
Évaluation et Rapport de situation du COSEPAC sur la grenouille à pattes rouges du Nord (Rana aurora) au Canada - 2015

©Photo : Kristiina Ovaska
Description longue de la bannière
Photo (vue du dessus) d'une grenouille à pattes rouges du Nord (Rana aurora) adulte. L'espèce a une peau lisse à légèrement rugueuse, un pli dorsolatéral sur chaque flanc, qui commence près de l'œil et se termine près de l'aine, des membres relativement longs par comparaison aux autres groupes de grenouilles, et des pattes palmées. Le dos est brunâtre, parsemé de petites taches noires aux contours indistincts, et la surface dorsale des membres porte souvent des barres noires. Un masque foncé, délimité vers le bas par une bande de couleur crème, s'étend généralement de l'œil à la mâchoire.
Table des matières
- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique
- Préface
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances de la population
- Menaces et facteurs limitatifs
- Facteurs limitatifs
- Menaces
- Description des menaces
- Fragmentation du paysage et effets cumulatifs
- Espèces envahissantes ou espèces indigènes autrement problématiques – impact moyen à élevé
- Corridors de transport et de service – impact moyen
- Développement résidentiel et commercial – faible impact
- Utilisation des ressources biologiques – exploitation forestière – faible impact
- Modifications des systèmes naturels – faible impact
- Pollution – faible impact
- Changements climatiques – impact inconnu
- Nombre de localités
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique du rédacteurs du rapport
- Collections examinées
Liste des figures
- Figure 1. Grenouille à pattes rouges du Nord de la Colombie-Britannique : A) adulte (île South Pender), B) têtard (ruisseau Vedder), C) grappe d'œufs (ruisseau Vedder). Photographies de Kristiina Ovaska.
- Figure 2. Aire de répartition canadienne de la grenouille à pattes rouges du Nord. Les mentions sont issues des données compilées pour le présent rapport de situations.
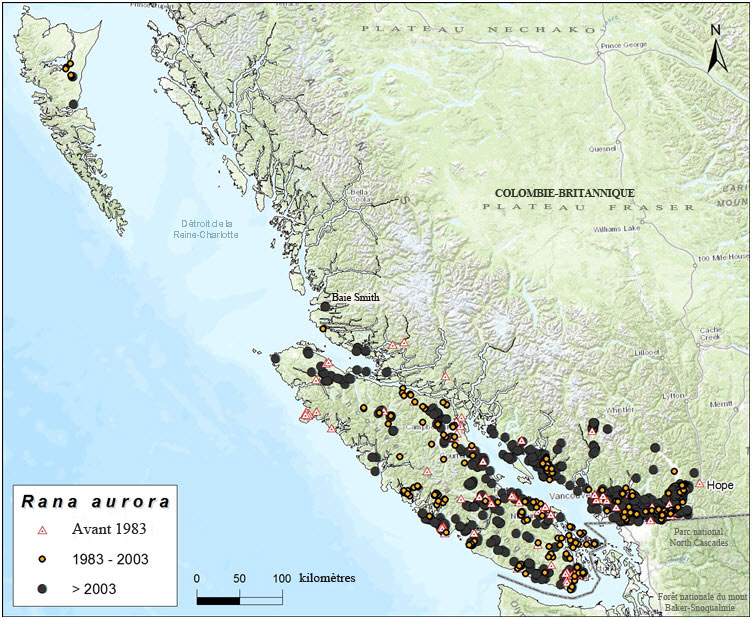
- Figure 3. Répartition canadienne du Grenouille à pattes rouges du Nord (Rana aurora) pour quatre périodes, montrant la zone d'occurrence, avant 2004 et dans la période 2004 2013.
- Figure 4. Distribution de la fréquence du nombre de grappes d'œufs de grenouille à pattes rouges du Nord observées de 2006 à 2013 dans les milieux humides occupés (N = 197) en Colombie-Britannique. Des comptes maximums par milieu humide par année ont été utilisés pour ceux qui ont fait l'objet d'un relevé sur plusieurs années. À noter que l'intervalle sur l'axe des x change entre les 3 premières barres, puis il devient constant. Notes : (i) Tous les relevés n'ont pas été menés au moment idéal ou dans des conditions idéales de détection des grappes d'œufs; (ii) Dans certains cas, plusieurs plans d'eau dans le même complexe de milieux humides sont présentés comme des points de données individuels même s'ils ne représentent pas des populations indépendantes.
Liste des tableaux
- Tableau 1. Nombre d'occurrences de la grenouille à pattes rouges du Nord dans différentes parties de son aire de répartition en Colombie-Britannique selon la période et le nombre d'occurrences documentées avant 1983
- Tableau 2. Activités de recherche menées depuis les années 1990 visant à établir l'aire de répartition canadienne, la zone d'occurrence et la zone d'occupation de la grenouille à pattes rouges du Nord
- Tableau 3. Nombre estimatif de grenouilles à pattes rouges du Nord adultes en état de se reproduire dans les milieux humides occupés par l'espèce qui ont fait l'objet de relevés des grappes d'œufs au printemps
- Tableau 4. Résumé des principales menaces pesant sur la grenouille à pattes rouges du Nord selon l'évaluation des menaces fondée sur les catégories de menaces de l'UICN
- Tableau 5. Aires protégées occupées par la grenouille à pattes rouges du Nord en Colombie-Britannique, au Canada, d'après les mentions historiques (indiquées par l'exposanth) et les mentions recueillies depuis 1983.
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2015. Évaluation et Rapport de situation du COSEPAC sur la grenouille à pattes rouges du Nord (Rana aurora) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xiii + 78 p. (Registre public des espèces en péril).
Rapport(s) précédent(s) :
COSEPAC. 2004. Évaluation et Rapport de situation du COSEPAC sur la grenouille à pattes rouges (Rana aurora) au Canada - Mise à jour. Comité sur la situation des espèces en péril au Canada. Ottawa. vii + 54 p. (Registre public des espèces en péril).
WAYE, H. 1999. COSEWIC status report on the red-legged frog Rana aurora in Canada in COSEWIC assessment and status report on the red-legged frog Rana aurora in Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. 1-31 p.
Le COSEPAC remercie Barbara Beasley d'avoir rédigé le rapport de situation sur la grenouille à pattes rouges du Nord (Rana aurora) au Canada, aux termes d'un marché conclu avec Environnement Canada. La supervision du rapport a été assurée par Kristiina Ovaska, co-présidente du Sous-comité de spécialistes des amphibiens et des reptiles du COSEPAC.
Pour obtenir des exemplaires supplémentaires, s'adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Northern Red-legged Frog Rana aurora in Canada.
Grenouille à pattes rouges du Nord -- Individu adulte de la grenouille à pattes rouges du Nord (Rana aurora) photographié à l'île South Pender, en Colombie-Britannique. Photo : Kristiina Ovaska.


Grenouille à pattes rouges du Nord
Rana aurora
La grenouille à pattes rouges du Nord (Rana aurora) appartient à la famille des Ranidés, ou « vraies grenouilles ». Avant 2004, le nom Rana aurora s'appliquait tant au R. aurora qu'au R. draytonii (grenouille à pattes rouges de la Californie), puis des données génétiques ont révélé qu'il s'agit d'espèces distinctes. Seul le R. aurora se trouve au Canada.
La grenouille à pattes rouges du Nord est de taille moyenne; les adultes ont généralement une longueur du museau au cloaque de 50 à 70 mm. Le dos est brun, parsemé de petites taches noires, et les pattes portent des barres noires. Chez les adultes, la coloration du dessous des pattes postérieures est rouge et visible à travers la peau translucide, d'où le nom commun de l'espèce.
La grenouille à pattes rouges du Nord joue des rôles écologiques importants tant à titre de consommateur d'invertébrés, notamment des insectes nuisibles, qu'à titre de proie pour des poissons, des reptiles, des oiseaux et des mammifères. Ses déplacements réguliers entre des milieux aquatiques et terrestres lui confèrent un rôle dans le transfert d'éléments nutritifs et d'énergie entre écosystèmes. L'espèce est sensible aux polluants, y compris les pesticides et les sous-produits azotés.
L'aire de répartition de la grenouille à pattes rouges du Nord s'étend depuis le sud-ouest de la Colombie-Britannique, vers le sud le long du littoral du Pacifique, jusqu'au nord-ouest de la Californie, à l'ouest des monts Cascades. Une population a été introduite dans l'île Chichagof, en Alaska, et une population d'origine inconnue est présente dans l'île Graham, en Colombie-Britannique. Les récentes mentions de la présence de la grenouille à pattes rouges du Nord confirment que l'espèce demeure répandue dans l'île de Vancouver et la partie continentale adjacente de la province. Les nouvelles mentions étendent son aire de répartition à l'ensemble de la Sunshine Coast (sud de la côte continentale au nord-ouest de la région métropolitaine de Vancouver) et du corridor Sea-to-Sky (route 99, de Vancouver à Whistler) et plus haut dans les vallées entourant les basses terres du Fraser. Il reste des populations dans quelques zones urbanisées de la région du Lower Mainland et de la vallée du Fraser, mais l'échec des recherches menées au cours des cinq dernières années laisse supposer qu'elles ont disparu de certaines parties des villes de Vancouver, Richmond et Delta, où elles étaient présentes par le passé. Des activités de recherche intensives menées dans le parc Stanley depuis les années 1970 sans que des individus y soient trouvés indiquent que l'espèce y est disparue.
La grenouille à pattes rouges du Nord a besoin de milieux aquatiques pour se reproduire et de milieux terrestres pour s'alimenter, à de faibles altitudes (généralement à moins de 500 m, bien que l'espèce puisse se trouver à une altitude aussi élevée que 1 040 m). Elle pond ses œufs sur les parties submergées des plantes tapissant des milieux humides permanents ou temporaires exposés au soleil, où l'eau est d'au moins 30 cm de profondeur, et le débit, faible. Le taux de survie des têtards est plus élevé dans les milieux humides temporaires montrant une structure complexe et abritant relativement peu de prédateurs en comparaison des milieux humides permanents. Les adultes et les juvéniles se dispersent jusqu'à 5 km des milieux humides vers des forêts humides, où ils trouvent refuge dans des terriers humides, sous de gros morceaux d'arbres abattus et dans la végétation du sous-étage. L'habitat d'hivernage se compose de refuges souterrains dans les forêts et les milieux humides.
L'habitat a connu une perte et une dégradation considérables dans le sud et l'est de l'île de Vancouver et la vallée du bas Fraser à cause de l'agriculture, de la construction de routes, de l'urbanisation, de l'introduction du ouaouaron et de l'exploitation forestière.
Les adultes se reproduisent à la fin de l'hiver ou au début du printemps, revenant souvent au même site de reproduction année après année. Les mâles produisent un appel sous l'eau. Les femelles pondent de 200 à 1 100 œufs agglutinés en une seule grappe. La mortalité précoce est relativement élevée, mais diminue à l'atteinte de la maturité; chez les adultes, le taux de survie annuel est estimé à 69 %. Les mâles atteignent généralement la maturité sexuelle à l'âge de 2 ans, alors que les femelles peuvent prendre jusqu'à 4 ans pour l'atteindre. La durée de vie maximale en captivité est de 15 ans. La durée d'une génération est estimée à 4 à 6 ans.
Les populations de grenouilles à pattes rouges du Nord en Colombie-Britannique comptent probablement plus de 100 000 adultes. La taille des populations reproductrices varie d'un site à l'autre, la plus grande se trouvant dans les secteurs boisés relativement peu développés de la côte ouest de l'île de Vancouver. Les grappes d'œufs (un indice du nombre de femelles reproductrices) dénombrées se chiffrent à plus de 1 400 par étang, mais il est probable qu'un nombre aussi élevé soit extrêmement rare. Des étangs ayant fait l'objet d'un relevé jusqu'à maintenant, 80 % abritaient moins de 100 grappes d'œufs, et 35 %, moins de 10. Une surveillance à long terme des populations a été lancée, mais n'a pas été effectuée suffisamment longtemps pour établir les tendances. Des disparitions et des déclins locaux ont été documentés. La population de Delta semble avoir diminué lentement depuis les années 1960, mais la surveillance est limitée. Dans un milieu humide du corridor Sea-to-Sky, il s'est produit un déclin estimatif de 73 à 92 % sur 3 ans par suite de la construction de l'autoroute et de la mortalité routière.
L'urbanisation, la mortalité routière, l'exploitation forestière, l'aménagement de barrages et la gestion de l'eau, l'introduction d'espèces envahissantes et de poissons, les maladies et la pollution constituent les menaces réputées peser sur la grenouille à pattes rouges du Nord. Dans la vallée du bas Fraser, la population humaine devrait doubler tous les 20 à 30 ans et augmenter également sur le littoral sud-est de l'île de Vancouver, dans des parties du corridor Sea-to-Sky et sur la Sunshine Coast. Une partie de l'habitat existant de l'espèce en zones urbaines et en zones rurales agricoles sera converti aux fins de la construction d'habitations et, par conséquent, l'habitat sera davantage fragmenté et exposé à la pollution. Les populations de ouaouarons devraient augmenter et s'étendre, ce qui intensifiera la compétition, la pression de prédation et les dommages à l'habitat de reproduction causés par les barrages, les activités de gestion de l'eau, les poissons introduits et les maladies. L'exploitation forestière pourrait entraîner la modification de l'habitat dans l'ensemble des vastes parties éloignées de l'aire de répartition de l'espèce. Les températures extrêmes, les tempêtes et les inondations résultant des changements climatiques exacerberont probablement la dégradation et la perte d'habitat causées par d'autres facteurs.
La grenouille à pattes rouges du Nord est classée « apparemment non en péril » (G4) à l'échelle mondiale. Au Canada, l'espèce est inscrite à titre d'espèce préoccupante à l'annexe 1 de la Loi sur les espèces en péril du gouvernement fédéral et sur la liste bleue provinciale des espèces en péril en Colombie-Britannique. À l'heure actuelle, environ 3 210 km2 ou 10 % de l'aire de répartition à moins de 500 m d'altitude et 16 % des occurrences de la grenouille à pattes rouges du Nord se trouvent dans des aires protégées. La majeure partie de l'aire de répartition et des occurrences se trouvent sur des terres provinciales non protégées et des terres forestières privées.
Durée d'une génération
Estimée d'après les données disponibles sur le taux de survie des adultes (voir Biologie).
- Entre 4 à 6 ans
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures?
Des déclins locaux ont été observés et des déclins à l'échelle de l'aire de répartition sont inférés d'après les tendances en matière d'habitat. Le nombre d'individus matures est tombé à zéro dans le parc Stanley entre les années 1970 et aujourd'hui. Des déclins ont été observés dans le territoire de la Corporation de Delta depuis les années 1960. Un déclin de 73 à 92 % sur 3 ans s'est produit au site de Pinecrest à cause de la route 99 (voir Fluctuations et tendances des populations).
- Inconnu
Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] (52-60 ans).
Les données sont insuffisantes pour estimer l'ampleur du déclin.
- Inconnu
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [10 dernières années ou 3 dernières générations].
Selon le BC Conservation Data Centre (2014), la tendance à court terme au cours des 10 dernières années ou des 3 dernières générations est une réduction de 10 à 50 %. Les données insuffisantes ne permettent toutefois pas d'estimer de façon fiable l'ampleur de la réduction.
- Inconnu
Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [10 prochaines années ou 3 prochaines générations].
Aucune donnée n'est disponible.
- Inconnu
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [10 années ou 3 générations] commençant dans le passé et se terminant dans le futur.
Aucune donnée n'est disponible. Comme les sites historiques n'ont pas été rééchantillonnés de façon uniforme, il est impossible d'estimer le nombre.
- Inconnu
Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé?
Les effets négatifs de nombreuses menaces, notamment le développement résidentiel, la construction de routes et la pollution, pourraient être atténués. Les effets de l'exploitation forestière sont réversibles au fil du temps si les forêts ont la possibilité d'atteindre la maturité et que des réserves riveraines de taille appropriée sont établies et que des parcelles de gros arbres sont maintenues dans les hautes terres à chaque rotation.
- Plus ou moins réversibles et comprises; n'ont pas cessé.
Y a-t-il des fluctuations extrêmes du nombre d'individus matures?
Les comptes des grappes d'œufs (indice du nombre de femelles reproductrices) ne montrent pas de fluctuations extrêmes sur des périodes de surveillance relativement courtes (< 10 ans), mais il est en général reconnu que l'effectif local des grenouilles qui se reproduisent dans des étangs fluctue considérablement.
- Inconnu
Superficie estimée de la zone d'occurrence
Calculée d'après toutes les observations connues.
- 75 625 km2
Indice de zone d'occupation (IZO) actuel en km2 (carrés de 2 km de côté) : 247 carrés = 988 km2
Calculé d'après toutes les observations connues.
- 2 588 km2 (discret) mais l'IZO réel est probablement plus élevé
La population totale est-elle gravement fragmentée?
La mer, les lacs, les rivières, les montagnes, les routes et les terres déboisées fragmentent les populations à l'échelle de l'aire de répartition canadienne de l'espèce, plus gravement dans les zones urbaines de la région du Lower Mainland et sur le côté sud-est de l'île de Vancouver. Toutefois, au moins 50 % de la population canadienne ne se trouve pas dans des parcelles d'habitat isolées.
- Non
Nombre de localitésTable Footnotea
Le développement résidentiel et commercial qui nécessite le drainage d'un milieu humide isolé et la coupe de la forêt environnante constitue une menace qui pourrait rapidement nuire à tous les individus du taxon présents dans un seul site. Le nombre total d'occurrences sensibles au développement résidentiel s'élève à 106. (Au moins 232 sites sont touchés par d'autres menaces; voir tableau 1).
- Inconnu, mais beaucoup plus élevé que 10Table Footnotea
Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence?
La zone d'occurrence connue d'après les mentions historiques n'a pas diminué. Les activités d'échantillonnage et la sensibilisation à l'espèce ont augmenté au fil du temps, tout comme la zone d'occurrence connue.
- Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation?
Les activités d'échantillonnage et la sensibilisation à l'espèce ont augmenté au fil du temps, tout comme l'IZO connu. Toutefois, dans la région du Lower Mainland, la modélisation de l'occupation prédit une occupation nulle des sites où la présence de l'espèce a été documentée par le passé (voir Zone d'occurrence et zone d'occupation).
- Oui (prévu)
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations?
Un déclin est inféré et prévu d'après les tendances en matière d'habitat et les menaces. Une population a disparu. Environ 72 % des localités pour lesquelles des mentions historiques sont disponibles n'ont pas été revisitées au cours des 30 dernières années, et les activités de recherche se sont limitées à celles qui avaient été revisitées.
- Oui (inféré et prévu)
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités Table Footnotea ?
- Oui (prévu)
Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat?
Une croissance démographique rapide dans la région du Lower Mainland, dans le corridor Sea-to-Sky, sur la Sunshine Coast et dans le sud-est de l'île de Vancouver cause un déclin continu de la superficie et de la qualité de l'habitat. Partout dans le reste de l'aire de répartition, l'exploitation forestière a réduit la qualité de l'habitat; 13 % des forêts aménagées de moins de 500 m d'altitude situées dans l'aire de répartition de l'espèce ont été coupées au cours des 20 dernières années, et l'exploitation forestière continue (voir Tendances en matière d'habitat et Menaces – Utilisation des ressources biologiques).
- Oui (observé, inféré et prévu)
Y a-t-il des fluctuations extrêmes du nombre de populations?
- Non
Y a-t-il des fluctuations extrêmes du nombre de localités?
- Non
Y a-t-il des fluctuations extrêmes de la zone d'occurrence?
- Non
Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation?
- Non
Population:
La taille cumulative des populations dans les milieux humides recensés s'élève à plus de 32 000 individus (voir Taille et tendances des populations – Abondance). Seule une petite partie de l'habitat occupé a été inventorié.
- Total
- Au moins 100 000
La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans].
Aucune analyse de viabilité des populations n'a été réalisée pour la Colombie-Britannique. Une analyse de la viabilité d'une sous population se trouvant près de Whistler a été réalisée pour évaluer le besoin d'autres mesures d'atténuation afin de réduire la mortalité sur les routes. Dans ces cas, il a été prédit que la sous-population allait disparaître d'ici 20 à 40 ans si aucune mesure d'atténuation n'était prise (voir Menaces – Routes).
- Données insuffisantes
Principales menaces
Développement résidentiel et commercial; espèces envahissantes (ouaouaron; maladies émergentes telles que la chytridiomycose); pollution; exploitation forestière; changements climatiques (sécheresses estivales; disparition précoce des sites de reproduction éphémères)
Autres menaces à impact faible ou inconnu : agriculture, ouvrages hydroélectriques de retenue
- Situation des populations de l'extérieur?
Statut aux États-Unis : N4 – « apparemment non en péril »; État de Washington : S4 – « apparemment non en péril »; Oregon : S3S4 – « vulnérable » à « apparemment non en péril »; Californie : S2? – « en péril », cote inexacte.
Une immigration a-t-elle été constatée ou est-elle possible?
Des individus de l'État de Washington pourraient immigrer dans le Lower Mainland de la Colombie-Britannique, mais la situation de l'espèce dans la région frontalière est inconnue. Une bonne partie de l'habitat est de mauvaise qualité (agriculture intensive) le long du corridor accessible de la frontière dans les basses terres entre Blaine et le mont Vedder
- Possible
Des individus immigrants seraient-ils adaptés pour survivre au Canada?
- Oui
Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants?
- Peut-être
La possibilité d'une immigration depuis des populations externes existe-t-elle?
- Peu probable
L'information concernant l'espèce est elle de nature délicate?
- Non
COSEPAC : Espèce désignée « préoccupante » en avril 1999. Réexamen et confirmation du statut en mai 2002, en novembre 2004, et en mai 2015.
Depuis la préparation du rapport de situation précédent du COSEPAC, en 2004, la grenouille à pattes rouges du Nord (Rana aurora) a été reconnue comme une espèce distincte de la grenouille à pattes rouges de la Californie (Rana draytonii). Il est maintenant confirmé qu'une population historique a disparu du Lower Mainland de la Colombie-Britannique (du parc Stanley Park; aucune mention depuis les années 1970 malgré les activités de recherche continues). La collecte récente de données et un modèle d'occupation donnent à penser que d'autres populations, à Delta et à Richmond, sont sur le point de disparaître de la province. Au cours de la même période, les activités de recherche accrues menées à l'extérieur du Lower Mainland ont révélé des populations jusqu'alors inconnues dans le corridor Sea-to-Sky, le long de la Sunshine Coast et de la côte centrale ainsi que dans l'île de Vancouver. Très peu de surveillance a été effectuée pour évaluer les tendances des populations; toutefois, la population d'un site a chuté de façon spectaculaire par suite de l'aménagement de routes et de la mortalité routière.
La grenouille à pattes rouges du Nord est prise en considération dans le cadre du processus d'évaluation environnementale de la Colombie-Britannique, qui inclut des mesures visant à éviter, à réduire et à surveiller les effets néfastes d'un projet d'une manière qui cadre avec le plan de gestion de l'espèce. Une ébauche de plan de gestion est en cours d'examen. La grenouille à pattes rouges du Nord est inscrite à la liste des espèces sauvages désignées et, sur les terres publiques provinciales, est gérée en vertu de la Forest and Range Practices Act de la Colombie-Britannique. La stratégie de gestion des espèces sauvages désignées (Identified Wildlife Management Strategy) prévoit des politiques, des procédures et des lignes directrices pour la gestion des espèces sauvages désignées, y compris la création de zones d'habitat d'espèces sauvages (« Wildlife Habitat Areas », ou WHA) et l'établissement de mesures générales connexes. La grenouille à pattes rouges du Nord est maintenant protégée dans 23 aires d'une superficie totale de 336 ha dans l'île de Vancouver, et la création d'autres aires est envisagée.
Aucune connaissance traditionnelle autochtone sur l'espèce n'était disponible.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces Préoccupantes de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Le Service canadien de la faune d'Environnement Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
La grenouille à pattes rouges du Nord appartient à la grande famille quasi cosmopolite des Ranidés, ou « vraies grenouilles (Amphibiens : Anoures : Ranidés : Rana : Rana aurora Baird et Girard, 1852). Huit espèces du genre Rana sont indigènes de la côte ouest de l'Amérique du Nord : Rana aurora, R. boylii (grenouille à pattes jaunes des contreforts), R. cascadae (grenouille des Cascades), R. draytonii (grenouille à pattes rouges de la Californie), R. muscosa (grenouille à pattes jaunes des montagnes), R. pretiosa (grenouille maculée de l'Oregon), R. luteiventris (grenouille maculée de Columbia) et R. sierra (grenouille à pattes jaunes de la Sierra Nevada). Le Rana aurora et le R. draytonii ont récemment été considérés comme deux espèces distinctes d'après des différences dans l'ADN mitochondrial (Shaffer et al., 2004), dans les séquences d'acides aminés des peptides de la peau (Conlon et al., 2006) ainsi que dans la morphologie, le comportement, la fréquence allélique et la structure des chromosomes (Hayes et Miyamoto, 1984; Green, 1986).
Les huit espèces qui sont restreintes à l'ouest de l'Amérique du Nord forment le groupe d'espèces de R. boylii, que les analyses moléculaires indiquent être un groupe monophylétique bien défini, vieux d'environ huit millions d'années (Macey et al,. 2001; Hillis et Wilcox, 2005). Les relations au sein du groupe d'espèces de R. boylii ne sont pas complètement élucidées, mais un récent séquençage de l'ADN mitochondrial donne à penser que le R. aurora, le R. cascadae et le R. muscosa peuvent être étroitement apparentés (Macey et al., 2001; Hillis et Wilcox, 2005).
La grenouille à pattes rouges du Nord est de taille moyenne; les adultes ont généralement une longueur du museau au cloaque qui va d'environ 50 mm à plus de 70 mm (Matsuda et al., 2006). Les femelles sont un peu plus grosses que les mâles et peuvent atteindre une longueur d'environ 100 mm (Nussbaum et al., 1983). À l'instar de la plupart des autres ranidés de l'Amérique du Nord, la grenouille à pattes rouges du Nord a une peau lisse à légèrement rugueuse, un pli dorsolatéral sur chaque flanc, qui commence près de l'œil et se termine près de l'aine, des membres relativement longs par comparaison aux autres groupes de grenouilles, et des pattes palmées. Le dos est brunâtre, parsemé de petites taches noires aux contours indistincts; la surface dorsale des membres porte souvent des barres noires (figure 1a). Un masque foncé, délimité vers le bas par une bande de couleur crème, s'étend généralement de l'œil à la mâchoire. La gorge et la poitrine sont grises ou blanches, avec des mouchetures noires variables, parfois rougeâtres, alors que le dessous des pattes postérieures et la partie inférieure du tronc sont rougeâtres, d'où le nom commun de l'espèce. La vivacité de la coloration rouge varie selon l'emplacement géographique et l'ontogénie (Altig et Dumas, 1972); le rouge peut être complètement absent chez les juvéniles ou se présenter comme une faible coloration rougeâtre ou jaunâtre sur le dessous des pattes. Les mâles adultes se distinguent des femelles par la présence, à longueur d'année, de callosités nuptiales sur les pouces.
© Photographs by Kristiina Ovaska
Long description for Figure 1a.
La grenouille adulte a une peau lisse à légèrement rugueuse, un pli dorsolatéral sur chaque flanc, qui commence près de l'œil et se termine près de l'aine, des membres relativement longs par comparaison aux autres groupes de grenouilles, et des pattes palmées. Le dos est brunâtre, parsemé de petites taches noires aux contours indistincts, et la surface dorsale des membres porte souvent des barres noires. Un masque foncé, délimité vers le bas par une bande de couleur crème, s'étend généralement de l'œil à la mâchoire.

© Photographs by Kristiina Ovaska
Long description for Figure 1.
La couleur des têtards va du havane au brun verdâtre. Le tronc, la queue et les nageoires sont généralement couverts de mouchetures ou de taches de couleur or ou laiton, et le ventre, blanc, a souvent une teinte rosâtre. La queue est relativement courte (environ 1,5 fois ou moins la longueur du corps), et la nageoire dorsale est relativement haute (plus haute que la musculature de la queue à son point le plus large), ce qui donne au têtard une apparence trapue.

© Photographs by Kristiina Ovaska
Long description for Figure 1c.
Les grappes d'œufs sont fixées à la végétation; elles sont grosses, gélatineuses et molles, et ont une apparence globulaire. Chaque œuf est recouvert d'une épaisse enveloppe gélatineuse.
Les œufs sont pondus en grosses grappes gélatineuses molles qui ont une apparence globulaire (de 10 à 20 cm de diamètre). Chaque œuf, recouvert d'une épaisse enveloppe gélatineuse, mesure 3 mm de diamètre. La couleur des têtards va du havane au brun verdâtre. Le tronc, la queue et les nageoires sont généralement couverts de mouchetures ou de taches de couleur or ou laiton, et le ventre, blanc, a souvent une teinte rosâtre. Les têtards peuvent être d'une taille relativement grande (jusqu'à 70 à 80 mm environ) immédiatement avant la métamorphose. La queue est relativement courte (environ 1,5 fois ou moins la longueur du corps), et la nageoire dorsale est relativement haute (plus haute que la musculature de la queue à son point le plus large), ce qui donne au têtard une apparence trapue (figure 1b; Corkran et Thoms, 1996).
