
Évaluation et Rapport de situation du COSEPAC sur le Pannaire jaune pâle Pannaria lurida au Canada - 2016

- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique des rédacteurs du rapport
- Collections examinées
- Figure 1. Répartition du Pannaria lurida s.l. aux États-Unis, d'après les mentions connues du Consortium of North American Lichen Herbaria.
- Figure 2. Occurrences où le Pannaria lurida a été récolté ou signalé au Canada, dans le Maine, aux États-Unis et à Saint-Pierre-et-Miquelon.
- Figure 3. Lieux de récolte de spécimens de lichens au Nouveau-Brunswick et en Nouvelle-Écosse. Ces données ont été recueillies du 19e siècle à aujourd'hui. Les points noirs qui se trouvent dans la mer représentent des lieux de collecte situés sur des îles.
- Figure 4. Occurrences connues du Pannaria lurida qui ont été revisitées, principalement en 2014 (voir le texte). Les points noirs représentent les occurrences de P. lurida qui sont encore existantes au Canada, et les points jaunes, les occurrences où le P. lurida n'a pas été retrouvé.
- Figure 5. Photo du Pannaria lurida poussant de manière luxuriante sur un érable rouge près de Chelsea, en Nouvelle-Écosse. D'autres lichens sont associés avec l'espèce, dont le Lobaria pulmonaria et le Pseudocyphellaria perpetua.
- Figure 6. Thalles de Pannaria lurida et de Lobaria pulmonaria poussant sur le tronc et les branches d'épinettes blanches (Picea glauca) tordues par le vent, à Halls Farm, dans la vallée de Codroy, dans l'ouest de Terre-Neuve.
- Figure 7. Carte de l'habitat prédit du P. lurida dans les Maritimes, au Canada. Les polygones rouges indiquent l'habitat idéal (probabilité de présence > 90 %). Il est à signaler que l'habitat est moins abondant qu'il ne le semble sur la carte, car les polygones ont été agrandis de façon à ce qu'ils soient plus clairement visibles.
- Figure 8. Structure et cycle vital du Pannaria lurida . Les ascospores sont simples (au centre), et les cyanobactéries, visibles dans la coupe du thalle (à droite), forment des chaînes de plus en plus comprimées vers la base de la surface photosynthétique du thalle.
- Figure 9. Caractère convenable actuel (à gauche) et futur (dans 100 ans, à droite) du climat pour le Pannaria lurida dans l'est de l'Amérique du Nord.
- Tableau 1. Occurrences connues du Pannaria lurida en Nouvelle-Écosse, à l'Île-du-Prince-Édouard, au Nouveau-Brunswick et à Terre-Neuve.
- Tableau 1 - Partie A. Occurrences du Pannaria lurida découvertes avant 1986. En l'absence d'une nouvelle visite, on ignore si l'espèce y est encore présente.
- Tableau 1 - Partie B. Occurrences du Pannaria lurida récemment découvertes. Le P. lurida est jugé comme encore présent à moins que l'occurrence ait été revisitée et que l'absence de l'espèce ait été constatée.
- Tableau 2. Nombre estimatif d'individus matures dans chacune des occurrences de Pannaria lurida.
- Tableau 3. Habitat et présence du Pannaria lurida et menaces qui pèsent sur l'espèce dans les occurrences revisitées. La plupart de ces occurrences ont été découvertes avant 1986.
- Annexe 1. Calculateur des menaces pour le Pannaria lurida
COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur lepannaire jaune pâle (Pannaria lurida) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xii + 47 p.
Le COSEPAC remercie David Richardson, Frances Anderson, Robert Cameron, Stephen Clayden, Christopher Pepper et Troy McMullin d'avoir rédigé le rapport de situation sur la pannaire jaune pâle (Pannaria lurida) au Canada, aux termes d'un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par René Belland, coprésident du Sous-comité de spécialistes des mousses et lichens du COSEPAC.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Wrinkled Shingle Lichen Pannaria lurida in Canada.
Pannaire jaune pâle (Pannaria lurida), gracieuseté de Frances Anderson.
La pannaire jaune pâle (Pannaria lurida) est un lichen foliacé qui forme des plaques ou des rosettes pouvant mesurer jusqu'à 10 cm de diamètre. Elle pousse presque toujours sur le tronc de feuillus. La face supérieure du thalle est gris brunâtre et ridée. Le symbiote photosynthétique est une cyanobactérie.
La pannaire jaune pâle est présente en Asie, en Australie, dans les îles du Pacifique, en Afrique et en Amérique. Trois sous-espèces ont été décrites. Ce serait la sous-espèce russellii qui serait présente au Canada et dans le nord-est des États-Unis. Toutefois, il est possible qu'il s'agisse d'une sous-espèce différente, mais aucune analyse moléculaire n'a été réalisée pour confirmer cette théorie.
Au Canada, 56 occurrences de pannaire jaune pâle sont connues : 49 en Nouvelle-Écosse, 4 au Nouveau-Brunswick, 2 à Terre-Neuve et 1 à l'Île-du-Prince-Édouard. Il existe probablement des occurrences qui n'ont pas encore été découvertes, particulièrement en Nouvelle-Écosse et peut-être au Nouveau-Brunswick, voire même à Terre-Neuve.
En Nouvelle-Écosse et au Nouveau-Brunswick, la pannaire jaune pâle colonise les feuillus matures, le plus souvent les érables rouges qui poussent à proximité, mais généralement pas à l'intérieur, des milieux mal drainés. Ainsi, l'espèce se rencontre sur les arbres situés près de la limite des marécages ou des plaines inondables boisés. La seule occurrence signalée à l'Île-du-Prince-Édouard se trouvait sur un thuya, tandis qu'à Terre-Neuve les occurrences ont été signalées sur des épinettes blanches poussant dans un milieu inhabituel, sur une falaise à proximité de la mer.
Les organes de fructification du champignon sont fréquents chez la pannaire jaune pâle et constituent la seule composante spécialisée de reproduction de l'espèce. Les spores qui sont éjectées de ces organes doivent atterrir sur le tronc d'un arbre mature, germer et rencontrer une souche de cyanobactérie compatible du genre Nostoc. Après qu'elle ait été enveloppée par le champignon, la cyanobactérie fournit à celui-ci des glucides et de l'azote, grâce à sa capacité de photosynthèse et de fixation de l'azote atmosphérique. Contrairement à de nombreuses autres espèces de lichens, la pannaire jaune pâle ne produit aucune structure de reproduction végétative. Cependant, la fragmentation du thalle et la fixation de ces fragments sur le tronc des arbres hôtes pourraient permettre la dispersion de l'espèce à une échelle très restreinte.
La population canadienne de pannaire jaune pâle compte environ 5 000 individus. Toutefois, certaines des occurrences connues n'ont pas été revisitées ou n'ont pas fait l'objet de dénombrements, et le nombre d'individus matures variait grandement d'un site à l'autre, de sorte que la population pourrait compter plus de 10 000 individus. Le nombre d'individus par occurrence allait de 1 à un peu plus de 2 400.
Au total, 56 occurrences sont connues au Canada. Des 24 occurrences découvertes avant 1986, 19 ont été revisitées, et l'espèce n'a pas été retrouvée dans 15 d'elles. En outre, l'espèce était absente de deux occurrences récemment découvertes en Nouvelle-Écosse et d'une à l'Île-du-Prince-Édouard. Ainsi, l'espèce était absente de 18 des 56 occurrences, ce qui représente un déclin de 32 %. On suppose que cette diminution du nombre d'occurrences correspond à un déclin équivalent du nombre d'individus matures.
D'après l'analyse du calculateur des menaces, l'impact global des menaces qui pèsent sur le P. lurida est « élevé à très élevé »; la principale menace actuelle est l'exploitation forestière, qui cause la disparition des arbres hôtes ainsi que des modifications du microclimat. Les activités d'exploitation forestière destinée à la production de bois d'œuvre, de bois de chauffage, de copeaux de bois et de biomasse ont des répercussions particulièrement graves, puisque l'espèce colonise généralement les arbres qui présentent une écorce rugueuse, caractère qui apparait après environ 50 ans. La récolte annuelle de bois de feuillus en Nouvelle-Écosse a doublé entre 1990 et 2000, et on s'attend à ce que cette hausse se poursuive, avec la récolte continue de feuillus. Un déclin semblable du nombre de forêts de feuillus anciennes a été observé au Nouveau-Brunswick, mais l'exploitation forestière y est principalement réalisée dans les sites mésiques en terrain élevé.
Les autres menaces d'une gravité moindre qui pèsent sur la pannaire jaune pâle sont les changements climatiques, la construction routière, le développement et la pollution. Une diminution des pluies, un prolongement des périodes de sécheresse estivales et une réduction du brouillard sont prévus en Nouvelle-Écosse, et ces facteurs pourraient entraîner une diminution du taux de croissance ou la mort de la pannaire jaune pâle. Les activités de construction routière ou de développement qui modifient le drainage et entraînent des changements de l'humidité dans les boisés qui environnent ou se trouvent à proximité de l'espèce pourraient nuire à la croissance ou causer la mort de celle-ci. Enfin, l'espèce est sensible au dioxyde de soufre et aux pluies acides. Le niveau de ces deux menaces a diminué au cours des dernières années, mais les émissions continues pourraient surpasser la capacité tampon de l'écorce des arbres hôtes et rendre celle-ci trop acide pour qu'elle soit colonisée par l'espèce.
La cote mondiale de la pannaire jaune pâle est de G3 (vulnérable) à G5 (non en péril). Aux États-Unis, aucune cote n'a été attribuée à l'espèce, ou sa situation n'a pas encore été évaluée. Au Canada, aucune cote n'a été attribuée à la pannaire jaune pâle.
En Nouvelle-Écosse, cinq occurrences sont protégées : une dans le parc national Kejimkujik, une dans le parc provincial Five Islands, une autre dans des terres communes municipales, et une dans une réserve naturelle provinciale. Au Nouveau-Brunswick, à la pointe Clark, une occurrence se trouve dans une aire naturelle protégée.
| Sujet | Information |
|---|---|
| Durée d'une génération (généralement, âge moyen des parents dans la population; indiquez si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN [2011] est utilisée) De 10 à 30 ans, la meilleure estimation étant de 17 ans. | 17 ans |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre d'individus matures? Oui, un déclin est inféré d'après la disparition continue des arbres hôtes de l'espèce, causée par l'exploitation forestière et les changements climatiques. | Oui |
| Pourcentage estimé de déclin continu du nombre total d'individus matures pendant [cinq ans ou deux générations (34 ans)]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] de [réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. Au total, 56 occurrences ont été découvertes avant 1986 ou récemment au Canada. Dix-neuf des 24 occurrences découvertes avant 1986 ont été revisitées, mais l'espèce n'a pas été retrouvée dans 15 d'entre elles. En outre, l'espèce n'était plus présente dans certaines des occurrences plus récemment découvertes, soit deux en Nouvelle-Écosse et une à l'Île-du-Prince-Édouard, ce qui signifie qu'elle est absente de 18 des 56 occurrences. Ainsi, au total 32 % des occurrences sont disparues. Cette diminution du nombre d'occurrences correspondrait à un déclin équivalent du nombre d'individus matures. | 32 % |
| Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations (50 ans)]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] de [réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
Est-ce que les causes du déclin sont
|
|
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? | Non |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 127 338 km2 |
| Indice de zone d'occupation (IZO) (Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté). | 120 km2 |
La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d'occupation totale se trouvent dans des parcelles d'habitat qui sont
|
|
| Nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) (utilisez une fourchette plausible pour refléter l'incertitude, le cas échéant) Il y a 56 occurrences connues de pannaire jaune pâle au Canada : 49 en Nouvelle-Écosse, 4 au Nouveau-Brunswick, 2 à Terre-Neuve-et-Labrador et une à l'Île-du-Prince-Édouard. Les occurrences sont dispersées, de sorte qu'il est peu probable qu'un seul phénomène menaçant puisse toucher plusieurs occurrences en même temps; le nombre de localités est donc considéré comme équivalent au nombre d'occurrences. |
56 |
| Y a-t-il un déclin [observé, inféré ou prévu] de la zone d'occurrence? L'espèce n'est plus présente à l'Île-du-Prince-Édouard. | Oui |
| Y a-t-il un déclin [observé, inféré ou prévu] de l'indice de zone d'occupation? Seize des dix-huit occurrences découvertes avant 1986 sont disparues. | Oui |
| Y a-t-il un déclin [observé, inféré ou prévu] du nombre de sous-populations? | On ne sait pas |
| Y a-t-il un déclin [observé, inféré ou prévu] du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) Les relevés n'ont pas permis de confirmer la présence de l'espèce dans 16 des 19 occurrences découvertes avant 1986 qui ont été revisitées ainsi que dans deux occurrences récemment découvertes en Nouvelle-Écosse et une à l'Île-du-Prince-Édouard. |
Oui |
| Y a-t-il un déclin [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat? Un déclin de l'habitat causé par l'exploitation forestière et les changements climatiques est observé et inféré en Nouvelle-Écosse et au Nouveau-Brunswick. | Oui |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Non |
| Sous-populations (utilisez une fourchette plausible) | Nbre d'individus matures |
|---|---|
| Terre-Neuve | 103 |
| Nouveau-Brunswick | 70 |
| Nouvelle-Écosse *Il est à signaler que le nombre d'individus a été extrapolé d'après le nombre médian de thalles par occurrence (voir la section Abondance) |
4 410 |
| Total : Un dénombrement a été effectué dans seulement un petit nombre d'occurrences récemment découvertes; un grand nombre d'individus peuvent parfois être présents sur un même arbre, et le nombre total d'individus matures pourrait être supérieur à 10 000. | 5 000 à > 10 000 |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Analyse non effectuée |
| Sujet | Information |
|---|---|
| Un calculateur des menaces a t il été rempli pour l'espèce? | Oui (annexe 1) Menaces principales :
|
| Sujet | Information |
|---|---|
| Statut des populations de l'extérieur les plus susceptibles de fournir des individus immigrants au Canada. | L'espèce est en déclin dans le nord des États-Unis. Il y avait deux occurrences dans le Maine, mais aucune d'elles n'a été retrouvée dans le cadre de relevés récents. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Possible, mais improbable |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui, dans la mesure où il reste des milieux propices |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Oui, pour l'instant |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Oui |
| Les conditions de la population source se détériorent elles? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Oui |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
On ne sait pas |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Non |
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Non |
| Sujet | Information |
|---|---|
| COSEPAC | Espèce désignée « menacée » en avril 2016. |
| Sujet | Information |
|---|---|
| Statut | Menacée |
| Code alphanumérique | A2b |
| Justification de la désignation | Ce lichen colonise des arbres feuillus matures, le plus souvent l'érable rouge, et on en connaît 56 occurrences dans les provinces de l'Atlantique. Les relevés n'ont pu confirmer si l'espèce était toujours présente à l'Île-du-Prince-Édouard, à l'une des deux occurrences à Terre-Neuve, à deux des quatre occurrences au Nouveau-Brunswick, ainsi qu'à plusieurs des 49 occurrences connues en Nouvelle-Écosse. Les menaces pesant sur l'espèce incluent l'exploitation forestière continue menant à l'élimination des arbres hôtes, ainsi que l'impact des changements climatiques, résultant en un climat humide convenable moindre. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Correspond au critère de la catégorie « espèce en voie de disparition », A2b. L'espèce semble aujourd'hui absente de 18 de 56 occurrences connues (déclin de 32 %). Les occurrences disparues sont réparties comme suit : 1 à Terre-Neuve, 1 à l'Île-du-Prince-Édouard, 2 au Nouveau-Brunswick et 14 en Nouvelle-Écosse. Dans certains cas, l'habitat a été détruit. Cette perte d'occurrences s'est traduite par une diminution considérable du nombre d'individus matures. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Sans objet. Même si l'IZO est très petit (120 km2), le nombre de localités dépasse le seuil établi, et il n'y a pas de fluctuations extrêmes. |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Sans objet. |
| Critère D (très petite population ou répartition restreinte) | Sans objet. |
| Critère E (Analyse quantitative) | Non effectuée. |
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Nom : Pannaria lurida (Mont.) Nyl. (Mém. Soc. Sci. Nat. Cherbourg 5:109,1857).
