
Évaluation et Rapport de situation du COSEPAC sur la Limace pygmée Kootenaia burkei au Canada - 2016

- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Facteurs limitatifs
- Menaces
- Changements climatiques et phénomènes météorologiques violents (impact de la menace : moyen à faible; portée : généralisée; gravité : modérée à légère) :
- Espèces exotiques (non indigènes) envahissantes (impact de la menace : moyen à faible; portée : restreinte; gravité : modérée à légère) :
- Exploitation forestière et récolte du bois (impact de la menace : faible; portée : restreinte; gravité : modérée) :
- Incendies et suppression des incendies (impact de la menace : faible; portée : petite; gravité : modérée) :
- Routes et voies ferrées (impact de la menace : faible; portée : petite; gravité : modérée à légère) :
- élevage de bétail (impact de la menace : faible; portée : petite; gravité : modérée à légère) :
- Effets cumulatifs
- Nombre de localités
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique des rédacteurs du rapport
- Collections examinées
- Figure 1. Une limace pygmée (Kootenaia burkei) de la Colombie-Britannique . La longueur de la limace était d'environ 10 mm.
- Figure 2. Répartition mondiale de la limace pygmée (Kootenaia burkei). Sources de données : Leonard et al. (2003); Hendricks et Maxwell (2005); Hendricks, comm. pers. (2013); Montana Government (2013); Tableau 1 (mentions canadiennes).
- Figure 3. Répartition canadienne de la limace pygmée. Source des mentions d'observation : Tableau 1.
- Figure 4. Survol des sites ayant fait l'objet de relevés de gastéropodes dans l'aire de répartition de la limace pygmée et autour de celle ci, dans le sud est de la Colombie-Britannique (voir le tableau 2 pour les sources de données dans la zone d'intérêt).
- Tableau 1. Mentions sur la répartition de la limace pygmée (Kootenaia burkei) au Canada.
- Tableau 2. Résumé des activités de recherche de gastéropodes terrestres effectuées dans le sud est de la Colombie-Britannique. Le nombre de sites de relevé ne se chevauchant pas a été obtenu à partir de cartes SIG de la zone d'intérêt délimitée à la figure 4.
- Tableau 3. Résumé de l'évaluation du calculateur des menaces de l'UICN pour la limace pygmée. Les catégories de menaces qui ne s'appliquaient pas à l'espèce ont été omises (d'où les écarts dans la numérotation des menaces).
- Annexe 1. Sommaire des sites recensés et des gastéropodes observés par Biolinx Environmental Research Ltd. (K. Ovaska et L. Sopuck) au cours de travaux sur le terrain réalisés aux fins du présent rapport de situation, en septembre 2013, ainsi que pour la limace gainée (Zacoleus idahoensis), en septembre 2014, dans la région de Kootenay, en Colombie-Britannique. Le ministère de l'Environnement de la Colombie-Britannique a fourni un soutien supplémentaire pour les relevés réalisés en 2014. [Note de la rédaction : Les coordonnées géographiques ont été enlevées du tableau ci dessous. On peut obtenir le tableau complet en communiquant avec le Secrétariat du COSEPAC.]
- Annexe 2. Résultats détaillés du calculateur des menaces pour la limace pygmée, obtenus lors de la conférence téléphonique de mai 2014.
COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur la limace pygmée (Kootenaia burkei) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xi + 63 p.
Le COSEPAC remercie Kristiina Ovaska et Lennart Sopuck d'avoir rédigé le rapport de situation sur la limace pygmée (Kootenaia burkei) au Canada. Le présent rapport a été préparé aux termes d'un marché conclu avec Environnement Canada, et sa supervision a été assurée par Joe Carney, coprésident du Sous-comité de spécialistes des mollusques du COSEPAC.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Pygmy Slug Kootenaia burkei in Canada.
Limace pygmée - Photo : Kristiina Ovaska.
La limace pygmée est la seule espèce du genre nouvellement décrit Kootenaia. Comme le sous-entend son nom commun, la limace pygmée est très petite, la longueur d'un adulte variant habituellement entre 9 et 14 mm. Sa couleur va du gris foncé au havane clair, et son manteau et sa queue sont densément couverts de mouchetures bleuâtres. Des taches foncées sont souvent présentes sur son manteau. Sa queue est arrondie (non carénée) avec une série de sillons longitudinaux obliques et parallèles, pouvant ressembler à de minces rayures foncées. La limace pygmée est une espèce endémique régionale qui se rencontre dans les forêts humides du nord du bassin du Columbia, où l'on trouve de nombreuses espèces végétales et animales uniques.
La répartition mondiale de la limace pygmée s'étend depuis le sud-est de la Colombie-Britanique, en passant par l'Idaho (Idaho Panhandle), jusqu'au nord-ouest du Montana. Au Canada, la limace pygmée est présente dans les zones des chaînes de montagnes Selkirk et Purcell à l'intérieur de la région de la chaîne Columbia, dans le sud-est de la Colombie-Britannique. L'espèce a été observée à 44 sites dans la province; le nombre de sites pourrait continuer à augmenter grâce aux activités de recherche accrues. Environ 36 % de son aire de répartition se trouve au Canada.
En Colombie-Britannique, les limaces pygmées sont présentes surtout dans la zone biogéoclimatique intérieure à thuya et à pruche, qui est l'une des zones les plus pluvieuses de l'intérieur de la province. Elles ont été observées dans les forêts mixtes et conifériennes humides, à des altitudes faibles à moyennes (de 580 m à 1 585 m), où elles sont communément associées aux milieux riverains le long de petits ruisseaux affluents. Les besoins clés en matière d'habitat semblent être caractérisés par un substrat très humide et par une grande quantité d'abris, notamment ceux qu'offrent les débris ligneux grossiers et les épaisses litières de feuilles. Des limaces pygmées ont été observées dans des peuplements de seconde venue de 40 à 50 ans et dans des peuplements anciens de plus de 200 ans. Le thuya géant et le peuplier de l'Ouest figurent parmi les espèces d'arbres courantes dans les sites occupés; la végétation du sous-étage est souvent composée d'espèces hydrophiles, comme la ronce parviflore, le bois piquant et l'athyrie fougère-femelle.
L'histoire naturelle de la limace pygmée est peu connue. Les limaces sont hermaphrodites, mais l'échange de sperme entre individus, plutôt que l'autofécondation, est probablement la norme. Elles pondent un petit nombre d'œufs, qui sont relativement gros (10 % ou plus de la longueur corporelle de l'adulte reproducteur). On sait que les limaces se nourrissent de lichens et de champignons, et probablement qu'elles consomment aussi de la matière organique en décomposition dans la couche d'humus. La plupart des observations en Colombie-Britannique et aux États-Unis ont été faites à l'automne, lorsque les limaces sont actives sur le parterre forestier. Il est probable que les jeunes et une proportion inconnue d'adultes hivernent. La durée d'une génération est d'environ 1 an. La petite taille de l'espèce pourrait lui permettre de se maintenir dans de petites parcelles d'habitat, pourvu que ses besoins en matière d'humidité et d'abri soient comblés. En général, les limaces présentent une faible capacité de dispersion si elles ne sont pas aidées par les humains ou transportées par le vent ou l'eau, mais aucun moyen de dispersion passif n'est connu pour la limace pygmée, ce qui exacerbe les effets de la fragmentation de l'habitat sur sa répartition dans le paysage.
On ne connaît ni la taille ni les tendances des populations de la limace pygmée. En Colombie-Britannique, les relevés effectués visaient principalement à clarifier la répartition de la limace pygmée et non pas à estimer son abondance. Les observations de l'espèce en Colombie-Britannique ont été effectuées durant la période 2007-2015, ce qui ne permet pas de dégager les tendances des populations.
La répartition canadienne de la limace pygmée témoigne probablement d'une expansion post-glaciaire à partir de refuges plus au sud. Sa répartition actuelle est probablement limitée vers le nord, par une courte saison de croissance et par les hivers longs et froids, et vers l'est et l'ouest, par des types de forêts plus secs. La faible capacité de dispersion et les besoins en matière d'habitat humide limitent la vitesse avec laquelle les limaces peuvent coloniser de nouveaux milieux.
Les populations de limaces pygmées sont menacées par les événements extrêmes liés aux changements climatiques, les espèces envahissantes introduites, les incendies et la suppression des incendies, l'exploitation forestière, les routes et l'élevage de bétail. On estime que les plus graves menaces pesant sur les limaces pygmées à l'échelle de leur aire de répartition canadienne sont les sécheresses et les inondations, dont la fréquence et la gravité devraient continuer d'augmenter selon les prévisions de scénarios de changement climatique. Parmi les espèces non indigènes envahissantes qui menacent les populations de limaces pygmées, on retrouve notamment les gastéropodes introduits, qui sont propagés par inadvertance par les humains, et qui se nourrissent d'espèces indigènes ou leur font compétition ainsi que d'autres prédateurs invertébrés, comme les carabes, qui peuvent être des prédateurs agressifs des limaces. On prévoit que la fréquence et la gravité des feux de végétation vont augmenter en raison des changements climatiques. En raison de leur faible mobilité, les gastéropodes ne peuvent pas échapper aux feux en se déplaçant et recolonisent lentement les brûlis. L'exploitation forestière est pratiquée dans l'ensemble de l'aire de répartition de la limace pygmée et continue de modifier et de fragmenter les habitats. Les effets de l'exploitation forestière sur les limaces pourraient être atténués, dans une certaine mesure, par les bandes riveraines boisées, qui sont requises le long des grands cours d'eau fréquentés par des poissons, ou que les entreprises forestières peuvent laisser volontairement le long des petits cours d'eau sans poissons, où ces bandes ne sont pas requises. Les chemins forestiers et les autres routes d'accès aux ressources continuent aussi de fragmenter les habitats.
La limace pygmée ne bénéficie ni d'une protection ni d'un statut officiels aux termes de la Loi sur les espèces en péril du gouvernement fédéral, de la Wildlife Act de la Colombie-Britannique ou d'autres lois. NatureServe attribue les cotes suivantes à la limace pygmée : cote mondiale - G2 (en péril); États-Unis - N2 (en péril); Canada - N1 (gravement en péril); Idaho - S2 (en péril); Montana - S1S2 (espèce gravement en péril à en péril); Colombie-Britannique - S1? (possiblement gravement en péril). En Colombie-Britannique, l'espèce figure sur la liste rouge provinciale des espèces en péril.
Dans l'aire de répartition canadienne de l'espèce, les terres protégées représentent environ 20 % du territoire et font partie notamment de parcs provinciaux, d'aires d'habitat faunique provinciales établies pour d'autres espèces et d'autres aires de conservation. Aucune mention d'observation de la limace pygmée n'a été répertoriée sur ces terres, exception faite d'un site dans une petite aire de conservation. La majeure partie de l'aire de répartition et des sites connus de l'espèce se trouvent sur des terres forestières provinciales.
| Sujet | Information |
|---|---|
| Durée d'une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN [2008] est utilisée) | Environ 1 an |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures? | Inconnu |
| Pourcentage estimé du déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. | Inconnu |
| Pourcentage [prévu ou présumé] [de changement, de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de changement, de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
| Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé? | S.o. |
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Probablement que non |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 15 552 km2 |
| Indice de zone d'occupation (IZO) [Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté.] | 180 km2 (distincts; 45 carrés de 2 km de côté); 1 160 km2 (continus le long de cours d'eau; 290 carrés de 2 km de côté) |
| La population totale est-elle gravement fragmentée? | Inconnu |
| Nombre de localités | > 20 compte tenu du nombre de sous-bassins occupés et des menaces posées par les changements climatiques et les phénomènes météorologiques violents ou les espèces envahissantes. |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | Inconnu |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | Inconnu |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations? | Inconnu |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Inconnu |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? | Oui, déclin observé, inféré et prévu de la qualité de l'habitat |
| Y a-t-il des fluctuations extrêmes du nombre de populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
| Population | Nombre d'individus matures |
|---|---|
| Total | Inconnu |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Non effectuée |
| Sujet | Information |
|---|---|
24. Un calculateur des menaces a-t-il été rempli pour l'espèce? Oui
|
Quels autres facteurs limitatifs sont pertinents? Capacité de dispersion faible; dépendance envers les micromilieux humides |
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur? | G2 (mondiale), N2 (États-Unis), S2 (Idaho), SIS2 (Montana) |
| Une immigration a-t-elle été constatée ou est-elle possible? | N'a pas été constatée, mais est possible. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Oui |
| Les conditions se détériorent-elles au Canada? | Oui |
| Les conditions de la population source se détériorent-elles? | Oui |
| La population canadienne est-elle considérée comme un puits? | Non |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Possible à long terme dans certaines zones près de la frontière, mais le taux serait très bas. |
| Sujet | Information |
|---|---|
| L'information concernant l'espèce estelle de nature délicate? | Non |
| Sujet | Information |
|---|---|
| COSEPAC | N'a pas été évaluée dans le passé. |
| Sujet | Information |
|---|---|
| Statut | Espèce préoccupante |
| Code alphanumérique | S.o. |
| Justification de la désignation | Au Canada, cette petite limace est confinée aux forêts humides du bassin Columbia nord de la Colombie-Britannique. Elle se trouve dans les forêts humides mixtes et de conifères et est communément associée aux habitats riverains le long de petits ruisseaux. Les exigences essentielles en matière d'habitat incluent une humidité élevée du substrat avec une abondance de débris ligneux et de litière de feuilles en guise d'abri. Les menaces incluent les routes existantes et nouvelles résultant en la fragmentation, des effets de lisière accrus, et des obstacles à la dispersion; la prédation et la compétition des espèces envahissantes; l'endommagement des zones riveraines associé au pâturage du bétail; la perte et la dégradation de l'habitat liées à l'exploitation forestière; et les conséquences prévues des changements climatiques, incluant un accroissement des conditions de sécheresse ainsi qu'une augmentation du nombre d'incendies de forêt et de leur gravité. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Ce critère ne s'applique pas, car aucune estimation de la taille ou des tendances des populations n'est disponible. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Sans objet. La zone d'occurrence (15 552 km2) satisfait au critère de la catégorie « espèce menacée » (< 20 000 km2) et l'IZO (180 km2) satisfait au critère de la catégorie « espèce en voie de disparition » (< 500 km2) et au critère « espèce menacée » (< 2 000 km2), la population n'est pas très fragmentée et le nombre de localités (> 20) dépasse les seuils, et il n'y a pas de fluctuations extrêmes. |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Ce critère ne s'applique pas. Aucune estimation de la taille ou des tendances des populations n'est disponible. |
| Critère D (très petite population totale ou répartition restreinte) | Ce critère ne s'applique pas. Il n'existe aucune estimation de la taille des populations, et le critère D2 pour « espèce menacée » ne s'applique pas, étant donné que l'IZO et le nombre de localités dépassent les seuils fixés. |
| Critère E (analyse quantitative) | Sans objet. Aucune analyse quantitative n'a été réalisée. |
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
La limace pygmée (Kootenaia burkei) a été décrite en 2003 d'après des spécimens du nord de l'Idaho (localité-type : ruisseau Little Bumblebee, forêt nationale Panhandle; Leonard et al., 2003). L'espèce est la seule représentante de son genre, qui fait partie de la grande famille cosmopolite des Arionidés (Leonard et al., 2003). Une autre classification réalisée par Bouchet et Rocroi (2005) hisse toutes les sous-familles d'Arionidés au statut de famille. Aucune de ces classifications n'est satisfaisante, car les études génétiques actuelles ne soutiennent la monophylétie ni de la famille des Arionidés, qui est plus inclusive, ni de ses sous-familles qui ont été étudiées (Backeljau, comm. pers., 2011).
De tous les autres genres nord-américains de la famille des Arionidés, le genre Kootenaia est celui qui est le plus étroitement apparenté au genre Prophysaon, mais des divergences génétiques et l'anatomie comparative permettent de les distinguer clairement (Leonard et al., 2003). Outre le genre Prophysaon, Burke (2013) a placé le genre Kootenaia et deux genres monotypiques découverts récemment, soit les genres Carionarion et Securicauda (Leonard et al., 2011), dans la famille des Anadenidés (Anadeninés dans d'autres classifications), mais sans fournir d'explication.
Voici la classification actuelle de l'espèce :
Embranchement : Mollusques
Classe : Gastéropodes
Sous-classe : Pulmonés
Ordre : Stylommatophores
Sous-ordre : Arionoïdés
Famille : Arionidés
(sous-famille : Anadeninés)
Genre : Kootenaia
Espèce : Kootenaia burkei
Le genre Kootenaia tire son nom de la Première Nation des Kootenays, qui occupaient par le passé les terres renfermant l'aire de répartition de l'espèce (Leonard et al., 2003). L'espèce a été nommée en l'honneur de Thomas Burke, un biologiste de la faune, qui a beaucoup travaillé sur les gastéropodes terrestres de la côte pacifique nord-ouest des États-Unis.
Comme le sous-entend son nom commun, la limace pygmée est très petite, la longueur en extension d'un adulte variant habituellement entre 9 et 14 mm (Leonard et al., 2003). En Colombie-Britannique, la longueur de limaces vivantes, y compris de jeunes limaces, variait entre 3 et 16 mm (Ovaska et Sopuck, données inédites, 2007-2015). Le corps de la limace pygmée est mince, et son manteau couvre environ la moitié de sa longueur totale (figure 1). Sa queue est arrondie (non carénée) avec une série de sillons longitudinaux obliques et parallèles. Les sillons se ramifient à l'extrémité de la queue, formant de petits polygones, caractéristiques de l'espèce (Burke, 2013). Les sillons sur la queue peuvent ressembler à de minces rayures foncées. La limace pygmée n'a pas de ligne d'abscission, comme celle observée chez les limaces-prophyses (espèce du genre Prophysaon).

Description longue de la figure 1
Photo d'une limace pygmée sur une feuille (vue dorsale). La limace pygmée adulte est petite (d'une longueur variant habituellement entre 9 et 14 mm). Sa couleur va du gris foncé au havane, et son manteau et sa queue sont densément couverts de mouchetures bleuâtres. Des taches foncées sont souvent présentes sur son manteau. Le corps de la limace pygmée est mince, et son manteau couvre environ la moitié de sa longueur totale. Sa queue est arrondie (non carénée) avec une série de sillons longitudinaux obliques et parallèles, pouvant ressembler à de minces rayures foncées.
Sa couleur va du gris foncé au havane clair, et son manteau et sa queue sont densément couverts de mouchetures bleuâtres. Des taches foncées sont souvent présentes sur son manteau, mais à l'occasion, celles-ci sont absentes (Leonard et al., 2003). En Colombie-Britannique, dans certains sites, on a seulement trouvé des limaces non tachetées, mais, dans la majorité des sites, on n'a trouvé que des limaces tachetées (Ovaska et Sopuck, données inédites, 2007-2015).
Sur le plan de l'anatomie, la limace pygmée se démarque des autres limaces de la famille des Arionidés par la région distale de son système reproducteur, sur lequel on se base couramment pour classifier et identifier les gastéropodes. Comparativement à celui d'autres Arionidés, la composante mâle du système reproducteur de la limace pygmée est de taille anormalement réduite. De plus, elle dépourvue d'un épiphallus libre et d'un canal déférent (Leonard et al., 2003).
La structure génétique des populations de limace pygmée est inconnue. En Colombie-Britannique, l'espèce a été observée dans des localités isolées, la plupart d'entre elles étant situées au sud de 49,4° de latitude N. Ces localités ainsi que celles se trouvant plus au nord (jusqu'à 50,5° de latitude N.) sont séparées par une distance minimale de 58 km (entre les sites 4 et 6). Les quatre localités les plus nordiques sont aussi relativement éloignées les unes des autres (42 km entre les sites 5 et 9; 29 km entre les sites 9 et 10; 38 km entre les sites 10 et 4). Les limaces des localités nordiques sont très probablement isolées les unes des autres et par rapport à celles plus au sud. Compte tenu des capacités de dispersion limitées des limaces et de leur affinité pour les milieux très humides, un échange génétique entre sous-populations est peu probable à l'extérieur d'un même ruisseau ou sous-bassin, et ce, même dans la partie sud de l'aire de répartition, où la répartition de l'espèce est probablement plus continue.
La limace pygmée est présente dans une zone relativement petite à l'intérieur d'une aire écologique nationale du COSEPAC (montagnes du Sud). Il n'y a pas de disjonctions de l'aire de répartition ni d'autres renseignements qui porteraient à croire à la présence d'unités distinctes et importantes sur le plan évolutionnaire au sein de la sous-population canadienne, mais la variabilité génétique, anatomique et écologique de l'espèce n'a pas fait l'objet d'études. L'espèce est donc traitée comme une seule unité désignable.
La limace pygmée est une espèce endémique régionale des forêts humides du nord du bassin du Columbia, une région où l'on trouve de nombreuses espèces végétales et animales uniques (Brunsfeld et al.,2001). Cette région s'étend du sud-est de la Colombie-Britannique et du nord-est de l'État de Washington, en passant par le nord de l'Idaho (Idaho Panhandle), jusqu'au nord-ouest du Montana. Appartenant à l'un des rares genres de limaces endémiques à l'ouest de l'Amérique du Nord, et en tant que seule représentante de son genre, la limace pygmée présente un intérêt scientifique pour l'étude de l'histoire glaciaire et des relations évolutionnaires.
La répartition mondiale de la limace pygmée s'étend depuis le sud-est de la Colombie-Britannique, en passant par le nord de l'Idaho (Idaho Panhandle), jusqu'au nord-ouest du Montana (figure 2). Dans le nord de l'Idaho, l'espèce a été observée à sept endroits (cinq localités, Leonard et al., 2003; Leonard, comm. pers., 2013; deux localités, Hendricks et Maxwell, 2005; Hendricks, comm. pers., 2013). Vingt-cinq observations de la limace pygmée ont été répertoriées dans la base de données du Montana Natural Heritage Program (à jour en date du 11 avril 2013; Montana Government, 2013). Elles ont été effectuées dans les comtés de Lincoln, de Sanders et de Mineral dans le nord-ouest de l'État. Même si on ne fait état d'aucune mention d'observation dans l'extrême nord-est de l'État de Washington, l'espèce pourrait y être présente compte tenu de la proximité des observations répertoriées en Colombie-Britannique par rapport à la frontière. Au Canada, l'espèce n'a été observée que dans la région de Kootenay (Colombie-Britannique). Environ 36 % de l'aire de répartition de l'espèce se trouve au Canada.

