
Évaluation et Rapport de situation du COSEPAC sur le Coccinelle à neuf points Coccinella novemnotata Canada - 2016

- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendance des populations
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique du rédacteur du rapport
- Collections examinées
- Figure 1. Coccinelle à neuf points (Coccinella novemnotata).
- Figure 2. Aire de répartition de la coccinelle à neuf points (Coccinella novemnotata).
- Figure 3. Zone d'occurrence et indice de zone d'occupation de la coccinelle à neuf points, établis d'après les données de collecte de spécimens de musée et les résultats de relevés récents (1897–2014).
- Figure 4. Zone d'occurrence et indice de zone d'occupation de la coccinelle à neuf points, établis d'après les données de collecte de spécimens de musée (1995–2004).
- Figure 5. Zone d'occurrence et indice de zone d'occupation de la coccinelle à neuf points établis d'après les données de collecte de spécimens de musée et les résultats de relevés récents (2005–2014).
- Figure 6. Activités de recherche ciblant la coccinelle à neuf points (Coccinella novemnotata) (points noirs).
- Figure 7. Chevauchement des sites visités dans le cadre des recherches ciblées (points orange) comportant un rayon de 18 km (points bleus) avec 287 sites connus où la présence de la coccinelle à neuf points (Coccinella novemnotata) avait déjà été observée antérieurement (points noirs).
- Figure 8. Chevauchement des sites visités dans le cadre des recherches ciblées (points orange) comportant un rayon de 120 km (cercles bleus) avec 729 sites connus où la présence de la coccinelle à neuf points (Coccinella novemnotata) avait déjà été observée antérieurement (points noirs).
- Figure 9. Variation de l'abondance relative de la coccinelle à neuf points (Coccinella novemnotata), une espèce indigène (rectangles blancs), et de la coccinelle à sept points (Coccinella septempunctata) (rectangles gris) et de la coccinelle asiatique (Harmonia axyridis) (rectangles noirs), deux espèces non indigènes, par rapport à celle de tous les Coccinellidés inclus dans la base de données récoltés en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec.
- Tableau 1. La base de données contient environ 1 061 mentions de coccinelles à neuf points capturées au Canada entre 1897 et 2014 (voir la section « Collections examinées »).
- Tableau 2. recherches ciblées effectuées en 2013–2014. d'une durée totale de 262,4 heures et étendues à 230 sites, ces recherches se sont soldées par la découverte de quatre individus, soit un à osoyoos (Colombie-Britannique) et trois à medicine hat (Alberta).
- Tableau 2a. Province de la Colombie-Britannique
- Tableau 2b. Province de l'Alberta
- Tableau 2c. Province de l'Ontario
- Tableau 2d. Province de Québec
- Tableau 2e. Province de Terre-Neuve-et-Labrador
- Tableau 2f. Province du Yukon
- Tableau 3a. Variation de l'abondance relative (AR) de la coccinelle à neuf points (CNP) par rapport aux coccinelles (Coccinellidés) indigènes et non indigènes récoltées en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec.
- Tableau 3b. Variation de l'abondance relative (AR) de la coccinelle à neuf points (CNP) par rapport aux coccinelles (Coccinellidés) indigènes récoltées en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec.
- Tableau 4a. Variation en pourcentage de l'abondance relative (AR) sur une période de deux décennies de la coccinelle à neuf points (CNP) par rapport à toutes les coccinelles (Coccinellidés) indigènes et non indigènes récoltées en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec.
- Tableau 4b. Variation en pourcentage de l'abondance relative (AR) sur une période de deux décennies de la coccinelle à neuf points (CNP) par rapport à toutes les coccinelles (Coccinellidés) indigènes récoltées en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec.
- Annexe 1. Résultats obtenus au moyen du calculateur des menaces de l'UICN pour la coccinelle à neuf points.
COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur la coccinelle à neuf points (Coccinella novemnotata) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xi + 62 p.
Le COSEPAC remercie Paul Grant d'avoir rédigé le rapport de situation sur la coccinelle à neuf points (Coccinella novemnotata) au Canada, aux termes d'un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Jennifer Heron, coprésidente du Sous-comité de spécialistes des arthropodes du COSEPAC.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Nine-spotted Lady Beetle Coccinella novemnotata in Canada.
Coccinelle à neuf points - Photo : John Acorn.
Description et importance de l'espèce sauvage
La coccinelle à neuf points (Coccinella novemnotata Herbst) est un petit coléoptère (4,7 à 7,0 mm) indigène en Amérique du Nord. Les adultes se reconnaissent facilement à leurs caractères morphologiques externes distinctifs : les élytres sont orange vif à rouges, avec une ligne foncée bordant la suture de chaque élytre. Les élytres sont généralement ornés de neuf points, mais la taille et le nombre de points peuvent varier. La tête et le pronotum sont noirs avec des marques blanches. Autrefois une des coccinelles les plus communes et les plus largement réparties en Amérique du Nord, cette espèce charismatique jouait un rôle important à titre d'agent de lutte biologique contre les pucerons et d'autres insectes nuisibles.
Répartition
La coccinelle à neuf points est une espèce largement répartie qui se rencontre dans la plupart des régions du sud du Canada bordant la frontière canado-américaine depuis l'île de Vancouver jusque dans le sud du Québec, et au nord, jusqu'à Quesnel en Colombie-Britannique, à Edmonton en Alberta, au lac Athabasca en Saskatchewan et à Roberval au Québec. Vers le sud, cette coccinelle est également présente dans la zone continentale des États-Unis presque jusqu'à la frontière mexicaine.
Habitat
La coccinelle à neuf points est une espèce généraliste en matière d'habitat qui consomme un large éventail de proies dans des habitats très divers. Elle fréquente les milieux agricoles, les jardins suburbains, les parcs, les forêts conifériennes, les forêts décidues, les prairies herbeuses, les prés, les zones riveraines et les aires naturelles isolées. Cette grande plasticité à l'égard de l'habitat témoigne de la capacité de l'espèce de s'adapter aux changements saisonniers de proies qui se produisent dans les différents types de végétation qu'elle fréquente.
Biologie
Le cycle vital de la coccinelle à neuf points comporte quatre grandes étapes : œuf, larve, nymphe et adulte. Cette espèce peut avoir deux générations par année. Les adultes de la génération printanière peuvent entrer en estivation pour échapper aux hautes températures estivales avant de redevenir actifs et de se reproduire au début de l'automne. Les adultes de la génération automnale se rassemblent pour hiberner et entrent en diapause et redeviennent actifs et se reproduisent lorsque les températures s'élèvent au début du printemps. Au Canada, la coccinelle à neuf points occupe une large niche écologique dans divers types d'habitats exposés à des régimes de température différents. On sait peu de choses sur sa capacité de dispersion en milieu naturel. En général, les coccinelles sont des insectes très mobiles qui se montrent peu fidèles à leur lieu de naissance et sont enclins à se disperser sur des distances allant de faibles à grandes. L'action conjuguée de la densité des proies et de diverses variables environnementales telles que la température, la vitesse du vent et les précipitations joue un rôle déterminant dans la dispersion de l'espèce. Cette espèce n'est pas migratrice. Elle est prédatrice tant au stade larvaire qu'à l'âge adulte et se nourrit principalement de pucerons. Elle est elle-même la proie de coccinelles introduites et d'autres invertébrés ainsi que de vertébrés et sert d'hôte à diverses espèces de parasitoïdes et de pathogènes.
Taille et tendances de la population
La vaste aire de répartition historique de la coccinelle à neuf points contraste vivement avec l'aire de répartition actuelle. Avant 1975, cette espèce était très largement répartie et une des espèces de coccinelles les plus fréquemment récoltées en Amérique du Nord. Depuis, cette espèce a subi un déclin et est rarement capturée en dépit des recherches ciblées dont elle est l'objet. L'abondance relative de la coccinelle à neuf points a continué de décliner au cours de la dernière décennie en comparaison de celle d'autres espèces de coccinelles.
Menaces et facteurs limitatifs
Les causes précises du déclin de la coccinelle à neuf points sont inconnues. Les menaces potentielles pesant sur l'espèce comprennent les interactions négatives avec des espèces non indigènes récemment établies, telles la coccinelle à sept points et la coccinelle asiatique, résultant de la compétition, de la prédation intraguilde et des effets indirects de l'introduction de pathogènes. Les effets directs et indirects des pesticides et des autres produits chimiques utilisés en agriculture pour lutter contre les pucerons, principales proies de l'espèce, ainsi que la perte d'habitat occasionnée par l'expansion urbaine, l'abandon des terres agricoles et d'autres perturbations humaines sont également considérés comme des menaces potentielles.
Protection, statuts et classements
Au Canada, aucune loi ne confère une protection à la coccinelle à neuf points et à sa résidence ou à son habitat. À l'échelle mondiale, NatureServe lui a attribué la cote de conservation G2 (en péril). Au Canada, aucune cote infranationale (provinces ou territoires) ne lui a été attribuée, mais au Québec, elle compte parmi les espèces susceptibles d'être désignées menacées ou vulnérables.
| Sujet | Information |
|---|---|
| Durée d'une génération | Deux générations par année. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures? | Oui. Un déclin continu est inféré d'après la réduction de l'abondance relative et la non-détection de l'espèce dans les sites où elle était autrefois commune. |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations]. | Inconnu. |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. | Oui. Réduction inférée de 70 % entre les périodes 1995–2004 et 2005–2014 d'après l'abondance relative de toutes les espèces de coccinelles (indigènes et non indigènes) récoltées. Réduction inférée de 62 % entre les périodes 1995–2004 et 2005–2014 d'après l'abondance relative uniquement des coccinelles indigènes récoltées. |
| Pourcentage [prévu ou présumé] [de changement, de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu. |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu. |
Est-ce que les causes du déclin sont
|
|
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Non. |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence 3 253 910 km2 (1897–2014). 559 510 km2 (1995–2004). |
716 847 km2 (2005–2014). |
| Indice de zone d'occupation (IZO) (toujours fournir une valeur établie à partir d'une grille à carrés de 2 km de côté). 1 308 km2 (1897–2014). 64 km2 (1995–2004). |
40 km2 (2005–2014). |
La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d'occupation totale se trouve dans des parcelles d'habitat qui sont
|
Non. La coccinelle à neuf points est un insecte mobile. C'est également une espèce généraliste en matière d'habitat qui n'est pas confinée à des parcelles d'habitat particulières ou séparées d'autres parcelles d'habitat par une distance excédant sa capacité de dispersion. |
| Nombre de « localités » (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) (utiliser une fourchette plausible pour refléter l'incertitude, le cas échéant). |
Sans objet. Il est impossible de calculer le nombre de localités pour cette espèce. Celle-ci a une très vaste aire de répartition et se montre peu fidèle à l'égard des sites, et les menaces qui pèsent sur elle ne sont pas entièrement comprises. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | Non. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | Oui. Déclin inféré de 37,5 %. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de sous-populations? | Probablement. Un déclin continu est inféré d'après la réduction de l'abondance relative et la non-détection de l'espèce dans les sites où elle était autrefois commune. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.)? |
Inconnu. Il est impossible de calculer le nombre de localités pour cette espèce. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? | Oui. Un déclin continu de la qualité de l'habitat est inféré. |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Peu probable. |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Inconnu. |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Peu probable. |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Peu probable. |
| Sous-populations (indiquer une fourchette plausible) | Nombre d'individus matures |
|---|---|
| - | Inconnu. |
| Total | Inconnu. |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Inconnue. |
| Sujet | Information |
|---|---|
| 8.1 Espèces exotiques (non indigènes) envahissantes, y compris des parasitoïdes/parasites et des pathogènes); 9.3 Effluents agricoles et sylvicoles, notamment les effluents contaminés par des pesticides topiques et systémiques; 2.1 Cultures annuelles et pérennes de produits autres que le bois, incluant l'intensification des cultures; 7.3 Autres modifications de l'écosystème causées par l'abandon des terres gérées et des terres agricoles et la succession naturelle subséquente dans ces milieux. |
Voir le tableau d'évaluation des menaces |
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur. | À l'extérieur du Canada, l'aire de répartition de l'espèce couvre l'ensemble des États-Unis, où les sous-populations ont également connu un déclin important. La dynamique source-puits de cette espèce est inconnue, mais celle-ci est capable de se disperser sur de grandes distances. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Oui. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui. |
| Y a-t-il suffisamment d'habitats disponibles au Canada pour les individus immigrants? | Probablement. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Inconnu. |
| Les conditions de la population source se détériorent-elles? | Inconnu. |
| La population canadienne est-elle considérée comme un puits? | Inconnu. |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Peu probable. La population a subi un déclin important à l'échelle de son aire de répartition aux États-Unis. |
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Non. |
| Sujet | Information |
|---|---|
| COSEPAC | Espèce désignée « en voie de disparition » en avril 2016. |
| Sujet | Information |
|---|---|
| Statut | Espèce en voie de disparition |
| Code alphanumérique | A2bce |
| Justification de la désignation | Cette espèce a déjà été commune et largement répartie dans l'ensemble du sud du Canada, depuis l'île de Vancouver jusque dans le sud du Québec, en passant par les Prairies. Depuis, elle a connu un déclin considérable et est maintenant rarement observée. Malgré des efforts de recherche ciblés au cours de la dernière décennie, l'abondance de l'espèce a diminué relativement à d'autres espèces de coccinelle. Les causes précises du déclin sont inconnues. Les menaces possibles comprennent l'introduction de coccinelles non indigènes, lesquelles pourraient affecter cette espèce indigène par la compétition, la prédation intraguilde, ou l'introduction d'agents pathogènes. D'autres menaces possibles incluent la diminution de la qualité de l'habitat causée par les effets indirects de l'utilisation de pesticides/substances chimiques associés à l'agriculture pour lutter contre leurs espèces proies, l'expansion urbaine, ainsi que l'abandon des terres agricoles et la succession naturelle subséquente. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Les critères A2bce sont satisfaits, car un déclin d'au moins 50 % du nombre d'individus matures au cours des dix dernières années est inféré. Les causes n'ont peut-être pas cessé, ne sont pas comprises et ne sont peut-être pas réversibles; (b) un déclin généralisé de l'abondance relative de l'espèce a été constaté; (c) il y a eu un déclin de l'IZO et de la qualité de l'habitat; (e) des taxons introduits (coccinelle à sept points et coccinelle asiatique), des pathogènes, des parasitoïdes/parasites et des polluants sont soupçonnés d'avoir contribué aux déclins. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Sans objet. L'espèce a une très vaste répartition, et la superficie de la zone d'occurrence est supérieure au seuil. Le critère relatif au nombre de localités n'est pas satisfait; la population n'est pas gravement fragmentée et ne subit pas des fluctuations extrêmes. |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Sans objet. On ne dispose pas de données suffisantes sur le nombre d'individus matures. |
| Critère D (très petite population totale ou répartition restreinte) | Sans objet. On ne dispose pas de données suffisantes sur le nombre d'individus matures. La population canadienne n'est pas restreinte sur le plan de l'IZO, ne satisfait pas au critère relatif au nombre de localités et n'est pas vulnérable à très court terme aux effets des activités humaines ou aux phénomènes stochastiques à l'échelle de son aire de répartition. |
| Critère E (analyse quantitative) | Sans objet. Les données disponibles sont insuffisantes pour faire des projections sur la population canadienne et établir la probabilité de disparition de l'espèce du pays ou de la planète à l'état sauvage. |
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Classe Insecta – Insectes
Sous-classe Pterygota – Ptérygotes ou insectes ailés
Ordre Coleoptera – Coléoptères
Sous-ordre Polyphaga – Polyphages : coccinelles, longicornes, charançons, taupins, lucioles, scarabées, staphylins, etc.
Superfamille Cucujoidea – Cucujoïdes : Coccinellidés, Cucujidés, Laemophloeidés, Silvanidés, Érotylidés, Nitidulidés
Famille Coccinellidae – Coccinellidés ou coccinelles
Sous-famille Coccinellinae
Tribu Coccinellini
Genre Coccinella
Espèce Coccinella novemnotata Herbst, 1793 – Coccinelle à neuf points
Nom scientifique : Coccinella novemnotata
Nom commun français : Coccinelle à neuf points
Noms communs anglais : Nine-spot ladybug, nine-spotted ladybug, nine-spotted lady bird beetle, Nine-spotted Lady Beetle
À l'échelle mondiale, la famille des Coccinellidés comprend environ 6 000 espèces réparties dans environ 360 genres (Vandenberg, 2002; Giorgi et Vandenberg, 2009). Au Canada, la famille est représentée par 60 genres contenant 161 espèces, dont neuf espèces adventices aujourd'hui bien établies (Hodek et al., 2012; Bousquet et al., 2013). La taxinomie, les caractères distinctifs et la répartition géographique de ces espèces au Canada sont bien connus (Dobzhansky, 1935; Watson, 1956; Brown, 1962; Brown et de Ruette, 1962; Belicek, 1976; Watson, 1976; Larochelle, 1979; Gordon, 1985; Vandenberg, 2002; Majka et McCorquodale, 2006; Acorn, 2007; Marriott et al., 2009; Majka et McCorquodale, 2010; Hodek et al., 2012; Bousquet et al., 2013).
Le genre Coccinella contient 15 espèces, dont la plupart se rencontrent en Amérique du Nord. Au Canada, 11 de ces espèces, dont le Coccinella novemnotata, sont indigènes, et deux espèces ont été introduites (ITIS, 2015).
La coccinelle à neuf points (Coccinella novemnotata) (figure 1) a été décrite pour la première fois comme une espèce distincte par Herbst (1793). L'espèce n'a fait l'objet d'aucune étude taxinomique par la suite, et la description originale est considérée comme encore valide. Aucune sous-espèce n'est reconnue.

Description longue de la figure 1
Photo d'une coccinelle à neuf points adulte sur une feuille. Les élytres (ailes antérieures durcies) de la coccinelle à neuf points sont orange pale à rouges et généralement ornés de neuf taches noires de dimensions variables, soit quatre taches sur chaque élytre et une tache centrale. La suture (ligne de jonction des deux élytres) est surlignée par une fine ligne foncée. La tête est large et noire, avec une bande pâle entre les yeux. La marge antérieure du pronotum est entièrement pâle, tandis que la portion postérieure du pronotum est noire.
Les coccinelles sont des insectes holométaboles. Leur cycle vital comporte quatre grandes étapes (œuf, larve, nymphe et adulte). Chacun de ces stades de développement est différent morphologiquement de celui qui le précède.
À l'âge adulte, la coccinelle à neuf points (4,7 à 7,0 mm) a les élytres (ailes antérieures durcies) orange pâle à rouges, habituellement avec neuf taches noires de taille variable, quatre sur chaque élytre et une au centre. Ces taches peuvent varier en nombre et en taille d'un individu à l'autre, au point de faire complètement défaut chez certains individus. Chez la coccinelle à neuf points, la suture (ligne de jonction des deux élytres) est surlignée par une fine ligne foncée. La tête est large et noire avec une bande pâle entre les yeux. La marge antérieure du pronotum est entièrement pâle, tandis que la portion postérieure du pronotum est noire (Gordon, 1985; Acorn, 2007) (figure 1). Les adultes ne montrent aucun dimorphisme sexuel (Stellwag et Losey, 2014). La coccinelle à neuf points se distingue facilement des autres espèces de coccinelles par la marge antérieure pâle de son pronotum et sa suture élytrale noirâtre.
Les œufs, jaunes à orange, de forme allongée et mesurant environ 1 mm de longueur, sont déposés en position dressée en masses compactes d'environ 18 œufs (Hodek et al., 2012). Les larves, en forme de diamant allongé, sont noires avec des taches régulières orange ou rouges sur les côtés et ont les pattes tronquées et parfois hérissées de soies raides. Chez les larves, les tergites (ou segments dorsaux) sont surmontés de projections en forme de tubercules hérissés d'une soie ou de structures semblables à des poils (Rees et al., 1994). Des descriptions détaillées et des clés d'identification des larves sont présentées dans Rees et al. (1994). Les nymphes sont habituellement jaunes à orange avec des marques noires (Hodek et al., 2012).
La structure spatiale et la variabilité des sous-populations de la coccinelle à neuf points n'ont encore jamais été étudiées au Canada. De la même façon, peu d'études génétiques ont été consacrées à l'espèce, et aucun signe apparent de structure génétique subspécifique n'est actuellement connu.
La variation allozymique a été étudiée chez diverses espèces de coccinelles non indigènes (n = 8) et indigènes (n = 6) en Amérique du Nord, dont la coccinelle à neuf points (Krafsur et al., 2005). Aux fins de cette étude, 38 spécimens de coccinelles à neuf points ont été récoltés dans trois régions de l'Amérique du Nord (Iowa, État de New York et Arkansas). Cette étude a révélé que la diversité allélique et le niveau d'hétérozygotie étaient similaires chez les espèces de coccinelles non indigènes et indigènes et, dès lors, qu'aucune relation évidente n'existait entre la colonisation fructueuse de nouveaux milieux et la diversité génétique (Krafsur et al., 2005). Cette étude a également révélé l'existence de taux de flux génétique élevés chez toutes les sous-populations de coccinelles étudiées (Krafsur et al., 2005). Enfin, toutes les espèces de coccinelles ont affiché une capacité de dispersion remarquable et un faible niveau de différenciation en sous-populations distinctes (Krafsur et al., 2005).