En Colombie-Britannique, la grenouille à pattes rouges du Nord peut être confondue avec les grenouilles maculées de l'Oregon et de Columbia, qui lui ressemblent par la forme de leur corps et la couleur rougeâtre du dessous de leurs pattes arrière et de la partie inférieure de leur tronc (Matsuda et al., 2006). La grenouille à pattes rouges du Nord est sympatrique avec la grenouille maculée de l'Oregon dans la vallée du bas Fraser, alors qu'elle est largement allopatrique avec la grenouille maculée de Columbia. Il est possible que les extrémités sud-est et nord de l'aire de répartition continentale de la grenouille à pattes rouges du Nord chevauchent l'aire de répartition de la grenouille maculée de Columbia (voir Aire de répartition canadienne). Les spécimens capturés dans ces régions doivent donc être examinés de près.
Aucune étude de la structure spatiale ou génétique des populations de grenouilles à pattes rouges du Nord présentes en Colombie-Britannique ou ailleurs n'a été réalisée. Une certaine différenciation est à prévoir entre les populations de l'île de Vancouver, du continent et des îles des détroits de Georgia et de Johnstone puisqu'elles sont géographiquement isolées les unes des autres par des étendues de mer. L'étendue de la pleine mer entre l'île de Vancouver et le continent est la plus petite (< 1 km) au niveau des îles extracôtières du détroit de Johnstone. Étant donné que l'espèce est restreinte à des altitudes relativement faibles, les chaînes de montagnes isolent probablement les populations trouvées dans les vallées des bassins versants de la côte ouest de l'île de Vancouver, de la Sunshine Coast (sud de la côte continentale au nord-ouest de la région métropolitaine de Vancouver), du corridor Sea-to-Sky (route 99, de Vancouver à Whistler) et du nord de la vallée du Fraser. Les recherches menées sur d'autres espèces de grenouilles ont révélé que les crêtes montagneuses ont un effet d'isolement sur le flux génique (Funk et al., 2005).
La grenouille à pattes rouges du Nord occupe une aire géographique restreinte dans une des provinces fauniques des amphibiens et des reptiles terrestres du COSEPAC (côte du Pacifique). Rien n'indique une variabilité génétique, écologique ou morphologique suffisante au sein de la population canadienne pour justifier l'établissement de plus d'une unité désignable.
La grenouille à pattes rouges du Nord a des besoins sur le plan spatial relativement grands, dépend de milieux terrestres et aquatiques et est associée avec des forêts humides, des berges de cours d'eau et des milieux humides. La présence de l'espèce indique donc la persistance de la connectivité de l'habitat à l'échelle du paysage, de la santé des écosystèmes et des valeurs rattachées aux espèces sauvages.
La grenouille à pattes rouges du Nord mange des invertébrés, y compris une variété d'insectes. Au stade de têtard ou d'adulte, elle est la proie d'invertébrés, de poissons, d'autres amphibiens, de reptiles, d'oiseaux et de mammifères. Ses déplacements réguliers entre des milieux aquatiques et terrestres lui attribuent un rôle dans le transfert d'éléments nutritifs et d'énergie entre écosystèmes.
Les grenouilles occupent une place importante dans la mythologie et l'art des Premières Nations Salish de la côte, de la Première Nation Haïda et d'autres groupes des Premières Nations de l'Ouest du Canada, mais on ne sait pas si la grenouille à pattes rouges du Nord en particulier a une signification spéciale pour les peuples autochtones.
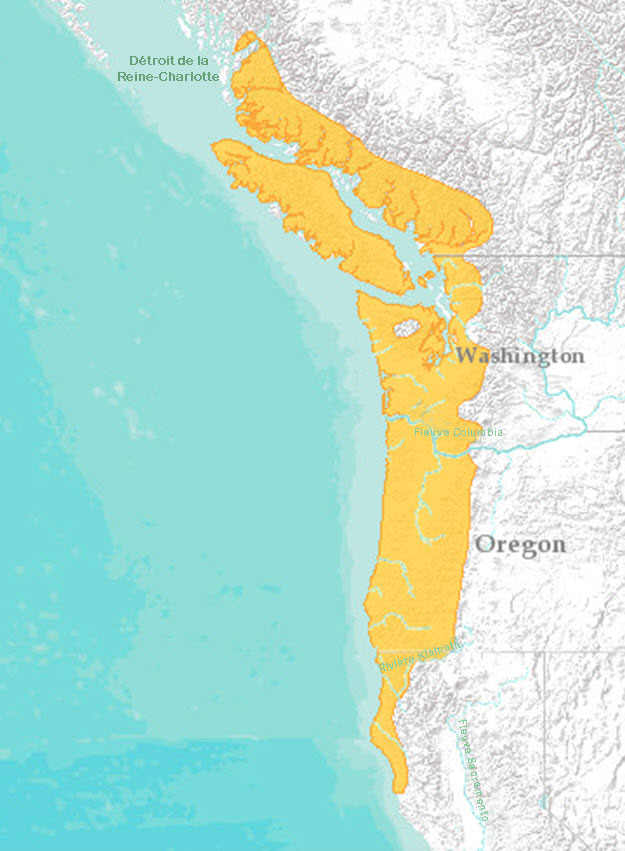
L’aire de répartition mondiale de la grenouille à pattes rouges du Nord s’étend depuis le sud-ouest de la Colombie-Britannique jusqu’au nord-ouest de la Californie (figure 2). L’espèce est présente partout dans l’ouest des États de Washington et de l’Oregon, à l’ouest des monts Cascades, jusqu’à la côte du Pacifique. Dans le nord-ouest de la Californie, la grenouille à pattes rouges du Nord se trouve aussi loin au sud que le comté de Mendocino (Shaffer et al., 2004). À cet endroit, son aire de répartition chevauche sur plusieurs kilomètres celle du R. draytonii, qui s’étend vers le sud jusqu’à la Basse-Californie, au Mexique. Une population isolée existe dans le sud-est de l’Alaska conséquemment à une récente introduction (MacDonald, 2010). Une population dans les îles de la Reine-Charlotte a aussi probablement été introduite. Environ 35 % de l’aire de répartition mondiale de la grenouille à pattes rouges du Nord se trouve au Canada.
Carte préparée par Jenny Wu, Secrétariat du COSEPAC.
Description longue pour la figure 2
Carte montrant l'aire de répartition mondiale du Grenouille à pattes rouges du Nord, qui est vaste, mais disjointe, dans les bassins versants du Mississippi et des Grands Lacs de l'est de l'Amérique du Nord. L'espèce est présente depuis l'Alabama et le Mississippi, au sud, jusqu'en Ontario et au Michigan, au nord, et de l'État de New York, à l'est, jusqu'en Oklahoma et au Minnesota, à l'ouest. Dans le bassin versant du Mississippi, sa répartition est continue à l'est du fleuve, mais disjointe à l'ouest de celui ci. Dans le bassin des Grands Lacs, on trouve des populations disjointes en Ontario, au Michigan et au Wisconsin.
Au Canada, la grenouille à pattes rouges du Nord se trouve dans le sud-ouest de la Colombie-Britannique, où elle est présente dans l’ensemble de l’île de Vancouver, dans plusieurs îles des détroits de Georgia et de Johnstone et sur les terres continentales adjacentes, à l’ouest de la chaîne Côtière (figure 3, tableau 1). Plus de 50 % de l’aire de répartition canadienne de l’espèce se trouve dans l’île de Vancouver. La plupart des mentions proviennent d’altitudes inférieures à 500 m, bien que l’espèce ait été signalée à des altitudes allant jusqu’à 1 040 m.
N/A
Description longue pour la figure 3
Carte de l'aire de répartition canadienne de la grenouille à pattes rouges du Nord, dans le sud ouest de la Colombie-Britannique, où l'espèce est présente dans l'ensemble de l'île de Vancouver, dans plusieurs îles des détroits de Georgia et de Johnstone et sur les terres continentales adjacentes, à l'ouest de la chaîne Côtière. Dans la zone continentale, l'aire de répartition s'étend depuis la vallée du bas Fraser jusqu'aux environs de Hope vers l'est, puis le long du corridor Sea-to-Sky vers le nord jusqu'à Whistler, et le long de la Sunshine Coast et de la côte centrale jusqu'au détroit de Smith. Plusieurs occurrences se trouvent à Haida Gwaii.
Description longue pour le tableau 1
Le nombre total d’occurrences (pour toutes les années) dans des aires protégées et le nombre estimatif de celles exposées à des menaces sont indiqués à la droite du tableau.
Une occurrence s’entend d’un ensemble de sites se trouvant dans un rayon de 1 km l’un de l’autre lorsqu’ils sont portés sur Google Earth. Les occurrences ont été considérées comme étant menacées par l’urbanisation, les routes pavées et l’exploitation forestière si ces utilisations des terres étaient visibles à moins de 1 km (< 500 m dans le cas des routes) d’un site d’occurrence sur Google Earth. Les occurrences ont été considérées comme menacées par des espèces envahissantes d’après un chevauchement avec la répartition cartographiée du ouaouaron (Govindarajulu, 2003; Govindarajulu, comm. pers., 2014) et des données de relevés récents (Murray et. al., sous presse; Mitchell et al., 2012). « I » - inconnu, « s.o. » - sans objet.
| Région du Lower | Sea-to-Sky | Sunshine Coast | Côte centrale | Île de Vancouver | Détroit de Johnstone | Détroit de Georgia | Total |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Baie Horseshoe à Caulfield | 0 | 1 | 1 | 2 | s.o. | 0 | 1 | 1 | 0 | U | 0 |
| Vancouver Nord – Lynn Valley à Deep Cove | 0 | 1 | 3 | 3 | s.o. | 1 | 2 | 3 | 0 | 0 | 0 |
| Parc Stanley | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 |
| Vancouver | 1 | 0 | 1 | 2 | 1 | 1 | 1 | 2 | 0 | 2 | 0 |
| Burnaby | 2 | 0 | 3 | 5 | 1 | 1 | 4 | 5 | 0 | 5 | 0 |
| Coquitlam | 0 | 4 | 5 | 8 | s.o. | 2 | 1 | 3 | 1 | U | 0 |
| Parc provincial Golden Ears | 0 | 1 | 2 | 3 | s.o. | 3 | 0 | 0 | 0 | 3 | 0 |
| Maple Ridge à Pitt Meadows | 2 | 2 | 3 | 6 | 2 | 0 | 2 | 3 | 3 | 6 | 1 |
| Mission (chutes Stave au lac Errock) | 2 | 0 | 14 | 14 | 0 | 1 | 1 | 9 | 1 | 14 | 11 |
| Lac Harrison (ouest) – Harrison Mills à Port Douglas | 0 | 2 | 6 | 7 | s.o. | 0 | 1 | 1 | 7 | 1 | 0 |
| Parc provincial Sasquatch | 1 | 3 | 2 | 3 | 0 | 3 | 0 | 0 | 0 | U | 0 |
| Agassiz à Harrison Hot Springs | 0 | 3 | 21 | 21 | s.o. | 0 | 2 | 14 | 0 | U | 18 |
| Laidlaw à Hope | 1 | 0 | 2 | 4 | 1 | 1 | 0 | 3 | 0 | U | 0 |
| Chilliwack-Lac Cultus | 4 | 12 | 31 | 37 | 2 | 4 | 4 | 26 | 4 | 12 | 16 |
| Abbotsford – secteur du mont Sumas | 0 | 3 | 15 | 17 | s.o. | 0 | 10 | 7 | 1 | U | 1 |
| Abbotsford - Flats | 1 | 2 | 7 | 7 | 0 | 2 | 6 | 7 | 0 | 7 | 7 |
| Aldergrove, Langley, Surrey | 3 | 11 | 18 | 25 | 2 | 3 | 6 | 25 | 0 | 25 | 20 |
| Surrey | 1 | 0 | 8 | 9 | 1 | 3 | 7 | 9 | 0 | 9 | 1 |
| Delta | 0 | 11 | 0 | 11 | s.o. | 0 | 0 | 0 | 0 | 11 | 1 |
| Richmond | 0 | 0 | 3 | 3 | s.o. | 0 | 3 | 3 | 0 | 3 | 1 |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Squamish à Calcheak | 1 | 0 | 11 | 11 | 0 | 2 | 1 | 8 | 6 | U | 0 |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Péninsule Sechelt (Gibson's à Doriston) | 0 | 12 | 9 | 18 | s.o. | 2 | 1 | 6 | 16 | U | 0 |
| Saltery Bay à Lund | 1 | 0 | 6 | 7 | 1 | 0 | 1 | 3 | 7 | 2 | 0 |
| Bassin de la Tzoonie | 0 | 0 | 2 | 2 | s.o. | 0 | 0 | 0 | 2 | 0 | 0 |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Inlet Loughborough | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Île North Broughton | 0 | 0 | 1 | 1 | s.o. | 0 | 0 | 0 | 1 | 0 | 0 |
| Inlet Kingcome | 2 | 0 | 0 | 2 | 2 | 0 | 0 | 0 | 1 | 0 | 0 |
| Lac Lottie, au nord du passage Well's | 0 | 0 | 2 | 2 | s.o. | 0 | 0 | 0 | 2 | 0 | 0 |
| Île Bramham | 0 | 1 | 0 | 1 | s.o. | 0 | 0 | 0 | 0 | 0 | 0 |
| Île Greaves | 0 | 0 | 2 | 2 | s.o. | 0 | 0 | 0 | 2 | 0 | 0 |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Metchosin a Shirley (Sooke) | 2 | 2 | 4 | 8 | 2 | 0 | 4 | 4 | 0 | 8 | 0 |
| Jordan River à Port Renfrew area | 0 | 5 | 15 | 18 | s.o. | 3 | 1 | 8 | 12 | 0 | 0 |
| Carmanah, Bamfield, ruisseau China | 1 | 3 | 16 | 20 | 1 | 6 | 1 | 2 | 13 | 0 | 0 |
| Colwood à la région de Cobble Hill | 8 | 10 | 8 | 20 | 5 | 3 | 6 | 9 | 6 | 20 | 1 |
| Duncan à la région de Crofton | 2 | 11 | 10 | 20 | 1 | 1 | 2 | 4 | 16 | 10 | 2 |
| Nanaimo au lac Cameron | 9 | 8 | 10 | 23 | 6 | 6 | 9 | 11 | 6 | 23 | 7 |
| Port Alberni | 5 | 2 | 9 | 16 | 5 | 0 | 2 | 7 | 11 | 16 | 3 |
| Détroit de Barkley (à l'ouest du chenal Trevor) à Tofino | 5 | 1 | 22 | 26 | 3 | 11 | 7 | 9 | 12 | 0 | 0 |
| Baie Clayoquot | 0 | 16 | 0 | 16 | s.o. | 0 | 0 | 0 | 15 | 0 | 0 |
| Bowser à Nootka | 2 | 6 | 18 | 24 | 2 | 7 | 3 | 8 | 9 | 10 | 5 |
| Rivière Tsolum au ruisseau Black | 1 | 2 | 5 | 7 | 0 | 1 | 0 | 4 | 4 | 4 | 3 |
| Campbell River à Sayward | 5 | 17 | 14 | 35 | 4 | 4 | 1 | 4 | 31 | 0 | 0 |
| Région de Woss | 1 | 5 | 5 | 10 | 1 | 1 | 0 | 3 | 8 | 0 | 0 |
| Kyuquot à la péninsule Brooks | 6 | 0 | 0 | 6 | 6 | 5 | 0 | 0 | 1 | 0 | 0 |
| Port McNeill à Shushartie et au cap Scott | 3 | 0 | 15 | 18 | 3 | 3 | 2 | 5 | 15 | 0 | 0 |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Île Malcolm | 0 | 0 | 2 | 2 | s.o. | 1 | 1 | 1 | 1 | 0 | 0 |
| Île Turnour | 0 | 0 | 1 | 1 | s.o. | 0 | 0 | 0 | 1 | 0 | 0 |
| Île Quadra | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 |
| Range | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Île Denman Island | 0 | 0 | 1 | 1 | s.o. | 1 | 0 | 0 | 1 | U | 0 |
| Île Texada Island | 0 | 0 | 3 | 3 | s.o. | 0 | 1 | 1 | 2 | 3 | 0 |
| Île Gambier Island | 0 | 0 | 1 | 1 | s.o. | 0 | 1 | 0 | 1 | U | 0 |
| Île Bowen Island | 0 | 0 | 1 | 1 | s.o. | 0 | 1 | 1 | 0 | U | 0 |
| Île Ruxton (près de Valdes) | 0 | 0 | 1 | 1 | s.o. | 0 | 1 | 0 | 0 | U | 0 |
| Île Galiano Island | 0 | 4 | 0 | 4 | s.o. | 1 | 4 | 4 | 1 | 0 | 0 |
| Île Saltspring Island | 0 | 5 | 1 | 6 | s.o. | 0 | 3 | 6 | U | 6 | 2 |
| Île Pender Island | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
| Île Saturna Island | 0 | 1 | 1 | 1 | s.o. | 1 | 1 | 1 | 0 | U | 0 |
| % du total échantillonné | Période Avant-1983 |
Période 1983 - 2003 |
Période >2003 - 2014 |
Période Total pour toutes les années |
Période Non documentée de nouveau depuis avant 1983 |
Protection du site / Menace Dans des aires protégées |
Protection du site / Menace Urbanisation |
Protection du site / Menace Routes |
Protection du site / Menace Exploitation forestière |
Protection du site / Menace Espèces envahissantes |
Protection du site / Menace Pollution |
|---|---|---|---|---|---|---|---|---|---|---|---|
| - | 76 | 168 | 341 | 525 | 55 | 86 | 106 | 232 | 222 | 202 | 100 |
| Tous les ans | 14 | 32 | 65 | - | - | 16 | 20 | 44 | 42 | 39 | 19 |
| Avant 1983 | - | - | - | - | 72 | - | - | - | - | - | - |
Dans la zone continentale, la grenouille à pattes rouges du Nord est présente depuis la vallée du bas Fraser jusqu'aux environs de Hope vers l'est, puis le long du corridor Sea-to-Sky vers le nord jusqu'à Whistler, et le long de la Sunshine Coast et de la côte centrale jusqu'au détroit de Smith, juste au nord du cap Caution. Des spécimens prélevés au parc Manning en 1945 (RBCM, nos 816 et 817) étaient incorrectement identifiés; il s'agit plutôt de grenouilles maculées de Columbia (inspection par K. Ovaska en août 2014). Plus loin au nord, il existe des mentions isolées près de Kitimat, sur la côte centrale (RBCM, nos 1199 et 1200). Les spécimens en question n'ont pu être retracés, et il est probable qu'ils aient été confondus avec la grenouille maculée de Columbia, qui se trouve dans la région. Un individu observé dans la région de Cariboo en 2004 (Packham, comm. pers., 2013) peut aussi être une grenouille maculée de Columbia.
La présence de la grenouille à pattes rouges du Nord dans l'archipel Haida Gwaii a été documentée pour la première fois en 2001, mais l'espèce y était probablement présente depuis bien avant cette année-là (Ovaska et al., 2002). Elle a été observée dans 10 sites près de Port Clements, dans l'île Graham, autant dans des secteurs habités qu'éloignés. En 2014, elle a été trouvée en abondance quelque 40 km plus loin, à l'extrémité sud de l'ile Graham, près de Skidegate (Ovaska, comm. pers., 2014). La grenouille à pattes rouges du Nord n'a pas été observée dans la réserve de parc national Gwaii Haanas, dans l'île South Moresby (Wojtaszek, comm. pers., 2013). Il est probable que la population de l'île Graham soit le résultat d'une introduction d'origine humaine, semblable à l'introduction délibérée de rainettes du Pacifique (Pseudacris regilla), qui sont aujourd'hui répandues dans l'archipel (Reimchen, 1991). Cependant, sans des recherches plus approfondies, la possibilité que la grenouille à pattes rouges du Nord soit une espèce indigène qui a été ignorée ne peut pas être écartée.
Les mentions obtenues depuis 2004 (à l'exception des mentions de Kitimat et de Haida Gwaii) prolongent la limite nord de l'aire de répartition canadienne connue de la grenouille à pattes rouges du Nord jusqu'à l'île Greaves (Powelson, comm. pers., 2013), soit juste au nord d'un site déjà répertorié dans l'île Bramham (Meggill, comm. pers., 2003). Trois spécimens de musée proviennent d'endroits situés au nord de Powell River (de la région de Kingcome et de l'inlet Loughborough, près de Powell River; CMC, nos 1879 et 1886A, B). L'identification de ces spécimens a été confirmée (en septembre 2003, par Francis Cook, chercheur émérite, Musée canadien de la nature). D'autres observations ont été signalées dans l'île North Broughton, à l'anse Caviar et au nord de Lund. La dernière décennie a vu une augmentation des activités de recherche et de surveillance visant les espèces sauvages en général, notamment dans l'île Calvert et aux environs de Bella Bella. Ces travaux n'ont pas produit de mentions de la grenouille à pattes rouges du Nord, mais des crapauds de l'Ouest (Anaxyrus boreas) et des salamandres foncées (Ambystoma gracile) ont été trouvés (Reynolds, comm. pers., 2014).
Avant 2003, seules quelques vieilles mentions (années 1940) de la présence de la grenouille à pattes rouges du Nord sur la côte Sunshine existaient. Par contre, les données recueillies au cours de la dernière décennie révèlent qu'elle est présente dans la péninsule Sechelt, depuis Gibsons jusqu'au nord de Lund (Mitchell et al., 2012). L'espèce est également présente dans le bassin de la rivière Tzoonie, qui s'étend à l'intérieur des terres depuis l'inlet Sechelt. De nombreux autres bassins versants côtiers n'ont pas fait l'objet de relevés ciblant les amphibiens.
Dans le corridor Sea-to-Sky, les inventaires effectués depuis 2007 ont permis d'ajouter des dizaines de sites à l'unique mention de la grenouille à pattes rouges du Nord datant de 1926, au lac Daisy. L'espèce est dispersée entre Squamish et Whistler dans des milieux humides et des forêts à des altitudes relativement faibles. Seuls quelques relevés ont été réalisés au nord de Whistler jusqu'à Pemberton, et ils n'ont pas produit de mentions de la présence de la grenouille à pattes rouges du Nord (Malt, 2011).
Il existait de nombreux sites historiques dans la vallée du bas Fraser, depuis la côte jusqu'à Hope (de l'ouest à l'est) et depuis la frontière avec les États-Unis jusqu'au pied de la chaîne Côtière (du sud au nord). Des relevés récents indiquent que l'espèce demeure répandue partout dans la région, sauf dans le parc Stanley (Stanley Park Ecology Society, 2010) et les zones urbaines, en particulier Vancouver, Richmond et Delta (Malt, 2013) (voir Activités de recherche). Quelques nouvelles mentions proviennent de vallées de bassins versants qui drainent la chaîne Côtière et se déversent dans les lacs Pitt, Slave et Henderson, situés sur la rive nord du Fraser, et de bassins versants qui drainent le nord de la chaîne Cascades près de Chilliwack. Ces mentions étendent l'aire de répartition historique au-delà des basses terres vers les vallées montagneuses.
Dans l'île de Vancouver, les mentions récentes prolongent l'aire de répartition de la grenouille à pattes rouges du Nord d'environ 60 km vers le nord-ouest, depuis Port Hardy jusqu'au cap Scott (B.C. Ministry of Environment, 2012; McCurdy, comm. pers., 2014). La péninsule Brooks n'a pas fait l'objet de nouveaux relevés ciblant les amphibiens depuis 1981. Il existe de nombreux nouveaux sites dans l'aire de répartition historique de l'espèce partout dans l'île, en particulier dans le nord et dans l'île Nootka, ainsi que dans plusieurs îles des détroits de Johnstone et de Georgia.
On estime la superficie de la zone d'occurrence au Canada à 75 625 km2, d'après la méthode du plus petit polygone convexe contenant toutes les occurrences connues (à l'exception d'Haida Gwaii, où l'espèce est probablement introduite). Cette estimation repose sur les mentions tirées de collections de musées, d'ensembles de données et de rapports de recherche publiés et inédits et d'observations fortuites. La zone d'occurrence inclut des milieux en haute altitude (> 1 100 m) et en mer qui ne constituent pas un habitat propice pour l'espèce.
L'indice de zone d'occupation (IZO) a été calculé selon le nombre de carrés de 2 km de côté occupés par l'espèce. L'IZO a été estimé à 2 588 km2, ce qui est probablement une sous-estimation, les activités de recherche étant incomplètes.
Les activités d'échantillonnage et la sensibilisation à l'espèce ont augmenté au fil du temps, ce qui a amélioré les connaissances sur la zone d'occurrence et l'IZO. La zone d'occurrence et l'IZO n'étaient pas indiqués dans le rapport de situation précédent, mais les calculs reposant sur les mentions d'avant 2003 (compilées dans le présent rapport) ont donné une zone d'occurrence de 67 368 km2 et un IZO de 2 588 km2. Les valeurs de la zone d'occurrence et de l'IZO ainsi obtenues sont respectivement 1,12 et 3,12 fois plus élevées que dans le cas où toutes les mentions (celles ci-dessus et celles obtenues depuis 2003) auraient été incluses. On ne sait pas si l'espèce continue d'être présente dans tous les sites déjà répertoriés. Dans certaines parties de l'aire de répartition de l'espèce (Vancouver, Richmond et Delta), l'IZO a diminué.
Plus de 2 500 mentions (groupées en 524 occurrences dans le tableau 1) ont été utilisées pour établir la répartition de la grenouille à pattes rouges du Nord, y compris la zone d'occurrence et l'IZO. La majorité de ces mentions (60 %) ont été obtenues après 2004, alors que moins de 200 ont été recueillies avant 1983. Comme ces chiffres l'indiquent, les activités de recherche de l'espèce ont été considérables ces dernières années (voir le tableau 2 pour une description des activités de recherche et des endroits où l'espèce a été trouvée).
