
Recommandations pour la qualité de l'eau potable au Canada : Document technique – Les virus entériques
Santé Canada, Ottawa,Ontario
Avril, 2019
Table des matières
Partie I. Vue d'ensemble et application
Partie II. Science et considérations techniques
- 4.0 Description et effets sur la santé
- 5.0 Sources et exposition
- 6.0 Méthodes d'analyse
- 7.0 Techniques de traitement
- 8.0 Évaluation des risques
- 9.0 Justification de la recommandation
- 10.0 Références
Annexe B : Tableaux
- Tableau B.1. Caractéristiques des virus entériques humains d'origine hydrique
- Tableau B.2. Présence des virus entériques dans les eaux de surface au Canada et aux États-Unis
- Tableau B.3. Présence de virus entériques dans les eaux souterraines au Canada et aux États-Unis
- Tableau B.4. Présence de virus entériques dans l'eau potable au Canada et aux É.-U.
- Tableau B.5. Sélection d'éclosions de virus entériques liées à l'eau potable

Télécharger le rapport complet
(PDF format, 1,35 Mo, 75 pages)
Organisation : Santé Canada
Type : Recommandations
Date publiée : 2019-04-12
Sujets connexes
Partie I. Vue d'ensemble et application
1.0 Recommandation
La recommandation pour les virus entériques dans l'eau potable est un objectif de traitement basé sur la santé correspondant à une élimination et (ou) une inactivation d'au moins 4 log des virus entériques. Selon la qualité de la source d'approvisionnement en eau, une élimination et (ou) inactivation logarithmique plus importante pourrait être nécessaire. Les méthodes actuellement disponibles pour la détection des virus entériques ne sont pas applicables pour une surveillance régulière. Des techniques de traitement et des mesures de protection des sources d'approvisionnement en eau reconnues pour leur efficacité à réduire le risque de maladies d'origine hydrique doivent être mises en œuvre et maintenues si la source d'approvisionnement en eau est susceptible d'être contaminée par des matières fécales ou si des virus entériques ont déjà causé des éclosions de maladies d'origine hydrique.
2.0 Sommaire
Les virus sont des microorganismes extrêmement petits qui sont incapables de se reproduire à l'extérieur d'une cellule hôtesse. En général, les virus sont spécifiques à l'hôte, ce qui signifie que les virus qui infectent des animaux ou des végétaux n'infectent habituellement pas les humains, même si quelques virus entériques ont été détectés à la fois chez les humains et chez les animaux. La plupart des virus infectent aussi certains types de cellules seulement chez un hôte. C'est pourquoi les effets qu'une infection virale a sur la santé varient énormément. Les virus qui peuvent se multiplier dans le tractus gastro-intestinal des humains ou des animaux sont appelés « virus entériques ». Il y a plus de 140 sérotypes de virus entériques qui peuvent infecter les humains.
Santé Canada a terminé récemment son examen des risques pour la santé découlant de la présence de virus entériques dans l'eau potable. Le présent document technique passe en revue et évalue tous les risques connus pour la santé qui sont associés à la présence de virus entériques dans l'eau potable. Il tient compte des nouvelles études et approches, ainsi que des limites de la méthodologie et de l'interprétation des méthodes d'analyse disponibles pour détecter les virus dans l'eau potable. D'après cet examen, la recommandation pour les virus entériques dans l'eau potable est un objectif de traitement basé sur la santé qui consiste à diminuer la teneur en virus entériques d'au moins 4 log (soit 99,99 %) par élimination et (ou) inactivation.
2.1 Effets sur la santé
Différentes maladies humaines sont associées aux virus entériques. La maladie gastro-intestinale est le principal effet des virus entériques sur la santé. Les virus entériques peuvent aussi causer des maladies aiguës graves comme la méningite, la poliomyélite et des maladies fébriles non spécifiques. Ils ont aussi été incriminés dans des maladies chroniques, comme le diabète sucré et le syndrome de fatigue chronique.
La période d'incubation et la gravité des effets sur la santé dépendent du virus qui a causé l'infection. La gravité des effets d'une infection virale sur la santé dépendra aussi des caractéristiques de la personne en cause (p. ex. âge, état de santé). En théorie, une seule particule virale infectieuse peut causer une infection; cependant, toute infection dépend de la capacité de la particule virale à se reproduire à l'intérieur des cellules de l'hôte. Pour beaucoup de virus entériques, on sait ou on suppose que le nombre de particules virales infectieuses nécessaires pour causer une infection est faible.
2.2 Exposition
Les virus entériques ne peuvent pas se multiplier dans l'environnement, mais ils peuvent survivre pendant des périodes prolongées (p. ex. deux à trois ans dans les eaux souterraines) et sont plus infectieux que la plupart des autres microorganismes. Les virus entériques sont excrétés dans les selles des personnes ou des animaux infectés et certains virus entériques peuvent aussi être excrétés dans l'urine. Les sources d'eau peuvent être contaminées par des matières fécales humaines qui empruntent toutes sortes de voies, y compris les effluents des stations de traitement des eaux usées, les fuites d'égouts sanitaires, les rejets de lagunes de traitement des eaux usées et des fosses septiques. Les virus peuvent aussi pénétrer dans le réseau de distribution pendant la construction de conduites principales, lorsque des activités d'exploitation ou d'entretien entraînent des fluctuations de pression dans le réseau ou lorsque des composants souterrains sont inondés.
On a détecté la présence de virus entériques dans les eaux de surface et dans des sources souterraines. Ils semblent très prévalents dans les eaux de surface et leur présence varie en fonction du temps et du lieu. Dans le cas des eaux souterraines, on a détecté des virus à la fois dans des aquifères confinés et des aquifères non confinés, et ils peuvent parcourir des distances importantes (c. à d. des centaines de mètres) en peu de temps (c. à d. en quelques heures ou jours). Les aquifères confinés comportent une couche de faible perméabilité qui peut bloquer le transport du virus. Ces aquifères peuvent toutefois demeurer quand même vulnérables à la contamination virale à cause de voies comme des fractures, des trous percés par des racines et d'autres interruptions qui permettent au virus de traverser la couche encaissante pour pénétrer dans l'aquifère sous-jacent. La présence de virus entériques dans les eaux souterraines n'est généralement pas continue et peut varier considérablement avec le temps. On a établi un lien entre la maladie et la consommation d'eau souterraine contaminée par des matières fécales qui n'est pas traitée ou est insuffisamment traitée.
2.3 Analyse et traitement
Une méthode de gestion des risques, tel l'approche de la source au robinet ou du plan de gestion de la sécurité sanitaire de l'eau, constitue la meilleure façon de diminuer les virus entériques et les autres microorganismes pathogènes d'origine hydrique dans l'eau potable. Ce genre d'approche nécessite une évaluation du système afin de : caractériser la source d'approvisionnement en eau; décrire les procédés de traitement qui sont en place; cerner les conditions qui pourraient donner lieu à une contamination; et mettre en place les mesures correctives nécessaires pour atténuer les risques. La détermination de la vulnérabilité d'une source d'approvisionnement en eau à la contamination par les matières fécales, surtout celles d'origine humaine (p.ex. fosses septiques, fuites d'égouts sanitaires), constitue un élément important d'une évaluation d'un système parce que la surveillance régulière de la présence de virus entériques dans l'eau potable n'est pas pratique pour le moment. La collecte d'échantillons de la source d'approvisionnement en eau et leur analyse pour détecter la présence de virus entériques sont toutefois importantes pour les services publics de distribution d'eau qui veulent procéder à une évaluation quantitative du risque microbien. Il existe des méthodes moléculaires et de culture cellulaire validées pour la détection des virus entériques.
Une fois établies les caractéristiques de la source d'approvisionnement en eau, il faudrait fixer des objectifs de diminution des microorganismes pathogènes, par inactivation ou élimination, et mettre en place des procédés de traitement efficaces pour diminuer la concentration de virus entériques dans l'eau potable traitée. Il existe plusieurs techniques efficaces pour diminuer le nombre de virus entériques dans l'eau potable; dans la plupart des cas, la désinfection primaire sera le procédé principal d'inactivation des virus. En règle générale, il convient de filtrer (ou d'employer une technique équivalente) et de désinfecter tout approvisionnement tiré des eaux de surface ou des eaux souterraines sous l'influence directe des eaux de surface (ESSIDES) pour atteindre les objectifs du traitement des virus entériques et des protozoaires. Dans les eaux de subsurface jugées vulnérables aux virus, l'élimination ou l'inactivation logarithmique devrait atteindre un facteur d'au moins 4 log.
L'absence de bactéries indicatrices (p. ex. Escherichia coli, coliformes totaux) ne signifie pas nécessairement qu'il n'y a pas de virus entériques. L'application et le suivi d'une approche de la source au robinet ou d'un plan de gestion de la sécurité sanitaire de l'eau, y compris la surveillance des procédés et de la conformité (p. ex. turbidité, méthode de désinfection, E. coli), sont importants pour vérifier si l'eau a été traitée adéquatement et si elle est, par conséquent, d'une qualité microbiologique acceptable. Dans le cas des eaux souterraines non traitées, les tests de détection de la présence de bactéries indicatrices sont utiles, mais pas nécessairement suffisants, pour évaluer le potentiel de contamination fécale, ce qui peut inclure les virus entériques. Les résultats des essais bactériologiques devraient être considérés conjointement avec une évaluation de vulnérabilité propre au site.
2.4 Évaluation quantitative du risque microbien
L'évaluation quantitative du risque microbien (ÉQRM) est un outil qui emploie les données sur la qualité des sources d'approvisionnement en eau, l'information sur les procédés de traitement et les caractéristiques particulières du microorganisme pathogène présent pour estimer la charge de morbidité liée à l'exposition aux microorganismes pathogènes dans l'eau potable. L'ÉQRM est généralement utilisée à deux fins : elle peut servir à établir des objectifs de diminution des microrganismes pathogènes dans le cadre de l'élaboration de recommandations pour la qualité de l'eau potable, comme dans ce document. L'ÉQRM peut également servir à établir l'ordre de priorité des risques propres à un site dans le cadre d'une approche de la source au robinet ou du plan de gestion de la sécurité sanitaire de l'eau.
L'ÉQRM tient compte de différents virus entériques dont les caractéristiques font de bons représentants de tous les virus pathogènes similaires. Un virus de référence est sélectionné parmi ces virus entériques. On présume que contrôler le virus de référence assurerait aussi le contrôle de tous les autres virus préoccupants similaires. De nombreux virus entériques ont été envisagés. Comme il n'y a aucun virus en particulier qui possède toutes les caractéristiques d'un virus de référence idéal, la présente évaluation des risques utilise des caractéristiques de plusieurs différents virus.
2.5 Considérations internationales
Les recommandations, normes ou directives relatives à la qualité de l'eau potable établies par des gouvernements étrangers ou des agences internationales peuvent varier en raison des connaissances scientifiques disponibles au moment de l'évaluation, ainsi que de l'utilisation de différentes politiques et approches.
Diverses organisations ont établi des recommandations ou directives pour les virus entériques. L'U.S. EPA impose généralement une élimination ou inactivation de virus entériques de 4 log dans les stations de production d'eau potable. L'Organisation mondiale de la santé recommande l'adoption de mesures de contrôle (p. ex. prévention de la contamination des sources d'approvisionnement d'eau, traitement adéquat) dans le cadre d'un plan de gestion de la sécurité sanitaire de l'eau, afin de réduire les risques potentiels de virus entériques. Ni l'Union européenne, ni l'Australie (NHMRC) n'ont établi une recommandation ou une directive pour la diminution des virus entériques dans l'eau potable.
3.0 application de la recommandation
Remarque : Des conseils précis concernant l'application des recommandations pour l'eau potable devraient être obtenus auprès de l'autorité compétente en matière d'eau potable dans le territoire concerné.
La mise en œuvre d'une méthode de gestion des risques pour les systèmes d'approvisionnement en eau potable, comme l'approche de la source au robinet ou le plan de gestion de la sécurité sanitaire de l'eau, devrait réduire l'exposition aux virus. Ces méthodes obligent à procéder à une évaluation d'un réseau qui consiste à caractériser la source d'approvisionnement en eau, à décrire les procédés de traitement qui empêchent ou réduisent la contamination, à mettre en évidence les conditions qui peuvent causer la contamination et à déterminer des mesures de contrôle à prendre pour atténuer les risques par les systèmes de traitement et de distribution au consommateur.
3.1 Évaluations des sources d'approvisionnement en eaupotable
Les évaluations des sources d'approvisionnement en eau devrait faire partie des évaluations périodiques des systèmes. Elles doivent porter sur la détermination de sources possibles de contamination fécale dans le bassin versants ou l'aquifère, les voies ou les événements possibles (risque faible à élevé) qui font que les virus entériques peuvent se retrouver dans la source d'approvisionnement en eau et les conditions susceptibles de produire des concentrations maximales de virus entériques. Pour les sources d'approvisionnement en eau de subsurface, ces évaluations devraient inclure idéalement une étude hydrogéologique et au moins une évaluation de la vulnérabilité de l'aquifère, l'intégrité du puits et une enquête sur les sources possibles de contamination fécale de la région. La surveillance des indicateurs microbiens permet de savoir si une source d'approvisionnement en eau souterraine peut être touchée par la contamination fécale. Dans le cas des eaux de subsurface jugées vulnérables à la contamination virale, l'élimination ou l'inactivation des virus entériques devrait atteindre au moins 4 log. Dans le cas des sources ESSIDES, un traitement supplémentaire peut s'imposer pour contrer d'autres contaminants microbiologiques comme les protozoaires entériques.
Là où la surveillance des virus est possible, on prélève généralement des échantillons à un endroit représentatif de la qualité de l'eau qui alimente le réseau de distribution d'eau potable, comme à l'admission d'une station de traitement des eaux ou, dans le cas des eaux souterraines, à chaque puits. Dans le cas des eaux de surface, il est recommandé de procéder à une analyse mensuelle pendant les quatre saisons pour déterminer les niveaux de référence et caractériser au moins deux événements météorologiques afin de comprendre les conditions des pointes. Comme la qualité des eaux de surface varie avec le temps, il peut être nécessaire de procéder à un échantillonnage plus intensif (c.-à-d. cinq échantillons par semaine) pour quantifier les concentrations de pointe. Dans le cas des eaux souterraines, y compris des aquifères confinés, la présence d'une contamination virale est difficile à prédire. On recommande une analyse mensuelle pendant les quatre saisons afin de caractériser adéquatement la présence d'une contamination virale. Les répercussions des conditions de pluie et de sécheresse sur les concentrations de virus dans les eaux souterraines doivent aussi être prises en compte, étant donné qu'elles peuvent représenter des périodes de risque élevé. Les puits qui sont utilisés sur une base saisonnière peuvent être assujettis à des exigences propres à chaque site, déterminées par l'autorité compétente. Si l'on compte utiliser des données de surveillance dans le cadre d'une ÉQRM, il faut en déterminer la viabilité et l'infectiosité, ainsi que l'efficience de la méthode utilisée sur le plan de la récupération.
3.2 Procédés de traitement appropriés
Une élimination ou une inactivation logarithmique d'au moins 4 log des virus entériques est recommandée pour toutes les sources d'approvisionnement en eau, y compris les sources d'approvisionnement en eau souterraines. Pour bon nombre de sources d'approvisionnement en eau, une diminution de plus de 4 log peut s'imposer. Une administration peut autoriser une diminution inférieure au minimum recommandé de 4 log pour une source d'approvisionnement en eau souterraine si l'évaluation du système d'approvisionnement en eau potable a confirmé que le risque de la présence de virus entériques est minime ou que l'aquifère assure une filtration locale adéquate.
L'élimination mécanique des virus (p. ex. par filtration naturelle ou technique) peut être difficile à cause de leur petite taille et des variations de leur charge superficielle. La désinfection constitue donc une barrière importante pour diminuer de façon appropriée la concentration de virus dans l'eau potable. Il est possible de neutraliser efficacement les virus en appliquant différentes techniques de désinfection, individuellement ou en combinaison. Pour déterminer le type et le degré de traitement indiqués, il faut tenir compte des fluctuations possibles de la qualité de l'eau, y compris de sa dégradation à court terme, ainsi que de la variabilité de l'efficacité du traitement. Des essais pilotes ou d'autres procédés d'optimisation peuvent aider à déterminer la variabilité du traitement.
Les petits réseaux d'eau souterraine qui fournissent de l'eau potable au public (c.-à-d. les réseaux semi-publics) et qui sont vulnérables à la contamination virale devraient faire l'objet d'un traitement visant à éliminer ou à inactiver les virus entériques. L'utilisation de systèmes certifiés de désinfection aux rayons ultraviolets (UV) exploités à une dose de 40 mJ/cm2 est efficace pour parvenir à une inactivation de 4 log de la plupart des virus entériques, à l'exception de l'adénovirus. Une dose de 186 mJ/cm2 n'est pas jugée nécessaire, puisque l'eau potable n'est pas une source principale d'exposition à ce virus au Canada, et qu'il n'a jamais été associé à une éclosion au pays. Les ménages qui possèdent un puits privé doivent en évaluer la vulnérabilité à la contamination par les matières fécales pour déterminer s'il faut le traiter. Les provinces et les territoires offrent des recommandations générales sur la construction, l'entretien, la protection et l'analyse des puits. Lorsqu'ils veulent déterminer précisément le risque de contamination virale, les propriétaires de puits privés devraient comprendre les enjeux relatifs à l'aménagement d'un puits, notamment leur construction, les caractéristiques du sol et l'emplacement du puits par rapport aux sources de contamination fécale (p. ex. fosses septiques, égouts sanitaires, déchets d'origine animale).
Si le propriétaire d'un puits privé est incapable de déterminer que le risque de virus entériques atteignant son puits est minimal, il devrait communiquer avec l'autorité compétente en matière d'eau potable dans le territoire concerné qui déterminera les options de traitement possibles.
3.3 Entretien et exploitation appropriés des réseaux de distribution
Des virus peuvent pénétrer dans un réseau de distribution au cours de la construction ou de la réparation de conduites principales, lorsque les activités périodiques d'exploitation et d'entretien provoquent des transitoires de pression (p. ex. utilisation de vannes ou de bornes-fontaines, démarrage ou arrêt de pompes) ou encore à la suite de l'immersion de purgeurs d'air ou de reniflards situés dans des chambres souterraines.
Les résiduels de désinfectant secondaires types seraient inefficaces lorsqu'il s'agit de neutraliser des virus dans le réseau de distribution. Il s'ensuit que le maintien de l'intégrité physique et hydraulique du réseau de distribution et la diminution du nombre de phénomènes de pression négative ou de basse pression sont les éléments essentiels d'une approche de la source au robinet ou du plan de gestion de la sécurité sanitaire de l'eau. Les réseaux de distribution dont l'entreposage est insuffisant ou inexistant (p.ex. réservoir, canalisation incendie) tendent à être plus vulnérables aux phénomènes transitoires de pression et les conduites principales en cul-de-sac ont tendance à subir des transitions plus importantes que les conduites principales en boucle fermée. Les chambres de vannes souterraines susceptibles d'être inondées devraient faire l'objet d'une inspection mensuelle et être drainées si elles sont inondées. Il est recommandé de surveiller régulièrement la qualité de l'eau du réseau de distribution (p. ex. les indicateurs microbiens, les résiduels de désinfectant, la turbidité, le pH), mettre en place des programmes d'exploitation et d'entretien (p. ex. rinçage de conduites, prévention des retours d'eau polluée, gestion des actifs) et pratiquer une hygiène rigoureuse pendant tous les travaux de construction, de réparation et d'entretien de conduitesafin que l'eau potable soit acheminée jusqu'au consommateur avec une qualité acceptable.
Partie II. Science et considérations techniques
4.0 Description et effets sur la santé
La taille des virus varie de 20 à 350 nm : ce sont donc les microorganismes les plus petits. Ils sont constitués d'un génome d'acide nucléique (soit de l'acide ribonucléique [ARN]ou l'acide désoxyribonucléique [ADN]) entouré d'une capsule protéique protectrice, la capside. Certains virus, dits virus enveloppés, ont aussi une enveloppe de lipoprotéines qui entoure la capside. Les virus non enveloppés n'ont pas cette enveloppe de lipoprotéines. Les virus peuvent se reproduire seulement dans une cellule hôtesse vivante. Même si le génome viral code les protéines structurelles virales et d'autres molécules nécessaires à la reproduction, les virus doivent compter sur le métabolisme cellulaire de l'hôte pour synthétiser ces molécules.
La reproduction virale dans les cellules hôtesses entraîne la production de virions infectieux et de nombreuses particules incomplètes non infectieuses (Payment et Morin, 1990). Le rapport entre les particules du virus comme tel et le nombre réel de virions infectieux varie de 10:1 à plus de 1000:1. Dans le contexte des maladies d'origine hydrique, un « virus » est donc défini comme une « particule du virus complet » ou « virion » infectieux avec son génome d'ADN ou d'ARN et son enveloppe protéique tels qu'ils existent hors de la cellule. Il s'agirait de la forme la plus simple dans laquelle un virus peut infecter un hôte. Dans l'environnement, les virions infectieux se dégraderont et perdront leur infectiosité, mais il est quand même possible de les voir au microscope électronique ou de les détecter par des méthodes moléculaires.
En général, les virus sont propres à l'hôte. Par conséquent, les virus qui peuvent infecter les humains n'infectent habituellement pas des hôtes non humains, comme des animaux ou des végétaux. L'inverse est vrai aussi : les virus qui infectent des animaux et des végétaux n'infectent habituellement pas les humains, même si l'on a détecté la présence d'un faible nombre de virus entériques à la fois chez les humains et les animaux (c.-à-d. virus zoonotiques). La plupart des virus infectent aussi seulement des types particuliers de cellules dans un hôte. Les types de cellules susceptibles dépendent du virus et, par conséquent, les effets sur la santé découlant d'une infection virale varient beaucoup, selon l'endroit où les cellules susceptibles se trouvent dans le corps. En outre, l'infection virale peut déclencher des réactions immunitaires qui produisent des symptômes non spécifiques. Les virus qui peuvent se multiplier dans le tractus gastro-intestinal des humains ou des animaux sont des « virus entériques ». Les virus entériques sont excrétés dans les selles des personnes infectées et certains virus entériques peuvent aussi être excrétés dans l'urine (Hilt et coll., 2014). Ces excrétions peuvent contaminer les sources d'approvisionnement en eau. Les virus non entériques, comme les virus respiratoires, ne sont pas considérés comme des microorganismes pathogènes d'origine hydrique puisqu'ilsne se transmettent pas facilement des personnes infectées aux sources d'approvisionnement en eau.
Il existe plus de 200 virus entériques reconnus (Haas et coll., 2014), dont 140 sérotypes sont reconnus pour infecter les humains (AWWA, 1999; Taylor et coll., 2001). Les maladies associées aux virus entériques sont variées. En plus de la gastroentérite, les virus entériques peuvent causer des maladies aiguës graves, comme la méningite, la poliomyélite et des maladies fébriles non spécifiques. Ils ont aussi été mis en cause dans l'étiologie de certaines maladies chroniques, comme le diabète sucré et le syndrome de la fatigue chronique.
Les virus entériques couramment associés aux maladies humaines d'origine hydrique comprennent les norovirus, le virus de l'hépatite a (VHA), le virus de l'hépatite E (VHE), les rotavirus et les entérovirus. Les caractéristiques de ces virus entériques, ainsi que leurs effets connexes sur la santé, sont abordés ci-dessous et résumés dans le tableau B.1 à l'annexe B. Le tableau B.1 comprend également certains virus entériques qu'on a peu souvent, ou de manière peu concluante, associés à des éclosions de maladies humaines liées à l'eau potable, mais qui peuvent pourtant causer des maladies.
4.1 Virus entériques couramment associés à des maladies humaines
4.1.1 Norovirus
Les norovirus sont des virus à aRN monocaténaire sans enveloppe d'un diamètre de 35 à 40 nm appartenant à la famille des Caliciviridae. Les norovirus sont actuellement subdivisés en sept génogroupes (GI à GVII) constitués de plus de 40 génotypes distincts (CDC, 2013a; Vinjé, 2015). On continue toutefois d'identifier de nouvelles variantes de norovirus et l'on en a détecté plus de 150 souches dans les eaux usées seulement (Aw et Gin, 2010; Kitajima et coll., 2012). Les génogroupes GI, GII et GIV contiennent les génotypes de norovirus habituellement associés aux maladies humaines (Verheof et coll., 2015), les norovirus du génogroupe II, et plus précisément le GII.4, causant plus de 90 % du total des cas sporadiques de gastroentérite aiguë chez les enfants (Hoa Tran et coll., 2013).
Même si la plupart des norovirus semblent propres à l'hôte, on a signalé des cas d'animaux infectés par des norovirus humains. On a, par exemple, isolé des variantes du GII chez des animaux de ferme (Mattison et coll., 2007; Chao et coll., 2012) et des chiens (Summa et coll., 2012), et c'est pourquoi on se demande si le norovirus est transmissible entre animaux et humains. On n'a pas signalé la présence de norovirus animaux chez les humains et l'on n'a pas détecté d'autres génogroupes, comme le GIII, le GV et le GVI, ailleurs que chez des hôtes non humains (Karst et coll., 2003; Wolf et coll., 2009; Mesquita et coll., 2010).
Les infections norovirales surviennent chez les nourrissons, les enfants et les adultes. La période d'incubation est de 12 à 48 h (CDC, 2013a). Les effets sur la santé des infections norovirales sont résolutifs et durent habituellement de 24 à 48 heures. Les symptômes comprennent les nausées, les vomissements, la diarrhée, les douleurs abdominales et la fièvre. Chez les personnes en santé, les symptômes sont généralement très désagréables mais ne mais ne posent pas de menace pour la vie. Chez les groupes vulnérables, comme les personnes âgées, la maladie est considérée plus grave. Teunis et coll. (2008) ont signalé que la dose infectieuse du norovirus était faible (≥ 18 particules virales). Schmidt (2015) a toutefois déterminé des limites de l'étude et conclu qu'il se peut que l'infectiosité soit surestimée (voir la section 8.3.1). Plusieurs études ont signalé que certaines personnes avaient une résistance naturelle à l'infection aux norovirus. On pense qu'il se peut qu'il manque à ces personnes un récepteur à la surface des cellules nécessaire pour lier le virus ou qu'elles aient une réponse immunitaire anamnestique qui empêche l'infection (Hutson et coll., 2003; Lindesmith et coll., 2003; Cheetham et coll., 2007). L'immunité à l'infection norovirale semble de courte durée, de l'ordre de plusieurs mois. Une estimation récente tirée d'un modèle de transmission indique toutefois que l'immunité peut durer des années (Simmons et coll., 2013).
Les norovirus sont excrétés à la fois dans les matières fécales et dans les vomissures des personnes infectées et ils peuvent être transmis par l'eau contaminée. Les personnes infectées peuvent excréter le norovirus avant d'avoir des symptômes et pendant deux semaines ou plus après leur disparition (Atmar et coll., 2008; aoki et coll., 2010). Les norovirus se transmettent aussi facilement par contact de personne à personne. On a associé un grand nombre des cas de gastroentérite norovirale à des groupes de personnes vivant dans un milieu fermé comme des écoles, des camps de vacances, des établissements et des bateaux de croisière. L'ingestion de particules en aérosol peut aussi causer l'infection (CDC, 2011; Repp et Keene, 2012). Les infections à norovirus sont très saisonnières et atteignent un pic le plus souvent au cours des mois d'hiver (Ahmed et coll., 2013).
4.1.2 Virus de l'hépatite
À ce jour, six types de virus de l'hépatite ont été identifiés (A, B, C, D, E et G), mais deux d'entre eux seulement, soit l'hépatite a (VHA) et l'hépatite E (VHE), semblent transmis par la voie fécale-orale et sont donc associés à la transmission d'origine hydrique. Bien que le VHA et le VHE puissent tous deux causer l'hépatite (inflammation du foie), pour laquelle il n'existe aucun traitement spécifique, il s'agit de deux virus distincts.
4.1.2.1 Virus de l'hépatite a
Le VHA est un petit virus à aRN monocaténaire de 27 à 32 nm de diamètre, sans enveloppe et à symétrie icosaédrique. Il appartient à la famille des Picornaviridae et il avait été classé à l'origine dans le genre Enterovirus. Cependant, ilprésente certaines propriétés particulières en termes de structure génétique et de reproduction, on l'a classé dans un nouveau genre, Hepatovirus, dont il est le seul membre (Carter, 2005).
La période d'incubation d'une infection à VHA varie de 15 à 50 jours, la moyenne s'établissant à environ 28 jours (CDC, 2015a). La dose infectieuse médiane pour le VHA est inconnue, mais on suppose qu'elle est faible (c.-à-d. 10 à 100 particules virales) (FDA, 2012). Les infections à VHA, communément appelées hépatites infectieuses, provoquent de nombreux symptômes, dont la fièvre, des malaises (fatigue), l'anorexie, des nausées et de la gêne abdominale, suivis de quelques jours de jaunisse. L'infection à VHA peut aussi causer au foie des dommages qui découlent de la réponse immunitaire de l'hôte à l'infection des hépatocytes par le VHA. Dans certains cas, les dommages au foie peuvent être mortels.
L'infection à VHA survient chez les enfants et les adultes. La maladie est habituellement résolutive, mais sa gravité augmente avec l'âge. Par exemple, on observe seulement des symptômes minimes ou aucun symptôme chez les plus jeunes enfants (Yayli et coll., 2002), mais dans une étude portant sur des cas d'infection à VHA chez des personnes âgées de 50 ans et plus, on a observé un taux de mortalité six fois plus élevé que le taux moyen de 0,3 % (Fiore, 2004). Le virus est excrété dans les selles des personnes infectées pendant jusqu'à deux semaines avant l'apparition des symptômes de l'hépatite et le virus se propage par la voie fécale-orale (Chin, 2000; Hollinger et Emerson, 2007; CDC, 2015a). Le VHA est aussi excrété dans l'urine des personnes infectées (Giles et coll., 1964; Hollinger et Emerson, 2007; Joshi et coll., 2014). La convalescence peut être prolongée (huit à dix semaines) et dans certains cas d'infection à VHA, les personnes infectées peuvent avoir des rechutes pendant jusqu'à six mois (CDC, 2015a).
Le taux d'incidence de VHA est la plus élevée en asie, en afrique, en amérique latine et au Moyen-Orient (Jacobsen et Wiersma, 2010). Au Canada, le taux l'incidence de VHA a diminué considérablement depuis l'introduction du vaccin contre le VHA en 1996 (ASPC, 2015a). Des études sur la séroprévalence ont signalé une prévalence nationale de 2 % et 20 % chez les enfants et les adultes d'origine canadienne non vaccinés, respectivement (Pham et coll., 2005; aSPC, 2015b). Les cas de VHA non liée aux voyages sont rares au Canada.
4.1.2.2 Virus de l'hépatite E
Le VHE est un virion sans enveloppe d'un diamètre de 27 à 34 nm à aRN polyadénylé monocaténaire qui appartient à la famille des Hepeviridae. Même si la plupart des virus entériques humains n'ont pas de réservoir non humain, on a signalé que le VHE est zoonotique (transmis des animaux aux humains, avec des réservoirs naturels non humains) (AWWA, 1999; Meng et coll., 1999; Wu et coll., 2000; Halbur et coll., 2001; Smith et coll., 2002; Smith et coll., 2013, 2014). Il y a quatre génotypes de VHE infectieux pour les humains. Les génotypes 1 et 2 sont transmis entre humains, tandis que les génotypes 3 et 4 semblent zoonotiques (transmis aux humains par le chevreuil, le porc et le sanglier) (Smith et coll., 2014). Ces génotypes ont été subdivisés de nouveau en au moins 24 sous-types (Smith et coll., 2013), mais cette classification est en révision (Smith et coll., 2014).
Sur le plan clinique, il est impossible de distinguer l'infection à VHE de l'infection à VHA. Les symptômes comprennent les malaises, l'anorexie, des douleurs abdominales, l'arthralgie, la fièvre et la jaunisse. La dose infectieuse médiane de VHE est inconnue. Sa période d'incubation varie de 15 à 60 jours, la moyenne s'établissant à 42 jours (CDC, 2015b). L'infection à VHE se résorbe habituellement d'une à six semaines après son apparition. Les virions sont excrétés dans les selles pendant une semaine ou plus après l'apparition des symptômes (Percival et coll., 2004). La maladie est le plus souvent déclarée chez les jeunes adultes et les adultes d'âge moyen (15 à 40 ans). Le taux de mortalité est de 0,5 à 3 %, sauf chez les femmes enceintes dont le taux de mortalité peut atteindre presque 20 à 25 % (Matson, 2004). Les maladies associées au VHE sont rares dans les pays industrialisés, la plupart des infections étant liées aux voyages à l'étranger.
4.1.3 Rotavirus
Les rotavirus sont des virus à aRN bicaténaire non enveloppés d'environ 70 nm de diamètre appartenant à la famille des Reoviridae. Ces virus ont été divisés en huit groupes sérologiques (A à H) (Marthaler et coll., 2012), dont trois (A, B et C) infectent les humains. Les rotavirus du groupe A sont subdivisés en sérotypes selon les caractéristiques de leurs protéines de surface, VP7 et VP4. Il y a 28 types de VP7 (appelés types G) et quelque 39 types de VP4 (les types P) qui produisent une grande diversité antigénique (Mijatovic-Rustempasic et coll., 2015, 2016). Même si la plupart des rotavirus semblent propres à l'hôte, des études révèlent leur potentiel de transmission zoonotique (Cook et coll., 2004; Kang et coll., 2005; Gabbay et coll., 2008; Steyer et coll., 2008; Banyai et coll., 2009; Doro et coll., 2015; Mijatovic-Rustempasic et coll., 2015, 2016), mais on croit que c'est plutôt rare et que la transmission n'est probablement pas pathogène (CDC, 2015c).
En général, les rotavirus causent la gastroentérite, y compris des vomissements et de la diarrhée. Le sujet peut vomir pendant une période allant jusqu'à 48 heures avant l'apparition de la diarrhée. La gravité de la gastroentérite peut varier de bénigne, durant moins de 24 heures, à grave, dans certains cas, qui peuvent être mortels. Chez les jeunes enfants, les manifestations extra-intestinales possibles, comme des symptômes respiratoires et des convulsions, sont attribuables au fait que l'infection est systémique plutôt que localisée à la muqueuse jéjunale (Candy, 2007). La période d'incubation dure généralement moins de 48 heures (CDC, 2015c). La maladie dure généralement de cinq à huit jours. La dose infectieuse médiane de rotavirus est estimée à six particules virales (Haas et coll., 1999). Le virus est excrété en nombre exceptionnellement élevé par les personnes infectées, jusqu'à 1011/g de selles (Doro et coll., 2015). Certains rotavirus peuvent aussi produire une protéine toxique qui peut provoquer la diarrhée lors du contact avec les cellules infectées (Ball et coll., 1996; Zhang et coll., 2000). Ce phénomène est inhabituel, puisque la plupart des virus n'ont pas d'effet toxique.
Le rotavirus du groupe a est endémique dans le monde entier et constitue le groupe le plus courant et le plus répandu. C'est la principale cause de diarrhée aiguë (et de la déshydratation qui en découle) chez les humains et chez plusieurs espèces animales (Estes et Greenberg, 2013). On qualifie les infections qu'il cause de diarrhée infantile, diarrhée hivernale, gastroentérite infectieuse non bactérienne aiguë et gastroentérite virale aiguë. Les enfants de six mois à deux ans, les nouveau-nés prématurés, les personnes âgées et les sujets dont le système immunitaire est compromis sont particulièrement vulnérables aux symptômes plus graves causés par l'infection à rotavirus du groupe a. Ce dernier est la principale cause de diarrhée grave chez les nourrissons et les enfants et il cause environ la moitié des cas qui nécessitent une hospitalisation, habituellement à cause de la déshydratation. Aux États-Unis, avant le lancement d'un vaccin antirotavirus en 2006, il y avait plus de trois millions de cas par année chez les enfants (Parashar et coll., 2006; Glass et coll., 2012). Des infections asymptomatiques peuvent survenir chez des adultes, ce qui donne au virus un autre moyen de se propager dans la collectivité. Dans les régions tempérées, la maladie associée au rotavirus se manifeste principalement au cours de l'hiver, tandis que sous les tropiques, elle est présente pendant toute l'année (Moe et Shirley, 1982; nakajima et coll., 2001; Estes et Kapikian, 2007). Les maladies causées par le rotavirus du groupe B, aussi appelé rotavirus de la diarrhée chez l'adulte, ont été limitées surtout à la Chine, où l'on a signalé des éclosions de diarrhée grave touchant des milliers de personnes (Ramachandran et coll., 1998). On a établi un lien entre le rotavirus du groupe C et des cas rares et sporadiques de diarrhée chez les enfants dans de nombreux pays et régions, dont l'Amérique du nord (Jiang et coll., 1995). Les premières éclosions ont été signalées au Japon et en angleterre (Caul et coll., 1990; Hamano et coll., 1999).
4.1.4 Entérovirus
Les entérovirus (EV) constituent un grand groupe de (plus de 250) virus appartenant au genre Enterovirus et à la famille des Picornaviridae. Au nombre des plus petits virus, ils sont constitués d'un aRN monocaténaire sans enveloppe, de 20 à 30 nm de diamètre et à symétrie icosaédrique. Le genre Entérovirus comporte 12 espèces dont sept ont été associées à la maladie humaine: EV-A à EV-D et rhinovirus (RV)-A, B et C (Tapparel et coll., 2013; Faleye et coll., 2016; The Pirbright Institute, 2016). On continue d'identifier d'autres sérotypes d'entérovirus.
La période d'incubation et les effets sur la santé des infections à entérovirus sont variés. La période d'incubation des entérovirus varie de deux à 35 jours (AWWA, 2006). De nombreuses infections à entérovirus sont asymptomatiques. Lorsque des symptômes sont présents, leur gravité peut varier de bénigne à mortelle. La virémie (c.-à-d. le passage du virus dans le sang) est fréquente, offrant un moyen de transporter les entérovirus jusqu'à différents organes cibles et provoquant un éventail de symptômes. Les symptômes bénins comprennent la fièvre, les malaises, l'irritation de la gorge, les vomissements, les éruptions cutanées et des maladies des voies respiratoires supérieures. La gastroentérite aiguë est peu courante. Les complications les plus graves comprennent la méningite, l'encéphalite, la poliomyélite, la myocardite et des maladies fébriles non spécifiques chez les nouveau-nés et les jeunes enfants (Rotbart, 1995; Roivainen et coll., 1998). Il y a d'autres complications comme la myalgie, le syndrome de Guillain-Barré, l'hépatite et la conjonctivite. Les entérovirus ont aussi été incriminés dans l'étiologie de maladies chroniques comme la myosite inflammatoire, la myocardiopathie dilatée, la sclérose latérale amyotrophique, le syndrome de la fatigue chronique et l'atrophie musculaire post-poliomyélite (Pallansch et Roos, 2007; Chia et Chia, 2008). Des études appuient aussi l'existence d'un lien entre l'infection à entérovirus et l'apparition du diabète sucré insulinodépendant (type 1) (Nairn et coll., 1999; Lönnrot et coll., 2000; Latinen et coll., 2014; Oikarinen et coll., 2014). Bien que de nombreuses infections à entérovirus sont asymptomatiques, on estime qu'environ 50 % des infections à virus Coxsackie a et 80 % des infections à virus Coxsackie B provoquent la maladie (Cherry, 1992). On a aussi signalé que le virus Coxsackie B est l'entérovirus non poliomyélitique qui a été associé le plus souvent à des maladies graves (Mena et coll., 2003). Les infections à entérovirus atteignent leur pic au cours de l'été et au début de l'automne (Nwachuku et Gerba, 2006; Pallansch et Roos, 2007).
Les entérovirus sont endémiques dans le monde entier, mais peu d'éclosions d'origine hydrique on été signalés; parmi celles-ci, une éclosion liée à l'eau potable au Bélarus (Amvrosieva et coll., 2001) et d'autres éclosions associées à des eaux à usage récréatif (Hauri et coll., 2005; Sinclair et coll., 2009). Le nombre élevé de sérotypes, la nature habituellement bénigne des infections et le fait qu'ils soient très transmissibles dans une collectivité par contact direct entre personnes, expliquent probablement pourquoi nous en savons si peu sur leur transmission par voie hydrique (Lodder et coll., 2015).
