
Criquet des Îles-de-la-Madeleine (Melanoplus madeleineae) : évaluation et rapport de situation du COSEPAC 2016
Titre officiel : Évaluation et Rapport de situation du COSEPAC sur le criquet des Îles-de-la-Madeleine (Melanoplus madeleineae) au Canada

Préoccupante
2016
Table des matières
- Table des matières
- Sommaire de l’évaluation
- Résumé
- Résumé technique
- Préface
- Description et importance de l’espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances de la population
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements
- Sources d’information
- Sommaire biographique des rédactrices du rapport
- Collections examinées
Liste des figures
- Figure 1. Criquet des Îles-de-la-Madeleine femelle. Spécimen recueilli sur l’île du Havre aux Maisons, aux îles de la Madeleine (Québec), le 11 août 2010. Spécimen recueilli par Paul Catling et versé dans la Collection nationale canadienne d’insectes, d’arachnides et de nématodes, à Ottawa (Ontario). Photo : Cory Sheffield.
- Figure 2. Criquet des Îles-de-la-Madeleine mâle. Spécimen recueilli sur l’île du Havre aux Maisons, aux îles de la Madeleine (Québec), le 11 août 2010. Spécimen recueilli par Paul Catling et versé dans la Collection nationale canadienne d’insectes, d’arachnides et de nématodes, à Ottawa (Ontario). Photo : Cory Sheffield.
- Figure 3. Emplacement des îles de la Madeleine par rapport au reste du Canada et de l’Amérique du Nord.
- Figure 4. La présence du criquet des Îles-de-la-Madeleine a récemment été confirmée à l’île du Havre aux Maisons et à l’île d’Entrée. Carte : GoogleEarth.
- Figure 5. Zone d’occurrence maximale et indice de zone d’occupation calculés pour le criquet des Îles-de-la-Madeleine. Remarque : le criquet n’a pas été observé dans tous les habitats des îles. Les îles ont une superficie de 205,5 km2.
- Figure 6. Activités de recherche du criquet des Îles-de-la-Madeleine en 2015.
- Figure 7. Pré maritime herbeux dégagé, habitat du criquet des Îles-de-la-Madeleine. Parmi les plantes en fleurs, on remarque la marguerite blanche (Leucanthemum vulgare). L’échouerie de la Pointe-Basse, Cap Alright (47,39265 N; 61,77428 O). Les spécimens ont été recueillis sur ce site en 2010 par Catling et ses collaborateurs (2013). Photo prise le 10 juillet 2015 par Jennifer Heron.
- Figure 8. Pré maritime herbeux dégagé, habitat du criquet des Îles-de-la-Madeleine. L’échouerie de la Pointe-Basse, Cap Alright (47,39265 N; 61,77428 O). Les spécimens ont été recueillis sur ce site en 2010 par Catling et ses collaborateurs (2013). Photo prise le 10 juillet 2015 par Jennifer Heron.
- Figure 9. Pré maritime herbeux dégagé où des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis et dont le code à barres a été établi, sur l’île du Havre aux Maisons (47,39715 N; 061,8453 O). Photo prise le 9 juillet 2015 par Jennifer Heron.
- Figure 10. Pré maritime herbeux dégagé où des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis et dont le code à barres a été établi, sur le chemin de la Belle-Anse, près du chemin des Caps, sur l’île du Cap aux Meules, aux îles de la Madeleine (47,39845 N; 061,93752 O). Photo prise le 10 juillet 2015 par Jennifer Heron.
- Figure 11. Pré maritime herbeux dégagé où des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis et dont le code à barres a été établi, sur le chemin de la Belle-Anse, près du chemin des Caps (47,39845 N; 061,93752 O). Photo prise le 10 juillet 2015 par Jennifer Heron.
- Figure 12. Carte des îles de la Madeleine présentant les secteurs d’habitat qui, selon les prévisions, sont menacés par la hausse du niveau de la mer attribuable aux changements climatiques. Carte tirée de Bernatchez et al. (2008).
Liste des tableaux
- Tableau 1. Registre des criquets des Îles-de-la-Madeleine conservés dans des musées et des collections. [Note des rédacteurs : Ce tableau a été modifié afin d’en supprimer les coordonnées géographiques. Pour obtenir le tableau complet, il faut communiquer avec le Secrétariat du COSEPAC.].
- Tableau 2. Activités de recherche du criquet des Îles-de-la-Madeleine, du 6 au 12 juillet 2015, pour un total de 40 heures-personne sur 24 sites. [Note des rédacteurs : Ce tableau a été modifié afin d’en supprimer les coordonnées géographiques. Pour obtenir le tableau complet, il faut communiquer avec le Secrétariat du COSEPAC.].
- Tableau 3. Résultats du calculateur des menaces de l’Union internationale pour la conservation de la nature – Partenariat pour les mesures de conservation (IUCN-CMP) pour le criquet des Îles-de-la-Madeleine.
Information sur le document
COSEPAC
Comité sur la situation
des espèces en péril
au Canada
![]()
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l’on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur le criquet des Îles-de-la-Madeleine (Melanoplus madeleineae) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xi + 49 p. (Registre public des espèces en péril site Web).
Note de production :
Le COSEPAC remercie Cory S. Sheffield et Jennifer Heron d’avoir rédigé le rapport de situation sur le criquet des Îles-de-la-Madeleine, Melanoplus madeleineae, au Canada, aux termes d’un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Paul Grant, coprésident du Sous-comité des spécialistes des arthropodes du COSEPAC.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Magdalen Islands Grasshopper Melanoplus madeleineae in Canada.
Illustration/photo de la couverture :
Criquet des Îles-de-la-Madeleine femelle photographié dans un pré dégagé sur l’île du Havre aux Maisons, aux Îles-de-la-Madeleine (Québec), avec l’île d’Entrée en arrière-plan, le 11 août 2010. – Photo : Paul Catling
COSEPAC Sommaire de l’évaluation
Sommaire de l’évaluation – novembre 2016
- Nom commun
- Criquet des Îles-de-la-Madeleine
- Nom scientifique
- Melanoplus madeleineae
- Statut
- Préoccupante
- Justification de la désignation
- Cette espèce endémique canadienne est restreinte aux îles de la Madeleine, au Québec, où elle a été observée dans sept des huit îles principales. Les menaces pesant sur l’espèce sont faibles, mais les activités récréatives, la mortalité attribuable à la circulation routière et la perte d’habitat due à une érosion côtière prévue pourrait avoir un impact sur l’espèce ou son habitat.
- Répartition
- Québec
- Historique du statut
- Espèce désignée « préoccupante » en novembre 2016.
COSEPAC Résumé
Criquet des Îles-de-la-Madeleine
Melanoplus madeleineae
Description et importance de l’espèce sauvage
Le criquet des Îles-de-la-Madeleine (Melanoplus madeleineae) est une espèce de grande taille (entre 21 et 29 mm) sans caractéristique distinctive, qui est endémique aux îles de la Madeleine, archipel situé dans la province de Québec, au Canada. Le principal trait qui permet de l’identifier est son fémur postérieur, dont la surface inférieure est pourpre et la surface extérieure est uniformément noire (c.-à-d. sans bande).
Le criquet des Îles-de-la-Madeleine est un héritage de la glaciation du Wisconsin. L’espèce est peut-être dérivée du criquet boréal, une espèce étroitement apparentée, plus petite, mais morphologiquement similaire, qui est absente des îles de la Madeleine, mais qui est très répandue sur le continent.
Répartition
L’aire de répartition mondiale et canadienne du criquet des Îles-de-la-Madeleine est restreinte aux îles de la Madeleine, archipel situé dans le golfe du Saint-Laurent, au Québec. L’archipel des îles de la Madeleine comporte huit îles principales et plusieurs autres îles plus petites et inhabitées. Ce criquet a été observé sur sept îles principales. Six de ces îles (île du Havre Aubert, île d'Entrée, île du Cap aux Meules, Grosse Île, île du Havre aux Maisons et île de la Pointe-aux-Loups) sont reliées par des bancs de sable et des petites îles, tandis que l’île d'Entrée est séparée d’environ 4 km du groupe principal. Cette espèce n’a pas été observée à l’île Brion, qui est située à environ 13 km de la Grosse Île.
Habitat
Le criquet des Îles-de-la-Madeleine est présent dans des prés maritimes dégagés et des milieux en pente abritant des communautés végétales (principalement des herbes ou du carex).
Biologie
Le criquet des Îles-de-la-Madeleine hiverne à l’état d’œuf; la nymphe éclot au début du printemps. L’espèce pourrait avoir la capacité de demeurer en diapause durant deux ans, ce qui a déjà été observé chez son taxon sœur, le criquet boréal. La croissance s’effectue par métamorphose graduelle : la larve traverse cinq stades nymphaux et elle est progressivement plus grosse et présente des caractéristiques morphologiques adultes plus prononcées au fil des mues. Les nymphes et les adultes partagent les mêmes habitudes alimentaires. Les adultes sont actifs de la mi-juillet à la fin de septembre, jusqu’au moment de l’accouplement et de la ponte. Les femelles pondent leurs œufs dans un substrat de sol mou, potentiellement sur le sol nu (p. ex. sentiers, chemins de terre, etc.). Comme la plupart des représentants des Mélanoplinés, cette espèce se nourrit probablement d’un vaste éventail de plantes hôtes, bien que ses préférences alimentaires (le cas échéant) soient inconnues.
Taille et tendances de la population
Il n’existe aucune information quant aux tendances et à la taille des sous-populations du criquet des Îles-de-la-Madeleine. Environ 80 spécimens ont été recueillis et conservés dans des musées en tant que spécimens témoins. Les relevés réalisés jusqu’à présent visaient surtout à consigner de nouvelles sous-populations, à recueillir des renseignements sur l’histoire naturelle et l’habitat et à étudier la génétique. Les relevés stratégiques de cette espèce ont surtout été réalisés à l’aide de filets fauchoirs dans des habitats convenables. Il est difficile de synchroniser les activités de collecte avec le moment culminant de l’émergence des adultes, ce qui donne à penser que le taux de détection et la perception de rareté varient considérablement.
Menaces et facteurs limitatifs
Plusieurs menaces potentielles planent sur le criquet des Îles-de-la-Madeleine. La mortalité routière est potentiellement élevée chez les criquets, là où les routes traversent un habitat convenable ou lorsque les adultes pondent sur des routes de terre qui leur offrent un substrat mou. Les activités récréatives comme la conduite de véhicules tout terrain sur des routes qui sillonnent les prés peuvent entraîner une mortalité directe, une compaction du sol et un piétinement des plantes dont se nourrissent les criquets. L’espèce peut également être la proie d’une foule de prédateurs, de parasites et de pathogènes naturels qui régulent les populations d’orthoptères. Dans l’ensemble, l’impact des menaces est considéré comme faible.
Protection, statuts et classements
Aucune loi fédérale ou provinciale ne protège en particulier le criquet des Îles-de-la-Madeleine, n’atténue les menaces propres aux criquets ou ne protège l’habitat de cette espèce. La cote de conservation mondiale est G2 (en péril), la cote de conservation nationale au Canada est N2 (en péril) et la cote provinciale au Québec est S2 (en péril). Environ 30 % de la superficie des îles principales des îles de la Madeleine est située sur les terres publiques. Par contre, on ignore dans quelle portion de ce territoire le criquet des Îles-de-la-Madeleine est présent.
Résumé technique
- Nom scientifique :
- Melanoplus madeleineae
- Nom français :
- Criquet des Îles-de-la-Madeleine
- Nom anglais :
- Magdalen Islands Grasshopper
- Répartition au Canada :
- Québec
Données démographiques
| Éléments du résumé technique | information |
|---|---|
| Durée d’une génération | De 1 à 2 ans |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d’individus matures? | Inconnu. |
| Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations]. | Inconnu. |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations]. | Inconnu. |
| Pourcentage [prévu ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu. |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu. |
| Est-ce que les causes du déclin sont a) clairement réversibles et b) comprises et c) ont effectivement cessé? | a. Sans objet. b. Sans objet. c. Sans objet. |
| Y a-t-il des fluctuations extrêmes du nombre d’individus matures? | Possible, oui. |
Information sur la répartition
| Éléments du résumé technique | information |
|---|---|
| Superficie estimée de la zone d’occurrence | 1 021 km2 (selon l’occupation par l’espèce sur toutes les îles où l’espèce a été observée). |
| Indice de zone d’occupation (IZO) (grille à carrés de 2 km x 2 km = 142 km2). |
568 km2 (toutefois, la superficie terrestre des îles s’élève à 205,5 km2) |
| La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d’occupation totale se trouvent dans des parcelles d’habitat qui sont a) plus petites que la superficie nécessaire au maintien d’une population viable et b) séparées d’autres parcelles d’habitat par une distance supérieure à la distance de dispersion maximale présumée pour l’espèce? | a. Non. b. Inconnu. |
| Nombre de localités (utilisez une fourchette plausible pour refléter l’incertitude, le cas échéant) (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
De 7 à 14 |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d’occurrence? | Non. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l’indice de zone d’occupation? | Non. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de sous-populations? | Non. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l’étendue ou la qualité] de l’habitat? | Oui. Déclin inféré de la qualité de l’habitat. |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non. |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non. |
| Y a-t-il des fluctuations extrêmes de la zone d’occurrence? | Non. |
| Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation? | Non. |
Nombre d’individus matures (dans chaque sous-population)
| Sous-populations (utilisez une fourchette plausible) | Nombre d’individus matures |
|---|---|
| Total | Inconnu. |
Analyse quantitative
| Éléments du résumé technique | information |
|---|---|
| La probabilité de disparition de l’espèce à l’état sauvage est d’au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Inconnu. Aucune donnée disponible pour réaliser l’analyse. |
Menaces
Un calculateur des menaces a-t-il été rempli pour l’espèce?
- 4.1 Routes et voies ferrées (impact faible)
- 6.1 Activités récréatives (impact faible)
- 8.1 Espèces exotiques (non indigènes) envahissantes (impact faible)
Quels autres facteurs limitatifs sont pertinents? Prédation et parasitisme.
