
Baleine noire de l'Atlantique Nord (Eubalaena glacialis) : évaluation et rapport de situation du COSEPAC 2013

En voie de disparition 2013
Table des matières
- Table des matières
- COSEPAC Sommaire de l'évaluation
- COSEPAC Résumé
- Résumé technique
- Préface
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Collisions avec des navires et enchevêtrement dans les engins de pêche
- Vulnérabilité aux collisions avec des navires
- Gestion de l'industrie du transport maritime
- Vulnérabilité à l'enchevêtrement dans les engins de pêche
- Gestion des activités de pêche dans les eaux états-uniennes
- Autres facteurs limitatifs potentiels
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique du ou des rédacteurs du rapport
- Collections examinées
- Communications Personnelles
Liste des figures
- Figure 1. Baleine noire de l'Atlantique Nord (illustration fournie par Scott Landry, Provincetown Center for Coastal Studies
- Figure 2. Observations (points noirs, 18492010) et répartition inférée à l'échelle du bassin (zones grises) des baleines noires dans l'Atlantique Nord. Les données sur les observations proviennent de la base de données du North Atlantic Right Whale Consortium (Right Whale Consortium, 2011) et n'ont pas été corrigées en fonction des activités d'observation et n'ont pas, non plus, fait l'objet d'un examen par les pairs. Les profils de répartition fondés sur ces données sont biaisés en fonction de l'endroit et du moment, où les observations aériennes et maritimes, qu'elles soient systématiques ou fortuites, ont été effectuées. Chaque point correspond à une observation, qui pourrait représenter plus d'un individu.
- Figure 3. Zones légalement désignées au Canada et aux États-Unis comme faisant partie de l'habitat essentiel de la baleine noire : la baie du cap Cod (BCC), le Grand chenal Sud (GCS) et les aires de mise bas situées au Sud-Est des États-Unis (SEEU, de la Floride à la Georgie) en vertu de la Endangered Species Act des États-Unis; et le bassin Roseway (BR) et le bassin de Grand Manan (BGM) en vertu de la Loi sur les espèces en péril.
- Figure 4. Répartition de la baleine noire de l'Atlantique Nord inférée à partir de données d'observation en eaux états-uniennes et canadiennes dans la période 1849-2010. Les données sur les observations proviennent de la base de données du North Atlantic Right Whale Consortium (Right Whale Consortium, 2011) et n'ont pas été corrigées en fonction des activités d'observation et n'ont pas, non plus, fait l'objet d'un examen par les pairs. Chaque point correspond à une observation, non pas à une baleine individuelle.
- Figure 5. Effectifs totaux des jeunes baleines noires de l'Atlantique Nord (jeunes de l'année) observés morts ou vivants chaque année, dans la période 1993-2010 (Pettis, 2010; Waring et al., 2011) Au moins quelquesuns des baleineaux morts auront aussi été observés vivants et, par conséquent, auront été « comptés » deux fois dans la figure.
- Figure 6. Nombre cumulatif de baleineaux (0-1 an) et d'individus excluant les baleineaux (> 1 an) de la baleine noire de l'Atlantique Nord qui ont été ajoutés au catalogue du New England Aquarium dans la période 1980-2009 (Hamilton et al., 2007). Il faut noter que le nombre de baleineaux ajoutés au catalogue ne correspond pas au nombre de baleineaux nés chaque année, parce que l'identification des baleineaux nouvellement nés est une tâche difficile (en raison des callosités qui ne sont pas encore complètement formées). Nombre de baleineaux sont identifiés seulement plus tard (dans la baie de Fundy, lorsqu'ils ont environ 6 mois), une fois que la disposition des callosités est mieux développée. Par conséquent, il n'est pas inhabituel qu'un baleineau né une année civile ne soit photo identifié pour la première fois que l'année civile suivante et qu'il soit encore classé comme baleineau à ce moment-là. Il faut également noter que, dans la catégorie des individus excluant les baleineaux, on retrouve seulement les animaux qui n'ont pas été observés pour la première fois, ni photo identifiés lorsqu'ils étaient des baleineaux. Le plafonnement de la courbe pour les individus excluant les baleineaux semble indiquer que tous (ou presque tous) ces individus ont maintenant été identifiés et que tous les nouveaux individus sont identifiés pour la première fois lorsqu'ils sont des baleineaux
- Figure 7. Graphique du haut : Nombre annuel de baleines noires de l'Atlantique Nord adultes (≥ 9 ans) présumées vivantes entre 1980 et 2009(en fonction des individus identifiés dans le catalogue d'identification photographique; Right Whale Consortium, 2011). Le nombre total réel est probablement plus grand que le nombre indiqué, parce qu'il est possible que le catalogue d'identification photographique n'inclue pas les individus adultes qui ne fréquentent ou n'occupent pas régulièrement les zones ayant fait l'objet de relevés. De plus, les individus qui n'ont pas été observés et identifiés pendant une période de six ans ou plus sont exclus, même si certains d'entre eux sont peut-être encore vivants. Graphique du bas : Relation linéaire représentant le nombre d'individus adultes présumés vivants entre 1997 et 2009 (période pendant laquelle environ 75-100 % du nombre total des individus adultes étaient présumés vivants)
- Figure 8. Nombre cumulatif de baleines noires de l'Atlantique Nord mâles et femelles adultes présumées mortes (selon la règle de six ans) et dont la mort a été confirmée (Right Whale Consortium, 2011). Cette figure montre que, récemment, la mort d'un plus grand nombre de femelles adultes a été confirmée et, aussi, que le nombre de mâles et de femelles présumés morts est nettement supérieur au nombre de mâles et de femelles dont la mort est confirmée.
Liste des tableaux
Liste des annexes
Information sur le document

Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2013. Évaluation et Rapport de situation du COSEPAC sur la baleine noire de l'Atlantique Nord (Eubalaena glacialis) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xi + 63 pp.
Rapport(s) précédent(s) :
COSEWIC. 2003. COSEWIC assessment and update status report on the North Atlantic Right Whale Eubalaena glacialis in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vii + 28 pp.
Gaskin, D.E. 1990. Update COSEWIC status report on the North Atlantic right whale Eubalaena glacialis in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 25 pp.
Gaskin, D.E. 1985. Update COSEWIC status report on the right whale Eubalaena glacialis in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 50 pp.
Hay, K.A. 1980. COSEWIC status report on the right whale Eubalaena glacialis in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 12 pp.
Note de production :
Le COSEPAC remercie Sarah M.E. Fortune et Andrew W. Trites d'avoir rédigé le rapport de situation sur la baleine noire de l'Atlantique Nord (Eubalaena glacialis) au Canada, aux termes d'un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Randall Reeves, coprésident du Sous comité de spécialistes des mammifères marins.
Pour obtenir des exemplaires supplémentaires, s'adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-953-3215
Téléc. : 819-994-3684
COSEPAC Courriel :
COSEPAC site Web
Also available in English under the title COSEWIC Assessment and Status Report on the North Atlantic Right Whale (Eubalaena glacialis) in Canada.
Illustration/photo de la couverture :
Baleine noire de l'Atlantique Nord -- illustration fournie par Scott Landry, Provincetown Center for Coastal Studies.
©Sa Majesté la Reine du chef du Canada, 2014.
No de catalogue CW69-14/328-2014F-PDF
ISBN 978-0-660-22072-7

COSEPAC Sommaire de l'évaluation
Sommaire de l'évaluation - novembre 2013
- Nom commun
- Baleine noire de l'Atlantique Nord
- Nom scientifique
- Eubalaena glacialis
- Statut
- En voie de disparition
- Justification de la désignation
- Cette espèce de baleine longévive, et dont la reproduction est lente, est presque disparue en raison de la chasse commerciale à la baleine, mais a été protégée de la chasse depuis 1935. Les baleines se retrouvant au Canada font partie d'une seule population globale de l'espèce, laquelle est endémique à l'océan nord-atlantique. Depuis 1990, la population totale a augmenté à un rythme d'environ 2,4 % par année. La population totale en 2010, incluant toutes les classes d'âge, a été estimée à 468 individus, dont 122 à 136 de femelles adultes. Le nombre d'individus matures, en tenant compte d'un sex-ratio en faveur des mâles parmi les adultes et d'un petit nombre de femelles incapables de se reproduire, est estimé se situer entre 245 et 272. Le taux de croissance de la population est inférieur à ce qui pourrait être prédit selon la biologie de l'espèce et est limité par les collisions avec des navires et par l'enchevêtrement dans des engins de pêche. Bien que des mesures aient été mises en œuvre au Canada et aux États-Unis pour réduire les collisions avec des navires, elles continuent de se produire, et on s'attend à ce que la circulation maritime augmente considérablement dans l'aire de répartition de l'espèce au cours des prochaines décennies. De plus, les femelles adultes semblent être plus susceptibles d'être heurtées que les mâles. Bien que des efforts limités aient également été réalisés pour réduire l'incidence et la gravité des enchevêtrements, ces situations n'en demeurent pas moins une cause importante de blessures et de mortalité.
- Répartition
- Océan Atlantique
- Historique du statut
- La baleine noire a été considérée comme une espèce distincte et a été désignée « en voie de disparition » en 1980. Réexamen et confirmation du statut en avril 1985 et en avril 1990. Division en deux espèces en mai 2003 afin de permettre une désignation séparée pour la baleine noire de l'Atlantique Nord. La baleine noire de l'Atlantique Nord a été désignée « en voie de disparition » en mai 2003 et en novembre 2013.
COSEPAC Résumé
Baleine noire de l'Atlantique Nord
Eubalaena glacialis
Description et importance de l'espèce sauvage
Les baleines noires de l'Atlantique Nord (Eubalaena glacialis sont des baleines à fanons de grande taille, mesurant environ 14 m (écart-type de ±0,15) et pesant environ 30 mt (écart-type de ±5,4) à leur taille maximale, selon des modèles de croissance standards appliqués à des données de nécropsies et de photogrammétrie. Néanmoins, les baleines noires peuvent atteindre une longueur de 18 m, les femelles étant environ 0,7 m plus longues que les mâles lorsqu'elles sont sexuellement matures. D'apparence trapue, elles ont des nageoires ressemblant à des pagaies et une grosse tête (environ le quart de leur longueur corporelle) et n'ont pas de nageoire dorsale. Leur large nageoire caudale, profondément entaillée, a une bordure postérieure lisse. La majeure partie du corps est noire, mais certains individus ont une pigmentation blanche sur le ventre et le menton. De grandes plaques saillantes de tissu épithélial (callosités) sont présentes sur la tête et le menton, au-dessus des yeux, derrière l'évent et le long de la lèvre inférieure.
Répartition
Dans l'ouest de l'Atlantique Nord, l'aire de répartition des baleines noires s'étend de la Floride à Terre-Neuve et au golfe du Saint-Laurent. Une partie de la population (principalement les femelles matures sur le plan reproducteur, les baleineaux et quelques jeunes baleines) migre, chaque année, des aires de mise bas hivernales au large de la Floride et de la Georgie (États-Unis) vers le Canada. Environ les deux tiers de la population se rassemblent habituellement à l'embouchure de la baie de Fundy et dans la plate-forme Néo-Écossaise durant l'été et l'automne, et des effectifs, mais peu nombreux, fréquentent deux zones dans le golfe du Saint-Laurent – une au nord et à l'est de la Gaspésie, et l'autre au sud-est de la Gaspésie, à l'embouchure de la baie des Chaleurs.
Habitat
Les baleines noires de l'Atlantique Nord nagent à des profondeurs variées et à des distances plus ou moins grandes de la côte, notamment dans les eaux côtières peu profondes, les eaux côtières profondes et les eaux extracôtières. Les baleines gestantes et allaitantes fréquentent les eaux côtières chaudes peu profondes au large de la Floride et de la Georgie, de novembre environ à avril. De janvier à la mi-mai, les baleines fréquentent des eaux plus froides et productives vers le nord (p. ex. la baie du cap Cod). Tous les groupes démographiques cherchent leur nourriture dans le Grand chenal Sud, au printemps et au début de l'été, et dans les zones côtières profondes, comme la baie de Fundy et la plate-forme Néo-Écossaise, de l'été à la fin de l'automne. Les baleines noires de l'Atlantique Nord dépendent, pour se nourrir, des processus environnementaux et océaniques (liés, entre autres, au vent, à la température, à la salinité, aux courants) qui entraînent la formation de fortes concentrations de copépodes. Par conséquent, la variabilité de ces paramètres peut modifier la qualité de leur habitat.
Biologie
Les baleines noires de l'Atlantique Nord sont des animaux filtreurs qui se nourrissent principalement de copépodes calanoïdes et, à l'occasion, d'Euphausiacés et de larves de balanes communes. Elles se nourrissent d'une variété d'espèces de copépodes durant l'hiver et de Calanus finmarchicus aux stades de développement où ils sont riches en huile, durant le printemps, l'été et l'automne.
La plupart des femelles matures sur le plan reproducteur donnent naissance à un seul petit, tous les 3 à 5 ans. L'âge de la première mise bas observée varie de 5 à 21 ans (âge moyen de 10 ans), et au moins 2 femelles ont continué à produire des baleineaux sur 31 ans d'observation. L'âge de la première reproduction pour les mâles est d'environ 15 ans. La durée d'une génération (l'âge moyen des femelles avec baleineau) était de 16,1 ans pour la population croissante de 2002 à 2009, mais aurait pu atteindre 35,7 ans (pré exploitation). La durée d'une génération varie donc de 16 à 36 ans.
Les baleines noires se reproduisent en groupes de parade nuptiale composés d'une femelle et de plusieurs mâles. L'intervalle entre les mises bas de 3 à 5 ans signifie qu'il y a environ une femelle en ovulation pour quatre mâles adultes, ce qui entraîne une forte compétition entre les mâles pour concrétiser les occasions d'accouplement. La gestation durerait environ 12 mois. Les baleineaux sont habituellement allaités pendant un an et auront atteint environ 75 % de leur taille adulte au sevrage. Quelques individus reconnaissables ont été aperçus sur plus de trois décennies, et l'individu le plus âgé répertorié aurait atteint au moins 70 ans.
Taille et tendances des populations
En 2010, un total de 468 baleines noires, dont 19 baleineaux de l'année et 449 individus excluant les baleineaux (individus de plus de 1 an), étaient vivantes ou présumées vivantes. Sur les 449 individus excluant les baleineaux, 49 % étaient des mâles; 35 % étaient des femelles; et 17 % étaient des individus de sexe indéterminé. La population totale d'individus adultes en 2010 se situait probablement entre 305 et 325. Le nombre estimé d'individus matures (compte tenu des femelles nullipares et du sex ratio biaisé en faveur des mâles) serait situé entre 245 et 272. Le taux moyen de croissance démographique de 1990 à 2007 était de 2,4 % par année. La population a augmenté d'environ 50 % entre 1990 et 2010, et la structure d'âge est demeurée plutôt constante. Le nombre de baleineaux aperçus durant les relevés aériens et les relevés effectués en bateau (1993-2010) variait de 1 à 39 par année.
Menaces et facteurs limitatifs
La mortalité causée par les collisions avec des navires et l'enchevêtrement dans les engins de pêche a limité le rétablissement de la population. L'habitat de la baleine noire de l'Atlantique Nord fait l'objet d'une forte circulation maritime, et les engins de pêche installés y sont très nombreux. Les autres facteurs pouvant limiter la croissance démographique sont peu connus.
Protection, statuts et classements
Le stock de l'ouest de baleines noires de l'Atlantique Nord a été gravement décimé par la chasse qui a commencé dans les années 1600 et s'est poursuivie jusqu'à ce que l'espèce se voit accorder une protection juridique dans les années 1930. L'espèce est inscrite comme menacée d'extinction sur la liste rouge de l'UICN et est désignée en voie de disparition aux termes de la Endangered Species Act des États-Unis. Au Canada, les baleines noires sont protégées en vertu du Règlement sur les mammifères marins pris en application de la Loi sur les pêches. La baleine noire de l'Atlantique Nord a été désignée comme espèce en voie de disparition en 2005, en vertu de la Loi sur les espèces en péril (LEP), après avoir été évaluée par le COSEPAC en 2003. L'espèce a été réévaluée comme étant en voie de disparition par le COSEPAC en novembre 2013.
Résumé Technique
Eubalaena glacialis
Baleine noire de l'Atlantique Nord
(baleine franche)
North Atlantic Right Whale
- Répartition au Canada (province/territoire/océan) :
- Océan Atlantique (au large de la Nouvelle Écosse, du Québec, du Nouveau Brunswick, de l'Île-du-Prince-Édouard et de Terre Neuve et Labrador)
Données démographiques
Durée d'une génération
Âge moyen des femelles avec baleineaux connues pour la période 2002 à 2009 (16,1 ans ± 1,54 d'écart-type; plage de 13,1 à 18,4 ans); il aurait pu, cependant, atteindre 35,7 ans (pré-exploitation) si on se fie à la modélisation de la population.
16,1 à 35,7 ans
Y a-t-il un déclin continu observé du nombre total d'individus matures?
Non
Pourcentage estimé de déclin continu du nombre total d'individus matures sur cinq ans ou deux générations.
Aucun déclin actuellement
Pourcentage observé d'augmentation du nombre total d'individus matures au cours des dix dernières années.
Si on suppose que la proportion d'individus matures est restée constante dans le temps, le nombre total d'individus matures a augmenté de ~24 % sur 10 ans (2001-2010) et de ~57 %sur 20 ans(1991-2010).)
Pourcentage prévu ou présumé d'augmentation du nombre total d'individus matures au cours des dix prochaines années ou trois prochaines générations.
Inconnu
Pourcentage observé d'augmentation du nombre total d'individus matures au cours de toute période de dix ans commençant dans le passé et se terminant dans le futur.
Augmentation de ~24 % au cours des 10 dernières années (2001-2010) et de ~57 %au cours des 20 dernièresannées (1991-2010)
Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé?
Il n'y a pas de déclin actuellement, mais la croissance démographique aurait été plus grande en l'absence de mortalité causée par les humains.
Y a-t-il des fluctuations extrêmes du nombre d'individus matures?
Non
Information sur la répartition
Superficie estimée de la zone d'occurrence á l'intérieur du territoire canadien
˜1,8 million de km2
Indice de zone d'occupation (IZO)
Voir le texte pour d'autres possibilités. Les valeurs suivantes s'appliquent à l'habitat essentiel faisant l'objet d'une désignation légale, autre que l'aire de mise bas au large du sud-est des États-Unis :
Baie de Fundy 924 km²
Bassin Roseway 3 648 km²
Baie de cap Cod 1 984 km²
Grand chenal Sud 9 264 km²
7 092 km2englobant seulement les aires de mise bas (sud-est des États-Unis)
La population totale est-elle gravement fragmentée?
Non
Nombre de localités Note de bas de page ∗
Sans objet
Y a-t-il un déclin continu de la zone d'occurrence?
Non
Y a-t-il un déclin continu de l'indice de zone d'occupation?
Non
Y a-t-il un déclin continu du nombre de populations?
Non
Y a-t-il un déclin continu du nombre de localités*?Note de bas de page∗?
Non
Y a-t-il un déclin continu de la superficie, de l'étendue ou de la qualité de l'habitat?
La qualité subit probablement un déclin.
Y a-t-il des fluctuations extrêmes du nombre de populations?
Non
Y a-t-il des fluctuations extrêmes du nombre de localités*?Note de bas de page∗
Non
Y a-t-il des fluctuations extrêmes de la zone d'occurrence?
Non
Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation?
No
| Population | Nombre d'individus matures |
|---|---|
| Ouest de l'Atlantique Nord [Cette estimation suppose que la population comptait au moins 449 individus excluant les baleineaux en 2010; elle a été obtenue en doublant le nombre de femelles qui auraient été vivantes et qui auraient eu au moins 1 baleineau durant leur vie.] |
245-272 |
| Est de l'Atlantique Nord [Une population vestige existe peut-être encore.] |
Inconnu, mais si cette population existe encore, ses effectifs sont trop peu nombreux pour avoir un impact. |
Analyse quantitative
La probabilité de disparition de l'espèce à l'état sauvage sur 20 ans ou 5 générations, ou sur 100 ans.
Inconnue
Menaces (réelles ou imminentes pour les populations ou leur habitat)
Les causes anthropiques de mortalité comprennent notamment, les collisions avec des navires et l'enchevêtrement dans les engins de pêche. Les menaces potentielles et les facteurs limitatifs incluent la pollution par le bruit, la pollution chimique, les parasites et la dégradation de l'habitat.
Immigration de source externe (immigration de l'extérieur du Canada)
Situation des populations de l'extérieur?
L'espèce existe en une seule population transfrontalière qui se déplace saisonnièrement entre le Canada et les États-Unis. Elle figure dans la liste des espèces en voie de disparition aux États-Unis. Une très petite population vestige existe peut-être dans l'est de l'Atlantique Nord.
Une immigration a-t-elle été constatée ou est-elle possible?
Improbable
Des individus immigrants seraient-ils adaptés pour survivre au Canada?
Probablement
Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants?
Inconnu
La possibilité d'une immigration de populations externes existe-t-elle?
Non
Historique du statut
- COSEPAC :
- La baleine noire a été considérée comme une espèce distincte et a été désignée « en voie de disparition » en 1980. Réexamen et confirmation du statut en avril 1985 et en avril 1990. Division en deux espèces en mai 2003 afin de permettre une désignation séparée pour la baleine noire de l'Atlantique Nord. La baleine noire de l'Atlantique Nord a été désignée « en voie de disparition » en mai 2003 et en novembre 2013.
Statut et justification de la désignation
- Statut:
- En voie de disparition
- Code alphanumérique :
- D1
- Justification de la désignation :
- Cette espèce de baleine longévive, et dont la reproduction est lente, est presque disparue en raison de la chasse commerciale à la baleine, mais a été protégée de la chasse depuis 1935. Les baleines se retrouvant au Canada font partie d'une seule population globale de l'espèce, laquelle est endémique à l'océan nord-atlantique. Depuis 1990, la population totale a augmenté à un rythme d'environ 2,4 % par année. La population totale en 2010, incluant toutes les classes d'âge, a été estimée à 468 individus, dont 122 à 136 de femelles adultes. Le nombre d'individus matures, en tenant compte d'un sex-ratio en faveur des mâles parmi les adultes et d'un petit nombre de femelles incapables de se reproduire, est estimé se situer entre 245 et 272. Le taux de croissance de la population est inférieur à ce qui pourrait être prédit selon la biologie de l'espèce et est limité par les collisions avec des navires et par l'enchevêtrement dans des engins de pêche. Bien que des mesures aient été mises en œuvre au Canada et aux États-Unis pour réduire les collisions avec des navires, elles continuent de se produire, et on s'attend à ce que la circulation maritime augmente considérablement dans l'aire de répartition de l'espèce au cours des prochaines décennies. De plus, les femelles adultes semblent être plus susceptibles d'être heurtées que les mâles. Bien que des efforts limités aient également été réalisés pour réduire l'incidence et la gravité des enchevêtrements, ces situations n'en demeurent pas moins une cause importante de blessures et de mortalité.
