
Évaluation et Rapport de situation du COSEPAC sur le Leptoge des terrains inondés (Leptogium rivulare) au Canada - 2015

- Table des matières
- COSEPAC - sommaire de l'évaluation
- COSEPAC - résumé
- Résumé technique
- Préface
- Historique du COSEPAC
- Mandat du COSEPAC
- Composition du COSEPAC
- Définitions (2015)
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Facteurs limitatifs
- Menaces
- Espèces exotiques/non indigènes envahissantes
- Changement climatique et sécheresses
- Gestion et utilisation de l'eau et exploitation de barrages
- Exploitation forestière et récolte du bois
- Effluents agricoles et sylvicoles
- Développement résidentiel et commercial
- Exploitation de mines et de carrières
- Polluants atmosphériques
- Nombre de localités du COSEPAC
- Protection, statuts et classements
- Remerciements
- Experts contactés
- Sources d'information
- Sommaire biographique des rédacteurs du rapport
- Collections examinées
- Figure 1. Deux thalles de Leptogiumn rivulare poussant sur l'écorce d'un arbre.
- Figure 2. Répartition mondiale du Leptogiumn rivulare, délimitée d'après les frontières politiques ou territoriales importantes. Les polygones en gris foncé représentent un pays, une région, un État, une province ou un comté où la présence du L. rivulare a été confirmée.
- Figure 3. Répartition connue du Leptogiumn rivulare en Amérique du Nord, sur une carte indiquant les aires écologiques du COSEPAC. Aux États-Unis, on trouve des occurrences au Wisconsin (WI), en Illinois (IL) et au Vermont (VT), mais l'espèce est présumée disparue dans ces deux derniers États.
- Figure 4. Lieux de récolte de spécimens de lichens en Ontario, au Québec et au Manitoba, d'après les données du Consortium of North American Lichen Herbaria. Les données ont été consignées pour les spécimens récoltés du 19e siècle jusqu'à aujourd'hui. La carte a été produite en septembre 2015.
- Figure 5. Mare printanière hébergeant le Leptogiumn rivulare dans le centre de l'Ontario (sous-population des basses terres de l'Ontario).
- Figure 6. Habitat riverain de la sous-population de Leptogiumn rivulare du lac postglaciaire Agassiz, en bordure de la rivière Attawapiskat, au Manitoba, où on peut voir des pierres éparses.
- Figure 7. Répartition du L. rivulare au Canada, divisée en trois groupes :(1) basses terres de l'Ontario, (2) Sud du Bouclier (3), zone anciennement recouverte par les lacs glaciaires Agassis et Ojibway. Il est à signaler que le groupe du Sud du Bouclier compte des occurrences isolées de L. rivulare à Wawa, à l'île Cockburn, à Temagemi et à Algonquin. Les secteurs où se sont concentrées les activités de recherche visant le L. rivulare (cercles hachurés) sont également indiqués sur la carte, en plus de l'occurrence des États-Unis qui se trouve la plus près des occurrences canadiennes, située au Wisconsin. Enfin, la carte montre la superficie approximative occupée par le lac postglaciaire Agassiz et sa combinaison possible avec le lac postglaciaire Ojibway (adaptée de Teller et al., 2002); les flèches grises indiquent les principales voies de drainage des lacs postglaciaires par rapport aux occurrences nordiques isolées de L. rivulare.
- Figure 8. Propagation annuelle de l'agrile du frêne (EAB) dans le sud de l'Ontario (à partir de Windsor et d'Ottawa), en fonction des peuplements de frênes présentant un taux de mortalité modéré à élevé. Les étoiles bleues indiquent les emplacements où l'agrile du frêne est présent mais où le taux de mortalité est inconnu. Les régions où l'agrile du frêne a endommagé et tué des peuplements de frênes sont indiquées par des points dont la couleur représente l'année de l'infestation : les points verts représentent le début de l'infestation, les points jaunes et orange, la période subséquente, et les points rouges, la vague actuelle d'infestation; les étoiles bleues indiquent les signalements les plus récents.
- Tableau 1. Sommaire des occurrences de la sous-population des basses terres de l'Ontario, du nombre d'individus qu'elles comptent et des menaces les plus imminentes qui pèsent sur elles. Le frêne est présent dans 29 occurrences et dominant dans 10 occurrences.
- Tableau 2. Sommaire des occurrences de la sous-population du Sud du Bouclier, du nombre d'individus qu'elles comptent et des menaces les plus imminentes qui pèsent sur elles. *Il est à signaler que l'occurrence de Wawa est disparue. Le frêne est présent dans 32 des 35 occurrences existantes, dominant dans 10 occurrences et codominant dans 7 autres occurrences.
- Tableau 3. Sommaire des occurrences de la sous-population du bassin du lac postglaciaire Agassiz, du nombre d’individus qu’elles comptent et des menaces les plus imminentes qui pèsent sur elles.
- Annexe 1. Tableau d'évaluation des menaces pour leleptogium rivulare
COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada

COSEPAC
Comité sur la situation
des espèces en péril
au Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2015. Évaluation et Rapport de situation du COSEPAC sur le leptoge des terrains inondés (Leptogium rivulare) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xiii + 54 p. (Registre public des espèces en péril).
Rapport(s) précédent(s) :
COSEPAC. 2004. Évaluation et Rapport de situation du COSEPAC sur le leptoge des terrains inondés (Leptogium rivulare) au Canada. Comité sur la situation des espèces en péril au Canada, Ottawa. vi + 33 p.
Le COSEPAC remercie Samuel R. Brinker, Chris Lewis, Graham Cameron et Shaun Thompson d'avoir rédigé le rapport de situation sur le leptoge des terrains inondés (Leptogium rivulare) au Canada, aux termes d'un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par David Richardson, coprésident du Sous-comité de spécialistes des mousses et lichens du COSEPAC.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel COSEPAC
Site Web COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Flooded Jellyskin Leptogium rivulare in Canada.
Illustration/photo de la couverture :
Leptoge des terrains inondés -- Photo gracieusement fournie par Chris Lewis.
Leptoge des terrains inondés Leptogium rivulare
Leleptoge des terrains inondés (Leptogium rivulare)est un petit lichen foliacé à thalle gris ou gris bleuâtre (à l'état sec) dont la surface devient gélatineuse lorsqu'elle est mouillée. Chaque thalle mesure jusqu'à 4 cm de diamètre et présente de nombreux petits organes de fructification (apothécies) brun rougeâtre sur sa surface. Le leptoge des terrains inondés est un cyanolichen, c'est-à-dire un lichen dont le symbiote photosynthétique est une cyanobactérie du genre Nostoc. Les cyanolichens fournissent des quantités importantes d'azote aux écosystèmes qui les hébergent. Le leptoge des terrains inondés est un des seuls macrolichens capables de tolérer une immersion saisonnière dans l'eau douce.
Le leptoge des terrains inondésest une espèce rare à l'échelle mondiale qui se rencontre dans les régions boréales et tempérées, dans les anciennes régions glaciaires de l'est de l'Amérique du Nord ainsi que de l'est, du centre et de l'ouest de l'Europe. Il se rencontre principalement entre les 45e et 60e parallèles nord. Aux États-Unis, le leptoge des terrains inondés a déjà été signalé en Illinois et au Vermont (potentiellement dans des refuges glaciaires), mentions les plus méridionales de l'espèce, mais il existe une seule mention récente de l'espèce, dans le centre du Wisconsin.
Au Canada, on connaît trois sous-populations de leptoge des terrains inondés. La plus grande sous-population, celle des basses terres de l'Ontario, est principalement limitée aux mares printanières boisées. La sous-population du Sud du Bouclier, deuxième en importance, se situe le long de la limite sud du Bouclier précambrien, près de l'interface avec les basses terres d'âge Paléozoïque, en Ontario et au Québec, et comprend des occurrences isolées à Wawa et à Temagami. La sous-population du bassin du lac postglaciaire Agassiz comprend des localités très éparpillées dans l'écorégion de la forêt boréale, dans le nord du Manitoba et l'Ontario. Un groupe d'occurrences qui se trouve près de Flin Flon, au Manitoba, constitue la mention la plus nordique (55°N) de l'espèce au Canada.
Au Canada, le leptoge des terrains inondés pousse uniquement dans les milieux humides et calcaires soumis à des inondations saisonnières. La sous-population des basses terres de l'Ontario est principalement limitée aux mares printanières boisées. La sous-population du Sud du Bouclier se rencontre également dans des marécages et des mares périodiquement inondés, le long de la limite sud du Bouclier précambrien, près de l'interface avec les basses terres d'âge Paléozoïque. La sous-population du bassin du lac postglaciaire Agassiz est petite et composée d'occurrences éparpillées sur un vaste territoire dans le nord de l'Ontario et du Manitoba, où l'espèce colonise des affleurements rocheux ou de gros blocs rocheux situés dans la zone inondable du rivage de lacs assis sur un substrat rocheux calcaire ou en bordure de cours d'eau ou de lacs qui connaissent des crues saisonnières et présentent des dépôts calcaires. Pour que le leptoge des terrains inondés prospère, l'eau doit avoir une faible charge en sédiments, le substrat doit être adéquat (arbre, arbuste ou roche), et la température doit être appropriée. Le leptoge des terrains inondés est généralement observé sur des frênes et, dans une moindre mesure, sur des érables, des ormes et des saules. L'ombrage partiel que fournissent les arbres et les grands arbustes semble être important pour le maintien d'un taux d'humidité élevé et de températures modérées durant les mois d'été. L'espèce ne tolère généralement pas l'ombre complète. La faible capacité de dispersion du leptoge des terrains inondés limite probablement la présence et l'abondance de l'espèce.
Le leptoge des terrains inondés produit généralement une abondance d'apothécies, et la reproduction sexuée est importante pour le maintien de l'espèce. Les spores sont dispersées de manière passive par le vent et peuvent possiblement être emportées par le courant. L'espèce ne produit aucun organe végétatif spécialisé, mais elle peut probablement se reproduire par fragmentation à petite échelle. La dispersion de l'espèce est probablement limitée par le fait que les conditions dont elle a besoin sont peu communes dans le paysage et que les spores ont besoin pour germer d'un substrat au pH convenable, d'une température, d'une luminosité et d'un taux d'humidité convenables ainsi que de la présence d'une cyanobactérie compatible qui permet la régénération d'un organisme symbiotique champignon-algue. Certains vecteurs biotiques, comme les oiseaux et les mammifères, pourraient constituer des moyens de dispersions, mais de manière peu fréquente.
Il est impossible d'évaluer les tendances en matière de répartition ou d'effectif pour le leptoge des terrains inondés au Canada, car on dispose de très peu de données historiques. Jusqu'à 2004, la seule population canadienne connue comprenait uniquement quatre occurrences. Une de ces occurrences, celle de Wawa, en Ontario, est historique et n'a pas été retrouvée, et il est probable qu'elle soit disparue à cause de la pollution atmosphérique et de la destruction de son habitat. Depuis 2004, l'intensification des activités de recherche et l'acquisition de connaissances sur les besoins en matière d'habitat du leptoge des terrains inondés ont permis une hausse du nombre d'occurrences connues, qui est actuellement de 76 (environ 352 000 individus). Il est probable que des occurrences additionnelles existent dans le nord de l'Ontario et au Manitoba et possiblement en Saskatchewan et dans le nord du Québec, dans les régions anciennement recouvertes par les lacs postglaciaires Agassiz et Ojibway. Cependant, la sous-population du lac Agassiz se compose de sites très éparpillés comptant chacun un petit nombre de thalles, de sorte qu'il est peu probable que des recherches supplémentaires dans ces autres régions mènent à une augmentation considérable de l'effectif total connu.
D'après le calculateur des menaces du COSEPAC, l'impact des menaces sur la survie du leptoge des terrains inondés est élevé. Depuis l'évaluation précédente de l'espèce par le COSEPAC, en 2004, la gravité et la portée des menaces ont changé. Actuellement, la principale menace pesant sur le leptoge des terrains inondés est l'agrile du frêne, qui tue toutes les espèces indigènes de frênes et se propage rapidement en Ontario et au Québec. Les frênes sont des hôtes importants dans une proportion considérable de la répartition du leptoge des terrains inondés. En fait, 99 % des thalles connus sont associés à des communautés végétales où le frêne est présent. Vingt des 76 occurrences connues (environ le quart de la population canadienne) se trouvent dans des milieux dominés par le frêne, et le frêne est un hôte codominant dans 7 autres occurrences. Compte tenu du rythme connu de propagation de l'agrile du frêne, les occurrences de leptoge des terrains inondés du sud de l'Ontario et du Québec seront probablement toutes touchées par cette menace au cours des 10 à 20 prochaines années. L'orme, autre hôte important pour le leptoge des terrains inondés dans les occurrences du centre de l'Ontario, est touché par la maladie hollandaise de l'orme, qui tue des arbres de façon continue dans la province.
Le changement climatique constitue une autre menace importante pour l'espèce, car il pourrait modifier les inondations saisonnières dans les mares printanières et le long des cours d'eau où les crues favorisent l'espèce ainsi que l'établissement des arbres et arbustes hôtes préférés par l'espèce. Environ 80 % des occurrences de leptoge des terrains inondés sont associées à des mares printanières, type de milieu qui risque de s'assécher et de devenir moins fréquent. En outre, la capacité de dispersion limitée du leptoge des terrains inondés accentue sa vulnérabilité au changement climatique, car plusieurs des occurrences sont petites et isolées dans des parcelles restantes de forêt renfermant des mares printanières.
Les barrages constituent une autre menace pour le leptoge des terrains inondés, car ils modifient les régimes d'inondations le long des rivières. La modification des régimes hydrologiques pourrait endommager ou détruire l'habitat du leptoge des terrains inondés. D'autres activités qui entraînent une modification des cours d'eau, de la qualité de l'eau ou de la végétation protectrice qui entoure les sites hébergeant l'espèce, comme l'exploitation forestière, l'exploitation minière, les carrières et le développement urbain, pourraient dégrader l'habitat et ainsi accroître l'exposition des individus au rayonnement solaire et au vent, réduire le taux d'humidité et augmenter l'érosion et la turbidité de l'eau.
L'inscription du leptoge des terrains inondés à la liste rouge mondiale a été proposée en janvier 2015. L'espèce a été désignée « menacée » par le COSEPAC en 2004 et a ensuite été inscrite à ce titre à l'annexe 1 de la Loi sur les espèces en péril du Canada. Un programme de rétablissement fédéral sur l'espèce a été publié en 2013. En outre, le leptoge des terrains inondés est désigné « espèce menacée » aux termes de la Loi de 2007 sur les espèces en voie de disparition de l'Ontario, qui confère une protection à l'espèce et à son habitat. Il est également protégé dans un parc provincial au Manitoba et dans neuf parcs provinciaux ou réserves de conservation en Ontario, qui englobent environ 4 % de la population canadienne totale. Le leptoge des terrains inondés ne bénéficie d'aucune protection juridique particulière au Québec.
| Sujet | Information |
|---|---|
| Durée d'une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN [2008] est utilisée) Inconnue dans le cas du leptoge des terrains inondés, mais elle serait probablement de l'ordre de 10-à-20-ans chez d'autres cyanolichens selon certaines recherches (COSEWIC, 2011a, b). |
10 à 20-années |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures? Des déclins ont été observés et signalés dans quelques occurrences bien connues où des ormes hôtes ont été tués par la maladie hollandaise de l'orme; les individus qui vivaient sur ces arbres meurent à mesure que l'écorce se désagrège. Toutefois, la plupart des occurrences n'ont jamais fait l'objet de relevés répétés. |
Oui |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] La plupart des occurrences n'ont jamais fait l'objet de relevés répétés. |
Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. Des relevés répétés doivent être réalisés. |
Inconnu |
| Pourcentage [prévu] [de réduction] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. Des déclins sont prévus en raison des infestations d'agrile du frêne dans 20-occurrences dominées par le frêne, qui hébergent 10-% du nombre total d'individus matures, ainsi que dans 7-autres occurrences où le frêne est un hôte codominant important, qui représentent 34-% additionnels de la population canadienne totale (voir les tableaux 1-et-2). |
Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. Les occurrences n'ont pas fait l'objet de relevés répétés. |
Inconnu |
| Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé? Les causes du déclin ne sont pas réversibles. La position officielle du Canada est que l'agrile du frêne ne peut pas être éradiqué, et il est également impossible d'éradiquer la maladie hollandaise de l'orme. |
Non |
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Probablement pas |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 1-861-848 km2 |
| Indice de zone d'occupation (IZO) 388 km2 en Ontario, 36 km2 au Manitoba et 12 km2 au Québec |
436 km2 |
| La population totale est-elle gravement fragmentée? Les occurrences les plus au nord sont fragmentées, mais elles représentent seulement une petite proportion de la population totale. |
Non |
| Nombre de localités (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN (février 2014) pour obtenir des précisions sur ce terme.) 37-localités définies en fonction des menaces suivantes-: agrile du frêne, barrages, extraction d'agrégats et expansion des zones urbaines/industrielles. |
37 |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | On ne sait pas |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | On ne sait pas |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations? L'espèce est disparue d'un site historique à Wawa, probablement à cause d'un barrage hydroélectrique qui a réduit l'inondation de l'habitat. |
Oui |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? Oui, un déclin a été inféré en fonction de la propagation de l'agrile du frêne. Toutefois, il est difficile d'évaluer l'ampleur de ce déclin, car on ne dispose pas de données historiques. |
Oui |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? Il est prévu que l'agrile du frêne touchera au moins 20-occurrences où le frêne rouge et le frêne noir dominent l'étage arboré. Les frênes représentent environ 50-% de l'étage arboré dans 7-autres occurrences, et des frênes sont présents mais représentent une composante mineure dans 37-autres occurrences (tableaux-1 et 2). En outre, l'orme d'Amérique, autre hôte de l'espèce, continue d'être touché par la maladie hollandaise de l'orme, ce qui entraîne une modification de l'habitat de l'espèce et une diminution du substrat disponible. |
Oui |
| Y a-t-il des fluctuations extrêmes du nombre de populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités*? | Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
| Sous-population | Nombre d'individus matures |
|---|---|
| Basses terres de l'Ontario (33 occurrences) | ~190-000 |
| Sud du Bouclier (36 occurrences) | ~160-000 |
| Bassin du lac postglaciaire Agassiz (7 occurrences) | ~2-000 |
| Total | ~352-000 |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans] | S/O |
La principale menace pesant sur le leptoge des terrains inondés est l'agrile du frêne, qui tue les arbres hôtes et entraîne ainsi la mort des individus de l'espèce. Lorsque les arbres meurent, le substrat ne convient plus à l'espèce. Le changement climatique constitue une autre menace importante, car on s'attend à ce qu'il cause un assèchement des conditions dans le sud de l'aire de répartition du leptoge des terrains inondés. Ce phénomène entraînera probablement une diminution de l'inondation des mares printanières ainsi qu'une modification des crues le long des cours d'eau où des inondations périodiques sont nécessaires au maintien de l'habitat. Les autres menaces sont la destruction de l'habitat associée à l'extraction d'agrégats, la modification des régimes d'inondation associée à la construction de barrages sur les rivières et la perte d'habitat associée au développement ou à l'étalement urbain et industriel. La récolte d'arbres à l'intérieur ou à proximité des mares printanières peut entraîner une augmentation de l'exposition au soleil, ce qui pourrait causer une hausse des taux d'évaporation et une diminution des taux d'humidité.