Description longue pour le tableau 2
| Région du Lower | Corridor Sea-to-Sky | Sunshine Coast | Côte centrale | Île de Vancouver | Îles du détroit de Georgia |
À noter que d'autres relevés généraux de milieux humides ont été menés dans certaines régions dans le cadre d'évaluations environnementales ou d'autres projets.
| Environs | Période | Description des activités de recherche et occurrence |
|---|---|---|
| Basses terres du Fraser | Fin des années 1990 | Relevés des grappes d'œufs dans plus de 94 étangs visant l'habitat de la grenouille maculée de l'Oregon (Rana pretiosa). Taux d'occupation par le R. aurora de 50 % (Haycock et Knopp, 1998). |
| Basses terres du Fraser | 1997–2000 | Grappes d'œufs prélevées le long du chemin Towne, à la périphérie de la prairie Sumas, aux fins d'études écotoxicologiques* (De Solla et al., 2002a,b; Loveridge, 2002). |
| Basses terres du Fraser | 2005–2009 | Relevés des grappes d'œufs dans plus de 100 sites sur une période d'au moins 28 jours pendant 5 ans. Données négatives non disponibles. *Les relevés des grappes d'œufs sur environ 1 km d'habitat situé dans des fossés sur le chemin Towne de la prairie Sumas en 2009 ont confirmé l'occupation récente (Knopp, comm. pers., 2013). |
| Basses terres du Fraser | 2010–2011 | Relevés des grappes d'œufs dans 43 étangs/bassins visant l'habitat de la R. pretiosa et de la tortue peinte de l'Ouest (Chrysemys picta). Taux d'occupation par le R. aurora de 47 % (Pearson, 2010, 2011). |
| Basses terres du Fraser | 2012 | Relevés des grappes d'œufs dans 119 étangs. Sites choisis d'après les données de base existantes, des représentations géographiques et écologiques et l'occurrence connue ou possible de la R. pretiosa et de la C. picta. Taux d'occupation par le R. aurora de 48 %. Les données ont permis de résumer la probabilité d'occupation : nulle pour les sites à Vancouver, Richmond et Delta; très faible (< 0,2) pour les sites d'Abbotsford et de la région combinée de Pitt Meadows, Port Coquitlam et Coquitlam; moyenne (~ 0,41) pour Chilliwack, Burnaby, Vancouver Nord et Maple Ridge; élevée (≥ 0,5) pour Surrey, Mission, Kent, Harrison et la région combinée de Anmore, Belcarra et Port Moody (carte de l'occupation à l'annexe 1) (Malt, 2013). |
| Chilliwack Lac Cultus |
2002–2011 | Relevés dans plus de 40 sites sur une période d'au moins 31 jours pendant 10 ans. Données négatives non disponibles (Knopp, comm. pers., 2013). Pose de pièges-fosses sur des terres du ministère de la Défense nationale à l'USS Chilliwack. Détails non disponibles. Capture de R. aurora (Hawkes, comm. pers., 2013). |
| Chilliwack | 2007 | Récupération de poissons et d'amphibiens dans des fossés (sur une distance de16 000 m) et 14 cours d'eau pendant l'été. Capture de 2 R. aurora dans 1 cours d'eau. Taux d'occupation de 7 % (Blair, 2007). |
| Abbotsford | 2005 | Récupération de poissons et d'amphibiens dans 8 ruisseaux et fossés au moyen d'une épuisette. Capture de R. aurora dans un fossé. Taux d'occupation de 12,5 % (City of Abbotsford, 2005) |
| Delta | 1990–2002 | Pose de pièges en entonnoir pour prélever des poissons et des amphibiens dans 145 plans d'eau (fossés et étangs) de 23 bassins versants. Taux d'occupation des bassins versants par le R. aurora de 43 %. Taux d'occupation des étangs non rapporté. L'espèce a été désignée « peu commune » (Rithaler, 2002a,b, 2003a,b; Danyluk, comm. pers., 2014). Relevé, à la fin de l'été et à l'automne 1999, dans une ferme située près de la route Annacis. Capture de 3 R. aurora juvéniles (Knopp, comm. pers., 2013). |
| Delta | 2012 | Les relevés par Malt (2013) (décrits ci-dessus) ciblaient 4 étangs dans 4 bassins versants à Delta. Taux d'occupation par le R. aurora de 0 %. Comme les coordonnées des sites exacts des observations réalisées de 1990 à 2002 n'étaient pas dans les bases de données et ne figuraient pas non plus sur les cartes archivées de la Corporation de Delta, la comparaison est limitée (Malt, 2013). |
| Parc Stanley Vancouver | 1998–1999 | Relevés standard du RISC, mais aucun détail présenté. Taux d'occupation par le R. aurora de 0 % (Stanley Park Ecology Society, 2010). |
| Parc Stanley Vancouver | 2007–2013 | Pose de 47 pièges en entonnoir dans 4 étangs (lac Beaver, système de biofiltration du Lost Lagoon, étang Beaver et étang Moose) pendant 3 nuits de 2007 à 2009, et effort semblable depuis. Installation de planches, 2 BioBlitz, plus de 100 heures de recherche. Taux d'occupation par le R. aurora de 0 % (Stanley Park Ecology Society 2010; Worcester, comm. pers., 2014). |
| Burnaby | 2011 | Récupération au moyen de pièges à ménés et d'épuisettes lors du drainage d'un bassin de confinement. Capture de 86 R. aurora (EBA, 2011). |
| Réservoir de Coquitlam | 2011 | Pose de pièges-fosses pour prélever des petits mammifères et des amphibiens. Relevés en zone limitée; 111 heures-personnes et 17 757 heures-pièges; 1 site. Capture de 13 R. aurora adultes (Golder and Associates, 2011a). |
| Bassin de la rivière Alouette | 2011–2012 | Relevés et pose de pièges en entonnoir dans 26 sites sur une période d'au moins 10 jours. Données négatives non disponibles (Knopp, comm. pers., 2013). Relevés des grappes d'œufs d'espèces en péril dans 6 étangs. Taux d'occupation par le R. aurora de 50 % (Mitchell, 2013). |
| Mission | 2010 | Pose de pièges en entonnoir et de pièges à ménés. Récupération. Capture de 4 R. aurora (DWB Consulting Services Ltd., 2010). |
| Ruisseau Roche, Vancouver Nord | 2011 | Pose de pièges-fosses pour prélever des musaraignes; 32 pièges × 22 vérifications; 40 pièges × 15 vérifications; 1 site. Capture de 3 R. aurora adultes et de 1 R. aurora d'âge inconnu (Dupuis, 2011). |
| Environs | Période | Description des activités de recherche et occurrence |
|---|---|---|
| Région de Whistler | 2004–2009 | Relevés des grappes d'œufs sur une période d'au moins 6 jours à Pinecrest et dans la zone autour du lac Lava; 27 sites. Données négatives non disponibles (Knopp, comm. pers., 2013). |
| Lac Alice Pinecrest Brandywine |
2007–2010 | Relevés répétés des grappes d'œufs, pose de pièges à ménés et relevés de durée limitée dans 55 étangs. Taux d'occupation par le R. aurora des étangs répartis dans les 3 régions de 60 %. Pose de pièges-fosses, marquage-recapture, relevés sur les routes à Pinecrest (Malt, 2011). |
| Squamish à Pemberton AGF | 2007–2010 | Relevés d'un jour dans le lac Brohm, le lac Cheakamus et l'aire de gestion de la faune proposée dans les milieux humides de la vallée de Pemberton. Espèce non décelée dans la vallée de Pemberton. Relevés ciblant le crapaud de l'Ouest (Anaxyrus boreas) dans la rivière Mamquam, la rivière Squamish et le lac Lucille. Capture de R. aurora à Mamquam (Malt, 2011). |
| Bassins de compensation à Pinecrest et bassins d'eaux pluviales à Cheakamus | 2012 | Relevés visuels depuis la rive, pose de pièges à ménés; 5 bassins × 8 vérifications des pièges; 4 bassins × 3 vérifications. Taux d'occupation de 0 % (Tayless, 2012). |
| Environs | Période | Description des activités de recherche et occurrence |
|---|---|---|
| Péninsule Sechelt | 2010–2011 | Relevés des grappes d'œufs dans 4 étangs et relevés d'été ciblant le C. picta. Taux d'occupation par le R. aurora de 100 % (Mitchell et al., 2012). |
| Powell River | 2010–2011 | Relevés des grappes d'œufs dans 6 étangs et relevés d'été ciblant le C. picta. Taux d'occupation par le R. aurora de 66 % (Mitchell et al., 2012). |
| Île Texada | 2010–2011 | Relevés des grappes d'œufs dans 9 étangs et relevés d'été ciblant le C. picta. Taux d'occupation par le R. aurora de 44 % (Mitchell et al., 2012). |
| Environs | Période | Description des activités de recherche et occurrence |
|---|---|---|
| Région de Bella Bella | 2009–2013 | Relevés dans des cours d'eau et des estuaires ciblant les saumons dans 60 sites de la région, à environ 40 km au nord et au sud de Bella Bella (de l'île Hunter à la péninsule Don) au cours des 5 dernières années. Aucune mention du R. aurora (Reynolds, comm. pers., 2014). |
| Environs | Période | Description des activités de recherche et occurrence |
|---|---|---|
| Sayward | 1997–2001 | Étude par radiotélémétrie pour observer le choix d'habitat et les déplacements dans les coupes à blanc et les peuplements forestiers (Chan-McLeod, 2003), et les blocs de coupe à rétention variable (Chan-McLeod et Wheeldon, 2004; Chan-McLeod et Moy, 2007). |
| Nord-est de l'île de Vancouver | 2002 | Relevés dans 113 petits milieux humides, dont 28 étangs éphémères asséchés, 50 % à une altitude de plus de 900 m. Taux d'occupation d'environ13 % (Wind, 2003). |
| District forestier de North-Central Island | 2006–2012 | Relevés des grappes d'œufs du R. aurora dans 58 milieux humides. Taux d'occupation de 45 % (B.C. Ministry of Environment, 2012). |
| Bassin de la rivière Campbell | 2001 | Relevés et pose de pièges-fosses dans 15 sites (pose de 3 ensembles de pièges-fosses dans chacun des 3 sites). Taux d'occupation de 53 %. Aucun R. aurora capturé dans les pièges-fosses (Garcia, 2001). |
| Rivière Salmon | 2003 | Pose de pièges-fosses (4 ouverts sur une période de 2,5 mois, vérifiés chaque semaine). Capture de 2 R. aurora (Materi et Forrest, 2004). |
| District forestier de Campbell River | 2006–2012 | Relevés des grappes d'œufs du R. aurora dans 35 milieux humides. Taux d'occupation de 40 % (B.C. Ministry of Environment, 2012). |
| Région de Quinsam | 2011–2012 | Relevés dans 5 étangs, 15,2 heures-personnes. Taux d'occupation par le R. aurora de 40 %. Récupération de grappes d'œufs dans 4 étangs; 0,48 ha, 5 heures × 2 personnes. Capture de 2 R. aurora dans un étang (Golder Associates, 2011b, 2012). |
| Baie Clayoquot | 1998–1999 | Pose de pièges en entonnoir et relevés sur les rives de 148 étangs de 6 unités d'aménagement de bassins versants. Taux d'occupation des étangs de 26 % et des bassins versants de 83 % (Baley et al., 2000). |
| Tofino, Ucluelet Pacific Rim |
2008–2013 | Relevés des grappes d'œufs dans 45 étangs. Taux d'occupation par le R. aurora de 84 % (reproducteurs) et de 89 % (tous les stades) (Beasley, 2011; Beasley, données inédites, 2014). |
| Sud-est de l'île de Vancouver | 2002 | Relevés dans 122 petits milieux humides, dont 44 étangs éphémères asséchés, la plupart à des altitudes de moins de 600 m. Taux d'occupation par le R. aurora d'environ 23 % (Wind, 2003). |
| Bassin de la rivière Englishman | 2008 | Relevés des grappes d'œufs et pose de pièges en entonnoir dans le lac Healy (20 pièges × 1 nuit) et le lac Shelton (25 pièges × 1 nuit). Occupation par le R. aurora dans les 2 sites (Wind, 2008b). |
| District forestier de South Island | 2006–2012 | Relevés des grappes d'œufs du R. aurora dans 104 milieux humides. Taux d'occupation par le R. aurora de 57 % (B.C. Ministry of Environment, 2012). |
| Lacs Nanaimo | 2007 | Relevés des grappes d'œufs et relevés d'été dans 68 milieux humides, dans 34 sites différents. Taux d'occupation par le R. aurora de 62 à 74 % (adultes) et de 4 à 19 % (reproducteurs). La première valeur de la plage indique le pourcentage dans les étangs avant la récolte, et la deuxième, le pourcentage après la récolte (Wind, 2008a). |
| Nanaimo | 2011 | Récupération d'amphibiens dans un complexe de milieux humides asséchés. Capture de 6 R. aurora (EcoDynamic Solutions, 2011). |
| Péninsule Saanich | 2002 | Information recueillie dans plus de 30 étangs pour établir la répartition du ouaouaron (Rana catesbeiana). R. aurora observés dans plusieurs étangs, mais la plupart de ces derniers n'ont pas été explorés à fond pour y établir la présence de l'espèce. Lors des activités intensives de marquage-recapture de ouaouarons dans 4 étangs, aucun R. aurora n'a été capturé dans l'un, un faible nombre l'a été dans 2, et un grand nombre l'a été dans le quatrième (Govindarajulu, 2003; Govindarajulu, comm. pers., 2014). |
| Bassin de la rivière Jordan | 2004–2005 | Relevés de durée limitée (5 dates, 2 à 3 personnes, durée totale de 57 heures) et relevés sur les routes (2 dates, 2 personnes, distance totale de 37 km). Capture de 45 R. aurora (Hawkes, 2005). |
| Bassin de la rivière Jordan | 2012 | Relevés des grappes d'œufs dans 2 étangs aménagés. Taux d'occupation par le R. aurora de 100 % (Tuttle, 2013). |
| Environs | Période | Description des activités de recherche et occurrence |
|---|---|---|
| Îles Gulf | 1996 | Relevés de durée limitée (de 0,5 à 6,3 h/site) dans 10 îles et installation d'abris artificiels dans 4 îles visant la couleuvre à queue fine (Contia tenuis). Taux d'occupation des îles (Galiano, Pender, Saltspring, Saturna) par le R. aurora de 40 % (Engelstoft et Ovaska, 1997, 1998). |
| Île Saltspring Baie Burgoyne | 2011 | Relevés de durée limitée lors d'une journée de BioBlitz. Capture de 3 R. aurora adultes (Briony Penn Associates, 2011). |
La majeure partie des activités récentes ont porté sur l'exploration de nouveaux endroits hors des limites de l'aire de répartition connue de l'espèce, notamment la Sunshine Coast (Mitchell et al., 2012) et le corridor Sea-to-Sky (Malt, 2011). Bien qu'aucun relevé ciblé n'ait été mené sur la côte centrale, des groupes universitaires et sans but lucratif ont effectué des recherches sur le saumon dans de nombreux cours d'eau de la région de Bella Bella au cours des dix dernières années (Reynolds, comm. pers., 2014), mais ils n'ont pas trouvé de grenouilles à pattes rouges du Nord.
Le ministère des Forêts, des Terres et de la Gestion des Ressources naturelles de la Colombie-Britannique a mené, dans l'île de Vancouver, les activités de recherche les plus intensives (B.C. Ministry of Environment, 2012). Il a réalisé un vaste recensement des individus reproducteurs de la grenouille à pattes rouges du Nord dans près de 200 milieux humides en forêts aménagées dans 3 districts forestiers. L'espèce a été trouvée dans la moitié des milieux recensés (soit dans 99 des 197 étangs, dont certains se trouvaient à proximité les uns des autres; tableau 2). Le but était de trouver les milieux humides boisés propices à protéger en vertu de la Forest and Range Practices Act de la Colombie-Britannique.
Plusieurs groupes sans but lucratif et le ministère des Transports et de l'Infrastructure de la Colombie-Britannique sont de plus en plus sensibilisés à la mortalité routière, qui tue un grand nombre d'amphibiens. Les relevés sur des routes et dans des milieux humides adjacents ont permis de découvrir de nombreuses occurrences de l'espèce (Blood et Henderson, 2000; Beasley, 2006; Materi, 2008; Clegg, 2011; Wind, 2012). Des études d'impact sur l'environnement et des activités de récupération menées par plusieurs consultants ont produit les mêmes observations.
Peu de relevés ont été réalisés dans les sites historiques. Le lac Marion (aussi appelé lac Jacobs) est l'un des 2 sites où la quasi–totalité des données démographiques sur la grenouille à pattes rouges du Nord ont été recueillies dans les années 1970 (Calef, 1973 a, b). Cette population n'a pas fait l'objet d'un nouveau relevé (Richardson, comm. pers., 2014), même si elle est présente dans la forêt expérimentale de l'Université de la Colombie-Britannique. Les mêmes plans d'eau d'un autre site de recherche historique dans la rivière Little Campbell (Licht, 1969) n'ont pas été visités de nouveau, mais les étangs voisins ont récemment été recensés par des volontaires du groupe A Rocha, sans but lucratif (Baylis, comm. pers., 2014). L'espèce continue de se reproduire à 600 m du site historique.
Un relevé d'envergure a été mené en 2012 pour modéliser l'occupation des milieux humides dans la région métropolitaine de Vancouver et la vallée du bas Fraser (Malt, 2013; annexe 1). Un milieu humide a été choisi au hasard dans chaque carré de 10 km de côté d'une grille superposée sur la région. On a mené un relevé dans les 64 milieux humides ainsi obtenus pour établir l'occupation par des grenouilles à pattes rouges du Nord en état de se reproduire (présence de grappes d'œufs). Des relevés ont également été menés dans 55 autres milieux humides dans le cadre de la surveillance communautaire continue dans le bassin de la rivière Little Campbell et de travaux en cours sur 2 autres espèces en péril, soit la grenouille maculée de l'Oregon et la tortue peinte de l'Ouest (Chrysemys picta) (Pearson, 2010, 2011, 2012; Mitchell et al., 2012). Ces relevés ont permis d'établir la présence de la grenouille à pattes rouges du Nord dans 57 milieux humides (voir Menaces et facteurs limitatifs : Développement résidentiel et commercial pour les résultats du modèle).
Il existe des occurrences historiques notables où l'espèce n'a pas été détectée dans le cadre de relevés récents, et ce, malgré un effort considérable. Ainsi, elle n'a pas été trouvée dans le parc Stanley depuis les années 1970. Ces six dernières années, les activités de recherche, menées sur plusieurs saisons, incluent des captures par piège en entonnoir, des relevés visuels des grappes d'œufs, la vérification de planches et des demandes de données d'observation auprès de naturalistes et d'autres utilisateurs du parc (Stanley Park Ecology Society, 2010; Worcester, comm. pers., 2014). Le degré des activités de recherche dans le parc Stanley est suffisant et permet d'affirmer que l'espèce y est disparue. Vancouver, Delta et Richmond sont trois autres zones d'occurrence historiques. Elles ont été échantillonnées en 2012 par des observateurs chevronnés, qui n'ont pas trouvé l'espèce lors des relevés des grappes d'œufs (Malt, 2013). Toutefois, comme les activités de recherche portaient sur seulement deux à quatre étangs par zone au cours d'une seule saison, un plus grand effort est requis avant de pouvoir tirer des conclusions définitives.
La grenouille à pattes rouges du Nord vit dans les forêts humides de basse altitude. Elle a besoin de milieux aquatiques pour se reproduire et de milieux terrestres pour s'alimenter, répartis dans une configuration spatiale lui permettant d'accomplir les différents stades de son cycle vital. L'hibernation peut avoir lieu dans le tapis forestier ou dans l'eau (Licht, 1969; Ritson et Hayes, 2000, in Hayes et al., 2008). Dans l'État de l'Oregon, des adultes ont hiverné dans un petit étang de 0,03 ha situé à 50 m d'un plus grand étang de 0,6 ha utilisé pour la reproduction (Hayes et Rombough, 2004). Autrement, les besoins précis de l'espèce en sites d'hivernage sont méconnus.
L'espèce a été observée à des altitudes variant du niveau de la mer à 860 m dans l'État de Washington et à 1 427 m dans l'État de l'Oregon (Leonard et al., 1993). En Colombie-Britannique, le site le plus élevé a été observé à 1 040 m (Wind, 2003), mais la plupart des mentions dans la province sont observées à des altitudes inférieures à 500 m. Dans la région de la baie Clayoquot, sur la côte ouest de l'île de Vancouver, Beasley et al. (2000) ont observé la grenouille à pattes rouges du Nord plus fréquemment dans les milieux humides situés à moins de 500 m d'altitude (taux d'occupation de 30 %) que dans ceux situés à plus de 500 m d'altitude (taux d'occupation de 14 %). Wind (2003) a mené, dans l'île de Vancouver, des relevés d'amphibiens dans 236 milieux humides à des altitudes variant entre le niveau de la mer et 1 200 m. L'altitude moyenne d'occurrence de l'espèce était de 515 m, mais la plupart des sites étaient à plus basse altitude (mode = 180 m).
La grenouille à pattes rouges du Nord se reproduit dans une variété de plans d'eau permanents et temporaires, comme des fondrières, des étangs, des fossés, des sources, des marais, les rives de grands lacs et les tronçons de rivières à courant faible (Blaustein et al., 1995, et références citées dans ce document). Une salinité supérieure à 4,5 % est létale pour les embryons, tout comme des eaux trop acides (pH ≤ 3,5) ou trop alcalines (pH ≥ 9), et les températures de l'eau supérieures à 21º C ne leur sont pas propices (Hayes et al., 2008, et références citées dans cet article). Une végétation émergente abondante est généralement présente aux sites de reproduction (Adams, 1999; Ostergaard et Richter, 2001; Pearl et al., 2005; Adams et al., 2011). Les femelles pondent leurs œufs dans des eaux calmes (vitesse du courant < 5 cm/sec), à des endroits ensoleillés pendant au moins une partie de la journée (Licht, 1969; Richter et Azous, 1995). Wind (2008a) a trouvé un grand nombre de grenouilles à pattes rouges du Nord adultes dans de petits étangs situés en milieu forestier dans la région des lacs Nanaimo, mais ceux-ci étaient très peu utilisés aux fins de reproduction. Les grenouilles se reproduisaient plus souvent dans certains de ces étangs après que la coupe du bois ait réduit le couvert forestier autour de ceux-ci.
Dans les basses terres du Puget Sound de l'État de Washington, l'espèce a le plus souvent été trouvée dans des milieux humides à faible pente et exposés au sud; ces seules caractéristiques pouvaient expliquer 63 % de la variation de l'occupation des milieux humides (Adams, 1999). La reproduction avait lieu dans des plans d'eau permanents qui avaient tendance à être de vastes milieux humides à structure complexe. Dans le même État, Ostergaard et Richter (2001) ont observé des individus se reproduisant dans des étangs de collecte des eaux pluviales (c.-à-d. de petits réservoirs naturels ou modifiés pour le stockage des eaux pluviales). La présence de l'espèce était positivement corrélée avec la complexité du milieu humide, laquelle correspond au rapport entre la couverture de végétation émergente et la superficie des eaux libres, ainsi qu'avec le pourcentage du couvert forestier environnant. Les grappes d'œufs étaient les plus nombreuses dans les étangs où le couvert forestier comptait pour plus de 30 % de la superficie à moins de 200 m des berges. Adams et al. (2011) ont établi que les milieux humides dont plus de 35 % des rives sont couvertes d'arbres de plus de 5 m de hauteur à moins de 5 m de la rive montraient la plus faible probabilité de connaître la disparition locale de l'espèce dans la vallée de la Willamette, en Oregon. Dans la baie Clayoquot, dans l'île de Vancouver, la grenouille à pattes rouges du Nord était plus fréquemment observée dans des tourbières ombrotrophes et minérotrophes que dans d'autres types de milieux humides comme les marécages, les marais et les parties peu profondes de grands plans d'eau (Beasley et al., 2000).
Adams (2000) a rapporté que le taux de survie des têtards de grenouille à pattes rouges du Nord dans des enclos expérimentaux était hautement variable d'un site à l'autre, mais tendait à être plus faible dans les milieux humides permanents que dans les milieux humides temporaires. Cette différence était peut-être due aux gradients d'habitat ou aux effets des prédateurs. Ces résultats semblent indiquer que les plans d'eau permanents, qui abritent plus de prédateurs, peuvent constituer un habitat puits plutôt qu'un habitat source pour les recrues à la population. Toutefois, il convient d'interpréter l'étude avec circonspection avant d'inférer le caractère propice de sites de reproduction d'après des données faisant uniquement état de la présence ou de l'absence de l'espèce puisque celles-ci ne révèlent rien sur les tendances de survie des larves et des individus nouvellement métamorphosés.
Les individus métamorphosés passent une bonne partie de leur vie dans des milieux terrestres, et des juvéniles et des adultes sont souvent observés en hautes terres, à proximité de petits milieux humides ou sur les berges boisées de cours d'eau (Blaustein et al., 1995, et références citées dans ce document). Chan-McLeod et Wheeldon (2004) ont observé des grenouilles utilisant des parcelles résiduelles de forêts anciennes (> 140 ans) de plus de 0,7 ha dans des blocs de coupe à rétention variable de 1 an. Des études radiotélémétriques indiquent que les adultes privilégient les forêts dont le sous-étage comporte une structure complexe et qu'ils sont souvent associés à des débris ligneux et au polystic à épées (Polystichum munitum) (Haggard, 2000; Ritson et Hayes, 2000, in Hayes et al., 2008; Schuett-Hames, 2004). Les individus radiopistés étaient relativement sédentaires après avoir atteint des milieux terrestres; ils se déplaçaient sur une distance de moins de 10 m par jour et parcouraient jusqu'à 80 m par saison (Hayes et al., 2008, et références citées dans cet article). Durant un été sec, les grenouilles radiopistées restées généralement près des berges de cours d'eau forestiers (Chan-McLeod, 2003; voir Déplacements et dispersion). Lorsque les conditions sont propices, ces grenouilles peuvent être observées sur le tapis forestier loin des plans d'eau; des individus ont été observés à une distance maximale de 400 m d'un plan d'eau pendant des nuits pluvieuses (Nussbaum et al., 1983; Beasley, données inédites, 2014).
La grenouille à pattes rouges du Nord occupe une variété de types et d'âges de forêts, mais semble plus abondante dans les peuplements anciens et humides (questions traitées dans Waye, 1999, et Blaustein et al., 1995; voir aussi MENACES ET FACTEURS LIMITATIFS pour connaître les liens avec la foresterie). Dans les monts Cascades de l'État de Washington, l'espèce était le plus abondante dans les peuplements mûrs (de 80 à 190 ans) et le moins abondante, dans les jeunes peuplements (de 55 à 75 ans) (Aubry et Hall, 1991). L'abondance était négativement corrélée avec l'élévation et l'inclinaison du terrain. Les captures étaient également associées à des conditions modérément humides dans les peuplements relativement anciens; les peuplements anciens et très humides semblent plutôt moins propices. D'après l'étude d'un ensemble plus récent de séries temporelles de données recueillies dans une forêt secondaire à dominance de douglas (Pseudotsuga menzeisii), Aubry (2000) a établi que l'espèce était plus abondante dans les peuplements approchant de l'âge de la récolte (de 50 à 70 ans) que dans les jeunes peuplements, où seules de rares captures ont été réalisées. Les peuplements approchant l'âge d'exploitabilité avaient un couvert dense et comptaient des arbres d'une hauteur de 30 à 45 m; la couche d'herbes et d'arbustes s'était rétablie, mais la quantité de gros débris ligneux était faible comparativement à celle de vieux peuplements. Selon une étude menée dans l'État de Washington et en Oregon, l'espèce était le plus abondante dans des milieux de faible altitude et relativement peu inclinés, mais il n'y avait aucune relation entre l'âge des peuplements (classés comme vieux, mûrs ou jeunes) et l'abondance (Bury et al., 1991). Il est probable que l'association entre l'espèce et l'âge des forêts varie selon l'emplacement géographique, le type de forêts, l'humidité ou d'autres conditions.
En Colombie-Britannique, les mentions de répartition et les observations anecdotiques laissent supposer que l'espèce est commune dans les forêts secondaires et qu'elle se trouve également dans des jardins de banlieue et des étangs saisonniers sur des terres agricoles et des pâturages adjacents à des aires boisées. Dans l'île de Vancouver, Wind (2003) a trouvé l'espèce dans des milieux humides situés autant dans des zones récemment déboisées (moins de 5 ans) que des peuplements plus vieux (plus de 6 à 120 ans et plus). L'abondance relative et le taux de survie n'ont pas été étudiés (voir MENACES ET FACTEURS LIMITATIFS pour un exposé sur les effets du déboisement sur l'espèce).
Les grenouilles à pattes rouges du Nord adultes s'éloignent sur une distance allant jusqu'à 4,8 km de leur site de reproduction (Hayes et al., 2001, 2007), tandis que les juvéniles peuvent se disperser sur plus de 500 m depuis le milieu humide où ils se sont développés (Beasley, données inédites, 2014). Les sentiers et les haltes temporaires incluent des forêts situées en terrain élevé, des ruisseaux, des zones riveraines, des zones de suintement, et des milieux humides à végétation émergente, arbustifs/broussailleux ou boisés (Hayes et al., 2008, et références citées dans cet article). Les déplacements des adultes radiopistés déplacés dans le cadre d'une expérience révèlent que les coupes à blanc constituent des obstacles par temps chaud et sec dans le nord de l'île de Vancouver (Chan-McLeod, 2003). Lorsque la perméabilité augmentait durant les jours pluvieux et à des températures plus basses, les individus pouvaient parcourir plus de 190 m (en ligne droite) à travers les coupes à blanc durant 2 à 3 jours pluvieux (Chan-McLeod, 2003). Les grenouilles n'empruntaient pas les rives des ruisseaux traversant des coupes à blanc (Chan-McLeod 2003) et n'utilisaient pas les parcelles boisées résiduelles comme relais à travers les blocs de coupe à rétention variable (Chan-McLeod et Moy, 2007). Elles étaient toutefois attirées par les petits cours d'eau en milieu boisé de plus de 1,8 m de largeur (Chan-McLeod, 2003) et, lorsqu'elles ont été placées dans des parcelles boisées résiduelles, elles sont restées le plus longtemps dans celles sillonnées par des cours d'eau (Chan-McLeod et Moy, 2007). Les grenouilles ont aussi effectué des déplacements directionnels délibérés vers des parcelles boisées résiduelles en fonction de la superficie et de la distance de la parcelle. Les déplacements sélectifs vers une parcelle boisée résiduelle de taille moyenne ne se produisaient pas si les grenouilles étaient à plus de 20 m de distance, mais elles se déplaçaient vers les grandes parcelles (> 0,8 ha) à 50 m de distance (Chan-McLeod et Moy, 2007).
Au cours du siècle dernier, les activités humaines ont mené à la modification des parcelles d'habitat de la grenouille à pattes rouges du Nord dans la plus grande partie de son aire de répartition en Colombie-Britannique. La dégradation et la perte d'habitat sont considérables dans le sud et l'est de l'île Vancouver, la vallée du bas Fraser, le corridor Sea-to-Sky et certaines parties de la Sunshine Coast. Le taux et l'irréversibilité de la perte d'habitat sont les plus élevés dans ces régions à cause de l'urbanisation, de l'agriculture intensive et de l'aménagement de routes. La propagation du ouaouaron (Lithobates catesbeianus), espèce introduite, a entraîné la dégradation de presque toutes les parcelles d'habitat fragmenté qui restent dans la vallée du bas Fraser et le sud-est de l'île de Vancouver.
L'altération de l'habitat dans les régions plus éloignées du nord et de l'ouest de l'île de Vancouver et sur la côte continentale au nord de Powell River est principalement le résultat de l'exploitation forestière sur des terres publiques. Les impacts des pratiques forestières assujetties à la Forest and Range Practices Act pourraient être moins graves et de plus courte durée que les impacts d'autres activités humaines, dans la mesure où les milieux humides sont protégés, l'aménagement de routes est fait avec rigueur de sorte à modifier le moins possible les régimes hydrologiques, les structures forestières telles que les arbres abattus sont gardées et la régénération naturelle des forêts peut suivre son cours.
Des cartes thématiques de base produites par Hectares B.C. (2014) ont été utilisées pour obtenir des valeurs récentes de la couverture terrestre. Des valeurs ont été calculées pour les altitudes de 500 m et moins dans chacune des 2 régions du ministère de l'Environnement (Ministry of Environment) de la Colombie-Britannique où la grenouille à pattes rouges du Nord se trouve. La région de l'île de Vancouver comprend l'île de Vancouver et les îles Gulf, ainsi qu'un secteur adjacent sur le continent, au nord de la Sunshine Coast. La région du Lower Mainland s'étend de Vancouver à Hope et Pemberton, et inclut la Sunshine Coast.