Classification : Le genre Pannaria regroupe quelque 80 espèces, dont 8 sont présentes en Amérique du Nord (Brodo et al., 2001; Ekman et al., 2014). Ce genre appartient à la famille des Pannariacées, qui comprend 300 espèces connues réparties entre 30 genres d'après de récentes analyses phylogénétiques (Ekman et al., 2014). La famille des Pannariacées est classée dans l'ordre des Peltigérales, dans la classe des Lécanoromycètes et dans l'embranchement des Ascomycètes.
Trois sous-espèces sont actuellement reconnues pour le Pannaria lurida s.l. :ssp. lurida, ssp. russellii et ssp. quercicola (Jørgensen, 2000). De récentes études portant sur les deux premières sous-espèces ont montré que celles-ci pouvaient être distinguées sur le plan phylogénétique (Ekman et al., 2014). Toutefois, il est difficile de distinguer avec certitude la sous-espèce russellii de la sous-espèce quercicola, qui a été signalée dans le sud de la Nouvelle-Angleterre et dans les Appalaches. Des illustrations en noir et blanc des trois sous-espèces sont présentées dans Jørgensen (2000).
Le Pannaria lurida ssp. russellii (Tuck.) P.M. Jørg. serait la seule sous-espèce présente au Canada et dans le nord des États-Unis (Jørgensen, 2000). Le spécimen type de la sous-espèce a été récolté à Hingham, au Massachusetts, aux États-Unis (J.L. Russell, 1844. Proc. Acad. Arts & Sci. 1848:1;227). Selon Jørgensen (2000, p. 691), la médulle de la sous-espèce ne présenterait généralement pas de réaction à la paraphénylènediamine (Pd). L'application de Pd induit une réaction (tissus virant au rouge) en présence de pannarine. Plus de 30 spécimens frais et spécimens d'herbier de P. lurida s.l., y compris ceux de l'herbier du New York Botanical Garden qui proviennent du sud des États-Unis, des Appalaches et de plus au nord, ont récemment été examinés. Les spécimens de P. lurida ssp. russellii réagissaient à la Pd dans la partie située immédiatement sous l'extrémité des lobes en croissance (Anderson, comm. pers., 2014). Cette réaction a également été observée chez des spécimens de P. lurida provenant de l'est du Canada conservés dans l'Herbier national, à Ottawa (McMullin, comm. pers., 2014). Même si la plupart des spécimens provenant du Canada et du Maine semblent similaires, ils varient quelque peu de la description de Jørgensen (2000) de la sous-espèce, par la couleur du thalle, la forme et la grosseur des spores, la texture de la surface supérieure et la couleur des rhizohyphes (voir la section Description morphologique). Cette variation donne à penser qu'il pourrait exister une autre sous-espèce au Canada et dans le Maine. Malheureusement, aucune analyse moléculaire n'a été réalisée pour vérifier cette théorie; ainsi, aux fins du présent rapport, on considère que le matériel canadien appartient au P. lurida, sans précision de la sous-espèce.
Le Pannaria lurida ssp. lurida (Mont.) Nyl. renferme de la pannarine dans l'ensemble de son thalle (réaction Pd+) et possède des lobes étroits, un hypothalle proéminent ainsi que des spores étroitement ellipsoïdes. Cette sous-espèce est subtropicale, se rencontrant en Floride et dans les États du golfe du Mexique.
Le Pannaria lurida ssp. quercicola (P.M. Jørg.) renferme lui aussi de la pannarine dans l'ensemble de son thalle, mais ses spores sont ellipsoïdes et pointus aux extrémités, et il se rencontre dans les forêts de feuillus à climat tempéré à chaud de l'est de l'Amérique du Nord et du Mexique (Jørgensen, 2000; Nash et al., 2002) et ailleurs dans le monde (voir la section Répartition mondiale, ci-dessous).
Nom commun : Le nom anglais de l'espèce, « Wrinkled Shingle Lichen » (NatureServe, 2014) fait référence aux rides couramment observées sur la face supérieure du thalle. D'autres noms anglais sont utilisés pour désigner l'espèce, dont « Veined Shingle Lichen » (Brodo et al. 2001) et, en Australie, « Matted Lichen » (ALA, 2014).
Le Pannaria lurida, est un lichen foliacé formant des plaques ou des rosettes mesurant 4 à 9 cm de diamètre avec des lobes de 1 à 4 mm de largeur. La face supérieure du thalle est gris brunâtre, ridée sur toute sa surface et souvent couverte d'une pruine, particulièrement près du contour. Les apothécies sont brun-rouge, présentent un rebord thallin et sont souvent ornées de poils ressemblant à des soies autour de leur base. Les asques renferment huit spores incolores simples, largement ellipsoïdes à ovoïdes mesurant environ 10-17 μm × 8-11 μm. La face inférieure du thalle est de couleur havane et se fixe au substrat au moyen de rhizohyphes havane clair à noir-bleu (Anderson, comm. pers., 2014). Le symbiote photosynthétique est une cyanobactérie du genre Nostoc, et sa disposition dans la médulle, entre les hyphes du champignon, fait en sorte que le thalle devient assez gélatineux lorsque mouillé comparativement à d'autres membres du même genre (voir la section Physiologie et adaptabilité). L'espèce est dépourvue d'isidies et de sorédies.
Composition chimique : Le thalle renferme une quantité plus ou moins importante de pannarine, une depsidone se composant d'un hétérocycle central à sept membres fusionné sur deux côtés à des cycles phényle hautement substitués (Blaser et Stoeckli-Evans, 1991; Huneck et Yoshimura, 1996). Ce composé lichénique se trouve souvent en plus forte concentration sur la marge des lobes (Brodo et al., 2001). Il est responsable du changement de couleur (rouge-orange) produit par l'essai chimique à la goutte au paraphénylènediamine. Les essais à la goutte K ou C ne donnent aucune réaction positive.
Le Pannaria lurida peut être confondu avec le Pannaria rubiginosa, mais le thalle de ce dernier ne présente pas de rides et ne devient pas gélatineux lorsque mouillé (Jørgensen, 2000). En outre, le Pannaria rubiginosa possède un hypothalle noir-bleu plus proéminent et présente fréquemment des lobules ronds sur sa marge (Brodo et al., 2001). Le thalle du P. lurida possède une teinte gris-brun distinctive.
On considère que l'espèce forme une seule unité désignable au Canada, car aucune analyse génétique n'a été réalisée à partir de matériel provenant d'Amérique du Nord, et aucune distinction morphologique ou autre différence n'a été observée pour appuyer l'existence de plus d'une unité désignable.
Le genre Pannaria est considéré comme un des groupes primitifs de cyanolichens. Le genre a évolué pendant le Crétacé, lorsque la côte du proto-Pacifique présentait des conditions tempérées et était rattachée à l'actuelle Amérique du Sud et aux zones terrestres du Pacifique Sud, dont une partie est devenue l'Australie (Galloway, 2008). En outre, des études phylogénétiques du symbionte (Nostoc) du genre Pannaria viennent appuyer cette origine ancienne (Elvebakk et al., 2008).
Outre au Canada,le Pannaria lurida s.l. a été signalé dans les pays et territoires français d'outre-mer suivants (nombre de mentions indiqué entre parenthèses à la suite du nom du pays) : Australie (75), Taïwan (3), Équateur (1), Fidji (4), Honduras (1), Indonésie (3), Japon (88), Kenya (6), Corée (7), Malawi (1), Mexique (2), Nouvelle-Calédonie (1), Nouvelle-Zélande (1), Papouasie-Nouvelle-Guinée (4), La Réunion (1), Afrique du Sud (1), Sri Lanka (2), Tanzanie (10), États-Unis (102) (Global Biogeographic Information Facility, 2014), Chili, Madagascar, Philippines, Samoa et Saint-Pierre-et-Miquelon (Jørgensen, 2003; Andreas Beck, comm. pers., 2014; GBIF, 2015).
Le Pannaria lurida s.l. se rencontre depuis les États de l'est du pays jusqu'en Floride et au Texas (Flenniken, 1999) vers le sud et jusqu'au désert de Sonora vers l'ouest (Nash et al., 2002; figure 1). Deux spécimens de P. lurida ssp. russellii ont été récoltés relativement récemment dans le Maine. Le premier a été récolté en 1997 à proximité de la frontière avec le Canada, à l'étang Jordan, dans l'île des Monts Déserts (J. Hinds, comm. pers., 2014), et le deuxième, près de Eagle Hill, à Steuben (A. Dibble, comm. pers.). Wolfgang Maass a signalé en 1981 deux occurrences dans le Maine, assez près de la frontière du Nouveau-Brunswick, à Princeton et à Wesley. Le P. lurida n'a pas été retrouvé dans ces sites lors des recherches menées en 2014. Dans la partie nord-est des États-Unis, un groupe (> 30) très actif de lichénologues professionnels et amateurs (Tuckerman Workshop) mène des relevés lichénologiques depuis plus de 20 ans. Ils ont signalé très peu d'occurrences de P. lurida dans cette région des États-Unis.
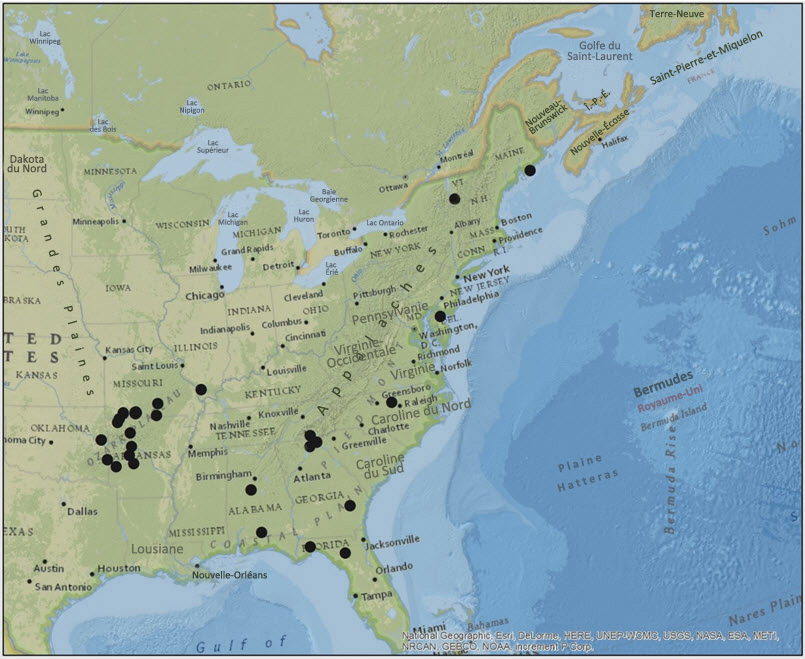
Description longue de la figure 1
Carte de la répartition de la pannaire jaune pâle aux États-Unis, où l'espèce est présente depuis les États de l'est du pays jusqu'en Floride et au Texas vers le sud et jusqu'au désert de Sonora vers l'ouest.
Le Pannaria lurida a été signalé pour la première fois à Terre-Neuve en 1956, en Nouvelle-Écosse en 1981, au Nouveau-Brunswick en 1986 et à l'Île-du-Prince-Édouard en 2009 (tableau 1 - partie A et tableau 1 - partie B). Les mentions de l'espèce par Lepage (1947, 1958) au Québec ont été discréditées (Maass et al., 1986). Enfin, le P. lurida a été signalé au 19e siècle dans les îles de Saint-Pierre-et-Miquelon, mais l'espèce n'a pas été retrouvée lors de récentes recherches dans cette région (Cameron, comm. pers., 2015).