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur Des spécimens de leptoge des terrains inondés ont été récoltés au 19e-siècle en Illinois et au Vermont, mais l'espèce est actuellement présente uniquement au Wisconsin (Bennet et Bartkowiak, 2013). |
- |
| Une immigration a-t-elle été constatée ou est-elle possible? | Probablement pas |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Inconnu, mais probable. |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? Oui, on trouve des milieux propices à l'espèce dans le nord de l'Ontario et du Manitoba, dans le centre de l'Ontario et le sud-est du Québec ainsi que dans certaines portions du nord-est de la Saskatchewan et du nordouest du Québec, dans les régions anciennement recouvertes par les lacs postglaciaires Agassiz et Ojibway. Dans les régions chevauchant la ceinture argileuse, l'envasement important dans la colonne d'eau pourrait constituer un facteur limitatif. |
Oui |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? Aux États-Unis, la population la plus proche se trouve dans le centre du Wisconsin, à environ 670-km à l'ouest d'une sous-population canadienne située en Ontario. |
Probablement pas |
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Non |
COSEPAC : Espèce désignée «-menacée-» en mai 2004. Réexamen du statut : l'espèce a été désignée «-préoccupante-» en novembre 2015
Depuis que le COSEPAC a évalué le leptoge des terrains inondés en 2004, des activités de recherche considérables ont permis une amélioration des connaissances sur la répartition et les effectifs de l'espèce dans le centre et l'est de l'Ontario, et des relevés ont permis une expansion de son aire de répartition connue dans le sud-ouest du Québec, le nord de l'Ontario et le sud-est du Manitoba. En outre, son aire de répartition connue dans le sud de l'Ontario s'est étendue vers l'ouest, jusqu'aux comtés de Grey et de Bruce, et vers le nord, jusqu'au parc provincial Algonquin. De plus, la présence de l'espèce dans le site historique de Temagami a été confirmée.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Leptogium rivulare (Ach.) Mont. in Gaudichaud, Bot. Voy. Monde La Bonite: 117 (1846). [nomen sed non planta]. Lichen rivulare Ach., Lich. Suec. Prodr. 131 (1798); type : Suède (H-Ach 1915B, lectotype désigné par Jørgensen et James [1983]).
Classification : Le genre Leptogium est classé dans la famille des Collématacées, dans l'ordre des Peltigérales, dans la classe des Lécanoromycètes et dans l'embranchement des Ascomycètes. Le Leptogium rivulare fait partie du complexe du Leptogium azureum et se distingue par ses asques à quatre spores et par son habitat particulier. En Amérique du Nord, l'espèce a longtemps été appelée L. crenatellum Tuck. (par exemple, Fink, 1935, Sierk, 1964), mais il s'agit d'un synonyme de L. rivulare, comme l'ont établi Jørgensen et James (1983), qui ont étudié la nomenclature de l'espèce. Les types pertinents ont été examinés par Sierk (1964), auteur d'une monographie du genre, et par le lichénologue norvégien Per Magnus Jørgensen (1994).
Le genre Leptogium est cosmopolite et comprend environ 180 espèces, dont 60 ont été signalées en Amérique du Nord (Esslinger, 2012), 30 au Canada, et 19 en Ontario, au Manitoba et au Québec.
Nom commun : Le nom anglais du leptoge des terrains inondés, « Flooded Jellyskin » décrit l'habitat de l'espèce (flooded) ainsi que le caractère gélatineux des lichens des genres Leptogium et Collema (jellyskin).
Le Leptogium rivulare est un petit lichen foliacé à thalle gris ou gris bleuâtre présentant de nombreuses petites apothécies brun rougeâtre clair (figure 1). Le thalle mesure 0,5 à 4 cm de largeur (Sierk, 1964) et comporte des lobes lisses dépourvus des plis, sorédies ou isidies qui caractérisent les espèces semblables, arrondis à plutôt allongés, larges de 0,7 à 3,5 mm. Les cortex supérieur et inférieur sont chacun constitués d'une seule couche de cellules arrondies. La moelle est extrêmement mince. Le photobionte est une cyanobactérie du genre Nostoc. À l'état humide, les lobes se gonflent d'eau et deviennent gélatineux et translucides, d'où le nom anglais « jellyskin » donné aux lichens de ce genre. Le lichen est habituellement fertile, et les apothécies peuvent être assez abondantes pour donner à l'ensemble du thalle un aspect brunâtre lorsqu'observé à une certaine distance. Cette fertilité exceptionnelle permet de distinguer facilement le L. rivulare du L. cyanescens (Rabenh.) Korber, très commun et à thalle beaucoup plus grand. Les apothécies du L. rivulare peuvent être superficielles ou rétrécies à la base, et leur diamètre est normalement de 0,4 à 0,6 mm, atteignant parfois 1,2 mm. Le disque est brun clair à brun rougeâtre, plat ou légèrement concave, à marges lisses, uniformes, relativement minces, avec une couche extérieure de tissu thalloïde gris. Les spores sont incolores, multicellulaires, submuriformes, avec trois (ou quatre) cloisons transversales et une seule cloison longitudinale, qui est parfois absente. Les spores sont elliptiques, mais légèrement pointues aux extrémités, et mesurent 15 à 21 μm sur 7,5 à 10 μm. Il y a toujours quatre spores par asque (Hinds et Hinds, 2007; Jørgensen, 2007; Brodo et al., 2001).
Composition chimique : Aucune substance lichénique n'a été signalée chez le L. rivulare (Brodo et al., 2001; Jørgensen, 2007).
Longue description de la figure 1
Photo de deux thalles de leptoge des terrains inondés (Leptogium rivulare) poussant sur l'écorce d'un arbre. Ce lichen foliacé à thalle gris bleuté possède des lobes lisses (dépourvus des plis, sorédies ou isidies qui caractérisent les espèces semblables) arrondis à quelque peu allongés. Le thalle présente à sa surface de nombreuses apothécies, qui peuvent être superficielles ou rétrécies à la base. Le disque est brun clair à brun rougeâtre, plat à légèrement concave, à marges lisses, uniformes, relativement minces, avec une couche extérieure de tissu thalloïde gris.
Il est possible que la sous-population de L. rivulare du bassin du lac postglaciaire Agassiz, qui pousse sur la roche, soit issue d'une population source différente de celle des populations du sud, qui colonisent l'écorce d'arbres (voir la section Répartition). Toutes les occurrences nordiques de L. rivulare sont associées à des régions inondées par le lac postglaciaire Agassiz (occurrencesde la rivière Attawapiskat, du lac Sailing, de la rivière Severn, du lac Stout-embouchure de la rivière Crooked, de Tri Lakes et du lac Whitefish) ou à ses principaux émissaires et voies interlacustres (occurrences du parc Algonquin, du lac Temagami et de Wawa). Après le retrait des glaciers, les niveaux d'eau ont baissé et le relèvement isostatique a causé une modification de l'écoulement, et les cours d'eau ont fini par adopter leur configuration actuelle, ce qui a peut-être isolé le L. rivulare dans des parcelles d'habitat propice reliques dans certaines régions; cette théorie viendrait contredire celles selon lesquelles il y a eu une dispersion aléatoire à longue distance. Graf (1997) ainsi que Harris et Keeny Marr (2009) ont décrit de façon relativement détaillée, pour d'autres espèces aquatiques, les voies migratoires postglaciaires à partir de refuges glaciaires, et ils ont établi un lien entre la répartition actuelle du Caltha natans, plante aquatique, et l'emplacement d'anciens lobes glaciaires associés au lac glaciaire Agassiz.
Les canaux creusés par les eaux de fonte glaciaire le long du rivage du lac glaciaire Agassiz, dans le nord de l'Ontario et certaines parties du nord et du sud du Manitoba, pourraient avoir servi au L. rivulare de voies migratoires vers le nord depuis des refuges plus au sud. Le fait que, au Manitoba, les individus des occurrences nordiques poussent uniquement sur la roche alors que ceux des populations plus au sud poussent sur l'écorce vient appuyer la notion de populations issues de différentes sources.
Au Canada, le L. rivulare est présent dans les aires écologiques nationales des Plaines des Grands Lacs et Boréale définies par le COSEPAC (2011c). La présence de l'espèce dans différentes régions écogéographiques pourrait donner à penser que certaines populations ont un caractère distinct, mais aucune donnée génétique ou morphologique n'indique qu'il existe une distinction importante sur le plan évolutif. Ainsi, aux fins de la présente évaluation, une seule unité désignable du L. rivulare est reconnueau Canada. Des recherches préliminaires menées par l'Université du Manitoba sur la phylogénie du photobionte du L. rivulare donnent à penser que le mycobionte du lichen s'associe à une seule souche de cyanobactérie dans l'est de l'Amérique du Nord (Piercey-Normore, comm. pers., 2015).
Le Leptogium rivulare est le résultat d'une relation symbiotique entre un champignon et une cyanobactérie du genre Nostoc. Les cyanolichens fournissent des quantités importantes d'azote aux écosystèmes qui les hébergent (Kallio, 1974; Becker et al., 1977; Campbell et Fredeen, 2004). Le L. rivulare pourrait constituer une importante source d'azote dans les mares printanières lorsqu'un grand nombre d'individus sont présents. En outre, les cyanolichens sont de bons indicateurs de la qualité de l'air et sont particulièrement sensibles au dioxyde de soufre et aux pluies acides (Gilbert, 1970; Ferry et al., 1973; Goward et Arsenault, 2000; Sigal et Johnston, 1986; Farmer et al., 1991; Richardson, 1991). Dans le monde, peu de macrolichens se sont adaptés pour survivre sous l'eau, dans les étangs, les rivières et les autres cours d'eau douce périodiquement inondés. La capacité du L. rivulare de coloniser cette niche spécialisée en fait une espèce unique dans le genre Leptogium.
Le Leptogium rivulareest un lichen à large répartition qui se rencontre dans les régions boréales et tempérées, principalement dans les anciennes régions glaciaires de l'est de l'Amérique du Nord, d'Europe et de certaines parties d'Asie centrale (figure 2). Les premières mentions de l'espèce remontent à 200 ans, en Scandinavie (Suède et Finlande) et en Russie (Jørgensen et James, 1983), et il y a un peu moins de 200 ans en France (Lettau, 1942). En janvier 2015, il a été proposé que le L. rivulare soit inscrit à la liste rouge mondiale. L'espèce est disparue en Finlande, en Estonie et en France et compte une seule mention au Bélarus, en Lithuanie et au Portugal. Elle est relativement abondante dans deux républiques de Russie, celles des Maris (15 mentions) et des Komis (30 mentions) (IUCN, 2015). Il existe également des mentions historiques de l'espèce en Europe de l'Est, soit en Estonie (Randlane, 1987), au Bélarus (Motiejūnaitė et Golubkov, 2005), en Lithuanie (Motiejūnaitė et al., 2011) et en Russie (Paukov et Teptina, 2012). Le L. rivulare a été signalé par Alstrup et Christensen (2006) comme nouvelle espèce en Afrique, en Tanzanie, mais cette mention était erronée, et le spécimen appartenait à un taxon d'Asie (Christensen, comm. pers., 2013).
Aux États-Unis, des spécimens de L. rivulare ont été récoltés au 19e siècle en Illinois et au Vermont, mais ces mentions sont maintenant considérées comme historiques, et l'espèce a récemment été signalée pour la première fois au Wisconsin par Bennet et Bartkowiak (2013). Selon Bennet et Bartkowiak (2013), un spécimen récolté en Géorgie (vers 1978) a été mal identifié, et l'identification d'un spécimen récolté en Oregon (vers 1974) est douteuse.
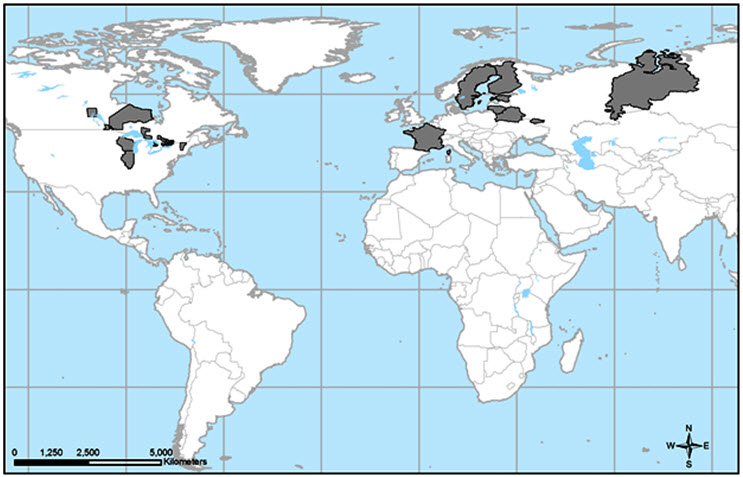
Longue description de la figure 2
Carte illustrant l'aire de répartition mondiale du leptoge des terrains inondés, qui se trouve dans les anciennes régions glaciaires de l'est de l'Amérique du Nord et de l'est, du centre et de l'ouest de l'Europe. L'espèce se rencontre principalement entre les 45e et 60e parallèles nord. Aux États-Unis, le leptoge des terrains inondés a été signalé dans le passé au sud jusqu'en Illinois et au Vermont, mais il existe une seule mention récente de l'espèce, dans le centre du Wisconsin.
Au Canada, trois sous-populations de leptoge des terrains inondés sont connues : celle des basses terres de l'Ontario, qui est principalement limitée aux mares printanières boisées; celle du Sud du Bouclier, se situe le long de la limite sud du Bouclier précambrien, près de l'interface avec les basses terres d'âge Paléozoïque, en Ontario et au Québec, et comprend des occurrences isolées à Wawa et à Temagami; celle du bassin du lac postglaciaire Agassiz, qui comprend des localités très éparpillées dans l'écorégion de la forêt boréale, dans le nord du Manitoba et l'Ontario.
Le Canada compte de loin la plus grande population de Leptogium rivulare dans le monde. L'espèce est présente uniquement dans les aires écologiques nationales des Plaines des Grands Lacs et Boréale (figure 3), au Manitoba, en Ontario et au Québec. Au Canada, l'aire du L. rivulare s'étend depuis Flin Flon, au Manitoba, point le plus au nord et à l'ouest, jusqu'à la rivière Attawapiskat, 1 000 km plus à l'est, à la limite sud-ouest des bassesterres de la baie d'Hudson, puis jusqu'au comté de Grey, 800 km plus au sud, dans le sud de l'Ontario, et enfin vers l'est, le long de la limite sud du Bouclier canadien, dans le centre de l'Ontario, depuis la région des lacs Kawartha jusqu'à la région de Gatineau, au Québec (figure 3). Dans ce vaste territoire, les occurrences situées dans le centre et l'est de l'Ontario, le long du Sud du Bouclier, sont les seules qui se trouvent relativement près les unes des autres; la majorité des autres sites connus sont isolés. L'aire de répartition canadienne totale du L. rivulare représente environ le tiers de l'aire mondiale connue de l'espèce.
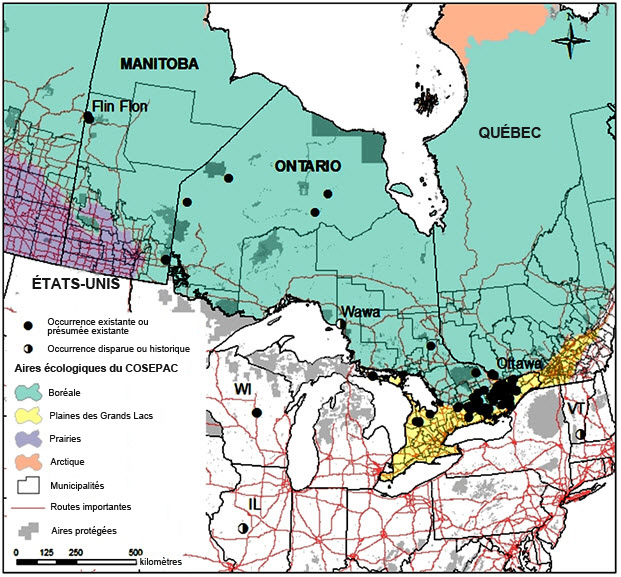
Longue description de la figure 3
Carte montrant la répartition canadienne connue du leptoge des terrains inondés ainsi que les aires écologiques du COSEPAC. Trois occurrences sont également indiquées aux États-Unis, au Wisconsin, en Illinois et au Vermont. Au Canada, le leptoge des terrains inondés est présent uniquement dans les aires écologiques nationales des Plaines des Grands Lacs et Boréale, au Manitoba, en Ontario et au Québec. Son aire s'étend depuis Flin Flon, au Manitoba, point le plus au nord et à l'ouest, jusqu'à la rivière Attawapiskat, 1 000 km plus à l'est, à la limite sud-ouest des bassesterres de la baie d'Hudson, puis jusqu'au comté de Grey, 800 km plus au sud, dans le sud de l'Ontario, et enfin vers l'est, le long de la limite sud du Bouclier canadien, dans le centre de l'Ontario, depuis la région des lacs Kawartha jusqu'à la région de Gatineau, au Québec.
La zone d'occurrence du L. rivulare au Canada est d'environ 1 861 848 km2. La zone d'occupation des populations canadiennes connues est estimée à 157 m2, dont environ 156 m2 en Ontario, 0,1 m2 au Manitoba et 0,6 m2 au Québec. L'indice de zone d'occupation, calculé selon une grille à carrés de 2 km de côté, s'élève à 436 km2 au Canada (388 km2 en Ontario, 36 km2 au Manitoba et 12 km2 au Québec).