Immigration de source externe (immigration de l’extérieur du Canada)
| Éléments du résumé technique | information |
|---|---|
| Situation des populations de l’extérieur les plus susceptibles de fournir des individus immigrants au Canada. | Sans objet. L’espèce est endémique aux îles de la Madeleine. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Sans objet. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Sans objet. |
| Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants? | Sans objet. |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
Sans objet. |
| Les conditions de la population source se détériorent-elles? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
Sans objet. |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) |
Sans objet. |
| La possibilité d’une immigration depuis des populations externes existe-t-elle? | Sans objet. |
Nature délicate de l’information sur l’espèce
| Éléments du résumé technique | information |
|---|---|
| L’information concernant l’espèce est-elle de nature délicate? | Non. |
Historique du statut
COSEPAC: Espèce désignée « préoccupante » en novembre 2016.
Statut et justification de la désignation :
| Éléments du résumé technique | information |
|---|---|
| Statut : | Préoccupante |
| Code alphanumérique : | Sans objet |
| Justification de la désignation : | Cette espèce endémique canadienne est restreinte aux îles de la Madeleine, au Québec, où elle a été observée dans sept des huit îles principales. Les menaces pesant sur l’espèce sont faibles, mais les activités récréatives, la mortalité attribuable à la circulation routière et la perte d’habitat due à une érosion côtière prévue pourraient avoir un impact sur l’espèce ou son habitat. |
Applicabilité des critères :
| Éléments du résumé technique | information |
|---|---|
| Critère A (déclin du nombre total d’individus matures) : |
Sans objet. Données insuffisantes. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) : |
Cette espèce satisfait presque à ce critère pour être désignée « menacée » en raison de sa petite aire de répartition, mais il a été impossible de déterminer avec certitude qu’elle satisfaisait au critère. |
| Critère C (nombre d’individus matures peu élevé et en déclin) : |
Sans objet. Données insuffisantes pour évaluer le déclin du nombre total d’individus matures. |
| Critère D (très petite population totale ou répartition restreinte) : |
Sans objet. Données insuffisantes pour affirmer que le nombre d’individus matures au sein de la population, l’indice de zone d’occupation et le nombre de localités sont supérieurs aux seuils. |
| Critère E (analyse quantitative) : |
Sans objet. Données existantes insuffisantes pour prévoir la probabilité de disparition du pays ou de la planète à l’état sauvage. |
Préface
Historique du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Mandat du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Composition du COSEPAC
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Définitions (2015)
- Espèce sauvage
- Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d’animal, de plante ou d’un autre organisme d’origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s’est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
- Disparue (D)
- Espèce sauvage qui n’existe plus.
- Disparue du pays (DP)
- Espèce sauvage qui n’existe plus à l’état sauvage au Canada, mais qui est présente ailleurs.
-
En voie de disparition (VD)
(Remarque : Appelée « espèce disparue du Canada » jusqu’en 2003.) - Espèce sauvage exposée à une disparition de la planète ou à une disparition du pays imminente.
- Menacée (M)
- Espèce sauvage susceptible de devenir en voie de disparition si les facteurs limitants ne sont pas renversés.
-
Préoccupante (P)
(Remarque : Appelée « espèce en danger de disparition » jusqu’en 2000.) - Espèce sauvage qui peut devenir une espèce menacée ou en voie de disparition en raison de l'effet cumulatif de ses caractéristiques biologiques et des menaces reconnues qui pèsent sur elle.
-
Non en péril (NEP)
(Remarque : Appelée « espèce rare » jusqu’en 1990, puis « espèce vulnérable » de 1990 à 1999.) - Espèce sauvage qui a été évaluée et jugée comme ne risquant pas de disparaître étant donné les circonstances actuelles.
-
Données insuffisantes (DI)
(Remarque :Autrefois « aucune catégorie » ou « aucune désignation nécessaire ».) - Une catégorie qui s’applique lorsque l’information disponible est insuffisante (a) pour déterminer l’admissibilité d’une espèce à l’évaluation ou (b) pour permettre une évaluation du risque de disparition de l’espèce.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu’en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d’Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Description et importance de l’espèce sauvage
Nom et classification
Embranchement : Arthropoda – Arthropodes
Sous-embranchement : Hexapoda – Hexapodes
Classe : Insecta – Insectes
Ordre : Orthoptera – Orthoptères (criquets, grillons et sauterelles)
Sous-ordre : Caelifera – Criquets
Superfamille : Acridoidea
Famille : Acrididae – Acrididés
Sous-famille : Melanoplinae – Mélanoplinés [en partie; l’espèce est souvent considérée comme étant membre de la sous-famille des Cyrtacanthacridinae]
Genre : Melanoplus
Espèce : Melanoplus madeleineae Vickery et Kevan 1978
Nom anglais : Magdalen Islands Grasshopper
Nom français : Criquet des Îles-de-la-Madeleine
Contexte taxonomique
Le Melanoplus madeleineae, ci-après appelé le criquet des Îles-de-la-Madeleine, a été décrit à partir de spécimens capturés en 1975 aux îles de la Madeleine, dans la province de Québec, et de spécimens historiques recueillis il y a fort longtemps, jusqu’en 1912 (Vickery et Kevan, 1978). L’espèce a été décrite par Vickery et Kevan (1978) en fonction de caractéristiques morphologiques permettant de l’identifier. Une récente analyse moléculaire appuie la validité taxonomique de cette espèce (Chapco et Litzenberger, 2002). Le criquet des Îles-de-la-Madeleine est étroitement apparenté (taxon sœur) au criquet boréal (M. borealis), bien que ce dernier ne soit pas présent aux îles de la Madeleine (Kevan et Vickery, 1978; Vickery et Kevan, 1985; Chapco et Litzenberger, 2002).
Le criquet des Îles-de-la-Madeleine appartient à la famille des Acrididae (Acrididés), qui est caractérisée par de courtes antennes. Cette famille est formée de trois sous-familles : les Melanoplinae, les Gomphocerinae et les Oedipodinae. Le criquet des Îles-de-la-Madeleine fait partie de la vaste sous-famille des Melanoplinae (Mélanoplinés), dont le régime alimentaire spécialisé est axé sur les plantes herbacées non graminoïdes et qui est caractérisée par un éperon (ou épine prosternale) situé sur la face ventrale, entre les pattes antérieures. La sous-famille des Melanoplinae comporte les criquets le plus couramment observés, notamment ceux du genre Melanoplus regroupant 239 espèces en Amérique du Nord (Arnett, 2000) et 42 espèces au Canada (Vickery et Kevan, 1985). Les deux autres sous-familles sont faciles à distinguer de la sous-famille des Melanoplinae, tant sur le plan morphologique que biologique.
Description morphologique
Le criquet des Îles-de-la-Madeleine se développe et grandit selon un processus de métamorphose graduelle comportant trois stades de vie : œuf, nymphe (formes immatures) et adulte. La métamorphose graduelle signifie que les nymphes ressemblent aux adultes et deviennent de plus en plus grandes au fil des mues.
Adultes
Le criquet des Îles-de-la-Madeleine (figures 1 et 2) est considéré comme étant un gros criquet (femelle : de 24 à 29 mm de longueur; mâle : de 21 à 25,5 mm de longueur). Comme les autres Mélanoplinés, l’espèce possède un éperon très évident situé sur la face ventrale, entre les pattes antérieures. Les antennes sont habituellement filiformes (c.-à-d. qu’elles ne sont pas aplaties, ni en forme de sabre). La tête de la majorité des Mélanoplinés, y compris celle du criquet des Îles-de-la-Madeleine, est orientée à la verticale (c.-à-d. qu’elle n’est pas fortement inclinée vers l’arrière, de l’extrémité à la base, comme c’est le cas chez les Acridinés) et n’est pas particulièrement grosse par rapport au corps. Les ailes antérieures sont légèrement pigmentées, mais ne présentent pas de bandes transversales, contrairement à la majorité des Locustinés. Les ailes postérieures ne sont pas pigmentées. La capacité de vol des Mélanoplinés varie beaucoup, même au sein d’un genre, et le criquet des Îles-de-la-Madeleine n’est probablement pas doté d’une puissante capacité de vol (Vickery et Kevan, 1985). Le corps de cette espèce est considéré comme étant peu dur par rapport aux corps sclérifiés des autres criquets.
Les caractéristiques distinctives du criquet des Îles-de-la-Madeleine sont notamment la couleur pourpre que l’on retrouve sur la surface inférieure du fémur postérieur, l’absence de bandes sur la surface extérieure du fémur postérieur et les taches dorées situées près des yeux (Vickery et Kevan, 1978; 1985) (figures 1 et 2). La couleur et la morphologie du criquet des Îles-de-la-Madeleine ressemblent beaucoup à celles de son plus proche parent, le criquet boréal, bien que ce dernier soit légèrement plus gros et que les genitalia des mâles n’aient pas la même forme (Vickery et Kevan, 1985). De plus, le criquet boréal n’est pas présent aux îles de la Madeleine (Vickery et Kevan, 1985).

Description longue pour la figure 1
Photo d’un spécimen de criquet des Îles-de-la-Madeleine femelle. Les traits distinctifs du criquet des Îles-de-la-Madeleine sont notamment la couleur pourpre de la surface inférieure de son fémur postérieur, l’absence de bandes sur la surface extérieure du fémur postérieur et les taches dorées situées près des yeux.

Description longue pour la figure 2
Photo d’un spécimen de criquet des Îles-de-la-Madeleine mâle. Les traits distinctifs du criquet des Îles-de-la-Madeleine sont notamment la couleur pourpre de la surface inférieure de son fémur postérieur, l’absence de bandes sur la surface extérieure du fémur postérieur et les taches dorées situées près des yeux.
Stades immatures
Les stades nymphaux (forme immature) ressemblent aux adultes, mais leurs ailes et leurs genitalia ne sont pas complètement développés et la sclérification de leur corps n’est pas terminée, des caractéristiques qui servent de confirmation taxonomique. Les stades immatures du criquet des Îles-de-la-Madeleine n’ont pas été décrits, et l’on ignore combien de mues il faut à la nymphe pour passer du stade de l’œuf au stade adulte. Toutefois, chez son proche parent, le criquet boréal, la nymphe traverse cinq stades nymphaux avant de devenir adulte, et l’on peut supposer que ce nombre est le même pour le criquet des Îles-de-la-Madeleine. Selon la description du criquet boréal, la larve du premier stade est surtout de couleur foncée, à l’exception d’une bande brune étroite au milieu du dos et d’un croissant pâle sur le côté de la tête et le pronotum. Du deuxième au cinquième stade nymphal, la couleur globale du corps de la nymphe est habituellement verdâtre ou brune. Les nymphes ont été recueillies au cours de la campagne de terrain de 2015 aux fins de la préparation du présent rapport de situation.
Œufs
Les œufs du criquet des Îles-de-la-Madeleine n’ont pas été décrits. Toutefois, chez son proche parent, le criquet boréal, les œufs sont allongés et habituellement bruns, et ils sont pondus en oothèques de 30 à 50 oeufs.
Structure spatiale et variabilité des populations
L’aire de répartition connue du criquet des Îles-de-la-Madeleine est limitée aux îles de la Madeleine, une petite région composée d’un grand nombre d’îles et d’îlots. Les îles principales (île du Havre Aubert, île du Cap aux Meules, île du Havre aux Maisons, Grosse Île, île de l’Est et île de la Grande Entrée) sont reliées entre elles par des dunes de sable ou d’autres petites portions de terre.
Les études confirment que le criquet des Îles-de-la-Madeleine est endémique à cette région, d’après les caractéristiques génétiques distinctives qui le différencient de son taxon sœur, le criquet boréal (Chapco et Litzenberger, 2002), lequel, selon les connaissances actuelles, n’est pas présent sur ces îles (Vickery et Kevan, 1985). Selon les estimations, l’événement géologique qui a séparé ces deux espèces se serait produit il y a environ 85 000 à 11 000 ans (Chapco et Litzenberger, 2002).
On en connaît peu sur la structure spatiale des sous-populations de criquet des Îles-de-la-Madeleine. On considère que l’espèce n’est pas dotée d’une puissante capacité de vol (Vickery et Kevan, 1985) et que sa capacité à migrer entre les îles est probablement limitée. Toutefois, cette espèce a récemment été observée sur l’île d’Entrée, qui est située à 4,8 km du groupe d’îles principales (Catling et al., 2013). Il est probable que cette espèce ait toujours été présente sur cette île, mais qu’elle n’ait jamais été détectée auparavant.
Unités désignables
Le criquet des Îles-de-la-Madeleine est évalué comme une seule unité désignable. L’espèce a été observée sur sept des quatorze îles de l’archipel des îles de la Madeleine et est endémique à ces îles. On ne dispose d’aucune information sur le caractère distinct ou l’importance au point de vue évolutif des sous-populations de ces sept îles. L’espèce est présente dans l’aire écologique de l’Atlantique du COSEPAC.
Importance de l’espèce
Rarement des spécimens de criquet des Îles-de-la-Madeleine ont été prélevés (Catling et al., 2013), et cette espèce est endémique aux îles de la Madeleine (Vickery et Kevan, 1978; 1985; Catling et al., 2013). On croit qu’elle représente un taxon unique dérivé du criquet boréal à la suite d’un isolement qui se serait produit au cours de la période de glaciation du Wisconsin (Cameron, 1962; voir Vickery et Kevan, 1985).
Bien qu’elles fassent partie du Plateau néo-écossais dans le nord-est de l’Amérique du Nord, les îles de la Madeleine ont été isolées du continent au moins depuis le retrait du dernier inlandsis du Wisconsin, il y a de cela environ de 85 000 à 11 000 ans (Cameron, 1962). On présume que le niveau de la mer était plus bas avant le retrait de cet inlandsis et que l’aire de répartition du criquet boréal s’étendait alors sur les masses continentales végétalisées reliées au continent. La hausse du niveau de la mer résultant du retrait de l’inlandsis a probablement isolé les populations de criquet boréal restantes sur les masses continentales plus élevées séparées des zones plus basses du Plateau néo-écossais, ce qui a favorisé une spéciation dépourvue de tout flux génétique avec les populations du continent. Des preuves moléculaires montrent une très étroite affinité entre le criquet des Îles-de-la-Madeleine et les populations continentales du criquet boréal. Hamilton (2002), Fortin et Larocque (2003) et Rémillard et ses collaborateurs (2016) décrivent en détail l’histoire glaciaire des îles de la Madeleine. Hamilton (2002) discute également de l’isolement et de la spéciation de l’entomofaune.