| Numéro | Province | Localité | Comté | Auteur(s) de la récolte | Date de la découverte | Localité revisitée O/N | Espèce encore présente |
|---|---|---|---|---|---|---|---|
| 1 | T.-N. 1 | Woodville, vallée de Codroy | - | Teuvo Ahti | 1956 | O | O |
| 2 | T.-N. 2 | Ruisseau Eels, Parc national du Gros-Morne | - | Teuvo Ahti | 1978 | O | N |
| 1 | N.-B. 1 | Rexton | Northumberland | Wolfgang Maass | 1986 | O | N |
| 2 | N.-B. 2 | Sackville | Westmorland | Wolfgang Maass | 1986 | O | N |
| 1 | N.-É. 1 | Parc provincial Five Islands | Colchester | H. Harries | 1998 | O | O |
| 2 | N.-É. 2 | Rivière St Andrews | Colchester | Wolfgang Maass | 1981 | N | ? |
| 3 | N.-É. 3 | Lac Midway | Digby | Wolfgang Maas | 1982 | O | N |
| 4 | N.-É. 4 | Rivière Sissiboo | Digby | Wolfgang Maas | 1981 | N | ? |
| 5 | N.-É. 5 | Gaetz Brook | Halifax | Wolfgang Maas | 1982 | O | N |
| 6 | N.-É. 6 | Murchyville | Halifax | Wolfgang Maas | 1982 | O | N |
| 7 | N.-É. 7 | Lac Porter | Halifax | Wolfgang Maas | 1982 | O | N |
| 8 | N.-É. 8 | Upper New Cornwall | Lunenburg | Wolfgang Maass | 1981 | O | N |
| 9 | N.-É. 9 | West Northfield | Lunenburg | Brodo et Angus | 19?? | O | N |
| 10 | N.-É. 10 | Greenfield | Queens | Wolfgang Maass | 1982 | O | N |
| 11 | N.-É. 11 | Parc national Kejimkujik | Queens | Wolfgang Maass | 1981 | O | O |
| 12 | N.-É. 12 | Parc national Kejimkujik | Queens | Wolfgang Maass | 1982 | O | O |
| 13 | N.-É. 13 | Kempt | Queens | Wolfgang Maas | 1982 | O | N |
| 14 | N.-É. 14 | Lac Rossignol | Queens | Wolfgang Maas | 1982 | O | N |
| 15 | N.-É. 15 | Meagher | Queens | Wolfgang Maas | 1981 | O | N |
| 16 | N.-É. 16 | Rivière Medway | Queens | Wolfgang Maas | 1982 | O | N |
| 17 | N.-É. 17 | Ponhook | Queens | Wolfgang Maass | 1981 | N | ? |
| 18 | N.-É. 18 | Summerville | Queens | Wolfgang Maass | 1981 | O | N |
| 19 | N.-É. 19 | Chemin Ohio | Shelburne | Wolfgang Maass | 1983 | N | ? |
| 20 | N.-É. 20 | North Kemptville | Yarmouth | Wolfgang Maass | 1981 | N | ? |
| Province | Localité | Comté | Auteur(s) de la récolte | Date de la découverte | Localité revisitée O/N | Espèce encore présente |
|---|---|---|---|---|---|---|
| Î.-P.-É. 1 | Pleasant View | Prince | Troy McMullin | 2009 | O | N |
| N.-B. 1 | Pointe Clark | Charlotte | Stephen Clayden | 2006/7 | N | O |
| N.-B. 2 | Pomeroy Ridge | Charlotte | Stephen Clayden | 2008 | O | O |
| N.-É. 1 | Lac Ruggles | Annapolis | Tom Neily | 2007 | N | O |
| N.-É. 2 | Hardwoodlands | Hants | Chris Pepper | 2014 | N | O |
| N.-É. 3 | Ruisseau Ash | Lunenburg | Frances Anderson | 2008 | N | O |
| N.-É. 4 | Beech Hill | Lunenburg | Frances Anderson | 2009 | N | O |
| N.-É. 5 | Chelsea | Lunenburg | Frances Anderson | 2014 | N | O |
| N.-É. 6 | Chemin Grimm | Lunenburg | Frances Anderson | 2008 | O | N |
| N.-É. 7 | Chutes Indian | Lunenburg | Frances Anderson | 2008 | O | N |
| N.-É. 8 | Indian Path | Lunenburg | Frances Anderson | 2004 | N | O |
| N.-É. 9 | Lapland | Lunenburg | Frances Anderson | 2014 | N | O |
| N.-É. 10 | Lac Rhodes | Lunenburg | Frances Anderson | 2009 | N | O |
| N.-É. 11 | Lac Shingle | Lunenburg | Tom Neily | 2007 | N | O |
| N.-É. 12 | West Clifford | Lunenburg | Troy McMullin | 2014 | N | O |
| N.-É. 13 | Beech Hill | Queens | Tom Neily | 2007 | N | O |
| N.-É. 14 | Lac Bon Mature | Queens | Tom Neily | 2007 | N | O |
| N.-É. 15 | Lac McGowan | Queens | Tom Neily | 2007 | O | O |
| N.-É. 16 | Medway | Queens | Frances Anderson | 2014 | N | O |
| N.-É. 17 | Rivière Grand | Richmond | Chris Pepper | 2014 | N | O |
| N.-É. 18 | Canada Hill | Shelburne | Tom Neily | 2005 - 2011 | N | O |
| N.-É. 19 | Ruisseau Eight Mile? | Shelburne | Chris Pepper | 2014 | N | O |
| N.-É. 20 | Granite Village | Shelburne | Tom Neily | 2012 | N | O |
| N.-É. 21 | Jordan Falls | Shelburne | Chris Pepper | 2014 | N | O |
| N.-É. 22 | Sable River | Shelburne | Tom Neily | 2010 | N | O |
| N.-É. 23 | Ruisseau Six Mile | Shelburne | Frances Anderson | 2007 | N | O |
| N.-É. 24 | Au sud du lac Misery | Shelburne | Tom Neily | - | N | O |
| N.-É. 25 | BaissièreTidney | Shelburne | Frances Anderson | 2008 | N | O |
| N.-É. 26 | Rivière Tidney | Shelburne | Tom Neily | 2005-2011 | N | O |
| N.-É. 27 | Lac Veitch | Shelburne | Chris Pepper | 2014 | N | O |
| N.-É. 28 | À l'ouest du lac Canada Hill | Shelburne | Tom Neily | 2007 | N | O |
| N.-É. 29 | Lac Rushy | Yarmouth | Tom Neily | 2008 | N | O |
La plupart des occurrences connues de Pannaria lurida se trouvent près de la côte de l'Atlantique en Nouvelle-Écosse, au sud d'Halifax, et vers le centre de la province jusqu'au Cap-Breton (figure 2). Au Nouveau-Brunswick, les occurrences se trouvent près du détroit de Northumberland, face à l'Île-du-Prince-Édouard, et dans le sud de la province, à la frontière avec l'État du Maine. À Terre-Neuve et à l'Île-du-Prince-Édouard, l'espèce a été observée uniquement sur les côtes ouest.
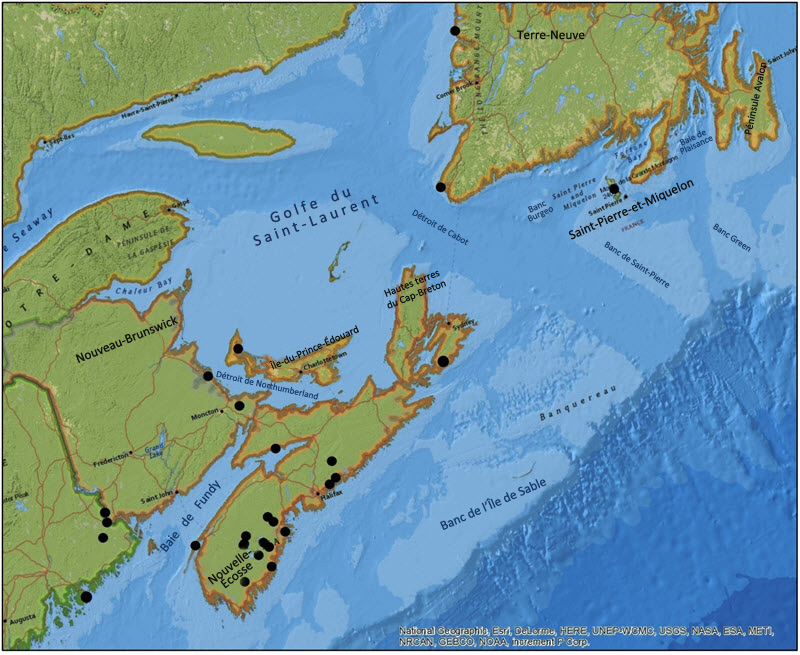
Description longue de la figure 2
Carte montrant les occurrences connues de pannaire jaune pâle au Canada, dans le Maine et à Saint-Pierre-et-Miquelon. Au Canada, l'espèce a été signalée dans l'île de Terre-Neuve, en Nouvelle-Écosse, au Nouveau-Brunswick et à l'Île-du-Prince-Édouard.
Des spécimens de lichens sont récoltés depuis longtemps dans l'est du Canada, comme en font état diverses sources, dont Goward et al. (1998) et Clayden (2010). Aucune des recherches menées avant le début des travaux de terrain réalisés pour la préparation du présent rapport ne ciblait le Pannaria lurida en particulier, mais l'espèce était bien connue des personnes ayant réalisé des relevés généraux (figure 3).
Au total, 56 occurrences de Pannaria lurida sont connues au Canada, dont 24 ont été découvertes avant 1986 (tableau 1A), année où Maass et al. (1986) ont terminé et publié leur description de l'espèce. L'intérêt pour le P. lurida a été ravivé en 2000, avec la publication de Jørgensen (2000). Les récoltes de spécimens ont repris dans les Maritimes en 2005, et l'intervalle d'environ deux générations (~34 ans) s'étant écoulé entre 1986 et les récoltes récentes constitue une division logique entre les occurrences découvertes avant 1986et récemment.
Dix-neuf des occurrences découvertes en 1986 ou antérieurement ont été revisitées pour la préparation du présent rapport, soit 15 en Nouvelle-Écosse, 2 au Nouveau-Brunswick et 2 à Terre-Neuve. Trente-deux occurrences de P. lurida ont été découvertes récemment (après 1986), dont cinq ont été revisitées. Les visites visaient à vérifier la présence de l'espèce et à dénombrer les individus, si possible. Les résultats de ces nouvelles visites sont présentés dans les tableaux 1A et 1B et la figure 4.
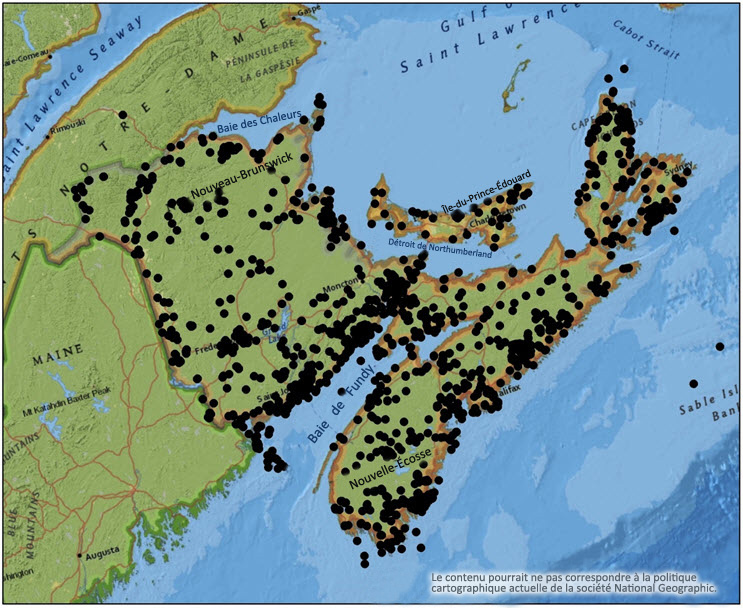
Description longue de la figure 3
Carte des lieux de récolte de spécimens de lichens au Nouveau-Brunswick, en Nouvelle-Écosse et à l'Île-du-Prince-Édouard, selon les données recueillies du 19e siècle à aujourd'hui.

Description longue de la figure 4
Carte des occurrences connues de pannaire jaune pâle qui ont été revisitées, principalement en 2014. Les symboles indiquent si l'espèce était encore présente ou si elle n'a pas été retrouvée.
Une seule occurrence de P. lurida a été signalée à Île-du-Prince-Édouard. L'espèce a été découverte dans cette province en 2009 par T. McMullin dans l'aire naturelle Pleasant View Cedars (McMullin et al., 2012). En 2014, T. McMullin a revisité l'occurrence trois fois à la recherche du P. lurida, mais sans succès. En outre, deux autres zones de l'île potentiellement propices à l'espèce ont été examinées : la réserve naturelle North Enmore et l'aire naturelle Townshend Woodlot. Ces deux régions présentent de relativement vastes zones boisées peuplées de feuillus anciens ainsi qu'un taux d'humidité élevé. La réserve naturelle North Enmore est le seul endroit sur l'île, hormis l'aire naturelle Pleasant View Cedars, qui héberge le P. rubiginosa, mais le P. lurida n'y a pas été trouvé. L'aire naturelle Townshend Woodlot a été visitée par de nombreux lichénologues (Selva en 1995 et 1997, McMullin en 2006 et 2014, Clayden en 2009, et Richardson en 2011), mais aucun n'y a trouvé le P. lurida.
Deux occurrences de Pannaria lurida sont connues dans l'île de Terre-Neuve.
Le P. lurida a été découvert par T. Ahti dans le sud-ouest de Terre-Neuve, à Halls Farm, dans la vallée de Codroy, le 7 juillet 1956. En 2007, M. Pitcher a tenté de retrouver le P. lurida; accompagné de J. Brazil, il a cherché l'espèce dans la forêt de conifères chétifs au sud de Halls Farm et dans la forêt riveraine bordant l'anse au nord du cap Woody, mais sans succès. Pitcher a réalisé de nouvelles recherches au nord de Halls Farm en 2008, accompagné de J. McCarthy. Ils ont retrouvé la population, sur des épinettes blanches (Picea glauca) matures, mais rabougries, sur la bordure d'une falaise maritime de grès, à l'ouest de Millville, dans la vallée de Codroy, dans l'ouest de Terre-Neuve. Cette occurrence a été revisitée en 2010 par Emilie Kissler. Celle-ci a observé le P. lurida sur les branches et le tronc de 14 arbres, principalement des épinettes blanches, et a dénombré 103 thalles au total.
T. Ahti a également trouvé le P .lurida dans un site près du ruisseau Eels, à l'embouchure du ruisseau Western, dans l'ouest de Terre-Neuve. Ce site était occupé par un boisé côtier d'Abies balsamea exposé aux vents et placé derrière des dunes de sable, à une altitude de 6 m. L'espèce poussait sur le Picea glauca. En 2014, Ahti et M. Piercey-Normore ont visité cette région, mais n'ont pas retrouvé le P. lurida. Ils ont également cherché le P. lurida près de l'anse Sally, mais n'y ont pas non plus trouvé l'espèce.
Le Species Status Advisory Committee (SSAC) de Terre-Neuve a jugé qu'il était nécessaire que des travaux de terrain supplémentaires soient réalisés. Le modèle heuristique mis au point par R. Cameron pour l'Erioderma pedicellatum a été appliqué par E. Kissler aux zones de Terre-Neuve que Cameron avait déjà ciblées comme étant les plus propices au P. lurida. Les zones où la présence de l'espèce a été prédite par le modèle comprennent la péninsule d'Avalon, la baie de l'Eau, la baie d'Espoir, la vallée de Codroy, la baie de Port au Port et la baie des Îles (Kissler, 2010). Des travaux de terrain ont ensuite été réalisés dans la vallée de Codroy, à Port-au-Port /Stephenville Crossing, à Three Mile Rock, dans la péninsule Northern et dans la région allant de Cow Head à Port aux Choix, mais aucune nouvelle population de P. lurida n'a été trouvée (Kissler, 2010). En outre, M. Pitcher a mené un relevé dans la région nord-est de la péninsule Avalon, au nord de St. John's, pour le compte de l'East Coast Trail Corporation. Il a également mené des relevés ciblant le P. lurida dans la région de Cow Head, dans une forêt ancienne, près d'un petit cimetière côtier, mais n'a pas trouvé l'espèce. En 2014, C. Hanel a visité un site sur la côte ouest de la péninsule Port au Port, le long d'une piste côtière, où les conditions étaient semblables à celles de l'occurrence de Halls Farm, mais n'y a pas trouvé le P. lurida.
Le P. lurida a également été signalé au 19e siècle dans les îles de Saint-Pierre et de Miquelon, situées à proximité de Terre-Neuve (Delamare et al., 1888; Le Gallo, 1952). Cette région a été revisitée en 2015, mais l'espèce n'y a pas été trouvée (Cameron, comm. pers., 2015).
Quatre occurrences de P. lurida sont connues au Nouveau-Brunswick : pointe Clark, Pomeroy Ridge, Rexton, et nord de Sackville. L'espèce n'était pas présente dans au moins deux de ces occurrences lorsque celles-ci ont été revisitées en 2014 (voir le tableau 1B). En outre, les participants du Tuckerman Workshop ont récolté des spécimens de lichen le long de la côte de l'embouchure de la baie de Fundy, mais n'ont pas trouvé l'espèce.
Des relevés lichénologiques généraux ont été menés en Nouvelle-Écosse, notamment par les participants du Tuckerman Workshop (voir la section Aire de répartition aux États-Unis), qui ont visité la province à deux occasions (pointe White et Parrsboro) ainsi que Terre-Neuve à une occasion (péninsule Avalon) au cours des dernières années et sont capables d'identifier le P. lurida.
Dans le cadre des travaux de terrain menés pour la préparation du présent rapport, les occurrences connues du P. lurida ont été revisitées en 2014. Le P. lurida a été observé pour la première fois en Nouvelle-Écosse en 1981 par W. Maass (Maass et al., 1986). Les notes qui figurent sur les emballages des spécimens de Maass fournissent des indications concernant les sites, mais les coordonnées sont manquantes, de sorte qu'il y a une certaine incertitude quant à l'emplacement exact des occurrences; les recherches ont donc été menées dans toute la région entourant le site présumé. De nouvelles visites ont été réalisées dans 19 des 24 occurrences signalées avant 1986 ainsi que dans 5 des occurrences découvertes plus récemment (tableau 1A).