Au Canada, trois sous-populations sont connues. La plus grande des souspopulations, celle des basses terres de l'Ontario, est principalement limitée aux mares printanières boisées situées dans des parcelles restantes de forêt décidue à sols issus de matériaux parentaux calcaires. La sous-population du Sud du Bouclier, deuxième en importance, se situe le long de la limite sud du Bouclier précambrien, près de l'interface avec les basses terres d'âge Paléozoïque, en Ontario et au Québec, et comprend des occurrences isolées à Wawa et à Temagami. Les individus de cette souspopulation poussent également dans des mares printanières boisées ou des forêts marécageuses de feuillus périodiquement inondées. La sous-population du bassin du lac postglaciaire Agassiz est petite et limitée à une échelle très locale, mais l'espèce est répandue dans l'écorégion de la forêt boréale du nord de l'Ontario et du Manitoba, où elle pousse sur la roche. Les occurrences de cette sous-population sont limitées aux bords des lacs et des grandes rivières où des sables, des limons et des argiles calcaires gaciolacustres sont exposés par endroits. Un groupe d'occurrences qui se trouve près de Flin Flon, au Manitoba, constitue le site le plus nordique (55°N) où le L. rivulare a été signalé au Canada.
Des recherches ciblant le Leptogiumn rivulare ont été menées par Irwin Brodo après la découverte de l'espèce près d'Ottawa, en Ontario (Brodo, 1971). Le L. rivulare n'a pas été repéré dans les quelque 60 sites semblant pouvoir convenir à l'espèce près de cette ville ou dans des sites plus éloignés situés près de l'île Upper Duck et des îles Petrie, en Ontario, de Chelsea, au Québec, et le long du cours moyen de la rivière Noire, au Québec. En outre, aucun nouveau site hébergeant l'espèce n'a été découvert dans le cadre d'autres recherches menées au Québec, à 160 km au nord-est d'Ottawa, et au lac Anima Nipissing, au nord du lac Temagami, en Ontario (COSEWIC, 2004). De 2001 à 2002, Robert Lee a cherché le L. rivulare dans plus de 60 sites, dans les régions adjacentes à l'occurrence connue d'Ottawa, mais sans succès. Plus tard, il a découvert deux nouveaux sites dans le comté de Lanark, alors que Brodo a trouvé une occurrence poussant sur la roche, près de Flin Flon, au Manitoba (COSEWIC, 2004).
Les activités de vérification sur le terrain associées à la préparation du présent rapport de situation ont ravivé l'intérêt pour le L. rivulare, et des activités de recherche supplémentaires ont mené à la découverte de nombreuses nouvelles occurrences et ont fourni de nouvelles connaissances sur les besoins en matière d'habitat du L. rivulare.
Les relevés lichéniques et les récoltes de spécimens de lichens ont été davantage ciblés en Ontario qu'ailleurs au Canada, grâce aux activités du personnel du Musée canadien de la nature et du ministère des Richesses naturelles de l'Ontario. En outre, de nombreux services aux visiteurs participent aux excursions sur le terrain (figure 4).
De 2005 à 2009, des relevés lichéniques ciblés et généraux ont mené à la découverte d'environ 20 occurrences jamais encore signalées dans l'est de l'Ontario, dans des mares printanières. Les relevés se sont intensifiés de 2010 à 2015, et le personnel du ministère des Richesses naturelles et des Forêts (MRNF) de l'Ontario a consacré plus de 470 heures-personnes à la recherche du L. rivulare (annexe 1) dans environ 176 sites distincts (séparés par au moins 1 km).
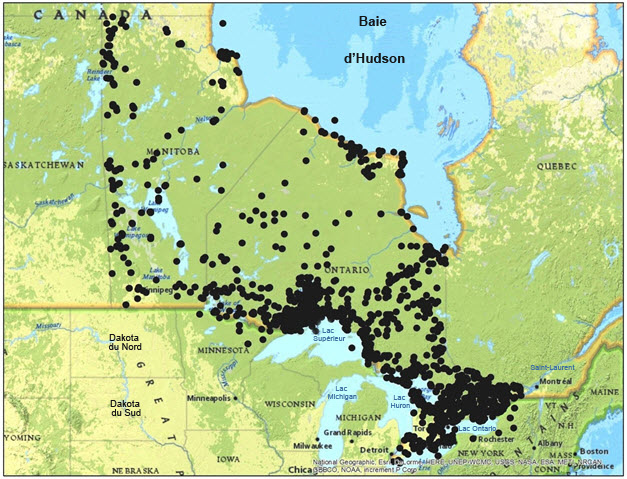
Longue description de la figure 4
Carte montrant les sites où des spécimens de lichens ont été récoltés en Ontario, au Québec et au Manitoba, d'après les données du Consortium of North American Lichen Herbaria. Les données correspondent aux spécimens récoltés du 19e siècle jusqu'à aujourd'hui.
En 2010, 16 nouveaux sites ont été signalés dans l'est de l'Ontario, dans le comté de Lanark et le nord du comté de Peterborough. Un site additionnel a été découvert dans le comté de Leeds et Grenville. En 2011, 16 autres sites répartis dans l'est et le centre de l'Ontario et 2 sites isolés situés dans le nord-ouest de l'Ontario, en bordure de la rivière Severn et du système du lac Stout-embouchure de la rivière Crooked, ont été trouvés dans le cadre du projet Far North Natural Heritage Fieldwork du MRNF. De plus, l'occurrence signalée en 1946 au lac Temagami a été retrouvée.
En 2012, les recherches se sont étendues dans les milieux propices des comtés de Renfrew, de Frontenac, de Hastings, de Peterborough, de Simcoe, de Grey et de Bruce, et 30 sites additionnels ont été signalés. Un nouveau site a été découvert au parc Algonquin. De plus, 15 heures-personnes ont été consacrées, à pied et en canot, à la recherche de l'espèce dans le cours inférieur de la rivière Magpie, près de Wawa, mais le L. rivulare n'a pas été retrouvé.
En 2013, des relevés ont été menés dans certaines portions de la municipalité régionale de Niagara, dans le comté de Haldimand et dans les districts de Nipissing et de Rainy River. Malgré les 19 jours de recherche, aucun site additionnel n'a été découvert. La présence de niveaux d'eau élevés a rendu difficile la réalisation des relevés dans certaines portions de la rivière Rainy, du lac des Bois et du lac Rainy. Les recherches menées le long de la rivière Opeongo donnent à penser que les milieux ne sont pas suffisamment riches à cet endroit, et les autres indicateurs de conditions propices aux espèces calciphiles n'ont pas été observés. Deux nouveaux sites ont été découverts dans des tronçons distincts de la rivière Attawapiskat, dans les basses-terres de la baie d'Hudson, dans le cadre du projet Far North Natural Heritage Fieldwork du MRNF. Aucune occurrence n'a été découverte durant les relevés menés dans les milieux propices du sud du comté de Grey.
En 2014, un site additionnel a été découvert dans le cadre d'un relevé botanique organisé par Conservation de la nature Canada à l'île Cockburn, au lac Huron, dans le district de Manitoulin. D'autres recherches ont été réalisées aux rivières Severn, Black Duck et Ekwan, dans le nord de l'Ontario, et le long de la rivière Pigeon, dans le nordouest de l'Ontario, à l'ouest de Thunder Bay, mais sans succès.
En 2015, des recherches ont été menées sans succès dans quatre sites du sud de l'Ontario, soit le parc provincial Rondeau et les aires de conservation de Kopegaron Woods et de Clear Creek, tous trois situés dans le comté d'Essex, ainsi qu'un marécage inondé, dans l'alvar Carden.
Les recherches ciblées ont été moins nombreuses au Manitoba, mais plusieurs récoltes de lichens en général ont été réalisées dans la plupart des parties de la province (figure 4).
En 2006, des relevés visant le L. rivulare ont été menés près de Flin Flon, où se trouve la seule occurrence alors déjà connue au Manitoba, signalée par Brodo. Les recherches ont été réalisées aux lacs Payuk, Neso, Twin, Athapapuskow, Schist, Naosap et Whitefishs. Aucun individu de l'espèce n'a été trouvé, probablement parce que le niveau de l'eau était trop élevé (Bazin, comm. pers., 2012). D'autres relevés y ont été menés en 2010 sur une période de quatre jours durant laquelle le niveau de l'eau était plus bas, et de nouvelles occurrences du L. rivulare ont été découvertes aux lacs Payuk, Neso, Twin et Whitefishs.
En 2012, des milieux potentiellement propices à l'espèce ont été repérés au lac Sailing, dans le sud-est du Manitoba, durant des relevés des oiseaux nicheurs. Un relevé en bonne et due forme a été réalisé à ce lac ainsi qu'à un certain nombre d'autres lacs interreliés, dont les lacs South Cross, North Cross et Nason. Un petit nombre d'individus de l'espèce ont été trouvés au lac Sailing, dans une baie abritée, le long d'un rivage boisé (Bazin, comm. pers., 2012).
En juin 2013, l'espèce a été cherchée sans succès sur la partie périodiquement immergée de l'écorce de frênes rouges (Fraxinus pennsylvanica) et d'érables à Giguère (Acer negundo) poussant dans une forêt en plaine inondable, en bordure d'une petite section de la rivière Assiniboine, dans le parc provincial Spruce Woods, le long du sentier du lac Marsh.
Au Québec, les activités de recherche ont été relativement limitées comparativement à en Ontario (figure 4). Des relevés ciblant le L. rivulare ont été réalisés en 2012 et 2013, principalement dans la région de l'Outaouais, pour un total 32 heures-personnes.Le premier spécimen confirmé de L. rivulare récolté au Québec a été trouvé dans un petit étang, à Aylmer. Une journée a été consacrée aux relevés dans les régions du parc de la Gatineau et du parc du Lac-Leamy. Les recherches réalisées dans quatre mares printanières au parc du Lac-Leamy ont mené à la découverte de deux sites hébergeant le L. rivulare, mais aucun individu de l'espèce n'a été trouvé dans le parc de la Gatineau. Le milieu y était peut-être trop acide, car aucun indicateur de conditions propices aux espèces calciphiles n'a été observé. L'espèce pourrait peut-être être trouvée dans des milieux de plus faible altitude présentant des affleurements de marbre (Thompson, comm. pers., 2012). Très récemment, une autre occurrence du L. rivulare a été signalée dans un secteur périodiquement inondé du parc du Bic, dans la MRC de Rimouski-Neigette, au Québec (Anderson, comm. pers., 2015).
Le Leptogium rivulare, comme la plupart des autres lichens du genre Leptogium, ne pousse que dans les milieux à substrat riche en composés alcalins ou à substrat recevant un apport en calcium. Le L. rivulare est une espèce spécialiste en matière d'habitat et se rencontre uniquement sur l'écorce ou la roche, dans les milieux humides, partiellement ombragés et périodiquement inondés par de l'eau douce présentant de très faibles charges de sédiments. Au Canada, on trouve des milieux qui sont à la fois calcaires et soumis à des inondations périodiques sur le rivage de plans d'eau alcalins (étangs, lacs, mares printanières) (figure 5) et de rivières dont le niveau fluctue de manière saisonnière (figure 6) et qui sont assis sur un substrat rocheux calcaire (c'est-à-dire de calcaire ou de dolomie) ou dont le bassin est constitué de matériaux parentaux calcaires (issus de dépôts glaciaires ou d'argiles lacustres déposées dans des lacs proglaciaires) (Brinker et Lewis, 2011). Les milieux qui :
- ne répondent pas à ces exigences,
- sont exposés à des concentrations accrues de polluants atmosphériques ou
- se trouvent dans des paysages fortement urbanisés ou agricoles recevant des charges accrues en sédiments (notamment dans certaines portions du sud du Manitoba et du sud de l'Ontario) ne semblent pas convenir au L. rivulare (S. Thompson, C. Lewis, G. Cameron, S. Brinker, obs. pers.).
Ces exigences expliquent probablement l'absence apparente de l'espèce dans la majeure partie du sud de l'Ontario, où les pluies acides ont entraîné une diminution considérable du pH du substrat et où les plans d'eau présentent généralement des charges élevées en sédiments, ainsi que dans plusieurs parties du Manitoba (voir la section « Facteurs limitatifs »).

Longue description de la figure 5
Photo montrant une mare printanière hébergeant le leptoge des terrains inondés dans un boisé du centre de l'Ontario. Plusieurs des arbres sont entourés d'eau.

Longue description de la figure 6
Photo montrant l'habitat riverain d'une occurrence de leptoge des terrains inondés, en bordure de la rivière Attawapiskat, au Manitoba. Le niveau de la rivière fluctue de façon saisonnière, et des pierres éparses sont visibles le long du rivage.
Dans la sous-population des basses terres de l'Ontario et la sous-population du Sud du Bouclier, en Ontario et au Québec, les individus poussent le plus souvent sur l'écorce d'arbres ou d'arbustes qui sont soumis à des inondations saisonnières. On trouve parfois des individus sur des pierres situées à proximité d'arbres colonisés par le L. rivulare. Toutefois, la superficie occupée par le L. rivulare sur la roche est minime comparativement à celle occupée sur l'écorce en Ontario. Au Manitoba, dans la souspopulation de L. rivulare du lac glaciaire Agassiz, les individus ont été observés uniquement sur la roche, sur les rivages de lacs alcalins, dans des sites généralement abrités des vents dominants et de l'action des vagues. Dans ces milieux, le L. rivulare pousse en compagnie de mousses et du Dermatocarpon luridum, autre espèce de lichen, sur des affleurements rocheux ou des pierres présentant des fissures, dans des zones périodiquement inondées de petites baies abritées (Bazin, comm. pers., 2012).
Selon le COSEPAC (2004), l'espèce préfère les arbres âgés dont l'écorce est rugueuse et recouverte de mousses. Toutefois, Robillard (2012) n'a trouvé aucun lien entre la présence de l'espèce et le diamètre des arbres, et dans le cadre des récents relevés menés près de Stony Swamp, le L. rivulare était abondant sur les gros arbres âgés à écorce rugueuse ainsi que sur de plus petits arbres à écorce lisse, avec ou sans bryophytes. Le L. rivulare se rencontre le plus souvent sur le frêne rouge (Fraxinus pennsylvanica) et le frêne noir (Fraxinus nigra) et est généralement plus abondant chez cette dernière espèce. Des individus ont été signalés sur des érables (Acer saccharinum, Acer ×freemanii, Acer rubrum), mais leur écorce semble présenter un pH plus faible et moins retenir l'humidité que celle des frênes, ce qui explique que le L. rivulare soit moins commun et que la diversité de lichens soit plus faible chez les érables (Brinker, comm. pers., 2015). En outre, l'espèce peut coloniser l'orme d'Amérique (Ulmus americana) et occasionnellement le peuplier baumier (Populus balsamifera) et le chêne à gros fruits (Quercus macrocarpa). L'espèce a également été observée sur le peuplier deltoïde (Populus deltoides), le caryer cordiforme (Carya cordiformis) et le bouleau jaune (Betula alleghaniensis). Il arrive parfois que le L. rivulare colonise des branches récemment tombées des arbres, mais il disparaît lorsque l'écorce se désintègre. Le L. rivulare a déjà été signalé, mais très rarement, sur une seule espèce de conifère, le Thuja occidentalis. De plus, le L. rivulare a occasionnellement été observé sur des arbustes, dont le cornouiller stolonifère (Cornus stolonifera), le céphalanthe occidental (Cephalanthus occidentalis) et l'aulne rugueux (Alnus incana ssp. rugosa), et il a été signalé une fois sur le nerprun bourdaine (Frangula alnus). Il a également été observé à de très rares occasions sur la vigne des rivages(Vitis riparia). Dans le nord de l'Ontario, l'espèce a été signalée sur le saule de Bebb (Salix bebbiana) et le saule satiné (Salix pellita), en bordure de rivières qui connaissent des crues saisonnières.
Les tendances associées aux milieux humides colonisés par le L. rivulare sont peu connues. De nombreux exemples de milieux humides hébergeant l'espèce subsistent, même dans les paysages fortement urbanisés ou dominés par les activités agricoles, car certaines parties de ces milieux sont souvent trop humides ou trop marginaux pour être exploités. Cependant, depuis la colonisation du sud de l'Ontario et du Québec, la superficie de milieux humides a connu un grave déclin, particulièrement dans l'est de l'Ontario, dans les régions du Niagara et de Toronto, où plus de 85 % des milieux humides ont été convertis pour d'autres utilisations (Ducks Unlimited, 2010). Dans le sud de l'Ontario, le comté d'Ottawa-Carleton et le sud des comtés de Frontenac, de Lennox et d'Addington sont les régions hébergeant le L. rivulare où le déclin des milieux humides a été le plus marqué (perte de 65 à 85 %; Ducks Unlimited, 2010). Dans les comtés de Hastings, de Bruce, de Lanark et de Leeds et Grenville, la diminution des superficies de milieux humides a été modérée (45 à 65 %) depuis la colonisation (Ducks Unlimited, 2010). Parmi les régions qui hébergent le L. rivulare,celles qui ont été les moins touchées par la diminution des superficies de milieux humides sont la ville de Kawartha Lakes ainsi que les comtés de Peterborough et de Grey, où les pertes ont été de 25 à 45 % (Ducks Unlimited, 2010). La dégradation de l'habitat (diminution de sa superficie et de sa qualité) se poursuit dans l'ensemble de cette région.
On trouve des barrages dans la plupart des parties de l'aire de répartition canadienne du L. rivulare depuis le milieu du 19e siècle, d'abord utilisés par les scieries, puis pour la production d'hydroélectricité et la lutte contre les inondations. Les projets proposés de construction de nouveaux barrages constituent une menace pour l'espèce, particulièrement dans le nord (voir la section « Menaces »).
Les pertes d'habitat ont été relativement plus faibles dans les sites du nord de l'Ontario et du Manitoba, où l'espèce pousse sur les rivages rocheux de lacs et de rivières, que dans les autres régions. Toutefois, ces milieux hébergent seulement une petite portion de la population totale de L. rivulare.
Le cycle vital du L. rivulare n'a fait l'objet d'aucune étude. Les individus de l'espèce sont généralement fertiles, sauf lorsqu'ils sont très jeunes. La reproduction sexuée joue un rôle primordial dans le maintien des populations de L. rivulare, car celui-ci ne produit aucune structure végétative (sorédies, isidies, schizidies, etc.). Le L. rivulare peut probablement se multiplier par fragmentation, lorsque les minuscules lobules se détachent du thalle parent et se retrouvent sur un substrat propice à proximité.
On en sait peu sur le taux de croissance et la longévité du L. rivulare. Un thalle de 2 cm de diamètre, peut-être issu d'un fragment de thalle provenant d'une colonie située à proximité, a été trouvé dans un site de l'est de l'Ontario, sur la tige d'un cornouiller stolonifère âgé de 4 ans (selon le nombre de cernes et le nombre de verticilles de cicatrices foliaires). Cette observation donne à penser que le lichen peut croître de 2,5 mm par année (Lewis, obs. pers.), ce qui se compare aux autres lichens foliacés arboricoles (Brodo et al., 2001).