La coccinelle à neuf points ne compte qu'une unité désignable au Canada. Aucune sous-espèce n'est reconnue. Même si cette espèce se rencontre dans de nombreuses zones écologiques, le degré de différenciation en sous-populations distinctes est faible (Krafsur et al., 2005).
Les coccinelles sont des espèces iconiques. Avant que ses populations commencent à décliner fortement, la coccinelle à neuf points était l'une des espèces de coccinelles les plus communes au Canada. À titre de prédateur de nombreuses espèces de pucerons et d'autres ravageurs herbivores, elle jouait un rôle économique important comme agent de lutte biologique dans les jardins et les cultures agricoles (Wheeler et Hoebeke, 1995; Hesler et al., 2012). Le déclin observé de cette espèce charismatique a suscité au sein du grand public un intérêt pour sa conservation et pour son rôle dans la fonction des écosystèmes (Evans, 2004; Harmon et al., 2007; Losey et al., 2007; Gardiner et al., 2011; Gardiner et al., 2012; Losey et al., 2012; Bahlai et al., 2013; Turnipseed et al., 2014; Ugine et Losey, 2014).
Des initiatives telles que le Lost Lady Bug Project, qui vise à aider les citoyens scientifiques à trouver des coccinelles à neuf points et à documenter leurs observations à l'échelle de l'aire de répartition de l'espèce en Amérique du Nord, témoignent du grand intérêt du public à l'égard de cette espèce et contribuent à mettre en lumière les changements touchant la composition des communautés de coccinelles qui se produisent à l'échelle des paysages.
Aucune connaissance traditionnelle autochtone ne semble rattachée à cette espèce.
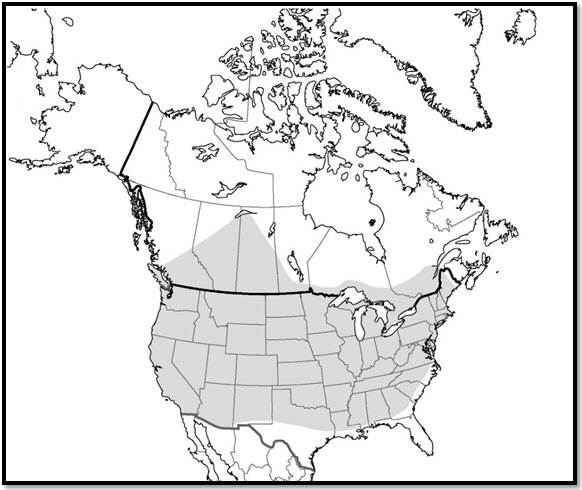
La coccinelle à neuf points est une espèce largement répartie qui se rencontre dans la plupart des régions du sud du Canada et dans la zone continentale des États-Unis jusqu'à la frontière mexicaine (Brown, 1962; Gordon, 1985) (figure 2).
Description longue de la figure 2
Carte illustrant la répartition mondiale de la coccinelle à neuf points, espèce largement répartie qui se rencontre dans la plupart des régions du sud du Canada et dans la zone continentale des États-Unis jusqu'à la frontière mexicaine. Au Canada, la coccinelle à neuf points est présente depuis l'île de Vancouver jusque dans le sud du Québec.
L'aire de répartition canadienne de la coccinelle à neuf points s'étend principalement dans le sud du Canada et les prairies depuis l'île de Vancouver jusque dans le sud du Québec (Brown, 1962; Gordon, 1985; Grant, données personnelles) (figure 2). Aux confins nordiques de son aire de répartition, la coccinelle à neuf points a été observée à proximité de Quesnel (Colombie-Britannique), d'Edmonton (Alberta), du lac Athabasca (Saskatchewan) et de Roberval (Québec). La carte de l'aire de répartition de la coccinelle à neuf points présentée dans Gordon (1985) contient une mention recueillie près du Grand lac des Esclaves, dans les Territoires du Nord-Ouest. Cette mention n'a pu être vérifiée et est considérée comme située à l'extérieur de l'aire de répartition connue de l'espèce. Cette espèce est peut-être présente dans le sud des Territoires du Nord-Ouest et du Yukon, mais cette hypothèse n'était corroborée par aucune mention confirmée au moment de l'élaboration du présent rapport (2016). L'aire de répartition canadienne de l'espèce est fondée sur des mentions de collecte historiques et récentes, mais plusieurs régions n'ont fait aucun relevé et certaines mentions sont passablement anciennes (> 50 ans).
Au cours des dix dernières années, treize mentions de l'espèce ont été enregistrées au Canada, soit dans deux sites à Cranbrook, un site à Kamloops, un site à Osoyoos et deux sites au lac Williams, en Colombie-Britannique; un site à Calgary, un site à Cardston, trois sites à Medicine Hat et un site à Steveville, en Alberta; un site à Mont-Saint-Hilaire, au Québec.
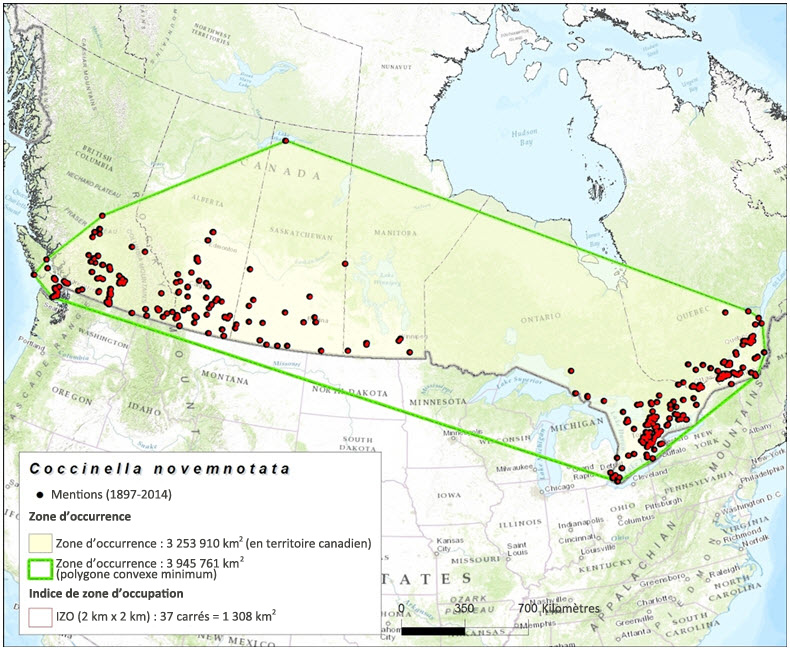
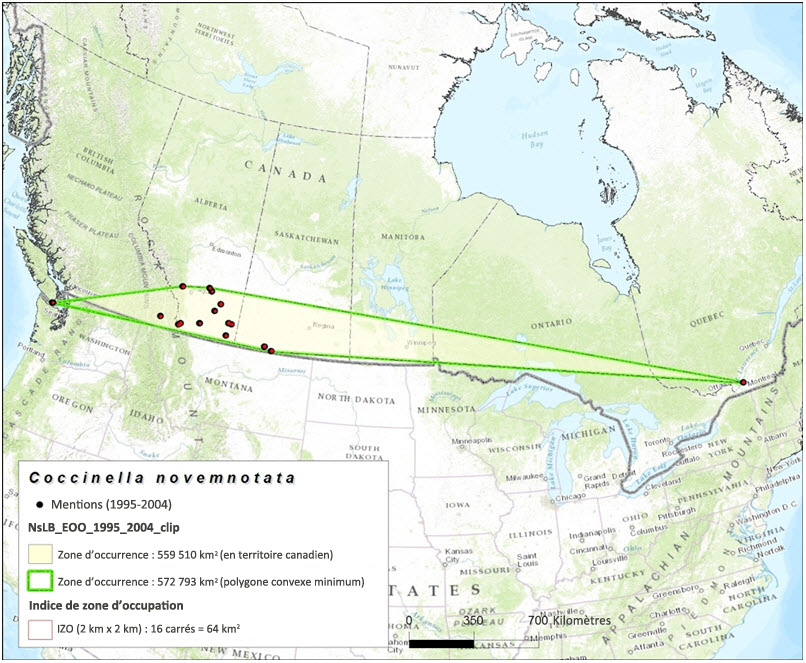
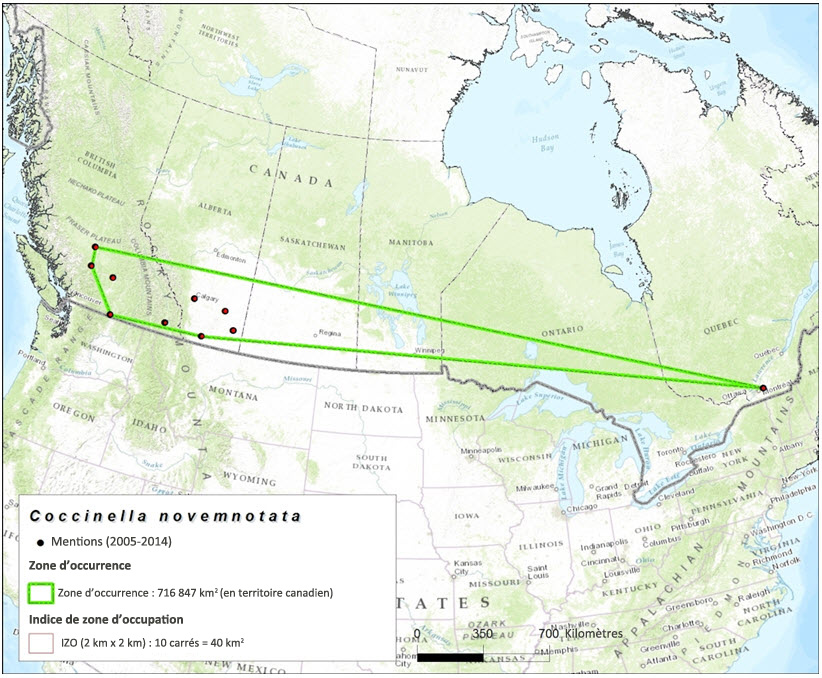
La zone d'occurrence de la coccinelle à neuf points est fondée sur les données de collecte de spécimens de musée et les mentions recueillies dans le cadre de relevés et versées dans une base de données. Établie selon la méthode du polygone convexe minimum à l'intérieur du territoire canadien, la zone d'occurrence pour la période 1897–2014 (toutes les mentions enregistrées dans la base de données) s'élève à 3 253,910 km2 (figure 3). La zone d'occurrence fondée sur les mentions enregistrées durant la période 1995-2004 s'établit à 559 510 km2 (figure 4). Enfin, la zone d'occurrence fondée sur les mentions enregistrées durant la période 2005-2014 s'établit à 716 847 km2 (figure 5).
L'indice de zone d'occupation (IZO) fondée sur les données de collecte de spécimens de musée et les résultats des relevés effectués durant la période 1897-2014 (toutes les mentions versées dans la base de données) s'élève à 1 308 km2 (figure 3). La valeur de l'IZO s'établit à 64 km2 (figure 4) pour la période 1995-2004, et à 40 km2 pour la période 2005–2014 (figure 5).
Description longue de la figure 3
Carte montrant la zone d'occurrence et l'indice de zone d'occupation (IZO) de la coccinelle à neuf points, établis d'après les données de collecte de spécimens de musée et les résultats de relevés récents (1897 à 2014). La zone d'occurrence de l'espèce au Canada s'établit à 3 253 910 km2, et l'IZO, à 1 308 km2.
Description longue de la figure 4
Carte montrant la zone d'occurrence et l'IZO de la coccinelle à neuf points, établis d'après les données de collecte de spécimens de musée (1995 à 2004). La zone d'occurrence de l'espèce au Canada s'établit à 559 510 km2, et l'IZO, à 64 km2.
Description longue de la figure 5
Carte montrant la zone d'occurrence et l'IZO de la coccinelle à neuf points, établis d'après les données de collecte de spécimens de musée et les résultats de relevés récents (2005 à 2014). La zone d'occurrence de l'espèce au Canada s'établit à 716 847 km2, et l'IZO, à 40 km2.
Les dates de capture des spécimens de musée et les mentions de collecte de l'espèce sont réparties entre 1897 et 2014. Une base de données comprenant presque 23 000 mentions de coccinelles (Coccinellidés), dont 1 061 mentions de coccinelles à neuf points, a été créée à partir des données de 26 collections amassées dans diverses régions du Canada (voir la section « Collections examinées »). Les mentions de coccinelles contenues dans cette base de données proviennent de toutes les provinces et de tous les territoires du Canada, mais celles de la coccinelle à neuf points ont été enregistrées uniquement en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec (tableau 1).
| Province/territoire | Nombre de Coccinellidés capturés | Nombre de coccinelles à neuf points capturées |
|---|---|---|
| Yukon | 527 | 0 |
| Territoires du Nord-Ouest | 90 | 0 |
| Nunavut | 1 | 0 |
| Colombie-Britannique | 7 017 | 247 |
| Alberta | 778 | 160 |
| Saskatchewan | 1 793 | 35 |
| Manitoba | 2 369 | 9 |
| Ontario | 6 715 | 331 |
| Québec | 1 950 | 279 |
| Nouveau-Brunswick | 658 | 0 |
| Nouvelle-Écosse | 686 | 0 |
| Île du Prince-Édouard | 65 | 0 |
| Terre-Neuve-et-Labrador | 87 | 0 |
| Total | 22 736 | 1 061 |
Les relevés n'ont pas été effectués de façon systématique ou exhaustive dans le temps et dans l'espace à l'échelle de l'aire de répartition de la coccinelle à neuf points. Pour de vastes pans de territoire et de longues périodes, on dispose de très peu de données. Au Canada, la majorité des recherches ont été effectuées dans des systèmes agricoles ou à proximité de centres urbains, et non pas dans des milieux naturels moins perturbés (Acorn, 2007; McCorquodale et al., 2011). Bien que les données de certaines collections amassées dans diverses régions du Canada n'aient pas encore été versées dans la base de données, l'identité des spécimens de nombreuses autres collections a été établie de manière fiable, et il est donc possible d'évaluer le statut historique des espèces de coccinelles au Canada (McCorquodale et al., 2011; Grant, données personnelles).
Durant la préparation du présent rapport de situation, les sites où l'espèce avait été observée récemment ont été revisités, et des relevés ciblés ont été effectués dans des régions possiblement sous-échantillonnées jusque-là, y compris des régions naturelles situées dans le nord de la Colombie-Britannique, de l'Alberta, du Yukon et des Territoires du Nord-Ouest (figure 6). Un total de 230 sites a été visité durant ces recherches, d'une durée totale de 262,4 heures (tableau 2). Pour un insecte aussi visible et facile à capturer, il s'agit d'un important effort de recherche par site. Toutefois, seulement quatre individus ont été trouvés dans deux sites où l'espèce avait déjà été observée antérieurement, soit trois à Medicine Hat (Alberta) et un à Osoyoos (BC).
La capacité de dispersion de la coccinelle à neuf points est inconnue. Toutefois, si l'on se fonde sur la capacité de dispersion potentielle (en présence de conditions optimales) d'autres espèces de coccinelles (voir la section « Déplacements et dispersion »), cette espèce peut probablement franchir des distances de 18 à 120 km en un seul vol (Jeffries et al., 2013). Ces distances de dispersion ont servi de fondement à l'estimation du chevauchement entre les sites visités dans le cadre des recherches ciblées et les sites où la présence de la coccinelle à neuf points avait déjà été observée antérieurement. Le rayon de 18 km tracé autour des 230 visités dans le cadre des recherches ciblées en 2013 et 2014 chevauchait 287 sites inclus dans la base de données et 729 sites comportant un rayon de 120 km (figure 7 et figure 8). Compte tenu de la vaste répartition et de la grande mobilité de cette espèce, cet effort de recherche représente une couverture relativement satisfaisante des sites connus.
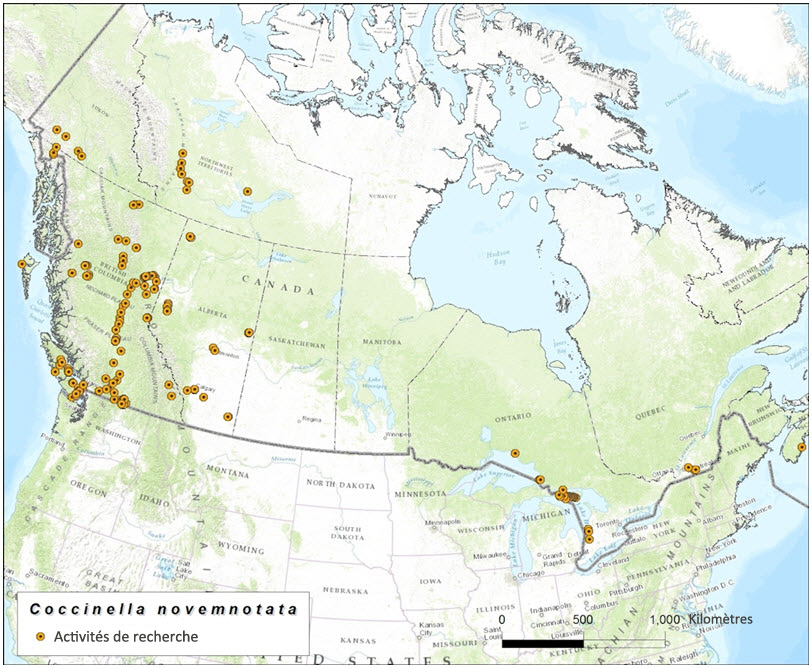
Description longue de la figure 6
Carte indiquant l'emplacement des recherches ciblant la coccinelle à neuf points effectuées en 2013 et 2014 en préparation du présent rapport.