Dans le monde, le Pannaria lurida s.l. a été observé dans une vaste gamme de milieux et de climats, allant de forêts tempérées au nord, jusqu'à l'extrémité ouest de Terre-Neuve (Ahti, comm. pers., 2014), à des mangroves côtières tropicales, en Australie (Stephens, 1979).
En Nouvelle-Écosse,le P. lurida se rencontre le plus fréquemment à proximité de milieux humides mal drainés dominés par des feuillus. Ces sites se trouvent près de la lisière de marécages boisés ou de plaines inondables riveraines, ou encore au pied de pentes modérées à abruptes. Quelques occurrences ont été signalées dans des peuplements de feuillus situés en terrain élevé, au sommet de pentes de moins de 100 m d'altitude. Seulement deux occurrences se trouvent à quelques km de la côte; les autres sont à plus de cinq km à l'intérieur des terres. Le couvert forestier est modérément dégagé (la densité du couvert a été évaluée pour 13 des 15 occurrences). L'espèce pousse sur l'écorce rugueuse d'arbres matures, principalement sur les côtés les plus exposés au soleil. L'érable rouge (Acer rubrum) constitue la principale espèce hôte, suivie en fréquence par les peupliers (Populus tremuloides, Populus balsamifera). L'espèce a également été observée sur le frêne noir (Fraxinus nigra), le frêne blanc (Fraxinus americana), l'érable à sucre (Acer saccharum), l'érable rouge (Quercus rubra) et le hêtre à grandes feuilles (Fagus grandifolia) (Maass et al., 1986). Le P. lurida a été signalé une seule fois sur un conifère (épinette blanche), en Nouvelle-Écosse, dans le parc provincial Five Islands, où l'espèce a récemment été observée. L'habitat y était très semblable à celui de l'occurrence de la vallée de Codroy, à Terre-Neuve (voir la section Activités de recherche, ci-dessus). En outre, ce site est très singulier, puisque les épinettes (examinés pour la dernière fois par H. Harries dans les années 1990) y étaient également colonisées par d'autres cyanolichens, dont le Collema subflaccidum, le Lobaria pulmonaria, le Pseudocyphellaria perpetua et le Sticta fuliginosa.
Au Nouveau-Brunswick, une occurrence se trouve dans une forêt mixte humide dominée par le Thuja occidentalis, l'Acer rubrum et le Fraxinus nigra et comptant quelques Abies balsamea et Picea mariana épars. Toutefois, au Nouveau-Brunswick et en Nouvelle-Écosse le P. lurida n'a jamais été observé sur un arbre du genre Thuja, alors qu'il a été observé uniquement sur des arbres de ce genre à l'Île-du-Prince-Édouard. Environ quinze forêts marécageuses à thuya ont fait l'objet de relevé lichénologiques intensifs au Nouveau-Brunswick (S. R. Clayden, données inédites). Même si certains cyanolichens, dont le Pannaria conoplea et le P. rubiginosa, sont fréquemment présents sur le tronc et/ou les branches de thuyas dans ces peuplements, ce n'est pas le cas du P. lurida. L'espèce se rencontre généralement dans des peuplements matures au Nouveau-Brunswick et en Nouvelle-Écosse, mais quelques occurrences ont été observées dansdes peuplements allant du stade de perchis au stade mature, et même dans quelques cas ayant dépassé la maturité. Il arrive qu'un grand nombre de thalles soient présents sur un même arbre (figure 5). Le diamètre à hauteur de poitrine (dhp) du tronc des arbres hôtes va de 8 à 50 cm. Le tronc de la majorité des arbres colonisés présente un dhp de 12 à 23 cm dans les marécages boisés, mais un plus grand diamètre (dhp de 22 à 36 cm) dans les sites légèrement mieux drainés. Dans les marécages boisés, le sous-étage comprend des eaux stagnantes, des sphaignes et des coussins de mousses ainsi que des arbustes, notamment l'Ilex verticillata et l'Alnus incana. Le P. lurida peut être le seul cyanolichen présent sur le tronc d'un arbre hôte ou pousser aux côtés d'une diversité d'autres espèces, le plus souvent le Lobaria pulmonaria, le Lobaria quercizans, le Collema subflaccidum/furfuraceum, le Leptogium cyanescens, le Pannaria conoplea, le Pannaria rubiginosa et le Pseudocyphellaria perpetua.
Le seul spécimen connu provenant de l'Île-du-Prince-Édouard a été récolté dans un peuplement de thuyas (Thuja occidentalis) matures (McMullin et al., 2012). On trouve de l'eau stagnante dans tout le peuplement, et le site est abrité par un dense couvert forestier et la forêt environnante, de sorte que le taux d'humidité y est élevé. Le peuplement est entouré d'une forêt mixte (conifère et feuillus) mature plus sèche. Le peuplement de thuyas se trouve à environ 500 m de la côte. Il héberge de nombreuses espèces de lichens qui sont rares et vulnérables à l'Île-du-Prince-Édouard, y compris les suivantes : Heterodermia speciosa, Leptogium saturninum, Menegazzia terebrata, Nephroma helveticum ssp. helveticum, N. laevigatum, Pannaria rubiginosa, Parmeliella triptophylla, Physconia grumosa, Protopannaria pezizoides et Scytinium subtile. Le Pannaria lurida a en outre été observé à proximité du Lobaria pulmonaria, du L. quercizans, du Collema subflaccidum et du Leptogium cyanescens.
À Terre-Neuve, le P. lurida se rencontre sur des promontoires côtiers situés à moins de 10 m du bord de la falaise, sur le tronc ou les branches d'épinettes blanches (Picea glauca), dans des boisés de conifères (figure 6). On présume que l'exposition au sel neutralise le pH de l'écorce des conifères. L'espèce pousse principalement sur le côté nord des troncs, et certains thalles sont parfois orientés vers l'océan. Les arbres colonisés par le P. lurida présentaient un dhp de 4 à 21 cm.Le P. lurida a été observé sur les branches et le tronc de 14 arbres. Le peuplement se trouvait à moins de 10 m d'une falaise de grès, à une altitude de 39 m. Le peuplement principal présentait des zones de suintement, et le parterre forestier était couvert de mousses hypnacées, particulièrement le Rhytidiadelphus triquetrus, ainsi que de graminées, mais hébergeait peu de fougères ou d'autres plantes (Kissler, 2010; Ahti in Jørgensen, 2000; Hanel, comm. pers., 2014).
Peu de renseignements sur l'habitat accompagnaient le spécimen provenant de Saint-Pierre-et-Miquelon, excepté qu'il avait été récolté dans l'île Miquelon, à Langlade, sur l'écorce d'un arbre, près de la rivière Belle, où il poussait avec l'Ulota drummondii et le Lobaria scrobiculata (Delamare, 1888).

Description longue de la figure 5
Photo de la pannaire jaune pâle poussant de manière luxuriante sur le tronc d'un érable rouge en Nouvelle-Écosse. La surface du tronc est presque entièrement recouverte de thalles de l'espèce.

Description longue de la figure 6
Photo de la pannaire jaune pâle et de la lobaire pulmonaire (Lobaria pulmonaria) poussant sur le tronc et les branches d'épinettes blanches tordues par le vent dans l'ouest de l'île de Terre-Neuve.
En Nouvelle-Écosse, au cours des 10 dernières années, un déclin des milieux propices au P. lurida a été causé par l'exploitation des forêts de feuillus. En 1996, le bois de feuillus représentait seulement environ 6 % du bois total récolté en Nouvelle-Écosse, mais en 2010, il représentait 15 %. En volume, la récolte annuelle de bois de feuillus en Nouvelle-Écosse est passée de 400 000 mètres cubes à plus de 800 000 mètres cubes de 1990 à 2000 (voir la section Menaces). Cette exploitation a entraîné une diminution considérable de la superficie de forêts peuplées d'arbres âgés de 60 ans et plus (NSDNR, 2008), catégorie d'arbres privilégiés par le P. lurida parce que leur écorce est rugueuse. Selon les projections de l'exploitation future, le bois de feuillus représentera 30 % des récoltes, pour un total annuel d'environ 1 900 000 m3 (Woodbridge Associates, 2011).
Il est important de déterminer si la superficie de milieux propices limite la présence du P. lurida au Canada. Une analyse de régression logistique a été élaborée en vue de fournir un modèle prédictif d'habitat pour le P. lurida. Les variables explicatives comprenaient les normales climatiques sur trente ans des précipitations annuelles et des températures en juillet (données d'Environnement Canada), les données SIG relatives à l'altitude et à la distance par rapport à la côte (tirées d'Atlas), la hauteur moyenne des peuplements forestiers, le pourcentage de feuillus, la catégorie de maturité des arbres, le type de couverture (feuillus, conifères, mixte) et la densité du couvert forestier (d'après les données SIG provinciales sur le couvert forestier des ministères des Ressources naturelles de la Nouvelle-Écosse et du Nouveau-Brunswick) ainsi que la distance par rapport au milieu humide le plus près (d'après la base de données SIG sur les milieux humides du ministère des Richesses naturelles). Les variables expliquées étaient les localités connues de P. lurda comme localités positives (n = 26) et les sites où des relevés visant les cyanolichens ont été réalisés, mais où le P. lurida n'a pas été trouvé, comme localités négatives (n = 77).
Le modèle prédictif d'habitat a été mis au point au moyen d'une procédure de régression descendante, puis l'indice de Moran a été utilisé pour évaluer l'autocorrélation spatiale des résidus. Il n'y avait aucune autocorrélation spatiale entre les résidus, et aucun ajustement n'a été fait. L'aire sous la courbe ROC était de 0,848 (la courbe ROC est une mesure courante de la performance d'un modèle; un résultat de 0,5 ne vaut pas mieux que le hasard et un résultat de 1,0 indique une concordance parfaite). Le modèle donnait un R2 de Nagelkerke de 0,426 et un R2 de McFadden de 0,301 (P = 0,001). L'altitude, les précipitations annuelles, la hauteur du peuplement forestier et la distance par rapport à un milieu humide constituaient des facteurs importants pour la prédiction de la présence du P. lurida. Les facteurs qui n'étaient pas significatifs dans le modèle étaient les températures en juillet, la densité du couvert forestier, le pourcentage de feuillus dans le peuplement, la maturité du peuplement et le type de couvert (feuillus, conifères, mixte). Ces facteurs ont été codés comme des données binaires dans le modèle, tandis que le pourcentage de feuillus était une variable continue. Le fait que le pourcentage de feuillus n'était pas significatif pourrait refléter des limitations quant à l'interprétation photographique. Par exemple, si le P. lurida est présent dans des sites où le pourcentage de feuillus est faible, le photointerprète n'inclura peut-être pas les feuillus comme composante du peuplement, et l'espèce d'arbre peut parfois être sujette à des confusions ou à des erreurs d'interprétation, par exemple, la pruche peut être confondue avec des feuillus. Il ne fait aucun doute que les feuillus sont importants pour la description de l'habitat du P. lurida, car l'espèce vit sur l'écorce de ce type d'arbres. Toutefois, il semble que le pourcentage de feuillus, selon l'évaluation réalisée au moyen des données actuellement disponibles, ne soit pas un bon prédicteur de la présence du P. lurida. Selon une matrice de confusion, le modèle prédisait la présence du P. lurida aux endroits où il est réellement présent dans une proportion de 58 % et prédisait correctement l'absence de l'espèce dans une proportion de 91 %. Le modèle a montré que l'habitat idéal (probabilité de présence > 90 %) se trouve à des altitudes de moins de 100 m, reçoit des précipitations annuelles de 1 400 à 1 600 mm, est occupé par des peuplements dont les arbres mesurent plus de 14 m de hauteur et se situe à moins de 1 200 m d'un milieu humide. Il y a environ 4 589 ha d'habitat idéal prédit (probabilité de présence > 90 %) en Nouvelle-Écosse, ce qui représente environ 0,0834 % du territoire de la province (figure 7). Il y a environ 391 ha d'habitat idéal prédit au Nouveau-Brunswick, mais aucun à l'Île-du-Prince-Édouard.
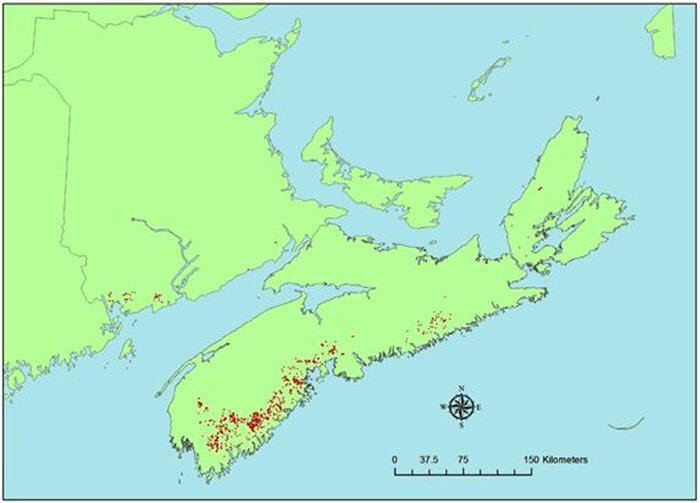
Description longue de la figure 7
Carte illustrant l'habitat prédit (probabilité de présence > 90 %) de la pannaire jaune pâle dans les Maritimes, au Canada.
Le Pannaria lurida se reproduit au moyen d'ascospores (figure 8) éjectées des apothécies, structures communes sur le thalle de l'espèce (voir la section Dispersion, ci-dessous). Une fois parvenues à maturité, les apothécies éjectent les spores, et celles-ci sont transportées par le vent et la pluie jusqu'à de nouveaux substrats, où elles produisent un nouveau thalle uniquement si elles rencontrent une cyanobactérie compatible du genre Nostoc.
La durée d'une génération varie de 10 ans chez les lichens à colonisation rapide, comme le Xanthoria parietina, à plus de 17 ans chez les espèces de lichens des forêts anciennes, comme le Lobaria pulmonaria (Scheidegger et Goward, 2002; Larsson et Gauslaa, 2010). Puisque le Pannaria lurida pousse principalement dans les forêts anciennes, une durée de 17 ans constitue une bonne estimation pour l'espèce.
Description longue de la figure 8
Diagramme de la structure et du cycle vital de la pannaire jaune pâle, montrant les ascospores simples ainsi qu'une coupe du thalle, à l'intérieur duquel on peut voir les cyanobactéries qui forment des chaînes de plus en plus comprimées vers la base de la surface photosynthétique du thalle.
Chez le P. lurida, les chaînes de cyanobactéries du genre Nostoc se gonflent considérablement lorsque mouillées, ce qui fait en sorte que le thalle devient assez gélatineux dans ces conditions (Jørgensen, 2000). L'eau supplémentaire ainsi stockée permet de prolonger la période de photosynthèse. La face inférieure du P. lurida est fixée au substrat par de denses rhizohyphes entrelacées. Selon certaines études, celles-ci améliorent la rapidité d'absorption de l'eau (Rundel, 1988).