On ne dispose d'aucune donnée sur la durée d'une génération chez l'espèce, mais des rechercher sur d'autres cyanolichens donnent à penser qu'elle pourrait être de l'ordre de 10 à 20 ans (Larsson et Gauslaa, 2011), et cette durée a été adoptée pour deux autres lichens du genre Leptogium évalués par le COSEPAC (COSEWIC, 2011a, b).
Les exigences physiologiques du L. rivulare n'ont pas fait l'objet d'études poussées. Il semble que l'ensemble des cyanolichens ait deux exigences. Premièrement, ils ont besoin d'eau sous forme liquide (la vapeur d'eau suffit à déclencher l'activité physiologique chez de nombreux lichens, mais non chez ceux du genre Leptogium) (COSEPAC, 2011a,b). Deuxièmement, ils ont besoin d'un substrat riche en composés alcalins et ayant généralement un pH de 5 ou plus (Gauslaa, 1985; Gauslaa, 1995; COSEWIC, 2011a,b). Les surfaces ayant un pH inférieur à 5 (notamment l'écorce de la plupart des conifères) ne peuvent probablement pas héberger de colonies de L. rivulare.
En outre, le L. rivulare semble avoir besoin d'un ensoleillement au moins partiel. Le couvert forestier des sites colonisés présente toujours des ouvertures, parce que les inondations périodiques limitent la densité du couvert ou que les sites se trouvent sur des rivages, face à des superficies d'eaux libres. Le rayonnement solaire est donc plus fort à ces endroits qu'aux alentours. L'ombrage excessif inhibe probablement la photosynthèse et la croissance chez l'espèce. De plus, la sédimentation excessive, qui laisse une couche de particules d'argiles sur l'écorce des arbres après les inondations, rend le milieu non propice à l'espèce, car les particules recouvrent alors les thalles et limitent leur photosynthèse.
Le L. rivulare semble tolérer les eaux stagnantes et courantes. Dans les mares printanières isolées, l'eau demeure stagnante et calme, alors que les individus situés sur les rivages des lacs peuvent être exposés aux vagues, et ceux qui poussent en bordure de grandes rivières sont soumis à des courants turbulents durant la fonte printanière.
L'espèce se disperse par ses spores sur de longues distances de manière passive. La fragmentation des thalles pourrait constituer un moyen de dispersion sur de courtes distances, les fragments étant probablement transportés par l'eau. Cependant, il est possible que la reproduction sexuée soit rarement couronnée de succès chez l'espèce, car il faut que les spores atterrissent sur un substrat convenable dans un milieu qui présente les conditions requises en ce qui a trait au pH, à la qualité de l'eau, à la température, à la luminosité, etc., et où on trouve la cyanobactérie compatible qui permet la régénération d'un organisme symbiotique champignon-algue. Ces exigences pourraient expliquer que de nombreuses occurrences du L. rivulare soient fragmentées. Le flux génétique entre les habitats pourrait être limité. D'autres mécanismes de dispersion physique, comme le vent, les amphibiens, les arthropodes, les mammifères, etc., pourraient parfois permettre le transport de spores ou de fragments de thalles jusque dans des milieux isolés, comme des mares printanières situées dans des parcelles de forêt restantes.
Le Leptogiumn rivulare a été observé en association avec un certain nombre d'autres espèces de lichens et de bryophytes. Dans le centre de l'Ontario, l'espèce pousse souvent aux côtés du leptoge cylindrique (Leptogium teretiusculum) et, moins fréquemment, du leptoge subtil (Leptogium subtile). Dans le nord de l'Ontario, le L. rivulare a été observé en compagnie du Bacidia rosella, espèce de lichen, dans deux sites. Au Manitoba, où le L. rivulare colonise la roche, l'espèce pousse souvent aux côtés du dermatocarpe jaune pâle (Dermatocarpon luridum).
Le L. rivulare semble avoir une faible capacité de compétition. Il est capable de survivre lorsqu'il est mélangé avec des bryophytes corticoles qui supplantent généralement les lichens, mais avec le temps, le L. rivulare peut être délogé par les bryophytes lorsqu'elles sont abondantes. Une certaine couverture de bryophytes pourrait en fait être profitable au L. rivulare, car elle réduit le rythme de la perte d'eau du substrat et créerait ainsi un microclimat plus humide. Les bryophytes corticoles observées aux côté du L. rivulare dans plus d'une occurrence sont l'Amblystegium riparium, l'A. varium, le Dichelyma pallescens, le Fontinalis novae-angliae, le Hynum linddbergii, le Leptodictyum riparium, le Leskea polycarpa et le Plagiothecium cavifolium.
Les relevés visant le L. rivulare ont été menés dans plusieurs types d'habitat distincts (voir la section « Besoins en matière d'habitat »). Les observateurs ont examiné les troncs d'arbres, les arbustes, les affleurements rocheux et les pierres, depuis le sol jusqu'à une hauteur de 2 m, selon la limite des inondations saisonnières. Les zones visées par les relevés ont été déterminées sur le terrain selon les évaluations intuitives d'experts; seules les zones accessibles (terre de la couronne, parcs municipaux, aires de conservation, etc.) ou pour lesquelles une permission d'entrée a été obtenue ont été visitées. Des demandes ont été présentées avant la réalisation des travaux de terrain dans les parcs provinciaux et les réserves de conservation, et un permis accordé aux termes de la Loi sur les espèces en péril (LEP) a été obtenu en vue de la réalisation des recherches sur les terres fédérales.
Les observateurs ont évalué la couverture spatiale des thalles de L. rivulare en superposant une de leurs mains aux thalles (main à plat avec doigts et pouce collés) et en consignant systématiquement le « nombre de mains » pour chaque site visité, puis en convertissant ce nombre en centimètres carrés (d'après la superficie de la main de l'observateur). Lorsque la couverture du L. rivulare était morcelée, les observateurs ont utilisé une de leurs mains déployée (main à plat avec doigts et pouce écartés au maximum) pour mesurer la superficie occupée. Lorsque les thalles étaient très clairsemés, les observateurs ont évalué la superficie occupée au mieux de leur capacité. Pour évaluer le nombre total d'individus, on a considéré que chaque thalle mesurait en moyenne 4,5 cm2, d'après les données de Sierk (1964) et de Hinds et Hinds (2007) et les observations effectuées sur le terrain.
L'effectif total de l'espèce au Canada est évalué à environ 352 000 individus, répartis entre 76 occurrences. Les occurrences ont été délimitées d'après les distances de séparation définies par NatureServe (2004), qui diffèrent de celle utilisée dans le programme de rétablissement fédéral sur l'espèce (Environment Canada, 2013), soit une distance de séparation de 1 km. Selon les normes de NatureServe, les distances de séparation tiennent compte de facteurs biologiques et physiques, comme la connectivité hydrologique et l'étendue des milieux non propices entre les occurrences.
Trois sous-populations ont été délimitées : 1) basses terres de l'Ontario, 2) Sud du Bouclier, 3) bassin du lac postglaciaire Agassiz. Chaque sous-population possède un ensemble distinct de caractéristiques en ce qui a trait à la géologie et au substrat, à la position géographique, à la couverture terrestre et à l'utilisation des terres (figure 7).
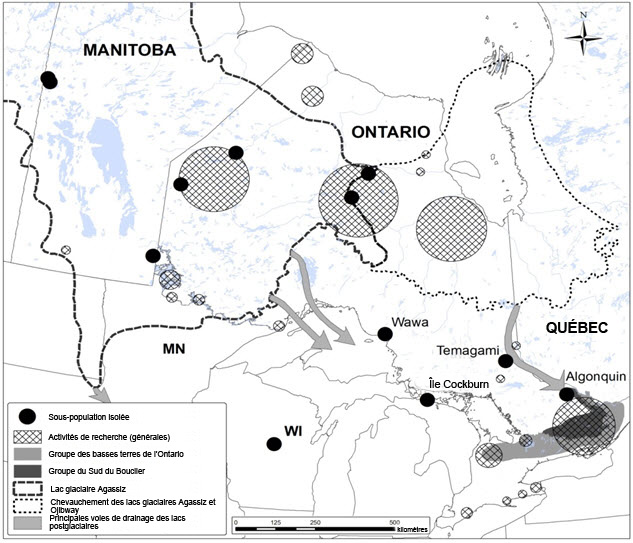
Longue description de la figure 7
Carte montrant la répartition du leptoge des terrains inondés au Canada, divisée en trois groupes : les basses terres de l'Ontario, le Sud du Bouclier et la zone anciennement recouverte par les lacs glaciaires Agassis et Ojibway. Le groupe du Sud du Bouclier compte des occurrences isolées à Wawa, à l'île Cockburn, à Temagami et à Algonquin. Sont également indiqués sur la carte les secteurs où se sont concentrées les activités de recherche visant le leptoge des terrains inondés et l'occurrence des États-Unis qui se trouve la plus près des occurrences canadiennes, située au Wisconsin. Enfin, la carte montre la superficie approximative occupée par le lac postglaciaire Agassiz et sa combinaison possible avec le lac postglaciaire Ojibway ainsi que les principales voies de drainage des lacs postglaciaires.
La sous-population des basses terres de l'Ontario comprend 33 occurrences et environ 190 000 individus, soit approximativement 54 % de la population canadienne totale (voir le tableau 1 ). Cette sous-population occupe le sud des comtés de Grey, de Bruce, de Peterborough, de Hastings et de Lanark, en Ontario, ainsi que la Communauté urbaine de l'Outaouais, au Québec. Elle se trouve sur des formations de calcaire et de dolomie recouvertes par des sols peu profonds ou des dépôts de surface plus épais issus de matériaux parentaux calcaires (notamment des tills pierreux), au sud du Bouclier canadien, dans le centre de l'Ontario. De plus, un certain nombre d'occurrences se trouvent le long de plaines inondables bordant des rivières importantes, y compris les rivières Otonabee et Moira, qui traversent et dont le bassin est constitué de vastes étendues de roche calcaire et/ou de tills calcaires exposés. Dans cette sous-population, le nombre moyen d'individus par occurrence est d'environ 5 700.
| Province | Nom de l'occurrence | cm2 | Nbre d'individus | Frêne présent / dominant |
Menace la plus imminente pesant sur l'occurrence | Protection |
|---|---|---|---|---|---|---|
| Québec | Aylmer | 1-600 | 356 | Oui / Oui | Agrile du frêne | Non |
| Québec | Parc du Lac-Leamy | 4698 | 1-044 | Oui / Non | - | Non |
| Ontario | Parc provincial Balsam Lake | 6-800 | 1-511 | Non / Non | - | Oui |
| Ontario | Chemin Birchview | 3920 | 871 | Oui / Non | - | Non |
| Ontario | Cassidy Block | 157-700 | 35-044 | Oui / Non | - | Non |
| Ontario | Charles Vanderwater C.A. | 16-335 | 3-630 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Coboconk | 30-000 | 6-667 | Oui / Non | Carrière | Non |
| Ontario | Forêt de Ferguson | 12 | 3 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Chemin Grey 19 | 4-200 | 933 | Oui / Non | Développement immobilier | Non |
| Ontario | Chemin Grey 3 | 1-120 | 249 | On ne sait pas | - | Non |
| Ontario | Propriété des Hubert | 5 | 1 | Oui / Non | - | Non |
| Ontario | Chemin Juniper Island | 280 | 62 | Oui / Non | - | Non |
| Ontario | Campus de Kemptville | 696 | 155 | Oui / Non | - | Non |
| Ontario | Kitley | 957 | 213 | Oui / Non | - | Non |
| Ontario | Forêt du comté de Lanark | 6-960 | 1-547 | Oui / Oui | - | Non |
| Ontario | Latta | 2-610 | 580 | Oui / Non | - | Non |
| Ontario | Baie Lost | 350 | 78 | Oui / Non | - | Non |
| Ontario | Forêt Marlborough | 32-712 | 7-269 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Chemin McCrea | 17-400 | 3-867 | Oui / Non | - | Non |
| Ontario | Rivière Mississippi | 696 | 155 | Non / Non | Barrages | Non |
| Ontario | Île Mississippi River | 6 | 1 | On ne sait pas | Barrages | Non |
| Ontario | Rivière Moira | 5-625 | 1-250 | Oui / Non | Barrages | Non |
| Ontario | Rivière Muskrat | 100 | 22 | Oui / Non | - | Non |
| Ontario | Forêt visée par une entente avec l'O.R.C.A. | 8-250 | 1-833 | Oui / Non | - | Non |
| Ontario | Rivière Otonabee | 1-485 | 330 | Oui / Non | Barrages | Non |
| Ontario | Quackenbush | 11-200 | 2-489 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Propriété Quinte-Stoco | 199-320 | 44-293 | Oui / Non | - | Non |
| Ontario | Carrière Rigby | 50-982 | 11-329 | Oui / Non | Carrière | Non |
| Ontario | Propriété des Sheffield | 400 | 89 | Oui / Oui | Parc proposé | Non |
| Ontario | Montée 30 Nord | 2800 | 622 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Hautes terres de South March | 3-480 | 773 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Marécage de Stoney | 280-000 | 62-222 | Oui / Non | Développement | Non |
| Ontario | Parc Allan de la SVCA | 4-060 | 902 | Oui / Oui | Agrile du frêne | Non |
| Total | - | 856-759 | 190-391 | - | - | - |
Cette sous-population comprend 36 occurrences (mais l'occurrence de Wawa, en Ontario, est probablement disparue) et environ 160 000 individus, soit approximativement 45 % de la population canadienne totale (voir le tableau 2 ). Les occurrences se trouvent principalement le long de la limite sud du bouclier Précambrien, près de l'interface avec les basses terres d'âge Paléozoïque (Chapman et Putnam, 1972), mais plusieurs d'elles se trouvent dans une zone où on trouve des sédiments argileux marins laissés par la mer de Tyrrell. Le principal groupe d'occurrences se trouve dans l'est et le centre de l'Ontario, depuis le nord du comté de Peterborough vers l'est jusqu'au nord des comtés de Hastings, Frontenac et Renfrew, avec des occurrences isolées à l'île Cockburn, au parc Algonquin, au lac Temagami et à Wawa (voir figure 7). Dans cette région, on trouve à plusieurs endroits de la roche métasédimentaire exposée riche en composés alcalins, notamment du marbre; la fréquence de cette roche diminue d'est en ouest dans cette région. Dans cette sous-population, le nombre moyen d'individus par occurrence est d'environ 7 500.
| Province | Nom de l'occurrence | cm2 | Nbre d'individus | Frêne présent / dominant |
Menace la plus imminente pesant sur l'occurrence | Protection |
|---|---|---|---|---|---|---|
| Ontario | 511 Nord | 522 | 116 | Oui / Non | - | Non |
| Ontario | Parc provincial Algonquin | 12-375 | 2-750 | Oui / Oui | Agrile du frêne | Oui |
| Ontario | Chemin Arcol | 174 | 39 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Arden | 83 | 18 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Lac Ashby White | ? | ? | On ne sait pas | - | Non |
| Ontario | Lac Billa | 10-370 | 2-304 | Oui / Partiellement | Agrile du frêne | Non |
| Ontario | Bloc 300 | 10-614 | 2-359 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Bloc 306 | 700 | 156 | Oui / Non | - | Non |
| Ontario | Bloc 402 | 4-524 | 1-005 | Oui / Non | - | Non |
| Ontario | Bloc 42 | 8-700 | 1-933 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Lac Blue | 1-000 | 222 | Oui / Non | - | Non |
| Ontario | Île Cockburn | 13-500 | 3-000 | Oui / Non | - | Oui |
| Ontario | Chemin Charlton | 4350 | 967 | Oui / Non | - | Non |
| Ontario | Région du lac Church | ? | ? | Oui / Non | - | Non |
| Ontario | Rivière Crowe | 26-350 | 5-856 | Oui / Partiellement | Agrile du frêne | Non |
| Ontario | Réserve de conservation Crowe River | 300 | 67 | Oui / Non | Barrages | Oui |
| Ontario | Long lac Darling | 420-210 | 93-380 | Oui / Partiellement | Agrile du frêne | Non |
| Ontario | Réserve naturelle provinciale Egan Chutes | 950 | 211 | Oui / Non | - | Oui |
| Ontario | Bloc forestier 40 | 348 | 77 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Parc provincial Frontenac | 825 | 183 | Oui / Non | - | Oui |
| Ontario | High Lonesome | 24-780 | 5-507 | Oui / Partiellement | - | Non |
| Ontario | Ruisseau Indian | 5 | 1 | Oui / Non | - | Non |
| Ontario | Lake Twp Ponds | 12-914 | 2-870 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Lavant | 23-580 | 5-240 | Oui / Partiellement | Agrile du frêne | Non |
| Ontario | Parc provincial Murphys Point | 26-625 | 5-917 | Oui / Partiellement | Agrile du frêne | Oui |
| Ontario | Lac Park | 9-457 | 2-102 | Oui / Non | - | Non |
| Ontario | Chemin Rapids | 10-312 | 2-292 | Oui / Partiellement | Agrile du frêne | Non |
| Ontario | Propriété des Smith | 17-400 | 3-867 | Oui / Non | - | Non |
| Ontario | Sherbrooke Sud | 10-850 | 2-411 | Oui / Non | - | Non |
| Ontario | Chemin Storms | 42-900 | 9-533 | Oui / Non | - | Non |
| Ontario | Tatlock | 250 | 56 | Oui / Non | Développement | Non |
| Ontario | Rivière Tay | 2 | 0 | Oui / Non | - | Non |
| Ontario | Temagami | 5220 | 1-160 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Wawa * | ? | ? | Oui / Oui | Barrage | Non |
| Ontario | Lac White | 19-360 | 4-302 | Oui / Oui | Agrile du frêne | Non |
| Ontario | Parc provincial Wolf Island | 1-400 | 311 | Oui / Oui | Agrile du frêne | Oui |
| Total | - | 720-950 | 160-212 | - | - | - |
Cette sous-population comprend 7 occurrences et environ 2 000 individus, soit approximativement 1 % de la population canadienne totale, et occupe un territoire allant des alentours de Flin Flon, dans le nord du Manitoba, jusqu'à la rivière Attawapiskat, 1 000 km plus à l'est, dans le nord de l'Ontario, et vers le sud jusqu'au lac Sailing, dans le sud-est du Manitoba (voir le tableau 3 ). La majeure partie de cette région est occupée par des sols podzoliques et brunisoliques reposant sur la roche-mère du Précambrien (Crins et al., 2009) et comprenant des matériaux parentaux siliceux ainsi que d'importants dépôts de tourbe. Ces conditions ne sont pas représentatives de l'habitat habituel du L. rivulare, en raison de leur acidité relative élevée. Toutefois, dans cette région, de vastes superficies ont été recouvertes par les lacs postglaciaires Agassiz et Ojibway, qui ont laissé à certains endroits des dépôts de limons calcaires glaciolacustes, d'argile et de moraines calcaires sableuses à loameuses contenant des galets (Crins et al., 2009). Les dépôts calcaires riches en composés alcalins qui se trouvent au fond des plans d'eau existant aujourd'hui ou sont exposés en bordure des lacs ou des voies interlacustres et des rivières ont une incidence sur la composition chimique de l'eau et en augmentent l'alcalinité et les concentrations en calcium; ces dépôts créent et maintiennent donc des conditions propices à l'espèce. Dans cette sous-population, le nombre moyen d'individus par occurrence est d'environ 200.
| Province | Nom de l'occurrence | cm2 | Nbre d'individus | Menace la plus imminente pesant sur l'occurrence | Protection |
|---|---|---|---|---|---|
| Ontario | Otoskwin-rivière Attawapiskat 1 | 7-425 | 1650 | - | Oui |
| Ontario | Otoskwin-rivière Attawapiskat 2 | 990 | 220 | - | Oui |
| Ontario | Rivière Severn | 300 | 67 | Projet de barrage | Non |
| Ontario | Lac Stout-embouchure de la rivière Crooked | 10 | 2 | - | Non |
| Ontario | Manitoba | - | - | - | - |
| Ontario | Tri-Lakes (lacs Payuk, Twin et Neso) | 36,5 | 8 | Concessions minières | Non |
| Ontario | Lac Whitefish | 13,5 | 3 | Concessions minières | Non |
| Ontario | Lac Sailing | 6 | 27 | - | Oui |
| Total | - | 8-781 | 1-977 | - | - |
On ne dispose d'aucune donnée quantitative pour les sites historiques, sauf dans le cas de l'occurrence historique de Wawa, en Ontario, signalée pour la première fois par Fabius LeBlanc en 1965. Vu le manque de données historiques et le grand nombre d'occurrences récemment découvertes, il est impossible d'évaluer le taux de déclin de la population. Cependant, des déclins sont prévus après les infestations d'agrile du frêne dans 20 occurrences dominées par le frêne, qui représentent 10 % de la population canadienne totale, ainsi que dans 7 autres occurrences où le frêne est une espèce codominante importante, qui représentent 34 % additionnels de la population canadienne totale ( tableau 2 ). Même si le taux de mortalité ne sera pas nécessairement de 100 % dans ces occurrences, on prévoit que l'agrile du frêne constituera un risque pour approximativement 44 % de la population canadienne au cours des 10 à 20 prochaines années.