Répartition
Aire de répartition mondiale canadienne
Les aires de répartition mondiale et canadienne du criquet des Îles-de-la-Madeleine sont restreintes aux îles de la Madeleine, dans le golfe du Saint-Laurent (Québec, Canada) (figure 3). Cet archipel est composé de huit îles principales et de plusieurs autres petites îles inhabitées; la superficie terrestre de l’archipel est de 205 km2 (répartie entre les 14 îles et îlots de l’archipel). Six des îles principales (île du Havre Aubert, île du Cap aux Meules, Grosse Île, île du Havre aux Maisons, Pointe aux Loups et île de la Grande Entrée) sont reliées par des bancs de sable, et le criquet des Îles-de-la-Madeleine est probablement présent sur toutes ces îles. Deux grosses îles ne sont pas reliées à ce groupe et sont séparées par l’océan des autres îles principales. On sait que l’île d’Entrée, située à 4,8 km de l’extrémité sud-est du groupe d’îles principales, possède une sous-population de criquet des Îles-de-la-Madeleine (Catling et al., 2013); l’île Brion est située à environ 13 km au nord de la Grosse Île, mais aucun relevé n’y a été effectué afin de vérifier la présence du criquet des Îles-de-la-Madeleine, comme c’est le cas pour plusieurs autres petites îles inhabitées qui sont considérées comme faisant partie de l’archipel (c.-à-d. Rocher aux Oiseaux, île aux Loups Marins, île Paquet et Rocher du Corps-Mort).
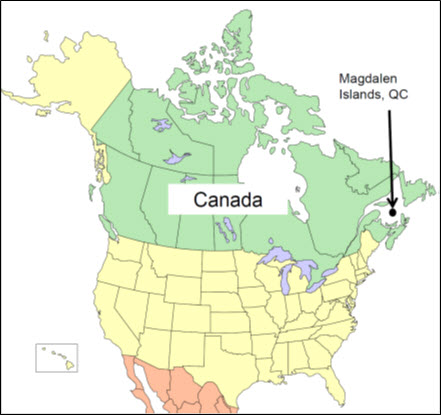
Description longue pour la figure 3
Carte de l’Amérique du Nord indiquant l’emplacement des îles de la Madeleine dans le golfe du Saint-Laurent, au Québec.
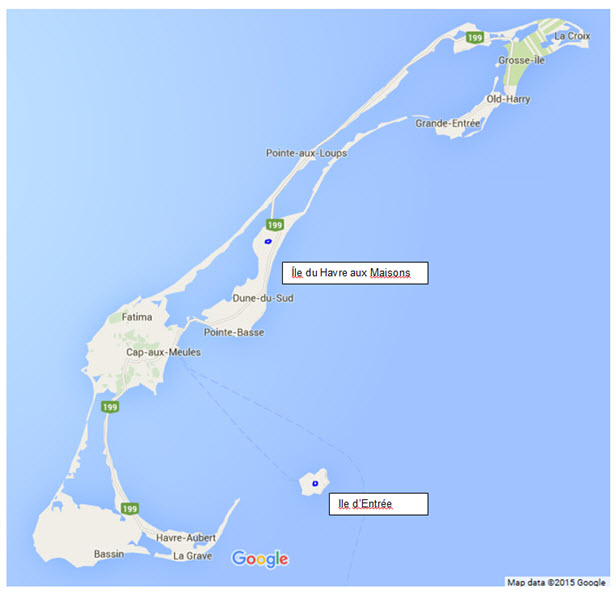
Description longue pour la figure 4
Carte indiquant l’emplacement des deux îles (Île du Havre aux Maisons et Île d’Entrée) de l’archipel des îles de la Madeleine où la présence du criquet des Îles-de-la-Madeleine a été récemment confirmée.
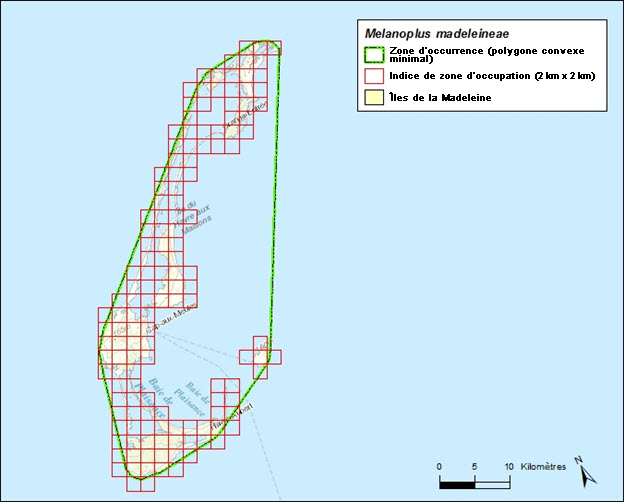
Description longue pour la figure 5
Carte illustrant la zone d’occurrence maximale du criquet des Îles-de-la-Madeleine et la grille à carrés ayant servi à calculer l’indice de zone d’occupation.

Description longue pour la figure 6
Carte montrant les sites (au Québec [îles de la Madeleine], à l’Île-du-Prince-Édouard et au Nouveau-Brunswick) qui où des relevés ont été effectués à la recherche de criquets des Îles-de-la-Madeleine en 2015 en vue du présent rapport.

Description longue pour la figure 7
Photo montrant l’habitat du criquet des Îles-de-la-Madeleine : un pré maritime herbeux dégagé. Parmi les plantes en fleurs, on remarque la marguerite blanche (Eucanthemum vulgare).

Description longue pour la figure 8
Photo montrant l’habitat du criquet des Îles-de-la-Madeleine : un pré maritime herbeux dégagé.

Description longue pour la figure 9
Photo d’un pré maritime herbeux dégagé où des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis et dont le code à barres a été établi.

Description longue pour la figure 10
Photo d’un pré maritime herbeux dégagé où des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis et dont le code à barres a été établi.

Description longue pour la figure 11
Photo d’un pré maritime herbeux dégagé où des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis et dont le code à barres a été établi.
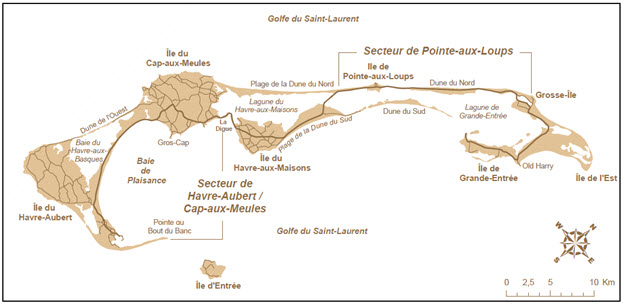
Description longue pour la figure 12
Carte des îles de la Madeleine montrant deux secteurs d’habitat susceptibles d’être inondés en raison de la hausse du niveau de la mer attribuable aux changements climatiques.
Zone d’occurrence et zone d’occupation
La superficie terrestre des îles de la Madeleine totalise 205,5 km2. La zone d’occurrence maximale pour le criquet des Îles-de-la-Madeleine est d’environ 1 021 km2, selon un polygone convexe minimal qui englobe toutes les îles de la Madeleine. L’indice de zone d’occupation (IZO), déterminé à l’aide d’une grille à carrés de 2 km x 2 km, s’élève à 568 km2. L’espèce n’est pas présente dans tout l’habitat qui est pris en compte dans ce calcul spatial, comme les boisés denses ou les plages de sable, ou encore les étendues marines qui n’offrent pas un habitat convenable pour l’espèce; par conséquent, la zone d’occupation biologique est probablement beaucoup plus petite.
Activités de recherche
Le criquet des Îles-de-la-Madeleine est considéré comme étant endémique aux îles de la Madeleine selon l’examen de nombreux spécimens de Melanoplus d’un peu partout au Canada (voir Vickery et Kevin, 1978; Vickery et Kevin, 1985). Il existe environ 80 spécimens témoins du criquet des Îles-de-la-Madeleine conservés dans des musées, dont les dates de collecte varient de 1912 à 2015 (tableau 1), y compris les spécimens signalés par Vickery et Kevan (1978), Chapco et Litzenberger (2002) et Catling et al. (2013).
| Nom du site de collecte | Nombre de spécimens | Sexe | Catégorie | Date de collecte | Collecteur(s) | Musée* | Référence |
|---|---|---|---|---|---|---|---|
| Île du Havre aux Maisons | 1 | - | - | 11 août2010 | P.M. Catling, B. Kostiuk | CNC | Catling et al., 2013 |
| Île du Havre aux Maisons | 1 | - | - | 13 août2010 | P.M. Catling, B. Kostiuk | CNC | Catling et al., 2013 |
| Île d’Entrée | 1 | - | - | 14 août2010 | P.M. Catling, B. Kostiuk | CNC | Catling et al., 2013 |
| Île du Havre aux Maisons, Paroissede Havre-aux-Maisons, entre Havre-aux-Maisons et Pointe-Basse | 1 | ♂ | Holotype | 9 septembre1975 | K.E. Kevan | ML | Vickery et Kevan, 1978 |
| Îlede la Grande Entrée, butesurplombant le bassin de l’île Boudreau | 1 | ♀ | Allotype | 9 septembre1975 | D.K. McE.Kevan | ML | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons, Paroissede Havre-aux-Maisons, entre Havre-aux-Maisons et Pointe-Basse | 2 | ♂ | Paratype | 8 septembre1975 | D.J. Luckin | ML | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons, Paroissede Havre-aux-Maisons, entre Havre-aux-Maisons et Pointe-Basse | 5 | ♀ | Paratype | 8 septembre1975 | D.J. Luckin | ML | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons, Paroissede Havre-aux-Maisons, entre Havre-aux-Maisons et Pointe-Basse | 1 | ♂ | Paratype | 8 septembre1975 | K.E. Kevan | ML | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons, Paroissede Havre-aux-Maisons, entre Havre-aux-Maisons et Pointe-Basse | 1 | ♂ | Paratype | 8 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons, Paroissede Havre-aux-Maisons, entre Havre-aux-Maisons et Pointe-Basse | 1 | ♀ | Paratype | 8 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Au sud-est duvillage de Havre-aux-Maisons | 1 | ♀ | Paratype | 11 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Cap Alright | 1 | ♀ | Paratype | 9 septembre1975 | D.J. Luckin | ML | Vickery et Kevan, 1978 |
| Île du Capaux Meules, Paroisse de Fatima, chemin Patton | 1 | ♂ | Paratype | 11 septembre1975 | D.J. Luckin | ML | Vickery et Kevan,1978 |
| Îledu Cap aux Meules, Paroisse deFatima, chemin Patton | 1 | ♀ | Paratype | 11 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Îledu Cap aux Meules, Paroisse deFatima, chemin Patton | 1 | ♀ | Paratype | 11 septembre1975 | K.E. Kevan | ML | Vickery et Kevan, 1978 |
| Île du Havre Aubert, Paroisse | 2 | ♀ | Paratype | 10 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Île du Havre Aubert, Cap Gridley | 1 | ♀ | Paratype | 10 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Au sud deVigneau | 1 | ♀ | Paratype | 10 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| Île aux Loups, au sud-ouest dePointe-aux-Loups | 1 | ♀ | Paratype | 9 septembre1975 | D.K. Kevan | ML | Vickery et Kevan, 1978 |
| « Îles de la Madeleine » | 1 | ♀ | Paratype | 29 juillet1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| « Îles de la Madeleine », août | 1 | ♀ | Paratype | « 1952 » | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| La Vernière | 1 | ♂ | Paratype | 26 juillet1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| La Vernière | 1 | ♀ | Paratype | 26 juillet1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| La Vernière | 1 | ♀ | Paratype | 5 août 1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| Île du Cap aux Meules | 2 | ♂ | Paratype | 29 juillet1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| Étangdu Nord | 1 | ♂ | Paratype | 29 juillet1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| Étangdu Nord | 2 | ♀ | Paratype | 29 juillet1952 | J.R. Beaudry | Université deMontréal | Vickery et Kevan, 1978 |
| Île aux Loups [Wolfe I.] | 1 | ♀ | Paratype | 21 août 1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery et Kevan, 1978 |
| Îlede la Grande Entrée [Coffin I.] | 1 | ♀ | Paratype | 17 août1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons[Alright I.] | 1 | ♂ | Paratype | 21 août1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery et Kevan, 1978 |
| Île du Havre Aubert[Basin I.] | 1 | ♂ | Paratype | « 1912 » | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery et Kevan,1978 |
| Île du Cap aux Meules[Grindstone I.] | 1 | ♂ | Paratype | 23 août 1912 | Bayard Long | Academy of Natural Sciences of Philadelphia | Vickery et Kevan, 1978 |
| Îlede la Grande Entrée [Coffin I.] | 1 | ♀ | Paratype | 17 août 1912 | Bayard Long | Musée royalde l’Ontario | Vickery et Kevan, 1978 |
| Île du Cap aux Meules[Grindstone I.] | 1 | ♂ | Paratype | 23 août 1912 | Bayard Long | Musée royalde l’Ontario | Vickery et Kevan, 1978 |
| Île du Havre aux Maisons; préadjacent à la plage | À terminer | immature | No Genbank? | 7 juillet2015 | C. Sheffieldet J. Heron | RSM | Sheffield etHeron, données personnelles |
| Chemin de laBelle-Anse, près du chemin des Caps | À terminer | immature | NoGenbank? | 10 juillet2015 | C. Sheffieldet J. Heron | RSM | Sheffield etHeron, données personnelles |
CNC = Collection nationale canadienne d’insectes, d’arachnides et de nématodes, Ottawa (Ont.).
ML = Musée Lyman, Université McGill, Montréal (Qc).
