la salamandre à petite bouche (soit LT et LTT). Au Canada, l'aire de répartition de ces salamandres unisexuées se limite au sud de l'Ontario.
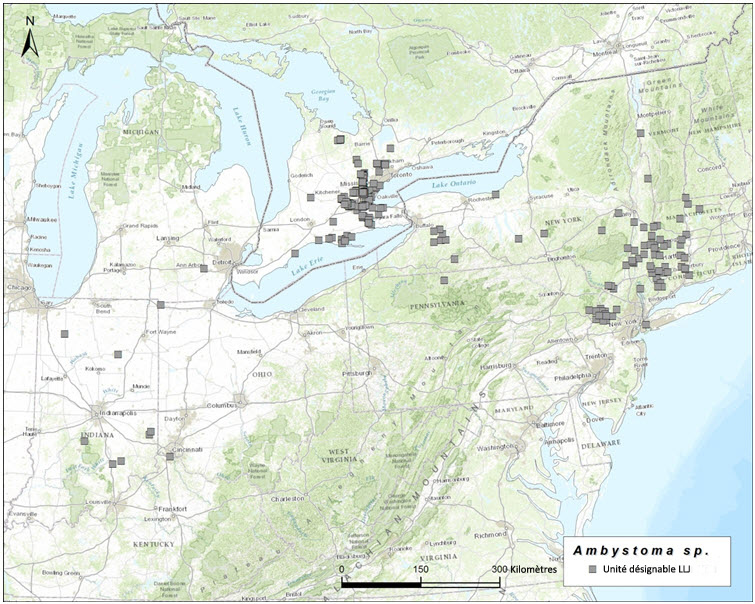
Description longue de la figure 6
Carte de l'aire de répartition mondiale de l'Ambystoma unisexué dépendant de la salamandre de Jefferson, qui a besoin de la salamandre de Jefferson (A. jeffersonianum) comme espèce donneuse de sperme. Ces populations contiennent des individus qui possèdent normalement une garniture chromosomique de la salamandre à points bleus et deux garnitures chromosomiques de la salamandre de Jefferson (soit LJJ). Au Canada, l'aire de répartition de ces salamandres unisexuées se limite au sud de l'Ontario.
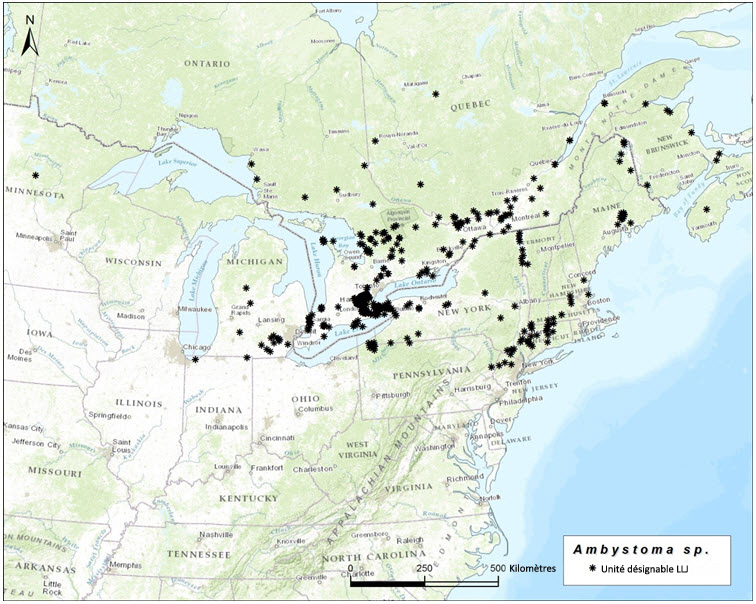
Description longue de la figure 7
Carte de l'aire de répartition mondiale de l'Ambystoma unisexué dépendant de la salamandre à points bleus, qui a besoin de la salamandre à points bleus (A. laterale) comme espèce donneuse de sperme. Ces populations contiennent des individus qui possèdent normalement deux garnitures chromosomiques de la salamandre à points bleus et une garniture chromosomique de la salamandre de Jefferson (soit LLJ). La salamandre à points bleus est largement répartie dans l'est du Canada, mais comparativement peu de populations ont fait d'objet d'analyses visant à déceler la présence d'individus unisexués; la répartition des salamandres unisexuées dépendantes de la salamandre à points bleus n'est donc pas bien comprise.
Zone d'occurrence et zone d'occupation
Les salamandres unisexuées ont besoin d'un donneur de sperme pour se reproduire, et des individus unisexués sont présents dans tous les étangs du Canada où se reproduisent des salamandres à petite bouche ou des salamandres de Jefferson. L'Ambystoma unisexué a toutefois été trouvé dans des étangs qui n'abritaient aucun donneur de sperme (COSEWIC, 2010) (p. ex. site 2 de Kitchener, tableau 1). Pour les espèces sauvages qui dépendent d'autres espèces, l'utilisation de valeurs biologiquement appropriées pour le taxon hôte est recommandée (annexe C dans COSEWIC, 2015); pour l'Ambystoma unisexué, les taxons hôtes sont les espèces donneuses de sperme. On a calculé que la salamandre à petite bouche (A. texanum) avait une zone d'occurrence de 43 km2 et un indice de zone d'occupation (IZO) de 12 km2 (COSEWIC, 2014). Cette estimation de la zone d'occurrence est fondée sur la superficie de l'île Pelée, qui est d'environ 43 km2, et l'IZO a été basé sur la superposition de mailles de grille à 2 km de côté sur les sites où des salamandres sont présentes sur l'île (COSEWIC, 2014). Selon la méthode du plus petit polygone convexe, qui a éliminé deux sites historiques, une zone d'occurrence de 20 km2 a été calculée par le Secrétariat du COSEPAC pour les salamandres unisexuées dépendantes de la salamandre à petite bouche. Comme l'IZO ne peut pas être plus grand que la zone d'occurrence, l'IZO doit aussi être établi à 20 km2.
La zone d'occurrence et l'IZO de la salamandre de Jefferson sont, respectivement, de 6 913 km2 et de 196 km2 (COSEWIC, 2010). La zone d'occurrence et l'IZO des salamandres unisexuées dépendantes de la salamandre de Jefferson ont quant à elles été établies à 24 626 km2 et à 748 km2, respectivement, d'après des données historiques et récentes. Les valeurs de la salamandre de Jefferson et des salamandres unisexuées dépendantes de la salamandre de Jefferson devraient être les mêmes puisque les individus unisexués ont besoin de la présence de salamandres de Jefferson pour se reproduire. Il pourrait exister un certain biais dans l'échantillonnage, parce qu'il est plus facile de trouver des individus unisexués, plus abondants, et parce que ces salamandres vivent longtemps (environ 30 ans) (voir Biologie - Cycle vital et reproduction) et peuvent exister dans des étangs où les chances de recrutement sont nulles. Étant donné que les individus unisexués dépendants de la salamandre de Jefferson ont besoin de salamandres de Jefferson pour se reproduire, on considère que la zone d'occurrence et l'IZO des individus unisexués dépendants de la salamandre de Jefferson sont les mêmes que ceux qui ont été calculés d'après les plus récentes données pour la salamandre de Jefferson. Les nouveaux calculs pour la salamandre de Jefferson, fondés sur les données les plus récentes (2004 à 2015), produisent des estimations de 9 457 km2 pour la zone d'occurrence et de 188 km2 pour l'IZO.
La salamandre à points bleus est largement répartie au Canada, et relativement peu d'étangs ont fait l'objet de recherches visant des salamandres unisexuées dépendantes de la salamandre à points bleus. Des individus unisexués ont toutefois été trouvés dans des populations de salamandres à points bleus dans l'ensemble de l'aire de répartition de l'espèce, ce qui a permis d'établir la zone d'occurrence à 671 668 km2. Au moyen de grilles à mailles de 2 km de côté, l'IZO a été calculé à 1 932 km2. Il est peu probable que les individus unisexués n'existent que dans les populations éloignées et isolées (p. ex. les populations du nord de l'Ontario et du Québec). Par conséquent, l'IZO doit être considéré comme une sous-estimation, et la zone d'occurrence connue pourrait aussi augmenter au fur et à mesure de l'obtention de nouvelles données.
Activités de recherche
Les salamandres du complexe bisexué-unisexué ne sont pas faciles à distinguer les unes des autres sans analyse génétique. L'élaboration de méthodes d'identification génétique de ces salamandres a été entreprise à l'Université de Guelph à la fin des années 1970. Depuis, les biologistes des États-Unis et du Canada envoient des spécimens à Guelph à des fins de confirmation génétique. Au départ, les identifications étaient faites par simple curiosité; d'autres collectes, nécessaires cette fois, ont plus tard été motivées par des préoccupations de conservation dans des régions où une espèce donneuse de sperme s'était vu attribuer une cote de conservation. Les salamandres unisexuées ont initialement été considérées comme de rares « individus hybrides ». Les séquences de l'ADN mitochondrial ont cependant révélé que les salamandres unisexuées n'étaient pas des hybrides (Hedges et al., 1992), et il a été constaté que les individus unisexués étaient plus communs et plus nombreux que les individus de l'espèce bisexuée avec laquelle ils vivent en sympatrie dans la majeure partie de leur aire de répartition (Bogart et Klemens, 1997). En fait, très peu de populations observées étaient exemptes d'individus unisexués (p. ex., Long Island, New York [Bogart et Klemens, 1997]; Île-du-Prince-Édouard [Lowcock, 1989]; populations du centre de la Pennsylvanie et de l'ouest de l'État de New York [Bogart et Klemens, 2008]).
Au Canada, les individus unisexués sont plus nombreux que les individus bisexués dans les populations de salamandres à petite bouche et de salamandres de Jefferson (tableau 1); c'est pourquoi des relevés aléatoires dans les étangs de reproduction suffisent habituellement à confirmer la présence d'individus unisexués. L'échantillonnage de salamandres unisexuées dépendantes de la salamandre à petite bouche sur l'île Pelée a principalement été effectué de 1983 à 1995 par Lawrence Licht (Ph. D.) (voir COSEWIC, 2004) (tableau 1). La majeure partie des activités de recherche en Ontario a été étendue de manière à mieux comprendre l'aire de répartition et l'habitat de la salamandre de Jefferson et des individus unisexués qui lui sont associés parce que les promoteurs et les entreprises d'extraction agrégats avaient besoin de connaître les sites abritant la salamandre de Jefferson, espèce inscrite à la liste de la Loi sur les espèces en péril (LEP) depuis 2003. Les biologistes employés par des entreprises d'experts-conseils (p. ex. Ecoservices, LGL, NRSI) et des biologistes du ministère des Richesses naturelles et des Forêts de l'Ontario (MRNFO) acheminent des échantillons de tissus à l'Université de Guelph aux fins de l'analyse génétique des individus qu'ils ont échantillonnés. Même si les experts-conseils et le MRNFO considèrent les identifications et les sites comme de l'information délicate, les chercheurs et l'équipe de rétablissement de la salamandre de Jefferson sont autorisés à les utiliser. Ces données sont importantes pour les évaluations de la situation de l'espèce par le COSEPAC, et sont incluses au tableau 1 à titre de données inédites, ainsi que dans les calculs et les cartes de la zone d'occurrence et de l'IZO. Beaucoup moins d'efforts ont été consacrés aux relevés visant la salamandre à points bleus et les individus unisexués qui lui sont associés. En Ontario, les experts-conseils et les membres de l'équipe de rétablissement de la salamandre de Jefferson recueillent des échantillons dans des zones situées à proximité de l'aire de répartition de la salamandre de Jefferson pour documenter l'« absence » de salamandres de Jefferson (p. ex. population de Cambridge au tableau 1). La densité des salamandres unisexuées qui coexistent avec les salamandres à points bleus est plus variable dans l'ensemble de l'aire de répartition. En effet, des ratios élevés entre individus unisexués et bisexués sont constatés au sein de certaines populations du sud de l'Ontario (tableau 1) et du sud du Québec (Nöel et al., 2011). Dans le centre de l'Ontario, Lowcock et al. (1991) ont observé une diminution de la proportion d'individus unisexués dans les populations de salamandres à points bleus, diminution qui va du sud au nord. L'échantillonnage de la même population dans le parc provincial Algonquin n'a révélé la présence que de salamandres à points bleus en 2010, mais un faible nombre d'individus unisexués a été trouvé en 2011 (tableau 1).
Par conséquent, par rapport aux aires de répartition connues des salamandres unisexuées dépendantes de la salamandre à petite bouche et de la salamandre de Jefferson, l'aire de répartition canadienne des salamandres unisexuées dépendantes de la salamandre à points bleus est peu comprise, mais pas entièrement inconnue. L'existence de salamandres unisexuées dépendantes de la salamandre à points bleus a été signalée en Nouvelle-Écosse par Gilhen (1978) et au Nouveau-Brunswick par Cook et Gorham (1979). Des collectes ont été effectuées par Les Lowcock dans les provinces des Maritimes, le sud du Québec et le centre de l'Ontario (Lowcock, 1989, 1991), et plusieurs autres collectes ont été faites par Jean-François Desroches au Québec (voir aussi Nöel et al., 2008, 2011). Les identifications de salamandres bisexuées et unisexuées ont été incluses dans un relevé herpétologique général effectué dans le sud de l'Ontario par l'organisme Hamilton Field Naturalists (Lamond, 1994). Ce relevé a permis d'identifier des individus unisexués et leurs espèces donneuses de sperme dans un grand nombre de populations du sud de l'Ontario. Des salamandres à points bleus et des salamandres unisexuées qui leur sont associées ont été échantillonnées dans le parc provincial Rondeau, dans la municipalité de Chatham-Kent, sur la rive nord du lac Érié, ainsi que dans des populations du parc provincial Algonquin et du secteur de Haliburton, dans le district de Nipissing (tableau 1).
Habitat
Besoins en matière d'habitat
On présume que l'Ambystoma unisexué utilise le même habitat que ses espèces donneuses de sperme. Ces salamandres sont présentes dans les forêts décidues ou mixtes des hautes terres (Klemens, 2000) qui contiennent des étangs de reproduction convenables, ou à proximité. Ces sites comprennent des dolines calcaires, des étangs de kettle et d'autres types de bassins naturels (Nyman, 1991). Les étangs de reproduction sont exempts de poissons prédateurs, souvent éphémères, et formés par le ruissellement printanier, l'eau souterraine ou des sources. Les salamandres unisexuées et leurs espèces bisexuées sympatriques passent l'hiver sous terre, sous la ligne de gel. Les étangs de reproduction des salamandres doivent contenir des points de fixation pour les œufs, et les étangs éphémères doivent exister aussi longtemps que nécessaire pour le développement des larves. Les œufs sont normalement fixés à des brindilles ou à des branches submergées, mais ils peuvent aussi être fixés à la végétation riveraine submergée ou à des herbes ou des carex submergés. Les salamandres à points bleus et les salamandres à petite bouche pondent leurs œufs un à un ou en petits groupes (Petranka, 1978). Les salamandres unisexuées pondent leurs œufs en masses de 20 à 50 œufs qui sont semblables aux masses d'œufs pondues par les salamandres de Jefferson (Bogart, 1982). Dans les étangs, les salamandres se nourrissent d'une variété d'invertébrés ainsi que de larves ou de têtards d'autres amphibiens.
Bériault (2005) a modélisé la qualité de l'habitat, la qualité de l'eau ainsi que le microhabitat et le macrohabitat utilisé par les salamandres de Jefferson et à points bleus. Tous les sites qu'elle a examinés contenaient, respectivement, des individus unisexués dépendants de la salamandre de Jefferson et de la salamandre à points bleus, et aucune distinction n'a été faite entre les individus bisexués et les individus unisexués. Elle a constaté que les larves d'Ambystomatidés n'étaient pas particulièrement vulnérables à un pH relativement faible, à d'autres paramètres chimiques de l'eau ou à la profondeur, à la température ou à la qualité de l'eau dans les étangs de l'Ontario. Le succès des larves peut toutefois être perturbé par une disponibilité réduite des proies qui, pour leur part, sont plus vulnérables à un pH faible que les larves de salamandres elles-mêmes (Sandinski et Dunson, 1992).
Mis à part les quelques jours où ils passent dans l'étang de reproduction, les adultes vivent et s'alimentent dans les boisés décidus ou mixtes. On trouve rarement les salamandres fouisseuses sur le tapis forestier, sauf lorsqu'elles migrent en direction ou en provenance d'un étang de reproduction. La majeure partie de l'information sur l'écologie terrestre des Ambystomatidés provient d'expériences réalisées au moyen de radioémetteurs implantés dans la cavité du corps des individus ciblés. Faccio (2003) a effectué le suivi de salamandres de Jefferson et de salamandres maculées au Vermont. Certains des individus pistés par Faccio étaient possiblement des individus unisexués dépendants de la salamandre de Jefferson, car les salamandres de Jefferson et leurs congénères unisexuées sont présentes au Vermont (Bogart et Klemens, 1997), mais quatre des huit salamandres de Jefferson en question étaient des mâles. Les salamandres utilisent des terriers horizontaux en été, mais utilisent des failles et des terriers verticaux en hiver (Faccio, 2003).
Au Canada, des salamandres unisexuées dépendantes de la salamandre de Jefferson ont fait l'objet d'une étude de radiopistage (Bériault, 2005). L'habitat de ces salamandres comprend les zones terrestres situées à 1 km ou plus d'un étang de reproduction, ainsi qu'une « zone centrale » avec un rayon d'environ 300 m autour d'un étang de reproduction. Les microhabitats comprennent les terriers de petits mammifères, les failles rocheuses, les souches d'arbres, la litière de feuilles, les rondins et les débris ligneux qui jonchent le tapis forestier. Les salamandres pistées utilisent de profondes failles rocheuses et des terriers de petits mammifères (Bériault, 2005). À l'heure actuelle, un rayon de 1 km autour des étangs de reproduction est protégé pour la salamandre de Jefferson et les individus unisexués qui lui sont associés (voir Protection, statuts et classements) afin de permettre l'expansion des populations, l'immigration et la dispersion. Avant 2010, les autorités en matière de planification utilisaient une « zone tampon » de 30 m autour des étangs de reproduction, et des superficies considérables d'habitat d'alimentation et d'hivernage ont été perdues. Ces pertes se reflètent dans la taille réduite des populations et dans la disparition de populations entières dans certains sites historiques (voir Fluctuations et tendances).