Description longue de la figure 2
Carte de la répartition mondiale de la limace pygmée, qui s'étend depuis le sud-est de la Colombie-Britannique, en passant par le nord de l'Idaho (Idaho Panhandle), jusqu'au nord-ouest du Montana.
| N° de site | Description du site | Alt. (m) | Zone CBE Note du tableaua | Date | Nbre de limaces trouvées | Temps de recherche (min.-pers.) | Habitat | Âge approx. du peuplement (années) | Source Note du tableaub |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Ruisseau Lost (aire de repos), au bord de la route 3, entre Salmo et Creston, (C.-B.) | 664 | ICHxw (près de la limite avec ICHdw1) | 22 sept. 2007 | 1 | 118 | Forêt mixte de seconde venue; forêt riveraine | 50 | Ovaska et Sopuck, 2009a (RBCM, non catalogué) |
| 2 | Ruisseau Hawkins, Yahk Meadow FSR, à env. 3 km de Yahk, C.-B. | 913 | ICHdw1 | 23 sept. 2007 | 8 | 132 | Forêt mixte de seconde venue; plaine inondable le long du ruisseau | 80 | Ovaska et Sopuck, 2009a (RBCM, 007-00077-001) |
| 3 | Yahk River FSR, le long d’un affluent du ruisseau Sunrise, C.-B. | 1 260 | ICHdm | 5 sept. 2008 | 1 | 70 | Forêt de conifères de seconde venue; forêt riveraine le long d’un ruisseau asséché | 50-100 | Ovaska et Sopuck, 2009a (photos) |
| 4 | Ruisseau Lemon, vallée de la Slocan, C.-B. | 705 | ICHdw1 | 3 sept. 2008 | 3 | 70 | Forêt mixte de seconde venue; zone riveraine le long d’un ruisseau | 60 | Ovaska et Sopuck, 2009a (photos) |
| 5 | Halfway River FSR, au sud de Galena (à l’est du lac Arrow), C.-B. | 781 | ICHmw2 | 8 oct. 2008 | 2 | 124 | Vieille forêt mixte; zone riveraine le long d’un ruisseau | 100 | Ovaska et Sopuck, 2009a (photos) |
| 6 | Chemin Marsh Creek, en provenance du chemin Champion Park (en prov. de la route 3), entre Fruitvale et Salmo, C.-B. | 1 090 | ICHdw1 | 9 oct. 2008 | 1 | 120 | Vieille forêt mixte; lisière d’une petite trouée du couvert forestier dans une dépression humide | Ovaska et Sopuck, 2009a (photos) | |
| 7 | Sundown Cr. FSR (embranchement), env. 5 km au sud-est de Moyie, C.-B. | 975 | ICHdw1 | 8 oct. 2009 | 1 | 60 | Forêt mixte de seconde venue; zone riveraine humide le long du ruisseau | 70 | Ovaska et Sopuck, 2009b (photos) |
| 8 | Ruisseau Sundown, au S.-O. de Moyie, C.-B. | 1 140 | ICHdw1 | 11 sept. 2010 | 16 | 123 | Forêt mixte de seconde venue; zone de suintement près d’un petit ruisseau | 50 | Ovaska et al., 2010 (RBCM, non catalogué) |
| 9 | Slewiskin (McDonald) FSR (site 2), au sud de Nakusp, C.-B. | 640 | ICHmw2 | 22 sept. 2013 | 1 | 60 | Forêt mixte de seconde venue; le long d’un ruisseau affluent à fort débit | 60-70 | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 10 | East Wilson Cr. FSR (site 2), au nord de New Denver, C.-B. | 581 | ICHdw1 | 22 sept. 2013 | 1 | 60 | Forêt mixte de seconde venue; le long d’un ruisseau affluent à fort débit | 40-50 | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 11 | Ruisseau Nine Mile. (site 2B), Pend d’Oreille, C.-B. | 608 | ICHxw | 23 sept. 2013 | 1 | 60 | Vieille forêt de conifères; forêt humide le long du ruisseau | 100+ | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 12 | Sheep Cr. FSR (site 1), au sud de Salmo, C.-B. | 1 179 | ICHmw2 | 24 sept. 2013 | 3 | 50 | Vieille forêt mixte; forêt riveraine le long d’un ruisseau affluent à fort débit | 100+ | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 13 | Sheep Cr. FSR (site 2), au sud de Salmo, C.-B. | 969 | ICHdw1 | 24 sept. 2013 | 2 | 40 | Forêt mixte de seconde venue; zone riveraine le long d’un ruisseau à fort débit | 80 | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 14 | Chemin Carroll Cr., à l’ouest de Yahk, C.-B. | 993 | ICHdw1 | 24 sept. 2013 | 1 | 50 | Vieille forêt de conifères; zone humide le long d’un ruisseau | 200+ | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 15 | Teepee Cr. FSR, au sud-est de Cranbrook, C.-B. | 1 125 | MSdk1 (près de la limite avec ICHmk4) | 25 sept. 2013 | 9 | 40 | Vieille forêt de conifères; zone humide adjacente au ruisseau | 100 | Travaux sur le terrain à l’appui de ce rapport de situation, par K. Ovaska et L. Sopuck (RBCM, non catalogué) |
| 16 | Meachen Cr. FSR (site 3 au ruisseau Fiddler), env. 14 km au sud du lac Mary, C.-B. | 1 284 | ESSFwm | 18 sept. 2014 | 4 | 40 | Versant orienté vers le nord d’un ravin où poussent de jeunes peupliers de l’Ouest le long d’un ruisseau affluent à fort débit | 60-70 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 17 | Hellroaring Cr. FSR (site 1), au sud du lac St. Mary, C.-B. | 1 304 | ESSFwm | 18 sept. 2014 | 1 | 60 | Lisière de peupliers de l’Ouest à côté de la route dans une forêt principalement de conifères en pente abrupte à mi-pente | 60-70 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 18 | Cherry Cr. FSR (site 3), près du lac Cherry, C.-B. | 1 229 | Limite entre ICHdm et MSdk1 (à la limite même) | 20 sept. 2014 | 2 | 50 | Plaine alluviale avec des aulnes près d’un ruisseau dans une forêt de conifères, peut-être inondée de façon saisonnière | 80-90 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 19 | Bloom Cr. FSR (site 3), C.-B. | 1 269 | MSdk1 | 20 sept. 2014 | 1 | 60 | Zone riveraine avec des aulnes le long d’un petit ruisseau (filet d’eau) dans une forêt de seconde venue | 60 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 20 | Yahk R. FSR (site 3), C.-B. | 1 111 | MSdk1 | 21 sept. 2014 | 1 | 40 | Site alluvial humide, inondé périodiquement, le long d’un grand cours d’eau de fort débit; trouée du couvert dans une forêt de conifères avec végétation du sous-étage abondante | 100 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 21 | Lamb Cr. FSR (site 1), à l’ouest de Moyie, C.-B. | 1 121 | ICHdm | 22 sept. 2014 | 1 | 40 | Plaine inondable riveraine humide dans un ravin étroit à l’intérieur d’un paysage de caractérisé par des abris formés par la coupe des arbres; plusieurs très grands peupliers de l’Ouest | 70 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 22 | Tate Cr. FSR (site 1), en prov. du Lamb Cr. FSR, C.-B. | 1 194 | ICHdm | 22 sept. 2014 | 5 | 40 | Bande riveraine boisée (largeur de 50 à 75 m) avec de grands peupliers de l’Ouest dans un sol riche de plaine inondable le long du ruisseau | 80 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 23 | Tate Cr. FSR (site 2), en prov. du Lamb Cr. FSR, C.-B. | 1 392 | ICHdm | 22 sept. 2014 | 2 | 50 | Ravin peuplé d’arbres le long d’un petit ruisseau (zone riveraine < 20 m) entouré d’une vieille forêt de conifères sur terrain élevé | 150 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 24 | Irishman R. FSR, près de Moyie, C.-B. | 971 | ICHdw1 | 22 sept. 2014 | 1 | 40 | Vieille forêt de conifères humide avec peu de végétation du sous-étage, sauf sous les trouées du couvert | 125+ | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 25 | Cold-Freeman FSR (site 2), en prov. du Hawkins FSR, à l’est de Yahk, C.-B. | 1 179 | ICHdm | 23 sept. 2014 | 2 | 40 | Zone riveraine humide le long d’un ruisseau avec des buttes et des dépressions, et des peupliers de l’Ouest dans une forêt de conifères de seconde venue | 70-80 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 26 | Skelly Cr. FSR (site 1), en prov. du Goat Cr. FSR, au nord-est de Creston, C.-B. | 944 | ICHdw1 | 24 sept. 2014 | 1 | 40 | Plaine inondable riveraine le long d’un ruisseau avec des peupliers de l’Ouest dans une forêt de seconde venue principalement de conifères | 60-70 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 27 | Skelly Cr FSR (site 2), en prov. du Goat Cr. FSR, au nord-est de Creston, C.-B. | 1 095 | ICHdm | 24 sept. 2014 | 1 | 56 | Zone riveraine étroite (largeur de 10 m) le long d’un ruisseau affluent à fort débit, entourée d’une forêt dense de conifères de seconde venue avec peu de végétation du sous-étage | 40-50 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 28 | Goat R. FSR (site 2), au N.-E. de Creston, C.-B. | 1 092 | ICHdw1 | 24 sept. 2014 | 1 | 40 | Dépression humide dans une forêt de seconde venue (ayant environ 60-70 ans) | 60-70 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 29 | Mt. Thompson FSR (site 1), à l’est de Creston, C.-B. | 855 | ICHxw | 24 sept. 2014 | 1 | 54 | Zone riveraine humide où poussent des peupliers de l’Ouest le long d’un ruisseau à fort débit dans une pente de conifères normalement sèche avec peu de végétation du sous-étage | 90-100 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 30 | Sanca Cr. FSR (site 1), au nord de Creston, C.-B. | 1 189 | ICHdw1 | 25 sept. 2014 | 5 | 40 | Zone riveraine étroite (largeur d’environ 10 m) le long d’un ruisseau affluent à fort débit dans un paysage normalement sec à prédominance de pins; zone humide rare | 40 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 31 | Sanca Cr. FSR (site 4; South Fork), au nord de Creston, C.-B. | 1 585 | ESSFdm | 25 sept. 2014 | 1 | 40 | Zone riveraine humide dans une plaine inondable le long d’un cours d’eau dans une vieille forêt de conifères; chablis abondants et gros rochers | 100+ | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 32 | Dodge Cr. FSR (site 2) au ruisseau Dodge, au sud de Creston, C.-B. | 1 325 | ICHmw4 | 26 sept. 2014 | 1 | 80 | Zone riveraine avec quelques peupliers de l’Ouest dans une jeune forêt; le paysage est normalement sec et a fait l’objet de coupe à blanc, et ne compte que quelques ruisseaux | 30-40 | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 33 | Monk Cr. FSR (site 1) | 1 411 | ESSFdm | 27 sept. 2014 | 2 | 60 | Vieille forêt de conifères humide avec un sol profond et productif et des arbustes dans une dépression | 100+ | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 34 | Route 6 vers Nelway (petit embranchement), au sud de Salmo, C.-B. | 668 | ICHdw1 | 27 sept. 2014; 24 sept. 2015 | 8; 8 | 126; 60 | Zone riveraine humide le long d’un cours d’eau dans une forêt de conifères de seconde venue | 40-60 | Min. de l’Env., 2015 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 35 | Lacs Champion (site 2), au nord de Trail, C.-B. | 1 079 | ICHdw1 | 28 sept. 2014 | 1 | 40 | Vieille forêt de conifères où le bois humide et en état de décomposition avancé est abondant et avec des parcelles d’arbustes dans des dépressions humides | 100+ | Min. de l’Env., 2014 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 36 | Archibald-Tillicum FSR (site 2), au sud-ouest de Salmo, C.-B. | 1 229 | ICHdw1 | 29 sept. 2014; 24 sept. 2015 | 5; 5 | 50; 120 | Zone riveraine humide avec des peupliers de l’Ouest le long d’un petit ruisseau dans un paysage forestier exploité dans une forêt de seconde venue | 50-60 | Min. de l’Env., 2015 et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 37 | Erie Cr. FSR (site 1), au nord d’Erie, au nord-ouest de Salmo, C.-B. | 991 | ICHdw1 | 29 sept. 2014 | 2 | 40 | Ravin humide où poussent des peupliers de l’Ouest le long d’un petit ruisseau dans le paysage d’une forêt sèche plus jeune (exploitée) | 60-70 | Min. de l’Env. (Ovaska et Sopuck, 2014) et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 38 | Erie Cr. FSR (site 2), au nord d’Erie, au nord-ouest de Salmo, C.-B. | 915 | ICHdw1 | 29 sept. 2014 | 1 | 40 | Plaine inondable riveraine humide avec des aulnes dans une vieille forêt de conifères | 100+ | Min. de l’Env. (Ovaska et Sopuck, 2014) et travaux sur le terrain pour le rapport du COSEPAC sur la limace gainée Note du tableauc (RBCM, non catalogué) |
| 39 | Chemin Beaver Lookout (en prov. du Archibald-Tillicum FSR), C.-B. | 987 | ICHdw1 | 24 sept. 2015 | 2 | 62 | Zone riveraine à côté d’un ruisseau à fort débit; petite clairière arbustive humide entre le chemin et le ruisseau | 40-50 | Min. de l’Env. (Ovaska et Sopuck, 2015); (RBCM, non catalogué) |
| 40 | Elmer Creek FSR, au sud-est de Creston, C.-B. | 1 013 | ICHdw1 | 24 sept. 2015 | 3 | 90 | Zone riveraine le long d’un petit cours d’eau affluent à fort débit | 40-50 | Min. de l’Env. (Ovaska et Sopuck, 2015); (RBCM, non catalogué) |
| 41 | American Creek FSR, près du ruisseau Hawkin, chemin Meadow, à l’est de Yahk, C.-B. | 1 135 | ICHdm | 25 sept. 2015 | 1 | 60 | Trouée du couvert avec végétation herbacée abondante sur un terrain en pente dans une forêt humide; zone de suintement sur la pente | 60-70 | Min. de l’Env. (Ovaska et Sopuck, 2015); (RBCM, non catalogué) |
| 42 | Randall Creek FSR, au sud du Hawkin Cr. FSR | 1 327 | ICHdm | 25 sept. 2015 | 2 | 90 | Pente herbeuse humide dans une trouée du couvert forestier (env. 30 m plus haut qu’un petit ruisseau ou une petite dépression) | 50-60 | Min. de l’Env. (Ovaska et Sopuck, 2015) |
| 43 | Chemin West Yahk, à l’est de Yahk | 1 189 | ICHdm | 25 sept. 2015 | 3 | 90 | Talus humide entre deux cours d’eau dans une vieille forêt | 80-90 | Min. de l’Env. (Ovaska et Sopuck, 2015); (RBCM, non catalogué) |
| 44 | Chemin West Yahk (site 3), à l’est de Yahk | 1 223 | ICHdm | 25 sept. 2015 | 2 | 60 | Habitat riverain le long d’un ruisselet dans une parcelle de forêt | 50-60 | Min. de l’Env. (Ovaska et Sopuck, 2015); (RBCM, non catalogué) |
Au Canada, la limace pygmée est présente dans les zones des chaînes de montagnes Selkirk et Purcell à l'intérieur de la région de la chaîne Columbia, dans le sud-est de la Colombie-Britannique (figure 3). Son aire de répartition s'étend entre le bras est du fleuve Columbia (lac Koocanusa), dans l'est, et les lacs Arrow, dans l'ouest. La mention la plus nordique (50,5° de latitude N.) se trouve 62 km au sud-est de Revelstoke du côté est du lac Upper Arrow.

Description longue de la figure 3
Carte de la répartition de la limace pygmée au Canada, là où elle est présente dans les zones des chaînes de montagnes Selkirk et Purcell à l'intérieur de la région de la chaîne Columbia, dans le sud-est de la Colombie-Britannique. Son aire de répartition s'étend entre le bras est du fleuve Columbia (lac Koocanusa), à l'est, et les lacs Arrow, à l'ouest. La mention la plus nordique (50,5 degrés de latitude nord) se trouve 62 km au sud-est de Revelstoke du côté est du lac Upper Arrow.
La limace pygmée a été découverte en Colombie-Britannique en 2007, et des relevés ciblés subséquents effectués de 2008 à 2015 ont permis de répertorier des sites supplémentaires. Il y a désormais des mentions d'observations dans 44 sites en Colombie-Britannique (un site est désigné comme tel s'il est situé à plus de 1 km d'un autre site; tableau 1). Tous les sites, sauf quatre, sont situés au sud de 49,58° de latitude N. et à moins de 64 km au nord de la frontière canado-états-unienne. De nouveaux sites continuent d'être trouvés grâce aux activités de recherche accrues; sept nouveaux sites ont été découverts en septembre 2013, 23 en 2014 dans le cadre de relevés ciblés liés à la préparation du présent rapport de situation et de celui sur la limace gainée (Zacoleus idahoensis), respectivement; et six ont été découverts en 2015 dans le cadre de relevés supplémentaires pour le ministère de l'Environnement de la Colombie-Britannique, mais l'aire de répartition connue ne s'est pas étendue de façon significative. Des sites additionnels existent probablement, particulièrement dans la portion sud de l'aire de répartition de l'espèce.
À l'aide de la méthode du plus petit polygone convexe, compte tenu des mentions d'occurrence connues, la zone d'occurrence est de 15 552 km2, la frontière avec les États-Unis correspondant à la limite méridionale du polygone.
L'indice de zone d'occupation (IZO) fondé sur une grille distincte pour chaque mention d'observation (ou groupe de mentions) est de 180 km2 (45 carrés de 2 km de côté). On pourrait obtenir un IZO plus réaliste en tenant compte des ruisseaux entiers lorsque ceux-ci font l'objet d'observations de l'espèce. Selon cette méthode, l'IZO continu est de 1 160 km2 (290 carrés de 2 km de côté) (calculs de la zone d'occurrence et de l'IZO effectués par Jenny Wu, Secrétariat du COSEPAC). D'autres sites non répertoriés pourraient exister, et contribueraient à l'accroissement de l'IZO.
Les activités de recherche menées dans la région de Kootenay, en Colombie-Britannique, avant les années 1990 sont peu documentées. Dans son étude sur les gastéropodes terrestres du bassin du Columbia, Forsyth (1999) n'a relevé que quatre dénombrements de courte durée, de 1905 à 1945, qui mentionnaient la présence de mollusques terrestres. Depuis le début des années 1990, des relevés de grande envergure ont été menés dans la région de Kootenay, et plus de 700 sites ont fait l'objet de relevés (tableau 2; figure 4). La plupart de ces relevés ciblaient spécifiquement les gastéropodes terrestres, sauf ceux réalisés par Copley et Copley qui étaient des relevés généraux des arthropodes au cours desquels tous les gastéropodes trouvés étaient recueillis et identifiés par la suite. Les relevés ont été effectués principalement à l'automne, qui est généralement la meilleure période pour repérer les gastéropodes terrestres, particulièrement les limaces, car les conditions (temps doux et pluvieux) sont propices à l'activité des gastéropodes, et la plupart des limaces sont matures, ce qui permet de les trouver plus facilement.