RSM = Royal Saskatchewan Museum, Regina (Sask.).
| Jour | Mois | Année | Temps de recherche (en minutes) pour deux personnes | Province | Lieu | Description de l’habitat | Criquets recueillis |
|---|---|---|---|---|---|---|---|
| 6 | Juillet | 2015 | 70 | N.-B. | Sortie 53,autoroute 15, sortie Cap Pelé | Accotement deroute jonché de fleurs, d’environ 20 à 30 mètres de largeur, sur un solhumide où il y avait beaucoup de plantes en fleurs. | - |
| 6 | Juillet | 2015 | 90 | Î.-P.-É. | Souris, àl’extérieur du village; près du Lighthouse and Beach Motel | Milieu humideherbeux adjacent à la plage; sol sablonneux | - |
| 7 | Juillet | 2015 | 18 | Î.-P.-É. | Souris, àl’extérieur du village; dans le fossé au bord de la route | Accotement deroute jonché de fleurs où l’on retrouve de grandes parcelles de lupin | - |
| 8 | Juillet | 2015 | 170 | Qc :îles de la Madeleine | Grosse Île etGrande Entrée | Au bout de laflèche; herbes des prés et de la dune de sable | Non |
| 8 | Juillet | 2015 | 90 | Qc :îles de la Madeleine | Forêtsd’épinettes et de sapins au bord de la route | Forêtsd’épinettes et de sapins au bord de la route | - |
| 9 | Juillet | 2015 | 270 | Qc :îles de la Madeleine | Étangdu Nord; propriétémunicipale | Milieu humideadjacent aux dunes de sable (océan) et à l’étang (eau douce); collecte dansdeux habitats – en bordure des dunes de sable et dans un habitat de terreshumides et de tourbières. | Oui |
| 9 | Juillet | 2015 | 220 | Qc : îlesde la Madeleine | Réservenaturelle de laGrande-Demoiselle – paysage culturel –site 12 | Oui(immature) | |
| 9 | Juillet | 2015 | 30 | Qc :îles de la Madeleine | Milieuherbeux dégagé adjacent à un stationnement dans un centre urbain | Accotementherbeux de route adjacent à un ravin où il y a un ruisseau à fort débit;plantes envahissantes et herbes | Oui |
| 9 | Juillet | 2015 | 90 | Qc :îles de la Madeleine | Plage, halteroutière | Halteroutière donnant accès à la plage et à un milieu herbeux près d’une dune desable au bord de l’eau; absence de pente (basse terre) | Oui(immature) |
| 10 | Juillet | 2015 | 70 | Qc :îles de la Madeleine | Pointe-Basse,Cap Alright | Pré plat(pente < 5 %; altitude : 26 m); Pré maritime de bassealtitude composé d’un entrecroisement de végétation plus courte et devégétation plus haute et de zones plus denses dominées par l’astragale. | - |
| 10 | Juillet | 2015 | 20 | Qc :îles de la Madeleine | Bordure deroute | Bord de routeherbeux où l’on trouve des coccinelles | Non |
| 10 | Juillet | 2015 | 70 | Qc :îles de la Madeleine | Pré herbeuxvallonneux | Pré herbeuxvallonneux en bordure de route; altitude : 54 m; un peu de rochesexposées | Oui(immature) |
| 10 | Juillet | 2015 | 210 | Qc :îles de la Madeleine | Cap àFidèle, site de la Côte – site 4 | Pré maritime– bon site pour les abeilles et autres hyménoptères; habitats ayant subid’importantes perturbations où l’on trouve des parcelles herbeuses adjacentesà un quai comportant un développement commercial (amarrage de bateaux) et unrestaurant; beaucoup de circulation et stationnement très fréquenté | Oui |
| 10 | Juillet | 2015 | 90 | Qc :îles de la Madeleine | Aire àpique-nique, cheminde la Belle-Anse, près du chemin des Caps | Pré degenévriers; végétation très courte et herbes; astragales épars, boutons d’or,trèfles rouges, carex et fraises | - |
| 10 | Juillet | 2015 | 120 | Qc :îles de la Madeleine | Pointeà Frank – site 6 | Près del’océan | - |
| 10 | Juillet | 2015 | Pièges lumineux | Qc :îles de la Madeleine | Réservenationale de faune de la pointe de l’Est; propriété du SCF | Habitat dedunes de sable dégagées contenant de la bruyère et d’autres plantes des dunes | - |
| 8 - 11 | Juillet | 2015 | Continue sur une période de 7 jours | Qc :îles de la Madeleine | Maisondu 47, chemin Vigneau, Havre-aux-Maisons | Cour arrièreherbeuse contenant du lupin et d’autres fleurs sauvages | Non |
| 11 | Juillet | 2015 | 108 | Qc :îles de la Madeleine | Baie du Havreaux Basques | Bord de plageen bordure de route; stationnement de halte routière | Oui (pas dansune cariçaie où l’habitat est probablement inondé, mais dans des zonesdégagées herbeuses sur sol sablonneux, plus élevées, qui ont peu de chanced’être inondées) |
| 11 | Juillet | 2015 | 120 | Qc :îles de la Madeleine | Cap au Sud;propriété privée du phare | Pré maritimeadjacent à l’océan (falaise abrupte > 40 m) où se trouve un phare. | Oui |
| 11 | Juillet | 2015 | 60 | Qc :îles de la Madeleine | La dune de l’ouest; Veillesur les Isles |
Dunes desable près de l’océan et adjacentes à un grand milieu humide/ marais;tourbière à canneberges; habitat plat | Oui (quelquesspécimens) |
| 11 | Juillet | 2015 | 42 | Qc :îles de la Madeleine | Intersection entre le chemin de la Montagne et le chemin des Buttes; zonesde gravier en bordure de route | Habitatforestier contenant diverses espèces de sapins (Abies spp.) | - |
| 11 | Juillet | 2015 | 78 | Qc :îles de la Madeleine | Adjacent à latour radio; intersection entre le chemin de la Montagne et le chemin du Grand-Pré | Pré dégagécontenant quelques conifères et forêt mixte | Oui |
| 11 | Juillet | 2015 | 96 | Qc :îles de la Madeleine | Cap àFidèle; site de laCôte – site 4; capture de Methocha! | Zone du quaifédéral du Canada; pré maritime – bon site pour les abeilles et autreshyménoptères; habitats ayant subi d’importantes perturbations où l’on trouvedes parcelles herbeuses adjacentes à un quai comportant un développementcommercial (amarrage de bateaux) et un restaurant; beaucoup de circulation etstationnement très fréquenté | Oui |
| 12 | Juillet | 2015 | 120 | Qc :îles de la Madeleine | Réservenationale de faune de la pointe de l’Est; propriété du SCF | Habitat dedunes de sables dégagées contenant de la bruyère et d’autres plantes desdunes | - |
Tableau 3. Résultats du calculateur des menaces de l’Union internationale pour la conservation de la nature – Partenariat pour les mesures de conservation (IUCN-CMP) pour le criquet des Îles-de-la-Madeleine.
- Nom scientifique de l’espèce ou de l’écosystème
- Criquet des Îles-de-la-Madeleine (Melanoplus madeleineae)
- Identification de l’élément
- -
- Code de l’élément
- -
- Date
- 2015-12-08
- Évaluateur(s) :
- Cory Sheffield et Jennifer Heron (auteurs du rapport); Angèle Cyr (Secrétariat du COSEPAC); Paul Grant (coprésident du Sous-comité des spécialistes des arthropodes); Nathalie Desrosiers (ministère des Forêts, de la Faune et des Parcs); Mathieu Morin (ministère des Forêts, de la Faune et des Parcs – gestionnaire pour les îles de la Madeleine); James Miskelly (membre du Sous-comité des spécialistes des arthropodes), John Klymko (membre du Sous-comité des spécialistes des arthropodes), Jessica Linton (membre du Sous-comité des spécialistes des arthropodes)
- Références :
- -
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l’intensité de leur impact : Maximum de la plage d’intensité |
Comptes des menaces de niveau 1 selon l’intensité de leur impact : Minimum de la plage d’intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 0 | 0 |
| D | Faible | 3 | 3 |
| - | Impact global des menaces calculé : | Faible | Faible |
- Impact global des menaces attribué :
- D = Faible
| # | Menace | Impact (calculé) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | Élevée (constante) | - |
| 1.1 | Habitations et zones urbaines | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | Élevée (constante) | Le développement immobilier est minimal aux îles de la Madeleine. La majorité des lots sont grands et les maisons n’occupent qu’une petite partie des parcelles de terrain. Des règlements municipaux sont en place pour réduire au minimum le développement immobilier dans les zones rurales des îles. |
| 2 | Agriculture et aquaculture (en anglais seulement) | Inconnu | Petite (1-10 %) | Inconnue | Élevée (constante) | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | Inconnu | Petite (1-10 %) | Inconnue | Élevée (constante) | L’utilisation des zones agricoles semi-naturelles, comme les champs de foin, par le criquet des Îles-de-la-Madeleine est inconnue. La création d’un habitat dégagé peut avoir eu un effet positif sur le criquet. Une superficie de 600 ha est cultivée pour la production de foin sur les îles de la Madeleine, et ce foin est récolté une ou deux fois par année (Morin, comm. pers., 2015). Les répercussions de la récolte du foin sur le criquet boréal, l’espèce la plus apparentée au criquet des Îles-de-la-Madeleine, donnent à penser que cette espèce est tolérante à la récolte du foin. |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | Sans objet. Le criquet n’est pas présent dans les zones boisées. |
| 2.3 | Élevage et élevage à grande échelle | Inconnu | Petite (1-10 %) | Inconnue | Élevée (constante) | La menace est présente. Il y a un certain niveau de broutage dans les pâturages des îles, mais la gravité de cet impact est inconnue. |
| 3 | Production d’énergie et exploitation minière (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Modérée (peut-être à court terme, < 10 ans) | - |
| 3.3 | Énergie renouvelable | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Modérée (peut-être à court terme, < 10 ans) | À l’heure actuelle, il n’y a aucune infrastructure ou installation de production d’énergie solaire ou éolienne à grande échelle aux îles. Toutefois, un projet d’installation de trois éoliennes est en cours d’examen par le gouvernement du Québec. L’emprise de l’infrastructure éolienne prévue est située directement dans l’habitat d’une plante en voie de disparition et l’habitat d’autres espèces en péril. On ignore si le criquet est présent sur cette même emprise. |
| 4 | Corridors de transport et de service (en anglais seulement) | Faible | Petite (1-10 %) | Élevée (31-70 %) | Élevée (constante) | - |
| 4.1 | Routes et voies ferrées | Faible | Petite (1-10 %) | Élevée (31-70 %) | Élevée (constante) | La menace est présente. Mortalité potentielle de criquets causée par les véhicules, surtout sur les chemins de terre où les criquets peuvent se reposer et où les femelles sont susceptibles de pondre. |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | - | - | - | - | - |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | Faible | Petite (1-10 %) | Modérée (11-30 %) | Élevée (constante) | - |
| 6.1 | Activités récréatives | Faible | Petite (1-10 %) | Modérée (11-30 %) |
Élevée (constante) | La menace est présente. Les îles de la Madeleine sont une destination touristique populaire pour les activités récréatives. Le nombre de visiteurs a augmenté au fil des ans (voir la section Menaces). |
| 7 | Modification du système naturel (en anglais seulement) | - | Pas une menace | Légère (1-10 %) |
Effet neutre ou avantage potentiel | - |
| 7.3 | Autres modifications de l’écosystème | - | Pas une menace | Légère (1-10 %) |
Effet neutre ou avantage potentiel | Effet neutre ou bénéfique. Un certain nombre de digues pourraient être construites pour prévenir l’érosion. De tels travaux stabiliseraient une partie des zones et préviendraient la perte d’habitat. |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | Faible | Généralisée (71-100 %) | Légère (1-10 %) |
Élevée (constante) | - |
| 8.1 | Espèces exotiques/non indigènes envahissantes | Faible | Généralisée (71-100 %) | Légère (1-10 %) |
Élevée (constante) | La menace est présente. Un grand nombre de prédateurs exotiques peuvent se nourrir de criquets (voir la section Menaces) |
| 9 | Pollution (en anglais seulement) | - | - | - | - | - |
| 10 | Phénomènes géologiques (en anglais seulement) | Non calculé (immédiateté inconnue) | Petite (1-10 %) | Extrême (71-100 %) |
Inconnue | - |
| 10.2 | Tremblements de terre et tsunamis | Non calculé (immédiateté inconnue) | Petite (1-10 %) | Extrême (71-100 %) |
Inconnue | La menace est peut-être présente. Les six îles principales sont reliées par des bancs de sable. Des tremblements de terre et des tsunamis pourraient endommager à court terme l’habitat de connexion. |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | Inconnu | Généralisée (71-100 %) | Inconnue | Modérée (peut-être à court terme, < 10 ans) | - |
| 11.1 | Déplacement et altération de l’habitat | Non calculé (en dehors de la période d’évaluation) | Petite (1-10 %) | Inconnue | Faible (peut-être à long terme, > 10 ans) | La possible hausse du niveau de la mer pourrait avoir des répercussions sur l’habitat de plages riveraines. |
| 11.2 | Sécheresses | Non calculé (en dehors de la période d’évaluation) | Généralisée (71-100 %) | Inconnue | Faible (peut-être à long terme, > 10 ans) | Les sécheresses extrêmes périodiques pourraient avoir des répercussions sur la végétation fourragère disponible pour le criquet des Îles-de-la-Madeleine. En période de sécheresse extrême, il se peut que les œufs en dormance n’éclosent pas en raison de conditions climatiques inadéquates. Cette menace est inconnue, car on ne connaît pas en détail le cycle biologique particulier de cette espèce. |
| 11.3 | Températures extrêmes | Inconnu | Généralisée (71-100 %) | Inconnue | Modérée (peut-être à court terme, < 10 ans) | Des températures extrêmement froides pourraient avoir des répercussions sur le criquet, de façon directe (mortalité) ou indirecte (diminution de l’abondance des plantes fourragères). Les modèles climatiques pour la région des Maritimes semblent indiquer que le climat deviendra encore plus froid et plus humide, ce qui met de la pression sur les tolérances physiologiques des criquets sur les îles. Cette menace est inconnue. |
| 11.4 | Tempêtes et inondations | Non calculé (en dehors de la période d’évaluation) | Inconnue | Inconnue | Faible (peut-être à long terme, > 10 ans) | Les îles de la Madeleine subissent des épisodes naturels de forts vents et de grosses tempêtes; ces phénomènes peuvent causer l’érosion de l’habitat. |
Les activités de collecte entomologique passées ont été rares aux îles de la Madeleine; des relevés ciblés ont été réalisés aux îles à quelques occasions seulement et pour quelques autres groupes d’insectes (p. ex., Kevan et Vickery, 1978; Hamilton, 2002). Un nombre relativement restreint de spécimens de criquet des Îles-de-la-Madeleine ont été recueillis (tableau 1) (Catling et al., 2013). L’espèce a été décrite en 1978, et 32 spécimens ont été inclus dans la série type, conservée au Musée d’entomologie Lyman (13 spécimens), à la Collection nationale canadienne (CNC) d’insectes, d’arachnides et de nématodes (2 spécimens), à l’Université de Montréal (10 spécimens), à l’Academy of Natural Sciences of Philadelphia (5 spécimens) et au Musée royal de l’Ontario (2 spécimens) (Vickery et Kevan, 1978). Au moment de la rédaction du présent rapport, environ 80 spécimens étaient conservés au Musée Lyman, dont un certain nombre provenaient de la série type d’autres établissements.