| Localité | Année | Durée | CNP* | Auteur(s) des relevés |
|---|---|---|---|---|
| Arras | 2013 | 90 | Non | Copley C; Copley D; Heron J; Gartner H |
| Arras | 2013 | 35 | Non | Copley C; Copley D; Heron J; Gartner H |
| Vallée de la rivière AshNonla | 2014 | 15 | Non | Heron J; |
| Attachie | 2013 | 462 | Non | Copley C; Copley D; Heron J; Gartner H |
| Attachie | 2013 | 90 | Non | Copley C; Copley D; Heron J; Gartner H |
| Brisco | 2014 | 15 | Non | Grant P |
| Chetwynd | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Chetwynd | 2013 | 90 | Non | Copley C; Copley D; Heron J; Gartner H |
| Clinton | 2013 | 140 | Non | Copley C; Copley D; Heron J; Gartner H |
| Comox | 2014 | 95 | Non | Heron J |
| Lac Coquihalla | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Delta | 2014 | 15 | Non | Heron J |
| Île Denman | 2014 | 15 | Non | Heron J |
| Île Denman | 2014 | 15 | Non | Heron J |
| Île Denman | 2014 | 15 | Non | Heron J |
| Île Denman | 2014 | 15 | Non | Heron J |
| Fairmont Hot Springs | 2014 | 15 | Non | Grant P |
| Fairmont Hot Springs | 2014 | 15 | Non | Grant P |
| Fort St. John | 2013 | 15 | Non | Copley C |
| Fort St. John | 2013 | 124 | Non | Copley C; Copley D; Heron J; Gartner H |
| Fort St. John | 2013 | 420 | Non | Copley C; Copley D; Heron J; Gartner H |
| Fort St. John | 2013 | 53 | Non | Copley C; Copley D; Heron J; Gartner H |
| Fort St. John | 2013 | 210 | Non | Copley C; Copley D; Heron J; Gartner H |
| Fort St. John | 2013 | 435 | Non | Copley C; Copley D; Heron J; Gartner H |
| Fort Ware | 2014 | 15 | Non | Robb B; Copley C; Copley D; |
| Île GaliaNon | 2014 | 30 | Non | Ott L |
| Grand Victoria | 2014 | 15 | Non | Heron J |
| Grand Victoria | 2014 | 15 | Non | Heron J |
| Grand Victoria | 2014 | 15 | Non | S/O |
| Haida Gwaii | 2014 | 60 | Non | McClaren E. |
| Haynes Lease | 2013 | 630 | Non | Sheffield C; Weston M; Heron J |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hazelton | 2014 | 60 | Non | Westcott L |
| Hixon | 2013 | 140 | Non | Copley C; Copley D; Heron J; Gartner H |
| Hope | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Hudson's Hope | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Hudson's Hope | 2013 | 74 | Non | Copley C; Copley D; Heron J; Gartner H |
| Hudson's Hope | 2013 | 255 | Non | Copley C; Copley D; Heron J; Gartner H; Cannings S |
| Hudson's Hope | 2013 | 360 | Non | Copley C; Copley D; Heron J; Gartner H |
| Parc prov. Inkameep | 2013 | 360 | Non | Sheffield C, Weston M; Heron J |
| Parc Iona Beach | 2014 | 30 | Non | Cesselli S; Turner S |
| Parc prov. Kakwa | 2014 | 115 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 5 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 10 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 10 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 15 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 10 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 60 | Non | Ramey B; Bev B |
| Parc prov. Kakwa | 2014 | 5 | Non | Ramey B; Bev B |
| Parc prov. Keily | 2014 | 15 | Non | Robb B; Copley C; Copley D; |
| Parc prov. Keily | 2014 | 15 | Non | Robb B; Copley C; Copley D; |
| Vallée du bas Fraser | 2014 | 30 | Non | S/O |
| Vallée du bas Fraser | 2014 | 30 | Non | S/O |
| Vallée du bas Fraser | 2014 | 30 | Non | S/O |
| Île Mayne | 2014 | 30 | Non | Dunn M |
| Île Mayne | 2014 | 30 | Non | Dunn M |
| Île Mayne | 2014 | 30 | Non | Dunn M |
| Île Mayne | 2014 | 30 | Non | Dunn M |
| Merritt | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Meziadin Junction | 2014 | 60 | Non | Westcott L |
| Okanagan Sud | 2013 | 180 | Non | Sheffield C; Gardiner L; Dyer O; Heron J |
| Mont Kobau | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Mont Kobau | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Mont Kobau | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Mont Kobau | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Mont Kobau | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Mont Kobau | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Nahatlach | 2013 | 60 | Non | Heron J, Lynch G |
| Nahatlach | 2013 | 30 | Non | Heron J, Lynch G |
| Nahatlach | 2013 | 30 | Non | Heron J, Lynch G |
| Nahatlach | 2013 | 30 | Non | Heron J, Lynch G |
| Nord de la Californie | 2014 | 60 | Non | Heron J |
| Nord de la Californie | 2014 | 150 | Non | Heron J |
| Nord de la Californie | 2014 | 30 | Non | Heron J |
| Nord de la Californie | 2014 | 15 | Non | Heron J; Sheffield C |
| Nord de la Californie | 2014 | 15 | Non | Heron J; Sheffield C |
| Nord de la Californie | 2014 | 15 | Non | Heron J; Sheffield C |
| Nord de la Californie | 2014 | 15 | Non | Heron J; Sheffield C |
| Nord de l'île de Vancouver | 2014 | 15 | Non | Copley C; Copley D; Heron J; Gartner H |
| Nord de l'île de Vancouver | 2014 | 15 | Non | Copley C; Copley D; Heron J; Gartner H |
| Okanagan Falls | 2014 | 75 | Non | Heron J; Burdock N |
| Osoyoos | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Osoyoos | 2014 | 15 | OUI 1 | Copley C; Copley D; Heron J; |
| Osoyoos | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Osoyoos | 2014 | 15 | Non | Copley C; Copley D; Heron J; |
| Osoyoos | 2013 | 120 | Non | Heron J; Sheffield C |
| Rivière Pine | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Rivière Pine | 2013 | 120 | Non | Copley C; Copley D; Heron J; Gartner H |
| Prince George | 2013 | 160 | Non | Copley C; Copley D; Heron J; Gartner H |
| Prince George | 2013 | 90 | Non | Copley C; Copley D; Heron J; Gartner H |
| Prince George | 2013 | 140 | Non | Copley C; Copley D; Heron J; Gartner H |
| Prince George | 2013 | 99 | Non | Copley C; Copley D; Heron J; Gartner H |
| Princeton | 2014 | 30 | Non | Heron J |
| Quesnel | 2013 | 180 | Non | Copley C; Copley D; Heron J; Gartner H |
| Quesnel | 2013 | 70 | Non | Copley C; Copley D; Heron J; Gartner H |
| Parc Prov. Keily | 2014 | 15 | Non | Copley C; Copley D; |
| Osoyoos | 2013 | 40 | Non | Heron J; Sheffield C |
| Parc Prov. Russel | 2014 | 15 | Non | Copley C; Copley D; |
| Parc Prov. Russel | 2014 | 15 | Non | Bennett R; Copley C; Copley D; |
| Parc Prov. Russel | 2014 | 15 | Non | Bennett R; Copley C; Copley D; |
| Sage Sparrow Grasslands | 2013 | 360 | Non | Heron J; Sheffield C |
| Smithers | 2014 | 60 | Non | Westcott L |
| Smithers | 2014 | 60 | Non | Westcott L |
| Smithers | 2014 | 60 | Non | Westcott L |
| Smithers | 2014 | 60 | Non | Westcott L |
| Smithers | 2014 | 60 | Non | Westcott L |
| Sooke | 2014 | 15 | Non | Grant P |
| South | 2014 | 15 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J |
| Okanagan-Sud | 2014 | 15 | Non | Heron J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J; Sandhu J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J; Sandhu J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J; Sandhu J |
| Okanagan-Sud | 2014 | 30 | Non | Heron J; Weston W; Bunge S; Pope B |
| Okanagan-Sud | 2013 | 280 | Non | Sheffield C; Gardiner L; Dyer O; Heron J |
| Okanagan-Sud | 2014 | 15 | Non | Heron J; Sandhu J |
| Parc prov. Strathcona | 2014 | 15 | Non | Bennett R; Copley C; Copley D; Heron J |
| Parc prov. Strathcona | 2014 | 15 | Non | Bennett R; Copley C; Copley D; Heron J |
| Sydney | 2014 | 60 | Non | Heron J; Gelling L |
| Tatton | 2013 | 128 | Non | Copley C; Copley D; Heron J; Gartner H |
| Taylor | 2013 | 40 | Non | Copley C; Copley D; Heron J; Gartner H |
| Région de Thompson | 2014 | 30 | Non | Letay S |
| Tsay Keh | 2014 | 15 | Non | Bennett R; Copley C; Copley D; |
| Tsay Keh | 2014 | 15 | Non | Bennett R; Copley C; Copley D; |
| Tsay Keh | 2014 | 15 | Non | Bennett R; Copley C; Copley D; |
| Tumbler Ridge | 2013 | 70 | Non | Copley C; Copley D; Heron J; Gartner H |
| Île de Vancouver | 2014 | 30 | Non | Casselli S; Turner S |
| Île de Vancouver | 2014 | 15 | Non | Heron J |
| Île de Vancouver | 2014 | 15 | Non | Heron J |
| Parc prov. du lac-Vaseux | 2013 | 60 | Non | Heron J; Sheffield C |
| Victoria | 2014 | 15 | Non | Heron J; Gelling L |
| Victoria | 2014 | 15 | Non | Grant P |
| Victoria | 2014 | 15 | Non | Grant P |
| Similkameen | 2013 | 80 | Non | Heron J; Sheffield C |
| Parc prov. Whiskers Point | 2013 | 10 | Non | Copley C; Copley D; Heron J; Gartner H |
| Parc prov. White Lake | 2013 | 315 | Non | Sheffield C; Dyer O; Heron J |
| Lac Williams | 2014 | 30 | Non | Coot K |
| Lac Williams | 2014 | 60 | Non | Coot K; Foot T |
| Lac Williams | 2013 | 132 | Non | Copley C; Copley D; Heron J; Gartner H |
| Lac Williams | 2013 | 80 | Non | Copley C; Copley D; Heron J; Gartner H |
| Localité | Année | Durée | CNP* | Auteur(s) des relevés |
|---|---|---|---|---|
| Calgary | 2014 | 15 | Non | Grant P |
| Calgary | 2014 | 15 | Non | Grant P |
| Calgary | 2014 | 15 | Non | Grant P |
| Calgary | 2014 | 15 | Non | Grant P |
| Calgary | 2014 | 15 | Non | Grant P |
| Calgary | 2014 | 15 | Non | Grant P |
| Calgary | 2014 | 15 | Non | Grant P |
| Cold Lake | 2014 | 15 | Non | Grant P |
| Cold Lake | 2014 | 15 | Non | Grant P |
| Cold Lake | 2014 | 15 | Non | Grant P |
| Cold Lake | 2014 | 15 | Non | Grant P |
| Conklin | 2014 | 15 | Non | Grant P |
| Conklin | 2014 | 15 | Non | Grant P |
| Conklin | 2014 | 15 | Non | Grant P |
| Conklin | 2014 | 15 | Non | Grant P |
| Conklin | 2014 | 15 | Non | Grant P |
| Edmonton | 2014 | 30 | Non | Anweiler G |
| Grande Prairie | 2014 | 15 | Non | Grant P |
| Grande Prairie | 2014 | 15 | Non | Grant P |
| Grande Prairie | 2014 | 15 | Non | Grant P |
| Grande Prairie | 2014 | 15 | Non | Grant P |
| Grande Prairie | 2014 | 15 | Non | Grant P |
| Ruisseau Mclean | 2014 | 15 | Non | Grant P |
| Medicine Hat | 2014 | 30 | Non | Leibel H |
| Medicine Hat | 2014 | 15 | OUI 3 | Buck M |
| Sherwood Park | 2014 | 30 | Non | Anweiler G |
| Sherwood Park | 2014 | 30 | Non | Anweiler G |
| Comté de Vulcan | 2014 | 30 | Non | Leibel H |
| Zama City | 2014 | 15 | Non | Grant P |
| Zama City | 2014 | 15 | Non | Grant P |
| Zama City | 2014 | 15 | Non | Grant P |
| Zama City | 2014 | 15 | Non | Grant P |
| Zama City | 2014 | 15 | Non | Grant P |
| Localité | Année | Durée | CNP* | Auteur(s) des relevés |
|---|---|---|---|---|
| Baie Providence, Île Manitoulin | 2014 | 240 | Non | Foster R; Harris A; Jones C |
| Baie Deans, île Manitoulin | 2014 | 270 | Non | Foster R; Harris A; Jones C |
| Baie Lonely, île Manitoulin | 2014 | 150 | Non | Foster R; Harris A; Jones C |
| Baie Square, Île Manitoulin | 2014 | 105 | Non | Foster R; Harris A; Jones C |
| Baie Dominion, Île Manitoulin | 2014 | 120 | Non | Foster R; Harris A; Jones C |
| Baie Shrigley, Île Manitoulin | 2014 | 165 | Non | Foster R; Harris A; Jones C |
| Baie Portage, Île Manitoulin | 2014 | 180 | Non | Foster R; Harris A; Jones C |
| Taskerville, Île Manitoulin | 2014 | 105 | Non | Foster R; Harris A; Jones C |
| Havre Murphy, Île Manitoulin | 2014 | 30 | Non | Foster R; Harris A; Jones C |
| Baie Misery, Île Manitoulin | 2014 | 180 | Non | Foster R; Harris A; Jones C |
| Baie Sand (Hensly), Île Manitoulin | 2014 | 96 | Non | Foster R; Harris A; Jones C |
| Baie Carroll Wood, Île Manitoulin | 2014 | 105 | Non | Foster R; Harris A; Jones C |
| Havre Burnt I., Île Manitoulin | 2014 | 210 | Non | Foster R; Harris A; Jones C |
| Île Great Duck, | 2014 | 180 | Non | Foster R; Harris A; Jones C |
| Baie Belanger, Île Manitoulin | 2014 | 105 | Non | Foster R; Harris A; Jones C |
| Baie Sand, île Cockburn | 2014 | 300 | Non | Foster R; Harris A; Jones C |
| Aéroport, île Cockburn | 2014 | 90 | Non | Foster R; Harris A; Jones C |
| Embouchure de la rivière Mississaugi | 2014 | 102 | Non | Foster R; Harris A; Jones C |
| Baie Pancake, lac Supérieur | 2014 | 210 | Non | Foster R; Harris A; Jones C |
| Baie Batchewana, lac Supérieur | 2014 | 60 | Non | Foster R; Harris A; Jones C |
| Dunes Pic River, lac Supérieur | 2014 | 48 | Non | Foster R; Harris A; Jones C |
| Parc prov. Point Farms, lac Huron | 2014 | 180 | Non | Foster R; Harris A; Jones C |
| Plage Black's Point, lac Huron | 2014 | 60 | Non | Foster R; Harris A; Jones C |
| Parc prov. Pinery, lac Huron | 2014 | 36 | Non | Foster R; Harris A; Jones C |
| Localité | Année | Durée | CNP* | Auteur(s) des relevés |
|---|---|---|---|---|
| Chemin Magenta | 2014 | 60 | Non | Bereczky V |
| Lac Gale GR11 | 2014 | 60 | Non | Bereczky V |
| Mont Saint-Hilaire | 2014 | 120 | Non | Bereczky V |
| Prairie mont Aki | 2014 | 120 | Non | Bereczky V |
| Localité | Année | Durée | CNP* | Auteur(s) des relevés |
|---|---|---|---|---|
| Rivière Jean-Marie | 2014 | 30 | Non | Allaire D |
| Fort Simpson | 2014 | 30 | Non | Allaire D |
| Fort Simpson | 2014 | 60 | Non | Allaire D |
| Wrigley | 2014 | 30 | Non | Allaire D |
| Wrigley | 2014 | 30 | Non | Allaire D |
| Fort Simpson | 2014 | 30 | Non | Allaire D |
| Wrigley | 2014 | 30 | Non | Allaire D |
| Wrigley | 2014 | 30 | Non | Allaire D |
| Fort Simpson | 2014 | 30 | Non | Allaire D |
| Fort Simpson | 2014 | 30 | Non | Allaire D |
| Localité | Année | Durée | CNP* | Auteur(s) des relevés |
|---|---|---|---|---|
| Nord | 2014 | 45 | Non | Heron J |
| Nord | 2014 | 15 | Non | Heron J; Sheffield C |
| Nord | 2014 | 15 | Non | Heron J; Sheffield C |
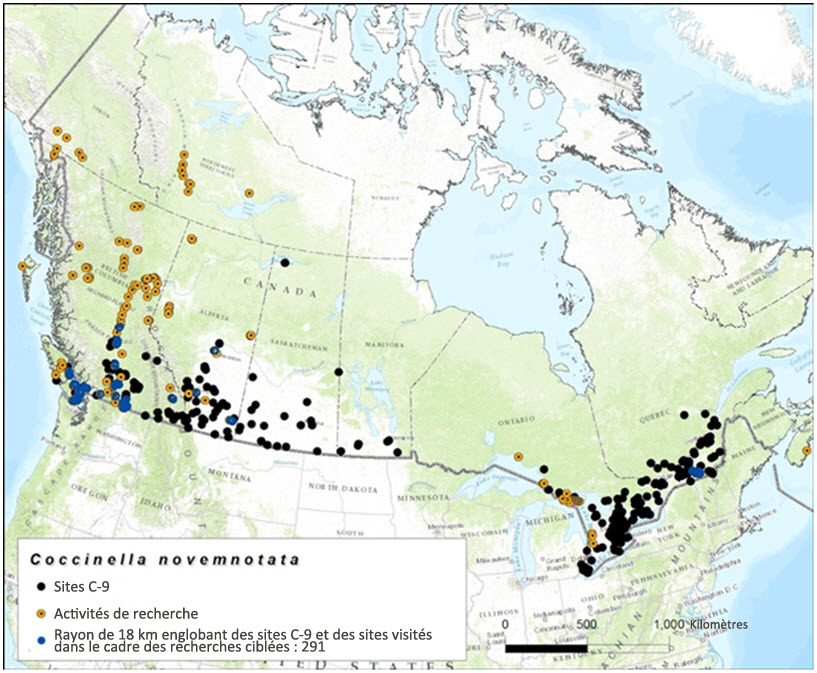
Description longue de la figure 7
Carte montrant le chevauchement entre les sites comportant un rayon de 18 km visités dans le cadre des recherches ciblant la coccinelle à neuf points en 2013 et 2014 et les sites inclus dans la base de données.
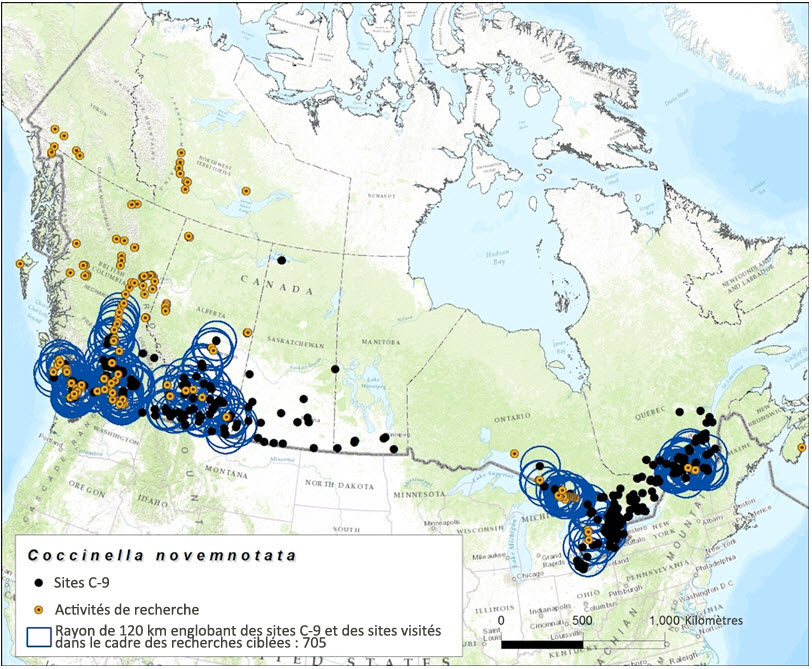
Description longue de la figure 8
Carte montrant le chevauchement entre les sites comportant un rayon de 120 km visités dans le cadre des recherches ciblant la coccinelle à neuf points en 2013 et 2014 et les sites inclus dans la base de données.
La coccinelle à neuf points est une espèce généraliste en matière d'habitat qui est reconnue comme fréquentant les milieux plantés d'arbustes ou de petits arbres intercalés de zones herbeuses ouvertes, mais pas les forêts à couvert complètement fermé. Sa présence a été documentée dans des milieux agricoles, des jardins suburbains, des parcs, des forêts conifériennes, des forêts décidues, des prairies, des prés, des zones riveraines et d'autres milieux naturels ouverts. En milieu agricole, la coccinelle à neuf points était autrefois une des espèces de coccinelles les plus abondantes dans les cultures de luzerne, de pomme de terre, de maïs, de soja et de coton (Wheeler et Hoebeke, 1995; Harmon et al., 2007; Losey et al., 2007; Gardiner et al., 2011). Elle était également communément observée sur différentes plantes cultivées dans les jardins, ainsi que sur les graminées, le trèfle et les mauvaises herbes (Wheeler et Hoebeke, 1995; Harmon et al., 2007; Losey et al., 2007; Gardiner et al., 2011). Elle se rencontre également sur un grand nombre de plantes non cultivées telles que les bouleaux, les pins, les épinettes, les érables, les sorbiers, les peupliers, les saules, les armoises, les pruniers et cerisiers, les aulnes, les chardons, les plantes de prairie et les plantes poussant en bordure de dunes (Wheeler et Hoebeke, 1995; Acorn, 2007; Harmon et al., 2007; Losey et al., 2007).
La coccinelle à neuf points se déplace dans différents milieux et types de végétation pour tirer profit des fluctuations saisonnières de la disponibilité des proies, et sa répartition est donc en grande partie dictée par la disponibilité des proies plutôt que par le type de milieu (Hagen, 1962; Hodek et Honěk, 1996; Sloggett et Majerus, 2000; Hodek et al., 2012).
Les adultes en hibernation se rassemblent généralement dans des microhabitats bien aérés, en particulier sous des pierres, dans des anfractuosités rocheuses, dans des buttes de graminées, dans la litière ou sous l'écorce des arbres (Hodek et al., 2012). Les larves se rencontrent généralement dans des milieux riches en proies et se nymphosent dans ces mêmes milieux.
L'aire de répartition canadienne de la coccinelle à neuf points est vaste et chevauche de nombreuses écozones et de nombreux types de milieux (Gordon, 1985). Cette espèce se disperse également fréquemment sur des distances allant de courtes à grandes et d'un type de végétation à un autre pour tirer profit des fluctuations saisonnières de l'abondance des proies. Aucune étude n'a permis d'établir un lien précis entre les tendances liées à l'habitat et les déclins des sous-populations de cette coccinelle. On ignore si des tendances particulières liées à l'habitat ont joué un rôle dans le déclin de cette espèce largement répartie et au régime alimentaire diversifié dans une grande partie de son aire de répartition connue au Canada.
Les impacts cumulés de la conversion à grande échelle de l'habitat pourraient toutefois avoir contribué aux déclins des sous-populations dans certaines parties de l'aire de répartition de l'espèce. L'expansion des grands centres urbains, y compris les régions métropolitaines de Vancouver, de Victoria et de Calgary, de même que l'exploitation intensive des paysages agricoles et d'autres pratiques industrielles ont entraîné un déclin cumulatif de la qualité de l'habitat et la perte d'habitat (Federal, Provincial and Territorial Governments of Canada, 2010; Javorek et Grant, 2011).
Au cours des dernières décennies, la capacité des paysages agricoles d'offrir aux espèces sauvages l'habitat dont elles ont besoin s'est considérablement amoindrie dans les différentes écozones du Canada (Federal, Provincial and Territorial Governments of Canada, 2010; Javorek et Grant, 2011). Une des causes de ce phénomène est l'utilisation plus intensive des terres agricoles, qui s'est traduite notamment par une dépendance accrue aux produits chimiques antiparasitaires. Ces produits sont soupçonnés d'avoir des effets négatifs directs sur les sous-populations de coccinelles à neuf points ou de leur nuire indirectement en réduisant la disponibilité de leurs proies.