La rareté apparente du L. rivulare aux États-Unis pourrait limiter la possibilité d'une immigration de source externe si la population canadienne connaissait un déclin. Il est probable que seulement une des trois occurrences des États-Unis existe actuellement, les deux autres n'ayant pas été observées depuis le 19e siècle. En outre, une grande distance sépare les occurrences canadiennes de l'occurrence des États-Unis la plus proche, qui se trouve dans le centre du Wisconsin (Bennet et Bartkowiak, 2013); elle se situe à environ 670 km à l'ouest de l'occurrence canadienne la plus proche, située en Ontario, et à 745 km au sud de l'occurrence manitobaine la plus proche. De vastes relevés lichénologiques ont été menés au Minnesota, dans la région adjacente au Canada, notamment par Wetmore (comm. pers., 2012) dans la région des lacs de la zone frontalière, mais aucune population de L. rivulare n'a été trouvée. Donc, il est possible, mais peu probable, qu'une immigration de source externe se produise.
La présence du L. rivulare au Canada est limitée par le besoin de l'espèce de milieux périodiquement inondés et partiellement ombragés à substrat riche en composés alcalins ou à substrat recevant un apport en calcium (figure 5).Dans le nord de l'Ontario et du Manitoba, le L. rivulare semble absent de la majeure partie de l'écozone boréale, car le substrat y est généralement trop acide vu la présence de roche-mère du Précambrien et de sols podzoliques et brunisoliques issus de matériaux parentaux siliceux (Crins et al., 2009). Malheureusement, il n'existe aucune carte à petite échelle de la géologie de surface ou de la végétation qui permettrait de cibler les régions du nord du Manitoba ou de l'Ontario où la composition chimique du sol et la couverture végétale pourraient être propices au L. rivulare. Les cartes physiques existantes sont trop brutes. Les affleurements calcaires sont très rares dans les régions nordiques (du moins en Ontario). De plus, les régions où on trouve des dépôts glaciaires calcaires n'ont pas été cartographiées en détail et ne sont pas accessibles par la route. D'après les travaux de terrain réalisés jusqu'à maintenant, le L. rivulare ne se rencontre que dans des localités très éparpillées sur un vaste territoire dans le nord du Manitoba et de l'Ontario et est rare dans cette région comparativement au sud, où les deux sous-populations comptent un effectif élevé.
D'après le calculateur des menaces du COSEPAC, l'impact global des menaces sur la survie du L. rivulare est élevé (voir l'annexe 1). Les principales menaces pesant sur le L. rivulare sont présentées ci-dessous.
La menace la plus grave et la plus imminente pesant sur le L. rivulare est associée à l'agrile du frêne (Agrilus planipennis), coléoptère xylophage exotique (Coleoptera: Buprestidae) originaire d'Extrême-Orient et arrivé en Amérique du Nord au début des années 1990. Il a été détecté pour la première fois à Détroit, au Michigan, et à Windsor, en Ontario, durant l'été 2002. Depuis, il s'est propagé dans 19 États ainsi que dans le sud de l'Ontario et du Québec (figure 8) (Paiero et al. 2012, EAB 2013). Les individus adultes ne causent aucun dommage à long terme aux arbres, mais les larves consomment l'écorce interne (phloème) de toutes les espèces indigènes de frênes (Fraxinus spp.), ce qui perturbe la capacité des arbres de transporter l'eau et les éléments nutritifs et finit par tuer les arbres hôtes infestés. En Amérique du Nord, les frênes meurent généralement 5 à 7 ans après le début de l'infestation. Les jeunes arbres peuvent mourir en 1 ou 2 ans (Knight et al., 2013). Depuis qu'il a été détecté pour la première fois, l'agrile du frêne a causé la mort de dizaines de millions de frênes, hôtes importants pour le L. rivulare en Ontario et au Québec, et soulève des craintes quant à l'avenir des frênes en Amérique du Nord (Poland et McCullough, 2006).
Au Canada, le frêne constitue l'espèce (substrat) dominante de la strate arborée dans 20 occurrences existantes de L. rivulare (10 dans la sous-population des basses terres de l'Ontario et 10 dans celle du Sud du Bouclier), et il est partiellement dominant (occupe environ la moitié de la strate arborée) dans 7 autres occurrences, toutes situées dans la sous-population du Sud du Bouclier. Ces 27 occurrences comptent environ 44 % du nombre total d'individus matures au Canada (10 % provenant d'occurrences où le frêne est l'hôte dominant, et 34 %, d'occurrences où le frêne est partiellement dominant). En fait, 99 % des thalles connus sont associés à des communautés végétales où le frêne est présent. L'impact de l'agrile du frêne sur le L. rivulare au cours des trois prochaines générations variera probablement d'un site à l'autre, en fonction de la prédominance du frêne dans l'habitat (Jönsson et Thor, 2012). Le frêne est un des arbres possédant l'écorce la plus riche en composés alcalins, et son écorce a une texture rugueuse particulière. En outre, il tolère les inondations et prospère dans les sols calcaires. Si les frênes disparaissent, d'autres hôtes pourront servir de substrat au L. rivulare, principalement les érables. Cependant, l'écorce des érables semble posséder un pH plus bas que celle des frênes, en plus d'avoir une moins bonne capacité de rétention de l'humidité et de présenter une plus faible diversité lichénique (Brinker, comm. pers., 2015; BLS, 2015). Ainsi, la disparition des frênes entraînerait un déclin considérable de la population totale de L. rivulare, ce qui serait grave, car le Canada compte de loin la plus grande population de l'espèce dans le monde.
Il est difficile de lutter contre l'agrile du frêne, car l'espèce passe la majeure partie de son cycle vital cachée sous l'écorce de ses arbres hôtes (Wang et al., 2010). La position officielle du Canada est que l'agrile du frêne ne peut pas être éradiqué (Marchant, 2007). L'insecte ne s'attaque pas aux semis de moins de 2,5 cm de diamètre, mais ceuxci ne peuvent pas fournir des superficies de substrat convenable ou les conditions microclimatiques que les grands arbres offrent au L. rivulare. Les infestations d'agrile du frêne débutent à petite échelle, s'étendent, puis ralentissent à mesure que le nombre d'arbres hôtes vivants diminue. Certaines pratiques sylvicoles, comme l'éclaircissement de faible intensité destiné à réduire le pourcentage global de frênes dans la strate arborée et à améliorer la croissance des arbres, pourraient réduire la vitesse de propagation (Streit et al., 2012) de l'agrile du frêne et donc profiter au L. rivulare. Cependant, il n'existe aucune donnée expérimentale pour confirmer cette théorie.
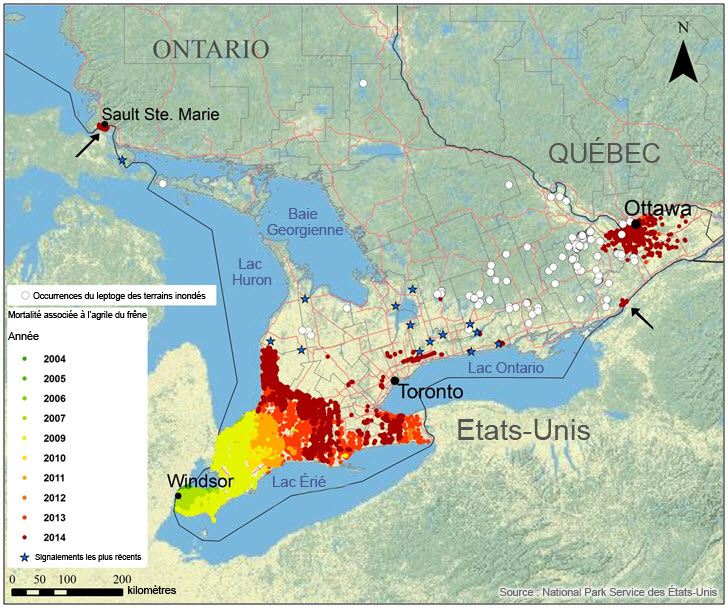
Longue description de la figure 8
Carte montrant la propagation annuelle de l'agrile du frêne dans le sud de l'Ontario, en fonction des peuplements de frênes présentant un taux de mortalité modéré à élevé causé par cet insecte. Les occurrences de leptoge des terrains inondés sont également indiquées sur la carte.
La vitesse de progression du front de l'infestation d'agrile du frêne varie d'un endroit à l'autre, mais dans les régions peuplées, cette vitesse est accrue par le déplacement par les humains de bois infesté. La vitesse de propagation moyenne a été évaluée à 10,6 km par an sur une période de 4 ans au Michigan, (Smitley et al., 2008), à jusqu'à 20 km par an sur une période de 10 ans en Ontario (P. Hodge, comm. pers., 2015), et à 30 km par an sur une période de 4 ans à Moscou (Straw et al., 2013). Selon ces vitesses de propagation, on peut penser que les 27 occurrences de L. rivulare qui se trouvent dans des peuplements forestiers où le frêne est une composante importante seront probablement touchées par des infestations d'agrile du frêne d'ici les 10 à 20 prochaines années et que la mort des arbres surviendra 5 à 7 ans après le début de l'infestation. D'après des études sur le vol de l'agrile du frêne, les individus adultes ont une capacité de dispersion de jusqu'à 5 km (Cappaert et al., 2005). Le front de l'infestation d'agrile du frêne peut se trouver à 30 km ou plus de la zone où l'insecte peut être détecté de façon fiable par des relevés visuels (Marchant, 2007). La figure 8 montre la progression de l'agrile du frêne dans le sud de l'Ontario de 2004 à 2014. Actuellement, sept occurrences de L. rivulare se trouvent à 10 km ou moins de peuplements où l'agrile du frêne a causé une mortalité modérée à élevée des frênes.
Le L. rivulare pousse couramment sur l'orme d'Amérique, qui est très sensible à la maladie hollandaise de l'orme, causée par l'Ophiostoma ulmi, champignon introduit. Le champignon est propagé par le scolyte de l'orme (Hylurgopinus rufipes) et le scolyte européen de l'orme (Scolytus multistriatus). La maladie hollandaise de l'orme a causé des taux de mortalité élevés dans l'ensemble de l'aire de répartition de l'orme d'Amérique. Les thalles de L. rivulare présents sur des ormes qui succombent à la maladie finissent par disparaître. La maladie hollandaise de l'orme a été signalée pour la première fois en Ontario il y a plus de cinquante ans. Les drageons ainsi que les semis issus des arbres infectés font en sorte que l'orme demeure un arbre commun. Cependant, lorsqu'ils atteignent un certain stade de maturité, ils sont réinfectés par le champignon. Généralement, les arbres sont infectés avant que leur écorce soit suffisamment crevassée pour pouvoir être colonisée par le L. rivulare. Cette modification de la dynamique de la population pourrait avoir une incidence sur les populations de L. rivulare, mais on ignore dans quelle mesure.
La consommation des thalles par les limaces du complexe d'espèces d'Arion subfuscus, espèces de taille moyenne probablement introduites d'Europe, constitue une menace pour les lichens dans l'est du Canada (Cameron, 2009) ainsi que pour le L. rivulare dans l'est de l'Ontario d'après le programme de rétablissement provincial sur l'espèce. Selon des observations, ces limaces consomment de préférence les apothécies du L. rivulare (Environment Canada, 2013). Diverses espèces du genre Arion, notamment les espèces du complexe d'A. subfuscus, se sont introduites dans des milieux situés à l'intérieur de l'aire de répartition canadienne du L. rivulare. Ce complexe pourrait comprendre au moins deux espèces cryptiques, l'A. fuscus et l'A. subfuscus, et leur répartition n'est pas bien comprise (Grimm et al., 2009).
En Ontario et au Québec, l'A. subfuscus oul'A. fuscus ainsi que des limaces indigènes des genres Philomycus et Pallifera ont été signalées dans le cadre de récents travaux d'identification. Il ne semble pas exister de données sur les préférences alimentaires précises de ces limaces en ce qui a trait aux lichens, mais les limaces du genre Arion sont des généralistes, et leur régime alimentaire comprend une diversité de champignons et de matières végétales mortes ou vivantes (Beyer et Saari, 1978; R. Forsyth, comm. pers., 2015). De nombreux nouveaux sites hébergeant le L. rivulare ont été signalés au cours des quatre dernières années, mais les observateurs n'ont pas accordé d'attention à l'abondance des limaces et aux dommages qu'elles pouvaient avoir causés. Des études doivent donc être réalisées à ce sujet.
Le Lyngbya wollei a été signalé pour la première fois au Manitoba durant les années 1990, dans plusieurs lacs (Government of Manitoba, 2007). Cette algue exotique forme d'épais tapis sur le fond des lacs ou des tapis flottants. Elle est collante, ce qui favorise sa dispersion par divers agents auxquels elle adhère (par exemple des remorques, des bateaux et des VTT) et lui permet de recouvrir les rivages rocheux. L'établissement de cette espèce dans les régions où le Leptogium rivulare est présent est préoccupant. Il faut réaliser d'autres travaux pour mieux connaître l'étendue de cette menace dans les sites hébergeant le L. rivulare au Manitoba. L'algue a été signalée dans le parc provincial du Whiteshell, mais sa présence à l'occurrence de L. rivulare du lac Sailing n'a pas été confirmée. Ainsi, elle ne constitue pas une menace immédiate pour le L. rivulare au Manitoba (Bazin, comm. pers., 2012).
Au cours des cent prochaines années, on prévoit que le changement climatique entraînera une hausse des températures moyennes à l'échelle planétaire par rapport à celles connues au cours des quelques derniers millions d'années (Crowley, 1990). Au Canada, les 10 années les plus chaudes ont toutes été enregistrées depuis 1998, et la décennie 2001-2010 a été la plus chaude jamais enregistrée (Feltmate et Thistlethwaite, 2013). Plus particulièrement dans le sud de l'Ontario et du Québec, on prévoit que la hausse des températures entraînera un assèchement des conditions, surtout durant les mois d'été, ce qui causera une diminution des niveaux d'eau dans la région des Grands Lacs (Feltmate et Thistlethwaite, 2013). On s'attend à ce que les enveloppes climatiques régionales (régions possédant des régimes climatiques semblables) subissent des changements radicaux, notamment en Ontario d'après la modélisation du climat réalisée par McKenney et al. (2010). Selon ces scénarios, les espèces ayant des exigences très précises en matière d'habitat et une capacité de dispersion limitée, comme le L. rivulare, seront plus vulnérables que les espèces généralistes quant à l'habitat ou que celles qui possèdent une bonne capacité de dispersion.
Le L. rivulare a besoin d'être immergé de manière périodique, ce qui le rend vulnérable aux dommages physiques causés par les phénomènes météorologiques extrêmes, comme les inondations et l'érosion par la glace, qui deviendront de plus en plus courants selon les prévisions. Par exemple, en 2004 et 2006 dans un site de l'est de l'Ontario, plusieurs mares printanières étaient encore remplies d'eau en octobre, alors qu'elles sont généralement asséchées à ce moment de l'année, en raison des précipitations automnales anormalement abondantes reçues juste avant la formation de la glace. Peu après, une épaisse couche de glace (10 à 20 cm) s'est formée, et la partie inférieure du tronc de nombreux arbres hébergeant le L. rivulare est ainsi restée immergée. Durant ces deux hivers, le niveau de l'eau a baissé à la mi-saison, de sorte que la glace n'était plus soutenue et s'est affaissée en frottant sur l'écorce, réduisant ainsi la couverture de mousses et de lichens. Chez certains arbres immergés sur une grande hauteur, jusqu'à 90 % de la couverture de L. rivulare a ainsi été détruite (Lewis, obs. pers., 2015).