La limace pygmée a été découverte par Biolinx Environmental Research Ltd. en 2007; leurs relevés au cours des années suivantes (2008, 2009, 2010, 2013, 2014 et 2015) ont permis de répertorier d'autres mentions d'observation de l'espèce. Les relevés de septembre 2013 et 2014 ont été effectués à l'appui du présent rapport de situation et de celui sur la limace gainée, respectivement, et visaient les milieux convenant à la limace pygmée; la limace gainée est présente dans des milieux semblables (voir l'annexe 1 pour les sites des relevés et les espèces trouvées). En 2013, dans le but de mieux délimiter la répartition de la limace pygmée en Colombie-Britannique, des relevés ont été réalisés dans des zones se trouvant à l'extérieur de son aire de répartition connue (vers l'ouest, le nord et l'est), les relevés étant graduellement effectués vers l'intérieur de l'aire de répartition principale présumée, dans la région de Kootenay-Ouest. Les relevés de 2013 ont permis de répertorier 7 nouvelles observations de la limace pygmée; ceux de 2014 ont permis de répertorier 23 nouvelles mentions (sites 9 à 38 dans le tableau 1 et la figure 3), mais n'ont accru que légèrement la zone d'occurrence. Des relevés supplémentaires financés par le ministère de l'Environnement de la Colombie-Britannique, réalisés en 2015 (Ovaska et Sopuck, 2015), ont permis de découvrir 6 nouveaux sites, mais n'ont pas permis d'accroître la zone d'occurrence.
| Année | Mois | Nbre de sites | Temps de recherche | Relevés effectués par : | Source ou projet Note du tableaud |
|---|---|---|---|---|---|
| 1998-1999 | Septembre (1 en juillet) | 40 | RBCM (Kelly Sendall, Phil Lambert) | Projet Living Landscape; dossier du RBCM | |
| 1990-2013 | Divers | 135 | Robert Forsyth | Base de données principale personnelle de R. Forsyth (à jour en date de 2013) et autres sites particuliers; y compris ceux de Flathead Bioblitz, 2012 | |
| 2007 | Juillet, septembre | 63 | 66,1 heures-personnes | Biolinx Environmental Research Ltd. (Kristiina Ovaska, Lennart Sopuck) | Ovaska et Sopuck, 2009a |
| 2008 | Septembre, octobre | 45 | 48 heures-personnes | Biolinx Environmental Research Ltd. (Kristiina Ovaska, Lennart Sopuck) | Ovaska et Sopuck, 2009a |
| 2009 | Octobre | 17 | 20,9 heures-personnes | Biolinx Environmental Research Ltd. (Kristiina Ovaska, Lennart Sopuck) | Ovaska et Sopuck, 2009b |
| 2009-2013 | Juillet-septembre | 96 | Claudia et Darren Copley | Fichiers de données de C. Copley | |
| 2008-2011 | Divers | 85 | Dwayne Lepitzki | Relevés en Alberta et en Colombie-Britannique; base de données personnelle de Lepitzki | |
| 2010 | Septembre | 56 | 67,9 heures-personnes | Biolinx Environmental Research Ltd. (Kristiina Ovaska, Lennart Sopuck) | Ovaska et al., 2010 |
| 2011 | Août, septembre | 29 | Jeff Nekola, Brian Coles, Michael Horsek | Relevés pour la Valhalla Wilderness Society; Nekola et al.,2011 | |
| 2012 | Août | 6 | Melissa Frey | Flathead Bioblitz; base de données du RBCM. Note : Les sites supplémentaires qui chevauchent ceux de Forsyth ont été exclus. | |
| 2013 | Septembre | 36 | 31,7 heures-personnes | Biolinx Environmental Research Ltd. (Kristiina Ovaska, Lennart Sopuck) | Travaux sur le terrain liés à la préparation du rapport de situation du COSEPAC sur la limace pygmée |
| 2013 | Juin | 14 | Dwayne et Brenda Lepitzki | Flathead Bioblitz; fichiers de données de Lepitzki | |
| 2014 | Septembre | 72 | 72,2 | Kristiina Ovaska et Lennart Sopuck | Relevés des gastéropodes pour le ministère de l'Environnement de la Colombie-Britannique et travaux sur le terrain liés à la préparation du rapport de situation du COSEPAC sur la limace gainée |
| 2015 | Septembre | 36 Note du tableaue | 38,5 heures-personnes | Biolinx Environmental Research Ltd (Kristiina Ovaska, Lennart Sopuck) | Relevés des gastéropodes effectués pour le ministère de l'Environnement de la Colombie-Britannique (Ovaska et Sopuck, 2015) |
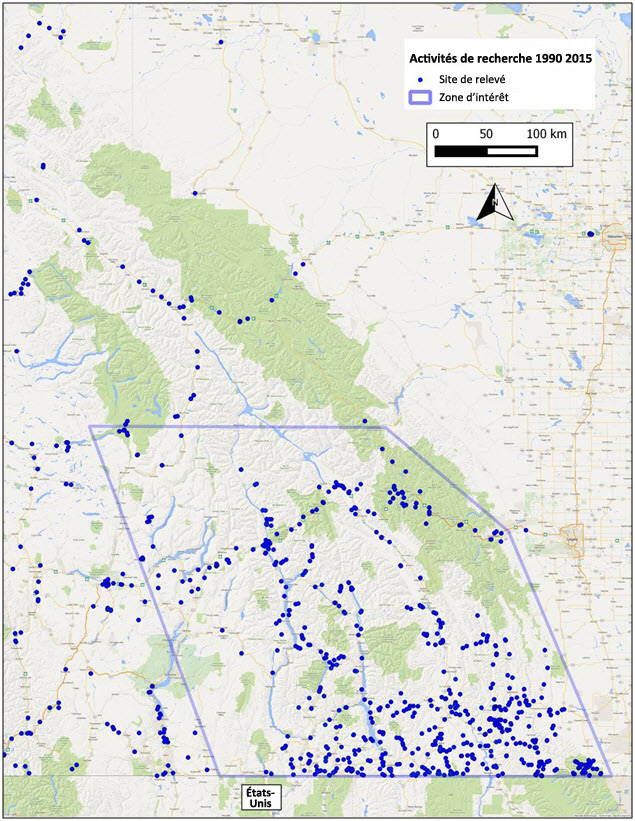
Description longue de la figure 4
Carte montrant les sites (indiqués par des symboles) ayant fait l'objet de relevés de gastéropodes dans l'aire de répartition de la limace pygmée et autour de celle-ci, dans le sud-ouest de la Colombie-Britannique de 1990 à 2015.
À l'échelle de son aire de répartition mondiale, la limace pygmée est présente dans les forêts mixtes et conifériennes humides, particulièrement dans les milieux riverains (Leonard et al., 2003; Hendricks et Maxwell, 2005; Ovaska et Sopuck, 2009a,b). Leonard et al. (2003) ont mentionné la proximité des sites occupés à des plans d'eau pérennes, probablement parce que ces derniers possèdent des substrats humides convenables. Un modèle de la répartition prévue pour le Montana, fondé sur l'analyse des caractéristiques biophysiques de 20 points connus et de 60 000 points de référence aléatoires, a montré que l'habitat hautement convenable se limitait essentiellement à des bandes de milieu riverain le long des cours d'eau (Montana Government, 2013). Les observations en Colombie-Britannique ont, elles aussi, été effectuées surtout dans les milieux riverains, souvent le long de ruisseaux à fort débit (tableau 1). La zone riveraine le long des ruisseaux était souvent confinée à des bandes étroites dans les ravins, mais, dans certains cas, on a trouvé des limaces dans des zones de suintement plus étendues sur un terrain plus plat. Parmi les plantes communes du sous-étage dans les sites occupés, on trouve la ronce parviflore (Rubus parviflorus), le bois piquant (Oplopanax horridum), l'athyrie fougère-femelle (Athyrium filix-femina), la salsepareille (Aralia nudicaulis), la linnée boréale (Linnaea borealis) et la tiarelle trifoliée (Tiarella trifoliata). Les limaces ne vivent pas dans les milieux humides ou rivages ouverts, comme les marais à quenouilles (Typha latifolia).
La limace pygmée est présente dans des endroits à faible et moyenne altitude dans l'Idaho (de 640 m à 700 m au-dessus du niveau de la mer; Leonard et al., 2003), au Montana (de 762 m à 1 372 m au-dessus du niveau de la mer pour la plupart des mentions; altitude indiquée en pieds, de 2 500 à 4 500 pi; Montana Government, 2013), et en Colombie-Britannique (de 571 m à 1 585 m au-dessus du niveau de la mer; tableau 1). Trente-six des 44 (81,8 %) mentions canadiennes renvoient à la zone biogéoclimatique intérieure à thuya et à pruche (ICH) (voir Meidinger et Pojar, 1991 pour la classification des zones). Deux sites se trouvent dans la zone montagnarde à épinette (MS), deux à la frontière entre les zones ICH et MS, et quatre dans la zone à épinette d'Engelmann et sapin subalpin (ESSF) (tableau 1). La zone ICH se trouve à des altitudes faibles à moyennes sur les versants inférieurs des montagnes de la chaîne Columbia, dans le sud-est de la Colombie-Britannique, et s'étend vers le sud, dans l'est de l'État de Washington, le nord de l'Idaho (Idaho Panhandle) et l'ouest du Montana (Ketcheson et al., 1991). Elle est délimitée en altitude, suivant un gradient d'élévation, par la zone ESSF. La zone ICH est caractérisée par des hivers frais et pluvieux et des étés chauds et secs, la majeure partie de l'humidité du sol provenant de la fonte des neiges. La saison de croissance (avec des températures au-dessus de 0 °C) dure de deux à cinq mois, selon la latitude et l'altitude. Il s'agit d'une des zones intérieures les plus pluvieuses de la Colombie-Britannique, ayant des caractéristiques en commun avec les forêts de conifères humides le long de la côte pacifique; on l'appelle parfois la zone humide de l'intérieur (Interior Wet Belt en anglais). Des forêts de conifères productives sur terrain élevé sont présentes partout dans le paysage, mais la topographie et les conditions du sol ont engendré une mosaïque de types de forêts plus humides et plus secs ayant une diversité relativement grande dans le sous-étage et l'étage supérieur.
En Colombie-Britannique, la limace pygmée a été observée dans des peuplements forestiers mixtes et conifériens d'âges variés, allant de peuplements de seconde venue de 40 à 50 ans à des peuplements anciens (> 200 ans) (tableau 1). Le thuya géant (Thuja plicata) était présent dans 74 %, le peuplier de l'Ouest (Populus trichocarpa) dans 61 %, l'épinette d'Engelmann (Picea engelmannii) dans 58 % et la pruche de l'Ouest (Tsuga heterophylla) dans 42 % des 38 sites occupés. D'autres espèces d'arbres étaient présentes, notamment le sapin grandissime (Abies grandis), le sapin gracieux (Abies amabilis), le sapin subalpin (Abies lasiocarpa), le bouleau à papier (Betula papyrifera), le mélèze de l'Ouest (Larix occidentalis) et, rarement, le pin tordu (Pinus contorta). Il y avait généralement une végétation du sous-étage modérée composée d'arbustes variés, comprenant souvent des espèces hydrophiles, comme la ronce parviflore et le bois piquant. Des parcelles de litière de feuilles plus profondes ou des quantités relativement importantes de débris ligneux grossiers (en moyenne, une couverture de 14 % à moins de 10 m des individus observés) couvraient habituellement le sol (Ovaska et Sopuck, données inédites, 2007-2014). Par temps humide, on a observé des limaces dans la litière de feuilles, souvent parmi les feuilles humides de peuplier de l'Ouest ou sous les débris ligneux du parterre forestier. Les sites avaient comme caractéristiques communes, entre autres, des substrats très humides et une couverture abondante pouvant abriter les limaces. La disponibilité de ces caractéristiques de micromilieu pourrait exercer une influence de première importance sur le type de forêt ou d'autres caractéristiques de l'habitat d'échelle grossière.
À l'intérieur de l'aire de répartition canadienne de la limace pygmée, la plupart des milieux convenables se trouvent sur des terres forestières provinciales et privées faisant l'objet d'une exploitation forestière continue. L'élimination de la couverture forestière, la construction de routes forestières et les activités de sylviculture associées à la foresterie ont eu la plus forte incidence sur la disponibilité d'habitat dans l'aire de répartition de l'espèce, et l'exploitation forestière continue de fragmenter et de modifier l'habitat. Quatre-vingt-quinze pour cent des sites connus abritant la limace pygmée se trouvent dans des paysages où des activités d'exploitation forestière sont en cours (voir la section Menaces).
La possibilité annuelle de coupe (PAC) établie pour les terres publiques dans les trois zones d'approvisionnement forestier englobant l'aire de répartition de l'espèce (lacs Arrow, lac Kootenay et lac Cranbrook) est demeurée relativement constante au cours des quatre dernières décennies (MFLNRO, 2014a). La plus récente PAC pour ces trois zones d'approvisionnement forestier, s'appliquant aux 5 à 10 prochaines années, semble indiquer qu'un taux de récolte légèrement plus bas sera maintenu. La majeure partie du bois récolté dans le passé provenait de forêts anciennes et de forêts en maturation se trouvant dans des zones ayant subi des perturbations naturelles. À l'avenir, une plus grande proportion de la récolte proviendra de peuplements de seconde venue en régénération. Dans les zones où des activités d'exploitation forestière ont eu lieu il y a 50 à 60 ans (principalement à des altitudes faibles à moyennes), les conditions dans les forêts en maturation pourraient permettre aux limaces de recoloniser quelques-unes des zones exploitées auparavant. Ces augmentations de la disponibilité de l'habitat ne compenseront que partiellement la dégradation continue de l'habitat causée par l'exploitation forestière. Il est toutefois important de noter qu'aucune exploitation forestière n'a lieu (ou alors elle est restreinte) dans les parcs, les aires protégées, près des eaux fréquentées par des poissons, dans les bassins hydrographiques approvisionnant une communauté, dans les zones d'aménagement de forêt ancienne, et dans les zones spéciales de gestion des ressources se trouvant un peu partout dans l'aire de répartition de l'espèce. On estime que, sur les terres disponibles pour la récolte dans la zone d'approvisionnement forestier du lac Kootenay, qui englobe plus de la moitié de l'aire de répartition de l'espèce, 42 % sont des terres forestières productives, après avoir tenu compte de ces aires de conservation et d'autres contraintes relatives à l'exploitation forestière (MFLNRO, 2014b). Du bois supplémentaire est récolté chaque année sur des terres privées et par les titulaires de permis d'exploitation de boisés sur des terres publiques (on n'a pas pu trouver de données quantitatives).
Le pâturage par les animaux d'élevage sur les terres forestières publiques est confiné principalement aux secteurs plus secs du sud et de l'est de l'aire de répartition de l'espèce (iMapBC, 2014). Les tenures de parcours sur les terres publiques sont gérées de façon à éviter le pâturage excessif, ce qui réduit potentiellement l'impact sur les zones riveraines.
La conversion des terres à des fins de développement résidentiel et industriel ainsi qu'à des fins agricoles a entraîné la perte permanente d'habitat des limaces, principalement sur les terres privées à basse altitude, en particulier le long de vallées fluviales, de rives lacustres et de routes. Toutefois, la densité de la population dans la région de Kootenay-Ouest est relativement faible comparativement à d'autres régions du sud de la Colombie-Britannique, comme la vallée de l'Okanagan et l'est du bassin du Columbia. Depuis 2001, le taux de croissance démographique dans la région de Kootenay-Ouest n'a été que de 1,3 % par décennie, la population atteignant 64 379 personnes en 2011 (Columbia Basin Rural Development Institute, 2012). Il y a de grands centres de population dans l'aire de répartition de l'espèce à Nelson et à Creston, et quelques collectivités nettement plus petites à Fruitvale, à Kaslo, à Nakusp, dans la vallée de la Slocan, près du lac Moyie et à Yahk. Les villes relativement grandes de Castlegar, de Trail et de Cranbrook se trouvent tout juste à l'extérieur de l'aire de répartition de l'espèce.
Des activités d'exploitation de mines et de carrières se déroulent également, mais ne touchent qu'un petit pourcentage de l'aire de répartition de la limace pygmée. Les concessions minières et de placer sont communes partout dans l'aire de répartition de l'espèce, particulièrement dans le sud (Trail, Nelson, Salmo, lac Moyie) et le centre (New Denver/Silverton), et plusieurs projets d'exploration minérale sont en cours (Grieve, 2010). Même si de nouvelles mines pourraient être exploitées à l'avenir, aucun projet de mine ne fait actuellement l'objet d'une évaluation dans l'aire de répartition de la limace pygmée (iMapBC, 2014). Cependant, tout juste au nord-est, près de Trout Lake, la réouverture et l'expansion d'une mine de molybdène sont en train d'être examinées. Une importante dégradation de l'habitat due à la pollution atmosphérique s'est produite au cours des 100 dernières années à proximité de la fonderie située à Trail, en périphérie de l'aire de répartition de l'espèce.
Des réservoirs aménagés pour la production hydroélectrique ont entraîné l'inondation de vastes zones d'habitat potentiel des limaces au cours du siècle dernier (lac Kootenay et Pend D'Oreille dans l'aire de répartition de l'espèce; lacs Arrow, lac Duncan et lac Koocanusa en périphérie). Plusieurs projets visant à mettre à niveau des centrales hydroélectriques sont en cours, mais aucun projet de création ou d'expansion des réservoirs à grande échelle n'est prévu dans un avenir rapproché (iMapBC, 2014). Les corridors de lignes de transport d'énergie sont relativement communs dans l'aire de répartition de l'espèce, et plusieurs autres corridors seront probablement aménagés pour accommoder l'expansion d'exploitations hydroélectriques. Plus de 20 projets d'installations hydroélectriques au fil de l'eau de plus petite échelle ont aussi été proposés ou sont approuvés (1 est en exploitation, 8 sont approuvés et les autres sont en train d'être évalués) dans l'aire de répartition de l'espèce (Wildsight, 2014).
Le développement récréatif, p. ex. les stations de ski, les centres touristiques et les terrains de camping, sont éparpillés dans l'aire de répartition de la limace pygmée, mais les infrastructures sont limitées pour l'instant. Aucun projet majeur de développement touristique ne fait actuellement l'objet d'une évaluation dans la région (iMapBC, 2014). La zone du projet de développement proposé du centre de villégiature Jumbo Glacier est située au nord-est de l'aire de répartition de l'espèce. Les activités récréatives se déroulent un peu partout dans la région et comprennent, entre autres, l'utilisation de véhicules tout-terrain, de motoneiges et de vélos de montagne.
La fragmentation de l'habitat qui s'est produite dans le passé et qui est toujours en cours en raison de toutes les activités humaines combinées, particulièrement à basse altitude, est préoccupante en ce qui concerne la limace pygmée. La fragmentation a été causée par l'intensification de l'exploitation forestière, l'augmentation de la fréquence des feux de végétation catastrophiques (attribuable à l'accumulation de combustibles à des niveaux non naturels), la création d'importants réservoirs hydroélectriques, la construction de routes, l'urbanisation et la conversion de terres pour l'agriculture. La zone biogéoclimatique intérieure à thuya et à pruche (ICH) est sujette à des perturbations périodiques dues aux incendies, mais dans une moindre mesure que les zones biogéoclimatiques plus sèches situées dans la partie sud de la région intérieure de la Colombie-Britannique (Biodiversity Guidebook, 1995). L'exploitation forestière, toutefois, élimine de façon sélective du bois de grande valeur dans les sites humides et productifs, diminuant ainsi le nombre de refuges disponibles pour les limaces après l'exploitation.
Dans le cadre du programme de résilience de Kootenay-Ouest (non daté), une série de documents abordant les changements climatiques et leurs répercussions sur la région de Kootenay-Ouest, en Colombie-Britannique, ont été publiés. La limace pygmée est présente surtout dans les sous-zones sud et centrale de Kootenay-Ouest et le long de la limite sud de la sous-zone nord. Tous les modèles et scénarios examinés laissent entrevoir une hausse des températures moyennes saisonnières, ces dernières présentant des augmentations progressives dans les années 2020, 2050 et 2080 (Utzig, 2012a). D'ici 2080, on prévoit que les hivers seront de 2 à 5 C° plus chauds et de 10 à 25 % plus pluvieux et que les étés seront de 3 à 7 °C plus chauds et jusqu'à 30 % plus secs que durant la période de référence (les modèles présentant la pire performance ayant été exclus). Les changements connexes qui ont des répercussions sur la limace pygmée sont notamment l'augmentation du stress hydrique survenant en été, l'augmentation potentielle des feux de végétation et des infestations d'insectes et des éclosions de maladies, qui réduiraient la couverture forestière, et les changements des régimes d'écoulement saisonniers des cours d'eau résultant de l'accumulation annuelle de neige réduite et des sécheresses estivales, qui modifieraient les zones riveraines où les limaces sont présentes. On prévoit également une augmentation de l'ampleur et de la fréquence des phénomènes extrêmes, comme les épisodes de pluie très intenses, les sécheresses graves et les tempêtes de vent (Utzig, 2012a).
Wang et al. (2012) ont étudié les effets des changements climatiques sur les 16 zones biogéoclimatiques de la Colombie-Britannique, qui sont fondées sur des gradients climatiques à grande échelle et sont largement utilisées pour classifier les écosystèmes dans la province (Meidinger et Pojar, 1991). Les modèles ont montré qu'un déplacement des enveloppes climatiques sur lesquelles repose ce système de classification en zones s'est produit depuis les années 1970 (Wang et al., 2012). Les projections futures (pour les années 2020, les années 2050 et les années 2080) des modèles, à l'échelle de la province, montrent une expansion considérable des forêts continentales humides de thuya et de pruche, des forêts types de la zone ICH, où l'on rencontre la limace pygmée, ce qui pourrait agrandir la zone allant jusqu'à tripler sa superficie d'ici 2080, la zone ICH devenant le type de forêt le plus commun dans la province à long terme (Wang et al., 2012). À l'échelle régionale, les projections sont plus complexes et subissent l'influence de la topographie et de facteurs locaux (Utzig, 2012b). Les modèles climatiques pour la région de Kootenay-Ouest indiquent que la zone ICH s'agrandira à long terme selon un des trois scénarios de changement climatique examinés (scénario « doux/humide (Warm/Moist) »), tandis qu'elle devrait, en grande partie, être déplacée par la zone côtière à pruche de l'Ouest et les écozones de transition côtières, qui sont aussi composées de forêts humides, selon le scénario « chaud/pluvieux (Hot/Wet) », et par les écosystèmes plus secs de prairie-steppe et de forêts de pins ponderosa, selon le scénario « très chaud/sec (Very Hot/Dry) » (tableau 3.1 et figure 3.5 dans Utzig, 2012b). L'expansion des types de forêts humides favorables à la limace pygmée est censée se produire principalement dans la partie nord de l'aire de répartition de l'espèce et/ou à des altitudes plus élevées (> 1 000 m au-dessus du niveau de la mer). Inversement, l'habitat convenable deviendrait plus petit dans la partie sud de l'aire de répartition et à plus basse altitude. Cependant, il est incertain si la limace pygmée pourra se déplacer vers le nord et plus en altitude au même rythme que le déplacement des écosystèmes pour tirer parti des milieux propices nouvellement disponibles. Les changements pourraient être entraînés, en grande partie, par les événements climatiques extrêmes, notamment les sécheresses ou les orages durant l'été, ainsi que par les infestations de ravageurs, les feux ou d'autres perturbations, plutôt que de se produire au cours d'une transition plus graduelle (Pojar, 2010; Utzig, 2012b); aussi, de nouvelles zones bioclimatiques pourraient émerger avec les nouvelles combinaisons de variables climatiques saisonnières (Utzig, 2012b), ce qui augmenterait l'imprévisibilité des projections.