Tendances en matière d'habitat
L'habitat des salamandres unisexuées dépendantes de la salamandre de Jefferson et de la salamandre à points bleus dans le sud de l'Ontario se limite à des zones boisées fragmentées et à de petites terres agricoles. Les activités d'aménagement des terres associées à l'urbanisation, à l'extraction d'agrégats et à l'exploitation des ressources donnent lieu à une perte continue d'habitat convenable ainsi qu'à une augmentation de la fragmentation de l'habitat, et, donc, à une hausse de l'isolement des populations. En plus d'entraîner la perte directe d'habitat, l'exploitation des ressources peut altérer la nappe phréatique et l'écoulement des eaux souterraines, ce qui a un effet négatif sur les régimes d'humidité dans les milieux humides et les substrats adjacents. La réduction ou la perte d'habitat d'alimentation et d'hivernage des populations de salamandres de Jefferson est attribuée au drainage des milieux humides et à l'exploitation des ressources (COSEWIC, 2010). Ces changements peuvent raccourcir l'hydropériode des étangs éphémères et, donc, mener à une perte d'habitat convenable. La réduction, la dégradation et la perte d'étangs de reproduction ont été documentées chez la salamandre à petite bouche et les salamandres unisexuées qui lui sont associées sur l'île Pelée (COSEWIC, 2004) (voir Menaces).
Biologie
La biologie générale des salamandres à petite bouche, de Jefferson et à points bleus a été résumée par Downs (1989) et Petranka (1998) et sur divers sites Web (p. ex. NatureServe, 2014 et AmphibiaWeb, 2014). Des observations du cycle vital ont été faites dans certaines populations où des salamandres unisexuées coexistent avec ces espèces, et où elles pourraient même ne pas avoir été remarquées (p. ex. Weller, 1980). De manière générale, on présume que les individus unisexués reproduisent le comportement normal et observable des larves, des juvéniles et des adultes femelles des espèces donneuses de sperme cooccurrentes.
Cycle vital et reproduction
La parade nuptiale des salamandres unisexuées avec les mâles de la salamandre à petite bouche et de la salamandre à points bleus sur l'île Pelée a été décrite (Licht, 1989; Licht et Bogart, 1990). La parade nuptiale des salamandres de Jefferson a été décrite par Mohr (1931). Une fois dans les étangs, les mâles courtisent les femelles et déposent des spermatophores sur le substrat (p. ex. feuilles, brindilles) au fond de l'étang. La femelle recueille un spermatophore au moyen des « lèvres » de son cloaque et, après 1-2 jours, elle pond plusieurs masses d'environ 30 œufs chacune sur des tiges ou sur la végétation submergée en périphérie de l'étang (Brodman, 1995). Bien que les salamandres de Jefferson mâles puissent courtiser les femelles de leur espèce ainsi que les femelles unisexuées, Dawley et Dawley (1986) ont montré que les salamandres de Jefferson mâles étaient capables distinguer les femelles de leur espèce des femelles unisexuées, et que les mâles utilisés dans leurs expériences préféraient les salamandres de Jefferson femelles. Les mâles de la salamandre à points bleus et de la salamandre de Jefferson s'accrochent aux femelles durant la parade, mais l'amplexus ne fait pas partie du processus d'accouplement des mâles de la salamandre à petite bouche sur l'île Pelée (Licht et Bogart, 1990).
La persistance des salamandres unisexuées avec leurs espèces donneuses de sperme pourrait être liée à une stratégie de reproduction des Ambystomatidés qui implique la compétition spermatique. Les Ambystomatidés mâles sont généralement plus communs que les femelles dans les étangs de reproduction qui ne comptent pas d'individus unisexués, et ils produisent beaucoup plus de spermatophores que ce qui serait nécessaire pour soutenir la population. Chez certaines espèces (p. ex. la salamandre maculée), des « champs » de spermatophores contenant plusieurs centaines de spermatophores peuvent être observés dans les étangs de reproduction, et les mâles déposent leurs spermatophores par-dessus ceux de leurs rivaux (Petranka, 1998). Les salamandres à petite bouche déposent leurs spermatophores relativement au hasard lorsque les mâles et les femelles sont à proximité (Licht et Bogart, 1990). Les mâles de la salamandre de Jefferson et de la salamandre à points bleus entreprennent des parades qui réservent des spermatophores aux femelles courtisées, mais les mâles produisent quand même beaucoup plus de spermatophores que nécessaire (~20 par femelle courtisée) et 2 fois plus de spermatophores lorsqu'ils courtisent des femelles conspécifiques que lorsqu'ils courtisent des salamandres unisexuées (Uzzell, 1969). Il a été avancé que les Ambystoma unisexués pourraient réduire la densité des populations de leurs espèces donneuses de sperme. Clanton (1934) a posé l'hypothèse voulant que les spermatophores constituent un facteur limitatif dans les populations dominées par des salamandres unisexuées, et que celles-ci aient le dessus sur les femelles de l'espèce donneuse de sperme, ce qui réduirait constamment la population de l'espèce donneuse de sperme jusqu'à ce que le recrutement cesse. Comme les salamandres sur l'île Pelée sont isolées sur cette île depuis quelque 5 000 ans (Calkin et Feenstra, 1985), elles représentent vraisemblablement une condition d'équilibre pour les salamandres unisexuées qui vivent avec des salamandres bisexuées donneuses de sperme. Le pourcentage de salamandres bisexuées et unisexuées sur l'île Pelée (table 1) est semblable à ce que l'on observe dans les populations comptant des individus bisexués et unisexués dans la zone continentale.
D'après le taux de croissance et la taille des individus qui se reproduisaient pour la première fois dans un étang du sud de l'Ontario, Weller (1980) a estimé que les salamandres de Jefferson mâles retournaient à l'étang de reproduction 22 mois après leur métamorphose. Les salamandres de Jefferson femelles et les salamandres unisexuées, cependant, avaient besoin d'au moins 34 mois avant de revenir. Des données indiquent aussi que les salamandres unisexuées dépendantes de la salamandre à petite bouche prennent généralement plus de temps pour atteindre la maturité sexuelle que les femelles de la salamandre à petite bouche (Licht et Bogart, 1989). Les individus unisexués triploïdes dépendants de la salamandre à points bleus atteignent aussi la maturité sexuelle plus lentement que les salamandres à points bleus; les salamandres unisexuées tétraploïdes ont besoin d'une année de plus (Lowcock et al., 1992). Weller (1980) a constaté que la fréquence de la reproduction variait en fonction de chaque individu. Des 26 salamandres de Jefferson mâles qui ont atteint et quitté les étangs de reproduction au cours de la première année de son étude, 12 % sont revenues au cours de chacune des 4 années suivantes, 4 % ne sont pas revenues avant 4 ans, et les autres sont revenues dans l'étang tout en sautant des années selon diverses combinaisons. Les femelles, y compris les salamandres de Jefferson et les salamandres unisexuées dépendantes de la salamandre de Jefferson, ont affiché des tendances similaires, avec 10 % de 206 femelles revenant chaque année dans l'étang, 6 % ne revenant pas avant 4 ans, et les autres revenant ou sautant des années selon diverses combinaisons.
Une mortalité élevée des œufs, observée pour de nombreuses masses d'œufs dans un étang de reproduction, signifie souvent la présence d'Ambystoma unisexués (Petranka, 1998). Piersol (1910) a observé une forte mortalité des masses d'œufs dans un étang de Toronto (Ontario) qui abritait probablement une population de salamandres de Jefferson affichant une forte fréquence d'individus unisexués, et Clanton (1934) a mentionné la mortalité des œufs dans le sud du Michigan, où des salamandres bisexuées vivaient avec des salamandres unisexuées. La période embryonnaire, de la ponte des œufs à l'éclosion, varie entre 3 et 14 semaines dans le complexe unisexué/bisexué, et dépend du moment de la ponte au cours de la saison ainsi que de la température de l'eau (Smith, 1983), avec une moyenne d'environ 28 jours (Brodman, 1995) au sein d'une population du nord de l'Ohio composée de salamandres de Jefferson et des individus unisexués qui leur sont associés. Un taux de survie embryonnaire de 60 à 88 % a été constaté par Cook (1983) chez la salamandre de Jefferson, d'après l'étude de cinq étangs dans le Massachusetts. Par contre, les œufs de salamandres unisexuées dépendantes de la salamandre à petite bouche affichent habituellement un très faible taux de survie embryonnaire (16 %) (Bogart et Licht, 1986).
Lorsque les œufs éclosent, les larves s'alimentent de zooplancton jusqu'à ce qu'elles soient assez grosses pour s'alimenter d'autres invertébrés, dont des nématodes, des hydracariens, des cladocères, des copépodes, des collemboles, des larves de moustiques et de chironomidés, des escargots et d'autres insectes (Smith et Petranka, 1987). Les larves sont souvent cannibales et mangent aussi des larves d'espèces d'Ambystoma sympatriques (Brandon, 1961; Smith et Petranka, 1987). La période larvaire varie de 2 à 4 mois, sans doute en fonction de la température de l'eau, de la nourriture disponible et de l'hydropériode (Downs, 1989). En Ontario, la métamorphose se produit entre la mi-juillet et la mi-septembre (Bogart, obs. pers.). Au début de novembre, les juvéniles atteignent en moyenne une longueur totale de 62 mm (Downs, 1989). D'après des études réalisées au Maryland (Thompson et al., 1980), en Ohio (Downs, 1989) et en Illinois (Mullen et Klueh, 2009), les taux de survie des larves et les taux de recrutement seraient très faibles (de 0 à 0,7 %); on connaît peu l'écologie des juvéniles. Downs (1989) a trouvé des juvéniles qui s'étaient éloignés jusqu'à 92 m de l'étang de reproduction en 10 jours.
Downs (1989) a effectué une étude de marquage-recapture dans laquelle il a estimé que de 10 à 18 % des adultes survivent durant 3 ans. Weller (1980) a estimé un taux de survie annuelle très élevé chez l'A. jeffersonianum, soit de 0,981 pour les femelles (y compris les salamandres unisexuées) et de 0,883 pour les mâles. D'après la squelettochronologie, Flageole et Leclair (1992) ont documenté que la plupart des individus dans la population examinée étaient âgés de 2 à 18 ans, mais que certains atteignaient 32 ans. Des données probantes sur la longévité des salamandres unisexuées proviennent de l'observation suivante de L. Licht : [traduction] « Une salamandre (sans doute LJJ) qui a été recueillie au printemps 1988 dans un étang de reproduction à proximité de Hamilton, en Ontario, est toujours en vie au moment d'écrire ces lignes. En présumant que l'espèce a besoin d'une période de 3 ans pour atteindre la maturité sexuelle et se reproduire, on peut conclure que cette salamandre a maintenant au moins 30 ans » (Licht, comm. pers., 2015).
La durée d'une génération chez la salamandre de Jefferson a été calculée à 11 ans d'après la formule suivante : âge à maturité + 1/mortalité, où mortalité = taux annuel de mortalité des adultes (Downs, 1989). Le taux de mortalité a été estimé d'après les données fournies pour la salamandre de Jefferson et les salamandres unisexuées qui lui sont associées (Weller, 1980; Downs, 1989) comme la moyenne de 2, 12 et 27 % = 14 %. Par conséquent : durée d'une génération = 4 + 1/0,14 = 11 ans. Les taux de mortalité n'ont pas été calculés pour les populations de salamandres unisexuées dépendantes de la salamandre à petite bouche ou de la salamandre à points bleus, et doivent varier en fonction des différentes menaces qui pèsent sur la survie des adultes (p. ex. routes et circulation routière à proximité des étangs de reproduction). La durée d'une génération chez la salamandre à petite bouche a été estimée à 3 ans (COSEWIC, 2004) d'après l'âge moyen estimé des adultes reproducteurs seulement, sans tenir compte de la mortalité. Les salamandres unisexuées dépendantes de la salamandre à petite bouche et de la salamandre à points bleus ont besoin de plus de temps que les salamandres bisexuées pour atteindre la maturité sexuelle (Licht et Bogart, 1989; Lowcock et al., 1992), et le taux de mortalité présumé des adultes est inconnu. Une durée de génération d'environ 8 ans serait plus réaliste pour ces salamandres unisexuées.
Physiologie et adaptabilité
Comme chez la plupart des amphibiens, les Ambystoma unisexués ont une peau perméable à l'eau et à l'oxygène. Pour éviter le dessèchement et l'anoxie, les juvéniles et les adultes se tiennent normalement dans des milieux frais et humides et ne se déplacent à la surface du sol que pendant les nuits pluvieuses ou humides. Dans leur aire de répartition, les Ambystoma unisexués entreprennent leurs activités de reproduction en même temps que leurs espèces donneuses de sperme, qui comptent souvent parmi les premières salamandres à se reproduire au printemps (Petranka, 1998); elles sont d'ailleurs actives à < 1 °C (Feder et al., 1982). L'espèce migre et se reproduit souvent lorsque les étangs sont encore couverts de glace et avant que le sol soit complètement dégelé (Bogart, obs. pers.). Les salamandres unisexuées ne possèderaient pas d'agents cryoprotecteurs : normalement, les adultes et les œufs gelés meurent (Pisapio et Bogart, données inédites).
Bien qu'on n'ait pas analysé le venin présent dans la peau des Ambystoma bisexués ou unisexués, on soupçonne qu'il joue un important rôle de défense contre les prédateurs. Les glandes à venin sont particulièrement concentrées dans la peau de la surface dorsale de la queue. Lorsque la salamandre rencontre un prédateur possible ou qu'elle est manipulée par un humain, elle présente la queue et l'élève en direction de la menace. La salamandre réagit habituellement à un prédateur en agitant la queue et en faisant suinter du venin (Ducey et Brodie, 1983; Brodie, 1989).
Dispersion et migration
Dans le sud de l'Ontario, on constate deux périodes de déplacement chez les salamandres unisexuées et leurs espèces donneuses de sperme : la dispersion des juvéniles nouvellement métamorphosés depuis les étangs jusqu'à la forêt environnante, qui a généralement lieu en juillet et en août, et la migration des adultes, qui quittent leurs sites d'hibernation au printemps pour aller se reproduire dans les étangs et en revenir (de la fin de mars jusqu'à la mi-avril).
La distance de migration entre l'étang de reproduction et l'habitat terrestre chez les salamandres unisexuées dépendantes de la salamandre de Jefferson peut dépasser 1 km (Bériault, 2005), mais la distance parcourue varie selon les individus et les populations. Quatre-vingt-dix pour cent des adultes suivis par radiotélémétrie résidaient dans un habitat convenable à moins de 300 m de leur étang de reproduction (Bériault, 2005). Durant leurs migrations, les salamandres peuvent traverser des milieux terrestres qu'on ne considère pas comme des milieux convenables, par exemple des champs agricoles, des plantations et des routes. Comme les salamandres unisexuées adultes et juvéniles migrent seulement les nuits pluvieuses ou humides, on peut les trouver dans ce genre de milieu lorsque les conditions ne leur permettent pas d'achever leur migration. La fidélité à l'étang, c.-à-d. que chaque individu retourne annuellement au même étang pour se reproduire, n'a pas été examinée chez les Ambystoma unisexués, mais elle a été confirmée chez plusieurs espèces d'Ambystoma, comme la salamandre maculée (Schoop, 1965), la salamandre de Jefferson (Douglas et Monroe 1981) et l'A. talpoideum (Raymond et Hardy, 1990).
Interactions intraspécifiques
Dans l'aire de répartition des Ambystoma unisexués, les espèces donneuses de sperme d'Ambystoma cooccurrentesprésentent des caractéristiques semblables et partagent des habitats semblables. Les larves d'Ambystomatidés s'alimentent d'autres larves d'Ambystomatidés, mais on ignore si les individus unisexués ont la capacité de choisir leurs proies ou de faire une distinction entre les larves bisexuées et unisexuées. La prédation exercée par des larves de la salamandre marbrée (A. opacum) sur les larves de salamandres bisexuées et unisexuées pourrait considérablement réduire les taux de survie (Cortwright, 1988). Les larves de la salamandre marbrée sont plus grosses parce que cette espèce se reproduit à l'automne plutôt qu'à l'été. Elles ne sont pas présentes au Canada, mais elles coexistent avec des salamandres unisexuées dans la zone continentale de l'Ohio ainsi que sur l'île Kelleys, toujours en Ohio. Les salamandres maculées coexistent avec des salamandres unisexuées au sein de nombreuses populations du Québec et de la zone continentale de l'Ontario. Nyman (1991) a observé le partitionnement écologique des larves de salamandres maculées et de salamandres unisexuées dans le New Jersey.On trouve communément des tritons verts (Notopthalmus viridescens) qui se reproduisent dans les mêmes étangs que les salamandres unisexuées. Rien n'indique que ces espèces ont un grave impact sur les salamandres unisexuées, mais il n'existe pas de données concernant une concurrence possible.