La vulnérabilité des espèces au changement climatique en Ontario (Brinker et Jones, données inédites) est actuellement évaluée au moyen de l'Indice de vulnérabilité au changement climatique de NatureServe (Young et al., 2010). Selon les résultats préliminaires obtenus, le L. rivulare serait modérément vulnérable aux effets des changements climatiques futurs en Ontario. Les principaux facteurs participant à sa vulnérabilité sont les suivants : 1) sa niche hydrologique spécialisée, car environ 80 % des occurrences sont associées à des mares printanières, qui devraient subir un assèchement et être moins fréquemment inondées à cause du changement climatique; 2) sa capacité de dispersion limitée et le fait qu'environ 20 % de la population de l'Ontario se trouve dans des paysages fortement modifiés par l'activité humaine; 3) sa dépendance à l'égard d'un régime de perturbation précis (inondations) pour le maintien de son habitat, car la fréquence et l'étendue de ces perturbations devraient changer à cause du changement climatique. La vulnérabilité des deux occurrences du Québec est semblable à celle des occurrences de l'Ontario, car elles se trouvent près les unes des autres, dans des habitats similaires. Les occurrences du nord de l'Ontario et du Manitoba pourraient être moins vulnérables, mais elles représentent actuellement moins de 1 % de la population canadienne totale.
La manipulation ou la régulation des niveaux d'eau qui modifient les régimes d'inondations et la profondeur de l'eau peuvent dégrader l'habitat du L. rivulare ainsi que la capacité physique de dispersion et de reproduction de l'espèce. Les barrages destinés à la production d'hydroélectricité ou à la régulation des niveaux d'eau et à la lutte contre les inondations sont fréquents dans la majeure partie de l'aire de répartition du L. rivulare, et la demande en hydroélectricité augmente avec la croissance de la population humaine et le désir de réduire la dépendance envers les combustibles fossiles. En Ontario, le barrage hydroélectrique construit sur la rivière Magpie a probablement grandement contribué à la disparition du L. rivulare dans le site de Wawa. Le barrage des chutes Steephill a été construit de 1911 à 1913, puis abandonné en 1927. Un nouveau barrage hydroélectrique dont la construction a été achevée en 1991 a mené à la création d'un réservoir qui a modifié la dynamique des crues de la rivière. Les crues naturelles ont pratiquement disparu en aval du barrage, ce qui a causé une diminution de la végétation riveraine (S. Brinker, obs. pers.). Les barrages constituent actuellement une menace pour plusieurs autres sites existants bordant des rivières en Ontario, y compris les sites des rivières Crowe, Otonobee, Moira et Mississippi, où le nombre d'individus est plus faible que dans les régions voisines où les crues ne font pas l'objet d'une régulation. Une occurrence située en bordure de la rivière Severn, dans le nord de l'Ontario, pourrait être touchée par un projet de barrage (D. Berube, comm. pers., 2012).
D'autres structures entraînant une modification des régimes d'inondation ou des niveaux d'eau, comme les ponceaux qui modifient la direction du ruissellement de surface ou les ouvrages de franchissement tels les routes ou les sentiers traversant des milieux humides, peuvent constituer des menaces, car elles fragmentent les milieux humides et modifient les hydropériodes et les débits.
En Ontario, les activités d'exploitation forestière qui éliminent des arbres à l'intérieur ou autour des mares printanières peuvent modifier les processus hydrologiques des mares et réduire les taux d'humidité à l'échelle locale, en plus de réduire les quantités de substrat disponibles si les arbres coupés sont des arbres hôtes. Le frêne rouge est une essence commerciale, mais d'autres arbres hôtes, comme le frêne noir et l'érable rouge, sont rarement ciblés pour l'exploitation. En outre, la construction de routes associée aux activités forestières peut endommager l'habitat si le tracé traverse les mares printanières ou passe trop près de celles-ci. Les sites qui se trouvent sur des terres de la Couronne sont protégés par une zone tampon de 30 m entourant les mares boisées, conformément au Forest Management Guide for conserving Biodiversity at the Stand and Site Scales de l'Ontario (OMNR 2010). Toutefois, la délimitation des mares peut être incorrecte si elle n'est pas réalisée à la bonne saison, et cette zone ne s'applique pas dans le cas des terres privées.
Les activités forestières réalisées sur les terres de la Couronne doivent respecter une orientation générale (présentée dans le Stand and Site Guide; OMNR, 2010) pour l'exploitation dans les marécages à frênes noirs, les mares boisées, les sources, les zones de suintement et les baissières ainsi qu'en ce qui a trait aux perturbations et aux routes. Ainsi, de manière générale, il y a peu de risque que ces activités aient des répercussions négatives sur des populations de L. rivulare non répertoriées dans des mares boisées. Le ministère des Richesses naturelles et des Forêts (MRNF) examine chaque année environ 150 demandes d'exploitation forestière visant des terres de la Couronne à l'intérieur de l'aire de milieux propices au L. rivulare (Cameron, comm. pers., 2013). Le personnel du MRNF doit alors réaliser des relevés de terrain pour déterminer si le L. rivulare est présent; si l'espèce est découverte, l'habitat occupé doit être délimité et protégé (voir OMNR, 2010, page 21 – Habitat Protection and Ownership). Puisque les activités forestières doivent respecter une orientation générale (présentée dans le Stand and Site Guide; OMNR, 2010) pour l'exploitation dans les marécages à frênes noirs, les mares boisées, les sources, les zones de suintement et les baissières ainsi qu'en ce qui a trait aux perturbations et aux routes, il y a peu de risques qu'un peuplement forestier colonisé par le L. rivulare soit coupé. Cependant, il est presque impossible pour le personnel de réaliser des relevés dans tous les nombreux sites exploités chaque année. Toutefois, les risques sont les plus élevés dans le cas des terrains privés, qui ne font pas l'objet de relevés de terrain du MRNF.
La qualité de l'eau de la plupart des ruisseaux et rivières du sud de l'Ontario est dégradée, ce qui contribue à l'absence apparente du L. rivulare au sud d'une ligne délimitant le sud des comtés de Hastings, de Peterborough et de Grey. Dans cette région, le ruissellement en milieux urbain et agricole a détérioré la qualité de l'eau. La hausse de la température de l'eau, les modifications de la composition chimique de l'eau et l'augmentation des charges de sédiments sont néfastes pour le L. rivulare. Si les eaux d'une rivière ou d'un lac ont une charge excessive de sédiments issus du ruissellement agricole, elles peuvent laisser après le retrait de l'eau un dépôt de limon sur le tronc des arbres et les roches dans les plaines inondables, ce qui nuit au L. rivulare (COSEWIC, 2004). La pollution de source ponctuelle ou de source diffuse qui entraîne une hausse des concentrations de phosphore, de nitrates, de métaux lourds et de sel constitue une menace pour les populations de L. rivulare, alors que l'eutrophisation de l'eau cause des proliférations d'algues et une modification du pH.
Les occurrences de Leptogiumn rivulare font l'objet d'une pression à des fins de développement, particulièrement dans le canton de Darling, en Ontario, où les propriétés privées sont subdivisées et aménagées, réduisant ainsi l'habitat de l'espèce (COSEWIC, 2004). À Stony Swamp, les activités d'utilisation des terres associées à l'expansion des milieux urbains et des infrastructures routières connexes a des répercussions négatives sur les régimes hydrologiques des étangs saisonniers qui se trouvent à l'extérieur des terrains gérés par la Commission de la capitale nationale (E. Katic, comm. pers., 2015). Un site est menacé par un projet immobilier dans le comté de Grey.
L'élimination d'arbres ou de roches pour le développement industriel ou récréatif (par exemple, aménagement de sentiers) et les activités d'arpentage (par exemple, dégagement d'une ligne de visée) entraînent également une perte d'habitat pour le L. rivulare. La construction de chalets près de certains lacs (Payuk, Twin et Whitefishs) au Manitoba pourrait avoir un impact sur les occurrences de L. rivulare situées sur le rivage de ces lacs, mais peu de nouvelles constructions ont été observées (Bazin, comm. pers., 2012).
L'altération des milieux et la perte d'habitat causées par les activités d'extraction d'agrégats (y compris les sablières, les gravières et les carrières de calcaire) menacent le L. rivulare et son habitat. De nombreuses occurrences existantes se trouvent sur des formations de calcaire et de marbre, qui sont d'importantes sources d'agrégats en Ontario. Des occurrences se trouvant sur des arbres dans un site près du Long lac Darling ont été éliminées par les activités d'une sablière privée (COSEWIC, 2004). De plus, des arbres colonisés se trouvent à l'intérieur des limites d'exploitation prévues de plusieurs carrières près de Buckhorn et de Coboconk, en Ontario (Lewis, obs. pers.). Au Manitoba, l'ensemble des lacs Payuk, Twin et Whitefishs et environ la moitié du lac Neso sont visés par des concessions minières (Bazin, comm. pers., 2012), et celles-ci menacent l'habitat de l'espèce. Cependant, même si la demande pour le calcaire de qualité est croissante, tous les projets pouvant menacer des sites connus hébergeant le L. rivulare en Ontario sont refusés par le MRNF.
Les cyanolichens, dont le L. rivulare, sont extrêmement sensibles aux effets de la pollution atmosphérique, particulièrement au dioxyde de soufre et aux pluies acides (COSEWIC, 2004), qui peuvent entraîner une diminution du pH de l'eau et du substrat. En Nouvelle-Angleterre, le L. rivulare aurait connu un déclin à cause de la pollution et de la destruction de son habitat (Hinds et Hinds, 2007). La disparition du L. rivulare au site de Wawa est probablement attribuable à une modification du niveau de l'eau (voir précédemment) combinée aux effets des panaches de dioxyde de soufre issus de l'usine d'Algoma Ore, qui faisait la transformation du minerai de sidérite de 1939 à 1998. Au Manitoba, il est probable que les occurrences de L. rivulare situées près de Flin Flon seront touchées par la pollution issue des activités minières et des fonderies. Ces activités ont dénudé le paysage et diminué le pH du substrat sur une vaste superficie. Les tendances associées aux polluants acidifiants sont présentées dans une série de rapports (voir par exemple Canadian Council of Ministers of the Environment, 2013).
Les localités du COSEPAC sont des zones particulières du point de vue géographique et écologique dans lesquelles un seul phénomène menaçant peut affecter rapidement tous les individus d'une occurrence. Une série de menaces pèsent actuellement sur l'espèce, dont l'agrile du frêne, le changement climatique et les sécheresses, la manipulation des niveaux d'eau, la pression associée au développement urbain et industriel ainsi que l'extraction d'agrégats. Il est impossible de prédire avec exactitude quelles occurrences seront touchées par un seul phénomène menaçant à un moment ou à un autre. Toutefois, globalement, le nombre total de localités peut être évalué à 37 d'après les menaces qui suivent. Agrile du frêne : Québec, Aylmer; Ontario, parc Algonquin, chemin Arcol, Arden, lac Billa, bloc 300, bloc 42, Charles Vanderwater C.A., rivière Crowe, forêt de Ferguson, bloc forestier 40, High Lonesome, Lake Twp. Ponds, forêt du comté de Lanark, Lavant, forêt Marlborough, pointe Murphys, Quackenbush (excluant le parc provincial), chemin Rapids, montée 30 Nord, hautes terres de South March, parc Allan de la SVCA, Wawa, lac White, île Wolf. Barrages : Ontario, réserve de conservation Crowe River, rivière Mississippi, île Mississippi River, rivière Moira, Wawa. Extraction d'agrégats : Ontario, Coboconk, Rigby; Manitoba, TriLakes (lacs Payuk, Neso et Twin). Expansion urbaine/industrielle : Ontario, chemin Grey 19, Tatlock, marécage de Stoney.
Les menaces pesant sur chaque occurrence sont indiquées dans les tableaux 1 à 3. La menace est la même pour plusieurs occurrences (agrile du frêne), mais il est peu probable que celle-ci touche plusieurs occurrences au cours d'une même année. Ainsi, on considère que le nombre total de localités connues est de 37.
L'inscription du Leptogium rivulare à la liste rouge mondiale a été proposée en janvier 2015 (IUCN, 2015), et NatureServe a attribué à l'espèce la cote G3 (vulnérable)/G5 (non en péril) à l'échelle mondiale (NatureServe, 2013). Le L. rivulare est inscrit à la liste rouge de la Suède et est considéré comme fortement menacé dans ce pays (ArtDatabanken, 2013). En Estonie, le L. rivulare est jugé comme gravement menacé d'extinction (eBiodiversity, 2013).
Au Canada, le Leptogium rivulare est coté N3 (vulnérable) à l'échelle nationale. Il a initialement été désigné « espèce menacée » au Canada par le COSEPAC en 2004 (COSEWIC, 2004). Le L. rivulare été inscrit à l'annexe 1 (partie 3) de la Loi sur les espèces en péril fédérale. Un programme de rétablissement fédéral a été finalisé en 2013 (Environment Canada, 2013). Celui-ci comprenait une première désignation de l'habitat essentiel pour tous les sites colonisés existants, soit au moment de la publication 40 sites répartis entre 21 occurrences en Ontario et 15 sites répartis entre 8 occurrences au Manitoba.
Ontario
Le Leptogiumn rivulare est actuellement désigné menacé aux termes de la Loi de 2007 sur les espèces en voie de disparition de la province. En vertu de cette loi, l'espèce et son habitat bénéficient d'une protection depuis le 30 juin 2013. En outre, la Loi sur les parcs provinciaux et les réserves de conservation (2006) confère au L. rivulare une protection dans les parcs provinciaux et les réserves de conservation.
Manitoba
Le Leptogiumn rivulare ne figure pas sur la liste des espèces en péril établie en vertu de la Loi sur les espèces en voie de disparition du Manitoba. L'occurrence du lac Sailing se trouve à l'intérieur du parc provincial du Whiteshell et est donc protégée aux termes de la Loi sur les parcs provinciaux (2013) du Manitoba.
Québec
Le L. rivulare ne bénéficie d'aucune protection juridique provinciale au Québec.
NatureServe (2013) a attribué les cotes infranationales suivantes au L. rivulare : S1 (gravement en péril) au Manitoba et en Ontario. La situation de l'espèce au Québec n'a pas encore été évaluée, car elle n'a été découverte que récemment dans cette province, et elle n'a été évaluée pour aucun État des États-Unis.
La situation générale du L. rivulare au Canada a été évaluée en 2010, et l'espèce a été désignée « en péril » en Ontario et au Manitoba ainsi qu'à l'échelle du Canada (CESCC, 2010).
Des 76 occurrences de L. rivulare, 9 bénéficient actuellement d'une protection, car elles se trouvent à l'intérieur de parcs provinciaux ou de réserves de conservation. L'occurrence du lac Sailing, au Manitoba, est située à l'intérieur du parc provincial Whiteshell. En Ontario, une occurrence se trouve dans la réserve de conservation de Crowe River, et les huit autres, dans les parcs provinciaux suivants : Algonquin, Balsam Lake, Egan Chutes, Frontenac, Murphys Point, Otoskwin-rivière Attawapiskat et Wolf Island.
Au Québec, l'occurrence d'Aylmer se trouve sur des terrains appartenant à la ville de Gatineau, et aucun développement n'y est actuellement autorisé, même si l'espèce n'y bénéficie d'aucune protection officielle. L'occurrence du lac Leamy est située à l'intérieur d'un parc de la Commission de la capitale nationale et est donc indirectement protégée par la planification de l'aménagement qui y est réalisée.
En Ontario, la majorité des autres occurrences se trouvent sur des terres de la Couronne ou des terrains municipaux et pourraient être protégées si elles étaient inscrites dans des processus officiels de planification. Selon le processus d'application de la Loi sur les ressources en agrégats de l'Ontario, la préparation d'un rapport sur l'environnement naturelest exigée, y compris l'évaluation de la présence d'espèces en péril ou d'autres espèces d'importance devant peut-être être protégées. En outre, selon l'Énoncé de principe provincial de la Loi sur l'aménagement du territoire de l'Ontario, aucun développement n'est autorisé à l'intérieur de l'habitat d'importance d'une espèce en voie de disparition ou menacée. Les municipalités reçoivent des demandes de rezonage, de divisibilité (demande de consentement) ou de permis de construction, et des études d'impact environnemental doivent souvent être réalisées.
Si le L. rivulare est trouvé dans le cadre des relevés de terrain, il pourrait être protégé par l'application de conditions ou le rejet de la demande. Conformément à la Loi sur les terres publiques, la cession de terres de la Couronne, l'aménagement de nouvelles routes ou de nouveaux sentiers ou la délivrance de permis d'utilisation des terres doivent être évalués par le ministère des Richesses naturelles et des Forêts dans le cadre d'une évaluation environnementale de portée générale relative à des projets d'intendance de ressources et de développement d'installations. Dans le cadre de ce processus, le projet est examiné par un biologiste des espèces en péril, qui peut demander la réalisation d'une évaluation des impacts. Le L. rivulare pourrait être trouvé durant les travaux de terrains réalisés pour cette évaluation, et il serait alors protégé conformément aux dispositions de la Loi sur les espèces en péril ou par le rejet de la demande.
En Ontario, l'Ontario Forest Management Guide for conserving Biodiversity at the Stand and Site Scales (OMNR, 2010) s'applique dans le cas les terres de la Couronne qui sont visées par des activités forestières et où le L. rivulare est présent, et il impose des restrictions relatives aux routes à l'échelle du secteur préoccupant, qui est plus vaste et peut englober de multiples mares et les portions de forêt les reliant. Pour protéger les espèces, ces lignes directrices (OMNR, 2010) prescrivent un secteur préoccupant correspondant à une zone d'un rayon de 30 m où la coupe est interdite autour de toute mare boisée hébergeant le L. rivulare. En outre, la forêt résiduelle doit être conservée dans une zone d'un rayon de 15 m autour des mares boisées adjacentes qui n'hébergent pas le L. rivulare.
Au Québec, toute activité réalisée dans les marécages ou sur les rivages de lacs et de cours d'eau est régie par le paragraphe 2 de l'article 22 de la Loi sur la qualité de l'environnement. Une analyse doit être réalisée avant que tous travaux puissent être exécutés dans un tel milieu, et un certificat d'autorisation doit être délivré. En outre, des lignes directrices précises s'appliquent aux activités forestières.
Un grand nombre de collègues ont aidé les rédacteurs du présent rapport à répertorier les occurrences et la répartition du Leptogium rivulare au Canada. Les rédacteurs du présent rapport remercient les personnes suivantes pour leur aide dans le cadre des travaux de terrain réalisés en Ontario : Robert Lee, Erica Oberndorfer, David Seburn, Linda Touzin, James Pagé, Julie Nadeau, Marie-Ange Gravel, Colin Jones, Daryl Couslon, Tobi Kiesewalter, Mike Burrell, Erica Barkley, Wasyl Bakowsky, Don Sutherland, Rob Craig, Mary Dillon et Meagan Robson. Ils remercient également Linda Ley pour ses identifications des spécimens de bryophytes. Des remerciements sont aussi adressés à Cassandra Robillard, qui a donné accès aux résultats des travaux qu'elle a réalisés à Stony Swamp (Ottawa), Don Faritosh, du MRNO, qui a fourni des ressources cartographiques pour la région de Temagami, ainsi que Glen Desy et Nathan Hanes, qui ont réalisé la cartographie près de Wawa.