Région de l'île de Vancouver : Les zones urbaines couvraient environ 3,3 % (environ 845 km2) de l'île en 2006 (Hectares BC, 2014). Celles du côté sud-est de l'île devraient prendre de l'expansion à mesure que les populations humaines continuent de croître. Par exemple, le district régional de Nanaimo prévoyait une augmentation du nombre d'habitants de 60 % d'ici 2036, accompagnée d'une demande de plus de 30 000 unités unifamiliales non attenantes, de 9 600 maisons en rangée et de 7 600 appartements (Urban Futures, 2007). Malgré des stratégies visant à accroître la densité des logements et de limiter l'étalement urbain, le district régional de la Capitale a connu, de 2001 à 2007, une augmentation de la superficie du territoire urbanisé de 3 % (421 ha) dans sa zone de confinement urbain et de 12 % (1 200 ha) à l'extérieur de celle-ci (Capital Regional District, 2008). L'autoroute à 4 voies aménagée entre Campbell River et Nanaimo en 2001 facilite le développement vers le nord.
L'exploitation forestière touche une grande partie des forêts de faible altitude (< 500 m) dans cette région. Environ 16 % (4 130 km2) ont récemment été déboisées (il y a moins de 20 ans) et 36 % (9 380 km2) sont de jeunes peuplements (moins de 140 ans) (Hectares B.C., 2014). De récents blocs de coupe sont visibles dans les grands massifs de peuplements de forêt ancienne au nord-ouest de l'inlet Knight (Google Earth, 2013). Une amélioration des parcelles d'habitat est prévue à mesure que les forêts de seconde venue mûrissent et que les milieux humides sont restaurés; toutefois, dans presque l'ensemble de l'île, l'exploitation de forêts de seconde venue à faible altitude est en cours, et les parcelles d'habitat régénérées subissent de nouveau une dégradation. Des zones d'habitat d'espèces sauvages (« Wildlife Habitat Areas », ou WHA) établies dans le cadre de la stratégie de gestion des espèces sauvages désignée (« Identified Wildlife Habitat Guidelines ») ont permis de protéger environ 336 ha d'habitat de reproduction et d'alimentation important de la grenouille à pattes rouges du Nord dans des forêts mûres de seconde venue et des forêts anciennes, et ces superficies devraient s'accroître légèrement au fil du temps (voir Protection et propriété de l'habitat).
Région du Lower Mainland : La grande région métropolitaine de Vancouver et la vallée du bas Fraser constituaient probablement une partie importante de l'aire de répartition de la grenouille à pattes rouges du Nord en Colombie-Britannique dans le passé. En effet, cette région comptait autrefois des parcelles d'habitat productives et étendues de forêts et de milieux humides de faible altitude. Depuis la colonisation de la région par les Européens, la majeure partie du couvert forestier a définitivement disparu, et les milieux humides, qui occupaient auparavant 10 % du territoire, n'en occupaient plus que 1 % en 1990 (Boyle et al., 1997). En 2006, les zones urbaines couvraient environ 13 % (1 260 km2) de la région, et les terres agricoles, 8 % supplémentaires (7 760 km2) (Hectares B.C., 2014). L'urbanisation et l'agriculture continuent de s'intensifier dans la région, qui connaît l'un des taux de croissance les plus rapides au Canada. La population humaine de la région du Lower Mainland a grimpé de 102 % entre 1971 et 2006 (Statistics Canada, 2009). Cette croissance ne se limite plus à Vancouver; elle touche aussi les collectivités de la vallée du bas Fraser entre Surrey et Chilliwack, ainsi que dans le corridor Sea-to-Sky. Il est prévu que les populations humaines continueront de doubler au cours des 20 à 30 prochaines années dans le district régional de la vallée du Fraser (Fraser Valley Regional District, 2004) et le district régional de Squamish-Lillooet (Squamish-Lillooet Regional District, 2008). Par conséquent, la pression en faveur de l'aménagement des terres, y compris celles abritant les derniers milieux humides et peuplements de forêt mûre, se fait intensément sentir. Bien que les stratégies de croissance incluent des objectifs en vue d'accroître les densités dans les zones de confinement urbain, l'étalement urbain devrait se poursuivre. Par exemple, la population de la région métropolitaine de Vancouver devrait croître de plus de 35 000 personnes par année, alors que le nombre d'unités résidentielles devrait augmenter, d'ici 2040, de 31 % dans les zones urbaines et de 1 % dans les aires rurales, agricoles, récréatives et de conservation (Metro Vancouver, 2014).
Sur la Sunshine Coast, le développement résidentiel le long de la côte entre Gibsons et Powell River a augmenté au cours de la dernière décennie, ce qui s'est traduit par la modification et la perte de milieux humides et du couvert forestier adjacent. Le développement résidentiel devrait continuer de s'accroître de 12,8 % au cours de la prochaine décennie (2015-2025; BC Stats, 2014). L'exploitation forestière touche également de grands secteurs, en particulier dans les parcelles d'habitat de faible altitude le long de la côte (Sunshine Coast Regional District, 2003; carte du couvert forestier).
La grenouille à pattes rouges du Nord a un cycle vital biphasique caractéristique des amphibiens à reproduction aquatique de l'hémisphère Nord. Les œufs sont pondus dans l'eau, passent au stade de larves aquatiques, qui se métamorphosent en grenouilles juvéniles capables de quitter le milieu aquatique. Les mâles et les femelles juvéniles passent respectivement de un à deux ans et de deux à quatre ans à s'alimenter dans les milieux terrestres et riverains avant d'atteindre la maturité sexuelle et de retourner en milieu aquatique pour se reproduire. En dehors de la saison de reproduction, les adultes sont principalement terrestres et peuvent être observés loin de tout plan d'eau.
Licht (1969, 1971, 1974) et Calef (1973a,b) ont étudié le taux de survie et la biologie de la reproduction de la grenouille à pattes rouges du Nord dans le sud de la Colombie-Britannique. Leurs études demeurent les plus détaillées à ce jour en ce qui concerne la biologie et l'histoire naturelle de l'espèce au Canada. Une étude par marquage-recapture menée à Umpqua, en Oregon, fournit les seules données démographiques connues pour l'espèce aux États-Unis (Hayes, comm. pers., 2012).
La grenouille à pattes rouges du Nord a une reproduction de type explosif (sensu Wells, 1977). Les adultes se rassemblent aux sites de reproduction pendant une courte période (de 2 à 4 semaines) à la fin de l'hiver ou au début du printemps (selon l'altitude), souvent immédiatement après la rupture des glaces. Les mâles lancent des appels distincts, et ils le font généralement sous l'eau. Par conséquent, le chant nuptial est à peine perceptible par l'oreille humaine, voire inaudible, à partir de la surface de l'eau (Licht, 1969). Le moment de la migration vers les lieux de reproduction et la période de ponte varient d'un endroit et d'une année à l'autre, en fonction de la température de l'air et de l'eau; il semble qu'une température de l'eau d'au moins 6 à 7 °C soit nécessaire pour la ponte, mais la température est souvent inférieure durant le développement embryonnaire (Licht, 1974; Brown, 1975). Dans le sud de la Colombie-Britannique, des accouplements ont été observés de janvier à avril, mais la période de reproduction est généralement terminée à la fin de mars (Licht, 1969; Calef, 1973b; Beasley, données inédites, 2014). Les mâles peuvent s'accoupler de multiples fois au cours d'une saison de reproduction, mais leur succès reproducteur semble très variable (Calef, 1973a). Les femelles adultes se reproduisent chaque année (Licht, 1974; Hayes, comm. pers., 2012). Chez les populations étudiées en Colombie-Britannique, les individus des deux sexes atteignent la maturité sexuelle à 3 ans ou plus (Licht, 1974). Chez les populations de l'Oregon, des mâles ont atteint la maturité sexuelle en 1 an (Hayes et Hayes, 2003); la probabilité de l'atteinte de la maturité sexuelle chez les femelles s'élevait à 74 % à 2 ans et à 26 % à 3 ans (Hayes, comm. pers., 2012).
Comme c'est le cas de la plupart des anoures qui se reproduisent dans l'eau, la fécondation est externe. Les femelles déposent leurs œufs en une volumineuse grappe gélatineuse (de 20 à 30 cm de diamètre), qu'elles fixent souvent à la végétation submergée (Leonard et al., 1993; figure 1b). La taille moyenne des grappes était de 680 œufs (plage de 243 à 935 œufs; Licht, 1974) dans des marais près de Vancouver, et de 531 + 19 œufs (moyenne + écart-type; Calef, 1973b), dans un autre site des basses terres continentales (lac Marion).
La durée de la période d'incubation et du stade larvaire dépend de la température et varie largement selon les conditions ambiantes. L'éclosion peut se produire dès le neuvième jour suivant la ponte (à une température constante de 18,3 °C; Storm, 1960), mais prend généralement beaucoup plus de temps. Dans le sud de la Colombie-Britannique, l'éclosion a lieu aussi tôt que la mi-mars ou pendant la première moitié de mai (Calef, 1973b). Le stade larvaire dure de 11 à 14 semaines environ (Calef, 1973b). La plupart des têtards se transforment entre le début de juillet et le début d'août, mais le moment exact de la métamorphose varie d'un endroit et d'une année à l'autre (Licht, 1969, Calef, 1973b). L'hivernation au stade de têtard a été observée en Oregon et en Colombie-Britannique (Corkran, comm. pers., 2007; Beasley, données inédites, 2014).
La grenouille à pattes rouges du Nord a une courbe de survie de type III, caractérisée par un taux de mortalité élevé des juvéniles. Le taux de survie annuel des individus après ce stade critique augmente considérablement. Chez cette espèce, le taux de mortalité est le plus élevé au stade de têtard. Le taux de mortalité des embryons et des individus métamorphosés est par contre relativement faible (Calef, 1973b; Licht, 1974). Dans un étang asséché, Licht (1974) a noté un taux de survie de plus de 90 % chez les embryons entre la ponte et l'éclosion, mais un taux inférieur à 1 % chez les têtards entre l'éclosion et la métamorphose. Il a par ailleurs observé un taux de survie de 5,3 % chez les têtards jusqu'à la métamorphose dans un plan d'eau permanent (la partie inondée de la rivière Little Campbell). Calef (1973b) a signalé un taux de survie de 5 % chez les têtards dans un autre site du Lower Mainland (lac Marion); les petits têtards étaient particulièrement vulnérables à la prédation et la plus grande mortalité s'est produite au cours des 3 à 4 premières semaines suivant l'éclosion. Le taux de survie estimé des recrues de la population de la rivière Little Campbell, au cours de leur première année en tant qu'individus métamorphosés, était de 52 %. Le taux de survie annuel des adultes (sexes confondus) était également élevé, soit de 69 % (Licht, 1974). En Oregon, Hayes (comm. pers., 2012) a rapporté des taux de survie des larves aussi faibles (de 3,2 à 5,5 %) et des taux de survie des adultes aussi élevés (femelles adultes, de 79 à 89 %; mâles adultes, de 67 à 80 %). D'après la plage des taux de survie, la durée d'une génération est estimée à 4 à 6 ans.
Si les infections fongiques et la dessiccation dues aux variations du niveau d'eau contribuent à la mortalité des embryons, la prédation est considérée comme la principale cause de mortalité des têtards (Calef, 1973b; Licht, 1974). Des expériences en enclos sur le terrain, où le nombre de prédateurs, en l'occurrence des tritons à peau rugueuse (Taricha granulosa), est contrôlé, confirment l'importance de la prédation comme facteur de mortalité des têtards (Calef, 1973b).
Les mâles adultes sont considérablement plus nombreux que les femelles aux sites de reproduction, mais en dehors de la saison de reproduction, le rapport des sexes semble équilibré (Calef, 1973a). La longévité des adultes dans la nature est estimée à 8 à 12 ans (Hayes et al., 2008); une longévité de 15 ans a été signalée en captivité (McTaggart Cowan, 1941). Les populations de nombreuses espèces d'anoures se reproduisant en milieu aquatique fluctuent beaucoup d'une année à l'autre (Pechmann et Wilbur, 1994). Dans une partie de la vallée de la Willamette, en Oregon, la disparition locale de la grenouille à pattes rouges du Nord et la colonisation d'étangs par l'espèce étaient fréquentes (Adams et al., 2011). Les données sur les fluctuations des populations de cette espèce au Canada se limitent à très peu de milieux humides et à une période de moins de 10 ans; elles ne révèlent pas de vastes fluctuations (Beasley, données inédites, 2014). Waye (1999) et Hayes et al. (2008) ont indiqué que les populations de grenouilles à pattes rouges survivent probablement à 1 ou 2 mauvaises années de recrutement grâce à la survie d'adultes sur plusieurs années.
La grenouille à pattes rouges du Nord est adaptée à la reproduction dans des conditions froides (Licht, 1971). Les adultes sont actifs dès le début du printemps, lorsque les températures de l'air et de l'eau sont basses. Les mâles peuvent lancer leur appel dans des eaux aussi froides que 4 ou 5 °C (Licht, 1971; Calef, 1973a; Brown, 1975). Les œufs peuvent supporter une exposition à de basses températures de cet ordre, bien que la ponte ait typiquement lieu dans des eaux légèrement plus chaudes. La fenêtre de tolérance thermique des jeunes embryons (jusqu'au stade de développement 11 de Gosner) est de 4 à 21o°C (Licht, 1971). Les températures létales, tant maximale que minimale, sont toutes deux les plus faibles chez le genre Rana en Amérique du Nord, et la tolérance de l'espèce ressemble le plus à celle de la grenouille des bois (Lithobates sylvaticus) de l'Alaska, adaptée au froid. La tolérance thermique des embryons augmente à mesure qu'ils se développent. Dans la nature, les œufs sont protégés par une masse gélatineuse et sont habituellement submergés. Ces 2 facteurs atténuent les fluctuations thermiques (Licht, 1971), mais les œufs d'une grappe les plus proches de la surface de l'eau meurent dans des conditions de gel (Beasley, données inédites, 2014).
La grenouille à pattes rouges du Nord n'est pas réputée être tolérante au gel, comme l'est la grenouille des bois et quelques autres anoures du Nord. Elle hiverne plutôt dans le fond des étangs ou sur le tapis forestier, vraisemblablement dans des parcelles de microhabitat protégées contre le gel.
Les grenouilles métamorphosées demeurent sur les rives du site de reproduction pendant des jours voire des semaines après la métamorphose (Licht, 1969). En Oregon, des individus métamorphosés ont quitté un site de reproduction de 1 à 86 jours après la métamorphose (Chelgren et al., 2008). Des juvéniles ont été capturés jusqu'à 500 m de leur lieu de naissance durant la période de dispersion en automne (Beasley, données inédites, 2014). Les déplacements des juvéniles plus âgés sont inconnus. Les résultats d'études par marquage-recapture et télémétrie indiquent que la distance parcourue par les adultes entre les sites de reproduction et les sites d'alimentation d'été varie énormément. En Oregon, 4 adultes ont été trouvés, entre mai et juillet 2002, à des distances en ligne droite de 3,5 à 4,8 km de leur point de capture de janvier et de février de la même année (Hayes et al., 2007). Dans l'État de Washington, 5 femelles adultes munies d'un radio-émetteur ont été suivies sur des distances relativement longues (jusqu'à 80 m par jour) au cours de la migration printanière. Elles se sont installées à une distance en ligne droite d'environ 312 m des sites de reproduction (Serra Shean, 2002). Lorsque les adultes arrivent en milieux terrestres, ils se déplacent généralement sur de courtes distances, bien que des déplacements plus longs aient été observés à l'occasion pendant cette période (Chan-McLeod, 2003; Chan-McLeod et Wheeldon, 2004; Chan-McLeod et Moy, 2007).
Les mâles ont manifesté une fidélité à certains sites de reproduction d'une année à l'autre; environ 20 % des mâles marqués une année donnée ont été recapturés l'année suivante au même endroit (Calef, 1973a). Qui plus est, environ 58 % des mâles recapturés sont retournés au même lit d'algues, alors que nombre d'autres occupaient des lits d'algues adjacents situés à moins de 100 m du lieu de capture initial. Comme les femelles passent moins de temps en milieux humides, leur fidélité aux sites a été peu documentée (Calef, 1973a).
Parmi les prédateurs des têtards de grenouille à pattes rouges du Nord figurent des poissons carnivores, comme la truite arc-en-ciel (Onchorynchus mykiss) introduite, des salamandres, dont le triton à peau rugueuse et la salamandre foncée, et divers invertébrés, comme les larves de libellules (Odonates) et le léthocère (Lethocerus americanus) (Calef, 1973b). Les sangsues sont des prédateurs des têtards et des œufs d'anoures (Licht, 1974). Le ouaouaron, espèce introduite, est un prédateur des larves et des adultes (Janicowski et Orchard, 2013). Plusieurs autres vertébrés et invertébrés se nourrissant de grenouilles métamorphosées et de têtards sont souvent présents dans les milieux aquatiques occupés par l'espèce, notamment le raton-laveur (Procyon lotor), le Grand Héron (Ardea herodias), le Martin-pêcheur d'Amérique (Megaceryle alcyon) et la couleuvre rayée (Thamnophis sirtalis) (Licht, 1974).
La grenouille à pattes rouges du Nord est l'hôte de divers parasites et autres organismes pathogènes. Les organismes pathogènes les plus importants décelés chez l'espèce sont probablement le Batrachochytrium dendrobatidis (Bd), champignon chytride isolé en Californie (Nieto et al., 2007), en Oregon, dans l'État de Washington (Pearl et al., 2007) et en Colombie-Britannique (Govindarajulu et al., 2013; Richardson et al., 2014), et les iridovirus isolés chez des têtards en Californie (Mao et al., 1999). L'iridovirus isolé chez des grenouilles à pattes rouges du Nord était identique à celui trouvé chez une espèce de poisson sympatrique, ce qui porte à croire que les poissons (indigènes et introduits) peuvent agir comme réservoirs du virus (Mao et al., 1999).
Le régime alimentaire des adultes et des juvéniles métamorphosés de la grenouille à pattes rouges du Nord se compose d'une grande variété de petits invertébrés, dont les araignées (Aranéides), les coléoptères (p. ex. Carabidés, Staphylinidés, Chrysomélidés, Curculionidés et Limnebiidés), les cicadelles (Cicadellidés), les punaises demoiselles (Nabidés) et les limaces (Arionidés) (Licht, 1986; Beasley, données inédites, 2014). Les têtards se nourrissent principalement d'algues vertes filamenteuses. Des expériences en enclos ont révélé que les têtards qui s'alimentaient ainsi modifiaient la composition et l'abondance du périphyton (Dickman, 1968). Selon Dickman (1968), l'alimentation des têtards pourrait amorcer une succession saisonnière du périphyton dans les plans d'eau, laquelle peut à son tour causer de nombreux effets dans les réseaux trophiques.
Des relevés visant à estimer les populations reproductrices de grenouille à pattes rouges du Nord ont été réalisés dans des sites de l'ensemble de l'île de Vancouver depuis 2006 et dans plusieurs milieux humides de la vallée du bas Fraser abritant d'autres espèces en péril (grenouille maculée de l'Oregon, tortue peinte de l'Ouest) depuis 2010 (tableau 2). Les dénombrements des grappes d'œufs constituent un moyen efficace d'obtenir un indice de la taille des populations parce que chaque femelle pond une seule grappe d'œufs par année et que les grappes d'œufs sont beaucoup plus faciles à compter que tout autre stade du cycle vital.
Dans le Lower Mainland, les grappes d'œufs de grenouille à pattes rouges du Nord ont été dénombrées en même temps que les grappes d'œufs de grenouille maculée de l'Oregon, espèce en voie de disparition. L'échantillonnage a été relativement minutieux puisque les sites ont été visités à répétition pendant toute la saison de reproduction (Pearson, 2010, 2011, 2012). Des dénombrements sur plusieurs jours ont aussi été effectués afin de couvrir les vastes superficies d'habitat de ponte dispersées à l'échelle du lac Swan (4 ha) et du lac Wood (2 ha), près d'Ucluelet, dans l'île de Vancouver. Les dénombrements réalisés dans d'autres sites de l'île de Vancouver, du Lower Mainland, de l'île Texada et de la Sunshine Coast ont pris une seule journée ou une partie de journée. Seuls les endroits accessibles ont été recensés à nombre de ces sites (McConkey, données inédites, 2008; Beasley, données inédites, 2014; Mitchell, comm. pers., 2014).
Deux études par marquage-recapture dans des sites de reproduction menées en Colombie-Britannique (Licht, 1969, 1971, 1974; Calef, 1973a, b) ont permis de recueillir de l'information sur la taille des populations au lac Marion (aussi appelé lac Jacobs), près de Maple Ridge (Calef, 1973a, b), et dans les marais de la rivière Little Campbell, à Surrey (Licht, 1969, 1974).
Les premières études sur la grenouille à pattes rouges du Nord menées dans 2 sites continentaux ont relevé plus de 600 grappes d'œufs dans le lac Marion (Calef, 1973 a, b) et moins de 40 grappes d'œufs dans des étangs près de la rivière Little Campbell (Licht, 1969, 1971, 1974). Les inventaires récents réalisés dans 197 milieux humides répartis à l'échelle de l'aire de répartition de l'espèce, mais principalement dans l'île de Vancouver, ont révélé que la plupart des sites contenaient relativement peu de grappes d'œufs (figure 4). Les observateurs ont trouvé 10 grappes ou moins dans 35 % des milieux humides et plus de 100 grappes dans seulement 20 % des sites recensés. Les dénombrements les plus élevés ont été obtenus dans des sites de la côte ouest de l'île de Vancouver (lac Pixie, près de Port Renfrew : 1 400; lac Swan et ruisseau « Lost Shoe 4 », près d'Ucluelet : 1 374 et 618, respectivement; passage Julia, dans la baie Barkley : 445) (B.C. Ministry of Environment, 2012; Beasley, 2011, comm. pers., 2013). Des dénombrements très élevés ont aussi été obtenus dans le Lower Mainland, en particulier dans les sites de reproduction de la grenouille maculée de l'Oregon (marécage Maria, chemin Chaplin : 678; marécage Maria : 404; marécage Mountain : 236; vallée de la Morris : 203) et dans d'autres sites de la vallée du Fraser (pied du mont Vedder, sur le chemin Town à Chilliwack : 392; ferme no 2 d'AAC : 285) (Pearson, 2010, 2011, 2012).
Il est possible que les sites affichant de faibles dénombrements abritent des portions d'une population plus grande, ou métapopulation, réparties dans de multiples étangs de reproduction, comme cela peut souvent être le cas chez les amphibiens (Marsh et Trenham, 2001). Ces sites peuvent aussi représenter de petites populations isolées mais persistantes.
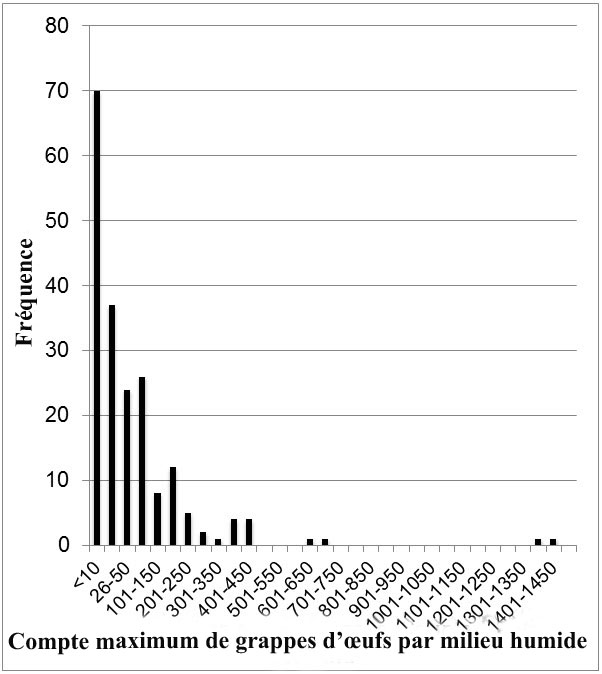
Sources des données : Beasley, 2011; Beasley, données inédites, 2014; B.C. Ministry of Environment, 2012; Malt, 2011; Mitchell et al., 2012; Mitchell, comm. pers., 2014; Pearson, 2010, 2011, 2012).
Long description for Figure 4
Graphique montrant la distribution de la fréquence du nombre de grappes d'œufs de grenouille à pattes rouges du Nord observées de 2006 à 2013 dans les milieux humides occupés (N = 197) en Colombie-Britannique. Trente-cinq pour cent des milieux humides renfermaient 10 grappes d'œufs ou moins, et seuls 20 % en renfermaient 100 ou plus (voir le corps du rapport).
La population canadienne totale de grenouille à pattes rouges du Nord peut être grossièrement estimée à bien au-delà de 100 000 adultes selon l'abondance connue de 12 603 grappes d'œufs dénombrées dans 197 milieux humides au cours de la dernière décennie (tableau 3), ce qui correspond à moins de la moitié des occurrences connues. Si chaque femelle reproductrice a besoin d'un mâle, alors le nombre d'adultes reproducteurs dans les sites recensés est d'au moins 32 446. Si seule une partie de la population de mâles adultes se reproduit chaque année, comme l'ont estimé Calef (1973a) et Licht (1969), la population de grenouilles à pattes rouges du Nord dans les sites recensés pourrait donc être de 3 à 6 fois plus grande que le nombre de grappes d'œufs.
Description longue pour le tableau 3
| Région | Période | Nombre de milieux humides occupés visés par un relevé | Nombre moyen de grappes d'œufs par année pour la régionTable Footnoteb | Plage du nombre de grappes d'œufs par année pour la région | Nombre estimatif d'adultes reproducteurs aux sites recensésTable Footnotec |
|---|---|---|---|---|---|
| Île de Vancouver | District forestier de Campbell River 2005–2012 |
14 | 537 | 388-719 | 1 438 |
| Île de Vancouver | Districts forestiers du North Island et du North-Central Island 2006–2012 |
29 | 360 | 233-493 | 986 |
| Île de Vancouver | District forestier du South Island 2006–2012 |
61 | 4 105 | 3 257–5 114 | 10 228 |
| Île de Vancouver | Ucluelet – Tofino 2008–2013 |
38 | 4 837 | 3 216–6 242 | 12 484 |
| Zone continentale | Région du Lower et vallée du Fraser 2010–2012 |
36 | 2 382 | 1 621–3 211 | 6 422 |
| Zone continentale | Corridor Sea-to-Sky 2007–2010 |
8 | 48 | 32–66 | 132 |
| Zone continentale | Sunshine Coast 2010–2012 |
6 | 285 | 242–328 | 656 |
| Détroit de Georgia | Île Texada 2010–2012 |
5 | 49 | 48–50 | 100 |
| TOTAL – C.-B. | - | 197 | 12 603 | - | 32 446 |
Longue description pour le tableau 4
| Numéro | Description | Portée Table Footnoted | Gravité Table Footnotee | Immédiateté Table Footnotef | Impact Table Footnoteg | Populations ou régions touchées |
|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | Petite | Extrême | Élevée | Faible | Lower Mainland et sud-est de l'île de Vancouver |
| 1.1 | Zones résidentielles et urbaines | Petite | Extrême | Élevée | Faible | Lower Mainland et sud-est de l'île de Vancouver |
| 4 | Corridors de transport et de service (en anglais seulement) | Grande | Modérée | Élevée | Moyen | À l'échelle de l'aire de répartition, sauf les parties éloignées de l'ouest de l'île de Vancouver et de la côte centrale |
| 4.1 | Routes et voies ferrées | Grande | Modérée | Élevée | Moyen | À l'échelle de l'aire de répartition, sauf les parties éloignées de l'ouest de l'île de Vancouver et de la côte centrale |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | Petite à restreinte | Modérée | Élevée | Faible | À l'échelle de l'aire de répartition, sauf les zones urbaines et agricoles |
| 5.3 | Exploitation forestière et récolte du bois | Petite à restreinte | Modérée | Élevée | Faible | À l'échelle de l'aire de répartition, sauf les zones urbaines et agricoles |
| 7 | Modification du système naturel (en anglais seulement) | Petite | Modérée | Élevée | Faible | Sud-est de l'île de Vancouver, Campbell River, vallée du Fraser |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | Petite | Modérée | Élevée | Faible | Sud-est de l'île de Vancouver, Campbell River, vallée du Fraser |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | Grande à généralisée | Modérée à élevée | Élevée | Moyen à élevé | À l'échelle de l'aire de répartition pour Bd; Lower Mainland et sud-est de l'île de Vancouver pour le ouaouaron |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | Grande à généralisée | Modérée à élevée | Élevée | Moyen à élevé | À l'échelle de l'aire de répartition pour Bd; Lower Mainland et sud-est de l'île de Vancouver pour le ouaouaron |
| 8.2 | Espèces indigènes problématiques | Petite à restreinte | Modérée à élevée | Élevée | Faible à moyen | À l'échelle de l'aire de répartition pour Bd; Lower Mainland et sud-est de l'île de Vancouver pour le ouaouaron |
| 9 | Pollution (en anglais seulement) | Petite | Faible à modérée | Élevée | Faible | Lower Mainland et vallée du bas Fraser |
| 9.3 | Effluents agricoles et sylvicoles | Petite | Faible à modérée | Élevée | Faible | Lower Mainland et vallée du bas Fraser |
| 9.5 | Polluants atmosphériques | Inconnue | Faible à modérée | Élevée | Faible | Lower Mainland et vallée du bas Fraser |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | Généralisée | Inconnue | Élevée | Inconnu | À l'échelle de l'aire de répartition, en particulier la zone côtière à douglas du sud-est de l'île de Vancouver |
| 11.1 | Déplacement et altération de l'habitat | Négligeable | Inconnue | Élevée | Négligeable | À l'échelle de l'aire de répartition, en particulier la zone côtière à douglas du sud-est de l'île de Vancouver |
| 11.2 | Sécheresses | Généralisée | Inconnue | Élevée | Inconnu | À l'échelle de l'aire de répartition, en particulier la zone côtière à douglas du sud-est de l'île de Vancouver |
| 11.3 | Températures extrêmes | Généralisée | Inconnue | Élevée | Inconnu | À l'échelle de l'aire de répartition, en particulier la zone côtière à douglas du sud-est de l'île de Vancouver |
| 11.4 | Tempêtes et inondations | Petite | Inconnue | Élevée | Inconnu | À l'échelle de l'aire de répartition, en particulier la zone côtière à douglas du sud-est de l'île de Vancouver |
Il s'est produit 2 déclins de population documentés, et des observations anecdotiques laissent entrevoir la survenue d'un troisième déclin, qui s'ajouterait à la disparition de la population du parc Stanley (Stanley Park Ecology Society, 2010). En premier lieu, Rithaler (2002a, 2003a) a conservé les mentions de reproduction de 1962 à 2002 et décrit qualitativement une diminution de l'abondance de la grenouille à pattes rouges du Nord dans la région circonscrite par la Corporation de Delta (Delta, Ladner, Tsawwassen). Aucune grappe d'œufs n'a été trouvée dans 4 sites lors d'un relevé récent (Malt, 2013) et, après plusieurs jours de recherche, un seul individu a été observé dans la partie la moins développée de la tourbière Burns (Robertson Environmental, 2013). En deuxième lieu, Malt (2012) a utilisé des données démographiques recueillies dans le cadre des opérations de récupération avant la construction et les a comparées à des données recueillies par la suite en utilisant la méthode marquage-recapture pour évaluer les effets de la modification du tracé de la route 99 jusqu'à Whistler en 2008. Il a estimé que, dans un des milieux humides fragmentés par la construction de l'autoroute, la population locale a chuté, passant de 357 individus en 2007 à 30 à 96 individus en 2010, soit un déclin de 73 à 92 %. En troisième lieu, il ne semble pas y avoir de grenouilles à pattes rouges du Nord dans les milieux humides de Victoria envahis par le ouaouaron, ou y en avoir très peu, alors qu'elle persiste en grand nombre dans les sites suburbains n'abritant pas cet envahisseur (Ovaska, comm. pers., 2014; Fraser, comm. pers., 2014).