Taille et tendances des populations
Activités et méthodes d'échantillonnage
Les populations d'Ambystomatidés sont efficacement échantillonnées au moyen de clôtures de déviation combinées à des pièges à fosse au printemps, alors que les adultes font leur entrée dans les étangs de reproduction (Heyer et al., 1994); les individus peuvent aussi être piégés dans les étangs de reproduction au moyen de pièges à ménés (figure 8) (tableau 1). De petits échantillons de tissus sont prélevés à des fins d'extraction de l'ADN, après quoi les salamandres sont relâchées. Avant 2004, c'est-à-dire avant que les méthodes d'analyse de l'ADN ne soient utilisées, un étang était désigné comme un étang de reproduction pour les membres du complexe bisexué/unisexué si la présence d'au moins quelques individus était confirmée sur le plan génétique grâce à l'analyse d'isozymes (Bogart, 1982). Les individus devaient être sacrifiés aux fins des analyses d'isozymes; c'est pourquoi celles-ci n'étaient pas acceptables pour déterminer les fréquences absolues des individus bisexués et unisexués dans les populations ou pour estimer les tendances au fil du temps. Des estimations des fréquences des individus bisexués/unisexués ont été obtenues pour quelques populations de salamandres à points bleus et de salamandres unisexuées dépendantes de la salamandre à points bleus d'après la ploïdie qui a été déterminée grâce à un faible échantillon de sang, par cytométrie en flux (Lowcock et al., 1991, 1992) (tableau 1). Depuis 2004, les loci d'ADN microsatellite offrent une meilleure méthode d'analyse génétique (Ramsden et al., 2006; Bogart et al., 2007), car on peut ainsi établir le génotype d'un grand nombre d'individus en relativement peu de temps (voir Identification génétique); des tendances n'ont toutefois été dégagées que pour un très faible nombre de populations, d'après le nombre de masses d'œufs observées au fil du temps. Les efforts soutenus des membres de l'équipe de rétablissement de la salamandre de Jefferson ont donné lieu à des relevés d'étangs de reproduction nouveaux et historiques en Ontario, en vue de trouver de nouveaux étangs occupés (voir Activités de recherche) et d'estimer les tendances en matière de population au fil du temps.

Description longue de la figure 8
Photo d'un piège à ménés faisant l'objet d'une inspection visant à trouver des salamandres dans l'aire de conservation de Hilton Falls à la fin mars (la glace a libéré les bords de l'étang).
Tableau 1. Fréquences des génomotypes trouvés au sein des sous-populations d'Ambystoma unisexués en Ontario où des échantillons de taille adéquate étaient disponibles pour estimer les distributions des fréquences d'individus bisexués et d'individus unisexués diploïdes ou polyploïdes. Les fréquences sont présentées selon le nombre d'individus de chaque génomotype et le pourcentage dans la sous-population (en parenthèses). Tous les génomotypes unisexués ont au moins une garniture chromosomique de l'A. laterale (L), et au moins une garniture ou un génome de l'A. jeffersonianum (J) ou de l'A. texanum (T). Les individus diploïdes ont 2 garnitures chromosomiques, les triploïdes en ont 3, les tétraploïdes en ont 4 et les pentaploïdes en ont 5.
| Population | (n) | LL | TT | LT | LLT | LTT | LLLT | LTTT | LLTT | Source |
|---|---|---|---|---|---|---|---|---|---|---|
| Île Pelée Note du tableaua | 1287 | 83 (6,4) | 191 (14,8) | 272 (21,1) | 474 (36,8) | 209 (16,2) | 22 (1,7) | 19 (1,5) | 17 (1,3) | COSEWIC (2004) |
| Sous-population | (n) | JJ | LJJ | LJJJ | LLJJ | LLJJJ | Source |
|---|---|---|---|---|---|---|---|
| Kitchener (site 1) Note du tableaub | 142 | 12 (8,45) | 111 (78,17) | 19 (13,38) | 0 | 0 | LGL (2007) (données inédites) |
| Kitchener (site 1) Note du tableaub | 191 | 15 (7,85) | 139 (72,77) | 36 (18,85) | 0 | 0 | LGL (2008) (données inédites) |
| Kitchener (site 2) Note du tableaub | 46 | 0 | 38 (82,6) | 5 (10,87) | 2 (4,35) | 1 (2,17) | NRSI (2009) (données inédites) |
| Kitchener (site 2) Note du tableaub | 16 | 0 | 20 (95,24) | 0 | 1 (4,76) | 0 | NRSI (2015) (données inédites) |
| Aire de conservation de Hilton Falls Note du tableaub | 520 | 168 (32,3) | 337 (64,8) | 15 (2,88) | 0 | 0 | Ramsden (2004) |
| Waterdown Note du tableaub | 118 | 11 (9,32) | 103 (87,29) | 4 (3,39) | 0 | 0 | OMNR (2007) (données inédites) |
| Erindale Note du tableauc | 2865 | 426 (14,9) | 2439 (85,13) | Weller (1980) |
| Sous-population | (n) | LL | LJ | LLJ | LLLJ | LLLLJ | Source |
|---|---|---|---|---|---|---|---|
| Cambridge Note du tableaud | 124 | 17 (13,7) | 6 (4,8) | 99 (79,8) | 2 (1,61) | 0 | Ecoservices (2007) (données inédites) |
| Parc provincial Rondeau Note du tableaue | 72 | 25 (34,7) | 8 (11,11) | 37 (51,39) | 2 (2,78) | 0 | S. Dobbyn (2004) (données inédites) |
| Parc provincial Algonquin Note du tableau d | 63 | 63 (100,0) | 0 | 0 | 0 | 0 | Wildlife Research Station (2010) (données inédites) |
| Parc provincial Algonquin Note du tableau d | 131 | 121 (92,4) | 0 | 10 (7,63) | 0 | 0 | Wildlife Research Station (2011) (données inédites) |
| Étang à castors de Haliburton Note du tableau b | 2646 | 650 (24,6) | 0 | 1714 (64,78) | 279 (10,54) | 3 (0,1) | Lowcock (1991) |
Abondance
Compte tenu de la difficulté d'établir une distinction entre les salamandres unisexuées et leurs espèces donneuses de sperme cooccurrentes, seules quelques études ont été réalisées pour identifier génétiquement de grands nombres d'individus dans des sous-populations en vue d'estimer l'abondance relative des individus bisexués et unisexués (tableau 1). Quelque 80 % des 1 287 salamandres échantillonnées sur l'île Pelée étaient des individus unisexués dépendants de la salamandre à petite bouche. Les salamandres unisexuées dépendantes de la salamandre de Jefferson sont plus nombreuses que les salamandres de Jefferson dans toutes les populations connues de l'Ontario, et certaines populations ne sont constituées que d'individus unisexués. L'absence de salamandres de Jefferson dans ces populations est sans doute attribuable en partie au faible effort d'échantillonnage et à la densité beaucoup plus élevée des salamandres unisexuées. Dans trois études dont la taille d'échantillon était supérieure à 100 individus, le pourcentage d'individus unisexués dépendants de la salamandre de Jefferson variait d'environ 67 à 92 % des individus échantillonnés. Les salamandres unisexuées dépendantes de la salamandre à points bleus représentaient 75 % de 2 646 salamandres trouvées dans un étang de la région de Haliburton (Ontario), mais n'ont pas été observées au sein d'autres populations de l'Ontario.
On ne dispose d'aucune estimation solide du nombre d'adultes que comptent les trois UD. La population dépendante de la salamandre à petite bouche est probablement très faible, avec possiblement moins de 1 000 adultes, d'après le faible nombre d'étangs de reproduction et les données limitées sur le plan des captures. Il pourrait y avoir moins de 2 500 salamandres de Jefferson adultes (COSEWIC, 2010). Les salamandres unisexuées dépendantes de la salamandre de Jefferson sont plus nombreuses que les individus de leur espèce donneuse de sperme, et représentent 60 à 95 % des sous-populations, soit plus de 10 000 adultes unisexués. Une forte incertitude est associée à la taille de la population de salamandres unisexuées dépendantes de la salamandre à points bleus, mais la population est sans doute très grande, comptant possiblement plus de 100 000 adultes.
Fluctuations et tendances
De manière générale, on présume que les salamandres fouisseuses (Ambystoma sp.) vivent assez longtemps (~ 30 ans) (voir Cycle vital et reproduction). Par conséquent, les salamandres unisexuées ont probablement plusieurs occasions de reproduction au cours de leur vie, ce qui pourrait compenser l'effet de facteurs extrinsèques, comme une vague de froid qui gèle des œufs ou un temps sec au printemps et à l'été qui assèche des étangs printaniers et tue des larves. Les fluctuations annuelles du nombre d'adultes qui se reproduisent peuvent dépendre des cohortes des années antérieures de « bon » ou de « mauvais » recrutement. Les tendances de la densité de population et les présomptions fondées sur la présence ou l'absence de détection d'individus ne peuvent être estimées qu'en effectuant des relevés annuels répétés dans les mêmes étangs et des relevés dans plusieurs étangs la même année.
D'après des relevés répétés sur une période d'environ 15 ans (1990 à 2005) en Ontario, on a estimé qu'aucune population de salamandres de Jefferson n'était plus grande qu'à l'époque où elle a été observée pour la première fois. La plupart des populations étaient en baisse, et certaines sont probablement disparues. On compte actuellement moins de 33 sous-populations existantes connues (définies ici comme le nombre d'étangs de reproduction existants connus) qui abritent des salamandres de Jefferson et les salamandres unisexuées qui leur sont associées. Quelques nouvelles sous-populations ont été trouvées depuis 2000, mais la persistance d'environ 25 sous-populations historiques ne peut pas être confirmée. De nombreux étangs de reproduction qui contenaient auparavant de nombreuses masses d'œufs en contiennent maintenant peu ou n'en contiennent pas. Certains étangs historiques ont été ensemencés de poissons prédateurs, d'autres s'assèchent maintenant trop vite pour permettre aux larves de compléter leur développement, et certains ont été détruits par des activités d'aménagement (COSEWIC, 2010).
Des observations annuelles documentant une réduction considérable du nombre de masses d'œufs, qui est passé de quelques centaines en 1979 à moins de 10 en 2006, portent à croire à un déclin de plus de 90 % durant cette période dans le cas de la salamandre de Jefferson et des salamandres unisexuées qui lui sont associées; la majorité des sites connus (plus de 20 étangs) a été échantillonnée (Bogart, obs. pers.). Weller (1980) a estimé que la population qu'il étudiait dans la région de Peel comptait 80 mâles reproducteurs et environ 838 femelles reproductrices (surtout des salamandres unisexuées). Weller a aussi compté les individus capturés lors de leur migration vers les étangs de reproduction. Il a constaté que leur nombre avait diminué entre le début et la fin de son étude de trois ans, passant de 624 en 1975 à 513 en 1976, puis à 324 individus en 1977. En 2003 et en 2004, des chercheurs ont examiné des sites de reproduction historiques le long de l'escarpement du Niagara, qui avaient été documentés en 1990 et en 1991. Dix-huit sites ont fait l'objet de recherches ciblant les masses d'œufs de salamandres de Jefferson et de salamandres unisexuées dépendantes de la salamandre de Jefferson, effectuées par le personnel du Programme de surveillance de l'escarpement du Niagara en Ontario. La présence de salamandres de Jefferson et de salamandres unisexuées dépendantes de la salamandre de Jefferson n'a été confirmée que dans trois sites (COSEWIC, 2010). Seules quelques-unes des sous-populations documentées de salamandres de Jefferson et des salamandres unisexuées qui leur sont associées ayant fait l'objet de relevés répétés en Ontario ont été stables pendant une période relativement longue d'après les relevés effectués en 1979, en 1981, en 1990 (NHIC, 1998) et de 2003 à 2005 (Bériault, 2005; Ramsden, 2008).
On ne dispose d'aucune donnée de relevé semblable pour la salamandre à petite bouche ou la salamandre à points bleus et les salamandres unisexuées qui leur sont associées. Il y a perte d'habitat de la salamandre à points bleus dans les aires aménagées et dans les zones où des milieux humides ont été drainés en Ontario et au Québec, mais la présence ou l'abondance de cette salamandre au fil du temps n'a pas été documentée. Les derniers relevés effectués pour évaluer la salamandre à petite bouche et les salamandres unisexuées qui lui sont associées sur l'île Pelée remontent aux années 1980 (COSEWIC, 2004), mais un relevé a été entrepris (2015) sur l'île par Dennis Murray (Ph. D.) et ses étudiants de l'Université Trent.
Immigration de source externe
Les salamandres unisexuées dépendantes de la salamandre à petite bouche sur l'île Pelée vivent sur une île isolée, et il n'existe aucune possibilité d'immigration de source externe en provenance de l'Ohio ou du Michigan, où d'autres individus unisexués dépendants de la salamandre à petite bouche vivent en petites populations isolées (figure 5). Les populations de salamandres de Jefferson et de salamandres unisexuées dépendantes de la salamandre de Jefferson les plus proches aux États-Unis se trouvent dans les comtés de Cattaraugus et de Wayne, dans l'État de New York (Bogart et Klemens, 2008) (figure 6). Vu les déplacements limités de ces salamandres, leur répartition actuelle et les obstacles à leur dispersion, une immigration provenant des États-Unis est très improbable. La plupart des salamandres unisexuées dépendantes de la salamandre à points bleus vivent probablement au Canada (figure 4), mais ces salamandres sont aussi présentes dans les États américains qui bordent le Canada entre le Maine et le Minnesota; c'est pourquoi il existe plusieurs régions où une dispersion au Canada depuis les États-Unis est possible.
Menaces et facteurs limitatifs
Facteurs limitatifs
La présence de populations d'Ambystoma unisexués est limitée par la présence d'une espèce donneuse de sperme acceptable vivant dans le même étang de reproduction.
Menaces
La classification des menaces présentée ci-dessous est fondée sur le système unifié de classification des menaces proposé par l'Union internationale pour la conservation de la nature (UICN) et le Partenariat pour les mesures de conservation (Conservation Measures Partnership, ou CMP) (voir Master et al. [2009] et CMP [2010] pour les détails). Les menaces peuvent être observées, inférées ou prévues à court terme. Dans le présent plan, les menaces sont caractérisées en fonction de leur portée (habituellement au cours des 10 prochaines années), de leur gravité (au cours des 10 prochaines années ou 3 générations, selon la plus longue de ces deux éventualités) et de leur immédiateté (menace en cours ou prévue). L'« impact » de la menace est calculé selon la portée et la gravité. L'impact des menaces pour les trois UD de salamandres unisexuées est présenté au tableau 2 (voir les tableaux complets aux annexes 2 à 4). Les menaces pesant sur chaque UD sont décrites ci-dessous, par ordre décroissant selon leur importance perçue.
| Numéro de la menace | Menace | UD dépendante de la salamandre à petite bouche | UD dépendante de la salamandre de Jefferson | UD dépendante de la salamandre à points bleus |
|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | Faible | Élevé | Négligeable |
| 1.1 | Zones résidentielles et urbaines | Faible | Élevé | Négligeable |
| 1.2 | Zones commerciales et industrielles | Non calculé | Faible | Négligeable |
| 1.3 | Zones touristiques et récréatives | Non calculé | Faible | Négligeable |
| 2 | Agriculture et aquaculture | Faible | Faible | Négligeable |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | Faible | Faible | Négligeable |
| 2.3 | Élevage de bétail | Non calculé | Faible | Négligeable |
| 3 | Production d'énergie et exploitation minière | Non calculé (immédiateté inconnue) | Élevé | Non calculé |
| 3.2 | Exploitation de mines et de carrières | Non calculé (immédiateté inconnue) | Élevé | Non calculé |
| 4 | Corridors de transport et de service | Faible | Élevé - moyen | Faible |
| 4.1 | Routes et voies ferrées | Faible | Élevé - moyen | Faible |
| 5 | Utilisation des ressources biologiques | Non calculé | Négligeable | Faible |
| 5.3 | Exploitation forestière et récolte du bois | Non calculé | Négligeable | Faible |
| 6 | Intrusions et perturbations humaines | Faible | Non calculé | Non calculé |
| 6.1 | Activités récréatives | Faible | Non calculé | Non calculé |
| 7 | Modifications des systèmes naturels | Élevé | Non calculé | Négligeable |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | Élevé | Non calculé | Négligeable |
| 7.3 | Autres modifications de l'écosystème | Faible | Non calculé | Non calculé |
| 8 | Espèces et gènes envahissants ou autrement problématiques | Inconnu | Faible | Négligeable |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | Inconnu | Faible | Négligeable |
| 9 | Pollution | Inconnu | Inconnu | Négligeable |
| 9.1 | Eaux usées domestiques et urbaines | Non calculé | Inconnu | Négligeable |
| 9.3 | Effluents agricoles et sylvicoles | Inconnu | Inconnu | Non calculé |
| 11 | Changements climatiques et phénomènes météorologiques violents | Non calculé (à l'extérieur de la période d'évaluation) | Non calculé | Non calculé |
| 11.2 | Sécheresses | Non calculé (à l'extérieur de la période d'évaluation) | Non calculé | Non calculé |
| Impact global des menaces calculé : | - | Élevé | Très élevé | Faible |
UD 1 : Salamandres unisexuées dépendantes de la salamandre à petite bouche (impact global des menaces calculé = élevé)
Modification du système naturel (impact « élevé »)
Le drainage des milieux humides représente probablement la plus grande menace pesant sur les salamandres à petite bouche et les salamandres unisexuées dépendantes de la salamandre à petite bouche. Les fossés de drainage modifient les niveaux d'eau dans les étangs de reproduction de salamandres. Le dragage des canaux et des fossés au bord de la route à la pointe Fish (un des sites occupés sur l'île Pelée) entraîne l'eau hors des étangs de reproduction. Le dragage peut avoir lieu dans toute l'île, et la seule population qui n'est pas touchée est celle de la carrière abandonnée de la pointe Sheridan. Les niveaux d'eau fluctuent dans le lac Érié, et les salamandres sont mieux adaptées aux niveaux élevés. Le déboisement et l'enlèvement des débris ligneux grossiers, particulièrement dans le secteur de la pointe Fish, peuvent avoir un effet négatif sur les jeunes (les adultes utilisent des terriers).
Corridors de transport et de service (impact « faible »)
Les étangs de reproduction des salamandres sur l'île Pelée sont situés à proximité des routes, et des individus traversent fréquemment ces routes durant la migration et la dispersion. La mortalité routière représente une menace pour les salamandres en migration et en dispersion. Même si la circulation est relativement faible durant la période de reproduction du printemps, celle qui est associée au tourisme a augmenté, particulièrement durant l'été, moment où les juvéniles quittent les étangs.