En favorisant la régénération du couvert arborescent, l'abandon des terres et des exploitations agricoles gérées peut également réduire la qualité des aires d'alimentation de la coccinelle à neuf points (Harmon et al., 2007; Bucknell et Pearson, 2007). Ce lent processus de succession naturelle s'est produit surtout dans l'est du Canada.
Bien que les changements à grande échelle de la disponibilité de l'habitat et des proies puissent être perçus comme une possible explication au déclin de l'espèce, aucune donnée ne permet d'établir un lien de causalité entre l'altération du paysage et la diminution de la densité des effectifs de la coccinelle (Elliott et Kieckheffer, 1990; Elliott et al., 1999; Harmon et al., 2007).
Les informations présentées ci-dessous sont tirées d'ouvrages généraux sur les coccinelles (Acorn, 2007; Hodek et al., 2012) et, dans certains cas, de publications traitant spécifiquement de la coccinelle à neuf points.
Les coccinelles sont des insectes holométaboles, c'est-à-dire à métamorphose complète. Leur cycle vital comporte donc quatre étapes : œuf, larve, nymphe, adulte. La coccinelle à neuf points peut avoir deux générations par année (McMullen, 1967) mais, comme chez la plupart des espèces de coccinelles, son cycle vital dépend souvent des conditions climatiques régionales (Hodek et al., 2012). La durée de vie adulte varie en fonction inverse de la température (Hodek et al., 2012). En laboratoire, des coccinelles à neuf points adultes ont vécu 62, 48 et 21 jours à 21 °C, 27 °C et 32 °C, respectivement (McMullen, 1967). Ces données portent à croire que les adultes vivent probablement plus longtemps dans les régions plus fraîches. Les adultes de la génération printanière peuvent entrer en estivation pour échapper aux températures estivales élevées puis redevenir actives et pondre leurs œufs au début de l'automne (McMullen, 1967; Hodek et al., 2012). Les adultes de la génération automnale se rassemblent pour entrer en diapause et hiberner et redeviennent actives et se reproduisent uniquement lorsque les températures s'élèvent au début du printemps suivant (McMullen, 1967; Hodek et al., 2012; Losey et al., 2012).
Chez la coccinelle à neuf points, la période de pré-oviposition (nombre de jours écoulés entre l'émergence de l'adulte [passage de la nymphe à l'adulte] et la première ponte) s'élève à environ 5 jours à 25 °C. La période d'oviposition ou de ponte qui suit dure environ 31 jours (Ugine et Losey, 2014). Durant cette période, les femelles peuvent pondre jusqu'à 690 œufs (Ugine et Losey, 2014). Les œufs sont déposés en position dressée, en masses compactes réunissant environ 18 œufs, sur diverses plantes susceptibles d'être infestées par des sous-populations de pucerons (Acorn, 2007; Hodek et al., 2012). De nombreuses femelles déposent également parmi les œufs fécondés des œufs non fécondés qui serviront de nourriture aux jeunes larves (Acorn, 2007).
Le développement de l'œuf à l'adulte dure environ 20 jours chez la coccinelle à neuf points, selon la température (Ugine et Losey, 2014). L'éclosion survient environ trois jours suivant la ponte (Ugine et Losey, 2014). Les larves passent par quatre stades avant de se nymphoser pour ensuite atteindre l'âge adulte (Losey et al., 2012). Elles mettent environ 4 à 5 jours à atteindre le troisième stade, puis 7 jours à boucler le quatrième stade pour se nymphoser. L'insecte passe ensuite environ 5 jours sous forme de nymphe avant d'atteindre l'âge adulte (Ugine et Losey 2014). Les élytres deviennent durs une journée après l'émergence (Losey et al., 2012). Les mâles repèrent les femelles en se fondant sur des signaux chimiques et visuels, et les deux sexes sont polyandres, s'accouplant avec plusieurs partenaires (Omkar et Srivastava, 2002; Srivastava et Omkar, 2004; Acorn, 2007).
Chez de nombreuses espèces de coccinelles, le sex-ratio s'établit approximativement à 1:1, et l'activité du tissu folliculaire des testicules s'amorce au stade nymphal, si bien que l'accouplement peut débuter peu de temps après l'émergence (Acorn, 2007; Hodek et al., 2012). Chez la coccinelle à neuf points, le sex-ratio est d'environ 56:44 en faveur des femelles, et les femelles sont en moyenne plus lourdes que les mâles (30,3 mg vs 25,6 mg), bien qu'elles soient approximativement de la même taille que ceux-ci (Smith, 1966). Le poids et la taille des coccinelles adultes sont également positivement corrélés à la disponibilité de nourriture, elle-même corrélée à la capacité des coccinelles d'hiberner avec succès (Smith, 1966). Les coccinelles qui ont accès à une quantité de nourriture limitée sont généralement plus petites et plus légères et présentent des taux de survie hivernale plus faibles (Smith, 1966).
La coccinelle à neuf points est une espèce aposématique, c'est-à-dire qu'elle présente une coloration vive destinée à dissuader ses prédateurs (Acorn, 2007). Bien que le phénomène ne soit pas documenté, cette espèce (comme d'autres espèces de coccinelles) peut émettre des alcaloïdes défensifs au niveau de ses articulations tibio-fémorales lorsqu'elle est provoquée (saignée réflexe) (Hodek et al., 2012). Une cinquantaine d'alcaloïdes différents ont été isolés dans la saignée réflexe des coccinelles (Laurent et al., 2005). La composition en alcaloïdes et les effets de ces composés sur les prédateurs varient d'une espèce à l'autre (Marples et al., 1989; Laurent et al., 2005; Hodek et al., 2012).
La coccinelle à neuf points occupe également une large niche écologique couvrant toute une gamme de régimes de température au Canada, résiste bien au froid et peut hiberner à l'âge adulte. Cette plasticité lui permet d'exploiter les fluctuations saisonnières d'abondance de ses proies dans divers types de milieux et de végétation (Hodek et al., 2012). Toutefois, il se peut que sa capacité d'adaptation soit limitée. La compétition avec d'autres espèces de coccinelles introduites pourrait expliquer les réductions de la taille corporelle observées récemment chez la coccinelle à neuf points (Losey et al., 2012) (voir la section « Relations interspécifiques »).
On sait peu de choses sur les taux de dispersion naturelle de la coccinelle à neuf points. De manière générale, les coccinelles sont des insectes très mobiles qui se montrent peu fidèles à leur lieu de naissance et qui se dispersent fréquemment sur des distances allant de courtes à grandes (van der Werf, 2000; Acorn, 2007; Hodek et al., 2012). La capacité de l'espèce de se disperser sur des distances relativement grandes explique les taux de flux génétique élevés observés entre les sous-populations (Krafsur et al., 2005) et permet aux individus de tirer profit des fluctuations de disponibilité de ses proies (Hodek et al., 2012).
Les facteurs favorisant la dispersion sont une combinaison de variables liées à la densité des proies et de variables environnementales telles que la température, la vitesse du vent et les précipitations (Ives et al., 1993; Hodek et Honěk, 1996; van der Werf, 2000; Cardinale et al., 2006; Krivan, 2008; Jeffries et al., 2013). Des études antérieures ont également montré que l'émigration des coccinelles diminue lorsque la densité des proies augmente (Ives, 1981; Ives et al., 1993; Elliott, 2000; van der Werf, 2000; Cardinale et al., 2006; Jeffries et al., 2013) et que la densité des coccinelles adultes est corrélée positivement à la densité des pucerons (Turchin et Kareiva, 1989; Hodek et Honěk, 1996; Osawa, 2000; Evans et Toler, 2007).
L'estimation des taux de dispersion sur des distances plus grandes est compliquée par la difficulté de suivre les insectes sur le terrain. Une étude utilisant des radars entomologiques verticaux a révélé que la majorité des coccinelles volent à une altitude de 150 à 479 m au-dessus du sol, peut-être pour profiter de températures plus fraîches et obtenir l'énergie accrue nécessaire pour atteindre des altitudes plus grandes (Jeffries et al., 2013). La vitesse de vol moyenne des coccinelles oscillait entre 31 km/h à 150 m au-dessus du sol à 59 km/h à 1 500 m au-dessus du sol (Jeffries et al., 2013). Dans le cadre d'expériences de vol utilisant des coccinelles « harnachées », des durées de vol moyenne de 36,5 minutes et maximale de plus de 2 heures ont été enregistrées (Jeffries et al., 2013). En extrapolant à partir de ces résultats, les chercheurs ont estimé qu'en présence de conditions météorologiques optimales, les coccinelles auraient pu parcourir 18 km en un seul vol (30 km/h pendant 36,5 minutes) et que les quelques individus volant à des altitudes et à des vitesses plus élevées (59 km/h pendant deux heures) auraient franchi 120 km en un seul vol (Jeffries et al., 2013).
La coccinelle à neuf points est une espèce généraliste au plan trophique et en matière d'habitat qui tire souvent profit des fluctuations d'abondance des pucerons dans de nombreux types de milieux (Hagen, 1962; Hodek et Honěk, 1996; Sloggett et Majerus, 2000). Tant les larves que les adultes se nourrissent principalement de nombreuses espèces de pucerons (Acorn, 2007; Hodek et al., 2012), mais aussi d'autres petits insectes et autres invertébrés et d'œufs (p. ex. tétranyques, charançon postiche de la luzerne, cicadelles, cochenilles, psylles, œufs de lépidoptères) et de sève, de nectar et de pollen (Wheeler et Hoebeke, 1995; Acorn, 2007; Hesler et al., 2012; Losey et al., 2012). De manière générale, les coccinelles peuvent être attirées par des densités de pucerons de moins de 10 individus par mètre carré et même par les composés volatils émis par les plantes blessées par des herbivores (Hodek et al., 2012).
La coccinelle à neuf points est elle-même victime de prédation intraguilde par d'autres espèces de coccinelles introduites (Turnipseed et al., 2014). Le déclin des sous-populations de coccinelles à neuf points coïncide largement avec l'introduction et la propagation de la coccinelle à sept points (Coccinella septempunctata) et de la coccinelle asiatique (Harmonia axyridis). Bien qu'il soit difficile d'établir un lien de causalité direct, divers mécanismes pourraient avoir contribué au déclin observé, notamment la compétition directe pour la nourriture, la prédation intraguilde et la propagation de nouveaux parasitoïdes ou pathogènes. La compétition avec les espèces de coccinelles introduites pour les pucerons et d'autres sources de nourriture est également soupçonnée d'être à l'origine de la réduction de la taille corporelle observée chez la coccinelle à neuf points (Losey et al., 2012) (voir la section « Menaces »). La réduction de la taille corporelle résultant d'une compétition avec les espèces de coccinelles introduites pourrait également compromettre la survie hivernale de l'espèce (Smith, 1966; Losey et al., 2012).
La coloration aposématique des coccinelles et leur capacité d'excréter par saignée réflexe des alcaloïdes défensifs ayant mauvais goût au niveau de leurs articulations tibio-fémorales contribuent à amoindrir la prédation par des vertébrés tels que les oiseaux (Laurent et al., 2005; Acorn, 2007; Hodek et al., 2012). Ces mécanismes de défense ne leur procurent toutefois qu'une protection partielle, car les coccinelles sont consommées par de nombreuses espèces de prédateurs vertébrés et invertébrés (Acorn, 2007; Hodek et al., 2012), et les araignées qui tissent des toiles en font souvent leurs proies (Nentwig, 1983; Richardson et Hanks, 2009; Sloggett, 2010).
De façon générale, les coccinelles sont parasitées par de nombreuses espèces de mouches (Tachinidés et Phoridés), de guêpes de la superfamille des Chalcidoïdes, d'acariens, de nématodes, de sporozoaires, de champignons et de bactéries (Wheeler et Hoebeke, 1995; Acorn, 2007; Bjornson, 2008; Roy et Cottrell, 2008; Hodek et al., 2012).
La guêpe Dinocampus coccinellae, de la famille des Braconidés, est le principal parasitoïde de nombreuses espèces de coccinelles, y compris la coccinelle à sept points et la coccinelle asiatique, et il peut causer des déclins substantiels chez les sous-populations de la coccinelle à neuf points (Ceryngier et Hodek, 1996; Abassi et al., 2001; Acorn, 2007 ; Hodek et al., 2012). Cette guêpe est présente sur tous les continents à l'exception de l'Antarctique ainsi que sur de nombreuses îles (Hodek et al., 2012). Il est difficile de déterminer l'aire de répartition naturelle d'origine de cette guêpe parce qu'il semble qu'elle ait atteint certaines des régions où elle est actuellement présente avec des coccinelles relâchées à titre d'agents de lutte biologique (Hodek et al., 2012).
La coccinelle à neuf points a également des relations interspécifiques avec des acariens parasites (c.-à-d. Coccipolipus hippodamiae), des champignons pathogènes (c.-à-d. Beauveria bassiana), des microsporidies (Nosématidés) et des bactéries. Tous ces organismes peuvent avoir une incidence néfaste sur sa valeur adaptative et sa survie hivernale (Cali et Briggs, 1967; Hurst et al., 1995; Barron et Wilson, 1998; Webberley et Hurst, 2002; Webberley et al., 2004).
Les collections d'insectes constituent d'importantes sources d'information sur la répartition géographique des espèces (Wiggins et al., 1991). Les spécimens conservés dans les collections canadiennes ont été capturés par des entomologistes professionnels, des étudiants et des entomologistes amateurs passionnés dans le cadre d'inventaires de la biodiversité, de relevés généraux ou ciblant des taxons précis, des études écologiques ou des études appliquées en agriculture ou en foresterie. Les données de ces collections ont permis de délimiter l'aire de répartition des diverses espèces de coccinelles et peuvent être utilisées à profit pour évaluer les variations temporelles de la répartition et de l'abondance si leurs forces et leurs faiblesses sont comprises et prises en compte (McCorquodale et al., 2011).
En raison des biais associés, il est difficile de documenter avec précision les changements liés à l'aire de répartition d'une espèce (Fortin et al., 2005; Elith et al., 2006; Koch et Strange, 2009). Une carte illustrant la répartition géographique d'une espèce peut montrer un rétrécissement de l'aire de répartition de cette dernière alors qu'elle reflète en réalité une diminution de la taille d'une sous-population, la probabilité de collecte de l'espèce considérée étant moindre en pareil cas (McCorquodale et al., 2011). En outre, les collections peuvent comporter des biais liés à la présence de séries temporelles et, par conséquent, ne pas refléter l'abondance réelle d'une espèce, les spécialistes cessant souvent de récolter des spécimens des espèces de coccinelles communes (McCorquodale et al., 2011). À l'inverse, les espèces nouvellement introduites et envahissantes peuvent être récoltées de manière disproportionnée par rapport à leur abondance relative réelle (McCorquodale et al., 2011).
Les tendances liées à l'abondance absolue sont également biaisées par l'effort de recherche. En conséquence, la détermination de l'abondance relative ou de la composition en pourcentage d'une espèce donnée par rapport à un nombre total d'espèces est une approche communément utilisée pour mesurer la taille des populations d'insectes et réduire les biais liés à l'effort de recherche. Il est possible d'obtenir une approximation de l'abondance de la coccinelle à neuf points en comparant ses mentions de collecte à celles de toutes les espèces de coccinelles (Coccinellidés) récoltées durant des périodes et sur des territoires similaires. Une comparaison similaire limitée cette fois aux seules espèces indigènes a également été effectuée. Comme les sous-populations des espèces non indigènes peuvent s'étendre et croître rapidement, l'inclusion des espèces non indigènes dans une telle analyse peut amplifier artificiellement l'importance des déclins. À l'inverse, comme de nombreuses espèces de coccinelles indigènes sont en déclin au Canada, leur inclusion dans les comparaisons visant à établir l'abondance relative de la coccinelle à neuf points pourrait entraîner une sous-estimation de l'ampleur des déclins subis par cette dernière.
Des ensembles de données multiples provenant de collections amassées dans diverses régions du Canada (voir la section « Collections examinées ») ont été utilisés pour suivre dans le temps l'évolution générale de la répartition géographique et de l'abondance de la coccinelle à neuf points. La base de données contient près de 23 000 mentions de Coccinellidés capturés entre 1895 et 2014, dont 1 061 mentions de coccinelles à neuf points amassées entre 1897 et 2014. McCorquodale et al. (2011) ont visité de nombreuses collections afin d'établir ou de confirmer l'identité des spécimens de Coccinellidés qui s'y trouvaient avant de procéder à la saisie des données de collecte dans la base de données. Subséquemment, d'autres données associées à d'autres spécimens de musée et à des spécimens récoltés dans le cadre de relevés ou pour des collections ont été versées dans la base de données durant la préparation de ce rapport (Grant, données personnelles). Les localités de collecte ont été géoréférencées de manière à permettre l'établissement de cartes de répartition à l'aide d'un logiciel SIG (système d'information géographique). Les données de latitude et de longitude, lorsqu'elles étaient précisées sur les étiquettes, ont été transcrites telles quelles dans la base de données, mais autrement, on a utilisé la latitude et la longitude correspondant au centre de la ville indiquée sur l'étiquette, sauf lorsqu'il était possible de déterminer une localité plus précise. En 2013 et en 2014, plus de 262,4 heures ont été consacrées à la conduite de recherches sur le terrain étendues aux 230 sites inclus dans la base de données (tableau 2).
Les méthodes suivantes ont été utilisées pour caractériser les changements dans le temps de la répartition de la coccinelle à neuf points et la couverture des activités de recherche :
- Changements liés à la zone d'occurrence établie par le COSEPAC au cours des dix dernières années (2005–2014) comparés aux changements survenus au cours de la décennie précédente (1995–2004) et à toutes les mentions incluses dans la base de données (1897–2014) (figures 3 à 5).
- Changements liés à l'indice de zone d'occupation (IZO) calculé par le COSEPAC au cours des dix dernières années (2005–2014) comparés aux changements survenus au cours de la décennie précédente (1995–2004) et à toutes les mentions incluses dans la base de données (1897–2014) (figures 3 à 5).
- L'effort de recherche a été combiné aux distances de dispersion potentielle de 18 km et de 120 km (Jeffries et al., 2013).
- L'abondance relative de la coccinelle à neuf points parmi les spécimens de musée a été déterminée par tranches de dix ans dans chacune des provinces et dans chacun des territoires où l'espèce a été observée. L'abondance relative de la coccinelle à neuf points a été estimée par comparaison du nombre de mentions de l'espèce avec celles de toutes les espèces de coccinelles indigènes et non indigènes récoltées, d'une part, et de toutes les espèces de coccinelles indigènes récoltées, d'autre part (figure 9; tableaux 3 et 4).
Ces données ont été complétées par des résultats de recherches publiés et des avis d'experts documentant les déclins des sous-populations et de l'aire de répartition de la coccinelle à neuf points en Amérique du Nord.
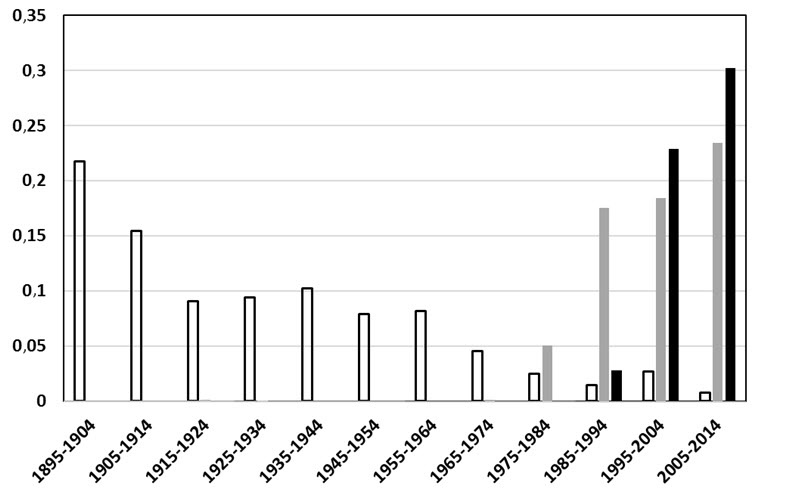
Description longue de la figure 9
Diagramme montrant la variation de l'abondance de la coccinelle à neuf points, de la coccinelle à sept points et de la coccinelle asiatique par rapport à celle de tous les Coccinellidés inclus dans la base de données récoltés en Colombie-Britannique, en Alberta, en Saskatchewan, au Manitoba, en Ontario et au Québec. Les données sont présentées par décennie, de 1895 à 2014.
Il est impossible d'estimer l'abondance d'insectes aussi largement répartis que la coccinelle à neuf points sur la base des données actuellement disponibles. Tel que mentionné précédemment, les changements touchant la zone d'occurrence, l'indice de zone d'occupation (IZO) et l'abondance relative seront utilisés pour mesurer le statut de conservation de l'espèce.