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 2 | 0 |
| D | Faible | 4 | 6 |
| - | Impact global des menaces calculé : | Élevé | Moyen |
| Menace | Menace | Impact (calculé) | Impact (calculé) | Portée (10 proch. années) | Gravité (10 ans ou 3 générations) | Immédiateté |
|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) |
| 1.1 | Zones résidentielles et urbaines | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) |
| 1.2 | Zones commerciales et industrielles | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | Élevée (continue) |
| 1.3 | Zones touristiques et récréatives | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) |
| 2 | Agriculture et aquaculture | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Élevée (continue) |
| 2.3 | Élevage de bétail | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 3 | Production d'énergie et exploitation minière | - | Négligeable | Négligeable (< 1 %) | Extrême à élevée (31-100 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) |
| 3.2 | Exploitation de mines et de carrières | - | Négligeable | Négligeable (< 1 %) | Extrême à élevée (31-100 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) |
| 4 | Corridors de transport et de service | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 4.1 | Routes et voies ferrées | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 4.2 | Lignes de services publics | - | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | Élevée (continue) |
| 5 | Utilisation des ressources biologiques | D | Faible | Restreinte (11-30 %) | Modérée (11-30 %) | Élevée (continue) |
| 5.2 | Cueillette de plantes terrestres | - | Négligeable | Restreinte à petite (1-30 %) | Négligeable (< 1 %) | Élevée (continue) |
| 5.3 | Exploitation forestière et récolte du bois | D | Faible | Restreinte (11-30 %) | Modérée (11-30 %) | Élevée (continue) |
| 6 | Intrusions et perturbations humaines | - | Négligeable | Restreinte (11-30 %) | Négligeable (< 1 %) | Élevée (continue) |
| 6.1 | Activités récréatives | - | Négligeable | Restreinte (11-30 %) | Négligeable (< 1 %) | Élevée (continue) |
| 7 | Modification des systèmes naturels | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | Élevée (continue) |
| 7.1 | Incendies et suppression des incendies | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | Élevée (continue) |
| 7.2 | Gestion et utilisation de l'eau et exploitation de barrages | - | Négligeable | Négligeable (< 1 %) | Élevée (31-70 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) |
| 8 | Espèces et gènes envahissants ou autrement problématiques | CD | Moyen à faible | Restreinte (11-30 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | CD | Moyen à faible | Restreinte (11-30 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 10 | Phénomènes géologiques | - | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) |
| 10.3 | Avalanches et glissements de terrain | - | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) |
| 11 | Changements climatiques et phénomènes météorologiques violents | CD | Moyen à faible | Généralisée (71-100 %) | Modérée à légère (1-30 %) | Élevée (continue) |
| 11.2 | Sécheresses | D | Faible | Généralisée (71-100 %) | Légère (1-10 %) | Élevée (continue) |
| 11.4 | Tempêtes et inondations | D | Faible | Restreinte à petite (1-30 %) | Modérée à légère (1-30 %) | Élevée (continue) |
Les régimes des feux dans le passé et les projections pour l'avenir, sous l'effet des changements climatiques, ont été examinés en détail pour la région de Kootenay-Ouest (Utzig et al., 2011). Au cours de la première moitié du 20e siècle, des feux se sont déclarés presque annuellement et ont brûlé des zones étendues, particulièrement dans la partie sud de la région, les superficies brûlées annuellement dépassant 30 000 ha certaines années (figures 2 et 3 dans Utzig et al., 2011). Un seuil semblait avoir été atteint aux alentours de 1940, la fréquence annuelle des feux ayant beaucoup diminué jusque dans les années 1980 et de légères augmentations s'étant produites par la suite. Cette diminution a été associée à une tendance au refroidissement au printemps et en été et aux efforts de suppression des incendies déployés dans la deuxième moitié du siècle. Les projections futures de tous les modèles montrent des augmentations de la superficie brûlée, les plus fortes augmentations se produisant dans la sous-région du nord. Compte tenu de l'incertitude, les sorties des différents modèles présentent une grande variabilité en ce qui concerne l'augmentation de la fréquence des incendies, mais, d'ici 2050, l'augmentation moyenne projetée pourrait être de jusqu'à 300 fois dans la sous-zone du nord, 30 fois dans la sous-zone du milieu et 15 fois dans la sous-zone du sud. Les augmentations projetées sont plus modestes d'ici 2020, et ce n'est que dans le nord qu'elles dépassent les valeurs historiques enregistrées depuis le début du 20e siècle (figure 9 dans Utzig et al., 2011).
Il existe peu d'information sur la biologie de la limace pygmée, outre les notes sur l'histoire naturelle dans la description de l'espèce (Leonard et al., 2003) et les renseignements relatifs aux mentions subséquentes sur la répartition au Montana et en Colombie-Britannique (Montana Government, 2013; Ovaska et Sopuck, données inédites, 2007-2014). On peut obtenir quelques renseignements généraux en lisant sur la biologie d'autres limaces de la famille des Arionidés.
La limace pygmée est hermaphrodite, c'est-à-dire qu'elle possède les organes reproducteurs femelles et mâles (Leonard et al., 2003). Cependant, comme chez la plupart des gastéropodes pulmonés, il y a probablement échange de sperme entre individus (Tompa, 1984); rien n'indique qu'il y ait autofécondation chez cette espèce.
La ponte n'a été observée qu'une seule fois; une limace de l'Idaho en captivité a pondu trois œufs en juillet (Leonard et al., 2003). Les œufs de forme ovale étaient grands (1 x 1,8 mm comparativement à la longueur du corps de l'adulte reproducteur, qui était de 9 mm) et ont éclos sept semaines plus tard, en septembre. Dans l'Idaho, des individus adultes ont été observés au printemps et à l'automne (Leonard et al., 2003). La base de données du Montana Natural Heritage Program contient 25 mentions d'observation de la limace pygmée de septembre à novembre, la plupart des mentions ayant été faites au mois d'octobre (Montana Government, 2013). En Colombie-Britannique, des individus adultes (compte tenu de la longueur du corps > 9 mm) et des jeunes ont été observés de septembre à octobre (tableau 1), mais seuls quelques relevés ciblés ont été effectués durant d'autres mois. Les jeunes limaces les plus petites avaient une longueur de 2-3 mm et étaient probablement nouvellement écloses.
Les jeunes limaces hivernent vraisemblablement, mais la proportion d'adultes qui le font est inconnue. La durée d'une génération est probablement de un an ou légèrement supérieure, compte tenu de la petite taille du corps des individus adultes et de la durée de vie relativement courte des limaces de la famille des Arionidés en général.
La limace pygmée est souvent associée aux milieux riverains, et semble avoir besoin d'un milieu où le taux d'humidité est élevé. On en sait très peu sur son degré de tolérance aux perturbations de l'habitat, mais il est fort probable que les activités humaines qui modifient l'hydrologie des sites occupés et qui entraînent l'assèchement ou l'inondation du sol forestier aient une incidence négative sur la limace pygmée. En raison de la petite taille de l'espèce et de sa capacité de tirer parti des micromilieux humides, des sous-populations de limaces pygmées pourraient persister dans de petites parcelles d'habitat qui subsistent, pourvu que leurs besoins en matière d'humidité soient comblés. Toutefois, il est peu probable que les parcelles d'habitat isolées d'où l'espèce est disparue soient recolonisées par l'immigration, du moins à court terme.
On ne connaît rien sur les déplacements et la dispersion des limaces pygmées. En général, les limaces terrestres ont une faible capacité de dispersion si elles ne sont pas aidées par les humains ou transportées par des moyens passifs tels que le vent ou l'eau (examen dans Cordeiro, 2004). On ne connaît aucun mode de transport passif pour la limace pygmée, mais il est possible que les limaces puissent involontairement se fixer sur la fourrure de mammifères, par exemple d'ours, comme on l'a présumé pour d'autres limaces (COSEWIC, 2012).
Leonard et al. (2003) mentionnent que des limaces ont été observées en train de se nourrir de lichens poussant sur les débris ligneux grossiers du sol forestier. Les limaces pourraient aussi consommer une grande quantité de champignons, comme on l'a rapporté pour d'autres limaces (Prophysaon coeruleum: McGraw et al., 2002), ce qui pourrait aider à la dispersion de leurs spores. En Colombie-Britannique, des limaces pygmées ont été observées sur des champignons qui poussaient sur des arbres abattus (Ovaska et Sopuck, données inédites, 2013; figure 1). Les limaces pygmées sont probablement la proie d'une variété d'invertébrés du sol forestier, comme les carabes (Coleoptera : Carabidae) et les chilopodes (Chilopoda). En raison de leur petite taille, les limaces ne sont probablement pas chassées activement par les petits mammifères; elles seraient plutôt consommées par les oiseaux s'alimentant sur le sol forestier.
En Colombie-Britannique, les relevés effectués visaient principalement à clarifier la répartition de la limace pygmée et non pas à estimer son abondance (voir Activités de recherche). Les méthodes d'échantillonnage impliquaient essentiellement un ou plusieurs observateurs marchant dans la zone d'intérêt et faisant des recherches sur le sol forestier, en se concentrant sur les micromilieux jugés importants pour les gastéropodes, notamment les grumes en décomposition, l'écorce arrachée, les souches, les roches, d'autres objets pouvant servir d'abris ou les refuges humides et les amas de litière humide de feuilles. Certains des relevés effectués, y compris tous ceux au cours desquels des limaces pygmées ont été observées, tenaient compte de la quantité de temps consacrée à la recherche intensive dans les micromilieux convenables sous la forme d'un indice d'effort de recherche (tableau 1 et références mentionnées).
La taille et les densités des populations sont inconnues, mais les limaces semblent être réparties de façon éparse dans le paysage, même dans les milieux apparemment convenables à l'espèce. La plupart des observations effectuées en Colombie-Britannique mentionnaient de 1 à 3 limaces par site, mais des groupes de 8 à 16 limaces, y compris des jeunes, ont été observés quatre fois (aux sites 2, 8 15 et 34 dans le tableau 1) à l'intérieur d'une zone d'environ 10 à 15 m de diamètre. Dans tous les sites, l'espèce était généralement repérée dans les premières 10 à 15 minutes de recherche, ou elle n'était pas décelée du tout.
La comparaison des mentions de la limace pygmée avec celles d'autres limaces forestières indigènes de la région de Kootenay, en Colombie-Britannique, apporte quelques éclaircissements sur sa rareté relative. Au cours des relevés de Biolinx Environmental Research Ltd. effectués en 2007-2014, on a constaté que la limace-sauteuse pâle (Hemphillia camelus) était répandue et abondante comparativement à la limace pygmée, que la limace à grand manteau (Magnipelta mycophaga; statut selon le COSEPAC : préoccupante) était plus répandue, mais qu'elle était présente seulement dans quelques sites épars, que le Prophysaon andersonii (Reticulate Taildropper) était moins présent (observations moins fréquentes), mais qu'il était largement réparti en Colombie-Britannique, sa répartition s'étendant bien au-delà de la région de Kootenay, et que la limace gainée (Zacoleus idahoensis; COSEWIC, 2016) n'était que rarement présente, dans la partie sud de l'aire de répartition de la limace pygmée.
La limace pygmée n'a été découverte que récemment (2007), et aucune donnée n'existe sur les fluctuations ou les tendances des populations au Canada et aux États-Unis.
Les mentions canadiennes de la limace pygmée les plus proches de la frontière avec les États-Unis se trouvent seulement 4 km au nord de la partie est de l'État de Washington (site 11 dans le tableau 1 et à la figure 3) et 7,6 km au nord de la partie ouest du Montana (site 2 dans le tableau 1 et à la figure 3). Aucune mention de l'espèce n'a été répertoriée dans l'État de Washington, mais l'habitat au sud de la frontière avec la Colombie-Britannique semble être relativement continu. Dans l'ouest du Montana, l'espèce a été observée à moins de 5 km environ de la frontière avec le Canada (Montana Government, 2013). Sur des images de Google Earth®, on aperçoit d'importantes zones de coupe à blanc dans le territoire situé entre les populations, du côté canadien, ce qui pourrait empêcher le déplacement des limaces. Cependant, des échanges d'individus avec les États-Unis pourraient se produire par les parties est et ouest de l'aire de répartition canadienne de la limace pygmée sur des périodes plus longues, dans la mesure où le permettent les limites de la faible capacité de dispersion des limaces et les obstacles représentés par l'utilisation humaine des terres, comme les routes, les zones de coupe à blanc et les agglomérations.
La limite nord de la répartition mondiale de la limace pygmée se trouve dans le sud-est de la Colombie-Britannique, où sa répartition témoigne probablement d'une expansion post-glaciaire à partir de refuges plus au sud. Son expansion vers le nord est probablement limitée par une courte saison de croissance et des hivers longs et froids. Des types de forêts plus secs vers l'est limitent probablement son expansion dans cette direction et expliquent l'absence de l'espèce dans la majeure partie de la région de Kootenay-Est. Une topographie complexe et la mosaïque composée de milieux secs et humides qui en résulte restreignent vraisemblablement sa répartition, tant à l'intérieur de bassins versants qu'entre ceux-ci. L'exploitation forestière a fragmenté encore plus l'habitat et réduit la perméabilité du paysage aux déplacements et au flux génétique. La faible capacité de dispersion et les besoins en matière d'habitat humide limitent la vitesse avec laquelle les limaces peuvent coloniser de nouveaux milieux.
Le calculateur des menaces de l'UICN (Master et al.,2009) a permis d'évaluer les menaces pesant sur la limace pygmée (tableau 3; annexe 2). On a tenu compte des menaces à l'échelle de la répartition canadienne complète de l'espèce pour faire entrer en ligne de compte les sites potentiels non répertoriés, en appliquant les menaces et l'utilisation des terres pour les sites connus. La méthode du calculateur des menaces consiste à donner une valeur à la portée, à la gravité et à l'immédiateté pour chacune des catégories de menaces standards, l'impact global des menaces étant ensuite calculé à partir de ces valeurs.
L'impact global des menaces pour la limace pygmée a été évalué comme étant « élevé à moyen », où cette plage d'impact témoigne de l'incertitude. Selon le scénario d'impact « élevé », il y a deux menaces à impact moyen et quatre menaces à faible impact, tandis que, selon le scénario d'impact « moyen », il y a six menaces à faible impact. Dans le cas des deux principales menaces, les « Changements climatiques et phénomènes météorologiques violents » et « Espèces exotiques (non indigènes) envahissantes », il existe beaucoup d'incertitude concernant la portée dans les dix prochaines années et la gravité de l'impact sur les populations de limaces. Les rubriques qui suivent correspondent aux catégories (ou aux sous-catégories) de menaces du calculateur des menaces, et sont présentées en ordre décroissant approximatif de leur importance perçue.
Les phénomènes météorologiques violents et la fréquence accrue des événements extrêmes découlant des changements climatiques ont été considérés comme des menaces ayant une portée généralisée pour la limace pygmée (71 à 100 % des populations touchées), parce que l'ensemble de l'aire de répartition canadienne de l'espèce subira probablement l'influence des mêmes régimes climatiques généraux. Cependant, les caractéristiques du relief et de l'habitat pourraient moduler les impacts sur les limaces entre bassins versants et entre les sites. Les principaux effets sur les limaces s'aggraveront probablement en raison des sécheresses et des inondations, la fréquence et la gravité de ces deux types d'événements étant censées augmenter selon les scénarios de changements climatiques (Utzig, 2012a). En raison de la dépendance des limaces pygmées envers les milieux très humides, les sécheresses graves et prolongées en été pourraient s'avérer particulièrement dévastatrices pour les sous-populations locales de limaces, tant directement parce qu'elles augmentent la mortalité, qu'indirectement parce qu'elles réduisent la durée de la période servant à la croissance et la la reproduction. Les sécheresses sur plusieurs années consécutives qui persistent jusqu'à tard dans l'automne devraient être particulièrement néfastes. La région de Kootenay, en Colombie-Britannique, a connu des conditions de sécheresse de niveau 2 (sec) en mai, en juin et en juillet 2015, cette période étant suivie de conditions de niveau 3 (très sec) du mois d'août au milieu du mois de septembre (BC Ministry of Forests, Lands and Natural Resource Operations, 2015). Quatre sites où l'espèce avait été observée dans le passé ont été revisités à la fin de septembre 2015, et des observations de l'espèce ont été faites dans deux de ces sites (Ovaska et Sopuck, 2015). La sécheresse pourrait avoir réduit l'abondance ou la détectabilité des limaces, mais la taille de l'échantillon est trop petite pour tirer des conclusions fiables. La détectabilité réduite de l'espèce était particulièrement évidente dans un site à l'extrémité est de l'aire de répartition de l'espèce, où neuf limaces pygmées avaient été observées en 2013, et aucune en 2015. On ne sait pas si les limaces étaient enfoncées plus en profondeur dans le substrat ou si elles avaient connu des déclins.
La fréquence accrue des inondations pourrait entraîner la mortalité ou le déplacement de limaces vivant près des cours d'eau et éroder les zones riveraines de la couche d'humus, éliminant ainsi des refuges. Même si les inondations peuvent être de courte durée le long des cours d'eau de montagne, leurs effets sont potentiellement plus dévastateurs lorsque les limaces vivent dans des milieux à relief plat qui peuvent rester inondés pendant des périodes plus longues.
La gravité des effets sur les limaces pygmées découlant des changements climatiques et des phénomènes météorologiques violents comporte beaucoup d'incertitudes, comme en témoigne la plage étendue de valeurs attribuées dans le calculateur des menaces. Cependant, une approche de précaution est nécessaire en raison de la nature potentiellement généralisée et grave de cette menace. À quelques exceptions près, les effets découlant des changements climatiques sur les gastéropodes terrestres n'ont pas été étudiés. Les études existantes se sont penchées principalement sur les déplacements d'habitat le long de gradients altitudinaux en Europe, et prévoient des rétrécissements des aires de répartition et des déclins des sous-populations des espèces vivant en altitude (Müller et al., 2009), et des déplacements vers une altitude plus élevée dans le cas des espèces vivant à plus basse altitude (Baur et Baur, 2013). Dans le cas des limaces pygmées, il est probable que les facteurs immédiats, comme les sécheresses entraînant des déplacements d'écosystèmes, soient plus importants que les déplacements eux-mêmes; compte tenu de leur faible capacité de dispersion et de leur dépendance envers les milieux humides, les limaces pourraient ne pas être capables de suivre les déplacements d'écosystèmes susceptibles de survenir.
Les gastéropodes non indigènes et les autres invertébrés représentent une menace pour les gastéropodes indigènes, car ils leur font compétition pour la nourriture et les abris, peuvent être leurs prédateurs ou modifier les processus écosystémiques et les milieux. Plus de 20 espèces de gastéropodes non indigènes ont été répertoriées en Colombie-Britannique (Forsyth, 2004). Même si on les retrouve principalement dans les zones perturbées, bon nombre d'entre elles se répandent dans les milieux forestiers. Les humains continuent de faciliter la propagation des gastéropodes introduits à l'échelle de la province; ces derniers sont présents dans la plupart des régions fréquentées par les humains, y compris les sites de pique-nique, les terrains de camping et les aires de repos le long des routes. Parmi les autres invertébrés introduits à grande échelle en Colombie-Britannique, on compte les carabes (Coleptera : Carabidae), qui peuvent se nourrir de gastéropodes (Symondson, 2004), et les vers de terre, qui peuvent réduire (voire éliminer) la couche d'humus par leur comportement, et entraîner des effets potentiellement néfastes sur les invertébrés indigènes du sol forestier (Addison, 2009). Dans la région de Kootenay-Ouest, des gastéropodes introduits ont été observés à deux des 38 sites connus de la limace pygmée (sites 1 et 10), et ils étaient probablement aussi présents à au moins deux autres sites, faciles d'accès par les utilisateurs à des fins récréatives (sites 6 et 11). L'accès accru des humains à l'arrière-pays en raison d'activités d'exploitation de ressources naturelles, et l'expansion du réseau routier contribueront à la propagation de ces gastéropodes et d'autres invertébrés introduits vers de nouvelles régions.
La gravité des effets associés aux espèces introduites sur la limace pygmée comporte un degré élevé d'incertitude, comme en témoigne la plage étendue de valeurs attribuées dans le calculateur des menaces. Les gastéropodes introduits représentent une menace pour les gastéropodes indigènes partout dans le monde (Mahtfeld, 2000), mais leurs effets sur les milieux terrestres sont généralement peu répertoriés. La faune des îles, où des espèces exotiques de prédateurs d'invertébrés et des espèces concurrentes, y compris d'autres gastéropodes, a été, en grande partie, responsable de la disparition d'espèces indigènes d'escargots terrestres (p. ex. à Hawaï : Hadfield et al., 1993; Pacifique Sud : Cowie, 2001) constitue une exception. En Colombie-Britannique, parmi les gastéropodes introduits, on retrouve des nécrophages/prédateurs, comme le Boettgerilla pallens et des espèces du genre Oxychilus, et des herbivores/détritivores, comme des espèces du genre Arion, qui peuvent devenir très abondantes dans les milieux convenables et pourraient posséder un avantage démographique par rapport aux espèces indigènes en compétition pour les ressources. Les carabes sont des prédateurs connus des gastéropodes terrestres dans les milieux naturels et perturbés, et les limaces constituent une grande part du régime alimentaire de bon nombre de Carabidés généralistes (Symondson, 2004). Malgré que les prédateurs des gastéropodes soient généralement des prédateurs spécialisés, la prédation sur les limaces ne semble pas nécessiter d'adaptations spécifiques chez les carabes. Au Royaume-Uni, les carabes, dont le régime est composé en grande partie de limaces sont de taille relativement grande, sont dotés de fortes pièces buccales, sont très mobiles et s'alimentent dans de nombreux milieux (Tod, 1970; cité dans Symondson, 2004). Pour se défendre contre les carabes, les limaces peuvent, entre autres, produire d'importantes quantités de mucus ou du mucus très visqueux, générer des répulsifs ou des substances chimiques toxiques dans leur mucus ou leurs tissus, et amputer leur queue (autotomie) (Symondson, 2004). D'après les connaissances existantes, la limace pygmée ne possède aucun de ces mécanismes et pourrait dépendre du camouflage et de sa petite taille pour éviter la prédation. Il se pourrait, toutefois, que la limace pygmée soit sans défense contre les prédateurs agressifs introduits, comme le carabe Carabus granulatus, qui a été observé en train de se nourrir de limaces indigènes (Hemphillia camelus et Prophysaon andersonii) dans la région de Kootenay-Ouest au cours de travaux sur le terrain réalisés pour le présent rapport; la capacité d'autotomie de la queue ne semblait apporter aucun avantage au P. andersonii (Ovaska et Sopuck, données inédites, 2013). Le site où le C. granulatus a été observé (site 2013-13 dans l'annexe 1) est situé à seulement 2,5 km d'un site connu de la limace pygmée.