Au cours des deux à six dernières années, quelques projets de surveillance à long terme ont été lancés dans différentes parties de l'aire de répartition de la grenouille à pattes rouges du Nord : la côte ouest de l'île de Vancouver (Beasley, 2011), la Sunshine Coast, l'île Texada et le Lower Mainland, dans des sites importants pour la tortue peinte de l'Ouest (Mitchell et al., 2012), et les sites de la vallée du Fraser importants pour la grenouille maculée de l'Oregon (Pearson, 2012). Les comptes de grappes d'œufs fluctuent légèrement d'une année à l'autre, mais il est trop tôt pour déceler des tendances.
Des herpétologues ont signalé des baisses d'effectif dans la partie sud de l'aire de répartition de l'espèce en Californie (Jennings et Hayes, 1994) et dans la vallée de la Willamette, en Oregon (Blaustein et al., 1994), depuis le milieu des années 1970. Des populations reproductrices persistent au fond de la vallée de la Willamette, et ce, malgré la modification étendue de l'habitat (Pearl, 2005). Un programme quinquennal de suivi (de 2004 à 2008) a révélé des probabilités d'occupation stables (intervalle de 0,38 à 0,43) dans le complexe de refuges de la vallée de la Willamette où, contrairement au reste de la région, la perte de milieux humides a été minime (Adams et al., 2011). Toutefois, il a aussi révélé un niveau considérable de disparition et de colonisation locales de milieux humides dans le complexe de refuges : à 17 reprises, la grenouille à pattes rouges du Nord n'a pas été détectée à un site où elle était présente l'année précédente et, à 13 reprises, elle l'a été à un site où elle n'avait pas été observée l'année précédente (Adams et al., 2011).
La grenouille à pattes rouges du Nord semble demeurer relativement commune dans au moins quelques régions de l'État de Washington (Adams et al., 1998, 1999), y compris dans des paysages modifiés par l'homme (Richter et Azous, 1995; Ostergaard et al., 2008).
La possibilité d'une immigration de source externe pour reconstituer des populations canadiennes grâce à la dispersion d'individus provenant de populations américaines voisines est limitée. Il existe plusieurs mentions de la présence de telles populations près de la frontière internationale, dans le Lower Mainland, mais la plupart datent d'avant 1960, et on ignore si ces populations existent encore. La dispersion depuis les États-Unis pourrait se produire à travers les basses terres situées à l'ouest de la vallée du Columbia, près du lac Cultus, mais cette région est grandement fragmentée et modifiée par les activités agricoles, les lotissements résidentiels et les routes. Il reste quelques secteurs boisés à proximité immédiate de la frontière. Immédiatement à l'est de la vallée du Columbia, les pics élevés de la chaîne des Cascades posent un obstacle à la dispersion.
Les limites septentrionales actuelles de l'aire de répartition de la grenouille à pattes rouges du Nord découlent probablement davantage de l'histoire glaciaire de la région que des limites de tolérance physiologique ou écologique de l'espèce, comme en témoignent deux populations nordiques isolées, introduites à Haida Gwaii et dans le sud-est de l'Alaska. Il se peut que l'espèce soit encore en voie d'étendre son aire de répartition vers le nord, le long du littoral du Pacifique, mais l'altitude (et peut-être aussi les interactions avec la grenouille maculée de Columbia ou d'autres espèces) peut constituer un obstacle à son expansion vers l'intérieur.
Une évaluation des menaces a été réalisée au moyen du calculateur des menaces de l'IUCN (Master et al., 2009). Chaque menace a été classée en fonction de la proportion de la population ou de l'aire de répartition canadiennes menacées (portée) au cours des dix prochaines années, l'ampleur de la menace pour ce qui est du déclin prévu de la population au cours des trois prochaines générations (gravité) et le caractère immédiat de la menace (immédiateté). L'évaluation de la portée reposait sur les renseignements sur l'utilisation des terres dans l'île de Vancouver tirés des cartes thématiques de base disponibles auprès de Hectares BC (2014) ainsi que sur les changements prévus d'utilisation des terres résumés dans les stratégies de croissance régionale de plusieurs districts régionaux situés dans l'aire de répartition de l'espèce. La combinaison des rangs pour ces trois mesures a été utilisée pour calculer l'impact global de la menace. Selon ce calcul, l'impact global des menaces sur l'espèce est « élevé », selon six menaces à impact allant de « faible » à « moyen », résumées au tableau 3 et décrites ci-dessous.
Les menaces pesant sur la grenouille à pattes rouges du Nord ont été évaluées pour l'ensemble de l'aire de répartition canadienne de l'espèce, mais il existe de nettes différences dans la portée et le niveau d'impact des menaces dans différentes parties de l'aire de répartition, notamment entre le Lower Mainland et le sud-est de l'île de Vancouver, et le reste de l'île de Vancouver et le secteur continental au nord de la Sunshine Coast (tableau 1).
Il a été établi que les conditions d'habitat dans les sites de reproduction et aux alentours ont un effet sur l'occurrence et la persistance des populations de grenouilles à pattes rouges du Nord (Richter et Azous, 1995; Adams, 1999; Ostergaard et al., 2008; Adams et al., 2011; Malt, 2013). L'aire de répartition canadienne de l'espèce chevauche les régions les plus peuplées connaissant la plus forte croissance démographique de la Colombie-Britannique, soit la vallée du bas Fraser et le sud et l'est de l'île de Vancouver (figure 3). Des grenouilles y sont encore présentes dans de nombreuses petites parcelles d'habitat restantes entourées de terres agricoles et de zones urbaines. La croissance continue de la population humaine mènera à une expansion de l'usage à des fins résidentielles et à l'intensification des pratiques agricoles qui, selon les prévisions, accélèreront la perte et la dégradation de l'habitat au cours de la prochaine décennie (voir Tendances en matière d'habitat). Dans les secteurs moins peuplés de l'aire de répartition de l'espèce (nord et ouest de l'île de Vancouver, Sunshine Coast et vers le nord le long de la côte), les activités forestières sont intensives et continuent de modifier et de fragmenter l'habitat. La succession forestière signifie qu'il se produit une progression des effets après le déboisement.
Bien que la Water Act et le Riparian Area Regulations provinciaux protègent les milieux humides et certaines zones riveraines (p. ex. des zones tampons de 5 m autour des cours d'eau offrant un habitat aux poissons), la majeure partie des milieux forestiers seront modifiés, et les parcelles restantes seront davantage fragmentées. La fragmentation perturbe les processus des métapopulations en limitant les déplacements entre les populations locales à l'échelle du paysage. Ailleurs, il a été établi que la fragmentation de l'habitat contribue à des disparitions et à des déclins localisés des amphibiens forestiers se reproduisant dans des étangs qui dépendent de la dispersion entre sous-populations à l'échelle du paysage (p. ex. Ambystoma maculatum; Gibbs, 1998). Green (2003), après avoir comparé les tendances des populations et les paramètres démographiques d'un grand nombre d'espèces et de populations d'amphibiens, a conclu que la fragmentation de l'habitat, en nuisant à la recolonisation, risque d'avoir des conséquences immédiates et catastrophiques sur les espèces à dispersion obligatoire manifestant des fluctuations démographiques importantes et une fréquence élevée d'extinction localisée, comme les amphibiens se reproduisant dans les étangs (traduction libre de la p. 341 de Green, 2003). Ces considérations s'appliquent à la grenouille à pattes rouges du Nord en général, bien que les détails des fluctuations et de la dynamique de ses populations dans l'espace et dans le temps soient inconnus.
Dans les zones urbaines et agricoles de la région du Lower Mainland et du sud et de l'est de l'île de Vancouver, l'essor constant de la population et l'expansion de l'aire de répartition du ouaouaron, espèce envahissante, viennent ajouter à la problématique. Le ouaouaron est un prédateur qui fait compétition aux espèces indigènes de grenouilles et propage des maladies. On le rencontre habituellement dans des milieux dégradés ainsi qu'en présence de poissons prédateurs introduits. Plusieurs chercheurs ont tenté de séparer l'influence de la contribution de chacun de ces facteurs au déclin des populations de grenouille à pattes rouges du Nord (Adams, 1999; Govindarajulu, 2004; Pearl et al., 2005; Adams et al., 2011). Il semble exister plusieurs effets indirects et interactions synergétiques.
L'aménagement de routes et de barrages, la gestion de l'eau, la présence de d'espèces végétales envahissantes, l'ensemencement de poissons de pêche sportive dans l'habitat de reproduction et la pollution attribuable aux effluents agricoles et forestiers constituent d'autres menaces anthropiques dans l'ensemble de l'aire de répartition de l'espèce. La menace posée par les maladies, en particulier une flambée de chytridiomycose, est une préoccupation majeure, la chytridiomycose ayant causé des déclins et des disparitions de populations d'anoures partout dans le monde. Les changements climatiques planétaires pourraient exacerber nombre de ces effets. Les interactions synergétiques entre divers facteurs, d'origine humaine et naturelle, sont probablement communes et peuvent toucher les amphibiens de manière imprévisible.
Les maladies épidémiques sont considérées comme une menace potentielle pour toutes les espèces d'amphibiens. La chytridiomycose, due au champignon chytride Batrachochytrium dendrobatidis (Bd), a été mis en cause dans les déclins et les disparitions rapides de populations locales d'amphibiens partout dans le monde (Lips et al., 2008), y compris l'ouest des États-Unis. Nieto et al. (2007) ont détecté, dans le nord de la Californie, le Bd chez des têtards de grenouille à pattes rouges du Nord. Pearl et al. (2007) ont détecté le Bd chez 28 % des amphibiens échantillonnés dans 43 % des sites répertoriés en Oregon et dans l'État de Washington. Sur 7 espèces ayant fait l'objet d'analyses de détection du Bd dans le cadre de l'étude, la grenouille à pattes rouges du Nord et le ouaouaron figuraient parmi les 4 espèces qui ont présenté des résultats positifs. Le Bd se trouve aussi chez des grenouilles à pattes rouges du Nord échantillonnées à l'échelle de la Colombie-Britannique, mais aucune éclosion de la maladie n'a été signalée (Govindarajulu et al., 2013; Richardson et al., 2014). Il est possible que les grenouilles à pattes rouges du Nord soient résistantes à une exposition chronique au Bd et que la probabilité d'une éclosion de la maladie soit faible, à moins que leur résistance diminue ou que de nouvelles souches du champignon émergent ou deviennent prévalentes. Les conditions futures résultant de changements climatiques pourraient mener à une diminution de la résistance et à l'émergence de nouvelles souches. Il a été établi que la variabilité de la température (variation diurne extrême) perturbe la fonction immunitaire des amphibiens (Raffel et al., 2006) et est liée à des éclosions de maladies chez d'autres espèces (Rohr et Raffel, 2010). Une expérience en mésocosme a révélé qu'une plus grande variabilité de la température avait un effet négatif sur la condition corporelle de têtards de grenouille à pattes rouges du Nord exposés à Bd (Hamilton et al., 2012).
Outre le Bd, les iridovirus causent des maladies qui sont létales pour les amphibiens (Daszak et al., 2003; Blaustein et al., 2012). Les iridovirus, associés à de nombreux phénomènes de mortalité massive d'amphibiens partout dans le monde, étaient prévalents lors de la plupart des mortalités massives d'amphibiens étudiées à la fin des années 1990 partout aux États-Unis (Green et al., 2002) et dans certaines parties du Canada (Saskatchewan et Ontario) (Bollinger et al., 1999; Greer et al., 2005). Des échantillons d'iridovirus recueillis à des fins de diagnostic n'ont pas été prélevés en Colombie-Britannique; la prévalence est donc inconnue (Govindarajulu, comm. pers., 2015). Un iridovirus a été isolé chez deux têtards de grenouille à pattes rouges du Nord prélevés dans le parc national Redwood, en Californie, l'un en 1994 et l'autre en 1996 (Mao et al., 1999) mais, autrement, sa présence chez l'espèce n'a pas été signalée. Étant donné que les changements environnementaux résultant de la perte et de la dégradation de l'habitat sont reconnus pour modifier la prévalence des iridovirus, les taux de transmission et la susceptibilité de l'hôte (Gray et al., 2009; Blaustein et al., 2012), la menace potentielle qu'ils posent ne peut pas être ignorée.
Le ouaouaron est présent dans la plus grande partie de l'aire de répartition de la grenouille à pattes rouges du Nord dans le Lower Mainland, tout comme sur la Sunshine Coast, dans le sud-est de l'île de Vancouver (de Victoria à Campbell River) et dans certaines des îles Gulf (Govindarajulu, 2003; Mitchell et al., 2012). Il a été introduit à Maple Ridge et Aldergrove dans les années 1940, et son aire de répartition actuelle dans la vallée du bas Fraser s'étend du parc Stanley, à Vancouver (Stanley Park Ecology Society, 2010), au lac Morris, près de Harrison Mills vers l'est (Murray et al., sous presse). La modélisation de l'occupation prévoit que l'aire de répartition de cette espèce envahissante chevauchera la plus grande partie de l'aire de répartition de la grenouille à pattes rouges du Nord dans la vallée du bas Fraser dans 70 ans (Murray, comm. pers., 2014). Le ouaouaron étend également rapidement son aire de répartition dans l'île de Vancouver et les îles Gulf depuis les années 1990 (Govindarajulu, 2004), et il se propage sur la Sunshine Coast (Mitchell et al., 2012). Dans l'île de Vancouver,on le rencontre maintenant de Sooke à Campbell River vers le nord et dans l'est, et jusqu'à Port Alberni dans l'ouest, ce qui représente environ 20 % de l'aire de répartition de la grenouille à pattes rouges du Nord dans l'île. Le ouaouaron a été observé dans les îles Saltspring, Pender, Lasqueti et Texada, et dans des secteurs de la péninsule Sechelt (Mitchell et al., 2012; Govindarajulu, comm. pers., 2014).
Il est supposé que l'introduction et la propagation du ouaouaron ont contribué à la baisse des effectifs de la grenouille à pattes rouges du Nord dans l'ensemble de son aire de répartition; toutefois, une incertitude perdure quant à l'importance de son effet (Pearl et al., 2005; Adams et al., 2011). À certains endroits en Colombie-Britannique, comme le parc Stanley et Delta, le ouaouaron semble avoir complètement déplacé la grenouille à pattes rouges du Nord (Rithaler, 2002b, 2003a; Stanley Park Ecology Society, 2010), probablement en raison de nombreux facteurs contributifs. Par exemple, dans la Corporation de Delta, dans le Lower Mainland, la modification de l'habitat, en particulier l'enlèvement de la végétation riveraine et l'approfondissement des chenaux, semble avoir favorisé la croissance des populations de ouaouarons et de grenouilles vertes (Lithobates clamitans) ainsi que la disparition de la grenouille à pattes rouges du Nord dans certains milieux humides (Rithaler, comm. pers., 2014). À d'autres endroits, la grenouille à pattes rouges du Nord semble cohabiter avec le ouaouaron, au moins à court terme, mais en nombre réduit (Govindarajulu, comm. pers., 2014). La grenouille à pattes rouges du Nord dispose de refuges de reproduction dans des étangs éphémères ou temporaires, alors que le ouaouaron et les poissons introduits sont limités aux plans d'eau permanents.
L'ensemencement de poissons indigènes et non indigènes de pêche sportive est couramment pratiqué dans l'ensemble de l'aire de répartition de la grenouille à pattes rouges du Nord en Colombie-Britannique (Wind, 2005; Freshwater Fisheries Society of BC, 2014). Des poissons non indigènes, comme des achigans (espèces du genre Micropterus), des crapets (espèces du genre Lepomis) et des perches (espèces du genre Perca) sont illégalement libérés, tandis que l'omble de fontaine (Salvelinus fontinalis) et la truite brune (Salmo trutta)étaient légalement ensemencés dans l'île de Vancouver et le Lower Mainland jusqu'à il y a environ 20 ans (Silvestri, comm. pers., 2014). Aujourd'hui, tous les ensemencements légaux dans ces régions se limitent à la truite arc-en-ciel et à la truite fardée (Oncorhynchus clarkii), dont la plupart des individus sont stérilisés au moyen de la technique du choc de pression, qui cause la triploïdie (Silvestri, comm. pers., 2014). Certains sites reçoivent des géniteurs non stérilisés. L'ensemencement illégal de poissons soulève les plus grandes préoccupations parce que les populations ne sont pas limitées. Les poissons se nourrissent de larves de grenouille à pattes rouges du Nord, ce qui réduit leurs populations, à moins que l'habitat en milieux humides soit suffisamment complexe pour offrir à ces dernières un refuge adéquat (Adams et al., 2011). Les poissons ensemencés peuvent aussi causer des maladies, notamment en introduisant des protozoaires parasites (Neite et al., 2007) ou des Saprolegnia (Kiesecker et al., 2001), organismes qui peuvent réduire le succès d'éclosion des œufs de grenouille à pattes rouges du Nord. Plusieurs études ont révélé des associations négatives entre la grenouille à pattes rouges du Nord et la présence de poissons non indigènes (Adams, 1999; Adams, 2000; Pearl et al., 2005).
Le réseau routier est tellement dense que 44 % des occurrences connues de la grenouille à pattes rouges du Nord se trouvent à moins de 500 m de routes (tableau 1). Aux altitudes inférieures à 500 m de l'ensemble de l'aire de répartition canadienne de l'espèce, 94, 77, 65 et 36 % des occurrences se trouvent respectivement à moins de 5, 1, 500 et 100 m d'une route (Hectares BC, 2014).
Les routes posent une menace pour les amphibiens de diverses manières. La construction de routes cause la perte d'habitat. Des routes sont souvent aménagées dans des secteurs de basses terres où les grenouilles sont les plus abondantes. Par exemple, dans le cas du prolongement et de l'amélioration de la route 99 dans le corridor Sea-to-Sky, près de Pinecrest, un nouveau tracé de 1,9 km a été aménagé à travers un vaste complexe de milieux humides occupés par la grenouille à pattes rouges du Nord et plusieurs autres espèces d'amphibiens (Golder Associates Ltd., 2008). Un total de 695 grenouilles à pattes rouges du Nord ont été récupérées dans la région avant la construction du nouveau tracé de la route (Golder Associates Ltd., 2006, 2007, 2008). Dans l'un des milieux humides ainsi fragmentés, il s'est produit une réduction estimée de la population allant de 73 à 92 % par rapport à ce qu'elle était en 2007, avant la construction, et à ce qu'elle était en 2010, après la construction (Malt, 2012). Deux autres grands projets d'élargissement de routes ont été exécutés récemment dans l'aire occupée par la grenouille à pattes rouges du Nord dans la région du Lower Mainland, soit l'élargissement de l'autoroute 1 à Port Mann et de la route périphérique sur la rive sud du Fraser. D'autres travaux devraient être effectués au cours des 10 prochaines années.
Le trafic pose une menace constante pour les populations qui traversent des routes pendant leurs migrations saisonnières. Il a été établi que les populations locales d'amphibiens partout dans le monde diminuent (Fahrig et al., 1995), deviennent génétiquement isolées (Reh, 1989; Vos et al., 2001) et même disparaissent (Cooke, 1995, in Puky, 2003) par suite de la mortalité sur les routes. La menace que la mortalité sur les routes exerce sur une population dépend du volume du trafic (Hels et Buchwald, 2001), de la largeur de la route ainsi que de la vitesse de déplacement et du nombre d'individus tentant de la traverser. Le volume du trafic sur les chemins de service, comme ceux empruntés pour l'exploitation forestière, est faible à la brunante et en soirée lorsque les grenouilles se déplacent. Toutefois, même des volumes relativement faibles, p. ex. 21 à 542 véhicules par heure, peuvent avoir des effets isolants sur des populations en raison de la mortalité sur les routes (Fahrig et al., 1995). La mortalité de la grenouille à pattes rouges du Nord a été documentée à plusieurs endroits : la route 4, près de Coombs (Blood et Henderson, 2000); le chemin des lacs Nanaimo, à Nanaimo (Wind, 2012); la route 4, dans la réserve de parc national Pacific Rim (Beasley, 2006); le chemin du lac Ryder, dans la vallée du Fraser (Clegg, 2011); le chemin Laburnum, à Qualicum (Materi, 2008); le chemin Lazo, à Comox (Wind, 2012); le chemin du lac Wake, près de Duncan (Wind, 2012); la nouvelle route Sea-to-Sky, à Pinecrest (Malt, 2012). Le nombre de cas de mortalité routière non rapportés est inconnu, mais il est probable qu'il est élevé compte tenu du vaste réseau routier sillonnant l'aire de répartition de l'espèce. On s'attend à ce que la circulation intense au site Pinecrest de l'autoroute Sea-to-Sky cause la disparition de la population locale de grenouille à pattes rouges du Nord d'ici 20 à 40 ans si le réseau actuel de passages inférieurs et de barrières n'est pas amélioré (Malt, 2012). Il est important de noter que les sites de la route Sea-to-Sky et du chemin Laburnum, à Qualicum, affichaient des taux de mortalité routière élevés malgré l'installation de tunnels pour amphibiens spécialement conçus pour assurer leur passage sécuritaire, jusqu'à ce que des barrières soient installées.
En plus de causer la perte d'habitat et la mortalité directe des amphibiens, les routes agissent comme obstacles aux déplacements par temps sec; en effet, les grenouilles à pattes rouges du Nord les traversent rarement à moins qu'il pleuve (Beasley, données inédites, 2014). Les sels de voirie et d'autres polluants se trouvant sur les routes se déversent dans les fossés et les milieux humides (voir Pollution. L'espèce séjourne, et parfois se reproduit, dans des étangs d'eaux pluviales en bord de route, où elle est vulnérable à l'ensemble des impacts négatifs liés aux routes.
La superficie urbanisée couvre actuellement 211 000 ha, soit 6 % de l'aire de répartition canadienne de l'espèce aux altitudes de moins de 500 m (Hectares BC, 2014). Environ 20 % des occurrences connues se trouvent à moins de 1 km de zones de développement résidentiel ou commercial existantes (tableau 1). Il est prévu que la construction domiciliaire prendra un rapide essor, en particulier dans la vallée du bas Fraser et à Squamish (voir Tendances en matière d'habitat). La conversion permanente des terres aux fins de la construction d'habitations est en cours à la périphérie de centres urbains et suburbains, dont Maple Ridge, Surrey-Sud, Delta, Mission, le mont Sumas, Chilliwack, Squamish et la Sunshine Coast. Il devient rare de voir de vastes bandes de forêts entourant des milieux humides, et les paysages hautement fragmentés sont de plus en plus courants.
Les activités accrues de développement résidentiel et commercial mènent à la perte irréversible d'habitat et contribuent à sa fragmentation. Les résultats d'un modèle d'occupation de la région du Lower Mainland confirment les baisses d'effectifs supposées de la grenouille à pattes rouges du Nord par suite de l'urbanisation. Malt (2013) a mené, au printemps 2012, des relevés systématiques visant à évaluer la présence d'individus reproducteurs dans 64 milieux humides de municipalités situées dans la région métropolitaine de Vancouver et la vallée du Fraser. Un relevé de 55 sites additionnels a été effectué dans le cadre de travaux en cours visant d'autres espèces en péril. Des grappes d'œufs de grenouille à pattes rouges du Nord ont été trouvées dans 48 % des sites (annexe 1). Les données ont été utilisées pour établir la probabilité d'occupation en fonction de l'utilisation des terres environnantes selon les données d'occupation du sol pour 2007-2008. La probabilité d'occupation diminuait à mesure que le pourcentage de l'espace urbain augmentait dans un rayon de 2 km autour de chaque site. La probabilité d'occupation était nulle pour les sites à Vancouver, Richmond et Delta (Malt, 2013). Ces résultats concordent avec les tendances régionales que montrent la baisse des effectifs de certaines espèces et la perte d'habitat dans la région Nord-Ouest du Pacifique américain (Hayes et al., 2008, et références citées dans cet article). Plusieurs études dont l'assemblage spécifique incluait la grenouille à pattes rouges du Nord ont démontré une corrélation négative entre la diversité des espèces d'amphibiens dans les milieux humides et le taux d'urbanisation des environs (Ostergaard et al., 2008).
Environ 80 % de la superficie totale se trouvant à une altitude égale ou inférieure à 500 m dans l'aire de répartition de la grenouille à pattes rouges du Nord est située dans des forêts aménagées. Ces forêts incluent des secteurs qui ont été coupés dans les 20 dernières années (13 %), d'autres coupés il y a entre 20 et 140 ans (38 %), de même que des forêts mûres de plus de 140 ans (29 %) (Hectares BC, 2014). Si l'exploitation forestière se poursuit au même rythme, 5 % de l'aire de répartition de l'espèce sera déboisée au cours de la prochaine décennie. Dans les récents plans d'aménagement du littoral, un taux de coupe durable a été fixé à environ 1 % du territoire de base par année. Si 50 % de la forêt aménagée de plus de 140 ans est située dans le territoire de base et qu'elle sera coupée au cours de la prochaine décennie, alors 1,5 % (soit 0,01/an × 10 ans × 0,5 × 0,29) de la superficie occupée par la grenouille à pattes rouges du Nord sera récoltée au cours de la prochaine décennie. Ce pourcentage constitue une sous-estimation puisque les forêts de 80 ans en maturation, et non seulement celles de plus de 140 ans, sont en voie d'être coupées.
L'enlèvement de la végétation et l'aménagement de routes liés à l'exploitation forestière peuvent mener à des changements du régime hydrologique des bassins versants qui, à leur tour, peuvent avoir un effet potentiellement bénéfique ou préjudiciable sur la qualité de l'habitat en milieux humides. L'hydropériode est plus longue dans certains milieux humides après la coupe (Wind, 2008a), alors que d'autres s'assèchent avant la fin du développement des larves (Beasley et al., 2000; Wind et Dunsworth, 2006). L'enlèvement d'arbres dans les zones riveraines bordant de petits étangs dans la région des lacs Nanaimo a mené à l'élimination de l'ombrage et à l'augmentation de la température de l'eau, incitant un grand nombre de grenouilles à pattes rouges du Nord à pondre des grappes d'œufs dans certains sites (Wind, 2008a). Or, on ne sait pas si les individus métamorphosés, quittant les étangs en direction des zones déboisées, survivraient sans l'humidité et le couvert végétal normalement présents en zones riveraines. Il n'existe actuellement aucune donnée permettant de répondre à cette question. Les résultats d'études de modélisation donnent à penser que les facteurs de stress qui agissent sur les individus métamorphosés et les juvéniles sont les plus susceptibles d'influer sur les fluctuations démographiques (Biek et al., 2002; Govindarajulu et al., 2005). Les petits milieux humides (< 0,5 ha) sont importants à titre d'habitat pour l'hydratation et l'alimentation durant les étés secs (Golder Associates Inc., 2007) ainsi que pour la reproduction. Une protection ne leur est toutefois pas accordée en Colombie-Britannique en vertu de la Forest and Range Practices Act. En fait, les petits milieux humides ne sont habituellement pas identifiés sur les cartes (Beasley et al., 2000; Wind, 2008a).