- Critère A :
- A (déclin du nombre total d'individus matures) : Sans objet. Pas de déclin au cours des trois dernières générations et aucun déclin prévu au cours des trois prochaines générations.
- Critère B :
- (petite aire de répartition, et déclin ou fluctuation) : Sans objet. La zone d'occurrence et l'indice de zone d'occupation sont trop grands.
- Critère C :
- (nombre d'individus matures peu élevé et en déclin) : Sans objet. Aucun déclin continu.
- Critère D :
- (très petite population totale ou répartition restreinte) : Correspond au critère D1 de la catégorie « espèce en voie de disparition », le nombre d'individus matures étant estimé à 245-272.
- Critère E :
- (analyse quantitative) : Aucune analyse applicable n'a été effectuée.
Préface
Le statut d'espèce en voie de disparition de la baleine noire de l'Atlantique Nord a été confirmé en 2003 par le COSEPAC. De nouvelles mesures de gestion visant à réduire la mortalité causée par les collisions avec des navires dans les eaux canadiennes et états-uniennes et par l'enchevêtrement dans les engins de pêche dans les eaux états-uniennes ont été mises en œuvre depuis la dernière évaluation. Néanmoins, les collisions avec des navires et l'enchevêtrement dans les engins de pêche continuent à provoquer la mort de baleines ou à leur infliger des blessures graves.
Le nombre d'individus qui seraient vraisemblablement vivants a augmenté considérablement depuis la dernière évaluation, sans aucun changement majeur de la structure/composition de la population en fonction de l'âge et du sexe. La population totale estimée a augmenté de 1990 à 2007 à un rythme d'environ 2,4 % par année. Les effectifs totaux estimés de la population de baleines en 2010 étaient de 468 (individus de tous âges, y compris 19 baleineauxnés cette année-là), et le nombre estimé d'individus matures, compte tenu des femelles considérées comme incapables de se reproduire et du sex ratio biaisé en faveur des mâles, variait entre 245 et 272.

Historique du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Mandat du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Composition du COSEPAC
Le COSEPAC est composé de membres de chacun des organismes responsable des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Définitions (2013)
- Espèce sauvage
- Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d'animal, de plante ou d'un autre organisme d'origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s'est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
- Disparue (D)
- Espèce sauvage qui n'existe plus.
- Disparue du pays (DP)
- Espèce sauvage qui n'existe plus à l'état sauvage au Canada, mais qui est présente ailleurs.
- En voie de disparition (VD)Note de bas de page*
- Espèce sauvage exposée à une disparition de la planète ou à une disparition du pays imminente.
- Menacée (M)
- Espèce sauvage susceptible de devenir en voie de disparition si les facteurs limitants ne sont pas renversés.
- Préoccupante (P)Note de bas de page**
- Espèce sauvage qui peut devenir une espèce menacée ou en voie de disparition en raison de l'effet cumulatif de ses caractéristiques biologiques et des menaces reconnues qui pèsent sur elle.
- Non en péril (NEP)Note de bas de page***
- Espèce sauvage qui a été évaluée et jugée comme ne risquant pas de disparaître étant donné les circonstances actuelles.
- Données insuffisantes (DI)Note de bas de page****
- Une catégorie qui s'applique lorsque l'information disponible est insuffisante (a) pour déterminer l'admissibilité d'une espèce à l'évaluation ou (b) pour permettre une évaluation du risque de disparition de l'espèce.
Le Service canadien de la faune d'Environnement Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Description et importance de l'espèce
Nom et classification
À l'échelle mondiale, il y a trois espèces reconnues de baleines franches – E. glacialisdans l'Atlantique Nord, E. australis dans l'hémisphère Sud et E. japonica dans le Pacifique Nord (Rosenbaum et al., 2000; IWC, 2001b; Perrin, 2013). Deux de ces trois espèces de baleines franches fréquentent les eaux canadiennes, l'E. japonicadans le Pacifique et l'E. glacialis dans l'Atlantique. Aucune sous-espèce n'est reconnue (Perrin, 2013). Le présent rapport porte sur la situation de l'E. glacialis – la baleine noire de l'Atlantique Nord. Au Canada, les deux noms communs utilisés en français sont « baleine noire » et « baleine franche ».
Description morphologique
Les baleines noires de l'Atlantique Nord sont des baleines à fanons de grande taille, mesurant environ 14 m (écart-type de ±0,15) et pesant environ 30 mt (écart-type de ±5,4) à leur taille maximale, selon des modèles de croissance standards appliqués à des données de nécropsies et de photogrammétrie (Fortune et al., 2012). Néanmoins, les baleines noires peuvent atteindre une longueur de 18 m (Reeves et Kenney, 2003). Elles sont légèrement dimorphes, la femelle étant environ 0,7 m plus longue que le mâle. D'apparence trapue, elles ont des nageoires ressemblant à des pagaies et une grosse tête (figure 1). Une épaisse couche de graisse (8-22 cm) leur sert d'isolant et leur permet de stocker de l'énergie (Miller et al., 2011). La majeure partie de leur corps est noire, mais certains individus ont une pigmentation blanche au ventre et au menton. Elles n'ont pas de nageoire dorsale et n'ont aucun sillon le long de la gorge. Les baleines noires ont une grande nageoire caudale, large d'environ 3 à 5 m, avec une bordure postérieure lisse.
Figure 1. Baleine noire de l'Atlantique Nord (illustration fournie par Scott Landry, Provincetown Center for Coastal Studies
La disposition des callosités (plaques saillantes de tissu épithélial) sur le rostre, derrière l'évent, au-dessus des yeux, aux coins du menton, et, parfois, le long de la lèvre et de la mâchoire inférieures (figure 1) est unique à chaque baleine noire. La disposition des callosités ne change pas avec le temps et peut être utilisée, tout comme les cicatrices sur le corps, pour identifier les individus (Payne et al., 1983; Kraus et al., 1986a; Crone et Kraus, 1990; Hamilton et Martin, 1999).
Le jet de l'évent en V d'une baleine noire peut atteindre une hauteur de 5 m et est une des caractéristiques permettant de distinguer l'espèce en mer. La baleine noire se nourrit dans les profondeurs ou près de la surface. Lorsqu'elle écrème la surface, elle utilise son rostre étroit et très arqué et sa mâchoire inférieure recourbée, qui sont particulièrement adaptés pour cette méthode d'alimentation. Une baleine noire compte environ 250 fanons noirs ou marron dans la mâchoire supérieure. Les fanons sont longs (de 2,0 à 2,8 m) et étroits (jusqu'à 18 cm de largeur), et ils ont chacun des franges fines comme des poils le long de leurs bords intérieurs.
Structure spatiale et variabilité des populations
L'aire de répartition des baleines noires de l'Atlantique Nord s'étend de la Floride à Terre-Neuveet au Saint-Laurent (figure 2). Les aires de mise bas hivernales (faisant partie de l'habitat essentiel désigné) sont situées dans les eaux côtières de la Floride et de la Georgie (figure 3). Les baleines qui utilisent les aires de mise bas au début de l'hiver migrent vers le nord, à la fin de l'hiver et au printemps, pour se nourrir dans la baie du cap Cod, le Grand chenal Sud, et la baie de Massachusetts (figure 3) Les baleines qui utilisent les aires de mise bas au début de l'hiver migrent vers le nord, à la fin de l'hiver et au printemps, pour se nourrir dans la baie du cap Cod, le Grand chenal Sud, et la baie de Massachusetts (figure 3) (Kraus et al., 1986b; Winn et al., 1986; Kenney et al., 2001). Ce ne sont pas tous les individus connus (particulièrement les mâles adultes) qui fréquentent ces aires durant l'hiver. Généralement, on ne voit pas les mâles adultes dans les aires de mise bas ou la baie du cap Cod, pendant l'hiver. En fait, les endroits où ils pourraient se trouver à ce moment-là demeurent en grande partie inconnus (Brown et al., 2001; Frasier et al., 2007a).
Durant l'été et l'automne, les baleines noires se rassemblent et se nourrissent à l'embouchure de la baie de Fundy (principalement à l'est de l'île Grand Manan) et dans le bassin Roseway entre le banc de Brown et le banc de Baccaro, du côté ouest de la plate-forme Néo-Écossaise (figures 3 et 4). À cette période, des effectifs peu nombreux se trouvent ailleurs dans la plate-forme Néo-Écossaise et le golfe du Saint-Laurent, principalement le long de la Basse-Côte-Nord et au sud et à l'est de la Gaspésie, à l'embouchure de la baie des Chaleurs. Une aire de reproduction probable a récemment été découverte au milieu du golfe du Maine (Cole et al., 2013). Des baleines noires ont également été aperçues sporadiquement dans le détroit du Danemark, près de l'Islande et en Norvège, aux Açores, aux Bermudes et dans le golfe du Mexique (Braham et Rice, 1984; Mead, 1986; Jacobsen et al., 2004; Mellinger et al., 2011; Silva et al., 2012).
Figure 2. Observations (points noirs, 1849-2010) et répartition inférée à l'échelle du bassin (zones grises) des baleines noires dans l'Atlantique Nord. Les données sur les observations proviennent de la base de données du North Atlantic Right Whale Consortium (Right Whale Consortium, 2011) et n'ont pas été corrigées en fonction des activités d'observation et n'ont pas, non plus, fait l'objet d'un examen par les pairs. Les profils de répartition fondés sur ces données sont biaisés en fonction de l'endroit et du moment, où les observations aériennes et maritimes, qu'elles soient systématiques ou fortuites, ont été effectuées. Chaque point correspond à une observation, qui pourrait représenter plus d'un individu.
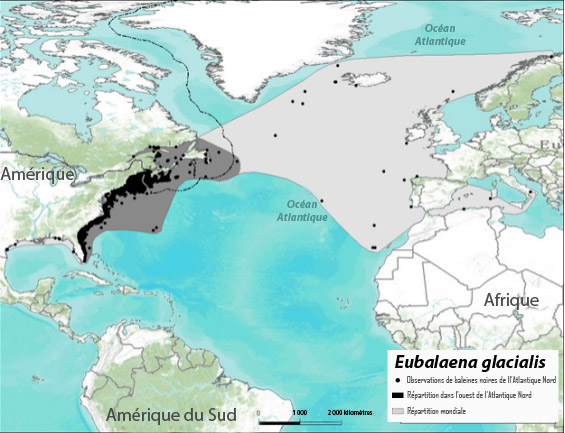
Description longue pour la figure 2
Carte montrant les emplacements où des baleines noires ont été observées (1849 2010) et la répartition inférée à l'échelle du bassin des baleines noires dans l'Atlantique Nord. Les points représentent les observations répertoriées. L'aire de répartition dans l'ouest de l'Atlantique Nord (de la Floride à Terre Neuve et Labrador et au golfe du Saint-Laurent) est indiquée en gris, tout comme la répartition mondiale.
Figure 3. Zones légalement désignées au Canada et aux États-Unis comme faisant partie de l'habitat essentiel de la baleine noire : la baie du cap Cod (BCC), le Grand chenal Sud (GCS) et les aires de mise bas situées au Sud-Est des États-Unis (SEEU, de la Floride à la Georgie) en vertu de la Endangered Species Act des États-Unis; et le bassin Roseway (BR) et le bassin de Grand Manan (BGM) en vertu de la Loi sur les espèces en péril.
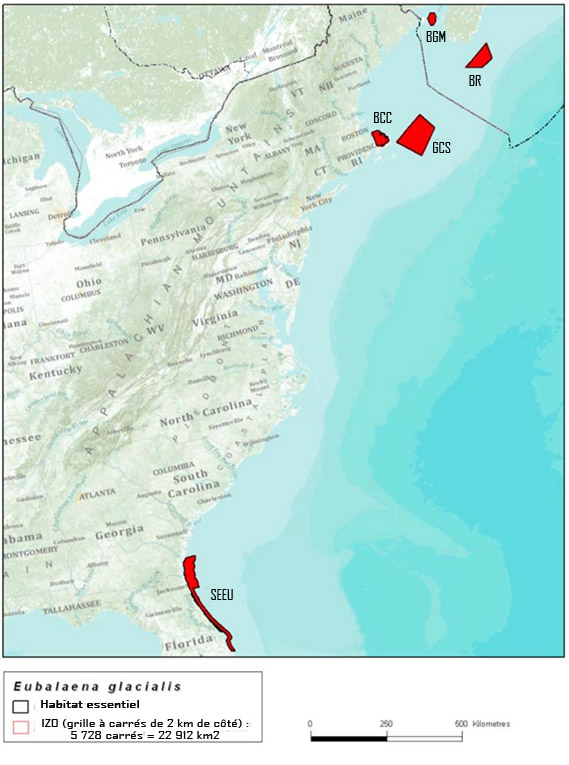
Description longue pour la figure 3
Carte montrant les zones légalement désignées au Canada et aux États-Unis comme faisant partie de l'habitat essentiel de la baleine noire. Ces zones (en rouge) comprennent la baie du cap Cod, le Grand chenal Sud et les aires de mise bas situées au Sud-Est des États-Unis (de la Floride à la Georgie) (zones désignées en vertu de la Endangered Species Act des États Unis) et le bassin Roseway et le bassin de Grand Manan (désignés en vertu de la Loi sur les espèces en péril).
Figure 4. Répartition de la baleine noire de l'Atlantique Nord inférée à partir de données d'observation en eaux états-uniennes et canadiennes dans la période 1849-2010. Les données sur les observations proviennent de la base de données du North Atlantic Right Whale Consortium (Right Whale Consortium, 2011) et n'ont pas été corrigées en fonction des activités d'observation et n'ont pas, non plus, fait l'objet d'un examen par les pairs. Chaque point correspond à une observation, non pas à une baleine individuelle.
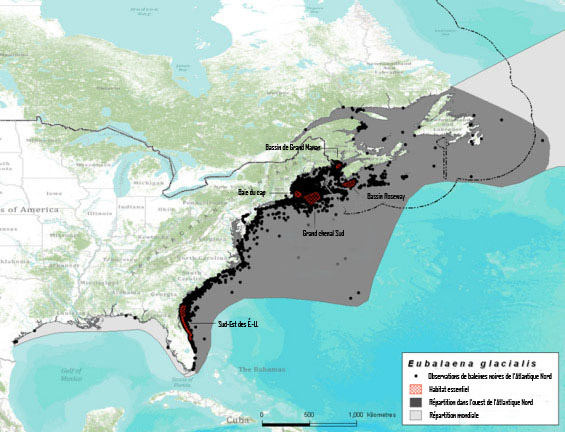
Description longue pour la figure 4
Carte montrant la répartition de la baleine noire de l'Atlantique Nord le long de la côte est de l'Amérique du Nord, inférée à partir de données d'observation en eaux états-uniennes et canadiennes dans la période 1849-2010. Les points représentent des observations répertoriées. Les zones d'habitat essentiel, la répartition dans l'ouest de l'Atlantique Nord et la répartition mondiale sont indiquées sur la carte.
Les baleines noires semblent se tenir en groupes séparés, selon leur âge, sexe et état reproducteur. Les probabilités de voir des femelles adultes dans certaines zones au large du Canada et des États-Unis sont plus faibles pendant les années entre les mises bas, comparativement aux années de mise bas (Brown et al., 2001). Les femelles avec des baleineaux retournent souvent à la même pouponnière estivale. Cette grande fidélité à l'égard des sites peut influer sur l'endroit où les jeunes, sevrés de leur mère, retournent pour se nourrir, les baleines se retrouvant ainsi dans des lieux d'estivage précis, selon leur lignée (Rastogi et al., 2004).
Aucune barrière géographique ne crée ni ne favorise la structure génétique ou l'isolement démographique marqué. L'ADN extrait de fanons et d'os de spécimens de musée semble indiquer que les baleines noires présentes dans l'est et l'ouest de l'Atlantique Nord ne sont pas génétiquement distinctes (Rosenbaum et al., 2000). Les observations de baleines noires de l'ouest de l'Atlantique Nord dans l'est de l'Atlantique Nord laissent supposer que le croisement est possible (Silva et al., 2012) – cependant, les baleines noires de l'est de l'Atlantique Nord ne montrent aucun signe de rétablissement, depuis que la chasse à la baleine a cessé (Braham et Rice, 1984) pas plus qu'il n'y a eu d'indice de recolonisation par des individus venant de l'ouest de l'Atlantique.
Unités désignables
Il n'y a aucune raison de reconnaître plus d'une unité désignable pour les baleines noires de l'Atlantique Nord au Canada.
Importance de l'espèce
Une part importante de l'aire de répartition connue de l'espèce se trouve en eaux canadiennes. Dans le passé (des années 1630 au début des années 1900), les baleines noires avaient une grande importance économique pour leur huile et leurs fanons (Reeves et al., 2007). Aujourd'hui, les baleines noires étant protégées contre la pêche commerciale, leur valeur économique au Canada provient du tourisme d'observation des baleines au Nouveau-Brunswick et en Nouvelle-Écosse. La baleine noire est l'un des mammifères marins les plus étudiés au monde, et elle possède une stratégie de reproduction intéressante, impliquant une compétition intense du sperme (Frasier et al., 2007a). Les organisations qui s'inquiètent au sujet des espèces en voie de disparition et de la conservation du milieu marin ont attiré l'attention du public sur le triste sort réservé aux baleines noires, ce qui a donné lieu à la prise de mesures réglementaires tant aux États-Unis qu'au Canada en vue de protéger ces animaux des collisions avec des navires et, aux États-Unis, pour les protéger de l'enchevêtrement dans les engins de pêche.
Répartition
Aire de répartition mondiale
Historiquement, les baleines noires de l'Atlantique Nord ont été présentes presque partout dans l'Atlantique Nord, mais pas dans les eaux arctiques ou tropicales. Leur aire de répartition comprend les eaux du golfe du Saint-Laurent et de l'Atlantique canadien (jusqu'au Labrador) et s'étend vers l'est jusqu'au sud du Groenland, de l'Islande et de la Norvège et vers le sud, le long de la côte européenne jusqu'au nord-ouest de l'Afrique (Schevill et Moore, 1983; Braham et Rice, 1984; Brown, 1986; IWC, 1986; Mead, 1986; Mitchell et al., 1986). Elles sont maintenant rares ou absentes dans la majeure partie de l'est de l'aire de répartition, où elles ont déjà été relativement communes, notamment dans le nord-ouest de l'Afrique, le golfe de Gascogne, et le nord-ouest de l'Europe jusqu'en Norvège et en Islande, vers l'ouest (Reeves et al., 2007). Dans l'ouest de l'Atlantique, elles sont toujours présentes dans la majeure partie de leur aire de répartition historique.
Des relevés effectués depuis la fin des années 1970 ont permis d'identifier six zones (cinq d'entre elles font partie de l'habitat essentiel désigné) qui ont une importance saisonnière pour les baleines noires, le long de la côte est de l'Amérique du Nord (figure 3). Quatre de ces zones se trouvent aux États-Unis (au sud-est des États Unis, dans la baie du cap Cod, le Grand chenal Sud et le banc Georges/golfe du Maine; Kraus et Kenney, 1991; NMFS, 1994) et deux, au Canada (bassin de Grand Manan dans la baie de Fundy et bassin Roseway dans la portion ouest de la plate forme Néo Écossaise [Brown et al., 2009]). Les baleines noires de l'Atlantique Nord sont aussi présentes dans Jeffreys Ledge, au large du Massachusetts (Weinrich et al., 2000) et leur présence a été signalée, à l'occasion, en grands nombres dans d'autres zones, par exemple dans le détroit de Rhode Island, au printemps de certaines années (p. ex. en 1998, en 2010 et en 2011; Kenney, 2010; données d'observations inédites du NMFS, 2010).
La mise bas des baleines noires de l'Atlantique Nord a lieu normalement dans des eaux relativement chaudes à l'extrémité sud de leur aire de répartition, de novembre à avril. La plupart des mises bas ont lieu dans les eaux côtières de la Georgie et de la Floride (Kraus et Brown, 1992), mais des observations récentes portent à croire que quelques-unes ont lieu ailleurs et, probablement, un peu plus tard dans l'année (Brown et al., 2001; Patrician et al., 2009).
Les paires mère-petit passent environ 1 à 2 mois dans les aires de mise bas (Fortune et al., 2013) avant de se diriger vers le nord, le long de la côte, pour se nourrir dans le Grand chenal Sud et la baie du cap Cod, au printemps. La plupart des baleines finiront par passer l'été à l'embouchure de la baie de Fundy, de la fin de juillet à la mi-octobre. Cependant, des baleines noires y ont été aperçues dès le mois de mai et jusqu'en décembre (Stone et al., 1988; Murison et Gaskin, 1989; Woodley et Gaskin, 1996; Baumgartner et Mate, 2003; Mellinger et al., 2007). Une deuxième pouponnière estivale et automnale pourrait exister entre le cap Farewell (Groenland) et l'Islande (Reeves et Mitchell, 1986; Knowlton et al., 1994).
On sait que des individus de tous les groupes démographiques sont présents dans l'aire de mise bas, mais les femelles gestantes et parturientes semblent y passer plus de temps que les autres baleines (Fortune et al., 2013). La plupart des baleines noires non gestantes passent l'hiver ailleurs – mais on ne connaît pas les endroits fréquentés par les femelles sans baleineaux (« au repos») (Kraus et Rolland, 2007). Le bassin Jordan, situé dans le golfe du Maine entre la plate-forme littorale du Nord et la plate-forme littorale de l'Est, semble être une importante aire de reproduction (Cole et al., 2013).
Aire de répartition canadienne
Dans les eaux canadiennes, on retrouve régulièrement des groupes de baleines noires à l'embouchure de la baie de Fundy et dans la plate forme Néo-écossaise (Mitchell et al., 1986; Winn et al., 1986) (figure 3). Des baleines noires ont aussi été aperçues dans les bassins profonds du cours inférieur du Saint-Laurent, près de la confluence du Saguenay (R. Michaud, comm. pers., 1998; de la Chenelière, comm. pers., 2010), près des îles Mingan au large de la Basse-Côte-Nord du Québec (R. Sears, comm. pers. en 1994, en 1995 et en 1998, mentionné ici, compte tenu du rapport de situation antérieur du COSEPAC), et près de Percé, en Gaspésie (N. Cadet, comm. pers.; J.F. Boulin, comm. pers.; cité dans Brown et al., 2009).