4.2 Virus entériques rarement ou potentiellement associés aux maladies humaines
Les adénovirus sont membres de la famille des adenoviridae et incluent les virus icosaédriques sans enveloppe d'un diamètre de 70 à 100 nm à aDN linéaire bicaténaire. Il existe actuellement sept espèces reconnues (A à G) d'adénovirus humains qui regroupent plus de 60 (séro)types (Robinson et coll., 2013). La plupart des isolats d'origine hydrique sont des types 40 et 41 (Mena et Gerba, 2009), mais on a isolé aussi d'autres sérotypes (Van Heerden et coll., 2005; Jiang, 2006; Hartmann et coll., 2013). La période d'incubation varie de 3 à 10 jours (Robinson et coll., 2007). Les adénovirus peuvent causer tout un éventail de symptômes. Les sérotypes 40 et 41 causent la majorité des gastroentérites liées à des adénovirus. Les adénovirus sont une cause courante de gastroentérite virale aiguë chez les enfants (Nwachuku et Gerba, 2006). Les infections sont généralement limitées aux enfants de moins de cinq ans (FSA, 2000; Lennon et coll., 2007) et sont rares chez les adultes. L'infection cause une diarrhée et des vomissements qui peuvent durer une semaine (ASPC, 2010). La charge virale des selles des personnes infectées est élevée (~106 particules/g de matières fécales) (Jiang, 2006). Cela contribue à la transmission par la voie fécale-orale, que ce soit par contact direct avec des objets contaminés ou par les eaux utilisées à des fins récréatives et, potentiellement,, par l'eau potable. Les adénovirus ont été mis en cause dans deux éclosions liées à l'eau potable (une en Finlande et une en albanie), sans toutefois en être la cause principale (Kukkula et coll., 1997; Divizia et coll., 2004). Aucune éclosion d'adénovirus liée à l'eau potable n'a été signalée au Canada (Schuster et coll., 2005; Murphy et coll., 2017). L'eau potable ne constitue pas la principale voie d'exposition aux adénovirus.
Les astrovirus font partie de la famille des astroviridae. Il y en a huit sérotypes (HAst1-8) et l'on continue d'en découvrir de nouveaux types (Finkbeiner et coll., 2009a, 2009 b; Kapoor et coll., 2009; Jiang et coll., 2013). Les astrovirus regroupent deux génogroupes (A et B) capables d'infecter des êtres humains (Carter, 2005). Les membres de cette famille comprennent des virus sans enveloppe de 28 à 30 nm de diamètre à aRN monocaténaire. L'infection à astrovirus provoque habituellement une diarrhée durant deux à trois jours et la période d'incubation initiale varie de un à cinq jours (Lee et coll., 2013). L'infection provoque généralement une diarrhée moins forte que celle que causent les rotavirus et ne cause pas de déshydratation importante. Les autres symptômes observés à la suite d'une infection à astrovirus comprennent les maux de tête, les malaises, les nausées, les vomissements et une fièvre légère (Percival et coll., 2004; Méndez et arias, 2007). Les sérotypes 1 et 2 sont courants durant l'enfance (Palombo et Bishop, 1996). Il se peut que les autres sérotypes (4 et plus) surviennent à l'âge adulte seulement (Carter, 2005). Les éclosions d'astrovirus chez les adultes sont rares, mais possibles (Oishi et coll., 1994; Caul, 1996; Gray et coll., 1997). En général, les personnes en santé acquièrent une bonne immunité contre la maladie et la réinfection est donc rare. Les infections à astrovirus atteignent généralement leur pic au cours de l'hiver et du printemps (Gofti-Laroche et coll., 2003).
Les sapovirus ont été identifiés la première fois chez de jeunes enfants au cours d'une éclosion de gastroentérite à Sapporo, au Japon (Chiba et coll., 1979) et sont de plus en plus reconnus comme cause d'éclosions de gastroentérite dans le monde entier (Chiba et coll., 2000; Farkas et coll., 2004; Johansson et coll., 2005; Blanton et coll., 2006; Gallimore et coll., 2006 : Phan et coll., 2006; Pang et coll., 2009). Comme les norovirus, ils font partie de la famille des Caliciviridae (Atmar et Estes, 2001). On a détecté la présence de sapovirus dans des eaux environnementales et des eaux usées non traitées et traitées au Japon (Hansman et coll., 2007; Kitajima et coll., 2010a), en Espagne (Sano et coll., 2011) et au Canada (Qui et coll., 2015). On n'en a toutefois pas détecté dans l'eau potable (Sano et coll., 2011).
Les aichivirus font partie de la famille des Picornaviridae. Comme les sapovirus, on les a identifiés pour la première fois dans des échantillons de selles provenant de patients atteints de gastroentérite au Japon (Yamashita et coll., 1991). On en a détecté depuis la présence dans les selles de personnes de plusieurs pays, y compris la France, le Brésil et la Finlande (Reuter et coll., 2011). Même si l'on a détecté la présence d'aichivirus dans des eaux usées non traitées et traitées (Sdiri-Loulizi et coll., 2010), on sait très peu de choses de leur présence dans les sources d'approvisionnement en eau.
Les polyomavirus sont membres de la famille des Polymaviridae qui inclut des espèces infectant les humains, y compris le polyomavirus BK et le polyomavirus JC. Même si l'on a détecté la présence de ces virus dans des eaux environnementales et des eaux usées (Vaidya et coll., 2002; Bofill-Mas et Girones, 2003; aWWA, 2006; Haramoto et coll., 2010), on n'en a pas encore documenté la transmission par l'eau. Le fait que le polyomavirus JC est aussi excrété dans l'urine appuie la possibilité de transmission par l'eau contaminée (Polo et coll., 2004). On a associé les polyomavirus à des maladies chez des personnes dont le système immunitaire est compromis, par exemple, la gastroentérite, des maladies respiratoires et d'autres maladies plus graves, y compris le cancer (AWWA, 2006).
Il importe de signaler que l'on continue de détecter et de reconnaître de nouveaux virus entériques.
5.0 Sources et exposition
Différentes unités de concentration de virus, définies à l'annexe a, sont utilisées dans cette section et dans l'ensemble du document. Ces unités sont celles déclarées par les auteurs de l'étude et reflètent la méthode de détection utilisée. Il est important de tenir compte du fait que l'infectiosité des virus détectés n'a pas toujours été évaluée. Compte tenu de la variation de ces méthodes d'étude, il est impossible de comparer facilement les concentrations, surtout lorsque des méthodes moléculaires et non moléculaires sont employées.
5.1 Sources
5.1.1 Sources de contamination
Les matières fécales humaines constituent la principale source des virus entériques humains dans l'eau. Les virus entériques sont excrétés en grande quantité dans les selles des personnes infectées (symptomatiques et asymptomatiques). Ils se diffusent facilement dans l'environnement par les matières fécales et sont transmissibles à d'autres personnes par la voie fécale-orale. Les personnes infectées peuvent excréter plus d'un billion (1012) de virus/g de matières fécales (Bosch et coll., 2008; Tu et coll., 2008). La présence de ces virus dans une population humaine est variable et reflète la conjoncture épidémique et endémique. On a signalé que les concentrations de virus entériques atteignent leur maximum dans les échantillons d'eaux usées au cours de l'automne et de l'hiver, ce qui donne à penser que le taux endémique de la maladie est plus élevé pendant cette période de l'année ou que les virus entériques survivent mieux à des températures froides. La contamination de sources d'approvisionnement en eau par des matières fécales peut emprunter diverses voies, y compris les effluents de stations de traitement des eaux usées, l'épandage d' eaux usées domestiques ou de boues station d'épuration sur des terres, les fuites d'égouts sanitaires, les effluents de fosses septiques et l'infiltration d'eaux de surface dans des aquifères souterrains (Vaughn et coll., 1983; Bitton, 1999; Hurst et coll., 2001; Powell et coll., 2003; Borchardt et coll., 2004; Bradbury et coll., 2013). Certains virus entériques (p. ex. VHA) peuvent aussi être excrétés dans l'urine des personnes infectées (voir la section 4.7).
On détecte couramment la présence de virus entériques d'origine humaine dans les eaux usées non traitées et traitées. Bradbury et coll. (2013) ont signalé des concentrations dans les eaux usées qui varient de 1,3 × 104 copies génomiques (CG)/L à 3,6 × 107 CG/L pour une concentration moyenne de 2,0 × 106 GC/L. Au cours d'une récente étude canadienne (Qiu et coll., 2015), les chercheurs ont analysé la présence de multiples virus entériques de source humaine dans tout le procédé de traitement des eaux usées : les concentrations moyennes dans les eaux usées non traitées ont varié de 46 à 70 copies d'équivalent génomique/L pour les entérovirus et les adénovirus respectivement. Même si les concentrations virales diminuent considérablement dans tout le procédé de traitement des eaux usées, on a quand même repéré des virus dans les rejets (Qui et coll., 2015). Ces constatations concordent avec celles d'autres chercheurs (Sedmak et coll., 2005; He et coll., 2011; Li et coll., 2011; Simmons et coll., 2011; Edge et coll., 2013; Hata et coll., 2013; Kitajima et coll., 2014; Kiulia et coll., 2015) et mettent en évidence le rôle que les rejets d'eaux usées peuvent jouer dans la contamination des eaux de surface.
Les virus entériques d'origine humaine peuvent aussi survivre au traitement dans les fosses septiques (Hain et O'Brien, 1979; Vaughn et coll., 1983). Scandura et Sobsey (1997) ont ensemencé des entérovirus dans quatre fosses septiques situées dans des sols sablonneux. On a détecté la présence de virus dans les eaux souterraines moins d'un jour après l'ensemencement et leur présence a persisté jusqu'à 59 jours (la plus longue période d'étude). Les concentrations ont varié de 8 à 908 unités formant plage/L. Les auteurs ont signalé une diminution des virus pouvant atteindre 9 log dans des conditions optimales (non précisées) et une contamination importante causée par les eaux usées dans les systèmes à sable à gros grains et à nappe phréatique élevée. Borchardt et coll. (2011) ont mesuré la concentration de norovirus dans les effluents de fosse septique (79 600 CG/L) et dans l'eau du robinet (34 à 70 CG/L) au cours d'une enquête sur une éclosion survenue à un restaurant. La fosse septique et le puits du restaurant étaient tous deux conformes aux codes du bâtiment de l'État, mais situés dans un environnement hydrogéologique très vulnérable (c.-à-d. aquifère de dolomite fracturé). Des essais au colorant ont confirmé que l'effluent de la fosse septique passait de la fosse (à cause d'un accessoire qui fuyait) et du champ d'infiltration au puits en six et 15 jours respectivement. Bremer et Harter (2012) ont procédé à une analyse probabiliste pour évaluer les effets de fosses septiques sur des puits privés. On a calculé que la probabilité que des puits fussent rechargés par des effluents de fosse septique variait de 0,6 % dans le cas des parcelles importantes (c.-à-d. 20 acres) à faible conductivité hydraulique à presque 100 % dans le cas des petites parcelles (c.-à-d. 0,5 acre) à conductivité hydraulique élevée. Dans le cas des terrains d'une acre, la probabilité a varié de 40 à 75 % pour des conductivités hydrauliques faibles à moyennes respectivement. Kozuskanich et coll. (2014) ont évalué la vulnérabilité d'un aquifère du substratum rocheux à la pollution produite par les fosses septiques dans un village de 500 personnes comptant sur les services locaux et ils ont constaté que la contamination des eaux souterraines par des eaux usées était omniprésente. Morrissey et coll. (2015) ont signalé que l'épaisseur du sol sous le champ d'infiltration de la fosse septique est un facteur critique qui joue sur la contamination des eaux souterraines.
Plusieurs études d'occurrence ont révélé la présence de virus entériques dans toutes sortes d'approvisionnements en eau comptant sur les services locaux (c.-à-d. puits et fosses septiques privés et semi-publics) (Banks et coll., 2001; Banks et Battigelli, 2002; Lindsey et coll., 2002; Borchardt et coll., 2003; Francy et coll., 2004; Trimper, 2010; allen, 2013). Banks et coll. (2001) ont analysé 27 sources d'approvisionnement en eau semi-publics dans un aquifère sablonneux semi-confiné et ont détecté la présence de virus dans trois puits (11 %). Banks et Battigelli (2002) ont signalé la présence de virus dans une source d'approvisionnement en eau semi-public sur 90 située dans un aquifère de roche cristalline confiné en utilisant des méthodes moléculaires. Aucun puits n'a produit de résultats positifs aux analyses par culture cellulaire. Lindsey et coll. (2002) ont analysé 59 sources d'approvisionnement en eau semi-publics dans divers aquifères du substratum rocheux non confinés (54 puits) et aquifères de sable et gravier non confinés (5 puits) : ils ont détecté de virus entériques dans cinq puits (8 %) en utilisant des méthodes de culture cellulaire. Borchardt et coll. (2003) ont analysé 50 puits privés dans sept districts hydrogéologiques, selon la saison, au cours d'une période d'un an. Ils ont détecté, au moyen de méthodes moléculaires, la présence de virus dans quatre puits (8 %) situés à proximité d'une fosse septique : un puits était situé dans un aquifère de sable et de gravier perméable, tandis que les trois autres se trouvaient dans un substratum rocheux fracturé recouvert d'un mort-terrain minime. Francy et coll. (2004) ont analysé 20 puits semi-publics situés dans des aquifères de sable et de gravier non confinés et confinés, dans le sud-est du Michigan, à cinq ou six reprises au cours d'une période de deux ans,. Ils ont repéré la présence de virus entériques dans sept puits (35 %) en utilisant des méthodes moléculaires ou de culture cellulaire. L'étude a aussi inclus le prélèvement d'échantillons dans des secteurs urbains. Les auteurs ont noté que les échantillons contenaient des virus plus souvent aux sites desservis par des fosses septiques qu'aux endroits desservis par des égouts sanitaires.
Trimper (2010) a analysé 23 puits privés forés dans des aquifères du substratum rocheux fracturé recouverts de moins de cinq mètres de mort-terrain dans trois communautés canadiennes (en Colombie-Britannique, en Ontario et à Terre-Neuve) où des fosses septiques locales étaient présentes. Chaque puits a fait l'objet de deux à quatre prélèvements sur une période de 10 mois. La présence de virus a été détectée au moyen des méthodes moléculaires dans 38 %, 67 % et 78 % des puits de chaque communauté. Les distances moyenne et maximale entre les puits et la fosse septique étaient de 32 et de 40 mètres, respectivement. Cette étude a permis de conclure que la contamination virale présente une menace importante pour les aquifères du substratum rocheux fracturé dans les régions rurales dotées de fosses septiques locales. En outre, l'étude indique que puisque les temps de déplacement des virus peuvent être très courts dans les aquifères du substratum rocheux, les marges minimales en pratique n'offrent peut-être pas une protection suffisante (Trimper, 2010; novakowski, 2015). Allen et coll. (2017) ont analysé 11 puits privés situés dans un aquifère du substratum fracturé, recouvert de till glaciaire ou de gravier, une fois par mois pendant huit mois. Dans cinq des onze puits (45 %), des virus ont été détectés par des méthodes moléculaires. La profondeur des puits variait de 24 à 74 mètres, des fosses septiques locales étant situées à environ 15 mètres des puits (distance maximale de 200 mètres). L'échantillonnage mensuel est suggéré pour obtenir une meilleure indication de la vulnérabilité des puits (Cherry et coll., 2006; allen, 2013).
Les fuites d'égouts sanitaires constituent aussi une importante source de virus entériques. On considère que les puits situés dans des secteurs sous lesquels passe un réseau souterrain d'égouts sanitaires sont exposés à un risque accru de contamination virale à cause des fuites des égouts sanitaires (Powell et coll., 2003; allen et coll., 2017). Borchardt et coll. (2004) ont analysé quatre puits municipaux dans un aquifère de sable et de gravier tous les mois de mars 2001 à février 2002 et déterminé que l'on détectait plus souvent la présence de virus entériques dans des puits situés dans des zones sous lesquelles passait un réseau d'égouts sanitaires que dans ceux qui sont situés dans un secteur sans égout sanitaire. Borchardt et coll. (2007) ont signalé des constatations semblables : deux puits municipaux sur trois tirant leur eau d'un aquifère du substratum rocheux confiné ont produit un résultat positif dans sept échantillons sur 20 analysés par des méthodes moléculaires. Les puits positifs pour le virus étaient situés dans une zone urbaine contenant de nombreuses conduites d'égouts proches tandis que le troisième puits était ouvert à des aquifères tant non confinés que confinés, mais n'était pas situé à proximité d'une source de matières fécales humaines; ce puits ne contenait pas de virus tout au long de la période d'étude. Allen et coll. (2017) ont signalé que cinq des huit puits d'approvisionnement en eau public (63 %) situés dans le substratum rocheux fracturé étaient contaminés par des virus à un moment donné durant une période de huit mois d'échantillonnage mensuel. Les puits étaient situés à une distance entre 10 et 100 mètres des égouts sanitaires. Bradbury et coll. (2013) ont signalé un lien temporel entre les sérotypes de virus présents dans les eaux usées et ceux qui se trouvent dans un aquifère confiné, ce qui semble indiquer que le transport entre les égouts et les réseaux d'eau souterraine est très rapide, soit de l'ordre de quelques jours à quelques semaines. Hunt et coll. (2014) ont attribué ce phénomène à des voies préférentielles comme les fractures dans la couche semi-perméable (c.-à-d. l'unité géologique subjacente de faible perméabilité), des puits alimentés par de multiples aquifères et un coulis mal posé dans les puits. L'échantillonnage ultérieur de ces puits dans des conditions de sécheresse a révélé une présence plus faible de virus (Gotkowitz et coll., 2016). Les auteurs ont remarqué que le programme d'échantillonnage précédent, mentionné dans l'étude de Bradbury et coll. (2013), s'était déroulé durant une période historique humide. La détection de virus a été associée à des précipitations (Bradbury et coll., 2013; Gotkowitz et coll., 2016). À l'opposé, allen (2013) a relevé que de faibles précipitations pouvaient entraîner des concentrations plus élevées de virus.
Les animaux peuvent être une source de virus entériques, mais ceux qu'on détecte chez les animaux ne sont généralement pas pathogènes pour les êtres humains, à quelques exceptions près. Comme nous l'avons déja mentionné, le VHE peut avoir un réservoir non humain. Jusqu'à maintenant, le VHE a posé problème dans les pays en développement et, par conséquent, la plupart des renseignements sur la présence du VHE dans des sources d'approvisionnement en eau proviennent d'études réalisées dans ces pays. Il y a peu d'information sur la présence du VHE dans l'eau et les eaux usées des pays industrialisés (Clemente-Casares et coll., 2003; Kasorndorkbua et coll., 2005). Gentry-Shields et coll. (2015) ont signalé récemment la présence de VHE dans un seul échantillon d'eau de surface prélevé à un endroit situé à proximité du champ pulvérisé d'une exploitation intensive d'engraissement d'animaux produisant surtout du porc en Caroline du nord (2015), ce qui laisse entendre qu'il y a peut-être un lien entre ces activités et la propagation du VHE.
5.1.2 Présence dans l'eau
Comme on l'a signalé ci-dessus, les virus entériques peuvent contaminer des sources d'approvisionnement en eau en empruntant toutes sortes de voies. La section qui suit décrit en détail les études qui ont examiné la présence de virus entériques dans les eaux de surface et les eaux souterraines, ainsi que sur l'eau potable. Il importe de signaler que ces données proviennent en grande partie d'études ciblées, puisque l'on n'effectu pas une surveillance régulière des virus entériquesdans les eaux de source ou les eaux potable; et qu'elles ne représentent peut-être pas la situation actuelle. Il importe aussi de tenir compte du fait qu'on a utilisé diverses méthodes de détection (c.-à-d. à base de culture cellulaire, moléculaire) (voir la section 6.0) et que l'on n'a pas toujours évalué l'infectiosité des virus détectés. Étant donné ces méthodes d'études qui varient, il n'est pas facile de comparer les données.
Plusieurs études ont signalé la présence de virus entériques dans les eaux de surface dans le monde entier, y compris au Canada (Sattar, 1978; Sekla et coll., 1980; Payment et coll., 1984, 2000; Raphael et coll., 1985a, 1985 b; Payment, 1989, 1991, 1993; Payment et Franco, 1993; Pina et coll., 1998, 2001; Sedmak et coll., 2005; Van Heerden et coll., 2005; EPCOR, 2010, 2011; Gibson et Schwab, 2011; Edge et coll., 2013; Corsi et coll., 2014; Pang et coll., 2014). Le tableau B.2 de l'annexe B done un aperçu de certaines études qui ont examiné la présence de virus entériques dans des sources d'eau de surface au Canada et aux États-Unis. Les virus entériques semblent très prévalents dans les eaux de surface et leur présence révèle une importante variabilité spatiotemporelle. Cette variabilité indique en grande partie si la source de pollution est continue ou découle d'un apport soudain de contamination fécale (voir la section 5.5). La prévalence des virus dans les eaux de surface dépend aussi de facteurs environnementaux comme l'ensoleillement, la température et la prédation (de Roda Husman et coll., 2004, 2009; Rutjes et coll., 2009; Lodder et coll., 2010) (voir la section 5.2.1).
On a détecté les virus entériques dans tout un éventail de sources d'eaux souterraines, en utilisant des techniques moléculaires ou de culture cellulaire, les taux de prévalence variant de moins de 1 % à 46 % (Abbaszadegan et coll., 1999, 2003; Banks et coll., 2001; Banks et Battigelli, 2002; Lindsey et coll., 2002; Borchardt et coll., 2003; Fout et coll., 2003; Francy et coll., 2004; Locas et coll., 2007, 2008; Hunt et coll., 2010; Gibson et Schwab, 2011; Borchardt et coll., 2012; allen, 2013; Bradbury et coll., 2013; Pang et coll., 2014). Le tableau B.3 de l'annexe B done un aperçu de certaines études qui ont examiné la présence de virus entériques dans des sources d'eaux souterraines du Canada et des États-Unis. On a détecté les virus dans différents types d'aquifères, y compris des puits semi-publics dans un aquifère sablonneux semi-confiné (Banks et coll., 2001) et un aquifère de roche cristalline confiné (Banks et Battigelli, 2002), ainsi que dans des puits municipaux profonds (220 à 300 m) dans un aquifère de grès et de dolomite confiné (Borchardt et coll., 2007; Bradbury et coll., 2013). Bradbury et coll. (2013) ont signalé que les concentrations de virus dans les puits municipaux profonds étaient en général aussi élevées sinon plus élevées que les concentrations de virus dans l'eau de lac. En général, il est possible de caractériser la présence de virus dans les eaux souterraines comme transitoire, intermittente ou éphémère, parce qu'il arrive souvent que les puits ne contiennent pas de virus dans deux échantillons séquentiels prélevés et la fréquence de détection est faible selon le nombre d'échantillons (Borchardt et coll., 2003; allen, 2013, 2017). Toutefois, la fréquence de détection des virus, par puits individuel, varie de 30 à 100 % (tableau B.3) et englobe un large éventail de types d'aquifères, ce qui indique que la vulnérabilité globale des puits à la contamination virale est élevée (Fout et coll., 2003; Francy et coll., 2004; Borchardt et coll., 2007; allen et coll., 2017).
Des études ciblées réalisées aux États-Unis ont permis de détecter la présence de virus entériques dans l'eau potable, y compris des eaux souterraines désinfectées aux rayons UV (Borchardt et coll., 2012; Lambertini et coll., 2011). Le tableau B.4 de l'annexe B présente certaines de ces études. Borchardt et coll. (2012) ont fait état de la présence de virus entériques dans presque 25 % des 1 200 échantillons d'eau du robinet provenant de 14 communautés comptant sur des eaux souterraines non traitées. L'adénovirus était le plus prévalent (157/1 204) des virus détectés, même si l'on en a constaté la présence à des concentrations d'un ou deux ordres de grandeur de moins que celles du norovirus et de l'entérovirus. L'entérovirus est celui qui présentait la concentration la plus élevée, qui atteignait en moyenne et au maximum 0,8 CG/L et 851 CG/L respectivement. Les auteurs ont pu montrer l'existence d'un lien entre la concentration moyenne de tous les virus et la présence des maladies gastro-intestinale aiguë (MGA) dans la communauté (voir la section 5.4.1). Au cours d'une étude d'accompagnement, Lambertini et coll. (2011) ont démontré que les virus entériques peuvent pénétrer dans des réseaux de distribution à la suite d'événements courants (p. ex. installation de tuyaux). On a détecté des entérovirus, norovirus GI et GII, adénovirus, rotavirus et des virus de l'hépatite A à la tête de puits après la désinfection aux rayons UV (dose minimale = 50 mJ/cm2) et aux robinets domestiques alimentés par des réseaux de distribution sans résiduel de désinfectant. On a détecté les virus dans 10,1 % des échantillons désinfectés aux rayons UV (95e centile de la concentration de virus ≤ 1,1 CG/L). Par ailleurs, on a détecté les virus dans 20,3 % des échantillons prélevés après la désinfection aux rayons UV (95e centile de concentration de virus ≤ 8,0 CG/L). Cette augmentation de la détection et de la concentration entre la désinfection aux rayons UV et les robinets domestiques a été attribuée au fait que les virus pénètrent directement dans le réseau de distribution (voir la section 5.4.1). Des études antérieures réalisées au Canada n'ont pas permis de détecter la présence de virus entériques dans l'eau traitée (Payment et Franco, 1993; Payment et coll., 1984).
5.2 Survie
Comme on l'a déjà mentionné, les virus ne peuvent se reproduire que dans une cellule hôtesse (c.-à-d. elles ne peuvent pas se reproduire dans l'environnement). Ils peuvent toutefois survivre pour de longues périodes (c.-à-d. de 2 à 3 ans; Banks et coll., 2001; Cherry et coll., 2006) et être transportés sur de longues distances (Keswick et Gerba, 1980; Pang, 2009; Hunt et Johnson, 2017).
La survie des virus dépend du temps qu'il leur faut pour perdre leur capacité d'infecter des cellules hôtesses (c.-à-d. phénomène d'inactivation) et de la vitesse à laquelle un virus se fixe en permanence aux particules de sol ou y sont adsorbés (Gerba, 1984; Yates et coll., 1985, 1987, 1990; Yates et Yates, 1988; Bales et coll., 1989, 1991, 1997; Schijven et Hassanizadeh, 2000; John et Rose, 2005). Les deux processus (inactivation et adsorption) sont propres à chaque type de virus (Goyal et Gerba, 1979; Sobsey et coll., 1986) et généralement indépendants l'un de l'autre (Yates et coll., 1987; Schijven et Hassanizadeh, 2000; John et Rose, 2005). Même si l'on sait que les concentrations de virus diminuent dans l'environnement, les phénomènes d'inactivation et d'adsorption sont très complexes et ne sont pas bien compris (Schijven et Hassanizadeh, 2000; azadpour-Keeley et coll., 2003; Gordon et Toze, 2003; Johnson et coll., 2011a; Hunt et coll., 2014; Bellou et coll., 2015). La grande raison de ce manque de compréhension réside dans le fait que les virus sont des particules colloïdales qui peuvent se déplacer comme particules en suspension indépendante ou en se fixant à d'autres particules colloïdales non vivantes comme l'argile ou à des macromolécules organiques (Robertson et Edberg, 1997). Il y a une autre difficulté : les vitesses de dégradation ne sont pas toujours linéaires (Pang, 2009). On a observé que la vitesse de dégradation des virus les plus résistants ralentit avec le temps (Page et coll., 2010).
Dans beaucoup des études d'évaluation des vitesses d'inactivation ou des caractéristiques d'adsorption des virus, y compris celles dont il est question ci-dessous, les chercheurs ont utilisé des substituts. Les substituts peuvent être un organisme, une particule ou une substance qu'on utilise pour étudier le sort d'un microorganisme pathogène dans un environnement naturel (c.-à-d. phénomènes d'inactivation ou d'adsorption) ou dans un environnement de traitement (c.-à-d. filtration ou désinfection) (Sinclair et coll., 2012). On a suggéré d'utiliser des bactériophages ou des coliphages comme substituts des virus (Stetlar, 1984; Havelaar, 1987; Payment et Franco, 1993). Le bactériophage PRD1 et le coliphage MS2, par exemple, ressemblent au rotavirus et au poliovirusNote de bas de page 1 respectivement par la forme et la grosseur (Azadpour-Keeley et coll., 2003). Les deux survivent longtemps et ont une faible tendance à l'adsorption (Yates et coll., 1985). Par contre, LeClerc et coll. (2000) ont signalé de nombreuses lacunes que présente l'utilisation de bactériophages ou de coliphages comme substituts de virus. Comme l'inactivation et l'adsorption varient considérablement selon le type de virus, on reconnaît généralement qu'aucun virus ou substitut ne peut servir à lui seul à décrire les caractéristiques de tous les virus entériques. Les méthodes moléculaires ou de culture cellulaire (voir la section 6.1.2) causent aussi de la confusion dans l'interprétation des résultats (de Roda Husman et coll., 2009). Une solution consiste à utiliser un éventail de microorganismes tirés d'eaux usées (Schijven et Hassanizadeh, 2000). Sinclair et coll. (2012) décrivent un processus de sélection de substituts représentatifs de systèmes naturels ou d'origine humaine.
5.2.1 Inactivation dans l'environnement
Les virus sont inactivés par des perturbations des protéines de leur enveloppe et la dégradation de leurs acides nucléiques. Des analyses critiques des facteurs qui jouent sur l'inactivation des virus indiquent que les facteurs les plus importants sont la température, l'adsorption à des particules et l'activité microbienne (Schijven et Hassanizadeh, 2000; Gordon et Toze, 2003; John et Rose, 2005).
L'inactivation des virus augmente en général à mesure que la température monte, mais cette tendance se produit principalement à des températures de plus de 20 °C (John et Rose, 2005). Le poliovirus incubé dans un milieu de conservation a été réduit de 2 log après 1 022 jours à 4 °C par rapport à 4 log après 200 jours à 22 °C (de Roda Husman et coll., 2009). Des expériences menées en laboratoire ont démontré l'infectiosité à long terme de certains virus dans des eaux souterraines stockées dans l'obscurité : rotavirus, jusqu'à sept mois (période d'analyse la plus longue) et astrovirus humain, au moins 120 jours, gardés dans les deux cas à 15 °C (Espinosa et coll., 2008); poliovirus et virus Coxsackie, au moins 350 jours à 4 °C (de Rosa Husman et coll., 2009); adénovirus, 364 jours à 12 °C (Charles et coll., 2009) et norovirus, au moins 61 jours à 12 °C (Seitz et coll., 2011). Il est possible de détecter des génomes de virus pendant des périodes beaucoup plus longues, soit : au moins 672 jours dans le cas de l'adénovirus gardé à 12 °C (Charles et coll., 2009) et au moins 1 266 jours dans celui du norovirus conservé à la température ambiante (Seitz et coll., 2011).
Gerba (1984) a signalé que les virus associés à des particules persistent en général plus longtemps, parce que la particule offre une protection contre les enzymes protéolytiques ou d'autres substances pouvant inactiver les virus. Cet effet dépend du type de virus et de la nature de la particule. Les particules d'argile sont particulièrement efficaces pour protéger les virus contre la décroissance naturelle en raison de leur forte capacité d'adsorption (Carlson et coll., 1968; Sobsey et coll., 1986). On signale que certains types de matières organiques (c.-à-d. protéines) protègent mieux les virus contre l'inactivation (Gordon et Toze, 2003).
Herrmann et coll. (1974) ont signalé que les virus sont inactivés plus rapidement en présence d'une microflore indigène, en particulier de bactéries protéolytiques comme Pseudomonas aeruginosa. Les auteurs ont observé une diminution de cinq log de la concentration de virus Coxsackie et de poliovirus dans un lac naturel après neuf et 21 jours respectivement. Par contre, on a observé une diminution de moins de deux log dans le cas des deux virus, pendant la même période, dans de l'eau de lac stérile. Gordon et Toze (2003) ont constaté que la présence de microflore indigène constituait la principale raison de l'inactivation du virus dans les eaux souterraines. On n'a observé aucune décroissance du poliovirus dans des eaux souterraines stériles à 15 °C, tandis que l'on en a constaté une d'un log après cinq jours dans des eaux souterraines non stériles. Dans le cas du virus Coxsackie, on a observé une diminution d'un log après 528 et 10,5 jours dans des eaux souterraines stériles et non stériles, respectivement.
Les virus ont tendance à survivre plus longtemps dans les eaux souterraines parce que les températures y sont plus basses, qu'ils y sont protégés contre les rayons du soleil et que l'activité microbienne est moindre (Keswick et coll., 1982; John et Rose, 2005). Banks et coll. (2001) indiquent que la survie des virus dans les eaux souterraines est estimée de façon prudente à trois ans, tandis que Cherry et coll. (2006) indiquent qu'une estimation raisonnable s'établit à un à deux ans. Hunt et coll. (2014) affirment que la présence de génomes de virus dans les eaux souterraines démontre des temps de déplacement dans les aquifères de deux à trois ans entre la source de contamination fécale et le puits. On a observé que les concentrations de virus dans les eaux de surface varient selon les saisons, les concentrations les plus élevées se présentant à des températures plus basses. Schijven et coll. (2013) laissent entendre que le phénomène peut être lié à la diminution de l'activité biologique attribuable à la température plus basse.
5.2.2 Adsorption et migration
De nombreuses recherches ont porté sur les mécanismes du phénomène d'adsorption (Carlson et coll., 1968; Bitton, 1975; Duboise et coll., 1976; Goyal et Gerba, 1979; Keswick et Gerba, 1980; Gerba et coll., 1981; Vaughn et coll., 1981; Gerba, 1984; Gerba et Bitton, 1984; Yates et coll., 1987; Yates et Yates, 1988; Bales et coll., 1989, 1991, 1995, 1997; Powelson et coll., 1991; Rossi et coll., 1994; Song et Elimelech, 1994; Loveland et coll., 1996, Pieper et coll., 1997; Sinton et coll., 1997; DeBorde et coll., 1998, 1999; Ryan et coll., 1999; Schijven et coll., 1999, 2002; Schijven et Hassanizadeh, 2000; Woessner et coll., 2001; Borchardt et coll., 2004; Pang et coll., 2005; Michen et Graule, 2010; Bradbury et coll., 2013; Harvey et coll., 2015). Le processus d'adsorption dans des environnements souterrains est contrôlé principalement par les interactions électrostatiques et hydrophobes (Bitton, 1975; Gerba, 1984). Les caractéristiques hydrologiques de l'aquifère, les caractéristiques de surface du virus en fonction de la chimie de l'eau et les propriétés physiques et chimiques des particules de sol individuelles (DeBorde et coll., 1999) sont tous des facteurs qui jouent un rôle dans la dynamique de l'adsorption.
Un pH bas et une force ionique élevée, conditions qui réduisent les forces de répulsion électrostatique entre le virus et la particule de sol, favorisent en général l'adsorption des virus (Bitton, 1975; Duboise et coll., 1976; Gerba, 1984). Les phases minérales à charge positive (p. ex. oxydes de fer, d'aluminium ou de manganèse) favorisent l'adsorption des virus parce que la plupart portent une charge négative dans les eaux naturelles (Bitton, 1975; Goyal et Gerba, 1979; Keswick et Gerba, 1980). Les particules d'argile offrent aussi des sites de liaison à forte charge positive qui augmentent considérablement la surface disponible pour l'adsorption du virus (Carlson et coll., 1968). Par contre, les sols argileux risquent de se contracter et de se fissurer, ce qui permet l'apparition de fractures qui favorisent le transport rapide des virus (Pang, 2009). On croit que la présence de matières organiques est à l'origine d'un grand nombre des incertitudes qui règnent au sujet du processus d'adsorption (Schijven et Hassanizadeh, 2000). La matière organique peut à la fois perturber les interactions hydrophobes et fournir des sites d'adsorption hydrophobes : tout dépend de la combinaison du sol et du type de virus (Gerba, 1984; Schijven et Hassanizadeh, 2000). Les substances humiques, par exemple, portent une charge négative comme les virus et leur font donc concurrence pour les mêmes sites d'adsorption (Powelson et coll., 1991; Pieper et coll., 1997).
Les conditions hydrauliques jouent aussi un rôle important dans l'adsorption des virus (Berger, 1994; azadpour-Keeley et coll., 2003). L'eau souterraine doit s'écouler assez lentement pour permettre au virus d'entrer en contact avec la particule de sol et d'y adhérer. Sinon, le virus reste dans l'eau et est transporté vers l'aval. Plusieurs chercheurs ont signalé que les virus peuvent parcourir des distances importantes en peu de temps, en empruntant des voies privilégiées, à cause de l'exclusion liée aux dimensions des pores. Ce phénomène signifie que des particules, comme les virus, sont transportées plus rapidement que la vitesse moyenne des eaux souterraines parce qu'elles doivent traverser des pores de plus grandes dimensions où les vitesses sont plus élevées (Bales et coll. 1989; Sinton et coll., 1997; Berger, 1994; DeBorde et coll., 1999; Cherry et coll., 2006; Bradbury et coll., 2013; Hunt et coll., 2014). Des vitesses d'écoulement de l'eau souterraine de l'ordre de 1 à 400 m/jour ont été signalées dans des aquifères du substratum rocheux au Canada (Novakowski et coll., 2006a; Belan, 2010; Praamsma, 2017; Persaud et coll., 2018). Les conditions de pompage des puits peuvent aussi créer d'importants gradients hydrauliques et des vitesses élevées dans les eaux souterraines. Le faible volume d'eau qui peut être libéré du substratum rocheux (entreposage particulier) créé un cône de rabattement qui s'étend beaucoup plus loin dans le cas des puits de substratum rocheux que ceux dans les aquifères en milieu poreux. Ces facteurs pourraient mener au transport rapide et étendu de virus dans les aquifères du substratum rocheux fracturé (Allen et coll., 2017). Ce phénomène est mis en évidence par Bradbury et coll. (2013), qui ont signalé un transport de virus de l'ordre de quelques semaines à partir d'une source de contamination jusqu'à des puits municipaux dont la profondeur variait de 220 à 300 mètres. En utilisant un essai colorimétrique, Levison et novakowski (2012) ont trouvé que dans les puits situés dans un substratum rocheux fracturé recouvert d'une couche minime de mort-terrain, la pénétration du soluté avait lieu en moins de 4 heures à des profondeurs variant entre 19 et 35 mètres. Cela démontre que le transport de soluté, et par conséquent, de virus, est rapide dans un substratum rocheux fracturé recouvert d'une couche minime de mort-terrain.
En général, le processus d'adsorption n'inactive pas les virus et l'adsorption est réversible (Carlson et coll., 1968; Bitton, 1975). Comme les interactions entre les virus et le sol sont très sensibles à la charge superficielle, tout changement de la qualité de l'eau qui suffit pour causer une inversion de charge entraînera la désorption de virus qui peuvent être infectieux (Song et Elimelech, 1994; Pieper et coll., 1997). Les changements de la qualité de l'eau qui peuvent provoquer une désorption comprennent une augmentation du pH, une diminution de la force ionique et la présence de matières organiques en quantités suffisantes (Carlson et coll., 1968; Duboise et coll., 1976; Bales et coll., 1993; Loveland et coll., 1996). Par exemple, lorsqu'un effluent septique alcalin se mélange à des eaux souterraines, l'élévation du pH permet le transport rapide des virus, particulièrement lorsque le débit est sous-saturé (Scandura et Sobsey, 1997). L'apport de pluie après une tempête peut réduire la force ionique, entraîner la désorption de virus et leur transport d'amont en aval. Les virus infectieux résorbés peuvent ainsi continuer à contaminer les sources d'approvisionnement en eau longtemps après la contamination initiale (Sobsey et coll., 1986; DeBorde et coll., 1999) (voir la section 5.5). La matière organique réduit la capacité des milieux souterrains d'adsorber des microorganismes pathogènes par fixation aux sites d'adsorption disponibles, ce qui bloque l'adsorption des microorganismes pathogènes. (Pang, 2009).
Les documents publiés signalent une plage importante de distances de transport de virus (U.S. EPA, 2006d). On a signalé des distances de transport d'environ 400 m dans le cas d'aquifères de sable et de gravier tandis qu'on a observé la distance la plus éloignée (1 600 m) dans une formation karstique. Les puits d'approvisionnement en eau s'alimentant dans des aquifères karstiques et de substratum rocheux fracturés sont très vulnérables à la contamination, parce que le débit des eaux souterraines et le transport des microorganismes pathogènes peuvent être extrêmement rapides, de l'ordre de quelques heures (Amundson et coll., 1988; Scandura et Sobsey, 1997; Powell et coll., 2003; Borchardt et coll., 2011; Levison et novakowski, 2012; Kozuskanich et coll., 2014). Il ne faut pas gérer les sources d'eaux souterraines dans les formations karstiques et de substratum rocheux fracturé de la même façon que les aquifères de sable et de gravier (Crowe et coll., 2003; novakowski, 2015).
