
Chauve-souris de Keen (Myotis keenii) évaluation et rapport de situation du COSEPAC: chapitre 3
Information sur l'espèce
Nom et classification
La chauve-souris de Keen (Myotis keenii; Merriam, 1895) appartient à l’ordre des Chiroptères et à la famille des Vespertilionidés. Findley (1972) a classé les Myotis nord-américains à longues oreilles que sont le M. keenii, le vespertilion nordique (Myotis septentrionalis), le vespertilion à longues oreilles (Myotis evotis) et le vespertilion du Sud-Ouest (southwestern bat, Myotis auriculus) dans le groupe evotis du sous-genre Myotis. Le vespertilion de Miller (Myotis milleri) et la chauve-souris à queue frangée (Myotis thysanodes) appartiennent peut-être aussi à ce groupe. Bien qu’ils aient en commun certains traits morphologiques et comportementaux, Manning a conjecturé que les Myotis à longues oreilles nord-américains ne formaient pas un groupe monophylétique. Reducker et al. (1983) ont cependant proposé, en se fondant sur les données d’études chromosomiques et électrophorétiques et d’analyses de séquences de l’ADN mitochondrial, que ce groupe était monophylétique. Toutefois, les analyses de l’ADNmt les plus récentes, réalisées par Dewey (données inédites), montrent que seuls le M. evotis, le M. keeni, le M. thysanodes et le M. milleri forment un groupe monophylétique.
La taxinomie du M. keenii et des espèces apparentées de Myotis à longues oreilles est complexe et l’histoire de la nomenclature prête à confusion. Ce taxon a été décrit et nommé pour la première fois par Merriam (1895), qui l’a considéré comme une sous-espèce du Vespertilio subulatus. La localité-type du spécimen type (USNM 729220) est Masset, dans l’île Graham des îles de la Reine-Charlotte (Colombie-Britannique). Miller (1897) a par la suite classé cette espèce dans le genre Myotis (Myotis subulatus keenii). Dans leur réexamen de la taxinomie des Myotis américains, Miller et Allen (1928) traitent le M. keenii comme une espèce distincte, Myotis keenii, comprenant deux sous-espèces allopatriques : une forme côtière (M. keenii keenii) et une forme du centre et de l’est (M. keenii septentrionalis). Une troisième sous-espèce, le M. keenii auriculus, a été reconnue par Findley (1960) lorsque ce dernier a associé le vespertilion à longues oreilles du sud-ouest des États-Unis à cette espèce, mais Genoways et Jones (1969) considèrent le groupe auriculus comme une espèce distincte (M. auriculus), ce que reconnaissent la plupart des taxinomistes. D’après une analyse multivariée des traits morphologiques, van Zyll de Jong (1979) a conclu que le M. k. keenii et leM. k. septentrionalis étaient des espèces distinctes reconnaissables à des caractéristiques de leur crâne, de leur dentition et de leur pelage. Les deux taxons ont été formellement reconnus comme des espèces distinctes par van Zyll de Jong (1985) et Jones et al. (1986).
Les taxinomistes (p. ex. Manning, 1993) considèrent généralement que parmi les espèces de Myotis à longues oreilles, le M. septentrionalis est le plus proche parent du M. keenii. Cependant, van Zyll de Jong et Nagorsen (1994) concluent, à partir de données morphologiques et biogéographiques, que c’est le M. evotis qui se rapproche le plus du M. keenii. En procédant à une analyse discriminante, ils ont constaté que le M. keenii de Haida Gwaii (ou îles de la Reine-Charlotte1) et le M. evotis des secteurs à l’est de la chaîne Côtière ne présentaient aucun chevauchement morphologique et que les spécimens de musée des régions côtières de la Colombie-Britannique pouvaient être associés avec assurance à l’une ou l’autre des espèces. Par contre, des spécimens de l’ouest du Washington présentaient un certain chevauchement dans leur morphologie. Ces auteurs supposent que les deux espèces sont parapatriques, le M. keenii se trouvant dans les régions côtières et le M. evotis se trouvant principalement à l’est des chaînes côtières. Ils présument que le M. keenii peut avoir évolué dans une zone refuge côtière. Disposant de peu d’échantillons venant de la vallée du bas Fraser, du sud-est de l’île de Vancouver et de l’ouest du Washington, où les aires de répartition des deux espèces se chevauchent, ces chercheurs n’ont pas été en mesure d’évaluer le degré d’introgression.