La majeure partie de la répartition canadienne de la limace pygmée se trouve sur des terres faisant l'objet d'exploitation forestière. Des zones étendues du paysage ont déjà subi des coupes à blanc et des coupes sélectives, et de nouvelles activités d'exploitation forestière continuent de dégrader l'habitat et de fragmenter l'aire de répartition de l'espèce, mais on manque de données sur la quantité d'habitat qui sera touchée au cours des dix prochaines années. L'exploitation de forêts de seconde venue en maturation a commencé dans la région, à un rythme inconnu en grande partie. Les effets de l'exploitation forestière sur les limaces découleraient de changements des régimes d'humidité et de température du sol forestier associés à l'élimination du couvert forestier, et de perturbations de la végétation du sous-étage et de la structure du sol forestier.
Si l'on se fonde sur une estimation visuelle à partir des couches forestières dans iMapBC (2014) et Google Earth (2014; images de 2003-2013), 95 % des sites connus de la limace pygmée se trouvent dans des paysages faisant actuellement l'objet d'exploitation forestière, mais on ne sait pas combien de nouveaux sites feront l'objet d'exploitation dans les dix prochaines années. Un site seulement (site 35 dans le tableau 1) est protégé de l'exploitation forestière. Les limaces pygmées pourront probablement persister dans de petites parcelles forestières ou à l'intérieur de bandes riveraines boisées dans les sites exploités, du moins à court terme, comme le montre leur présence dans ce genre de milieux dans des paysages récemment exploités. Il est toutefois concevable qu'il y ait un délai avant que les effets complets de l'exploitation forestière récente ne se manifestent, et la viabilité à long terme des sous-populations dans ces milieux demeure inconnue.
Dans les paysages exploités, la limace pygmée pourrait être protégée dans les bandes riveraines boisées. Ces bandes sont requises pour les grands cours d'eau fréquentés par des poissons aux termes de la Forest and Range Practices Act de la Colombie-Britannique, mais elles ne sont pas requises pour les plus petits cours d'eau sans poissons (S6) le long desquels on rencontre généralement la limace pygmée; ces bandes ne sont pas requises non plus pour d'autres entités de drainage non classées, comme les zones de suintement. Cependant, certaines entreprises forestières exploitées dans la région de Kootenay laissent volontairement des bandes le long de tous les cours d'eau, peu importe leur taille ou leur statut (Stuart-Smith, comm. pers., 2014). Malgré ces efforts volontaires, de nombreux cours d'eau de petite taille seront probablement touchés, ce qui accroît la portée de cette menace. Outre les bandes riveraines boisées, il y a généralement une zone d'une largeur de 7 m où la machinerie est interdite le long des ruisseaux, même si des arbres peuvent être coupés dans cette zone. L'habitat de la limace pygmée le long de ruisseaux dans des ravins escarpés se trouverait protégé, parce que le terrain est habituellement trop abrupt pour la récolte de bois (Stuart-Smith, comm. pers., 2014).
Les incendies sont dangereux pour les gastéropodes terrestres, car ils causent leur mortalité directe et, fait peut-être plus important encore, modifient l'habitat en réduisant le nombre d'abris et les sources de nourriture disponibles à court et à long terme (Jordan et Hoffman Black, 2012). En raison de leur mobilité généralement faible, les gastéropodes ne peuvent pas échapper aux incendies en se déplaçant et recolonisent lentement les zones brûlées. Dans la région de Kootenay-Ouest, on prévoit que les feux seront plus fréquents et plus graves en raison des changements climatiques (voir Tendances en matière d'habitat). L'étendue et l'intensité des incendies sont censées avoir une forte incidence sur l'avenir des populations de gastéropodes; les effets seront le plus prononcés probablement lorsque l'incendie couvre une vaste zone continue et s'étend profondément dans le sol, tandis que les effets d'incendies mineurs, discontinus et moins graves seront probablement moins dévastateurs. Dans le deuxième cas, les gastéropodes pourraient survivre dans des refuges souterrains ou dans des parcelles d'habitat non brûlées, qui pourraient constituer des sources pour la recolonisation une fois l'habitat régénéré. Les zones riveraines le long de petits ruisseaux où vivent des limaces pourraient être protégées, dans une certaine mesure, des incendies qui balaient le paysage, particulièrement dans les ravins escarpés et les pentes exposées au nord; on a observé la présence de zones riveraines non brûlées le long de cours d'eau dans des zones récemment touchées par des incendies étendus, au cours de travaux sur le terrain menés aux fins du présent rapport (Ovaska et Sopuck, données inédites, 2007-2013).
Plusieurs études ont rapporté les effets négatifs d'incendies sur la richesse spécifique et/ou l'abondance de gastéropodes terrestres (examen dans Jordan et Hoffman Black, 2012). Les escargots semblent être particulièrement vulnérables (Anderson, 2004; Duncan, 2005), mais des effets sur les limaces ont aussi été rapportés (Duncan, 2005). Dans le sud-ouest de l'Oregon, des brûlages dirigés de faible intensité ont réduit la superficie de l'aire de répartition ainsi que l'abondance de quatre espèces de gastéropodes terrestres (Duncan, 2005). Les effets étaient plus graves sur les escargots que sur les limaces (p. ex. limace-prophyse bleu-gris [Prophysaon coeruleum]), mais les limaces n'étaient plus présentes dans plus du quart des sites qui les abritaient, selon des relevés effectués avant les brûlages. L'auteur croit que, dans les sites où les limaces persistent, ces dernières survivent dans des fissures profondes du substrat rocheux grossier ou dans d'autres refuges souterrains. Il pense aussi que la répartition de micromilieux permettant les déplacements verticaux est importante pour la viabilité à long terme des populations de limaces dans le paysage.
Il y a de nombreux chemins forestiers dans l'aire de répartition de la limace pygmée, et une route importante se trouve à moins de 1 km de quatre sites connus (sites 1, 7 ,10 et 34). Un examen visuel du paysage dans un rayon de 1 km autour de chaque site connu, à l'aide d'iMapBC (2014), a permis d'estimer que la densité routière était « élevée » dans six sites (sites 10, 11, 14,18, 22 et 32), et faible dans sept sites (sites 6, 12, 13, 26, 27, 34 et 35); les 25 autres sites ont été évalués comme présentant une densité routière « moyenne ».
Le nombre de nouvelles routes en lien avec l'exploitation forestière et d'autres types d'extraction des ressources va probablement augmenter au cours des dix prochaines années avec l'expansion de ces activités vers de nouvelles zones. Les effets négatifs sur les limaces causés par ces nouvelles routes découlent de la perte d'habitat dans le corridor routier, et des effets de lisière pouvant s'étendre loin dans la forêt, de l'altération possible des régimes de drainage, de l'assèchement dû à l'augmentation du vent et au rayonnement solaire, et de la fragmentation de l'habitat par les obstacles s'opposant aux déplacements. Les effets négatifs sur les limaces pourraient aussi être exacerbés par la circulation sur les routes existantes, qui lève de la poussière jusque dans la forêt alentour, ou par la réouverture de routes dans des zones exploitées auparavant. Les corridors routiers pourraient aggraver les effets des sécheresses en créant des effets de lisière. Cependant, étant donné que les routes d'accès aux ressources ne sont généralement pas placées le long de cours d'eau, leurs effets sur la limace pygmée dans les milieux riverains sont circonscrits aux traverses de cours d'eau. C'est pourquoi la portée de cette menace a été évaluée comme étant « petite » (1-10 % des limaces touchées) et restera près de la limite inférieure de la plage de valeurs (environ 1 %).
Les animaux d'élevage ne sont généralement pas en liberté dans les versants boisés denses et escarpés caractéristiques de la région de Kootenay-Ouest, et les tenures de pâturage dans l'aire de répartition de la limace pygmée autorisent principalement l'utilisation de forêts claires plus sèches situées au sud et à l'est (iMapBC, 2014). Toutefois, dans les endroits où les bovins et autres animaux d'élevage sont en liberté, on les trouve généralement concentrés dans les zones riveraines, où ils peuvent avoir une incidence sur l'habitat des limaces, car ils compactent le sol et éliminent la végétation du sous-étage. Des signes de la présence de bovins ont été observés dans cinq (13 %) des sites connus occupés par la limace pygmée ou près de ceux-ci (annexe 2).
Les effets cumulatifs sont le résultat d'interactions additives et synergétiques entre deux ou plusieurs menaces, qui pourraient accroître le niveau d'impact global des menaces. Dans le cas de la limace pygmée, les effets cumulatifs risquent d'être plus graves en raison d'interactions entre les menaces suivantes : changements climatiques et phénomènes météorologiques violents, incendies et suppression des incendies, et exploitation forestière. L'accroissement de la fréquence et de la gravité des sécheresses estivales prolongées aura vraisemblablement pour effet d'aggraver les effets de l'exploitation forestière (récente et prévue) et des feux de végétation sur l'habitat des limaces. Par exemple, les étroites bandes riveraines boisées qui auraient normalement soutenu des sous-populations viables de la limace pygmée pourraient ne plus le faire si les sécheresses durent plus longtemps et sont plus fréquentes. Les sécheresses graves augmenteront probablement la fréquence, la superficie et l'intensité des incendies, ce qui pourrait entraîner la perte de sous-populations des zones locales. Ces deux interactions exacerberaient la fragmentation de l'habitat et l'isolement des sous-populations de limaces pygmées. Toute activité qui accroît l'accès par les humains, comme les routes d'accès aux ressources, aggrave le potentiel d'introduction ou de propagation d'espèces non indigènes envahissantes de gastéropodes et d'autres invertébrés. On s'attend aussi à ce que les changements climatiques et les perturbations de la forêt favorisent leur propagation, les impacts sur les espèces indigènes de gastéropodes demeurant, en grande partie, inconnus et non suivis, mais potentiellement graves.
Les plus graves menaces potentielles pesant sur la limace pygmée, selon l'analyse du calculateur des menaces, découlent des changements climatiques et des phénomènes météorologiques violents et des espèces envahissantes introduites. Si on tient compte de chaque bassin versant occupé comme d'une localité distincte, où toutes les limaces pourraient être touchées par un seul événement menaçant (sécheresse grave), on compte au moins 28 localités; le nombre exact dépend de la façon dont on délimite les sous-bassins et de quels sites sont combinés; de plus, il pourrait y avoir d'autres bassins versants où l'espèce est présente. Même s'il est probable que les sécheresses seront de grande échelle et toucheront l'ensemble de la région, il pourrait être plus approprié d'évaluer les effets sur les limaces à l'échelle des bassins versants, selon l'importance de l'exploitation forestière dans le paysage, la largeur des bandes riveraines boisées, la disponibilité de débris ligneux grossiers et d'autres conditions propres aux sites qui ont une incidence sur les refuges des limaces et les régimes d'humidité du sol forestier. De même, les effets des espèces envahissantes introduites pourraient être évalués à l'échelle d'un bassin versant, étant donné que ces espèces peuvent se répandre dans l'ensemble d'un bassin versant après leur introduction initiale.
Actuellement, la limace pygmée ne bénéficie ni d'une protection ni d'un statut officiels aux termes de la Loi sur les espèces en péril du gouvernement fédéral, de la Wildlife Act de la Colombie-Britannique ou d'autres lois.
Les cotes mondiales, nationales et infranationales de NatureServe (2013) pour la limace pygmée sont les suivantes : cote mondiale - G2 (en péril); États-Unis - N2 (en péril); Canada - N1 (gravement en péril); Idaho - S2 (en péril); Montana - S1S2 (gravement en péril à en péril; Colombie-Britannique - S1? (possiblement gravement en péril). En Colombie-Britannique, l'espèce figure sur la liste rouge provinciale des espèces en péril. Au Montana, la limace pygmée est désignée « espèce préoccupante » (Species of Concern) (Montana Government, 2013).
Environ 10 % de l'aire de répartition canadienne de la limace pygmée se trouve à l'intérieur de parcs provinciaux, notamment les parcs provinciaux Valhalla, Kokanee Glacier, West Arm, Lockhart Creek, Kianuko, Stagleap, Champion Lakes et Kootenay Lake (estimation visuelle de la couche des aires protégées dans iMapBC). En mars 2014, le projet de loi 4, une modification à la Parks Act, a été adopté par le gouvernement de la Colombie-Britannique. Le projet de loi visait à permettre le forage d'exploration, l'extraction d'échantillons de minerai et la construction de routes dans les parcs de la Colombie-Britannique. Environ 5 % de l'aire de répartition se trouve à l'intérieur d'aires d'habitat faunique provinciales établies pour d'autres espèces ou dans d'autres aires de conservation (renseignements tirés d'iMapBC). De plus, l'aire de conservation Darkwoods, une vaste propriété privée (55 000 ha) achetée par Conservation de la nature Canada en 2008, située entre les lacs Nelson et Kootenay et adjacente au parc provincial West Arm et à l'aire de gestion de la faune Midge Creek, chevauche aussi une partie de l'aire de répartition principale de la limace pygmée. Cette aire de conservation englobe 4 % de l'aire de répartition de l'espèce, de sorte qu'on atteint un pourcentage total de 19 % de terres protégées. Une autre partie (25 %) de l'aire de répartition se trouve dans des bassins versants approvisionnant une communauté et qui sont protégés dans une certaine mesure. La limace pygmée n'a été observée dans aucune des zones ci-dessus, exception faite du site 11, qui se trouve dans une petite aire de conservation associée à un aménagement hydroélectrique de BC Hydro. De plus, le site 35 se trouve dans le parc provincial des lacs Champion et le site 6, tout juste à côté de ce parc. Les relevés limités effectués dans l'aire de conservation Darkwoods (Copley, données inédites, 2010-2012) n'ont pas permis de repérer des limaces pygmées.
La majeure partie de l'aire de répartition et la plupart des observations de la limace pygmée se trouvent sur des terres forestières provinciales. En tant qu'espèce figurant sur la liste rouge provinciale des espèces touchées par les pratiques forestières et de parcours, la limace pygmée est potentiellement candidate à la gestion aux termes de l'Identified Wildlife Management Strategy de la Forest and Range Practices Act de la Colombie-Britannique. Toutefois, elle n'est pas inscrite comme espèce sauvage désignée à l'heure actuelle; c'est pourquoi aucune mesure de gestion spécifique n'existe ni n'est requise. Les réserves riveraines autour des cours d'eau fréquentés par des poissons, qui sont requises aux termes de la Loi, pourraient aider la limace pygmée à survivre dans les zones d'exploitation forestière, mais une telle protection n'est pas requise autour des plus petits cours d'eau sans poissons (S6). Il convient de noter, cependant, que certaines entreprises forestières laissent volontairement des bandes protégées le long de tous les cours d'eau, y compris ceux classés S6 (Stuart-Smith, comm. pers. 2014).
Durant la préparation du présent rapport, les rédacteurs ont contacté les personnes suivantes - nous les remercions toutes pour les renseignements qu'elles nous ont fournis :
Secrétariat du COSEPAC :
Neil Jones
Julie Perrault
Sonia Schnobb
Jenny Wu
Service canadien de la faune :
Syd Cannings
David Cunnigton
Rhonda Millikin
Parcs Canada :
Patrick Nantel
Représentants du gouvernement de la Colombie-Britannique :
Ted Antifeau
Dave Fraser
Jennifer Heron
Conservation Data Centre de la Colombie-Britannique :
Lea Gelling
- Claudia Copley, Collections Manager (arthropodes), Royal B.C. Museum, Victoria (Colombie-Britannique)
- Robert Forsyth, Research Associate, Royal B.C. Museum, Victoria (Colombie-Britannique)
- Heidi Gardner, Collection Manager (mollusques), Royal British Columbia Museum, Victoria (Colombie-Britannique)
- Paul Hendricks, Montana Natural Heritage Program
- William Leonard, biologiste, Olympia (Washington)
- Dwayne Lepitzki, biologiste, Banff (Alberta)
De plus, les personnes suivantes ont participé à la conférence téléphonique portant sur le calculateur des menaces visant l'espèce, qui s'est déroulée en février 2014 : Ian Adams, Joe Carney, Dave Fraser, Andrew Hebda, Gerry Mackie, Rob McQuarry, Dwayne Lepitzki, Julie Perrault, Kristiina Ovaska, Lennart Sopuck, Kari Stuart-Smith, Charlene Strelaeff. Jenny Wu a préparé les cartes et fourni les calculs de la zone d'occurrence et de l'IZO. Le financement du présent rapport a été assuré par Environnement Canada.
Addison, J.A. 2009. Distribution and impacts of invasive earthworms in Canadian forest ecosystems. Biological Invasions 11:59-79.
Anderson, T. 2004. Callused Vertigo (Vertigo authuri) : a technical conservation assessment. USDA Forest Service, Rocky Mountain Region [Disponible en anglais seulement, consulté en janvier 2014].
Backeljau, T., comm. pers. 2011. Correspondance par courriel avec R. Forsyth, février 2011, professeur, Département des invertébrés, Section de la malacologie, Institut royal des sciences naturelles de Belgique, Bruxelles, Belgique.
Baur, B. et A. Baur. 2013. Snails keep the pace: shift in upper elevation limit on mountain slopes as a response to climate warming. Canadian Journal of Zoology 91:596-597.
BC Ministry of Forests, Lands and Natural Resource Operations. 2015. British Columbia Drought 2015: 2015 Drought Levels at a Glance. NRS GeoHub [Disponible en anglais seulement, consulté en décembre 2015].
Biodiversity Guidebook. 1995. British Columbia Ministry of Forest, Lands and Natural Resource Operations, Forest Practices Code, Biodiversity Guidebook [Disponible en anglais seulement, consulté en janvier 2014].
Bouchet, P. et J.P. Rocroi. 2005. Classification and nomenclator of gastropod families. Malacologia 47:1-397.
Brunsfeld, S.J., J. Sullivan, D.E. Soltis et P.S. Soltis. 2001. Comparative phylogeography of Northwestern North America: A synthesis. P. 319–339, in J. Silvertown et J. Antonovics (eds.). Integrating Ecological and Evolutionary Processes in a Spatial Context. Blackwell Science, Oxford.
Burke, T. 2013. Snails and Slugs of the Pacific Northwest. Oregon State University Press, Cornvallis, Oregon. 337 p.
Columbia Basin Rural Development Institute. 2012. The last 10 years: Growth corridors stabilize population in the Basin Boundary (PDF; 225 KB) [Disponible en anglais seulement, consulté en janvier 2014].
Copley, C. Fichiers de données inédites 2010-2012. Collection manager, Royal British Columbia Museum, Victoria, Colombie-Britannique.
Cordeiro, J. 2004. Population/occurrence delineation – terrestrial snails. NatureServe Explorer : An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. [Disponible en anglais seulement, consulté en décembre 2013].
COSEWIC. 2012. COSEWIC status report on Magnum Mantleslug Magnipelta mycophaga in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 41 pp. (Également disponible en français : COSEPAC. 2012. Évaluation et rapport de situation du COSEPAC sur la limace à grand manteau (Magnipelta mycophaga) au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa, 47 p.).
COSEWIC. 2016. COSEWIC status report on Sheathed Slug Zacoleus idahoensis in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. ? p.
Cowie, R.H. 2001. Invertebrate invasions on Pacific islands and the replacement of unique native faunas: a synthesis of the land and freshwater snails. Biological Invasions 3:119-136.
Duncan, N. 2005. Monitoring of sensitive mollusk populations following low-intensity wildfire in old growth coniferous forest. Unpublished report prepared for USDI Bureau of Land Management, Roseburg District Office, Oregon 97470, USA. 12 p.
Forsyth, R.G. 1999. Distribution of nine new or little-known exotic land snails in British Columbia. The Canadian Field-Naturalist 113:559-568.
Forsyth, R.G. 2004. Land Snails of British Columbia. Royal British Columbia Museum: Victoria, Colombie-Britannique, Canada. 18 p.
Google Earth. 2014. Imagery of the Kootenay Region of British Columbia taken from 2003-2013 [Disponible en anglais seulement, consulté en janvier 2014].
Grieve, D. 2010. Exploration and mining in the Kootenay-Boundary region, British Columbia (PDF; 1.6 MB) [Disponible en anglais seulement, consulté en janvier 2014].
Hadfield, M.G., S.E. Miller et A.H. Carwile. 1993. The decimation of endemic Hawaiian tree snails by alien predators. American Zoologist 33:610-622.
Hendricks, P., comm. pers. 2013. Correspondance par courriel avec K. Ovaska, décembre 2013. Montana Natural Heritage Program.
Hendricks, P. et B.A. Maxwell. 2005. USFS Northern Region 2005 Land Mollusk Inventory: a Progress Report. (Agreement #05-CS-11015600-033) (PDF; 3.3 MB). Rapport inédit, 52 p. [Disponible en anglais seulement, consulté en janvier 2014].
iMapBC. 2014. British Columbia Government, Data BC, Geographic Services, iMapBC 2.0 [Disponible en anglais seulement, consulté en janvier 2014].
Jordan, S.F. et S. Hoffman Black. 2012. Effects of forest land management on terrestrial mollusks: a literature review. Unpublished report prepared for USDA Forest Service, Region 6, and USDI Oregon/Washington Bureau of Land Management by the Xerces Society for Invertebrate Conservation, Portland, Oregon, 87 p.
Ketcheson, M.V., T.F. Braumandl, D. Meidinger, G. Utzig, D.A. Demarchi et B.M. Wikeem. 1991. Chapter 11: Interior Cedar -- Hemlock Zone. P. 168-181, in D. Meidinger and J. Pojar (eds.). Ecosystems of British Columbia. BC Ministry of Forests, Victoria, British Columbia. [Disponible en anglais seulement, consulté en décembre 2013].
Leonard, W.P., comm. pers. 2013. Correspondance par courriel avec Kristiina Ovaska, octobre 2013, biologiste, Olympia, Washington.
Leonard, W.P., L. Chichester, J. Baugh et T. Wilke. 2003. Kootenaia burkei, a new genus and species of slug from northern Idaho, United States (Gastropoda: Pulmonata: Arionidae). Zootaxa 355:1-16.
Leonard, W.P., L. Chichester, C.H. Richart et T.A. Young. 2011. Securicauda hermani and Carinacauda stormi, two new genera and species of slug from the Pacific Northwest of the United States (Gastropoda: Stylommatophora: Arionidae), with notes on Gliabates oregonia Webb 1959. Zootaxa 2746:43-56.