5.3 Exposition
Les virus entériques sont transmis par la voie fécale-orale. Les vecteurs peuvent comprendre l'eau, les aliments (en particulier les crustacés et les salades), les vecteurs passifs (objets inanimés comme les poignées de porte qui, une fois contaminés par un virion infectieux, facilitent le transfert du virus pathogène à un hôte) et le contact de personne à personne. Les virus entériques peuvent aussi se propager par les aérosols. Le norovirus, par exemple, devient aérosolisé au cours de vomissements qui peuvent entraîner le rejet de jusqu'à 30 millions de virus au cours d'un seul épisode de vomissements (Caul, 1994; Marks et coll., 2000; Marks et coll., 2003; Lopman et coll., 2012; Tung-Thompson et coll., 2015). Une mauvaise hygiène est également un facteur qui contribue à la propagation des virus entériques. De plus, l'incidence élevée d'infections à rotavirus, surtout chez les jeunes enfants, porte certains chercheurs à croire que les rotavirus peuvent aussi se propager par la voie respiratoire (Kapikian et Chanock, 1996; Chin, 2000). Pour beaucoup des virus entériques susmentionnés, des éclosions sont survenues à la fois par transmission de personne à personne et par des sources communes, soit des aliments contaminés, des approvisionnements en eau potable contaminée ou des eaux employées à des fins récréatives.
5.4 Maladies d'origine hydrique
Comme on le signale à la section 4.0, certains sérotypes ou génotypes de virus entériques sont associés le plus couramment à des maladies humaines. Dans le cas des norovirus, on associe les génogroupes GI, GII et GIV aux maladies humaines et les infections atteignent habituellement leur pic culminant au cours de l'hiver. Le rotavirus du groupe a est endémique dans le monde entier et constitue le groupe le plus courant et le plus répandu. Le rotavirus du groupe B existe principalement en Chine. On a établi un lien entre celui du groupe C et des cas rares et sporadiques de diarrhée chez les enfants dans beaucoup de pays, y compris en amérique du nord (Jiang et coll., 1995). Dans le cas des entérovirus, on a identifié plusieurs types infectieux parmi les maladies humaines. Les infections à entérovirus atteindraient leur pic au cours de l'été et au début de l'automne (Nwachuku et Gerba, 2006; Pallansch et Roos, 2007). Les sérotypes 40 et 41 causent la majorité des gastroentérites liées à des adénovirus. On associe les deux génogroupes A et B d'astrovirus aux maladies humaines (Carter, 2005) et les infections atteignent leur point culminant à l'hiver et au printemps.
L'exposition aux virus entériques par l'eau peut produire à la fois un taux endémique de maladies dans la population et des éclosions de maladies d'origine hydrique.
5.4.1 Maladies endémiques
Pour diverses raisons, les taux de maladies infectieuses endémiques, notamment les maladies d'origine hydrique, sont nettement sous-déclarés et sous-diagnostiqués (Majowicz et coll., 2004; MacDougall et coll., 2008; Gibbons et coll., 2014). Il est encore plus compliqué de déterminer l'incidence « réelle » des maladies endémiques d'origine hydrique du fait que, au Canada, il n'existe aucun système de surveillance national propre à ces maladies (Pons et coll., 2015). Plusieurs groupes (Payment et coll., 1991, 1997; Hellard et coll., 2001; Colford et coll., 2005, 2006, 2009; Messner et coll., 2006) ont tenté d'estimer le fardeau de la maladie associé à l'eau, y compris l'eau potable. L'estimation du fardeau des maladies au Canada est présentée ici. Les statistiques mentionnées ci-dessous sont des estimations mathématiques calculées à l'aide de modèles d'évaluation des risques et ne reflètent pas la véritable incidence.
Le fardeau annuel des maladies gastro-intestinale aiguë (MGA) endémiques au Canada provenant de toutes les sources d'approvisionnement en eau (c.-à-d. aliments, eau, animaux, de personne à personne) est estimé à 20,5 millions de cas (0,63 cas/année-personne) (Thomas et coll., 2013). On estime qu'environ 1,7 % (334 966) de ces cas, soit 0,015 cas/année-personne, sont associés à la consommation d'eau du robinet des réseaux municipaux qui desservent plus de 1 000 personnes au Canada (Murphy et coll., 2016a). Plus de 29 millions (84 %) de Canadiens comptent sur de tels réseaux, 73 % (environ 25 millions) comptant sur une source d'eau de surface, 1 % (0,4 million) sur une source d'eau souterraine soumise à l'influence directe des eaux de surface (ESSIDES) et les 10 % restants (3,3 millions), sur une source d'eau souterraine (Statistique Canada, 2013a, 2013 b). Murphy et coll. (2016a) ont calculé que les systèmes comptant sur des sources d'eau de surface traitée seulement au chlore ou au dioxyde de chlore, ou sur des sources ESSIDES non traitées ou traitées au minimum, ou sur des sources d'eau souterraine non traitée, causent la majeure partie du fardeau de MGA (c.-à-d. 50 121 des cas estimés ou 0,047 cas/année-personne) associées aux réseaux municipaux qui desservent plus de 1 000 personnes. Par contre, on a établi un lien entre quelque 0,007 cas/année-personne (ou 15 991 cas estimés) et des systèmes comptant sur des sources d'approvisionnement en eau subissant peu d'effet et dotées de multiples procédés de traitement. Les auteurs ont aussi calculé que plus de 35 % (ou 122 608 cas estimés) des 335 000 cas de MGA étaient attribuables au réseau de distribution.
On calcule que 103 230 (0,51 %) cas de MGA par année, soit 0,003 cas/année-personne, sont attribuables à la présence d'agents microbien pathogènes dans l'eau potable provenant de réseaux privés et de petits réseaux communautaires d'alimentation en eau au Canada (Murphy et coll., 2016b). Les puits privés sont à l'origine de plus de 75 % (environ 78 073) de ces cas estimatifs, ce qui représente 0,027 cas/année-personne. Les systèmes d'eau de petites collectivités qui sont alimentés par des eaux souterraines ont causé 13 034 cas supplémentaires de MGA par année, l'incidence la plus élevée, soit 0,027 cas/année-personne, se produisant dans des réseaux où l'eau n'est pas traitée. On a par contre attribué à des systèmes d'eau de petites collectivités alimentés par des sources d'eaux de surface 12 123 cas annuels estimatifs de MGA, l'incidence la plus élevée, soit 0,098 cas/année-personne, étant signalée dans le cas des systèmes sans traitement. Les auteurs ont calculé que ces cas prédits de MGA sont attribués en majorité au norovirus. Plus précisément, sur les 78 073 cas calculés de MGA/année causés par la consommation d'eau potable provenant de puits privés non traités au Canada, on estime que le norovirus a causé plus de 70 % des cas symptomatiques (c.-à-d. 55 500). Tout comme dans le cas des puits privés, le norovirus a causé la grande majorité des cas calculés associés à la consommation d'eau potable provenant de systèmes d'eau de petites collectivités, ce qui représente 83 % (10 869) des cas calculés, ainsi que de systèmes d'eau de petites collectivités alimentés par des eaux de surface (9 003 cas, > 74 %). Au total, ces estimations indiquent que les Canadiens desservis par des systèmes d'eau de petites collectivités alimentés en eaux de surface non traitées ou insuffisamment traitées risquent davantage d'être exposés à des agents microbiens pathogènes, en particulier au norovirus, et risquent donc davantage d'être victimes de MGA d'origine hydrique.
Des études ont révélé la présence de virus entériques dans toutes sortes de sources d'eaux souterraines (voir la section 5.1.2), mais on sait toutefois peu de choses au sujet de la contribution de ces virus aux maladies d'origine hydrique dans la communauté. Borchardt et coll., (2012) ont estimé l'incidence de MGA dans 14 communautés, servant de 1 300 à 8 300 personnes, alimentées par des eaux souterraines non traitées. On a analysé des échantillons d'eau du robinet pour détecter la présence d'adénovirus, d'entérovirus et de norovirus (voir la section 5.1.2), et les ménages ont consigné les symptômes de MGA dans des journaux de santé. On a déclaré plus de 1 800 épisodes d'affections MGA et 394 057 jours-personne de suivi. L'incidence de MGA pour tous les âges s'est établie à 1,71 épisode/année-personne et elle a été la plus élevée chez les enfants de ≤ 5 ans (2,66 épisodes/année-personne). Borchardt et coll. (2012) ont déterminé qu'il y avait un lien entre trois mesures sommaires de contamination virale et l'incidence de MGA, soit la concentration moyenne, la concentration maximale et le pourcentage d'échantillons positifs. Ces liens étaient particulièrement solides dans le cas du norovirus et l'on n'a pas établi de lien entre l'exposition à l'adénovirus et des cas de MGA. Afin d'essayer de caractériser encore davantage le lien entre la présence de virus et les affections entériques, les auteurs ont calculé la fraction de MGA attribuables aux virus présents dans l'eau du robinet des communautés en se basant sur l'évaluation quantitative du risque microbien (ÉQRM) (voir la section 8.0). Ils ont déterminé que de 6 à 22 % des cas deMGA dans ces communautés étaient attribuables aux virus entériques (Borchardt et coll., 2012).
Lambertini et coll. (2011, 2012) ont estimé le risque de MGA causées par la contamination virale du réseau de distribution dans les 14 réseaux municipaux d'approvisionnement en eaux souterraines étudiés par Borchardt et coll. (2012). Au cours de leur étude, les chercheurs ont mis en œuvre la désinfection aux rayons UV (sans résiduel de chlore). Les virus entériques ont été dénombrés à la tête de puits, après la désinfection aux rayons UV (dose minimale = 50 mJ/cm2) et aux robinets domestiques. Les auteurs ont observé une augmentation de la détection et de la concentration des virus entre la désinfection aux rayons UV et le robinet domestique et ils ont attribué cette constatation aux virus pénétrant dans le réseau de distribution. Ils ont estimé le risque de MGA à cause de la contamination du réseau de distribution et ils ont déterminé qu'il variait de 0,0180 à 0,0611 épisode/année-personne.
5.4.2 Éclosions
Des éclosions d'origine hydrique causées par les virus entériques ont été signalées au Canada et dans le monde entier (Hafliger et coll. 2000; Boccia et coll. 2002; Parshionikar et coll. 2003; Hoebe et coll. 2004; nygard et coll. 2004; Kim et coll. 2005; Yoder et coll. 2008; Larsson et coll. 2014; Kauppinen et coll., 2017; Murphy et coll., 2017). Certaines de ces éclosions sont décrites en détail au tableau B.5 de l'annexe B. On ne connaît pas la prévalence réelle des éclosions, principalement à cause de la sous-déclaration et du sous-diagnostic. Des systèmes intégrés de surveillance des éclosions et d'intervention sont essentiels à la compréhension de ces éclosions.
Le norovirus est un des virus entériques les plus souvent signalés en amérique du nord et dans le monde entier. Au Canada, les éclosions de norovirus sont à déclaration obligatoire à l'échelle nationale depuis 2007. (ASPC, 2015a). Il n'y a toutefois pas de renseignements sur les sources et c'est pourquoi on ne sait pas trop combien de cas déclarés sont attribuables à l'eau.
Au Canada, entre 1974 et 2001, on a signalé 24 éclosions et 1 382 cas confirmés de maladies d'origine hydrique causées par des virus entériques (Schuster et coll., 2005). Dix de ces éclosions ont été attribuées au VHA, 12 à des norovirus et deux à des rotavirus (O'Neil et coll., 1985; Santé et Bien-être social Canada, 1990; Santé Canada, 1994, 1996; INSPQ, 1994, 1998, 2001; Boettger, 1995; Beller et coll., 1997; De Serres et coll., 1999; Todd, 1974-2001; BC Provincial Health Officer, 2001). Il y a aussi eu 138 éclosions d'étiologie inconnue, dont une partie pourrait être attribuable à des virus entériques, ainsi qu'une seule éclosion mettant en cause de multiples virus pathogènes. Sur les dix éclosions attribuées au VHA d'origine hydrique, quatre ont été causées par la contamination de sources publics d'approvisionnement en eau potable, deux par la contamination de sources d'approvisionnement en eau semi-publicsNote de bas de page 2 et les quatre autres, par la contamination d'approvisionnements en eau privés. Seulement quatre des 12 éclosions d'origine hydrique déclarées attribuables à des infections à norovirus au Canada sont survenues dans des approvisionnements en eau publics et les autres ont été attribuées à des approvisionnements en eau semi-publics. Les deux éclosions à rotavirus ont découlé de la contamination d'approvisionnements en eau potable semi-publics. La contamination des sources d'approvisionnement en eau par les eaux usées, et un échec de traitement (p. ex. eaux de surface non filtrées ou mal filtrées, utilisation exclusive du chlore) constitue un des principaux facteurs qui expliquent ces éclosions (Schuster et coll., 2005). Des événements météorologiques ont eu tendance à exacerber ces problèmes. Les répercussions de ces événements météorologiques sur la contamination virale sont abordées dans la section 5.5. Aucune éclosion virale d'origine hydrique n'a été déclarée au Canada depuis 2001.
Aux États-Unis, entre 1991 et 2002, on a signalé 15 éclosions et 3 487 cas confirmés de maladies virales d'origine hydrique, dont 12 éclosions et 3 361 cas ont été attribués à des norovirus, une éclosion et 70 cas, à un « petit virus rond », et deux éclosions et 56 cas, au VHA (Craun et coll., 2006). Pendant cette période, on a déclaré aussi 77 éclosions représentant 16 036 cas d'étiologie inconnue. Des virus entériques ont probablement causé un pourcentage important de ces éclosions (Craun et coll., 2006). Entre 2003 et 2012, les Centers for Disease Control and Prevention (CDC) des États-Unis ont déclaré 138 éclosions de maladies infectieuses liées à la consommation d'eau potable (Blackburn et coll., 2004; Liang et coll., 2006; Yoder et coll., 2008; Brunkard et coll., 2011; CDC, 2013 b, 2015d) qui ont causé 8 142 cas de maladie. Les virus entériques ont été impliqués comme la seule cause dans 13 (9,4 %) de ces éclosions, causant 743 cas de maladie. Le norovirus a causé 10 éclosions (sur 13) tandis que le VHA a été impliqué dans les autres. On a aussi impliqué le norovirus dans trois éclosions mixtes (c.-à-d. éclosions comportant de multiples causes). On a établi un lien entre ces éclosions et 1 818 cas de maladie. Les éclosions virales ont été attribuées en grande majorité à la consommation d'eaux souterraines non traitées ou insuffisamment traitées, comme l'ont signalé d'autres chercheurs (Hynds et coll., 2014a,b; Wallender et coll., 2014).
Les éclosions d'origine hydrique attribuables à des norovirus sont courantes dans le monde entier (Brugha et coll., 1999; Brown et coll., 2001; Boccia et coll., 2002; anderson et coll., 2003; Carrique-Mas et coll., 2003; Maunula et coll., 2005; Hewitt et coll., 2007; Gunnarsdóttir et coll., 2013; Giammanco et coll., 2014). Il se produit aussi des éclosions de VHA partout dans le monde (De Serres et coll., 1999; Hellmer et coll., 2014). On signale aussi fréquemment des liens entre des sources d'eaux souterraines et des éclosions de norovirus et de VHA (Häfliger et coll., 2000; Maurer et Stürchler, 2000; Parchionikar et coll., 2003; Kauppinen et coll., 2017). De grandes épidémies de VHE d'origine hydrique sont survenues dans des pays en développement (Guthmann et coll., 2006), mais aucune n'a été signalée au Canada ni aux États-Unis (Purcell, 1996; Chin, 2000). Des astrovirus et des adénovirus ont aussi été impliqués dans des éclosions causées par l'eau potable même s'ils n'en étaient pas la cause principale (Kukkula et coll., 1997; Divizia et coll., 2004).
5.5 Effet des conditions environnementales
La concentration de virus dans une source d'approvisionnement en eau dépend d'un grand nombre de conditions et de phénomènes environnementaux dont beaucoup ne sont pas bien définis ou ne sont pas transférables entre bassins versants. Les conditions environnementales qui peuvent faire varier la qualité de l'eau comprennent les précipitations, la fonte des neiges, la sécheresse, les incidents en amont (dans le cas des eaux de surface), l'agriculture et la faune (Dechesne et Soyeux, 2007). Les phénomènes météorologiques extrêmes, comme les crues soudaines et les ouragans, dont la fréquence et la gravité devraient augmenter sous l'effet des changements climatiques, peuvent aussi avoir des répercussions importantes sur la qualité de l'eau (Casteel et coll., 2006; Sinigalliano et coll., 2007; Lapointe et coll., 2012; Maslova, 2017).
Plusieurs études ont signalé des éclosions de maladies d'origine hydrique après des précipitations extrêmes (Curriero et coll., 2001; Thomas et coll., 2006; nichols et coll., 2009; new Zealand Department of Internal affairs, 2017). Les éclosions sont souvent classées comme des événements se produisant dans une source d'approvisionnement comme les eaux de surface ou les eaux souterraines, mais on ignore si la catégorie des eaux souterraines comprend toutes les sources d'approvisionnement souterraines ou seulement les réseaux d'eaux souterraines correctement classées. Curriero et coll. (2001) ont évalué le lien entre les précipitations et les maladies d'origine hydrique aux États-Unis et ils ont constaté que les éclosions étaient précédées de précipitations dépassant le 90e centile. Les éclosions attribuables à la contamination des eaux de surface étaient les plus significatives en cas de précipitations extrêmes survenant au cours du mois de l'éclosion, tandis que les éclosions attribuables aux eaux souterraines montraient que les précipitations extrêmes étaient les plus significatives deux mois avant l'éclosion. Au Canada, Thomas et coll. (2006) ont déclaré que les précipitations dépassant le 93e centile augmentaient le risque d'éclosion d'un facteur de 2,3. Une étude réalisée en angleterre a conclu que le risque d'éclosion était lié à deux situations : de faibles précipitations au cours des trois semaines précédentes et des précipitations excessives au cours de la semaine précédant l'éclosion (Nichols et coll., 2009). Une éclosion importante de gastroentérite survenue à Havelock north, en nouvelle-Zélande, au cours de laquelle 5 500 des 14 000 résidants sont tombés malades et qui s'est soldée par trois décès, a été liée à une contamination des sources d'eaux souterraines non traitées découlant de fortes pluies (New Zealand Department of Internal affairs, 2017).
Drayna et coll. (2010) ont signalé un lien important entre les précipitations et les visites à l'urgence pédiatrique dans une communauté desservie par des eaux de surface traitées : le nombre de visites augmentait de 11 % quatre jours après une pluie. Uejio et coll. (2014) ont constaté qu'il y avait un lien entre une précipitation extrême et une augmentation des cas de MGA chez les enfants buvant de l'eau souterraine municipale non traitée. Le risque relatif d'avoir une MGA augmentait pour passer en moyenne de 1 à 1,4 au cours des semaines pendant lesquelles les précipitations atteignaient 3,3 cm comparativement aux semaines sans précipitation. Au cours des semaines où les précipitations ont dépassé 9 cm, le risque relatif passait à 2,4. On n'a constaté aucun lien dans le cas des eaux municipales traitées ou des collectivités desservies par des puits privés. Les auteurs ont posé en hypothèse que l'aire d'alimentation des puits domestiques non traités comportait plusieurs fosses septiques desservant une population peu nombreuse, tandis que l'aire d'alimentation des puits municipaux non traités subissait l'effet de fuites provenant d'égouts sanitaires contenant des matières fécales d'une population importante, ce qui accroîtrait la probabilité d'une contamination virale.
On a établi un lien entre les précipitations et la détection des virus dans les eaux de surface (Fong et Lipp, 2005; Fong et coll., 2005; Rijal et coll., 2009; Edge et coll., 2013; Corsi et coll., 2014; Hata et coll., 2014), une diminution des concentrations (Dorner et coll., 2007) ou l'absence de changement (Choi et Jiang, 2005; Sidhu et coll., 2012). Les causes de l'incohérence de ces constatations ne sont pas claires. Des études ont signalé que les retombées sont propres aux sites et liées à divers facteurs, dont l'hydromorphologie et les sources de contamination fécale (Westrell et coll., 2006a; Dechesne et Soyeux, 2007; Corsi et coll., 2014). Corsi et coll. (2014) ont aussi posé en hypothèse que ces différences sont attribuables à la façon dont les auteurs de diverses études ont défini des variables prédictrices des précipitations.
5.6 Vulnérabilité des eaux souterraines
On reconnaît de plus en plus que des virus entériques peuvent contaminer les eaux souterraines (Schijven et coll., 2010). En général, les puits de surface et les puits forés dans les aquifères du substratum rocheux fracturé, sont très vulnérables à lacontamination fécale (Conboy et Goss, 2000; allen et coll., 2017). Environ 70 % des aquifères régionaux qui fournissent de l'eau potable au Canada sont du substratum rocheux fracturé (RNCan, 2005), et d'autres recherches seront nécessaires pour mieux comprendre la fréquence, l'ampleur et les facteurs associés à la survenue des virus au Canada (Novakowski, 2015; allen et coll., 2017). Les puits situés dans les aquifères sablonneux épais et protégés dotés de nappes phréatiques profondes peuvent présenter un risque plus faible, car le processus d'adsorption atténue les virus (Schijven et coll., 2006; novakowski, 2015).
On a détecté des virus entériques dans divers types d'aquifères, ce qui fait ressortir la complexité que présente le transport de virus dans la subsurface. Des études ont relevé la présence de virus entériques dans des échantillons d'eaux souterraines prélevés de divers aquifères : aquifères de sable semi-confinés (Banks et coll., 2001); aquifères de roche cristalline confinés (Banks et Battigelli, 2002); aquifères du substratum rocheux non confinés (Liberman et coll., 2002; Lindsey et coll., 2002; Borchardt et coll., 2003; Locas et coll., 2007; Trimper et coll., 2010; allen, 2013); aquifères de sable et gravier perméables (Lindsey et coll., 2002; Borchardt et coll., 2003); aquifères du substratum rocheux fracturé confinés (Powell et coll., 2003; Borchardt et coll., 2007; Bradbury et coll., 2013); aquifères de sable et de gravier alluvionnaires (Borchardt et coll., 2004); aquifères de sable et de gravier confinés (Francy et coll., 2004); et aquifères karstiques (Johnson et coll., 2011b).
Outre le matériau constituant l'aquifère, il faut tenir compte de bien d'autres aspects au moment d'évaluer si les eaux souterraines risquent d'être contaminées par des virus (Banks et coll., 2001; Cherry et coll., 2006; Schijven et coll., 2010; Hynds et coll., 2012). Le tableau 1 présente les facteurs qui augmentent considérablement le risque potentiel d'une contamination virale dans les eaux souterraines, comme bien des études le précisent (Conboy et Goss, 2000; Cherry et coll., 2006; novakowski et coll., 2006b; novakowski, 2015; allen et coll., 2017; Hunt et Johnson, 2017; Praamsma, 2017). En ce qui concerne la conception et la construction des puits, Praamsma (2017) a constaté que les puits installés dans le substratum rocheux étaient vulnérables à la contamination de surface causée par la présence de fractures verticales et horizontales interconnectées à au plus 10 mètres de l'élévation du sol en présence d'un mort-terrain minime. Les auteurs ont suggéré que des tubages de puits plus profonds pourraient réduire la vulnérabilité des puits. Dans le même ordre d'idées, Jackson et Heagle (2016) ont remarqué que les puits domestiques dotés de tubages qui se terminent au-dessus de la première couche semi-perméable continue permettent aux contaminants provenant des fosses septiques et d'autres sources de contamination de pénétrer dans le puits. Allen et coll. (2017) ont démontré que la probabilité de détection des virus était positivement corrélée à la longueur d'intervalle ouvert du puits (p. ex. la longueur de roc dégagé sans tubage de puits), parce que les puits ayant des intervalles ouverts plus longs capturent plus de trajets d'écoulement de l'eau souterraine. En général, les puits construits avec des tubages plus profonds qui traversent une couche semi-perméable épaisse et continue et qui disposent d'une prise d'eau sous la couche semi-perméable pourraient êtres sensibles à la contamination. Toutefois, il devient de plus en plus évident que les puits construits d'après les normes actuelles de l'industrie sont vulnérables à la contamination, selon le milieu hydrogéologique (Novakowski et coll., 2006b; 2015; allen et coll., 2017; Whelan et coll., 2017).
La capacité d'une couche semi-perméable de protéger les puits d'approvisionnement en eau contre la contamination virale dépend de son intégrité. Les voies préférées comme les fractures, les trous de racines ou d'autres ruptures, compromettent l'intégrité de beaucoup de couches semi-perméables. Il est possible de détecter la présence de virus dans des aquifères confinés parce que la taille extrêmement petite des virus entériques (< 100 nm) comparativement à la taille probable des fractures de la couche semi-perméable (5 à 50 µm) limite la capacité d'une couche semi-perméable de jouer le rôle de barrière qui bloque le transport des virus (Cherry et coll., 2006). Bradbury et coll. (2013) ont démontré la vulnérabilité d'un aquifère de grès confiné aux conditions environnementales, car ils ont constaté qu'il y avait un lien entre la détection de virus et les précipitations et la fonte des neiges. Gleeson et coll. (2009) ont aussi signalé le rechargement extrêmement rapide et localisé d'un aquifère du substratum rocheux fracturé recouvert d'une mince couche de mort-terrain à cause de la fonte des neiges.
Dans le cas des aquifères confinés, il est difficile de prédire si une couche semi-perméable protège contre la contamination virale. Cherry et coll. (2006) ont recommandé de procéder à des études hydrogéologiques et techniques dans le contexte de la mise en valeur de sources d'eaux souterraines, afin de réunir suffisamment d'information pour établir et caractériser les voies préférées à grande vitesse qui traversent la couche semi-perméable. Des outils d'échantillonnage et de surveillance sont disponibles, mais ils sont toutefois difficiles et coûteux de mettre en œuvre (Bradbury et coll., 2006). Borchardt et coll. (2007) ont conclu qu'en l'absence de modèles robustes de transport microbien, il est préférable de supposer que l'eau souterraine tirée d'un aquifère confiné est aussi vulnérable à la contamination microbienne que celle d'un aquifère non confiné. Hynds et coll. (2012) ont conclu qu'à cause de l'importance des voies de contamination localisée, les évaluations hydrogéologiques ne suffisent pas à elles seules pour évaluer le risque de contamination, particulièrement dans le cas des puits mal conçus, construits ou entretenus.
Étant donné la complexité, l'incertitude et les coûts liés aux évaluations de la vulnérabilité des eaux souterraines, il peut s'avérer plus efficace pour la protection de la santé publique de supposer que la contamination par des agents pathogènes est présente au moins de temps en temps, et d'établir des barrières de sécurité en conséquence (p. ex. désinfection) (Hunt et Johnson, 2017). Des évaluations complètes de systèmes demeurent importantes pour comprendre les facteurs qui contribuent à la contamination de l'eau par tous les pathogènes entériques – bactéries et protozoaires, ainsi que les virus.
| Facteur(sans ordre particulier) | Commentaire |
|---|---|
| Emplacement de la source de matières fécales |
|
| Profondeur de la nappe phréatique |
|
| pH des eaux souterraines |
|
| Matériaux constituant l'aquifère |
|
| Force ionique et précipitation |
|
| Matières organiques dissoutes |
|
| Survie des virus |
|
| Pompage |
|
| Épaisseur du mort-terrain |
|
| Conception, construction et emplacement des puits |
|
5.6.1 Transport des virus
Pang (2009) a passé en revue les publications sur le transport des virus dans les milieux souterrains et a calculé des taux d'élimination à partir d'expériences menées sur le terrain et de grosses carottes de sol intactes dans toutes sortes de conditions. Les taux d'élimination étaient liés à la durée de la contamination, sans compter le débit et les caractéristiques particulières du microbe et des milieux souterrains. Les taux d'élimination des virus (voir le tableau 2) sont représentatifs de conditions de débit saturées. Une élimination supplémentaire peut survenir à travers la zone non saturée si la distance verticale est suffisante entre la source de contamination fécale et la nappe phréatique. La capacité d'élimination des microorganismes pathogènes des aquifères contaminés diminue considérablement en grande partie parce que la matière organique se fixe aux sites d'adsorption disponibles, ce qui bloque l'adsorption des microorganismes pathogènes. L'auteur a prévenu qu'il faut faire preuve de prudence lorsqu'on calcule des marges de recul, car les taux d'élimination peuvent ralentir en fonction de la distance, en particulier dans le cas des aquifères de sable à grain fin et des aquifères où il y a apport continu d'effluents (p. ex. fosses septiques, égouts sanitaires qui fuient, apport artificiel géré). Des renseignements supplémentaires sont disponibles dans l'étude de Pang (2009).
Schijven et Hassanizadeh (2002) ont calculé des marges de recul pour une diminution de 9 log afin de protéger six aquifères sablonneux anoxiques contre des fuites d'égouts sanitaires. Les marges de recul calculées ont varié de 153 à 357 m (moyenne = 228 m). Yates et Yates (1989) ont encouragé une approche basée sur les marges de recul afin d'éviter de supposer que les caractéristiques hydrogéologiques d'un secteur sont constantes. Par contre, DeBorde et coll. (1999) ont signalé que la détermination des marges de recul dans tout un éventail de conditions hydrogéologiques a généralement échoué à cause de l'incertitude liée aux caractéristiques de l'aquifère et aux caractéristiques de chaque virus. L'approche de rechange vise avant tout à appliquer un traitement approprié pour réduire le risque au lieu de quantifier les diminutions logarithmiques.
| Aquifère | Vélocité | Taux d'élimination de virus (log/m) |
Distance calculée pour produire une diminution de 4 lognote a (m) de virus | Conditions de l'étude |
|---|---|---|---|---|
| Sable | < 2 m/j | 100 | 4 | Aquifères de sable de pierre ponce |
| Sable | < 2 m/j | 10-1 – 10-2 | 40 – 400 | Non indiquées |
| Sable et gravier | < 3 m/j | 10-1 | 40 | Distance < 17 m |
| Sable et gravier, y compris FB | < 3 m/j | 10-1 – 10-2 | 40 – 400 | Distance < 177 m |
| Sable et gravier | < 3 m/j | 10-3 | 4 000 | Distance = 183 à 970 m; contaminationnote b |
| Sable et gravier | < 3 m/j | 10-4 | 40 000 | Distance = 210 à 2 930 m; contaminationnote b |
| Gravier sablonneux | > 11 m/j | 10-2 – 10-3 | 400 – 4 000 | Distance < 163 m; proprenote c |
| Gros gravier | > 50 m/j | 10-2 | 400 | Proprenote c |
| Gros gravier | > 50 m/j | 10-3 | 4 000 | Contaminationnote b |
| Till argileux fracturé et saprolite de schiste fracturé | note d | 100 – 10-1 | 4 – 40 | Proprenote c |
| Gneiss fracturé | note d | 10-1 – 10-2 | 40 – 400 | Proprenote c |
| Calcaire fracturé | note d | 10-2 | 400 | Contaminationnote b |
| Craie fissurée | note d | 10-2 – 10-3 | 400 – 4 000 | Contaminationnote b |
| Calcaire karstique | note d | 10-1 – 10-2 | 40 – 400 | Distance < 85 m |
| Calcaire karstique | note d | 10-3 | 4 000 | Distance = 1 250 m; contaminationnote b |
| Calcaire karstique | note d | 10-4 | 40 000 | Distance - 5 000 m; contaminationnote b |
|
||||
5.7 Liens avec les organismes indicateurs
Aux fins du présent document, le terme « indicateur » désigne un microorganisme dont la présence dans l'eau indique une contamination. Un « substitut » s'entend d'un organisme, d'une particule ou d'une substance qu'on utilise pour évaluer le sort d'un microorganisme pathogène dans un environnement naturel (p. ex. processus d'adsorption) ou de traitement (p. ex. désinfection de l'eau potable).
Les organismes indicateurs qui, selon l'approche de la source au robinet ou le du plan de salubrité gestion de la sécurité sanitaire de l'eau, font régulièrement l'objet au Canada d'une surveillance visant à vérifier la qualité de l'eau potable sont E. coli et les coliformes totaux. La présence d'E. coli dans l'eau indique que celle-ci est contaminée par des matières fécales et qu'elle présente donc un risque élevé pour la santé, que l'on y observe ou non la présence d'organismes pathogènes particuliers, comme les virus entériques. Cependant, l'absence d'E. coli ne témoigne pas nécessairement de l'absence de virus entériques. Les coliformes totaux ne sont pas propres aux matières fécales et ne peuvent donc pas servir à révéler une contamination fécale (ni la présence possible d'agents pathogènes entériques). Les coliformes totaux révèlent plutôt des problèmes généraux liés à la qualité de l'eau. Les documents techniques des recommandations sur E. coli et les coliformes totaux (Santé Canada, 2012a, 2012 b) renferment plus de renseignements sur le rôle d'E. coli et des coliformes totaux dans la gestion de la qualité de l'eau.
5.7.1 Sources d'eau de surface
Plusieurs études ont porté sur le lien entre les microorganismes indicateurs et la présence ou l'absence de virus entériques de source humaine dans des sources d'eau de surface et ont présenté des résultats contradictoires. Dans certains cas, on a établi une corrélation entre la présence d'E. coli ou de C. perfringens et celle de virus entériques dans des eaux de surface polluées par des matières fécales humaines (Payment et Franco, 1993; Payment et coll., 2000; Ashbolt et coll., 2001; Hörman et coll., 2004), tandis que dans d'autres, on n'a observé aucun lien entre l'un ou l'autre des indicateurs et les virus entériques (Griffin et coll., 1999; Jiang et coll., 2001; Dorner et coll., 2007; Edge et coll., 2013). Le même phénomène vaut dans le cas des bactériophages. Des études ont signalé un lien entre les bactériophages et la présence de virus entériques dans les eaux de surface (Skraber et coll., 2004; Ballester et coll., 2005; Haramoto et coll., 2005a), tandis que d'autres ne l'ont pas fait (Hot et coll., 2003; Hörman et coll., 2004; Choi et Jiang, 2005). Des chercheurs ont aussi observé à la fois un lien (Haramoto et coll., 2005) et une absence de lien avec des bactéries coliformes (Skraber et coll., 2004; Ballester et coll., 2005; Choi et Jiang, 2005). On a aussi signalé, dans le cas des entérocoques, des observations contradictoires résumées par Lin et Ganesh (2013).
Wu et coll. (2011) ont étudié les données publiées pendant 40 ans sur les corrélations entre les indicateurs et les microorganismes pathogènes et ont conclu que, sauf dans le cas des coliphages F-spécifiques et des adénovirus, aucune autre paire d'indicateurs et microorganismes pathogènes n'a démontré de lien statistiquement significatif. Cette absence générale de corrélation est vraisemblablement attribuable à divers facteurs, notamment aux différents taux de survie dans l'environnement, au lieu échantillonnéet aux différences méthodologiques de l'analyse de l'eau (Payment et Pintar, 2006). Toutefois, Wu et coll. (2011) ont souligné que les indicateurs non fécaux et, en particulier, les coliformes totaux étaient plus souvent corrélés aux microorganismes pathogènes qu'à E. coli, ce qui met en évidence que la surveillance des coliformes totaux en parallèle avec E. coli pourrait être encore plus utile dans les analyses de la qualité de l'eau. Les caractéristiques des bassins versants, notamment les sources et les degrés de contamination fécale, ainsi que des facteurs géochimiques, peuvent influer sur la corrélation entre les indicateurs fécaux et les virus, ce qui explique les différences propres à chaque site (Wilkes et coll., 2009; Payment et Locas, 2011). Ces observations ont soulevé des questions importantes concernant le bien-fondé de l'utilisation des indicateurs traditionnels comme prédicteurs de la contamination des eaux de surface par les virus. Elles ont également mis en évidence la nécessité d'exercer une surveillance régulière des virus entériques dans les eaux de surface afin de mieux comprendre le risque pour la santé publique.
5.7.2 Sources d'eau souterraine
Des études ciblées ont porté sur la présence de virus entériques dans des eaux souterraines (voir la section 5.1.2). Dans le contexte de la surveillance des virus, certaines de ces études ont porté aussi sur la présence de divers microorganismes indicateurs fécaux. Une étude sur des puits privés aux États-Unis a révélé que 8 % des puits analysés par réaction en chaîne de la polymérase (PCR) étaient positifs pour un virus entérique ou plus, mais aucun des puits contaminés ne contenait d'indicateurs de contamination fécale (c.-à-d. E. coli, entérocoques, coliphages) et 25 % seulement des puits touchés par les virus étaient positifs sur le plan des coliformes totaux (Borchardt et coll., 2003). Plusieurs autres études réalisées aux États-Unis n'ont signalé aucun lien entre la détection d'un organisme indicateur et celle de virus entériques dans un échantillon d'eaux souterraines (Abbaszadegan et coll., 1998, 2003; Borchardt et coll., 2004). Abbaszadegan et coll., (2003) ont analysé la présence de virus et d'indicateurs microbiens dans des échantillons d'eaux souterraines de 35 États américains. Les sources d'eaux souterraines étaient situées dans divers lieux hydrogéologiques et certaines se trouvaient sous l'influence des eaux de surface. Dans l'ensemble, il n'y avait pas de corrélation importante entre les indicateurs microbiens et la présence de virus. Par contre, dans le cas des sites ayant fait l'objet d'un échantillonnage répété, il y avait une probabilité accrue que les échantillons ayant produit des résultats positifs pour les virus donnent également des résultats positifs pour les indicateurs microbiens. On a établi plus souvent un lien entre les indicateurs non fécaux (c.-à-d. coliformes totaux et endospores aérobies) et des échantillons contenant des virus qu'avec E. coli et les entérocoques (Abbaszadegan et coll., 2003). Au cours d'une enquête portant sur une importante éclosion de maladies gastro-intestinales attribuables aux eaux souterraines sur l'île South Bass, en Ohio, on n'a observé aucun lien entre virus et indicateur, mais on a détecté des entérocoques et E. coli à des fréquences et en concentrations presque identiques (Fong et coll., 2007; O'Reilly et coll., 2007). Borchardt et coll. (2007) ont signalé la présence de virus entériques dans un aquifère confiné au Wisconsin (É.-U.), mais on n'a pas détecté de bactéries de coliformes fécaux dans les échantillons d'eau positifs pour le virus.
Une méta-analyse a démontré que les bactériophages (coliphages somatiques et à ARN-F) étaient de mauvais indicateurs de la présence ou de l'absence de virus, qu'ils étaient peu nombreux et moins souvent présents que les indicateurs bactériens (Payment et Locas, 2011), ce qui donne à penser que les coliphages sous-estiment la présence de virus entériques. Une méta-analyse plus récente (Hynds et coll., 2014b) a appuyé cette constatation et n'a démontré aussi aucun lien entre les virus entériques et E. coli ou bien les coliformes totaux, mais de faibles liens entre les virus entériques et d'autres indicateurs (p. ex. entérocoques). Fout et coll. (2017) ont procédé à une méta-analyse des données brutes tirées de 12 études internationales des eaux souterraines portant sur 718 réseaux publics d'approvisionnement en eau potable situés dans divers environnements hydrogéologiques. On a évalué les corrélations entre la présence du virus et celle de l'indicateur dans les échantillons et dans les puits en plus de tenir compte de la méthode de détection utilisée (c.-à-d. culture cellulaire par rapport à méthode moléculaire). Les auteurs ont constaté que les virus cultivables étaient statistiquement associés à tous les indicateurs (E. coli, entérocoques, phages F-spécifiques, phages somatiques et spores), sauf pour les coliformes totaux, à l'échelle de l'échantillon; la plus forte association concerne E. coli et les phages somatiques. Par contre, il n'y avait aucune corrélations statistiquement significative entre les virus détectés à l'aide des méthodes moléculaires, appelées virus PCR, et les indicateurs, à l'échelle de l'échantillon (Fout et coll., 2017). À l'échelle des puits, le phage somatique était l'indicateur le plus significativement associé au virus cultivable. Dans le cas du virus PCR, une faible corrélation a été observée pour les coliformes totaux, E. coli et le phage somatique à l'échelle des puits. Dans l'ensemble, les auteurs ont déterminé que les corrélations entre les indicateurs et les virus détectés par des méthodes moléculaires étaient toujours plus faibles que celles entre les virus cultivables. Se fondant sur des analyses supplémentaires, les auteurs ont également déterminé que tous les indicateurs présentaient une faible sensibilité et des faibles valeurs prédictives positives de la présence du virus, peu importe que l'on utilise des approches fondées sur la culture cellulaire ou la méthode moléculaire. Cela signifie que les indicateurs sont souvent absents en présence de virus (c.-à-d. que le taux de faux négatifs est élevé). Abbaszadegan et coll. (2003), Allen (2013) et Fout (2017) ont tous observé que l'échantillonnage répété des indicateurs améliorait leur valeur prédictive positive et permettait de mieux comprendre la sensibilité d'un puits à la contamination virale. Fout et coll. (2017) ont également noté que la force des associations entre les indicateurs microbiens et les virus variait selon le milieu hydrogéologique.
Comme il n'y a pas de lien constant entre les indicateurs et les virus entériques dans les eaux souterraines et comme on a détecté la présence de virus entériques dans toutes sortes de sources d'eaux souterraines, y compris des aquifères confinés, il importe d'évaluer les systèmes pour déterminer la vulnérabilité de tous les aquifères à la contamination virale. La détection des indicateurs peut fournir de l'information quant à savoir si une source d'eau souterraine peut être touchée par la contamination fécale, mais elle doit être considérée à la lumière d'autres facteurs (voir la section 5.6) au moment d'évaluer la vulnérabilité des eaux souterraines à la contamination virale.