Les relevés de criquet réalisés en 1999 et en 2000 ont permis de recueillir huit autres spécimens de criquet des Îles-de-la-Madeleine, qui ont par la suite été utilisés dans une étude génétique de cette espèce et d’espèces étroitement apparentées (voir Chapco et Litzenberger, 2002). Toutefois, à l’heure actuelle, on ignore si ce matériel a été détruit dans le cadre de cette étude. Catling et al. (2013) ont recueilli trois autres spécimens en 2010, dont les deux qui sont présentés aux figures 1 et 2.
Des relevés sur le terrain ont été effectués durant la préparation du présent rapport de situation, du 6 au 12 juillet 2015, pour un total de 40 heures-personne sur 23 sites (20 sites aux îles de la Madeleine). Les relevés ont été réalisés par récolte à la main et par filet fauchoir dans des habitats herbeux dégagés, dans des bordures de routes adjacentes à des zones agricoles, dans des zones dégagées sablonneuses à végétation clairsemée et des milieux boisés dégagés. Tous les criquets recueillis ont été aspirés hors du filet fauchoir et placés dans l’éthanol à 70 % jusqu’à ce qu’ils soient examinés.
Environ 280 criquets ont été recueillis dans l’ensemble des sites en 2015. Les criquets recueillis ont été dénombrés pour chaque site de relevé. Tous les spécimens recueillis étaient immatures; il a donc été impossible d’arriver à une identification fiable à l’aide des clés taxonomiques, qui sont fondées sur les caractéristiques des adultes. Ces spécimens immatures ont été séparés en groupes d’espèces morphologiques, puis ont été divisés en sous-échantillons pour le codage à barres de l’ADN. Le codage à barres de l’ADN produit une courte séquence génétique (c.-à-d. le code à barres de l’ADN) à partir d’une portion standard du génome mitochondrial des spécimens, ce qui permet de comparer ce « code à barres » à celui d’autres espèces ou spécimens figurant dans le système Barcodes of Life Data (BOLD). On présume que chaque espèce possède sa propre séquence de code à barres de l’ADN (traditionnellement, les spécimens biologiques sont identifiés à l’aide des caractéristiques morphologiques). Le code à barres standard pour presque tous les groupes d’animaux est une région de 648 paires de bases du gène mitochondrial c oxidase 1 (« CO1 »). Sur chaque spécimen de criquet sélectionné, une seule patte (ou une portion de patte pour les spécimens plus gros) a été retirée et envoyée à l’Institut de biodiversité de l’Ontario, à Guelph (Ontario), pour le traitement et le séquençage des tissus. Les séquences de code à barres de l’ADN, les images des spécimens et toutes les données concernant la collecte des spécimens ont été téléchargées dans le système Barcodes of Life Data (BOLD), sous le projet « Magdalen Islands Grasshopper » (MAGIG). De plus, on a procédé au codage à barres de l’ADN de deux spécimens adultes récemment recueillis (fournis par Paul Catling); le code à barres de l’ADN de ces spécimens a servi de code à barres standard pour le criquet des Îles-de-la-Madeleine.
Le codage à barres de l’ADN ne remplace pas la taxonomie traditionnelle, mais il peut aider à vérifier l’identification et avoir comme deuxième utilité de déterminer s’il est nécessaire d’investir du temps et des ressources afin d’améliorer la taxonomie d’un groupe d’espèces. Il peut également être utile pour faire une association entre spécimens immatures et adultes. Dans le cas des criquets, le moment où les relevés sont effectués peut mener à des données erronées concernant l’abondance relative, y compris pour le criquet des Îles-de-la-Madeleine. Des relevés antérieurs peuvent avoir été menés après le moment culminant de l’émergence des adultes (Catling et al., 2013). Par conséquent, le codage à barres de l’ADN peut servir d’outil pour les relevés lorsqu’il est difficile d’évaluer à quel moment survient le moment culminant de l’émergence des adultes.
Selon les résultats du codage à barres de l’ADN, au moins sept des 280 spécimens recueillis en 2015 étaient des criquets des Îles-de-la-Madeleine. Les spécimens de criquet des Îles-de-la-Madeleine pour lesquels le codage à barres de l’ADN a été établi ont été capturés uniquement à Cap-aux-Meules (sur l’escarpement rocheux du chemin de la Belle-Anse, près du chemin des Caps) et à l’île du Havre aux Maisons (au bord de la plage) (tableau 1). Ces chiffres représentent moins de 3 % du nombre total de criquets recueillis, ce qui donne à penser que l’espèce est relativement rare. En comparaison, le criquet voyageur (Melanoplus sanguinipes) représentait plus de 80 % de tous les spécimens pour lesquels le codage à barres de l’ADN a été établi. La variation intraspécifique du gène CO1 est considérée comme étant faible, avec un résultat de 0,16 %. Il a été possible d’établir le code à barres de l’ADN de plusieurs autres espèces de criquet du genre Melanoplus recueillis en 2015 aux îles de la Madeleine. Par contre, en raison de la faible variation interspécifique du gène CO1 entre ces espèces et les espèces du continent et en raison des erreurs taxonomiques présumées de la bibliothèque de référence du système Barcodes of Life Data pour les spécimens du genre Melanoplus, il a été impossible de procéder à une identification exacte de ces spécimens en se fondant uniquement aux codes à barres de l’ADN. Cependant, il a été déterminé que ces spécimens n’étaient pas des criquets des Îles-de-la-Madeleine.
Habitat
Besoins en matière d’habitat
Étant donné le faible nombre de relevés et d’activités de collecte, il est difficile de caractériser avec exactitude l’habitat de l’espèce. Catling et al. (2013) ont décrit les sites de collecte, en général, comme étant des prés dégagés constitués d’une flore indigène, entre autres des espèces des genres Carex, Juncus et Eupatorium.
Habitat d’alimentation et d’abri
Les spécimens recueillis jusqu’à présent ont été prélevés dans des prés dégagés où la végétation est dense. En 2015, des spécimens immatures ont été recueillis à deux sites qui ont élargi la description de l’habitat, qui inclut désormais des zones herbeuses contenant beaucoup d’herbacées non graminoïdes et d’herbes non indigènes. En 2015, des spécimens immatures de criquet des Îles-de-la-Madeleine ont été recueillis à deux sites situés dans des habitats différents (tableau 1). L’habitat du premier site de collecte, situé à Cap-aux-Meules (sur le chemin de la Belle-Anse, près du chemin des Caps), était un pré herbeux en expansion adjacent à un escarpement sablonneux (à environ 30 m au-dessus du niveau de la mer). L’habitat de l’île du Cap aux Meules état situé sur des sols sablonneux bien drainés et était constitué d’une diversité moyenne de plantes herbacées basses, comme le liondent d’automne (Leontodon autumnalis), le sumac (Rhus spp.), la carotte sauvage (Daucus carota), la renoncule (Ranunculus spp.), la fraise (Fragaria spp.), le trèfle rouge (Trifolium repria), le trèfle rampant (Trifolium repens), la marguerite blanche (Leucanthemum vulgare), l’épervière (Hieracium spp.), le carvi commun (Carum carvi) et bien d’autres espèces de composées jaunes, dont le salsifis (Trapopogon spp.). La vesce jargeau (Vicia cracca), une espèce exotique, était également présente.
Le deuxième site de collecte de 2015 était situé sur l’île du Havre aux Maisons en bordure de la plage, à moins de 5 m au-dessus du niveau de la mer, près d’un habitat de dunes (au-dessus des dunes et de la ligne de marée haute) où l’on retrouvait des plantes herbacées non graminoïdes. Le site contenait un certain nombre des plantes susmentionnées, mais était dominé par la gesse maritime (Lathyrus japonicus), l’ammophile à ligule courte (Ammophila breviligulata) et d’autres herbes hautes.
Sites de ponte et habitat d’hivernage
Le criquet des Îles-de-la-Madeleine hiverne probablement au stade d’œuf (Vickery et Kevin, 1985) dans un substrat sablonneux mou situé dans des zones dégagées à végétation herbacée dense. Le sol des îles de la Madeleine a une forte teneur en sable, auquel s’ajoutent des sédiments de gravier et de grès. Il a été démontré que les sites de ponte choisis par les femelles gravides contribuent à la bonne condition de la progéniture chez les criquets (Fielding, 2011).
Habitat improbable
Dans le cadre des relevés de 2015, aucun criquet n’a été observé dans les communautés d’avant-dunes de sable dégagées et les communautés végétales éparses de milieux dunaires. L’absence de criquet des Îles-de-la-Madeleine et d’autres criquets dans cet habitat peut être indirectement mesurée par les résultats d’une étude sur le régime alimentaire du Pluvier siffleur (Charadrius melodus) sur les îles de la Madeleine (Shaffer et Laport, 1994). L’absence de ce criquet, et de tout autre orthoptère, dans les fientes de cet oiseau donne à penser qu’un régime alimentaire composé de criquets n’est pas commun dans les habitats côtiers d’alimentation de cet oiseau de rivage (Shaffer et Laport, 1994). Cependant, la présence d’insectes au vol rapide, comme les cicindèles (sous-famille des Cicindelinae) dans le régime alimentaire de certains Pluviers siffleurs (Shaffer et Laport, 1994; Majka et Shaffer, 2008) semble indiquer que les criquets, surtout les femelles gravides qui se déplacent lentement, sont probablement des proies. De plus, aucun criquet des Îles-de-la-Madeleine n’a été observé dans les milieux boisés denses.
Tendances en matière d’habitat
La flore et la faune des îles de la Madeleine sont jugées stables depuis les 8 000 à 11 000 dernières années (Hamilton, 2002). À une échelle de temps géologique, l’archipel peut avoir déjà fait partie d’un refuge glaciaire plus gros durant la période de glaciation du Wisconsin (Hamilton, 2002). La majorité des îles de la Madeleine sont maintenant habitées, et si la population humaine résidente est demeurée relativement stable (de 12 000 à 14 000 personnes) depuis les années 1960 (Fortin et Larocque, 2003), la population des îles augmente pour atteindre 50 000 personnes durant la saison touristique estivale.
Historiquement, et selon les preuves archéologiques, les îles de la Madeleine ont été visitées par les Micmacs, qui allaient probablement y chasser l’abondante sous-population de morses (Odobenus rosmarus) qui résidait sur les îles à une certaine époque. Avant les premiers établissements permanents, la végétation des îles de la Madeleine était principalement constituée de forêts denses de conifères (c. à d. de sapins baumiers [Abies balsamea] et d’épinettes [Picea spp.]), bien que ces arbres étaient probablement rabougris et tordus par le vent, formant une « forêt de lutins ». Des zones marécageuses et des crêtes de sable basses étaient intercalées entre ces forêts (Hamilton, 2002). Les six îles principales de l’archipel ont été victimes de déforestation à compter de la fin des années 1800 et du début des années 1900. À l’heure actuelle, les images de GoogleEarth (2015) montrent que la majorité de la végétation sur l’île du Havre Aubert, l’île du Havre aux Maisons, l’île du Cap aux Meules et l’île d’Entrée a été déboisée et est maintenant constituée de prés dégagés ou de pâturages; il reste peu de forêts de conifères. Le criquet des Îles-de-la-Madeleine n’a pas été observé dans des forêts de conifères et, par conséquent, l’accroissement des prés dégagés pourrait avoir été bénéfique pour l’espèce. Par la suite, une portion de ces zones déboisées a été convertie en grands terrains résidentiels ruraux (Hamilton, 2002).
Biologie
Les renseignements sur la biologie et l’histoire naturelle du criquet des Îles-de-la-Madeleine présentés ici sont un résumé des références générales sur la biologie des criquets (Preston-Mafham, 1990) et d’une espèce qui lui est étroitement apparentée, le criquet boréal (Vickery et Kevan, 1985).
Cycle vital et reproduction
Le criquet des Îles-de-la-Madeleine possède un cycle vital typique du genre Melanoplus (Pladt, 2002) et de la majorité des autres criquets. L’espèce hiverne au stade d’œuf (Vickery et Kevan, 1985). Chez une espèce similaire, le criquet boréal, l’œuf reste en diapause durant deux ans (Pladt, 2002; Fielding, 2008), et cette durée pourrait être la même pour le criquet des Îles-de-la-Madeleine.
Comme tous les insectes qui se développent par métamorphose graduelle, plusieurs stades nymphaux se succèdent et, au fil des mues, les larves de chaque stade nymphal grossissent et présentent des caractéristiques morphologiques adultes plus prononcées. Le criquet boréal traverse cinq stades nymphaux, et c’est probablement aussi le cas pour le criquet des Îles-de-la-Madeleine. Immédiatement après l’éclosion, la larve du criquet quitte le sol et grimpe sur des surfaces surélevées avoisinantes où elle effectue sa première mue; elle entre alors dans son premier stade nymphal. Durant ce premier stade, la nymphe est très petite (environ de 2 à 3 mm de longueur) et est dépourvue de bourgeons alaires apparents. Au fil des stades nymphaux (immatures), les bourgeons alaires se développent. La taille (et la forme) des bourgeons alaires sont des indicateurs distinctifs qui permettent de déterminer le stade de développement.