Au Québec, les rédacteurs remercient Claude Roy, lichénologue consultant, et Jean Gagnon, de la Direction du patrimoine écologique et des parcs du Ministère du Développement durable, de l'Environnement, de la Faune et des Parcs, pour l'information et les conseils qu'ils ont fournis pour la sélection des sites ainsi que pour leur aide précieuse sur le terrain.
De plus, les rédacteurs du présent rapport remercient le Centre d'information sur le patrimoine naturel, la Direction du Grand Nord du ministère des Richesses naturelles et des Forêts ainsi que l'Accord Canada-Ontario, qui ont financé une partie du coût des travaux sur le terrain, en plus de fournir un appui non financier. Ils remercient également tous les experts nommés ci-dessous pour leur aide.
Frances Anderson, lichénologue consultante, Nouvelle-Écosse
Ron Bazin, biologiste spécialiste des espèces sauvages, Service canadien de la faune
Jim Bennet, professeur auxiliaire, University of Wisconsin
Curtis Bjӧrk, lichénologue consultant, Colombie-Britannique, et agrégé de recherche à l'herbier de l'University of Idaho
Irwin Brodo, lichénologue chercheur émérite, Musée canadien de la nature, Ottawa
Robert Cameron, écologiste, Protected Areas and Ecosystems Branch, Nova Scotia Environment
Stephen Clayden, conservateur de la botanique, Musée du Nouveau-Brunswick
Jennifer Doubt, conservatrice de la botanique, Musée canadien de la nature, Ottawa
Louise Dumouchel, Unité d'évaluation des risques phytosanitaires, Agence canadienne d'inspection des aliments
Colin Freebury, associé de recherche, Musée canadien de la nature, Ottawa
Trevor Goward, naturaliste et lichénologue consultant, Colombie-Britannique
Robert Lee, lichénologue consultant
James Lendemer, adjoint à la recherche et à la conservation, New York Botanical Garden
Janet Marsh, lichénologue consultante, Alberta
R. Troy McMullin, boursier de recherches postdoctorales, Institut de la biodiversité de l'Herbier de l'Ontario, département de biologie intégrative, Université de Guelph
Michele Piercey-Normore, professeure et conservatrice des cryptogames, Université du Manitoba
Clifford Wetmore, professeur émérite, University of Minnesota
Anderson, F. comm. pers. 2015. A New Occurrence for L. rivulare in Quebec at Parc du Bic, MRC Rimouski Neigette, 17/8/2015.
Alstrup, V. et S.N. Christensen. 2006. New records of lichens with cyanobacteria from Tanzania and Kenya. Cryptogamie, Mycologie 27(1): 57–68.
ArtDatabanken. 2013. Swedish Species Information Centre (Anglais seulement, consulté en juin 2013).
Bailey, R.H. et P.W. James. 1979. Birds and the dispersal of lichen propagules. Lichenologist 11(1):105-106.
Bazin, R., comm. pers. 2012. Correspondance par courriel adressée à Sam Brinker concernant l'emplacement des relevés visant le leptoge des terrains inondés réalisés au Manitoba et les niveaux d'eau et conditions de l'habitat observés, biologiste spécialiste des espèces sauvages, Service canadien de la faune, Winnipeg, Manitoba.
Becker, V.E., J.E. Reeder et R. Stetler. 1977. Biomass and habitat of nitrogen fixing lichens in an oak forest in the North Carolina Piedmont. Bryologist: 80: 93-99.
Bennet, J.P. et M.E. Bartkowiak. 2013. Three New Species of Leptogium in Wisconsin. Evansia: 30 (2): 72- 75
Berube, D., comm. pers. 2012. Correspondance par courriel adressée à S. Brinker concernant la description de l'habitat du leptoge des terrains inondés le long de la rivière Severn et la construction du barrage Muskrat, Biodiversity Species at Risk Biologist, district de Sioux Lookout, ministère des Richesses naturelles, Ontario.
Beyer, W. et D. Saari. 1978. Activity and ecological distribution of the slug, Arion subfuscus (Draparnaud) (Stylommotophora, Arionidae). American Midland Naturalist, 100/2: 359-367.
BLS 2015. The Importance of Ash Trees to Lichens (Anglais seulement)
Brinker, S.R. 2010-2013. Observations personnelles inédites de Sam Brinker faites durant les travaux de terrain menés pour la préparation de la mise à jour du rapport de situation du COSEPAC sur le leptoge des terrains inondés, botaniste, Centre d'information sur le patrimoine naturel, Peterborough, Ontario.
Brinker, S.R. et C.J. Lewis. 2013. Improving our knowledge of lichens. Natural Heritage Information Centre (NHIC) Newsletter 18: 3-5.
Brodo, I.M., S.D. Sharnoff et S. Sharnoff. 2001. Lichens of North America. Yale University Press, 795 pp.
Cameron, G. 2010-2013. Observations personnelles inédites de Graham Cameron faites durant les travaux de terrain menés pour la préparation de la mise à jour du rapport de situation du COSEPAC sur le leptoge des terrains inondés, biologiste des espèces en péril, ministère des Richesses naturelles, Bancroft, Ontario.
Cameron, G., comm. pers. 2013. Correspondance avec Sam Brinker concernant les répercussions de l'exploitation forestière dans l'habitat du leptoge des terrains inondés, biologiste des espèces en péril, ministère des Richesses naturelles, Bancroft.
Cameron, R. 2009. Are non-native gastropods a threat to endangered lichens? Canadian Field-Naturalist 123(2): 169–171
Campbell, J. et A.L. Fredeen. 2004. Lobaria pulmonaria abundance as an indicator of macrolichen diversity in Interior Cedar-Hemlock forests of east-central British Columbia. Canadian Journal of Botany 82: 970-982.
Canadian Council of Ministers of the Environment, 2013. 2010-2011 PROGRESS REPORT ON THE CANADA-WIDE ACID RAIN STRATEGY FOR POST-2000. PN 1490, ISSN 1911-1541. (Également disponible en français : Conseil canadien des ministres de l'environnemen. 2013. Rapport d'étape 2010-2011 sur la Stratégie pancanadienne sur les émissions acidifiantes après l'an 2000, PN 1491, ISSN 1911-155X.)
Canadian Endangered Species Conservation Council (CESCC). 2011. Wild Species 2010: The General Status of Species in Canada. National General Status Working Group: 302 pp. PDF (consulté en octobre 2013). (Également disponible en français : Conseil canadien pour la conservation des espèces en péril (CCCEP). 2011. Espèces sauvages 2010 : la situation générale des espèces au Canada, Groupe de travail national sur la situation générale, 323 p, PDF.)
CFIA (Canadian Food Inspection Agency). 2013. Areas Regulated for the Emerald Ash Borer (consulté en novembre 2013). (Également disponible en français : ACIA (Agence canadienne d'inspection des aliments). 2013. Zones réglementées à l'égard de l'agrile du frêne.)
CFIA (Canadian Food Inspection Agency). 2014. RMD-13-01 Regulated Areas for Emerald Ash Borer (EAB) (Agrilus planipennis Fairmaire) (consulté le 26 mai 2014). (Également disponible en français : ACIA (Agence canadienne d'inspection des aliments). 2014. DGR-13-01 : Régions réglementées à l'égard de l'agrile du frêne (Agrilus planipennis Fairmaire).)
Cappaert, D., D.C. McCullough, T.M. Poland et N.W. Siegert. 2005. Emerald Ash Borer in North America: A research and regulatory challenge. American Entomologist 51:152–165.
Chapman, L.J. et D.F. Putnam. 1984. The Physiography of Southern Ontario. 3rd ed. Ontario Ministry of Natural Resources, Ontario Geological Survey. Special volume 2.
Christensen, S., comm. pers. 2013. Correspondance par courriel adressée à Sam Brinker concernant la répartition du Leptogium rivulare en Tanzanie, Associate Professor, Aarhus University, Denmark.
2012. Consortium of North American Lichen Herbaria. (Anglais seulement, consulté en juin 2013).
COSEWIC. 2004. COSEWIC assessment and status report on the Flooded Jellyskin Leptogium rivulare in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa, Ontario. vi + 30 pp. (Également disponible en français : COSEPAC. 2004. Évaluation et Rapport de situation du COSEPAC sur le leptoge des terrains inondés (Leptogium rivulare) au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa, vi + 33 p.)
COSEWIC. 2011a. COSEWIC assessment and status report on the Batwing Vinyl Lichen Leptogium platynum in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. ix + 22 pp. (Également disponible en français : COSEPAC. 2011a. Évaluation et Rapport de situation du COSEPAC sur le leptoge à grosses spores (Leptogium platinum) au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa, ix + 24 p.)
COSEWIC. 2011b. COSEWIC assessment and status report on the Peackock Vinyl Lichen Leptogium polycarpum in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. ix + 24 pp. (Également disponible en français : COSEPAC. 2011b. Évaluation et Rapport de situation du COSEPAC sur le leptoge à quatre spores (Leptogium polycarpum) au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa, x + 27 p.)
COSEWIC. 2011c. Status Reports, Guidelines for Recognizing Designatable Units, Approved by COSEWIC in November 2011. (consulté le 13 janvier 2012). (Également disponible en français : COSEPAC. 2011c. Rapports de situation – Lignes directrices pour reconnaître les unités désignables, Approuvées par le COSEPAC en novembre 2011.)
COSEWIC. 2012. Instructions for the Preparation of COSEWIC Status Reports (consulté le 13 janvier 2012). (Également disponible en français : COSEPAC. 2012. Instructions pour la préparation des rapports de situation du COSEPAC.)
Crins, W.J., P. A. Gray, P.W.C. Uhlig et M. Wester. 2009. The Ecosystems of Ontario, Part 1: Ecozones and Ecoregions. Ontario Ministry of Natural Resources, Peterborough, Ontario, Inventory, Monitoring and Assessment SIB TER IMA TR-01, 71 pp.
Crowley, T.J. 1990. Are there any satisfactory geologic analogs for a future greenhouse warming? Journal of Climate 3: 1282-1292.
Ducks Unlimited. 2010. Southern Ontario Wetland Conversion Analysis. Final Report. 23 pp. + appendices. PDF (consulté en novembre 2013).
eBiodiversity. 2013. Interface for taxa found in Estonia . (Anglais seulement, consulté en novembre 2013).
EAB Information. 2013. (Anglais seulement, consulté en octobre 2013).
Environment Canada. 2013. Recovery Strategy for the Flooded Jellyskin Lichen (Leptogium rivulare) in Canada. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa. iv + 23 pp. (Également disponible en français : Environnement Canada. 2013. Programme de rétablissement du leptoge des terrains inondés (Leptogium rivulare) au Canada, Série de Programmes de rétablissement de la Loi sur les espèces en péril, Environnement Canada, Ottawa, vii + 26 p.)
Esslinger, T. L. 2012. A cumulative checklist for the lichen-forming, lichenicolous and allied fungi of the continental United States and Canada. North Dakota State University (Anglais seulement, consulté en juin 2013).
Farmer, A.M., J.W. Bates et J.N.B. Bell. 1991. Seasonal variations in acidic pollutant inputs and their effects on the chemistry of stemflow, bark and epiphyte tissues in three oak woodlands in N.W. Britain. New Phytologist 118: 441-451.
Feltmate, B. et J. Thistlethwaite. 2013. Climate Change Adaptation: A Priorities Action Plan for Canada. University of Waterloo. 122 pp.
Ferry, B.W., M.S. Baddeley et D.L.H. Hawksworth. 1973. Air Pollution and Lichens. University of Toronto Press, 389 pp.
Fink, B. 1935. The Lichen Flora of the United States. University of Michigan Press, Ann Arbor.
Forsyth, R.G., comm. pers. 2013. Correspondance par courriel adressée à Sam Brinker concernant la limace Arion subfuscus en Ontario, Research Associate, Royal BC Museum, Victoria, Colombie-Britannique.
Gauslaa, Y. 1985. The ecology of Lobaria pulmonaria and Parmelia caperata in Quercus dominated forests in south-west Norway. Lichenologist 17: 117–140.
Gauslaa, Y. 1995. The Lobarion, an epiphytic community of ancient forests threatened by acid rain. Lichenologist 27: 59–76.
Goward, T. et A. Arsenault. 2000. Cyanolichens and conifers: implications for global conservation. Forest Snow and Landscape Research 75(3): 303-318.
Gilbert, O.L. 1970. Further studies on the effect of sulphur dioxide on lichens and bryophytes. New Phytologist 69: 605–627.
Goudie, A. 2006 Human impact on the natural environment. Blackwell, Oxford.
Government of Manitoba. 2007. Manitoba's Water Protection Handbook: Everyone's Responsibility. Manitoba Water Stewardship, Winnipeg, Manitoba. 64 pp.
Graf, D.L. 1997. Northern redistribution of freshwater pearly mussels (Bivalvia: Unionoidea) during Wisconsin deglaciation in the southern glacial Lake Agassiz region: a review. American Midland Naturalist 138: 37-47.
Grimm, W.F., R.G. Forsyth, F.W. Schueler et A. Karstad. 2009. Identifying Land Snails and Slugs in Canada: Introduced Species and Native Genera. Canadian Food Inspection Agency, Ottawa, Ontario, Canada. 166 pp. (Également disponible en français : Grimm, W.F., R.G. Forsyth, F.W. Schueler et A. Karstad. 2009. Identification des escargots et des limaces terrestres au Canada : espèces introduites et genres indigènes, Agence canadienne d'inspection des aliments, Ottawa (Ontario), Canada, 168 p.)
Harris, A.G. et J. Keeney Marr. 2009. Caltha natans Pallas (Ranunculaceae) new for Michigan and Thunder Bay District Ontario. Michigan Botanist 48: 72-77.
Hinds, J.W. et P.L. Hinds. 2007. The Macrolichens of New England. New York Botanical Garden Press, New York.
Hodge, P., comm. pers. 2013. Correspondance avec Sam Brinker concernant l'agrile du frêne en Ontario et le rythme de sa propagation ainsi que les taux de mortalité des frênes, Forest Health Technician Specialist, Direction des sciences et de la recherche, ministère des Richesses naturelles, Ontario.
IUCN 2015. Leptogium rivulare - The Global Fungal Red List Initiative. iucn.ekoo.se/iucn/species_view/393482/. Jan 15, 2015, IUCN Specialist Group.
Jönsson, M.T. and G. Thor 2012. Estimating coextinction risks from epidemic tree death: affiliate lichen communities among diseased host tree populations of Fraxinus excelsior. PLoS ONE 7(9): e45701. doi:10.1371/.0045701
Jorgensen, J.M. 1994. Further notes on the European Taxa of the genus Leptogium, with emphasis on the small species. Lichenologist 26 (1) : 1-29.
Jørgensen, J.M. 2007. Massalongiaceae. – Nordic Lichen Flora Vol. 3. Cyanolichens. Gӧtborg (2nd Edition). 223 pp. (pg 74).
Jørgensen, P.M. et P.W. James. 1983. Studies on some Leptogium species of Western Europe. Lichenologist 15: 109-125.
Kallio, P. 1974. Nitrogen fixation in subarctic lichens. Oikos, 25: 194-198.
Katic, E., comm. pers. 2011. Correspondance avec le Service canadien de la faune, février 2011, gestionnaire, Gestion des ressources naturelles et des terrains, Division de la ceinture de verdure, Commission de la capitale nationale, Ottawa, Ontario, cité dans Environment Canada. 2013. Recovery Strategy for the Flooded Jellyskin Lichen (Leptogium rivulare) in Canada. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa. iv + 23 pp.
Knight, K.S., J.P. Brown et R.P. Long. 2013 Factors affecting the survival of ash (Fraxinus spp.) trees infested by Emerald Ash Borer (Agrilus planipennis). Biological Invasions 15, 371–383.
Laarson, P. et Y. Gauslaa. 2011. Rapid juvenile development in old forest lichens. Botany 89(1): 65-72.
Lattman, H., L. Lindblom, J. Mattson, P. Milberg, M. Skage et S. Eckman. 2009. Estimating the dispersal capacity of the rare lichen Cliostomum currugatum. Biological Conservation 142(8):1870-1878.
Lettau G. 1942. Flechten aus Mitteleuropa VII. Repertorium Specierum novarum Regni vegetabilis119(5): 265–348. In P.M. Jørgensen. 1994. Further notes on European taxa of the lichen genus Leptogium, with emphasis on the small species. Lichenologist 26: 1-29
Lewis, C.J. 2010-2013. Observations personnelles inédites de Chris Lewis effectuées durant les travaux de terrain pour la préparation de la mise à jour du rapport de situation du COSEPAC sur le leptoge des terrains inondés, Permits and Agreements Analyst, ministère des Richesses naturelles, Peterborough, Ontario.
Manitoba Provincial Parks Act. 2013. (consulté en janvier 2013). (Également disponible en français : Manitoba. Loi sur les parcs provinciaux. 2013.)
Marchant, K.R. 2007. Managing the Emerald Ash Borer in Canada. Proceedings – Interagency Research Forum on Gypsy Moth and other Invasive Species. Canadian Food Inspection Agency, Guelph, Ontario.
McKenney, D.W., J.H. Pedlar, K. Lawrence, P.A. Gray, S.J. Colombo et W.J. Crins. 2010. Current and Projected Future Climatic Conditions for Ecoregions and Selected Natural Heritage Areas in Ontario. Ontario Ministry of Natural Resources. Applied Research and Development Branch. III. Series: Climate change research report; CCRR-16. 42 pp.
Michiwawa. 2011. The Wawa History Album. Sinter plant photos 4. (Anglais seulement, consulté en novembre 2013).
Motiejūnaitė, J. et V. V. Golubkov. 2005. Cyanolichens of freshwater aquatic and subaquatic habitats in Lithuania and Belarus. Botanica Lithuanica 11(1): 35–40.
Motiejūnaitė, J., W. von Brackel, D. Stončius et Z. Preikša. 2011. Contribution to the Lithuanian flora of lichens and allied fungi. III. Botanica Lithuanica 17(1): 39-46.
NatureServe Explorer. 2004. Habitat-based Plant Element Occurrence Delimitation Guidance, 1 October 2004. consulté en
NatureServe. 2013. NatureServe Explorer: An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. (Anglais seulement, consulté le 27 mars 2013).
NHIC (National Heritage Information Centre), Ontario Ministry of Natural Resources. 2002.
Ontario Endangered Species Act.2007. (consulté le 20 janvier 2013). (Également disponible en français : Ontario. Loi de 2007 sur les espèces en voie de disparition. 2007.).