5.7.3 Eau potable
En général, la surveillance d'organismes indicateurs dans l'eau potable traitée a pour but de vérifier l'efficacité du traitement. Comme on l'a mentionné, il n'y a pas (toujours) de lien entre les organismes indicateurs d'usage courant (p. ex. E. coli, coliformes totaux, entérocoques) et la présence de virus entériques dans les sources d'approvisionnement en eau.; de sorte que de tels indicateurs ne peuvent pas servir à indiquer la présence de ces virus dans l'eau potable traitée Il est difficile d'utiliser ces indicateurs bactériens pour prédire la présence ou l'absence de virus entériques dans l'eau potable traitée puisqu'ils ont des taux d'élimination différents par des procédés mécaniques et résistent moins aux désinfectants que les virus entériques (Havelaar et coll., 1985; Payment et coll., 1985; Hijnen et Medema, 2010; Santé Canada, 2012a). En dépit de ces limites, l'utilisation simultanée des indicateurs bactériens et des données sur le rendement du traitement (substitut) (section 7.0) permet d'obtenir des renseignementssur le caractère adéquat du traitement de l'eau potable. En particulier, la présence d'E. coli dans l'eau à la sortie d'une station de traitement signifie que le traitement est inadéquat et qu'il y a un risque accru que des agents pathogènes, y compris des virus entériques, soient présents dans l'eau potable traitée.
5.7.4 Autres indicateurs
Comme il n'y a pas d'indicateur constamment fiable de la présence de virus dans des sources d'approvisionnement en eau ou dans l'eau potable, on a proposé la détection directe de virus. L'adénovirus, le polyvirus de marbrure commun du poivre (PMMV) et le virus Torque Teno (TTV) sont au nombre des virus dont on a évalué la capacité prédictive. Pour un aperçu d'autres virus, consulter Lin et Ganesh (2013).
Étant donné leurs concentrations élevées et constantes et leur grande résistance à la désinfection aux rayons UV (voir la section 7.1.3.2), on a proposé d'utiliser les adénovirus comme indicateur possible (Albinana-Gimenez et coll., 2009). Leur faible concentration dans les sources d'approvisionnement en eau et leurs difficultés de détection limitent toutefois leur utilité comme indicateur (Symonds et Breitbart, 2015). Comme l'adénovirus, le PMMV, organisme pathogène du poivrier, est aussi présent en grandes concentrations dans les eaux usées, ainsi que dans les eaux usées traitées, et c'est pourquoi on a proposé de l'utiliser comme indicateur de la présence de virus entériques dans les sources d'approvisionnement en eau (Rosario et coll., 2009). On ne sait toutefois pas trop s'il y a un lien entre leur présence et celle de virus entériques infectieux (Symonds et Breitbart, 2015). Le TTV est très prévalent chez les humains, même s'il ne semble pas pathogène (Biagini, 2004). Le TTV a de nombreux traits en commun avec les virus entériques, y compris des modes de transport et des voies de transmission semblables (Nishizawa et coll., 1997; Abe et coll., 1999; Bendinelli et coll., 2001; Vaidya et coll., 2002; Haramoto et coll., 2005b; Diniz-Mendes et coll., 2008). On a donc suggéré de l'utiliser comme indicateur de la présence de virus entériques (Griffin et coll., 2008; Plummer et Long, 2013; Plummer et coll., 2014). Des études récentes ont signalé une faible occurrence de TTV dans des échantillons d'eaux usées, ainsi que des fluctuations de leur présence (Plummer et Long, 2013; Plummer et coll., 2014), ce qui indique qu'il se peut que le TTV ne soit pas un indicateur convenable.
6.0 Méthodes d'analyse
6.1 Détection des virus entériques
Il existe des méthodes normalisées de récupération et de détection des virus entériques (APHA et coll., 1998; U.S. EPA, 1996, 2001c, 2012; ASTM, 2004). Elles exigent toutefois de l'équipement de laboratoire spécialisé et du personnel très bien formé. Le traitement des échantillons coûte aussi relativement cher, et c'est pourquoi la surveillance routinière des virus entériques dans l'eau n'est pas practicable. Malgré cela, ces méthodes ont été validées et peuvent être utilisées par les laboratoires qui sont en mesure d'assurer la surveillance des virus entériques.. Dans les sections qui suivent, nous donnons un aperçu de ces méthodologies, ainsi que des informations sur des progrès récents en matière de la détection des virus, qui ont servi en contexte de recherche.
6.1.1 Concentration de l'échantillon
Dans le cas des eaux non traitées, on prélève les échantillons le plus souvent à proximité et en profondeur de la prise d'eau potable afin d'obtenir un échantillon représentatif de la source d'approvisionnement en eau. Les échantillons d'eau sont filtrés sur le terrain et ensuite expédiés sur de la glace à un laboratoire qui les traitera le plus rapidement possible (idéalement, dans les 24 heures). Le volume d'eau filtrée dépend de la concentration de virus prévue dans l'eau (c.-à-d. propre à un site) : plus la concentration des virus prévue est faible, plus le volume des échantillons doit être important. Les méthodes actuelles recommandent de filtrer quelques centaines de litres d'eau de surface et 1 500 litres ou plus d'eau souterraine (U.S. EPA, 2012; Cashdollar et coll., 2013; Fout et coll., 2015).
Deux méthodes de filtration traditionnelles servent à déterminer la concentration initiale des virus : la filtration par adsorption et la filtration par exclusion (ultrafiltration). La filtration par adsorption peut employer des filtres électropositifs comme ceux que prescrit la méthode 1615 (U.S. EPA, 2012) de l'Environmental Protection Agency (EPA) des États-Unis, des filtres chargés négativement (Beuret, 2003; Haramoto et coll., 2004; Fuhrman et coll., 2005; Villar et coll., 2006), des membranes de nitrocellulose (Hsu et coll., 2006) ou des filtres en laine de verre (Lambertini et coll., 2008). Au pH ambiant, la plupart des virus entériques sont chargés négativement et, par conséquent, ils sont capturés par un support filtrant électropositif. Pour adsorber des virus au moyen d'un support filtrant chargé négativement, il faut ajouter à l'échantillon un cation, comme du chlorure de magnésium, et il peut être nécessaire de rajuster le pH de l'échantillon pour obtenir un pH acide. Comme les virus s'adsorbent au support filtrant, il faut les éluer par la suite du filtre à l'aide d'une solution alcaline qui modifie la charge superficielle des particules virales de façon à ce qu'elles soient éluées dans la solution. Les éluants contiennent couramment de l'extrait de bœuf, de la glycine, du bouillon tryptose phosphate ou de l'hydroxyde de sodium dans les solutions (Katayama et coll., 2002; Hörman et coll., 2004; Brassard et coll., 2005; Villar et coll., 2006).
Les méthodes d'exclusion par la taille, comme l'ultrafiltration, sont indépendantes du pH et ont l'avantage de ne pas nécessiter d'étape d'élution (Olszewski et coll., 2005). À cause de la taille extrêmement petite des pores du filtre requis, l'encrassement est toutefois fréquent. D'habitude, on ne peut filtrer qu'environ 20 litres d'eau à la fois (Griffin et coll., 2003), bien qu'on utilise jusqu'à 200 litres dans certains laboratoires (Francy et coll., 2013). On continue d'optimiser les méthodes d'ultrafiltration, notamment pour récupérer simultanément des protozoaires, des bactéries et des virus (Morales-Morales et coll., 2003; Hill et coll., 2005, 2007; Liu et coll., 2012; Kahler et coll., 2015).
On a signalé un vaste éventail de taux de récupération pour différents virus tirés de l'eau, selon les méthodes de filtration et de concentration utilisées (Albinana-Gimenez et coll., 2009; Karim et coll., 2009). Dans le cas de la filtration par absorption utilisant différents filtres électropositifs, les taux de récupération déclarés varient de 0,016 % à 182 % (Huang et coll., 2000; Kittigul et coll., 2001; Albinana-Gimenez et coll., 2009; Karim et coll., 2009; Li et coll., 2010). De même, les taux de récupération varient lorsqu'on utilise des systèmes d'ultrafiltration différents (Paul et coll., 1991; Juliano et Sobsey, 1998; Soule et coll., 2000; Jiang et coll., 2001; Winona et coll., 2001; Olszewski et coll., 2005). Il s'ensuit qu'il est généralement recommandé de traiter en parallèle des échantillons d'eau ensemencés afin de mieux comprendre la présence réelle de virus. La concentration initiale de l'échantillon d'eau est habituellement suivie d'une concentration secondaire pour ramener le volume de l'échantillon à 1 à 2 ml de manière à produire un concentré suffisant pour détecter des virus. Les méthodes de concentration secondaires comprennent la floculation organique, la précipitation par polyéthylèneglycol et l'ultracentrifugation.
6.1.2 Méthodes de détection
Après la concentration de l'échantillon, il est possible d'utiliser tout un éventail de méthodes de détection. La plupart des méthodes de détection courantes comprennent des méthodes de culture cellulaire, des méthodes moléculaires (c.-à-d. basées sur la réaction PCR) ou une combinaison des deux [p. ex. test intégré de culture cellulaire et PCR quantitative (ICC-qPCR)]. Le choix de la méthode de détection à utiliser dépend de tout un éventail de facteurs propres au site, ainsi que du virus préoccupant. Certains virus sont difficiles ou impossibles à cultiver et c'est pourquoi on ne peut utiliser la méthode de culture cellulaire. La section qui suit décrit d'autres facteurs dont il faut tenir compte.
6.1.2.1 Culture cellulaire
Dans le passé, la culture cellulaire était la technique la plus répandue pour détecter des virus et elle demeure la meilleure méthode pour déterminer la présence de virus infectieux dans l'eau. La capacité de détecter des virus infectieux dans des échantillons d'eau est importante si l'on veut prédire les risques pour la santé du public et elle est essentielle pour réaliser une ÉQRM. Les virus entériques ne se reproduisent toutefois pas tous en culture cellulaire ou ne produisent pas tous un effet cytopathogène (ECP) clair (c.-à-d. une plage) nécessaire à la détection visuelle de l'infectiosité. Cette méthode peut sous-estimer la concentration virale dans un échantillon. Si certains virus se développent rapidement (p. ex. en quelques jours), les analyses par culture cellulaire peut prendre plusieurs semaines pour confirmer les résultats négatifs ou pour détecter les virus à croissance lente. De plus, l'agrégation des virus dans un échantillon peut entraîner qu'une plage donnée soit infectée par plus d'un virus, ce qui sous-estime la concentration virale (Teunis et coll., 2005). D'autres problèmes méthodologiques ont aussi une incidence sur les analyses par culture cellulaire, comme l'incapacité de préserver les cellulles en monocouche suffisamment longtemps pour que certains virus à croissance lente produisent une plage visible, et la présence de virus entériques à croissance rapide, qui peuvent entraîner une sous-estimation de la concentration des virus à croissance lente (Irving et Smith, 1981; Fong et Lipp, 2005). Il n'y a actuellement pas de lignée cellulaire universelle pour effectuer des cultures de tous les virus entériques.
6.1.2.2 Méthodes moléculaires
On a aussi utilisé des méthodes moléculaires pour détecter les virus entériques. Une brève description de certaines de ces méthodes suit.
La plupart des méthodes moléculaires de détection des virus entériques reposent sur la réaction PCR. Cette technique consiste à provoquer la lyse des virus pour libérer leur acide nucléique (ADN ou ARN) et ensuite introduire des amorces ciblant des régions codantes précises et amplifier ces régions. L'électrophorèse en gel d'agarose, la coloration au bromure d'éthidium et l'examen visuel à la lumière ultraviolette servent à déterminer la présence d'un signal PCRpositif. Les résultats sont habituellement déclarés sous forme de nombre de copies génomiques (cg) d'un virus/L.
Les méthodes de détection par PCR offrent des avantages importants par rapport aux méthodes de culture cellulaire : elles sont rapides (résultats connus en moins de 24 heures), hautementsensibles et si elles sont bien conçues, elles sont très spécifiques comparativement à la culture cellulaire. On a mis au point des méthodes de détection par PCR pour la plupart des principaux virus entériques d'intérêt dans le cas de la transmission d'origine hydrique. Il est possible de les appliquer à la détection simultanée de nombreux virus, par la réaction PCR multiplex (Fout et coll., 2003, Lee et coll., 2008). Le principal inconvénient des méthodes basées sur la PCR réside dans le fait qu'elles ne permettent pas de déterminer si les virus sont viables ou infectieux. Cela signifie qu'il ne faudrait pas interpréter les copies génomiques comme une mesure de la viabilité ou de l'infectiosité du virus. Les méthodes basées sur la PCR peuvent aussi être inhibées par des composés environnementaux courants, comme des acides humiques et fulviques, des métaux lourds et des composés phénoliques (Fong et Lipp, 2005). Il est possible d'extraire les inhibiteurs des échantillons, mais il faut à cette fin procéder à un traitement supplémentaire qui entraîne une perte de sensibilité. Il faut tenir compte de ces limites dans l'interprétation des résultats de la réaction PCR. On a mis au point des variantes de la méthode classique qui servent à détecter des microorganismes pathogènes, la plus courante étant la PCR quantitative (q)PCR (aussi appelée PCR en temps réel). La réaction qPCR est une PCR modifiée utilisant des sondes oligonucléotidiques et des colorants qui provoquent la fluorescence lorsqu'ils sont liés à un acide nucléique de virus. La région cible d'un virus étant amplifiée, la fluorescence est mesurée en temps réel, ce qui permet de quantifier les produits de la PCR. Cette méthode comporte plusieurs avantages par rapport à la PCR classique, dont l'élimination de l'analyse consécutive à la réaction (c.-à-d. que l'électrophorèse sur gel n'est pas nécessaire), un plus grand débit, la diminution de la probabilité de contamination (c.-à-d. système de tubes fermés) et la capacité de quantifier les virus au moyen d'une courbe standard (Smith et Osborn, 2009). La qPCR offre d'autres avantages uniques, dont sa capacité de différencier les types de virus, et la détection simultanée de différents microorganismes (c.-à-d. multiplexage) (Marion et coll., 2014; Bonilla et coll., 2015).
Les méthodes intégrant la culture cellulaire et la réaction PCR permettent de raccourcir le temps de traitement (comparativement à la culture cellulaire seule), d'améliorer la détection des virus qui ne forment pas l'ECP ou en forment en partie seulement (au moyen de la culture cellulaire traditionnelle) et de détecter des virus infectieux. Il est aussi possible de combiner des méthodes de culture cellulaire à des méthodes immunologiques (p. ex. cytométrie de flux) pour améliorer la détection des virus (Bosch et coll., 2004; Cantera et coll., 2010; Li et coll., 2010). L'amélioration de la sensibilité de l'analyse est un autre avantage qu'offre la combinaison de la culture cellulaire à des méthodes immunologiques ou moléculaires (Payment et Trudel, 1993; Jothikumar et coll., 2000; Hurst et coll., 2001; Payment, 2001, 2007; Reynolds et coll., 2001; Greening et coll., 2002; Ko et coll., 2003), car les cellules infectées amplifient la quantité de virus et offrent ainsi davantage de matière cible à détecter. Dans le cas de la réaction ICC-qPCR, par exemple, il y a amplification de l'acide nucléique du virus après que les virus ont été cultivés dans la culture cellulaire pendant une période variant d'à peine cinq heures jusqu'à quelques jours. La culture vise à produire plus de virus pour l'amplification par la réaction PCR et à accroître ainsi la sensibilité. En outre, comme les échantillons sont dilués dans un milieu de culture cellulaire, on croit que l'inhibition de la réaction PCR est réduite au minimum. On croit aussi que la toxicité de la culture cellulaire (effets cytotoxiques causés par la présence de toxines plutôt que de virus qui inactivent habituellement les cellules en monocouche) est réduite au minimum puisqu'il est possible d'interrompre l'analyse par congélation et de détecter les virus avant la mort cellulaire (Reynolds, 2004). Tout comme la culture cellulaire traditionnelle, la réaction ICC-qPCR est limitée à la détection des virus qui se reproduisent en culture cellulaire (c.-à-d. des virus cultivables). La réaction ICC-qPCR a été appliquée avec succès à la détection de virus entériques infectieux dans des échantillons environnementaux (Xagoraraki et coll., 2007; Balkin et coll., 2010; Dong et coll., 2010; Rigotto et coll., 2010; Ming et coll., 2011; Pang et coll., 2012; Fongaro et coll., 2013, 2015; Ogorzaly et coll., 2013a). Cette méthode a été utilisée aussi au cours d'études portant sur la désinfection, et en particulier celles qui visaient à évaluer l'impact des rayons UV sur l'adénovirus (Gerrity et coll., 2008; Li et coll., 2009; Mayer et coll., 2010; Ryu et coll., 2015). Certaines études signalent des estimations comparables de l'inactivation logarithmique au milieu de culture cellulaire et de la réaction ICC-qPCR (Gerrity et coll., 2008; Ryu et coll., 2015), mais dans d'autres cas, les chercheurs ont observé des taux beaucoup plus élevés (p. ex. d'un facteur de trois) d'inactivation par culture cellulaire (Li et coll., 2009; Mayer et coll., 2010).
On a aussi étudié des méthodes indépendantes de la culture cellulaire pour détecter la présence de virus infectieux. L'immunocapture-qPCR, par exemple, a été appliquée avec succès à la détection de particules virales intactes (c.-à-d. qui peuvent être infectieuses) (Haramoto et coll., 2010; Ogorzaly et coll., 2013b). D'autres recherches s'imposent toutefois pour évaluer l'utilité de cette méthode.
6.2 Détection d'indicateurs viraux
Comme on l'a indiqué ci-dessus, les méthodes de détection des virus dans l'eau ne sont pas pratiques à des fins de surveillance régulière. On a donc proposé d'utiliser divers indicateurs (voir la section 5.7) pour révéler la présence de virus entériques dans l'eau (Deere et coll., 2001; OMS, 2004). Les indicateurs utilisés le plus couramment sont Escherichia coli (E. coli), les coliformes totaux, les entérocoques, les spores de Clostridium perfringens et les coliphages.
6.2.1 E. coli
Escherichia coli est l'indicateur microbien le plus souvent utilisé pour révéler la contamination fécale de sources d'approvisionnement en eau. Le document de Santé Canada (2006a) fournit plus de détails sur les méthodes de détection d'E. coli.
6.2.2 Coliformes totaux
Bien qu'il ne s'agisse pas d'un indicateur de la contamination fécale, les coliformes totaux sont utiles comme indicateurs de la qualité globale de l'eau. Le document de Santé Canada (2006 b) fournit plus de détails sur les méthodes de détection des coliformes totaux.
6.2.3 Entérocoques
Les entérocoques peuvent servir d'indicateur de la contamination fécale et, indirectement, de la présence de virus (U.S. EPA, 2000; Ashbolt et coll., 2001). Des méthodes normalisées de détection des entérocoques dans l'eau ont été publiées (APHA et coll., 1998; U.S. EPA, 2002a,b). Des trousses commerciales pour la détection de ces indicateurs sont aussi disponibles.
6.2.4 Clostridium perfringens
Les spores de Clostridium perfringens sont des indicateurs de contamination fécale récente et passée, mais elles ne sont pas aussi nombreuses que les coliformes dans les matières fécales ou dans l'eau contaminée. Ces spores servent aussi d'indicateurs de l'efficacité du traitement (voir les sections 6.3 et 7.0). Des méthodes de détection normalisées pour C. perfringens ont été publiées (ASTM, 2002; HPA, 2004).
6.2.5 Coliphages
Les coliphages peuvent servir à la surveillance microbienne des réseaux d'eau souterraine (U.S. EPA, 2006d) et à l'évaluation de l'efficacité du traitement de l'eau (voir les sections 6.3 et 7.0). Des méthodes normalises de détection des coliphages dans l'eau ont été publiées (Mooijman, 2001, 2005; U.S. EPA, 2001a,b; APHA et coll., 2005).
6.3 Détection de substituts
On a proposé divers paramètres substituts pour évaluer le sort et le transport des virus dans l'environnement (voir la section 5.2) et l'efficacité du traitement (c.-à-d. diminution des virus entériques par filtration naturelle ou artificielle). Les spores de C. perfringens et les bactériophages sont les substituts les plus courants. On utilise généralement trois types de bactériophages comme indicateurs : les coliphages somatiques, les bactériophages à ARN-F propres au mâle (aussi appelés coliphages F-spécifiques) et les phages Bacteroides (c.-à-d. les phages qui infectent Bacteroides fragilis, B. thetaiotaomicron et la souche GB-124 de Bacteroides) (Grabow, 2001; Armon, 2015). Aux États-Unis, des méthodes normalisées de détection des coliphages somatiques et propres au mâle ont été mises au point (U.S. EPA, 2001a,b). L'Organisation internationale de normalisation (ISO) a aussi publié des méthodes normalisées (série ISO 10705) pour la détection des bactériophages (Mooijman et coll., 2001, 2005).
7.0 Techniques de traitement
Le principal objectif du traitement est de diminuer le nombre d'organismes causant des maladies et les facteurs de risque connexes pour les ramener à un niveau acceptable ou sécuritaire. Il est possible d'y parvenir grâce à une ou à plusieurs méthodes de traitement qui éliminent mécaniquement ces organismes ou les inactivent. Pour assurer au mieux l'enlèvement ou l'inactivation des microorganismes pathogènes, il faut comprendre l'importance relative de chacun des procédés. Certains systèmes d'approvisionnement en eau utilisent plusieurs procédés redondants, de sorte que le traitement reste adéquat même en cas de faille de l'un des procédés. Dans les autres réseaux, tous les procédés en place doivent bien fonctionner pour que le degré de traitement requis soit obtenu. Par exemple, de nombreux approvisionnements en eau souterraine reposent uniquement sur la désinfection pour inactiver les virus. Pour ces systèmes, l'échec du procédé de désinfection pourrait entraîner l'éclosion d'une maladie d'origine hydrique. Puisque les méthodes d'analyse disponibles rendent difficile le contrôle régulier des microorganismes pathogènes dans l'eau potable traitée, il faut se concentrer sur la caractérisation des risques associés aux sources d'approvisionnement en eau et veiller à ce que des procédés de traitement efficaces soient en place afin d'assurer la salubrité de l'eau potable. Il faut aussi mettre en place si possible des mesures de protection des sources d'approvisionnement en eau, notamment assurer le traitement des eaux usées, afin de diminuer au minimum la contamination fécale.
L'approche de la source au robinet, y compris la protection des bassins versants ou des têtes de puits, est une méthode universellement acceptée de diminution des virus entériques et des autres microorganismes pathogènes d'origine hydrique dans l'eau potable (O'Connor, 2002; CCME, 2004; OMS, 2012). Il faut également que les opérateurs aient reçu une formation pour assurer en tout temps le bon fonctionnement des procédés de traitement (Smeets et coll., 2009).
La section 7.1 traite des technologies de traitement à l'échelle municipale qui peuvent éliminer ou inactiver les virus entériques dans l'eau potable. Les systèmes résidentiels peuvent avoir une capacité nominale de traiter des volumes plus importants que ceux qui sont nécessaires à une seule résidence et peuvent donc servir ainsi dans de petits systèmes, comme il est mentionné dans la section 7.2. Il faut obtenir de l'autorité compétente chargée de la protection de l'eau potable de l'information précise sur les technologies applicables aux petits réseaux.
7.1 Traitement à l'échelle municipale
Il existe tout un éventail de technologies d'élimination mécanique et d'inactivation disponibles pour diminuer efficacement les virus entériques afin d'atteindre les objectifs du traitement approprié de l'eau potable. Nous abordons brièvement dans ce document des méthodes possibles de traitement des virus, mais d'autres renseignements figurent déjà dans d'autres sources (U.S. EPA, 1991; Deere et coll., 2001; LeChevallier et Au, 2004; Hijnen et Medema, 2010; AWWA, 2011a). Il est essentiel d'atteindre les cibles en matière d'élimination mécanique et d'inactivation avant que l'eau ne parvienne au premier consommateur du réseau de distribution.
Les procédés d'élimination mécanique, comme la technologie de filtration, se voient attribuer un crédit d'« élimination logarithmique » pour la diminution des niveaux de virus entériques lorsque ces procédés atteignent certaines limites de turbidité de l'eau à la sortie des filtres, comme l'explique la section 7.1.2. Les procédés de désinfection primaire font partie des méthodes d'inactivation. Les crédits d''« inactivation logarithmique » sont calculés à l'aide des concepts de la désinfection présentés à la section 7.1.3. On fait le total de l'élimination logarithmique et du crédit d'« inactivation logarithmique » pour calculer la '«diminution logarithmique» totale des virus dans le cas du procédé de traitement évalué. Comme la désinfection primaire est un procédé important pour inactiver les virus, il est essentiel de comprendre les plages de calcul établies et la façon dont les différents paramètres peuvent nuire à l'efficacité de la désinfection primaire (voir la section 7.1.3). La désinfection secondaire sert à maintenir, dans le réseau de distribution, une concentration résiduelle de désinfectant de manière à prévenir la recroissance microbienne et sert de sentinelle pour les changements de la qualité de l'eau. Même si aucun crédit d'inactivation logarithmique n'est accordé aux procédés de désinfection secondaire, la surveillance des résiduels de désinfectant peut fournir un avertissement précoce que des contaminants sont entrés dans le réseau de distribution (voir la section 7.1.4). Les spécialistes de l'industrie encouragent le recours à la surveillance continue en ligne des résiduels de chlore dans le réseau de distribution (AWWA, 2015).
À cause des difficultés causées par l'analyse routinière des virus entériques, on a déterminé que d'autres microorganismes, comme les spores de Clostridium perfringens et les bactériophages, pourraient servir de substitut pour évaluer l'efficacité des procédés de traitement (Payment et Franco, 1993; Havelaar et coll., 1995; Nasser et coll., 1995; Hijnen et Medema, 2010). On utilise souvent des substituts dans des essais pour déterminer l'efficacité et des études pilotes pour calculer l'élimination logarithmique ou l'inactivation des virus entériques par un procédé de traitement précis. Plusieurs études ont démontré que les bactériophages remplacent bien les virus entériques et qu'on a utilisé autant les bactériophages somatiques que les bactériophages à ARN F-spécifique dans tout un éventail d'évaluations du traitement de l'eau potable. Ces microorganismes comprennent notamment les bactériophages MS2, Qß, F2, PRD-1 et ɸX174 (Mesquita et Emelko, 2012). L'utilisation de ces microorganismes comme substituts pour évaluer l'efficacité de la station de traitement à grande échelle a toutefois été limitée, car leur concentration dans les sources d'eau est généralement insuffisante pour qu'ils soient utiles afin de vérifier régulièrement l'efficacité du traitement (Payment et Locas, 2008). Sinclair et coll. (2012) ont signalé que le choix d'un substitut approprié dépend de tout un éventail de facteurs, dont le type de virus entériques trouvé dans l'eau de source et le procédé de traitement à évaluer. Il importe de signaler que l'utilisation de différentes méthodes (voir la section 6.0) peut produire différentes évaluations de l'efficacité du traitement.
Étant donné l'incertitude associée à l'utilisation de l'élimination logarithmique et les crédits d'inactivation logarithmique calculés à partir de publications scientifiques, il est recommandé que chaque réseau de distribution d'eau potable procède à une évaluation propre au site de l'efficacité du traitement (Smeets, 2011). Les essais en laboratoire et à l'échelle pilote peuvent servir à optimiser les procédés, et les essais avec traceur et la modélisation hydraulique à l'échelle réelle peuvent fournir des estimations plus précises du temps de contact à utiliser pour les calculs d'inactivation de la désinfection primaire. Hijnen (2011) a mis au point une méthode générique qu'il est possible d'utiliser pour effectuer une évaluation propre au site de la capacité de diminution logarithmique du procédé de traitement d'une station en particulier. La méthode repose sur l'établissement du rapport entre la diminution logarithmique des virus entériques (ou d'un virus en particulier) et la diminution logarithmique d'indicateurs/substituts contrôlés de façon routinière comme E. coli ou les spores de C. perfringens à partir des données recueillies à une station de traitement à grande échelle. Comme il est impossible dans bien des cas de surveiller directement les virus, l'auteur a proposé d'utiliser la détermination de la diminution logarithmique des indicateurs fécaux bactériens (basée sur l'analyse de volumes importants) comme estimation approximative de la diminution logarithmique des virus. On peut aussi utiliser des essais pour déterminer l'efficacité et des essais pilotes simulant des conditions à grande échelle pour réunir des données quantitatives sur la diminution logarithmique des virus entériques ou des substituts qui sont propre au site.
7.1.1 Degré de traitement nécessaire
Pour déterminer le degré de traitement nécessaire, il faut caractériser adéquatement les sources d'approvisonnement en eau. La caractérisation des sources d'approvisionnement en eau comprend généralement plusieurs étapes , dont la délimitation de la zone de la source d'approvisionnement en eau, le repérage des menaces qui pèsent sur la source d'approvisionnement en eau et des risques qu'elle encourt, ainsi que la mesure de la qualité microbiologique, physique, chimique et radiologique de la source d'approvisionnment (OMS, 2012, 2017). Il est également important de surveiller les changements saisonniers pour que les services publics de distribution de l'eau potable produisent constamment de l'eau traitée de haute qualité, quelles que soient les conditions de l'eau brute (Valade et coll., 2009; Huck et Sozański, 2011).
Dans la mesure du possible, il faut caractériser les concentrations de virus dans les sources d'approvisionnement en eau en s'appuyant sur un échantillonnage et une analyse à l'échelle réelle. Il faut tenir compte des conditions normales, comme de la surveillance guidée par les événements, notamment le ruissellement printanier, les tempêtes et les déversements d'eaux usées. Les résultats de l'analyse doivent tenir compte du taux de récupération pour la méthode d'analyse et de la viabilité des microorganismes pathogènes afin de produire l'évaluation la plus précise de la présence de microorganismes pathogènes infectieux dans les sources d'approvisionnement en eau (Emelko et coll., 2008, 2010; Schmidt et Emelko, 2010; Schmidt et coll., 2010). À de nombreux endroits, il se peut qu'il soit impossible de prélever des échantillons pour détecter la présence de virus entériques et l'on peut recourir à la caractérisation des sources d'approvisionnement, y compris les paramètres de la qualité de l'eau qui peuvent renseigner sur le risque ou le niveau de contamination fécale, pour déterminer le degré de traitement nécessaire. On trouve ailleurs (OMS, 2012) plus de renseignements sur la caractérisation des risques dans les petits systèmes. Pour déterminer le procédé de traitement le plus approprié, il faut procéder à une évaluation du site et effectuer un essai pilote ou une analyse convenables. Des situations météorologiques plus instables rattachants aux changements climatiques rendront le choix d'un procédé adéquat encore plus important (Huck et Coffey, 2004).
Il faut évaluer les sources d'eau de subsurface pour déterminer si l'approvisionnement peut être contaminé par des virus entériques (voir la section 5.6) et des protozoaires (Santé Canada, 2012c). Les évaluations hydrogéologiques propres au site devraient au moins répertorier les sources de matières fécales dans le secteur de la source d'approvisionnement en eau (p. ex. déchets animaux et humains domestiques, faune, pressions exercées par les activités récréatives) et définir le sous-sol (p. ex. profondeur de l'aquifère, zones de protection, composition des couches de subsurface, conditions de la trajectoire d'écoulement préférentielle, risques liés aux précipitations). Dans le cas des sources jugées vulnérables aux virus, l'élimination ou l'inactivation des virus doit atteindre au moins 4 log (99,99%). L'autorité responsable en matière d'eau potable dans le secteur de compétence concerné peut exiger une surveillance supplémentaire des indicateurs ou des substituts dans le cadre de l'évaluation des risques (Mesquita et Emelko, 2015).
Des technologies de traitement devraient être en place pour obtenir une élimination ou une inactivation des virus entériques d'au moins 4 log (99,99 %) dans le cas des sources vulnérables à la contamination virale. Un tel traitement permet de ramener une concentration de 1 virus/100 L d'eau d'une source d'approvisionnement à 1 × 10−4 virus/100 L, ce qui satisfait à l'objectif sanitaire de 10−6 années de vie corrigées de l'incapacité (AVCI)/personne par année (voir la section 8.0 pour en savoir davantage sur les AVCI). L'eau brute peut toutefois, dans bien des cas, contenir une concentration beaucoup plus élevée de virus, et un traitement supplémentaire peut s'imposer pour respecter les exigences sanitaires en matière d'élimination logarithmique ou d'inactivation des virus. Il est important de souligner que le traitement actuel peut fournir une inactivation virale supérieure à 4 log. À titre d'exemple, les stations de traitement des eaux de surface qui assurent la désinfection au chlore libre résiduel pour obtenir une inactivation de 0,5 log de Giardia produisent généralement une inactivation des virus d'au moins 8 log (Santé Canada, 2018). Il importe de surveiller le procédé (p. ex. turbidité, dose de désinfectant et résiduel, pH, température et débit) pour vérifier que l'eau a été convenablement traitée et qu'elle est, par conséquent, d'une qualité microbiologique acceptable.
7.1.2 Élimination mécanique
Il est possible d'éliminer mécaniquement les virus par diverses technologies, dont la filtration avec procédé chimique, la filtration lente sur sable, la filtration sur diatomées et la filtration sur membrane ou une autre technique de filtration approuvée. La filtration sur berge est une méthode de filtration passive d'élimination des microorganismes que l'autorité qui a compétence sur l'eau potable pourrait aussi envisager d'utiliser.
En général, l'élimination mécanique des virus entériques peut être difficile à cause de leur petite taille (20 à 350 nm) et des variations du point isoélectrique et de l'hydrophobicité des différents virus qui peuvent être présents dans la source d'approvisionnement en eau (Boudaud et coll., 2012). À cause des différences au niveau de la taille, de la forme et du point isoélectrique entre les virus entériques et leurs substituts, leur élimination peut varier selon le procédé de traitement. Mayer et coll. (2015) et Michen et Graule (2010) ont signalé des points isoélectriques de certains virus entériques et substituts.
La surveillance aux stations de traitement à grande échelle indique que l'élimination mécanique des virus entériques varie et que la désinfection constitue par conséquent une importante barrière de traitement des virus entériques d'origine hydrique (Bell et coll. 1998). Hijnen et Medema (2010) présentent une critique détaillée des études réalisées sur l'élimination mécanique des virus.
7.1.2.1 Filtration avec procédé chimique
La coagulation vise à déstabiliser (c.-à-d. à en neutraliser la charge) les particules colloïdales (y compris les organismes pathogènes) afin qu'elles s'agglomèrent bien pendant la floculation et soient par la suite éliminées par clarification (sédimentation ou flottation à l'air dissous). Les appareils à contact de boues, y compris les procédés de floculation lestée de sable, combinent la coagulation, la floculation et la clarification en un seul procédé. La filtration sur milieu granulaire peaufine le travail en éliminant d'autres petites particules colloïdales que la clarification n'a pas éliminées (s'il y a lieu). Pour que la filtration soit efficace, les particules colloïdales doivent être déstabilisées pendant la coagulation. C'est pourquoi la combinaison coagulation, clarification et filtration sur milieu granulaire est reconnue comme un procédé de traitement physico-chimique et couramment appelée filtration avec procédé chimique. Les stations de filtration directe n'ont pas d'étape de clarification et les stations de filtration en ligne n'ont pas d'étape de floculation ni de clarification (AWWA, 2011a; MWH, 2012). Outre l'élimination physique des virus par filtration avec procédé chimique, une recherche récente a montré que les virus peuvent aussi perdre leur infectiosité après une exposition à des espèces hydrolysant l'aluminium au cours de la coagulation (Matsushita et coll., 2011).
Les publications ont décrit des études à échelle réelle analysant l'élimination des virus entériques ou des substituts par filtration avec procédé chimique (Stetler et coll., 1984; Payment et coll., 1985, Payment et Franco, 1993; Havelaar et coll., 1995). On a aussi réalisé plusieurs études pilotes et des études en laboratoire au cours desquelles on a injecté des virus entériques ou des substituts dans l'eau non traitée pour déterminer l'efficacité de l'élimination (Rao et coll., 1988; Bell et coll., 1998; Gerba et coll., 2003; Hendricks et coll., 2005; Abbaszadegan et coll., 2007, 2008; Mayer et coll., 2008; Shin et Sobsey, 2015). Ces études ont analysé l'élimination de tout un éventail de virus entériques, y compris les entérovirus, les norovirus, les rotavirus, l'adénovirus et les virus de l'hépatite, ainsi que des substituts. L'élimination logarithmique mentionnée pour ces études a démontré que le procédé de clarification réalisait une élimination logarithmique de 0,6 à 3,2, suivie d'une élimination supplémentaire de 0,5 à 3,8 par le procédé de filtration. L'élimination totale (c.-à-d. par clarification et filtration) a varié de 1,0 à 5,1, la majorité des stations de traitement parvenant à réaliser une élimination logarithmique de 2,2 à 3,8. En général, l'élimination des virus entériques au moyen de la filtration avec procédé chimique peut varier énormément selon le degré d'optimisation du procédé et le type de virus présent dans la source d'approvisionnement en eau (Bell et coll., 1998; LeChevallier et Au, 2004). Hijnen et Medema (2010) ont effectué une analyse détaillée des études mentionnées ci-dessus, ainsi que d'autres études. Les auteurs ont procédé à une analyse statistique des données sur l'élimination des virus entériques, notamment en pondérant les données basées sur l'ordre de grandeur du procédé et le type de microorganisme, et ils ont déterminé que la filtration avec procédé chimique peut produire une élimination logarithmique moyenne de 3,0. Cette étude a toutefois mis aussi en évidence la variation importante observée dans l'élimination des virus entre les études analysées, ce qui indique les limites de l'application des valeurs moyennes d'élimination logarithmique pour estimer l'efficacité des procédés de traitement de l'eau potable.
Du point de vue opérationnel, on a signalé que la dose de coagulant, le type de coagulant, le pH et la température constituent des variables importantes dans l'élimination des virus (LeChevallier et Au, 2004, Hendricks et coll., 2005; Hijnen et Medema, 2010). Hendricks et coll. (2005) ont signalé une diminution de la dose d'alun de 26 mg/L à 13 mg/L dans une station pilote de filtration avec procédé chimique afin de diminuer l'élimination logarithmique de MS2, qui est tombée de 3,0 à 1,0, et une diminution de l'élimination logarithmique de ɸX174 qui est passée de 5,10 à 1,5. Les auteurs ont signalé que les éliminations logarithmiques étaient presque nulles dans le cas du MS2 et du ɸX174 lorsque le procédé n'était pas dosé à l'alun. Ces résultats correspondent à ceux d'autres études qui ont signalé l'élimination minime ou nulle des virus ou des substituts lorsque la filtration n'est pas précédée d'une coagulation (Nasser et coll., 1995; Hijnen et coll., 2010). On a aussi étudié l'élimination des virus entériques dans des conditions de coagulation améliorées au moyen de doses de coagulant plus élevées. Abbaszadegan et coll. (2007) et Mayer et coll. (2008) ont signalé des éliminations logarithmiques maximales variant de 1,75 à 3,0 dans le cas des adénovirus, du calicivirus félin, du virus Coxsackie, de l'échovirus et du poliovirus dans le cadre d'essais de floculation au cours desquels on a optimisé l'élimination du COD en utilisant 40 mg/L de FeCl3 à un pH variant de 5 à 6. Les auteurs ont signalé que l'élimination des virus s'est améliorée en général à mesure que la dose de coagulant augmentait et que le pH diminuait. La température a aussi un effet sur l'élimination des particules, y compris les virus. En général, la formation de flocs et l'efficacité de l'élimination des particules diminuent à mesure que la température baisse (AWWA, 2011a).
Des études ont aussi démontré qu'il faut un effluent de filtration de 0,1 UTN ou moins pour maximiser la diminution des microorganismes pathogènes (Harrington et coll., 2001, 2003; Xagoraraki et coll., 2004). Dans le cadre d'études pilotes réalisées par Xagoraraki et coll. (2004), on a maximisé l'élimination logarithmique moyenne du bactériophage MS2 (médiane 3,2, plage de 2,5 à 3,6) lorsque la turbidité de l'eau était inférieure à 0,2 UTN comparativement aux éliminations (médiane de 2,4, plage de 0,25 à 3,5) obtenues lorsque la turbidité était inférieure à 0,3 UTN. Les périodes de maturation du filtre et de fin de vie du filtre sont aussi considérées comme des périodes de vulnérabilité à l'apparition des microorganismes pathogènes dans l'eau filtrée (Amburgey et coll., 2003; Soucie et Sheen, 2007). C'est pourquoi il faut surveiller les filtres de près et les laver soigneusement à contre-courant pour optimiser leur efficacité. Il est recommandé de ne pas faire circuler de nouveau l'eau du lavage à contre-courant des filtres dans la station de traitement sans d'abord lui faire subir un traitement supplémentaire. La recirculation de l'eau de lavage à contre-courant des filtres a été reconnue comme un facteur déterminant d'éclosions de Cryptosporidium au Royaume-Uni et pourrait avoir exacerbé l'éclosion de Cryptosporidium survenue à Milwaukee (Hrudey, 2017).