Le premier stade nymphal correspond au début de l’alimentation et de la croissance; la larve mange et grandit, puis elle mue quelques jours plus tard. Au deuxième stade nymphal, le corps de la larve est un peu plus long, mais les bourgeons alaires se développent peu et consistent en deux minuscules rabats. Une fois de plus, une mue survient après quelques jours (le nombre de jours varie selon la température), et la larve du troisième stade est plus longue et ses bourgeons alaires se développent de façon plus prononcée (c.-à-d. une nervation est visible). La nymphe du quatrième stade est encore plus grosse et possède de petits bourgeons alaires triangulaires qui présentent des lignes saillantes claires (nervures) s’étendant par-dessus le premier segment de l’abdomen. Ces nymphes ont un corps plus charnu. Ce stade correspond à la principale période d’alimentation chez le criquet. Le dernier stade immature (le cinquième) survient lorsque l’individu est presque mature. Les bourgeons alaires sont beaucoup plus longs et ressemblent à une aile entièrement développée. Ce stade dure généralement environ une semaine, après quoi le criquet muera en un adulte possédant des ailes totalement développées (une journée entière est nécessaire pour que les ailes durcissent). Ces individus se reproduisent dans un délai d’une semaine. Une fois que la diapause de l’œuf prend fin, le développement jusqu’au stade adulte prend généralement environ un mois, selon l’espèce et les conditions météorologiques locales.
Les criquets des Îles-de-la-Madeleine adultes sont actifs de la fin de juillet jusqu’à la mi-septembre (Vickery et Kevan, 1985). Chez le criquet boréal, la femelle commence à pondre de 8 à 10 jours après l’accouplement (Pladt, 2002), mais on ignore quel est ce délai chez le criquet des Îles-de-la-Madeleine. Les femelles pondent leurs œufs en oothèques de 30 à 50 œufs, à une profondeur de 1 à 3 cm sous la surface du substrat, ou contre des roches, des racines de plantes ou d’autres microsites similaires. Selon l’espèce et la géographie, les œufs éclosent à la fin du printemps si le sol est chaud, ou au début de l’été (à la mi-juin). Au sein d’une même sous-population, l’éclosion ne se produit pas de manière simultanée et peut s’étendre sur une période de deux semaines ou plus. Avec une si petite quantité de données, il est difficile d’évaluer la période d’activité maximale des adultes de cette espèce.
Physiologie et adaptabilité
Aucune donnée n’est disponible sur la physiologie et l’adaptabilité propres au criquet des Îles-de-la-Madeleine. Il est probable que les sous-populations de criquet des Îles-de-la-Madeleine survivraient si elles étaient établies dans les régions côtières du continent, bien que la parenté génétique entre cette espèce et le criquet boréal (espèces qui sont géographiquement isolées) puisse mener à une possible hybridation. Cependant, Vickery et Kevan (1985) font observer qu’une telle hybridation est peu probable en raison de la différence entre les genitalia mâles des deux espèces.
Déplacements et dispersion
On en connaît peu sur la capacité de déplacement et de dispersion du criquet des Îles-de-la-Madeleine, mais les adultes sont considérés comme n’étant pas dotés d’une puissante capacité de vol (Vickery et Kevan, 1985). À l’heure actuelle, sa présence a été signalée sur six des principales îles interreliées des îles de la Madeleine et, plus récemment, sur l’île d’Entrée, située à 4 km des îles principales. Au-delà de cette île, la masse de terre la plus près est l’Île-du-Prince-Édouard (à 105 km plus au sud), et la dispersion vers cette masse de terre est considérée comme improbable. On ignore si les adultes ou les stades immatures peuvent migrer d’une île à l’autre (même si elles sont reliées par des bancs de sable).
Relations interspécifiques
Le criquet des Îles-de-la-Madeleine peut être la proie d’un large éventail de vertébrés et d’invertébrés (Preston-Mafham, 1990); la proportion de prédation de chaque taxon dépend de l’habitat et du lieu géographique. Les prédateurs vertébrés sont considérés comme le principal facteur ayant stimulé l’évolution de certaines défenses chimiques chez les orthoptères (Preston-Mafham, 1990). Par conséquent, on peut supposer que les vertébrés sont des prédateurs importants dans la majorité des milieux terrestres.
Il y a une faible diversité de vertébrés terrestres sur les îles de la Madeleine, et l’on ignore quelles sont les répercussions de la prédation par les vertébrés sur le criquet des Îles-de-la-Madeleine. On ignore également dans quelle mesure les mammifères indigènes sont des prédateurs du criquet des Îles-de-la-Madeleine; un tel comportement de prédation a été observé uniquement chez quelques espèces de mammifères terrestres (Cameron, 1962), dont deux sont indigènes : le campagnol des prés (Microtus pennsylvanicus) et la souris sylvestre (Peromyscus maniculatus). En raison de renseignements contradictoires, il est difficile de déterminer s’il faut considérer les chauves-souris comme des espèces indigènes ou exotiques; cependant, les espèces de chauves-souris de l’Est du Canada ne sont pas considérées comme des prédateurs importants des criquets (Brigham, comm. pers., 2015). Le campagnol des prés n’est probablement pas lui non plus un prédateur important des criquets. Toutefois, la souris sylvestre inclut les criquets dans son régime alimentaire. Le renard roux (Vulpes vulpes) n’est pas considéré comme étant une espèce indigène des îles de la Madeleine, malgré des signalements confirmés de sa présence sur les îles remontant à la fin des années 1800 (Cory, 1878). Le renard roux est un prédateur connu des criquets (Catling, 1988; Read et Bowen, 2001). De plus, il n’y a aucun amphibien et aucun reptile sur les îles de la Madeleine.
Les oiseaux sont des prédateurs importants des criquets, et environ 320 espèces d’oiseaux habitent sur les îles de la Madeleine; une telle diversité signifie qu’il y a une foule de prédateurs possibles (Preston-Mafham, 1990). Aucun orthoptère n’a été trouvé dans le cadre d’une étude du régime alimentaire (fondée sur l’examen du contenu des fientes) du Pluvier siffleur sur les îles de la Madeleine (Shaffer et Laport, 1994). Ces données donnent également à penser que les criquets ne sont peut-être pas abondants dans l’habitat du pluvier. Le régime alimentaire du Pluvier siffleur inclut sans doute plusieurs arthropodes très mobiles (Shaffer et Laport, 1994; Majka et Shaffer, 2008), ce qui porte à croire que ces oiseaux seraient capables de capturer des criquets immatures et adultes s’ils étaient présents et abondants.
La présence du criquet des Îles-de-la-Madeleine a été observée dans des boulettes de régurgitation du Courlis corlieu (Numenius phaeopus) sur la Grosse Île. Le Courlis corlieu et les autres courlis sont des prédateurs généralistes des insectes (Skeel et Mallory, 1996) et ne sont pas des prédateurs spécialisés des criquets. Cependant, les criquets constituent peut-être une importante source de nourriture éphémère pour le Courlis corlieu dans les milieux herbeux, comme les Prairies canadiennes (Ray Poulin, comm. pers., 2016).
Un grand nombre de prédateurs généralistes des arthropodes se nourrissent d’orthoptères, dont les araignées (Aranae), les coléoptères prédateurs (p. ex. cicindèles et carabes [Coleoptera : Carabidae]), les asiles (Diptera : Asilidae), les fourmis et un grand nombre de guêpes prédatrices (Hymenoptera) (Preston-Mafham, 1990). Les larves des espèces du genre Epicauta, dans la famille des méloés (Meloidae), sont des prédateurs spécialisés des œufs de criquet (Pinto, 1991). À certains endroits, elles ont des répercussions importantes et peuvent détruire près de 10 % des oothèques de criquets (Parker et Wakeland, 1957). On ignore si elles ont des répercussions importantes en tant que prédateur du criquet des Îles-de-la-Madeleine, car aucune espèce d’Epicauta n’a été signalée sur les îles de la Madeleine.
Chez les criquets, les relations interspécifiques particulières sont établies avec leurs parasites et parasitoïdes. Plusieurs groupes de Diptera (familles des Sarcophagidae, des Anthomyiidae, des Tachinidae et des Nemestrinidae) et d’Hymenoptera (p. ex. les Scelionidae) sont des parasitoïdes des criquets (Smith, 1940; Smith et Finlayson, 1950; York et Prescott, 1952; Rees, 1985; Przybyszewski et Capinera, 1991; Laws et Joern, 2012).
Les acariens sont également des parasites importants des criquets (Branson, 2003), et sont considérés comme étant potentiellement importants pour les futures initiatives de lutte contre les ravageurs visant des espèces problématiques (Belovsky et al., 1997). On trouve couramment des acariens sur les ailes des criquets adultes, mais ils semblent avoir peu de répercussions. Par contre, certaines espèces d’acariens sont des prédateurs des œufs de criquet et peuvent avoir une incidence sur les sous-populations de criquets (Belovsky et al., 1997). L’effet des acariens sur la majorité des espèces de criquet, y compris le criquet des Îles-de-la-Madeleine, est peu documenté.
Biron et ses collaborateurs (2005) signalent qu’un ver capillaire juvénile (Nematomorpha) parasite les orthoptères et modifie le comportement des criquets adultes, les incitant à sauter dans l’eau douce, ce qui les tue. Divers nématodes sont des parasites connus des criquets, et certains sont utilisés comme agents de lutte biologique (Baker et Capinera, 1997). Toutefois, aucune documentation précise ne confirme la présence de ces parasites chez le criquet des Îles-de-la-Madeleine.
Les pathogènes fongiques et bactériens sont des facteurs importants qui ont des effets sur les orthoptères (Bucher et Stephens, 1957; Streett et McGuire, 1990; Glogoza et Weiss, 2010), et une espèce (probablement celle qui a les répercussions les plus importantes), le champignon Entomopthora grylli, semble être un pathogène spécialisé des criquets et des grillons (Pickford et Riegert, 1964; Erlandson et al., 1988). Les stades nymphaux semblent être plus susceptibles de contracter une infection fongique que les adultes (Kistner et Belovsky, 2013). Un autre pathogène fongique pourrait avoir un effet important : Nosema locustae (Ewen et Mukerji, 1980). Toutefois, la situation de ces pathogènes chez les sous-populations de criquet des Îles-de-la-Madeleine n’est pas documentée.
Taille et tendances de la population
Activités et méthodes d’échantillonnage
Les relevés visant le criquet des Îles-de-la-Madeleine ont été axés sur le signalement de nouvelles sous-populations, sur les renseignements concernant l’histoire naturelle et l’habitat et sur la collecte de matériel pour les études génétiques. Les relevés ont été effectués principalement le long de transects dans des parcelles d’habitat convenable et à l’aide d’un filet fauchoir, à la recherche de criquets. Catling et al. (2013) ont indiqué qu’il était difficile de faire correspondre les activités de collecte avec le moment culminant de l’émergence des adultes, ce qui porte à croire que le succès de détection pourrait varier considérablement. Les activités de collecte ont été effectuées du 8 au 15 juillet, au moment où la majorité des espèces de criquet présentes sur les îles étaient immatures (nous avons recueilli des nymphes des stades un à cinq). En raison des problèmes d’identification des criquets immatures, la technique du codage à barres de l’ADN a été utilisée pour identifier les espèces recueillies aux divers sites et en estimer l’abondance.
Abondance
Il existe peu de données qui permettent d’estimer l’abondance du criquet des Îles-de-la-Madeleine sur les sites existants. Dans la plupart des cas, un ou deux individus ont été observés ou recueillis. Les résultats du codage à barres de l’ADN semblent indiquer que le criquet des Îles-de-la-Madeleine est relativement rare par rapport aux autres espèces des îles; en effet, cette espèce constituait moins de 3 % de tous les spécimens de Melanoplus observés ou recueillis.
Fluctuations et tendances
Les fluctuations naturelles des sous-populations de criquet sont attribuables à des facteurs comme les parasites, les prédateurs, l’abondance de nourriture et les conditions météorologiques des années précédentes. Pour certaines espèces, les fluctuations de la taille des populations peuvent également être attribuables à des variations du taux de survie des œufs en fonction du site de ponte choisi (Fielding, 2011). Les renseignements disponibles ne permettent pas d’évaluer les fluctuations ou les tendances des sous-populations chez le criquet des Îles-de-la-Madeleine en particulier. Toutefois, l’espèce peut subir des fluctuations extrêmes, si l’on se fie aux renseignements recueillis chez son taxon sœur, le criquet boréal, ainsi que chez d’autres espèces de criquet du genre Melanoplus (Mulkern, 1980).
Immigration de source externe
Le criquet des Îles-de-la-Madeleine est endémique aux îles de la Madeleine; l’immigration de source externe du Canada est donc impossible.
Menaces et facteurs limitatifs
Le calculateur des menaces de l’Union internationale pour la conservation de la nature – Partenariat pour les mesures de conservation (2006) (IUCN-CMP) a été utilisé pour repérer et classer les menaces qui pèsent sur le criquet des Îles-de-la-Madeleine (Salafsky et al., 2008; Master et al., 2009). L’impact global des menaces pour l’espèce est faible.
Les menaces ayant un impact faible sont les routes et les voies de circulation, où les véhicules peuvent causer de la mortalité chez les criquets qui traversent les routes ou qui tentent de pondre dans le substrat des routes de terre; les activités récréatives, notamment la conduite de véhicules tout terrain dans les prés; ainsi que les espèces envahissantes et d’autres espèces problématiques reconnues comme étant des prédateurs du criquet.