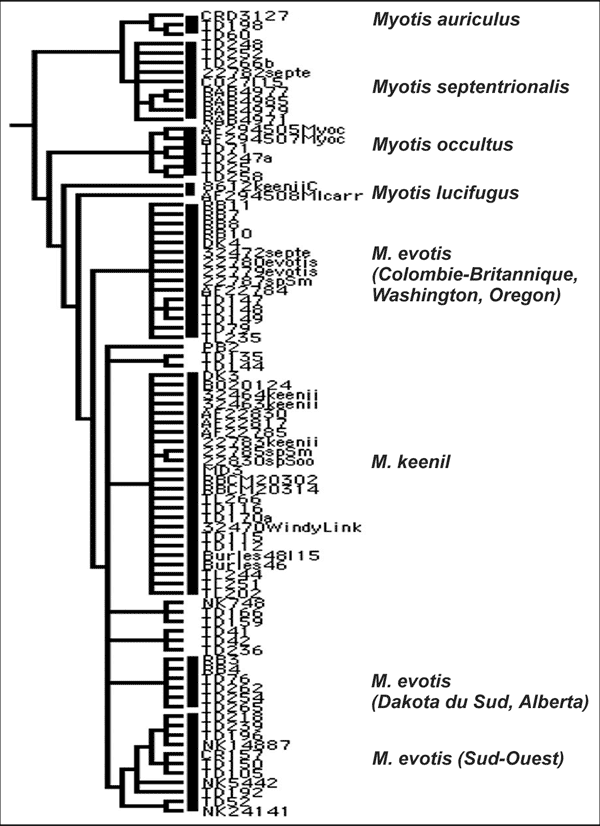
Les résultats d’une analyse récente de l’ADNmt (gène du cytochrome b) par Tanya Dewey (données inédites) confirment la parenté étroite entre le M. keenii et le M. evotis, ainsi que la relation lointaine de ces deux taxons avec le M. septentrionalis (figure 1). La divergence entre les séquences génétiques du M. keenii et du M. septentrionalis dépasse les 10 p. 100, ce qui répond aux critères de différenciation entre espèces (Bradley et Baker, 2001). Dewey note également l’existence de trois lignées relativement distinctes dans le groupe M. keenii/evotis : un clade correspondant à l’aire actuelle du M. keenii, un clade evotis présent dans le sud de la Colombie-Britannique, dans l’État du Washington et l’Oregon, ainsi qu’un clade evotis que l’on trouve surtout de l’Alberta jusqu’au Dakota du Sud. Les clades M. evotis et M. keenii sont largement allopatriques, mais, des individus de ces deux groupes ont été trouvés au même endroit dans les monts Skeena, et les trois clades sont représentées dans la vallée de la Skagit. Les divergences de séquence entre ces lignées vont de 0,8 p. 100 à 3,3 p. 100, ce qui, d’après Bradley et Baker (2001), s’inscrit parfaitement dans la fourchette de divergence intraspécifique. Les résultats de Dewey semblent donc indiquer que le M. evotis et le M. keenii sont conspécifiques, le M. keenii constituant une sous-espèce côtière. Cependant, d’ici à ce que soit réalisée une étude systématique intégrant les données moléculaires (ADN nucléaire et mitochondrial) et morphologiques, nous recommandons que le M. keenii soit considéré comme une unité taxinomique distincte aux fins de conservation et de gestion.
On désigne aussi cette espèce du nom commun de vespertilion de Keen. Les noms communs anglais sont Keen’s bat, Keen’s myotis et Keen’s long-eared myotis. Nous ne connaissons aucun nom autochtone spécifique à cette espèce de chauve-souris. Les Haïdas, par exemple, appellent les chauves-souris collectivement GudGadu Gamhlgaal (prononcer « goutt gaadou gâm hl gaal »), ce qui se traduit par « animal qui se suspend la tête en bas » (B. Wilson, comm. pers.). D’après les consultations menées auprès d’autres groupes autochtones, aucun souvenir culturel ne semble rattaché aux chauves-souris (N. Crookes, comm. pers.; H. Morven, comm. pers.).