OMNR (Ontario Ministry of Natural Resources) 2010. Forest Management Guide for Conserving Biodiversity at the Stand and Site Scales.
Paiero, S.M., M.D. Jackson, A. Jewiss-Gaines, T. Kimoto, B.D. Gill et S.A. Marshall. 2012. Field Guide to the Jewel Beetles (Coleoptera: Buprestidae) of Northeastern North America. Canadian Food Inspection Agency, Ottawa, Canada. 411 pp. (Également disponible en français : Paiero, S.M., M.D. Jackson, A. Jewiss-Gaines, T. Kimoto, B.D. Gill et S.A. Marshall. 2012. Guide des buprestes (coleoptera : buprestidae) du nord-est de l'Amérique du Nord, Agence canadienne d'inspection des aliments, Ottawa, Canada, 420 p.)
Paukov, A.G. et A.Y. Teptina. 2012. New records of lichens from Middle Urals, Russia. Folia Cryptogamica Estonica 49: 39-43.
Pinceel, J., K. Jordaens, N. Van Houtte, G. Bernon et T. Backeljau. 2005. Population genetics and identity of an introduced terrestrial slug: Arion subfuscus s.l. in the north-east USA (Gastropoda, Pulmonata, Arionidae). Genetica 125: 155-171.
Provincial Parks and Conservation Reserves Act.2006. (consulté le 20 janvier 2013). (Également disponible en français :Loi de 2006 sur les parcs provinciaux et les réserves de conservation. 2006. .)
Randlane, T. 1987. Leptogium rivulare (ACH.) MONT. - A new rare lichen species in Estonia. Folia Cryptogamica Estononica 25: 8-11.
Richardson, D.H.S. 1991. Lichens as biological indicators – recent developments. In: Jeffrey, D.W. et B. MADDEN (eds). Bioindicators and environmental management. Toronto, Academic Press. 263–272.
Riley, J.L. 2003. Flora of the Hudson Bay Lowland and its Postglacial Origins. NRC Press, Ottawa, Ontario, Canada. 236 pp.
Robillard, C. 2012. Distribution patterns of the threatened Flooded Jellyskin lichen Leptogium rivulare in Stony Swamp, Ontario. Unpublished thesis. University of Ottawa and the Canadian Museum of Nature. 14 pp.
Sierk, H.A. 1964. The genus Leptogium in North America north of Mexico. Bryologist 67: 245-317.
Sigal, L.L. et J.W. Johnstone. 1986. Effects of acid rain and ozone on nitrogen fixation and photosynthesis in the lichen Lobaria pulmonaria (L.) Hoffm. Environmental and Experimental Botany 26: 59–64.
Species at Risk Act. 2002. PDF (consulté le 20 janvier 2012). (Également disponible en français : Loi sur les espèces en péril. 2002.)
Straw, N.A., D.T. Williams, O. Kulinch et Y.I. Gninenko. 2013. Distribution, impact and rate of spread of Emerald Ash Borer Agrilus planipennis (Coleoptera: Buprestide) in the Moscow region of Russia. Forestry 86: 515-522.
Streit, M.A., T. Scarr and L Farintosh. 2012. Preparing for Emerald Ash Borer: A Landowner's Guide to Managing Ash Forests. Leeds-Grenville Stewardship Council and Eastern Ontario Model Forest. 14pp.
Teller, J.T., D.W. Leverington et J.D. Mann. 2002. Freshwater outbursts to the oceans from glacial Lake Agassiz and their role in climate change during the last deglaciation. Quaternary Science Reviews 21:879–887.
Thompson, S. 2010-2013. Observations personnelles inédites de Shaun Thompson effectuées durant les travaux de terrain pour la préparation de la mise à jour du rapport de situation du COSEPAC sur le Leptogium rivulare, District Ecologist, ministère des Richesses naturelles de l'Ontario, Kemptville, Ontario.
Thompson, S., comm. pers. 2012. Correspondance par courriel adressée à Sam Brinker et Chris Lewis concernant les relevés visant le leptoge des terrains inondés menés au Québec. District Ecologist, ministère des Richesses naturelles de l'Ontario, Kemptville, Ontario.
Wang X.Y, Z.Q. Yang, J.R. Gould, Y.N. Zhang, G.J. Liu et E.S. Liu. 2010. The biology and ecology of the emerald ash borer, Agrilus planipennis, in China. Journal of Insect Science 10:128.
Wetmore, C.M., comm. pers. 2012. Correspondance par courriel adressée à Chris Lewis concernant la présence du leptoge des terrains inondés dans la région du lac Supérieur et dans le nord du Minnesota, Professor Emeritus, University of Minnesota.
Young, B., E. Byers, K. Gravuer, K. Hall, G. Hammerson et A. Redder. 2010. Guidelines for Using the NatureServe Climate Change Vulnerability Index Release 2.01 – Canada. NatureServe, Arlington, VA. 48 pp.
Samuel R. Brinker est botaniste au Centre d'information sur le patrimoine naturel de l'Ontario de la Direction des sciences et de la recherche du ministère des Richesses naturelles et des Forêts. Il réalise des relevés botaniques et collabore avec des partenaires provinciaux et fédéraux à des inventaires de base, des suivis et des évaluations ciblées des espèces rares. De plus, il compile, examine et tient à jour les données sur les cotes de conservation provinciales et la présence des espèces rares de plantes vasculaires et de lichens. Il est l'auteur de nombreux rapport de situation et sommaires du statut de l'espèce fédéraux sur des plantes et des lichens. M. Brinker est détenteur d'un baccalauréat en études environnementales de l'Université de Waterloo. Il a occupé différents postes au ministère des Richesses naturelles de l'Ontario et a été biologiste-conseil durant de nombreuses années.
Chris Lewis a commencé à étudier les lichens en 2003, alors qu'il terminait un baccalauréat en biologie à l'Université de Trent, à Peterborough, en Ontario. Depuis, il s'intéresse de plus en plus aux divers aspects de la taxinomie des lichens et de l'écologie de leur répartition, et il a été auteur ou coauteur d'environ 20 rapports traitant des lichens et d'environ 5 articles publiés dans des revues scientifiques. Actuellement, Chris travaille comme biologiste en gestion au district de Peterborough du ministère des Richesses naturelles et des Forêts de l'Ontario. Il s'intéresse tout particulièrement à l'écologie des lichens rares ainsi qu'à la taxonomie des lichens d'Ontario. La majeure partie de ses spécimens de lichens sont déposés dans les principaux herbiers d'Amérique du Nord, dont ceux de l'Université de Guelph (OAC), du Musée royal de l'Ontario (ROM), du Musée canadien de la nature (CANL) et du New York Botanical Garden (NYBG). Chris a siégé au Sous-comité de spécialistes des lichens du COSEPAC et travaille depuis 2010 comme chercheur associé au Musée canadien de la nature, à Ottawa.
Graham Cameron est biologiste en gestion au district de Bancroft du ministère des Richesses naturelles et des Forêts de l'Ontario. Il a réalisé des relevés visant de nombreuses espèces en péril, dont le leptoge des terrains inondés et la physconie pâle. Récemment, il a participé à une étude sur l'écologie spatiale de la tortue mouchetée, dans le cadre de laquelle données télémétriques sur les déplacements de plus de 30 individus de l'espèce ont été recueillies sur trois saisons de terrain. Il a travaillé au parc Kawartha Highlands Signature Site, au parc provincial The Pinery et au Centre écologique du Canada du parc provincial Samuel de Champlain. Graham détient un diplôme technique en gestion des écosystèmes du Sir Sanford Fleming College, à Lindsay, en Ontario, ainsi qu'un baccalauréat ès sciences avec spécialisation en biologie et en sciences de l'environnement de l'Université de Trent.
Shaun Thompson est actuellement biologiste en gestion au ministère des Richesses naturelles et des Forêts de l'Ontario. Auparavant, il a été écologiste du district de Kemptville durant 20 ans. Il a été président et coprésident de plusieurs équipes de rétablissement des espèces en péril; il a notamment été président de l'Équipe de rétablissement du leptoge des terrains inondés et s'est chargé de la coordination de l'ébauche du programme de rétablissement pour cette espèce. Shaun a réalisé des études et des relevés visant de nombreuses espèces en péril en Ontario, dont des végétaux et des animaux, et il a consacré un temps considérable aux relevés visant le leptoge des terrains inondés, particulièrement dans l'est de l'Ontario. En outre, Shaun a travaillé pour Parcs Ontario à titre de gardien et de directeur de parc ainsi que de technicien des poissons et de la faune et compte 32 années de service au ministère des Richesses naturelles de l'Ontario. Il est diplômé de l'Université Carleton (baccalauréat en biologie) et du Sir Sandford Fleming College (technique en ressources forestières et fauniques).
Les ressources liées au L. rivulare contenues dans les herbiers et sites Web suivants ont été consultées :
Musée national du Canada, Ottawa - CANL
Site Web du Consortium of North American Lichen Herbaria
University of Minnesota Herbarium – MINN
New York Botanical Garden -- NYBG
University of Guelph Herbarium – OAC
Royal Ontario Museum – ROM
University of Manitoba Herbarium – WIN
De plus, les lichénologues énumérés dans la section « Remerciements » ont été consultés concernant les spécimens de L. rivulare qui font partie de leurs collections privées ou qu'ils ont déposés dans des herbiers officiels.
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de nniveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de nniveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 3 | 1 |
| D | Faible | 2 | 4 |
| - | Impact global des menaces calculé : | Élevé | Élevé |
| Numéro de menace | Menace | Impact (calculé) | Impact (calculé) Définition |
Portée (10-prochaines années) |
Gravité (10-ans ou 3-gén.) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | - | Négligeable | Négligeable (<1-%) |
Extrême (71-100-%) | Élevée (Continue) |
- |
| 1.1 | Zones résidentielles et urbaines | - | Négligeable | Négligeable (<1-%) |
Extrême (71-100-%) |
Élevée (Continue) |
L'expansion des zones urbaines a des répercussions négatives sur l'espèce à Stony Swamp, en Ontario. Dans le canton de Darline, en Ontario, des propriétés sont subdivisées et aménagées, réduisant ainsi les milieux naturels où l'espèce pousse. Au Manitoba, la construction de chalets autour de lacs où le L.-rivulare est présent pourrait avoir un impact sur l'habitat de l'espèce. |
| 1.2 | Zones commerciales et industrielles | - | Négligeable | Négligeable (<1-%) |
Extrême (71-100-%) |
Inconnue | L'expansion des zones commerciales et industrielles dans la région d'Ottawa, y compris un projet d'installation de gestion des déchets, pourrait menacer certaines populations de leptoge des terrains inondés, mais on ignore l'immédiateté de cette menace. |
| 1.3 | Zones touristiques et récréatives | - | Négligeable | Négligeable (<1-%) |
Modérée - Légère (1-30-%) |
Modérée (possiblement à court terme, < 10-ans /3-gén.) | L'aménagement de sentiers dans les parcs et les aires de conservation perturbe l'habitat et nécessite l'élimination d'arbres hôtes; cette menace est particulièrement marquée dans la région d'Ottawa. Le leptoge des terrains inondés est désigné espèce en péril en Ontario, de sorte que l'aménagement de sentiers ne sera plus autorisé dans les endroits où l'espèce a été signalée, et aucun l'élargissement de sentier ne pourra être réalisé près des sites existants dans les parcs et les aires sauvages protégées. |
| 3 | Production d'énergie et exploitation minière | - | Négligeable | Négligeable (<1-%) |
Extrême (71-100-%) |
Élevée (Continue) |
- |
| 3.2 | Exploitation de mines et de carrières | - | Négligeable | Négligeable (<1-%) |
Extrême (71-100-%) |
Élevée (Continue) |
L'altération des milieux, la perte d'habitat et la modification des niveaux d'eau causées par l'extraction d'agrégats menacent l'espèce au Long lac Darling et à Coboconk, en Ontario. En outre, des concessions minières aux lacs Payuk, Twin, Whitefish et Neso, au Manitoba, menacent l'habitat de l'espèce. Cependant, même si la demande pour le calcaire de grande qualité est croissante, aucune extraction ne serait autorisée dans les sites où le leptoge des terrains inondés serait trouvé, et tout projet de carrière qui menacerait l'espèce serait rejeté par le MRNO, par exemple en vertu de la LEVD. |
| 3.3 | Énergie renouvelable | - | - | - | - | - | L'espèce pourrait être touchée par des projets d'énergie renouvelable, notamment des projets d'énergie solaire et d'éoliennes, mais l'impact et l'immédiateté de cette menace sont inconnus. |
| 4 | Corridors de transport et de service | - | Négligeable | Négligeable (<1-%) |
Élevée - Modérée (11-70-%) |
Élevée (Continue) |
- |
| 4.1 | Routes et voies ferrées | - | Négligeable | Négligeable (<1-%) |
Élevée - Modérée (11-70-%) |
Élevée (Continue) |
L'aménagement de chemins associé à la construction de chalets, à l'extraction d'agrégats, à l'expansion des zones urbaines et industrielles et à la création de campings dans le parc provincial Murphys Point peut perturber les régimes hydriques des terres environnantes et par conséquent avoir un impact négatif sur l'espèce. Il existe des contraintes réglementaires destinées à atténuer cette menace et à limiter ses répercussions sur les terres publiques, mais ces contraintes ne s'appliquent pas aux terrains privés. |
| 4.2 | Lignes de services publics | - | - | - | - | - | L'installation et l'entretien des lignes de services publics pourraient avoir des répercussions sur le leptoge des terrains inondés, mais il est impossible d'évaluer l'impact et l'immédiateté de cette menace à l'heure actuelle. |
| 4.3 | Voies de transport par eau | - | - | - | - | - | - |
| 4.4 | Corridors aériens | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques | D | Faible | Petite (1-10-%) |
Élevée (31-70-%) |
Élevée (Continue) |
- |
| 5.3 | Exploitation forestière et récolte du bois | D | Faible | Petite (1-10-%) |
Élevée (31-70-%) |
Élevée (Continue) |
Des arbres sont coupés pour la production de bois de chauffage et de bois d'œuvre en Ontario, et cette activité peut réduire les taux d'humidité et la persistance des mares saisonnières essentielles à l'espèce. |
| 6 | Intrusions et perturbations humaines | - | Négligeable | Négligeable (<1-%) |
Négligeable (<1-%) |
Élevée (Continue) |
- |
| 6.3 | Travail et autres activités | - | Négligeable | Négligeable (<1-%) |
Négligeable (<1-%) |
Élevée (Continue) |
Récolte de spécimens destinés à la recherche et à la définition de l'espèce. |
| 7 | Modifications des systèmes naturels | CD | Moyen - Faible | Restreinte - Petite (1-30-%) |
Élevée - Légère (1-70-%) |
Élevée (Continue) |
- |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | CD | Moyen - Faible | Restreinte - Petite (1-30-%) |
Élevée - Légère (1-70-%) |
Élevée (Continue) |
La manipulation des niveaux d'eau par les installations hydroélectriques sur les rivières Crowe, Otonobee, Moira, Mississippi et Severn, en Ontario, modifient et pourraient continuer de modifier les crues saisonnières, qui sont nécessaires au maintien de la santé et à la persistance de l'espèce. |
| 8 | Espèces et gènes envahissants ou autrement problématiques | C | Moyen | Restreinte (11-30-%) |
Élevée (31-70-%) |
Élevée (Continue) |
- |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | C | Moyen | Restreinte (11-30-%) |
Élevée (31-70-%) |
Élevée (Continue) |
Au cours des dix prochaines années, on s'attend à ce que l'agrile du frêne se propage à l'ensemble du sud et de l'est de l'Ontario, où se trouve la majeure partie de la population canadienne du leptoge des terrains inondés. Au cours des trois prochaines générations, l'impact de la mort des frênes dans la zone touchée par l'agrile du frêne variera d'un site à l'autre, selon la prédominance des frênes et l'abondance initiale du leptoge des terrains inondés. La propagation continue de la maladie hollandaise de l'orme constitue une autre menace, mais de moindre importance; cette maladie tue les ormes, troisième essence colonisée par l'espèce après le frêne et l'érable. De plus, une limace exotique envahissante, l'Arian subfuscus, endommage les thalles de l'espèce en Ontario. Enfin, au Manitoba, le Lyngbya wollei, cyanobactérie exotique, forme d'épais tapis ou des masses flottantes qui peuvent recouvrir les thalles de l'espèce durant les inondations périodiques et ainsi empêcher leur photosynthèse et leur croissance. |
| 9 | Pollution | D | Faible | Petite (1-10-%) |
Légère (1-10-%) |
Élevée (Continue) |
- |
| 9.3 | Effluents agricoles et sylvicoles | D | Faible | Petite (1-10-%) |
Légère (1-10-%) |
Élevée (Continue) |
L'eau des ruisseaux et rivières qui inonde périodiquement l'habitat de l'espèce peut endommager les thalles si elle contient des déjections animales ou des sédiments agricoles. |
| 9.5 | Polluants atmosphériques | - | Inconnu | Grande - Restreinte (11-70-%) |
Inconnue | Élevée (Continue) |
Le dioxyde de soufre et les pluies acides peuvent causer des dommages au leptoge des terrains inondés, mais l'espèce pousse sur l'écorce d'arbres dont le pH est élevé, dans des régions où la poussière issue de la roche calcaire devrait améliorer le pouvoir tampon de l'écorce et du lichen. Ainsi, l'impact de cette menace est probablement moindre dans le cas de la présente espèce que dans celui de nombreux autres cyanolichens qui poussent dans d'autres types de milieux. |
| 11 | Changements climatiques et phénomènes météorologiques violents | CD | Moyen - Faible | Restreinte (11-30-%) |
Élevée - Modérée (11-70-%) |
Modérée (possiblement à court terme, < 10-ans /3-gén.) | - |
| 11.2 | Sécheresses | CD | Moyen - Faible | Restreinte (11-30-%) |
Élevée - Modérée (11-70-%) |
Modérée (possiblement à court terme, < 10-ans /3-gén.) | Une hausse des températures est prévue en raison des changements climatiques, ce qui aura probablement de graves répercussions sur l'espèce, qui pousse dans des mares printanières et en bordure de cours d'eau et de lacs sujets à des crues périodiques. Quatre vingt pour cent des occurrences poussent dans des mares printanières. Les sécheresses prolongées entraîneront l'assèchement des mares et des rivages. L'impact de tels changements climatiques est incertain, car on s'attend à ce que ceux-ci se fassent sentir à long terme (>10-ans); toutefois, s'ils survenaient plus rapidement, leur impact sur les populations de leptoge des terrains inondés pourrait être modérée à élevé. |
Classification des menaces d'après l'IUCN-CMP, Salafsky et al. (2008).