Mahtfeld, K. 2000. Impact of introduced gastropods on molluscan communities, northern North Island. Conservation Advisory Science Notes No. 277.Department of Conservation, Wellington, Nouvelle-Zélande. 18 p.
Master, L., D. Faber-Langendoen, R. Bittman, G.A. Hammerson, B. Heidel, J. Nichols, L. Ramsay et A. Tomaino. 2009. NatureServe conservation status assessments: factors for assessing extinction risk. NatureServe, Arlington, Virginia, 57 p.
McGraw, R., N. Duncan et E. Cazares. 2002. Fungi and other items consumed by the Blue-Gray Taildropper Slug (Prophysaon coeruleum) and the Papillose Taildropper Slug (Prophysaon dubium). The Veliger 45:261-264.
Meidinger, D. et J. Pojar. 1991. Ecosystems of British Columbia. BC Ministry of Forests, Victoria, British Columbia. 330 p.
MFLNRO. 2014a. BC Ministry of Forest, Lands and Natural Resource Operations. Current Allowable Annual Cut (AAC) for Timber Supply Areas (TSA) [Disponible en anglais seulement, consulté en janvier 2014].
MFLNRO. 2014b. BC Ministry of Forest, Lands and Natural Resource Operations. Timber harvesting land base for the Kootenay Lake TSA [Disponible en anglais seulement, consulté en janvier 2014].
Montana Government. 2013. Montana Field Guide: Pygmy Slug – Montana Field Guide: Pygmy Slug - Kootenaia burkei [Disponible en anglais seulement, consulté en janvier 2014].
Müller, J., C. Bässler, C. Strätz, B. Klöcking et R. Brand. 2009. Molluscs and climate warming in a low mountain range national park. Malacologia 51:89-109.
NatureServe. 2013. NatureServe Explorer : An online encyclopedia of life [web application]. Version 7.1. NatureServe, Arlington, Virginia. [Disponible en anglais seulement, consulté en janvier 2014].
Nekola, J., B. Coles et M. Horsak. 2011. Land snail biodiversity assessment for the Selkirk Mountains Park region in southeastern British Columbia. Rapport inédit préparé pour la Valhalla Wilderness Society. 24 p. [Disponible en anglais seulement, consulté en janvier 2014].
Ovaska, K. et L. Sopuck, données inédites. 2007-2013. Données recueillies au cours de relevés des gastéropodes terrestres dans la région Kootenay, dans le cadre de divers projets réalisés par Biolinx Environmental Research Ltd., Sidney, Colombie-Britannique.
Ovaska, K. et L. Sopuck. 2009a. Surveys for terrestrial gastropods at risk in southeastern British Columbia in 2008, and synthesis with 2007 data. Rapport inédit préparé par Biolinx Environmental Research Ltd. pour le ministère de l'Environnement de la Colombie-Britannique, Victoria, Colombie-Britannique, 92 p.
Ovaska, K. et L. Sopuck. 2009b. Surveys for terrestrial gastropods at risk within Ktunaxa Traditional Territory, October 2009. Rapport inédit préparé par Biolinx Environmental Research Ltd. pour le ministère de l'Environnement de la Colombie-Britannique, Victoria, Colombie-Britannique, 27 p.
Ovaska, K. et L. Sopuck. 2014. Terrestrial gastropod surveys in the Kootenay Region, September 2014. Report prepared by Biolinx Environmental Research Ltd. for the Ministry of Environment, Victoria, Colombie-Britannique, 46 p.
Ovaska, K. et L. Sopuck. 2015. Terrestrial gastropod surveys in the in the Kootenay Region, British Columbia, September 2015. Inédit. Préparé par Biolinx Environmental Research Ltd. aux termes du contrat PA16-JHQ-073 pour le ministère de l'Environnement de la Colombie-Britannique, Wildlife Science Section, Vancouver, Colombie-Britannique, 51 p.
Ovaska, K., L. Sopuck et J. Heron. 2010. Terrestrial gastropod surveys on private and municipal land in the Kootenay region, British Columbia, B.C. Ministry of Environment, Terrestrial Conservation Science Section, University of British Columbia Campus, 315-2202 Main Mall, Vancouver, Colombie-Britannique, 47 p.
Pojar, J. 2010. A new climate for conservation (PDF; 8.9 MB). Nature, carbon and climate change in British Columbia. Report prepared for the Working Group on Biodiversity, Forests and Climate. [Disponible en anglais seulement, consulté en janvier 2014].
Stuart-Smith, K., comm. pers. 2014. Conversation téléphonique avec K. Ovaska durant la conférence téléphonique sur le calculateur des menaces. Le 5 février 2014, biologiste, CanFor, Cranbrook, Colombie-Britannique.
Symondson, W.O.C. 2004. Coleoptera (Carabidae, Staphylinidae, Lampyridae, Drilidae and Silphidae) as predators of terrestrial gastropods. P. 37-84, in G.M. Barker (ed.). Natural Enemies of Terrestrial Molluscs. CABI Publishing, Wallingford, Royaume-Uni.
Tod, M.E. 1970. The significance of predation by soil invertebrates on field populations of Agriolimax reticulatus (Gastropoda, Limacidae). Thèse de doctorat, University of Edinburgh, United Kingdom. Cité dans Symondson (2004).
Tompa, A.S. 1984. Land snails (Stylommatophora). P. 47-140, in A.S. Tompa, N.H. Verdonk et J.A.M. van den Biggelaar (eds.). The Mollusca, 7: Reproduction. Academic Press, London and New York.
Utzig, G. 2012a. Climate Change Projections for the West Kootenays. Report # 3 from the West Kootenay Climate Vulnerability and Resilience Project. [consulté en janvier 2014].
Utzig, G. 2012b. Ecosystem and Tree Species Bioclimate Envelope Modeling for the West Kootenays. Report #5 from the West Kootenay Climate Vulnerability and Resilience Project. [consulté en janvier 2014].
Utzig, G., J. Boulanger et R.F Holt. 2011. Climate Change and Area Burned: Projections for the West Kootenays. Report #4 from the West Kootenay Climate Vulnerability and Resilience Project. [consulté en janvier 2014].
Wang, T., E.M. Campbell, G.A. O'Neill et S.N. Aitken. 2012. Projecting future distributions of ecosystem climate niches: uncertainties and management applications. Forest Ecology and Management 279:128-140.
West Kootenay Resilience Program. Sans date. Resilience and climate change: adaptation potential for ecological systems and forest management in the West Kootenays. [consulté en janvier 2014].
Wildsight. 2014. Rivers at Risk. [consulté en janvier 2014].
Kristiina Ovaska, Ph.D., M.Sc., a obtenu son doctorat en biologie à l'Université de Victoria (University of Victoria) et a ensuite réalisé des études postdoctorales en biologie des populations à l'Université de la Colombie-Britannique (University of British Columbia) et en comportement animal à l'Université McGill. Elle est actuellement partenaire chez Biolinx Environmental Research Ltd., biologiste auprès de l'Habitat Acquisition Trust et chercheure associée au Royal British Columbia Museum. Son expérience avec les gastéropodes terrestres englobe des recherches sur les effets des pratiques forestières, des études sur les profils d'abondance et de répartition des espèces en péril, et de nombreux relevés dans différentes parties de la Colombie-Britannique, dont la région de Kootenay. Elle a rédigé des rapports de situation et des documents de rétablissement ainsi que des lignes directrices en matière de pratiques de gestion exemplaires pour des gastéropodes terrestres. Ses photographies de gastéropodes ont paru dans le « Land Snails of British Columbia » de la série Royal BC Museum Handbook, dont l'auteur est R. Forsyth. Elle est l'auteure de plus de 40 publications dans des revues scientifiques à comité de lecture, dont plusieurs articles sur des gastéropodes terrestres.
Lennart Sopuck, M.Sc., biologiste agréé, a étudié de nombreuses espèces sauvages au cours des 30 dernières années. Son domaine d'expertise englobe l'évaluation et l'atténuation des effets de diverses activités humaines sur les espèces sauvages, dont les espèces en péril. Avec Mme Ovaska, il est partenaire chez Biolinx Environmental Research Ltd. et a mené bon nombre de relevés et de projets de recherche sur les gastéropodes terrestres de la Colombie-Britannique. Il est coauteur de plusieurs rapports de situation et programmes de rétablissement, d'un plan d'action visant de multiples espèces, et de documents de gestion visant des espèces de gastéropodes terrestres.
Les collections du Royal British Columbia Museum ont été consultées, mais aucun spécimen n'a été examiné.
| No du site | Description du site | Alt. (m) | Type d’habitat | Âge du peuple-ment (années) | Date | Effort de recherche (min. - pers.) | Espèces observées (nbre d’individus) |
|---|---|---|---|---|---|---|---|
| 2013-1 | Echo Lake Recr. Site, Akolkolex R. FSR, C.-B. | 859 | Forêt de conifères de seconde venue | 80 | 20 sept. 2013 | 50 | Arion rufus (1), Euconulus fulvus (1), Hemphillia camelus (1), Vertigo sp. (1) |
| 2013-2 | Akolkolex-Dumont FSR, C.-B. | 600 | Forêt ancienne de conifères; site où le sol est humide et riche | 200 | 20 sept. 2013 | 60 | Arion sp. (7), Cryptomastix mullani (2), Discus sp. (1), Discus whitneyi (1), Euconulus fulvus (1), Microphysula ingersollii (2), Nesovitrea sp. (1), Vitrina pellucida (3), Zonitoides sp. (1) |
| 2013-3 | Akolkolex FSR (site 1), C.-B. | 646 | Forêt mixte de seconde venue; le long d’un petit ruisseau | 50 | 20 sept. 2013 | 60 | Discus whitneyi (12), Euconulus fulvus (1), Hemphillia camelus (1), Microphysula ingersollii (3), Nesovitrea sp. (2), Punctum randolphii (1), Vertigo sp. (1), Vitrina pellucida (4) |
| 2013-4 | Akolkolex FSR (site 2), C.-B. | 635 | Forêt mixte de seconde venue; le long d’un petit ruisseau | 60-70 | 20 sept. 2013 | 60 | Deroceras laeve (1), Discus sp. (1), Hemphillia camelus (8), Microphysula ingersollii (1) |
| 2013-5 | Ruisseau Little Fish (près de), au bord de la route 23, au sud de Revelstoke, C.-B. | 560 | Forêt mixte de seconde venue; dépression humide | 30-40 | 20 sept. 2013 | 40 | Allogona ptygophora (1), Discus whitneyi (6), Nesovitrea sp. (3), Zonitoides arboreus (1) |
| 2013-6 | Eagle Bay Recr. Site au bord du Shelter Bay FSR, en bordure du lac Arrow, C.-B. | 451 | Forêt de conifères de seconde venue; bande étroite de milieu riverain le long d’un petit ruisseau | 70 | 20 sept. 2013 | 90 | Prophysaon andersoni (3), Zonitoides arboreus (2) |
| 2013-7 | Lac Catherine, à l’ouest du lac Upper Arrow, C.-B. | 833 | Forêt mixte de seconde venue; sur la rive d’un lac | 40-50 | 21 sept. 2013 | 60 | Allogona ptygophora (9), Euconulus fulvus (3), Hemphillia camelus (6), Zonitoides sp. (3) |
| 2013-8 | Fosthall/Mosquito Lake FSR, à l’ouest du lac Upper Arrow, C.-B. | 700 | Forêt de conifères ancienne; dépression humide | 100 | 21 sept. 2013 | 60 | Aucune |
| 2013-9 | Mosquito Lake Recr. Site, à l’ouest du lac Upper Arrow, C.-B. | 682 | Forêt mixte ancienne; bande de forêt étroite qui reste sur la rive d’un lac | 100 | 21 sept. 2013 | 40 | Euconulus pratica, Vertigo sp., Zonitoides sp. |
| 2013-10 | Mosquito Cr. FSR, à l’ouest du lac Upper Arrow, C.-B. | 612 | Jeune forêt mixte de seconde venue; le long d’un petit ruisseau | 30 | 21 sept. 2013 | 60 | Cryptomastix mullani (2), Hemphillia camelus (9), Microphysula ingersollii (1), Nesovitrea sp. (1), Vertigo sp. (1), Vitrina pellucida (3) |
| 2013-11 | Steven's Cr. Recr. Site, à l’ouest du lac Upper Arrow, C.-B. | 842 | Forêt mixte de seconde venue; le long d’un ruisseau à fort débit | 40-50 | 21 sept. 2013 | 60 | Cryptomastix mullani (4), Euconulus fulvus (1), Hemphillia camelus (1), Nesovitrea sp. (1), Punctum randolphii (5), Vertigo sp. (1) |
| 2013-12 | Whatshan R. FSR. (près de l’extrémité est du lac Whatshan), C.-B. | 693 | Forêt mixte de seconde venue; site humide | 60 | 21 sept. 2013 | 40 | Cryptomastix mullani (3), Discus whitneyi (4), Euconulus fulvus (2), Hemphillia camelus (1), Nesovitrea sp. (2), Zonitoides sp. (1) |
| 2013-13 | Parc provincial McDonald Cr. à l’est du lac Arrow, C.-B. | 456 | Forêt mixte de seconde venue; terrain de camping (zone perturbée) | 70 | 21 sept. 2013 | 120 | Allogona ptygophora (1), Arion rufus (2), Cepaea nemoralis (20), Hemphillia camelus (1), Prophysaon andersoni (22) |
| 2013-14 | Slewiskin (McDonald) FSR (site 1), au sud de Nakusp, C.-B. | 745 | Forêt ancienne principalement composée de conifères; le long d’un affluent à fort débit | 100 | 22 sept. 2013 | 40 | Discus whitneyi (5), Euconulus fulvus (3), Vertigo sp. (1), Zonitoides sp. (2), Zonitoides arboreus (1) |
| 2013-15 | Slewiskin (McDonald) FSR (site 2), au sud de Nakusp, C.-B. | 640 | Forêt mixte de seconde venue; le long d’un ruisseau affluent à fort débit | 60-70 | 22 sept. 2013 | 60 | Allogona ptygophora (1), Cryptomastix mullani (3), Discus whitneyi (1), Kootenaia burkei (1), Microphysula ingersollii (2), Nesovitrea sp. (1), Planigyra clappi (6), Punctum randolphii (1), Vertigo sp. (8), Vitrina pellucida (3), Zonitoides arboreus (3) |
| 2013-16 | East Wilson Cr. FSR (site 1), au nord de New Denver, C.-B. | 673 | Jeune forêt mixte de seconde venue; le long d’un ruisseau affluent à fort débit | 40 | 22 sept. 2013 | 50 | Discus whitneyi (20; pourrait comprendre des Radiodiscus), Euconulus fulvus (2), Microphysula ingersollii (1), Nesovitrea sp. (3), Punctum randolphii (1), Radiodiscus abietum (1), Vertigo sp. (10), Zonitoides arboreus (1) |
| 2013-17 | East Wilson Cr. FSR (site 2), au nord de New Denver, C.-B. | 581 | Forêt mixte de seconde venue; le long d’un ruisseau affluent à fort débit | 40-50 | 22 sept. 2013 | 60 | Arion sp. (2), Cryptomastix mullani (3), Discus whitneyi (2), Euconulus fulvus (2), Kootenaia burkei (1), Punctum randolphii (5), Vertigo sp. (10), Zoogenetes harpa (10) |
| 2013-18 | Kane Cr. FSR, à l’est de New Denver, C.-B. | 829 | Forêt mixte de seconde venue; le long d’un ruisseau affluent à fort débit | 70-80 | 22 sept. 2013 | 60 | Arion sp. (3), Euconulus fulvus (1), Hemphillia camelus (2), Nesovitrea sp. (1), Oreohelix sp. (1), Radiodiscus abietum (1), Vertigo sp. (1), Zonitoides arboreus (1) |
| 2013-19 | Keen Cr. FSR, à l’ouest de Kaslo, C.-B. | 758 | Vieille forêt de conifères; zone humide le long d’un ruisseau | 200+ | 22 sept. 2013 | 50 | Discus sp. (1), Euconulus fulvus (3), Pristiloma sp. (1), Vertigo sp. (3), Zonitoides sp. (1) |
| 2013-20 | Parc provincial Kokanee Cr., C.-B. | 558 | Forêt mixte de seconde venue; forêt perturbée dans un terrain de camping | 80 | 22 sept. 2013 | 80 | Arion rufus (3), Cepaea nemoralis (50), Cryptomastix mullani (1), Limax maximus (1) |
| 2013-21 | Sentinel Mtn FSR, à l’est de Castlegar, C.-B. | 526 | Forêt mixte de seconde venue; lisière de forêt en bordure de la route | 50 | 23 sept. 2013 | 50 | Arion sp. (6), Arion intermedius (10), Cryptomastix mullani (9), Deroceras reticulatum (8), Discus whitneyi (25), Euconulus fulvus (4), Nesovitrea sp. ovitrea sp. (2), Vertigo sp. (1), Vitrina pellucida (7), Zonitoides sp. (1) |
| 2013-22 | Ruisseau Murphy, au sud de Castlegar, C.-B. | 437 | Jeune forêt mixte de seconde venue; site perturbé à côté d’un ruisseau | 30 | 23 sept. 2013 | 40 | Cryptomastix mullani (4), Nesovitrea sp. (7), Punctum randolphii (1), Zonitoides arboreus (2) |
| 2013-23 | Ruisseau Casino, au sud-est de Trail, C.-B. | 1 070 | Forêt mixte de seconde venue; zone de suintement dans une forêt sèche | 70 | 23 sept. 2013 | 60 | Anguispira kochi (3), Cryptomastix mullani (4), Euconulus fulvus (2), Punctum randolphii (2), Vertigo sp. (5), Zonitoides arboreus (2) |
| 2013-24 | Chemin Seven Mile, Pend d’Oreille, C.-B. | 521 | Forêt de conifères de seconde venue; forêt riveraine perturbée | 60 | 23 sept. 2013 | 40 | Allogona ptygophora (4), Deroceras reticulatum (4), Euconulus fulvus (1), Haplotrema vancouverense (2), Microphysula ingersollii (1), Prophysaon andersoni (1), Punctum randolphii (2), Vertigo sp. (2) |
| 2013-25 | Ruisseau Nine Mile (site 1), Pend d’Oreille, C.-B. | 703 | Jeune peuplement mixte de seconde venue; forêt riveraine perturbée | 30 | 23 sept. 2013 | 40 | Cryptomastix mullani (1), Deroceras laeve (2), Discus whitneyi (2), Euconulus fulvus (1), Oreohelix sp. (1) |
| 2013-26 | Ruisseau Nine Mile (site 2A), Pend d’Oreille, C.-B. | 618 | Vieille forêt de conifères; forêt humide le long d’un ruisseau | 100 | 23 sept. 2013 | 60 | Allogona ptygophora (5), Cryptomastix mullani (4), Euconulus fulvus (1), Kootenaia burkei (1), Oreohelix sp. (2), Zacoleus idahoensis (2) |
| 2013-27 | Sheep Cr. FSR (site 1), au sud de Salmo, C.-B. | 1 179 | Vieille forêt mixte; forêt riveraine le long d’un ruisseau affluent à fort débit | 100+ | 24 sept. 2013 | 50 | Discus whitneyi (1), Euconulus fulvus (15), Kootenaia burkei (3) |
| 2013-28 | Sheep Cr. FSR (site 2), au sud de Salmo, C.-B. | 969 | Forêt mixte de seconde venue; zone riveraine le long d’un ruisseau à fort débit | 80 | 24 sept. 2013 | 40 | Discus whitneyi (1), Euconulus fulvus (20), Hemphillia camelus (3), Kootenaia burkei (2), Prophysaon andersoni (2), Vertigo sp. (1), Zonitoides arboreus (3) |
| 2013-29 | Ezekiel - Corn Cr. FSR, au sud-ouest de Creston, C.-B. | 841 | Forêt de conifères de seconde venue; le long d’un ruisseau à fort débit | 70-80 | 24 sept. 2013 | 40 | Allogona ptygophora (1), Anguispira kochi (1), Cryptomastix mullani (1), Discus whitneyi (1), Euconulus fulvus (5), Microphysula ingersollii (1) |
| 2013-30 | Spider-Kid Cr. FSR, à l’est de Creston, C.-B. | 961 | Forêt mixte plus ancienne; le long d’un ruisseau à fort débit | 100+ | 24 sept. 2013 | 70 | Cryptomastix mullani (3), Discus sp. (2), Euconulus fulvus (2), Hemphillia camelus (2), Vertigo sp. (5), Zonitoides sp. (1) |
| 2013-31 | Carroll Cr. Road, à l’ouest de Yahk, C.-B. | 993 | Vieille forêt de conifères; zone humide le long d’un ruisseau | 200+ | 24 sept. 2013 | 50 | Kootenaia burkei (1), Zacoleus idahoensis (1) |
| 2013-32 | Gold Cr. FSR, à l’est de Cranbrook, C.-B. | 1 199 | Forêt de conifères de seconde venue; rive humide dans un paysage sec | 80 | 25 sept. 2013 | 40 | Deroceras laeve (1), Discus sp. (1), Euconulus fulvus (2), Vertigo sp. (3), Zonitoides arboreus (1) |
| 2013-33 | Teepee Cr. FSR, au sud-est de Cranbrook, C.-B. | 1 125 | Vieille forêt de conifères; zone humide adjacente à un ruisseau | 100 | 25 sept. 2013 | 40 | Discus whitneyi (1), Euconulus fulvus (1), Kootenaia burkei (9), Vertigo sp. (1) |
| 2013-34 | Plumbob Cr. FSR, au sud-est de Cranbrook, C.-B. | 1 059 | Forêt mixte de seconde venue; dépression humide et zone riveraine le long d’un ruisseau à débit lent | 80 | 25 sept. 2013 | 40 | Discus whitneyi (1), Euconulus fulvus (4), Oreohelix sp. (2) |
| 2013-35 | Caven Cr. FSR, env. 4 km à l’ouest du lac Koocanusa, C.-B. | 810 | Forêt de conifères de seconde venue; dépression humide dans un paysage sec | 80 | 25 sept. 2013 | 40 | Euconulus fulvus (3), Zonitoides arboreus (2) |
| 2014-1A | Jonction des ruisseaux Wait et Lost Dog, env. 20 km au nord-est de Kimberley, C.-B. | 867 | Fond d’un ravin dans une forêt plus sèche | 20 | 16 sept. 2014 | 170 | Euconulus fulvus (10), Microphysula ingersollii (5), Zonitoides arboreus (1) |
| 2014-1B | Jonction des ruisseaux Wait et Lost Dog, env. 20 km au nord-est de Kimberley, C.-B. | 849 | Lit d’un ruisseau affluent dans une plaine inondable (sèche) | 15 | 16 sept. 2014 | 60 | Deroceras reticulatum (25), Euconulus fulvus (1), Nesovitrea sp. (3), Zonitoides arboreus (1) |