On observe régulièrement des baleines noires se nourrissant, allaitant et interagissant socialement, pendant l'été et l'automne, à l'embouchure de la baie de Fundy entre le Nouveau-Brunswick et la Nouvelle-Écosse (Kraus et al., 1982; Goodyear, 1996). Des baleines sont également observées se nourrissant et interagissant socialement dans la partie ouest de la plate forme Néo-écossaise, environ 50 km au sud de la Nouvelle-Écosse (Stone et al., 1988; Kraus et Brown, 1992; Brown et al., 1995) (Stone et al., 1988; Kraus et Brown, 1992; Brown et al., 1995). Les vocalisations de baleines noires dans la plate forme Néo-écossaise sont le plus nombreuses entre les mois d'août et d'octobre, mais elles sont aussi entendues, dans une certaine mesure, de juillet jusqu'à la fin de décembre, ce qui porte à croire que quelques individus, à tout le moins, sont encore présents au début de l'hiver (Mellinger et al., 2007).
Aucune baleine noire n'a été signalée, depuis plus de quatre siècles (c.-à-d. depuis les années 1600), dans le détroit de Belle Isle, entre le Labrador et Terre-Neuve, où elles auraient coexisté, jusqu'à un certain point, avec les baleines boréales (Balaena mysticetus) (Aguilar, 1986; Cumbaa, 1986). Même si on a longtemps pensé que les baleines noires y étaient chassées pendant l'été, et que les baleines boréales y étaient chassées, elles, de la fin de l'automne jusqu'à la fin du printemps (Cumbaa, 1986; Reeves et Mitchell, 1986), des analyses d'ADN extrait de matière osseuse trouvée dans les sites de pêche de baleiniers basques (p. ex. à Red Bay, au Labrador) indiquent que c'étaient surtout des baleines boréales qui y étaient capturées et que le détroit de Belle Isle n'était pas un important habitat pour les baleines noires (Rastogi et al., 2004). De plus, seules des preuves limitées indiquent que des baleines noires auraient été chassées dans la baie de Fundy (Reeves et Barto, 1985) et dans la plate forme Néo-écossaise (voir la chasse à la baleine noire dans le golfe du Saint Laurent mentionnée dans Mitchell et al., 1986). La répartition de l'espèce peut avoir changé au fil du temps, ou le profil d'utilisation actuel de l'habitat pourrait être celui d'une population vestige utilisant seulement la limite sud d'une aire de répartition auparavant plus étendue (Kenney et al., 2001).
Compte tenu de données historiques sur la chasse à la baleine et de données d'observations récentes, on estime que la zone d'occurrence canadienne est d'environ 1,8 million de km2. On peut estimer l'indice de zone d'occupation (IZO) de plusieurs façons : a) en utilisant seulement l'habitat essentiel désigné au Canada (le bassin de Grand Manan à l'embouchure de la baie de Fundy et le bassin Roseway dans la plate forme Néo-écossaise, les deux étant des aires d'alimentation) : environ 4 500 km2; b) en utilisant toutes les principales aires d'alimentation désignées légalement comme faisant partie de l'habitat essentiel en eaux canadiennes (le bassin de Grand Manan, le bassin Roseway) et désignées comme faisant partie de l'habitat essentiel aux États-Unis (la baie du cap Cod, le Grand chenal Sud) : environ 15 820 km2; c) en utilisant seulement l'habitat essentiel légalement désigné au large du sud-est des États-Unis (aires de mise bas) : 7 092 km2. La dernière façon convient le mieux à la définition de la plus petite zone essentielle à n'importe quel stade du cycle vital pour la survie de l'espèce, qui n'est pas nécessairement au Canada.
La proportion de la population de baleines noires de l'Atlantique Nord qui fréquente les eaux canadiennes, pendant une année donnée, est incertaine et varie probablement d'une année à l'autre. En général, on estime qu'environ 60 % des femelles allaitantes amènent leur baleineau à la baie de Fundy (Frasier et al., 2007b), tandis que le reste des femelles passent la saison d'alimentation estivale/automnale ailleurs (Malik et al., 1999). Il est possible que tous les individus de la population de l'Atlantique Nord viennent ou passent dans les eaux canadiennes à un moment donné, la plupart des années.
La répartition et l'abondance des baleines noires de l'Atlantique Nord sont évaluées dans les eaux états-uniennes depuis 1978 et, dans les eaux canadiennes, depuis 1979. Des relevés aériens et maritimes ont été effectués régulièrement durant l'été et l'automne, lorsqu'il y a de fortes concentrations d'individus à l'embouchure de la baie de Fundy et dans le bassin Roseway (tableau 1). Ces relevés ont commencé dans le cadre du Cetacean and Turtle Assessment Program (CETAP, 1982), mené en eaux états-uniennes et canadiennes. Les relevés ciblant la répartition et la démographie des baleines noires dans les eaux canadiennes ont été effectués principalemenent par le New England Aquarium, à partir de 1980 dans la baie de Fundy (annuellement) et de 1981 dans le bassin Roseway (fréquemment, mais pas annuellement) (Hamilton et al., 2007). Parmi les autres relevés aériens en eaux canadiennes, on retrouve ceux effectués par l'University of Rhode Island en 1987 et en 1989 (dans le cadre d'un projet de pose de radioémetteurs) et en 1992 (à la recherche d'individus qui n'étaient pas dans le Grand chenal Sud). En 1998, East Coast Ecosystems a réalisé des relevés de la plate forme Néo-écossaise et des environs de la baie de Fundy. La National Oceanic and Atmospheric Administration (NOAA) des États-Unis a aussi effectué des relevés aériens et maritimes dans la baie de Fundy et la plate forme Néo-écossaise, certaines années. Depuis 2006, des relevés ont été réalisés de façon collaborative au sud de la Gaspésie par le Canadian Whale Institute, le Centre d'études et de protection de la baleine noire du Saint-Laurent et le New England Aquarium.
Outre les relevés maritimes et aériens, des hydrophones autonomes ont été déployés pour détecter la présence de baleines noires dans les zones historiquement importantes, comme le sont le bassin Roseway et la plate forme Néo-écossaise (Brown et al., 1995; Mellinger et al., 2007). Des études au moyen de bouées acoustiques ont aussi été réalisées dans la baie de Fundy (Laurinolli et al., 2003), et les déplacements et vocalisations d'individus ont été surveillés dans les eaux canadiennes au moyen d'étiquettes de collecte de données à ventouse (p. ex. Nowacek et al., 2004), d'appareils enregistreurs de la profondeur (p. ex. Baumgartner et Mate, 2003) et d'étiquettes émettrices (p. ex. Mate et al., 1997; Baumgartner et Mate, 2005).
Habitat
Besoins en matière d'habitat
Les baleines noires de l'Atlantique Nord nagent à des profondeurs variées et à des distances plus ou moins grandes de la côte, notamment dans les eaux côtières peu profondes, les eaux côtières profondes et les eaux extracôtières (figure 4). Les femelles gestantes et allaitantes fréquentent les eaux côtières peu profondes dans les parties sud de l'aire de répartition de novembre environ à avril. La chaleur relative (environ 20 ºC) des eaux côtières au large de la côte sud-est des États-Unis pourrait permettre aux baleineaux de consacrer moins d'énergie à la thermorégulation et plus à la croissance (Keller et al., 2006). La migration vers ces eaux pourrait aussi réduire les risques de prédation par le grand requin blanc (Carcharodon carcharias) et l'épaulard (Orcinus orca) (Kenney, 2002; Reeves et Kenney, 2003; Ford et Reeves, 2008; Hamilton et Cooper, 2010; Cassoff et al., 2011).
On connaît quatre principales aires d'alimentation : la baie du cap Cod (Massachusetts), le Grand chenal Sud (Massachusetts), le bassin de Grand Manan et le bassin Roseway. De janvier à la mi-mai, les baleines noires se rassemblent pour se nourrir dans les eaux peu profondes de la baie du cap Cod, où les vents, les courants et la stratification thermique entraînent la concentration de zooplancton en nappes denses à la surface (DeLorenzo Costa et al., 2006). Elles se nourrissent aussi dans le Grand chenal Sud (Massachusetts) d'avril à juillet (Kenney et al., 1986), et dans la baie de Fundy et le bassin Roseway de la fin de juillet à la mi-octobre (Baumgartner et al., 2003). Il s'agit de bassins profonds entourés d'eaux peu profondes, et les copépodes sont concentrés dans ces zones en raison de la convergence, de la remontée des eaux froides et d'autres courants (Wishner et al., 1988; Kenney et al., 1995; Kenney et Wishner, 1995).
Pour que les baleines réussissent à se nourrir adéquatement, on estime qu'un seuil de densité de proies suffisant est nécessaire (Kenney et al., 1986; Mayo et al., 2001). On sait que, dans le bassin de Grand Manan, les baleines noires plongent à des profondeurs de 90-150 m (près du plancher océanique) dans les zones où la biomasse de copépodes est élevée. Cependant, on n'a jamais observé de baleine en train de se nourrir dans les zones où les concentrations de proies sont inférieures à 820 organismes/m3 (170 mg/m3(Murison et Gaskin, 1989). Les densités de particules mesurées en présence de baleines noires en quête de nourriture variaient d'environ 2 000 à 21 000 organismes/m3 (une moyenne d'environ 7 500 organismes/m3) dans la baie de Fundy (Baumgartner et Mate, 2003) et étaient supérieures à 1 000 organismes de zooplancton/m3 dans la baie du cap Cod (Mayo et Marx, 1990).
Tendances en matière d'habitat
L'activité humaine a détérioré l'habitat de la baleine noire de plusieurs façons. D'importantes augmentations de la circulation maritime, par exemple, ont accru le risque de collisions avec les navires (Hackett, 2003; Ward-Geiger et al., 2005) et ont contribué à l'augmentation du bruit de fond (Parks et al., 2011). Le masquage des vocalisations diminue la capacité des baleines noires à communiquer (Clark et al., 2009). Des perturbateurs du système endocrinien trouvés dans les tissus de baleines noires pourraient provoquer le dysfonctionnement du système reproducteur (Woodley et al., 1991; Weisbrod et al., 2000; Kraus et al., 2007).
Il existe des données selon lesquelles les changements environnementaux liés à l'oscillation nord-atlantique (NAO) affecteraient la qualité de l'habitat des baleines noires. La NAO a une incidence sur la vitesse et la direction du vent, la température de l'air, les chutes de pluie et l'intensité, la fréquence et la trajectoire des tempêtes (Greene et Pershing, 2000; Visbeck et al., 2001), qui, à leur tour, contrôlent l'advection des copépodes Calanus finmarchicus, la principale proie des baleines, vers les aires d'alimentation, comme le bassin de Grand Manan (Greene et Pershing, 2000; Greene et al., 2003; Greene et al., 2004). La disponibilité de C. finmarchicus varie selon la phase, positive ou négative, de la NAO, l'abondance et la prévisibilité étant plus élevées pendant les années de phase positive de la NAO (Conversi et al., 2001; Turner et al., 2006). Par conséquent, les conditions de l'habitat (c.-à-d. les densités de proies) fluctueront probablement plus fréquemment dans les prochaines années, étant donné qu'on prévoit que la périodicité de la NAO (variation entre les phases positive et négative) augmentera (Greene et Pershing, 2004).
Biologie
Cycle vital et reproduction
On pense que les femelles mettent bas dans les eaux côtières du sud-est des États-Unis, principalement entre Brunswick (Georgie) et Jacksonville (Floride), entre les mois de novembre et d'avril (Kraus et al., 1986b; Firestone et al., 2008). Les femelles parturientes et allaitantes restent dans les aires de mise bas de 1 à 2 mois (Fortune et al., 2013), puis elles quittent vers les aires d'alimentation nordiques avec leurs baleineaux du début de mars à la mi-mars (Winn et al., 1986; Kraus et Kenney, 1991; Kraus et al., 1993; Firestone et al., 2008). L'allaitement dure environ une année, mais peut s'étendre à une deuxième année (Hamilton et Cooper, 2010). Les femelles sont fécondées de nouveau une ou plusieurs années après le sevrage de leur baleineau (Knowlton et al., 1994). La gestation dure environ 12 mois (Best, 1994), ce qui laisse supposer que la reproduction a lieu durant l'hiver, lorsque les mâles adultes et les femelles sans baleineaux sont dispersés le long de la côte est des États-Unis (entre les aires de mise bas et au moins aussi loin que la partie centrale du golfe du Maine, vers le nord, et ce, jusqu'en février, voire plus tard; Cole et al., 2013).
L'âge de la première mise bas observée, déterminé à partir d'études de données de photo-identification, a servi d'indicateur de la maturité sexuelle chez les femelles et varie de 5 à 21 ans, l'âge moyen étant de 10 ans (Kraus et al., 2007). Cependant, l'âge présumé de la maturité sexuelle est probablement surestimé, en raison de la mortalité néonatale et périnatale et de la couverture limitée des relevés (Browning et al., 2010). La durée de vie reproductive des baleines noires est d'au moins 31 ans, compte tenu d'observations de 2 femelles (Kraus et al., 2007). L'âge moyen de toutes les femelles ayant des baleineaux (c.-à-d. la durée moyenne d'une génération) de 2002 à 2009 était de 16,1 ans(±1,54 d'écart type, plage de 13,1 à 18,4ans; données inédites calculées par le Right Whale Consortium, 2011). Cependant, on s'attend à ce que cet âge augmente avec le ralentissement de la croissance de la population et à mesure que l'âge de toutes les femelles plus âgées devient connu. La modélisation des populations semble indiquer qu'un âge moyen de 35,7 ans pour les femelles ayant un baleineau est approprié dans le cas d'une population stationnaire en parfaite condition, en supposant une durée de vie reproductive de 10 à 69 ans et une mortalité ou sénescence reproductive uniforme de 1 % (Taylor et al., 2007). Cette estimation de la durée d'une génération est probablement élevée, mais de meilleures estimations de la durée de vie reproductive prévue pour les baleines noires de l'Atlantique Nord ne sont pas disponibles. Par conséquent, la durée d'une génération pour cette population varie entre 16 et 36 ans.
Le taux de natalité (basé sur 106 femelles adultes et 19 baleineaux nés en 2010) était de 0,18 baleineau par femelle mature (Hamilton et Knowlton, 2011), et le nombre moyen de baleineaux nés entre 1993 et 2010 était de 17,5 (plage de 1 à 39 baleineaux nés par année).
L'examen post-mortem et l'analyse des hormones fécales sont les seuls moyens (Rolland et al., 2007) de déterminer l'âge de la maturité sexuelle chez le mâle. Des mâles de tous âges ont été observés dans des groupes en parade nuptiale (Kraus et Hatch, 2001), mais des analyses génétiques indiquent que l'âge de la première paternité serait d'environ 15 ans, ce qui porte à croire que la compétition empêche les jeunes mâles de contribuer à la reproduction (Frasier et al., 2007a).
On pense que la stratégie de reproduction des baleines noires serait partiellement fondée sur la compétition du sperme. Les mâles ont un pénis pouvant atteindre jusqu'à 2,3 m de long et des testicules dont le poids combiné peut atteindre environ 972 kg (Brownell et Ralls, 1986; Frasier et al., 2007a) – il s'agit des plus grands testicules et du plus grand rapport entre le poids des testicules et le poids corporel observés chez les mammifères, et d'un des plus grands rapports entre la longueur du pénis et celle du corps (Brownell et Ralls, 1986; Atkinson, 2002; Frasier et al., 2007a). En supposant que la grandeur des testicules et la longueur exceptionnelle du pénis sont indicateurs d'une stratégie d'accouplement impliquant la rivalité (Ginsberg et Huck, 1989; Gomendio et al., 1998; Frasier et al., 2007a), on peut penser que de toutes les espèces de baleines et probablement de tous les mammifères, c'est chez la baleine noire qu'on retrouve la plus intense compétition du sperme (Frasier et al., 2007a). Les observations de groupes en parade nuptiale appuient cette hypothèse (même si la majorité des groupes en parade nuptiale sont observés en dehors de la période de reproduction présumée; Parks et al., 2007a).
L'intervalle entre les mises bas de 3 à 5 ans (Kraus et al., 2007) signifie qu'il y a environ une femelle en ovulation pour quatre mâles adultes, ce qui entraîne une forte compétition entre les mâles pour l'accès à une femelle. Les groupes en parade nuptiale peuvent comprendre 40 baleines, voire plus, car de nombreux mâles essaient de s'accoupler avec la femelle convoitée (Kraus et Hatch, 2001). Les femelles suscitent l'attention des mâles par leurs vocalisations (Parks et Tyack, 2005). Les mâles semblent se faire concurrence pour les meilleures positions afin de pouvoir profiter de chaque occasion d'accouplement qui se présente lorsque la femelle respire (Kraus et Hatch, 2001). La femelle peut s'accoupler environ 60 fois pendant un épisode de parade avec plusieurs mâles, et, parfois, il y a double intromission (Mate et al., 2005; Parks et Tyack, 2005; Frasier et al., 2007a). Ces observations appuient davantage l'existence d'un système de reproduction basé sur la compétition du sperme (Frasier et al., 2007a).
Une femelle adulte ne donne naissance qu'à un seul baleineau à la fois, l'intervalle entre les mises bas étant très variable (intervalle moyen de 3 à 5 ans et plage de 2 à 13 ans; Kraus et al., 2007). Le sex ratio à la naissance est probablement 50:50. Cependant, en 2010, 49 % des individus excluant les baleineaux étaient des mâles, 35 % étaient des femelles et 17 % étaient de sexe indéterminé (Hamilton et Knowlton, 2011). Le sex ratio dans le cas des individus dont le sexe est connu était biaisé en faveur des mâles (58:42); il serait près de 50:50 seulement si tous les individus de sexe indéterminé étaient des femelles. Depuis les années 1990, les jeunes ont représenté de façon constante 26 à 31 % de la population des individus connus ayant plus d'un an. Les adultes constituent 65 à 69 % de la population, et le 4 à 6 % restant correspond à des individus d'âge inconnu (Hamilton et al., 1998; Hamilton et al., 2010; Hamilton et Knowlton, 2011).
Le taux de reproduction (mesuré en fonction du nombre de baleineaux nés) est très variable. Depuis 1990, le nombre de baleineaux observés chaque année varie de 1 à 39, ce qui est plus variable que les prévisions fondées uniquement sur le hasard (Kraus et al., 2007; Waring et al., 2011) (figure 5). Cela s'est avéré vrai surtout entre deux périodes (1993-1995 et 1998-2000), lorsque le nombre de naissances a été particulièrement bas (Kraus et al., 2007). Plusieurs années avec un taux de reproduction élevé ont suivi ces années de faible taux (Kraus et al., 2007). Selon certains, les fluctuations océaniques qui influent sur l'abondance de la principale proie (Calanus finmarchicus) ont à terme une incidence sur le succès reproducteur des baleines noires (Greene et Pershing 2004; Kraus et al., 2007). Cependant, les modèles actuels sont incapables de prédire les taux de reproduction futurs avec exactitude, de sorte qu'il est impossible de prévoir les tendances avec confiance.
Figure 5. Effectifs totaux des jeunes baleines noires de l'Atlantique Nord (jeunes de l'année) observés morts ou vivants chaque année, dans la période 1993-2010 (Pettis, 2010; Waring et al., 2011). Au moins quelques-uns des baleineaux morts auront aussi été observés vivants et, par conséquent, auront été « comptés » deux fois dans la figure.
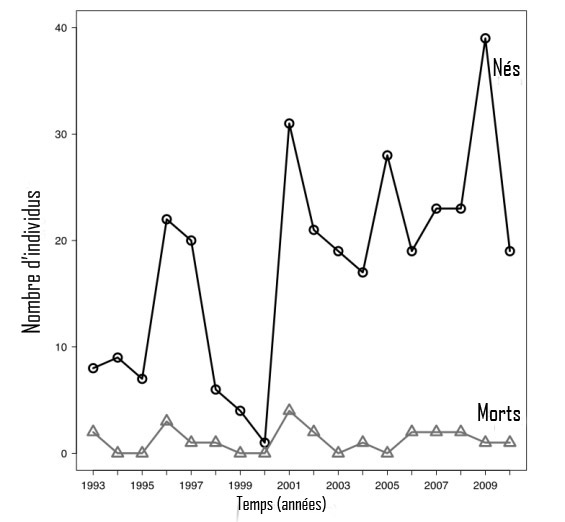
Description longue pour la figure 5
Graphique montrant les tendances des effectifs totaux des jeunes baleines noires de l'Atlantique Nord (jeunes de l'année) observés morts ou vivants chaque année, dans la période 1993-2010. Le nombre de baleineaux qui naissent est très variable. Depuis 1990, le nombre de baleineaux répertoriés annuellement s'est situé entre 1 et 39.
Les eaux côtières du sud-est des États-Unis sont largement reconnues comme étant la zone où la majorité des baleineaux naissent (Kraus et Brown, 1992). Cependant, des observations récentes de femelles accompagnées de leur très jeune baleineau ayant des caractéristiques de nouveau-né (p. ex. la petite taille de la tête, une dépression marquée du rostre et des callosités moins bien formées) semblent indiquer qu'une deuxième aire de mise bas pourrait exister plus loin, vers le nord, probablement au large du Massachusetts (Patrician et al., 2009). Des observations dans le passé d'au moins deux petits baleineaux dans la baie du cap Cod concordent avec la présence d'une aire de mise bas locale (Watkins et Schevill, 1982; Schevill et al., 1986). La présence d'une telle zone aiderait à expliquer pourquoi 25 % des femelles actives sur le plan reproducteur (c.-à-d. des femelles allaitantes) n'ont pas été observées dans l'aire de mise bas du sud-est des États-Unis de 1980 à 1992 (Brown et al., 2001).
Des taux de mortalité naturelle n'ont pas été calculés. La longévité moyenne est inconnue, mais certains adultes reconnaissables ont été aperçus sur plus de trois décennies, et la baleine noire de l'Atlantique Nord la plus âgée répertoriée aurait eu environ 70 ans lorsqu'elle a été aperçue pour la dernière fois (Kraus et Rolland, 2007).
Physiologie et adaptabilité
La croissance rapide et l'accumulation de graisse sont deux mécanismes physiologiques qui pourraient améliorer la probabilité de survie des jeunes et contribuer au succès reproducteur. Selon des modèles de croissance, les jeunes baleines noires de l'Atlantique Nord atteignent environ 75 % de leur longueur asymptotique au cours de leur première année de vie, avant le sevrage (Fortune et al., 2012). Cette rapide croissance postnatale pourrait favoriser la survie des jeunes, car elle réduit le risque de prédation et optimise la taille de la tête et de la bouche et fait ainsi augmenter le succès de la quête de nourriture, immédiatement après le sevrage. La croissance accélérée réduirait aussi les coûts de thermorégulation, car elle fait diminuer le rapport surface/volume des baleineaux.