Les activités forestières modifient les milieux terrestres de plusieurs manières. L'enlèvement du couvert forestier entraîne une réduction de l'humidité, de plus grandes fluctuations de la température et une augmentation de la vitesse des vents au niveau du tapis forestier (Chen et al.,1990, 1992). Le compactage et la perturbation mécanique du sol réduisent la quantité de bois mort sur pied, l'épaisseur de la litière de feuilles et le nombre de terriers. Ces changements physiques altèrent les ressources alimentaires (c.-à-d. l'abondance des invertébrés; voir Addison et al., 2003), les refuges utilisés pendant le jour, les abris contre les prédateurs et les hibernacles pour les amphibiens (Hayes, et al., 2008). Chan-McLeod (2003) a démontré que les coupes à blanc de moins de 12 ans faisaient obstacle aux déplacements des grenouilles à pattes rouges du Nord par temps sec (voir Habitat de migration et de dispersion). Les coupes à blanc existantes deviendront plus hospitalières à mesure que le couvert forestier se reconstitue (Chan-McLeod, 2003) et que les forêts vieillissent (Aubry et Hall, 1991; Aubry, 2000); toutefois, la menace récurrente de l'exploitation forestière se manifeste sur un cycle de révolution de 80 ans (ou moins).
Les répercussions négatives de l'exploitation forestière peuvent être atténuées dans une certaine mesure par l'ajustement de la configuration spatiale des parterres de coupe ainsi que de la superficie et de l'emplacement des parcelles boisées résiduelles (Chan-McLeod et Moy, 2007). Ainsi, les impacts de l'exploitation forestière future dépendront de la probabilité que la superficie et la configuration du couvert forestier maintenu, à l'échelle du peuplement et du paysage, assure une protection et une connectivité suffisantes de l'habitat.
Les aménagements hydroélectriques créent des bassins de retenue qui peuvent inonder les vallées fluviales, les milieux humides et les milieux forestiers secs. Par exemple, le bassin versant de la rivière Jordan, dans l'île de Vancouver, a été endigué à la fin du 20e siècle, ce qui a donné lieu à la création de grands réservoirs qui ont altéré plus de 90 ha d'habitat de reproduction propice de la grenouille à pattes rouges du Nord (Hawkes, 2005). Les lacs Kettle, Buttle et Elsie (perte de 72 ha) ainsi que la rivière Campbell ont aussi été endigués. La fluctuation des niveaux d'eau empêche l'établissement de milieux riverains peu profonds offrant la végétation émergente nécessaire à la ponte; par exemple, le niveau de l'eau dans le réservoir Division du bassin versant de la rivière Jordan présente des variations allant jusqu'à 9,6 m durant la période de reproduction critique de la grenouille à pattes rouges du Nord (Hawkes, 2005). En outre, des ouvrages de gestion de l'eau sont utilisés pour prévenir les inondations et améliorer l'habitat de la sauvagine. De nombreuses digues du Lower Mainland sont vétustes et doivent être remplacées, par exemple les digues du parc régional Minnekhada et des milieux humides de Codd. Le non-remplacement de ces ouvrages pourrait entraîner l'intrusion d'eau de mer, qui détruirait l'habitat d'eau douce de la grenouille à pattes rouges du Nord. La gestion de l'eau en faveur de la sauvagine peut aller à l'encontre de celle en faveur des amphibiens. Par exemple, dans le marais du lac Mount, le niveau de l'eau a été abaissé afin d'améliorer les habitats d'alimentation et de nidification de la sauvagine, mais cela a mené à une perte d'habitat de reproduction de la grenouille à pattes rouges.
Des plantes envahissantes, comme l'alpiste roseau (Phalaris arundinacea) et la salicaire commune (Lythrum salicaria), se sont rapidement propagées dans la région côtière de la Colombie-Britannique. L'alpiste roseau se rencontre dans tous les milieux humides du Lower Mainland, qui constitue plus de 10 % de l'aire de répartition de la grenouille à pattes rouges du Nord.
La grenouille à pattes rouges du Nord est exposée à la pollution provenant d'une gamme de sources, dont les eaux usées urbaines, les effluents industriels, agricoles et forestiers et les polluants atmosphériques. Les mares, les étangs et autres milieux humides qui servent d'habitat agissent comme puits pour divers polluants, ce qui expose les amphibiens se reproduisant en milieu aquatique à des contaminants pendant les premiers stades critiques du cycle vital (Vitt et al., 1990). Les produits chimiques sont rapidement absorbés par la peau des amphibiens et la couche gélatineuse des œufs. Il a été établi que les conditions des eaux pluviales (métaux et chlorure dans les sédiments issus des sels de voirie) sont toxiques pour les embryons et les larves des ranidés (Snodgrass et al., 2008).
Des pesticides organochlorés ont été appliqués à grande échelle dans la vallée du Fraser dans les années 1970 (Finizio et al., 1998), avant d'être remplacés par des pesticides organophosphorés (De Solla et al., 2002a). L'atrazine est utilisée sur le maïs de grande culture presque partout dans la vallée (Belzer et al., 1998; Pearson, comm. pers., 2011), et les résidus peuvent rester dans les sols et les plans d'eau (Top, 1996; Environment Canada, 2011a). Les fermes non biologiques utilisent le glyphosate pour lutter contre les mûriers le long des clôtures et préparer les champs au printemps, en plus de l'appliquer sur le maïs Round Up Ready et parfois directement le long des voies d'eau (Pearson, comm. pers., 2011; Environment Canada, 2011a). À part l'application commerciale d'herbicides et de pesticides, on rapporte le ruissellement de nitrates, de nitrites et de phosphates et le transport de lixiviats par les eaux souterraines, deux phénomènes à la hausse dans la vallée du Fraser à cause de l'augmentation spectaculaire des densités de bétail (Schindler et al., 2006; Environment Canada, 2011b).
De nombreux pesticides et engrais agricoles sont toxiques pour les amphibiens, causant des effets mutagènes et des anomalies du développement (Bonin et al.,1997), et réduisant la disponibilité des aliments. Dans le cadre d'une expérience, le succès d'éclosion des œufs de grenouille à pattes rouges du Nord placés dans des fossés agricoles autour de Sumas Prairie, en Colombie-Britannique, était beaucoup plus faible (jusqu'à 9 et 34 % au cours de chacune de 2 années, respectivement) que celui des œufs placés dans des sites de référence moins exposés au ruissellement agricole (85 % ou plus) (De Solla et al., 2002a). Par rapport aux sites de référence, les concentrations d'ammoniac et de phosphate total ainsi que la demande biochimique en oxygène (DBO) étaient plus élevées dans les fossés agricoles (De Solla et al., 2002a), alors que les teneurs en pesticides organochlorés et en polychlorobiphényles (PCB) étaient semblables et non toxiques (De Solla et al., 2002b; Loveridge et al., 2007). En laboratoire, des analyses de sous-produits azotés issus d'engrais agricoles utilisés dans la vallée de la Williamette ont révélé que les larves de grenouille à pattes rouges du Nord étaient sensibles au sulfate d'ammonium et aux ions d'ammonium dérivés de composés apparentés (Schuytema et Nebeker, 1999; Nebeker et Schuytema, 2000). Il a été établi que le nitrate d'ammonium et le sulfate d'ammonium réduisent le taux de croissance et causent la mort des larves de grenouille à des concentrations plus faibles que celles généralement appliquées, lesquelles sont par ailleurs inférieures aux critères de qualité de l'eau de l'Environmental Protection Agency des États-Unis visant l'eau destinée à la consommation humaine et les poissons d'eaux chaudes (Marco et al., 1999). L'atrazine est un perturbateur endocrinien qui féminise les grenouilles léopards (Lithobates pipiens) mâles à des concentrations bien inférieures à celles normalement appliquées en laboratoire et dans la nature (Hayes et al., 2003). Dans le nord-ouest de la Californie, des grenouilles à pattes rouges du Nord mâles adultes et subadultes ont produit un biomarqueur qui laisse deviner une exposition à des composés féminisants (Bettaso et al., 2002, in Hayes et al., 2008). Il est probable que de nombreux perturbateurs endocriniens existent dans les effluents urbains et agricoles et qu'ils n'ont pas encore fait l'objet d'analyses visant à déterminer leurs effets sur les amphibiens (Hayes et al., 2008).
Une portion (moins de 30 %) de l'habitat de la grenouille à pattes rouges du Nord devrait subir un aménagement sylvicole intensif, dans le cadre duquel des herbicides seront peut-être utilisés. Les amphibiens en général sont sensibles aux effets des herbicides à base de glyphosate utilisés en foresterie pour la préparation des sites et le dégagement de conifères (Govindarajulu, 2008). Bien que les lignes directrices sur l'épandage protègent la plupart des plans d'eau et des zones riveraines, ces herbicides peuvent être pulvérisés sur des ruisseaux asséchés et des étangs temporaires. Par exemple, en 2009, une entreprise forestière a appliqué du glyphosate par hélicoptère sur un peuplement situé près de 2 ruisseaux à saumons et de 1 milieu humide de l'inlet Port Neville, dans l'île de Vancouver (Sowden, comm. pers., 2011). Des études en laboratoire révèlent que le Diuron™, herbicide couramment utilisé, peut ralentir le développement des membres et réduire la survie de la grenouille à pattes rouges du Nord à des concentrations supérieures à celles utilisées en conditions normales de pulvérisation sur le terrain, mais pouvant être observées dans de petites zones d'accumulation d'eau où l'herbicide se retrouve après l'application (Schuytema et Nebeker, 1998).
On en sait peu sur l'étendue des polluants atmosphériques en Colombie-Britannique, mais plusieurs insecticides organophosphorés, notamment le diazinon et le malathion, ont été trouvés dans des échantillons d'air prélevés récemment, soit en 2005, à Abbotsford (Raina et al., 2010). En Californie, des pesticides agricoles aéroportés ont été mis en cause dans le déclin de populations de grenouilles à pattes rouges de la Californie (Davidson et al., 2002).
La majeure partie de l'aire de répartition de la grenouille à pattes rouges du Nord est comprise dans la zone biogéoclimatique côtière à pruche de l'Ouest, qui devrait s'élargir au cours des 35 prochaines années en raison des changements climatiques, principalement vers de plus hautes altitudes (Wang et al., 2012). D'ici 2050, la zone biogéoclimatique côtière à douglas (d'une superficie actuelle de 0,25 million d'ha, soit environ 6 % de l'aire de répartition de l'espèce à 500 m d'altitude ou moins dans l'île de Vancouver) rétrécira de 19 % par rapport à sa superficie actuelle et s'agrandira de 16 % dans d'autres régions (Wang et al., 2012), ce qui entraînera une faible perte nette (- 3 %). Il est possible que la grenouille à pattes rouges du Nord change d'habitat, compte tenu de sa capacité de se déplacer sur de longues distances (voir Habitat de migration et de dispersion). Elle pourrait, par exemple, se déplacer vers de plus hautes altitudes à mesure que la température augmente, mais cela est incertain, étant donné que l'espèce se trouve habituellement à des altitudes inférieures à 500 m, en terrain relativement plat (voir Altitude et Habitat terrestre d'alimentation).
À mesure que la température augmente et que les phénomènes météorologiques deviennent extrêmes à cause des changements climatiques, les modèles prévoient des hivers plus humides et des étés plus secs dans l'aire de répartition canadienne de l'espèce. Les perturbations hydrologiques peuvent avoir des effets mixtes sur les populations d'amphibiens (Walls et al., 2013). Des hivers plus humides pourraient se révéler bénéfiques pour la grenouille à pattes rouges du Nord en lui offrant plus de parcelles d'habitat de reproduction pendant la période de ponte (de février à avril). Des étés plus secs pourraient aussi se révéler bénéfiques si des plans d'eau permanents deviennent des étangs éphémères qui abritent moins de prédateurs. Il pourrait cependant y avoir des conséquences négatives. Les milieux humides côtiers pourraient être engorgés d'eau à répétition par suite de l'élévation du niveau de la mer et de la hausse de la fréquence des ondes de tempête hivernale (Beckmann et al., 1997). Dans les milieux humides éphémères, les sécheresses estivales peuvent raccourcir les hydropériodes, de sorte que les larves n'aient pas le temps de se développer, et augmenter la mortalité accrue des individus métamorphosés en dispersion, la distance entre les milieux humides étant accrue.
Un modèle des changements prévus des températures estivales maximales dans l'ensemble de l'aire de répartition canadienne actuelle de la grenouille à pattes rouges du Nord indique que, d'ici 2080, 45 % des occurrences se trouveront dans des milieux limitatifs du point de vue thermique (Gerick et al., 2014). Des fluctuations extrêmes de la température, le gel printanier ou un temps trop chaud trop tôt dans la saison pourraient être néfastes pour les embryons ou les larves. Les changements combinés de la température et de l'hydropériode auront une incidence néfaste sur le taux de développement des larves, leur taille à la métamorphose et leur probabilité de survie jusqu'à ce stade, mais les résultats dépendront de la mesure à laquelle la température accrue peut compenser l'assèchement plus rapide et la plasticité du taux de développement (O'Regan et al., 2014). Des expériences en réservoirs de stockage de l'eau destinée au bétail ont révélé que les larves de grenouille à pattes rouges du Nord réagissent à des températures élevées et à l'assèchement rapide en accélérant leur développement et en devançant leur métamorphose. La taille à la métamorphose avait peu changé parce que le réchauffement avait entraîné une poussée de croissance du périphyton jusqu'à un niveau permettant de satisfaire les besoins métaboliques plus élevés des têtards. Le taux de mortalité était faible parce que l'hydropériode expérimentale était suffisamment longue pour permettre à l'espèce de se développer. Dans des conditions où la température varie davantage et où l'assèchement est plus rapide que dans les conditions expérimentales, O'Regan et al. (2014) prévoient que la grenouille à pattes rouges du Nord subirait de plus graves effets létaux.
Les autres conséquences possibles des changements climatiques dépendent des interactions avec d'autres menaces. La probabilité d'une éclosion de chytridiomycose pourrait augmenter suite à une plus grande variabilité des températures extrêmes (c.-à-d. des températures plus extrêmes entre le jour et la nuit) (Hamilton et al., 2012). Des températures à la hausse pourraient se traduire par une eutrophisation accrue des eaux chaudes réceptrices de ruissellements agricoles (en particulier dans les basses terres du Fraser) et un taux d'invasion et une productivité plus élevés du ouaouaron (Compass Resource Management, 2007). Des eaux plus chaudes mèneront aussi à l'expansion de l'aire de répartition de poissons indigènes et de poissons exotiques d'eaux chaudes (Chu et al., 2005; Rahel et Olden, 2008). L'exposition au rayonnement UV-B pourrait augmenter si les niveaux d'eau chutent et, bien que le rayonnement UV-B ambiant n'ait pas réduit le taux d'éclosion d'œufs de grenouille à pattes rouges du Nord (Blaustein et al., 1996; Ovaska et al., 1997), la croissance et le développement plus lents des larves ont été associés à l'exposition expérimentale des embryons (Belden et Blaustein, 2002a).
Une seule menace pourrait détruire un nombre inconnu, mais élevé, de localités fréquentées par la grenouille à pattes rouges du Nord, compte tenu de la distribution des occurrences connues et des types de menaces (tableau 1). L'exploitation forestière menace 222 occurrences dispersées dans les parties éloignées de l'aire de répartition de l'espèce. L'aménagement de routes et l'aménagement de terrains aux fins de la construction d'habitations et d'activités commerciales menacent respectivement 232 et 106 occurrences dans les régions dominées par l'occupation humaine. Ces dernières menaces agiront beaucoup plus rapidement, comme l'indique le déclin de population qui s'est produit lorsque le tracé de la route 99 a été modifié au sein d'un milieu humide de reproduction de grenouilles à pattes rouges du Nord. Il est peu probable que les effets des changements climatiques et les phénomènes météorologiques violents, comme les sécheresses prolongées, perturberont rapidement une grande partie de l'aire de répartition de l'espèce, du moins à court terme.
Au Canada, le COSEPAC a évalué la grenouille à pattes rouges du Nord en 1999 et l'a désignée « espèce préoccupante ». Le statut a été réexaminé et confirmé en 2002, en 2004 et en mai 2015. L'espèce est inscrite sur la liste fédérale des espèces préoccupantes de l'annexe 1 de la Loi sur les espèces en péril (LEP). La LEP n'impose pas d'exigences immédiates en ce qui concerne la protection de l'habitat des espèces ainsi désignées. La grenouille à pattes rouges du Nord est toutefois considérée comme un élément du processus d'évaluation environnementale qui exige que les effets néfastes possibles d'un projet soient décrits et, si le projet est exécuté, que des mesures soient prises pour éviter ou réduire et suivre ces effets néfastes. De telles mesures doivent être conformes au plan de gestion qui s'applique à une espèce préoccupante (Species at Risk Public Registry, 2013). Une ébauche de plan de gestion visant la grenouille à pattes rouges du Nord est en cours d'examen (Govindarajulu, comm. pers., 2015).
La collecte, la possession et le commerce de toutes les espèces de vertébrés indigènes, y compris les amphibiens, sont interdits aux termes de la Wildlife Act de la Colombie-Britannique. L'efficacité de la loi pour ce qui est de protéger les grenouilles est limitée parce qu'elle est difficile à appliquer et ne couvre pas les dommages à l'habitat.
La grenouille à pattes rouges du Nord est cotée G4 (« apparemment non en péril », désignation en juin 2008) à l'échelle mondiale (Hammerson, 2008). La cote N4 (« apparemment non en péril », désignation en novembre 1996) lui a été attribuée à l'échelle des États-Unis. Son statut dans divers États est le suivant : Californie, S2? (« en péril »?); Oregon, S3S4 (« vulnérable à la disparition ou à l'extinction » à « apparemment non en péril »); État de Washington, S4 (« apparemment non en péril ») (NatureServe, 2013). La cote S3S4 indique qu'une espèce est exposée à un risque de disparition modéré dans le territoire concerné à cause d'une aire de répartition limitée, d'un petit nombre de populations ou d'occurrences, de baisses d'effectifs marquées, de menaces graves ou d'autres facteurs.
Le statut de conservation S3S4 a été attribué à la grenouille à pattes rouges du Nord en Colombie-Britannique (désignation en décembre 2010) (B.C. Conservation Data Centre, 2014). Elle figure sur la liste bleue provinciale des espèces en péril. Les espèces figurant sur cette liste sont des taxons préoccupants qui ont des caractéristiques qui les rendent particulièrement sensibles ou vulnérables aux activités humaines ou aux phénomènes naturels (BC Species and Ecosystems Explorer, 2013). La grenouille à pattes rouges du Nord a été inscrite sur la liste des espèces sauvages désignées (« Identified Wildlife ») incluse dans la version 2004 de la stratégie de gestion des espèces sauvages désignées (« Identified Wildlife Management Strategy »). Cette version contient des lignes directrices précises sur la gestion de l'habitat de la grenouille à pattes rouges du Nord et des lignes directrices sur les zones d'espèces sauvages (« Wildlife Habitat Areas », ou WHA) en terres forestières.
Environ 16 % des occurrences documentées de l'espèce se trouvent dans des aires protégées (tableau 1). L'espace compris dans ces parcs et réserves écologiques couvre une superficie de 1 900 à 3 013 km2 à des altitudes de moins de 500 m et à moins de 1 000 m, respectivement (tableau 4 et tableau 5). Environ 13 % du territoire dans l'île de Vancouver est protégé. Un pourcentage total de 24 % de l'île appartient à des intérêts privés, dont 75 % sont des terres forestières privées (van Kooten, 1995; Sierra Club, 2003). La majeure partie des terres forestières privées et des ensembles urbains et agricoles sont situés dans le quadrant sud-est de l'île de Vancouver. Cette région semble importante pour la grenouille à pattes rouges du Nord en raison de l'abondance de milieux humides et de forêts de faible altitude. Pourtant, elle renferme relativement peu d'aires protégées. En 2003, la réserve de parc national des Îles-Gulf a été établie au large de la côte sud-est de l'île de Vancouver. Le nouveau parc comprend principalement des terres qui étaient déjà protégées en tant que parcs régionaux ou provinciaux, mais aussi de nouvelles acquisitions. Plusieurs secteurs du parc renferment un habitat pour la grenouille à pattes rouges du Nord. Les parties ouest et nord de l'île de Vancouver sont constituées en grande partie de terres publiques, utilisées principalement par des sociétés forestières à des fins de récolte, bien que 2 grandes zones soient protégées au sein des parcs provinciaux Brooks-Peninsula et Cape-Scott. De plus, une étroite bande est protégée dans la réserve de parc national Pacific Rim; elle s'élargit à la hauteur de Nitnat et rejoint le parc provincial Carmanah-Walbran. Le secteur de la plage Long de la réserve de parc national Pacific Rim est divisé par une route, site marqué par des taux chroniques de mortalité routière chez l'espèce (Beasley, 2006).
Dans le Lower Mainland, une minime partie du territoire est protégée, et la plus grande superficie se compose de petits parcs entourés de lotissements urbains et ruraux. Un grand nombre des sites réputés être fréquentés par la grenouille à pattes rouges du Nord se trouvent dans ces parcs. La protection de ces terres n'empêche malheureusement pas la dégradation et la fragmentation de l'habitat (Waye, 1999).
La plus grande partie du territoire de la Sunshine Coast et de la côte centre-sud fait l'objet d'un zonage permettant des activités forestières, mais un certain nombre de nouvelles aires protégées ont été créées le long de la côte centre-sud en 2006 et 2007. Les plus grandes sont les aires de conservation Ugwiwey/Cape Caution (102 km2), Lockhard-Gordon (245 km2), Tsa-latl/Smokehouse (378 km2), Phillips Estuary/?Nacinuxw (14 km2) et Palemin/Estero Basin (29 km2). Aucun relevé visant la grenouille à pattes rouges du Nord n'a été mené dans ces parcs, mais ceux-ci chevauchent la zone d'occurrence de l'espèce.
La Forest and Range Practices Act de la Colombie-Britannique prévoit quelques dispositions pour protéger l'habitat de la grenouille à pattes rouges du Nord par le biais de règlements sur les pesticides et la largeur des zones riveraines et des mesures générales visant les espèces sauvages (« General Wildlife Measures ») énoncées dans la stratégie de gestion des espèces sauvages désignées. Par exemple, le règlement sur la gestion des zones riveraines exige que des zones tampons de couvert forestier non perturbé soient maintenues autour des grands milieux humides et des cours d'eau. Mais ces dispositions ne visent pas les petits milieux humides (< 0,5 ha); or, ce sont ces petits milieux humides qui sont grandement utilisés par la grenouille à pattes rouges du Nord.
La stratégie de gestion des espèces sauvages désignées comprend des lignes directrices sur la protection et la gestion de la grenouille à pattes rouges du Nord qui prévoient l'établissement de WHA et des mesures générales les espèces sauvages connexes. L'île de Vancouver compte actuellement 23 WHA pour la grenouille à pattes rouges du Nord, d'une superficie totale de 336 ha (tableau 5). La création de 8 autres WHA dans l'île a été proposée, et le recensement mené sur la Sunshine Coast pourrait appuyer l'établissement de WHA dans cette région (McConkey, comm. pers., 2015). De plus, plusieurs WHA mises en place pour d'autres espèces, notamment le Guillemot marbré (Brachyramphus marmoratus), protègent aussi l'habitat de la grenouille à pattes rouges du Nord.
Description longue pour le tableau 5
| Environs | ||||||
|---|---|---|---|---|---|---|
| Nord de l'île de Vancouver | District de Campbell River, île de Vancouver | Sud de l'île de Vancouver | Lower Mainland | Corridor Sea-to-Sky | Sunshine Coast | Ensemble de l'aire de répartition en C.-B. |
| Site | Type de protection | Superficie (ha) |
|---|---|---|
| Étang Beacon, île Malcolm | WHA Table Footnotei | 3 |
| Rivière Marble, près du lac Alice | WHA Table Footnotei | 4 |
| Rivière Marble | PP Table Footnotei | 1 419 |
| Rivière Nimpkish | ER Table Footnotei | 18 |
| Cap Scott | PP Table Footnotei | 22 294 |
| Péninsule BrooksTable Footnoteh | PP Table Footnotei | 51 631 |
| Total | - | 75 369 |
| Site | Type de protection | Superficie (ha) |
|---|---|---|
| Brewster Bridge, Lac Brewster | WHA Table Footnotei | 1,6 |
| Diver 1, à l'ouest du lac Brewster | WHA Table Footnotei | 4,7 |
| Martha 1, près du lac Martha | WHA Table Footnotei | 15,8 |
| Marais Sorenson's, près de | WHA Table Footnotei | 11,9 |
| Kendrick, près du ruisseau Gunpowder | WHA Table Footnotei | 20,5 |
| Pointe Maquinna, île Nootka | WHA Table Footnotei | 15,5 |
| Total | - | 70 |
| Site | Type de protection | Superficie (ha) |
|---|---|---|
| Lac Swan, près d'Ucluelet | WHA Table Footnotei | 18,6 |
| Corrigan, près de Port Alberni | WHA Table Footnotei | 13 |
| Corrigan 6.5, près de Port Alberni | WHA Table Footnotei | 37,6 |
| Ruisseau Cous, près de Port Alberni | WHA Table Footnotei | 10,7 |
| Flora, près de Nitnat | WHA Table Footnotei | 3 |
| Plateau Noyse | WHA Table Footnotei | 52 |
| Ruisseau Upper Canoe Pass, près de l'inlet Effingham | WHA Table Footnotei | 4,5 |
| Passage Julia 1, près de l'inlet Effingham | WHA Table Footnotei | 10,4 |
| Passage Julia 2 | WHA Table Footnotei | 4,6 |
| Passage Julia 3 | WHA Table Footnotei | 9,4 |
| Ruisseau Rosemond, près de Jordan Ridge | WHA Table Footnotei | 33 |
| Ruisseau Loss Creek West, près de Jordan Ridge | WHA Table Footnotei | 26 |
| Ruisseau Loss Creek East | WHA Table Footnotei | 16 |
| Ruisseau McVicar, près de Jordan Ridge | WHA Table Footnotei | 9 |
| Ruisseau Pete Wolfe, près de Jordan Ridge | WHA Table Footnotei | 12 |
| Carmanah - Walbran | PP Table Footnotei | 16 450 |
| Rivière Cowichan | PP Table Footnotei | 1 414 |
| Île NewcastleTable Footnoteh | PP Table Footnotei | 336 |
| Rivière EnglishmanTable Footnoteh | PP Table Footnotei | 97 |
| Rivière EnglishmanTable Footnoteh | RP Table Footnotei | 207 |
| Parc GoldstreamTable Footnoteh | PP Table Footnotei | 477 |
| Lac Kennedy | PP Table Footnotei | 241 |
| Pacific Rim, Sentier de la Côte-Ouest, Cap Beale, Plage Long | NPR Table Footnotei | ~ 27 900 |
| Strathcona | PP Table Footnotei | ~ 43 800 - ~ 133 000 Table Footnotej |
| Plage Miracle | PP Table Footnotei | 137 |
| Chutes Little Qualicum | PP Table Footnotei | 440 |
| Ruisseau Rosewall | PP Table Footnotei | 54.3 |
| Île Denman | CC Table Footnotei | ~ 191 |
| Parc Westwood-Lake, Nanaimo | MP Table Footnotei | 50 |
| Îles Gulf, Pender et Saturna | NPR Table Footnotei | ~ 3 600 |
| Lac Thetis, Saanich | RP Table Footnotei | 834 |
| Francis King, Saanich | RP Table Footnotei | 200 |
| Tourbière Rithet's, Saanich | CA Table Footnotei | 42 |
| Goldstream, Saanich | PP Table Footnotei | 477 |
| Total | - | 97 207 – 186 407 |
| Site | Type de protection | Superficie (ha) |
|---|---|---|
| Canyon Lynn, Vancouver Nord | MP Table Footnotei | 250 |
| Lac Burnaby, Burnaby | RP Table Footnotei | 3,11 |
| Île Douglas, Port Coquitlam | FRIR Table Footnotei | 187 |
| Minnekhada, Coquitlam | RP Table Footnotei | 175 |
| Golden Ears | PP Table Footnotei | 8 450 - 30 500 Table Footnotej |
| Milieux humides du ruisseau Silverdale, Mission | MP Table Footnotei | 46 |
| Sasquatch | PP Table Footnotei | 1 217 |
| F.H. Barber, près de Hope | PP Table Footnotei | 8,5 |
| Bridal Veil, Chilliwack | PP Table Footnotei | 32 |
| Milieux humides du lac Cheam, Chilliwack | RP Table Footnotei | 107 |
| Lac Cultus, Chilliwack | PP Table Footnotei | 2 729 |
| Ruisseau Willibrand, Abbotsford | MP Table Footnotei | 41,5 |
| Sentier Clearbrook, Abbotsford | MP Table Footnotei | 9,2 |
| Île Brae, près de Langley | RP Table Footnotei | 69 |
| Parc Latimer, Langley | MP Table Footnotei | 16,8 |
| Campbell Valley, Langley | RP Table Footnotei | 450,6 |
| Parc régional Aldergrove | RP Table Footnotei | 166,5 |
| Parc Hawthorn, Surrey | MP Table Footnotei | 27,8 |
| Forêt urbaine Green Timbers, Surrey | MP Table Footnotei | 239,7 |
| Parc Crescent, Plage Crescent | MP Table Footnotei | 46,8 |
| Réserve naturelle de Delta, Tourbière Burns | ECA Table Footnotei | 2 042 |
| Total | - | 16 314 - 38 364 |
| Site | Type de protection | Superficie (ha) |
|---|---|---|
| Lac Alice, Squamish | PP Table Footnotei | 396 |
| Chutes Brandywine, près de Whistler | PP Table Footnotei | 420 |
| Total | - | 816 |
| Site | Type de protection | Superficie (ha) |
|---|---|---|
| Smuggler Cove Marine, Sechelt | PP Table Footnotei | 185 |
| Baie Sargeant | PP Table Footnotei | 142 |
| Total | - | 327 |
| Site | Grand Total |
|---|---|
| Ensemble de l'aire de répartition en C.-B. | 190 103 - 301 353 |
Les administrations municipales et régionales des régions du Lower Mainland, de l'île de Vancouver et de la Sunshine Coast ont préparé des plans d'aménagement des terres, des règlements administratifs et des règlements de zonage qui offrent un certain degré de protection à l'habitat en milieux humides. Le ministère de la Protection de l'eau, des terres, et de l'air de la Colombie-Britannique a préparé un ensemble de pratiques de gestion bénéfiques s'appliquant aux amphibiens et aux reptiles dans l'ensemble de la province (Ovaska et al., 2003).
