
Isoète prototype (Isoetes prototypus): évaluation et rapport de situation du COSEPAC: chapitre 6
Biologie
Cycle vital et reproduction
On ne sait à peu près rien des différents stades du cycle vital de l’Isoetes prototypus, ni sur les besoins de chaque stade. La figure 9 illustre le cycle vital général du genre Isoetes.
Il ne semble y avoir aucune publication sur la reproduction ou le cycle vital de l’Isoetes prototypus. Selon W. Carl Taylor (comm. pers., 2003), il existe très peu d’informations sur la reproduction et le cycle vital du genre Isoetes en général.
D’après Caplen et Werth (2000), « il n’existe aucune information sur l’histoire naturelle de la reproduction sexuée des Isoetes » [traduction], mais des observations directes de la reproduction sexuée ont déjà été faites in vitro.
On ignore pratiquement tout des facteurs qui ont une incidence sur la survie de l’Isoetes prototypus, la structure d’âge et la stabilité de ses populations ainsi que ses taux de reproduction et de mortalité. Les feuilles de la plante sont persistantes, et on suppose que la plante est vivace, mais les observations in situ ont toutes été réalisées en été et au début de l’automne.
L’étude de Cobb (1963) constitue une des descriptions les plus complètes du cycle vital et de la reproduction des plantes du genre Isoetes. La plante
communément appelée « isoète » est en fait le sporophyte, qui produit deux types de spores : de grandes spores femelles, les mégaspores, et de petites spores mâles, les microspores. Ces spores sont portées par des sporanges situés dans une cavité de la base en cuillère de certaines feuilles (généralement surtout les feuilles médianes et périphériques). La plante produit généralement les deux types de spores, mais les mégaspores et les microspores sont habituellement logées dans des sporanges distincts; chez certaines espèces, les deux types de spores peuvent se trouver dans le même sporange. Jusqu’à 100 mégaspores, ou parfois plus, peuvent se développer dans un même sporange, alors que le nombre de microspores par sporange peut atteindre plusieurs centaines de milliers. En général, les spores arrivent à maturité vers la fin de l’été et sont libérées lorsque les tissus du sporange se dégradent ou se brisent.
Figure 9. Cycle vital des Isoetes (tiré de Stern, 1985, avec la permission de McGraw-Hill Education).
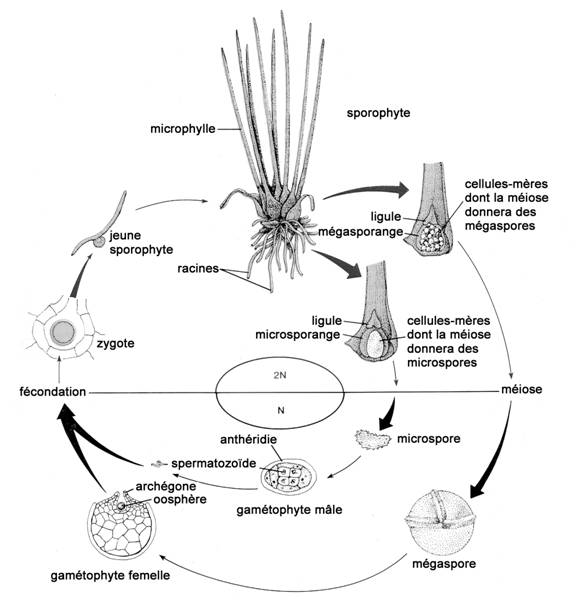
Les microspores et les mégaspores donnent naissance à des gamétophytes (ou prothalles) mâles et femelles distincts. Le gamétophyte mâle produit généralement quatre minuscules spermatozoïdes motiles et allongés portant quatre cils, soit deux à chaque extrémité. Le gamétophyte femelle est rond et multicellulaire et porte des oosphères (gamètes femelles) sur sa face dorsale. Chaque oosphère est située à la base d’une petite structure en forme d’entonnoir, l’archégone, qui s’ouvre lorsque l’oosphère est prête à être fécondée et se referme après la fécondation. Le jeune sporophyte se développe directement à partir d’une oosphère fécondée (zygote).
On estime que la reproduction des Isoetes est presque exclusivement sexuée, en raison de leur nature hétérosporée. Cependant, Caplen et Werth (2000) ont avancé que la grande proximité des mégasporanges et des microsporanges, associée à une libération graduelle des spores par dégradation du tissu sporangial, pourrait permettre un taux appréciable de fécondation entre gamétophytes issus d’un même sporophyte, ce qui équivaut à l’autopollinisation observée chez certaines plantes phanérogames.
Bien que de nombreux hybrides d’Isoetes aient été signalés récemment dans les publications scientifiques et que certains résultats préliminaires semblent indiquer que l’I. prototypus serait un des ancêtres de l’I. tuckermanii, de l’I. acadiensis et de l’I. lacustris (Taylor, comm. pers., 2003), aucun hybride dont un des parents serait l’I. prototypus n’a été observé à ce jour.