En résumé, la plupart des stations de filtration avec procédé chimique qui fonctionnent bien et sont optimisées pour éliminer les particules et les matières organiques naturelles (MON) devraient pouvoir produire une élimination moyenne de 2 log dans le cas des virus entériques (Payment et coll., 1985, Payment et Franco, 1993; Hendricks et coll., 2005; Santé Canada, 2012d). Pour y parvenir, il est essentiel que les étapes de coagulation et de floculation qui précèdent soient optimisées. Il faut procéder à des jars-tests pour optimiser le procédé de coagulation (U.S. EPA, 2004; AWWA, 2011b). Il peut aussi être utile de surveiller la charge superficielle nette des particules de la surface après la coagulation lorsque la qualité de la source d'approvisionnement en eau varie beaucoup (Conio et coll., 2002; Newcombe et Dixon, 2006; AWWA, 2011b; Kundert, 2014; McVicar et coll., 2015; Sharp, 2015).
7.1.2.2 Filtration lente sur sable
Au cours de la filtration lente sur sable, l'eau non traitée traverse lentement par gravité un lit de sable poreux submergé. Pendant la filtration, il y a croissance biologique dans le lit de sable et le substrat de gravier. Des bactéries et d'autres matières provenant de la source d'approvisionnement en eau s'accumulent alors à la surface pour former un « schmutzdecke », qui est un film biologique constitué d'une diversité d'organismes, tels que les algues, les protozoaires et les bactéries. Lorsque l'eau non traitée traverse le lit de sable, des mécanismes physiques, chimiques et biologiques éliminent les virus de l'eau (Anderson et coll., 2009). La fixation des virus au média filtrant et aux biofilms, ainsi que l'inactivation des virus par des procédés biologiques comme la prédation, sont reconnues comme principaux mécanismes d'élimination (Anderson et coll., 2009; Hijnen et Medema, 2010). Il est recommandé de procéder à un essai pilote pour vérifier que la filtration lente sur sable parviendra à traiter une source d'approvisionnement en eau (Bellamy et coll., 1985; Logdon et coll., 2002).
De nombreuses études pilotes, ainsi qu'un nombre limité d'études à échelle réelle portant sur des substituts ou des virus entériques, ont démontré que la filtration lente peut produire des éliminations de l'ordre de 0,6 à quatre log (Poytner et Slade, 1977; Slade, 1978; McConnell et coll., 1984; Hijnen et coll., 2004a; Anderson et coll., 2009; Hijnen et Medema, 2010; Bauer et coll., 2011; Schijven et coll., 2013).
Des études pilotes ont démontré que la température, l'âge du schmutzdecke et les charges hydrauliques constituent des paramètres opérationnels importants qui ont une incidence sur l'élimination des virus par filtration lente sur sable. Poynter et Slade (1977) ont signalé une élimination du poliovirus 1 pouvant atteindre un facteur de 4 log dans un filtre contenant 600 mm de sable, où le taux de charge hydraulique atteint 4,8 m/jour (0,2 m/h) et les températures varient de 16 à 18 °C. L'élimination est tombée à moins de 1 log lorsque le taux de charge a atteint 12 m/jour (0,5 m/h) et les températures ont oscillé entre 5 et 9 °C. Anderson et coll. (2009) ont signalé des taux d'élimination de MS2 variant de 1,5 à 2,2 log à la fois à l'eau froide (3 à 10 °C) et à l'eau plus chaude (21 à 22 °C) lorsque le taux de charge hydraulique s'établissait à 0,1 m/h (épaisseur du lit = 0,4 m). Les éliminations sont tombées entre 0,2 et 1,3 log lorsque le taux de charge hydraulique est passé à 0,4 m/h. Schijven et coll. (2013) ont montré récemment que l'âge du schmutzdecke constitue aussi un important paramètre opérationnel dans l'élimination des virus par filtration lente sur sable. Des essais pilotes réalisés à des températures se situant entre 10 et 16 °C et à un taux de filtration de 0,03 m/h ont révélé que l'élimination du MS2 passait de 1,6 log lorsque le schmutzdecke avait 4 jours à 3,1 log après 553 jours.
Slade (1978) a rapporté des concentrations de virus entériques dans l'eau traitée et non traitée provenant de deux filtres à sable lents à grande échelle et constaté des efficacités d'élimination similaires à celles signalées dans les études pilotes susmentionnées. Les éliminations de virus entériques (virus Coxsackie et poliovirus) ont varié de 1 à 2 log dans des filtres où l'épaisseur du lit variait de 0,3 à 0,45 m, les débits s'établissaient entre 1,12 et 4,15 m/jour (0,05 et 0,17 m/h) et les températures oscillaient entre 6 et 11 °C. On a observé une élimination de moins de 2 log dans plusieurs conditions, y compris aux températures plus basses et à des débits plus élevés (Slade, 1978).
En résumé, des filtres à sable à filtration lente conçus et exploités comme il se doit peuvent produire efficacement une élimination moyenne de 2,0 log dans le cas des virus entériques. Il est toutefois important de suivre les impacts potentiels des événements suivants lors de l'opération du filtre : températures plus basses, les augmentations du taux de charge hydraulique et le raclage d'entretien du filtre.
7.1.2.3 Filtration sur membrane
On utilise actuellement quatre types de membranes sous pression pour traiter l'eau potable : la microfiltration (MF), l'ultrafiltration (UF), la nanofiltration (NF) et l'osmose inverse (OI). Les membranes sont en général classées selon le type de substances qu'elles éliminent, la pression de fonctionnement et le diamètre des pores ou le seuil de rétention des molécules (SRM). Les membranes pour la MF et l'UF sont des membranes à basse pression qui sont utilisées habituellement pour éliminer des particules et des organismes pathogènes. La petite taille des virus entériques peut toutefois diminuer l'élimination comparativement à d'autres microorganismes pathogènes plus gros (p. ex. Cryptosporidium et Giardia). Les membranes NF et OI sont des membranes haute pression qui servent à éliminer des matières organiques (p. ex. MON dissoutes) et inorganiques (p. ex. sodium, chlorure, calcium et magnésium). Les grandes catégories de membranes, leurs dimensions et les substances éliminées sont analysées dans Kawamura (2000), AWWA (2005, 2011a) et MWH (2012).
Les mécanismes d'élimination des virus au moyen de membranes MF et UF comprennent l'exclusion en fonction de la taille, la répulsion électrostatique entre le virus et la membrane et l'adsorption du virus sur la membrane (Jacangelo et coll., 1995). L'exclusion en fonction de la taille constitue le principal mécanisme d'élimination lorsque les pores de la membrane sont plus petits que les virus présents dans la source d'approvisionnement en eau. Lorsque les pores de la membrane sont plus gros que le virus, la répulsion électrostatique et l'adsorption dominent. L'efficience de l'élimination dépendra à la fois des caractéristiques du virus et de celles de la membrane, y compris la charge superficielle, l'hydrophobicité, ainsi que la taille et la forme du virus (El Hadidy et coll., 2013). La filtration et l'adsorption attribuées à la présence d'une couche de particules qui se forme sur les membranes au cours du fonctionnement (agglutination) et l'encrassement irréversible jouent aussi un rôle dans l'élimination des virus au moyen de membranes MF/UF (Jacangelo et coll., 1995; ElHadidy et coll., 2013).
Les différences au niveau de la solubilité ou de la diffusivité constituent le principal mécanisme d'élimination des virus entériques à l'aide de membranes NF et OI. Comme les membranes NF et OI ne sont pas poreuses, elles peuvent toutefois filtrer aussi les microorganismes et les éliminer de l'eau d'alimentation (U.S. EPA, 2005), mais elles ne sont pas conçues pour que l'élimination soit vérifiable (ANSI/AWWA, 2010; Alspach, 2018).
On détermine habituellement le potentiel d'élimination des virus entériques au moyen d'un système de filtration à membrane en particulier en soumettant le système à une épreuve de provocation avec des substituts dans des conditions où l'élimination peut se rapprocher de celle des virus entériques. Les bactériophages MS2, PRD1 et Qß sont les substituts utilisés le plus souvent pour soumettre les membranes à des essais pour déterminer l'efficacité et ils ont produit le gros des données sur l'élimination présentées ci-dessous. Les efficiences de l'élimination tirées d'essais pilotes et à grande échelle menés à l'aide de substituts sont résumées dans U.S. EPA (2001d, 2005), AWWA (2005) et Antony et coll. (2012) et nous les abordons brièvement ci-dessous. Des données limitées sur l'élimination des virus entériques de l'eau potable au moyen de procédés à membrane sont aussi disponibles (Jacangelo et coll., 2006).
Les membranes MF ne constituent pas une barrière physique définitive pour les virus en raison du diamètre de leurs pores, qui varie habituellement de 0,05 à 5 µm. Des études pilotes ont toutefois révélé des éliminations de virus substituts (c.-à-d. MS2, Qß et PRD1) des eaux de surface, des eaux souterraines et de l'eau déionisée tamponnée qui variaient de 0,2 à 3,0 log en utilisant tout un éventail de membranes MF (Adham et Jacangelo, 1994; Jacangelo et coll., 1995; Jacangelo et coll., 1997; Kruithof et coll., 1997). À cause du manque d'uniformité dans l'élimination des virus par MF, plusieurs chercheurs ont étudié l'utilisation de la coagulation avant la MF afin d'améliorer l'élimination des virus. Des études ont démontré qu'il est possible d'éliminer les substituts à un niveau de 4 log ou plus lorsque la microfiltration est précédée d'une coagulation (Zhu et coll., 2005a, b; Fiksdal et Leiknes, 2006; Matsushita et coll., 2005, 2013).
Les membranes UF ont des pores d'un diamètre de 0,005 à 0,05 µm et peuvent, dans la plupart des cas, éliminer un grand nombre (> 4 log) de virus. On a décrit dans les publications un grand nombre d'études pilotes au cours desquelles on a utilisé tout un éventail de sources d'approvisionnement en eau, de matériaux pour les membranes et de modes d'exploitation. Les données présentées dans ces études indiquent que les membranes UF (SRM de 10 à 500 kilodaltons) peuvent produire une élimination variant de 3 à > 7 log de MS2 (Jacangelo, 1991; Jacangelo et coll., 1991; Adham et coll., 1995; Kruithof et coll., 1997; Jacangelo et coll., 2006). Se basant sur des données tirées de plus de 17 études pilotes, l'U.S. EPA (2001d) a signalé que des systèmes UF à membrane SRM de plus de 100 kilodaltons (kD) pouvaient souvent diminuer les concentrations des MS2 au-dessous de la limite de détection. Kruithof et coll. (2001) ont soumis à une épreuve de provocation une station de traitement des eaux de surface par ultrafiltration à grande échelle (15 MGD ou 68 190 m3/jour) et ont démontré des éliminations logarithmiques de MS2 variant de 4,8 à > 5,4 log. Jacangelo et coll. (2006) ont constaté que le MS2 constituait un bon substitut du poliovirus, du VHA et du calicivirus félin au cours de l'essai de membranes UF sur banc d'essai. Ces tests ont démontré que les membranes UF qui ont un SRM de 10 kD et de 100 kD pouvaient éliminer de 3 à > 5 log de poliovirus 1, de VHA et de calicivirus félin. Les données sur l'élimination à grande échelle des virus entériques par UF sont limitées. Qui et coll. (2015) ont signalé une élimination de 4,6 à 7,0 log dans le cas de plusieurs virus entériques, y compris le norovirus, le rotavirus, l'entérovirus et l'adénovirus, au moyen de membranes UF (pores de 0,04 µm de diamètre) dans une station de traitement des eaux usées à grande échelle.
On n'utilise pas habituellement les membranes NF et OI pour éliminer les particules et c'est pourquoi il y a moins de données disponibles pour démontrer leur capacité d'élimination des virus. Lovins et coll. (2002) ont indiqué une élimination variant de 5,4 à 6,8 log du MS2 au moyen de membranes NF (SRM de 100 à 300 daltons) dans une station pilote. De même, des essais pilotes réalisés par Lozier et coll. (2003) ont démontré une élimination > 6 log du MS2 au moyen de membranes OI (SRM non disponible) et variant de 3 à 5,5 log au moyen de membranes NF. Kruithof et coll. (2001) ont démontré une élimination de 3,0 à 4,8 log des MS2 au moyen de membranes OI au cours d'une épreuve de provocation réalisée dans une station de traitement à grande échelle.
Comme on l'a dit ci-dessus, des membranes UF, NF et OI intactes peuvent éliminer les virus dans une proportion de plus de 4 log. Des défaillances de l'intégrité peuvent toutefois compromettre l'efficacité des membranes et c'est pourquoi il est crucial d'en contrôler régulièrement l'intégrité. Plusieurs études ont démontré que des membranes endommagées (p. ex. piqûre dans la membrane, joint torique fissuré, etc.) peuvent diminuer de 0,4 à 3,8 log l'élimination logarithmique du MS2 (Kruithof et coll., 2001; Kitis et coll., 2003; El Hadidy et coll., 2014). On a aussi signalé que des couches d'encrassement peuvent diminuer en partie la capacité des membranes d'éliminer les virus même en présence de fissures (Kitis et coll., 2003; El Hadidy et coll., 2014). On trouve dans Guo et coll. (2010a) et Antony et coll. (2012) des analyses détaillées de méthodes de surveillance de l'intégrité du procédé à membrane.
En pratique, l'utilisation de la filtration à membrane pour produire des crédits d'élimination logarithmique des virus est limitée à cause des difficultés que pose l'utilisation de méthodes courantes de tests d'intégrité directs afin de détecter une défaillance qui est de la grosseur du virus (U.S. EPA, 2005; Antony et coll., 2012). On trouve dans d'autres sources (Jacangelo et coll., 1997; Lozier et coll., 2003; U.S. EPA, 2005; Kumar et coll., 2007; Guo et coll., 2010a, Pype et coll., 2016) une analyse des diverses méthodes de vérification directe et indirecte de l'intégrité dans les systèmes à membrane à basse et à haute pression. Il existe en outre des méthodes d'analyse normalisées (ASTM, 2010, 2014). Une minuscule défaillance de l'intégrité non détectée par des méthodes existantes de vérification de l'intégrité pourrait laisser passer des virus, ce qui a une incidence sur la qualité du filtrat. Des méthodes existantes de vérification de l'intégrité donnent de bons résultats dans le cas des systèmes à membrane à basse ou à haute pression, mais seulement dans le cas des particules dont la taille dépasse 0,4 à 1 μm (c.-à-d. bactéries et protozoaires) (U.S. EPA, 2005; Antony et coll., 2012). Par exemple, pour qu'un essai d'étanchéité ait une résolution suffisante pour détecter une défaillance de la taille du virus dans une membrane à fibre creuse de polymère, la pression transmembranaire qu'il faudrait appliquer provoquerait la rupture de la membrane (U.S. EPA, 2005). Même si l'élimination des virus démontrée par des essais pour déterminer l'efficacité auxquelles on soumet des systèmes à membrane peut être très efficiente, il est difficile de vérifier cette élimination pendant l'utilisation des membranes. C'est pourquoi Antony et coll. (2012) ont déterminé que dans la plupart des cas, il est difficile d'attribuer aux membranes une élimination de > 2 log dans le cas des virus. Des technologies émergentes comme les tests d'intégrité à impulsions utilisant un colorant fluorescent ou des sondes nanométriques peuvent permettre de vérifier l'élimination de > 2 log dans le cas des virus (Pyper et coll., 2016).
En résumé, la filtration sur membrane est un procédé de traitement de l'eau potable bien établi qui permet d'éliminer certains virus. À l'heure actuelle, la démonstration de l'intégrité de la membrane et la vérification de l'élimination logarithmique des virus comportent des limites. Il faut donc communiquer avec l'autorité responsable en matière d'eau potable dans le secteur de compétence concerné pour déterminer si l'élimination logarithmique des virus serait attribuée pour la filtration sur membrane. Les services d'approvisionnement en eau qui envisagent de recourir à la filtration sur membrane doivent savoir que l'épreuve de provocation est généralement effectuée par des fabricants de membranes faisant appel à un tiers. Par conséquent, les résultats de la certification par un tiers devraient être demandés auprès du fabricant.
La nouvelle norme 419 de la NSF International/American National Standards Institute (NSF/ANSI) comprend des exigences de performance minimales pour l'élimination de Cryptosporidium par filtration sur membrane, mais ne comporte aucune exigence de ce type pour l'élimination des virus. Par conséquent, les membranes ne peuvent pas être certifiées à cette fin. Toutefois, les procédures d'analyse font appel à des essais pour déterminer l'efficacité utilisant le coliphage MS-2, qui est considéré comme un substitut prudent de Cryptosporidium. Lorsque cette procédure d'essai est utilisée, les fabricants peuvent inclure à leur liste l'efficacité d'élimination des virus obtenue pour les membranes UF certifiée pour l'élimination de Cryptosporidium selon cette norme (NSF/ANSI, 2015).
Il convient de signaler que toutes les membranes s'encrassent lorsqu'elles sont utilisées et que, par conséquent, le flux (débit par unité de surface) pour une différence de pression donnée peut diminuer considérablement (AWWA, 2005; MWH, 2012). Pour venir à bout de l'encrassement des membranes, il faut procéder à un lavage à contre-courant régulier et à un nettoyage chimique périodique, réalisé à l'aide de produits spécialement formulés. Santé Canada recommande fortement que les produits de nettoyage soient certifiés par un organisme de certification accrédité selon la norme 60 de la NSF/ANSI (NSF/ANSI, 2017). Lorsqu'il n'est plus possible de ramener le flux à des conditions acceptables, il faut remplacer les membranes (Alspach et coll., 2014).
7.1.2.4 Filtration in situ
Le passage dans le sol par filtration in situ naturelle est considéré comme une barrière importante pour les microorganismes. Si l'on suppose que la trajectoire de l'écoulement et le temps de transport sont suffisants, la filtration in situ naturelle peut améliorer la qualité microbienne de l'eau suffisamment pour protéger la santé publique (Schijven et coll., 2002). Des études publiées font état de diminutions logarithmiques des virus dans différents contextes naturels, dont la filtration sur berge de cours d'eau, et le schmutzdecke au fond d'un étang (c.-à-d. filtration sur berge d'étendue d'eau). La filtration sur berge de rivière ou de lac oblige à installer des puits d'approvisionnement en eau verticaux ou horizontaux près d'une rivière ou d'un lac afin d'utiliser la berge et l'aquifère adjacent comme filtre naturel pour éliminer les particules, les microorganismes pathogènes et d'autres contaminants (Kuehn et Mueller, 2000; Ray et coll., 2002a, 2002 b; Sharma et Amy, 2011; Harvey et coll., 2015). Dans le cas de beaucoup d'études, on a utilisé des coliphages ou des bactériophages non pathogènes pour mesurer les distances de transport (voir la section 5.2). Ces substituts ont tendance à ne pas adsorber aussi fortement que les virus pathogènes, parce que leur point isoélectrique est en général plus bas. Ils sont aussi faciles à mesurer et il est possible de les injecter dans l'environnement (Bales et coll., 1989; Havelaar et coll., 1993; Schijven et Hassanizadeh, 2000; Tufenkji et Emelko, 2011).
Havelaar et coll. (1995) ont signalé une diminution de 4 log des entérovirus et des réovirus dans des puits situés de 25 à 30 m des rivières. Les auteurs ont signalé que la diminution était plus importante que prévu dans le cas des déplacements sur de courtes distances et de brève durée (2 à 10 semaines). L'effet bénéfique a été attribué à l'élimination de solides en suspension qui adsorbent la majorité des virus. Schijven et coll. (1998) ont fait état d'une diminution de 3,8 log de bactériophages à ARN F-spécifiques dans des puits situés à 2 m de bassins d'infiltration des eaux de surface. Les auteurs ont attribué la diminution à la petite taille des grains de sable et à une grande efficience de collision. Schijven et coll. (1999) ont fait état d'une diminution de 3 log des bactériophages PRD1 et des coliphages MS2 dans des puits situés à 2,4 m de bassins d'infiltration des eaux de surface et une diminution supplémentaire de 5 log dans le cas des puits situés à moins de 30 m. Les auteurs ont signalé que l'adsorption constituait le principal procédé à cause de la présence de fer extractible par oxalate et d'une forte concentration de cations divalents. Les auteurs ont conclu qu'il serait possible de porter la diminution à 8 log à moins de 30 m, ce qui correspond à un temps de déplacement d'environ 25 jours. Borchardt et coll. (2004) n'ont pu établir de lien entre la présence de virus et l'apport en eau de surface dans des puits situés à diverses distances de la rivière Mississippi, parce que tous les puits contenaient des virus sans égard à la contribution des eaux de surface. Le puits situé le plus près de la rivière et recevant le plus fort volume d'eau de surface a produit des échantillons contenant des virus dans quatre cas sur 12 échantillons prélevés en un an tandis que deux puits situés plus loin de la rivière auxquels les eaux de surface n'ont apporté aucune contribution détectable ont produit des échantillons contenant des virus dans cinq et six échantillons sur 12 respectivement. Dans le cas d'un autre puits recevant un volume intermédiaire d'eau de surface, neuf échantillons sur 12 contenaient des virus. On a signalé que le puits situé le plus près de la rivière se trouvait dans un secteur non résidentiel tandis que les autres se trouvaient dans des secteurs résidentiels contenant un réseau souterrain d'égouts sanitaires. Les auteurs ont conclu que le transport du virus dans les puits était probablement attribuable à l'infiltration des eaux de surface et à des fuites des égouts sanitaires. Ils ont prévenu qu'il se peut que les puits situés dans les secteurs urbains risquent davantage d'être contaminés par des virus à cause de fuites des égouts sanitaires.
Harvey et coll. (2015) ont signalé une diminution de > 3 log du coliphage MS2 dans un film biologique de 25 cm d'épaisseur au fond d'un étang. Les auteurs ont conclu que le film biologique réussissait très efficacement à éliminer les virus. Dizer et coll. (2004) ont tiré des conclusions semblables au sujet des bienfaits d'un film biologique.
7.1.2.5 Crédits d'élimination mécanique logarithmique selon les procédés de traitement
Les stations de traitement de l'eau potable qui respectent les limites de turbidité établies dans le document technique de la recommandation sur la turbidité (Santé Canada, 2012d) peuvent appliquer les crédits d'élimination moyens des virus entériques qui sont présentés au tableau 3. On peut aussi fonder les taux d'élimination logarithmique sur l'efficacité observée ou des études pilotes. En ce qui concerne la filtration in situ (p. ex. filtration sur berge de rivière ou de lac), il faudrait consulter l'autorité responsable en matière d'eau potable dans le secteur de compétence concerné pour connaître les exigences propres au site. Même si la littérature démontre que la filtration in situ a la capacité de diminuer les concentrations de virus, la surveillance de la qualité de l'eau peut être nécessaire durant l'évaluation des risques pour déterminer les objectifs de diminution logarithmique. Sinclair et coll. (2012) décrivent un processus de sélection d'un ou de plusieurs paramètres de surveillance pour les systèmes naturels ou d'origine humaine.
Il est possible d'atteindre les objectifs globaux du traitement des virus en combinant les crédits d'élimination mécanique logarithmique et les crédits de désinfection (voir la section 7.1.3). Par exemple, s'il faut diminuer globalement les virus de 6 log (99,9999 %) dans le cas d'un approvisionnement en eau de surface et si la filtration classique en élimine 2 log, il faut alors obtenir la diminution de 4 log qui reste en utilisant un autre procédé comme la désinfection, tout en cherchant à atteindre les objectifs du traitement dans le cas des protozoaires entériques. De même, une source ESSIDES devrait atteindre les objectifs de traitement dans le cas des virus entériques et des protozoaires entériques en combinant l'élimination mécanique logarithmique (naturelle ou technique) et les crédits de désinfection. Les sources d'approvisionnement en eau d'alimentation souterraines jugées vulnérables seulement parviendraient habituellement à une diminution de 4 log des virus basée sur les crédits de désinfection (voir la section 7.1.3).
| Procédé de traitement | Crédit d'élimination de virusTableau 3 - note a (log10) |
|---|---|
| Filtration classique | 2,0 |
| Filtration directe | 1,0 |
| Slow sand filtration | 2,0 |
| Filtration sur diatomées | 1,0 |
| Microfiltration | Les membranes de microfiltration peuvent être admissibles au crédit d'élimination des virus lorsqu'elles sont précédées d'une étape de coagulation. L'efficacité de l'élimination est démontrée par des tests de provocation et vérifiée par des tests de vérification directe de l'intégritéTableau 3 - note b |
| Ultrafiltration | L'efficacité de l'élimination est démontrée par des tests de provocation et vérifiée par des tests de vérification directe de l'intégritéTableau 3 - note b |
| Nanofiltration et osmose inverse | L'efficacité de l'élimination est démontrée par des tests de provocation et vérifiée par des tests de vérification directe de l'intégritéTableau 3 - note c |
| Filtration in situ | Exigences propres à chaque site, déterminées par l'autorité compétente en matière d'eau potableTableau 3 - note d |
|
|
7.1.3 Inactivation
Il est possible d'utiliser la désinfection primaire pour produire une diminution logarithmique des virus entériques et elle est en général effectuée après les traitements qui retirent les particules et les MON. Cette stratégie aide à assurer l'inactivation efficiente des pathogènes et réduit au minimum la formation de sous-produits de désinfection (SPD). Il est important de signaler que, dans la description de la désinfection microbienne de l'eau potable, le terme « inactivation » indique que le microorganisme pathogène n'est pas infectieux et ne peut se répliquer dans un hôte convenable, mais qu'il peut encore être présent. L'inactivation des virus par désinfection subit l'effet de tout un éventail de facteurs, dont le type et l'état physiologique du virus et le type de désinfectant. En outre, les paramètres chimiques et physiques de la qualité de l'eau comme le pH, la température, les éléments constituants inorganiques et organiques, sans oublier les conditions hydrauliques comme la conception du réacteur et les conditions de mélange, ont aussi un effet sur l'efficience de la désinfection (Sobsey, 1989).
Cinq désinfectants sont couramment utilisés pour traiter l'eau potable : le chlore libre, la monochloramine, l'ozone, le dioxyde de chlore et les rayons UV. À l'exception de ces derniers, basés sur la radiation électromagnétique, ce sont tous des agents oxydants. Les désinfectants chimiques inactivent les microorganismes en détruisant ou en endommageant la structure de leurs cellules, leur métabolisme, leur biosynthèse et leur croissance, alors que les rayons UV endommagent l'acide nucléique des pathogènes, ce qui les empêche de se reproduire et les rend par conséquent incapables de compléter des cycles infectieux.
Le chlore libre est le produit chimique le plus souvent utilisé pour la désinfection primaire, parce qu'il est facile de s'en procurer, qu'il est relativement peu coûteux et qu'il fournit un résiduel qui peut également être utilisé pour la désinfection secondaire afin de maintenir la qualité de l'eau dans le réseau de distribution. Le chlore libre est un oxydant efficace pour l'inactivation des virus entériques (U.S. EPA, 1991). À titre exemple, une concentration modérée de chlore de 0,5 mg/L et un temps de contact de 15 minutes permettent d'obtenir une inactivation des virus de 4 log à 7 °C. L'utilisation de la monochloramine est en général limitée à la désinfection secondaire (c.-à-d. l'entretien résiduel) car son pouvoir oxydant est faible. L'ozone et le dioxyde de chlore sont des désinfectants efficaces contre les virus entériques, mais ils coûtent habituellement plus cher et sont plus difficiles à appliquer, en particulier dans les petits systèmes. L'ozone se dégrade toutefois facilement après l'application et ne produit pas de résiduel pour la désinfection secondaire (Santé Canada, 2008). La désinfection aux rayons UV est efficace pour inactiver la plupart des virus entériques à l'exception de l'adénovirus, dont l'inactivation nécessite une forte dose. Les UV ne fournissent aucun résiduel pour la désinfection secondaire.
7.1.3.1 Désinfection chimique
Il est possible de prédire l'efficacité des désinfectants chimiques en fonction de la concentration résiduelle d'un désinfectant en particulier et des facteurs qui jouent sur son efficacité, surtout la température, le pH, le temps de contact et le degré de désinfection requis (U.S. EPA, 1991). Cette relation est appelée couramment « concept CT », où CT représente le produit de « C » (concentration résiduelle d'un désinfectant, en mg/L) par « T » (temps de contact avec le désinfectant, en minutes) pour un microorganisme en particulier dans des conditions définies (p. ex. température et pH). Pour tenir compte de la dégradation du désinfectant, on détermine habituellement la concentration résiduelle à la sortie de la chambre de contact au lieu d'utiliser la dose appliquée ou la concentration initiale.
On calcule généralement le temps de contact T en utilisant une valeur T10 qui est définie comme le temps de séjour durant lequel l'eau atteint ou dépasse le temps de contact requis dans une proportion de 90 %. On peut estimer cette valeur T10 en multipliant la durée de rétention (c.-à-d. le volume du réservoir divisé par le débit) par l'efficacité hydraulique de la chambre de contact. L'U.S. EPA (1991) fournit les facteurs de déflexion pour des exemples de chambres de contact. L'exposition médiane à la désinfection (CT50) et les réacteurs à fonctionnement continu reliés en série (N-CSTR) sont d'autres méthodes permettant d'évaluer l'efficacité de la désinfection (Tfaily et coll., 2015). On peut aussi réaliser un essai de traçage hydraulique pour déterminer le temps de contact réel dans les conditions de débit de la station. Comme la valeur T dépend des caractéristiques hydrauliques liées à la construction de la station de traitement, il est plus efficace d'améliorer les caractéristiques hydrauliques (c.-à-d. augmenter le facteur d'efficacité hydraulique de la chambre de contact) que d'augmenter la dose de désinfectant pour respecter les exigences relatives au facteur CT. L'amélioration de l'efficacité hydraulique d'une chambre de contact au chlore peut s'avérer une stratégie de contrôle importante pour combler toute lacune dans les crédits d'inactivation des virus et améliorer la fiabilité du procédé de désinfection (Douglas, 2016).
Le document de l'U.S. EPA (1991) présente des tableaux CT pour l'inactivation de 2 log, 3 log et 4 log des virus. Le tableau 4 présente certaines valeurs CT pour l'inactivation de 4 log (99,99 %) des virus entériques avec le chlore, la chloramine, le dioxyde de chlore et l'ozone. Ces valeurs indiquent que la chloramine est un désinfectant beaucoup plus faible que le chlore libre, le dioxyde de chlore et l'ozone, puisqu'il faut des concentrations ou des temps de contact beaucoup plus élevés pour atteindre le même taux d'inactivation. La chloramine n'est donc pas recommandée comme désinfectant primaire.
| Température(°C) | Valeurs CT (mg-min/L) pour une inactivation de 99,99 % (4 log) | |||
|---|---|---|---|---|
| Chlore libre (Cl2) | Chloramine (NH2Cl) |
Dioxyde de chlore (ClO2) |
Ozone (O3) |
|
| 5 | 8 | 1988 | 33,4 | 1,2 |
| 20 | 3 | 746 | 12,5 | 0,5 |
|
||||
Des études portant sur plusieurs virus entériques révèlent des degrés différents de résistance aux désinfectants chimiques (Engelbrecht et coll., 1980; Payment et coll., 1985; Hoff, 1986; Sobsey et coll., 1988; Payment et Armon, 1989; U.S. EPA, 1989; AWWA, 1999, 2011a; Thurston-Enriquez et coll., 2003a, 2005a,b). Ces études ont révélé que le VHA résistait davantage à l'inactivation au dioxyde de chlore et à l'ozone que d'autres types de virus. Le VHA résistait plus au chlore libre que le rotavirus et l'adénovirus 40, mais la vulnérabilité du virus Coxsackie B5 et du poliovirus de type 1 variait considérablement d'une étude à l'autre et l'on a indiqué à l'occasion qu'ils étaient plus résistants que le VHA. Les objectifs de désinfection et les tableaux des valeurs CT sont donc fondés sur des valeurs s'appliquant au VHA à l'exception de l'ozone, qui repose sur l'inactivation du poliovirus (U.S. EPA, 1991). Le tableau 5 présente les valeurs CT tirées de diverses études où divers désinfectants chimiques ont produit une inactivation de 2 log (99 %) de plusieurs virus.
Comme il est impossible de cultiver le norovirus humain, on a utilisé des substituts comme le calicivirus félin et le norovirus murin pour calculer les valeurs CT requises afin d'inactiver efficacement le norovirus humain dans l'eau potable. Lim et coll. (2010) ont signalé des valeurs CT de 0,141 et de 0,071 mg-min/L dans le cas du norovirus murin lorsqu'on utilise du chlore et du dioxyde de chlore (5 °C et pH de 7,2). Les auteurs ont signalé que ces résultats indiquent que le norovirus murin est beaucoup moins résistant que d'autres virus entériques (p. ex. poliovirus de type 1). Ils ont de plus laissé entendre que les valeurs CT recommandées couramment pour l'inactivation du virus au moyen du chlore et du dioxyde de chlore (tableau 5) suffisent pour produire une diminution de 4 log du norovirus humain. Des dosages RT-PCR qui ont démontré que le norovirus est moins résistant à la désinfection au chlore que le poliovirus type 1 appuient ces conclusions (Shin et Sobsey, 2008; Kitajima et coll., 2010b).
Les caractéristiques physiques de l'eau, comme la température, le pH et la turbidité, peuvent avoir un effet important sur l'inactivation et l'élimination des pathogènes. Les taux d'inactivation, par exemple, doublent ou triplent par tranche de 10 °C d'élévation de la température. Lorsque les températures approchent 0 °C, comme c'est souvent le cas en hiver au Canada, l'efficacité de la désinfection diminue et il faut augmenter la concentration de désinfectant ou le temps de contact (CT) avec celui-ci, ou les deux, pour atteindre le même degré d'inactivation. L'efficacité de certains désinfectants varie également selon le pH. Lorsque l'on emploie du chlore libre, une augmentation du pH de 6 à 10 réduit de 8 à 10 fois le taux d'inactivation (U.S. EPA, 1991). De même, la désinfection par la chloramine diminue à mesure que le pH augmente (Cromeans et coll., 2010; Kahler et coll., 2011). Par contre, Thurston-Enriquez et coll. (2005a) ont signalé que le dioxyde de chlore est 1,9 et 19,3 fois plus efficace à un pH de 8 qu'à un pH de 6 pour l'adénovirus 40 et le calicivirus félin, respectivement. On a fait état de résultats semblables pour d'autres virus entériques désinfectés au dioxyde de chlore (Alvarez et O'Brien, 1982; Noss et Olivieri, 1985). On a démontré que le pH avait peu d'effet sur l'efficacité de l'inactivation des virus par l'ozone, même si un pH plus élevé a un effet sur la stabilité de l'ozone et, par conséquent, augmente la demande en ozone.
La diminution de la turbidité est un préalable important pour l'inactivation des virus et d'autres microorganismes. La désinfection chimique peut être inhibée, car les particules auxquelles ils sont liés peuvent protéger les virus et d'autres microorganismes. De plus, la turbidité neutralise l'action des désinfectants et réduit l'efficacité de la désinfection chimique. Selon LeChevallier et coll. (1981), le pouvoir désinfectant du chlore libre diminue de huit fois lorsque la turbidité passe de 1 à 10 UTN. On a observé des effets négatifs des virus liés à des particules sur les procédés de désinfection au cours de plusieurs études (Templeton et coll., 2008). Le document technique de la recommandation sur la turbidité (Santé Canada, 2012d) traite plus en détail de l'effet de la turbidité sur l'efficience du traitement.
Virus |
Valeurs CT pour une inactivation de 2 log (99 %) | |||
|---|---|---|---|---|
| Chlore libre (Cl2) pH 6–7 |
Chloramine (NH2Cl) pH 8–9 |
Dioxyde de chlore (ClO2) pH 6–7 |
Ozone (O3) pH 6–7 |
|
| Poliovirus 1note a,note b,note c | 1,1–6 | 768–3740 | 0,2–6,7 | 0,1–0,2 |
| Rotavirusnote c | 0,01–0,05 | 3806–6476 | 0,2–2,1 | 0,006–0,06 |
| Virus de l'hépatite Anote d | 0,7–1,18 | 428–857 | <0,17–2,8 | 0,5 |
| Virus Coxsackie B5note a,note b,note d,note f | 1,7–12 | 550 | n.d. | n.d. |
| Adénovirus 40note e,note f | 0,02–2,4 | 360 | 0,25 | 0,027 |
|
||||
La désinfection chimique peut entraîner la formation de sous-produits, dont certains présentent un risque pour la santé. Le chlore, le désinfectant utilisé le plus couramment, réagit avec la MON pour former des trihalométhanes (THM) et des acides haloacétiques (AHA), ainsi que d'autres composés organiques halogénés (Rook, 1976; Krasner et coll., 2006). Les systèmes qui utilisent la monochloramine et, dans une moindre mesure, ceux qui utilisent le chlore libre peuvent aussi former de la N-diméthylnitrosamine (NDMA). Du bromate peut se former dans les systèmes qui utilisent des solutions d'hypochlorite produites sur les lieux ou achetées dans le commerce (Santé Canada, 2016). L'utilisation de dioxyde de chlore et d'ozone peut aussi entraîner la formation de SPD inorganiques, comme le chlorite/chlorate et le bromate respectivement. Il faut tenir compte de l'effet éventuel des SPD lorsqu'on choisit un désinfectant chimique, mais il est essentiel que les tentatives de diminution de la formation de sous-produits ne nuisent pas à l'efficacité de la désinfection. Cette condition est cruciale, car la certitude de contracter une maladie humaine causée par une désinfection inefficace dépasse largement celle des effets indésirables sur la santé causés par l'exposition de l'eau potable à des SPD (Hrudey, 2017). Les recommandations appropriées offrent plus de renseignements à ce sujet (Santé Canada, 2006, 2008a, 2008 b, 2011, 2016).
7.1.3.2 Désinfection aux rayons ultraviolets
Dans le cas de la désinfection aux rayons UV, le produit de l'intensité lumineuse « I » (mesurée en mW/cm2 ou W/cm2) et du temps « T » (mesuré en secondes) est la dose calculée (fluence) en mJ/cm2 pour un microorganisme donné. Cette relation est appelée concept IT. La désinfection aux rayons UV s'effectue à l'aide de lampes à basse pression (BP) qui émettent des rayons UV essentiellement sur une seule longueur d'onde (~254 nm) (monochromatiques), ou de lampes à pression moyenne (PM), qui émettent des rayons sur un spectre plus large (polychromatique). Les diodes électroluminescentes aux ultraviolets (DEL-UV) constituent une nouvelle technologie pour le traitement des eaux aux rayons UV (Wright et coll., 2012). Dans une critique d'études publiées sur l'application des DEL-UV, Song et coll. (2016) ont toutefois conclu qu'il faut une méthode normalisée de détermination de la dose de rayons UV émise par les DEL-UV afin de réduire les données incohérentes et incomparables sur la dose-réponse actuellement disponible dans les publications.
On a effectué de nombreuses études en laboratoire dans des conditions bien précises pour déterminer la sensibilité des virus aux rayons UV monochromatiques (BP). Le tableau 6 résume la plage des doses de rayons UV requises pour produire divers niveaux d'inactivation au moyen de lampes BP. Les données indiquent que dans la plupart des cas, sauf dans celui de l'adénovirus, il est possible de produire une inactivation de 4 log des virus en appliquant une dose de rayons UV BP de 40 mJ/cm2 (Chang et coll., 1985; Arnold et Rainbow, 1996; Meng et Gerba, 1996; AWWA, 2011a; U.S. EPA, 2000; Cotton et coll., 2001; Hofmann, 2015). En se fondant sur une analyse de données dose-réponse UV BP publiées, Hijnen et Medema (2010) ont signalé, pour le poliovirus, le rotavirus, le calicivirus, le virus de l'hépatite A et le virus Coxsackie, des constantes du taux d'inactivation variant de 0,10 à 0,19 mJ/cm2. Ils ont aussi signalé des inactivations maximales de 4,1 à 5,7 log à des doses de rayons UV de 28 à 50 mJ/cm2 (Hijnen et Medema, 2010). Chevrefils et coll., Hijnen et Medema (2010) et Hofmann (2015) présentent des tableaux détaillés sur les doses de rayons UV et les constantes du taux d'inactivation pour divers virus.