La couche de graisse pourrait aussi améliorer le succès reproducteur des femelles adultes qui ne se nourrissent pas pendant qu'elles sont dans les aires de mise bas. Des mesures échographiques indiquent que les femelles allaitantes catabolisent la graisse et regarnissent leurs réserves après le sevrage de leur baleineau (Miller et al., 2011). La couche de graisse est la plus épaisse quelques mois avant le début de la gestation, et les femelles allaitantes sont plus maigres (en termes d'épaisseur de la couche de graisse) que les femelles non allaitantes, non gestantes (Miller et al., 2011).
On sait que l'épaisseur de la couche de graisse est corrélée avec la disponibilité des proies et le succès reproducteur (Miller et al., 2011). La variabilité annuelle des intervalles entre les mises bas concorde avec les changements de l'épaisseur de la couche de graisse observés dans le milieu des années 1990, et les fluctuations de réserves de graisse coïncident avec les changements environnementaux à grande échelle qui contrôleraient la qualité et la quantité des proies. On a observé une diminution de la couche de graisse après le déclin dramatique de l'indice NAO en 1996. Inversement, l'épaisseur de la couche de graisse a augmenté pendant les années de disponibilité modérée des ressources.
Des tendances semblables relativement à l'épaisseur de la couche de graisse et à la disponibilité des proies ont été observées chez des individus immatures. Par exemple, la couche de graisse des baleines d'un an était significativement plus épaisse en 2002 qu'elle ne l'était en 1998 (Miller et al., 2011). Plusieurs hypothèses ont été émises pour expliquer la variabilité de l'épaisseur de la couche de graisse chez les jeunes baleines, notamment une meilleure nutrition pendant l'allaitement; une durée d'allaitement plus longue, des baleineaux d'un an ayant été observés près de leur mère bien après qu'ils aient eu un an, et les différences en matière d'abondance du Calanus finmarchicus entre les années 1998 et 2002 (Hamilton et Cooper, 2010; Miller et al., 2011).
Déplacements et dispersion
Les baleines noires commencent leur migration vers le nord à la fin de l'hiver et au début du printemps. Selon les modèles statistiques de migration, les paires mère-petit arrivent à la pointe de Long Island (New York) environ 21 à 24 jours après avoir quitté les eaux côtières de la Floride (en supposant qu'elles se déplacent à 2,8 3,1 km/h; Firestone et al., 2008). Des baleines noires de toutes les classes d'âge se réunissent pour se nourrir et interagir socialement dans la baie du cap Cod, au milieu de l'hiver et au printemps, et dans le Grand chenal Sud, à l'est du cap Cod (Winn et al., 1986; Hamilton et Mayo, 1990; Kenney et al., 1995) au printemps. En juin et en juillet, les baleines se déplacent vers les aires d'alimentation situées à l'embouchure de la baie de Fundy et dans l'ouest de la plate forme Néo-Écossaise, où elles se nourrissent de façon intense, en août-septembre. (Mitchell et al., 1986; Winn et al., 1986; Baumgartner et Mate, 2003; Mellinger et al., 2007). Les périodes de résidence dans la baie de Fundy durant l'été et l'automne sont variables et peuvent dépendre des conditions de l'océan et de la qualité des proies (Baumgartner et Mate, 2005).
Des baleines reconnaissables individuellement ont été répertoriées dans le golfe du Saint-Laurent ainsi que dans le bassin du Labrador au cours de l'été (Knowlton et al., 1992; Hamilton et Martin, 1999). Elles sont présentes à tout le moins occasionnellement pendant l'été dans l'est de la plate forme Néo-écossaise (Mitchell et al., 1986), dans l'estuaire du Saint-Laurent près de la confluence du Saguenay (1998), au large des îles Mingan le long de la Basse-Côte-Nord du Québec (1994, 1995 et 1998) et près de l'embouchure de la baie des Chaleurs, au sud de la Gaspésie (1995-1998 et 2000-2006). En 2001, une baleine noire a été trouvée morte dans le golfe du Saint-Laurent, près des îles de la Madeleine (données inédites, New England Aquarium; Brown et al., 2009), et une autre baleine (prise dans les engins de pêche) a été suivie par satellite; elle s'est déplacée le long de l'est de la plate forme Néo-écossaise, puis est entrée dans le golfe du Saint-Laurent, s'est rendue jusqu'aux îles de la Madeleine, puis est revenue vers la plate forme Néo-écossaise pour enfin se diriger vers le sud, jusqu'au golfe du Maine (données inédites, Provincetown Center for Coastal Studies; Brown et al., 2009).
Une migration vers le sud commence en octobre, certains individus passant dans le golfe du Maine et au large du cap Cod (Winn et al., 1986). Des attroupements de baleines noires sont parfois observés à l'automne, à Jeffreys Ledge (Weinrich et al., 2000), à Cashes Ledge et à Platts Bank (P. Clapham, comm. pers. 2003) (Waring et al., 2011).
Quelques déplacements transatlantiques ont été répertoriés. Au cours de l'automne de 1999, un mâle adulte a parcouru 5 700 km de la baie du cap Cod jusqu'au nord de la Norvège en 117 joursenviron (49 km/jour ou 2 km/h)(Jacobsen et al., 2004). Aucune baleine noire n'avait été aperçue en eaux norvégiennes depuis 1926 (Jonsgard, 1977; Smith et al., 2006). Un autre individu répertorié dans le catalogue de l'ouest de l'Atlantique Nord a été observé dans les Açores, le 9 janvier 2009(Silva et al., 2012).
Alimentation et interactions interspécifiques
Les baleines noires se nourrissent principalement de copépodes calanoïdes et, à l'occasion, d'Euphausiacés et de larves de balanes communes (Mayo et Marx, 1990). Durant l'hiver, elles se nourrissent dans la baie du cap Cod de nombreuses espèces de copépodes (p. ex. Calanus finmarchicus, Centropages typicus, Centropages hamatuset Pseudocalanus spp.). Cependant, au cours du printemps dans la baie du cap Cod et pendant l'été et au début de l'automne dans la baie de Fundy, elles se nourrissent presque exclusivement de C. finmarchicus aux stades de développement où ils sont riches en huile (Murison et Gaskin, 1989; Mayo et Marx, 1990; Mayo et al., 2001; Baumgartner et al., 2003; Baumgartner et Mate, 2003).
Le rorqual boréal (Balaenoptera borealis) (Baumgartner et al., 2011), le requin pèlerin (Cetorhinus maximus) (p. ex. dans Sims et Quayle, 1998), le hareng (Clupea harengus) (p. ex. dans Checkley Jr, 1982) et le lançon (Ammodytes spp.) (p. ex. dans Monteleone et Peterson, 1986) ont des régimes alimentaires semblables à celui des baleines noires et sont considérés comme d'éventuels compétiteurs. Les espèces comme le C. finmarchicus peuvent se cacher des poissons planctonophages, tels le hareng et le lançon, et des baleines planctonophages, tel le rorqual boréal, dans les zones peu éclairées. La capacité des baleines noires à repérer et à exploiter les « bancs » de copépodes à des profondeurs considérables porte à croire que le lançon, le hareng, le rorqual boréal et la baleine noire sont principalement en compétition pour les copépodes dans les eaux de surface (Baumgartner et al., 2011).
Taille et tendances des populations
Activités et méthodes d'échantillonnage
Le nombre d'individus dont on sait (ou présume) qu'ils sont vivants est considéré comme la meilleure estimation de la taille de la population, mais il faut, toutefois, considérer cette estimation comme un minimum (p. ex. dans Waring et al., 2011). De façon générale, les chercheurs ont conclu que, pour cette population, il n'y a pas moyen de générer un nombre maximal crédible avec une estimation connexe de la plage d'incertitude (Hamilton et al., 2007). Le nombre d'individus vivants comprend les baleines marquées individuellement qui ont été photographiées dans les six dernières années et comparées (au moyen de la disposition des callosités) avec les données de la base d'identification, tenue à jour par le New England Aquarium (Hamilton et al., 2007). Les individus qui n'ont pas été aperçus pendant plus de six ans sont présumés morts. On reconnaît qu'un petit nombre des baleines « présumées mortes » sont peut être encore vivantes, et aussi que certaines baleines « irrégulières » n'ont probablement pas été photographiées (Hamilton et al., 2007).
Les activités d'échantillonnage annuelles réalisées dans l'habitat essentiel désigné de la baleine noire ont été nombreuses, même si elles n'ont pas été réparties de façon uniforme. Certaines zones, p. ex. le bassin Roseway, ne font pas l'objet de relevés chaque année (tableau 1), et le nombre de relevés effectués chaque année est différent d'une zone à l'autre. En particulier, l'échantillonnage a été nettement plus intensif dans les aires de mise bas (de nombreux relevés y sont effectués hebdomadairement) que dans les aires d'alimentation (la baie du cap Cod a fait l'objet d'un relevé environ par semaine au cours des dernières années). La majorité des activités, spécialement celles menées dans les aires de mise bas, étaient des activités de surveillance aérienne dans le but d'informer les navigateurs de la position en temps quasi réel des baleines noires et de les aider ainsi à éviter les collisions navires-baleines.
| Zone des relevés | Institution | Période |
|---|---|---|
| Du Cap Hatteras (É.-U.) à la plate-forme Néo-Écossaise | Cetacean and Turtle Assessment Program, University of Rhode Island | 1979 à 1982 |
| Baie de Fundy | New England Aquarium | 1980 à 2011 |
| Bassin Roseway | New England Aquarium | 1981, 1983 à 1985, 1986 à 1991, 2004 à 2005 2009 à 2010 |
Des relevés visuels suivant des plans systématiques ont été effectués dans la baie de Fundy depuis 1980 et dans le bassin Roseway depuis 1981 (Brown et al., 2007), avec des données manquantes pour les années où des relevés n'ont pas été effectués (tableau 1). Les activités d'échantillonnage ont couvert une zone environ trois fois plus grande dans le bassin de Grand Manan dans la baie de Fundy que dans le bassin Roseway (90 812 km ont fait l'objet de relevés systématiques dans la baie de Fundy et 28 563 km ont fait l'objet de relevés dans le bassin Roseway entre octobre 1978 et 2010). Les critères suivants ont été appliqués à ces estimations des activités d'échantillonnage : 1) les données ont été recueillies de façon systématique (relevés aériens ou maritimes); 2) l'ensemble de données complet a été soumis au North Atlantic Right Whale Consortium; 3) au moins un observateur était officiellement au poste; 4) la visibilité était d'au moins 3,7 km et l'état de la mer était de niveau 4 ou meilleur à l'échelle Beaufort; et, dans le cas des relevés aériens, 5) l'altitude était inférieure à 365,76 m (comm. pers., Kenney, 2011; Correspondance par courriel adressée à S. Fortune, septembre, 2011, Associate Marine Research Scientist, Professor-in-Residence, University of Rhode Island, Rhode Island, États-Unis).
Abondance
Tel qu'il est indiqué dans le Manuel des opérations et des procédures du COSEPAC (août 2013, annexe C, p. 149), le nombre d'individus matures est le nombre d'individus connus, estimés ou inférés comme étant capables de se reproduire. Cela signifie qu'il faut, lorsque c'est possible, corriger le nombre total d'individus adultes vivants pour tenir compte des individus qui ne se reproduiront pas. En ce qui concerne les baleines noires de l'Atlantique Nord, il faut prendre en considération deux classes d'individus adultes. Premièrement, il ne faut pas compter les individus matures qui ne produiront jamais de nouvelles recrues (p. ex. les individus stériles ou sénescents sur le plan de la reproduction). Deuxièmement, en raison du sex ratio biaisé chez les individus adultes (plus de mâles que de femelles), il convient d'utiliser des estimations basses pour le nombre d'individus matures afin de tenir compte de cette situation (Manuel des opérations et des procédures du COSEPAC, p. 149).
Le nombre total de baleines noires de l'Atlantique Nord que l'on croyait vivantes en 2010 était de 468. Ce nombre comprenait 19 baleineaux de l'année et 449 individus excluant les baleineaux (individus de plus de 1 an) (Pettis, 2010; Hamilton et Knowlton, 2011) (figure 5). Un petit nombre de jeunes n'étaient pas reconnaissables individuellement et n'ont donc pas été inclus dans le catalogue ni dans l'estimation de la taille de la population totale (probablement moins de 10 individus, compte tenu des données d'années précédentes). En 2010, la population de 468 baleines noires comptait 4 % de baleineaux (n = 19), 27 % de jeunes baleines (ayant de 1 à 8 ans) (n = 124), 65 % de baleines adultes (ayant 9 ans ou plus) (n = 305) et 4 % d'individus d'âge indéterminé (n = 20) (Hamilton et Knowlton, 2011). Ces derniers sont probablement des individus plus âgés qui n'ont pas été observés pendant l'année de leur naissance, mais l'information disponible est insuffisante pour déterminer s'il s'agit de baleines adultes ou de jeunes baleines. Par conséquent, la population totale d'adultes en 2010 se situait probablement entre 305 et 325 individus.
Compte tenu de la forme de la courbe de découverte (le nombre d'individus excluant les baleineaux ajoutés annuellement à la base de données d'identification photographique a diminué de façon constante au fil du temps; figure 6), il semble que la majorité de la population ait été identifiée à l'aide de photographies. Il convient de noter que ni la figure 6 ni la figure 7 ne doivent être interprétées comme représentant le nombre réel ou supposé d'individus vivants dans une année donnée, entre 1980 et 2009. Le plafonnement de la courbe dans le bas de la figure 6 semble plutôt indiquer que les chercheurs ont découvert tous les individus adultes de la population, tandis que la courbe croissante dans le haut de la figure 6 indique que les nouveaux individus ajoutés au catalogue sont presque tous de nouvelles recrues – c. à d. des baleineaux. Ces courbes de découverte ne sont pas des estimations de l'abondance et ne tiennent pas compte des individus qui sont morts quelque temps après leur « découverte » et ont donc été enlevés du catalogue. Sur les 449 individusexcluant les baleineaux, présumés vivants en 2010, 49 %étaient des mâles, 35 % étaient des femelles et 17 %étaient des individus de sexe indéterminé (Hamilton et Knowlton, 2011).
Figure 6. Nombre cumulatif de baleineaux (0-1 an) et d'individus excluant les baleineaux (> 1 an) de la baleine noire de l'Atlantique Nord qui ont été ajoutés au catalogue du New England Aquarium dans la période 1980-2009 (Hamilton et al., 2007). Il faut noter que le nombre de baleineaux ajoutés au catalogue ne correspond pas au nombre de baleineaux nés chaque année, parce que l'identification des baleineaux nouvellement nés est une tâche difficile (en raison des callosités qui ne sont pas encore complètement formées). Nombre de baleineaux sont identifiés seulement plus tard (dans la baie de Fundy, lorsqu'ils ont environ 6 mois), une fois que la disposition des callosités est mieux développée. Par conséquent, il n'est pas inhabituel qu'un baleineau né une année civile ne soit photo identifié pour la première fois que l'année civile suivante et qu'il soit encore classé comme baleineau à ce moment-là. Il faut également noter que, dans la catégorie des individus excluant les baleineaux, on retrouve seulement les animaux qui n'ont pas été observés pour la première fois, ni photo identifiés lorsqu'ils étaient des baleineaux. Le plafonnement de la courbe pour les individus excluant les baleineaux semble indiquer que tous (ou presque tous) ces individus ont maintenant été identifiés et que tous les nouveaux individus sont identifiés pour la première fois lorsqu'ils sont des baleineaux.
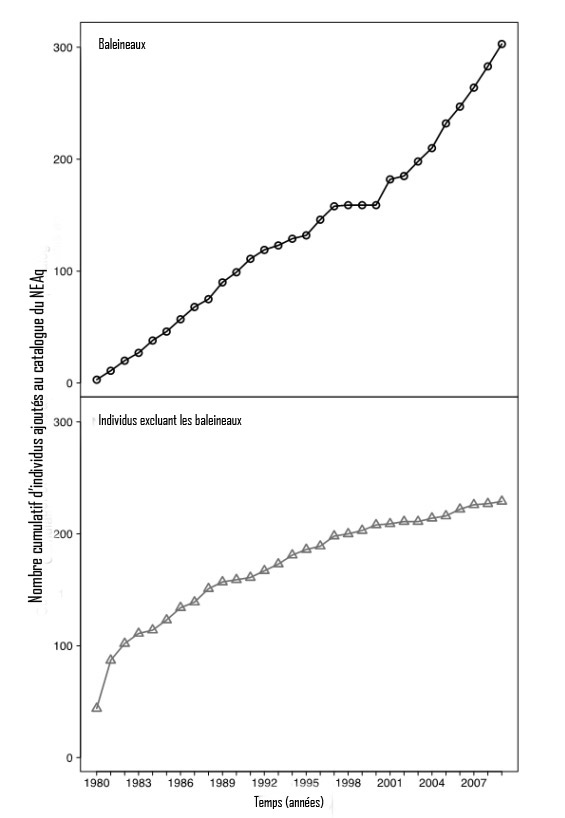
Description longue pour la figure 6
Deux graphiques montrant la tendance du nombre cumulatif de baleineaux (0-1 an; graphique du haut) et d'individus excluant les baleineaux (> 1 an; graphique du bas) de la baleine noire de l'Atlantique Nord qui ont été ajoutés au catalogue du New England Aquarium dans la période 1980-2009. Le nombre d'individus adultes nouvellement identifiés a peu fluctué d'une année à l'autre, tandis que le nombre de baleineaux nouvellement identifiés a augmenté de façon plus marquée, sauf dans la période de 1993 à 1995.
Figure 7. Graphique du haut : Nombre annuel de baleines noires de l'Atlantique Nord adultes (≥ 9 ans) présumées vivantes entre 1980 et 2009 (en fonction des individus identifiés dans le catalogue d'identification photographique; Right Whale Consortium, 2011). Le nombre total réel est probablement plus grand que le nombre indiqué, parce qu'il est possible que le catalogue d'identification photographique n'inclue pas les individus adultes qui ne fréquentent ou n'occupent pas régulièrement les zones ayant fait l'objet de relevés. De plus, les individus qui n'ont pas été observés et identifiés pendant une période de six ans ou plus sont exclus, même si certains d'entre eux sont peut-être encore vivants. Graphique du bas : Relation linéaire représentant le nombre d'individus adultes présumés vivants entre 1997 et 2009 (période pendant laquelle environ 75-100 % du nombre total des individus adultes étaient présumés vivants).
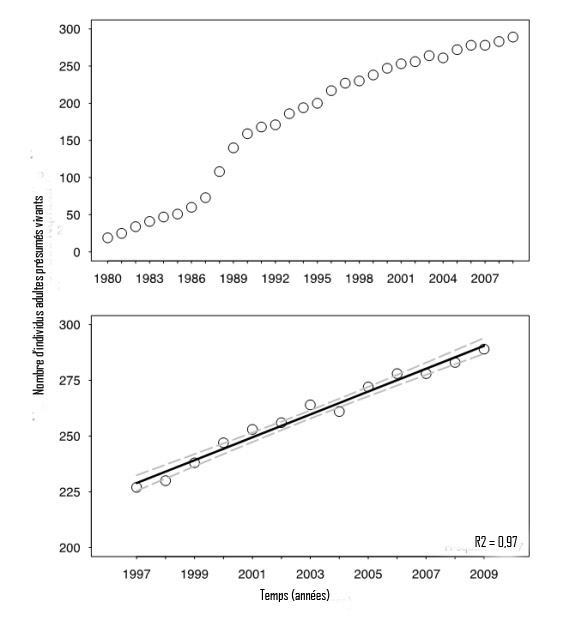
Description longue pour la figure 7
Deux graphiques montrant la tendance du nombre de baleines noires de l'Atlantique Nord adultes (9 ans ou plus) présumées vivantes. Le graphique du haut montre le nombre annuel de baleines adultes présumées vivantes entre 1980 et 2009, en fonction des individus identifiés dans le catalogue d'identification photographique. Le graphique du bas montre la relation linéaire représentant le nombre d'individus adultes présumés vivants entre 1997 et 2009.
Le nombre réel de baleines noires matures vivantes en 2010 est inférieur à la somme de tous les individus pouvant être adultes (c.-à-d. 305 plus les 20 individus catalogués d'âge indéterminé = 325 individus), parce que le sex ratio est biaisé de façon significative en faveur des mâles (58 %:42 %). Quelques-uns des mâles adultes peuvent donc être considérés comme un surplus en ce qui concerne les besoins en matière de reproduction de la population.
Si on applique le sex ratio des animaux de sexe connu au nombre maximal de 325 individus adultes estimés vivants en 2010, cela porte à croire qu'il y avait 188 mâles matures et 136 femelles matures; la taille effective de la population serait donc de 272 individus matures si on corrige le nombre de mâles pour qu'il soit égal ou inférieur au nombre de femelles (= 2 x 136 femelles). Cependant, le nombre d'individus matures sera plus bas si certaines des femelles matures sont nullipares, c.-à-d. incapables de se reproduire. On croit qu'environ 10 % des femelles adultes (12 sur 125) étaient nullipares dans la période 1989 2003 (Browning et al., 2010). Si 10 % des femelles n'ont jamais mis bas, cela réduit la taille effective de la population de 272 à 245 baleines noires matures (= 2 x 122,5 femelles). Cependant, l'hypothèse selon laquelle 10 % des femelles de plus de 10 ansseraient nullipares pourrait être trop élevée, étant donné que les taux de mise bas dans la population étaient relativement bas dans la période 1989-2000 (figure 5) et que 11 des 12 femelles nullipares identifiées (1989-2003) ont été aperçues dans les aires de mise bas, ce qui indiquerait qu'elles ont probablement été fécondées, mais qu'elles ont, soit aborté, soit perdu leur baleineau en route vers les aires de mise bas (Browning et al., 2010). Il est donc raisonnable de supposer que la taille effective de la population aurait varié entre 245 et 272 individus matures en 2010.
Fluctuations et tendances
La structure d'âge de la population de baleines noires de l'Atlantique Nord est restée relativement stable au fil du temps, et le nombre d'individus adultes nouvellement identifiés a peu fluctué d'une année à l'autre (figure 6). Parmi tous les individus excluant les baleineaux (individus ayant plus de 1 an) qu'on retrouve dans le catalogue d'identification photographique, les jeunes baleines ont représenté avec constance de 26 à 31 % de la population depuis les années 1990; les individus adultes, eux, ont représenté de 65 à 69 % de la population, le 4 à 6 % restant représentant les individus d'âge indéterminé (Hamilton et al., 1998; Hamilton et Cooper, 2010; Hamilton et al., 2010; Hamilton et Knowlton, 2011).