Figure 1. Relations phylogénétiques chez des espèces de Myotis à longues oreilles de l’ouest de l’Amérique du Nord. Arbre de parcimonie de consensus dérivé d’une analyse de 765 paires de bases du cytochrome b et de 225 caractères de parcimonie parmi 187 taxons terminaux. Haplotypes en double fusionnés et arbre élagué afin de montrer seulement les échantillons pertinents. Tiré de T. Dewey (données inédites).
Description
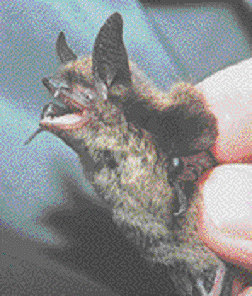
Le M. keenii est une petite chauve-souris (figure 2) au pelage dorsal brun foncé avec une tache foncée imprécise sur l’épaule (van Zyll de Jong, 1985; Nagorsen et Brigham, 1993). Le pelage ventral tire sur le chamois. Une petite frange de poils est visible sur le bord extérieur de la membrane caudale. Les oreilles sont longues (généralement > 16 mm); le calcar (éperon cartilagineux sur l’os du talon) n’a pas de carène bien distincte. Le crâne se caractérise par un rostre relativement étroit et une région frontale fortement inclinée. La formule dentaire est : incisives 2/3, canines 1/1, prémolaires 3/3, molaires 3/3. Les mesures types du corps des adultes (fourchette entre parenthèses) sont : envergure de 241 mm (de 224 à 262), longueur de l’oreille de 17 mm (de 13 à 20), longueur du tragus de 9 mm (de 6 à 12), longueur de l’avant-bras de 36,2 mm (de 33,8 à 39,5), masse corporelle de 5,1 g (de 3,8 à 6,7).
Figure 2. Photographie d’une chauve-souris de Keen (Myotis keenii) capturée dans la réserve de parc national de Gwaii Haanas, Haida Gwaii (Colombie-Britannique). Photo : D.W. Burles.
Le M. keenii ressemble beaucoup au M. evotis. L’identification des animaux vivants est problématique, ce qui constitue un obstacle majeur aux études sur le terrain. En moyenne, les avant-bras, les métacarpiens et les oreilles du M. keenii sont plus courts, mais ces mesures chevauchent celles du M. evotis (van Zyll de Jong et Nagorsen, 1994). En outre, Burles (2001) a démontré que la longueur de l’oreille était un trait variable sujet à d’importantes erreurs de mesure. Les clés d’identification contenues dans van Zyll de Jong (1985) et Nagorsen et Brigham (1993) utilisent la longueur d’oreille dépassant le nez quand l’oreille est repliée vers l’avant (< 5 mm chez le M. keenii; > 5 mm chez le M. evotis) comme critère servant à distinguer les espèces, mais ce trait semble en fait varier grandement entre les individus. Burles (données inédites) a mesuré le dépassement de l’oreille au-delà du nez dans un important échantillon de M. keenii de Haida Gwaii et constaté que cette mesure variait entre 2 et 9 mm. Il conclut que ce n’est pas un caractère taxinomique valable. La couleur de l’oreille, autre trait employé dans les clés de van Zyll de Jong (1985) et Nagorsen et Brigham (1993), semble varier elle aussi. Nagorsen (2002) conclut que le M. keenii et le M. evotis ne peuvent se distinguer avec certitude par leurs traits externes. Les mesures crâniennes et dentaires prises sur des spécimens de musée sont plus fiables, la longueur des rangées de dents étant le meilleur discriminateur unique entre les deux espèces (van Zyll de Jong et Nagorsen, 1994). Néanmoins, ces mesures montrent un certain chevauchement entre les deux taxons, et l’identification exacte requiert une analyse multivariée de mesures crâniennes et dentaires.
Populations importantes à l’échelle nationale
On ne reconnaît au M. keenii aucune sous-espèce et il n’existe pas de population importante à l’échelle nationale.