Intrusions et perturbations humaines (impact « faible »)
Des perturbations humaines ont lieu dans le secteur de la pointe Fish, qui est très fréquentée par les touristes. Les visiteurs qui photographient et cherchent des salamandres dans les bois et les étangs peuvent accidentellement piétiner des individus et leur habitat. D'autres visiteurs fréquentent aussi les habitats des salamandres pour observer des oiseaux et faire des randonnées.
Développement résidentiel et commercial (impact « faible »)
Une certaine expansion résidentielle a lieu sur l'île Pelée. Bien qu'il n'y ait aucun projet connu dans l'habitat des salamandres, cette possibilité persiste. Compte tenu de la petite taille de la population de salamandres, même un seul projet pourrait être problématique. La gravité de l'impact dépend du drainage du milieu humide (gravité extrême); si seul l'habitat terrestre était touché, la gravité serait réduite.
Agriculture (impact « faible »)
Un étang de reproduction est situé à proximité d'un champ agricole, et contient environ 5 % de la population. Il est possible que l'expansion du champ ou d'autres activités d'aménagement agricole ait lieu dans ce site, mais aucun plan n'est connu pour le moment.
Espèces introduites (impact « inconnu »)
L'introduction de dindons sauvages (Meleagris gallopavo) représente une menace potentielle pour la salamandre à petite bouche et les salamandres unisexuées qui lui sont associées sur l'île Pelée (Hamill, 2014; COSEWIC, 2014). Quelque 25 couples reproducteurs ont été relâchés en 2002, et une grande population est maintenant présente sur l'île. Les dindons peuvent modifier l'état du tapis forestier, au détriment des salamandres (particulièrement les juvéniles); il est aussi possible que les dindons mangent les salamandres.
UD 2 : Salamandres unisexuées dépendantes de la salamandre de jefferson (impact global des menaces calculé = très élevé)
Développement résidentiel et commercial (impact « élevé »)
La forêt carolinienne (y compris sa faune caractéristique) atteint la limite nord de sa répartition dans le sud de l'Ontario, mais la majeure partie de cette forêt en Ontario a été défrichée, d'abord à des fins agricoles, puis à des fins de développement urbain. Les pertes d'habitat se poursuivent à cause du développement résidentiel, particulièrement dans la région de Hamilton et le long de l'escarpement du Niagara. Lorsque les étangs de reproduction sont remblayés ou drainés, des disparitions se produisent à l'échelle locale. Les voies migratoires entre un étang de reproduction et l'habitat terrestre peuvent être bloquées par le développement (p. ex. clôtures anti-érosion). La cause la plus probable des faibles effectifs de la salamandre de Jefferson et des salamandres unisexuées qui lui sont associées au Canada est probablement le manque d'habitat convenable (milieux terrestres et étangs de reproduction).
Production d'énergie et exploitation minière (impact « élevé »)
L'extraction d'agrégats et l'exploitation de carrières représentent un problème important pour la salamandre de Jefferson et les salamandres unisexuées qui lui sont associées sur l'escarpement du Niagara. En plus de la perte d'étangs de reproduction et d'habitat terrestre, les modifications hydrologiques découlant de l'exploitation de carrières sous la nappe phréatique peuvent réduire l'hydropériode d'un étang de reproduction de sorte que celui-ci s'assèche avant la métamorphose des larves.
Corridors de transport et de service (impact « élevé - moyen »)
Le sud de l'Ontario est caractérisé par un dense réseau routier. Des salamandres sont souvent écrasées par des véhicules sur des routes lorsqu'ils migrent à destination ou en provenance de leur étang de reproduction (Beebee, 2013). D'après les données provenant de 500 étangs de reproduction de salamandres maculées au Massachusetts, Gibbs et Shriver (2005) ont modélisé l'importance démographique de la mortalité routière. Ils ont constaté qu'un risque annuel de mortalité routière supérieur à 10 % pouvait entraîner la disparition d'une population locale; dans le cas d'une mortalité routière de 20 à 30 %, une population serait disparue en 25 ans. Les bordures de chaussée et les bouches d'égout peuvent agir comme des barrières ou des pièges, respectivement. La mortalité routière est considérable dans certaines régions du sud de l'Ontario, malgré les tentatives d'atténuation (p. ex. fermeture de routes à proximité des sites de reproduction). On s'attend à ce que la mortalité routière ait de graves répercussions sur les populations locales de salamandres de Jefferson et sur les salamandres unisexuées qui leur sont associées.
Espèces envahissantes et problématiques (impact « faible »)
Les poissons introduits, le zooplancton envahissant et les plantes aquatiques envahissantes (Phragmites australis) ont été pris en compte dans cette catégorie. Les Ambystomatidés ne coexistent pas bien avec les poissons prédateurs, et de nombreux sites de reproduction où il semble que l'espèce n'existe plus ont été ensemencés de poissons. Le zooplancton introduit est en voie de devenir un problème à l'échelle de l'écosystème dans le sud de l'Ontario, car les arthropodes indigènes sont réticents à s'en nourrir, et les proies des salamandres pourraient être touchées. Mis à part l'introduction de poissons prédateurs, on ne dispose d'aucune information à savoir si ces autres menaces sont problématiques pour les salamandres.
Pollution (impact inconnu)
Les routes constituent souvent une source de pollution chimique (p. ex. sels, métaux et produits de combustion) qui dégrade les milieux aquatiques et terrestres adjacents. Les effets nocifs des sels épandus pour déglacer les routes peuvent s'étendre sur des distances considérables dans les milieux humides, et on a montré que ces effets nuisent à la salamandre maculée (Turtle, 2000; Karraker et al., 2008; Collins et Russell, 2009).
UD 3 : Salamandres unisexuées dépendantes de la salamandre à points bleus (impact global des menaces calculé = faible)
Même si les salamandres unisexuées dépendantes de la salamandre à points bleus sont soumises à la plupart des mêmes menaces que les salamandres unisexuées dépendantes de la salamandre de Jefferson dans le sud de l'Ontario, ces populations représentent un faible pourcentage (de 5 à 10 %) de l'aire de répartition de la salamandre à points bleus. En raison de leur grande aire de répartition et de l'occurrence d'une proportion considérable de cette aire dans des régions éloignées des activités et du développement humains, la portée des menaces est beaucoup moins grande que pour les salamandres unisexuées dépendantes de la salamandre de Jefferson; c'est pourquoi l'impact des menaces est plus faible.
Nombre de localités
On présume que chaque étang de reproduction, ou chaque réseau d'étangs si ces derniers se trouvent dans une zone de 1 km, constitue une localité telle que définie par le COSEPAC, c'est-à-dire une zone particulière du point de vue écologique et géographique dans laquelle un seul phénomène menaçant peut affecter rapidement tous les individus du taxon présent. Dans la plupart des cas, le phénomène menaçant serait la perte d'un étang de reproduction convenable, causée par des changements aux propriétés hydrologiques ou par le drainage associé au développement ou à d'autres activités humaines.
On compte très peu de populations de salamandres dépendantes de la salamandre à petite bouche au Canada. L'île contient six étangs de reproduction historiques (Bogart et Licht, 1991), et chacun d'entre eux peut être considéré comme une localité. Cependant, deux étangs ont probablement disparu entre 1990 et 2003 (COSEWIC, 2004). De même, le nombre de localités de la salamandre de Jefferson (environ 33) (COSEWIC, 2010) serait moins élevé que le nombre de localités des salamandres unisexuées dépendantes de la salamandre de Jefferson, car les populations historiques et récentes sont incluses à la figure 6 et, dans certains étangs, seuls des individus unisexués dépendants de la salamandre de Jefferson ont été trouvés (COSEWIC, 2010). On a présumé que le recrutement avait cessé chez les salamandres de Jefferson ou chez les salamandres unisexuées dépendantes de la salamandre de Jefferson dans les étangs uniquement peuplés de salamandres unisexuées selon les derniers relevés. Les localités des salamandres de Jefferson (environ 33), ou 30 localités récentes, seraient les mêmes que celles des salamandres unisexuées dépendantes de la salamandre de Jefferson (COSEWIC, 2015; annexe C). Le grand nombre de localités qui est estimé pour les salamandres unisexuées dépendantes de la salamandre à points bleus témoigne de la répartition étendue des salamandres à points bleus au Canada (figure 7).
Protection, statuts et classements
Statuts et protection juridiques
Les salamandres de Jefferson, à points bleus et à petite bouche sont problématiques sur le plan des décisions en matière de conservation et de gestion, car elles coexistent avec des individus unisexués qui n'ont normalement pas reçu de cote de conservation (Kraus, 1995). L'État du Connecticut a désigné les complexes A. jeffersonianum et A. laterale espèces préoccupantes (special concern) parce qu'il est difficile d'établir une distinction entre les salamandres unisexuées et les espèces bisexuées A. jeffersonianum et A. laterale (Klemens, 2000). Le Connecticut a désigné espèces menacées les populations pures d'A. laterale diploïdes dans l'est de l'État, mais n'a accordé aucun statut à l'espèce A. jeffersonianum comme telle (seulement au « complexe » A. jeffersonianum) (Klemens, 2000).
Un programme de rétablissement fédéral a été proposé (Environment Canada, 2015), mais n'a pas encore été approuvé. En Ontario, les « individus polyploïdes dominés par jeffersonianum » sont des individus unisexués qui ont besoin des mâles de la salamandre de Jefferson, soit l'équivalent des salamandres unisexuées dépendantes de la salamandre de Jefferson. Ces individus unisexués reçoivent la même protection de l'habitat que la salamandre de Jefferson aux termes de la Loi de 2007 sur les espèces en voie de disparition (LEVD) (Government of Ontario, 2007) sous forme d'un règlement sur l'habitat, qui est entré en vigueur le 18 février 2010 (ESA, 2007; règlement 242/08, art. 28). La protection comprend une zone de 300 m autour des étangs de reproduction et une zone de 1 km entre un étang de reproduction et d'autres étangs de reproduction connexes ou des étangs de reproduction potentiels qui permettraient la dispersion. Jusqu'ici, aucun règlement semblable sur l'habitat n'a été adopté pour les salamandres unisexuées du Canada qui vivent avec la salamandre à petite bouche (en voie de disparition) sur l'île Pelée ou pour les salamandres unisexuées qui dépendent de la salamandre à points bleus. Les salamandres unisexuées reçoivent une autre protection juridique en vertu de diverses lois sur la protection des poissons et des autres espèces sauvages dans plusieurs provinces. Par exemple, la Loi sur la pêche sportive et la chasse du Nouveau-Brunswick définit un « animal de la faune » comme tout animal ou oiseau vertébré, et il est interdit de capturer ou de prendre en captivité, de garder en captivité ou de relâcher de la captivité tout animal de la faune. La Loi de 1997 sur la protection du poisson et de la faune de l'Ontario offre une protection semblable aux espèces sauvages de l'Ontario. En vertu de cette loi, la salamandre de Jefferson, la salamandre à points bleus, la salamandre maculée et la salamandre à petite bouche sont aussi désignées espèces spécialement protégées, et la protection additionnelle dont jouissent ces espèces s'applique aussi aux Ambystoma unisexués.
Statuts et classements non juridiques
L'Ambystoma unisexué ne figure pas dans les bases de données de l'UICN ou de NatureServe. Le Centre d'information sur le patrimoine naturel (CIPN) de l'Ontario attribue des cotes de conservation infranationales à deux catégories d'Ambystoma unisexués : les salamandres de Jefferson X à points bleus (Jefferson X Blue-spotted Salamander) dominées par le génome de la salamandre de Jefferson (pop. d'Ambystoma hybrides 1), soit l'équivalent des salamandres unisexuées dépendantes de la salamandre de Jefferson, ont reçu la cote S2 (et font l'objet d'un suivi par le CIPN); les salamandres de Jefferson X à points bleus dominées par le génome de la salamandre à points bleus (pop. d'Ambystoma hybrides 2), soit l'équivalent des salamandres unisexuées dépendantes de la salamandre à points bleus, ont reçu la cote S4 (et ne font l'objet d'aucun suivi par le CIPN).
Protection et propriété de l'habitat
La plupart des salamandres unisexuées qui dépendent de la salamandre à petite bouche sur l'île Pelée vivent dans un habitat visé par des mesures de protection du patrimoine naturel qui appartient au MRNFO, à l'Office de protection de la nature de la région d'Essex ou à Conservation de la nature Canada. Environ le tiers des populations connues de salamandres unisexuées dépendantes de la salamandre de Jefferson se trouvent dans des milieux convenables (forêts abritant de petits étangs) situés dans des parcs provinciaux le long de l'escarpement du Niagara, dans la ceinture de verdure de l'Ontario ou sur des terres gérées par des offices de protection de la nature. Le seul fait que ces milieux se trouvent dans un parc provincial, une aire de conservation ou le long de l'escarpement du Niagara ne garantit pas leur protection, car ces endroits sont utilisés à plusieurs fins récréatives. Certains petits étangs permanents situés sur des terres privées ou dans des aires de conservation ont été ensemencés de poissons prédateurs afin d'offrir des possibilités de pêche récréative (Bogart et Cook, 1991). Les individus unisexués dépendants de la salamandre à points bleus peuvent recevoir une protection générale s'ils vivent dans des parcs provinciaux ou dans des aires de conservation en Ontario, au Nouveau-Brunswick, en Nouvelle-Écosse et au Québec. En Ontario, la Déclaration de principes provinciale faite en vertu de la Loi sur l'aménagement du territoire fournit une certaine protection aux habitats fauniques importants, qui peuvent inclure des étangs de reproduction de salamandres.
Remerciements et experts contactés
Le rédacteur du rapport remercie les membres de l'équipe d'élaboration du programme de rétablissement de la salamandre de Jefferson qui ont travaillé fort pour trouver des individus dans des populations historiques et de nouvelles occurrences de l'espèce en Ontario. Il remercie en particulier Emma Followes, l'ex-coordinatrice de l'équipe de rétablissement, John Pisapio, qui a collaboré aux activités de recherche, et Melinda Thompson, qui rassemble les données recueillies pour l'équipe de rétablissement. Le rédacteur a eu la chance de travailler avec de nombreux étudiants intéressés et dévoués à l'Université de Guelph, tant sur le terrain qu'au laboratoire, ainsi qu'avec plusieurs collègues des États-Unis, qui ont envoyé des spécimens et des échantillons de tissus ayant joué un rôle déterminant dans l'amélioration de notre compréhension des salamandres unisexuées. Plusieurs experts-conseils en biologie ont collaboré étroitement avec le MRNFO et l'équipe de rétablissement de la salamandre de Jefferson pour fournir des données importantes. Le rédacteur remercie particulièrement Alison Featherstone, Christen Harrison, Karl Konze, Jessica Linton, Al Sandilands et Gwendolyn Weeks. Des écologistes à l'emploi de divers offices de protection de la nature ont fourni des données sur des populations historiques de l'espèce et en ont trouvé de nouvelles occurrences. Bon nombre d'entre eux sont membres de l'équipe de rétablissement, et le rédacteur tient particulièrement à remercier Brenda Van Ryswyk, de l'Office de protection de la nature de la région d'Halton. Il est reconnaissant envers Francis Cook (Musée canadien de la nature), Ross MacCulloch et Amy Lathrop (Musée royal de l'Ontario), qui conservent des collections de spécimens de référence et qui l'ont constamment encouragé et soutenu. Le rédacteur remercie tout particulièrement Jenny Wu, du Secrétariat du COSEPAC, qui a élaboré les cartes et calculé la superficie des différentes zones aux fins du rapport.
Le financement du présent rapport a été assuré par le Service canadien de la faune d'Environnement Canada.
Experts contactés
- Ruben Boles, Gestion des normes et populations d'espèces, Service canadien de la faune, Environnement Canada.
- Michael J. Oldham, Centre d'information sur le patrimoine naturel (CIPN) de l'Ontario, ministère des Richesses naturelles.
Membres de l'équipe de rétablissement de la salamandre de Jefferson
- Karine Bériault, biologiste des espèces en péril, ministère des Richesses naturelles de l'Ontario, bureau régional de Vineland.
- Kim Barrett, écologiste principale, Conservation Halton.
- Emma Followes, écologiste de district, ministère des Richesses naturelles de l'Ontario, district d'Aurora.
- Ron Gould, biologiste des espèces en péril, ministère des Richesses naturelles de l'Ontario, district d'Aylmer.
- Lisa Grbinicek, planificatrice environnementale, spécialiste en surveillance écologique, programme de surveillance de l'escarpement du Niagara de l'Ontario, Commission de l'escarpement du Niagara.
- Sue Hayes, coordonnatrice de projet, inventaires des études en milieu terrestre, Office de protection de la nature de Toronto et de la région.
- Anne Marie Laurence, spécialiste en surveillance écologique, programme de surveillance de l'escarpement du Niagara de l'Ontario, Commission de l'escarpement du Niagara.
- Heather Lynn, écologiste du patrimoine naturel, Credit Valley Conservation.
- Bob Murphy, conservateur principal, Centre pour la biodiversité et la biologie de la conservation, Musée royal de l'Ontario.
- John Pisapio, biologiste de la faune, ministère des Richesses naturelles de l'Ontario, district d'Aurora.
- Tony Zammit, écologiste, Office de protection de la nature de Grand River.
Sources d'information
AmphibiaWeb. 2014. Information on amphibian biology and conservation (University of California, Berkeley, California.) [Disponible en anglais seulement, consulté le 30 octobre 2014].
Avise, J.C. 2008. Clonality: The Genetics, Ecology and Evolution of Sexual Abstinence in Vertebrate Animals. Oxford University Press, New York.
Austin, N.E. et J.P. Bogart. 1982. Erythrocyte area and ploidy determination in the salamanders of the Ambystoma jeffersonianum complex. Copeia 1982:485-488.