Le nombre de baleineaux qui naissent est très variable. Depuis 1990, le nombre de baleineaux répertoriés annuellement s'est situé entre 1 et 39 (Kraus et al., 2007; Waring et al., 2011) (figure 5). Ce nombre a été particulièrement bas de 1993 à 1995 et de 1998 à 2000. Ces périodes de faible production de baleineaux ont été suivies de plusieurs années où le taux de reproduction a été élevé (Kraus et al., 2007). Le succès reproducteur peut dépendre de la disponibilité de la nourriture (Greene et Pershing, 2004; Kenney, 2007; Kraus et al., 2007; Hlista et al., 2009; Browning et al., 2010). Malgré la variabilité interannuelle du taux de natalité, il y a eu une augmentation globale du nombre de baleineaux ajoutés annuellement dans le catalogue d'identification photographique (figure 6).
La population (individus ayant plus de 1 an) semble avoir subi des phases de croissance et de décroissance depuis les années 1980. On estime qu'elle a augmenté d'environ 2,5 % (erreur-type de 0,3 %) par année de 1986 à 1992 (Knowlton et al., 1994). Cependant, plus tard dans les années 1990, il semble qu'elle aurait quelque peu diminué, ou du moins cessé d'augmenter au rythme précédent, en raison d'une production de baleineaux relativement faible et d'un taux élevé de mortalité des individus adultes (Caswell et al., 1999; Fujiwara et Caswell, 2001; Kraus et al., 2007).
En 2003, on comptait 322 individusconnus (excluant les baleineaux) (Right Whale Consortium, 2011), ce nombre ayant augmenté à 396 en 2007 (Waring et al., 2011), à 417 en 2009 (Hamilton et al., 2010) et à 449 en 2010 (Hamilton et Knowlton, 2011) (figure 7). Le nombre d'individus excluant les baleineaux aurait donc augmenté d'environ 28 %depuis 2003 (figure 7). Cependant, le nombre d'individus matures n'a probablement augmenté que de 22 à 27 % environ (en supposant qu'il y avait 222 à 238 individus matures en 2003). L'effectif total (comprenant tous les individus) aurait donc doublé entre 1990 et 2010.
Le taux brut moyen d'augmentation (compte tenu des variations annuelles du nombre minimal d'individus vivants) a été de 2,4 % entre 1990 et 2007 (Waring et al., 2011). Plus récemment, Waring et al.(2013) ont obtenu un taux de croissance de la population de 2,6 % de 1990 à 2009. Ces augmentations sont inférieures aux taux annuels d'augmentation de deux populations de baleines franches australes : 6,8 % en Afrique du Sud (1971-1998; n = 3,104 baleines en 1997; Best et al., 2001; IWC, 2001a) et 6,9 % en Argentine (1971-1990; Cooke et Rowntree, 2001).
La taille de la population de baleines noires de l'Atlantique Nord avant qu'elles ne soient exploitées est inconnue. La détermination rétroactivement de la taille de la population à partir des données historiques sur la chasse à la baleine s'avère difficile (Reeves et al., 2007). Entre 1634 et 1950, au moins 5 500 (maximum d'environ 11 000) baleines noires ont été tuées dans l'ouest de l'Atlantique Nord, et on en déduit que les effectifs étaient probablement d'au moins quelques milliers avant que l'exploitation ne commence au début des années 1600 (Reeves et al., 2007).
Des analyses de l'ADN mitochondrial (ADNmt) de plus de 200 baleines échantillonnées dans l'ouest de l'Atlantique Nord semblent indiquer qu'il n'y a, actuellement, que six groupes matriarcaux (ou haplotypes d'ADNmt) et que la taille de la population a subi un effet de goulot d'étranglement (Malik et al., 1999; McLeod et al., 2010). Cet effet aurait pu se produire avant la pêche par les baleiniers basques dans les années 1500, compte tenu du petit nombre d'os de baleines noires récupérés dans les sites archéologiques de stations de pêche de baleiniers basques (Rastogi et al., 2004; McLeod et al., 2010). Neuf de ces stations, utilisées au cours du 16e siècle, étaient situées le long du Québec et du Labrador dans le détroit de Belle-Isle (Barkham, 1978; Braham et Rice, 1984). Des analyses génétiques ont été effectuées sur 21 humérus déterrés à Red Bay, au Labrador, dans un galion basque (coulé en 1565). Sur ces 21 humérus, 1 provenait d'une baleine noire et 20 de baleines boréales (Rastogi et al., 2004). Ce résultat signifierait que la petite taille actuelle de la population dans l'ouest de l'Atlantique Nord n'est pas un phénomène récent (Rastogi et al., 2004; Frasier et al., 2007a; Frasier et al., 2007b; McLeod et al., 2010).
L'effet de goulot d'étranglement génétique a probablement eu lieu avant les années 1500, à la suite d'un changement des conditions environnementales, qui a modifié la qualité et la quantité des proies (Frasier et al., 2007b). Selon certains, la dernière glaciation majeure (qui remonterait à 300 000 ans et aurait duré jusqu'à il y a 10 000 ans) est responsable du changement des conditions biotiques et abiotiques (Frasier et al., 2007b). Même si une telle hypothèse ne peut pas être vérifiée, des observations récentes de baleines noires en mauvais état de santé et montrant des signes d'une faible diversité génétique portent à croire que des facteurs nutritionnels et génétiques pourraient être en train d'influer sur le taux de rétablissement de la population (Frasier et al., 2007b; Brown et al., 2009).
Immigration de source externe
Les baleines noires qui passent le printemps, l'été et l'automne, à se nourrir dans les eaux canadiennes appartiennent à la même population que les baleineaux qu'on rencontre au sud-est des États-Unis. Les baleines noires sont considérées comme essentiellement disparues de l'est de l'Atlantique Nord. Par conséquent, il n'y a aucune possibilité d'une immigration de source externe.
Menaces et facteurs limitatifs
Collisions avec des navires et enchevêtrement dans les engins de pêche
Parmi les facteurs limitant la croissance des populations, ce sont les collisions avec des navires et les enchevêtrements dans des engins de pêche qui sont le mieux documentés. Une moyenne de 2,6 baleines par année ont subi des blessures graves ou sont mortes, soit à cause de collisions avec des navires ou d'enchevêtrements, entre les années 2005 et 2009 (Waring et al., 2011). Ces données sont fondées sur l'inspection de carcasses et constituent probablement une estimation minimale de la mortalité réelle causée par l'humain. Cela équivaut à environ 0,6 % du nombre total de baleines noires de l'Atlantique Nord que l'on croyait vivantes en 2010. Selon les résultats de nécropsies effectuées sur les carcasses de 75 baleines noires, entre 1970 et 2007, 48 % (36 individus) des baleines sont mortes d'une cause anthropique (11 % à cause d'enchevêtrements dans des engins de pêche et 37 % à cause de collisions avec des navires), 24 % (18 individus) étaient des baleineaux nouvellement nés et 28 %(21 individus) des baleines sont mortes de causes inconnues (Knowlton et Kraus, 2001; Moore et al., 2004; Brown et al., 2009). Dans la période de 1987 à 2006, 25 %de toutes les morts confirmées dues à des collisions avec des navires se sont produites en eaux canadiennes (Brown et al., 2009).
Sur tous les cas de mortalité dus à des collisions avec des navires, 75 % ont eu lieu entre 1991 et 2007 (Brown et al., 2009), ce qui représente la moitié de toutes les morts connues de baleines noires dans cette période. Depuis 2002, 71 % de la mortalité de baleines noires adultes documentée a été causée par des collisions avec des navires et 29 % par l'enchevêtrement dans des engins de pêche – et environ un tiers de la mortalité documentée dans le cas des baleineaux a été attribuée à des facteurs anthropiques (Moore et al., 2004). Il est probable que ces valeurs sous-estiment la mortalité totale de cause humaine, parce que tous les cas ne sont pas documentés (particulièrement les cas d'enchevêtrement). Par exemple, il est plus probable que les carcasses des baleines gravement enchevêtrées vont couler, parce que leur couche de graisse est plus petite. La modélisation démographique semble indiquer que la mortalité (nombre de baleines noires mortes par année) a augmenté de 3 à 5 % entre 1980 et 1998, et que la majorité de l'augmentation touchait des femelles adultes (Caswell et al., 1999; Fujiwara et Caswell, 2001; Moore et al., 2007).
La mortalité de cause humaine semble être plus élevée chez les femelles que chez les mâles (Fujiwara et Caswell, 2001). Une explication possible est que les femelles adultes sont plus vulnérables à la mortalité de cause humaine, comme les collisions avec des navires (p. ex. figure 8), parce qu'elles tendent à passer plus de temps à la surface de la mer, lorsqu'elles allaitent et sont en compagnie de leur baleineau. Ce faisant, il est plus probable qu'elles croisent des navires (Baumgartner et Mate, 2003). Six des 8 baleines trouvées mortes en 2004 et en 2005 étaient des femelles adultes; 3 d'entre elles étaient sur le point de mettre bas et une autre était sexuellement mature (Kraus et al., 2005). Au moins 4 des baleines sont mortes à cause de collisions avec des navires ou d'enchevêtrement dans des engins de pêche (Kraus et al., 2005). En moyenne, les femelles produisent 5,25 baleineaux au cours de leur vie – ainsi, la perte de ces baleines équivalait à perdre 21 individus (Kraus et al., 2005).
Figure 8. Nombre cumulatif de baleines noires de l'Atlantique Nord mâles et femelles adultes présumées mortes (selon la règle de six ans) et dont la mort a été confirmée (Right Whale Consortium, 2011). Cette figure montre que, récemment, la mort d'un plus grand nombre de femelles adultes a été confirmée et, aussi, que le nombre de mâles et de femelles présumés morts est nettement supérieur au nombre de mâles et de femelles dont la mort est confirmée.
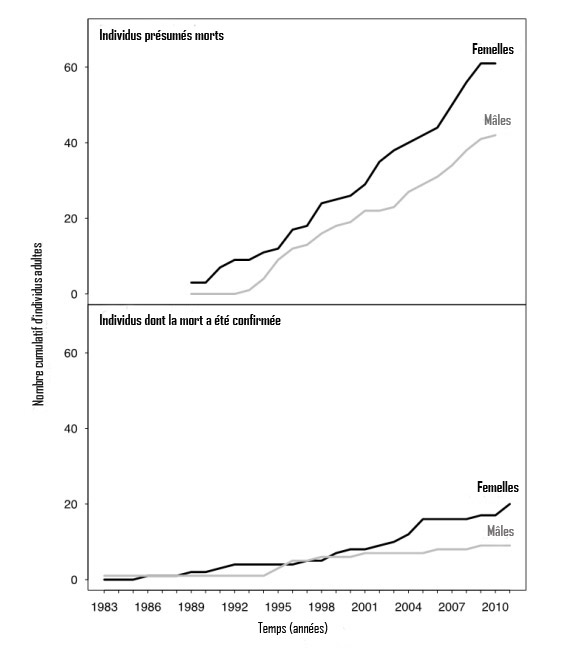
Description longue pour la figure 8
Deux graphiques montrant le nombre cumulatif de baleines noires de l'Atlantique Nord mâles et femelles adultes présumées mortes (graphique du haut) et dont la mort a été confirmée (graphique du bas).Les graphiques montrent que, récemment, la mort d'un plus grand nombre de femelles adultes a été confirmée et, aussi, que le nombre de mâles et de femelles présumés morts est nettement supérieur au nombre de mâles et de femelles dont la mort est confirmée.
Le risque de mortalité anthropique n'est pas distribué uniformément dans l'aire de répartition. Par exemple, on croit que le risque est moins élevé dans les zones qui ont été désignées comme habitat essentiel au Canada par le gouvernement fédéral ou qui ont été ainsi désignées aux États-Unis. Diverses mesures de conservation ont été mises en œuvre dans ces zones pour atténuer les risques de collision avec des navires, y compris des vitesses réduites dans les eaux états-uniennes et des routes maritimes décalées au Canada. De plus, des règlements ont été mis en œuvre aux États-Unis afin de réduire les cas d'enchevêtrement dans des engins de pêche. Les baleines qui se déplacent entre les zones spécialement désignées ou passent du temps à l'extérieur de celles-ci courent probablement un risque accru d'enchevêtrement ou de collision avec des navires. Une pétition pour modifier l'habitat essentiel existant aux États-Unis de façon à inclure l'« habitat migratoire » fait l'objet d'examen depuis 2010 (NMFS, 2010).
Vulnérabilité aux collisions avec des navires
Les collisions avec des navires sont la principale cause de mortalité documentée chez la baleine noire de l'Atlantique Nord (Reeves et al., 1978; Kraus et al., 2005; Moore et al., 2007) et ont probablement lieu plus fréquemment que les statistiques des nécropsies ne le suggèrent. Environ 7 % des baleines noires de l'Atlantique Nord connues individuellement ont des cicatrices ou des blessures attribuables à des collisions avec des navires. Néanmoins, il est difficile d'estimer les effets de ces blessures sur la morbidité, la productivité et la longévité (Brown et al., 2009).
La majorité des collisions mortelles et graves avec des navires, toutes les espèces de grandes baleines comprises, mettent en cause de grands navires de plus de 80 m de longueur (Laist et al., 2001). L'activité maritime a augmenté au cours des 50 dernières années, en grande partie à cause de l'expansion du commerce international (Waters et al., 2000; Hackett, 2003), et l'on prévoit qu'elle augmentera dans les ports de la côte atlantique des États-Unis d'environ 47 200 escales en 2000 à environ 93 500 escales en 2020 (Hackett, 2003; Ward-Geiger et al., 2005). Les baleines noires rencontrent souvent des navires à l'entrée de ports commerciaux et de bases militaires (Ward-Geiger et al., 2005). Les collisions mortelles avec des navires ont été signalées principalement dans les routes maritimes où près de celles-ci et dans les zones côtières où les baleines noires se regroupent (Knowlton et Kraus, 2001; Ward-Geiger et al., 2005).
Le comportement des baleines noires les rend particulièrement vulnérables aux collisions avec des navires. Plusieurs études de baleines noires munies d'enregistreurs acoustiques à capteurs multiples dans la baie de Fundy ont montré qu'elles ne réagissent pas aux navires ou aux bruits des navires qui s'approchent (Nowacek et al., 2004; Vanderlaan et Taggart, 2007). Cela semble indiquer que les baleines noires sont habituées au bruit des navires. Lors d'une étude expérimentale de signaux acoustiques conçus pour avertir les baleines et prévenir les collisions avec des navires, les baleines sont remontées à la surface là où le risque de collision était grandement accru (Nowacek et al., 2004). Le fait que les baleines se nourrissent à la surface augmente aussi les risques qu'elles soient heurtées par des navires. Durant le printemps, à la baie du cap Cod, les baleines noires passent jusqu'à 84 % de leur temps à se nourrir dans les quelques mètres supérieurs de la colonne d'eau (entre 0,5 et 2,5 m; Parks et al., 2011).
La vitesse des navires influe sur le risque qu'une collision se produise et sur la gravité des blessures subies (Kite-Powell et al., 2007; Vanderlaan et Taggart, 2007). Selon des modèles de régression logistique, presque toutes les collisions seraient mortelles lorsque la vitesse du navire dépasse 15 nœuds (Vanderlaan et Taggart, 2007). Les probabilités modélisées de blessures mortelles sont de 80 % à des vitesses de 15 nœuds et diminuent à 20 % à 8,6 nœuds. Pour réduire de 50 % les risques de blessures mortelles, il faut réduire les vitesses en deçà de 11,8 nœuds.
Gestion de l'industrie du transport maritime
Des mesures ont été prises par les gouvernements du Canada et des États-Unis afin de réduire la probabilité et la gravité des collisions avec des navires (Kraus et al., 2005, Vanderlaan et Taggart, 2009). Notamment, ils ont désigné l'habitat essentiel de la baleine noire, rendu obligatoire le signalement de la position du navire (mesure adoptée par l'Organisation maritime internationale [OMI]; Silber et al., 2002), mis en œuvre des modifications obligatoires aux itinéraires des navires avec des dispositifs de séparation du trafic de l'OMI (OMI, 2003; 2006), appliqué des limites obligatoires de vitesse des navires (NMFS, 2008) et recommandé des « zones à éviter » (ZAE) (OMI, 2007; 2008). La mise en application de la zone à éviter dans le bassin Roseway constitue probablement la mesure la plus importante prise par le Canada pour réduire le risque de collisions mortelles avec des navires pour les baleines noires (Vanderlaan et Taggart, 2009). La zone à éviter volontairement est une aire d'alimentation de forte densité, et la recommandation est en vigueur du 1er juin au 31 décembre (OMI, 2007; Vanderlaan et Taggart, 2009). Même s'il s'agit seulement d'une recommandation et qu'elle n'est donc pas obligatoire, les navigateurs la respectent dans une proportion variant de 57 % à 87 % (Vanderlaan et Taggart, 2009). On estime que ce taux élevé de conformité a réduit le risque de collisions mortelles pour les baleines noires dans le bassin Roseway d'environ 82 %(Vanderlaan et Taggart, 2009).
Les routes maritimes ont également été modifiées pour atténuer le risque de collisions avec des navires. En 2003, les 112 km de routes maritimes allant de l'entrée de la baie de Fundy jusqu'au port de Saint John (Nouveau-Brunswick) ont été décalés afin de réduire le chevauchement entre les routes maritimes et les zones d'alimentation à forte densité de baleines noires pendant l'été et l'automne (Knowlton et Brown, 2007). À cause de cette mesure, le temps de traversée des navires a été prolongé, mais la probabilité de collision entre les navires et les baleines noires a été réduite (Knowlton et Brown, 2007).
Aux États-Unis, les routes maritimes qui desservent Boston ont été décalées de 12° et leur largeur a été diminuée de 2,8 km (1,5 NM) (Knowlton et Brown, 2007) pour éviter les attroupements de rorquals à bosse et de baleines noires. Une zone à éviter saisonnière (recommandation) a été créée en 2009 pour les navires (jauge brute de 300 tonneaux ou plus) dans le Grand chenal Sud, pendant les périodes où les baleines noires y sont le plus abondantes (NMFS, 2011). De plus, des routes maritimes recommandées ont été établies en 2006 pour la Floride, la Georgie et le Massachusetts (NMFS, 2006), et des limites de vitesse obligatoires de 10 nœuds (pour les navires de 65 pieds ou plus) ont été établies dans les zones de gestion saisonnières (Seasonal Management Areas), telles les aires de mise bas (sud-est des États-Unis), la voie migratoire (entre la Floride et le Massachusetts) et les aires d'alimentation dans le golfe du Maine (la baie du cap Cod, au large de Race Point, et le Grand chenal Sud; Knowlton et Brown, 2007; NMFS, 2011). Le NOAA Fisheries Service désigne aussi des zones de gestion dynamique (Dynamic Management Areas) en fonction des données d'observation de baleines noires, en annonçant des zones de limite de vitesse à respecter sur une base volontaire pour réduire le nombre de collisions dans les zones où les baleines noires sont présentes.
Malgré les mesures de conservation qui ont été mises en œuvre pour réduire le risque de mortalité et de blessures graves dues aux collisions avec des navires, dix individus portant des nouvelles blessures graves dues à des collisions ont été aperçus en 2009-2010 (Hamilton et al., 2010; Hamilton et Knowlton, 2011). De plus, un traversier rapide reliant Yarmouth (Nouvelle-Écosse) à Portland (Maine) devrait être remis en service après une interruption de 5 ans, ce qui augmentera le risque de collisions pour les baleines noires.
Vulnérabilité à l'enchevêtrement dans les engins de pêche
Les baleines noires de l'Atlantique Nord sont vulnérables à l'enchevêtrement dans les engins de pêche, parce qu'elles fréquentent des zones de pêche intense, où des engins à haut risque sont déployés (p. ex. des casiers avec des filins à bouée dans la colonne d'eau et des filets avec des lignes-mères ancrées) (Johnson et al., 2007). Plus de 83 % des individus photo-identifiés portent des marques causées par l'enchevêtrement (Johnson et al., 2007; Knowlton et al., 2012). Lorsque la tête d'une baleine se trouve enchevêtrée, cela peut perturber son alimentation et même la faire mourir de faim. Ce genre d'enchevêtrement est plus commun dans le cas des baleines noires que dans celui d'autres espèces de baleines à fanons (Johnson et al., 2005). Qui plus est, lorsque la tête est enchevêtrée, il est particulièrement difficile d'utiliser des techniques de démêlement, parce que les sauveteurs courent un plus grand risque d'être frappés par la queue de la baleine qui bat l'eau pendant qu'ils essaient de la libérer (tandis que, lorsque c'est la queue qui est enchevêtrée, les sauveteurs peuvent se placer à une bonne distance derrière la queue de l'animal).
Même si aucun changement des engins de pêche n'a été rendu obligatoire au Canada, les risques d'enchevêtrement pourraient être plus faibles dans les eaux canadiennes, comme la baie de Fundy, que dans les eaux états-uniennes, en raison de la manière selon laquelle il est recommandé d'installer les lignes-mères dans la pêche canadienne du homard (Homarus americanus) au casier (Brillant et Trippel, 2010). Sur 61 incidents d'enchevêtrement analysés (30 impliquant des rorquals à bosse et 31, des baleines noires), les filins à bouée et les lignes-mères étaient les types les plus communs d'engins de pêche (81 %)impliqués (Johnson et al., 2005). En eaux états-uniennes, les cordages sont habituellement installés 3 m au-dessus du plancher océanique, où ils présentent le risque le plus important d'enchevêtrement pour les baleines (Brillant et Trippel, 2010). Dans la baie de Fundy, cependant, les cordages sont posés près du fond, mais on continue quand même à y signaler des enchevêtrements (Johnson et al., 2007). On sait que les baleines noires se nourrissent sur le plancher océanique, p. ex. à des profondeurs d'environ 200 m, dans la baie de Fundy (Baumgartner et al., 2007; p. 155), et elles sont souvent aperçues avec de la boue sur la tête (Kraus et Rolland, 2007, illustration en couleur 8). Les relevés de baleines noires et les données sur le déploiement des engins de pêche au Canada semblent indiquer que c'est la pêche du homard qui représente la plus grande menace pour les baleines noires, au printemps et à l'automne, lors de la migration vers les zones canadiennes désignées comme faisant partie de l'habitat essentiel, ou à partir de celles-ci (bassin de Grand Manan et bassin Roseway;Vanderlaan et al., 2011)
La pêche au homard n'est pas le seul type de pêche au Canada qui pose un risque d'enchevêtrement pour les baleines noires. Le plus grand risque d'enchevêtrement est associé à la pêche à la ligne des poissons de fond durant l'été, lorsque les baleines noires se trouvent à l'intérieur de zones de l'habitat essentiel pendant plusieurs mois (Vanderlaan et al., 2011). La menace relative d'enchevêtrement pour les baleines noires durant l'été (de juillet à octobre) est de 42 % dans le cas de la pêche à la ligne des poissons de fond et de seulement 9,6 % dans le cas de la pêche extracôtière du homard au casier (Vanderlaan et al., 2011). Compte tenu des données sur les cicatrices dues à l'enchevêtrement recueillies de 1993 à 2004, 87 ± 29 incidents d'enchevêtrement laissant des cicatrices se produisent annuellement, et il existe environ 1 % de chances, chaque année, qu'un enchevêtrement mortel se produise dans l'habitat essentiel de la baleine noire désigné au Canada (Vanderlaan et al., 2011).