La cyanobactérie du genre Nostoc qui compose le Pannaria lurida fournit à son symbionte des acides aminés, par fixation de l'azote atmosphérique, par l'entremise d'un processus nécessitant des phosphates. La diversité et l'abondance des cyanolichens peuvent augmenter avec l'augmentation des concentrations de phosphates, phénomène observé après la fertilisation d'une forêt montagnarde à Hawaï (Benner et Vitousek, 2007). Toutefois, la hausse des concentrations de phosphates n'a pas entraîné une augmentation de la fixation d'azote chez des espèces en particulier. Le Nephroma helveticum présentait le taux de fixation d'azote le plus faible, le Pseudocyphellaria crocata le taux le plus élevé, et le Pannaria lurida un taux intermédiaire (Benner et Vitousek, 2012).
Certains facteurs qui interfèrent avec le prélèvement de phosphates, comme les pluies acides et la pollution par le dioxyde de soufre, ont un effet inverse et inhibent la fixation de l'azote (Nash, 2008; voir la section Menaces, ci-dessous).
L'espèce n'est pas consommée de façon considérable par les limaces et autres gastropodes (de légers dommages ont été observés dans 4 des 15 occurrences), ce qui pourrait s'expliquer par sa production de pannarine. La pannarine est un des produits secondaires qui ont une variété de fonctions chez les lichens (Rankovic, 2015).
L'adaptabilité de chacune des sous-espèces du P. lurida n'a pas été étudiée.
Le Pannaria lurida ne possède pas de structures de reproduction asexuée spécialisées, comme des isidies ou des sorédies. La fragmentation du thalle peut permettre une dispersion à l'échelle locale sur un arbre, mais le thalle plutôt épais (généralement 106 à 200 μm d'épaisseur) ne se détache et ne se disperse probablement pas facilement, en raison de la masse de rhizohyphes. L'espèce se disperse donc principalement par ses ascospores (figure 8), qui sont éjectées à quelques centimètres dans les airs et peuvent être transportées par le vent sur des distances considérables. Dans les occurrences connues, la production d'apothécies semble commune. Dans les occurrences où des données ont été recueillies, un plus grand nombre de thalles possédant au moins six apothécies (22 arbres) a été observé que de thalles en possédant moins de six (8 arbres). La production de spores devrait donc être adéquate pour la dispersion. Cependant, pour qu'un nouveau thalle soit produit, les spores doivent rencontrer une cyanobactérie viable compatible lorsqu'elles atterrissent sur leur substrat, qui doit normalement être l'écorce d'arbres présentant une texture rugueuse et un pH adéquats.
Les lichens peuvent être colonisés par des champignons lichénicoles; plusieurs de ces champignons ont été observés chez le genre Pannaria, mais aucun n'a été découvert jusqu'à maintenant chez le P. lurida (Hawksworth, 1983; Ihlen et Wedin, 2008; Lawrey et Diederich, 2015).
Avant le début des travaux de terrain menés en 2014, les rédacteurs ont consulté les spécimens des herbiers du Musée de la Nouvelle-Écosse, du Musée du Nouveau-Brunswick et du Musée canadien de la nature. En outre, le Consortium of North American Lichen Herbaria (en ligne) ainsi que les participants du Tuckerman Workshop et les membres d'une liste de diffusion sur les lichens ont été consultés pour le recensement des mentions de l'espèce, particulièrement en Amérique du Nord.
Des travaux de terrain de grande envergure ont été réalisés par les rédacteurs du présent rapport durant l'été et l'automne 2014. Ils visaient principalement à visiter les sites où le P. lurida avait déjà été signalé et à chercher des milieux propices à l'espèce dans les zones où sa présence pouvait être prévue. Les rédacteurs ont en outre examiné l'habitat et en ont pris des photographies et ont consigné toute menace observée. Dans chaque site visité, ils rédacteurs ont évalué l'abondance en comptant les thalles et en consignant l'abondance des organes de reproduction, au moyen d'un formulaire normalisé. Ils ont également pris des photos des thalles de P. lurida et ont récolté des spécimens. Certains de ces spécimens ont été déposés dans l'herbier CANL, à Ottawa, et les autres dans l'herbier NBM, à Saint John.
Au Canada, 56 occurrences de pannaire jaune pâle sont connues : 49 en Nouvelle-Écosse, 4 au Nouveau-Brunswick, 2 à Terre-Neuve et 1 à l'Île-du-Prince-Édouard. Les relevés n'ont pas permis de confirmer la présence de l'espèce à l'Île-du-Prince-Édouard, dans une occurrence à Terre-Neuve et dans deux occurrences au Nouveau-Brunswick.
Des données sur le nombre de thalles ont été recueillies dans 11 occurrences (tableau 2). La population totale de P. lurida au Canada serait de 4 583 individus matures, d'après une estimation fondée sur une extrapolation des occurrences dénombrées (voir ci-dessous). L'extrapolation a été réalisée à partir du nombre médian de thalles par arbre. Sur le plan statistique, il est plus approprié d'utiliser le nombre médian que le nombre moyen dans les cas où les données sont fortement asymétriques (Baldi et Moore, 2014). Dans le cas présent, une occurrence, qui compte environ 2 446 thalles, est disproportionnée par rapport aux autres occurrences, qui comptent chacune 2, 7, 20, 40, 50, 73, 103, 191, et 248 thalles (voir le tableau 2 et les renseignements ci-dessous).
Une grande variation a été observée quant au nombre d'arbres colonisés et au nombre de thalles dans les diverses occurrences. Le P. lurida a été trouvé sur un seul arbre dans le cas de quatre occurrences, et sur deux à plus de 10 arbres dans les autres occurrences. Le nombre de thalles par arbre va d'un arbre colonisé par un seul thalle à deux arbres colonisés par un très grand nombre de thalles. En général, les thalles pouvaient être dénombrés sur chaque arbre, mais dans des conditions favorables, les thalles occupent de grandes superficies de l'écorce. L'abondance de l'espèce sur ces arbres donne à penser que des fragments de thalles se sont détachés puis rattachés ailleurs sur l'écorce ou que la surface du tronc de ces arbres hébergeait des souches compatibles du genre Nostoc, qui ont permis la colonisation.
| Province | Occurrence | Comté | Espèce hôte | Nbre de thalles | Occurrence revisitée | Espèce présente |
|---|---|---|---|---|---|---|
| N.-B. | Pointe Clark | Charlotte | Frêne, peuplier | 50 Note du tableaua | N | O |
| N.-B. | Pomeroy Ridge | Charlotte | Frêne | 20 | O | O |
| N.-É. | Hardwoodlands | Lunenburg | Peuplier | 40 | N | O |
| N.-É. | Chelsea | Lunenburg | Érable rouge | 3 arbres recouverts de grandes colonies ( Note du tableaub1 122 + 1 290 + 34) | N | O |
| N.-É. | Chemin Grimm | Lunenburg | Érable rouge | 7 | O | O |
| N.-É. | Indian Falls | Lunenburg | Peuplier | 145 + trois arbres recouverts de grandes colonies (34+15+54) | O | O |
| N.-É. | West Clifford | Lunenburg | Érable rouge | 11 + 1 arbre recouvert d'une grande colonie ( Note du tableaub180) | N | O |
| N.-É. | Medway | Queens | - | 2 | N | O |
| T.-N.-L. | Vallée de Codroy, Woodville | - | Épinette blanche | 103 | O | O |
| N.-É. | Parc provincial Five Islands | Colchester | Épinette blanche | 2 | O | O |
| N.-É. | Parc national Kejimkujik | Queens | Frêne, chêne, peuplier | 73 | O | O |
Dans trois occurrences (sur sept arbres) en Nouvelle-Écosse, les thalles étaient abondants au point qu'il était difficile de distinguer et de dénombrer les colonies (tableau 2, figure 5). Au chemin Chelsea, il y avait trois arbres recouverts de thalles. Deux de ces arbres présentaient un dhp respectif de 20 et 23 cm, et la couverture de thalles s'étendait du sol jusqu'à une hauteur de 5 m sur les arbres; le troisième arbre présentait un dhp de 12 cm, et son tronc était ceinturé de thalles sur une hauteur de 25 cm. À Indian Falls, il y avait trois arbres présentant de grandes colonies de lichens. À West Clifford, l'espèce a été observée sur un arbre ayant un dhp de 16 cm et recouvrait son tronc à partir du sol jusqu'à une hauteur de plus d'un mètre.
En se fondant sur un diamètre moyen des thalles de 5 cm et une distance entre les thalles de 1 cm, on a évalué le nombre de thalles sur ces troncs en divisant la superficie d'écorce colonisée (respectivement 5 027, 31 420, 36 133, 94 et 2 894 cm2) par 28 cm2. Ce calcul a donné l'effectif estimatif du P. lurida dans les trois occurrences, soit de 2 446 dans celle de Chelsea, 191 dans celle d'Indian Falls et 248 dans celle de West Clifford (tableau 2). En additionnant ces résultats aux données recueillies dans les autres occurrences dans lesquelles un dénombrement a été effectué, la population totale de P. lurida est de 3 180 thalles, et le nombre médian de thalles par occurrence est de 61. Si on utilise ce chiffre pour estimer l'effectif des 23 occurrences récemment découvertes qui n'ont pas été revisitées, la population totale de ces occurrences serait de 1 403 thalles de P. lurida, pour une population canadienne totale de 4 583 individus matures. Cette population se compose d'environ 100 thalles au Nouveau-Brunswick, 103 thalles à Terre-Neuve et le reste des thalles en Nouvelle-Écosse. L'espèce est très abondante dans certaines occurrences, et des dénombrements ont été effectués uniquement dans un petit nombre des occurrences récemment découvertes, de sorte que la population totale de P. lurida pourrait être supérieure à 10 000 individus matures. Pour cette raison, la variation de l'abondance a été évaluée en fonction du déclin du nombre d'occurrences.L'espèce n'était plus présente dans 18 des 56 occurrences découvertes avant 1986 ou plus récemment, ce qui représente une diminution de 32 %. Parmi les occurrences disparues, on compte une des deux occurrences de Terre-Neuve, la seule occurrence de l'Île-du-Prince-Édouard, deux des quatre occurrences connues au Nouveau-Brunswick, et le reste des occurrences en Nouvelle-Écosse. On suppose que cette diminution du nombre d'occurrences correspond à un déclin équivalent du nombre d'individus matures.
Alors que 32 des 56 occurrences connues au Canada ont été découvertes au cours des dix dernières années en raison d'une augmentation des relevés lichénologiques (tableau 2), la disparition d'au moins 18 occurrences a été constatée depuis la découverte de l'espèce au Canada. Des 24 occurrences découvertes avant 1986 en Nouvelle-Écosse, 15 ont été revisitées, mais l'espèce n'a pas été retrouvée dans 12 de celles-ci. En outre, l'espèce était absente de certaines des occurrences récemment découvertes qui ont été revisitées : deux en Nouvelle-Écosse, deux au Nouveau-Brunswick et une à l'Île-du-Prince-Édouard.
Le Pannaria lurida n'a pas été retrouvé dans deux occurrences du Maine situées à proximité de la frontière du Nouveau-Brunswick, ce qui donne à penser que la disparition des deux occurrences au Nouveau-Brunswick pourrait s'inscrire dans une tendance (figure 4). Le P. lurida a déjà été signalé au Vermont, au Massachusetts, au Rhode Island et au Connecticut, mais il n'y a pas été revu depuis 1950 et y est considéré comme menacé de disparition (Hinds et Hinds, 2007).
Les populations de P. lurida ne subissent pas de fluctuations extrêmes.
Une telle immigration est peu probable, puisque la localité existante la plus proche se trouve à l'île Mount Desert, dans le Maine, à 200 km au sud-ouest du milieu propice le plus proche, situé à Pomeroy Ridge, au Nouveau-Brunswick, où il y a actuellement une occurrence de P. lurida.
D'après l'analyse du calculateur des menaces, l'impact global des menaces qui pèsent sur le P. lurida est « élevé à très élevé » (annexe 1). La principale menace actuelle est l'exploitation forestière et la récolte du bois, qui entraîne la destruction des arbres hôtes et la disparition des conditions humides dans les milieux mal drainés dont l'espèce a besoin. Les autres menaces, d'une gravité moindre, sont les changements climatiques, le développement résidentiel, la construction routière et la pollution atmosphérique. De plus amples renseignements sur les menaces qui pèsent sur le P. lurida sont présentés ci-dessous et dans le tableau 3.