Beebee T.J.C. 2013. Effects of road mortaility and mitigation measures on amphibian populations. Conservation Biology 27:657-668.
Bériault, K.R.D. 2005. Critical Habitat of Jefferson Salamanders in Ontario: An Examination through Radiotelemetry and Ecological Serveys. Mémoire de maîtrise, University of Guelph. 69 pp.
Bi, K. et J.P. Bogart. 2006. Identification of intergenomic recombinations in unisexual salamanders of the genus Ambystoma by genomic in situ hybridization. Cytogenetics and Genome Research 112:307-312.
Bi, K. et J. P. Bogart. 2010. Time and time again: unisexual salamanders (genus Ambystoma) are the oldest unisexual vertebrates. BMC Evolutionary Biology 10:238 (DOI: 10.1186/1471-2148-10-238).
Bi, K, J.P. Bogart et J. Fu. 2008. The prevalence of genome replacement in unisexual salamanders of the genus Ambystoma (Amphibia, Caudata) revealed by nuclear gene genealogy. BMC Evolutionary Biology 8:158.
Bishop, S.C. 1947. Handbook of Salamanders. Ithaca, New York. Comstock.
Bogart, J.P. 1982. Ploidy and genetic diversity in Ontario salamanders of the Ambystoma jeffersonianum complex revealed through an electrophoretic examination of larvae. Canadian Journal of Zoology 60:848-855.
Bogart, J.P. 2003. Genetics and polyploidy of hybrid species. Pp 109-134 in D.M. Sever (ed.). Reproductive Biology and Phylogeny of Urodela, vol. 1. M/s Science, Enfield, New Hampshire.
Bogart, J. P., J. Bartoszek, D.W.A. Noble et K. Bi. 2009. Sex in unisexual salamanders: discovery of a new sperm donor with ancient affinities. Heredity 103:483-493.
Bogart, J.P. et W.J. Cook 1991. Ambystoma survey on the Niagara Escarpment. 1991. Unpubl. report prepared for the Niagara Escarpment Branch of the Ontario Heritage Fund. 46pp.
Bogart, J.P. et K. Bi. 2013. Genetic and genomic interactions of animals with different ploidy levels. Cytogenetic and Genome Research 140: 117-136.
Bogart, J.P., K. Bi, J. Fu, D.W.A. Noble et J. Niedzwieki. 2007. Unisexual salamanders (genus Ambystoma) present a new reproductive mode for eukaryotes. Genome 50:119-136.
Bogart, J.P., R.P. Elinson et L.E. Licht. 1989. Temperature and sperm incorporation in polyploidy salamanders. Science 246:1032-1034.
Bogart, J.P. et M.W. Klemens. 1997. Hybrids and genetic interactions of mole salamanders (Ambystoma jeffersonianum and A. laterale) (Amphibia: Caudata) in New York and New England. American Museum of Natural History Novitates 3218:1-78.
Bogart, J.P. et M.W. Klemens. 2008. Additional distributional records of Ambystoma laterale, A. jeffersonianum (Amphibia: Caudata) and their unisexual kleptogens in northeastern North America. American Museum of Natural History Novitates 3627:1-58.
Bogart, J.P. et L.E. Licht. 1986. Reproductive biology and the origin of polyploids in hybrid salamanders of the genus Ambystoma. Canadian Journal of Genetics and Cytology 28:605-617.
Bogart, J.P. et L.E. Licht. 1987. Evidence for the requirement of sperm in unisexual salamander hybrids (genus Ambystoma). Canadian Field Naturalist 101:434-436.
Bogart, J.P. et L.E. Licht. 1991. Status Report on the Small-mouthed Salamander Ambystoma texanum in Canada. Report for the Committee on the Status of Endangered Wildlife in Canada (COSEWIC). 19 pp.
Bogart, J.P., L.E. Licht, M.J. Oldham et S.J. Darbyshire. 1985. Electrophoretic identification of Ambystoma laterale and Ambystoma texanum as well as their diploid and triploid interspecific hybrids (Amphibia: Caudata) on Pelee Island, Ontario. Canadian Journal of Zoology 63:340-347.
Bogart, J.P., L.A. Lowcock, C.W. Zeyl et B.K. Mable. 1987. Genome constitution and reproductive biology of hybrid salamanders, genus Ambystoma, on Kelleys Island in Lake Erie. Canadian Journal of Zoology 65:2188-2201.
Brandon, R.A. 1961. A comparison of the larvae of five northeastern species of Ambystoma (Amphibia, Caudata). Copeia 1961:377-383.
Brodie, E.D., III. 1989. Individual variation in antipredator response of Ambystoma jeffersonianum to snake predators. Journal of Herpetology 23:307-309.
Brodman, R. 1995. Annual variation in breeding success of two syntopic species of Ambystoma salamanders. Journal of Herpetology 29:111-113.
Calkin, P.E. et B.H. Feenstra. 1985. Evolution of the Erie-basin Great Lakes. Pp. 149-170, in P.F. Karrow et P.E. Calkin (eds.). Quaternary Evolution of the Great Lakes. Geoleological Association of Canada. Special Paper 30. St. Johns, Newfoundland.
Charney, N.D. 2012. Relating hybrid advantage and genome replacement in unisexual salamanders. Evolution 66:1387-1397.
Clanton, W. 1934. An unusual situation in the salamander Ambystoma jeffersonianum (Green). Occasional Papers of the Museum of Zoology, University of Michigan No. 290:1-14.
CMP (Conservation Measures Partnership). 2010. Threats taxonomy. [Disponible en anglais seulement, consulté en août 2014].
Collins, S.J. et R.W. Russell. 2009. Toxicity of road salt to Nova Scotia amphibians. Environmental Pollution 157:320-324.
Cook, R.P. 1983. Effects of acid precipitation on embryonic mortality of Ambystoma salamanders in the Connecticut Valley of Massachusetts. Biological Conservation 27:77-88.
Cook, F.R. 1984. Introduction to Canadian Amphibians and Reptiles. National Museum of Natural Sciences, National Museums of Canada, Ottawa, Ontario. 200pp (Également disponible en français : Cook, F.R. 1984. Introduction aux amphibiens et reptiles du Canada. Musée national des sciences naturelles, Musées nationaux du Canada, Ottawa (Ontario). 211 p.).
Cook, F.R. et S.W. Gorham. 1979. The occurrence of the triploid form in populations of the Blue-spotted Salamander, Ambystoma laterale, in New Brunswick. Journal of the New Brunswick Museum 1979:154-161.
Cortwright, S.A. 1988. Intraguild predation and competition: an analysis of net growth shifts in larval amphibian prey. Canadian Journal of Zoology 66:1813-1821.
COSEWIC. 2000. COSEWIC assessment and status report on Jefferson Salamander (Ambystoma jeffersonianum) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 20 pp. (Également disponible en français : COSEPAC. 2000. Évaluation et Rapport de situation du COSEPAC sur la salamandre de Jefferson (Ambystoma jeffersonianum) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. v + 22 p.)
COSEWIC. 2004. COSEWIC assessment and update status report on the Small-mouthed Salamander Ambystoma texanum in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. v + 20 pp. (Également disponible en français : COSEPAC. 2004. Évaluation et Rapport de situation du COSEPAC sur la salamandre à nez court (Ambystoma texanum) au Canada - Mise à jour. Comité sur la situation des espèces en péril au Canada. Ottawa. v + 20 p.)
COSEWIC. 2010. COSEWIC assessment and status report on the Jefferson SalamanderAmbystoma jeffersonianum in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. xi + 38 pp. (Également disponible en français : COSEPAC. 2010. Évaluation et Rapport de situation du COSEPAC sur la salamandre de Jefferson(Ambystoma jeffersonianum) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xii + 40 p.)
COSEWIC. 2014. COSEWIC status appraisal summary on Small-mouthed Salamander (Ambystoma texanum) in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 10 pp (i-x). (Également disponible en français : COSEPAC. 2014. Sommaire du statut de l'espèce du COSEPAC sur la salamandre à petite bouche (Ambystoma texanum) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xi p.)
COSEWIC. 2015a. Definitions and Abbreviations. Approved by COSEWIC in November 2013. Minor revisions approved by COSEWIC in November 2015. Committee on the Status of Endangered Wildlife in Canada. [consulté en avril 2016]. (Également disponible en français : COSEPAC. 2015a. Définitions et abréviations. Approuvées par le COSEPAC en novembre 2013. Comité sur la situation des espèces en péril au Canada.)
COSEWIC. 2015b. COSEWIC guidelines for naming wildlife species. Approved by COSEWIC in April 2010. Minor revisions approved by COSEWIC in November 2013 and 2015. Committee on the Status of Endangered Wildlife in Canada. [consulté en avril 2016]. (Également disponible en français : COSEPAC. 2015b. Lignes directrices du COSEPAC pour l'appellation des espèces sauvages. Approuvées par le COSEPAC en avril 2010. Révisions mineures approuvées par le COSEPAC en novembre 2013 et 2015. Comité sur la situation des espèces en péril au Canada.)
Crother, B.I. (ed.). 2012. Scientific and Standard English Names of Amphibians and Reptiles of North America North of Mexico, with Comments Regarding Confidence in Our Understanding, 7th Edition. Herpetological Circular No. 39. Society for Study of Amphibians and Reptiles, Salt Lake City, Utah. 101 pp.
Dawley, R.M. et J.P. Bogart, 1989. Evolution and Ecology of Unisexual Vertebrates. Bulletin 466, New York State Museum, Albany, New York. 302 pp.
Dawley, E.M. et R.M. Dawley. 1986. Species discrimination by chemical cues in a unisexual-bisexual complex of salamanders. Journal of Herpetology 20(1):114-116
Dobbyn, S. 2004 (données inédites). Correspondance par courriel de J. Bogart adressée à Sandy Dobbyn. 30 novembre 2004. Natural Heritage Education Specialist, Rondeau Provincial Park. RR#1 Kent Road 17, Morpeth, Ontario.
Douglas, M.E. et B.L. Monroe, Jr. 1981. A comparative study of topographical orientation in Ambystoma (Amphibia: Caudata). Copeia 1981:460-463.
Downs, F. 1989. Ambystoma jeffersonianum. Pp. 87-172, in R.A. Pfingsten et F.L. Downs. (eds.). Salamanders of Ohio. Ohio Biological Survey Bulletin, New Series Vol. 7, No. 2.
Ducey, P.K. et E.D. Brodie, Jr. 1983. Salamanders respond selectively to contacts with snakes: survival advantages of alternative antipredator strategies. Copeia 1983:1036-1041.
Ecoservices 2007 (données inédites). Correspondance par courriel de J. Bogart adressée à Les Minch. 27 avril 2007. Principle/Senior Ecologist, Ecoservices Inc. Ecological and Environmental Consulting Services, 1-265 Frobisher Drive, Waterloo, Ontario.
Environment Canada. 2015. Recovery Strategy for the Jefferson Salamander (Ambystoma jeffersonianum) in Canada [Proposed]. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa. 26 pp. + Annexes. (Également disponible en français : Environnement Canada. 2015. Programme de rétablissement de la salamandre de Jefferson (Ambystoma jeffersonianum) au Canada [Proposition]. Série de programmes de rétablissement de la Loi sur les espèces en péril. Environnement Canada, Ottawa. 28 p. + annexe.)
Faccio, S.D. 2003. Post breeding emigration and habitat use by Jefferson and Spotted salamanders in Vermont. Journal of Herpetology 37:479-489.
Feder, M.E., J.F. Lynch, H.B. Shaffer et D.B. Wake. 1982. Field body temperatures of tropical and temperate zone salamanders. Smithsonian Herpetological Information Service 52:1-23.
Flageole, S. et R. Leclair, Jr. 1992. Étude démographique d'une population de salamandres (Ambystoma maculatum) à l'aide de la méthode squeletto-chronologique. Canadian Journal of Zoology 70:740-749.
Gibbs, J.P. et W.G. Shriver. 2005. Can road mortality limit populations of pool-breeding amphibians? Wetlands Ecology and Management 13:281-289.
Gilhen, J. 1974. Distribution, natural history and morphology of the Blue-spotted salamanders, Ambystoma laterale and A. tremblayi in Nova Scotia. Nova Scotia Museum Curatorial Report 22:1-38.
Greenwald, K.R. et H.L. Gibbs. 2012. A single nucleotide polymorphism assay for the identification of unisexual Ambystoma salamanders. Molecular Ecology Resources 12:354-362.
Government of Ontario. 2007. Endangered Species Act., 2007, S.O. 2007. (Également disponible en français : Gouvernement de l'Ontario. 2007. Loi de 2007 sur les espèces en voie de disparition, L.O. 2007.).
Hamill, S.E. 2014. DRAFT Recovery Strategy for the Small-mouthed Salamander (Ambystoma texanum) in Ontario. Ontario Recovery Strategy Series. Prepared for the Ontario Ministry of Natural Resources, Peterborough, Ontario. vii + 18 pp.
Hedges, S.B., J.P. Bogart et L.R. Maxson. 1992. Ancestry of unisexual salamanders. Nature 356:708-710.
Heyer, W.R., M.A. Donnelly, R.W. McDiarmid, L.C. Hayek et M.S. Foster 1994. Measuring and Monitoring Biological Diversity. Standard Methods for Amphibians. Smithsonian Institution Press, Washington DC.
Hubbs, C.L. et L.C. Hubbs 1932. Apparent parthenogenesis in nature, in a form of fish of hybrid origin. Science 76:628-630.
Jefferson Salamander Recovery Team. 2010. Recovery Strategy for the Jefferson Salamander (Ambystoma jeffersonianum) in Ontario. Ontario Recovery Strategy Series. Prepared for the Ontario Ministry of Natural Resources, Peterborough, Ontario. Vi + 27 pp.
Julian, S.E., T.L. King et W.K. Savage. 2003. Novel Jefferson salamander, Ambystoma jeffersonianum, microsatellite DNA markers detect population structure and hybrid complexes. Molecular Ecology Notes 3:95-97.
Karraker, N.E., J.P. Gibbs et J.R. Vonesh. 2008. Impacts of road deicing salt on the demography of vernal pool-breeding amphibians. Ecological Applications 18:724-734.
Klemens. M.W. 2000. Amphibians and reptiles in Connecticut: a checklist with notes on conservation status, identification, and distribution. DEP Bulletin 32: 1-96, Hartford, Connecticut: Connecticut Department of Environmental Protection, Environmental and Geographic Information Center.
Kraus, F. 1985a. A new unisexual salamander from Ohio. Occasional Papers of the Museum of Zoology, University of Michigan 709:1-24
Kraus, F. 1985b. Unisexual salamander lineages in northwestern Ohio and southern Michigan: a study of the consequences of hybridization. Copeia 1985:309-324.
Kraus, F. 1995. The conservation status of unisexual vertebrate populations. Conservation Biology 9:956-959.
Licht, L.E. 1989. Reproductive parameters of unisexual Ambystoma on Pelee Island, Ontario. Pp. 209-217, in R. Dawley et J.P. Bogart (eds.). Evolution and Ecology of Unisexual Vertebrates. New York State Museum Bull. 466. Albany, New York.
Licht, L.E. comm. pers. 2015. Correspondance par courriel adressée à J. Bogart. 12 janvier 2015. Professeur de zoologie (retraité), York University, Toronto, Ontario.
Licht, L.E. et J.P. Bogart. 1987. Ploidy and developmental rate in a salamander hybrid complex. Evolution 41:918-920.
Licht, L.E. et J.P. Bogart. 1989. Growth and sexual maturation in diploid and polyploidy salamanders (genus Ambystoma). Canadian Journal of Zoology 67:812-818.
Licht, L.E. et J.P. Bogart. 1990. Courtship behavior of Ambystoma texanum on Pelee Island, Ontario. Journal of Herpetology 24:450-452.
Lamond, W.G. 1994. The Reptiles and Amphibians of the Hamilton area. An Historical Summary and the Results of the Hamilton Herpetofaunal Atlas. Hamilton Naturalist Club, Hamilton, Ontario. 174 pp.
LGL 2007 (données inédites). Correspondance par courriel de J. Bogart adressée à Allison Featherstone. 26 avril 2007. Senior Planning Ecologist, Manager Cambridge Office LGL Limited environmental research associates 445 Thompson Drive, Unit 2 Cambridge Ontario N1T 2K7.
LGL. 2008 (données inédites). Correspondance par courriel de J. Bogart adressée à Allison Featherstone. 18 novembre 2008. Senior Planning Ecologist, Manager Cambridge Office LGL Limited environmental research associates 445 Thompson Drive, Unit 2 Cambridge Ontario N1T 2K7.
Logier, E.B.S. et G.C. Toner. 1961. Check List of the Amphibians & Reptiles of Canada & Alaska. Life Science Division, Royal Ontario Museum, Toronto. Contribution No. 53:1-92.
Lowcock, L.A. 1989. Biogeography in hybrid complexes of Ambystoma: the interpretation of unisexual-bisexual genetic data in space and time. Pp. 180-207, in R. Dawley et J.P. Bogart (eds.). Evolution and Ecology of Unisexual Vertebrates. New York State Museum Bull. 466. Albany, New York.
Lowcock, L.A. 1991. Evolution, ecology and the nature of bisexual-unisexual communities: The Ambystoma laterale-jeffersonianum complex in Central Ontario. Thèse de doctorat. University of Toronto, Ontario. Xviii + 197 pp.
Lowcock, L.A. 1994. Biotype, genomotype, and genotype: variable effects of polyploidy and hybridity on ecological partitioning in a bisexual-unisexual community of salamanders. Canadian Journal of Zoology 72:104-117.
Lowcock, L.A., H. Griffith et R.W. Murphy. 1991. The Ambystoma laterale-jeffersonianum complex in Central Ontario: Ploidy structure, sex ratio and breeding dynamics in bisexual-unisexual communities. Copeia 1991:69-88.