Comme l'indique le tableau 6, les adénovirus résistent beaucoup plus que d'autres virus entériques à la désinfection aux rayons UV monochromatiques (BP) (Eischeid et coll., 2009; Cotton et coll., 2001; Thurston-Enriquez et coll., 2003b; Nwachuku et coll., 2005; Guo et coll., 2010b; Rochelle et coll., 2011; Beck et coll., 2014; Sangsanont et coll., 2014). Les données tirées de ces études indiquent qu'il faut des doses variant de 82 à 261 mJ/cm2 pour produire une inactivation de 4 log (99,99 %) de l'adénovirus 2, 5, 40 et 41 dans une eau tamponnée sans demande de chlore. Il semble que les virus à ADN bicaténaire, comme les adénovirus, résistent davantage aux rayons UV que les virus à ARN monocaténaire (comme le VHA) (Meng et Gerba, 1996; Calgua et coll., 2014). Guo et coll. (2010 b) ont remarqué que la résistance de l'adénovirus aux rayons UV est attribuable non pas à la résistance aux dommages initiaux causés à l'ADN par les rayons UV, mais plutôt à la capacité de la cellule infectée de réparer l'ADN de l'adénovirus pour le rendre de nouveau virulent.
Plusieurs études ont démontré une inactivation plus importante de l'adénovirus, du rotavirus et des calicivirus par les rayons UV à PM comparativement aux rayons à BP (Malley et coll., 2004; Linden et coll., 2009; Beck et coll., 2014). Linden et coll. (2007) ont produit une inactivation de 3 log de l'adénovirus 40 au moyen d'une source de rayons UV polychromatiques à une dose d'environ 30 mJ/cm2 et à des longueurs d'onde de 220 et 228 nm. On a obtenu une inactivation de 4 log des rotavirus à une dose moyenne de rayons UV de 40 mJ/cm2. Beck et coll. (2014) ont démontré que des doses variant de 9 à 25 mJ/cm2 à des longueurs d'onde de 210, 220 et 230 nm pouvaient produire une inactivation de 4 log de l'adénovirus 2. D'autres études ont montré qu'il faut des doses variant de 22 à 117 mJ/cm2 pour inactiver l'adénovirus 2, 5, 40 ou 41 au moyen de lampes à PM (Eischeid et coll., 2009; Linden et coll., 2007, 2009; Shin et coll., 2009; Guo et coll., 2010b; Sangsanont et coll., 2014).
Il est démontré que l'adénovirus est plus sensible que des microbes de validation type (c.-à-d. MS2) aux rayons UV à faible longueur d'onde (Linden et coll., 2015). Il peut donc être utile pour les services publics qui utilisent des lampes à PM d'inclure de basses longueurs d'onde dans leur calcul de l'efficacité de la désinfection aux rayons UV. Beck et coll. (2014) ont calculé des facteurs de correction des spectres d'intervention et des méthodes de mise en œuvre pour aider ces services publics à atteindre les objectifs d'inactivation des virus nécessaires pour se conformer à la réglementation, mais les technologies actuelles à capteurs UV ne permettent pas de surveiller les basses longueurs d'onde dans un réacteur UV. On est en train de mettre au point une technologie pour faciliter la surveillance de basses longueurs d'onde UV.
Dans les réseaux municipaux d'approvisionnement en eau au Canada, une dose de rayons UV de 40 mJ/cm2 suffit habituellement pour produire une inactivation de 4 log de nombreux virus entériques, y compris le poliovirus, le rotavirus, les calicivirus, le virus de l'hépatite A et les virus Coxsackie. Toutefois, une dose de rayons UV de 40 mJ/cm2 administrée à l'aide de lampes à BP ne produirait pas une inactivation logarithmique des adénovirus. Il est possible de combiner les rayons UV à d'autres technologies d'inactivation (c.-à-d. stratégie à désinfectants multiples) pour obtenir une diminution de 4 log des virus. Au cours d'une étude en laboratoire, par exemple, Baxter et coll. (2007) ont constaté qu'une concentration de 0,22 mg/L de chlore conjuguée à une période de contact variant d'une minute dans une eau sans demande de chlore (pH = 8,5, T = 5 °C) produisait une inactivation de 4 log de l'adénovirus. Rattanakul et coll. (2015) ont signalé qu'il est possible de produire une inactivation de 4 log de l'adénovirus 5 à des doses de rayons UV de lampes à BP pouvant atteindre 50 mJ/cm2 suivies d'une dose de chlore de 0,15 mg/L combinée à une durée de contact de 40 secondes (pH = 7,2, T = 20 °C).
Par ailleurs, l'autorité responsable en matière d'eau potable dans le secteur de compétence concerné peut choisir un virus entérique comme le rotavirus comme organisme cible (c.-à-d. celui qu'on trouve au tableau 6), parce que même si l'adénovirus a été détecté dans l'eau, on n'a pas établi de lien avec les cas de MGA (Borchardt et coll., 2012). Il faut donc communiquer avec l'autorité responsable en matière d'eau potable dans le secteur de compétence concerné pour confirmer quel virus devrait être ciblé pour le traitement par rayons UV, puisque cela aura une incidence sur les exigences relatives à la dose.
| Virus | Doses de rayons UV requises (mJ/cm2) | |||
|---|---|---|---|---|
| 1-log | 2-log | 3-log | 4-log | |
| Virus de l'hépatite A | 4,1–5,5 | 8,2–13,7 | 12,3–22 | 16,4–29,6 |
| Virus Coxsackie B5, B6 | 6,9–9,5 | 13,7–18 | 20,6–27 | 36 |
| Poliovirus de type 1 | 4,0–8 | 8,7–15,5 | 14,2–29 | 20,6–38 |
| Rotavirus SA-11Footnote b, RVH-Wa | 7,1–10 | 14,8–26 | 23–44 | 36–61 |
| Adénovirus 2, 5, 40, 41 | 10–76 | 26–137 | 39–199 | 51–261 |
|
||||
En pratique, la dose de rayons UV administrée dans les stations de traitement à grande échelle dépend d'un certain nombre de facteurs, dont le profil hydraulique du réacteur, le débit, le facteur de transmittance des rayons UV de l'eau, l'intensité des rayons UV, la puissance lumineuse des lampes, l'emplacement des lampes, l'âge des lampes, l'encrassement et la cinétique de l'inactivation des microbes (U.S. EPA, 2006 b; Bolton et Cotton, 2008). Il faut effectuer des tests de validation visant à déterminer les conditions de fonctionnement dans lesquelles le réacteur produira la dose de rayons UV. Plusieurs stratégies différentes d'essais de validation des rayons UV sont disponibles et abordées dans ONORM (2001, 2003) et dans U.S. EPA (2006 b). Ces stratégies sont basées sur des tests biodosimétriques visant à déterminer l'inactivation logarithmique de microorganismes de provocation en particulier pour un réacteur donné en combinaison avec des liens fluence-réponse connus. À partir de ces données, il est possible de déterminer une fluence équivalente correspondante pour un système en particulier. Il faut également tenir compte des conditions de températures minimales, car une température basse peut réduire l'intensité lumineuse « I » et modifier les seuils de déclenchement de l'alarme. Le système doit être bien conçu pour ces conditions, sinon une hausse de valeur « T » peut être nécessaire dans des conditions de températures minimales (Oosterveld, 2017).
Il faut constamment surveiller le réacteur au moyen de capteurs étalonnés périodiquement pour vérifier qu'il reste dans les limites des conditions validées et qu'il produit la dose requise. Il est possible de vérifier encore davantage l'efficacité de systèmes UV à grande échelle en contrôlant l'inactivation de spores environnementales de clostridium sulfitoréducteur (Hijnen et coll., 2004b). Il faut aussi tenir compte de questions opérationnelles afin que l'efficacité du réacteur ne soit pas compromise (p. ex. mise en marche, panne, encrassement et nettoyage des lampes, entretien des capteurs UV) (U.S. EPA, 2006 b). En outre, l'accumulation de matières sur les gaines de quartz (c.-à-d. l'encrassement) peut nuire au processus de désinfection en absorbant le rayonnement qui, autrement, servirait à inactiver les agents pathogènes (Wait et coll., 2005, 2007; Wait et Blatchley, 2010). Le fer et le calcium ont tendance à constituer la majeure partie de la matière encrassante, bien que l'aluminium, le manganèse et le zinc puissent également y contribuer (Wait et Blatchley, 2010). Selon Wait et coll. (2005), la teneur en minéraux ne peut prédire à elle seule le risque d'encrassement, car le potentiel redox influe considérablement sur le processus d'encrassement. Wait et coll. (2005, 2007) recommandent que les réacteurs UV soient placés avant les étapes d'oxydation pour minimiser l'encrassement ou après l'élimination des particules précipitées. Des essais pilotes peuvent être nécessaires pour évaluer le potentiel d'encrassement. Il faut enlever les matières encrassantes pour maintenir le réacteur UV à une performance élevée (Bolton et Cotton, 2008).
La désinfection par les rayons UV est en général utilisée après les procédés d'enlèvement des particules, comme la filtration, afin de minimiser la protection des organismes pathogènes par des particules en suspension. Plusieurs études ont analysé l'effet des particules sur l'efficacité de la désinfection aux rayons UV et dans la plupart des cas, on a conclu que les variations de turbidité de 10 UTN et moins n'avaient pas d'effet sur la réaction des microorganismes à la dose de rayons UV (Christensen et Linden, 2002; Batch et coll., 2004; Mamane-Gravetz et Linden, 2004; Passantino et coll., 2004). On a constaté toutefois que la présence de particules d'acide humique et de coagulants a un effet considérable sur l'efficacité de la désinfection aux rayons UV, l'inactivation obtenue étant plus faible en présence de ces substances (Templeton et coll., 2005). Templeton et coll. (2007) ont constaté que l'inactivation des MS2 était moins importante pendant les périodes de maturation et de fin de vie du filtre, au cours desquelles la turbidité dépassait 0,3 UTN. Les auteurs ont aussi signalé que dans des échantillons d'influents non filtrés (plage de 4,4 à 9,4 UTN), la désinfection par rayons UV des phages en présence de flocons d'acide humique diminuait d'une proportion statistiquement significative (environ 0,5 log) comparativement à l'eau ne contenant pas de particule (Templeton et coll., 2005, 2007).
Templeton et coll. (2005) ont constaté aussi que les particules organiques qui absorbent les rayons UV (c.-à-d. les substances humiques) protégeaient les phages MS2 liés aux particules contre les rayons UV, alors que les particules inorganiques de kaolinton ne les protégeaient pas. Templeton et coll. (2008) ont conclu que les caractéristiques des particules des virus liés à des particules (taille, structure, composition chimique) et le type de désinfectant appliqué étaient plus pertinents que la turbidité. Les servicespublics devraient par conséquent s'efforcer de maintenir des conditions de filtration optimales en amont de la désinfection par les UV (Templeton et coll., 2007).
Pour les approvisionnements en eau souterraine dont la teneur en fer est élevée, Templeton et coll. (2006) ont constaté que le précipité d'oxyde de fer dans les échantillons d'eau souterraine oxydés par l'air pouvait nuire à la désinfection par les rayons UV. Les auteurs ont remarqué que les particules d'oxyde de fer qui causent la turbidité dans l'eau non traitée (2,7 UTN) ont fait baisser de 0,2 log l'inactivation de MS2 à une dose de rayons UV de 40 mJ/cm2 (Templeton et coll., 2006). Les auteurs ont signalé que le fait que les particules de fer ont produit un effet à une turbidité relativement basse indique que certains types de particules inorganiques peuvent protéger les virus contre l'inactivation par les rayons UV.
On s'attend à ce que les rayons UV entraînent une formation minime de SPD (Peldszus et coll., 2000; Hijnen et coll., 2006; Bolton et Cotton, 2008). Wang et coll. (2015) ont toutefois fait état de la formation de chlorate et de bromate pour les procédés d'oxydation avancée (POA) utilisant les rayons UV et le chlore (UV > 1 000 mJ/cm2; chlore libre = 5 à 10 mg/L), et les rayons UV et le peroxyde d'hydrogène (données non présentées). Selon les publications, des nitrites se forment aussi à partir des nitrates. Sharpless et Linden (2001) ont cependant fait état de la formation de moins de 0,1 mg/L d'azote sous forme de nitrite à partir d'une concentration d'azote-nitrates de 10 mg/L lorsque la dose de rayons UV utilisée atteignait 400 mJ/cm2. Les auteurs ont conclu que la formation de nitrite est peu susceptible de représenter un risque pour la santé pendant la désinfection aux rayons UV à l'aide de lampes à PM. Comme pour les désinfectants chimiques, il faut tenir compte de l'incidence éventuelle des SPD lorsque les rayons UV sont utilisés. Il est toutefois essentiel que les tentatives de diminution de la formation de SPD ne nuisent pas à l'efficacité de la désinfection. Il est possible d'obtenir plus d'information auprès de Santé Canada (2008a, 2015).
7.1.3.3 Utilisation de multiples désinfectants
Une stratégie à désinfectants multiples faisant appel à au moins deux étapes de désinfection primaire (c.-à-d. combinaison séquentielle de désinfectants) parvient à inactiver les virus entériques, ainsi que d'autres microorganismes, présents dans l'eau potable. Il est possible d'utiliser les rayons UV et le chlore libre, par exemple, de façon complémentaire au cours de la désinfection afin d'inactiver les protozoaires, les virus et les bactéries. Comme les rayons UV sont très efficaces pour inactiver les protozoaires (mais le sont moins contre certains virus) et comme le chlore est très efficace pour inactiver les bactéries et les virus (mais l'est moins contre les protozoaires), la stratégie à désinfectants multiples permet d'utiliser des doses plus faibles de chlore. Cette stratégie réduit donc la formation de SPD. Dans certaines stations de traitement, on utilise de l'ozone pour éliminer les composés à l'origine du goût et de l'odeur de l'eau, puis on procède à une désinfection au chlore. On pourrait alors considérer que l'ozone et le chlore pourraient permettre d'atteindre les objectifs globaux du traitement : tout dépend de facteurs comme la conception hydraulique du contacteur d'ozone et la présence d'ozone résiduel à la prise d'eau de l'effluent du contacteur.
Les approvisionnements qui utilisent du chlore liquide conformément à une stratégie à désinfectants multiples devraient utiliser des solutions d'hypochlorite homologuées conformes à la norme 60 de NSF International (NSF)/American National Standards Institute (ANSI) (NSF/ANSI, 2017) et suivre les recommandations relatives à la manutention et à l'entreposage de l'hypochlorite qui sont présentées dans le document de Santé Canada (2016).
7.1.4 Réseau de distribution
Réseau de distribution
Un réseau de distribution bien entretenu constitue un élément crucial dans l'approvisionnement d'une eau potable propre (Fisher et coll., 2000). Plusieurs études ont toutefois signalé que la contamination consécutive au traitement dans le réseau de distribution peut être à l'origine d'une partie des cas de MGA endémiques (Payment et coll., 1991, 1997; Hunter et coll., 2005; Nygård et coll., 2007). Lambertini et coll. (2012) ont démontré clairement que les virus pénètrent directement dans les réseaux de distribution et causent des cas de MGA (voir la section 5.4.1). Fox et coll. (2016) ont démontré que des contaminants situés à l'extérieur d'une petite fuite (5 mm de diamètre) dans une conduite sous pression pouvaient y pénétrer et être transportés à l'intérieur du réseau lorsque des pressions transitoires négatives surviennent. Ces pressions transitoires faibles et négatives peuvent découler des activités d'exploitation et d'entretien du réseau de distribution ou d'événements imprévus comme une panne d'électricité ou le bris d'une conduite. Les pressions transitoires faibles et négatives permettent également aux contaminants de pénétrer dans le réseau de distribution par des raccordements croisés ou des retours d'eau provenant d'installations domestiques, industrielles ou institutionnelles (Gullick et coll., 2004).
Les virus peuvent pénétrer dans un réseau de distribution durant la réalisation de travaux (p. ex. réparation d'une rupture de canalisation ou installation de conduites neuves) ou dans le contexte des activités normales d'exploitation et d'entretien qui créent des transitoires de pression (p. ex. manipulation de vannes/bornes, d'incendie, démarrage/arrêt de pompes) et causer des maladies (voir la section 5.1.2) (LeChevallier et coll., 2003; Nygård et coll., 2007; Lambertini et coll., 2011). D'après Lambertini et coll. (2011), l'installation de conduites dans le réseau de distribution a été étroitement associée à une contamination virale. Les virus peuvent pénétrer dans le réseau de distribution par des points de fuite, des joints défectueux ou d'autres ouvertures (LeChevallier et coll., 2003). Les purgeurs d'air et les reniflards situés dans des chambres souterraines susceptibles d'être inondées sont une autre voie d'intrusion des pathogènes (Kirmeyer et coll., 2001; LeChevallier et coll., 2003; Ebacher et coll., 2012). Les biofilms des réseaux de distribution peuvent aussi accumuler et libérer des virus pathogènes infectieux pour l'être humain à la suite d'un événement de contamination (Quignon et coll., 1997; Storey et Ashbolt, 2003; Skraber et coll., 2005; Wingender et Flemming, 2011; Kauppinen et coll., 2012). Les facteurs qui jouent sur l'intrusion des virus dans le réseau de distribution comprennent la durée d'un événement de faible pression ou de pression négative, l'importance de l'orifice de fuite (ou tout autre voie d'entrée) et la concentration de virus dans le sol ou l'eau adjacents à la conduite de distribution d'eau, qui peut s'avérer être importante dans le cas d'une fuite d'un égout sanitaire (Teunis et coll., 2010). Les réseaux de distribution dont l'entreposage flottant est insuffisant ou inexistant sont généralement plus vulnérables aux phénomènes transitoires de pression (LeChevallier et coll., 2011). Les conduites principales en cul-de-sac ont tendance à subir des phénomènes transitoires plus considérables que les conduites d'eau en boucle fermée (Gullick et coll., 2004). La capacité d'un désinfectant secondaire d'inactiver l'intrusion virale dépend du type de résiduel (c.-à-d. chlore libre ou chloramine) et de la concentration et de la demande de résiduel générée par l'eau ou le sol qui pénètre dans le réseau de distribution. On a signalé que les résiduels de désinfectants secondaires types sont inefficaces pour inactiver les virus dans le réseau de distribution (Payment, 1999; Betanzo et coll., 2008).
Des virus peuvent pénétrer dans le réseau et se fixer aux biofilms des conduites, s'accumuler dans le réseau de distribution et se détacher par la suite. Des mesures de protection des sources d'approvisionnement en eau, l'optimisation du traitement, le maintien de l'intégrité physique et hydraulique du réseau de distribution et la diminution du nombre de phénomènes de pression négative ou de basse pression sont essentiels pour limiter l'entrée des virus dans le réseau de distribution (Karim et coll., 2003; Yang et coll., 2011). Les chambres de vannes souterraines devraient faire l'objet d'une inspection mensuelle et être drainées si elles sont inondées (Ebacher et coll., 2012). Dans les systèmes fermés sans entreposage surélevé, l'installation de réservoirs hydropneumatiques ou l'agrandissement de conduites principales peut être nécessaire pour minimiser les impacts causés par les transitoires de pression (LeChevallier et coll., 2011). La modélisation hydraulique des réseaux de distribution peut indiquer les points du réseau les plus susceptibles aux événements de pressions négatives ou de basses pressions (Teunis et coll., 2010; LeChevallier et coll., 2011). Il est recommandé de surveiller régulièrement la qualité de l'eau du réseau de distribution (p. ex. indicateurs microbiens, résiduels de désinfectant, turbidité, pH), mettre en place des programmes d'exploitation et d'entretien (p. ex. rinçage de conduites, prévention des retours d'eau polluée, gestion des actifs) et pratiquer une hygiène rigoureuse durant pendant tous les travaux de construction, de réparation et d'entretien de conduites, afin que l'eau potable soit acheminée jusqu'au consommateur avec une qualité acceptable. (Kirmeyer et coll., 2001, 2014). On a aussi signalé que suite à des travaux de réparation de conduites, l'ouverture d'une seule vanne afin de permettre la distribution d'eau provenant du secteur isolé permettait de réduire le risque (Blokker et coll., 2014). On recommande aussi de surveiller les pressions dans le réseau de distribution afin d'optimiser le l'opération du réseau (Feffer et coll., 2016). On trouvera dans les études de LeChevallier et coll. (2011) d'autres conseils pour aider les services d'approvisionnement en eau à élaborer des pratiques exemplaires pour gérer les effets des pressions transitoires sur la qualité de l'eau. Des normes de désinfection visant à assurer des conditions hygiéniques pendant la construction ou la réparation de conduites d'eau sont également disponibles (ANSI/AWWA, 2014).
7.2 Échelle résidentielle
Le traitement de l'eau potable à l'échelle municipale est conçu pour diminuer la présence d'organismes pathogènes et les risques pour la santé connexes jusqu'à un niveau acceptable ou sécuritaire. C'est pourquoi il n'est habituellement pas nécessaire d'utiliser, dans les résidences, des dispositifs de traitement de l'eau, puisque l'eau qui les alimente a déjà été traitée par les installations municipales; il s'agit essentiellement d'un choix individuel.
Les eaux souterraines constituent une source courante d'eau potable pour les approvisionnements en eau individuels (puits privées) ou de petite taille. Comme nous l'avons vu aux sections 5.1.2 et 5.5, on a détecté des virus dans de nombreux différents types d'aquifères, y compris les aquifères karstique, de sable, de gravier et du substratum rocheux fracturé confinés et non confinés. Dans la plupart des cas, on a déterminé que des sources de matières fécales souterraines comme des égouts ou des fosses septiques qui fuient constituent la source probable de contamination virale de ces types de puits. Le Tableau 1 présente la liste des facteurs qui ont une incidence sur la probabilité de contamination virale des eaux souterraines. Des études ont aussi révélé que des virus peuvent survivre et parcourir des centaines à des milliers de mètres dans les eaux souterraines selon le type de matériau qui constitue l'aquifère (p. ex., sable à gros grains, gravier, roc fracturé). Des données épidémiologiques ont en outre établi un lien entre la consommation d'eau souterraine non traitée contaminée par des matières fécales et des maladies entériques. Par conséquent, les petits approvisionnements d'eau souterraine potable (c.-à-d. réseaux semi-publics) destinés à une population vulnérable à la contamination virale devraient être traités de façon à éliminer ou inactiver les virus entériques.
De nombreux puits privés approvisionnant des ménages tirent leur eau d'aquifères peu profonds ou du substratum fracturé (Conboy et Goss, 2000: Novakowski, 2015). On signale qu'au Canada, plus de 40 % des puits ruraux ont subi les répercussions des activités agricoles ou des sources de contamination fécales (Gouvernement du Canada, 2006). De plus, des études ont révélé la présence de virus dans 35 à 78% des puits privés dotés de fosses septiques (Francy et coll., 2004; Trimper et coll., 2010; Allen et coll., 2017). Même s'il est difficile pour les propriétaires d'évaluer en détail la vulnérabilité de leur puits à la contamination virale, des mesures peuvent être prises pour qu'ils soient davantage au fait des risques de contamination d'un puits par des matières fécales. Des gouvernements provinciaux et territoriaux offrent des conseils généraux sur la construction, l'entretien, la protection et l'analyse de puits. Lorsqu'ils se penchent plus particulièrement sur le potentiel de contamination virale, les propriétaires de puits doivent connaître la façon dont les puits sont construits, le type de matériau d'aquifère entourant le puits, ainsi que l'emplacement du puits par rapport aux sources de contamination fécale (p. ex. fosses septiques, égouts sanitaires, déchets animaux) (voir le tableau 1). Il est possible de tirer ces renseignements de dossiers fournis au propriétaire au cours de la construction du puits et de la fosse septique, ainsi que les rapports de forage, de cartes sur la vulnérabilité des aquifères et d'évaluations des eaux souterraines régionales que l'on peut généralement obtenir des gouvernements provinciaux et territoriaux. S'il n'y a pas suffisamment d'information disponible pour déterminer la vulnérabilité d'un puits à la contamination virale, il faudrait alors mettre en place un traitement pour réduire les risques. En général, l'eau de surface n'est pas recommandée pour l'approvisionnement en eau privé ou semi-public, sauf si elle est dûment filtrée et désinfectée, et si sa qualité est contrôlée.
L'analyse de l'eau du puits peut fournir aux propriétaires d'autres renseignements qu'ils peuvent utiliser conjointement avec l'information sur la construction de puits, l'emplacement et les caractéristiques de l'aquifère pour déterminer plus facilement si leur puits peut être vulnérable à la contamination virale. On recommande plus particulièrement d'analyser les paramètres qui peuvent indiquer une possible contamination de l'eau du puits par une fosse septique ou par des eaux de surface à cause de la mauvaise intégrité du puits. Les propriétaires de puits privés devraient analyser périodiquement (au minimum, deux fois par année) leurs puits pour en déterminer les paramètres bactériologiques (p. ex., coliformes totaux et E. coli). Il convient de réaliser des analyses plus souvent si le puits est peu profond ou se trouve dans un aquifère du substratum rocheux (voir plus haut) et si une fosse septique est présente sur le terrain de la propriété ou tout près. La présence de coliformes totaux et d'E. coli dans de l'eau de puits témoigne de la possibilité d'une vulnérabilité accrue d'un puits à la contamination fécale (voir la section 5.7.2) (Abbaszadegan et coll., 2003; Allen, 2013; Fout et coll., 2017). Santé Canada (2012a, b) donne de l'information sur la façon d'interpréter les résultats d'analyse de détection de coliformes totaux et d'E. coli.
D'autres paramètres de la qualité de l'eau peuvent aussi servir à évaluer la vulnérabilité d'un puits à la contamination. La présence de concentrations de nitrate (NO3-N) et de chlorure (Cl–) dépassant les concentrations de fond (> 0,90 à 2,0 mg NO3-N/L; > 10 mg/L Cl–) peut illustrer l'effet des effluents de fosse septique sur la qualité de l'eau de puits (Roberston et coll., 1989; Harman et coll., 1996; Santé Canada, 2013; Robertson et coll., 2013; Schaider et coll., 2016). Des études ont aussi établi un lien entre la présence de nitrate et de chlorure (au-dessus des concentrations de fond) et celle des virus entériques dans des puits privés (Borchardt et coll., 2003; Francy et coll., 2004). C'est pourquoi il est utile d'analyser périodiquement ces paramètres pour déterminer si les effluents de fosse septique ont une incidence sur un puits.
L'analyse de la présence des paramètres ci-dessus fournit des données permettant de déterminer si un puits peut être contaminé par des matières fécales; leur absence n'indique toutefois pas qu'il n'y a pas de pathogènes viraux. Les résultats de cette analyse doivent être examinés conjointement avec d'autres renseignements recueillis par le propriétaire, comme la construction du puits et son emplacement, le type d'aquifère et l'emplacement de la source de contamination fécale, pour déterminer si le puits est vulnérable à la contamination fécale.
Là où un traitement s'impose, il existe divers moyens de traiter les eaux de la source d'approvisionnement afin de produire de l'eau potable de grande qualité exempte de pathogènes. Ces moyens comprennent la filtration et la désinfection à l'aide de composés chlorés ou de techniques de remplacement, comme l'utilisation des rayons UV. Ces technologies s'apparentent aux procédés de traitement municipales, mais elles sont appliquées aussi à plus petite échelle. Par exemple, l'installation d'un système de traitement par rayons UV reconnu pour fournir une dose de 40 mJ/cm2 (voir plus bas) est un moyen efficace pour réduire les risques liés à des virus entériques souvent associés à des maladies humaines au Canada. On peut aussi recourir à d'autres procédés, comme la distillation, mais ils ne conviennent qu'aux petits systèmes d'approvisionnement en eau ou à des systèmes d'approvisionnement en eau individuels. La plupart de ces technologies ont été intégrées dans des dispositifs au point d'entrée (PE), qui traitent toute l'eau pénétrant dans le système, ou de dispositifs au point d'utilisation (PU), qui traitent l'eau à un seul point, par exemple, au robinet de la cuisine. Il ne faut pas installer au point d'entrée des technologies destinées au point d'utilisation, car l'eau traitée peut être corrosive pour les éléments de plomberie interne.
Santé Canada ne recommande pas de marques particulières de dispositifs de traitement de l'eau potable, mais le Ministère conseille vivement aux consommateurs d'utiliser seulement des dispositifs homologués par un organisme de certification agréé comme étant conformes à la norme NSF/ANSI appropriée. Les normes NSF/ANSI visent à protéger l'eau potable en aidant à garantir l'innocuité des matériaux et l'efficacité des produits qui entrent en contact avec l'eau potable.
Les organismes de certification garantissent la conformité d'un produit ou service aux normes en vigueur. Au Canada, le Conseil canadien des normes (CCN) a accrédité les organismes ci-dessous, qu'il autorise ainsi à homologuer les dispositifs de traitement de l'eau potable qui respectent les normes NSF/ANSI pertinentes (CCN, 2018) :
- Canadian Standards Association International (www.csa-international.org);
- NSF International (www.nsf.org);
- Water Quality Association (www.wqa.org); (en anglais seulement)
- Underwriters Laboratories Inc. (www.ul.com);
- Bureau de normalisation du Québec (www.bnq.qc.ca);
- International Association of Plumbing & Mechanical Officials (www.iapmo.org).(en anglais seulement)
On peut obtenir une liste à jour des organismes de certification agréés auprès du CCN (2018).
Un laboratoire doit régulièrement procéder à des analyses de détection d'E. coli et de coliformes totaux dans l'eau pénétrant dans le dispositif de traitement, de même que dans l'eau traitée, pour vérifier l'efficacité du dispositif en question. La capacité d'élimination des dispositifs de traitement diminue avec le temps et l'utilisation, et il faut les entretenir ou les remplacer. Les consommateurs doivent vérifier la longévité prévue des éléments de leur dispositif de traitement, suivre les recommandations du fabricant, et établir un calendrier d'entretien clair. Il faut inspecter les dispositifs de traitement et les entretenir selon le calendrier d'entretien et les recommandations du fabricant.
Les systèmes semi-publics et privés qui ont recours à la désinfection utilisent généralement le chlore ou les rayons UV en raison de leur disponibilité et de leur facilité d'utilisation relative. Le colmatage et l'encrassement de la surface de la lampe UV constituent toutefois un problème courant lorsque l'on applique les rayons UV à l'eau brute de dureté moyenne ou élevée, comme les eaux souterraines. Un filtre de prétraitement est souvent nécessaire pour réduire l'entartrage et l'encrassement et atteindre la qualité d'eau optimale requise pour assurer le bon fonctionnement du système UV selon les spécifications du fabricant. De plus, il est essentiel de nettoyer et de remplacer régulièrement la lampe conformément aux instructions du fabricant afin d'assurer le bon fonctionnement de l'appareil. On peut également avoir recours à des mécanismes spéciaux pour le nettoyage de la lampe UV ou à des adoucisseurs d'eau pour éviter les problèmes d'entartrage.
Les approvisionnements privés et semi-publics qui utilisent des systèmes UV peuvent se reporter à la norme NSF/ANSI 55 (NSF/ANSI, 2016b), qui définit des critères de rendement pour deux catégories de dispositifs homologués : les dispositifs de classe A et les dispositifs de classe B. Les dispositifs de traitement de classe A conformes à la norme NSF/ANSI 55 sont conçus pour produire une dose de rayons UV d'au moins 40 mJ/cm2 nécessaire à l'inactivation de microorganismes, y compris les bactéries, les virus, les oocytes de Cryptosporidium et les kystes de Giardia, dans l'eau contaminée. Ces systèmes permettent une diminution de 4 log de la plupart des virus (Tableau 6) et conviennent à cette utilisation. Il convient toutefois de signaler qu'ils ne sont pas conçus pour traiter les eaux usées ni les eaux contaminées par des eaux usées non traitées et qu'il faut les installer dans une eau limpide. Il importe de signaler que les systèmes de classe B conformes à la norme NSF/ANSI 55 sont conçus pour fournir une dose de rayons UV d'au moins 16 mJ/cm2 et ne permettent pas de diminuer de 4 log la concentration de la plupart des virus (Tableau 6). Les systèmes de classe B ne sont pas conçus pour désinfecter de l'eau microbiologiquement insalubre. Ils sont homologués seulement pour le traitement bactéricide supplémentaire d'approvisionnements publics en eau potable désinfectée ou d'autres approvisionnements en eau potable qui ont été analysés et sont jugés acceptables pour la consommation humaine. Les systèmes de classe B réduisent seulement les microorganismes non pathogènes naturellement présents dans l'eau.
Certaines provinces et certains territoires peuvent exiger que les systèmes semi-publics fournissent une dose de rayons UV de 186 mJ/cm2 en présence d'adénovirus (p. ex. lorsqu'il y a une influence des eaux usées). Les systèmes UV conçus conformément aux spécifications du UV Disinfection Guidance Manual de l'EPA des É.-U. sont disponibles sur le marché et peuvent fournir une dose de 186 mJ/cm2 pour les approvisionnements allant jusqu'à 24 gal US/min > facteur de transmission UV de 68 % (U.S EPA, 2006b).
Les approvisionnements privés et semi-publics qui utilisent du chlore liquide doivent utiliser des solutions d'hypochlorite homologuées qui satisfont à la norme NSF/ANSI 60 (NSF/ANSI, 2017) et suivre les recommandations relatives à la manutention et à l'entreposage de l'hypochlorite énoncées par Santé Canada (2016).
Les membranes d'osmose inverse ont des pores plus petits que les virus et, par conséquent, peuvent agir comme barrières physiques. La norme NSF/ANSI 58 (NSF/ANSI, 2016b) ne renferme toutefois aucune allégation de diminution des virus. Les membranes d'ultrafiltration ont des pores d'un diamètre de 0,005 à 0,05 µm et peuvent aussi constituer une barrière physique pour les virus, même s'il n'existe actuellement aucune norme NSF/ANSI portant sur les réseaux d'ultrafiltration à l'échelle résidentielle (p. ex. point d'utilisation). Concernant les systèmes d'approvisionnement en eau potable nécessitant une capacité accrue, il faut noter qu'un certain nombre de systèmes d'ultrafiltration certifiés conformes à la norme NSF/ANSI 419 (NSF/ANSI, 2015) ont été soumis à des essais de diminution virale. Toutefois, à l'instar des systèmes municipaux (voir la section 7.1.2), les tests d'intégrité directs n'ont pas une résolution suffisante pour vérifier l'élimination logarithmique des virus. Il importe de contacter l'autorité responsable en matière d'eau potable dans le secteur de compétence concerné pour confirmer l'acceptabilité ou l'applicabilité de ces systèmes.
8.0 Évaluation des risques
L'évaluation quantitative du risque microbien (ÉQRM) est un procédé qui utilise des modèles mathématiques, des données sur la qualité des sources d'approvisionnement en eau, des renseignements portant sur les procédés de traitement et les caractéristiques de l'agent pathogène pour estimer la charge de morbidité liée à une exposition à des microorganismes particuliersdans une source d'approvisionnement eneau potable. L'ÉQRM peut être utilisée de deux façons, soit pour la gestion d'un système d'eau potable dans le cadre d'une approche de la source au robinet ou, comme c'est le cas pour le présent document, pour appuyer l'élaboration de recommandations sur la qualité de l'eau potable.ÉQRM
D'autres renseignements et recommandations sur l'utilisation de l'ÉQRM dans le cadre d'une approche de la source au robinet sont publiés ailleurs (OMS, 2016; Santé Canada, 2018). Le présent document technique n'aborde que l'utilisation de l'ÉQRM pour l'élaboration d'une recommandation sur la qualité de l'eau potable pour les ÉQRMvirus entériques. ÉQRMÉQRMapprovisionnement
8.1 Objectifs basés sur la santé
Les objectifs basés sur la santé sont les buts à atteindre pour garantir la salubrité de l'eau potable. Au Canada, on tente souvent de réduire les dangers microbiologiques en cherchant à répondre à deux types d'objectifs : les objectifs de qualité de l'eau et les objectifs de traitement basés sur la santé. La recommandation bactériologique concernant E. coli constitue un exemple d'objectif de qualité de l'eau; elle établit une concentration maximale acceptable pour ce microorganisme dans l'eau potable (Santé Canada, 2012a). Les objectifs detraitement basés sur la santé précisent à quelle diminution des organismes pathogènes il faut parvenir à l'aide de mesures comme les procédés de traitement (voir la section 7.0). Les objectifs de traitement aident à choisir les procédés de traitement et devraient être définis en fonction de la qualité des sources d'approvisionnement en eau (voir la section 8.3.2). En raison du large éventail de microorganismes pathogènes, il est difficile d'évaluer tous les risques potentiels. C'est pourquoi les objectifs de traitement sont généralement définis en fonction de catégories de microorganismes (p. ex. bactéries, virus, protozoaires) plutôt que dd'organismes pathogènes particuliers. Pour les virus entériques, l'objectif de traitement basé sur la santéest une élimination ou inactivation d'au moins 4 log des virus. Pour bon nombre de sources d'approvisionnement, il devrait peut-être parvenir à une élimination ou à une inactivation logarithmique plus grande pour que le niveau de risque demeure acceptable.8.2 Niveaux de risque de référence
Le niveau de risque de référence est la charge de morbidité rattachée à l'eau potable qui est estimée tolérable ou acceptable. Cette valeur sert à établir les objectifs de traitement basés sur la santé.
Les niveaux de risque sont exprimés de plusieurs façons. Dans les Directives de qualité pour l'eau de boisson de l'OMS (OMS, 2017), on utilise les années de vie corrigées de l'incapacité (AVCI) comme unité de mesure du risque. Les AVCI permettent d'obtenir une valeur tenant compte du risque de maladie ou de blessure, et de la répercussion des effets connexes sur la santé (Murray et Lopez, 1996a; Havelaar et Melse, 2003). Les directives de l'OMS (2011) adoptent 10−6 AVCI/personne par année comme niveau de risque de référence. Le document National Guidelines for Water Recycling (NRMMC-EPHC, 2006) de l'Australie mentionne la même valeur. Par contre, d'autres organismes établissent des niveaux de risque microbien acceptable en s'appuyant sur le risque d'infection, sans tenir compte de la probabilité ou de la gravité des effets connexes sur la santé. L'EPA des É.-U. et les Pays-Bas, par exemple, utilisent un objectif en matière de santé correspondant à un risque d'infection annuel inférieur à 1/10 000 personnes (10−4) (Regli et coll., 1991; VROM, 2005).
L'évaluation des risques effectuée dans le présent document technique estime la charge de morbidité à l'aide des AVCI. Cette unité de mesure offre plusieurs avantages : elle prend en compte à la fois le nombre d'années perdues en raison d'un décès et le nombre d'années vécues avec une incapacité (comparativement à l'individu moyen en bonne santé dans la région) et permet de déterminer les effets sur la santé d'un seul type d'organisme pathogène. L'utilisation des AVCI permet également de comparer les effets sur la santé de divers organismes pathogènes et, peut-être, de différents dangers microbiologiques et de certains dangers chimiques. Bien qu'aucune unité de mesure commune de la santé n'ait été acceptée à l'échelle internationale, les AVCI sont utilisées par de nombreux groupes, et l'on dispose de données publiées et évaluées par des pairs. Dans la présente évaluation des risques, le niveau de référence de l'OMS (2011), soit 10−6 AVCI/personne par année, est retenu comme niveau de risque acceptable.
8.3 Évaluation quantitative du risque microbien
Tel qu'il est indiqué plus haut, le but de ce document est d'appuyer l'élaboration d'un objectif de traitement basé sur la santé pour les virus entériques dans l'eau potable. L'ÉQRM constitue un outil important à cette fin. L'ÉQRM suit une stratégie commune d'évaluation des risques, qui comporte quatre volets : l'identification des dangers, l'évaluation de l'exposition, l'évaluation de la relation dose-réponse et la caractérisation des risques. Dans ce cas-ci, le risque est déjà caractérisé par un niveau de référence de 1 × 10-6 AVCI/personne par année. Cette évaluation des risques examine la performance de traitement nécessaire pour diminuer la concentrations de virus entériques dans l'eau à la source à un niveau qui permet d'atteindre le résultat sur la santé, compte tenu d'une qualité donnée de la source d'approvisionnement de l'eau, de conditions d'exposition établies et de renseignements précis sur la dose-réponse.