| Occurrence, nom de l'auteur de la découverte et date de découverte | Habitat | Présence du Pannaria. lurida | Menaces observées ou inférées | Régime foncier | Autres cyanolichens trouvés | Hôte du P. lurida | Nom du dernier observateur et date |
|---|---|---|---|---|---|---|---|
| Région de Pleasant View Cedars, Troy McMullin, 2009 |
Boisé dominé par le thuya, espèce trouvée sur un seul thuya. | Non retrouvé | Aucune menace évidente | - | Diverses autres espèces de lichens rares ont été observées. | Sans objet | Troy McMullin |
| Occurrence, nom de l'auteur de la découverte et date de découverte | Habitat | Présence du Pannaria. lurida | Menaces observées ou inférées | Régime foncier | Autres cyanolichens trouvés | Hôte du P. lurida | Nom du dernier observateur et date |
|---|---|---|---|---|---|---|---|
| Vallée de Codroy, Teuvo Ahti, 1956 |
Sur un Picea glauca rabougri, mais mature, en bordure d'une falaise maritime de grès, à l'ouest de Millville, dans la vallée de Codroy, à l'ouest de Terre-Neuve. | Thalles sains | - | Terres privées | L. scrobiculata, L. pulmonaria, Leptogium cyanescens, Collema subflaccidum et Parmeliella triptophylla | Épinette blanche | Emilie Kissler, 2010 |
| Ruisseau Eels, près du parc national du Gros-Morne Teuvo Ahti, 1978 |
Boisé côtier à Abies balsamea exposé aux vents, derrière des dunes de sable, à une altitude de 6 m. | Non retrouvé | - | Parc national | - | - | Teuvo Ahti et Michele Piercey-Normore, 2014 |
| Occurrence, nom de l'auteur de la découverte et date de découverte | Habitat | Présence du Pannaria. lurida | Menaces observées ou inférées | Régime foncier | Autres cyanolichens trouvés | Hôte du P. lurida | Nom du dernier observateur et date |
|---|---|---|---|---|---|---|---|
| Pointe Clark, rivière St. Croix, comté de Charlotte, Stephen Clayden, 2006 et 2007 |
Forêt mixte humide dominée par le Thuja occidentalis, l'Acer rubrum et le Fraxinus nigra, avec Abies balsamea et Picea mariana épars. | Occurrence non revisitée | - | À l'intérieur de l'aire naturelle protégée Clark Point, Nouveau-Brunswick | - | Frêne noir | Stephen Clayden, 2007 |
| Pomeroy Ridge Stephen Clayden, 2008 |
Forêt bosselée humide clairsemée à Thuja occidentalis, à Abies balsamea, à Acer rubrum, à Fraxinus nigra, à Alnus incana et à Ilex verticillata, et zone humide située à proximité comprenant ces espèces ainsi que le Populus balsamifera. | Thalles sains | - | Terres privées | - | Frêne noir, érable rouge, peuplier baumier | Stephen Clayden et Kendra Driscoll, 2014 |
| Rivière Chockpish nord, au sud de Rexton, Wolfgang Maass, 1983 |
Forêt mixte à Thuja très humide, avec Abies, Acer et Fraxinus (d'après l'étiquette du spécimen) | Occurrence non retrouvée | La région a fait l'objet d'une exploitation intensive. Plusieurs zones ont été examinées | Terres privées | Collema leptaleum, C. furfuraceum, L. cyanescens, L. pulmonaria et L. quercizans | Sans objet | Frances Anderson, 2014 |
| Chemin Aboujagane, au nord de Sackville, Hinrich Harries et Wolfgang Maass, 1979 |
Forêt moussue et herbeuse humide à Abies, Picea et Acer rubrum, près de la bordure d'un marécage à Alnus incana (d'après l'étiquette du spécimen) | Occurrence non retrouvée | En 2014, de longues bandes occupées par des boisés matures humides bordaient les deux côtés du chemin. La forêt avait été exploitée au-delà de ces bandes. Exploitation forestière en cours dans la région. | Terres privées | Sans objet | Sans objet | Frances Anderson, 2014 |
| Occurrence, nom de l'auteur de la découverte et date de découverte | Habitat | Présence du Pannaria. lurida | Menaces observées ou inférées | Régime foncier | Autres cyanolichens trouvés | Hôte du P. lurida | Nom du dernier observateur et date |
|---|---|---|---|---|---|---|---|
| Upper New Cornwall, comté de Lunenburg, Wolfgang Maass, 1981 |
Un site se trouvait en bordure d'une plaine inondable, face au sud, le long d'un ruisseau effluent du lac Caribou. La flore de cyanolichens était minime. Un deuxième site plus au sud présentait des arbres plus grands et une plus grande variété de cyanolichens, |
Occurrence non retrouvée | Malgré la présence d'érables et de bouleaux matures parmi les conifères, il n'y avait aucun signe du P. lurida. | Terres privées | L. cyanescens, L. pulmonaria, L. scrobiculata. Deuxième site comprenant P. rubiginosa, Pannaria conoplea, Fuscopannaria ahlneri |
Sans objet | Frances Anderson, 2014 |
| Summerville, comté de Queens, Wolfgang Maass, 1981 |
Étroite bande d'arbres anciens bordant la route, vestige du type de forêt qui se trouvait à cet endroit auparavant. | Occurrence non retrouvée | Occurrence exploitée il y a environ 10 ans et maintenant occupée par de jeunes arbres. | Terres privées | Cyanolichens présents : P. conoplea et Lobaria sp. | Sans objet | Frances Anderson, 2014 |
| Parc national Kejimkujik, comté de Queens (2 occurrences), Wolfgang Maass, 1981, 1982 |
Boisé à chêne, à frêne et à peuplier dans des zones plates humides dominées par les feuillus, le long du chemin de terre menant au lac Georges. | Thalles sains | - | Parc national | - | Chêne, frêne, peuplier | Troy McMullin et Frances Anderson, 2014 |
| Greenfield, comté de Queens, Wolfgang Maass, 1982 |
La forêt a été exploitée d'un côté de la route, et l'autre côté est occupé par un milieu humide principalement peuplé de conifères. Des lichens du genre Lobaria étaient présents sur les quelques érables dans le milieu humide. | Occurrence non retrouvée | Un côté de la route menant à Greenfield a été exploité il y a environ 5 ans. | Terres privées | Lobaria sp. | Sans objet | Frances Anderson, 2014 |
| Kempt, comté de Queens, Wolfgang Maass, 1982 |
Les boisés ne semblaient pas prometteurs. | Occurrence non retrouvée | Les boisés ne semblaient pas prometteurs. | Terres privées | - | Sans objet | Frances Anderson, 2014 |
| Midway, comté de Digby, Wolfgang Maass, 1982 |
Le lac Midway est long de 1,9 km. Arbres matures, y compris des érables et des frênes. Quelques cyanolichens étaient présents dans le peuplement bordant le chemin, à l'extrémité marécageuse du lac. | Occurrence non retrouvée | Les boisés dans l'est ont été partiellement exploités, mais il y subsistait un peuplement d'Arbres important. | Terres privées | - | Sans objet | Frances Anderson, 2014 |
| Lac Rossignol, comté de Queens, Nouvelle-Écosse, Wolfgang Maass, 1982 |
Il y avait quelques érables rouges matures dans la zone exploitée, dans un petit ravin, et le reste des arbres avaient été coupés. | Non retrouvé | Les arbres ont été coupés d'un côté de la route vers 2005. L'autre côté est plus élevé et plus sec, et il est dominé par le pin blanc. | Anciens terrains de la Bowater Mersey | - | Sans objet | Frances Anderson, 2014 |
| Rivière Medway, comté de Queens, Wolfgang Maass, 1982 |
Quantités variables de boisés. Une partie est mature et l'autre en régénération. | Non retrouvé | Peu de signes d'exploitation forestière, mais on trouve des chalets et des habitations sur plus de 10 km le long de la route. | Terres privées | - | Sans objet | Frances Anderson, 2014 |
| Gaetz Brook, près du lac Scotts, Wolfgang Maass, 1982 |
Région fortement perturbée. Quelques arbres anciens sont encore présents, mais un lotissement a été construit il y a quelques années. | Occurrence non retrouvée | Une grande portion des arbres des anciens boisés a été complètement coupée en vue de ce qui ressemble à un lotissement. | Terres privées | - | Sans objet | Christopher Pepper, 2014 |
| Lac Porters, chemin Grande Lake par la route 107, à l'est de Halifax, Wolfgang Maass, 1982 |
Quelques boisés anciens intacts, mais principalement composés de conifères. Plusieurs des feuillus anciens étaient tombés, probablement durant l'ouragan Juan. | Occurrence non retrouvée | La région environnante a été en grande partie brûlée durant un grand feu de forêt en 2008, et une importante zone de coupe à blanc a été observée. | Terres privées | Quelques cyanolichens communs | - | Christopher Pepper, 2014 |
| Murchyville, au nord-est de Halifax, Wolfgang Maass, 1982 |
Un petit groupe de feuillus a été examiné attentivement. Le site était probablement favorable à une certaine époque, et il y a un petit ruisseau et une source. Pulmonaria et L. quercizans. | Occurrence non retrouvée | La zone a en majeure partie subi d'une coupe à blanc, et les thalles semblaient souffrir de dessèchement. | Terres privées | Un certain nombre de cyanolichens étaient présents, y compris le Collema subflaccidum, le Leptogium cyanescens et des Lobaria | - | Robert Cameron, 2014 |
| West Northfield, comté de Lunenburg, Irwin Brodo, 19?? |
Les boisés humides étaient principalement composés de conifères. | Occurrence non retrouvée | Des coupes importantes ont été réalisées au cours des dix dernières années. | Terres privées | - | Sans objet | Frances Anderson, 2014 |
| Indian Falls, comté de Lunenburg, Frances Anderson, 2008 |
Zone humide dégagée principalement plate traversée par un chemin de gravier menant à un parc municipal nommé « Indian Falls on the LaHave River ». | Thalles sains, sur neuf arbres | - | - | - | Érable rouge | Frances Anderson, 2014 |
| Au sud du chemin Grimm, Middle LaHave, comté de Lunenburg, 2008 |
Des érables étaient nichés contre une pente orientée vers le sud, à la limite d'une tourbière à épinette noire. | Thalles sains présents | Aucun signe de perturbation dans le site ou à proximité. | - | ?? | Érable rouge | Frances Anderson, 2014 |
| Lac McGowan, comté de Queens, Tom Neily, 2007 |
- | Occurrence non retrouvée | Reconfiguration des routes et bâtiments dans la région. | Écloserie du lac McGowan | - | Sans objet | Frances Anderson et Troy McMullin, 2014 |
L'exploitation forestière réalisée dans le passé a entraîné la disparition de certaines occurrences connues de Pannaria lurida en Nouvelle-Écosse et au Nouveau-Brunswick. L'exploitation forestière s'est élargie et comprend maintenant la récolte pour la production de biomasse et de copeaux de bois, ce qui a causé une augmentation de la récolte de feuillus. En outre, des baux de coupe forestière ont récemment été approuvés pour de vastes zones dans le sud et l'est de la Nouvelle-Écosse.
En Nouvelle-Écosse, le P. lurida se rencontre fréquemment à proximité de sites mal drainés, mais ne pousse pas nécessairement à l'intérieur de ces sites. Ce type de sites se trouve près de la limite de marécages boisés et de plaines inondables riveraines, ou au pied de pentes modérées à abruptes. Ainsi, le sol est généralement plus sec dans ces zones, et les arbres qui y poussent sont plus susceptibles d'être coupés que les érables qui poussent à l'intérieur des marécages (voir la section Habitat, ci-dessus). L'élimination des arbres sur lesquels pousse le P. lurida entraîne une diminution du nombre de thalles et a une incidence sur l'ensemble de la population. Cette menace est particulièrement préoccupante dans le cas du P. lurida, car seulement quelques arbres hébergent plus de 2 400 thalles, soit plus de 50 % du nombre de thalles connus au Canada. Selon Cameron et al. (2013), l'exploitation forestière réalisée à proximité ou à l'intérieur d'un paysage cause une diminution des taux d'humidité. Les coupes à blanc peuvent avoir un effet négatif sur les populations de P. lurida, mais si les activités d'exploitation sont réalisées à une échelle locale et selon une orientation convenable, elles peuvent être tolérées par l'espèce (voir la section Corridors de transport et de service).
Des activités d'exploitation forestière destinées à la production de pâtes et papier, de bois d'œuvre, de bois de chauffage et de biomasse sont actuellement réalisées en Nouvelle-Écosse. La récolte totale de bois a augmenté de manière constante entre le début des années 1980 et le milieu des années 2000 et a atteint son maximum en 2005, avec 6,9 millions de mètres cubes. Depuis, elle a commencé à diminuer et était de 4,1 millions de mètres cubes en 2009 (Woodbridge Associates, 2011) et de 3,6 millions de mètres cubes en 2014 (Province of NS, 2015). Cette tendance se reflète dans la récolte de bois de feuillus et celle de bois de conifères. La récolte de bois de feuillus a atteint un sommet en 2004, avec environ 890 000 m2, puis a atteint sa valeur la plus basse en dix ans en 2011, avec 474 000 m2 (NSDNR, 2012). Toutefois, on prévoyait une hausse de la récolte de bois de feuillus (Woodbridge Associates, 2011). Depuis 2011, une hausse constante de la récolte annuelle de bois de feuillus a en effet été constatée, et 705 000 m2 ont été exploités en 2014 (Province of Nova Scotia, 2015).
La possibilité annuelle de coupe (PAC) de bois de feuillus en Nouvelle-Écosse en 2005 a été évaluée à un peu moins de 2 000 000 m3 (NSDNR, 2008). Même si la récolte a été bien inférieure à la PAC, les récoltes passées et le taux actuel de récolte de bois de feuillus risquent d'avoir de graves répercussions sur l'habitat du P. lurida dans la province (voir aussi la section Tendances en matière d'habitat). Un déclin a été observé dans les forêts d'âge supérieur à 61 ans (figure 4; NSDNR, 2008), ce qui est préoccupant, puisqu'une période de 50 ans est probablement nécessaire avant que les nouveaux arbres présentent une écorce mature constituant un substrat convenant au P. lurida. En outre, Northern Pulp a récemment présenté une demande d'accès à 500 000 tonnes métriques vertes de fibres sur des terres de la Couronne dans le sud de la Nouvelle-Écosse, où le P. lurida est fréquent. Jusqu'à maintenant, le gouvernement a limité l'accès de l'entreprise aux 125 000 tonnes métriques vertes supplémentaires qui avaient déjà été convenues (Gorman et Zaccagna, 2014). Le reste du volume sera probablement prélevé sur des terres à bois privées. Plusieurs des sites dans lesquels le P. lurida est présent ou pourrait coloniser des arbres se trouvent sur des terres à bois privées dans cette région, de sorte que cette exploitation pourrait avoir de graves répercussions sur les populations de l'espèce.
Au Nouveau-Brunswick, il y a eu un déclin de la superficie de forêt de feuillus ancienne d'environ 20 % des années 1980 à aujourd'hui. Cette donnée est fondée sur une analyse des données d'inventaire forestier récoltées dans les terres de la Couronne et les petites forêts privées en tenure franche, qui représentent ensemble plus de 80 % des terres forestières de la province (NB DNR, données inédites). Des coupes en bandes ont été réalisées dans des forêts inéquiennes à un stade avancé de succession composées de feuillus tolérants à l'ombre, et des coupes sélectives secondaires commencent à être réalisées, de sorte que les effets négatifs de cette menace augmenteront probablement (Clayden, 2014). Au Nouveau-Brunswick, la récolte de forêts de feuillus anciennes est principalement réalisée dans des sites mésiques en terrain élevé, mais des occurrences connues et potentielles de P. lurida poussant dans des sites humides sont également touchées. Trois des quatre occurrences connues se trouvent (ou se trouvaient) sur des feuillus poussant dans des clairières ou en bordure de forêts marécageuses à thuya occidental (Thuja occidentalis). De vastes relevés lichénologiques ont été réalisés dans ce type de communauté dans les Maritimes et en Gaspésie (Clayden et al., 2011 et données inédites). L'autre occurrence de P. lurida au Nouveau-Brunswick se trouvait dans une forêt humide mixte où il n'y avait aucun thuya; cette localité et une des localités se trouvant dans un marécage à thuya, toutes deux situées dans l'est du Nouveau-Brunswick, ont subi une exploitation forestière ou ont été perturbées par ce type d'exploitation dans les années suivant leur découverte. En effet, le P. lurida n'a pas été retrouvé dans ces localités en 2014 (F. Anderson, comm. pers., 2014; voir l'annexe 2).
Les changements climatiques devraient entraîner une hausse des températures estivales et causer des périodes de pluies fortes entrecoupées de sécheresses, ce qui pourrait avoir des répercussions sur les populations de l'espèce en Nouvelle-Écosse. Une diminution de la fréquence du brouillard pourrait particulièrement nuire aux occurrences du comté de Shelburne. Ces changements touchant le régime d'humidité constituent probablement une menace pour le P. lurida. L'espèce semble limitée aux milieux généralement humides (voir la section Habitat, ci-dessus). Les occurrences se trouvent principalement au-dessus de la limite du brouillard, sauf plusieurs des occurrences du comté de Shelburne, sur la côte Atlantique de la Nouvelle-Écosse, au sud d'Halifax.
Les modèles bioclimatiques sont couramment utilisés pour évaluer l'effet des changements climatiques sur les espèces. Le modèle bioclimatique BIOCLIM est bien connu et a été utilisé pour caractériser le climat convenant actuellement au P. lurida et le climat futur potentiel. Le modèle BIOCLIM comprend jusqu'à 35 paramètres climatiques à l'intérieur de l'aire de répartition connue d'une espèce et évalue le caractère convenable du climat actuel et futur (Beaumonta et al., 2005). Les données climatiques mondiales tirées de Hijmans et al.. (2005) ont été intégrées au logiciel DIVA-GIS pour la modélisation. Selon le scénario climatique futur pour l'est de l'Amérique du Nord, il y aura une hausse de 2 °C des températures annuelles moyennes et une augmentation de 10 % des précipitations annuelles (Govindamsay et al., 2003).