Lowcock, L.A., H. Griffith et R.W. Murphy. 1992. Size in relation to sex, hybridity, ploidy, and breeding dynamics in central Ontario populations of the Ambystoma laterale-jeffersonianum complex. Journal of Herpetology 26:46-53.
Lowcock, L.A., L.E. Licht et J.P. Bogart. 1987. Nomenclature in hybrid complexes of Ambystoma (Urodela: Ambystomatidae): no case for the erection of hybrid "species". Systematic Zoology: 36:328-336.
Lowcock, L.A. et R.W. Murphy 1991. Pentaploidy in a hybrid vertebrate demonstrates enhanced tolerance of multiple chromosome sets. Experientia 47:490-493
Macgregor, H.C. et T.M. Uzzell. 1964. Gynogenesis in salamanders related to Ambystoma jeffersonianum. Science 143:1043-1045.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay et A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk. NatureServe, Arlington, Virginia. [Disponible en anglais seulement, consulté en août 2014].
Minton, S.A. 1954. Salamanders of the Ambystoma jeffersonianum complex in Indiana. Herpetologica 10:173-179.
Mohr, C.E. 1931. Observations on the early breeding habits of Ambystoma jeffersonianum in central Pennsylvania. Copeia 1931:102-104.
Mullen, S. J. et S. Klueh. 2009. Demographics of a geographically isolated population of a threatened salamander (Caudata: Ambystomatidae) in central Illinois. Herpetolocial Conservation and Biology 4:261- 269.
NatureServe. 2014. NatureServe Explorer: An online encyclopedia of life [application Web] Version 7.1. NatureServe, Arlington, Virginia. [Disponible en anglais seulement, consulté le 30 octobre 2014].
Noël S., J. Dumoulin, M. Ouellet, P. Galois et F-J Lapointe. 2008. Rapid identification of salamanders from the Jefferson complex with taxon-specific primers. Copeia 2008:158-161.
Noël, S., P. Labonté et F-J. Lapointe. 2011. Genotype frequencies and genetic diversity in urban and protected populations of Blue-spotted salamanders (Ambystoma laterale) and related unisexuals. Journal of Herpetology 45:294-299.
NRSI. 2009 (données inédites). Correspondance par courriel de J. Bogart adressée à Jessica Linton. 24 juin 2009. Terrestrial and Wetland Biologist, Natural Resource Solutions Inc., 225 Labrador Drive, Unit 1, Waterloo, Ontario.
Nyman, S. 1991. Ecological aspects of syntopic larvae of Ambystoma maculatum and the A. laterale-jeffersonianum complex in two New Jersey ponds. Journal of Herpetology 25:505-509.
OMNR. 2007 (données inédites). Correspondance par courriel de J. Bogart adressée à John Pisapio. 20 juin 2007. Wildlife Biologist, Ontario Ministry of Natural Resouces, Aurora District.
Petranka, J.W. 1998. Salamanders of the United States and Canada. Smithsonian Institution Press. 587pp.
Phillips, C.A., T. Uzzell, C.M. Spolski, J.M. Serb, R.E. Szafoni et T.R. Pollowy. 1997. Persistent high levels of polyploidy in salamanders of the Ambystoma jeffersonianum complex. Journal of Herpetology 31:530-535.
Piersol, W.H. 1910. Spawn and larvae of Ambystoma jeffersonianum. American Naturalist 44:732-738.
Ramsden, C. 2008. Population genetics of Ambystoma jeffersonianum and sympatric unisexuals reveal signatures of both gynogenetic and sexual reproduction. Copeia 2008:586-594.
Ramsden, C., K. Bériault et J.P. Bogart. 2006. A non-lethal method of identification for Ambystoma laterale, A. jeffersonianum and sympatric unisexuals. Molecular Ecology Notes 6:261-264.
Raymond, L.R. et L.M. Hardy. 1990. Demography of a population of Ambystoma talpoideum (Caudata: Ambystomatidae) in northwestern Louisiana. Herpetologica 46:371-382.
Reed, D.H., J.J. O'Grady, B.W. Brook, J.D. Ballou et R. Frankam. 2003. Estimates of minimum viable population sizes for vertebrates and factors influencing those estimates. Biological Conservation 113:23-34.
Sandinski, W.J. et W.A. Dunson. 1992. A multilevel study of effects of low pH on amphibians in temporary ponds. Herpetologica 26:413-422.
Schoop, C.R. 1965. Orientation of Ambystoma maculatum: movement to and from breeding pools. Science 149:558-559.
Schultz, R.J. 1969. Hybridization, unisexuality, and polyploidy in the teleost Poeciliopsis (Poeciliidae) and other vertebrates. American Naturalist 103:605-619.
Schultz, R.J. 1977. Evolutionary ecology of unisexual fishes. Pp. 277-331, in M.K. Hecht, W.C. Steere et B. Wallace (eds.). Evolutionary Biology, Vol. 10. Plenum Press, New York.
Smith, C.K. 1983. Notes on breeding period, incubation period, and egg masses of Ambystoma jeffersonianum (Green) (Amphibia: Caudata) from the southern limits of its range. Brimleyana 9:135-140.
Smith, C.K., et J.W. Petranka. 1987. Prey size distributions and size-specific foraging success of Ambystoma larvae. Oecologia 71:239-244.
Spolski, C., C.A. Phillips et T. Uzzell. 1992. Antiquity of clonal salamander lineages revealed by mitochondrial DNA. Nature 356:706-708.
Taylor, A.S. et J.P. Bogart, 1990. Karyotypic analyses of four species of Ambystoma (Amphibia, Caudata) which have been implicated in the production of all-female hybrids. Genome 33:837-844.
Thompson, E.L., J.E. Gates et G.S. Taylor. 1980. Distribution and breeding habitat selection of the Jefferson salamander, Ambystoma jeffersonianum, in Maryland. Journal of Herpetology 14:113-120.
Turtle, S.L. 2000. Embryonic survivorship of the spotted salamander (Ambystoma maculatum) in roadside and woodland vernal pools in southeastern New Hampshire. Journal of Herpetology 34:60-67.
Uzzell, T.M. 1964. Relations of the diploid and triploid species of the Ambystoma jeffersonianum complex (Amphibia, Caudata). Copeia 1964:257-300.
Uzzell, T. 1969. Notes on spermatophore production by salamanders in the Ambystoma jeffersonianum complex. Copeia 1969:602-612.
Vrijenhoek R.C., R.M. Dawley, C.J. Cole et J.P. Bogart. 1989. A list of the known unisexual vertebrates. Pp. 19-23, in R. Dawley et J.P. Bogart (eds.). Evolution and Ecology of Unisexual Vertebrates. New York State Museum Bull. 466. Albany, New York.
Weller, W. F. 1980. Migration of the salamanders Ambystoma jeffersonianum (Green) and A. platineum (Cope) to and from a spring breeding pond, and the growth, development and metamorphosis of their young. Mémoire de maîtrise. University of Toronto, Ontario. 248 pp.
Wilbur, H.M. 1971. The ecological relationship of the salamander Ambystoma laterale to its all-female, gynogenetic associate. Evolution 25:168-179.
Wilbur, H.M. 1976. A sequential sampling procedure for identifying triploid salamanders. Copeia 1976:391-392.
Wildlife Research Station. 2010 (données inédites). Correspondance par courriel de J. Bogart adressée à Patrick Moldowan. 21 mars 2010. Étudiant diplômé, Department of Biology, Lakehead University, Thunder Bay, Ontario.
Wildlife Research Station. 2011 (données inédites). Correspondance par courriel de J. Bogart adressée à Patrick Moldowan. 1er février 2011. Étudiant diplômé, Department of Biology, Lakehead University, Thunder Bay, Ontario.
Williams, R.N. et J.A. DeWoody. 2004. Fluorescent dUTP helps characterize 10 novel tetranucleotide microsatellites from an enriched salamander (Ambystoma texanum) genomic library. Molecular Ecology Notes 4:17-19.
Sommaire biographique du rédacteur du rapport
James (Jim) Bogart est professeur émérite au Département de biologie intégrative de l'Université de Guelph. Il étudie l'évolution au niveau des espèces par des travaux de biologie moléculaire, de cytogénétique et d'écologie, et s'intéresse particulièrement aux amphibiens polyploïdes. Il étudie les Ambystoma unisexués et leurs espèces donneuses de sperme depuis 1975. Il a conseillé plusieurs étudiants de cycles supérieurs qui ont étudié divers aspects de ces salamandres au Canada et aux États-Unis.
Collections examinées
La plupart des salamandres unisexuées et des espèces donneuses de sperme qui leur sont associées provenant de l'est des États-Unis qui ont été examinées se trouvent à l'American Museum of Natural History (AMNH). Les données et les numéros de catalogue pour ces spécimens sont présentés dans Bogart et Klemens (1997, 2008). Les spécimens de l'Ohio proviennent de collections de l'Université de Dayton et de l'Université d'État de l'Ohio. Les spécimens provenant de l'Indiana sont conservés par R. Brodman (Ph. D.), du Saint Joseph's College, à Rensselaer, en Indiana. Les échantillons du Michigan sont entreposés au Museum of Vertebrate Zoology à l'Université du Michigan. Les spécimens de référence canadiens sont gardés au Musée royal de l'Ontario ou au Musée canadien de la nature. Une collection d'échantillons gelés de tissus et d'ADN est cataloguée et gardée à l'Université de Guelph.
Annexe 1. Hausse et baisse de la ploïdie
Hausse de la ploïdie
Le niveau de ploïdie typique chez les Ambystoma unisexués est triploïde, mais des individus diploïdes, tétraploïdes et pentaploïdes ont été trouvés dans leur grande aire de répartition. Les individus pentaploïdes sont très rares (Lowcock et Murphy, 1991), et les individus tétraploïdes sont plus communs que les individus diploïdes (Phillips et al., 1997; Bogart et Klemens, 1997, 2008) (tableau 1). Dans des croisements en laboratoire (Bogart et al., 1989), des femelles triploïdes ont donné naissance à des petits triploïdes et tétraploïdes, et des femelles tétraploïdes ont produit des petits tétraploïdes et pentaploïdes. On a constaté une augmentation considérable du nombre de petits tétraploïdes nés de mères triploïdes lorsqu'on a fait passer la température de 6 °C à 15 °C. Ces données montrent que la hausse de la ploïdie représente un phénomène assez courant chez les Ambystoma unisexués et qu'elle pourrait être liée à des facteurs extrinsèques, comme la température. Quand on vérifie plusieurs loci microsatellites chez la progéniture d'une femelle unisexuée, on constate que les individus gynogénétiques présentent le même génotype multilocus (MLG) que leur mère (clones). La progéniture à ploïdie élevée dispose d'un allèle supplémentaire dans les loci où le génome issu du sperme possède des allèles différents. Il est possible de génotyper le mâle donneur de sperme d'une femelle unisexuée triploïde d'après la progéniture tétraploïde qui est produite (Bogart et Bi, 2013).
Les gènes des individus bisexués ont probablement été triés par de nombreuses générations de sélection naturelle et, en théorie, devraient représenter les « meilleurs » gènes pour l'environnement contemporain. Théoriquement, en s'appropriant de tels gènes (et chromosomes), les salamandres unisexuées cooccurrentes pourraient contourner de longues périodes de sélection naturelle, et être ainsi mieux adaptées à la compétition avec les salamandres bisexuées. La reproduction gynogénétique subséquente éviterait la perte de 50 % de « bons gènes » à laquelle on peut s'attendre dans le cadre de la méiose normale. Bien que les données empiriques montrent qu'il existe une certaine hausse génomique dans la plupart des populations de salamandres unisexuées (Bogart, 2003), la hausse de la ploïdie a probablement une limite supérieure : probablement la pentaploïdie (Lowcock et Murphy, 1991).
Baisse de la ploïdie et remplacement génomique
Le problème de l'augmentation constante des niveaux de ploïdie par le processus de hausse de la ploïdie pourrait être atténué dans le cas où des jeux de chromosomes peuvent être éliminés en plus d'être ajoutés. D'un point de vue évolutionnaire, il pourrait être avantageux pour un individu unisexué de conserver un jeu de chromosomes qui offre un avantage sélectif au sein d'une population contemporaine et d'éliminer un jeu de chromosomes désavantageux. Les données sur les microsatellites (Bogart et al. 2007, 2009; Ramsden, 2008; Nöel et al., 2011) montrent que les salamandres unisexuées diploïdes, triploïdes et tétraploïdes ont en commun des allèles d'ADN microsatellite avec leurs espèces donneuses de sperme sympatriques. Il importe de noter, car il s'agit sans doute d'un point important, que tous les génomotypes unisexués (Bogart, 2003; Bogart et al., 2009), qui sont plus de 20, contiennent au moins un jeu de chromosomes de la salamandre à points bleus (A. laterale). Le génome de l'A. laterale a été ciblé par Bi et al. (2008), qui en ont établi la généalogie d'après les séquences d'un marqueur d'ADN nucléaire spécifique et variable de l'A. laterale (L-G1C12). Ils ont constaté que les salamandres unisexuées avaient toujours le même haplotype que l'A. laterale dans les populations sympatriques dans la majeure partie de l'aire de répartition des salamandres unisexuées. Une autre hypothèse, voulant qu'un génome « ancien » de l'Ambystoma laterale persiste dans l'ensemble des diverses combinaisons de génomes unisexués, a été rejetée.
Bogart et Bi (2013) pensent que la baisse de la ploïdie et le remplacement génomique sont liés. Un tel lien nécessite (parfois) la sympatrie entre salamandres à points bleus et une autre espèce donneuse de sperme pour produire des individus tétraploïdes symétriques (ploïdie accrue). Les événements possibles sont illustrés à la figure 9. Ces conditions et biotypes sont présents sur l'île Pelée (tableau 1). En théorie, la production d'individus unisexués diploïdes LT (ploïdie réduite), par un individu tétraploïde symétrique LLTT pourrait se faire en omettant simplement l'étape préméiotique d'endoduplication, nécessaire pour la méiose triploïde (Bogart, 2003). L'incorporation de sperme dans un œuf LT à ploïdie non réduite a pour effet de hausser la ploïdie au niveau triploïde et explique le remplacement génomique. Un génome L (d'une femelle de niveau A) est remplacé par un nouveau génome L ou T chez les individus de niveau D. Un tel scénario pourrait aussi expliquer la baisse de la ploïdie et le remplacement génomique chez les individus unisexués qui coexistent avec les salamandres à points bleus et de Jefferson dans les régions continentales.
Le remplacement génomique a des conséquences sur les unités désignables (UD). On pourrait affirmer que l'Ambystoma unisexué représente une seule UD, et que les individus gagnent et perdent des génomes au hasard dans leur aire de répartition. Avec des modèles de simulation, Charney (2012) a effectué le suivi du résultat prévu pour les salamandres unisexuées qui avaient subi une hausse de la ploïdie et un remplacement génomique au fil du temps. Il a conclu que les génomes nucléaires finiraient par devenir impossibles à distinguer de ceux des hôtes sur une période relativement courte, déterminée par la fréquence du remplacement génomique qu'il considère comme égale à la fréquence de la hausse de la ploïdie. Le résultat logique serait des salamandres de Jefferson ou à points bleus diploïdes ou triploïdes possédant un ADNmt unisexué. Le fait que de tels individus n'ont jamais été trouvés, selon Charney, résulterait d'un effort d'échantillonnage inadéquat.
S'il est vrai que l'ADN extrait aux fins de l'identification des génomotypes bisexués et unisexués n'est pas toujours utilisé pour l'analyse des séquences d'ADNmt, les données sur ces séquences sont disponibles dans l'aire de répartition des salamandres unisexuées et de leurs hôtes bisexués (Hedges et al., 1992; Bogart, 2003; Bogart et al., 2007, 2009; Nöel et al., 2008, 2011; Bi et Bogart, 2010; Greenwald et Gibbs, 2012; Bi et Bogart, données inédites). Si le mécanisme de remplacement génomique exige des individus tétraploïdes symétriques, comme l'ont avancé Bogart et Bi (2013) (voir figure 9), un individu unisexué utiliserait le « mauvais » donneur de sperme. Par exemple, un individu unisexué LJJ, qui utilisait habituellement l'A. jeffersonianum comme donneur de sperme, a été inséminé de sperme d'un A. laterale. La progéniture gynogénétique serait toujours LJJ, mais la progéniture à ploïdie accrue serait LLJJ. Si les salamandres de Jefferson disparaissaient et les salamandres à points bleus persistaient, on pourrait s'attendre à ce que la fréquence des individus LLJ augmente par rapport à celle des LJJ, mais il faudrait probablement beaucoup de temps avant que les LLJ évincent les LJJ. En Ontario, il y a très peu de régions où les salamandres à points bleus et les salamandres de Jefferson sont sympatriques ou parapatriques, et c'est dans de telles régions, généralement perturbées, que l'on trouve les rares individus unisexués LLJJ et LJ. Les salamandres de Jefferson et les salamandres à points bleus ont des préférences différentes en matière d'habitat (Petranka, 1998), ce qui pourrait expliquer pourquoi on les trouve rarement ensemble; cette réalité s'étend probablement aux UD unisexuées qui leur sont associées et que l'on trouve aussi très rarement ensemble.
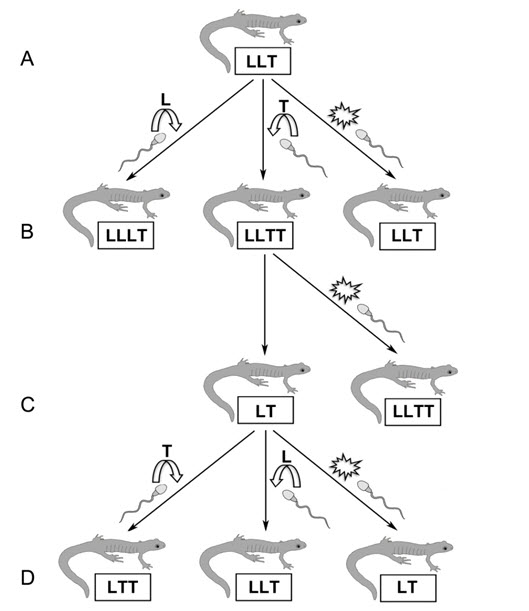
Description longue de la figure 9
Diagramme illustrant les événements hypothétiques pouvant expliquer la baisse et le remplacement génomiques chez les salamandres unisexuées de l'île Pelée (Ontario).