| 2014-1C | Jonction des ruisseaux Wait et Lost Dog, env. 20 km au nord-est de Kimberley, C.-B. | 856 | Zone riveraine le long d’un ruisseau dans un pré sur des terres consacrées à l’élevage | S.O. | 15 sept. 2014 | 40 | Deroceras reticulatum (12), Vitrina pellucida (1), Zonitoides nitidus (3) |
| 2014-2A | Kimberley Nature Park (site 1), Kimberley, C.-B. | 1 117 | Zone riveraine le long d’un petit ruisseau à fort débit, dans une forêt ombragée | 70 | 16 sept. 2014 | 50 | I (2), Discus sp. (1), Euconulus fulvus (2), Hemphillia camelus (2), Zonitoides arboreus (2) |
| 2014-2B | Kimberley Nature Park (site 2), Kimberley, C.-B. | 1 114 | Zone riveraine le long d’un petit ruisseau à fort débit sous des peupliers de l’Ouest, dans une forêt ombragée | 70 | 16 sept. 2014 | 30 | Arion circumscriptus (1), Discus whitneyi (3), Euconulus fulvus (3), Nesovitrea sp. (6), Vitrina pellucida (1), Zonitoides arboreus (2) |
| 2014-2C | Kimberley Nature Park (site C, lac Elmer), Kimberley, C.-B. | 1 144 | Zone riveraine le long d’un petit ruisseau se déversant dans le lac Elmer, dans une forêt ombragée | 60 | 16 sept. 2014 | 30 | Discus whitneyi (5), Euconulus fulvus (12), Zonitoides arboreus (2) |
| 2014-3 | Norbury Provincial Park, au nord-est de Cranbrook, C.-B. | 849 | Parcelle d’habitat humide dans un terrain boisé, dans une dépression de basses terres | 50 | 17 sept. 2014 | 70 | Deroceras laeve (3), Discus whitneyi (8), Euconulus fulvus (2), Punctum randolphii (1), Vitrina pellucida (15), Zonitoides nitidus (38) |
| 2014-4 | Plaines Bummers (site 1), au nord-est de Cranbrook, C.-B. | 767 | Lisière de forêt dans une plaine inondable de la riv. Kootenay; parcelle de peupliers de l’Ouest (dont certains sont grands) et taillis d’arbustes | 60 | 17 sept. 2014 | 50 | Deroceras laeve (4), Discus whitneyi (2), Euconulus fulvus (1), Euconulus pratica (4), Nesovitrea sp. (1), Zonitoides nitidus (1) |
| 2014-5 | Aire de repos sur la route 95A (zone du ruisseau Lost Dog), env. 10 km à l’est de Kimberley, C.-B. | 892 | Plaine inondable d’une rivière; peuplements denses d’épinettes le long d’une riv.; inondations périodiques | 100 | 17 sept. 2014 | 60 | Deroceras laeve (1), Deroceras reticulatum (2), Discus whitneyi (5), Euconulus fulvus (2) |
| 2014-6 | Chutes du ruisseau Meachen. (site 1), au sud du lac St. Mary, C.-B. | 1 100 | Ravin le long d’une rivière; beaucoup de chablis | 100 | 18 sept. 2014 | 60 | Hemphillia camelus (1), Vertigo sp. (6) |
| 2014-7 | Meachen Cr. FSR (site 2), env. 11 km au sud du lac St. Mary, C.-B. | 1 208 | Ravin escarpé couvert de mousses d’un ruisseau affluent à fort débit du ruisseau Meachen; substrat rocheux le long du ruisseau | 100+ | 18 sept. 2014 | 50 | Hemphillia camelus (5), Vertigo sp. (5) |
| 2014-8 | Meachen Cr. FSR (site 3 au ruisseau Fiddler), env. 14 km au sud du lac Mary, C.-B. | 1 284 | Versant exposé au nord d’un ravin couvert de jeunes peupliers de l’Ouest le long d’un ruisseau affluent à fort débit; parcelles de litière de feuilles profonde sous des peupliers de l’Ouest | 60-70 | 18 sept. 2014 | 40 | Euconulus fulvus (1), Kootenaia burkei (4), Microphysula ingersollii (1), Vertigo sp. (7) |
| 2014-9 | Meachen Cr. FSR (site 4), au sud du lac St. Mary, C.-B. | 1 457 | Suintement sur le versant nord | - | 18 sept. 2014 | 50 | Hemphillia camelus (8) |
| 2014-10 | Meachen Cr. FSR (site 5), au sud du lac St. Mary, C.-B. | 1 567 | À mi-pente d’un versant de forêt descendant vers une riv.; site humide, mais pas riverain | 150+ | 18 sept. 2014 | 52 | Hemphillia camelus (2) |
| 2014-11 | Hellroaring Cr. FSR (site 1), au sud du lac St. Mary, C.-B. | 1 304 | Lisière de peupliers de l’Ouest à côté de la route dans un versant de forêt abrupt à mi-pente | 60-70 | 18 sept. 2014 | 60 | Discus whitneyi (3), Kootenaia burkei (1), Microphysula ingersollii (3), Punctum randolphii (1), Vertigo sp. (1), Vitrina pellucida (1) |
| 2014-12 | Hellroaring Cr. FSR (site 2), au sud du lac St. Mary, C.-B. | 1 372 | étroite zone riveraine le long d’un ruisseau affluent à fort débit traversant une ancienne zone de coupe à blanc; parcelle de forêt ancienne traversée par une route suivant le ruisseau (côté opposé de la route par rapport à la zone d’étude) | 20 | 18 sept. 2014 | 68 | Euconulus fulvus (2), Hemphillia camelus (9), Magnipelta mycophaga (1), Microphysula ingersollii (2), Punctum randolphii (2), Vertigo sp. (1), Zonitoides sp. (1) |
| 2014-13 | Gold Cr. FSR (site 1), env. 35 km au sud de Cranbrook, C.-B. | 1 113 | Terrain plat dans une plaine inondable le long d’un ruisseau, où poussent de grandes épinettes | 100+ | 19 sept. 2014 | 40 | Deroceras laeve (8), Deroceras reticulatum (5), Discus whitneyi (12), Euconulus fulvus (4), Microphysula ingersollii (1), Zonitoides arboreus (2) |
| 2014-14 | Gold Cr. FSR (site 2), à l’ouest du lac Koocanusa, C.-B. | 972 | Terrain plat bien drainé le long d’un ruisseau | 70 | 19 sept. 2014 | 40 | Discus whitneyi (1) |
| 2014-15 | Wickman Cr. FSR (site 1), au bord du Yahk R. FSR, à l’ouest du lac Koocanusa, C.-B. | 1 159 | Plaine inondable riveraine le long d’un ruisseau et lisière de forêt en terrain élevé (lisière d’aulnes) | 40 | 19 sept. 2014 | 50 | Deroceras laeve (3), Discus whitneyi (8), Euconulus fulvus (1), Microphysula ingersollii (1), Vitrina pellucida (1) |
| 2014-16 | Wickman Cr. FSR (site 2), au bord du Yahk R. FSR, à l’ouest du lac Koocanusa, C.-B. | 1 184 | Peuplement de peupliers de l’Ouest le long d’un ruisseau dans une dépression humide | 40 | 19 sept. 2014 | 50 | Discus whitneyi (2), Euconulus fulvus (2), Hemphillia camelus (3), Microphysula ingersollii (3), Vertigo sp. (2), Zonitoides sp. (1) |
| 2014-17A | Cherry Cr. FSR (site 1), près du lac Cherry, C.-B. | 1 231 | Forêt d’arbres rabougris sur le versant exposé au sud, à l’extrémité sud du lac | 40-50 | 19 sept. 2014 | 70 | Discus whitneyi (2), Zacoleus idahoensis (2), Zonitoides sp. (1) |
| 2014-17B | Cherry Cr. FSR (site 2), à l’extrémité sud-ouest du lac Cherry, C.-B. | 1 221 | Site alluvial riche à côté d’un ruisseau (embouchure/ décharge du lac); faisant l’objet d’une coupe sélective | 80-90 | 20 sept. 2014 | 44 | Hemphillia camelus (3) |
| 2014-17C | Cherry Cr. FSR (site 3), près du lac Cherry, C.-B. | 1 229 | Plaine alluviale à côté d’un ruisseau, peut-être inondée de façon saisonnière; site humide | 80-90 | 20 sept. 2014 | 50 | Discus whitneyi (3), Euconulus fulvus (2), Kootenaia burkei (2), Nesovitrea sp. (1), Zonitoides arboreus (2) |
| 2014-19 | Bloom Cr. FSR (site 1), C.-B. | 1 213 | Ravin dans une forêt de conifères | 100+ | 20 sept. 2014 | 50 | Discus whitneyi (4), Euconulus fulvus (1), Hemphillia camelus (1), Zonitoides arboreus (3) |
| 2014-20 | Bloom Cr. FSR (site 2), C.-B. | 1 246 | Zone riveraine le long d’un ruisseau affluent à fort débit dans une forêt normalement sèche; site ombragé exposé au sud | 50-60 | 20 sept. 2014 | 80 | Euconulus fulvus (1), Hemphillia camelus (11), Microphysula ingersollii (1), Vertigo sp. (2) |
| 2014-21 | Bloom Cr. FSR (site 3), C.-B. | 1 269 | Zone riveraine le long d’un petit ruisseau (filet d’eau) dans une forêt de seconde venue | 60 | 20 sept. 2014 | 60 | Discus whitneyi (2), Euconulus fulvus (1), Hemphillia camelus (5), Kootenaia burkei (1), Punctum randolphii (1), Vertigo sp. (1) |
| 2014-22 | Yahk R. FSR (site 1; près du ruisseau Blacktail), C.-B. | 1 595 | Suintement le long d’un petit ruisseau dans une forêt d’épinettes sur un versant exposé au nord | 120+ | 20 sept. 2014 | 70 | Hemphillia camelus (2), Microphysula ingersollii (3), Punctum randolphii (1), Zacoleus idahoensis (1) |
| 2014-23 | Ruisseau Gilnockie. (site récr.), au bord du Yahk R. FSR, C.-B. | 1 051 | Plaine inondable riveraine à côté d’un ruisseau | 80 | 21 sept. 2014 | 40 | Deroceras reticulatum (1), Euconulus fulvus (1), Hemphillia camelus (1), Zonitoides arboreus (1) |
| 2014-24 | Yahk R FSR (site 2), C.-B. | 1 105 | Zone riveraine d’un ruisseau affluent à débit lent; principalement zone de coupe à blanc, un certain degré de coupe sélective (avec de vieux arbres d’environ 70 ans) | 20 | 21 sept. 2014 | 40 | Allogona ptygophora (2), Oreohelix strigosa (1) |
| 2014-25 | Yahk R. FSR (site 3), C.-B. | 1 111 | Site humide alluvial périodiquement inondé le long d’un plus grand cours d’eau de fort débit (rivière Yahk); trouée du couvert forestier avec une végétation du sous-étage abondante | 100 | 21 sept. 2014 | 40 | Allogona ptygophora (4), Euconulus fulvus (2), Kootenaia burkei (1), Magnipelta mycophaga (1), Microphysula ingersollii (1), Oreohelix sp. (1), Zonitoides arboreus (1) |
| 2014-26 | Yahk R. FSR (site 4), C.-B. | 1 216 | Forêt riveraine le long d’un petit cours d’eau affluent | 60-70 | 21 sept. 2014 | 50 | Discus whitneyi (1) |
| 2014-27 | Yahk R. FSR (site 5), C.-B. | 1 147 | Forêt riveraine le long d’un ruisseau; zone de coupe à blanc de l’autre côté du ruisseau | 80-100 | 21 sept. 2014 | 110 | Aucune |
| 2014-28 | Yahk R. FSR (site 6) sur le Malpas Cr. FSR, C.-B. | 1 323 | Peuplement ancien et humide de conifères, avec des petites trouées du couvert forestier, et dépressions à végétation herbacée | 100+ | 21 sept. 2014 | 50 | Euconulus fulvus (1), Hemphillia camelus (2), Microphysula ingersollii (1), Vertigo sp. (1) |
| 2014-29 | Yahk R FSR (site 7), C.-B. | 1 627 | Forêt riveraine le long d’un ruisseau affluent dans une forêt normalement sèche, à prédominance de pins | 80 | 21 sept. 2014 | 40 | Aucune |
| 2014-30 | Yahk R. FSR (site 8), C.-B. | 1 612 | Parcelle d’arbres dans un ravin | 60-70 | 21 sept. 2014 | 60 | Zacoleus idahoensis (1) |
| 2014-32 | Lamb Cr. FSR (site 1), à l’ouest de Moyie, C.-B. | 1 121 | Plaine inondable riveraine humide dans un ravin étroit (à un seul côté) à l’intérieur d’un paysage de coupe d’abri; plusieurs peupliers de l’Ouest très grands | 70 | 22 sept. 2014 | 40 | Deroceras laeve (1), Discus whitneyi (12), Euconulus fulvus (1), Kootenaia burkei (1), Microphysula ingersollii (1), Punctum randolphii (1), Vertigo sp. (4) |
| 2014-33 | Tate Cr. FSR (site 1), au bord du Lamb Cr. FSR, C.-B. | 1 194 | Bande riveraine (largeur de 50 à 75 m) sur un sol riche de plaine inondable le long d’un ruisseau | 80 | 22 sept. 2014 | 40 | Discus whitneyi (1), Hemphillia camelus (1), Kootenaia burkei (5), Vertigo sp. (1), Vitrina pellucida (1) |
| 2014-34 | Tate Cr. FSR (site 2), au bord du Lamb Cr. FSR, C.-B. | 1 392 | Ravin peuplé d’arbres le long d’un petit ruisseau (zone riveraine < 20 m) et forêt de conifères sur un terrain élevé alentour | 150 | 22 sept. 2014 | 50 | Euconulus fulvus (2), Hemphillia camelus (2), Kootenaia burkei (2), Microphysula ingersollii (1), Vertigo sp. (3) |
| 2014-35 | Tate Cr. FSR (site 3), au bord du Lamb Cr. FSR, C.-B. | 1 384 | Forêt de conifères humide avec peu de végétation du sous-étage, sauf dans les trouées du couvert, et vieux chemin/sentier traversant le site; forêt de transition entre les zones ICH et ESSF | 125+ | 22 sept. 2014 | 40 | Euconulus fulvus (1), Hemphillia camelus (1), Microphysula ingersollii (2), Vertigo sp. (1), Zonitoides arboreus (2) |
| 2014-36 | Irishman R. FSR, près de Moyie, C.-B. | 971 | Plaine inondable d’un ruisseau dans une zone de thuyas, contiguë à une forêt ancienne le long d’un ruisseau | 60-70 | 22 sept. 2014 | 40 | Discus whitneyi (1), Hemphillia camelus (1), Kootenaia burkei (1) |
| 2014-37 | Hawkins-Canuck Cr. FSR (site 1), à l’est de Yahk, C.-B. | 1 041 | Ravin peu profond et petit ruisseau intermittent; site humide exposé au nord avec végétation herbacée abondante | 70 | 23 sept. 2014 | 70 | Allogona ptygophora (7), Anguispira kochi (6), Discus whitneyi (12), Hemphillia camelus (2), Magnipelta mycophaga (2), Prophysaon andersoni (45), Punctum randolphii (1) |
| 2014-38 | Hawkins-Canuck Cr. FSR (site 2), à l’est de Yahk, C.-B. | 1 222 | Ravin peu profond avec ruisseau mouvant et étroite zone riveraine | 50-60 | 23 sept. 2014 | 40 | Discus whitneyi (15), Euconulus fulvus (3), Hemphillia camelus (4), Punctum randolphii (1), Vertigo sp. (4) |
| 2014-39 | American Cr. FSR, près du ruisseau Hawkins et du ch. Meadow, à l’est de Yahk, C.-B. | 1 135 | Trouée du couvert avec végétation herbacée abondante sur un terrain en pente dans une forêt humide; zone de suintement (en grande partie asséchée) sur le site | 60-70 | 23 sept. 2014 | 60 | Anguispira kochi (60), Discus sp. (20), Oreohelix strigosa (4), Prophysaon andersoni (1), Zacoleus idahoensis (1) |
| 2014-40 | Ch. West Yahk, à l’ouest de Yahk, C.-B. | 1 150 | Fond d’un ravin, petit ruisseau affluent (du ruisseau Hawkins) et forêt alentour | 100+ | 23 sept. 2014 | 80 | Allogona ptygophora (1), Discus whitneyi (10), Euconulus fulvus (3), Vitrina pellucida (1), Zacoleus idahoensis (4) |
| 2014-41 | Cold-Freeman FSR (site 1), près du Hawkins FSR, à l’est de Yahk, C.-B. | 1 277 | Lisière de forêt et ravin le long d’un petit ruisseau, parallèle à la route | 70-80 | 23 sept. 2014 | 40 | Deroceras laeve (3), Hemphillia camelus (2), Microphysula ingersollii (1) |
| 2014-42 | Cold-Freeman FSR (site 2), près du Hawkins FSR, à l’est de Yahk, C.-B. | 1 179 | Zone riveraine humide avec des buttes et des dépressions le long d’un ruisseau | 70-80 | 23 sept. 2014 | 40 | Discus whitneyi (3), Euconulus fulvus (2), Kootenaia burkei (2) |
| 2014-43 | Goat R. FSR (site 1), au nord-est de Creston, C.-B. | 849 | Zone riveraine étroite près d’un ruisseau affluent à fort débit | 70 | 24 sept. 2014 | 40 | Discus whitneyi (3), Euconulus fulvus (2), Microphysula ingersollii (3), Punctum randolphii (2) |
| 2014-44 | Skelly Cr. FSR (site 1), au bord du Goat Cr. FSR, au nord-est de Creston, C.-B. | 944 | Plaine inondable riveraine le long d’un ruisseau | 60-70 | 24 sept. 2014 | 40 | Discus whitneyi (1), Kootenaia burkei (1), Microphysula ingersollii (2), Punctum randolphii (1), Vertigo sp. (1) |
| 2014-45 | Skelly Cr. FSR (site 2), au bord du Goat Cr. FSR, au nord-est de Creston, C.-B. | 1 095 | Zone riveraine étroite (largeur de 10 m) le long d’un ruisseau affluent à fort débit, entourée d’une forêt dense de conifères avec peu de végétation du sous-étage | 40-50 | 24 sept. 2014 | 56 | Discus sp. (2), Kootenaia burkei (1), Microphysula ingersollii (1), Vertigo sp. (1) |
| 2014-46 | Goat R. FSR (site 2), au N.-E. de Creston, C.-B. | 1 092 | Dépression humide à moins de 50 m environ d’une rivière | 60-70 | 24 sept. 2014 | 40 | Deroceras laeve (1), Euconulus fulvus (2), Kootenaia burkei (1), Microphysula ingersollii (3), Vertigo sp. (1), Zonitoides arboreus (2) |
| 2014-47 | Mt. Thompson FSR (site 1), à l’est de Creston, C.-B. | 855 | Zone riveraine humide le long d’un ruisseau à fort débit sur un versant de conifères normalement sec avec peu de végétation du sous-étage | 90-100 | 24 sept. 2014 | 54 | Anguispira kochi (1), Discus whitneyi (5), Euconulus fulvus (1), Kootenaia burkei (1), Punctum randolphii (1) |
| 2014-48 | Mt. Thompson FSR (site 2), à l’est de Creston, C.-B. | 1 538 | Zone de suintement dans un ravin/une trouée du couvert | 150+ | 24 sept. 2014 | 40 | Discus whitneyi (1), Euconulus fulvus (1), Vitrina pellucida (1) |
| 2014-49 | Sanca Cr. FSR (site 1), au nord de Creston, C.-B. | 1 189 | Zone riveraine étroite (largeur d’environ 10 m) le long d’un ruisseau affluent à fort débit dans un paysage normalement sec à prédominance de pins; zone humide rare | 40 | 25 sept. 2014 | 40 | Hemphillia camelus (1), Kootenaia burkei (5), Microphysula ingersollii (1), Nesovitrea sp. (1), Punctum randolphii (2) |
| 2014-50 | Sanca Cr. FSR (site 2), au nord de Creston, C.-B. | 1 339 | Versant couvert de conifères dans une forêt ancienne; petite zone de suintement au site | 100+ | 25 sept. 2014 | 40 | Discus whitneyi (4), Euconulus fulvus (4), Hemphillia camelus (1), Punctum randolphii (1), Vertigo sp. (1), Zonitoides arboreus (2) |
| 2014-51 | Sanca Cr. FSR (site 3; South Fork), au nord de Creston, C.-B. | 1 360 | Zone riveraine le long d’un petit ruisseau affluent dans une forêt ancienne de conifères au fond d’une vallée | 150+ | 25 sept. 2014 | 44 | Hemphillia camelus (2), Vertigo sp. (1) |
| 2014-52 | Sanca Cr. FSR (site 4; South Fork), au nord de Creston, C.-B. | 1 585 | Zone riveraine humide dans une plaine inondable le long d’un cours d’eau dans une vieille forêt; chablis abondants et grands rochers | 100+ | 25 sept. 2014 | 40 | Euconulus fulvus (2), Kootenaia burkei (1) |
| 2014-53 | Lac Duck (site 1), vallée de Creston, C.-B. | 544 | Peuplement de peupliers de l’Ouest sur une plaine inondable le long de la rivière Kootenay | 50-60 | 25 sept. 2014 | 30 | Allogona ptygophora (100+), COCLU (1), Deroceras reticulatum (1), Oreohelix strigosa (100+) |
| 2014-55 | Dodge Cr. FSR (site 1), au sud de Creston, C.-B. | 1 052 | Zone riveraine étroite le long d’un petit ruisseau affluent asséché et forêt alentour sur un terrain élevé | 30-40 | 26 sept. 2014 | 46 | Anguispira kochi (13), Discus whitneyi (1), Hemphillia camelus (2), Oreohelix strigosa (1), Vitrina pellucida (1) |
| 2014-56 | Dodge Cr. FSR (site 2) au ruisseau Dodge, au sud de Creston, C.-B. | 1 325 | Zone riveraine dans une jeune forêt près des eaux d’amont du ruisseau Dodge; le paysage est normalement sec avec des zones de coupe à blanc et quelques ruisseaux seulement | 30-40 | 26 sept. 2014 | 80 | Anguispira kochi (4), Discus sp. (3), Euconulus fulvus (3), Hemphillia camelus (7), Kootenaia burkei (1), Microphysula ingersollii (1), Punctum randolphii (1), Vertigo sp. (50), Vitrina pellucida (1) |
| 2014-57 | Blazed Cr./Jersy Cr. FSR en provenance de la route 3, à l’ouest de Creston, C.-B. | 1 102 | Vieille forêt de conifères le long d’un ruisseau à fort débit | 100+ | 26 sept. 2014 | 44 | Anguispira kochi (3), Cryptomastix mullani (1), Discus whitneyi (2), Hemphillia camelus (1), Nesovitrea sp. (1), Oreohelix sp. (1), Zonitoides arboreus (1) |
| 2014-58 | Maryland FSR (site 1) en provenance de la route 3, à l’ouest de Creston, C.-B. | 1 508 | Forêt subalpine claire; très humide | 100+ | 26 sept. 2014 | 54 | Hemphillia camelus (2), Pristiloma chersinella (1) |
| 2014-59B | Lac Boundary par le Boundary L. FSR, à l’ouest de Creston, C.-B. | 1 288 | Forêt ancienne humide avec zones de suintement près des rives du lac | 150+ | 26 sept. 2014 | 50 | Deroceras laeve (1), Discus whitneyi (1), Hemphillia camelus (1), Pristiloma chersinella (1) |
| 2014-59C | Lac Boundary, à l’ouest de Creston, C.-B. | 1 288 | Forêt ancienne humide avec zones de suintement près des rives du lac | 150+ | 26 sept. 2014 | 130 | Hemphillia camelus (2) |
| 2014-60 | Maryland Cr. FSR, à l’ouest du lac Boundary, C.-B. | 1 300 | Vieux peuplement de conifères humide près d’un ruisseau | 125+ | 27 sept. 2014 | 40 | Discus whitneyi (1), Hemphillia camelus (2), Vertigo sp. (1) |