Selon le modèle BIOCLIM, les plus grandes zones dont le climat convient à l'espèce se trouvent actuellement aux États-Unis. Dans l'est du Canada, la Nouvelle-Écosse renferme les plus vastes zones présentant un climat propice, et ces zones sont marginales à l'Île-du-Prince-Édouard et petites au Nouveau-Brunswick et à Terre-Neuve. Les scénarios climatiques (projections sur 100 ans) semblent indiquer que les zones présentant un climat propice aux États-Unis, en Nouvelle-Écosse et à l'Île-du-Prince-Édouard diminueront considérablement. Une certaine augmentation des zones à climat peu à moyennement propice est prévue pour le nord de la Nouvelle-Écosse et de Terre-Neuve (figure 9).
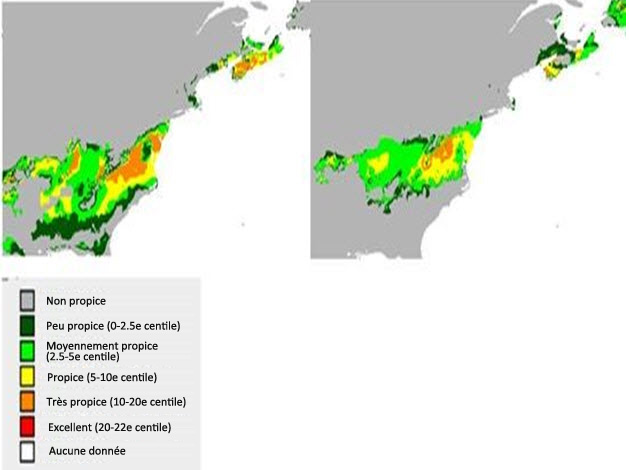
Description longue de la figure 9
Deux cartes illustrant le caractère convenable actuel et futur (dans 100 ans) du climat pour la pannaire jaune pâle dans l'est de l'Amérique du Nord. Le caractère convenable du climat est représenté par une échelle en six points allant de non propice à excellent.
Les cyanolichens sont extrêmement sensibles à la pollution atmosphérique par le dioxyde de soufre et aux pluies acides (Richardson et Cameron, 2004; Cameron et Richardson, 2006). La pollution atmosphérique transfrontalière touche encore le Canada atlantique. Selon l'Évaluation scientifique 2008 des dépôts acides au Canada (Environnement Canada, 2004), les dépôts acides totaux ont diminué dans l'est du Canada (CCME 2013), mais de grands secteurs du sud du Nouveau-Brunswick et du sud de la Nouvelle-Écosse continueront de recevoir des dépôts acides (CCME, 2008). C'est particulièrement le cas pour le sud de la Nouvelle-Écosse et les côtes est de la Nouvelle-Écosse et du Nouveau-Brunswick, où le P. lurida est présent. Les précipitations acides dans ces régions pourraient surpasser la capacité tampon de l'écorce des arbres, ce qui ferait en sorte que celle-ci ne conviendrait plus à l'espèce ou au symbionte cyanobactérien. L'écorce des arbres possède un pouvoir tampon intrinsèque, qui est plus élevé dans le cas des érables que dans celui des conifères. L'exposition continue aux pluies acides pourrait finir par faire en sorte que la capacité tampon des arbres ne suffise plus et que le substrat devienne rapidement trop acide pour les cyanolichens sensibles. Les thalles juvéniles sont particulièrement vulnérables à cette menace chimique (Nieboer et al., 1984)
Développement résidentiel et commercial
Un certain nombre d'occurrences du P. lurida, particulièrement celles du comté de Lunenburg, se trouvent dans des régions propices à la construction de chalets et de maisons, car elles sont attrayantes et se trouvent à proximité de routes. L'espèce serait disparue de Gaetz Brook, dans le comté d'Halifax, à cause de la construction d'un nouveau chemin et de maisons. Une portion encore plus grande du boisé a été coupée pour l'agrandissement d'un lotissement. Une autre occurrence dans la région de la rivière Medway, dans le comté de Queens, constitue une région active de construction de chalets.
Corridors de transport et de service
La construction de chemins associés aux parcs éoliens, à l'exploitation minière et au développement résidentiel menacent le P. lurida en éliminant des arbres et en modifiant les régimes d'humidité. Cette activité constitue particulièrement une menace lorsqu'elle cause un drainage et modifie le régime d'humidité dans les boisés environnants qui hébergent ou pourraient héberger le P. lurida. La construction de chemins associée à la fracturation et aux activités connexes constitue une autre menace, mais cette activité est actuellement en suspens en Nouvelle-Écosse et au Nouveau-Brunswick. La circulation routière ne semble pas constituer une menace pour les occurrences de P. lurida qui auraient subsisté à la modification du régime hydrologique causé par la construction du chemin. Parfois, les effets de bordure peuvent mener à la disparition de l'espèce. Les répercussions de la construction routière sur le régime d'humidité à proximité d'arbres dépendent de plusieurs facteurs, comme l'orientation du chemin par rapport au soleil et aux vents dominants. Par exemple, la disparition de l'occurrence de Gaetz Brook semble avoir été causée par d'importants travaux de construction routière et le dégagement d'une zone destinée à l'agrandissement d'un lotissement. Par ailleurs, l'occurrence de Chelsea, où les thalles de P. lurida sont abondants, se trouve à 20 m d'une route pavée à deux voies utilisée par des camions de transport du bois et pour la circulation régulière depuis au moins 50 ans. De même, l'occurrence de West Clifford, qui héberge également une abondance de thalles, se trouve à moins de 20 m de la route 325, couramment empruntée comme voie d'entrée et de sortie de Bridgewater.
Les principales menaces pouvant causer la disparition de l'espèce sont la disparition des forêts matures associée à l'exploitation forestière, les changements climatiques et la pollution atmosphérique. Il semble peu probable qu'un seul phénomène menaçant puisse toucher simultanément plus d'une occurrence. La meilleure estimation que l'on puisse faire consiste donc à considérer que le nombre de localités est égal au nombre total d'occurrences connues, soit 56. Même si certaines occurrences signalées dans le passé sont aujourd'hui disparues, il existe probablement des occurrences qui n'ont pas encore été découvertes, particulièrement en Nouvelle-Écosse. Toutefois, il est peu probable que le nombre d'occurrences encore inconnues soit supérieur au nombre d'occurrences actuellement connues; l'estimation la plus plausible serait donc que le nombre maximal de localités est inférieur à 100.
Cote mondiale : G3 (vulnérable) à G5 (non en péril).
Dernière évaluation de la cote mondiale : 11 décembre 2000.
Dernière modification de la cote mondiale : 21 janvier 2001.
Cote mondiale arrondie : G4 (apparemment non en péril; NatureServe, 2014).
Cote nationale : NNR (non classée).
Caroline du Nord : SNR. Pennsylvanie : SNR (non encore évaluée).
Cote nationale : NNR (non classée).
Québec : SNR (non encore évaluée).
En Nouvelle-Écosse, deux occurrences sont protégées dans le parc national Kejimkujik, une troisième dans le parc provincial Five Island, une quatrième dans des terres communes municipales, et une cinquième dans une réserve naturelle provinciale. Au Nouveau-Brunswick, à la pointe Clark, une occurrence se trouve dans une aire naturelle protégée. Toutes les autres occurrences au Canada se trouvent sur des terres de la Couronne ou des terres privées; hormis les occurrences au Nouveau-Brunswick, aucune de ces occurrences ne bénéficie d'une protection officielle. Au Nouveau-Brunswick, des zones tampons d'au moins 30 m doivent être laissées intactes autour de tous les milieux humides et cours d'eau (Loi sur l'assainissement de l'eau et Loi sur les terres et forêts de la Couronne du Nouveau-Brunswick). De même, la préservation de l'habitat des espèces sauvages est requise par la gestion des forêts de la Couronne (Loi sur les terres et forêts de la Couronne du Nouveau-Brunswick). La description de zones cibles et de peuplements/paysages a été établie pour les forêts anciennes de feuillus tolérants et les forêts anciennes mixtes. Depuis 1992, les plans de gestion des forêts comprennent l'identification spatiale des zones nécessaires à l'atteinte des cibles en matière d'habitat selon les configurations de peuplements et de paysages appropriées.
Un grand nombre de collègues ont aidé les rédacteurs du présent rapport à répertorier les occurrences et la répartition du P. lurida au Canada et aux États-Unis. Les rédacteurs du présent rapport remercient les personnes suivantes pour l'aide qu'elles leur ont fournie et le temps qu'elles leur ont consacré :
Les rédacteurs du rapport remercient Jean Gagnon et Claude Roy pour l'information qu'ils leur ont fournie concernant les mentions de l'espèce dans la province et à Saint-Pierre-et-Miquelon.
Les rédacteurs du rapport remercient Claudia Hanel, John McCarthy, Shelley Pardy et Mac Pitcher.
Les rédacteurs du rapport remercient particulièrement Tom Neily, qui a donné accès aux données qu'il a recueillies sur le P. lurida dans le cadre des vastes relevés lichénologiques qu'il a réalisés en Nouvelle-Écosse, ainsi que les membres de l'Équipe de rétablissement des cyanolichens de Nouvelle-Écosse.
Les membres du Tuckerman Lichen Workshop ainsi qu'un grand nombre de personnes, énumérées ci-dessous, ont aidé les rédacteurs du présent rapport à répertorier les occurrences et la répartition du P. lurida.
Canada : Irwin Brodo, Jennifer Doubt.
Finlande : Teuvo Ahti.
Allemagne : Andreas Beck.
Norvège : Yngvar Gauslaa, Einar Timdal, Per Magnus Jørgensen.
États-Unis : Alison Dibble, Walter Fertig, Scott LaGreca, Alan Friday, Dick Harris, Jim Hinds, Jason Hollinger, James Lawrey, James Lendemer, Michaela Schmull, Erin Tripp, Larry St. Clair.
Royaume-Uni : Mark Seaward, Rebecca Yahr, Heidi Doring, David Hawksworth.
ALA (Atlas of Living Australia) 2014. Pannaria lurida : Matted Lichen, disponible en anglais seulement
Baldi, B. et Moore, D.S. 2014. The practice of statistics in the life sciences. W.H. Freeman, New York, 3rd edition.
Beaumonta, L.J., L. Highesa et M. Poulsenb. 2005. Predicting species distributions: use of climatic parameters in BIOCLIM and its impact on predictions of species' current and future distributions. Ecological Modelling 186 (2): 251-270.
Benner, J.W. et Vitousek, P.M. 2007. Development of a diverse epiphyte community in response to phosphorus fertilization. Ecology Letters 10:628-636.
Benner, J.W. et Vitousek, P.M. 2012. Cyanolichens: a link between the phosphorus and nitrogen cycles in a Hawaiian montane forest. Journal of Tropical Ecology 28:73-81.
Clayden, S. R., Driscoll, K. E., Selva, S. B. et Sabine, D. L. 2011. Lichens and allied fungi of old wet cedar (Thuja occidentalis) forests in northeastern North America: indicators of environmental change. P. 116 in Proceedings, Plant Adaptation to Environmental Change, Plant Canada 2011 conference, July 17–21, 2011. Halifax, NS.
Czeczuga, B. 1988. Carotenoids. In Handbook of Lichenolgy (M. Galun Ed.) Vol III,
CRC Press, Boca Raton, pp 25 -34.
Blaser, D. et Stoeckli-Evans, H. 1991. Structure of Pannarin: a natural depsidone. Acta Crystallographica Section C 47:2624-2626.
Brodo, I.M., Sharnoff, S.D. et Sharnoff, S. 2001. Lichens of North America. Yale University Press, New Haven, 795 pp.
Cameron, R.P. et Richardson, D.H.S. 2006. Occurrence and abundance of epiphytic cyanolichens in Nova Scotia protected areas. Opuscula Philolichenum 3:5-14.
Cameron, R.P., T. Neily et H. Clapp. 2013. Forest harvesting impacts on mortality of an endangered lichen at the landscape and stand scales. Canadian Journal of Forest Research 43: 507-511.
Layden, S.R. 2010. Lichens and allied fungi of the Atlantic Maritime Ecozone. In Assessment of species diversity in the Atlantic Maritime Ecozone. (D.F. McAlpine et I.M. Smith eds.) NRC Press, Ottawa, pp. 153-178.
Clayden, S.R. 2014. Old tolerant hardwood forests in New Brunswick going down fast. New Brunswick Naturalist pp. 60-66.
CCME 2008. A national picture of acid deposition critical loads for forest soils in Canada. Canadian Council of Ministers of the Environment. (Également disponible en français : CCME. 2008. Charges critiques de dépôts acides dans les sols forestiers, profil de la situation au Canada, Conseil canadien des ministres de l'environnement.)
CCME 2013. Progress report on the Canada-wide acid rain strategy for post 2000. Canadian Council of Ministers of the Environment. (Également disponible en français : CCME. 2013. Rapport d'étape sur la Stratégie pancanadienne sur les émissions acidifiantes après l'an 2000, Conseil canadien des ministres de l'Environnement.)
Delamare, E., Renauld, F. et Cardot, J. 1888. Florule de l'île Miquelon: Amerique du Nord. W.G. Farlow, Lynon, p. 74.
Ekman, S., Wedin, M., Lindblom, L. et Jørgensen, P.M. 2014. Extended phylogeny and revised generic classification of the Pannariaceae (Peltigerales, Ascomycota). Lichenologist 46(5)627-656.
Elvebakk, A., Papaefthimiou, D., Robertson, E.H. et Liaimer, A. 2008. Phyologenetic patters among Nostoc cyanbionts with BI- and Tripartite lichens of the genus Pannaria. Journal of Phycology 44:1049-1059.
Flenniken, D.G. 1999. The macrolichens of West Virginia. Carlisle, Walnut Creek, p. 147.
Gorman, M. and Zaccagna, R. 2014. No Bail - out for Northern Pulp. Chronicle Herald Disponible en anglais seulement, January 30.
Galloway, D.J. 2008. Lichen Biogeography. In Lichen Biology (T.H. Nash ed). 2nd Edit. Cambridge University Press, Cambridge, pp 315-335.
GBIF (Global Biogeographic Information Facility) Disponible en anglais seulement, 2014. Pannaria lurida.
Govindasamy, B., Duffy, P.B. et Coquard, J. 2003. High-resolution simulations of global climate, part 2: effects of increased greenhouse cases. Climate Dynamics 21: 391– 404.
Goward, T., Brodo, I.M. et Clayden, S.R. 1998. Rare Lichens of Canada. Committee on the Status of Endangered Wildlife in Canada, Ottawa, 74pp.
Hawksworth, D.L. 1983. A key to the lichen-forming, parasitic, parasymbiotic and saprophytic fungi occurring on lichens in the British Isles. Lichenologist 15(1):1-44.
Hijmans, R.J., S.E. Cameron, J.L. Parra, P.G. Jones et A. Jarvis, 2005. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965-1978.
Hinds, J.W et Hinds, P.L. 2007. The Macrolichens of New England, New York Botanical Garden Press, New York.
Huneck, S. et Yoshimura, I. 1996. Identification of lichen Substances. SpringerVerlag, Heidelburg, p. 345.
Ihlen, P.G. et Wedin, M. 2008. An annotated key to the lichenicolous Ascomycota (including mitosporic morphs) of Sweden. Nova Hedwigia 86(3-4):275-365.
Jørgensen, P.M. 2000. Survey of the lichen family Pannariaceae on the American continent North of Mexico. Bryologist 103(4):670-704.