Menace 4. Corridors de transport et de service (impact faible)
4.1 Routes et voies ferrées
Les véhicules qui circulent sur les routes peuvent écraser les criquets et avoir un impact sur les sous-populations de criquet qui utilisent l’habitat adjacent au réseau routier (Catling et al., 2013). De plus, les femelles gravides, qui se déplacent lentement, ont tendance à sauter pour se déplacer d’un endroit à un autre et à faire une pause entre ces sauts; durant cette pause, elles sont susceptibles d’être écrasées par un véhicule. Le substrat de nombreuses routes moins utilisées sur les îles de la Madeleine possède une forte teneur en sable, mélangé à du gravier. On sait que les femelles pondent dans ces substrats mous, ce qui rend les œufs et les femelles plus vulnérables à la mortalité routière. La circulation routière est plus dense durant les mois d’été, lorsque les touristes viennent gonfler la population des îles.
La perspective de travaux d’accroissement des bordures de route et d’élargissement des routes est improbable, car la majorité de cette infrastructure routière est en place depuis des dizaines d’années. Par contre, les routes font continuellement l’objet de travaux de réparation, à la suite de grosses tempêtes ou d’inondations.
Menace 6. Intrusions et perturbations humaines (impact faible)
6.1 Activités récréatives
Les îles de la Madeleine sont une destination touristique populaire durant les chauds mois d’été, soit de juin à septembre, et l’économie des îles est liée au récréotourisme. Le récréotourisme a beaucoup augmenté au cours des 40 dernières années. Dans les années 1970, entre 9 000 et 18 000 touristes venaient visiter les îles chaque année; dans les années 1990, ce nombre a augmenté pour atteindre 30 000 visiteurs par année; en 2002, il y a eu 54 000 visiteurs; de 2003 à 2005, le nombre de visiteurs était supérieur à 50 000 par année; enfin, en 2012, on a dénombré pas moins de 62 200 visiteurs (Tourisme Îles de la Madeleine, 2012). À compter de 2012, des croisières internationales ont commencé à faire une halte aux îles de la Madeleine, ce qui a fait croître le nombre de touristes qui visitent les îles chaque été.
La plupart des activités récréatives ont lieu dans les écosystèmes de plages côtiers, bien qu’à certains endroits, pour accéder aux plages, il faut passer par les prés dégagés où des criquets sont susceptibles d’être présents. Les îles de la Madeleine possèdent un vaste réseau de parcs de prés dégagés, de belvédères et d’escarpements protégés qui sont des destinations populaires pour la randonnée et les promenades avec un chien. Certains de ces milieux ne sont pas protégés et sont vulnérables à des activités plus intenses, comme le vélo de montagne. Le piétinement et l’accroissement du nombre de randonneurs sur les sentiers d’un certain nombre de ces sites font en sorte que les criquets risquent davantage d’être écrasés (Catling et al., 2013). On sait que le criquet boréal passe la majorité de sa vie adulte à lézarder (se chauffer au soleil) sur des sols nus, et si le criquet des Îles-de-la-Madeleine a un comportement similaire, les répercussions sur les populations locales seront bien supérieures aux estimations actuelles (p. ex. les adultes pourraient utiliser les sentiers dégagés et les routes pour lézarder ou s’en servir comme site de ponte).
À l’heure actuelle, l’impact global des activités récréatives est considéré comme étant faible, mais ses activités pourraient constituer une plus grande menace si le tourisme se développe, surtout le tourisme lié aux bateaux de croisières internationales.
Menace 8. Espèces et gènes envahissants ou autrement problématiques (impact faible)
8.1 Espèces exotiques (non indigènes) envahissantes
L’Étourneau sansonnet (Sturnus vulgaris) pourrait constituer une menace à impact faible pour le criquet des Îles-de-la-Madeleine (Catling et al., 2013). En effet, cette espèce est présente en grand nombre sur les îles et ne cesse de s’y multiplier, et l’on sait qu’elle se nourrit de criquets et d’autres insectes (Wood, 1973). D’autres espèces d’étourneaux sont reconnues comme étant des prédateurs importants des criquets, et il a même été suggéré de s’en servir comme possible agent de lutte biologique (Ji et al., 2008). La propagation et l’envahissement graduels de plantes exotiques pourraient modifier l’abondance et la composition des plantes à fleurs qui sont disponibles en tant que source de nourriture pour les criquets.
Les chats domestiques ou errants (Felis catus) peuvent également être des prédateurs importants des criquets (Errington, 1936; McMurry et Sperry, 1941; Konecny, 1987; Catling, 1988; Read et Bowen, 2001), et la fréquence de prédation des criquets par les chats est plus élevée au cours des mois d’été, lorsque les criquets sont plus gros et plus abondants (Molsher et al., 1999).
D’autres mammifères problématiques ont été introduits et se sont établis sur les îles de la Madeleine. Il s’agit notamment du rat surmulot (Rattus norvegicus) (Cameron, 1962), de l’écureuil roux (Sciurus vulgaris), du lièvre d’Amérique (Lepus americanus) (Dinets, 2015) et du vison d’Amérique (Neovison vison) (Tourisme Îles de la Madeleine, 2016). Le coyote (Canis latrans) a été observé pour la première fois sur les îles en 2001 (Naughton, 2012).
Des insectes envahissants, comme la fourmi rouge (Myrmica rubra), pourraient avoir un impact sur le criquet des Îles-de-la-Madeleine, en se nourrissant d’œufs et d’adultes lorsque les femelles gravides sont lentes et lourdes. On sait que la fourmi supplante et réduit les assemblages fauniques d’invertébrés à proximité du nid et sur le territoire d’alimentation. À l’heure actuelle, la fourmi rouge n’a pas été signalée sur les îles de la Madeleine. Il existe un certain potentiel d’introduction d’une espèce parente, le criquet boréal, par transport anthropique (p. ex. par le transport de boutures végétales). L’introduction de cette espèce de criquet mènerait probablement à un croisement, à une hybridation et à une compétition avec le criquet des Îles-de-la-Madeleine. D’autres relevés d’insectes dans les habitats où la présence du criquet est connue sont nécessaires afin de détecter tout changement.
Autres facteurs
Les facteurs limitatifs qui s’appliquent au criquet des Îles-de-la-Madeleine sont notamment les suivants :
Capacité de déplacement limitée
Les femelles ne sont pas dotées d’une puissante capacité de vol et leurs mouvements plus lents peuvent rendre l’espèce vulnérable au piétinement et à la prédation.
Courte saison de croissance
Les îles de la Madeleine offrent une courte saison de croissance; en effet, durant les mois de juillet et août, on y enregistre des températures maximales de 29 °C et 31 °C et des températures minimales de 7,5 °C et 6,6 °C, respectivement. Les conditions climatiques difficiles réduisent le temps dont dispose le criquet pour compléter son cycle vital, y compris pour se disperser dans de nouveaux milieux.
Régimes climatiques fluctuants
Les criquets dépendent de la température ambiante pour réchauffer leur corps et se déplacer, et la température estivale fraîche et brumeuse n’est peut-être pas optimale pour le déplacement des criquets. Les variations annuelles des régimes climatiques pourraient avoir une incidence sur le nombre d’individus des sous-populations en raison d’une diminution des activités d’alimentation lors de conditions climatiques extrêmes. Des vents forts peuvent également accroître le niveau d’érosion à des endroits où le sol est exposé (p. ex. répercussions sur les œufs de criquet) et réduire les comportements des déplacements d’alimentation et de reproduction du criquet.
Nombre de localités
Il est difficile d’évaluer le nombre de localités occupées par le criquet des Îles-de-la-Madeleine. L’espèce se déplace lentement, bien qu’elle soit probablement assez mobile pour repeupler les habitats convenables au fil du temps. Les principales menaces, celles qui sont les plus graves et les plus plausibles pour l’espèce, sont : 4.1 la mortalité routière; 6.1 les activités récréatives qui ont un impact négatif sur l’habitat du criquet ou qui entraîne sa mortalité directe, et 8.1 la prédation du criquet. Ces menaces varient sur les différentes îles de la Madeleine, et par conséquent le nombre de localités se situe probablement dans une fourchette de sept îles (îles où la présence a été confirmée) à 14 îles (l’ensemble des îles et des îlets de l’archipel).
Protection, statuts et classements
Statuts et protection juridiques
À l’heure actuelle, aucune loi fédérale ou provinciale ne protège spécifiquement le criquet des Îles-de-la-Madeleine, n’atténue les menaces pour les criquets ou ne protège l’habitat de ces espèces.
La présence du criquet des Îles-de-la-Madeleine ou d’un habitat convenable pour cette espèce est possible dans la Réserve nationale de faune de la pointe de l’Est, bien que cette réserve nationale de faune n’ait pas fait l’objet d’un inventaire exhaustif visant cette espèce. Si l’espèce était repérée dans cette réserve nationale de faune, elle serait protégée sous le régime du Règlement sur les réserves d’espèces sauvages (C.R.C., ch. 1609) du gouvernement du Canada.
Au Québec, cette espèce n’est pas inscrite à la liste de la Loi sur les espèces menacées ou vulnérables (RLRQ, c. E-12.01) (LEMV) et ne bénéficie pas d’une protection au titre de la Loi sur la conservation et la mise en valeur de la faune (RLRQ, c. C -61.1) (LCMVF). De plus, cette espèce ne figure pas sur la liste des espèces susceptibles d’être désignées menacées ou vulnérables. Cette liste est établie conformément à la Loi sur les espèces menacées ou vulnérables (RLRQ, c. E-12.01) (LEMV).
Statuts et classements non juridiques
Les cotes de conservation attribuées au criquet des Îles-de-la-Madeleine sont les suivantes :
- Cote mondiale : G2 (en péril) (dernière mise à jour : décembre 2000)
- Cote nationale : N2 (en péril) (dernière mise à jour : décembre 2000)
- Cote provinciale : Québec S2 (en péril)
Protection et propriété de l’habitat
La Société de conservation des Îles-de-la-Madeleine (SCÎM) est une organisation non gouvernementale (ONG) de conservation établie aux îles de la Madeleine. Cette ONG travaille avec des propriétaires fonciers privés à des initiatives visant à protéger les espèces en péril et l’habitat et à préserver les écosystèmes naturels des îles de la Madeleine (Déraspe, comm. pers., 2015). Jusqu’à présent, l’organisation a protégé plus de 222 hectares d’habitat aux quatre coins de l’archipel (SCÎM, 2015); on ignore toutefois si le criquet est présent dans cet habitat. Attention FragÎles est également une organisation non gouvernementale de conservation établie aux îles de la Madeleine, qui travaille à des projets de collaboration avec les citoyens locaux, les planificateurs et les visiteurs afin d’assurer la protection des éléments des îles qui revêtent une valeur écologique (Attention FragÎles, 2010).
Environ 30 % de la superficie terrestre totale des îles de la Madeleine est constituée de terres privées, y compris les routes principales et les bancs de sable qui relient les îles ensemble. On ignore dans quelle mesure cet habitat est convenable pour le criquet des Îles-de-la-Madeleine ou si cette espèce y est présente.
Remerciements
Les personnes suivantes ont transmis des connaissances et fourni de précieux renseignements et avis : Paul Catling, Véronique Déraspe, Nathalie Desrosiers, Isabelle Gauthier, Mathieu Morin, John Klymko, Rémi Hebert et Andy Hamilton. Les auteurs du rapport souhaitent également remercier le musée royal de la Saskatchewan (Royal Saskatchewan Museum), qui a consacré du temps et des ressources pour le parachèvement du présent rapport, notamment une partie du financement requis pour le codage à barres de l’ADN des spécimens.
Les auteurs du rapport souhaitent remercier Marie Fortier d’avoir obtenu les permis nécessaires pour effectuer les relevés dans la Réserve nationale de faune de la pointe de l’Est. Ils tiennent également à remercier les conservateurs de spécimens et les gestionnaires des collections de leur avoir donné accès aux spécimens. Merci à Paul Grant (coprésident du Sous-comité des spécialistes des arthropodes) pour son soutien rédactionnel et technique, à Angèle Cyr (Secrétariat du COSEPAC) pour son soutien technique et à Jenny Wu (Secrétariat du COSEPAC) pour son soutien à la cartographie.
La photographie en couverture a été prise le 11 août 2010 par Paul Catling : Criquet des Îles-de-la-Madeleine femelle photographié dans un pré dégagé sur l’île du Havre-aux-Maisons, aux Îles-de-la-Madeleine (Québec), avec l’île d’Entrée en arrière-plan. Elle a été initialement publiée dans Catling et al. (2013).
Experts contactés
Catling, Paul. Chercheur. Collection nationale canadienne d’insectes, d’arachnides et de nématodes, Agriculture et Agroalimentaire Canada. Ottawa (Ontario).
Déraspe, Véronique. Directrice, Société de conservation des Îles-de-la-Madeleine (SCÎM), 350, chemin Principal, bureau 202, Cap-aux-Meules (Québec).
Desrosiers, Nathalie. Ministère des Forêts, de la Faune et des Parcs, Québec.
Fortier, Marie. Spécialiste des aires protégées, Environnement Canada, Québec (Québec).
Gauthier, Isabelle. Coordonnatrice provinciale des espèces fauniques menacées et vulnérables, ministère des Forêts, de la Faune et des Parcs, Québec.
Hamilton, Andrew. Chercheur. Collection nationale canadienne d’insectes, d’arachnides et de nématodes, Agriculture et Agroalimentaire Canada. Ottawa (Ontario).
Morin, Mathieu. Direction de la gestion de la faune de la Gaspésie-Îles-de-la-Madeleine, ministère des Forêts, de la Faune et des Parcs, Sainte-Anne-des-Monts (Québec).
Lonsdale, Owen. Gestionnaire des collections. Collection nationale canadienne d’insectes, d’arachnides et de nématodes, Agriculture et Agroalimentaire Canada. Ottawa (Ontario).
Wheeler, Terry. Musée d’entomologie Lyman, Département des sciences des ressources naturelles, Université McGill, Campus Macdonald, Montréal (Québec).
Sources d’information
Arnett, R.H. 2000. American Insects: A Handbook of the Insects of America North of Mexico. CRC Press.