Annexe 2. Tableau d'évaluation des menaces, y compris les commentaires, pour l'Ambystoma unisexué dépendant de la salamandre à petite bouche (UD 1).
Tableau d'évaluation des menaces
- Nom scientifique de l'espèce ou de l'écosystème
- Ambystoma unisexué, population dépendante de l'A. texanum
- Date
- 2/2/2015
- Évaluateur(s) :
- Jim Bogart (rédacteur du rapport), Kristiina Ovaska (facilitatrice), Bev McBride (Secrétariat du COSEPAC), Mary Sabine, Leslie Anthony, Isabelle Gauthier, Yohann Dubois, Joe Crowley
- Références :
- Rapport de situation du COSEPAC (ébauche de 2015)
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 1 | 1 |
| C | Moyen | 0 | 0 |
| D | Faible | 4 | 4 |
| - | Impact global des menaces calculé : | Élevé | Élevé |
- Impact global des menaces attribué :
- Élevé
- Commentaires sur l'impact global des menaces
- Durée d'une génération - 8 ans (24 ans pour 3 générations); env. 1 000 adultes; zone d'occurrence et IZO = 20 km 2
| Menace | Menace | Impact (calculé) | Impact (calculé) | Portée (10 prochaines années) | Gravité (10 ans ou 3 générations) | Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | D | Faible | Petite (1-10 %) | Élevée (31-70 %) | Modérée (peut-être à court terme, < 10 ans ou 3 générations) | - |
| 1.1 | Zones résidentielles et urbaines | D | Faible | Petite (1-10 %) | Élevée (31-70 %) | Modérée (peut-être à court terme, < 10 ans ou 3 générations) | Une certaine expansion résidentielle a lieu sur l'île Pelée. Bien que cette expansion ne touche aucun habitat connu des salamandres, une telle possibilité persiste. Comme la population est très faible, même un seul projet dans l'habitat des salamandres pourrait être problématique (portée de > 1 %). La gravité des effets dépend du drainage des étangs de reproduction (extrême); si seul l'habitat terrestre est touché, la gravité est réduite. |
| 1.2 | Zones commerciales et industrielles | - | - | - | - | - | - |
| 1.3 | Zones touristiques et récréatives | - | - | - | - | - | L'aménagement d'un nouveau sentier est possible (pointe Lighthouse), mais d'autres expansions sont peu probables. |
| 2 | Agriculture et aquaculture | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Modérée (peut-être à court terme, < 10 ans ou 3 générations) | - |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Modérée (peut-être à court terme, < 10 ans ou 3 générations) | L'étang Middle, au centre de l'île, est situé à proximité d'un champ agricole (l'étang est riche et abrite probablement 5 % de la population de salamandres sur l'île). D'autres aménagements agricoles sont possibles. |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | - |
| 2.3 | Élevage de bétail | - | - | - | - | - | - |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | - |
| 3 | Production d'énergie et exploitation minière | - | Non calculé (immédiateté inconnue) | Petite (1-10 %) | Extrême (71-100 %) | Inconnue | - |
| 3.1 | Forage pétrolier et gazier | - | - | - | - | - | - |
| 3.2 | Exploitation de mines et de carrières | - | Non calculé (immédiateté inconnue) | Petite (1-10 %) | Extrême (71-100 %) | Inconnue | Une ancienne carrière abritant des salamandres (étang North) pourrait redevenir active dans l'avenir, mais de tels plans sont inconnus. |
| 3.3 | Énergie renouvelable | - | - | - | - | - | - |
| 4 | Corridors de transport et de service | D | Faible | Petite (1-10 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 4.1 | Routes et voies ferrées | D | Faible | Petite (1-10 %) | Élevée (31-70 %) | Élevée (continue) | La plupart des étangs à salamandres sont situés dans l'intérieur de l'île, plutôt qu'en périphérie, où se trouve la majorité des routes. Parmi les répercussions des routes, on compte les obstacles aux migrations de reproduction et la mortalité routière. Il y a des étangs de chaque côté de la route dans le sud de l'île, et des salamandres qui traversent peuvent être tuées (~ 5 % de la population utilise ces étangs). Les répercussions sur la population sont incertaines, mais elles pourraient être graves. La circulation routière est relativement faible sur l'île Pelée durant la période de reproduction. |
| 4.2 | Lignes de services publics | - | - | - | - | - | - |
| 4.3 | Voies de transport par eau | - | - | - | - | - | - |
| 4.4 | Corridors aériens | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques | - | - | - | - | - | - |
| 5.1 | Chasse et capture d'animaux terrestres | - | - | - | - | - | Rien n'indique qu'une collecte ciblée a lieu; voir Activités récréatives |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | - |
| 5.3 | Exploitation forestière et récolte du bois | - | - | - | - | - | - |
| 5.4 | Pêche et récolte de ressources aquatiques | - | - | - | - | - | - |
| 6 | Intrusions et perturbations humaines | D | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | - |
| 6.1 | Activités récréatives | D | Faible | Petite (1-10 %) | Légère (1-10 %) | Élevée (continue) | Ce facteur n'est problématique que dans le secteur de la pointe Fish où se rendent les touristes; ces zones sont très utilisées, mais probablement pas durant la période de reproduction lorsque les salamandres sont les plus vulnérables. Des perturbations (piétinement des salamandres et de leur habitat) résultent de la présence de personnes photographiant et recherchant les salamandres dans les boisés et les étangs; les observateurs d'oiseaux et d'autres touristes se rendent aussi dans les habitats des salamandres. |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | - |
| 6.3 | Travail et autres activités | - | - | - | - | - | - |
| 7 | Modifications des systèmes naturels | B | Élevé | Grande (31-70 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | - |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | B | Élevé | Grande (31-70 %) | Élevée (31-70 %) | Élevée (continue) | Il s'agit probablement de la plus grande menace pour cette population. Les fossés de drainage modifient les niveaux d'eau dans les étangs à salamandres : p. ex. le dragage de canaux a lieu juste à côté de la route à la pointe Fish, et entraîne l'eau des étangs de reproduction; ces perturbations peuvent aussi avoir lieu dans toute l'île (la seule population non touchée est celle de la carrière nord). Le niveau d'eau du lac Érié varie grandement, et les salamandres sont mieux adaptées aux niveaux élevés. L'habitat ne fait pas l'objet d'une protection juridique contre les modifications liées au dragage et au drainage en Ontario, sauf aux termes de la Loi de 2007 sur les espèces en voie de disparition en ce qui concerne l'Ambystoma texanum (en voie de disparition). |
| 7.3 | Autres modifications de l'écosystème | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | Élevée (continue) | Le déboisement et l'enlèvement des débris ligneux grossiers ont surtout lieu dans le secteur de la pointe Fish. On s'attend principalement à ce que cette menace touche les salamandres juvéniles (les adultes utilisent des terriers). De plus, les dindons introduits modifient les conditions du tapis forestier, et leurs effets sont largement inconnus (la prédation par les dindons est prise en compte sous Espèces introduites). Remarque : la perte de populations d'espèces donneuses de sperme dans le cadre d'événements stochastiques n'est pas prise en compte ici; une telle perte accompagnerait probablement d'autres menaces qui pèsent tant sur les individus unisexués que sur les donneurs de sperme. |
| 8 | Espèces et gènes envahissants ou autrement problématiques | - | Inconnu | Généralisée (71-100 %) | Inconnue | Élevée (continue) | - |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | - | Inconnu | Généralisée (71-100 %) | Inconnue | Élevée (continue) | La prédation possible par les dindons sauvages introduits est préoccupante. On ne dispose d'aucune information sur l'infection par le champignon chytride ou d'autres maladies. Le roseau commun (Phragmites australis) envahissant est peu susceptible de toucher les salamandres. |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | - | - |
| 8.3 | Matériel génétique introduit | - | - | - | - | - | - |
| 9 | Pollution | - | Inconnu | Restreinte (11-30 %) | Inconnue | Élevée (continue) | - |
| 9.1 | Eaux usées domestiques et urbaines | - | - | - | - | - | - |
| 9.2 | Effluents industriels et militaires | - | - | - | - | - | - |
| 9.3 | Effluents agricoles et sylvicoles | - | Inconnu | Restreinte (11-30 %) | Inconnue | Élevée (continue) | Ruissellement en provenance des vignobles et des champs agricoles (pâturages?), particulièrement autour de l'étang Middle. La portée et la gravité de la menace sont incertaines. Il y a aussi de l'incertitude quant aux produits chimiques et aux quantités utilisées. |
| 9.4 | Déchets solides et ordures | - | - | - | - | - | - |
| 9.5 | Polluants atmosphériques | - | - | - | - | - | - |
| 9.6 | Apports excessifs d'énergie | - | - | - | - | - | - |
| 10 | Phénomènes géologiques | - | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | - | - |
| 11 | Changements climatiques et phénomènes météorologiques violents | - | Non calculé (à l'extérieur de la période d'évaluation) | Généralisée (71-100 %) | Élevée (31-70 %) | Faible (peut-être à long terme, > 10 ans ou 3 générations) | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | - |
| 11.2 | Sécheresses | - | Non calculé (à l'extérieur de la période d'évaluation) | Généralisée (71-100 %) | Élevée (31-70 %) | Faible (peut-être à long terme, > 10 ans ou 3 générations) | L'assèchement hâtif des étangs de reproduction est préoccupant, et aggrave les répercussions du prélèvement d'eau par les humains. Le niveau d'eau du lac Érié (voir Gestion de l'eau) fluctue pour des raisons inconnues. Tous les sites de reproduction des salamandres sauf un sont des étangs temporaires, et seraient touchés par les changements climatiques. On ne sait pas exactement s'il y a des prévisions propres à la région. |
| 11.3 | Températures extrêmes | - | - | - | - | - | - |
| 11.4 | Tempêtes et inondations | - | - | - | - | - | - |
Classification des menaces d'après l'IUCN-CMP, Salafsky et al. (2008).
Glossaire
- Impact
- Mesure dans laquelle on observe, infère ou soupçonne que l'espèce est directement ou indirectement menacée dans la zone d'intérêt. Le calcul de l'impact de chaque menace est fondé sur sa gravité et sa portée et prend uniquement en compte les menaces présentes et futures. L'impact d'une menace est établi en fonction de la réduction de la population de l'espèce, ou de la diminution ou de la dégradation de la superficie d'un écosystème. Le taux médian de réduction de la population ou de la superficie pour chaque combinaison de portée et de gravité correspond aux catégories d'impact suivantes : très élevé (déclin de 75 %), élevé (40 %), moyen (15 %) et faible (3 %). Inconnu : catégorie utilisée quand l'impact ne peut être déterminé (p. ex. lorsque les valeurs de la portée ou de la gravité sont inconnues); non calculé : l'impact n'est pas calculé lorsque la menace se situe en dehors de la période d'évaluation (p. ex. l'immédiateté est insignifiante/négligeable ou faible puisque la menace n'existait que dans le passé); négligeable : lorsque la valeur de la portée ou de la gravité est négligeable; n'est pas une menace : lorsque la valeur de la gravité est neutre ou qu'il y a un avantage possible.
- Portée
- Proportion de l'espèce qui, selon toute vraisemblance, devrait être touchée par la menace d'ici 10 ans. Correspond habituellement à la proportion de la population de l'espèce dans la zone d'intérêt (généralisée = 71-100 %; grande = 31-70 %; restreinte = 11-30 %; petite = 1-10 %; négligeable = < 1 %).
- Gravité
- Au sein de la portée, niveau de dommage (habituellement mesuré comme l'ampleur de la réduction de la population) que causera vraisemblablement la menace sur l'espèce d'ici une période de 10 ans ou de 3 générations (extrême = 71-100 %; élevée = 31-70 %; modérée = 11-30 %; légère = 1-10 %; négligeable = < 1 %; neutre ou avantage possible = > 0 %).
- Immédiateté
- Élevée = menace toujours présente; modérée = menace pouvant se manifester uniquement dans le futur (à court terme [< 10 ans ou 3 générations]) ou pour l'instant absente (mais susceptible de se manifester de nouveau à court terme); faible = menace pouvant se manifester uniquement dans le futur (à long terme) ou pour l'instant absente (mais susceptible de se manifester de nouveau à long terme); insignifiante/négligeable = menace qui s'est manifestée dans le passé et qui est peu susceptible de se manifester de nouveau, ou menace qui n'aurait aucun effet direct, mais qui pourrait être limitative.
Annexe 3. Tableau d'évaluation des menaces, y compris les commentaires, pour l'Ambystoma unisexué dépendant de la salamandre de jefferson (UD 2).
Tableau d'évaluation des menaces
- Nom scientifique de l'espèce ou de l'écosystème
- Ambystoma unisexué, population dépendante de l'A. jeffersonianum
- Identification de l'élément
- xxxxxx
- Code de l'élément
- xxxxxx
- Date
- 2/2/2015
- Évaluateur(s) :
- Jim Bogart (rédacteur du rapport), Kristiina Ovaska (facilitatrice), Bev McBride (Secrétariat du COSEPAC), Mary Sabine, Leslie Anthony, Isabelle Gauthier, Yohann Dubois, Joe Crowley
- Références :
- Rapport de situation du COSEPAC (ébauche de 2015)
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 3 | 2 |
| C | Moyen | 0 | 1 |
| D | Faible | 2 | 2 |
| - | Impact global des menaces calculé : | Très élevé | Très élevé |
- Impact global des menaces attribué :
- Très élevé
- Commentaires sur l'impact global des menaces
- Durée d'une génération - 11 ans (33 ans pour 3 générations); < 10 000 adultes
| Menace | Menace | Impact (calculé) | Impact (calculé) | Portée (10 prochaines années) | Gravité (10 ans ou 3 générations) | Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | B | Élevé | Grande (31-70 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 1.1 | Zones résidentielles et urbaines | B | Élevé | Grande (31-70 %) | Élevée (31-70 %) | Élevée (continue) | La perte d'habitat causée par l'aménagement résidentiel, particulièrement dans la région de Hamilton et le long de l'escarpement du Niagara, est problématique. Il est difficile de protéger l'habitat dans ces secteurs. L'impact moyen est élevé (l'habitat terrestre ou aquatique est touché). Si des étangs de reproduction sont drainés, des disparitions auront lieu à l'échelle locale. |
| 1.2 | Zones commerciales et industrielles | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | Le groupe ne connaissait l'existence d'aucune zone de projet imminent, mais le développement industriel est une possibilité. |
| 1.3 | Zones touristiques et récréatives | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | Modérée (peut-être à court terme, < 10 ans ou 3 générations) | Certaines activités d'aménagement ont probablement lieu, mais on n'en possède aucun exemple. Les répercussions proviennent de la perte d'habitat. Les terrains de golf, l'infrastructure des parcs et les secteurs aménagés semblables auraient vraisemblablement un impact moins élevé que les projets domiciliaires en raison de la réglementation applicable (p. ex. sur le plan de l'emplacement, protection des zones riveraines, etc.), et les modifications de l'habitat associées à cette menace sont souvent moins intenses que pour l'aménagement résidentiel (p. ex. les étangs peuvent être conservés dans les terrains de golf; cependant les herbicides et pesticides risquent de s'accumuler dans ces étangs, ce qui réduit leur caractère convenable). |
| 2 | Agriculture et aquaculture | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | Les taux actuels de transformation des terres aux fins de l'agriculture sont faibles dans le sud de l'Ontario. Aux endroits où une telle transformation a lieu, cependant, l'impact est élevé parce que les étangs sont habituellement drainés et transformés en terres cultivées, notamment pour le maïs et le soja. |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | La transformation de parcelles de feuillus en parcelles de conifères peut avoir lieu après les coupes, ce qui réduit la quantité d'habitat disponible, mais ce facteur n'est pas considéré comme une menace pour le moment en raison de sa très faible portée. |
| 2.3 | Élevage de bétail | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | Élevée (continue) | La portée est semblable à celle d'autres utilisations agricoles, mais la gravité est plus faible. Le bétail élevé en liberté peut avoir des conséquences sur les salamandres et leur habitat en utilisant les étangs comme source d'eau et en piétinant la végétation riveraine, les salamandres et leurs œufs. |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | - |
| 3 | Production d'énergie et exploitation minière | B | Élevé | Généralisée - grande (31-100 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 3.1 | Forage pétrolier et gazier | - | - | - | - | - | - |
| 3.2 | Exploitation de mines et de carrières | B | Élevé | Généralisée - grande (31-100 %) | Élevée (31-70 %) | Élevée (continue) | Extraction d'agrégats : l'exploitation de carrières est très problématique sur l'escarpement du Niagara. |
| 3.3 | Énergie renouvelable | - | - | - | - | - | - |
| 4 | Corridors de transport et de service | BC | Élevé - moyen | Grande (31-70 %) | Élevée - modérée (11-70 %) | Élevée (continue) | - |
| 4.1 | Routes et voies ferrées | BC | Élevé - moyen | Grande (31-70 %) | Élevée - modérée (11-70 %) | Élevée (continue) | Le sud de l'Ontario présente un dense réseau routier, et la grande majorité des sous-populations est exposée aux routes. Les conséquences découlent de la mortalité routière durant les migrations de reproduction et, dans une moindre mesure, des obstacles aux déplacements. On sait que la mortalité routière a lieu dans certaines régions du sud de l'Ontario malgré les tentatives d'atténuation de cette menace, comme la fermeture de routes à proximité de certains sites. La mortalité routière peut avoir de graves répercussions sur les populations locales. |
| 4.2 | Lignes de services publics | - | - | - | - | - | - |
| 4.3 | Voies de transport par eau | - | - | - | - | - | - |
| 4.4 | Corridors aériens | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 5.1 | Chasse et capture d'animaux terrestres | - | - | - | - | - | - |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | - |
| 5.3 | Exploitation forestière et récolte du bois | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | Dans le sud-ouest de l'Ontario, quelques petits boisés se trouvent sur des terres privées. L'habitat n'est plus convenable une fois que les troncs d'arbres ont été enlevés (au moins une population a disparu après des coupes). Toutefois, il y a peu d'exploitation forestière le long de l'escarpement du Niagara, où se trouvent la plupart des populations de salamandres. |
| 5.4 | Pêche et récolte de ressources aquatiques | - | - | - | - | - | - |
| 6 | Intrusions et perturbations humaines | - | - | - | - | - | - |
| 6.1 | Activités récréatives | - | - | - | - | - | Des véhicules tout-terrain endommagent parfois les étangs, habituellement tard dans l'été. Cette menace ne cible pas nécessairement les mares printanières, mais peut tuer des salamandres récemment métamorphosées. |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | - |
| 6.3 | Travail et autres activités | - | - | - | - | - | Les salamandres ne sont plus tuées dans le cadre de la recherche. |
| 7 | Modifications des systèmes naturels | - | - | - | - | - | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | - |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | - | - | - | - | - | Ce facteur n'est pas considéré comme problématique pour ces salamandres (mis à part le drainage des étangs aux fins du développement, qui est pris en compte sous la menace 1). |
| 7.3 | Autres modifications de l'écosystème | - | - | - | - | - | Remarque : La perte de populations donneuses de sperme à la suite d'événements stochastiques n'est pas prise en compte ici; une telle perte accompagnerait probablement d'autres menaces qui pèsent tant sur les individus unisexués que sur les donneurs de sperme. |
| 8 | Espèces et gènes envahissants ou autrement problématiques | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | D | Faible | Petite (1-10 %) | Extrême (71-100 %) | Élevée (continue) | Les poissons introduits, le zooplancton envahissant (introduit avec les poissons), les maladies émergentes et les plantes aquatiques envahissantes (Phragmites australis) ont été pris en compte. Les salamandres ne coexistent pas bien avec les poissons prédateurs. Le zooplancton introduit est en voie de devenir un grave problème à l'échelle de l'écosystème dans le sud de l'Ontario (les arthropodes indigènes ne s'en nourrissent pas). Mis à part l'introduction de poissons prédateurs, on ignore si ces autres menaces représentent un problème pour les salamandres. |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | - | - |
| 8.3 | Matériel génétique introduit | - | - | - | - | - | - |
| 9 | Pollution | - | Inconnu | Petite (1-10 %) | Inconnue | Élevée (continue) | - |
| 9.1 | Eaux usées domestiques et urbaines | - | Inconnu | Petite (1-10 %) | Inconnue | Élevée (continue) | Le ruissellement des routes (p. ex. sels) est pris en compte dans cette catégorie. Les milieux naturels sont protégés à un certain degré par des bandes tampons, mais certains étangs sont situés juste à côté des routes. On ne dispose d'aucune donnée sur les effets sur les salamandres; c'est pourquoi la gravité est « inconnue ». |
| 9.2 | Effluents industriels et militaires | - | - | - | - | - | - |
| 9.3 | Effluents agricoles et sylvicoles | - | Inconnu | Petite (1-10 %) | Inconnue | Élevée (continue) | Le ruissellement de produits chimiques agricoles (pesticides, herbicides, engrais) a été pris en compte. On ne dispose d'aucune information sur les effets de ce ruissellement sur les salamandres. |
| 9.4 | Déchets solides et ordures | - | - | - | - | - | - |
| 9.5 | Polluants atmosphériques | - | - | - | - | - | - |
| 9.6 | Apports excessifs d'énergie | - | - | - | - | - | - |
| 10 | Phénomènes géologiques | - | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | - | - |
| 11 | Changements climatiques et phénomènes météorologiques violents | - | - | - | - | - | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | - |
| 11.2 | Sécheresses | - | - | - | - | - | L'assèchement hâtif occasionnel des mares printanières à la suite de sécheresses prolongées ne représente probablement pas un grave problème pour ces salamandres longévives qui connaissent possiblement plusieurs périodes de reproduction. Cependant, plusieurs années de sécheresse auraient des conséquences sur les populations. Il est peu probable que ce facteur devienne problématique pour les salamandres au cours des 10 prochaines années, mais il pourrait prendre de l'importance à l'avenir. |
| 11.3 | Températures extrêmes | - | - | - | - | - | Fluctuations de température - un printemps chaud suivi de périodes de gel peut tuer les salamandres dans les étangs de reproduction. |
| 11.4 | Tempêtes et inondations | - | - | - | - | - | - |
Classification des menaces d'après l'IUCN-CMP, Salafsky et al. (2008).