8.3.1 Détermination des dangers
Au Canada, les virus entériques les plus préoccupants pour la santé humaine dans les sources d'eau potable sont les norovirus, les rotavirus, les virus de l'hépatite, les entérovirus et les adénovirus. Même si tous les virus entériques qui peuvent nuire à la santé humaine sont connus, les évaluations des risques tiennent rarement compte de chacun d'eux. L'évaluation porte seulement sur des virus entériques précis qui, en raison de leurs caractéristiques, sont représentatifs de tous les virus pathogènes semblables. On suppose que si le traitement est efficace contre le virus de référence, il le sera contre tous les virus préoccupants semblables. Un virus de référence représente le pire scénario possible, soit une combinaison des éléments suivants : fréquence élevée, forte concentration et longue survie dans les sources d'approvisionnement en eau, faible élimination et forte résistance à l'inactivation pendant le traitement et pathogénicité élevée pour tous les groupes d'âge. On a envisagé de nombreux virus entériques comme virus de référence, notamment des rotavirus, des adénovirus et des norovirus. Aucun de ces virus ne possède toutes les caractéristiques d'un virus de référence idéal. Les adénovirus représentent l'un des pires scénarios d'inactivation virale durant le traitement par rayons UV, mais ils sont moins prévalents dans la population que les norovirus ou les rotavirus. Les norovirus constituent une cause importante de gastroentérite virale dans tous les groupes d'âge; un modèle dose-réponse publié est disponible (Teunis et coll., 2008). Le modèle soulève toutefois beaucoup de débats, et certains sont d'avis qu'il surestime l'infectiosité des norovirus (Schmidt, 2015). Les rotavirus constituent une cause courante d'infections chez les enfants et peuvent causer de graves répercussions; un modèle dose-réponse est disponible. Les rotavirus sont toutefois plus sensibles au traitement que d'autres virus entériques. Puisqu'aucun virus ne comporte toutes les caractéristiques d'un virus de référence idéal, cette évaluation des risques repose sur les caractéristiques de plusieurs virus différents. Le modèle dose-réponse et les données d'inactivation virale par rayons UV sont fondés sur des études portant sur le rotavirus. Les valeurs CT pour les désinfectants chimiques reposent toutefois sur le VHA et le poliovirus (U.S. EPA, 1991, 1999), car il s'agit des meilleurs renseignements actuellement disponibles sur la désinfection chimique des virus entériques communément présents dans les eaux de surface et les eaux souterraines.
8.3.2 Évaluation de l'exposition
L'exposition correspond à la dose quotidienne d'agents pathogènes ingérés par un consommateur. La consommation d'eau potable est la principale voie d'exposition qui ait été prise en compte dans la présente évaluation des risques. Pour évaluer l'exposition, il faut connaître ou estimer à le volume d'eau ingéré et la concentration de virus entériques dans l'eau potable.
8.3.2.1 Estimation des concentrations dans les sources d'approvisionnement en eau
Le processus d'ÉQRM a été utilisé en sens inverse afin de guider l'élaboration d'objectifs de traitement basés sur la santé. Autrement dit, on a utilisé le processus d'ÉQRM pour répondre à la question suivante : quelle est la concentration moyenne de virus entériques dans la source d'approvisionnement en eau qui seraient associée à un niveau de risque de référence de 10-6 AVCI par personne par année (section 8.2), compte tenu d'un volume moyen d'eau ingéré (section 8.3.2.3) et des diminutions par le traitement relevées pour le système d'eau potable (section 8.3.2.3)?
Il a été démontré que les concentrations moyennes permettent de quantifier adéquatement les objectifs de traitement pour l'exposition par l'eau potable (Petterson et coll., 2015). Pour déterminer les concentrations moyennes dans les sources d'approvisionnement en eau, il faut se demander si la présence des virus est continue ou intermittente, s'il existe des tendances saisonnières et quelle est l'incidence des phénomènes rares, comme les sécheresses ou les inondations, sur les concentrations. Les pics à court terme des concentrations de virus peuvent accroître considérablement les risques de maladie et même déclencher des éclosions de maladies d'origine hydrique. Les programmes de surveillance doivent donc tenir compte de ces facteurs pour saisir la variabilité qui se produit dans la source d'approvisionnement en eau (Dechesne et Soyeux, 2007). Selon le projet Microrisk, il faut procéder à une analyse mensuelle pendant un an pour établir les niveaux de référence et caractériser ensuite au moins deux phénomènes complets afin de comprendre les conditions dans lesquelles les pics sont survenus. À cause de la variabilité temporelle des virus dans les eaux de surface, il peut être nécessaire d'intensifier l'échantillonnage (c.-à-d. cinq échantillons par semaine) pour quantifier les concentrations de pointe (Westrell et coll., 2006a). Il convient aussi de signaler que dans le cas des sources de rivière contenant un volume élevé d'effluents de la stations de traitement des eaux usées, des contaminations de pointe sont possibles en période de faible débit (Deschesne et Soyeux, 2007). Outre la surveillance, il faut réaliser l'analyse de l'incertitude qui aide à évaluer les concentrations estimées dans les sources d'approvisionnement en eau (Petterson et coll., 2015). Le document de Santé Canada (2018) contient d'autres renseignements sur la façon de contrôler ou d'estimer les concentrations de pathogènes. La détermination des concentrations dans les sources d'approvisionnement en eau doit aussi tenir compte d'autres facteurs comme le taux de récupération déterminé par des méthodes de détection des virus, taux qui est bien inférieur à 100 %, et l'infectiosité des virus détectés pour les humains.
8.3.2.2 Diminutions par le traitement
Dans cette évaluation du risque, on a analysé différentes combinaisons de diminutions logarithmiques obtenues grâce à des procédés de traitement et de concentrations de virus entériques dans les eaux de source d'approvisionnement, puis on les a comparées à une valeur de référence définie pour le risque. Il importe de remarquer que de nombreux facteurs peuvent avoir une incidence sur le traitement (voir la section 7.0). On suppose que les virus qui n'ont pas été éliminés ou inactivés pendant le traitement sont encore infectieux et pathogènes.
8.3.2.3 Volume d'eau ingéré
En ce qui concerne le volume d'eau consommée, seule l'eau du robinet non bouillie est prise en compte, puisque l'ébullition entraîne l'inactivation d'organismes pathogènes et une surestimation de l'exposition (Gale, 1996; Payment et coll., 1997; OMS, 2017). Au Canada, chaque personne consomme en moyenne environ 1 L d'eau du robinet non bouillie par jour (Statistique Canada, 2004, 2008). Dans la présente évaluation des risques, on utilise donc une consommation moyenne de 1 L d'eau par personne par jour pour déterminer l'exposition. Cette estimation est comparable aux tendances de consommation observées dans d'autres pays industrialisés (Westrell et coll., 2006b; Mons et coll., 2007). L'OMS (2011) recommande également d'utiliser une valeur de consommation estimative de 1 L d'eau du robinet non bouillie. On peut alors utiliser la concentration d'eau potable traitée et le volume d'eau ingérée pour déterminer l'exposition (c.-à-d. la dose de virus consommée par personne par jour).
8.3.3 Évaluation de la relation dose-réponse
Dans l'évaluation de la relation dose-réponse, on utilise des modèles dose-réponse pour estimer la probabilité d'infection (Pinfection) et le risque de développement d'une maladie après une exposition à des virus. On peut aussi utiliser cette relation dose-réponse, comme dans le cas de la présente évaluation, pour estimer la concentration d'un virus associée à un niveau de risque précis dans des conditions déterminées.
Dans la présente évaluation des risques, on utilisele modèle dose-réponse pour les rotavirus. Les données sur la relation dose-réponse du rotavirus sont caractérisées par les résultats obtenus par le biais du modèle bêta-Poisson (Haas et coll., 1999). Le modèle bêta-Poisson de la relation dose-réponse pose les hypothèses suivantes :
- une seule virion peut causer une infection (aucun seuil);
- la dose de l'organisme pathogène dans l'échantillon d'eau est distribuée selon l'équation de Poisson (c.-à-d. aléatoirement) et à une concentration moyenne connue (d);
- la probabilité que chaque pathogène consommé survive à l'interaction hôte-pathogène et cause une infection suit une distribution bêta comportant des paramètres de forme α et β;
- le nombre de pathogènes consommés qui survivent à l'interaction hôte-pathogène causant une infection suit une distribution binomiale.
On trouve des renseignements supplémentaires sur la dérivation de l'approximation réalisée au moyen du modèle dans Schmidt et coll. (2013).
Les paramètres α et β sont tirés d'études de la relation dose-réponse menées auprès de sujets en bonne santé; il se peut donc qu'ils ne représentent pas de façon adéquate les effets sur des sous-groupes sensibles, comme les personnes immunodéprimées, les jeunes enfants et les personnes âgées (Ward et coll., 1986). On estime la dose quotidienne de microorganismes ingérés par une personne au moyen des informations provenant de l'évaluation de l'exposition (voir la section 8.3.2). La probabilité annuelle de survenue d'une infection chez une personne est estimée au moyen de l'équation 1. Pour la présente évaluation des risques, on présume qu'il n'y a aucune propagation secondaire de l'infection.
Pinfection/année = 1 − (1 − Pinfection)365
Les personnes infectées ne développeront pas toutes une maladie clinique. Le risque annuel pour une personne est estimé au moyen de l'équation 2 :
Risque de maladie = Pinfection/année x S x I
où :
Pinfection/année = probabilité annuelle d'infection (obtenue à l'aide de l'équation 1)
S = proportion de la population sensible aux infections
I = proportion des personnes qui contractent une maladie symptomatique suite d'une infection
La proportion de la population qui est vulnerable aux infections et aux maladies varie selon le type de virus entériques étudié. Dans le cadre la présente évaluation des risques, le rotavirus représente tous les virus entériques qui peuvent être présents dans l'eau potable, y compris ceux qui s'y trouvent en plus grande concentration ou auxquels la majorité de la population peut être sensible (p. ex. norovirus). Puisqu'il est défini comme virus de référence, les taux de vaccination du rotavirus ne sont pas pris en compte; on suppose que 100 % de la population est sensible à l'infection (c.-à-d qu'on suppose que S=1). Les infections ne provoquent pas toutes une maladie symptomatique. Selon des données américaines, 88 % des personnes infectées par un rotavirus contractent une maladie symptomatique (Havelaar et Melse, 2003).
L'évaluation utilise les AVCI comme unité de mesure commune du risque pour convertir le risque annuel de maladie pour une personne en charge de morbidité par personne. Le principal avantage cité de l'AVCI comme unité de mesure de la santé publique est son caractère globale, qui combine les années de vie perdues (AVP) et les années vécues avec une incapacité (AVI) pour calculer la charge de morbidité. Le calcul des AVCI s'effectue ainsi :
AVCI = AVI + AVP
où :
AVI = la somme de [(fraction de l'effet) × (durée) × (poids de la gravité)] pour chaque effet sur la santéjouant un rôle dans la morbidité;
AVP = [(espérance de vie) − (âge au moment du décès)] × poids de la gravité
Pour les rotavirus, la gravité des effets sur la santé varie de la diarrhée légère à la diarrhée grave, voire un décès. La charge de morbidité que représente la gastroentérite causée par une infection à rotavirus attribuable à l'eau potable est de 8,28 AVCI/1 000 cas (8,28 × 10−3 AVCI/cas) (tableau 7).
| Effet sur la santé | Fraction de l'effetnote a | Durée de la maladienote b | Poids de la gravité note c | AVCI/cas | |
|---|---|---|---|---|---|
| Morbidité (AVI) | Diarrhée légère Diarrhée grave |
0,50 0,49 |
0,01918 (7 jours) 0,01918 (7 jours) |
0,067 0,39 |
6,43 × 10−4 3,74 × 10−3 |
| Mortalité (AVP) | Décès | 0,0001 | Espérance de vienote d; âge au décèsenote e |
1 | 3,90 × 10−3 |
| Charge de morbidité | 8,28 × 10−3 | ||||
|
|||||
Au moyen de cette charge de morbidité (AVCI/cas) et du risque annuel de maladie pour une personne, on peut estimée la charge de morbidité en AVCI/personne par année comme suit :
Charge de morbidité = risque de maladie × charge de morbidité (AVCI/cas)
(AVCI/personne par année)
où :
- Risque de maladie = valeur calculée au moyen de l'équation 2;
- Charge de morbidité (AVCI/cas) = 8,28 × 10−3
Comme indiqué précédemment, puisquela charge de morbidité est établie comme étant égale au niveau de risque de référence, les calculs AVCI sont utilisés pour convertir le niveau de risque de référence en valeurs pour le modèle dose-réponse pour rotavirus.
8.3.4 Caractérisation des risques
Dans la présente évaluation des risques, l'étape de la caractérisation des risques sert à établir un objectif minimal de traitement basé sur la santé permettant d'atteindre le niveau de risque de référence.
Comme l'illustre la figure 1, à mesure que la concentration de virus augmente dans la source d'approvisionnement en eau, la diminution logarithmique doit être importante pour que le traitement permette toujours d'atteindre le niveau de risque de référence. Par exemple, lorsque la concentration de virus est de 1/100 L dans la source d'approvisionnement en eau et que la station de traitement parvient constamment à une diminution d'au moins 4 log, la charge de morbidité pour la population répond au niveau de référence de 10-6 AVCI/personne par année (moins de 1 cas/1 000 personnes par année). Les sources d'approvisionnement en eau au Canada, y compris les eaux souterraines, peuvent toutefois présenter des concentrations virales de plus de 1/100 L (voir la section 5.0) et c'est pourquoi une diminution logarithmique encore plus importante s'exigepour atteindre le niveau acceptable. Il importe en outre de tenir compte de l'effet des conditions environnementales sur les concentrations dans la source d'approvisionnement en eau (c.-à-d. événements de pointe), car elles peuvent exiger une diminution logarithmique encore plus importante pour atteindre le niveau acceptable. Il est important que les fournisseurs d'eau potable tiennent compte de ces phénomènes de pics dans leur évaluation d'un site, afin de comprendre pleinement les risques éventuels qu'ils entraînent pour leur eau potable (Santé Canada, 2018). Le degré de traitement atteint doit par conséquent tenir compte non seulement des conditions de fonctionnement normales, mais également des variations possibles de la qualité de l'eau ou de l'efficacité du traitement.
Selon les données limitées dont nous disposons sur les virus entériques dans les sources d'approvisionnement en eau au Canada, un objectif de traitement basé sur la santé correspondant à une diminution de 4 log des virus entériques représente une exigence minimale. Il peut toutefois être nécessaire d'obtenir un plus fort taux d'élimination ou d'inactivation pour atteindre l'objectif de traitement.. Il faut évaluer le site pour déterminer quel taux de diminution virale doit être obtenu pour une source d'approvisionnement en eau donnée. Une surveillance des sources d'approvisionnement en eau pour y détecter la présence des virus entériques approvisionnementpermettra d'obtenir la meilleure évaluation du site soit. Cependant, s'il est impossible d'effectuer des mesures, les informations tirés d'évaluations du système , de même que les renseignements sur d'autres paramètres de la qualité de l'eau, peuvent servir à estimer le risque et le degré de contamination fécale des sources d'approvisionnement en eau.approvisionnement. Ces renseignements peuvent ensuite aider à déterminer si la concentration des virus entériques présents dans l'eau exige plus que le traitement minimal..
Il est également important de comprendre les diminutions logarithmiques auxquelles la station de traitement peut parvenir lorsqu'elle fonctionne dans des conditions optimales, ainsi que les répercussions des perturbations à court et à long terme du traitement sur les diminutions logarithmiques. En comprenant les variations de la qualité de l'eau à la source et de l'eau à la station de traitement, et en planifiant en conséquence, il est possible to crée un système plus robuste pouvant comporter des marges de sécurité. Il est également important de tenir compte de l'incertitude relative à l'ÉQRM, afin que le traitement en place produise une eau de qualité acceptable. Une analyse de sensibilité effectuée au moyen d'un modèle d'ÉQRM peut aussi aider à déterminer les points de contrôle critiques et leurs limites. D'autres renseignements sur les évaluations propres aux sites et l'utilisation de l'ÉQRM comme outil dans une approche de la source au robinet sont donnés ailleurs (Santé Canada, 2018).
Figure 1 - Objectif de traitement basé sur la santé pour les virus entériques , pour atteindre un niveau de risque acceptable de 10-6 AVCI/personne par année, pour une consommation quotidienne de 1 L d'eau potable, comme il a été établi au moyen de la méthode d'ÉQRM. Cette figure n'est pas indiquée aux fins d'examen(s) ou de comparaison(s) avec des données de surveillance des virus entériques.
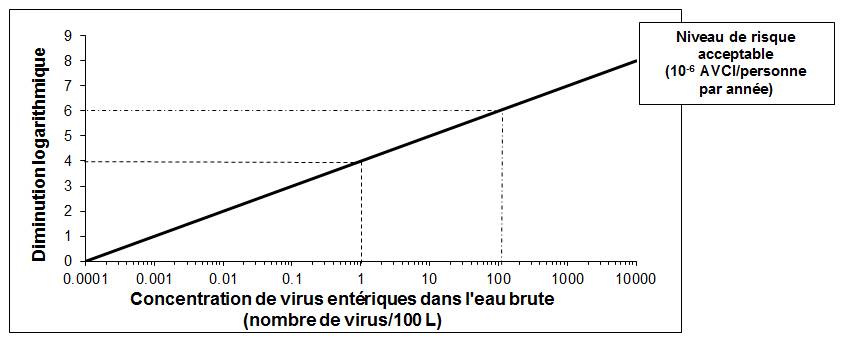
Figure 1 - Équivalent textuel
Text equivalent : Le degré de traitement requis pour atteindre un niveau de risque acceptable selon une consommation de 1 litre, pour des concentrations de virus entériques dans l'eau brute allant de 0,0001 virus pour 100 litres à 10 000 virus pour 100 litres, est présenté sous forme de graphique. L'axe des x correspond aux concentrations de virus entériques pour 100 litres d'eau brute, selon une échelle logarithmique. L'axe des y correspond à l'élimination logarithmique, selon une échelle linéaire. La relation entre les valeurs de l'axe des x et celles de l'axe des y, pour un niveau de risque de 10-6 AVCI par personne par année, forme une diagonale. Sur le graphique, la ligne pointillée horizontale part de l'axe des y à une valeur d'élimination d'environ 4 log, puis croise la diagonale à une valeur d'environ 1 virus entérique pour 100 litres d'eau brute. La diagonale indique que, lorsque les concentrations de virus dans l'eau brute augmentent, l'élimination logarithmique requis pour maintenir 10-6 DALY par personne et par an augmente également.
8.4 Considérations internationales
Des gouvernements et des organismes internationaux à toute les échelles se fient de plus en plus aux ÉQRM pour prendre des décisions éclairées au sujet des risques sanitaires que représentent les pathogènes contenus dans l'eau potable. L'OMS, la Commission européenne, les Pays-Bas, l'Australie et les États-Unis ont tous réalisé des progrès importants au niveau de la validation et des méthodes d'ÉQRM (Staatscourant, 2001; Medema et coll., 2006; NRMMC-EPHC, 2006; U.S. EPA, 2006a,b; OMS, 2017). Ces organismes et ces gouvernements ont adopté des approches qui utilisent l'ÉQRM pour éclairer l'établissement des objectifs en matière de santé (c.-à-d. les niveaux de risque de référence ou les niveaux de maladie) et la gestion des risques (p. ex. plans de salubrité de l'eau décrits dans OMS, 2017). L'U.S. EPA (2014) et la Commission européenne (Medema et coll., 2006), dans le cadre du projet Microrisk, ont tous deux publié des documents d'orientation sur l'ÉQRM de l'eau potable.
Les Pays-Bas et l'U.S. EPA ont fourni deux exemples d'approches réglementaires fondées sur les ÉQRM. Aux Pays-Bas, les fournisseurs d'eau doivent, conformément à l'approche employée par l'OMS, réaliser une ÉQRM propre au site pour tous les systèmes d'approvisionnement en eau de surface afin de déterminer si ceux-ci peuvent se conformer à un niveau de risque précisé. Les autorités hollandaises peuvent aussi exiger une ÉQRM dans le cas des approvisionnements en eau souterraine vulnérables. Aux États-Unis, l'EPA a évalué récemment les risques pour la santé attribuables aux pathogènes d'origine hydrique au moyen d'ÉQRM et a appliqué ces données pour établir des exigences nationales relatives à l'efficacité du traitement (U.S. EPA, 2006a, 2006c). En général, pour les réseaux de distribution d'eau potable, il faut parvenir à une élimination ou une inactivation des virus entériques d'au moins 4 log pour contrer le risque qu'ils représentent (U.S. EPA, 2006a, 2006c).
Santé Canada et le Comité fédéral-provincial-territorial sur l'eau potable ont adopté la même approche que l'OMS (2011) en fournissant des objectifs de rendement fondés sur les ÉQRM comme exigences minimales. L'ÉQRM particulière à un site est recommandée également comme outil qu'il est possible d'utiliser dans le cadre d'une approche de la source au robinet. L'ÉQRM propre au site offre plusieurs avantages, notamment 1) la capacité de comparer les risques posés par des groupes d'agents pathogènes représentatifs (p. ex. virus, protozoaires, bactéries) dans une évaluation globale; 2) la transparence des hypothèses; 3) la possibilité de tenir compte de la variabilité et de l'incertitude liées aux estimations; 4) l'élimination des facteurs de sécurité cachés (que les organismes de réglementation peuvent appliquer consciemment à la fin du processus, s’ils le désirent); 5) l’établissement des points de contrôle critiques et des limites propres au site au moyen d’analyses de la sensibilité et 6) les répercussions claires des effets de la gestion d’un système sur un paramètre de santé publique. Le document de Santé Canada (2018) contient d’autres renseignements sur l’utilisation de l’ÉQRM pour l’évaluation d’un site en particulier.
9.0 Justification de la recommandation
Plus de 140 types de virus entériques peuvent infecter les humains. Ces microorganismes pathogènes sont excrétés dans les matières fécales, et parfois dans l'urine, des personnes et des animaux infectés, et peuvent être présents dans les sources d'approvisionnement en eau. Leur présence dans les sources d'approvisionnement en eau varie au fil du temps et peut être considérablement influencée par des conditions météorologiques extrêmes ou par des déversements ou des événements perturbateurs (c.-à-d. augmentations des concentrations de virus associées à ces événements). La meilleure façon de prévenir la présence de virus entériques en concentrations dangereuses dans l'eau potable consiste à appliquer l'approche de la source au robinet.Cette approche englobe la protection des sources d'approvisionnement en eau et le traitement adéquat, assurés grâce au contrôle approprié des procédés, avant la vérification de l'absence de microorganismes indicateurs dans l'eau traitée.
Fixer des objectifs de traitement basés sur la santé permet d'assurer la protection de la santé publique. Or, pour fixer ces objectifs, il faut établir le niveau de risque acceptable. Le Comité fédéral-provincial-territorial sur l'eau potable a retenu le niveau de risque de référence utilisé par l'Organisation mondiale de la Santé, soit 10-6 AVCI/personne par année. Il s'agit d'une décision de gestion des risques qui offre un équilibre entre, d'une part, la charge de morbidité estimée pour les virus entériques et, d'autre part, le manque d'informations sur la fréquence de la prévalence de ces organismes pathogènes dans les sources d'approvisionnement en eau, les limites de la surveillance des maladies et les variations de l'efficacité des différentes procédés de traitement de l'eau.
L'approche basée sur les ÉQRM utilisée dans le présent document montre que si une source d'approvisionnement en eau présente une concentration virale de 1/100 L, le traitement de l'eau (c.-à-d. élimination ou inactivation) devrait constamment diminuer d'au moins 4 log la concentration des virus afin de respecter le niveau de référence de 10-6AVCI/personne par année. Par conséquent, dans le cas des sources vulnérables à la contamination virale, l'élimination ou l'inactivation d'au moins 4 log des virus entériques a été établie comme objectif de traitement basé sur la santé. Bon nombre de sources d'approvisionnement en eau au Canada, y compris d'eau souterraine, peuvent nécessiter une diminution supérieure à l'objectif de traitement minimal pour respecter le niveau de risque de référence. Il est recommandé, dans la mesure du possible, de protéger les bassins versants ou les aquifères qui servent de sources d'approvisionnement en eau potable, en réduisant au minimum la contamination par les matières fécales.
10.0 Références
Abbaszadegan, M., Lechevallier, M. et Gerba, C. (2003). Occurrence of viruses in US groundwaters. J. Am. Water Works Assoc., 95(9) : 107–120+12.
Abbaszadegan, M., Mayer, B.K., Ryu, H. et Nwachuku, N. (2007). Efficacy of removal of CCL viruses under enhanced coagulation conditions. Environ. Sci. Technol., 41(3) : 971–977.
Abbaszadegan, M., Monteiro, P., Nwachuku, N., Alum, A. et Ryu, H. (2008). Removal of adenovirus, calicivirus, and bacteriophages by conventional drinking water treatment. J. Environ. Sci. Health Part A Toxic Hazard. Subst. Environ. Eng., 43(2) : 171–177.
Abbaszadegan, M., Stewart, P. et LeChevallier, M. (1999). A strategy for detection of viruses in groundwater by PCR. Appl. Environ. Microbiol., 65(2): 444-449.
Abbaszadegan, M., Stewart, P.W., LeChevallier, M.W. et Gerba, C.P. (1998). Application of PCR technologies for virus detection in groundwater. American Water Works Association, 90740, Denver, Colorado.
Abe, K., Inami, T., Asano, K., Miyoshi, C., Masaki, N., Hayashi, S., Ishikawa, K.-., Takebe, Y., Win, K.M., El‑Zayadi, A.R., Han, K.-. et Zhang, D.Y. (1999). TT virus infection is widespread in the general populations from different geographic regions. J. Clin. Microbiol., 37(8) : 2703–2705.
Adham, S.S. et Jacangelo, J.G. (1994). Assessing the reliability of low pressure membrane systems for microbial removal. Dans : Proceedings of the ASCE Critical Issues in Water and Wastewater Treatment Conference, Boulder, Colorado.
Adham, S.S., Jacangelo, J.G. et Laine, J-M. (1995). Low pressure membranes: assessing integrity. J. Am. Water Works Assoc., 87(3) : 62–75.
Ahmed, S.M., Lopman, B.A. et Levy, K. (2013). A systematic review and meta-analysis of the global seasonality of norovirus. Plos One, 8(10).
Albinana-Gimenez, N., Clemente-Casares, P., Calgua, B., Huguet, J.M., Courtois, S. et Girones, R. (2009). Comparison of methods for concentrating human adenoviruses, polyomavirus JC and noroviruses in source waters and drinking water using quantitative PCR. J. Virol. Methods, 158(1–2) : 104–109.
Albinana-Gimenez, N., Miagostovich, M.P., Calgua, B., Huguet, J.M., Matia, L. et Girones, R. (2009). Analysis of adenoviruses and polyomaviruses quantified by qPCR as indicators of water quality in source and drinking-water treatment plants. Water Res., 43(7) : 2011–2019.
Allen, A. S. (2013). Vulnerability of a fractured bedrock aquifer to emerging sewage-derived contaminants and their use as indicators of virus contamination. Mémoire de maîtrise, Université de Guelph, Guelph (Ontario).
Allen, A., Borchardt, M.A., Kieke Jr., B.A., Dunfield, K., et Parker, B.L. (2017). Virus occurrence in private and public wells in a fractured dolostone aquifer in Canada. Hydrogeol J., 25:1117-1136.
Alspach, B. (2018). Pathogen rejection in potable reuse: The role of NF/RO and importance of integrity testing. J. Am. Water Works Assoc., 110(3): 39–44.
Alspach, B., Delphos, P., Pressman, J., Beaty, J., Cooke, T., Voutchkov, N., Schaefer, J., Noack, R., Marascia, F. et Konstanski, D. (2014). Metrics and methods for MF/UF system optimization. . Dans : Proceedings of the Membrane Technology Conference. American Water Works Association, Denver, Colorado.
Alvarez, M.E. et O'Brien, R.T. (1982). Mechanisms of inactivation of poliovirus by chlorine dioxide and iodine. Appl. Environ. Microbiol., 44(5) : 1064–1071.
Amburgey, J.E., Amirtharajah, A., Brouckaert, B.M. et Spivey, N.C. (2003). An enhanced backwashing technique for improved filter ripening. J. Am. Water Works Assoc., 95(12) : 81–94.
Amundson, D., Lindholm, C., Goyal, S.M. et Robinson, R.A. (1988). Microbial pollution of well water in southeastern minnesota. J. Environ. Sci. Health Part A Environ. Sci. Eng., 23(5) : 453–468.
Amvrosieva, T.V., Titov, L.P., Mulders, M., Hovi, T., Dyakonova, O.V., Votyakov, V.I., Kvacheva, Z.B., Eremin, V.F., Sharko, R.M., Orlova, S.V., Kazinets, O.N. et Bogush, Z.F. (2001). Viral water contamination as the cause of aseptic meningitis outbreak in Belarus. Cent. Eur. J. Public Health, 9(3) : 154–157.
Anderson, A.D., Heryford, A.G., Sarisky, J.P., Higgins, C., Monroe, S.S., Beard, R.S., Newport, C.M., Cashdollar, J.L., Fout, G.S., Robbins, D.E., Seys, S.A., Musgrave, K.J., Medus, C., Vinjé, J., Bresee, J.S., Mainzer, H.M. et Glass, R.I. (2003). A waterborne outbreak of norwalk-like virus among snowmobilers - Wyoming, 2001. J. Infect. Dis., 187(2) : 303–306.
Anderson, W.B., Deloyde, J.L., Van Dyke, M.I. et Huck, P.M. (2009). Influence of design and operating conditions on the removal of MS2 bacteriophage by pilot-scale multistage slow sand filtration. J. Water Supply Res. Technol. Aqua, 58(7) : 450–462.
ANSI/AWWA. (2010). ANSI/AWWA B110-09 – Membrane systems. American Water Works Association, Denver, Colorado.
ANSI/AWWA. (2014). ANSI/AWWA C652-14 – Disinfecting water mains. American Water Works Association, Denver, Colorado.
Antony, A., Blackbeard, J. et Leslie, G. (2012). Removal efficiency and integrity monitoring techniques for virus removal by membrane processes. Crit. Rev. Environ. Sci. Technol., 42(9) : 891–933.
Aoki, Y., Suto, A., Mizuta, K., Ahiko, T., Osaka, K. et Matsuzaki, Y. (2010). Duration of norovirus excretion and the longitudinal course of viral load in norovirus-infected elderly patients. J. Hosp. Infect., 75(1) : 42–46.
APHA, AWWA et WEF (1998). Standard methods for the examination of water and wastewater. 20th edition. American Public Health Association, American Water Works Association and Water Environment Federation, Washington, DC.
APHA, AWWA et WEF (2005). Standard methods for the examination of water and wastewater. 21st edition. American Public Health Association, American Water Works Association and Water Environment Federation, Washington, DC.
Armon, R.H. (2015). Ch. 35: Indicators of waterborne viruses. Dans : Environmental indicators. Armon, R.H. et Hanninen, O. (éditeurs). Springer, Dordrecht, Pays-Bas.
Arnold, W.R. et Rainbow, A.J. (1996). Host cell reactivation of irradiated adenovirus in UV-sensitive chinese hamster ovary cell mutants. Mutagenesis, 11(1) : 89–94.
Ashbolt, N.J., Grabow, W.O.K. et Snozzi, M. (2001). Indicators of microbial water quality. Dans : Water quality: Guidelines, standards and health—Assessment of risk and risk management for water-related infectious disease. Fewtrell, L. et Bartram, J. (éditeurs). IWA Publishing pour le compte de l'Organisation mondiale de la Santé, Londres, Royaume-Uni.
ASPC (2010). Fiche technique santé-sécurité : agents pathogènes – Adénovirus (sérotypes 40 et 41). Disponible à : https://www.canada.ca/fr/sante-public/services/biosecurite-biosurete-laboratoire/fiches-techniques-sante-securite-agents-pathogenes-evaluation-risques/adenovirus-serotypes-40-41.html
ASPC (2015a). Liste actuelle des maladies à déclaration obligatoire à l'échelle nationale. Disponible à : http://maladies.canada.ca/declaration-obligatoire/liste-maladies
ASPC (2015b). Guide canadien d'immunisation : Partie 4 – Agents d'immunisation active : vaccin contre l'hépatite A. Disponible à : https://www.canada.ca/fr/sante-public/services/publications/vie-saine/guide-canadien-immunisation-partie-4-agents-immunisation-active/page-6-vaccin-contre-hepatite-a.html
ASTM (2004). ASTM D244-92: Standard practice for recovery of enteroviruses from waters. American Society for Testing and Materials, Philadelphia, Pennsylvanie.
ASTM (2010). Standard practice for integrity testing of water filtration membrane systems. Designation: D6908-06 (Reapproved 2010). ASTM International, West Conshohocken, Pennsylvanie.
ASTM (2014). Standard practices for detecting leaks in reverse osmosis and nanofiltration devices. Designation: D3923-08 (Reapproved 2014). ASTM International, West Conshohocken, Pennsylvanie.
Atmar, R.L. et Estes, M.K. (2001). Diagnosis of noncultivatable gastroenteritis viruses, the human caliciviruses. Clin. Microbiol. Rev., 14(1) : 15–37.
Atmar, R.L., Opekun, A.R., Gilger, M.A., Estes, M.K., Crawford, S.E., Neill, F.H. et Graham, D.Y. (2008). Norwalk virus shedding after experimental human infection. Emerg. Infect. Dis., 14(10) : 1553–1557.
Aw, T.G. et Gin, K.Y. (2010). Environmental surveillance and molecular characterization of human enteric viruses in tropical urban wastewaters. J. Appl. Microbiol., 109(2) : 716–730.
AWWA (1999). Waterborne pathogens. AWWA manual of water practices M48. American Water Works Association, Denver, Colorado.
AWWA (2005). Microfiltration and ultrafiltration membranes for drinking water. AWWA manual of water practices M53. American Water Works Association, Denver, Colorado.
AWWA (2006). Waterborne pathogens. AWWA manual of water practices M48. 2nd edition. AWWA manual of water practices M48. American Water Works Association, Denver, Colorado.
AWWA (2011a). Water quality and treatment: A handbook of community water supplies. 6th edition. Edzwald, J.K. (ed.). McGraw-Hill, New York, New York.
AWWA (2011b). Operational control of coagulation and filtration processes: Manual of water supply practices – M37. American Water Works Association, Denver, Colorado.
AWWA (2015). Disinfection residual strategy panel memorandum: Panel findings. July 31, 2015. American Water Works Association, Denver, Colorado.
Azadpour-Keeley, A., Faulkner, B.R. et Chen, J. (2003). Movement and longevity of viruses in the subsurface. U.S. Environmental Protection Agency, Cincinnati, Ohio. (Rapport no EPA/540/S-03/500).
Baer, G.M. et Walker, J.A. (1977). Studies of an outbreak of acute hepatitis A: I. complement level fluctuation. J. Med. Virol., 1(1) : 1–7.
Bales, R.C., Gerba, C.P., Grondin, G.H. et Jensen, S.L. (1989). Bacteriophage transport in sandy soil and fractured tuff. Appl. Environ. Microbiol., 55(8) : 2061–2067.
Bales, R.C., Hinkle, S.R., Kroeger, T.W., Stocking, K. et Gerba, C.P. (1991). Bacteriophage adsorption during transport through porous media: Chemical perturbations and reversibility. Environ. Sci. Technol., 25(12) : 2088–2095.
Bales, R.C., Li, S., Maguire, K.M., Yahya, M.T. et Gerba, C.P. (1993). MS-2 and poliovirus transport in porous media: Hydrophobic effects and chemical perturbations. Water Resour. Res., 29(4) : 957–963.
Bales, R.C., Li, S., Yeh, T.-.J., Lenczewski, M.E. et Gerba, C.P. (1997). Bacteriophage and microsphere transport in saturated porous media: Forced-gradient experiment at Borden, Ontario. Water Resour. Res., 33(4) : 639–648.
Bales, R.C., Shimin, L., Maguire, K.M., Yahya, M.T., Gerba, C.P. et Harvey, R.W. (1995). Virus and bacteria transport in a sandy aquifer, Cape Cod, Massachusetts. Groundwater, 33(4) : 653–661.
Balkin, H.B. et Margolin, A.B. (2010). Detection of poliovirus by ICC/qPCR in concentrated water samples has greater sensitivity and is less costly using BGM cells in suspension as compared to monolayers. Virol. J., 7 : 282–422.
Ball, J.M., Tian, P., Zeng, C.Q., Morris, A.P. et Estes, M.K. (1996). Age-dependent diarrhea induced by a rotaviral nonstructural glycoprotein. Science, 272(5258) : 101–104.
Ballester, N.A., Fontaine, J.H. et Margolin, A.B. (2005). Occurrence and correlations between coliphages and anthropogenic viruses in the Massachusetts Bay using enrichment and ICC-nPCR. J. Water. Health, 3(1) : 59–68.
Banks, W.S.L. et Battigelli, D.A. (2002). Occurrence and distribution of microbiological contamination and enteric viruses in shallow ground water in Baltimore and Harford Counties, Maryland. U.S.G.S., Water resources investigations report 01-4216.
Banks, W.S.L., Klohe, C.A. et Battigelli, D.A. (2001). Occurrence and distribution of enteric viruses in shallow ground water and factors affecting well vulnerability to microbiological contamination in Worcester and Wicomico Counties, Maryland. U.S.G.S., Water resources investigations report 01-4147.
Banyai, K., Bogdan, A., Domonkos, G., Kisfali, P., Molnar, P., Toth, A., Melegh, B., Martella, V., Gentsch, J.R. et Szucs, G. (2009). Genetic diversity and zoonotic potential of human rotavirus strains, 2003-2006, Hungary. J. Med. Virol., 81(2) : 362–370.
Batch, L.F., Schulz, C.R. et Linden, K.G. (2004). Evaluating water quality effects on UV disinfection of MS2 coliphage. J. Am. Water Works Assoc., 96(7) : 75–87.
Bauer, R., Dizer, H., Graeber, I., Rosenwinkel, K.H. et Lopez-Pila, J.M. (2011). Removal of bacterial faecal indicators, coliphages and enteric adenoviruses from waters with high faecal pollution by slow sand filtration. Water Res., 45(2) : 439–452.
Baxter, C.S., Hofmann, R., Templeton, M.R., Brown, M. et Andrews, R.C. (2007). Inactivation of adenovirus types 2, 5, and 41 in drinking water by UV light, free chlorine, and monochloramine. J. Environ. Eng., 133(1) : 95–103.
BC Provincial Health Officer (2001). Report on the health of British Columbians. Public Health Officer's annual report 2000: Drinking water quality in British Columbia—The public health perspective. Ministry of Health Planning, Victoria, (C.-B.).
Beck, S.E., Rodriguez, R.A., Linden, K.G., Hargy, T.M., Larason, T.C. et Wright, H.B. (2014). Wavelength dependent UV inactivation and DNA damage of adenovirus as measured by cell culture infectivity and long range quantitative PCR. Environ. Sci. Technol., 48(1) : 591–598.
Beer, K.D., Gargano, J.W., Roberts, V.A., Hill, V.R., Garrison, L.E., Kutty, P.K., Hilborn, E.D., Wade, T.J., Fullerton, K.E. et Yoder, J.S. (2015). Surveillance for waterborne disease outbreaks associated with drinking water - United States, 2011-2012. MMWR Morb. Mortal. Wkly. Rep., 64(31) : 842–848.
Belan, K.P. (2010). Characterizing a fractured rock aquifer with hydraulic testing at a contaminated municipal well using flexible liner methods and depth discrete monitoring. Mémoire de maîtrise, Université de Guelph, Guelph (Ontario).
Bell, K., LeChevallier, M., Abbaszadegan, M., Amy, G., Sinha, S., Benjamin, M. et Ibrahim, E. (1998). Enhanced and optimized coagulation for particulate and microbial removal. AWWA Research Foundation and American Water Works Association, Denver, Colorado.
Bellamy, W.D., Hendricks, D.W. et Logsdon, G.S. (1985). Slow sand filtration: Influences of selected process variables. J. Am. Water Works Assoc., 77(12) : 62–66.
Beller, M., Ellis, A., Lee, S.H., Drebot, M.A., Jenkerson, S.A., Funk, E., Sobsey, M.D., Simmons III, O.D., Monroe, S.S., Ando, T., Noel, J., Petric, M., Middaugh, J.P. et Spika, J.S. (1997). Outbreak of viral gastroenteritis due to a contaminated well: International consequences. J. Am. Med. Assoc., 278(7) : 563–568.
Bellou, M.I., Syngouna, V.I., Tselepi, M.A., Kokkinos, P.A., Paparrodopoulos, S.C., Vantarakis, A. et Chrysikopoulos, C.V. (2015). Interaction of human adenoviruses and coliphages with kaolinite and bentonite. Sci. Total Environ., 517 : 86–95.
Bendinelli, M., Pistello, M., Maggi, F., Fornai, C., Freer, G. et Vatteroni, M.L. (2001). Molecular properties, biology, and clinical implications of TT virus, a recently identified widespread infectious agent of humans. Clin. Microbiol. Rev., 14(1) : 98–113.
Berger, P. (1994). Regulation related to groundwater contamination: The draft groundwater disinfection rule. Dans : Groundwater contamination and control. Zoller, U. (éditeurs). M. Dekker, New York, p. 645–659.
Betanzo, E.W., Hofmann, R., Hu, Z., Baribeau, H. et Alam, Z. (2008). Modeling the impact of microbial intrusion on secondary disinfection in a drinking water distribution system. J. Environ. Eng., 134(4) : 231–237.
Beuret, C. (2003). A simple method for isolation of enteric viruses (noroviruses and enteroviruses) in water. J. Virol. Methods, 107(1) : 1–8.