Attention FragÎles. 2010. Fighting together against erosion. When the citizens of the Magdalen Islands each do their part to slow erosion of the dunes [PDF, 4.04 Mo]. Available at [consulté le 15 décembre 2015]. (Également disponible en français : Attention FragÎles. 2010. Lutter ensemble contre l’érosion. Quand chaque citoyen et chaque citoyenne des Îles-de-la-Madeleine mettent leur grain de sable pour ralentir l’érosion des dunes [PDF, 4.64 Mo].)
Attention FragÎles. 2015. Consultations publiques : Évaluation environnementale stratégique sur l’ensemble de la filière des hydrocarbures. Présenté au ministère de l’Énergie et des Ressources naturelles [PDF, 890 Ko]. 18 novembre 2015. Site Web : [consulté le 14 décembre 2015]
Baker, G.L. et J.L. Capinera. 1997. Nematodes and nematomorphs as control agents of grasshoppers and locusts. Memoirs of the Entomological Society of Canada 171: 157-211.
Belovsky, G.E., D. Branson, J. Chase, J. Barker et G. Hammond. 1997. Mites and Nematode Parasites of Grasshoppers. In: Grasshopper Integrated Pest Management User Handbook. USDA/APHIS/PPQ, Tech. Bull. 1809: 1-3, Boise, ID.
Bernatchez P., C. Fraser, S. Friesinger, Y. Jolivet, S. Dugas, S. Drejza et A. Morissette 2008. Sensibilité des côtes et vulnérabilité des communautés du golfe du Saint-Laurent aux impacts des changements climatiques. Laboratoire de dynamique et de gestion intégrée des zones côtières, Université du Québec à Rimouski. Rapport de recherche remis au Consortium OURANOS et au FACC, 256 pages.
Biron, D.G., L. Marché, F. Ponton, H.D. Loxdale, N. Galéotti, L. Renault, C. Joly et F. Thomas. 2005. Behavioural manipulation in a grasshopper harbouring hairworm: a proteomics approach. Proceedings of the Royal Society of London B: Biological Sciences 272(1577): 2117-2126.
Branson, D.H. 2003. Effects of a parasite mite on life-history variation in two grasshopper species. Evolutionary Ecology Research 5: 397-409.
Brigham, M. 2015. University of Regina. Communication personnelle à Cory Sheffield.
Bucher, G.E. et J.M. Stephens. 1957. A disease of grasshoppers caused by the bacterium Pseudomonas aeruginosa (Schroeter) Migula. Canadian Journal of Microbiology 3(4): 611-625.
Cameron, A.W. 1962. Mammalian zoogeography of the Magdalen Islands Archipelago, Québec. Journal of Mammalogy 43: 505-514.
Catling, P.C. 1988. Similarities and contrasts in the diets of foxes, Vulpes vulpes, and cats, Felis catus, relative to fluctuating prey populations and drought. Australian Wildlife Research 15: 307-317.
Catling, P.M., D.F. McAlpine, C.I. Adam, G. Belliveau, D. Doucet, A.D. Fairweather et A.W. Thomas. 2013. New and noteworthy records of Orthoptera and allies in the Maritimes and the Îles-de-la-Madeleine, Québec. Canadian Field-Naturalist 127: 332-337.
Chapco, W. et G. Litzenberger. 2002. A molecular phylogenetic study of two relict species of melanopline grasshoppers. Genome 45: 313-318.
Dinets, V. 2015. Peterson Field Guide to Finding Mammals in North America. Houghton Mifflin Harcourt Publishing Company.
Erlandson, M.A., D.L. Johnson et O.O. Olfert. 1988. Entomophaga grylli (Fresenius) infections in grasshopper (Orthoptera: Acrididae) populations in Saskatchewan and Alberta, 1985–1986. Canadian Entomologist 120: 205-209.
Errington, P.L. 1936. Notes on food habits of southern Wisconsin house cats. Journal of Mammalogy, 17: 64-65.
Ewen, A.B. et M.K. Mukerji. 1980. Evaluation of Nosema locustae (Microsporidia) as a control agent of grasshopper populations in Saskatchewan. Journal of Invertebrate Pathology 35: 295-303.
Fielding, D.J., 2008. Diapause traits of Melanoplus sanguinipes and Melanoplus borealis (Orthoptera: Acrididae). Annals of the Entomological Society of America 101: 439-448.
Fielding, D.J. 2011. Oviposition site selection by the grasshoppers Melanoplus borealis and M. sanguinipes (Orthoptera: Acrididae). Journal of Orthoptera Research 20: 75-80.
Fielding, D.J. et L.S. Defoliart. 2010. Embryonic developmental rates of northern grasshoppers (Orthoptera: Acrididae): implications for climate change and habitat management. Environmental Entomology 39:1643-1651.
Fortin, J.-C. et P. Larocque. 2003. Histoire des Îles-de-la-Madeleine. Presses de l’Université Laval.
Glogoza, P.A. et M.J. Weiss. 2010. Grasshopper biology and management [PDF, 70.0 Ko]. Manuscrit inédit.
Hamilton, K.G.A. 2002. Îles-de-la-Madeleine (Magdalen Is.): a glacial refugium for short-horned bugs (Homoptera: Auchenorrhyncha)? Le Naturaliste Canadien 126: 25-40.
Ji, R., S.J. Simpson, F. Yu, Q.X. He et C.J. Yun. 2008. Diets of migratory rosy starlings (Passeriformes: Sturnidae) and their effects on grasshoppers: implications for a biological agent for insect pests. Biological Control 46: 547-551.
Kevan, D.K.M. et V.R. Vickery. 1978. The orthopteroid insects of the Magdalen Islands with notes from adjacent regions [Canada]. Annals of the Entomological Society of Québec 22: 193-204.
Kistner, E.J. et G.E. Belovsky. 2013. Susceptibility to disease across developmental stages: examining the effects of an entomopathogen on a grasshopper (Orthoptera: Acrididae) pest. Journal of Orthoptera Research 22: 73-77.
Konecny, M.J. 1987. Food habits and energetics of feral house cats in the Galapagos Islands. Oikos 50: 24-32.
Laws A.N. et A. Joern. 2012. Variable effects of dipteran parasitoids and management treatment on grasshopper fecundity in a tallgrass prairie. Bulletin of Entomological Research102: 123-130.
LEMV (Liste des espèces susceptibles d’être désignées menacées ou vulnérables). [consulté le 8 septembre 2016]
Magdalen Islands Conservation Society. 2015. [consulté le 14 décembre 2015]. (Également disponible en français : Société de conservation des Îles-de-la-Madeleine. 2015. Site Web : )
Majka, C.G. et F. Shaffer. 2008. Beetles (Coleoptera) in the diet of Piping Plovers in the Iles de la Madeleine, Québec, Canada. Wader Study Group Bull 115: 77–83.
McMurry, F.B. et C.C. Sperry. 1941. Food of feral house cats in Oklahoma, a progress report. Journal of Mammalogy 22: 185-190.
Molsher, R., A. Newsome et C. Dickman. 1999. Feeding ecology and population dynamics of the feral cat (Felis catus) in relation to the availability of prey in central-eastern New South Wales. Wildlife Research 26: 593-607.
Mulkern, G.B. 1980. Population fluctuations and competitive relationships of grasshopper species (Orthoptera: Acrididae). Transactions of the American Entomological Society 106:1-41.
NatureServe. 2015. NatureServe Explorer: An online encyclopedia of life[application Web]. Version 7.1. NatureServe, Arlington, Virginia. Available [consulté le 8 octobre 2015]. [en anglais seulement]
Parker, J.R. et C.C. Wakeland. 1957. Grasshopper egg pods destroyed by larvae of bee flies, blister beetles, and ground beetles. US Department of Agriculture Technical Bulletin No. 1165.
Pickford, R. et P.W. Riegert. 1964. The fungus disease caused by Entomophthora grylli Fres., and its effects on grasshopper populations in Saskatchewan in 1963. Canadian Entomologist 96: 1158-1166.
Pinto, J.D. 1991. The taxonomy of North American Epicauta (Coleoptera: Meloidae), with a revision of the nominate subgenus and a survey of courtship behavior. University of California Publications in Entomology 110: 1-372.
Preston-Mafham, K. 1990. Grasshoppers and Mantids of the World. Facts on File, New York. 192 pp.
Przybyszewski, J. et J.L. Capinera. 1991. Patterns of parasitism among shortgrass prairie grasshopper (Orthoptera: Acrididae) populations. Journal of the Kansas Entomological Society 64: 5-17.
Read, J. et Z. Bowen. 2001. Population dynamics, diet and aspects of the biology of feral cats and foxes in arid South Australia. Wildlife Research 28: 195-203.
Rees, N.E. 1985. Suitability of selected North American grasshopper species as hosts for grasshopper parasites from Pakistan. Agriculture, Ecosystems & Environment 12: 157-163.
Rémillard, A.M., G. St-Onge, P. Bernatchez, J.P. Buylaert, A.S. Murray et B. Vigneault, 2016. Chronology and stratigraphy of the Magdalen Islands archipelago from the last glaciation to the early Holocene: new insights into the glacial and sea-level history of eastern Canada. Boreas. 10.1111/bor.12179. ISSN 0300-9483.
Shaffer, F. et P. Laporte. 1994. Diet of Piping Plovers on the Magdalen Islands, Quebec. Wilson Bulletin 106: 531-536.
Skeel, M.A. et E.P. Mallory. 1996. Whimbrel (Numenius phaeopus). In The Birds of North America, No. 219 (A. Poole and F. Gill, eds.). The Academy of Natural Sciences, Philadelphia, PA, and the American Ornithologists' Union, Washington, D.C.
Smith, C.W. 1940. An Exchange of Grasshopper Parasites between Argentina and Canada with Notes on Parasitism of native Grasshoppers. Report of the Entomology Society of Ontario 70: 57-62.
Smith, R.W. et T.U. Finlayson. 1950. Larvae of dipterous parasites of nymphal and adult grasshoppers. Canadian Journal of Research 28: 81-117.
Streett, D.A. et M.R. McGuire. 1990. Pathogenic diseases of grasshoppers. Pp. 483-516 In: R.F. Chapman and A. Joern (eds). Biology of grasshoppers. John Wiley & Sons.
Tourism Îles de la Madeleine. 2016. [consulté le 8 septembre 2016] (Également disponible en français : Tourisme Îles de la Madeleine. 2016.)
Tourisme Îles de la Madeleine. 2012. Un bilan 2012 positif pour Tourisme Îles de la Madeleine. [consulté le 8 septembre 2016].
Vickery, V. R. et D.K.M. Kevan. 1985. The grasshoppers, crickets, and related insects of Canada and adjacent regions. Biosystematics Research Institute, Ottawa, Ontario. Publication Number 1777. 918 pp.
Vickery, V.R. 1989. The biogeography of Canadian Grylloptera and Orthoptera. Canadian Entomologist 121: 389-424.
Vickery, V.R. et D.K.E. Kevan. 1978. A new species of Melanoplus (Orthoptera: Acrididae: Melanoplinae) from the Magdalen Islands, Québec. Annals of the Entomological Society of Québec 22: 188-192.
Wood, J. 1973. Arthropoda which have been found in starling (Sturnus vulgaris) faeces. New Zealand Entomologist 5: 159-162.
York, G.T. et H.W. Prescott. 1952. Nemestrinid parasites of grasshoppers. Journal of Economic Entomology 45: 5-10.
Sommaire biographique des rédactrices du rapport
Cory S. Sheffield étudie les abeilles et la pollinisation depuis 1993, année où il était étudiant au baccalauréat spécialisé à l’Université Acadia, à Wolfville (Nouvelle-Écosse). Il a mené ensuite des études supérieures (M.Sc.) sur les interactions entre les insectes et les plantes à l’Université Acadia, ainsi qu’à Agriculture et Agoalimentaire Canada (AAC), à Kentville (Nouvelle-Écosse), de 1994 à 2006. Il a mené des études supérieures (Ph. D.) à l’Université de Guelph, en Ontario, tout en continuant à travailler à AAC. Ces études ont porté sur les abeilles de la Nouvelle-Écosse, notamment sur leur diversité et leur rôle dans la pollinisation des cultures. Durant cette période, il a publié, en collaboration avec plusieurs auteurs, des articles portant sur la redécouverte d’Epeoloides pilosulus en Nouvelle-Écosse, alors qu’on pensait que l’espèce avait disparu dans la province. Il a par la suite entrepris des études postdoctorales à l’Université York (Ontario), sur la taxonomie et le codage à barres de l’ADN des abeilles, avant d’accepter un poste d’associé de recherche en taxonomie des abeilles au sein de l’Initiative canadienne sur la pollinisation (CANPOLIN). Il œuvre aujourd’hui à titre de chercheur scientifique et de conservateur en zoologie des invertébrés au musée royal de la Saskatchewan (Royal Saskatchewan Museum), à Regina (Saskatchewan). Son intérêt pour les abeilles demeure toujours aussi vif, et il a publié des articles sur la taxonomie des abeilles du Canada et de l’Amérique du Nord, sur l’utilité du codage à barres pour l’identification des abeilles, sur la physiologie des abeilles, sur la contribution des abeilles à la pollinisation et sur la diversité des abeilles canadiennes.
Jennifer Heron s’intéresse à des sujets liés à la conservation des invertébrés depuis le début de sa maîtrise (M.Sc.) en 1998. Elle travaille actuellement à titre de spécialiste provinciale des invertébrés au ministère de l’Environnement de la Colombie-Britannique. Elle dirige et gère la stratégie provinciale de conservation des invertébrés, y compris l’élaboration et la mise en œuvre de la réglementation, des politiques, des procédures et des normes provinciales visant à assurer la conservation et le rétablissement des espèces d’invertébrés en péril, de leur habitat et des écosystèmes, et à éviter que ces espèces deviennent en péril. Elle travaille avec d’autres spécialistes des invertébrés à l’élaboration de stratégies de planification du rétablissement et attribue les cotes de conservation aux groupes d’invertébrés. Elle collabore avec les groupes de conservation et d’intendance locaux afin d’atteindre des objectifs communs de sensibilisation du public.
Collections examinées
Collection nationale canadienne d’insectes, d’arachnides et de nématodes, Ottawa (Ontario).
Musée Lyman, Université McGill, Montréal (Québec).