Glossaire
- Impact
- Mesure dans laquelle on observe, infère ou soupçonne que l'espèce est directement ou indirectement menacée dans la zone d'intérêt. Le calcul de l'impact de chaque menace est fondé sur sa gravité et sa portée et prend uniquement en compte les menaces présentes et futures. L'impact d'une menace est établi en fonction de la réduction de la population de l'espèce, ou de la diminution ou de la dégradation de la superficie d'un écosystème. Le taux médian de réduction de la population ou de la superficie pour chaque combinaison de portée et de gravité correspond aux catégories d'impact suivantes : très élevé (déclin de 75 %), élevé (40 %), moyen (15 %) et faible (3 %). Inconnu : catégorie utilisée quand l'impact ne peut être déterminé (p. ex. lorsque les valeurs de la portée ou de la gravité sont inconnues); non calculé : l'impact n'est pas calculé lorsque la menace se situe en dehors de la période d'évaluation (p. ex. l'immédiateté est insignifiante/négligeable ou faible puisque la menace n'existait que dans le passé); négligeable : lorsque la valeur de la portée ou de la gravité est négligeable; n'est pas une menace : lorsque la valeur de la gravité est neutre ou qu'il y a un avantage possible.
- Portée
- Proportion de l'espèce qui, selon toute vraisemblance, devrait être touchée par la menace d'ici 10 ans. Correspond habituellement à la proportion de la population de l'espèce dans la zone d'intérêt (généralisée = 71-100 %; grande = 31-70 %; restreinte = 11-30 %; petite = 1-10 %; négligeable = < 1 %).
- Gravité
- Au sein de la portée, niveau de dommage (habituellement mesuré comme l'ampleur de la réduction de la population) que causera vraisemblablement la menace sur l'espèce d'ici une période de 10 ans ou de 3 générations (extrême = 71-100 %; élevée = 31-70 %; modérée = 11-30 %; légère = 1-10 %; négligeable = < 1 %; neutre ou avantage possible = > 0 %).
- Immédiateté
- Élevée = menace toujours présente; modérée = menace pouvant se manifester uniquement dans le futur (à court terme [< 10 ans ou 3 générations]) ou pour l'instant absente (mais susceptible de se manifester de nouveau à court terme); faible = menace pouvant se manifester uniquement dans le futur (à long terme) ou pour l'instant absente (mais susceptible de se manifester de nouveau à long terme); insignifiante/négligeable = menace qui s'est manifestée dans le passé et qui est peu susceptible de se manifester de nouveau, ou menace qui n'aurait aucun effet direct, mais qui pourrait être limitative.
Annexe 4. Tableau d'évaluation des menaces, y compris les commentaires, pour l'Ambystoma unisexué dépendant de la salamandre à points bleus (UD 3).
Tableau d'évaluation des menaces
- Nom scientifique de l'espèce ou de l'écosystème
- Ambystoma unisexué, population dépendante de l'A. laterale
- Date
- 2/2/2015
- Évaluateur(s) :
- Jim Bogart (rédacteur du rapport), Kristiina Ovaska (facilitatrice), Bev McBride (Secrétariat du COSEPAC), Mary Sabine, Leslie Anthony, Isabelle Gauthier, Yohann Dubois, Joe Crowley
- Références :
- Rapport de situation du COSEPAC (ébauche de 2015)
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 0 | 0 |
| D | Faible | 2 | 2 |
| - | Impact global des menaces calculé : | Faible | Faible |
- Impact global des menaces attribué :
- Faible
- Commentaires sur l'impact global des menaces
- Durée d'une génération - 11 ans (33 ans pour 3 générations); > 1 000 000 d'adultes; zone d'occurrence : 671 668 km²; IZO : 1 932 km² (d'après l'ébauche de rapport de situation du COSEPAC)
| Menace | Menace | Impact (calculé) | Impact (calculé) | Portée (10 prochaines années) | Gravité (10 ans ou 3 générations) | Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 1.1 | Zones résidentielles et urbaines | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | L'aire de répartition est grande, et la plupart des régions ne sont pas touchées par l'aménagement résidentiel; ce facteur n'est problématique que dans le sud de l'Ontario et du Québec. |
| 1.2 | Zones commerciales et industrielles | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | Le groupe ne disposait d'aucun exemple précis, mais le développement est probable. Là encore, seule une très faible proportion de la grande aire de répartition serait touchée. |
| 1.3 | Zones touristiques et récréatives | - | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | Modérée (peut-être à court terme, < 10 ans ou 3 générations) | Une perte d'habitat découlant de l'aménagement de terrains de golf, d'installations de parcs et de terrains de camping pourrait avoir lieu, mais on ne dispose d'aucun exemple précis. |
| 2 | Agriculture et aquaculture | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | Il n'y a pas d'activités agricoles dans le nord de l'aire de répartition. La portée est très petite compte tenu de la grande aire de répartition de cette UD. |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | - |
| 2.3 | Élevage de bétail | - | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | Élevée (continue) | Cette menace est possible, mais elle n'est pas problématique dans la majeure partie de l'habitat. Il y a quelques fermes laitières au Nouveau-Brunswick, mais aucune expansion importante n'est probable; idem pour le Québec. |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | - |
| 3 | Production d'énergie et exploitation minière | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 3.1 | Forage pétrolier et gazier | - | - | - | - | - | - |
| 3.2 | Exploitation de mines et de carrières | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | L'extraction d'agrégats est problématique surtout dans le sud de l'Ontario, mais n'est pas considérée comme menaçante dans le reste de l'aire de répartition. |
| 3.3 | Énergie renouvelable | - | - | - | - | - | - |
| 4 | Corridors de transport et de service | D | Faible | Petite (1-10 %) | Élevée - modérée (11-70 %) | Élevée (continue) | - |
| 4.1 | Routes et voies ferrées | D | Faible | Petite (1-10 %) | Élevée - modérée (11-70 %) | Élevée (continue) | Les répercussions découlent de la mortalité routière durant les migrations de reproduction ainsi que des obstacles aux déplacements. Ce facteur est problématique surtout dans le sud de l'Ontario (probablement 5 à 10 % de l'aire de répartition de cette UD); sinon, la portée de cette menace est négligeable. |
| 4.2 | Lignes de services publics | - | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | Élevée (continue) | Un projet de pipeline traversant le Nouveau-Brunswick depuis St. John est proposé et pourrait être construit dans les 10 prochaines années; les coupes à blanc pour les corridors de transport d'énergie se poursuivent, ce qui fragmente l'habitat et modifie les propriétés hydrologiques des étangs. Seule une très faible proportion de l'aire de répartition est touchée. |
| 4.3 | Voies de transport par eau | - | - | - | - | - | - |
| 4.4 | Corridors aériens | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques | D | Faible | Restreinte (11-30 %) | Modérée (11-30 %) | Élevée (continue) | - |
| 5.1 | Chasse et capture d'animaux terrestres | - | - | - | - | - | - |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | - |
| 5.3 | Exploitation forestière et récolte du bois | D | Faible | Restreinte (11-30 %) | Modérée (11-30 %) | Élevée (continue) | La plupart des populations depuis le Bouclier canadien jusqu'au nord de l'aire de répartition sont situées sur des terres de foresterie exploitées, mais on ne sait pas exactement quelle superficie fera l'objet de coupes au cours des 10 prochaines années puisque les activités de foresterie reposent sur des cycles à long terme. En Ontario, on ne pratique habituellement pas de grandes coupes à blanc; les coupes sont plutôt sélectives. Normalement, les plans d'eau sont protégés. Selon l'échelle et le type des coupes, l'exploitation forestière n'est pas nécessairement néfaste pour les salamandres. |
| 5.4 | Pêche et récolte de ressources aquatiques | - | - | - | - | - | L'utilisation de salamandres comme appâts à poissons est illégale, et n'est pas considérée comme problématique. |
| 6 | Intrusions et perturbations humaines | - | - | - | - | - | - |
| 6.1 | Activités récréatives | - | - | - | - | - | - |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | - |
| 6.3 | Travail et autres activités | - | - | - | - | - | - |
| 7 | Modifications des systèmes naturels | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | - |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) | Bon nombre des plus grands étangs sont d'anciens étangs à castors qui existent depuis des décennies sinon plus. L'enlèvement des digues de castors peut assécher un étang ou changer les caractéristiques de celui-ci, au détriment des salamandres. Bien que l'enlèvement des digues de castors puisse avoir un grave impact à l'échelle locale, dans l'ensemble de la population, la portée de cette menace est négligeable. |
| 7.3 | Autres modifications de l'écosystème | - | - | - | - | - | Remarque : la perte de populations donneuses de sperme à la suite d'événements stochastiques n'est pas prise en compte ici; une telle perte accompagnerait probablement d'autres menaces qui pèsent tant sur les individus unisexués que sur les donneurs de sperme. |
| 8 | Espèces et gènes envahissants ou autrement problématiques | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | - |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) | Les poissons introduits, le zooplancton envahissant (introduit avec les poissons), les maladies émergentes et les plantes aquatiques envahissantes (Phragmites australis) ont été pris en compte. De manière générale, il y a moins d'introductions dans les régions du nord que dans le sud. L'empoissonnement a surtout lieu dans les grands plans d'eau (exempts de salamandres), mais des introductions illégales dans les étangs à salamandres peuvent avoir lieu occasionnellement (aucune donnée à ce sujet). Les salamandres ne coexistent pas bien avec les poissons prédateurs. Le zooplancton introduit est en voie de devenir un grave problème à l'échelle de l'écosystème dans le sud de l'Ontario (les arthropodes indigènes ne s'en nourrissent pas). On ne sait pas si le zooplancton ou les maladies émergentes représentent un problème pour cette population. |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | - | - |
| 8.3 | Matériel génétique introduit | - | - | - | - | - | - |
| 9 | Pollution | - | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (continue) | - |
| 9.1 | Eaux usées domestiques et urbaines | - | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (continue) | - |
| 9.2 | Effluents industriels et militaires | - | - | - | - | - | - |
| 9.3 | Effluents agricoles et sylvicoles | - | Négligeable | Négligeable (< 1 %) | Inconnue | Élevée (continue) | - |
| 9.4 | Déchets solides et ordures | - | - | - | - | - | - |
| 9.5 | Polluants atmosphériques | - | - | - | - | - | - |
| 9.6 | Apports excessifs d'énergie | - | - | - | - | - | - |
| 10 | Phénomènes géologiques | - | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | - | - |
| 11 | Changements climatiques et phénomènes météorologiques violents | - | - | - | - | - | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | - |
| 11.2 | Sécheresses | - | - | - | - | - | Ce facteur n'est pas considéré comme problématique pour les 10 prochaines années. |
| 11.3 | Températures extrêmes | - | - | - | - | - | - |
Glossaire
- Impact
- Mesure dans laquelle on observe, infère ou soupçonne que l'espèce est directement ou indirectement menacée dans la zone d'intérêt. Le calcul de l'impact de chaque menace est fondé sur sa gravité et sa portée et prend uniquement en compte les menaces présentes et futures. L'impact d'une menace est établi en fonction de la réduction de la population de l'espèce, ou de la diminution ou de la dégradation de la superficie d'un écosystème. Le taux médian de réduction de la population ou de la superficie pour chaque combinaison de portée et de gravité correspond aux catégories d'impact suivantes : très élevé (déclin de 75 %), élevé (40 %), moyen (15 %) et faible (3 %). Inconnu : catégorie utilisée quand l'impact ne peut être déterminé (p. ex. lorsque les valeurs de la portée ou de la gravité sont inconnues); non calculé : l'impact n'est pas calculé lorsque la menace se situe en dehors de la période d'évaluation (p. ex. l'immédiateté est insignifiante/négligeable ou faible puisque la menace n'existait que dans le passé); négligeable : lorsque la valeur de la portée ou de la gravité est négligeable; n'est pas une menace : lorsque la valeur de la gravité est neutre ou qu'il y a un avantage possible.
- Portée
- Proportion de l'espèce qui, selon toute vraisemblance, devrait être touchée par la menace d'ici 10 ans. Correspond habituellement à la proportion de la population de l'espèce dans la zone d'intérêt (généralisée = 71-100 %; grande = 31-70 %; restreinte = 11-30 %; petite = 1-10 %; négligeable = < 1 %).
- Gravité
- Au sein de la portée, niveau de dommage (habituellement mesuré comme l'ampleur de la réduction de la population) que causera vraisemblablement la menace sur l'espèce d'ici une période de 10 ans ou de 3 générations (extrême = 71-100 %; élevée = 31-70 %; modérée = 11-30 %; légère = 1-10 %; négligeable = < 1 %; neutre ou avantage possible = > 0 %).
- Immédiateté
- Élevée = menace toujours présente; modérée = menace pouvant se manifester uniquement dans le futur (à court terme [< 10 ans ou 3 générations]) ou pour l'instant absente (mais susceptible de se manifester de nouveau à court terme); faible = menace pouvant se manifester uniquement dans le futur (à long terme) ou pour l'instant absente (mais susceptible de se manifester de nouveau à long terme); insignifiante/négligeable = menace qui s'est manifestée dans le passé et qui est peu susceptible de se manifester de nouveau, ou menace qui n'aurait aucun effet direct, mais qui pourrait être limitative.