| 2014-61 | Monk Cr. FSR, à l’ouest de Creston, C.-B. | 1 411 | Vieille forêt de conifères humide avec un sol profond et productif dans une dépression | 100+ | 27 sept. 2014 | 60 | Discus whitneyi (5), Kootenaia burkei (2), Magnipelta mycophaga (1), Pristiloma chersinella (5), Punctum randolphii (2) |
| 2014-62 | Parc provincial Stagleap (par l’entrée du Monk Cr. FSR), Kootenay Pass, C.-B. | 1 960 | Forêt ancienne de haute altitude, très humide | 150+ | 27 sept. 2014 | 58 | Hemphillia camelus (1), Vitrina pellucida (1) |
| 2014-64 | Ch. Rosebud Lake, au sud de Salmo, C.-B. | 810 | Zone riveraine d’arbustes le long d’un petit ruisseau dans un ravin à l’intérieur d’un paysage forestier, jeune en grande partie, exploité | 20 | 27 sept. 2014 | 48 | Allogona ptygophora (1), Arion circumscriptus (1), Arion rufus (2) Cochlicopa lubrica (1), Deroceras laeve (2), Discus whitneyi (8), Euconulus fulvus (1), Vertigo sp. (1), Zonitoides sp. (3) |
| 2014-65 | Route 6 vers Nelway (petit embranchement), au sud de Salmo, C.-B. | 668 | Zone riveraine humide le long d’un cours d’eau | 40-60 | 27 sept. 2014 | 126 | Discus whitneyi (1), Haplotrema vancouverense (1), Kootenaia burkei (8), Nesovitrea sp. (2) |
| 2014-66A | Lacs Champion (site 1), au nord de Trail, C.-B. | 1 072 | Forêt ancienne humide où passe un petit ruisseau | 120+ | 28 sept. 2014 | 140 | Anguispira kochi (3), Cryptomastix mullani (5), Discus whitneyi (10), Euconulus fulvus (4), Nesovitrea sp. (1), Punctum randolphii (2), Zonitoides arboreus (1) |
| 2014-66B | Lacs Champion (site 2), au nord de Trail, C.-B. | 1 079 | Forêt ancienne où le bois humide et en état de décomposition avancé est abondant | 100+ | 28 sept. 2014 | 40 | Discus whitneyi (1), Kootenaia burkei (1) |
| 2014-67 | Chemin Nine Mile, au sud de Fruitvale, C.-B. | 850 | Forêt perturbée dans une dépression humide | 40-50 | 28 sept. 2014 | 50 | Cryptomastix mullani (2), Discus whitneyi (10), Euconulus fulvus (6), Hemphillia camelus (2), Nesovitrea sp. (1), Oreohelix strigosa (18), Prophysaon andersoni (12), Zonitoides arboreus (1) |
| 2014-68 | Bear Cr. FSR, au nord de Fruitvale, C.-B. | 724 | Site humide perturbé dans un ravin avec un petit ruisseau dans le paysage d’une forêt plus sèche | 40-50 | 28 sept. 2014 | 40 | Anguispira kochi (16), Cryptomastix mullani (3), Discus whitneyi (16), Hemphillia camelus (1), Nesovitrea sp. (1), Zonitoides arboreus (1) |
| 2014-69 | Bear Cr. FSR (site 2), au nord de Fruitvale, C.-B. | 821 | Ravin avec un petit ruisseau au fond, dans une parcelle de forêt de conifères de seconde venue | 50-60 | 28 sept. 2014 | 40 | Deroceras laeve (1), Discus whitneyi (2), Euconulus fulvus (4), Zonitoides arboreus (1) |
| 2014-70 | Parc provincial King George VI, par la route 22, au sud de Rossland, C.-B. | 693 | Lisière de forêt humide près d’un ruisseau asséché | 80 | 28 sept. 2014 | 40 | Allogona ptygophora (3), Arion circumscriptus (30), Cryptomastix mullani (2), Euconulus fulvus (10), Oreohelix strigosa (1), Prophysaon andersoni (23), Zonitoides arboreus (1) |
| 2014-71 | Archibald-Tillicum FSR (site 1), au sud-ouest de Salmo, C.-B. | 879 | Forêt humide ombragée avec de vieilles grosses souches dans une dépression le long d’un ruisseau | 70 | 29 sept. 2014 | 70 | Discus whitneyi (3), Haplotrema vancouverense (1), Hemphillia camelus (1) |
| 2014-72 | Archibald-Tillicum FSR (site 2), au sud-ouest de Salmo, C.-B. | 1 229 | Zone riveraine humide le long d’un petit ruisseau dans un paysage forestier exploité | 50-60 | 29 sept. 2014 | 50 | Euconulus fulvus (3), Hemphillia camelus (2), Kootenaia burkei (5), Microphysula ingersollii (2), Vertigo sp. (10) |
| 2014-73 | Erie Cr. FSR (site 1), au nord d’Erie, au nord-ouest de Salmo, C.-B. | 991 | Ravin humide le long d’un petit ruisseau dans le paysage d’une jeune forêt sèche (exploitée) | 60-70 | 29 sept. 2014 | 40 | Discus whitneyi (2), Euconulus fulvus (2), Kootenaia burkei (2), Nesovitrea sp. (3), Punctum randolphii (2), Zonitoides arboreus (1) |
| 2014-74 | Erie Cr. FSR (site 2), au nord d’Erie, au nord-ouest de Salmo, C.-B. | 915 | Forêt de plaine d’inondation riveraine humide | 100+ | 29 sept. 2014 | 40 | Hemphillia camelus (3), Kootenaia burkei (1) |
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 2 | 0 |
| D | Faible | 4 | 6 |
| - | Impact global des menaces calculé : | Élevé | Moyen |
| # | Menace | Impact (calculé) |
Impact (description) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | élevée (continue) | - |
| 1.1 | Habitations et zones urbaines | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | élevée (continue) | 1 site (sur 15, 7 % des sites) près d’une ville pourrait faire l’objet d’expansion; sur l’ensemble de l’aire de rép., le développement résidentiel sera probablement très faible dans l’habitat des limaces, malgré que les vallées fluviales pourraient être ciblées. |
| 1.2 | Zones commerciales et industrielles | - | Négligeable | Négligeable (< 1 %) | Extrême (71-100 %) | élevée (continue) | 2 sites (13 %) touchés; une expansion pourrait avoir lieu dans la zone Roseberry ou toucher l’infrastructure liée au barrage de la riv. Pend d’Oreille |
| 1.3 | Tourisme et espaces récréatifs | - | Négligeable | Négligeable (< 1 %) | élevée (31-70 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) | Aucun nouveau plan de développement n’est connu |
| 2 | Agriculture et aquaculture (en anglais seulement) | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | élevée (continue) | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | - | Négligeable | Négligeable (< 1 %) | élevée (31-70 %) | élevée (continue) | Aucun exemple précis connu dans l’habitat des limaces, mais l’expansion des prairies de fauche pourrait avoir lieu; l’habitat des limaces se trouve dans des régions où le sol est productif. |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | - |
| 2.3 | Élevage et élevage à grande échelle | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | élevée (continue) | élevage de bétail surtout dans les zones plus sèches de la région de Kootenay-Ouest; les bovins sont généralement concentrés dans les zones riveraines; leur présence a une incidence sur les plantes du sous-étage et les zones riveraines, car ils compactent le sol et éliminent la végétation; des bovins sont présents dans la zone du ruisseau Hawkins, mais il n’y a pas de bovins élevés en liberté dans la plupart des secteurs de l’aire de répartition de l’espèce. Très peu de tenures de pâturage dans l’aire de répartition de la limace pygmée, surtout dans la partie sud-est de l’aire. |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | - |
| 3 | Production d'énergie et exploitation minière (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) | Extrême à élevée (31-100 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) | - |
| 3.1 | Forage pétrolier et gazier | - | - | - | - | - | Pas de forage ni d’extraction pétrolière et gazière dans l’aire de répartition de la limace pygmée à l’heure actuelle. |
| 3.2 | Exploitation de mines et de carrières | - | Négligeable | Négligeable (< 1 %) | Extrême à élevée (31-100 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) | Il y a des concessions d’exploitation minière souterraine dans le voisinage immédiat de 3 sites (sur 15; 20 % des sites), mais les probabilités que ces mines soient exploitées dans les dix prochaines années sont faibles; beaucoup d’exploration, mais les probabilités que de nouvelles mines soient exploitées sont faibles. Dans le passé, il y a eu beaucoup d’activités d’exploration minière, mais si on porte un regard vers l’avenir, la portée de cette menace est probablement < 1 %. |
| 3.3 | Énergie renouvelable | - | - | - | - | - | Possible sur certaines crêtes, mais aucun exemple connu. |
| 4 | Corridors de transport et de service (en anglais seulement) | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | élevée (continue) | - |
| 4.1 | Routes et voies ferrées | D | Faible | Petite (1-10 %) | Modérée à légère (1-30 %) | élevée (continue) | Tous les sites sont près de routes (à moins de 1 km), et il y a expansion des routes vers de nouvelles zones en raison de l’extraction des ressources. Les effets sur les limaces sont causés par la perte d’habitat (corridors et en raison des effets de lisière), les changements possibles des régimes de drainage, la fragmentation de l’habitat (obstacles aux déplacements) liée aux nouvelles routes, et la dégradation de l’habitat due à la circulation sur les routes existantes (p. ex. qui lève de la poussière). Les nouvelles routes d’accès aux ressources sont rarement, sinon jamais, construites le long de ruisseaux, mais les traversent. Par conséquent, la quantité d’habitat des limaces qui est perdue est petite. La valeur de la portée de cette menace se situe près de la limite minimale (environ 1 %) et pourrait être négligeable. |
| 4.2 | Lignes de services publics | - | Négligeable | Négligeable (< 1 %) | Modérée (11-30 %) | élevée (continue) | Il y a des lignes de transport d’électricité dans 4 sites (sur 15; 27 % des sites). Les effets sont causés par le déboisement lié à l’expansion des lignes existantes ou à la construction de nouvelles lignes; la probabilité que de nouvelles lignes (exception faite de lignes mineures vers des maisons individuelles) soient construites est faible. Les activités de maintenance, comme le débroussaillage, ne constituent pas une menace, parce que la perte d’habitat a déjà eu lieu. La gravité est plus élevée qu’elle ne l’est pour les routes, en raison de l’empreinte et des effets de lisière plus importants. |
| 4.3 | Transport par eau | - | - | - | - | - | - |
| 4.4 | Trajectoires de vol | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | D | Faible | Restreinte (11-30 %) | Modérée (11-30 %) | élevée (continue) | - |
| 5.1 | Chasse et prélèvement d'animaux terrestres | - | - | - | - | - | - |
| 5.2 | Cueillette de plantes terrestres | - | Négligeable | Restreinte à petite (1-30 %) | Négligeable (< 1 %) | élevée (continue) | Cueillette de champignons; principalement dans les brûlis récents, mais aussi dans la forêt; certains sites se trouvent près des zones de cueillette. |
| 5.3 | Exploitation forestière et récolte du bois | D | Faible | Restreinte (11-30 %) | Modérée (11-30 %) | élevée (continue) | De l’exploitation forestière (coupe à blanc, coupe sélective) se déroule à moins de 1 km de toutes les mentions d’observation pour tous les sites; certaines activités ont eu lieu dans le passé (entre les années 1960 et les années 1980), mais des activités de récolte des forêts de seconde venue se déroulent dans certains sites, à tout le moins; il est difficile d’obtenir des renseignements sur les tendances pour les dix prochaines années. Les bandes riveraines atténuent les effets dans une certaine mesure si elles sont laissées en bordure des petits ruisseaux où les limaces sont présentes. Des bandes riveraines boisées ne sont pas requises pour les petits ruisseaux sans poissons (S6), mais certaines entreprises forestières laissent volontairement de telles bandes le long de ces ruisseaux (Kari Stuart Smith, comm. pers., 2013). Malgré ces efforts volontaires, de nombreux cours d’eau de petite taille seront probablement touchés, ce qui accroît la portée de cette menace. Il y a aussi, généralement, une zone d’une largeur de 7 m où la machinerie est interdite le long des ruisseaux, mais des arbres peuvent être coupés dans cette zone. Dans le cas des entités de drainage non classées (comme les zones de suintement), il n’est pas nécessaire de laisser de telles bandes. Les ravins auraient des bandes boisées, parce que le relief y est habituellement trop abrupt pour la récolte. La valeur de la portée est passée de « petite à restreinte » à « restreinte » en raison des commentaires fournis (le 24 juin 2014), compte tenu de l’absence d’exigences relatives aux bandes riveraines boisées dans le cas des petits cours d’eau et des zones de suintement. |
| 5.4 | Pêche et récolte des ressources aquatiques | - | - | - | - | - | - |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | - | Négligeable | Restreinte (11-30 %) | Négligeable (< 1 %) | élevée (continue) | - |
| 6.1 | Activités récréatives | - | Négligeable | Restreinte (11-30 %) | Négligeable (< 1 %) | élevée (continue) | Les activités récréatives touchent six sites connus (sur 15; 40 % des sites), compte tenu de la proximité par rapport aux points de départ de sentiers et aux lieux où d’autres activités récréatives sont possibles; portée réduite étant donné que les sites sont, en grande partie, éloignés des zones très fréquentées (p. ex. parcs très montagneux). Impacts causés par l’utilisation de VTT et de motoneiges (compactage du sol et dommages à la végétation); la randonnée pédestre sur les sentiers a peu d’impact, voire pas du tout. |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | - |
| 6.3 | Travaux et autres activités | - | - | - | - | - | - |
| 7 | Modification du système naturel (en anglais seulement) | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | D | Faible | Petite (1-10 %) | Modérée (11-30 %) | élevée (continue) | La zone ICH est une zone relativement humide, et les événements pouvant entraîner la disparition de peuplements entiers sont rares. Des incendies pourraient se déclarer dans les secteurs plus secs de cette zone. Les rares fois où des incendies se déclarent, ils ont tendance à être particulièrement dévastateurs en raison des mesures de suppression des incendies et des changements climatiques. Les incendies semblent atteindre des températures plus élevées et devenir de plus en plus graves dans la région. Les produits ignifuges utilisés pour combattre les incendies peuvent aussi s’avérer nocifs pour les limaces, mais aucune donnée n’est disponible. |
| 7.2 | Barrages, gestion et utilisation de l'eau | - | Négligeable | Négligeable (< 1 %) | élevée (31-70 %) | Modérée (peut-être à court terme, < 10 ans/3 gén.) | Un site connu se trouve près d’un barrage hydroélectrique, lequel pourrait éventuellement faire l’objet d’une expansion; la variation des niveaux d’eau pourrait aussi avoir une incidence sur ce site. Les projets d’installations au fil de l’eau pourraient avoir une incidence sur les limaces, mais aucun plan d’un projet prévu n’a été trouvé dans iMap dans l’aire de répartition de la limace pygmée (note : les données d’iMap ne sont pas complètes; plusieurs projets sont approuvés ou proposés dans l’aire de répartition de l’espèce, conformément à l’ébauche de rapport révisée). |
| 7.3 | Autres modifications de l'écosystème | - | Inconnu | Inconnue | - | - | Les régimes sylvicoles modifient la composition des forêts, p. ex. plantations de douglas de Menzies, principalement. |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | CD | Moyen à faible | Restreinte (11-30 %) | Modérée à légère (1-30 %) | élevée (continue) | - |
| 8.1 | Espèces exotiques/non indigènes envahissantes | CD | Moyen à faible | Restreinte (11-30 %) | Modérée à légère (1-30 %) | élevée (continue) | Des gastéropodes introduits sont présents ou potentiellement présents dans 4 sites occupés (sur 15; 27 % des sites); d’autres macro-invertébrés introduits, y compris des prédateurs, comme les carabes, pourraient aussi être présents dans ces sites. Le Carabus granulatus, un coléoptère carabidé introduit, a été observé en train de dévorer des limaces indigènes dans la région de Kootenay-Ouest, à quelques kilomètres seulement d’un site connu de la limace pygmée (Ovaska et Sopuck, données. inédites, 2013). Il existe une grande incertitude quant à l’impact des espèces introduites. |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | - | - |
| 8.3 | Introduction de matériel génétique | - | - | - | - | - | - |
| 9 | Pollution (en anglais seulement) | - | - | - | - | - | - |
| 9.1 | Eaux usées domestiques et urbaines | - | - | - | - | - | - |
| 9.2 | Effluents industriels et militaires | - | - | - | - | - | - |
| 9.3 | Effluents agricoles et forestiers | - | Inconnu | Inconnue | Inconnue | - | Déversement de carburant lié à la lutte contre un feu de forêt 2 km en aval du site du ruisseau Lemon (site 4), le 26 juillet 2013. En général, aucun pesticide ou herbicide n'est utilisé dans le cadre de l’exploitation forestière dans la région. Engrais : Ils sont épandus à l’occasion dans les zones nouvellement boisées, mais il ne s’agit pas d’une pratique courante. |
| 9.4 | Détritus et déchets solides | - | - | - | - | - | - |
| 9.5 | Polluants atmosphériques | - | Inconnu | - | - | - | - |
| 9.6 | Énergie excessive | - | - | - | - | - | - |
| 10 | Phénomènes géologiques (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | élevée (continue) | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | élevée (continue) | Par exemple : Un important glissement de terrain s’est produit à Johnsons Landing, en 2012, au bord du lac Kootenay, tout juste au nord-est de l’aire de répartition connue de l’espèce. |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | CD | Moyen à faible | Généralisée (71-100 %) | Modérée à légère (1-30 %) | élevée (continue) | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | Le réchauffement climatique entraînera l’expansion de la zone ICH, mais les limaces ne pourront peut-être pas en profiter à cause du rythme auquel le changement se produira. |
| 11.2 | Sécheresses | D | Faible | Généralisée (71-100 %) | Légère (1-10 %) | élevée (continue) | Il s’agit probablement de la principale menace pesant sur les limaces. On prévoit que les sécheresses estivales seront plus graves et prolongées. Beaucoup d’incertitudes entourent la portée et la gravité de cette menace. Même si le régime climatique évoluera à l’échelle de la région et que les sécheresses toucheront également la région en entier, les effets sur les limaces présentes à divers emplacements de l’aire de répartition seront différents, en raison de différences dans les régimes d’humidité dues aux caractéristiques hydrologiques et topographiques, et de la disponibilité des refuges. Il faut étudier les effets à l’échelle des bassins versants. |
| 11.3 | Températures extrêmes | - | - | - | - | - | Aux limites septentrionales de l’aire de répartition, en Colombie-Britannique; cela ne constitue probablement pas un problème. |
| 11.4 | Tempêtes et inondations | D | Faible | Restreinte à petite (1-30 %) | Modérée à légère (1-30 %) | élevée (continue) | Les inondations posent un problème, compte tenu de l’affinité des limaces pour les milieux riverains. Cependant, celles-ci ont probablement une certaine capacité de survivre aux inondations, qui sont un événement saisonnier survenant naturellement. Les crues printanières pourraient être plus intenses à l’avenir, bien que probablement de courte durée, et pourraient déplacer les limaces. Dans les sites où le terrain est plat, les inondations pourraient entraîner la disparition de populations locales. |
Détails de la page
- Date de modification :