Des organisations de conservation (Canards Illimités, The Land Conservancy, Pacific Parkland Foundation, Nature Trust of BC) jouent un rôle important dans l'acquisition, la protection et la restauration de milieux humides et de milieux terrestres adjacents dans le sud-ouest de la Colombie-Britannique. Quelques projets, tels que ceux visant les milieux humides du lac Cheam et de l'île Codd, les tourbières Burns et Blaney ainsi que le marais Pitt-Addison, peuvent être de taille suffisante pour protéger les milieux humides et le couvert forestier adjacent propices à la grenouille à pattes rouges du Nord.
Environnement Canada a financé la préparation du présent rapport de situation. Kristiina Ovaska a dirigé et rigoureusement révisé chacune des ébauches et, avec Lennart Sopuck, rédigé le rapport de situation de 2004, qui forme une partie du présent « document évolutif ». Le calcul des menaces était fondé sur des discussions menées par Purnima Govindarajulu, auxquelles ont participé Laura Matthias, Darryn McConkey, Aimee Mitchell, Kristiina Ovaska, Monica Pearson, Kym Welstead et Elke Wind en 2011, de même que Dave Fraser et Kristiina Ovaska en 2014. Jenny Wu a créé la carte de répartition en Colombie-Britannique et calculé la zone d'occurrence ainsi que l'indice de zone d'occupation. L'IUCN, Conservation International et NatureServe ont fourni les données de la carte de l'aire de répartition mondiale élaborée dans le cadre du projet d'évaluation mondiale des amphibiens (« Global Amphibian Assessment »). Gerald Schreiber a aidé à recueillir l'information sur la superficie des aires protégées. Plusieurs personnes ont généreusement fourni des données sur les sites et les activités de recherche; elles figurent dans la section « Experts contactés » et/ou « Sources d'information » ci-dessous. Purnima Govindarajulu, David Green, Elizabeth Jordan, Joshua Malt, Dennis Murray, Marie-France Noël et Cindy Paszkowski ont apporté de précieux commentaires et suggestions pour améliorer le rapport.
Cook, Francis, chercheur émérite, Musée canadien de la nature, Ottawa (Ontario).
Demarchi, Diana, Ministry of Environment, Victoria (Colombie-Britannique).
Evelyn, Michelle, responsable de projet, Sunshine Coast Wildlife Project, Gibsons (Colombie-Britannique).
Filion, Alain, Secrétariat du COSEPAC, Gatineau (Québec).
Fraser, David, Species at Risk Specialist, B.C. Ministry of Environment, Victoria (Colombie-Britannique).
Gelling, Leah, B.C. Conservation Data Centre, Ministry of Environment, Victoria (Colombie-Britannique).
Govindarajulu, Purnima, Herpetofauna Specialist, B.C. Ministry of Environment, Victoria (Colombie-Britannique).
Hanke, Gavin, conservateur – collection des amphibiens. Royal B.C. Museum, Victoria (Colombie-Britannique).
Hawkes, Virgil, Senior Wildlife Biologist, LGL Limited Environmental Research Associates, Victoria (Colombie-Britannique).
Howes, Briar, Conservation et gestion des espèces, Parcs Canada, Gatineau (Québec).
Knopp, Denis, BC's Wild Heritage, Chilliwack (Colombie-Britannique).
Malt, Joshua, Ecosystem Biologist, Ministry of Forests, Lands and Natural Resource Operations, Surrey (Colombie-Britannique).
McConkey, Darryn, Senior Ecosystem Biologist, Ministry of Forests, Lands and Natural Resource Operations, Nanaimo (Colombie-Britannique).
Mitchell, Aimee, Coastal Painted Turtle Project, Vancouver (Colombie-Britannique).
Murray, Rylee, chercheur de premier cycle, Biological Sciences, Simon Fraser University, Burnaby (Colombie-Britannique).
Nernberg, Dean, agent responsable des espèces en péril, Défense nationale, Ottawa (Ontario).
Palen, Wendy, professeure adjointe, Biological Sciences, Simon Fraser University, Burnaby (Colombie-Britannique).
Steigerwald, Michèle, gestionnaire adjointe des collections, Collection d'amphibiens et de reptiles, Musée canadien de la nature, Ottawa (Ontario).
Stipec, Katrina, B.C. Conservation Data Centre, Ministry of Environment, Victoria (Colombie-Britannique).
Wind, Elke, E. Wind Consulting, Nanaimo (Colombie-Britannique).
Adams, M.J. 1999. Correlated factors in amphibian decline: exotic species and habitat change in western Washington. Journal of Wildlife Management 63:1162–1171.
Adams, M.J. 2000. Pond permanence and the effects of exotic vertebrates on anurans. Ecological Applications 10:559–568.
Adams, M.J., R.B. Bury et S.A. Swarts. 1998. Amphibians of the Fort Lewis Military Reservation, Washington: sampling techniques and community patterns. Northwestern Naturalist 79:12-18.
Adams, M.J., C.A. Pearl, S.K. Galvan et B. McCreary. 2011. Non-native species impacts on pond occupancy by an anuran. Journal of Wildlife Management 75:30-35.
Adams, M.J., S.D. West et L. Kalmbach. 1999. Amphibian and reptile surveys of U.S. navy lands on the Kitsap and Toandos Peninsulas, Washington. Northwestern Naturalist 80:1-7.
Addison, J.A., J.A. Trofymow et V.G. Marshall. 2003. Abundance, species diversity and community structure of Collembola in successional coastal temperate forests on Vancouver Island, Canada. Applied Soil Ecology 24:233-246.
Altig, R. et P.C. Dumas. 1972. Rana aurora. In Reimer, W.J. (ed.), Catalogue of American Amphibians and Reptiles 160. American Society of Ichthyologists and Herpetologists.
Aubry, K.B. 2000. Amphibians in managed, second-growth Douglas-fir forests. Journal of Wildlife Management 64:1041–1052.
Aubry, K.B. et P.A. Hall. 1991. Terrestrial amphibian communities in the southern Washington Cascade Range. Pp. 327–338, in L.F. Ruggiero, K.B. Aubry, A.B. Carey and M.H. Huff (eds.). Wildlife and Vegetation of Unmanaged Douglas-Fir Forests. General Technical Report PNW-GTR-285. [consulté en juillet 2014].
Baird, S.F. et C. Girard. 1852. Descriptions of new species of reptiles, collected by the U.S. Exploring Expedition under the command of Capt. Charles Wilkes, U.S.N. First part -- Including the species from the western coast of America. Proceedings of the Academy of Natural Sciences of Philadelphia 6:174–177.
Baylis, A., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Juillet 2014. Biologiste en conservation, A Rocha Canada, Surrey, British Columbia.
BC Stats. 2014. Population estimates. BC Ministry of Technology, Innovation and Citizens' Services. [consulté en décembre 2014].
Beasley, B. 2006. A study of the incidence of amphibian road mortality between Ucluelet and Tofino, British Columbia. Wildlife Afield 3:1 Supplement:23-28.
Beasley, B. 2011. Wetland surveys for breeding amphibians within the Clayoquot Biosphere Reserve Region 2008-2011. Interim Monitoring Report. Association of Wetland Stewards for Clayoquot and Barkley Sounds. Ucluelet, British Columbia. 31 p. + annexes.
Beasley, B., données inédites. 2014. Données inédites provenant du projet SPLAT et de la surveillance des milieux humides de la région s’étendant entre Ucluelet et Tofino. Biologiste, Association of Wetland Stewards for Clayoquot and Barkley Sounds, Ucluelet, British Columbia.
Beasley, B., C. Addison et K. Lucas. 2000. Clayoquot Sound amphibian inventory 1998-1999. Report for the Ministry of Environment, Lands and Parks, Vancouver Island Region Office, Nanaimo, B.C. and the Long Beach Model Forest Society, Ucluelet, British Columbia. 67 p. + annexes.
Beckmann, L., Dunn, M. et K. Moore. 1997. Effects of climate change on coastal systems in BC and the Yukon. Pp 8:1-26 in E. Taylor and B. Taylor (eds.), Responding to Global Climate Change in British Columbia and the Yukon, Vancouver, British Columbia.
Belden, L.K. et Blaustein, A.R. 2002. Exposure of red-legged frog embryos to ambient UV-B radiation in the field negatively affects larval growth and development. Oecologia (Berlin) 130:551–554.
Belzer, W., C. Evans et A. Poon. 1998. Atmospheric concentrations of agricultural chemicals in the Lower Fraser Valley. Fraser River Action Plan, Environment Canada, Vancouver, British Columbia. 61 p. Fichier PDF [consulté en juillet 2014].
Bettaso, J.B., H.H. Welsh, Jr. et B.D. Palmer. 2002. Northern red-legged frogs and endocrine disrupting compounds (EDCs). Froglog 52:1. Cité dans Hayes, M.P., T. Quinn, K.O. Richter, J.P. Schuett-Hames, et J.T. Serra Shean. 2008. Maintaining lentic-breeding amphibians in urbanizing landscapes: the case study of the Northern Red-legged Frog (Rana aurora). Pp. 445-461 in J.C. Mitchell, R.E. Jung Brown et B. Bartholomew (eds.). Urban Herpetology. Herpetological Conservation Vol. 3.
Biek, R., W.C. Funk, B.A. Maxell et L.S. Mills. 2002. What is missing in amphibian decline research: insights from ecological sensitivity analysis. Conservation Biology 16:728-734.
Blair, D. 2007. The City of Chilliwack 2007 Ditch Maintenance Program Environmental Monitor’s Report. Prepared for The City of Chilliwack. Nova Pacific Environmental. 27 p. + annexe. EcoCat: Ecological Reports Catalogue. [consulté en juillet 2014].
Blaustein, A.R., Beatty, J.J., Olson, D.H. et Storm, R.M. 1995. The biology of amphibians and reptiles in old-growth forests in the Pacific Northwest. USDA Forest Service, General Technical Report PNW-GTR-337. 98 p.
Blaustein, A.R., S.S. Gervasi, P.T.J. Johnson, J.T. Hoverman, L.K. Belden, P.W. Bradley et G.Y. Xie. 2012. Ecophysiology meets conservation: understanding the role of disease in amphibian population declines. Philosophical Transactions of the Royal Society B367:1688-1707.
Blaustein, A.R., PD. Hoffman, J.M. Kiesecker et J.B. Hays. 1996. DNA repair activity and resistance to solar UV-B radiation in eggs of the Red-legged Frog. Conservation Biology 10:1398–1402.
Blaustein, A.R., D.B. Wake et W.P. Sousa. 1994. Amphibian declines: judging stability, persistence, and susceptibility of populations to local and global extinctions. Conservation Biology 8:60–71.
Blood, D.A. et C. Henderson. 2000. Traffic-caused mortality of amphibians on Highway 4A, Vancouver Island, and potential mitigation. Rapport inédit préparé par D.A. Blood and Associates, Nanaimo, British Columbia. 14 p.
Bollinger T.K., J.H. Mao, D. Schock, R.M. Brigham et V.G. Chinchar. 1999. Pathology, isolation, and preliminary molecular characterization of a novel iridovirus from tiger salamanders in Saskatchewan. Journal of Wildlife Diseases 35:413–429.
Bonin, J., M. Ouellet, J. Rodrigue, J-L. DesGranges, F. Gagné, T.F. Sharbel et L.A. Lowcock. 1997. Measuring the health of frogs in agricultural habitats subjected to pesticides. Pp. 246-257 in D.M. Green (ed.). Amphibians in Decline: Canadian Studies of a Global Problem. Herpetological Conservation. Number One. Society for the Study of Amphibians and Reptiles.
Boyle, C.A., L. L.avkulich, H. Schreier et E. Kiss. 1997. Changes in land cover and subsequent effects on lower Fraser Basin ecosystems from 1827 to 1990. Journal of Environmental Management 21:185-196.
Briony Penn Associates. 2011. Burgoyne Bay Bioblitz Report. July 23rd and 24th, 2011. Burgoyne Bay Bioblitz Report [consulté en novembre 2013].
B.C. Conservation Data Centre. 2014. Conservation Status Report: Rana aurora. British Columbia Ministry of Environment. [consulté en juillet 2014].
B.C. Species and Ecosystems Explorer. 2013. [consulté en décembre 2013].
B.C. Ministry of Environment. 2012. Amphibian Inventory – Region 1 – Nanaimo – MOE, 2006 – ongoing. Données inédites. Species Inventory Web Explorer [consulté en novembre 2013].
Brown, H.A. 1975. Reproduction and development of the red-legged frog, Rana aurora, in northwestern Washington. Northwest Science 49:241–252.
Bury, R.B., P.S. Corn et K.B. Aubry. 1991. Regional patterns of terrestrial amphibian communities in Oregon and Washington. Pp. 341–350 in L.F. Ruggiero, K.B. Aubry, A.B. Carey et M.H. Huff (eds.). Wildlife and Vegetation of Unmanaged Douglas-Fir Forests.. General Technical Report PNW-GTR-285. [consulté en juillet 2014].
Calef, G.W. 1973a. Spatial distribution and "effective" breeding population of red-legged frogs (Rana aurora) in Marion Lake, British Columbia. Canadian Field-Naturalist 87:279–284.
Calef, G.W. 1973b. Natural mortality of tadpoles in a population of Rana aurora. Ecology 54:741–758.
Capital Regional District. 2008. State of the Region Report. 2008 Regional Growth Strategy Five-Year Monitoring Review. fichier PDF [consulté en décembre 2013].
Chan-McLeod, A.A. 2003. Factors affecting the permeability of clearcuts to red-legged frogs. Journal of Wildlife Management 67:663–671.
Chan-McLeod, A.A. et A. Moy. 2007. Evaluating residual tree patches as stepping stones and short-term refugia for Red-legged Frogs. Journal of Wildlife Management 71:1836-1844.
Chan-McLeod, A.A. et B. Wheeldon. 2004. Rana aurora (Northern Red-legged Frog), habitat and movement. Herpetological Review 35:375.
Chelgren, N.D., D.K. Rosenberg, S.S. Heppell et A.I. Gitelman. 2008. Individual variation affects departure rate from the natal pond in an ephemeral pond-breeding anuran. Canadian Journal of Zoology 86:260-267.
Chen, J., J.F. Franklin et T.A. Spies. 1990. Microclimatic pattern and basic biological responses at the clearcut edges of old-growth Douglas-fir stands. Northwest Environmental Journal 6:424-425.
Chen, J., J.F. Franklin et T.A. Spies. 1992. Contrasting microclimates among clearcut, edge, and interior of old-growth Douglas-fir forest. Agricultural and Forest 63:219-237.
Chu, C., N.E. Mandrak et C.K. Minns. 2005. Potential impacts of climate change on the distributions of several common and rare freshwater fishes in Canada. Diversity and Distributions 11:299-310.
City of Abbotsford. 2005. Scientific collection summary report form. Fish collection for salvage prior to and during instream works. EcoCat: Ecological Reports Catalogue. [consulté en novembre 2013].
Clegg, S. 2011. Assessing the impacts of vehicular mortality of migrating amphibians near Ryder Lake, B.C. Report for the Fraser Valley Conservancy. Abstract. Pp. 30-31 in Herpetofauna and Roads Workshop Program, Vancouver Island University, Nanaimo, B.C. February 22-23, 2011. Fichier PDF [consulté en juillet 2014].
Compass Resource Management. 2007. Major impacts: climate change. Technical Subcommittee Component Report for the Biodiversity BC Technical Subcommittee for the Report on the Status of Biodiversity in B.C. Fichier PDF [consulté en juillet 2014].
Conlon, J.M., N. Ali-Ghafari, L. Coquet, J. Leprince, T. Jouenne, H. Vaudry et C. Davidson. 2006. Evidence from peptidomic analysis of skin secretions that the red-legged frogs, Rana aurora draytonii and Rana aurora aurora, are distinct species. Peptides 27:1305-1312.
Cooke, S.A. 1995. Road mortality of common toads (Bufo bufo) near a breeding site 1974 – 1994. Amphibia – Reptilia 87-90 in Pukey, M. 2003. Amphibian mitigation measures in Central-Europe. Pp. 433-449 in C.L. Irwin, P. Garrett et K.P. McDermott (eds.), 2003 Proceeding of the International Conference on Ecology and Transportation, Centre for Transportation and the Envronment, North Carolina State University, Raleigh (Caroline du Nord).
Corkran, C.C., comm. pers. 2007. Correspondance personnelle avec B. Beasley. Consultant en matière d’espèces sauvages, Portland (Oregon).
Corkran, C.C. et Thoms, C. 1996. Amphibians of Oregon, Washington, and British Columbia. Lone Pine Publishing, Vancouver, British Columbia.
Danyluk, A., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Janvier 2014. Agent principal en environnement, Corporation de Delta, Delta, British Columbia.
Daszak, P.A., A. Cunningham et A.D. Hyatt. 2003. Infectious disease and amphibian population declines. Diversity and Distributions 9:141-150.
Davidson, C., H.B. Shaffer et M.R. Jennings. 2002. Spatial tests of the pesticide drift, habitat destruction, UV-B, and climate-change hypotheses for California amphibian declines. Conservation Biology 16:1588-1601.
De Solla, S.R., K.E. Pettit, C.A. Bishop, K.M. Cheng et J.E. Elliott. 2002a. Effects of agricultural runoff on native amphibians in the Lower Fraser River Valley, British Columbia, Canada. Environmental Toxicology and Chemistry 21:353-360.
De Solla, S.R., C.A. Bishop, K.E. Pettit et J.E. Elliott. 2002b. Organochloride pesticides and polychlorinated biphenyls (PCBs) in eggs of red-legged frogs (Rana aurora) and northwestern salamanders (Ambystoma gracile) in an agricultural landscape. Chemosphere 46:1027-1032.
Dickman, M. 1968. The effect of grazing by tadpoles on the structure of a periphyton community. Ecology 49:1188–1190.
Dupuis, L. 2011. Summary of Pacific Water Shrew Survey along Roche Creek. Technical Memorandum to the District of North Vancouver and data set. [consulté en novembre 2013].
DWB Consulting Services Ltd. 2010. Wildlife and trapping report. Highway 7 – Nelson Street Intersection Improvement Project. Wildlife Species Inventory Report 4971.
EBA. 2011. Results of Wildlife Salvage during containment pond valve maintenance, Burnaby. Wildlife Species Inventory Report 4779.
EcoDynamic Solutions. 2011. Letter to B.C. Ministry of Environment RE: Amphibian Salvage, Rutherford Road, Nanaimo. Wildlife Species Inventory Report 4773.
Engelstoft, C. et K. Ovaska. 1997. Sharp-tailed Snake inventory within the Coastal Douglas Fir Biogeoclimatic Zone, June – November 1996. Final report. Alula Biological Consulting, Victoria, British Columbia. 36 p. + annexes.
Engelstoft C. et K. Ovaska. 1998. Sharp-tailed Snake study on the Gulf Islands and southeastern Vancouver Island, March – November 1997. Final report. Alula Biological Consulting, Victoria, British Columbia. 46 p + annexes.
Environment Canada. 2011a. Presence and levels of priority pesticides in selected Canadian aquatic ecosystems. Water Science and Technology Directorate. Government of Canada. 102 pp. Fichier PDF [consulté en juillet 2014]. [Également disponible en français : Environnement Canada. 2011a. Présence et concentrations des pesticides prioritaires dans certains écosystèmes aquatiques canadiens. Direction des sciences et de la technologie de l’eau. Gouvernement du Canada. 120 p. Fichier PDF.]
Environment Canada. 2011b. Water quality status and trends of nutrients in major drainage areas of Canada: Technical Summary. Government of Canada. 49 pp. Fichier PDF [consulté en juillet 2014]. [Également disponible en français : Environnement Canada. 2011b. États et tendances de la qualité de l'eau pour les nutriments dans les bassins versants importants du Canada : sommaire technique. Gouvernement du Canada. 59 p. Fichier PDF.]
Fahrig, L., J.H. Pedlar, S.E. Pope, P.D. Taylor et J.F. Wegner. 1995. Effect of road traffic on amphibian density. Biological Conservation 73:177-182.
Finizio, A., T.F. Bidleman et S.Y. Szeto. 1998. Emission of chiral pesticides from an agricultural soil in the Fraser Valley, British Columbia. Chemosphere 36:345–355.
Fraser, D., comm. pers. 2014. Échanges par téléphone avec B. Beasley. Spécialiste des espèces en péril, B.C. Ministry of Environment, Victoria, British Columbia.
Fraser Valley Regional District. 2004. Choices for our future: regional growth strategy for the Fraser Valley Regional District 2004. Fichier PDF [consulté en janvier 2014].
Freshwater Fisheries Society of BC. 2014. Fish Stocking Reports. [consulté en juillet 2014].
Funk, W.C., M.S. Blouin, P.S. Corn, B.A. Maxell, D.S. Pilliod, S. Amishm et F.W. Allendorf. 2005. Population structure of Columbia spotted frogs (Rana luteiventris) is strongly affected by the landscape. Molecular Ecology 14:483-496.
Garcia, P. 2001. Campbell River watershed preliminary amphibian survey. 2001. Rapport inédit préparé pour le BC Hydro Fish and Wildlife Bridge Coastal Compensation Fund. 15 p. et ensemble de données [consulté en novembre 2013].
Gerick, A.A., R.G. Munshaw, W.J. Palen, S.A. Combes et S. O’Regan. 2014. Thermal physiology and species distribution models reveal climate vulnerability of temperate amphibians. Journal of Biogeography 41:713-723.
Gibbs, J.P. 1998. Distribution of woodland amphibians along a forest fragmentation gradient. Landscape Ecology 113:263–268.
Golder Associates Ltd. 2006. Autumn 2006 Pinecrest Amphibian Salvage. Report to Hatfield Consultants Ltd., Vancouver Ouest, British Columbia.
Golder Associates Ltd. 2007. Pinecrest Amphibian Salvage 2007. Report to Hatfield Consultants Ltd., Vancouver Ouest, British Columbia.
Golder Associates Ltd. 2008. Fall Post-salvage Amphibian Monitoring at Pinecrest Sea-to-Sky Highway. Report to Hatfield Consultants Ltd., West Vancouver, British Columbia.
Golder Associates Ltd. 2011a. Pacific Water Shrew and amphibian salvages, Coquitlam: Permit SU11-69099. . Report to B.C. Ministry of Forests, Lands and Natural Resource Operations. 7 p. et ensemble de données [consulté en novembre 2013].
Golder Associates Ltd. 2011b. Reporting requirement for Wildlife Act Permit NA11-71742. Report to B.C. Ministry of Forests, Lands and Natural Resource Operations. Wildlife species inventory report 4746. 6 p.
Golder Associates Ltd. 2012. Reporting requirement for Wildlife Act Permit NA12-76350. Report to B.C. Ministry of Forests, Lands and Natural Resource Operations. Wildlife species inventory report 4883. 5 p.
Google Earth 7.0.3.8542. (February 26, 2013). Knight Inlet to Smith Sound, BC, Canada. 9U 647443 m E 5650501 m N, Eye alt. 57.5 km. Borders and labels; place layers. Image Landsat, 2014 Digital Globe [consulté le 12 juillet 2014].
Govindarajulu, P. 2003. Survey of bullfrogs Rana catesbeiana in British Columbia. http://web.uvic.ca/bullfrogs [consulté en août 2003].
Govindarajulu, P. 2004. Introduced Bullfrogs (Rana catesbeiana) in British Columbia: impacts on native Pacific Treefrogs (Hyla regilla) and Red-legged Frogs (Rana aurora). Thèse de doctorat, University of Victoria, Victoria, Bristish Columbia.
Govindarajulu, P., R. Altwegg et B.R. Anholt. 2005. Matrix model investigation of invasive species control: bullfrogs on Vancouver Island. Ecological Applications 15:2161-2170.
Govindarajulu, P. 2008. Literature review of impacts of glyphosate herbicide on amphibians: what risks can the silvicultural use of this herbicide pose for amphibians in B.C. Wildlife Report No. R-28. B.C. Ministry of Environment, Victoria, British Columbia.
Govindarajulu, P., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Janvier 2014. Small Mammal and Herpetofauna Specialist, B.C. Ministry of Environment, Victoria, British Columbia.
Govindarajulu, P., comm. pers. 2015. Correspondance par courriel adressée à B. Beasley. Janvier 2015. mall Mammal and Herpetofauna Specialist, B.C. Ministry of Environment, Victoria, British Columbia.
Govindarajulu, P., C. Nelson, J. LeBlanc, W. Hintz et H. Schwantje. 2013. Batrachochytrium dendrobatidis surveillance in British Columbia 2008-2009, Canada. British Columbia Ministry of Environment. EcoCat: Ecological Reports Catalogue. http://a100.gov.bc.ca/pub/acat/public/viewReport.do?reportId=34795 [consulté en novembre 2013].
Gray, M.J., D.L. Miller et J.T. Hoverman. 2009. Ecology and pathology of amphibian ranaviruses. Diseases of Aquatic Organisms 87:243-266.
Green, D.E., K.A. Converse et A.K. Schrader. 2002. Epizootiology of sixty-four amphibian morbidity and mortality events in the USA, 1996-2001. Annals of the New York Academy of Sciences 969:323-339.
Green, D.M. 2003. The ecology of extinction: population fluctuation and decline in amphibians. Biological Conservation 11:331–343.
Green, D.M. 1986. Systematics and evolution of western North American Frogs allied to Rana aurora and Rana boylii: electrophoretic evidence. Systematic Zoology 35:283-296.
Green, D.M. (ed.). 2012. Noms français standardisés des amphibiens et des reptiles d’Amérique du Nord au nord du Mexique. SSAR Herpetological Circulars 40. 63 p.
Greer A.L., M. Berrill et P.J. Wilson. 2005. Five amphibian mortality events associated with ranavirus infection in south central Ontario, Canada. Diseases of Aquatic Organisms 67:9–14
Haggard, J.A. 2000. A radio telemetric study of the movement patterns of adult northern red-legged frogs (Rana aurora aurora) at Freshwater Lagoon, Humboldt County, California. Mémoire de maîtrise, Humboldt State University, Arcata, California. 102 p.
Hamilton, P.T., J.M.L. Richardson, P. Govindarajulu et B.R. Anholt. 2012. Higher temperature variability increases the impact of Batrachochytrium dendrobatidis and shifts interspecific interactions in tadpole mesocosms. Ecology and Evolution 2:2450-2459.
Hammerson, G. 2008. Rana aurora. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.2. www.iucnredlist.org [consulté en décembre 2013].
Hawkes, V.C. 2005. Distribution of Red-legged Frog (Rana aurora aurora) breeding habitat in the Jordan River Watershed, Vancouver Island, British Columbia. LGL Project EA1667. Unpublished report by LGL Limited environmental research associates for BC Hydro Fish and Wildlife Bridge Coastal restoration Program, Burnaby BC. iv + 39 p. + annexes.
Hawkes, V.C., comm. pers. 2013. Correspondance par courriel adressée à B. Beasley. Senior Wildlife Biologist, LGL Limited Environmental Research Associates, Victoria, British Columbia.
Haycock, R. D. et D. Knopp. 1998. Amphibian survey with special emphasis on the Oregon spotted frog (Rana pretiosa). Selected wetland sites: Fraser River Lowlands and corridors to the interior Plateau. Draft Report. Ministry of Environment, Lands and Parks. Cité dans B.C. Conservation Data Centre. 2010. Conservation Status Report: Rana aurora. http://a100.gov.bc.ca/pub/eswp/esr.do?id=19586 [consulté en janvier 2014].
Hayes, M.P., comm. pers. 2012. Correspondance par courriel adressée à B. Beasley. Mai 2012. Chercheur scientifique, Washington Department of Fish and Wildlife, Olympia, Washington.
Hayes, M.P. et C.B. Hayes. 2003. Rana aurora aurora: Juvenile growth; Male size at maturity. Herpetological Review 34:233-234.
Hayes, M.P. et M.M. Miyamoto. 1984. Biochemical, behavioral and body size differences between Rana aurora aurora and R. a. draytoni. Copeia 4:1018-1022.
Hayes, M.P., C.A. Pearl et C.J. Rombough. 2001. Rana aurora aurora. Movement. Herpetological Review 32:35-36.
Hayes, M.P., T. Quinn, K.O. Richter, J.P. Schuett-Hames et J.T. Serra Shean. 2008. Maintaining lentic-breeding amphibians in urbanizing landscapes: the case study of the Northern Red-legged Frog (Rana aurora). P. 445-461 in J.C. Mitchell, R.E. Jung Brown et B. Bartholomew (eds.). Urban Herpetology. Herpetological Conservation Vol. 3. Society for the Study of Amphibians and Reptiles. Salt Lake City (Utah).