D'après toutes les données et tous les résultats des relevés versés dans la base de données (1897 à 2014), la zone d'occupation de la coccinelle à neuf points s'élève à 3 253 910 km2, et l'IZO, à 1 308 km2 (figure 3). Pour la période 1995-2004, la zone d'occurrence a été estimée à 559 510 km2, et l'IZO, à 64 km2 (figure 4). Au cours de la dernière décennie (2005 à 2014), la zone d'occurrence a augmenté à 716 847 km2, mais l'IZO a chuté à 40 km2 (figure 5). Par rapport à la décennie précédente, il s'agit d'une augmentation de 28 % dans le cas de la zone d'occurrence et d'une diminution de 37,5 % dans celui de l'IZO. La coccinelle à neuf points est largement répartie au Canada et est une espèce très mobile, et aucun relevé couvrant l'ensemble de sa répartition n'a été effectué pour une période donnée. Les tendances liées à la répartition géographique de l'espèce pourraient refléter des lacunes dans la qualité de la couverture des relevés ou dans la détection plutôt qu'une expansion ou une contraction de l'aire de répartition.
Historiquement, la coccinelle à neuf points était largement répartie et se rencontrait dans le sud de la Colombie-Britannique, de l'Alberta, de la Saskatchewan, du Manitoba, de l'Ontario et du Québec. Dans ces régions, elle était l'une des espèces de coccinelles les plus fréquemment récoltées avant 1975 (Brown, 1940; Gordon, 1985). Par la suite, son abondance a décliné de façon substantielle (Acorn, 2007; McCorquodale et al., 2011). Au cours de la décennie précédente (1995 à 2004), 65 mentions de l'espèce ont été enregistrées, et l'abondance relative de l'espèce par rapport à toutes les espèces de coccinelles (Coccinellidés) indigènes et non indigènes récoltées s'établissait à 0,027 (tableau 3a). Au cours des dix dernières années (2005 à 2014), seulement 13 mentions ont été enregistrées en Colombie-Britannique, en Alberta et au Québec (tableau 3a). Durant cette décennie, la coccinelle à neuf points n'a pas été observée en Saskatchewan, au Manitoba ou en Ontario, où elle se rencontrait autrefois. L'abondance relative de la coccinelle à neuf points a donc chuté de -0,019 pour s'établir à seulement 0,008, ce qui correspond à un déclin de 70,7 % à l'échelle nationale au cours des dix dernières années (tableau 4a).
Comme les sous-populations des espèces non indigènes peuvent s'étendre et croître rapidement, l'inclusion de ces espèces peut entraîner une surestimation artificielle de l'ampleur des déclins. En conséquence, aux fins du calcul de l'abondance relative de la coccinelle à neuf points, le nombre de mentions de l'espèce a également été comparé au nombre de mentions des espèces indigènes seulement (les mentions des espèces non indigènes ont été exclues de la comparaison). À l'inverse, comme de nombreuses espèces de coccinelles indigènes sont en déclin au Canada, leur inclusion aux fins du calcul de l'abondance relative de la coccinelle à neuf points peut entraîner une sous-estimation des déclins subis par cette dernière. En comparaison des coccinelles indigènes, l'abondance relative de la coccinelle à neuf points a chuté de 0,057 (1995-2004) à 0,022 (2005-2014) (tableau 3b), ce qui représente un déclin de 62 % à l'échelle nationale au cours des dix dernières années (tableau 4b. Dès lors, le déclin subi par la coccinelle à neuf points à l'échelle nationale est probablement supérieur à 62 % et pourrait être aussi élevé que 70,7 %. Depuis les dix dernières années et dans une vaste majorité des régions comprises dans son aire de répartition, la coccinelle à neuf points est aujourd'hui absente ou continue de décliner, et ses effectifs sont probablement extrêmement faibles.
| Variation | Décennie | C.-B. | Alb. | Sask. | Man. | Ont. | Qc | Total | Variation de l'AR Note du tableaua |
|---|---|---|---|---|---|---|---|---|---|
| Toutes | 1895-1904 | 78 | 2 | 0 | 9 | 32 | 3 | 124 | - |
| CNP | 1895-1904 | 5 | 1 | 0 | 2 | 16 | 3 | 27 | - |
| AR | 1895-1904 | 0,064 | 0,500 | 0,000 | 0,222 | 0,500 | 1,000 | 0,218 | - |
| Toutes | 1905-1914 | 108 | 27 | 16 | 52 | 80 | 8 | 291 | -0,063 |
| CNP | 1905-1914 | 11 | 7 | 0 | 1 | 18 | 8 | 45 | -0,063 |
| AR | 1905-1914 | 0,102 | 0,259 | 0,000 | 0,019 | 0,225 | 1,000 | 0,155 | -0,063 |
| Toutes | 1915-1924 | 495 | 40 | 0 | 124 | 95 | 16 | 770 | -0,064 |
| CNP | 1915-1924 | 18 | 20 | 0 | 1 | 18 | 13 | 70 | -0,064 |
| AR | 1915-1924 | 0,036 | 0,500 | 0 | 0,008 | 0,189 | 0,813 | 0,091 | -0,064 |
| Toutes | 1925-1934 | 1415 | 79 | 5 | 74 | 201 | 156 | 1930 | 0,003 |
| CNP | 1925-1934 | 61 | 27 | 1 | 0 | 24 | 68 | 181 | 0,003 |
| AR | 1925-1934 | 0,043 | 0,342 | 0,200 | 0,000 | 0,119 | 0,436 | 0,094 | 0,003 |
| Toutes | 1935-1944 | 340 | 50 | 112 | 46 | 160 | 159 | 867 | 0,009 |
| CNP | 1935-1944 | 8 | 2 | 0 | 0 | 12 | 67 | 89 | 0,009 |
| AR | 1935-1944 | 0.024 | 0.040 | 0.000 | 0.000 | 0.075 | 0.421 | 0,103 | 0,009 |
| Toutes | 1945-1954 | 816 | 30 | 111 | 350 | 707 | 172 | 2186 | -0.024 |
| CNP | 1945-1954 | 25 | 9 | 3 | 2 | 100 | 33 | 172 | -0.024 |
| AR | 1945-1954 | 0.031 | 0.300 | 0.027 | 0.006 | 0.141 | 0.192 | 0,079 | -0.024 |
| Toutes | 1955-1964 | 770 | 144 | 82 | 121 | 1077 | 201 | 2395 | 0.003 |
| CNP | 1955-1964 | 21 | 43 | 5 | 1 | 75 | 51 | 196 | 0.003 |
| AR | 1955-1964 | 0,027 | 0,299 | 0,061 | 0,008 | 0,070 | 0,254 | 0,082 | 0.003 |
| Toutes | 1965-1974 | 224 | 79 | 338 | 45 | 648 | 372 | 1706 | -0,037 |
| CNP | 1965-1974 | 6 | 13 | 9 | 1 | 30 | 18 | 77 | -0,037 |
| AR | 1965-1974 | 0,027 | 0,165 | 0,027 | 0,022 | 0,046 | 0,048 | 0,045 | -0,037 |
| Toutes | 1975-1984 | 543 | 66 | 563 | 402 | 1637 | 232 | 3443 | -0,020 |
| CNP | 1975-1984 | 19 | 7 | 8 | 0 | 36 | 16 | 86 | -0,020 |
| AR | 1975-1984 | 0,035 | 0,106 | 0,014 | 0,000 | 0,022 | 0,069 | 0,025 | -0,020 |
| Toutes | 1985-1994 | 874 | 18 | 283 | 759 | 658 | 196 | 2788 | -0,011 |
| CNP | 1985-1994 | 28 | 7 | 2 | 1 | 2 | 0 | 40 | -0,011 |
| AR | 1985-1994 | 0,032 | 0,389 | 0,007 | 0,001 | 0,003 | 0,000 | 0,014 | -0,011 |
| Toutes | 1995-2004 | 563 | 50 | 178 | 331 | 1153 | 158 | 2433 | 0,012 |
| CNP | 1995-2004 | 39 | 18 | 7 | 0 | 0 | 1 | 65 | 0,012 |
| AR | 1995-2004 | 0,069 | 0,360 | 0,039 | 0,000 | 0,000 | 0,006 | 0,027 | 0,012 |
| Toutes | 2005-2014 | 791 | 193 | 105 | 56 | 242 | 276 | 1663 | -0,019 |
| CNP | 2005-2014 | 6 | 6 | 0 | 0 | 0 | 1 | 13 | -0,019 |
| AR | 2005-2014 | 0,008 | 0,031 | 0,000 | 0,000 | 0,000 | 0,004 | 0,008 | -0,019 |
| Décennie | C.-B. | Alb. | Sask. | Man. | Ont. | Qc | Total | Variation de l'AR Note du tableaua | |
|---|---|---|---|---|---|---|---|---|---|
| Toutes | 1895-1904 | 67 | 2 | 0 | 9 | 32 | 3 | 113 | - |
| CNP | 1895-1904 | 5 | 1 | 0 | 2 | 16 | 3 | 27 | - |
| AR | 1895-1904 | 0,075 | 0,500 | 0,000 | 0,222 | 0,500 | 1,000 | 0,239 | - |
| Toutes | 1905-1914 | 96 | 27 | 16 | 51 | 80 | 8 | 278 | -0,077 |
| CNP | 1905-1914 | 11 | 7 | 0 | 1 | 18 | 8 | 45 | -0,077 |
| AR | 1905-1914 | 0,115 | 0,259 | 0,000 | 0,020 | 0,225 | 1,000 | 0,162 | -0,077 |
| Toutes | 1915-1924 | 452 | 37 | 0 | 122 | 95 | 16 | 722 | -0,065 |
| CNP | 1915-1924 | 18 | 20 | 0 | 1 | 18 | 13 | 70 | -0,065 |
| AR | 1915-1924 | 0,040 | 0,541 | 0,000 | 0,008 | 0,189 | 0,813 | 0,097 | -0,065 |
| Toutes | 1925-1934 | 1333 | 79 | 5 | 73 | 201 | 156 | 1847 | 0,001 |
| CNP | 1925-1934 | 61 | 27 | 1 | 0 | 24 | 68 | 181 | 0,001 |
| AR | 1925-1934 | 0,046 | 0,342 | 0,200 | 0,000 | 0,119 | 0,436 | 0,098 | 0,001 |
| Toutes | 1935-1944 | 337 | 50 | 112 | 46 | 159 | 158 | 862 | 0,005 |
| CNP | 1935-1944 | 8 | 2 | 0 | 0 | 12 | 67 | 89 | 0,005 |
| AR | 1935-1944 | 0,024 | 0,040 | 0,000 | 0,000 | 0,075 | 0,424 | 0,103 | 0,005 |
| Toutes | 1945-1954 | 801 | 30 | 111 | 349 | 707 | 172 | 2170 | -0,024 |
| CNP | 1945-1954 | 25 | 9 | 3 | 2 | 100 | 33 | 172 | -0,024 |
| AR | 1945-1954 | 0,031 | 0,300 | 0,027 | 0,006 | 0,141 | 0,192 | 0,079 | -0,024 |
| Toutes | 1955-1964 | 741 | 144 | 82 | 121 | 1061 | 199 | 2348 | 0,004 |
| CNP | 1955-1964 | 21 | 43 | 5 | 1 | 75 | 51 | 196 | 0,004 |
| AR | 1955-1964 | 0,028 | 0,299 | 0,061 | 0,008 | 0,071 | 0,256 | 0,083 | 0,004 |
| Toutes | 1965-1974 | 217 | 80 | 339 | 45 | 618 | 294 | 1593 | -0,035 |
| CNP | 1965-1974 | 6 | 13 | 9 | 1 | 30 | 18 | 77 | -0,035 |
| AR | 1965-1974 | 0,028 | 0,163 | 0,027 | 0,022 | 0,049 | 0,061 | 0,048 | -0,035 |
| Toutes | 1975-1984 | 509 | 67 | 563 | 391 | 1447 | 149 | 3126 | -0,021 |
| CNP | 1975-1984 | 19 | 7 | 8 | 0 | 36 | 16 | 86 | -0,021 |
| AR | 1975-1984 | 0,037 | 0,104 | 0,014 | 0,000 | 0,025 | 0,107 | 0,028 | -0,021 |
| Toutes | 1985-1994 | 687 | 19 | 245 | 634 | 333 | 115 | 2033 | -0,008 |
| CNP | 1985-1994 | 28 | 7 | 2 | 1 | 2 | 0 | 40 | -0,008 |
| AR | 1985-1994 | 0,041 | 0,368 | 0,008 | 0,002 | 0,006 | 0,000 | 0,020 | -0,008 |
| Toutes | 1995-2004 | 295 | 37 | 151 | 240 | 355 | 59 | 1137 | 0,037 |
| CNP | 1995-2004 | 39 | 18 | 7 | 0 | 0 | 1 | 65 | 0,037 |
| AR | 1995-2004 | 0,132 | 0,486 | 0,046 | 0,000 | 0,000 | 0,017 | 0,057 | 0,037 |
| Toutes | 2005-2014 | 337 | 91 | 33 | 12 | 71 | 54 | 598 | -0,035 |
| CNP | 2005-2014 | 6 | 6 | 0 | 0 | 0 | 1 | 13 | -0,035 |
| AR | 2005-2014 | 0,018 | 0,066 | 0,000 | 0,000 | 0,000 | 0,019 | 0,022 | -0,035 |
| Province | Nombre de captures, 1995-2004 Nombre de coccinelles indigènes et non indigènes |
Nombre de captures, 1995-2004 Nombre de CNP |
Nombre de captures, 1995-2004 AR |
Nombre de captures, 2005-2014 Nombre de coccinelles indigènes et non indigènes |
Nombre de captures, 2005-2014 Nombre de CNP |
Nombre de captures, 2005-2014 AR |
Variation de l'AR en % par rapport à la décennie précédente |
|---|---|---|---|---|---|---|---|
| C.-B. | 563 | 39 | 0,069 | 791 | 6 | 0,008 | -89,0 |
| Alb. | 50 | 18 | 0,360 | 193 | 6 | 0,031 | -91,4 |
| Sask. | 178 | 7 | 0,039 | 105 | 0 | 0,000 | -100,0 |
| Man. | 331 | 0 | 0,000 | 56 | 0 | 0,000 | 0,0 |
| Ont. | 1153 | 0 | 0,000 | 242 | 0 | 0,000 | 0,0 |
| Qc | 158 | 1 | 0,006 | 276 | 1 | 0,004 | -42,8 |
| Total | 2433 | 65 | 0,027 | 1663 | 13 | 0,008 | -70,7 |
| Province | Nombre de captures, 1995-2004 Nombre de coccinelles indigènes |
Nombre de captures, 1995-2004 Nombre de CNP |
Nombre de captures, 1995-2004 AR |
Nombre de captures, 2005-2014 Nombre de coccinelles indigènes |
Nombre de captures, 2005-2014 Nombre de CNP |
Nombre de captures, 2005-2014 AR |
Variation de l'AR en % par rapport à la décennie précédente |
|---|---|---|---|---|---|---|---|
| C.-B. | 295 | 39 | 0,132 | 337 | 6 | 0,018 | -86,5 |
| Alb. | 37 | 18 | 0,486 | 91 | 6 | 0,066 | -86,4 |
| Sask. | 151 | 7 | 0,046 | 33 | 0 | 0,000 | -100,0 |
| Man. | 240 | 0 | 0,000 | 12 | 0 | 0,000 | 0,0 |
| Ont. | 355 | 0 | 0,000 | 71 | 0 | 0,000 | 0,0 |
| Qc | 59 | 1 | 0,017 | 54 | 1 | 0,019 | 9,3 |
| Total | 1137 | 65 | 0,057 | 598 | 13 | 0,022 | -62,0 |
Le déclin de l'abondance relative de la coccinelle à neuf points coïncide avec une augmentation du nombre de captures d'espèces non indigènes telles que la coccinelle à sept points et la coccinelle asiatique (figure 9).
McCorquodale et al., (2011) a également passé en revue les données issues de la littérature et les données de collecte des spécimens capturés au Québec et en Ontario pour déterminer les variations dans le temps de l'abondance relative et de l'aire de répartition d'un sous-ensemble de 10 espèces indigènes. Il a centré son étude sur des régions pour lesquelles il disposait de données complètes et de haute qualité pour la période correspondant à l'arrivée des espèces non indigènes au Canada. Les spécimens utilisés dans cette étude ont été récoltés par des étudiants du niveau universitaire et dans le cadre de relevés généraux et fournissent une indication raisonnable de l'abondance relative des espèces considérées à l'échelle locale. Dans le cadre de cette étude, McCorquodale et al. (2011) ont inféré que l'aire de répartition et l'abondance relative de la coccinelle à neuf points ont chuté d'environ 18 % avant 1960 à moins de 0,05 % après 1980, période durant laquelle le nombre de captures de coccinelles non indigènes a augmenté. L'abondance relative d'autres espèces de coccinelles indigènes comme la coccinelle maculée (Coleomegilla maculata lengi) a toutefois augmenté après l'arrivée des coccinelles non indigènes (McCorquodale et al., 2011). Ces tendances contradictoires donnent à croire que les déclins observés chez la coccinelle à neuf points ne peuvent être attribués à des biais d'échantillonnage (McCorquodale et al., 2011).
Considérées collectivement, les données sur les changements de l'abondance relative et les cartes illustrant l'évolution de l'aire de répartition des coccinelles au Canada donnent à croire que l'effort de collecte a été suffisamment élevé pour refléter adéquatement l'évolution des populations de coccinelles et confirmer que ces déclins sont réels et non liés à des artéfacts causés par un échantillonnage inadéquat. Elles indiquent également que la coccinelle à neuf points, même si elle persiste à des densités très faibles, a continué de décliner au cours des dix dernières années à l'échelle de son aire de répartition au Canada.
Le déclin de la coccinelle à neuf points dans le nord-est des États-Unis a été documenté par Wheeler et Hoebeke (1995). Ces auteurs citent diverses études démontrant que l'espèce était autrefois commune dans de nombreuses régions du nord-est des États-Unis entre les années 1950 et 1970 mais y a rarement été rencontrée après 1985. Des relevés intensifs de coccinelles effectués en Iowa révèlent que la coccinelle à neuf points y était commune et largement répartie avant 1980 mais est aujourd'hui très rare ou a peut-être même disparu de cet État (Hesler, 2009). Au Minnesota, elle était abondante avant 1980, mais des recherches récentes donnent à croire qu'elle est aujourd'hui absente dans la majorité des régions de l'État ou qu'elle s'y maintient sous le seuil de détection (Koch, 2011). Harmon et al. (2007) ont passé en revue la littérature publiée et les mentions colligées par l'United States Department of Agriculture (USDA) et ont conclu que l'abondance relative des sous-populations de la coccinelle à neuf points a diminué de façon significative aux États-Unis et au Canada depuis les années 1970.
De façon générale, l'abondance relative des assemblages d'espèce de coccinelles indigènes par rapport aux assemblages d'espèces non indigènes au Canada et aux États-Unis a chuté de 68 % après 1986 (Harmon et al., 2007). Une étude similaire réalisée au Michigan sur une période de 24 ans, entre 1989 et 2012, a révélé que les assemblages de coccinelles étaient de plus en plus dominés par des espèces non indigènes, 71 % des coccinelles capturées étant non indigènes (Bahlai et al., 2013). Gardiner et al. (2009) ont noté que deux espèces non indigènes, la coccinelle à sept points et la coccinelle asiatique, représentaient à elles seules jusqu'à 90 % des communautés de coccinelles associées aux champs de soja au Michigan, au Wisconsin et en Iowa. Selon Tumminello et al. (2015), le déclin de la coccinelle à neuf points peut être attribué à l'établissement, à la propagation et à l'augmentation des sous-populations de la coccinelle à sept points. Bien que les causes des déclins observés chez les espèces indigènes, y compris chez la coccinelle à neuf points, à l'échelle de leur aire de répartition demeurent incertaines, ces déclins sont très évidents et bien réels.
En bref, cette coccinelle autrefois commune semble aujourd'hui très rare ou se maintenir sous le seuil de détection dans de nombreuses régions comprises dans son aire de répartition. De nombreuses études ont fait état de déclins continus de l'abondance relative et de contractions d'aires de répartition à l'échelle de l'aire de répartition de la coccinelle à neuf points au Canada et aux États-Unis (Staines et al., 1990; Wheeler et Hoebeke, 1995; Marshall, 1999; Stephans, 2002; Acorn, 2007; Harmon et al., 2007; Hesler et Kieckhefer, 2008; Fothergill et Tindall, 2010; Skinner et Domaine, 2010; Evans et al., 2011; Koch 2011; McCorquodale et al., 2011).
Les fluctuations naturelles des sous-populations de coccinelles sont liées à la dispersion, à la disponibilité des proies, aux conditions climatiques et à la survie hivernale. Toutefois, les coccinelles, dont la coccinelle à neuf points, ne subissent pas de fluctuations extrêmes (Acorn, 2007; Harmon et al., 2007; McCorquodale et al., 2011). Le déclin récent et soutenu de cette espèce à l'échelle de son aire de répartition mondiale et dans de nombreuses régions dispersées donne à croire que le phénomène ne résulte probablement pas d'une fluctuation naturelle.
La coccinelle est une espèce largement répartie dont l'aire de répartition s'étend d'une côte à l'autre aux États-Unis. Comme il s'agit d'une espèce très mobile qui se disperse facilement, il est possible que des sous-populations puissent se disperser et recoloniser des zones où l'espèce a subi un déclin si de l'habitat convenable est disponible. Toutefois, cette espèce a également subi un déclin aux États-Unis, et les raisons de ce déclin demeurent inconnues. Une immigration de source externe est donc jugée peu probable.