Aucune mesure de gestion n'a été prise au Canada pour réduire les risques d'enchevêtrement de baleines noires dans les engins de pêche. Cependant, le Fonds mondial pour la nature, par exemple, a élaboré un programme volontaire avec les pêcheurs afin de réduire le nombre de lignes de pêche dans l'eau. De plus, le MPO et la Grand Manan Fisherman's Association ont élaboré un plan d'atténuation pour réduire les interactions avec les engins de pêche au homard.
Gestion des activités de pêche dans les eaux états-uniennes
Aux États-Unis, l'Atlantic Large Whale Take Reduction Team (ALWTRT), mise en place par le NMFS en 1996, a élaboré le plan ALWTRP (Atlantic Large Whale Take Reduction Plan) en 1997 pour réduire le taux de blessures graves et de mortalité dû à l'enchevêtrement dans les engins de pêche au casier et au filet maillant (Johnson et al., 2007). Le plan ALWTRP a été publié dans le Federal Register des États-Unis en tant que décision finale provisoire, et les règlements ont été mis à jour en 1997 et en 2000. Trois décisions importantes ont été publiées par le NMFS en 2002, qui ont conduit à la modification additionnelle des engins de pêche commerciale. Parmi ces décisions, il y avait la mise au point d'une méthode pour limiter temporairement les activités de pêche dans les endroits où des regroupements imprévus de baleines noires sont observés (gestion de zones dynamiques ou Dynamic Area Management) et l'instauration de zones restreintes de façon permanente en fonction des regroupements annuels prévisibles des baleines noires (gestion de zones saisonnières ou Seasonal Area Management). Les mesures temporaires de gestion de zones saisonnières (p. ex. dans la baie du cap Cod) limitent les engins pouvant être utilisés pendant les périodes d'abondance maximale des baleines noires. De plus, une décision finale publiée par le NMFS en 2007 a agrandi la zone restreinte au sud-est des États-Unis et y a interdit la pêche au filet maillant durant la saison de mise bas des baleines noires (NMFS, 2007).
Malgré les récentes mesures de conservation, l'enchevêtrement dans des engins de pêche continue à se produire. En 2009 et en 2010, 12 cas d'enchevêtrement de baleines noires ont été répertoriés (Hamilton et al., 2010; Hamilton et Knowlton, 2011) et au moins 3 des baleines en sont mortes. De plus, des blessures graves pour 3 baleines ayant été enchevêtrées précédemment ont été signalées en 2009 (Hamilton et al., 2010; Hamilton et Knowlton, 2011).
Autres facteurs limitatifs potentiels
Par ailleurs, la croissance démographique pourrait également être limitée par les parasites, la maladie, les contaminants, les activités industrielles, les effets génétiques et démographiques liés à une population de petite taille ainsi que par le stress nutritionnel.
Des échantillons fécaux, recueillis principalement dans la baie Fundy, ont révélé plusieurs aspects concernant la santé des baleines noires relatifs aux parasites et aux microbes, aux métabolites hormonaux (p. ex. les hormones de stress de reproduction), aux contaminants de l'environnement et aux protéines immunes des viscères (Rolland et al., 2007). Il est tout particulièrement intéressant de noter que les matières fécales contiennent deux parasites, Giardia et Cryptosporidium, à des niveaux nettement plus élevés que ceux observés chez d'autres mammifères marins (Rolland et al., 2007). Ces parasites peuvent causer des maladies gastrointestinales chez les humains et les animaux terrestres, mais leur impact sur la santé des baleines noires demeure inconnu (Rolland et al., 2007). L'origine de ces organismes n'est pas claire : a) ils ont peut-être été introduits par des sources ponctuelles, comme les décharges d'eaux usées; b) ils sont peut-être propres à certaines espèces particulières de mammifères marins; et/ou c) ils font peut-être l'objet d'un cycle naturel à l'intérieur du milieu marin.
Des biopsies de la couche de graisse de baleines noires ont révélé que cette couche contient de faibles concentrations de perturbateurs endocriniens (p. ex. du dichlorodiphényltrichloroéthane [DDT] et des biphényles polychlorés [BPC]), dont on sait qu'ils contribuent au dysfonctionnement de la reproduction chez certains autres mammifères (Woodley et al., 1991; Weisbrod et al., 2000; Kraus et al., 2007). Ces contaminants liposolubles pourraient avoir des effets défavorables particulièrement sur les baleineaux qui dépendent des réserves de graisse de leur mère pendant l'allaitement (Kraus et al., 2007). Des concentrations même faibles de ces produits chimiques persistants peuvent nuire au développement initial des organes reproducteurs et des systèmes endocrinien, immunitaire et neurologique (Colborn et al., 1993; Rolland et al., 1995; Kraus et al., 2007). Cependant, les effets réels, s'il y en a, de l'exposition aux contaminants sur les baleines noires de l'Atlantique Nord demeurent inconnus.
Un faible niveau de pollution, la circulation maritime, les rejets en mer et le dragage ont tous été mentionnés dans les plans de rétablissement en tant que facteurs qui dégradent l'habitat de la baleine noire. Toutefois, aucune étude n'a, à ce jour, déterminé comment ou dans quelle mesure ces facteurs nuisent à l'habitat.
Le bruit sous-marin soulève aussi des inquiétudes. Le bruit de fond s'est intensifié dans la baie de Fundy, en raison de l'augmentation de la circulation maritime (Parks et al., 2011), et a probablement provoqué des changements dans la façon de vocaliser des baleines noires (Parks et al., 2007b; Parks et al., 2011). Le bruit ambiant ne cesse généralement d'augmenter dans les océans et a presque certainement compliqué la communication entre les baleines à fanons (Clark et al., 2009). Les effets du masquage des vocalisations (c. à d. la perte d'espace de communication) à cause du bruit des navires seraient plus importants pour les baleines noires de l'Atlantique Nord que pour les autres espèces de baleines à fanons (p. ex. les rorquals communs et les rorquals à bosse), parce que leurs vocalisations ne sont pas aussi fortes (Clark et al., 2009). À cause du milieu plus bruyant où elles se retrouvent, les baleines noires ont tendance à augmenter l'amplitude de leurs vocalisations pour pouvoir continuer à communiquer (Parks et al., 2011).
Le développement de l'énergie éolienne et de l'énergie marémotrice pourrait avoir une incidence sur les baleines noires. Des projets de ce genre ont été proposés dans les détroits de Nantucket et de Rhode Island, où des baleines noires ont été observées durant le printemps de 2010 (Kenney, 2010; Leeney et al., 2010), et à plusieurs autres endroits le long de la côte est des États-Unis, aux abords desquels les baleines noires peuvent passer au cours de leur migration. Les travaux de construction associés à des projets d'énergie renouvelable pourraient causer des perturbations physiques et acoustiques (y compris les levés sismiques pour les évaluations de sites) et augmenter les risques de collisions avec des navires (Madsen et al., 2006; Leeney et al., 2010). Des inquiétudes sont aussi soulevées en ce qui concerne l'exploration des ressources pétrolières et gazières extracôtières qui serait sur le point de commencer au large de la côte est des États Unis, augmentant les risques pesant sur ces baleines.
La croissance démographique pourrait également être limitée par la qualité et la quantité de proies disponibles. Des individus dont l'état corporel a été jugé médiocre ont été observés dans la période 1991 1999, pendant laquelle le taux de reproduction de la population de baleines noires a été plus bas que dans le passé (Pettis et al., 2004). Des changements de l'épaisseur de la couche de graisse, observés dans le milieu des années 1990, ont été corrélés avec des changements des intervalles entre les mises bas et des processus atmosphère océan de grande échelle, qui contrôlent la qualité et la quantité de proies disponibles (Miller et al., 2011). Des liens semblables ont été faits entre les températures de la surface de la mer et le succès annuel des mises bas pour les baleines franches australes (Leaper et al., 2006). On prévoit que l'oscillation nord atlantique (NAO) fluctuera plus fréquemment à l'avenir (Greene et Pershing, 2004), ce qui pourrait rendre les conditions d'alimentation plus variables pour les baleines noires de l'Atlantique Nord.
Parmi les facteurs, autres que la disponibilité des proies, qui ont probablement aussi réduit la performance en matière de reproduction durant la période 1989 2003, on retrouve les faibles niveaux de variabilité génétique et la consanguinité, ainsi que l'exposition aux polluants et aux toxines (Kraus et al., 2007). Néanmoins, il n'a pas été établi si les taux de reproduction sont restés inférieurs au niveau potentiel depuis 2003. Malgré cela, le nombre annuel de baleineaux nés durant les années 2000 a été supérieur, en moyenne, au nombre de baleineaux nés durant les années 1990 (figure 5).
Protection, statuts et classements
Statuts et protection juridiques
La baleine noire est protégée à l'échelle internationale de la pêche commerciale aux termes de la Convention internationale pour la réglementation de la chasse à la baleine et du commerce aux termes de la Convention sur le commerce international des espèces de faune et de flore sauvages menacées d'extinction (CITES). Aux États-Unis, les baleines noires sont protégées en vertu de lois rigoureuses, notamment la Endangered Species Act (ESA) et la Marine Mammal Protection Act (MMPA). Le National Marine Fisheries Service (NMFS) est responsable de la conservation des baleines noires à l'échelle nationale et a publié un plan de rétablissement en 1991 pour les baleines noires de l'Atlantique Nord et du Pacifique Nord ainsi qu'un plan révisé en 2005 visant seulement les baleines noires de l'Atlantique Nord (NMFS, 1991; 2005). Ce plan identifie les menaces et décrit les mesures requises pour diminuer les risques d'extinction et augmenter les perspectives de rétablissement des populations (NMFS, 1991; Brown et al., 2009). Le NMFS publie des rapports annuels d'évaluation des stocks pour les mammifères marins et établit un niveau de « prélèvement biologique potentiel » pour chacun d'entre eux (récemment 0,8 individus par année; Waring et al., 2011).
Au Canada, les baleines noires sont protégées en vertu du Règlement sur les mammifères marins relevant de la Loi sur les pêches, et Pêches et Océans Canada (MPO) est l'organisme responsable de la conservation des baleines noires (p. ex. de réglementer les activités de pêche, de produire des directives pour l'observation des baleines dans la baie de Fundy). La baleine noire de l'Atlantique Nord a été évaluée comme étant « en voie de disparition » par le COSEPAC en 2003 et a été inscrite comme telle dans la LEP en 2005 (décret modifiant les annexes 1 à 3 de la Loi sur les espèces en péril, 2005). L'espèce a été réévaluée comme étant en voie de disparition par le COSEPAC en novembre 2013.
Le MPO et le Fonds mondial pour la nature Canada ont publié conjointement le Plan canadien de rétablissement de la baleine noire de l'Atlantique Nord en 2000 (Anonyme, 2000), et un programme de rétablissement canadien a ensuite été publié, en 2009, conformément aux lignes directrices de la LEP (Brown et al., 2009). Le programme de rétablissement justifie la désignation du bassin de Grand Manan dans la baie de Fundy et du bassin Roseway dans la portion sud-ouest de la plate-forme Néo-Écossaise à titre d'habitat essentiel aux termes de la LEP (Brown et al., 2009). L'objectif de rétablissement provisoire est d'atteindre « une tendance à la hausse de l'abondance sur trois générations » en réduisant la mortalité anthropique, les blessures (collisions avec des navires et enchevêtrement dans des engins de pêche) et les perturbations (présence de navires et exposition aux contaminants) et en améliorant les connaissances sur les caractéristiques du cycle de vie, les taux de reproduction bas, l'habitat et les menaces pouvant nuire au rétablissement par le biais de recherches, de collaboration et d'élaboration d'activités d'éducation et d'intendance (Brown et al., 2009).
Statuts et classements non juridiques
L'Union internationale pour la conservation de la nature (UICN) estime que la baleine noire de l'Atlantique Nord est menacée d'extinction et l'a inscrite sur sa liste rouge (Reilly et al., 2012). Selon NatureServe (2001), sa cote mondiale est G1, et sa cote mondiale arrondie (au moyen de l'algorithme d'évaluation de la cote de l'espèce) est G1 Gravement en péril (dernière évaluation en 2006), ce qui signifie que l'espèce fait face à un risque très élevé d'extinction à cause de ses effectifs extrêmement bas, qui n'ont pas augmenté de façon significative, et ce, malgré les mesures de protection prises (tableau 2). Au niveau national, on a attribué la cote N1 à l'espèce, tant aux États-Unis (dernière évaluation en 1997) qu'au Canada (dernière évaluation en 2011), ce qui signifie que l'espèce est gravement en péril.
| Pays | État/Province/Territoire | Cote | Définition |
|---|---|---|---|
| Canada | Labrador | SNR | Espèce non classée |
| Canada | Nouveau-Brunswick | S1 | Gravement en péril |
| Canada | Île de Terre-Neuve | SH | Peut-être disparue |
| Canada | Nouvelle-Écosse | S1 | Gravement en péril |
| Canada | Île-du-Prince-Édouard | SH | Peut-être disparue |
| Canada | Québec | S1 | Gravement en péril |
| United States | Delaware | SXB | Repr. peut-être plus possible |
| États-Unis | Floride | S1 | Gravement en péril |
| États-Unis | Georgie | S1 | Gravement en péril |
| États-Unis | Maine | SNR | Espèce non classée |
| États-Unis | Maryland | SNA | Espèce non classée |
| États-Unis | Massachusetts | S1 | Gravement en péril |
| États-Unis | New Jersey | S1 | Gravement en péril |
| États-Unis | État de New York | SNA | Espèce non classée |
| États-Unis | Caroline du Nord | SNA | Espèce non classée |
| États-Unis | Rhode Island | SU | Non classable |
| États-Unis | Texas | S1 | Gravement en péril |
Protection et propriété de l'habitat
Les milieux importants constituant l'habitat pour l'alimentation et la mise bas des baleines noires de l'Atlantique Nord ont été protégés dans les eaux canadiennes et états-uniennes. Au Canada, le MPO a désigné 2 zones d'habitat essentiel – le bassin de Grand Manan dans l'embouchure de la baie de Fundy et le bassin Roseway dans la portion sud-ouest de la plate-forme Néo-Écossaise (Brown et al., 2009) (figure 3). Aux États-Unis, le NMFS a désigné 3 zones d'habitat essentiel : la baie du cap Cod, le Grand chenal Sud et les eaux côtières du sud-est des États-Unis, de Brunswick (Georgie) à Jacksonville (Floride), jusqu'à 15 milles marins au large, et de Jacksonville à Sebastian Inlet (Floride), jusqu'à 5 millesmarins de la côte (NMFS, 1994) (figure 3). La mise en place d'une protection spéciale de l'habitat migratoire fait l'objet d'examen aux États-Unis (NMFS, 2010).
Remerciements et experts contactés
La majeure partie des connaissances sur les baleines noires qui fréquentent les eaux canadiennes provient de chercheurs affiliés au New England Aquarium et de nombreuses organisations qui contribuent à la base de données des observations de la baleine noire (Right Whale Sightings Database) et au catalogue d'identification photographique (PhotoID Catalog) (notamment, la Blue Ocean Society, le Florida Fish and Wildlife Research Institute, le Georgia Department of Natural Resources, le New England Aquarium, le Northeast Fisheries Science Center, le Provincetown Center for Coastal Studies, la University of North Carolina Wilmington, le Whale Center of New England, le Wildlife Trust, la Woods Hole Oceanographic Institution, la Cornell University, l'Oregon State University, le NOAA Fisheries, le ministère des Pêches et des Océans et la Dalhousie University).
Le North Atlantic Right Whale Consortium a permis l'accès aux données d'observations et d'identification des baleines noires, qui sont conservées à l'University of Rhode Island (R. Kenney) et au New England Aquarium (P. Hamilton). P. Hamilton a fourni des données supplémentaires expressément pour ce rapport. Les données relatives aux observations et à l'identification des baleines noires en eaux canadiennes ont été fournies au Consortium par les organismes et organisations suivants : Associated Scientists of Woods Hole, Canadian Whale Institute, Cornell Laboratory of Ornithology, Dalhousie University Department of Oceanography, East Coast Ecosystems Research Organization, Pêches et Océans Canada/Institut océanographique de Bedford, Pêches et Océans Canada/Division des poissons de mer à la Station biologique de St. Andrews, Pêches et Océans Canada/Bureau des espèces en péril de la région des Maritimes, Pêches et Océans Canada/Gestion des ressources, Centre de recherche sur la vie marine de Grand Manan, Groupe de recherche et d'éducation sur les mammifères marins, Fonds international pour la protection des animaux, Station de recherche des îles Mingan (MICS pour Mingan Island Cetacean Study), Musée du Nouveau-Brunswick, Right Whale Project du New England Aquarium/John H. Prescott, Marine Laboratory, NOAA Fisheries/Northeast Regional Office and Northeast Fisheries Science Center, musée d'histoire naturelle de la Nouvelle-Écosse, Oregon State University, Provincetown Center for Coastal Studies, Trent University/Natural Resources DNA Profiling and Forensic Centre, University of Guelph, University of Prince Edward Island/Atlantic Veterinary College, University of Rhode Island, Fonds mondial pour la nature Canada, Woods Hole Oceanographic Institution.
R. Kenney et J. Wu ont préparé les cartes de répartition, et les autorités fédérales, provinciales et territoriales ont fourni des renseignements supplémentaires. Les examens scientifiques ont été fournis par R. Reeves, J. Watson, T. Frasier, M. Brown, S. Kraus et R. Kenney.
Experts contactés
COSEPAC
- Monique Goit, Secrétariat du COSEPAC, Gatineau (Québec)
- Karen Timm, Secrétariat du COSEPAC, Gatineau (Québec)
- Jenny Wu, spécialiste de la gestion de données et de la cartographie, Secrétariat du COSEPAC, Gatineau (Québec)
- Neil Jones, agent de projet scientifique et coordonnateur des CTA, Secrétariat du COSEPAC, Gatineau (Québec)
Gouvernement fédéral
- Diane Amirault-Langlais, Service canadien de la faune, Région de l'Atlantique
- Robert Anderson (Ph.D.), chercheur scientifique, Musée canadien de la nature, Partenariat fédéral en matière d'information sur la biodiversité, Ottawa (Ontario)
- Briar Howes (Ph.D.), Soutien scientifique, Programme des espèces en péril, Parcs Canada, Gatineau (Québec)
- Patrick Nantel (Ph.D.), biologiste de la conservation, Programme des espèces en péril, Direction de l'intégrité écologique, Parcs Canada, Gatineau (Québec)
Gouvernements provinciaux et territoriaux
- David Fraser, Scientific Authority Assessment A/Manager, SEARS-Species & Ecosystems at Risk Section, Ministry of Environment de la Colombie-Britannique, Victoria (Colombie-Britannique)
- Pascal Giasson, gestionnaire, Programme des espèces en péril, Direction de la pêche sportive et de la chasse, ministère des Ressources naturelles, Fredericton (Nouveau-Brunswick)
- Shelley Moores, Senior Manager, Endangered Species and Biodiversity Wildlife Division, ministère de l'Environnement et de la Conservation, Corner Brook (Terre-Neuve-et-Labrador).
- Susan E. Squires (Ph.D.), Ecosystem Management Ecologist, Biodiversity and Endangered Species, Wildlife Division, ministère de l'Environnement et de la Conservation, Corner Brook (Terre-Neuve-et-Labrador)
- J. Sherman Boates (Ph.D.), gestionnaire, Biodiversity, Wildlife Division, ministère des Ressources naturelles, Kentville (Nouvelle-Écosse)
- Mark. F. Elderkin, biologiste provincial (espèces en péril), Wildlife Division, ministère des Ressources naturelles, Kentville (Nouvelle-Écosse)
- Annie Lévesque, biologiste en conservation, Ministère des Ressources naturelles, Direction de l'expertise sur la faune et ses habitats, Québec
Ministère des Pêches et des Océans
- David Millar, président, Canadian Recovery Network for the North Atlantic Right Whale, Nouvelle-Écosse
Centres de données sur la conservation
- S. Blaney, Centre de données sur la conservation du Canada atlantique, Sackville (Nouveau-Brunswick)
- Annie Paquet, Centre de données sur le patrimoine naturel du Québec, Ministère des Ressources naturelles et de la Faune du Québec, Québec
- Stefen Gerriets, Centre de données sur la conservation du Canada atlantique
Conseils de gestion de la faune
- Marck O'Connor, directeur de la gestion de la faune, Conseil de gestion des ressources fauniques de la région marine du Nunavik, Inukjuak (Québec)
- Adamie Padlayat, directeur exécutif, Conseil de gestion des ressources fauniques de la région marine du Nunavik, Inukjuak (Québec)
Sources d'information
Aguilar, A. 1986. A review of old Basque whaling and its effects on the right whales(Eubalaena glacialis) of the North Atlantic, Report of the International Whaling Commission (Special Issue)10:191-200.
Anonyme. 2000. Plan canadien de rétablissement de la baleine noire de l'Atlantique Nord, préparé par l'Équipe de retablissement de la baleine noire pour le Fonds mondial pour la nature Canada et le ministère des Pêches et des Océans,106 p.
Atkinson, S. 2002. Male reproductive systems, p. 700-704, in W.F. Perrin, B. Wursig et J.G.M. Thewissen (éd.), Encyclopedia of marine mammals, Academic Press, San Diego (Californie), États-Unis
Barkham, S. 1978. The Basques: filling a gap in our history between Jacques Cartier and Champlain, Canadian Geographical Journal96:8-19.
Baumgartner, M.F., T.V.N. Cole, R.G. Campbell, G.J. Teegarden et E.G. Durbin. 2003. Associations between North Atlantic right whales and their prey, Calanus finmarchicus, over diel and tidal time scales, Marine Ecology Progress Series264:155-166.