Hayes, M.P. et C.J. Rombough. 2004. Rana aurora. Predation. Herpetological Review 35:375-376.
Hayes, M.P., C.J. Rombough et C.B. Hayes. 2007. Rana aurora aurora (Northern Red-legged Frog), movement. Herpetological Review 38:192-193.
Hayes, T.B., K. Haston, M. Tsui, A. Hoang, C. Haeffele et A. Vonk. 2003. Atrazine-induced hermaphroditism at 0.1 ppb in American Leopard Frogs (Rana pipiens): laboratory and field evidence. Environmental Health Perspectives 111:568-575.
Hectares BC. 2014. Vancouver Island, Land Use/Cover, Baseline Thematic Mapping. http://www.habc.org/app/habc/HaBC.html [consulté en juin 2014].
Hels, T. et E. Buchwald. 2001. The effect of road kills on amphibian populations. Biological Conservation 99:331-340.
Hillis, D.M. et T.P. Wilcox. 2005. Phylogeny of the New World true frogs (Rana). Molecular Phylogenetics and Evolution 34:299-314.
IUCN, Conservation International, and NatureServe, 2008. Rana aurora. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2014.3. http://www.iucnredlist.org/details/58553/0 [consulté en janvier 2014].
Jancowski, K. et S.A. Orchard. 2013. Stomach contents from invasive American bullfrogs Rana catesbeiana (= Lithobates catesbeianus) on southern Vancouver Island, British Columbia, Canada. NeoBiota 16:17–37.
Jennings, M.R. et M.P. Hayes. 1994. Amphibian and reptile species of special concern in California. Final Report to the California Department of Fish and Game. Rancho Cordova (Californie). 255 p.
Kiesecker, J.M., A.R. Blaustein et C.L. Miller. 2001. Transfer of a pathogen from fish to amphibians. Conservation Biology 15:1064-1070.
Knopp, D., comm. pers. 2013. Correspondance par courriel adressée à B. Beasley. Septembre 2013. BC’s Wild Heritage, Chilliwack, British Columbia.
Leonard, W.P., H.A. Brown, L.L.C. Jones, K.R. McAllister et R.M. Storm. 1993. Amphibians of Washington and Oregon. Seattle Audubon Society, Seattle (État de Washington).
Licht, L.E. 1969. Comparative breeding behavior of the red-legged frog, (Rana aurora aurora) and the western spotted frog (Rana pretiosa pretiosa) in southwestern British Columbia. Canadian Journal of Zoology 47:1287–1299.
Licht, L.E. 1971. Breeding habits and embryonic thermal requirements of the frogs, Rana aurora aurora and Rana pretiosa pretiosa in the Pacific northwest. Ecology 52:116–124.
Licht, L.E. 1974. Survival of embryos, tadpoles, and adults of the frogs Rana aurora aurora and Rana pretiosa sympatric in southwestern British Columbia. Canadian Journal of Zoology 52:613–627.
Licht, L.E. 1986. Food and feeding behavior of sympatric red-legged frogs, Rana aurora and spotted frogs, Rana pretiosa, in southwestern British Columbia. Canadian Field-Naturalist 100:22–31.
Lips, K.R., J. Diffendorfer, J.R. Mendelson III et M.W. Sears. 2008. Riding the wave: reconciling the roles of disease and climate change in amphibian declines. PloS Biol 6:e72.
Loveridge, A.R. 2002. The effects of agricultural effluent on Red-legged Frogs (Rana aurora aurora) and Threespine Stickleback (Gasterosteus aculeatus) in the Elk Creek and Sumas Prairie Watersheds, British Columbia, Canada. Mémoire de maîtrise, Simon Fraser University, Burnaby, British Columbia. ix + 125 p.
Loveridge, A.R., C.A. Bishop, J.E. Elliot et C.J. Kennedy. 2007. Polychlorinated biphenyls and organochlorine pesticides bioaccumulated in Green Frogs, Rana clamitans, from the Lower Fraser Valley, British Columbia, Canada. Bulletin of Environmental Contamination and Toxicology 79:315–318.
Macey, J.R., J.L. Strasburg, J.A. Brisson, V.T. Vredenburg, M. Jennings et A. Larson. 2001. Molecular phylogenetics of western North American Frogs of the Rana boylii Species Group. Molecular Phylogenetics and Evolution 19:131-143.
MacDonald, S.O. 2010. The amphibians and reptiles of Alaska: a field handbook. Version 2.0 – May 2010. Alaska Natural Heritage Program. Fichier PDF.
Malt, J. 2011. Sea to Sky Corridor Amphibian Inventory: Implications for Parks and Species at Risk. January 21, 2011. Rapport inédit. British Columbia Ministry of Forests, Lands and Natural Resource Operations, Surrey, British Columbia.
Malt, J. 2012. Assessing the effectiveness of amphibian mitigation on the Sea to Sky Highway. Population-level effects and best management practices for minimizing highway impacts. Final Report October 2012. 33 p. British Columbia Ministry of Forests, Lands and Natural Resource Operations, Surrey, British Columbia.
Malt, J. 2013. Red-legged Frog (Rana aurora) occupancy monitoring – Metro Vancouver and Fraser Valley. Analyse de données inédite. 11 mars 2013. B.C. Ministry of Forests, Lands and Natural Resource Operations, Surrey, British Columbia.
Mao, J., D.E. Green, G. Fellers et V.G. Chinchar. 1999. Molecular characterization of iridoviruses isolated from sympatric amphibians and fish. Virus Research 63:45-52.
Marco, A., C. Quichano et A.R. Blaustein. 1999. Sensitivity to nitrate and nitrite in pond-breeding amphibians from the Pacific Northwest, USA. Environmental Toxicology & Chemistry 18:2836-2839.
Marsh, D.M. et P.C. Trenham. 2001. Metapopulation dynamics and amphibian conservation. Conservation Biology 15:40-49.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay et A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk. NatureServe, Arlington, Virginia. 57 p.
Materi, J. et C. Forrest. 2004. Salmon River diversion wildlife overpass pilot project (BCRP Project No.01.W.08). Final Report. Ursus Environmental, Nanaimo, British Columbia. EcoCat: Ecological Reports Catalogue [consulté en novembre 2013].
Materi, J. 2008. Progress Report: Rupert Road post-construction wetland and amphibian monitoring in 2008. Prepared by Ursus Environmental, Parksville, British Columbia. for the Town of Qualicum Beach, Ursus Environmental, Nanaimo, British Columbia.
Matsuda, B., D. Green et P. Gregory. 2006. Amphibians and Reptiles of British Columbia. Royal British Columbia Museum Handbook. 266 p.
McConkey, D. Données inédites. 2008. British Columbia Ministry of Environment 2006-2008 Vancouver Island Red-legged Frog Inventory and Forestry Management Recommendations for Red-legged Frogs. British Columbia Ministry of Environment Ecosystems Branch, Nanaimo, British Columbia.
McConkey, D., comm. pers. 2015. Correspondance par courriel adressée à B. Beasley. Janvier 2015. Senior Ecosystem Biologist, British Columbia Ministry of Forests, Lands and Natural Resource Operations, Nanaimo, British Columbia.
McCurdy, A., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Juillet 2014. Biologiste, Ucluelet Aquarium, Ucluelet, British Columbia.
McTaggart Cowan, I. 1941. Longevity of the red-legged frog. Copeia 1941:48.
Meggill, W., comm. pers. 2003. Correspondance adressée à K. Ovaska. Août 2003. Adjoint de recherche, Department of Zoology, University of British Columbia, Vancouver, British Columbia.
Metro Vancouver. 2014. Vancouver 2040: Shaping Our Future. Regional Growth Strategy Bylaw No. 1136, 2010. Fichier PDF [consulté en janvier 2015].
Mitchell, A.M. 2013. Identify, conserve and restore populations of priority species at risk and their associated habitats within the Alouette River Watershed. Final Report 2012-2013. FWCP Report No. 12.W.ALU.01. April 2013. 19 p. et ensemble données [consulté en novembre 2013].
Mitchell, A., V. Kilburn et C. Currie. 2012. Red-legged Frog (Rana aurora) report 2012. Sunshine Coast / Lower Mainland Survey Results. The South Coast Western Painted Turtle Recovery Project. Vancouver, British Columbia.
Mitchell, A., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Janvier 2014. Biologiste, Western Painted Turtle Project, Vancouver, British Columbia.
Murray, R., V. Popescu, W. Palen et P. Govindarajulu. Sous presse. Relative performance of ecological niche and occupancy models for predicting invasions by patchily-distributed species. Biological Invasions.
NatureServe. 2013. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. http://www.natureserve.org/explorer [consulté en décembre 2013].
Nebeker, A.V. et G.S. Schuytema. 2000. Effects of ammonium sulfate on the growth of larval northwestern salamanders, red-legged and Pacific treefrog tadpoles, and juvenile fathead minnows. Bulletin of Environmental Contamination and Toxicology 64:271-278.
Nieto, N.C., M.A.Camann, J.E. Foley et J.O. Reiss. 2007. Disease associated with integumentary and cloacal parasites in tadpoles of northern red-legged frog Rana aurora aurora. Diseases of Aquatic Organisms 78:61-71.
Nussbaum, R.A., E.D. Brodie, Jr. et R.M Storm. 1983. Amphibians & Reptiles of the Pacific Northwest. Northwestern University Press of Idaho, Moscow. 332 p.
O’Regan, S.M., W.J. Palen et S.C. Anderson. 2014. Climate warming mediates negative impacts of rapid pond drying for three amphibian species. Ecology 95:845-855.
Ostergaard, E.C. et K.O. Richter. 2001. Stormwater ponds as surrogate wetlands for assessing amphibians as bioindicators. Fichier PDF [consulté en août 2003].
Ostergaard, E.C., K.O. Richter et S.D. West. 2008. Amphibian use of stormwater ponds in the Puget Lowlands. Pp. 259-270 in J.C. Mitchell, R.E. Jung Brown et B. Bartholomew (eds.) Urban Herpetology. Herpetological Conservation Vol. 3. Society for the Study of Amphibians and Reptiles. Salt Lake City (Utah).
Ovaska, K., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Juin 2014. Écologiste, Biolinx Environmental Research Ltd., Victoria, British Columbia.
Ovaska, K., T.M. Davis et I.N. Flamarique. 1997. Hatching success and larval survival of the frogs Hyla regilla and Rana aurora under ambient and artificially enhanced solar ultraviolet radiation. Canadian Journal of Zoology 75:1081-1088.
Ovaska, K., L. Hyatt et L. Sopuck. 2002. Rana aurora. Geographic distribution. Herpetological Review 33:318.
Ovaska, K., L. Sopuck, C. Engelstoft, L. Matthias, E. Wind et J. MacGarvie. 2003. Best Management Practices for Amphibians and Reptiles in Urban and Rural Environments in British Columbia. Prepared for BC Ministry of Water, Land and Air Protection, Nanaimo, British Columbia. Fichier PDF [consulté en juillet 2014].
Packham, R., comm. pers. 2013. Échanges par téléphone avec B. Beasley. Décembre 2013. Biologiste à la retraite, B.C. Ministry of Environment, Région de Cariboo.
Pearl, C.A. 2005. Rana aurora (Baird and Girard, 1852[b]): Northern Red-legged Frog. Pp. 528-530 in M. Lannoo (ed.). Amphibian Declines: The Conservation Status of United States Species. University of California Press, Berkeley, California.
Pearl, C.A., M.J. Adams, N. Leuthold et R.B. Bury. 2005. Amphibian occurrence and aquatic invaders in a changing landscape: implications for wetland mitigation in the Willamette Valley, Oregon, U.S. Wetlands 25:76-88.
Pearl, C.A., E.L. Bull, D.E. Green, J. Bowerman, M.J. Adams, A. Hyatt et W.H. Wente. 2007. Occurrence of the amphibian pathogen Batrachochytrium dendrobatidis in the Pacific Northwest. Journal of Herpetology 41:145-149.
Pearson, M. 2010. Inventory and long-term monitoring of Oregon Spotted Frog and Red-legged Frog egg masses in the Fraser Valley Lowlands. Spring 2010. Unpublished report prepared for the B.C. Ministry of Environment, Oregon Spotted Frog Recovery Team, South Coast Conservation Program, and B.C. Conservation Program. Balance Ecological, Vancouver, British Columbia. 36 p.
Pearson, M. 2011. Inventory and long-term monitoring of Oregon Spotted Frog and Red-legged Frog egg masses in the Fraser Valley Lowlands. Spring 2011. Unpublished report prepared for the B.C. Ministry of Environment, Oregon Spotted Frog Recovery Team, South Coast Conservation Program, and B.C. Conservation Program. Balance Ecological, Vancouver, British Columbia. 18 p.
Pearson, M., comm. pers. 2011. Échanges par téléphone avec B. Beasley. Novembre 2011.Biologiste, Balance Ecological Ltd., Vancouver, British Columbia.
Pearson, M. 2012. Inventory and long-term monitoring of Oregon Spotted Frog and Red-legged Frog egg masses in the Fraser Valley Lowlands. Spring 2012. Unpublished report prepared for the B.C. Ministry of Environment, Oregon Spotted Frog Recovery Team, South Coast Conservation Program, and B.C. Conservation Program. Balance Ecological, Vancouver, British Columbia. 18 p.
Pechmann, J.H.K. et H.M. Wilbur 1994. Putting declining amphibian populations in perspective: natural fluctuations and human impacts. Herpetologica 50:65–84.
Powelson, A., comm. pers. 2013. Correspondance par courriel adressée à B. Beasley. Décembre 2013. Resource Investment Officer, Ministry of Forests, Lands and Natural Resource Operations, Victoria, British Columbia.
Raffel. T.R., J.R. Rohr, J.M. Kiesecker et P.J. Hudson. 2006. Negative effects of changing temperature on amphibian immunity under field conditions. Functional Ecology 20:819-828.
Rahel, F.J. et J.D. Olden. 2008. Assessing the effects of climate change on aquatic invasive species. Conservation Biology 22:521-533.
Raina, R., P. Hall et L. Sun. 2010. Occurrence and relationship of organophosphorus insecticides and their degradation products in the atmosphere in western Canada agricultural regions. Environmental Science and Technology 44:8541-8546.
Reh, W. 1989. Investigations into the influences of roads on the genetic structure of populations of the common frog Rana temporaria. Pp. 101-103 in Langton, T.E.S. (ed.). Amphibians and Roads. Proceedings of the Toad Tunnel Conference, Rendsburg, Federal Republic of Germany, 7-8 January 1989. 202 p.
Reimchen, T.E. 1991. Introduction and dispersal of the Pacific treefrog, Hyla regilla, on the Queen Charlotte Islands, British Columbia. The Canadian Field-Naturalist 105:288–290.
Reynolds, J., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Janvier 2014. Professeur, Tom Buell BC Leadership Chair in Aquatic Conservation, Simon Fraser University, Burnaby, British Columbia.
Richardson, J., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Janvier 2014. Professeur, Department of Forest and Conservation Sciences, University of British Columbia, Vancouver, British Columbia.
Richardson, J.M.L., P. Govindarajulu et B.R. Anholt. 2014. Distribution of the disease pathogen Batrachochytrium dendrobatidis in non-epidemic amphibian communities of western Canada. Ecography 37:883-893.
Richter, K.O. et A.L. Azous. 1995. Amphibian occurrence and wetland characteristics in the Puget Sound Basin. Wetlands 15:306-312.
Rithaler, R.C. 2002a. Fish and amphibian spawning timing in Delta, British Columbia. Environmental Services Scientific/Technical Series. The Corporation of Delta,, British Columbia.
Rithaler, R.C. 2002b. Delta Watersheds: Fish and Amphibian distributions. (1:25,000 map; created 13 October 2001, amended 28 January 2002). Disponible auprès de la Corporation de Delta, 4500, cr. Clarence Taylor, Delta, British Columbia, V4K 3E2.
Rithaler, R.C. 2003a. Amphibians in Delta. (1:90,000 map; created 12 August 2002, amended 18 August 2003). Disponible auprès de la Corporation de Delta, 4500, cr. Clarence Taylor, Delta, British Columbia, V4K 3E2.
Rithaler, R.C. 2003b. Delta Timing Schedules for In-stream Works. (1:90,000 map). Disponible auprès de la Corporation de Delta, 4500, cr. Clarence Taylor, Delta,, British Columbia, V4K 3E2.
Rithaler, R.C., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Février 2014. Biologiste et ancien agent en environnement, Corporation de Delta, British Columbia.
Ritson, P.I. et M.P. Hayes. 2000. Late season activity and overwintering in the northern red-legged frog (Rana aurora aurora). Final report to the U.S. Fish and Wildlife Service, Portland, OR. 21 pp. Cité dans Hayes, M.P., T. Quinn, K.O. Richter, J.P. Schuett-Hames et J.T. Serra Shean. 2008. Maintaining lentic-breeding amphibians in urbanizing landscapes: the case study of the Northern Red-legged Frog (Rana aurora). Pp. 445-461 in J.C. Mitchell, R.E. Jung Brown et B. Bartholomew (eds.) Urban Herpetology. Herpetological Conservation Vol. 3. Society for the Study of Amphibians and Reptiles. Salt Lake City (Utah).
Robertson Environmental 2013. MK Delta Lands 10770-72nd Avenue, Highway 91 West, and adjacent lands. Baseline bio-inventory report 2008, 2009, and 2012 in support of rezoning application. Final Report. January 2013. Robertson Environmental Services Limited, Langley, British Columbia. 140 p.
Rohr, J.R. et T.R. Raffel. 2010. Linking global climate and temperature variability to widespread amphibian declines putatively caused by disease. Proceedings of the National Academy of Sciences. USA 107:8269-8274.
Salafsky, N., D. Salzer, A.J. Stattersfield, C. Hilton-Taylor, R. Neugarten, S.H.M. Butchart, B. Collen, N. Cox, L.L. Master, S. O’Connor et D. Wilkie. 2008. A standard lexicon for biodiversity conservation: unified classifications of threats and actions. Conservation Biology 22:897-911.
Schindler, D.W., P.J. Dillon et H. Schreier. 2006. A review of anthropogenic sources of nitrogen and their effects on Canadian aquatic ecosystems. Biogeochemistry 79:25-44.
Schmutzer, A.C., M.J. Gray, E.C. Burton et D.L. Miller. 2008. Impacts of cattle on amphibian larvae and the aquatic environment. Freshwater Biology 53:2613-2625.
Schuett-Hames, J.P. 2004. Northern red-legged frog (Rana aurora aurora) terrestrial habitat use in the Puget Low-lands of Washington. Mémoire de maîtrise, Evergreen State College, Olympia, Washington. 119 p.
Schuytema, G.S. et A.V. Nebeker. 1998. Comparative toxicity of diuron on survival and growth of Pacific treefrog, bullfrog, red-legged frog, and African clawed frog embryos and tadpoles. Archives of Environmental Contamination and Toxicology 34:370-376.
Schuytema, G.S. et A.V. Nebeker. 1999. Effects of ammonium nitrate, sodium nitrate and urea on red-legged frogs, Pacific treefrogs and African clawed frogs. Bulletin of Environmental Contamination and Toxicology 63:357-364.
Serra Shean, J.T.S. 2002. Post-breeding movements and habitat use by the Northern Red-legged Frog, Rana aurora aurora, at Dempsey Creek, Thurston County, Washington. Mémoire de maîtrise ès sciences. Evergreen State College, Olympia, Washington. 89 p.
Shaffer, H.B., G.M. Fellers, S.R. Voss, J.C. Oliver et G.B. Pauly. 2004. Species boundaries, phylogeography and conservation genetics of the red-legged frog (Rana aurora/draytonii) complex. Molecular Ecology 13:2667-2677.
Sierra Club, 2003. Vancouver Island : How much has been logged? [consulté en septembre 2003].
Silvestri, S., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Juillet 2014. Fisheries Biologist, Fish and Wildlife Branch, Ministry of Forests, Lands and Natural Resource Operations, Nanaimo, British Columbia.
Snodgrass, J.W., R.E. Casey, D. Joseph et J.A. Simon. 2008. Microcosm investigations of stormwater pond sediment toxicity to embryonic and larval amphibians: variation in sensitivity among species. Environmental Pollution 154:291-297.
Sowden, P., comm. pers. 2011. Correspondance par courriel adressée à B. Beasley. Novembre 2011. Vétérinaire à la retraite et naturaliste, Sayward, British Columbia.
Squamish-Lillooet Regional District. 2008. Regional Growth Strategy BYLAW NO. 1062, 2008. Schedule “A”. Fichier PDF [consulté en janvier 2014].
Stanley Park Ecology Society. 2010. State of the park report for the ecological integrity of Stanley Park. Stanley Park Ecology Society, Vancouver, British Columbia.
Statistics Canada. 2009. Ecoregion profile: Lower Mainland of British Columbia. http://www.statcan.gc.ca/pub/16-002-x/2009004/article/11031-eng.htm [consulté en décembre 2013 ]. [Également disponible en français : Statistique Canada. 2009. Profil d’écorégion : basses-terres continentales de la Colombie-Britannique. http://www.statcan.gc.ca/pub/16-002-x/2009004/article/11031-fra.htm.]
Storm, R.M. 1960. Notes on the breeding biology of the red-legged frog (Rana aurora aurora. Herpetologica 16:251–259.
Sunshine Coast Regional District. 2003. Sunshine Coast Habitat Atlas; forest cover map. Fichier PDF [consulté en juillet 2014].
Tayless, E. 2012. Resort Municipality of Whistler Wildlife Permit SU12-79065 Amphibian Sampling Data Report. Resort Municipality of Whistler, Whistler, British Columbia. 10 p.
Top, V. 1996. Dissipation and mobility of atrazine, simazine, diazinon and chlorphyrifos in the Ryder soil of the Lower Fraser Valley, British Columbia. Mémoire de maîtrise, University of British Columbia, Vancouver, British Columbia. 126 p. + annexes.
Tuttle, K.N. 2013. Continued monitoring of the constructed wetland at Diversion Reservoir, Jordon River Watershed, Southern Vancouver Island. LGL Report EA3285. Unpublished report by LGL Limited Environmental Research Associates, Sidney, B.C., for BC Hydro, Fish and Wildlife Compensation Program (Coastal), British Columbia. 38 p. + annexes.
Urban Futures. 2007. Population and housing change in the Nanaimo Region, 2006 to 2036. Fichier PDF [consulté en décembre 2013].
van Kooten, G.C. 1995. Modeling public forest land use tradeoffs on Vancouver Island. Journal of Forest Economics 1: 191-218. Fichier PDF [consulté en octobre 2003].
Vitt, L.J., Wilbur, H.M., Caldwell, J.P. et Smith, D.C. 1990. Amphibians as harbingers of decay. BioScience 40:418.
Vos, C.C., A.G. Antonisse-De Jong, P.W. Goedhart et M.J.M. Smulders. 2001. Genetic similarity as a measure for connectivity between fragmented populations of the moor frog (Rana arvalis). Heredity 86:598-608.
Walls, S.C., W.J. Barichivich et M.E. Brown. 2013. Drought, deluge and declines: the impact of precipitation extremes on amphibians in a changing climate. Biology 2:399-418.
Wang, T., E.M. Campbell, G.A. O’Neill et S.N. Aitken. 2012. Projecting future distributions of ecosystem climate niches: Uncertainties and management applications. Forest Ecology and Management 279:128-140.
Waye, H. 1999. COSEWIC Status Report on the Northern Red-Legged Frog, Rana aurora. Committee on the Status of Endangered Wildlife in Canada. 28 p.
Wells, K.D. 1977. The social behaviour of anuran amphibians. Animal Behaviour 25:666–693.
Wind, E. 2003. Aquatic-breeding amphibian monitoring program. Analysis of small wetland habitats on Vancouver Island. Annual Progress Report 2002. Rapport inédit préparé pour le Weyerhaueser BC Coastal Group, Nanaimo, British Columbia.
Wind, E. 2005. Effects of nonnative predators on aquatic ecosystems. Rapport inédit préparé pour le Ministry of Water, Land and Air Protection, Victoria, British Columbia.
Wind, E. 2008a. Pre- and post-harvest amphibian and small wetland study. Year 4. Rapport inédit prépapré pour Western Forest Products Inc. et Island Timberlands Limited Partnership.
Wind, E. 2008b. ECVI Storage Feasibility Project. Environmental Assessment Component for Shelton and Healy Lakes. E. Wind Consulting. Nanaimo, British Columbia.
Wind, E. 2012. Amphibian road surveys and mitigation assessments at three sites on Vancouver Island. Rapport inédit préparé pour l’Environmental Management Section, Ministry of Transportation and Infrastructure, Victoria, B.C. E. Wind Consulting, Nanaimo, British Columbia.
Wind, E. et G. Dunsworth. 2006. Does group retention harvesting protect amphibian habitat? Extended Abstract presented at the Variable Retention Forestry Science Forum, April 21-22, 2004. BC Journal of Ecosystems and Management 7:13-16.
Wojtaszek, G., comm. pers. 2013. Correspondance par courriel adressée à B. Beasley. Septembre 2013. Chef de l’équipe des écosystèmes, Réserve de parc national Gwaii Haanas, Skidegate, British Columbia.
Worcester, R., comm. pers. 2014. Correspondance par courriel adressée à B. Beasley. Janvier 2014. Conservation Programs Manager, Stanley Park Ecology Society, Vancouver, British Columbia.
Barbara Beasley détient un doctorat sur les stratégies d'accouplement des hirondelles obtenu à l'Université Simon Fraser (Simon Fraser University) en 1994. Elle travaille à titre de consultante indépendante en biologie et d'instructrice à temps partiel au Bamfield Marine Sciences Centre et à l'Université Quest (Quest University). Mme Beasley a coordonné un projet d'inventaire de trois ans, de 1997 à 2000, visant à évaluer l'abondance relative des amphibiens dans des forêts d'âge divers dans le cadre du processus de planification de l'utilisation des terres de la Colombie-Britannique pour la baie Clayoquot. Ses recherches à long terme portent sur la documentation de la mortalité routière des amphibiens, y compris la grenouille à pattes rouges du Nord, et l'analyse de l'efficacité des mesures d'atténuation. Mme Beasley a récemment fondé l'Association of Wetland Stewards for Clayoquot and Barkley Sounds, organisme sans but lucratif faisant la promotion de la conservation des amphibiens par la recherche, la surveillance et la sensibilisation du public sur la côte ouest de l'île de Vancouver.
Aucun spécimen n'a été examiné, mais des données ont été tirées de dossiers détenus dans les établissements suivants et consultés le 17 novembre 2013 par le biais du portail de données HerpNET (http://www.herpnet.org).
University of Alberta Museum of Zoology
University of Colorado Museum
Royal Ontario Museum, Toronto, Ontario
American Museum of Natural History, New York
University of Louisiana at Monroe
University of British Columbia Beaty Biodiversity Museum
Museum of Vertebrate Zoology, University of California, Berkeley
California Academy of Sciences, San Francisco
Smithsonian National Museum of Natural History, Washington, D.C.
Michèle Steigerwald a fourni les mentions du Musée canadien de la nature. Kristiina Ovaska a vérifié deux mentions douteuses du Royal British Columbia Museum et confirmé que les spécimens associés à ces mentions étaient des Rana lutreiventris et non des Rana aurora.
Carte indiquant l'occupation des sites de reproduction de la grenouille à pattes rouges du Nord dans les milieux humides de la grande région métropolitaine de Vancouver et de la vallée du Fraser au printemps 2012, de même que la couverture terrestre.
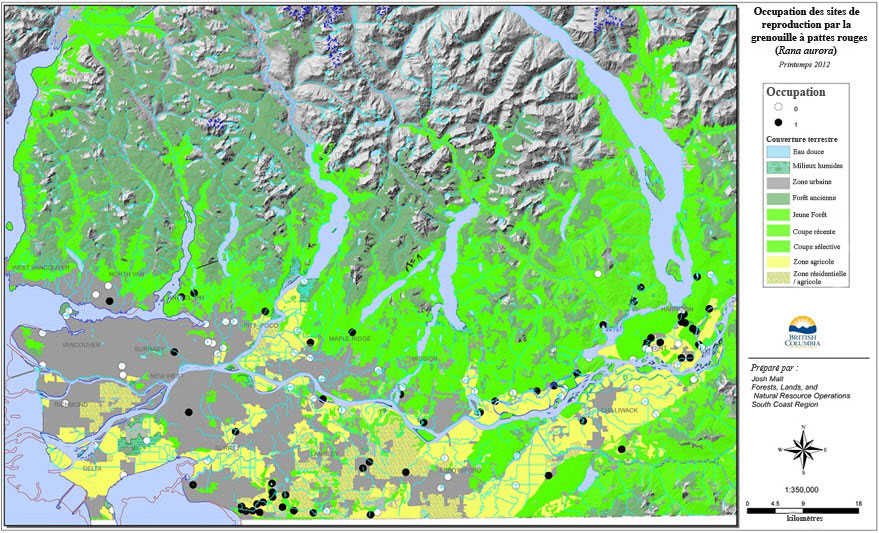
Source: Malt, données inédites, 2013; figure utilisée avec permission
Longue description pour l'annexe 1
Carte indiquant l'occupation des sites de reproduction de la grenouille à pattes rouges du Nord dans les milieux humides de la grande région métropolitaine de Vancouver et de la vallée du Fraser au printemps 2012, de même que la couverture terrestre.