La classification des menaces pesant sur la coccinelle à neuf points (Salafsky et al., 2008; Master et al., 2009) a été réalisée à l'aide du système de classification des menaces proposé par l'Union internationale pour la conservation de la nature (UICN) et le Partenariat pour les mesures de conservation (Conservation Measures Partnership, ou CMP) (2006) (IUCN–CMP). Le résultat de l'évaluation des menaces indique un impact global très élevé à élevé (annexe 1). Les menaces sont énumérées ci-après en ordre décroissant d'impact.
Espèces exotiques (non indigènes) envahissantes (8.1)
Il est largement admis que l'introduction accidentelle et intentionnelle d'espèces non indigènes peut avoir des impacts négatifs sur la flore et la faune (New, 1995; Cottrell et Shapiro-Ilan, 2003; Evans, 2004; Snyder et Evans, 2006; Finlayson et al., 2008; Kenis et al., 2008; Kajita et Evans, 2009; Crowder et Snyder, 2010; Smith et Gardiner, 2013; Ugine et Losey, 2014; Tumminello et al., 2015). Au cours du siècle dernier, des prédateurs généralistes d'insectes ont été introduits par inadvertance ou intentionnellement hors de leur aire de répartition indigène à titre d'agents de lutte biologique (Obrycki et Kring, 1998; Evans et al., 2011). Au moins 179 espèces de coccinelles non indigènes ont été introduites en Amérique du Nord seulement, et neuf de ces espèces, dont la coccinelle à sept points et la coccinelle asiatique, se sont établies avec succès au Canada (Gordon, 1985; Gordon et Vandenberg, 1991; Harmon et al., 2007; Evans et al., 2011; McCorquodale et al., 2011; Bousquet et al., 2013). Ces espèces sont encore couramment offertes à titre d'agents de lutte biologique et continuent d'être relâchées.
L'altération des habitats ou les relations interspécifiques avec des espèces non indigènes sont souvent la cause de déclins importants de l'aire de répartition et de l'abondance d'espèces de coccinelles indigènes (New, 1995; Cottrell et Shapiro-Ilan, 2003; Evans, 2004; Snyder et Evans, 2006; Finlayson et al., 2008; Kenis et al., 2008; Kajita et Evans, 2009; Crowder et Snyder, 2010; Smith et Gardiner, 2013; Ugine et Losey, 2014; Tumminello et al., 2015).
L'invasion de l'Amérique du Nord par la coccinelle à sept points et la coccinelle asiatique a été mise en cause dans le déclin généralisé des sous-populations de la coccinelle à neuf points et d'autres espèces de coccinelles indigènes (Wheeler et Hoebeke, 1995; Elliott et al., 1996; Marshall, 1999; Ellis et al., 1999; Brown, 2003; Cottrell et Shapiro-Ilan, 2003; Turnock et al., 2003; Hesler et al., 2004; Acorn, 2007; Harmon et al., 2007; Hesler et Kieckhefer, 2008; Fothergill et Tindall, 2010; Skinner et Domaine, 2010; Evans et al., 2011; Losey et al., 2012; Comont et al., 2013; Turnipseed et al., 2014; Ugine et Losey, 2014; Tumminello et al., 2015). Les interactions négatives résultant de la compétition, de la prédation intraguilde ou d'effets indirects tels que l'introduction de pathogènes constituent le facteur le plus souvent évoqué pour expliquer les déclins des sous-populations d'espèces indigènes (Schaefer et al., 1987; Ehler, 1990; Cottrell et Shapiro-Ilan, 2003; Louda et al., 2003; Evans, 2004; Lucas, 2005; Snyder et Evans, 2006; Lucas et al., 2007; Kenis et al., 2008; Riddick et al., 2009; Evans et al., 2011; Turnipseed et al., 2014; Ugine et Losey, 2014; Tumminello et al., 2015).
Compétition et prédation intraguilde :
L'aire de répartition de la coccinelle à neuf points a rapidement décliné par suite de l'établissement et de la dispersion de la coccinelle à sept points et de la coccinelle asiatique et de la croissance de leurs sous-populations en Amérique du Nord (Turnipseed et al., 2014; Ugine et Losey, 2014; Tumminello et al., 2015). Avant les années 1980, la coccinelle à neuf points était une espèce relativement commune en Amérique du Nord (Gordon, 1985; Tumminello et al., 2015). Il faudra toutefois attendre jusqu'au milieu des années 1990, soit près de 20 ans après l'arrivée et la propagation des espèces de coccinelles non indigènes, pour que le déclin de ses sous-populations soit largement reconnu (Wheeler et Hoebeke, 1995; McCorquodale et al., 2011).
Hodek et Michaud (2008) ont fait valoir que la coccinelle à sept points est un compétiteur efficace en présence de conditions très diversifiées. Sa capacité de concurrencer avec succès pour sa nourriture et de s'accoupler et de pondre ses œufs en présence de conditions diversifiées lui permet de présenter un bon rendement général plutôt que d'être l'espèce la mieux adaptée en présence de conditions bien particulières (Hodek et Michaud, 2008; McCorquodale et al., 2011). Losey et al. (2012) ont montré que la compétition par exploitation avec la coccinelle à sept points entraîne une diminution de la disponibilité des proies et une réduction de la taille corporelle chez la coccinelle à neuf points. Autre appui à l'hypothèse d'une compétition par exploitation selon laquelle toutes les espèces concurrentes ont accès à une ressource limitée, Hoki et al., (2014) ont montré que la coccinelle à sept points est plus vorace, présente un taux d'attaque des pucerons plus élevé et consacre moins de temps à la manipulation de ses proies (pucerons) que la coccinelle à neuf points. Tumminello et al., (2015) ont également étudié l'impact de la compétition par exploitation et de la prédation intraguilde et ont conclu que la coccinelle à sept points est probablement responsable du déplacement de la coccinelle à neuf points hors de son aire de répartition d'origine parce qu'elle se développe plus rapidement et est plus grande que cette dernière, présente des taux d'attaque plus élevés et montre un fort taux de prédation intraguilde ciblant cette dernière.
D'autres études ont également montré que les larves de la coccinelle à neuf points ont plus de chance d'atteindre le stade adulte lorsqu'elles sont élevées en compagnie de larves d'autres espèces indigènes plutôt qu'en compagnie de larves de la coccinelle à sept points, les taux de prédation des œufs ou des larves par les espèces indigènes étant faibles (Turnipseed et al., 2014). Des résultats similaires ont été observés chez d'autres espèces de coccinelles indigènes et introduites (Obrycki et al., 1998; Michaud, 2002; Sato et al., 2004; Snyder et al., 2004; Lucas et al., 2007; Pell et al., 2008; Gardiner et al., 2011; Hodek et al., 2012; Smith et Gardiner 2013). La prédation intraguilde joue également un rôle important en empêchant la recolonisation par les espèces indigènes, et les femelles évitent également les sites de ponte où des prédateurs intraguilde sont présents (Ruzicka, 1997; Hodek et al., 2012). L'établissement de la coccinelle à sept points dans les paysages agricoles a également entraîné des déclins documentés d'espèces de coccinelles indigènes et de la densité des pucerons (Alyokhin et Sewell, 2004; Evans, 2004).
En dépit des déclins documentés des sous-populations d'espèces de coccinelles indigènes au Canada (voir par exemple Turnock et al., 2003) et de l'introduction et de la dispersion d'espèces de coccinelles non indigènes en Amérique du Nord (voir par exemple Wheeler et Stoops, 1996; Lucas et al., 2007), les liens entre les espèces non indigènes et les causes des déclins ne sont pas claires. Par exemple, Acorn (2007) et Harmon et al., (2007) ont fait valoir qu'on ne dispose pas des preuves directes nécessaires pour affirmer que la compétition ou d'autres interactions avec les espèces non indigènes récemment introduites sont responsables des déclins observés chez les espèces indigènes. Même si les tendances observées semblent conforter l'impression selon laquelle la coccinelle à sept points et la coccinelle asiatique pourraient avoir un impact négatif par compétition par exploitation et prédation intraguilde sur la coccinelle à neuf points, d'autres mécanismes incluant l'introduction de parasitoïdes et de pathogènes pourraient également être en cause. (Losey et al., 2012).
Parasites, parasitoïdes, pathogènes et champignons :
Les espèces de coccinelles non indigènes peuvent également avoir un impact indirect sur les espèces indigènes en favorisant l'introduction et la transmission de nouveaux ennemis naturels tels que des parasitoïdes, des parasites et des pathogènes (Bjornson, 2008). Les coccinelles servent d'hôtes à divers parasitoïdes (c.-à-d. guêpe de la famille des Braconidés), acariens parasites (c.-à-d. Coccipolipus hippodamiae), nématodes, protozoaires, de champignons pathogènes (c.-à-d. Beauveria bassiana), microsporidies (Nosématidés), et de nombreuses bactéries peuvent avoir un impact négatif sur la valeur adaptative des coccinelles et compromettre leur survie hivernale (Cali et Briggs, 1967; Hurst et al., 1995; Ceryngier et Hodek, 1996; Barron et Wilson, 1998; Webberley et Hurst, 2002; Cottrell et Shapiro-Ilan, 2003; Webberley et al., 2004; Bjornson, 2008; Roy et Cottrell, 2008; Riddick et al., 2009; Bjornson et al., 2011). Bien que les effets de ces ennemis naturels sur la coccinelle à neuf points soient incertains, il est établi que les espèces indigènes sont souvent plus sensibles aux pathogènes exotiques que les espèces non indigènes (Cottrell et Shapiro-Ilan, 2003). Obrycki (1989) a montré que les espèces de coccinelles indigènes sont plus sensibles au Braconidé parasitoïde que les espèces non indigènes telles que la coccinelle asiatique. Cottrell et Shapiro-Ilan (2003) ont également indiqué que les coccinelles indigènes sont plus sensibles que la coccinelle asiatique au champignon pathogène Beauveria bassiana. Une plus grande sensibilité des espèces indigènes aux pathogènes exotiques pourrait donc conférer un avantage intraguilde aux coccinelles non indigènes et pourrait avoir contribué au déclin de la coccinelle à neuf points.
Effluents agricoles et sylvicoles (9.3)
Bien que les coccinelles affichent une plus grande tolérance aux pesticides que leurs proies (Gesraha, 2007), la pollution causée par les produits agrochimiques utilisés pour la lutte contre les insectes ravageurs peut avoir un impact direct sur les coccinelles non ciblées lorsque celles-ci entrent en contact avec ces produits ou avec leurs résidus, inhalent des substances volatiles ou consomment des proies, du nectar ou du pollen contaminés par des insecticides (Smith et Krischik, 1999; Youn et al., 2003; Singh et al., 2004; Moser et al., 2008; Moser et Obrycki 2009; Eisenback et al., 2010). Ces produits peuvent également avoir un effet indirect sur les populations de coccinelles indigènes en éliminant leurs sources de nourriture (Hodek et al., 2012; Bahlai et al., 2015). La zoophytophagie, comportement omnivore observé chez les espèces principalement prédatrices qui consomment également des matières végétales (pollen, nectar, tissus foliaires), accroît la fécondité et réduit le temps de développement (Coll, 1998; Patt et al., 2003; Moser et Obrycki, 2009). Ce comportement peut toutefois avoir des effets néfastes si les matières végétales ingérées sont protégées chimiquement par des insecticides (Moser et Obrycki, 2009). La sensibilité des coccinelles aux insecticides varie selon les espèces et le type de pesticide, les manifestations d'une exposition à des concentrations non létales d'insecticides allant d'effets létaux aigus à une réduction de la fécondité induite par des effets sur le comportement ou la reproduction (Theiling et Croft, 1988). De nombreux insectes prédateurs exposés à plus d'un composé présentent des effets néfastes synergiques, même si les composés incriminés sont individuellement inoffensifs (Petersen, 1993).
Dans les paysages urbains et agricoles, les sous-populations de coccinelles peuvent être menacées par de nombreux pesticides, en particulier des néonicotinoïdes, des régulateurs de la croissance des insectes et des pyréthroïdes à large spectre, lesquels sont généralement plus nocifs pour les coccinelles que les organophosphates (Kumar et Bhatt, 2002; Moser et Obrycki, 2009). Certains régulateurs de croissance des insectes tels que la buprofézine et le pyriproxyfène n'ont habituellement pas d'effet toxique aigu chez les coccinelles, mais ils peuvent compromettre leur développement et réduire leur fécondité (Olszak et al., 1994). Les néonicotinoïdes forment une classe de pesticides systémiques qui circulent et s'accumulent dans les tissus des plantes, y compris le pollen et le nectar. Ces composés sont très efficaces contre les phytoravageurs, en particulier les pucerons, mais il a été démontré qu'ils ont des effets néfastes pour d'autres insectes à des concentrations de l'ordre de quelques parties par milliard (ppb) (Smith et Krischik, 1999; Marletto et al., 2003). Les néonicotinoïdes peuvent également être utilisés pour le traitement des semences avant la plantation pour protéger les semis contre les ravageurs des feuilles et des racines en début de saison. Dans une étude, 72 % des larves de coccinelles asiatiques (Harmonia axyridis) exposées à des semis traités avec des néonicotinoïdes ont présenté des symptômes neurotoxiques (tremblements, paralysie et perte de coordination), et seulement 7 % d'entre elles se sont rétablies (Moser et Obrycki, 2009). L'utilisation de néonicotinoïdes peut donc avoir des effets néfastes chez les espèces non ciblées, en particulier chez les espèces zoophytophages.
Autres modifications de l'écosystème (7.3)
L'abandon des terres gérées et des exploitations agricoles dans l'est du Canada pourrait avoir contribué au déclin de la coccinelle à neuf points (Bucknell et Pearson, 2007; Harmon et al., 2007). L'expansion urbaine et l'abandon des terres agricoles peuvent entraîner une réduction de l'abondance des sources de nourriture pour cette espèce (Harmon et al., 2007). Bien que ces changements à grande échelle touchant l'habitat et la disponibilité de nourriture puissent être évoqués en guise d'explication au déclin de l'espèce, aucune donnée ne permet d'établir un lien de cause à effet entre les changements qui se sont produits à l'échelle du paysage et la réduction de la densité des coccinelles (Elliott et Kieckheffer, 1990; Elliott et al., 1999; Harmon et al., 2007).
La perte d'habitat et la réduction de la qualité de l'habitat se poursuivent de façon soutenue à l'échelle de l'aire de répartition de l'espèce (Federal, Provincial and Territorial Governments of Canada, 2010; Javorek et Grant, 2011). L'homogénéisation des paysages agriculturaux et les changements touchant les pratiques agricoles, en particulier la dépendance accrue à l'égard des engrais et des pesticides, jouent probablement aussi un rôle dans le déclin des espèces indigènes à l'échelle locale (Wheeler et Hoebeke, 1995; Bianchi et al., 2007; Evans et al., 2011). Cette question a déjà été abordée précédemment dans la section traitant de la pollution (Menace 9).
La plantation de plantes génétiquement modifiées (GM) résistantes aux insectes (p. ex. maïs GM exprimant les toxines de Bacillus thuringiensis (Bt)) a été considérée comme un risque potentiel pour les coccinelles parce que la toxine de la bactérie est présente dans le pollen (Harwood et al., 2007) mais pas chez les pucerons (Hodek et al., 2012). Même si la majorité des études ont indiqué que la consommation de pollen de maïs Bt ne semble avoir aucun effet sur les paramètres de la valeur adaptative chez les coccinelles (Duan et al., 2002; Lundgren et Wiedenmann, 2002; Porcar et al., 2010), d'autres ont montré qu'elle pouvait réduire la fécondité et retarder le développement (Moser et al., 2008).
La perte d'habitat et la réduction de la qualité de l'habitat occasionnées par l'intensification du développement résidentiel et commercial pourraient avoir joué un rôle dans le déclin de cette espèce à l'échelle locale. Les espaces verts et les jardins locaux dans les secteurs urbanisés de taille plus modeste pourraient toutefois continuer de procurer de l'habitat à la coccinelle à neuf points.
Il est impossible à ce stade-ci de calculer le nombre de localités abritant l'espèce. Le terme « localité » désigne une zone écologiquement ou géographiquement distincte à l'intérieur de laquelle un seul événement menaçant peut toucher rapidement tous les individus du taxon considéré. La coccinelle à neuf points présente une aire de répartition très vaste et est très mobile, et les menaces auxquelles elle est exposée demeurent incertaines. En l'absence d'indications très claires sur les menaces qui pèsent sur l'espèce à l'échelle de son aire de répartition, le terme « localité » ne peut être utilisé, et les sous-critères qui réfèrent au nombre de localités ne sont pas satisfaits.
En ce qui a trait au nombre de sites, treize mentions de l'espèce ont été enregistrées au Canada au cours des dix dernières années dans neuf sites différents, soit deux sites à Cranbrook, un site à Kamloops, un site à Osoyoos et deux sites au lac Williams, en Colombie-Britannique; un site à Calgary, un site à Cardston, trois sites à Medicine Hat et un site à Steveville, en Alberta; et un site à Mont-Saint-Hilaire, au Québec. Compte tenu de la vaste aire de répartition de la coccinelle à neuf points et de sa grande capacité de dispersion, il est raisonnable de penser qu'elle est présente dans d'autres sites à l'échelle de son aire de répartition.
Aucune loi fédérale ou provinciale ne protège la coccinelle à neuf points, n'atténue les menaces qui pèsent sur l'ensemble des coccinelles ou ne protège les sites de nidification ou l'habitat de cette espèce.
Au Québec, cette espèce n'est actuellement pas désignée menacée ou vulnérable (LEMV, 2015), mais elle est toutefois inscrite sur la Liste des espèces menacées ou vulnérables (LEMV, 2015). Elle est par conséquent considérée comme une espèce en péril et bénéficie d'une protection en vertu des articles 22 et 31.1 de la Loi sur la qualité de l'environnement (LRQ, c. Q-2).
Cette espèce est classée G2 (en péril) à l'échelle mondiale, mais elle est non classée à l'échelle nationale au Canada et aux États-Unis (NatureServe, 2014). Elle est également non classée dans la plupart des États, provinces et territoires où elle se rencontre, mais elle est considérée comme possiblement disparue (possibly extirpated) en Alberta (SNR), en Ontario (SH), au Connecticut (SH) et en Floride (SH).
Cotes de conservation nationales pour le Canada (Conseil canadien pour la conservation des espèces en péril [CESCC, 2010]) : Sensible au Canada et, à l'échelle infranationale, dans les Territoires du Nord-Ouest, en Colombie-Britannique, en Alberta et en Saskatchewan; possiblement en péril au Manitoba; disparue de l'Ontario et du Québec.
Liste rouge de l'UICN (2013) : Aucune cote de conservation.
Cette espèce n'a pas été évaluée ni classée sous le régime de l'Endangered Species Act des États-Unis.
Compte tenu de la vaste aire de répartition et de la large niche écologique de la coccinelle à neuf points au Canada, plusieurs zones d'habitat convenable se trouvent sur des terres privées en milieu agricole et urbain ainsi que sur des terres publiques et dans des aires protégées.
L'aire de répartition canadienne de la coccinelle à neuf points chevauche un grand nombre de parcs provinciaux et nationaux et d'aires protégées.
Le rédacteur remercie Jennifer Heron d'avoir supervisé l'élaboration du présent rapport, de même qu'Angèle Cyr (Secrétariat du COSEPAC). Il tient également à souligner la contribution des personnes suivantes : David McCorquodale (Université du Cap-Breton); John Acorn (Université de l'Alberta); Isabelle Gauthier (Ministère des Forêts, de la Faune et des Parcs); John Losey (Université Cornell); Cory Sheffield (Royal Saskatchewan Museum); Suzanne Carriere, Danny Allaire et Nicholas Larter (gouvernement des Territoires du Nord-Ouest); Mary Sabine (gouvernement du Nouveau-Brunswick); Barb Sharanowski (Université du Manitoba); Gilles Boiteau (Agriculture et Agroalimentaire Canada); Ken Millard et Lisa Ott (Galiano Conservancy Association); Claudia Copley (Royal British Columbia Museum); Darren Copley et Rob Cannings (Royal British Columbia Museum), Syd Cannings (Service canadien de la faune), ainsi que Gary Anweiler, Heather Leibel, Mattias Buck, Heidi Gartner, Robb Bennett, Erica McClaren, Berry Wijdeven, Mark Weston, Lynn Westcott, Sandy Cessellie, Bill Ramey, Bev Ramey, Michael Dunn, Geoff Lynch, Nick Burdock, Jeevan Sandu, Sylvia Latay, Rick Howie, Lea Gelling, Kathy Coot, Tom Foot, Karen Durovich, Sara Kalnay-Watson, Al Harris, Rob Foster, Vincent Bereczki, Bruce Bennett et les responsables du Lost Ladybug Project.
Abassi, S., M.A. Birkett, J. Pettersson, J.A. Pickett, L.J. Wadhams et C.M. Woodcock. 2001. Response of the ladybird parasitoid Dinocampus coccinellae to toxic alkaloids from the seven-spot ladybird, Coccinella septempunctata. Journal of Chemical Ecology 27:33-43.
Acorn, J. 2007. Ladybugs of Alberta: Finding the spots and connecting the dots. The University of Alberta Press, Edmonton, Alberta.