Baumgartner, M.F., et B.R. Mate. 2003. Summertime foraging ecology of North Atlantic right whales, Marine Ecology Progress Series264:123-135.
Baumgartner, M.F., et B.R. Mate. 2005. Summer and fall habitat of North Atlantic right whales (Eubalaena glacialis) inferred from satellite telemetry, Canadian Journal of Fisheries and Aquatic Sciences 62:527-543.
Baumgartner, M.F., N.S.J. Lysiak, C. Schuman, J. Urban-Rich et F.W. Wenzel. 2011. Diel vertical migration behavior of Calanus finmarchicus and its influence on right and sei whale occurrence, Marine Ecology Progress Series 423:167-184.
Best, P.B. 1994. Seasonality of reproduction and the length of gestation in southern right whales Eubalaena australis, Journal of Zoology 232:175-189.
Best, P.B., A. Brandão et D.S. Butterworth. 2001. Demographic parameters of southern right whales off South Africa, Journal of Cetacean Research and Management (Special Issue) 2:161-169.
Braham, H.W., et D.W. Rice. 1984. The right whale, Balaena glacialis, Marine Fisheries Review 46:38-44.
Brillant, S.W., et E.A. Trippel. 2010. Elevations of lobster fishery groundlines in relation to their potential to entangle endangered North Atlantic right whales in the Bay of Fundy, Canada, ICES Journal of Marine Science67:355-364.
Brown, M.W., J.M. Allen et S.D. Kraus. 1995. The designation of seasonal right whale conservation zones in the waters of Atlantic Canada, p. 90-98, in N. Shackell et M. Willison (éd.), Marine protected areas and sustainable fisheries, Science and Management of Protected Areas Association, Wolfville(Nouvelle-Écosse) Canada.
Brown, M.W., S. Brault, P.K. Hamilton, R.D. Kenney, A.R. Knowlton, M.K. Marx, C.A. Mayo, C.K. Clay et S.D. Kraus. 2001. Sighting heterogeneity of right whales in the western North Atlantic, 1980-1992, Journal of Cetacean Resource Management (Special Issue)2:245-250.
Brown, M.W., S.D. Kraus, C.K. Slay et L.P. Garrison. 2007. Surveying for discovery, science and management, p. 105-137, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Brown, M.W., D. Fenton, K. Smedbol, C. Merriman, K. Robichaud-LeBlanc et J. Conway. 2009. Programme de rétablissement de la baleine noire (Eubalaena glacialis) de l'Atlantique Nord dans les eaux canadiennes de l'Atlantique, Série de Programmes de rétablissement de la Loi sur les espèces en péril, Ottawa, Canada, vi + 72 p.
Brown, S.G. 1986. Twentieth-century records of right whales (Eubalaena glacialis) in the northeast Atlantic Ocean, Report of the International Whaling Commission (Special Issue)10:121-127.
Brownell, R.L.J., et K. Ralls. 1986. Potential sperm competition in baleen whales, Report of the International Whaling Commission (Special Issue)8:97-112.
Browning, C.L., R.M. Rolland et S.D. Kraus. 2010. Estimated calf and perinatal mortality in western North Atlantic right whales (Eubalaena glacialis), Marine Mammal Science 26:648-662.
Cassoff, R.M., K.E. Moore, M.A. William, S.G. Barco, D.S. Rotstein et M.J. Moore. 2011. Lethal engtanglement in baleen whales (Supplement), Diseases of Aquatic Organisms 96:175-185.
Caswell, H., M. Fujiwara et S. Brault. 1999. Declining survival probability threatens the North Atlantic right whale, Proceedings of the National Academy of the Sciences, 96:3308-3313, États-Unis.
Checkley Jr, D.M. 1982. Selective feeding by Atlantic herring (Clupea harengus) larvae on zooplankton in natural assemblages, Marine Ecology Progress Series 9:245-253.
Clark, C.W., W.T. Ellison, B.L. Southall, L. Hatch, S.M. Van Parijs, A. Frankel et D. Ponirakis. 2009. Acoustic masking in marine ecosystems: intuitions, analysis, and implication, Marine Ecology Progress Series395:201-222.
Colborn, T., F. vom Saal et A. Soto. 1993. Developmental effects of endocrine disrupting chemicals in wildlife and humans, Environmental Health Perspectives 101:378-384.
Cole, T.V.N., P. Hamilton, A.G. Henry, P. Duley, R.M. Pace III, B.N. White et T. Frasier. 2013. Evidence of a North Atlantic right whale Eubalaena glacialis mating ground, Endangered Species Research 21:55-64.
Conversi, A., S. Piontkovski et S. Hameed. 2001. Seasonal and interannual dynamics of Calanus finmarchicus in the Gulf of Maine (northeastern US shelf) with reference to the North Atlantic Oscillation, Deep-Sea Research II 48:519-530.
Cooke, J.G., et V.J. Rowntree. 2001. Estimates of demographic parameters for southern right whales (Eubalaena australis) observed off Peninsula Valdes, Argentina, Journal of Cetacean Research and Management (Special Issue) 2:125-132.
COSEPAC. 2003. Évaluation et Rapport de situation du COSEPAC sur la baleine noirede l'Atlantique Nord (Eub alaena glacialis) au Canada – Mise à jour. Comité sur la situation des espèces en péril au Canada. Ottawa. viii + 30 p.
Crone, M.J., et S.D. Kraus. 1990. Right whales (Eubalaena glacialis), in the Western North Atlantic: A catalog of indentified individuals, New England Aquarium, Boston (Massachusetts) États-Unis, 223 p.
Cumbaa, S.L. 1986. Archaeological evidence of the 16th century Basque right whale fishery in Labrador, Report of the International Whaling Commission (Special Issue) 10:198-190.
de la Chenelière, V., comm. pers. 2010. Communication orale entre Diane Gagnon et Veronique de la Chenelière, février 2010, biologiste, Groupe de recherche et d'éducation sur les mammifères marins, Tadoussac (Québec), Canada.
DeLorenzo Costa, A., E.G. Durbin, C.A. Mayo et E.G. Lyman. 2006. Environmental factors affecting zooplankton in Cape Cod Bay: implications for right whale dynamics, Marine Ecology Progress Series 323:281-298.
Firestone, J., S.B. Lyons, C. Wang et J.J. Corbett. 2008. Statistical modeling of North Atlantic right whale migration along the mid-Atlantic region of the eastern seaboard of the United States, Biological Conservation 141:221-232.
Ford, J.K.B., et R.R. Reeves. 2008. Fight or flight: antipredator strategies of baleen whales, Mammal Review 38:50-86.
Fortune, S.M., A.W. Trites, W.L. Perryman, M.J. Moore, H.M. Pettis et M.S. Lynn. 2012. Growth and rapid early development of North Atlantic right whales (Eubalaena glacialis), Journal of Mammalogy 93:1342-1354.
Fortune, S.M., A.W. Trites, C.A. Mayo, D.A.S. Rosen et P.K. Hamilton. 2013. Energetic requirements of North Atlantic right whales and the implications for species recovery, Marine Ecology Progress Series478:253-272
Frasier, T.R., P.K. Hamilton, M.W. Brown, L.A. Conger, A.R. Knowlton, M.K. Marx, C.K. Slay, S.D. Kraus et B.N. White. 2007a. Patterns of male reproductive success in a highly promiscuous whale species: the endangered North Atlantic right whale, Molecular Ecology 16:5277-5293.
Frasier, T.R., B.A. McLeod, R.M. Gillett, M.W. Brown et B.N. White. 2007b. Right whales past and present as revealed by their genes, p. 200-231, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Fujiwara, M., et H. Caswell. 2001. Demography of the endangered North Atlantic right whale, Nature 414:537-541.
Ginsberg, J.R., et U.W. Huck. 1989. Sperm competition in mammals, Trends in Ecology and Evolution 5:74-79.
Gomendio, M., A.H. Harcourt et E.R.S. Rolan. 1998. Sperm competition in mammals, inT.R. Birkhead et A.P. Moller (éd.), Sperm competition and sexual selection, Academic Press, San Diego (Californie), États-Unis.
Goodyear, J.D. 1996. Significance of feeding habitats of North Atlantic right whales based on studies of diel behaviour, diving, food ingestion rates, and prey, thèse de doctorat, Guelph University, Guelph (Ontario), Canada, 269 p.
Greene, C.H., et A.J. Pershing. 2000. The response of Calanus finmarchicus populations to climate variability in the Northwest Atlantic: basin-scale forcing associated with the North Atlantic Oscillation, ICES Journal of Marine Science57:1536-1544.
Greene, C.H., A.J. Pershing, R.D. Kenney et J.W. Jossi. 2003. Impact of climate variability on the recovery of endangered North Atlantic right whales, Oceanography 16:98-103.
Greene, C.H., et A.J. Pershing. 2004. Climate and the conservation biology of North Atlantic right whales: the right whale at the wrong time? Frontiers in Ecology and Environment 2:29-34.
Greene, C.H., A.J. Pershing, B.C. Monger, M.C. Benfield, E.G. Durbin et M.C. Casas. 2004. Supply-side ecology and the response of zooplankton to climate-driven changes in North Atlantic Ocean circulation, Oceanography 17:60-71.
Hackett, B. 2003. National dredging needs study of U.S. ports and harbors: update 2000, in A report submitted to the U.S. Army Corps of Engineers, Institute for Water Resources, U.S. Army Corps of Engineers, Alexandria (Virginie), États-Unis, 143 p.
Hamilton, P.K., et C.A. Mayo. 1990. Population characteristics of right whales (Eubalaena glacialis) observed in Cape Cod and Massachusetts Bays, 1978 1986, Report of the International Whaling Commission (Special Issue) 12:203 208.
Hamilton, P.K., A.R. Knowlton, M.K. Marx et S.D. Kraus. 1998. Age structure and longevity in North Atlantic right whales Eubalaena glacialis and their relation to reproduction, Marine Ecology Progress Series171:285-292.
Hamilton, P.K., et S.M. Martin. 1999. A catalog of identified right whales from the North Atlantic: 1935-1997, New England Aquarium, Boston (Massachusetts), États-Unis, 27 p.
Hamilton, P.K., A.R. Knowlton et M.K. Marx. 2007. Right whales tell their own stories: The photo-identification catalog, p. 75-104, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Hamilton, P.K., et L.A. Cooper. 2010. Changes in North Atlantic right whale (Eubalaena glacialis) cow-calf association times and use of the calving ground: 1993-2005, Marine Mammal Science26:896-916.
Hamilton, P.K., A.R. Knowlton et S.D. Kraus. 2010. Maintenance of the North Atlantic right whale catalog: 1 January-31 December 2009, New England Aquarium, Boston (Massachusetts), États-Unis, 31 p.
Hamilton, P.K., et A.R. Knowlton. 2011. Maintenance of the North Atlantic right whale catalog: 1 January-31 December 2010. New England Aquarium, Boston (Massachusetts), États-Unis, 34 p.
Hlista, B.L., H.M. Sosik, L.V. Martin Traykovski, R.D. Kenney et M.J. Moore. 2009. Seasonal and interannual correlations between right whale distribution and calving success and chlorophyll concentrations in the Gulf of Maine, États-Unis, 394:289-302.
IWC. 1986. Report of the workshop on the status of right whales, Report of the International Whaling Commission (Special Issue) 10:33.
IWC. 2001a. Report of the workshop on the comprehensive assessment of right whales: a worldwide comparison, Journal of Cetacean Research and Management (Special Issue) 2:1-60.
IWC. 2001b. Report of the scientific committee, Journal of Cetacean Research and Management (Suppl.) 3:1:76.
Jacobsen, K.O., M. Marx et N. Oien. 2004. Two-way trans-Atlantic migration of a North Atlantic right whale (Eubalaena glacialis), Marine Mammal Science 20:161-166.
Johnson, A., G. Salvador, J. Kenney, J. Robbins, S. Landry et P. Clapham. 2005. Fishing gear involved in entanglements of right and humpback whales, Marine Mammal Science 21:635-645.
Johnson, A.J., S.D. Kraus, J.F. Kenney et C.A. Mayo. 2007. The entangled lives of right whales and fisherman: can they coexist? In S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Jonsgard, Å. 1977. Tables showing the catch of small whales (including minke whales) caught by Norwegians in the period 1938-75, and large whales caught in different North Atlantic waters in the period 1868-1975, Report of the International Whaling Commission 27:397-405.
Keller, C.A., L.I. Ward-Geiger, W.B. Brooks, C.K. Slay, C.R. Taylor et B.J. Zoodsma. 2006. North Atlantic right whale distribution in relation to sea-surface temperature in the southeastern United States calving grounds, Marine Mammal Science 22:426-445.
Kenney, R.D., M.A.M. Hyman, R.E. Owen, G.P. Scott et H.E. Winn. 1986. Estimation of prey densities required by western North Atlantic right whales, Marine Mammal Science 2:1-13.
Kenney, R.D., H.E. Winn et M.C. Macaulay. 1995. Cetaceans in the Great South Channel, 1979-1989: right whale (Eubalaena glacialis), Continental Shelf Research 15:385-414.
Kenney, R.D., et K.F. Wishner. 1995. The South Channel Ocean Productivity Experiment, Continental Shelf Research 15:373-384.
Kenney, R.D., C.A. Mayo et H.E. Winn. 2001. Migration and foraging strategies at varying spatial scales in western North Atlantic right whales: a review of hypotheses, Journal of Cetacean Research and Management (Special Issue) 2:251-260.
Kenney, R.D. 2002. North Atlantic, North Pacific, and southern right whales (Eubalaena glacialis, E. japonica, and E. australis), in W.F. Perrin, B. Würsig et J.G.M. Thewissen (éd.), Encyclopedia of marine mammals, Academic Press, San Diego (Californie), ÉTATS UNIS.
Kenney, R.D. 2007. Right whales and climate change: facing the prospect of a greenhouse future, p. 436-459, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), ÉTATS UNIS.
Kenney, R.D. 2010. Right whales in Rhode Island Sound: April 2010, Right Whale News18:5-10.
Kenney, R.D., comm. pers. 2011. Correspondance par courriel adressée à S. Fortune, septembre 2011, Associate Marine Research Scientist, Professor-in-Residence, University of Rhode Island (Rhode Island), États-Unis.
Kite-Powell, H.L., A.R. Knowlton et M.W. Brown. 2007. Modeling the effect of vessel speed on right whale ship strike risk, Project report for NOAA/NMFS, Project NA04NMF47202394:8.
Knowlton, A.R., J. Sigurjónsson, J.N. Ciano et S.D. Kraus. 1992. Long-distance movements of North Atlantic right whales (Eubalaena glacialis), Marine Mammal Science:397-405.
Knowlton, A.R., S.D. Kraus et R.D. Kenney. 1994. Reproduction in North Atlantic right whales (Eubalaena glacialis), Canadian Journal of Zoology 72:1297-1305.
Knowlton, A.R., et S.D. Kraus. 2001. Mortality and serious injury of northern right whales (Eubalaena glacialis), Journal of Cetacean Resource Management (Special Issue)2:193-208.
Knowlton, A.R., et M.W. Brown. 2007. Running the gauntlet: right whales and vessel strikes, p. 409-435, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Knowlton, A.R., P.K. Hamilton, M.K. Marx, H.M. Pettis et S.D. Kraus. 2012. Monitoring North Atlantic right whale Eubalaena glacialis entanglement rates: a 30 yr retrospective, Marine Ecology Progress Series 466:293-302.
Kraus, S.D., J.H. Prescott, P.V. Turnbull et R.R. Reeves. 1982. Preliminary notes on the occurrence of the North Atlantic right whale (Eubalaena glacialis) in the Bay of Fundy, Report of the International Whaling Commission32:407-411.
Kraus, S.D., K.E. Moore, C.A. Price, M.J. Crone, W.A. Watkins, H.E. Winn et J.H. Prescott. 1986a. The use of photographs to identify individual North Atlantic right whales Eubalaena glacialis, Report of the International Whaling Commission (Special Issue)10:145-151.
Kraus, S.D., J.H. Prescott, A.R. Knowlton et G.S. Stone. 1986b. Migration and calving of right whales (Eubalaena glacialis) in the western North Atlantic, Report of the International Whaling Commission (Special Issue)10:139-144.
Kraus, S.D., et R.D. Kenney. 1991. Information on right whales (Eubalaena glacialis) in three proposed critical habitats in United States waters of the western North Atlantic Ocean, Report T-75133740, Marine Mammal Commisson, Washington D.C., États-Unis, 77 p.
Kraus, S.D., et M.W. Brown. 1992. A right whale conservation plan for the waters of Atlantic Canada, p. 79-85, in J.H.M. Willison, S. Bondrupnielsen, C. Drysdale, T. B. Herman, N.W.P.Munro et T.L. Pollock (éd.), Science and the management of protected areas, Elsevier Science, Londres (Royaume Uni).
Kraus, S.D., R.D. Kenney, A.R. Knowlton et J.N. Ciano. 1993. Endangered right whales of southwestern North Atlantic, in Final Report, Minerals Management Service Contract No. 14-35-0001-30486, Edgerton Research Laboratory, New England Aquarium, Boston (Massachusetts), États-Unis, 69 p.
Kraus, S.D., et J.J. Hatch. 2001. Mating strategies in the North Atlantic right whale (Eubalaena glacialis), Journal of Cetacean Resource Management (Special Issue)2:237-244.
Kraus, S.D., M.W. Brown, H. Caswell, C.W. Clark, M. Fujiwara, P.K. Hamilton, R.D. Kenney, A.R. Knowlton, S. Landry, C.A. Mayo, W.A. McLellan, M.J. Moore, D.P. Nowacek, D.A. Pabst, A.J. Read et R.M. Rolland. 2005. North Atlantic right whales in crisis, Science 309:561-562.
Kraus, S.D., R.M. Pace III et T.R. Frasier. 2007. High investment, low return: the strange case of reproduction in Eubalaena glacialis, p. 172-199, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Kraus, S.D., et R.M. Rolland. 2007. Right whales in the urban ocean, p. 1-38, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), États-Unis.
Laist, D.W., A.R. Knowlton, J.G. Mead, A.S. Collet et M. Podesta. 2001. Collisions between ships and whales, Marine Mammal Science17:35-75,
Leaper, R., J. Cooke, P. Trathan, K. Reid, V. Rowntree et R. Payne. 2006. Global climate drives southern right whale (Eubalaena australis) population dynamics, Biology Letters2:289-292.
Leeney, R.H., O.C. Nichols, L. Sette, S. Wood LaFonda et P. Huges. 2010. Marine megavertebrates and fishery resources in the Nantucket Sound--Assessing impacts of marine renewable energy installations on marine megavertebrates--Recommendations for the proposed Muskeget Channel Tidal Energy Project, in Report to Harris Miller Miller & Hanson Inc., septembre 2010, Provincetown Center for Coastal Studies, Provincetown(Massachusetts), États-Unis, 24 p.
Madsen, P.T., M. Wahlberg, J. Tougaard, K. Lucke et P. Tyack. 2006. Wind turbine underwater noise and marine mammals: implications of current knowledge and data needs, Marine Ecology Progress Series309:279-295.
Malik, S., M.W. Brown, S.D. Kraus, A.R. Knowlton, P.K. Hamilton et B.N. White. 1999. Assessment of mitochondrial DNA structuring and nursery use in the North Atlantic right whale (Eubalaena glacialis), Canadian Journal of Zoology 77:1217 1222.
Mate, B., P. Duley, B. Lagerquist, F. Wenzel, A. Stimpert et P. Clapham. 2005. Observations of a female North Atlantic right whale (Eubalaena glacialis) in simultaneous copulation with two males: supporting evidence for sperm competition, Aquatic Mammals 31:157-160.
Mate, B.R., S.L. Nieukirk et S.D. Kraus. 1997. Satellite-monitored movements of the northern right whale, Journal of Wildlife Management61:1393-1405.
Mayo, C.A., et M.K. Marx. 1990. Surface foraging behaviour of the North Atlantic right whale, Eubalaena glacialis, and associated zooplankton characteristics, Canadian Journal of Zoology68:2214-2220.
Mayo, C.A., B.H. Letcher et S. Scott. 2001. Zooplankton filtering efficiency of the baleen of a North Atlantic right whale, Eubalaena glacialis, Journal of Cetacean Research and Management (Special Issue) 2:225-229.
McLeod, B.A., M.W. Brown, T.R. Frasier et B.N. White. 2010. DNA profile of a sixteenth century western North Atlantic right whale (Eubalaena glacialis), Conservation Genetics11:339-345.
Mead, J.G. 1986. Twentieth-century records of right whales (Eubalaena glacialis) in the northwestern North Atlantic, Report of the International Whaling Commission (Special Issue)10:109-119.
Mellinger, D.K., S.L. Nieukirk, H. Matsumoto, S.L. Heimlich, R.P. Dziak, J. Haxel, M. Fowler, C. Meinig et H.V. Miller. 2007. Seasonal occurrence of North Atlantic right whale (Eubalaena glacialis) vocalizations at two sites on the Scotian Shelf, Marine Mammal Science23:856-867.
Mellinger, D.K., S.L. Nieukirk, K. Klinck, H. Klinck, R.P. Dziak, P.J. Clapham et B. Brandsdóttir. 2011. Confirmation of right whales near a nineteenth-century whaling ground east of southern Greenland, Biology Letters, doi:10.1098/rsbl.2010.1191.
Miller, C.A., D. Reeb, P.B. Best, A.R. Knowlton, M.W. Brown et M.J. Moore. 2011. Blubber thickness in right whales Eubalaena glacialis and Eubalaena australis related with reproduction, life history status and prey abundance, Marine Ecology Progress Series438:267-283.
Mitchell, E., V.M. Kozicki et R.R. Reeves. 1986. Sightings of right whales, Eubalaena glacialis, on the Scotian Shelf, 1966-1973, Report of the International Whaling Commission (Special Issue)10:831-101.
Moore, M.J., A.R. Knowlton, S.D. Kraus, W.A. McLellan et R.K. Bonde. 2004. Morphometry, gross morphology and available histopathology in North Atlantic right whale (Eubalaena glacialis) mortalities (1970-2002), Journal of Cetacean Research and Management 6:199-214.
Moore, M.J., W.A. McLellan, D. Pierre-Yves, R.K. Bonde et A.R. Knowlton. 2007. Right whale mortality: a message from the dead to the living, p. 359-379, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard University Press, Cambridge (Massachusetts), ÉTATS UNIS.
Murison, L.D., et D.E. Gaskin. 1989. The distribution of right whales and zooplankton in the Bay of Fundy, Canada, Canadian Journal of Zoology67:1411-1420.
NatureServe. 2001. NatureServe Explorer (en anglais seulement): an online encyclopedia of life , version 7.1, NatureServeArlington (Virgina), : [consulté le 10 juin 2013; ].