Jørgensen, P.M. 2003. Notes on African Pannariaceae (lichenized ascomycetes). Lichenologist 35(1):11-20.
Kissler, E. 2010. SSAC interim species status report on Pannaria lurida (Wrinkled Shield Lichen), Terre-Neuve.
Larsson, P. et Gauslaa, Y. 2010. Rapid juvenile development in old forest lichens. Botany 89:65-72.
Lawrey, J. D. et P. Diederich. 2015. Lichenicolous fungi - worldwide checklist, including isolated cultures and sequences available Disponible en anglais seulement, [3/21/2015].
Le Gallo, P.C. 1952. Lichens des îles St. Pierre et Miquelon. Revue bryologique & lichenologique, 21(1-2):144-172.
Lepage, E. 1947. Les lichens, les mousses et les hépatiques du Québec. Le Naturaliste Canadien 74:225-240.
Lepage, E. 1958. Premiere supplement au catalogue des lichens du Québec. Le Naturaliste Canadien 85:169-198.
Maass, W.S.G., Hoisington, B.L. et Harries, H. (1986) Pannaria lurida in Atlantic Canada. Proceedings of the Nova Scotian Institute of Science 36:131-135.
McMullin, R.T., Clayden, S.R. Selva, S.B., Curley, R. et Schram, L.J. 2012. A Lichen Checklist for Prince Edward Island, Canada. Evansia 29(1): 4-25.
Nash, T.H. 2008. Nitrogen, its metabolism and potential contribution. In Lichen Biology, 2nd edit. (T.H. Nash Ed.) Cambridge University Press, Cambridge, pp.216-233.
Nash, T.H., Ryan, B.D., Gries, C et Bungartz, F. 2002. Lichen flora of the greater Sonoran Desert Region. Arizona State University. Vol. 1, pp. 304-306.
NatureServe. 2014.
Nieboer, E., MacFarlane, J.D. et Richardson, D.H.S.1984. Modifications of plant cell buffering capacities by gaseous air pollutants. In: Gaseous air pollutants and plant metabolism (ed. M. Koziol & F.R. Whatley), Butterworths, London, 313-330.
NSDNR 2008. (Nova Scotia Department of Natural Resources). State of the Forest Report 19952005. Report FOR 2008-3. Nova Scotia Department of Natural Resources, Halifax, 40pp.
Province of Nova Scotia 2015. (Nova Scotia Department of Natural Resources) Registry of Buyers Annual Report (Disponible en anglais seulement, consulté le 17 février 2016).
Rankovic, B. 2015. Lichen secondary metabolites: Bioactive properties and pharmaceutical potential. Springer, Cham, 202pp.
Richardson, D.H.S. et Cameron, R.P. 2004. Cyanolichens: their response to pollution and possible management strategies for their conservation in Northeastern North America. Northeastern Naturalist 11:1-22.
Rundel, P.W. 1988. Water relations. In Handbook of Lichenolgy (M. Galun Ed.) Vol II, CRC Press, Boca Raton, pp17-36.
Scheidegger, C. et Goward, T. 2002. Monitoring lichens for conservation: red lists and conservation action plans. In monitoring with lichens – Monitoring lichens (P.L. Nimis, C. Scheidegger et P.A. Walseley, eds.) Kluwer Academic, Dordrecht pp. 163-181.
Schneider, A. 1897. A textbook of general lichenology with descriptions and figures of the general occurring in the northeastern United States. Willard Clute, Binghamton.
Stevens, G.N. 1979. Distribution and related ecology of macrolichens on mangroves on the east Australian coast. Lichenologis 11(3):295-305.
Woodbridge Associates. 2011. Economic impact analysis of timber management and supply changes on Nova Scotia's forest industry (PDF; 1.5 MB). (Disponible en anglais seulement), Prepared for Government of Nova Scotia Department of Natural Resources. 58 pp.
David Richardson est doyen émérite à l'Université Saint Mary's. Il étudie les lichens depuis 1963 et est l'unique auteur de deux livres sur le sujet : The Vanishing Lichens et Pollution Monitoring with Lichens. Il a également écrit plus de 20 chapitres de livres et 100 articles scientifiques sur divers aspects de la lichénologie. Il a étudié les lichens en Australie, au Canada, en Irlande et au Royaume-Uni.
Frances Anderson est agrégée de recherche au Nova Scotia Museum of Natural History, à Halifax. Elle étudie les lichens de Nouvelle-Écosse sur le terrain depuis plus de huit ans, et elle possède une vaste expérience des relevés lichénologiques. Elle a produit une liste des macrolichens de la province, disponible sur le site du Musée de la Nouvelle Écosse, dans la section « Curatorial Reports ».
Robert Cameron étudie les lichens depuis plus de dix ans. Ses premiers travaux ont porté sur les effets des pratiques forestières sur ces organismes, dans le cadre d'une maîtrise en biologie à l'Université Acadia. Depuis quelques années, Robert Cameron s'intéresse aux effets de la pollution atmosphérique sur les lichens, aux cyanolichens des forêts côtières et plus particulièrement à l'érioderme boréal. Il est actuellement l'écologiste de la Protected Areas Branch du Department of Environment and Labour de la Nouvelle-Écosse, où il est chargé du programme de recherches sur les zones protégées.
Stephen Clayden est conservateur de l'herbier de la Section de botanique et de mycologie du Musée du Nouveau-Brunswick. Il a commencé à s'intéresser aux lichens durant ses études à l'Université Mount Allison, et il a étudié la dynamique des communautés riches en lichens dans le nord-ouest du Québec dans le cadre de sa maîtrise à l'Université de Montréal. Sa recherche de doctorat, réalisée au Kings College, à Londres, au Royaume-Uni, portait sur le cycle vital des espèces du genre Rhizocarpon. Il a étudié les lichens dans le Canada atlantique et au Québec et a corédigé Rare Lichens of Canada: A Review and Provisional Listing avec Trevor Goward et Irwin Brodo
Christopher Pepper est naturaliste et collabore avec le Mersey Tobeatic Research Institute depuis plus de deux ans dans le cadre du projet sur l'érioderme boréal.
Troy McMullin est chercheur postdoctoral à l'Université de Guelph. Il a commencé à étudier les lichens en 2004 dans le cadre de sa maîtrise à l'Université Dalhousie, au cours de laquelle il s'est intéressé aux lichens présents dans les forêts acadiennes anciennes. Il a étudié la conservation et la gestion des lichens dans la forêt boréale en Ontarion durant son doctorat et étudie encore ce sujet actuellement. En tant qu'expert-conseil, Troy collabore régulièrement avec diverses organisations menant des inventaires lichénologiques et de la surveillance écologique visant les lichens. Il a étudié les lichens de la Californie à Haïda Gwaii et jusque dans les Maritimes et les Everglades. En outre, Troy est gestionnaire des collections de lichens de l'herbier de l'Institut de la biodiversité de l'Ontario.
Musée canadien de la nature, Ottawa
Consortium of North American Lichen Herbaria (CNALH) (Disponible en anglais seulement)
Herbier Louis-Marie, Université Laval, Québec
Musée de la Nouvelle-Écosse, Halifax
Musée du Nouveau-Brunswick, Saint John
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 1 | 0 |
| B | Élevé | 0 | 1 |
| C | Moyen | 3 | 0 |
| D | Faible | 2 | 5 |
| - | Impact global des menaces calculé : | Très élevé | Élevé |
| # | Menace | Impact (calculé) |
Impact (description) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | D | Faible | Petite (1-10 %) |
Modérée (11-30 %) |
Élevée (continue) |
- |
| 1.1 | Habitations et zones urbaines | D | Faible | Petite (1-10 %) |
Modérée (11-30 %) |
Élevée (continue) |
Un certain nombre d'occurrences se trouvent à proximité de chemins et dans des zones propices à l'aménagement de lots à bâtir et au développement résidentiel, particulièrement à Lunenburg. La disparition de l'occurrence de Gaetz Brook, près de Halifax, aurait été causée par la construction routière et domiciliaire. Dans cette occurrence, une proportion de terres boisées encore plus grande a été coupée, apparemment pour l'agrandissement d'un lotissement. Une autre occurrence dans la région de la rivière Medway, dans le comté de Queens, fait activement l'objet d'une construction de chalets. |
| 1.2 | Zones commerciales et industrielles | - | - | - | - | - | - |
| 1.3 | Tourisme et espaces récréatifs | - | - | - | - | - | - |
| 2 | Agriculture et aquaculture (en anglais seulement) | - | - | - | - | - | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | - | - | - | - | - | - |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | - |
| 2.3 | Élevage et élevage à grande échelle | - | - | - | - | - | - |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | - |
| 3 | Production d'énergie et exploitation minière (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) |
Inconnue | Modérée (possiblement à court terme, < 10 ans) |
- |
| 3.1 | Forage pétrolier et gazier | - | Négligeable | Négligeable (< 1 %) |
Inconnue | Modérée (possiblement à court terme, < 10 ans) |
La fracturation serait probablement réalisée principalement dans le nord de la Nouvelle-Écosse et serait probablement négligeable. Cette activité est également incluse dans la section « Routes et voies ferrées ». |
| 3.2 | Exploitation de mines et de carrières | - | - | - | - | - | - |
| 3.3 | Énergie renouvelable | - | - | - | - | - | - |
| 4 | Corridors de transport et de service (en anglais seulement) | D | Faible | Petite (1-10 %) |
Modérée (11-30 %) |
Élevée (continue) |
- |
| 4.1 | Routes et voies ferrées | D | Faible | Petite (1-10 %) |
Modérée (11-30 %) |
Élevée (continue) |
La construction de chemins associée aux parcs éoliens, à l'exploitation minière et au développement résidentiel et l'élimination d'arbres et les modifications du régime d'humidité qui en découlent auraient une incidence sur l'espèce. Les activités de construction routière liées à la fracturation pourraient également avoir des répercussions, mais font actuellement l'objet d'un gel. |
| 4.2 | Lignes de services publics | - | - | - | - | - | - |
| 4.3 | Transport par eau | - | - | - | - | - | - |
| 4.4 | Trajectoires de vol | - | - | - | - | - | - |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | AB | Très élevé-élevé | Généralisée-grande (31-100 %) |
Extrême-élevée (31-100 %) |
Élevée (continue) |
- |
| 5.1 | Chasse et prélèvement d'animaux terrestres | - | - | - | - | - | - |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | - |
| 5.3 | Exploitation forestière et récolte du bois | AB | Très élevé-élevé | Généralisée-grande (31-100 %) | Extrême-élevée (31-100 %) |
Élevée (continue) |
L'exploitation forestière réalisée dans le passé a entraîné la disparition d'occurrences connues de Pannaria lurida en Nouvelle-Écosse et au Nouveau-Brunswick. L'exploitation forestière s'est élargie et comprend maintenant la récolte pour la production de biomasse et de copeaux de bois, ce qui a causé une augmentation de la récolte de feuillus. En outre, des baux de coupe forestière ont récemment été approuvés pour de vastes zones dans le sud et l'est de la Nouvelle-Écosse. Portée : la fourchette de valeurs dénote une incertitude. * Les renseignements pour le Nouveau-Brunswick donnent à penser que la gravité se rapproche davantage de 30 % dans cette province et que de nouveaux sites pourraient être découverts. Toutefois, la majorité des occurrences de l'espèce se trouvent en Nouvelle-Écosse, ce qui explique la large plage de valeurs. L'impact en Nouvelle-Écosse augmentera considérablement, puisque le gouvernement vient d'approuver des baux de récolte supplémentaires. L'espèce est plus sensible aux changements du microclimat que l'anzie mousse-noire. |
| 5.4 | Pêche et récolte des ressources aquatiques | - | - | - | - | - | - |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | - | Négligeable | Négligeable (< 1 %) |
Légère (1-10 %) |
Élevée (continue) |
- |
| 6.1 | Activités récréatives | - | - | - | - | - | - |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | - |
| 6.3 | Travaux et autres activités | - | Négligeable | Négligeable (< 1 %) |
Légère (1-10 %) |
Élevée (continue) |
Récolte de lichens pour des recherches scientifiques. |
| 7 | Modification du système naturel (en anglais seulement) | - | - | - | - | - | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | - |
| 7.2 | Barrages, gestion et utilisation de l'eau | - | - | - | - | - | - |
| 7.3 | Autres modifications de l'écosystème | - | - | - | - | - | - |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | - | - | - | - | - | Très peu de dommages causés par des limaces, qu'elles soient exotiques ou indigènes, ont été signalés pour l'espèce. |
| 8.1 | Espèces exotiques/non indigènes envahissantes | - | - | - | - | - | - |
| 8.2 | Espèces indigènes problématiques | - | - | - | - | - | - |
| 8.3 | Introduction de matériel génétique | - | - | - | - | - | - |
| 9 | Pollution (en anglais seulement) | CD | Moyen-faible | Généralisée (71-100 %) |
Modérée-légère (1-30 %) |
Élevée (continue) |
- |
| 9.1 | Eaux usées domestiques et urbaines | - | - | - | - | - | - |
| 9.2 | Effluents industriels et militaires | - | - | - | - | - | - |
| 9.3 | Effluents agricoles et forestiers | - | - | - | - | - | - |
| 9.4 | Détritus et déchets solides | - | - | - | - | - | - |
| 9.5 | Polluants atmosphériques | CD | Moyen-faible | Généralisée (71-100 %) |
Modérée-légère (1-30 %) |
Élevée (continue) |
Les pluies acides pourraient surpasser la capacité tampon de l'écorce des feuillus hôtes, rendant ainsi le substrat non convenable à la croissance de cyanolichens ou à celle de la cyanobactérie à laquelle les spores du champignon doivent s'associer pour permettre la régénération de nouveaux thalles. Gravité : la fourchette de valeurs dénote une incertitude. * Plus près de 30 %. * Les pluies acides sont une menace importante pour l'espèce, puisque sa population se concentre dans le sud de la Nouvelle-Écosse, à proximité des sources de ce type de pollution. Il y a moins de précipitations à l'intérieur des terres, mais les pluies acides y constituent tout de même un facteur. |
| 9.6 | Énergie excessive | - | - | - | - | - | - |
| 10 | Phénomènes géologiques (en anglais seulement) | - | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | - | - |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | CD | Moyen-faible | Restreinte (11-30 %) |
Modérée-légère (1-30 %) |
Modérée-faible | La hausse prévue des températures estivales, combinée à des périodes de fortes pluies entrecoupées de sécheresses, pourrait toucher toutes les populations de l'espèce en Nouvelle-Écosse, tandis qu'une réduction de la fréquence du brouillard pourrait avoir une incidence sur les occurrences du comté de Shelburne en particulier. Selon les modèles, la superficie de la zone où le climat est propice à l'espèce pourrait augmenter légèrement au Nouveau-Brunswick, au Cap-Breton et à Terre-Neuve. Combinaison de 11.1 à 11.4 * À l'échelle mondiale, l'espèce semble avoir une assez bonne capacité d'adaptation et se rencontre dans un certain nombre de types de milieux. * Selon les modèles de changements climatiques, les changements climatiques sont probablement nuisibles à l'espèce, mais ces modèles semblent indiquer que, à long terme, la superficie de milieux propices pourrait augmenter. |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | - |
| 11.2 | Sécheresses | - | - | - | - | - | - |
| 11.3 | Températures extrêmes | - | - | - | - | - | - |
| 11.4 | Tempêtes et inondations | - | - | - | - | - | - |