Alyokhin, A. et G. Sewell. 2004. Changes in a lady beetle community following the establishment of three alien species. Biological Invasions 6:463-471.
Bahlai, C.A., M. Colunga-Garcia, S.H. Gage et D.A. Landis. 2013. Long-term functional dynamics of an aphidophagous coccinellid community remain unchanged despite repeated invasions. PLoS ONE 8:1-11.
Bahlai, C.A., W. van der Werf, M. O'Neal, L. Hemerik et D.A. Landis 2015. Shifts in dynamic regime of an invasive lady beetle are linked to the invasion and insecticidal management of its prey. Ecological Applications 25:1807-1818.
Barron, A. et K. Wilson. 1998. Overwintering survival in the seven spot ladybird, Coccinella septempunctata (Coleoptera: Coccinellidae). European Journal of Entomology 95:639-642.
Belicek, J. 1976. Coccinellidae of western Canada and Alaska with analyses of the transmontane zoogeographic relationships between the fauna of British Columbia and Alberta (Insecta: Coleoptera: Coccinellidae). Quaestiones Entomologicae 12:283-409.
Bianchi, F.J., A., Honĕk et W. van der Werf. 2007. Changes in agricultural land use can explain population decline in a ladybeetle species in the Czech Republic: evidence from a process-based spatially explicit model. Landscape Ecology 22:1541-1554.
Bjornson, S. 2008. Natural enemies of the convergent lady beetle, Hippodamia convergens Guérin-Méneville: their inadvertent importation and potential significance for augmentative biological control. Biological Control 44:305-311.
Bjornson, S., J. Le, T. Saito et H. Wang. 2011. Ultrastructure and molecular characterization of a microsporidium, Tubulinosema hippodamiae, from the convergent lady beetle, Hippodamia convergens Guérin-Méneville. Journal of Invertebrate Pathology 106:280-288.
Bousquet, Y., P. Bouchard, A.E. Davies et D.S. Sikes. 2013. Checklist of beetles (Coleoptera) of Canada and Alaska. Second edition. ZooKeys 360:1-44.
Brown, M.W. 2003. Intraguild responses of aphid predators on apple to the invasion of an exotic species, Harmonia axyridis. BioControl 48:141-153.
Brown, W.J. 1940. Notes on the American distribution of some species of Coleoptera common to the European and North American continents. The Canadian Entomologist 72:65-78.
Brown, W.J. 1962. A revision of the forms of Coccinella L., occurring in America north of Mexico (Coleoptera: Coccinellidae). The Canadian Entomologist 94:785-808.
Brown, W.J. et R. de Ruette. 1962. An annotated list of the Hippodamiini of Northern America, with a key to the genera (Coleoptera: Coccinellidae). The Canadian Entomologist 94:643-652.
Bucknell, D. et C.J. Pearson. 2007. A spatial analysis of land-use change and agriculture in eastern Canada. International Journal of Agricultural Sustainability 4:22-38.
Cali, A. et J.D. Briggs. 1967. The biology and life history of Nosema tracheophila sp. n. (Protozoa: Cnidospora: Microsporidea) found in Coccinella septempunctata Linnaeus (Coleoptera: Coccinellidae). Journal of Invertebrate Pathology 9:515-522.
Canadian Endangered Species Conservation Council (CESCC). 2011. Wild Species 2010: The General Status of Species in Canada (PDF; 4.8 MB). National General Status Working Group: 302 pp. (Également disponible en français : Conseil canadien pour la conservation des espèces en péril (CCCEP). 2011. Espèces sauvages 2010 – La situation générale des espèces au Canada (PDF; 4.9 Mo). Groupe de travail national sur la situation générale. 323 p.
Cardinale, B.J., J.J. Weis, A.E. Forbes, K.J. Tilmon et A.R. Ives. 2006. Biodiversity as both a cause and consequence of resource availability: a study of reciprocal causality in a predator-prey system. Journal of Animal Ecology 75:497-505.
Ceryngier, P. et I. Hodek. 1996. Enemies of the Coccinellidae. Pp. 319-350, in Hodek, I. et A. Honěk (eds). Ecology of Coccinellidae. Kluwer Academic, Dordecht.
Coll, M. et M. Guershon. 2002. Omnivory in terrestrial arthropods: mixing plant and prey diets. Annual Review of Entomology 47:267-297.
Comont, R.F., H.E. Roy, R. Harrington, C.R. Shortall et B.V. Purse. 2013. Ecological correlates of local extinction and colonisation in the British ladybird beetles (Coleoptera: Coccinellidae). Biological Invasions 16:1805-1817.
Cottrell, T.E. et D.I. Shapiro-Ilan. 2003. Susceptibility of a native and an exotic lady beetle (Coleoptera: Coccinellidae) to Beauveria bassiana. Journal of Invertebrate Pathology 84:137-144.
Crowder, D.W. et W.E. Snyder. 2010. Eating their way to the top? Mechanisms underlying the success of invasive insect generalist predators. Biological Invasions 12:2857-2876.
Dobzhansky, T. 1935. A list of Coccinellidae of British Columbia. Journal of the New York Entomological Society 43:331-336.
Duan, J.J., G. Head, M.J. McKee et al. 2002. Evaluation of dietary effects of transgenic corn pollen expressing Cry3Bb1 protein on a non-target ladybird beetle, Coleomegilla maculata. Entomologia Experimentalis et Applicata 104:271-280.
Ehler, L.E. 1990. Introduction strategies in biological control of insects. Pp 111-134, in Mackauer, M., L.E. Ehler, et J. Roland (ed.). Critical issues in biological control. Intercept Ltd, Andover.
Eisenback, B.M., S.M. Salom, L.T. Kok et A.F. Lagalante. 2010. Lethal and sublethal effects of imidacloprid on hemlock woolly adelgid (Hemiptera: Adelgidae) and two introduced predator species. Journal of Economic Entomology 103:1222-1234.
Elith, J., C.H. Graham, R.P. Anderson, M. Dudik, S. Ferrier et al. 2006. Novel methods improve prediction of species' distributions from occurrence data. Ecography 29:129-151.
Elliott, N.C. et R.W. Kieckheffer. 1990. A 13-year study of the aphidophagous insects of alfalfa. Prairie Naturalist 22:87-96.
Elliott, N. 2000. Adult coccinellid activity and predation on aphids in spring cereals. Biological Control 17:218-226.
Elliott, N.C., R.W. Kieckhefer, J.H. Lee et B.W. French. 1999. Influence of within-field and landscape factors on aphid predator populations in wheat. Landscape Ecology 14:239-252.
Elliott, N.C., R.W. Kieckhefer et W.C. Kauffman. 1996. Effects of an invading coccinellid on native coccinellids in an agricultural landscape. Oecologia 105:537-544.
Ellis, D.R., D.E. Prokrym et R.G. Adams. 1999. Exotic lady beetle survey in northeastern United States: Hippodamia variegata and Propylea quatuordecimpunctata (Coleoptera: Coccinellidae). Entomological News 111:73-84.
Evans, E.W. et T.R. Toler. 2007. Aggregation of polyphagous predators in response to multiple prey: ladybirds (Coleoptera: Coccinellidae) foraging in alfalfa. Population Ecology 49:29-36.
Evans, E.W., A.O. Soares et H. Yasuda. 2011. Invasions by ladybugs, ladybirds, and other predatory beetles. BioControl 56:597-611.
Evans, E.W. 2004. Habitat displacement of North American ladybirds by an introduced species. Ecology 85:637-647.
Federal, Provincial and Territorial Governments of Canada. 2010. Canadian Biodiversity: Ecosystem Status and Trends 2010. Canadian Councils of Resource Ministers. Ottawa, ON. vi + 142 pp. (Également disponible en français : Les gouvernements fédéral, provinciaux et territoriaux du Canada. 2010. Biodiversité canadienne : état et tendances des écosystèmes en 2010. Conseils canadiens des ministres des ressources. Ottawa (Ont.) vi + 148 p.).
Finlayson, C.J., K.M. Landry et A.V. Alyokhin. 2008. Abundance of native and non-native lady beetles (Coleoptera: Coccinellidae) in different habitats in Maine. Annals of the Entomological Society of America 101:1078-1087.
Fortin, M.J., T.H. Keitt, B.A. Maurer, M.L. Taper, D.M. Kaufman et T.M. Blackburn. 2005. Species' geographic ranges and distributional limits: pattern analysis and statistical issues. Oikos 108:7-17.
Fothergill, K. et K.V. Tindall. 2010. Lady beetle (Coleoptera: Coccinellidae: Coccinellinae) occurrences in southeastern Missouri agricultural systems: differences between 1966 and present. The Coleopterists Bulletin 64:379-382.
Gardiner, M.M., L.L. Allee, P.M.J. Brown, J.E. Losey, H.E. Roy et al. 2012. Lessons from lady beetles: accuracy of monitoring data from US and UK citizen-science programs. Frontiers in Ecology and the Environment: Online Preprint. doi:10.1890/110185.
Gardiner, M.M., Landis, D.A., Gratton, C., Schmidt, N., O'Neal, M., Mueller, E., Chacon, J., Heimpel, G.E. et DiFonzo, C.D. 2009. Landscape composition influences patterns of native and exotic lady beetle abundance. Diversity and Distributions 15:554–564.
Gardiner, M.M., M.E. O'Neal et D.A. Landis. 2011. Intraguild predation and native lady beetle decline. PloS ONE 6:e23576.
Gesraha, M.A. 2007. Impact of some insecticides on the Coccinellid predator, Coccinella undecimpunctata L. and its aphid prey, Brevicoryne brassicae L. Egyptian Journal of Biological Pest Control 17:65-69.
Giorgi, A. et J. Vandenberg. 2009. Coccinellidae. Lady beetles, ladybird beetles, ladybugs. Version 09, Novembre 2009 (en construction) in The Tree of Life Web Project.
Gordon, R.D. 1985. The Coccinellidae (Coleoptera) of America north of Mexico. Journal of New York Entomological Society 95:1-912.
Gordon, R.D. et N. Vandenberg. 1991. Field guide to recently introduced species of Coccinellidae (Coleoptera) in North America, with revised key to North America genera of Coccinellini. Proceedings of the Entomological Society of Washington 93:845-864.
Hagen, K.S. 1962. Biology and ecology of predaceous Coccinellidae. Annual Review of Entomology 7:289-326.
Harmon, J.P., E. Stephens et J. Losey. 2007. The decline of native coccinellids (Coleoptera: Coccinellidae) in the United States and Canada. Journal of Insect Conservation 11:85-94.
Harwood, J.D., R.A. Samson et J.J. Obrycki. 2007. Temporal detection of Cry1Ab-endotoxins in coccinellid predators from fields of Bacillus thuringiensis corn. Bulletin of Entomological Research 97:643-648.
Hesler, L.S. 2009. An annotated checklist of the lady beetles (Coleoptera: Coccinellidae) of Iowa, U.S.A. Insecta Mundi 91:1-10.
Hesler, L.S. et R.W. Kieckhefer. 2008. Status of exotic and previously common native coccinellids (Coleoptera) in south Dakota landscapes. Journal of the Kansas Entomological Society 81:29-49.
Hesler, L.S., G. McNickle, M. Catangui, J. Losey, E. Beckendorf, L. Stellwag, D. Brandt et P. Bartlett. 2012. Method for continuously rearing coccinella lady beetles (Coleoptera: Coccinellidae). The Open Entomology Journal 6:42-48.
Hesler, L.S., R.W. Kieckhefer et M.A. Catangui. 2004. Surveys and field observations of Harmonia axyridis and other Coccinellidae (Coleoptera) in eastern and central South Dakota. Transactions of the American Entomological Society 130:113-133.
Hodek I, H.F. van Emden et A. Honěk. 2012. Ecology and behaviour of the ladybird beetles (Coccinellidae). Wiley-Blackwell. Kindle Edition.
Hodek, I. et A. Honěk. 1996. Ecology of Coccinellidae. Kluwer Academic Publishers, Dordecht, The Netherlands.
Hodek, I. et J.P. Michaud. 2008. Why is Coccinella septempunctata so successful? (A point-of-view). European Journal of Entomology 105:1-12.
Hoki, E., J.E. Losey et T.A. Ugine. 2014. Comparing the consumptive and non-consumptive effects of a native and introduced lady beetle on pea aphids (Acyrthosiphon pisum). Biological Control 70:78-84.
Hurst, G.D.D., R.G. Sharpe, A.H. Broomfield et al. 1995. Sexually transmitted disease in a promiscuous insect, Adalia bipunctata. Ecological Entomology 20:230-236.
ITIS (Integrated Taxonomic Information System). 2015. Extrait [novembre 2015] de la base de données en ligne (Disponible en anglais seulement).
Ives, A.R., P. Kareiva et R. Perry. 1993. Response of a predator to variation in prey density at three hierarchical scales lady beetles feeding on aphids. Ecology 74:1929-1938.
Ives, P.M. 1981. Estimation of coccinellid numbers and movement in the field. The Canadian Entomologist 113:981-997.
Javorek, S.K. et M.C. Grant. 2011. Trends in wildlife habitat capacity on agricultural land in Canada, 1986-2006. Canadian Biodiversity: Ecosystem Status and Trends 2010, Technical Thematic Report No. 14. Canadian Councils of Resource Ministers. Ottawa, ON. vi + 46 pp. (Également disponible en français : Javorek, S.K. et M.C. Grant., 2011. Tendances de la capacité d'habitat faunique des terres agricoles du Canada, de 1986 à 2006. Biodiversité canadienne : état et tendances des écosystèmes en 2010, Rapport technique thématique nº 14. Conseils canadiens des ministres des ressources. Ottawa, (Ont.). vi + 51 p.).
Jeffries, D.L., J. Chapman, H.E. Roy, S. Humphries, R. Harrington, P.M.J. Brown et L.J. Handley. 2013. Characteristics and drivers of high-altitude ladybird flight: insights from vertical-looking entomological radar. PloS ONE 8:e82278.
Kajita, Y. et E.W. Evans. 2010. Alfalfa fields promote high reproductive rate of an invasive predatory lady beetle. Biological Invasions 12:2293-2302.
Kenis, M., M.A. Auger-Rozenberg, A. Roques, L. Timms, C. Pere, M.J.W. Cock, J. Settele, S. Augustin et C. Lopez-Vaamonde. 2008. Ecological effects of invasive alien insects. Biological Invasions 11:21-45.
Koch, J.B. et J.P Strange. 2009. Constructing a species database and historic range maps for North American bumblebees (Bombus sensu stricto Latreille) to inform conservation decisions. Uludag Bee Journal 9:97-108.
Koch, R.L. 2011. Recent detections of a rare native lady beetle, Coccinella novemnotata (Coleoptera : Coccinellidae), in Minnesota. Great Lakes Entomologist 44:196-199.
Krafsur, E.S., J.J. Obrycki et J.D. Harwood. 2005. Comparative genetic studies of native and introduced Coccinellidae in North America. European Journal of Entomology 102:469-474.
Krivan, K. 2008. Dispersal dynamics: Distribution of lady beetles (Coleoptera: Coccinellidae). European Journal of Entomology 105:405-409.
Kumar, S. et R.I. Bhatt. 2002. Pyrethroid-induced resurgence of sucking pests in the mango ecosystem. Journal of Applied Zoological Research 13:107-111.
Larochelle, A. 1979. Les Coléoptères Coccinellidae du Québec. Cordulia (Supplement) 10:1-111.
Laurent, P., J.C. Braekman, and D. Daloze. 2005. Insect chemical defense. Topics in Current Chemistry 240:167-229.
LEMV (Liste des espèces susceptibles d'être désignées menacées ou vulnérables). 2015. (consulté en novembre 2015).
Losey, J., J. Perlman et E.R. Hoebeke. 2007. Citizen scientist rediscovers rare Nine-spotted Lady Beetle, Coccinella novemnotata, in eastern North America. Journal of Insect Conservation 11:415-417.
Losey, J., J. Perlman, J. Kopco, S. Ramsey, L. Hesler, E. Evans, L. Allee et R. Smyth. 2012. Potential causes and consequences of decreased body size in field populations of Coccinella novemnotata. Biological Control 61:98-103.
Lost Lady Bug Project. 2016. (Disponible en anglais seulement). Consulté le 17 mai 2016.
Louda, S.M., R.W. Pemberton, M.T. Johnson et P.A. Follett. 2003. Non-target effects - the achilles' heel of biological control? Retrospective analyses to reduce risk associated with biocontrol introductions. Annual Review of Entomology 48:365-396.
Lucas, E. 2005. Intraguild predation among aphidophagous predators. European Journal of Entomology 102:351-364.
Lucas, E., C. Vincent, G. Labrie, G. Chouinard, F. Fournier, F. Pelletier, N.J. Bostanian, D. Coderre, M.P. Mignault et P. Lafountaine. 2007. The multicolored Asian ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae) in Québec agroecosystems ten years after its arrival. European Journal of Entomology 104:737-743.
Lundgren, J. G. et R.N. Wiedenmann. 2002. Coleopteran-specific Cry3Bb toxin from transgenic corn pollen does not affect the fitness of a nontarget species, Coleomegilla maculata DeGeer (Coleoptera, Coccinellidae). Environmental Entomology 31:1213-1218.
Majka, C.G. et D.B. McCorquodale. 2006. The Coccinellidae (Coleoptera) of the maritime provinces of Canada: new records, biogeographical notes, and conservation concerns. Zootaxa 1154:49-68.
Majka, C.G. et D.B. McCorquodale. 2010. Ladybird beetles (Coleoptera: Coccinellidae) of the Atlantic Maritime Ecozone. Chapter 21. Pp. 439-452. in McAlpine, D.F. et I.M. Smith. (eds). Assessment of species diversity in the Atlantic Maritime Ecozone. NRC Research Press, Ottawa, Canada.
Marletto, F., A. Patetta et A. Manino 2003. Laboratory assessment of pesticide toxicity to bumble bees. Bulletin of Insectology 56:155-158.
Marples, N.M., P.M. Brakefield et R.J. Cowie. 1989. Differences between the 7-spot and 2-spot ladybird beetles (Coccinellidae) in their toxic effects on a bird predator. Ecological Entomology 14:79-84.
Marriott, S.M, D.J. Giberson et D.B. McCorquodale. 2009. Changes in the status and geographic ranges of Canadian lady beetles (Coccinellidae) and the selection of candidates for risk assessment. Part I Foundation Report. Report to COSEWIC Arthropods Species Specialist Committee. 53 pp.
Marshall, S. 1999. Alien invasions, Ontario's ever changing bug landscape. Seasons. Spring 1999:26-29.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay et A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk. NatureServe, Arlington, VA.
McCorquodale, D.B., D.J. Giberson et S.M. Marriott. 2011. Changes in the status and geographic ranges of Canadian lady beetles (Coleoptera: Coccinellidae: Coccinellinae) and the selection of candidate species for risk assessment. Part 3: Final Report. Committee on the Status of Endangered Wildlife in Canada, Ottawa.
McMullen, R.D. 1967. The effects of photoperiod, temperature and food supply on rate of development and diapause in Coccinella novemnotata. The Canadian Entomologist 99:578-586.
Michaud, J.P. 2002. Invasion of the Florida citrus ecosystem by Harmonia axyridis (Coleoptera: Coccinellidae) and asymmetric competition with a native species, Cycloneda sanguinea. Environmental Entomology 31:827-835.
Moser, S.E., J.D. Harwood et J.J. Obrycki. 2008. Larval feeding on Bt-hybrid and non-Bt corn seedlings by Harmonia axyridis (Coleoptera, Coccinellidae) and Coleomegilla maculata (Coleoptera, Cocinellidae). Environmental Entomology 37:525-533.
Moser, S.E. et J.J. Obrycki. 2009. Non-target effects of neonicotinoid seed treatments; mortality of coccinellid larvae related to zoophytophagy. Biological Control 51:487-492.
Natureserve. 2014. Natureserve Explorer: An online encyclopedia of life [application Web]. Version 7.1. Natureserve, Arlington, Virginia. (Disponible en anglais seulement, consulté le 15 décembre 2014).
Nentwig, W. 1983. The prey of web-building spiders compared with feeding experiments (Araneae: Aranelidae, Linyphiidae, Pholcidae, Agelenidae). Oecologia 56:132-139.
New, T.R. 1995. Introduction to Invertebrate Conservation Biology. Oxford University Press, New York. 194 pp.
Obrycki, J.J. 1989. Parasitization of native and exotic coccinellids by Dinocampus coccinellae (Schrank) (Hymenoptera: Braconidae). Journal of Kansas Entomological Society 62:211-218.
Obrycki, J.J., K.L. Giles et A.M. Ormord. 1998. Interactions between an introduced and indigenous coccinellid species at different prey densities. Oecologia 117:279-285.
Obrycki, J.J. et T.J. Kring. 1998. Predaceous Coccinellidae in biological control. Annual Review of Entomology 43:295-321.
Olszak, R.W., B. Pawlik et R.Z. Zajac. 1994. The influence of some insect growth-regulators on mortality and fecundity of the aphidophagous coccinellids Adalia bipunctata L. and Coccinella septempunctata L. (Col-Coccinellidae). Journal of Applied Entomology 117:58-63.