NMFS. 1991. Final recovery plan for the northern right whale (Eubalaena glacialis), Prepared by the recovery team for the National Marine Fisheries Service, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Silver Spring (Maryland), États-Unis, 88 p.
NMFS. 1994. Designated critical habitat; Northern right whale, Federal Register 59:28794-28834.
NMFS. 2005. Recovery plan for the North Atlantic right whale (Eubalaena glacialis), U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service, Office of Protected Resources, Silver Spring (Maryland), États-Unis, 138 p.
NMFS. 2006. Recommended shipping routes to reduce ship strikes to North Atlantic Right Whales (en anglais seulement), [consulté le 10 juin 2013; ].
NMFS. 2007. Taking of marine mammals incidental to commerical fishing operations: Atlantic large whale take reduction plan regulations, Federal Register 72:57104 57194.
NMFS. 2008. Endangered fish and wildlife; final rule to implement speed restrictions to reduce the threat of ship collisions with North Atlantic right whales,. Federal Register 73:60173 60191.
NMFS. 2010. Endangered and threatened wildlife and designating critical habitat for the Endangered North Atlantic right whale, Federal Register 75:61690-61691.
NMFS. 2010. Unpublished NMFS sightings data (en anglais seulement).
NMFS. 2011. Site Web en anglais seulement [consulté le 15 septembre 2011; ].
Nowacek, D.P., M.P. Johnson et P.L. Tyack. 2004. North Atlantic right whales (Eubalaena glacialis) ignore ships but respond to alerting stimuli, Proceedings of the Royal Society of London Series B-Biological Sciences 271:227-231.
OMI. 2003. New and amended traffic separation schemes, REF. T2/2.07, COLREG.2/Circ.52 IMO, Londres, Royaume Uni.
OMI. 2006. New and amended traffic separation schemes, REF T2/2.7.1, COLREG.2/Circ.58 IMO, Londres, Royaume Uni.
OMI. 2007. Routing measures other than traffic separation schemes, REF.T2/2.7, COLREG.11/Circ.262 IMO, Londres, Royaume Uni.
OMI. 2008. Routing measures other than traffic separation schemes, Nav 54/3/1 IMO, Londres, Royaume Uni.
Parks, S.E., et P.L. Tyack. 2005. Sound production by North Atlantic right whales (Eubalaena glacialis) in surface active groups, Journal of the Acoustical Society of America117:3297-3306.
Parks, S.E., M.W. Brown, L.A. Conger, P.K. Hamilton, A.R. Knowlton, S.D. Kraus, C.K. Slay et P.L. Tyack. 2007a. Occurrence, composition, and potential functions of North Atlantic right whale (Eubalaena glacialis) surface active groups, Marine Mammal Science 23:868-887.
Parks, S.E., C.W. Clark et P.L. Tyack. 2007b. Short and long-term changes in right whale calling behavior: The potential effects of noise on acoustic communication, Journal of the Acoustical Society of America 122:3725-3731.
Parks, S.E., J.D. Warren, K. Stamieszkin, C.A. Mayo et D. Wiley. 2011. Dangerous dining: surface foraging of North Atlantic right whales increases risk of vessel collisions, Biology Letters:doi 10.1098/rsbl.2011.0578.
Patrician, M.R., I.S. Biedron, H.C. Esch, F.W. Wenzel, L.A. Cooper, P.K. Hamilton, A.H. Glass et M.F. Baumgartner. 2009. Evidence of a North Atlantic right whale calf (Eubalaena glacialis) born in northeastern US waters, Marine Mammal Science 25:462-477.
Payne, R., O. Brazier, E.M. Dorsey, J.S. Perkins, V.J. Rowntree et A. Titus. 1983. External features in southern right whales (Eubalaena australis) and their use in identifying individuals, p. 371-445, in R. Payne (éd.), Communication and behavior of whales, AAAS Selected Symposium 76, Westview Press, Boulder (Colorado), ÉTATS UNIS.
Perrin, W.F. 2013. Balaenidae, World Cetacea Database (en anglais seulement), [consulté le 18 juin 2013;].
Pettis, H.M., R.M. Rolland, P.K. Hamilton, S. Brault, A.R. Knowlton et S.D. Kraus. 2004. Visual health assessment of North Atlantic right whales (Eubalaena glacialis) using photographs, Canadian Journal of Zoology 82:8-19.
Pettis, H.M. 2010. North Atlantic right whale consortium 2010 annual report card (en anglais seulement), [consulté le 10 janvier 2011; .
Rastogi, T., M.W. Brown, B.A. McLeod, T.R. Frasier, R. Grenier, S.L. Cumbaa, J. Nadarajah et B.N. White. 2004. Genetic analysis of 16th-century whale bones prompts a revision of the impact of Basque whaling on right and bowhead whales in the western North Atlantic, Canadian Journal of Zoology 82:1647-1654.
Reeves, R.R., J.G. Mead et S.K. Katona. 1978. The right whale, Eubalaena glacialis, in the western North Atlantic, Report of the International Whaling Commission28:303-312.
Reeves, R.R., et M.F. Barto. 1985. Whaling in the Bay of Fundy, Whalewatcher 19:14 18.
Reeves, R.R., et E. Mitchell. 1986. American pelagic whaling of right whales in the North Atlantic, Report of the International Whaling Commission (Special Issue) 10:221-254.
Reeves, R.R., et R.D. Kenney. 2003. Baleen whales: right whales and allies, Balaena spp, p. 425-463, in G.A. Feldhamer, B.C. Thompson et J.A. Chapman (éd.), Wild mammals of North America: Biology, management, and conservation, second edition, Johns Hopkins University Press, Baltimore (Maryland), États-Unis.
Reeves, R.R., T.D. Smith et E.A. Josephson. 2007. Near-annihilation of a species: right whaling in the North Atlantic, p. 39-74, in S.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Havard University Press, Cambridge (Massachusetts), ÉTATS UNIS.
Reilly, S.B., J.L. Bannister, P.B. Best, M. Brown, R.L. Brownell Jr., D.S. Butterworth, P.J. Clapham, J. Cooke, G. Donovan, J. Urbán et A.N. Zerbini. 2012. Union internationale pour la conservation de la nature[consulté le 10 juillet 2012].
Right Whale Consortium. 2011. The North Atlantic Right Whale sightings and identification database [consulté le 21 octobre 2011], New England Aquarium, Boston (Massachusetts), États-Unis.
Rolland, R., M. Gilbertson et T. Colborn. 1995. Environmentally induced alterations in development: a focus on wildlife, Environmental Health Perspectives 103 (Supplement) 4:3-5.
Rolland, R.M., K.E. Hunt, G.J. Doucette, L.G. Rickard et S.K. Wasser. 2007. The inner whale: hormones, biotoxins and parasites, p. 232-266, inS.D. Kraus et R.M. Rolland (éd.), The urban whale: North Atlantic right whales at the crossroads, Harvard Univeristy Press, Cambridge (Massachusetts), ÉTATS UNIS.
Rosenbaum, H.C., R.L. Brownell, M.W. Brown, C. Schaeff, V. Portway, B.N. White, S. Malik, L.A. Pastene, N.J. Patenaude, C.S. Baker, M. Goto, P.B. Best, P.J. Clapham, P. Hamilton, M. Moore, R. Payne, V. Rowntree, C.T. Tynan, J.L. Bannister et R. Desalle. 2000. World-wide genetic differentiation of Eubalaena: questioning the number of right whale species, Molecular Ecology 9:1793-1802.
Schevill, W.E., et K.E. Moore. 1983. Townsend's unmapped North Atlantic right whales(Eubalaena glacialis), Breviora 476:1-8.
Schevill, W.E., W.A. Watkins et K.E. Moore. 1986. Status of Eubalaena glacialis off Cape Cod, Report of the International Whaling Commission (Special Issue) 10:79-82.
Silber, G.K., L.I. Ward, R. Clarke Lt., K.L. Schumacher et A.J. Smith. 2002. Ship traffic patterns in right whale critical habitat: year one of the mandatory ship reporting system, in NOAA Technical Memorandum NMFS-OPR-20, National Marine Fisheries Service, Office of Protected Resources, Silver Spring (Maryland), ÉTATS UNIS, 27 p
Silva, M.A., L. Steiner, I. Cascão, M. João Cruz, R. Prieto, T. Cole, P. Hamilton et M. Baumgartner. 2012. Winter sighting of a known western North Atlantic right whale in the Azores, Journal of Cetacean Research and Management12:65-69.
Sims, D.W., et V.A. Quayle. 1998. Selective foraging behaviour of basking sharks on zooplankton in a small-scale front, Nature 393:460-464.
Smith, T.D., K. Barthelmess et R.R. Reeves. 2006. Using historical records to relocate a long-forgotten summer feeding ground of North Atlantic right whales, Marine Mammal Science22:723-734.
Stone, G.S., S.D. Kraus, J.H. Prescott et K.W. Hazard. 1988. Significant aggregations of the endangered right whale Eubalaena glacialis on the continental shelf of Nova Scotia Canada, Canadian Field Naturalist 102:417-474.
Taylor, B.L., S.J. Chivers, J. Larese et W.F. Perrin. 2007. Generation length and percent mature estimates for IUCN assessments of cetaceans, Administrative Report LJ-07-01, National Marine Fisheries Service, Southwest Fisheries Science Center, rapport inédit, 24 p.
Turner, J.T., D.G. Borkman et C.D. Hunt. 2006. Zooplankton of Massachusetts Bay, USA, 1992–2003: relationships between the copepod Calanus finmarchicus and the North Atlantic Oscillation, Marine Ecology Progress Series 311:115-124.
Vanderlaan, A.S.M., et C.T. Taggart. 2007. Vessel collisions with whales: the probability of lethal injury based on vessel speed, Marine Mammal Science 23:144-156.
Vanderlaan, A.S.M., et C.T. Taggart. 2009. Efficacy of a voluntary area to be avoided to reduce risk of lethal vessel strikes to endangered whales, Conservation Biology 23:1467-1474.
Vanderlaan, A.S.M., R.K. Smedbol et C.T. Taggart. 2011. Fishing-gear threat to right whales (Eubalaena glacialis) in Canadian waters and the risk of lethal entanglement, Canadian Journal of Fisheries and Aquatic Sciences 68:2174 2193.
Visbeck, M.H., J.W. Hurrell, L. Polvani et H.M. Cullen. 2001. The North Atlantic Oscillation: past, present, and future, Proceedings of the National Academy of Sciences of the USA 98:12876-12877.
Ward-Geiger, L.I., G.K. Silber, R.D. Baumstark et T.L. Pulfer. 2005. Characterization of ship traffic in right whale critical habitat, Coastal Management 33:263-278.
Waring, G.T., E. Josephson, K. Maze-Foley et P.E. Rosel. 2011. U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessment--2010, National Marine Fisheries Service, Woods Hole (Massachusetts), États-Unis, 598 p.
Waring, G.T., E. Josephson, K. Maze-Foley et P.E. Rosel. 2013. U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments - 2012,. in NOAA Technical Memorandum NMFS-NE-223, Woods Hole (Massachusetts), États-Unis, 598 p.
Waters, J.K., R.H. Mayer et D.L. Kriebel. 2000. Shipping trends analysis for impacts of navigation trends on channel usage and design, in A final draft report prepared for the Institute of Water Resources, Department of Naval Architecture and Ocean Engineering, Annapolis (Maryland), États-Unis, 88 p.
Watkins, W.A., et W.E. Schevill. 1982. Observations of right whales, Eubalaena glacialis, in Cape Cod waters, Fishery Bulletin 80:875-880.
Weinrich, M.T., R.D. Kenney et P.K. Hamilton. 2000. Right whales (Eubalaena glacialis) on Jeffreys Ledge: a habitat of unrecognized importance? Marine Mammal Science 16:326-337.
Weisbrod, A.V., D. Shea, M.J. Moore et J.J. Stegeman. 2000. Organochlorine exposure and bioaccumulation in the endangered Northwest Atlantic right whale (Eubalaena glacialis) population, Environmental Toxiology and Chemistry Journal 19:654-666.
Winn, H.E., C.A. Price et P.W. Sorensen. 1986. The distributional biology of the right whale (Eubalaena glacialis) in the western North Atlantic, Report of the International Whaling Commission (Special Issue) 10:129-138.
Wishner, K., E. Durbin, A. Durbin, M. Macaulay, H. Winn et R. Kenney. 1988. Copepod patches and right whales in the Great South Channel off New England, Bulletin of Marine Science 43:825-844.
Woodley, T.H., M.W. Brown, S.D. Kraus et D.E. Gaskin. 1991. Organochlorine levels in North Atlantic right whale (Eubalaena glacialis) blubber, Archives of Environmental Contamination and Toxicology 21:141-145.
Woodley, T.H., et D.E. Gaskin. 1996. Environmental characteristics of North Atlantic right and fin whale habitat in the lower Bay of Fundy, Canada, Canadian Journal of Zoology 74:75-84.
Sommaire biographique du ou des rédacteurs du rapport
Sarah M.E. Fortune est candidate au doctorat à l'Université de la Colombie-Britannique. Ses travaux portent sur l'étude de l'écologie de l'alimentation des baleines boréales et des baleines noires de l'Atlantique Nord. Dans le passé, elle a travaillé pour le programme sur l'habitat de la baleine noire de l'Atlantique Nord (North Atlantic Right Whale Habitat Program, C. Mayo) au Provincetown Center for Coastal Studies, où elle a étudié l'écologie de l'alimentation des baleines noires dans la baie du cap Cod. Elle a effectué les travaux de recherche pour sa maîtrise sur la croissance et l'énergétique de la baleine noire de l'Atlantique Nord à l'Université de la Colombie-Britannique et a étudié l'énergétique des mammifères marins comme adjointe à la recherche au sein de la Marine Mammal Research Unit. Mme Fortune poursuit sa collaboration avec des chercheurs du New England Aquarium, de la Woods Hole Oceanographic Institution, du Provincetown Center for Coastal Studies et du Northwest Fisheries Science Center.
Andrew Trites (Ph.D.) est professeur et directeur de l'Unité des mammifères marins (Marine Mammal Unit) du Centre des pêches (Fisheries Centre) de l'Université de la Colombie-Britannique. Il est aussi directeur de recherche au Consortium de recherche sur les mammifères marins des Universités du Pacifique Nord, associé de recherche à l'aquarium de Vancouver (Vancouver Aquarium), membre du groupe de spécialistes sur les mammifères marins pour le COSEPAC et membre du comité consultatif sur les oiseaux et les mammifères marins de l'Organisation des sciences de la mer pour la Pacifique Nord (PICES). M. Trites étudie les mammifères marins du Pacifique Nord depuis plus de 30 ans. Il effectue des recherches sur des animaux en captivité, des études en milieu naturel et des modélisations pour simulation. De plus, il forme des étudiants des cycles supérieurs et collabore avec des chercheurs spécialisés dans des disciplines comme la nutrition, l'écologie, la physiologie, l'anthropologie et l'océanographie.
Collections examinées
Les bases de données d'identification et des observations de la baleine noire de l'Atlantique Nord (Right Whale Consortium, 2011) ont été consultées pour préparer ce rapport. Voici les principaux organismes ayant contribué à ces bases de données : Blue Ocean Society, Fish and Wildlife Research Institute, Georgia Department of Natural Resources, New England Aquarium, Northeast Fisheries Science Center, Provincetown Center for Coastal Studies, University of North Carolina Wilmington, Whale Center of New England, et Wildlife Trust. La base de données des observations est archivée à l'University of Rhode Island (R. Kenney), et la base de données d'identification, au New England Aquarium (P. Hamilton). Dans le présent rapport, on renvoie à cette collection de données inédites par la mention « Right Whale Consortium, 2011 » .
Communications Personnelles
Rapports de situation du COSEPAC antérieurs :
- Natalie Cadet--Observations Littoral Percé
- Philip Clapham--NMFS, Northeast Science Center
- Robert Michaud--Groupe de recherche sur les mammifères marins
- Rosaline Rolland--New England Aquarium
- Richard Sears--Mingan Island Cetacean Society
- Bradley White--Natural Resources and DNA Profiling and Forensic Centre, Trent University
Rapport de situation du COSEPAC actuel :
- Veronique de la Chenelière--Groupe de recherche et d'éducation sur les mammifères marins
- Robert Kenney--University of Rhode Island
Annexe 1. UICN--Tableau d'évaluation des menaces
Tableau d'évaluation des menaces
- Nom scientifique de l'espèce ou de l'écosystème :
- Baleine noire de l'Atlantique Nord (baleine franche), Eubalaena glacialis
- Identification de l'élément :
- -
- Code de l'élément :
- -
- Date :
- -
- Évaluateur(s) :
- -
- Références :
- -
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de nniveau 1 selon l'intensité de leur impact Maximum de la plage d'intensité | Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 0 | 0 |
| D | Faible | 4 | 4 |
| - | Impact global des menaces calculé : | Moyen | Moyen |
Impact global attribué :
-
Justification de l'ajustement de l'impact :
-
Commentaires sur l'impact global des menaces :
-
Justification de l'ajustement de l'impact :
-
| Nombre | Menace | Impact (calculé) Critère |
Impact (calculé) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | - | - | - | - | - | - |
| 1.1 | Habitations et zones urbaines | - | - | - | - | - | - |
| 1.2 | Zones commerciales et industrielles | - | - | - | - | - | - |
| 1.3 | Tourisme et espaces récréatifs | - | - | - | - | - | - |
| 2 | Agriculture et aquaculture (en anglais seulement) | - | - | - | - | - | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | - | - | - | - | - | - |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | - |
| 2.3 | Élevage et élevage à grande échelle | - | - | - | - | - | - |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | - |
| 3 | Production d'énergie et exploitation minière (en anglais seulement) | - | Impact | Inconnue | Inconnue | Élevée | - |
| 3.1 | Forage pétrolier et gazier | - | - | Restreinte | Inconnue | Élevée | - |
| 3.2 | Exploitation de mines et de carrières | - | - | - | - | - | - |
| 3.3 | Énergie renouvelable | - | - | Restreinte | Inconnue | Élevée | - |
| 4 | Corridors de transport et de service (en anglais seulement | D | Faible | Généralisée à grande | Légère | Élevée | - |
| 4.1 | Routes et voies ferrées | - | - | - | - | - | - |
| 4.2 | Lignes de services publics | - | - | - | - | - | - |
| 4.3 | Transport par eau | D | Faible | Généralisée à grande | Légèret | Élevée | Presque tous les individus traversent les routes maritimes durant la migration. Chaque année, 1,6 animal meurt ou est blessé gravement (1,2 en eaux états-uniennes et 0,4 en eaux canadiennes). C'est < 1 % de la population (plus précisément 0,35 %). Si on applique ce taux de mortalité sur les 10 prochaines années, on obtient une perte totale de 16 individus (3,5 % des 449 individus excluant les baleineaux, vivants en 2010). |
| 4.4 | Trajectoires de vol | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | D | Faible | Généralisée à grande | Légère | Élevée | - |
| 5.1 | Chasse et prélèvement d'animaux terrestres | - | - | - | - | - | - |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | - |
| 5.3 | Exploitation forestière et récolte du bois | - | - | - | - | - | - |
| 5.4 | Pêche et récolte de ressources aquatiques | D | Faible | Généralisée à grande | Légère | Élevée | Environ 1 animal meurt ou est gravement blessé chaque année par les engins de pêche (0,4 en eaux états-uniennes et 0,6 en eaux canadiennes). C'est < 1 % de la population (0,22 % de la population de 2010). À ce rythme, 10 animaux vont mourir au cours des 10 prochaines années, ce qui correspond à environ 2,2 % de la population actuelle. |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | D | Faible | Généralisée | Légère | Élevée | - |
| 6.1 | Activités récréatives | D | Faible | Généralisée | Légère | Élevée | Les collisions avec les bateaux de plaisanciers constituent probablement un problème pour cette espèce, tout comme l'observation des baleines au Canada. |
| 6.2 | Guerres, troubles civils et exercices militaires | - | - | - | - | - | Les exercices militaires pourraient être un problème, mais aucune donnée n'a été publiée à ce sujet. |
| 6.3 | Travaux et autres activités | D | Faible | Généralisée | Légère | Élevée | L'impact des navires de recherche fait l'objet de préoccupations. Un seul cas de collision avec un navire de recherche a été répertorié, mais le navire n'effectuait pas des travaux de recherche à ce moment-là. |
| 7 | Modification du système naturel (en anglais seulement) | - | - | - | - | - | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | - |
| 7.2 | Barrages, gestion et utilisation de l'eau | - | - | - | - | - | - |
| 7.3 | Autres modifications de l'écosystème | - | - | - | - | - | - |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | - | - | Inconnue | Inconnue | Inconnue | - |
| 8.1 | Espèces exotiques/non indigènes envahissantes | - | - | Inconnue | Inconnue | Inconnue | Cela pourrait constituer un problème, mais aucune donnée n'est disponible. |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | - | - |
| 8.3 | Introduction de matériel génétique | - | - | - | - | - | - |
| 9 | Pollution (en anglais seulement) | - | - | Généralisée | Inconnue | Élevée | - |
| 9.1 | Eaux usées domestiques et urbaines | - | - | Généralisée | Inconnue | Élevée | Cela pourrait constituer un problème, mais il n'y a pas de données à ce sujet. |
| 9.2 | Effluents industriels et militaires | - | - | Inconnue | Inconnue | Élevée | Présence de contaminants dans la couche de graisse; les effets sont inconnus. |
| 9.3 | Détritus et déchets solides | - | - | - | - | - | - |
| 9.4 | Détritus et déchets solides | - | - | Inconnue | Inconnue | Inconnue | - |
| 9.5 | Polluants atmosphériques | - | - | Généralisée | Inconnue | Inconnue | Les émanations des moteurs (bateaux et véhicules motorisés) pourraient constituer un problème, mais aucune donnée n'est disponible. |
| 9.6 | Énergie excessive | - | - | - | - | - | - |
| 10 | Phénomènes géologiques (en anglais seulement) | - | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | - | - |
| 11 | Changements climatiques et phénomènes météorologiques violents (en anglais seulement) | D | Faible | Généralisée | Légère | Élevée | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | - |
| 11.2 | Sécheresses | - | - | - | - | - | - |
| 11.3 | Températures extrêmes | D | Faible | Généralisée | Légère | Élevée | Les fluctuations de la température auront probablement une incidence sur le bassin de proies, et des changements positifs et négatifs sont prévus à l'avenir. |
| 11.4 | Tempêtes et inondations | - | - | - | - | - | - |
Classification des menaces d'après l'IUCN-CMP, Salafsky et al. (2008)