Herbivores
On ignore si l’espèce est une source de nourriture pour des animaux, mais on a observé des Canards colverts broutant d’autres espèces d’Isoetes (Brunton et Britton, 1999).
Physiologie
Les besoins physiologiques de l’I. prototypus sont peu connus, outre l’information concernant son habitat contenue dans le présent rapport. Aucune étude exhaustive de l’écologie de l’espèce n’a été entreprise.
Déplacements et dispersion
Les populations des espèces aquatiques du genre Isoetes sont souvent isolées les unes des autres, puisque les cours d’eau et les plans d’eau qu’elles occupent ne sont pas interreliés (Caplen et Werth, 2000). On n’a évidemment jamais observé la dispersion de propagules d’Isoetes sur le terrain, mais il est possible que celles-ci soient transportées par le courant des cours d’eau ou par des animaux comme le castor, le rat musqué ou la sauvagine (Caplen et Werth, 2000; Brunton et Britton, 1999), voire l’orignal. Par exemple, Brunton et Britton (1999) ont observé des Canards colverts broutant d’autres espèces du genre Isoetes et des Plongeons huards déracinant des Isoetes et s’en nourrissant peut-être. Brunton (comm. pers, 2004) avance en outre que les poissons et les tortues seraient des candidats au moins aussi probables comme causes de perturbation et comme vecteurs de dispersion.
Il semble exister très peu d’information sur la dispersion de l’Isoetes prototypus.
Cependant, dans de nombreux lacs, la présence de l’espèce a été révélée par celle d’individus déracinés flottant près des rives ou échoués sur la berge. Brunton et Britton (1993) ont avancé que ces plantes pourraient être déracinées par le passage de bateaux à moteur, les lignes à pêche, les ancres, les baigneurs, la sauvagine se nourrissant au fond, les rats musqués, les reptiles et les amphibiens en hibernation et/ou les poissons en fraie. D.L. Sabine (comm. pers., 2004) estime que les orignaux pourraient jouer un rôle dans le déracinement d’individus d’I. prototypus, puisqu’on trouve des sentiers d’orignal bien marqués jusqu’à une profondeur de 1,5 à 2,0 m dans certains lacs du Nouveau-Brunswick où l’I. prototypus est présent (le site 12, par exemple). Brunton (comm. pers., 2002) a également avancé que la formation de poches gazeuses et/ou l’inversion saisonnière des eaux pourrait être responsable du déracinement d’isoètes. Au lac Holland, des individus d’I. prototypus peuvent être aisément déracinés par les courants que génèrent les palmes de plongée, et on trouve parfois des tapis partiellement déracinés aux abords de sources subaquatiques. Certains des individus déracinés flottant à la surface n’ont plus de corme ni de racines mais portent encore souvent des microspores et des macrospores (Goltz, observation personnelle). Il est possible que la dispersion par le vent et le courant des individus flottant à la surface se solde par le dépôt de spores et la formation de nouvelles colonies dans les lacs abritant déjà l’I. prototypus.
Relations interspécifiques
L’Isoetes prototypus semble confiné aux lacs oligotrophes, où il pousse généralement dans des sédiments mous, floculeux et vaseux.
Nous avons pu confirmer que l’I. prototypus peut former de grands peuplements monospécifiques, mais pousse souvent parmi des peuplements d’autres espèces et d’hybrides d’Isoetes, en particulier l’I. lacustris et l’I. x harveyi, ou parmi de denses tapis d’Eriocaulon aquaticum. Aucun hybride dont un des parents serait l’I. prototypus n’a été découvert à ce jour, y compris lors de nos travaux de terrain des étés 2003 et 2004. Le meilleur moyen de détecter de tels hybrides serait de compter les chromosomes, mais très peu d’individus vivants ont été récoltés à cette fin. Il est cependant fort probable que l’espèce s’hybride, ou du moins qu’elle le faisait dans le passé, puisque les analyses d’ADN effectuées par W. Carl Taylor semblent indiquer que le génome de l’I. prototypus aurait contribué à celui de l’I. lacustris et peut-être à ceux de l’I. acadiensis et de l’I. tuckermanii.
Adaptabilité
On ignore tout de la capacité de l’Isoetes prototypus de s’adapter aux perturbations ou aux changements et de sa tolérance à une détérioration de son habitat. La plante pousse généralement à des profondeurs supérieures à 1,5 m, dans des lacs frais, oligotrophes et alimentés par des sources; on présume donc que la pollution des eaux, l’eutrophisation, la baisse du niveau de l’eau et les fluctuations de la température de l’eau pourraient nuire à l’espèce.