Omkar et S. Srivastava, 2002. The reproductive behaviour of an aphidophagous ladybeetle, Coccinella septempunctata (Coleoptera: Coccinellidae). European Journal of Entomology 99:465-470.
Osawa, N. 2000. Population field studies on the aphidophagous ladybird beetle Harmonia axyridis (Coleoptera:Coccinellidae): resource tracking and population characteristics. Population Ecology 42:115-127.
Patt, J.M., S.C. Wainright, G.C. Hamilton, D. Whittinghill, K. Bosley, J. Dietrick et J.H. Lashomb. 2003. Assimilation of carbon and nitrogen from pollen and nectar by a predaceous larva and its effects on growth and development. Ecological Entomology 28:717-728.
Pell, J.K., J. Baverstock, H.E. Roy, R.L. Ware et M.E.N. Majerus. 2008. Intraguild predation involving Harmonia axyridis: a review of current knowledge and future perspectives. BioControl 53:147-168.
Petersen, L.S. 1993. Effects of 45 insecticides, acaricides and molluscicides on the rove beetle Aleochara bilineata (Col.: Staphylinidae) in the laboratory. Entomophaga 38:371-382.
Porcar, M., I. Garcia-Robles, L. Dominguez-Escriba et A. Latorre. 2010. Effects of Bacillus thuringiensis Cry1Ab and Cry3Aa endotoxins on predatory Coleoptera tested through artificial diet-incorporation bioassays. Bulletin of Entomological Research 100:297-302.
Rees, B.E., D.M. Anderson, R.D. Gordon et D. Bouk. 1994. Larval key to genera and selected species of North American Coccinellidae (Coleoptera). Proceedings of the Entomological Society of Washington 96:387-412.
Richardson, M.L. et L.M. Hanks. 2009. Partitioning of niches among four species of orb-weaving spiders in a grassland habitat. Environmental Entomology 38:651-656.
Riddick, E.W., T.E. Cottrell et K.A. Kidd. 2009. Natural enemies of the Coccinellidae: Parasites, pathogens, and parasitoids. Biological Control 51:306-312.
Roy, H.E. et T. Cottrell. 2008. Forgotten natural enemies: interactions between coccinellids and insect-parasitic fungi. European Journal of Entomology 105:391-398.
Ruzicka, Z. 1997. Recognition of oviposition-deterring allomones by aphidophagous predators (Neuroptera: Chrysopidae, Coleoptera: Coccinellidae). European Journal of Entomology 94:431-434.
Salafsky, N., D. Salzer, A.J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O'Connor et D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conservation Biology 22:897-911.
Sato, S. et A.F.G. Dixon. 2004. Effect of intraguild predation on the survival and development of three species of aphidophagous ladybirds: Consequences for invasive species. Agricultural and Forest Entomology 6:21-24.
Schaeffer, P.W., R.J. Dysart et H.B. Specht. 1987. North American distribution of Coccinella septempunctata (Coleoptera: Coccinellidae) and its mass appearance in coastal Delaware. Environmental Entomology 16:368-373.
Singh, S.R., K.F.A. Walters, G.R. Port et P. Northing. 2004. Consumption rates and predatory activity of adult and fourth instar larvae of the seven spot ladybird, Coccinella septempunctata (L.), following contact with dimethoate residue and contaminated prey in laboratory arenas. Biological Control 30:127-133.
Skinner, B. et E. Domaine. 2010. Rapport sur la situation de la coccinnelle à neuf points (Coccinella novemnotata) au Québec. Ministère des Ressources naturelles et de la Faune du Québec. Faune Québec. 37 pp.
Sloggett, J.J. 2010. Predation of ladybird beetles by the orb-web spider Araneus diadematus. BioControl 55:631-638.
Sloggett, J.J. et M.E.N. Majerus. 2000. Habitat preferences and diet in the predatory Coccinellidae (Coleoptera): An evolutionary perspective. Biological Journal of the Linnaean Society 70:63-88.
Smith, B.C. 1966. Variation in weight, size, and sex ratio of Coccinellid adults (Coleoptera: Coccinellidae). The Canadian Entomologist 98:639-644.
Smith, C.A. et M.M. Gardiner. 2013. Biodiversity loss following the introduction of exotic competitors: does intraguild predation explain the decline of native lady beetles? PLoS ONE 8:e84448.
Smith, S.F. et V.A. Krischik. 1999. Effects of systemic imidacloprid on Coleomegilla maculata (Coleoptera, Coccinellidae). Environmental Entomology 28:1189-1195.
Snyder, W.E. et E.W. Evans. 2006. Ecological effects of invasive arthropod generalist predators. Annual Review of Ecology, Evolution and Systematics 37:95-122.
Snyder, W.E., G.M. Clevenger et S.D. Eigenbrode. 2004. Intraguild predation and successful invasion by introduced ladybird beetles. Oecologia 140:559-565.
Srivastava, S. et Omkar. 2004. Age-specific mating and reproductive senescence in the seven-spotted ladybird, Coccinella septempunctata. Journal of Applied Entomology 128:452-458.
Staines, C.L., M.J. Rothchild et R.B. Trumble. 1990. A survey of the Coccinellidae (Coleoptera) associated with nursery stock in Maryland. Proceedings of the Entomological Society of Washington 92:310-313.
Stellwag, L. et E. Losey. 2014. Sexual dimorphism in North American coccinellids: sexing methods for species of Coccinella L. (Coleoptera: Coccinellidae) and implications for conservation research. The Coleopterists Bulletin 68:271-281.
Stephens, E.J. 2002. Apparent extirpation of Coccinella novemnotata in New York State: Optimizing sampling methods and evaluating explanations for decline. MSc Thesis. Cornell University, Ithaca NY USA.
Theiling, K.M. et B.A. Croft. 1988. Pesticide side effects on arthropod natural enemies: a database summary. Agriculture, Ecosystems and Environment 21:191-218.
Tumminello, G., T.A. Ugine et J.E. Losey. 2015. Intraguild interactions of native and introduced coccinellids: The decline of a flagship species. Environmental Entomology 44:64-72.
Turchin, P. et P. Kareiva. 1989. Aggregation in Aphis varians - an effective strategy for reducing predation risk. Ecology 70:1008-1016.
Turnipseed, R.K., T.A. Ugine et J.E. Losey. 2014. Effect of prey limitation on competitive interactions between a native lady beetle, Coccinella novemnotata , and an invasive lady beetle, Coccinella septempunctata (Coleoptera: Coccinellidae). Environmental Entomology 43:969-976.
Turnock, W.J., I.L. Wise et F.O. Matheson. 2003. Abundance of some native coccinellines (Coleoptera: Coccinellidae) before and after the appearance of Coccinella septempunctata. The Canadian Entomologist 135:391-404.
Ugine, T.A. et J.E. Losey. 2014. Development times and age-specific life table parameters of the native lady beetle species Coccinella novemnotata (Coleoptera: Coccinellidae) and its invasive congener Coccinella septempunctata (Coleoptera: Coccinellidae). Physiological Ecology 43:1067-1075.
van der Werf, W., E.W. Evans et J. Powell. 2000. Measuring and modelling the dispersal of Coccinella septempunctata (Coleoptera: Coccinellidae) in alfalfa fields. European Journal of Entomology 97:487-493.
Vandenberg, N.J. 2002. Coccinellidae Latreille 1807. Pp. 371-389, in Arnett, R.H., M.C. Thomas, P.E. Skelley et J.H. Frank (éd.). American Beetles, Volume 2 Polyphaga: Scarabaeoidea through Curculionoidea, CRC Press, Boca Raton, FL.
Watson, W.Y. 1956. A study of the phylogeny of the genera of the tribe Coccinellini (Coleoptera). Contributions of the Royal Ontario Museum Life Sciences Division 42:1-52.
Watson, W.Y. 1976. A review of the genus Anatis Mulsant (Coleoptera: Coccinellidae). The Canadian Entomologist 108:935-944.
Webberley, K.M., G.D.D. Hurst, R.W. Husband et al. 2004. Host reproduction and an STD: causes and consequences of Coccipolipus hippodamiae distribution on coccinellids. Journal of Animal Ecology 73:1-10.
Webberley, K. M. et G.D.D. Hurst. 2002. The effect of aggregative overwintering on an insect sexually transmitted parasite system. Journal of Parasitology 88:707-712.
Wheeler, A.G., and E.R. Hoebeke. 1995. Coccinella novemnotata in northeastern North America: historical occurrence and current status (Coleoptera: Coccinellidae). Proceedings of the Entomological Society of Washington 97:701-716.
Wheeler, A.G. et C.A. Stoops. 1996. Status and spread of the Palaearctic lady beetles Hippodamia variegata and Propylea quatuordecimpunctata (Coleoptera: Coccinellidae) in Pennsylvania, 1993-1995. Entomological News 107:291-298.
Wiggins, G.B., S.A. Marshall et J.A. Downes. 1991. The importance of research collections of terrestrial arthropods. Brief for the Biological Survey of Canada (Terrestrial Arthropods) (Disponible en anglais seulement).
Youn, Y.N., M.J. Seo, J.G. Shin, C. Jang et Y.M. Yu. 2003. Toxicity of greenhouse pesticides to multicolored Asian lady beetles, Harmonia axyridis (Coleoptera: Coccinellidae). Biological Control 28:164-170.
Paul Grant (Ph.D.) est un entomologiste passionné qui, au fil des ans, a étudié de nombreux groupes d'insectes, dont les libellules, les papillons diurnes, les sauterelles et les coléoptères. Ses recherches sont actuellement centrées sur la bioacoustique des insectes. Il s'intéresse tout particulièrement aux facteurs limitant et favorisant une communication efficace, aux relations prédateurs-proies et à l'utilisation des sons produits par les insectes aux fins de la surveillance des espèces et de leur habitat. Paul Grant est également membre du Grasshopper Specialist Group (GSG) de l'Union internationale pour la conservation de la nature et de la Species Survival Commission (UICN / SSC) et coprésident du Sous-comité de spécialistes des arthropodes du Comité sur la situation des espèces en péril au Canada (COSEPAC).
- Acadia University Note1
- Centre de foresterie de l'Atlantique, Fredericton Note1
- Service canadien des forêts, Terre-Neuve Note1
- Collection nationale canadienne, Ottawa
- Musée canadien de la nature Note1
- Collection Claude-Chantal, Québec
- Collection d'insectes du Québec
- Collection Ouellet-Robert, Québec
- Centre de foresterie des Grands Lacs Note1
- Greg Pohl, comm. pers., 2014
- Insectarium de Montréal
- Collection de l'Insectarium René-Martineau, Québec
- John Acorn, comm. pers., 2014
- Centre de foresterie des Laurentides Note1
- Lost Lady Bug Project
- Musée d'Entomologie Lyman
- Nova Scotia Agricultural College Note1
- Royal Alberta Museum
- Royal BC Museum
- Musée royal de l'Ontario Note1
- Université Laval
- University of Alberta
- University of British Columbia
- University of Guelph Note1
- University of Manitoba
- Université du Nouveau-Brunswick Note1
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 2 | 0 |
| C | Moyen | 0 | 1 |
| D | Faible | 2 | 3 |
| - | Impact global des menaces calculé : | Très élevé | Élevé |
| # | Menace | Impact (calculé) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | Négligeable | Petite (1-10 %) | Négligeable (<1 %) | Élevée (continue) | - |
| 1.1 | Habitations et zones urbaines | Négligeable | Petite (1-10 %) | Négligeable (<1 %) | Élevée (continue) | La perte d'habitat et la réduction de la qualité de l'habitat occasionnées par l'intensification du développement résidentiel pourraient avoir joué un rôle dans le déclin de cette espèce à l'échelle locale. Les espaces verts et les jardins locaux dans les secteurs urbanisés de taille plus modeste pourraient toutefois continuer de procurer de l'habitat à la coccinelle à neuf points. |
| 1.2 | Zones commerciales et industrielles | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) |
Élevée (continue) | La perte d'habitat et la réduction de la qualité de l'habitat occasionnées par l'expansion des projets de développement commercial pourraient avoir joué un rôle dans le déclin de cette espèce à l'échelle locale, mais de façon globale, cette menace est considérée comme négligeable. |
| 1.3 | Tourisme et espaces récréatifs | Négligeable | Négligeable (< 1 %) |
Négligeable (< 1 %) | Élevée (continue) | La menace posée par la perte d'habitat et la réduction de la qualité de l'habitat occasionnées par les activités récréatives et touristiques est considérée comme faible car la plupart des aires réservées aux activités récréatives comportent des espaces ouverts convenant aux coccinelles. |
| 2 | Agriculture et aquaculture (en anglais seulement) | Faible | Petite (1-10 %) | Légère (1-10 %) |
Élevée (continue) | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | Faible | Petite (1-10 %) | Légère (1-10 %) |
Élevée (continue) | La perte d'habitat et la réduction de la qualité de l'habitat se poursuivent de façon soutenue à l'échelle de l'aire de répartition de l'espèce (Federal, Provincial and Territorial Governments of Canada, 2010; Javorek et Grant, 2011). L'homogénéisation des paysages agriculturaux et les changements touchant les pratiques agricoles, en particulier la dépendance accrue à l'égard des engrais et des pesticides, jouent probablement aussi un rôle dans le déclin des espèces indigènes à l'échelle locale (Wheeler et Hoebeke, 1995; Bianchi et al., 2007; Evans et al., 2011). Cette question est abordée ci-dessous dans la section traitant de la pollution (Menace 9). |
| 2.2 | Plantations pour la production de bois et de pâte | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | Négligeable. En général, la faible diversité floristique des plantations pour la production de bois et de pâte se traduit par une réduction de l'abondance des proies (pucerons). Les plantations pour la production de bois et de pâte sont peu nombreuses dans les prairies. |
| 2.3 | Élevage et élevage à grande échelle | Négligeable | Petite (1-10 %) | Négligeable (< 1 %) | Élevée (continue) | Négligeable. Le broutage peut avoir un impact direct sur les coccinelles par consommation directe, mais ce facteur n'est vraisemblablement pas important. De vastes étendues dans le sud de l'Alberta sont broutées, et la coccinelle à neuf points a été observée dans certaines de ces régions. Le broutage peut également avoir un impact bénéfique en favorisant la création de milieux ouverts. |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | Sans objet. |
| 3 | Production d'énergie et exploitation minière (en anglais seulement) | - | - | - | - | - |
| 3.1 | Forage pétrolier et gazier | - | - | - | - | Sans objet. Les éventuels inconvénients sont probablement compensés par les avantages potentiels. Les routes et les lignes sismiques et les structures linéaires ouvertes qui permettent la croissance de graminées et la régénération peuvent favoriser la création d'un habitat convenable et la dispersion de l'espèce. De façon globale, cette activité ne représente probablement pas une menace et est peut-être même bénéfique. |
| 3.2 | Exploitation de mines et de carrières | - | - | - | - | Sans objet. L'exploitation de carrières peut être bénéfique pour l'espèce car elle peut favoriser la création d'un habitat convenable. |
| 3.3 | Énergie renouvelable | - | - | - | - | Sans objet. Les voies d'accès et les milieux perturbés peuvent avoir des effets bénéfiques et ne représentent vraisemblablement pas une menace puisque cette espèce préfère les milieux ouverts. |
| 4 | Corridors de transport et de service (en anglais seulement) | - | - | - | - | - |
| 4.1 | Routes et voies ferrées | - | - | - | - | Effet potentiellement bénéfique (voir la menace 3). Les routes et les voies ferrées ne représentent vraisemblablement pas une menace puisque l'espèce préfère les milieux ouverts. |
| 4.2 | Lignes de services publics | - | - | - | - | Effet potentiellement bénéfique (voir la menace 3). Les lignes de services publics ne représentent vraisemblablement pas une menace puisque l'espèce préfère les milieux ouverts. |
| 4.3 | Transport par eau | - | - | - | - | Sans objet. |
| 4.4 | Trajectoires de vol | - | - | - | - | Sans objet. |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | - | - | - | - | - |
| 5.1 | Chasse et prélèvement d'animaux terrestres | - | - | - | - | Sans objet. Cette espèce n'est pas récoltée en milieu naturel à des fins de lutte biologique. La plupart des espèces récoltées à ces fins proviennent des États-Unis ou sont issues d'élevages. |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | Sans objet. |
| 5.3 | Exploitation forestière et récolte du bois | - | - | - | - | Sans objet. Les coupes à blanc auraient vraisemblablement un impact positif à court terme pour cette espèce. La majeure partie de l'aire de répartition de l'espèce se trouve dans des prairies et dans des milieux plus ouverts. |
| 5.4 | Pêche et récolte des ressources aquatiques | - | - | - | - | Sans objet. |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | - | - | - | - | - |
| 6.1 | Activités récréatives | - | - | - | - | Sans objet. |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | Sans objet. |
| 6.3 | Travaux et autres activités | - | - | - | - | Sans objet. |
| 7 | Modification du système naturel (en anglais seulement) | Faible | Restreinte-petite (1-30 %) |
Modérée-légère (1-30 %) | Élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | Sans objet. En général, les incendies créent des milieux ouverts et favorisent la succession de plantes à fleurs. Les incendies auraient donc probablement un impact positif dans certaines régions. |
| 7.2 | Barrages, gestion et utilisation de l'eau | - | - | - | - | Sans objet. |
| 7.3 | Autres modifications de l'écosystème | Faible | Restreinte-petite | Modérée-légère (1-30 %) | Élevée (continue) | L'abandon des terres gérées et des exploitations agricoles, principalement dans l'est de l'Ontario, pourrait contribuer au déclin de la coccinelle à neuf points. Ces terres sont soumises à la succession forestière naturelle. L'expansion urbaine et l'abandon des terres agricoles ont pour effet de limiter la disponibilité des sources de nourriture pour la coccinelle à neuf points. |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | Élevé-moyen | Généralisée (71-100 %) |
Grave-modérée (11-70 %) |
Élevée (continue) | - |
| 8.1 | Espèces exotiques/non indigènes envahissantes | Élevé-moyen | Généralisée (71-100 %) |
Grave-modérée (11-70 %) |
Élevée (continue) | Des prédateurs généralistes d'insectes ont été introduits par inadvertance ou intentionnellement hors de leur aire de répartition indigène à titre d'agents de lutte biologique depuis la fin du dix-neuvième siècle. L'altération de l'habitat ou les interactions avec des espèces non indigènes sont souvent la cause de déclins importants de l'aire de répartition ou de l'abondance d'insectes indigènes. L'invasion de l'Amérique du Nord par la coccinelle à sept points (Coccinella septempunctata) a été mise en cause dans le déclin généralisé des sous-populations de la coccinelle à neuf points et d'autres espèces de coccinelles indigènes. Les interactions négatives avec les espèces non indigènes récemment arrivées résultant de la compétition, de la prédation intraguilde ou d'effets indirects tels que l'introduction de pathogènes constituent le facteur le plus souvent évoqué pour expliquer les déclins de l'espèce. Les mauvaises herbes introduites pourraient également avoir un impact négatif pour les espèces de coccinelles indigènes. Les pathogènes représentent potentiellement une grande menace pour cette espèce. |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | Sans objet. Aucune espèce indigène d'oiseau ou de prédateur de coléoptères n'est considérée comme problématique. |
| 8.3 | Introduction de matériel génétique | - | - | - | - | Sans objet. |
| 9 | Pollution (en anglais seulement) | Élevé-faible | Grande- Restreinte (11-70 %) |
Grave-modérée (11-70 %) |
Élevée (continue) | - |
| 9.1 | Eaux usées domestiques et urbaines | - | - | - | - | Sans objet. |
| 9.2 | Effluents industriels et militaires | - | - | - | - | Sans objet. |
| 9.3 | Effluents agricoles et forestiers | Élevé-faible | Grande- restreinte (11-70 %) |
Grave-modérée (11-70 %) |
Élevée (continue) | En plus d'avoir un impact direct sur les coccinelles, les pesticides utilisés dans les régions agricoles peuvent avoir un impact indirect négatif sur la disponibilité des sources de nourriture en éliminant les pucerons qui s'attaquent aux plantes cultivées. |
| 9.4 | Détritus et déchets solides | - | - | - | - | Sans objet. |
| 9.5 | Polluants atmosphériques | - | - | - | - | Sans objet. |
| 9.6 | Énergie excessive | - | - | - | - | Sans objet. |
| 10 | Phénomènes géologiques (en anglais seulement) | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | Sans objet. |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | Sans objet. |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | Sans objet. |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | - | - | - | - | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | Inconnu. Il peut y avoir un déplacement de l'habitat. Ce facteur ne devrait pas avoir un impact important au cours des dix prochaines années, mais il pourrait avoir des répercussions plus graves à plus long terme. |
| 11.2 | Sécheresses | - | - | - | - | Inconnu. Les arbres stressés sont plus susceptibles d'être infestés par les pucerons et d'autres insectes ravageurs. Il est donc difficile d'apprécier la gravité de la menace posée par les sécheresses. |
| 11.3 | Températures extrêmes | - | - | - | - | Inconnu. Les gels tardifs peuvent avoir un impact sur les plantes, les pucerons et les coccinelles. Certains facteurs de stress ont des conséquences positives pour les pucerons, tandis que d'autres ont un impact négatif. |
| 11.4 | Tempêtes et inondations | - | - | - | - | Sans objet. |