Biagini, P. (2004). Human circoviruses. Vet. Microbiol., 98(2) : 95–101.
Bitton, G. (1975). Adsorption of viruses onto surfaces in soil and water. Water Res., 9(5–6) : 473–484.
Bitton, G. (1999). Wastewater microbiology. 2e édition. Wiley-Liss, New York, New York.
Blackburn, B.G., Craun, G.F., Yoder, J.S., Hill, V., Calderon, R.L., Chen, N., Lee, S.H., Levy, D.A. et Beach, M.J. (2004). Surveillance for waterborne-disease outbreaks associated with drinking water–United States, 2001-2002. MMWR Surveill. Summ., 53(8) : 23–45.
Blanton, L.H., Adams, S.M., Beard, R.S., Wei, G., Bulens, S.K., Widdowson, M.A., Glass, R.I. et Monroe, S.S. (2006). Molecular and epidemiologic trends of caliciviruses associated with outbreaks of acute gastroenteritis in the United States, 2000-2004. J. Infect. Dis., 193(3) : 413–421.
Bloch, A.B., Stramer, S.L., Smith, J.D., Margolis, H.S., Fields, H.A., McKinley, T.W., Gerba, C.P., Maynard, J.E. et Sikes, R.K. (1990). Recovery of hepatitis A virus from a water supply responsible for a common source outbreak of hepatitis A. Am. J. Public Health, 80(4) : 428–430.
Blokker, M., Smeets, P. et Medema, G. (2014). QMRA in the drinking water distribution system. Procedia Eng., p. 151‑159.
Boccia, D., Tozzi, A.E., Benvon, C., Rizzo, C., Russo, T., Buttinelli, G., Caprioli, A., Marziano, M.L. et Ruggeri, F.M. (2002). Waterborne outbreak of norwalk-like virus gastroenteritis at a tourist resort, Italy. Emerg. Infect. Dis., 8(6) : 563–568.
Boettger, B. (1995). Kaslo waterborne disease outbreak. B.C. Health Dis. Surveill, 4 : 122–125.
Bofill-Mas, S. et Girones, R. (2003). Role of the environment in the transmission of JC virus. J. Neurovirol., 9 Suppl 1 : 54–58.
Bolton, J.R. et Cotton, C.A. (2008). The ultraviolet disinfection handbook. American Water Works Association, Denver, Colorado.
Bonilla, J.A., Bonilla, T.D., Abdelzaher, A.M., Scott, T.M., Lukasik, J., Solo-Gabriele, H.M. et Palmer, C.J. (2015). Quantification of protozoa and viruses from small water volumes. Int. J. Environ. Res. Public. Health., 12(7) : 7118–7132.
Borchardt, M.A., Bertz, P.D., Spencer, S.K. et Battigelli, D.A. (2003). Incidence of enteric viruses in groundwater from household wells in Wisconsin. Appl. Environ. Microbiol., 69(2) : 1172–1180.
Borchardt, M.A., Bradbury, K.R., Alexander, E.C., Kolberg, R.J., Alexander, S.C., Archer, J.R., Braatz, L.A., Forest, B.M., Green, J.A. et Spencer, S.K. (2011). Norovirus outbreak caused by a new septic system in a dolomite aquifer. Ground Water, 49(1) : 85–97.
Borchardt, M.A., Bradbury, K.R., Gotkowitz, M.B., Cherry, J.A. et Parker, B.L. (2007). Human enteric viruses in groundwater from a confined bedrock aquifer. Environ. Sci. Technol., 41(18) : 6606–6612.
Borchardt, M.A., Haas, N.L. et Hunt, R.J. (2004). Vulnerability of drinking-water wells in La Crosse, Wisconsin, to enteric-virus contamination from surface water contributions. Appl. Environ. Microbiol., 70(10) : 5937–5946.
Borchardt, M.A., Spencer, S.K., Kieke, B.A., Lambertini, E. et Loge, F.J. (2012). Viruses in nondisinfected drinking water from municipal wells and community incidence of acute gastrointestinal illness. Environ. Health Perspect., 120(9) : 1272–1279.
Bosch, A., Guix, S., Sano, D. et Pinto, R.M. (2008). New tools for the study and direct surveillance of viral pathogens in water. Curr. Opin. Biotechnol., 19(3) : 295–301.
Bosch, A., Pinto, R.M., Comas, J. et Abad, F.X. (2004). Detection of infectious rotaviruses by flow cytometry. Methods Mol. Biol., 268 : 61–68.
Boudaud, N., Machinal, C., David, F., Freval-Le Bourdonnec, A., Jossent, J., Bakanga, F., Arnal, C., Jaffrezic, M.P., Oberti, S. et Gantzer, C. (2012). Removal of MS2, qbeta and GA bacteriophages during drinking water treatment at pilot scale. Water Res., 46(8) : 2651–2664.
Bradbury, K.R., Borchardt, M.A., Gotkowitz, M., Spencer, S.K., Zhu, J. et Hunt, R.J. (2013). Source and transport of human enteric viruses in deep municipal water supply wells. Environ. Sci. Technol., 47(9) : 4096–4103.
Bradbury, K.R., Gotkowitz, M.B., Hart, D. J., Eaton, T.T., Cherry, J.A., Parker, B.L. et Borchardt, M.A. (2006). Contaminant transport through aquitards: Technical guidance for aquitard assessment. Awwa Research Foundation, Denver, CO.
Brassard, J., Seyer, K., Houde, A., Simard, C. et Trottier, Y.L. (2005). Concentration and detection of hepatitis A virus and rotavirus in spring water samples by reverse transcription-PCR. J. Virol. Methods, 123(2) : 163–169.
Bremer, J.E. et Harter, T. (2012). Domestic wells have high probability of pumping septic tank leachate. Hydrol. Earth Syst. Sci., 16(8) : 2453–2467.
Brown, C.M., Cann, J.W., Simons, G., Fankhauser, R.L., Thomas, W., Parashar, U.D. et Lewis, M.J. (2001). Outbreak of norwalk virus in a Carribbean island resort: Application of molecular diagnostics to ascertain the vehicle of infection. Epidemiol. Infect., 126(3) : 425–432.
Brugha, R., Vipond, I.B., Evans, M.R., Sandifer, Q.D., Roberts, R.J., Salmon, R.L., Caul, E.O. et Mukerjee, A.K. (1999). A community outbreak of food-borne small round-structured virus gastroenteritis caused by a contaminated water supply. Epidemiol. Infect., 122(1) : 145–154.
Brunkard, J.M., Ailes, E., Roberts, V.A., Hill, V., Hilborn, E.D., Craun, G.F., Rajasingham, A., Kahler, A., Garrison, L., Hicks, L., Carpenter, J., Wade, T.J., Beach, M.J., Yoder Msw, J.S. et CDC (2011). Surveillance for waterborne disease outbreaks associated with drinking water-United States, 2007-2008. MMWR Surveill. Summ., 60(12) : 38–68.
Candy, D.C. (2007). Rotavirus infection: A systemic illness? PLoS Med., 4(4) : e117.
Cantera, J.L., Chen, W. et Yates, M.V. (2010). Detection of infective poliovirus by a simple, rapid, and sensitive flow cytometry method based on fluorescence resonance energy transfer technology. Appl. Environ. Microbiol., 76(2) : 584–588.
Carlson, G.F.,Jr, Woodard, F.E., Wentworth, D.F. et Sproul, O.J. (1968). Virus inactivation on clay particles in natural waters. J. Water Pollut. Control Fed., 40(2) : R89–106.
Carrique-Mas, J., Andersson, Y., Petersén, B., Hedlund, K.O., Sjögren, N. et Giesecke, J. (2003). A norwalk-like virus waterborne community outbreak in a Swedish village during peak holiday season. Epidemiol. Infect., 131(1) : 737–744.
Carter, M.J. (2005). Enterically infecting viruses: Pathogenicity, transmission and significance for food and waterborne infection. J. Appl. Microbiol., 98(6) : 1354–1380.
Cashdollar, J.L., Brinkman, N.E., Griffin, S.M., McMinn, B.R., Rhodes, E.R., Varughese, E.A., Grimm, A.C., Parshionikar, S.U., Wymer, L. et Fout, G.S. (2013). Development and evaluation of EPA method 1615 for detection of enterovirus and norovirus in water. Appl. Environ. Microbiol., 79(1) : 215–223.
Casteel, M.J., Sobsey, M.D. and Mueller, J.P. (2006). Fecal contamination of agricultural soils before and after hurricane-associated flooding in North Carolina. J. Environ. Sci. Health Part A Toxic Hazard. Subst. Environ. Eng., 41(2): 173-184.
Caul, E.O. (1994). Small round structured viruses: airborne transmission and hospital control. Lancet, 343(8908) : 1240–1242.
Caul, E.O. (1996). Viral gastroenteritis: Small round structured viruses, caliciviruses and astroviruses. part II. the epidemiological perspective. J. Clin. Pathol., 49(12) : 959–964.
Caul, E.O., Ashley, C.R., Darville, J.M. et Bridger, J.C. (1990). Group C rotavirus associated with fatal enteritis in a family outbreak. J. Med.Virol, 30: 201–205.
CCME (2004). De la source au robinet : guide d'application de l'approche à barrières multiples pour une eau potable saine. Conseil canadien des ministres de l'environnement, Winnipeg (Manitoba).
CCN (2016). Répertoire des organismes de certification accrédités de produits, de processus et de services, Conseil canadien des normes, Ottawa (Ontario). Disponible à : www.scc.ca/fr/accreditation/certification-de-produits-procedes-et-services/repertoire-des-organismes-de-certification-accredites.
CDC, (2011). Updated norovirus outbreak management and disease prevention guidelines. MMWR Recomm Rep., 60(RR-3) : 1–18.
CDC (2013a). Norovirus clinical overview. Disponible à : www.cdc.gov/norovirus/hcp/clinical-overview.html.
CDC (2013b). Surveillance for waterborne disease outbreaks associated with drinking water and other nonrecreational water – United States, 2009-2010. MMWR Morb. Mortal. Wkly. Rep., 62(35) : 714–720.
CDC (2015a). Viral hepatitis – hepatitis A information. Disponible à : www.cdc.gov/hepatitis/hav/havfaq.htm.
CDC (2015b). Chapter 19: Rotavirus. Dans : Epidemiology and prevention of vaccine-preventable diseases. Hamborsky, J., Kroger, A. et Wolfe, S. (éditeurs). 13th. Public Health Foundation, Washington, DC.
Chang, J.C., Ossoff, S.F., Lobe, D.C., Dorfman, M.H., Dumais, C.M., Qualls, R.G. et Johnson, J.D. (1985). UV inactivation of pathogenic and indicator microorganisms. Appl. Environ. Microbiol., 49(6) : 1361–1365.
Chao, D.Y., Wei, J.Y., Chang, W.F., Wang, J. et Wang, L.C. (2012). Detection of multiple genotypes of calicivirus infection in asymptomatic swine in Taiwan. Zoonoses Public. Health., 59(6) : 434–444.
Charles, K.J., Shore, J., Sellwood, J., Laverick, M., Hart, A. et Pedley, S. (2009). Assessment of the stability of human viruses and coliphage in groundwater by PCR and infectivity methods. J. Appl. Microbiol., 106(6) : 1827-1837.
Cheetham, S., Souza, M., McGregor, R., Meulia, T., Wang, Q. et Saif, L.J. (2007). Binding patterns of human norovirus-like particles to buccal and intestinal tissues of gnotobiotic pigs in relation to A/H histo-blood group antigen expression. J. Virol., 81(7) : 3535–3544.
Cherry, J.D. (1992). Enteroviruses: Poloviruses (poliomyelitis), coxsackieviruses, echoviruses, and enteroviruses. Dans : Textbook of pediatric infectious diseases. Feigin, R.D. (éditeurs). 3rd. W.B. Saunders Co., Philadelphie, Pennsylvanie., p. 3535–3544.
Cherry, J.A., Parker, B.L., Bradbury, K.R., Eaton, T.T., Gotkowitz, M.B., Hart, D.J. et Borchardt, M.A. (2006). Contaminant transport through aquitards: A state-of-the-science review. Awwa Research Foundation, Denver, Colorado.
Chevrefils, G., Caron, É., Wright, H., Sakamoto, G., Payment, P., Barbeau, B. et Cairns, B. (2006). UV dose required to achieve incremental log inactivation of bacteria, protozoa and viruses. J. Int. UV Assoc. (IUVA) News, 8 : 38–45.
Chia, J.K. et Chia, A.Y. (2008). Chronic fatigue syndrome is associated with chronic enterovirus infection of the stomach. J. Clin. Pathol., 61(1) : 43–48.
Chiba, S., Nakata, S., Numata-Kinoshita, K. and Honma, S. (2000). Sapporo virus: history and recent findings. J. Infect. Dis., 181(Suppl. 2) : S303–S308.
Chiba, S., Sakuma, Y., Kogasaka, R., Akihara, M., Horino, K., Nakao, T. et Fukui, S. (1979). An outbreak of gastroenteritis associated with calicivirus in an infant home. J. Med. Virol., 4(4) : 249–254.
Chin, J. (2000). Control of communicable diseases manual. 17th edition.. American Public Health Association, Washington, DC.
Choi, S. et Jiang, S.C. (2005). Real-time PCR quantification of human adenoviruses in urban rivers indicates genome prevalence but low infectivity. Appl. Environ. Microbiol., 71(11) : 7426–7433.
Christensen, J. et Linden, K. (2002). New findings regarding the impacts of suspended particles on UV disinfection of drinking water. Dans : Proceedings of the Annual Conference of the American Water Works Association, New Orleans, LA. 16 au 29 juin.
Clemente-Casares, P., Pina, S., Buti, M., Jardi, R., Martín, M., Bofill-Mas, S. et Girones, R. (2003). Hepatitis E virus epidemiology in industrialized countries. Emerg. Infect. Dis., 9(4) : 448–454.
Colford, J.M., Wade, T.J., Sandhu, S.K., Wright, C.C., Lee, S., Shaw, S., Fox, K., Burns, S., Benker, A., Brookhart, M.A., Van Der Laan, M. et Levy, D.A. (2005). A randomized, controlled trial of in-home drinking water intervention to reduce gastrointestinal illness. Am. J. Epidemiol., 161(5): 472-482.
Colford, J.M., Roy, S.L., Beach, M.J., Hightower, A., Shaw, S.E. et Wade, T.J. (2006). A review of household drinking water intervention trials and an approach to the estimation of endemic waterborne gastroenteritis in the united states. J. Water Health, 4(SUPPL. 2): 71-88.
Colford, J.M., Hilton, J.F., Wright, C.C., Arnold, B.F., Saha, S., Wade, T.J., Scott, J. et Eisenberg, J.N.S. (2009). The sonoma water evaluation trial: A randomized drinking water intervention trial to reduce gastrointestinal illness in older adults. Am. J. Public Health, 99(11): 1988-1995.
Conboy, M.J. et Goss, M.J. (2000). Natural protection of groundwater against bacteria of fecal orgin. J. Contam. Hydrol., 43:1-24.
Conio, O., Chioetto, M. et Hargesheimer, E. (2002). Organic monitors. Dans : Online monitoring for drinking water utilities. Hargesheimer, E., Conio, O. and Papovicova, J. (éditeurs). AWWA Research Foundation, Denver, Colorado, p. 163–202.
Cook, N., Bridger, J., Kendall, K., Gomara, M.I., El-Attar, L. et Gray, J. (2004). The zoonotic potential of rotavirus. J. Infect., 48(4) : 289–302.
Cornwell, D.A. et MacPhee, M.J. (2001). Effects of spent filter backwash recycle on cryptosporidium removal. J. Am. Water Works Assoc., 93(4) : 153–162.
Corsi, S.R., Borchardt, M.A., Spencer, S.K., Hughes, P.E. et Baldwin, A.K. (2014). Human and bovine viruses in the Milwaukee River watershed: Hydrologically relevant representation and relations with environmental variables. Sci. Total Environ., 490 : 849–860.
Cotton, C.A., Linden, K.G., Schmelling, D.C., Bell, C. et Landis, H. (2001). The development of the UV dose tables for LT2ESWTR implementation. First International Congress on Ultraviolet Technologies, Washington, DC.
Craun, M.F., Craun, G.F., Calderon, R.L. et Beach, M.J. (2006). Waterborne outbreaks reported in the United States. J. Water. Health., 4 Suppl 2 : 19–30.
Cromeans, T.L., Kahler, A.M. et Hill, V.R. (2010). Inactivation of adenoviruses, enteroviruses, and murine norovirus in water by free chlorine and monochloramine. Appl. Environ. Microbiol., 76(4) : 1028–1033.
Crowe, A.S., Schaefer, K.A., Kohut, A., Shikaze, S.G. et Ptacek, C.J. (2003). Science de l'eau et politiques : qualité des eaux souterraines, Conseil canadien des ministres de l'Environnement, Winnipeg, Manitoba.
Curriero, F.C., Patz, J.A., Rose, J.B. et Lele, S. (2001). The association between extreme precipitation and waterborne disease outbreaks in the United States, 1948-1994. Am. J. Public Health, 91(8) : 1194–1199.
De Serres, G., Cromeans, T.L., Levesque, B., Brassard, N., Barthe, C., Dionne, M., Prud'homme, H., Paradis, D., Shapiro, C.N., Nainan, O.V. et Margolis, H.S. (1999). Molecular confirmation of hepatitis A virus from well water: Epidemiology and public health implications. J. Infect. Dis., 179 : 37–43.
DeBorde, D.C., Woessner, W.W., Kiley, Q.T. and Ball, P. (1999). Rapid transport of viruses in a floodplain aquifer. Water Res., 33(10) : 2229–2238.
DeBorde, D.C., Woessner, W.W., Lauerman, B. et Ball, P.N. (1998). Virus occurrence and transport in a school septic system and unconfined aquifer. Ground Water, 36(5) : 825–834.
Dechesne, M. et Soyeux, E. (2007). Assessment of source water pathogen contamination. J. Water Health., 5 Suppl 1 : 39–50.
Deere, D., Stevens, M., Davison, A., Helm, G. et Dufour, A. (2001). Management strategies. Fewtrell, L. et Bartram, J. (éditeurs). IWA Publishing, Londres, Royaume-Uni.
De Roda Husman, A.M., Bijkerk, P., Lodder, W., van den Berg, H., Pribil, W., Cabaj, A., Gehringer, P., Sommer, R., and Duizer, E. (2004). Calicivirus Inactivation by nonionizing (253.7-nanometer-wavelength UV and ionizing (gamma) radiation. Appl. Environ. Microbiol., 70(9): 5089-5093.
De Roda Husman, A.M., Lodder, W.J., Rutjes, S.A., Schijven, J.F. et Teunis, P.F.M. (2009). Long-term inactivation study of three enteroviruses in artificial surface and groundwaters, using PCR and cell culture. Appl. Environ. Microbiol., 75(4) : 1050–1057.
Di Giovanni, G.D., Hashemi, F.H., Shaw, N.J., Abrams, F.A., LeChevallier, M.W. et Abbaszadegan, M. (1999). Detection of infectious cryptosporidium parvum oocysts in surface and filter backwash water samples by immunomagnetic separation and integrated cell culture-PCR. Appl. Environ. Microbiol., 65(8) : 3427–3432.
Diniz-Mendes, L., Paula, V.S., Luz, S.L. et Niel, C. (2008). High prevalence of human torque teno virus in streams crossing the city of Manaus, Brazilian Amazon. J. Appl. Microbiol., 105(1) : 51–58.
Divizia, M., Gabrieli, R., Donia, D., Macaluso, A., Bosch, A., Guix, S., Sanchez, G., Villena, C., Pinto, R.M., Palombi, L., Buonuomo, E., Cenko, F., Leno, L., Bebeci, D. et Bino, S. (2004). Waterborne gastroenteritis outbreak in Albania. Water Sci. Technol., 50(1) : 57–61.
Dizer, H., Grutzmacher, G., Bartel, H., Wiese, H.B., Szewzyk, R. et Lopez-Pila, J.M. (2004). Contribution of the colmation layer to the elimination of coliphages by slow sand filtration. Water Sci. Technol., 50(2) : 211–214.
Dong, Y., Kim, J. et Lewis, G.D. (2010). Evaluation of methodology for detection of human adenoviruses in wastewater, drinking water, stream water and recreational waters. J. Appl. Microbiol., 108(3) : 800–809.
Dorner, S.M., Anderson, W.B., Gaulin, T., Candon, H.L., Slawson, R.M., Payment, P. et Huck, P.M. (2007). Pathogen and indicator variability in a heavily impacted watershed. J. Water Health, 5(2) : 241–257.
Doro, R., Farkas, S.L., Martella, V. et Banyai, K. (2015). Zoonotic transmission of rotavirus: Surveillance and control. Expert Rev. Anti Infect. Ther., 13(11) : 1337–1350.
Douglas, I. (2016). Personal communication. Ottawa, Ontario.
Drayna, P., McLellan, S.L., Simpson, P., Li, S-H. et Gorelick, M.H. (2010). Association between rainfall and pediatric emergency department visits for acute gastrointestinal illness. Environ. Health Perspect., 118(10) : 1439–1443.
Duboise, S.M., Moore, B.E. et Sagik, B.P. (1976). Poliovirus survival and movement in a sandy forest soil. Appl. Environ. Microbiol., 31(4) : 536–543.
Ebacher, G., Besner, M.C., Clément, B. et Prévost, M. (2012). Sensitivity analysis of some critical factors affecting simulated intrusion volumes during a low pressure transient event in a full-scale water distribution system. Water Res., 46(13): 4017–4030.
Edge, T.A., Khan, I.U., Bouchard, R., Guo, J., Hill, S., Locas, A., Moore, L., Neumann, N., Nowak, E., Payment, P., Yang, R., Yerubandi, R. et Watson, S. (2013). Occurrence of waterborne pathogens and escherichia coli at offshore drinking water intakes in Lake Ontario. Appl. Environ. Microbiol., 79(19) : 5799–5813.
Eischeid, A.C. (2009). Fundamental mechanisms in the extreme UV resistance of adenovirus. Mémoire de doctorat. Université Duke. Disponible à : http://dukespace.lib.duke.edu/dspace/bitstream/handle/10161/1256/D_Eischeid_Anne_a_200904.pdf?sequence=1
ElHadidy, A.M., Peldszus, S. et Van Dyke, M.I. (2013). An evaluation of virus removal mechanisms by ultrafiltration membranes using MS2 and fx174 bacteriophage. Sep. Purif. Technol., 120 : 215–223.
ElHadidy, A.M., Peldszus, S. et Van Dyke, M.I. (2014). Effect of hydraulically reversible and hydraulically irreversible fouling on the removal of MS2 and fX174 bacteriophage by an ultrafiltration membrane. Water Res., 61 : 297–307.
Emelko, M.B., Schmidt, P.J. et Reilly, P.M. (2010). Particle and microorganism enumeration data: Enabling quantitative rigor and judicious interpretation. Environ. Sci. Technol., 44(5) : 1720–1727.
Emelko, M.B., Schmidt, P.J. et Roberson, J.A. (2008). Quantification of uncertainty in microbial data - reporting and regulatory implications. J. Am. Water Works Assoc., 100(3) : 94–104+14.
Engelbrecht, R.S., Weber, M.J., Salter, B.L. et Schmidt, C.A. (1980). Comparative inactivation of viruses by chlorine. Appl. Environ. Microbiol., 40(2) : 249–256.
EPCOR (2010). Annual quality assurrance report. Edmonton (Alberta).
EPCOR (2011). Annual quality assurrance report. Edmonton (Alberta).
Espinosa, A.C., Mazari-Hiriart, M., Espinosa, R., Maruri-Avidal, L., Méndez, E. et Arias, C.F. (2008). Infectivity and genome persistence of rotavirus and astrovirus in groundwater and surface water. Water Res., 42(10–11) : 2618–2628.
Estes, M.K et Kapikian, A.Z. (2007). Rotaviruses. 5th. Knipe, D.M. et Howley, P.M. (éditeurs). Lippincott Williams et Wilkins, Philadelphie, Pennsylvanie.
Estes, M.K. et Greenberg, H.B. (2013). Rotaviruses. In: Fields virology. Knipe, D.M. et Howley, P.M. (éditeurs). 6th. Wolters Kluwer Health/Lippincott Williams et Wilkins, Philadelphie, Pennsylvanie., p. 1347–1401.
Faleye, T.O.C., Adewumi, M.O. et Adeniji, J.A. (2016). Defining the enterovirus diversity landscape of a faecal sample: a methodological challenge? Viruses, 8(1).
Farkas, T., Zhong, W.M., Jing, Y., Huang, P.W., Espinosa, S.M., Martinez, N., Morrow, A.L., Ruiz-Palacios, G.M., Pickering, L.K. et Jiang, X. (2004). Genetic diversity among sapoviruses. Arch. Virol., 149(7) : 1309–1323.
FDA (2012). Hepatitis A virus. Dans : Bad bug book, foodborne pathogenic microorganisms and natural toxins. Deuxième édition. U.S. Food and Drug Administration. p. 174.
Feffer, A., Baker, D., Moore, R., Wagner, T. et Gresehover, B. (2016). Remote data monitoring improves distribution system efficiency and maintenance. Opflow, 42(2) : 10–14.
Fiksdal, L. et Leiknes, T. (2006). The effect of coagulation with MF/UF membrane filtration for the removal of virus in drinking water. J. Membr. Sci., 279(1–2) : 364–371.
Finkbeiner, S.R., Holtz, L.R., Jiang, Y., Rajendran, P., Franz, C.J., Zhao, G., Kang, G. et Wang, D. (2009a). Human stool contains a previously unrecognized diversity of novel astroviruses. Virol. J., 6 : 161.
Finkbeiner, S.R., Le, B.M., Holtz, L.R., Storch, G.A. et Wang, D. (2009b). Detection of newly described astrovirus MLB1 in stool samples from children. Emerg. Infect. Dis., 15(3) : 441–444.
Fiore, A.E. (2004). Hepatitis A transmitted by food. Clin. Infect. Dis., 38(5) : 705–715.
Fisher, I., Angles, M., Chandy, J., Cox, P., Warnecke, M., Kastl, G. et Jegatheesan, V. (2000). Biofilms - a sticky situation for drinking water? Water (Australia), 27(2) : 33–37.
Fong, T.T. et Lipp, E.K. (2005). Enteric viruses of humans and animals in aquatic environments: Health risks, detection, and potential water quality assessment tools. Microbiol. Mol. Biol. Rev., 69(2) : 357–371.
Fong, T.T., Griffin, D.W. et Lipp, E.K. (2005). Molecular assays for targeting human and bovine enteric viruses in coastal waters and their application for library-independent source tracking. Appl. Environ. Microbiol., 71(4) : 2070–2078.
Fong, T.T., Mansfield, L.S., Wilson, D.L., Schwab, D.J., Molloy, S.L. et Rose, J.B. (2007). Massive microbiological groundwater contamination associated with a waterborne outbreak in Lake Erie, South Bass Island, Ohio. Environ. Health Perspect., 115(6) : 856–864.
Fongaro, G., Nascimento, M.A., Rigotto, C., Ritterbusch, G., da Silva, A.D., Esteves, P.A. et Barardi, C.R. (2013). Evaluation and molecular characterization of human adenovirus in drinking water supplies: Viral integrity and viability assays. Virol. J., 10 : 166.
Fongaro, G., Padilha, J., Schissi, C.D., Nascimento, M.A., Bampi, G.B., Viancelli, A. et Barardi, C.R. (2015). Human and animal enteric virus in groundwater from deep wells, and recreational and network water. Environ. Sci. Pollut. Res. Int., 22(24) : 20060–20066.
Fout, G.S., Borchardt, M.A., Kieke, B.A. et Karim, M.R. (2017). Human virus and microbial indicator occurrence in public-supply groundwater systems: Meta-analysis of 12 international studies. Hydrogeol. J., 25(4): 903-919.
Fout, G.S., Cashdollar, J.L., Varughese, E.A., Parshionikar, S.U. et Grimm, A.C. (2015). EPA method 1615. measurement of enterovirus and norovirus occurrence in water by culture and RT-qPCR. I. collection of virus samples. J. Vis. Exp., (97). doi(97) : 10.3791/52067.
Fout, G.S., Martinson, B.C., Moyer, M.W.N. et Dahling, D.R. (2003). A multiplex reverse transcription-PCR method for detection of human enteric viruses in groundwater. Appl. Environ. Microbiol., 69(6) : 3158–3164.
Francy, D.S., Bushon, R.N., Stopar, J., Luzano, E.J. et Fout, G.S. (2004). Environmental factors and chemical and microbiological water-quality constituents related to the presence of enteric viruses in ground water from small public water supplies in Southeastern Michigan. U.S.G.S., Scientific investigations report 2004–5219.
Francy, D.S., Stelzer, E.A., Brady, A.M., Huitger, C., Bushon, R.N., Ip, H.S., Ware, M.W., Villegas, E.N., Gallardo, V. et Lindquist, H.D. (2013). Comparison of filters for concentrating microbial indicators and pathogens in lake water samples. Appl. Environ. Microbiol., 79(4) : 1342–1352.
FSA (2000). A report of the study of infectious intestinal disease in England. Food Standards Agency, HMSO, Londres, Royaume-Uni.
Fuhrman, J.A., Liang, X. et Noble, R.T. (2005). Rapid detection of enteroviruses in small volumes of natural waters by real-time quantitative reverse transcriptase PCR. Appl. Environ. Microbiol., 71(8) : 4523–4530.
Gabbay, Y.B., Borges, A.A., Oliveira, D.S., Linhares, A.C., Mascarenhas, J.D.P., Barardi, C.R.M., Simões, C.M.O., Wang, Y., Glass, R.I. et Jiang, B. (2008). Evidence for zoonotic transmission of group C rotaviruses among children in Belém, Brazil. J. Med. Virol., 80(9) : 1666–1674.
Gale, P. (1996). Developments in microbiological risk assessment models for drinking water-a short review. J. Appl. Bacteriol., 81(4) : 403–410.
Gallimore, C.I., Iturriza-Gomara, M., Lewis, D., Cubitt, D., Cotterill, H. et Gray, J.J. (2006). Characterization of sapoviruses collected in the united kingdom from 1989 to 2004. J. Med. Virol., 78(5) : 673–682.
Gelting, R., Sarisky, J., Selman, C., Otto, C., Higgins, C., Bohan, P.O., Buchanan, S.B. et Meehan, P.J. (2005). Use of a systems-based approach to an environmental health assessment for a waterborne disease outbreak investigation at a snowmobile lodge in Wyoming. Int. J. Hyg. Environ. Health, 208(1–2) : 67–73.
Gerba, C.P. (1984). Applied and theoretical aspects of virus adsorption to surfaces. Adv. Appl. Microbiol., 30 : 133–168.
Gerba, C.P. et Bitton, G. (1984). Microbial pollutants: Their survival and transport pattern to groundwater. Dans : Groundwater pollution microbiology. Bitton, G. et Gerba, C.P. (éditeurs). John Wiley & Sons, New York, New York, p. 65–88.
Gerba, C.P., Goyal, S.M., Cech, I. et Bogdan, G.F. (1981). Quantitative assessment of the adsorptive behavior of viruses to soils. Environ. Sci. Technol., 15(8) : 940–944.
Gerba, C.P., Goyal, S.M., LaBelle, R.L. et Bodgan, G.F. (1979). Failure of indicator bacteria to reflect the occurrence of enteroviruses in marine waters. Am. J. Public Health, 69(11) : 1116–1119.
Gerba, C.P., Riley, K.R., Nwachuku, N., Ryu, H. et Abbaszadegan, M. (2003). Removal of encephalitozoon intestinalis, calicivirus, and coliphages by conventional drinking water treatment. J. Environ. Sci. Health. A. Tox. Hazard. Subst. Environ. Eng., 38(7) : 1259–1268.
Gerrity, D., Ryu, H., Crittenden, J. et Abbaszadegan, M. (2008). UV inactivation of adenovirus type 4 measured by integrated cell culture qPCR. J. Environ. Sci. Health. A. Tox. Hazard. Subst. Environ. Eng., 43(14) : 1628–1638.
Giammanco, G.M., Di Bartolo, I., Purpari, G., Costantino, C., Rotolo, V., Spoto, V., Geraci, G., Bosco, G., Petralia, A., Guercio, A., Macaluso, G., Calamusa, G., De Grazia, S., Ruggeri, F.M., Vitale, F., Maida, C.M. et Mammina, C. (2014). Investigation and control of a norovirus outbreak of probable waterborne transmission through a municipal groundwater system. J. Water Health, 12(3) : 452–464.
Gibbons, C.L., Mangen, M.-.J., Plass, D., Havelaar, A.H., Brooke, R.J., Kramarz, P., Peterson, K.L., Stuurman, A.L., Cassini, A., Fèvre, E.M. and Kretzschmar, M.E. (2014). Measuring underreporting and under-ascertainment in infectious disease datasets: A comparison of methods. BMC Public Health, 14(1).
Gibson, K.E. et Schwab, K.J. (2011). Detection of bacterial indicators and human and bovine enteric viruses in surface water and groundwater sources potentially impacted by animal and human wastes in lower Yakima Valley, Washington. Appl. Environ. Microbiol., 77(1) : 355–362.
Giles, J.P., Liebhaber, H., Krugman, S. et Lattimer, C. (1964). Early viremia and viruria in infectious hepatitis. Virology, 24 : 107–108.
Glass, R.I., Parashar, U., Patel, M., Tate, J., Jiang, B. and Gentsch, J. (2012). The control of rotavirus gastroenteritis in the united states. Trans. Am. Clin. Climatol. Assoc., 123: 36-52; discussion 53.
Gleeson, T., Novakowski, K. et Kurt Kyser, T. (2009). Extremely rapid and localized recharge to a fractured rock aquifer. J. Hydrol., 376(3–4) : 496–509.
Gofti-Laroche, L., Gratacap-Cavallier, B., Demanse, D., Genoulaz, O., Seigneurin, J.M. et Zmirou, D. (2003). Are waterborne astrovirus implicated in acute digestive morbidity (E.M.I.R.A. study)? J. Clin. Virol., 27(1) : 74–82.
Gordon, C. et Toze, S. (2003). Influence of groundwater characteristics on the survival of enteric viruses. J. Appl. Microbiol., 95(3) : 536–544.
Gotkowitz, M.B., Bradbury, K.R., Borchardt, M.A., Zhu, et Spencer, S.K. (2016). Effects of climate and sewer condition on virus transport to groundwater. Environ. Sci. Technol., 50(16): 8497–8504.
Gouvernement du Canada (2006). La gestion de l'eau douce au Canada. IV. L'eau souterraine. Service d'information et de recherche parlementaires, Division des sciences et des technologies. Bibliothèque du Parlement.
Goyal, S.M. et Gerba, C.P. (1979). Comparative adsorption of human enteroviruses, simian rotavirus, and selected bacteriophages to soils. Appl. Environ. Microbiol., 38(2) : 241–247.
Grabow, W.O.K. (2001). Bacteriophages: update on application as models for viruses in water. Water SA, 27(2) : 251–268.
Gray, J.J., Green, J., Cunliffe, C., Gallimore, C., Lee, J.V., Neal, K. et Brown, D.W.G. (1997). Mixed genogroup SRSV infections among a party of canoeists exposed to contaminated recreational water. J. Med. Virol., 52(4) : 425–429.
Gray, J.J., Wreghitt, T.G., Cubitt, W.D. et Elliot, P.R. (1987). An outbreak of gastroenteritis in a home for the elderly associated with astrovirus type 1 and human calicivirus. J. Med. Virol., 23(4) : 377–381.
Greenberg, H.B., Wyatt, R.G. et Kapikian, A.Z. (1979). Norwalk virus in vomitus. Lancet, 1(8106) : 55.
Greening, G.E., Hewitt, J. et Lewis, G.D. (2002). Evaluation of integrated cell culture-PCR (C-PCR) for virological analysis of environmental samples. J. Appl. Microbiol., 93(5) : 745–750.
Griffin, D.W., Donaldson, K.A., Paul, J.H. et Rose, J.B. (2003). Pathogenic human viruses in coastal waters. Clin. Microbiol. Rev., 16(1) : 129–143.
Griffin, D.W., Gibson, C.J., Lipp, E.K., Riley, K., Paul, J.H. et Rose, J.B. (1999). Detection of viral pathogens by reverse transcriptase PCR and of microbial indicators by standard methods in the canals of the Florida Keys. Appl. Environ. Microbiol., 65(9) : 4118–4125.
Griffin, J.S., Plummer, J.D. et Long, S.C. (2008). Torque teno virus: An improved indicator for viral pathogens in drinking waters. Virol. J., 5 : 112.
Gullick, R.W., LeChevallier, M.W., Svindland, R.C. et Friedman, M.J. (2004). Occurrence of transient low and negative pressures in distribution systems. J. Am. Water Works Assoc., 96(11): 52–66.
Gunnarsdottir, M.J., Gardarsson, S.M. et Andradottir, H.O. (2013). Microbial contamination in groundwater supply in a cold climate and coarse soil: Case study of norovirus outbreak at lake Mývatn, Iceland. Hydrol. Res., 44(6) : 1114–1128.
Guo, H., Chu, X. et Hu, J. (2010b). Effect of host cells on low- and medium-pressure UV inactivation of adenoviruses. Appl. Environ. Microbiol., 76(21) : 7068–7075.
Guo, H., Wyart, Y., Perot, J., Nauleau, F. et Moulin, P. (2010a). Low-pressure membrane integrity tests for drinking water treatment: A review. Water Res., 44(1) : 41–57.
Guthmann, J.P., Klovstad, H., Boccia, D., Hamid, N., Pinoges, L., Nizou, J.Y., Tatay, M., Diaz, F., Moren, A., Grais, R.F., Ciglenecki, I., Nicand, E. et Guerin, P.J. (2006). A large outbreak of hepatitis E among a displaced population in Darfur, Sudan, 2004: The role of water treatment methods. Clin. Infect. Dis., 42(12) : 1685–1691.
Haas, C.N., Rose, J.B. et Gerba, C.P. (1999). Quantitative microbial risk assessment. John Wiley, New York, New York.
Haas, C.N., Rose, J.B. et Gerba, C.P. (2014). Quantitative microbial risk assessment. Second edition. John Wiley, New York, New York.
Häfliger, D., Hubner, P. et Luthy, J. (2000). Outbreak of viral gastroenteritis due to sewage-contaminated drinking water. Int. J. Food Microbiol., 54(1–2) : 123–126.
Haines, K.E. et O'Brien, R.T. (1979). The survival of enteric viruses in septic tanks and septic tank drain fields. New Mexico Water Resources Research Institute, New Mexico State University, Nouveau-Mexique.
Halbur, P.G., Kasorndorkbua, C., Gilbert, C., Guenette, D., Potters, M.B., Purcell, R.H., Emerson, S.U., Toth, T.E. et Meng, X.J. (2001). Comparative pathogenesis of infection of pigs with hepatitis E viruses recovered from a pig and a human. J. Clin. Microbiol., 39(3) : 918–923.
Hamano, M., Kuzuya, M., Fujii, R., Ogura, H., Mori, T., Nakayama, T., Yuen, E., Katayama, K., Mitsunobu, Y. et Inoue, K. (1999). Outbreak of acute gastroenteritis caused by human group C rotavirus in a primary school. Jpn. J. Infect. Dis., 52(4) : 170–171.
Hansman, G.S., Sano, D., Ueki, Y., Imai, T., Oka, T., Katayama, K., Takeda, N. et Omura, T. (2007). Sapovirus in water, Japan. Emerg. Infect. Dis., 13(1) : 133–135.
Haramoto, E., Katayama, H. et Ohgaki, S. (2004). Detection of noroviruses in tap water in Japan by means of a new method for concentrating enteric viruses in large volumes of freshwater. Appl. Environ. Microbiol., 70(4) : 2154–2160.
Haramoto, E., Katayama, H., Oguma, K. et Ohgaki, S. (2005a). Application of cation-coated filter method to detection of noroviruses, enteroviruses, adenoviruses, and torque teno viruses in the Tamagawa River in Japan. Appl. Environ. Microbiol., 71(5) : 2403–2411.
Haramoto, E., Katayama, H., Oguma, K., Yamashita, H., Nakajima, E. et Ohgaki, S. (2005b). One-year monthly monitoring of torque teno virus (TTV) in wastewater treatment plants in Japan. Water Res., 39(10) : 2008–2013.
Haramoto, E., Kitajima, M., Katayama, H. et Ohgaki, S. (2010). Real-time PCR detection of adenoviruses, polyomaviruses, and torque teno viruses in river water in Japan. Water Res., 44(6) : 1747–1752.
Harman, J., Robertson, W.D., Cherry, J.A. et Zanini, L. (1996). Impacts on a sand aquifer from an old septic system: Nitrate and phosphate. Ground Water, 34(6) : 1105–1114.
Harrington, G.W., Chen, H., Harris, A.J., Xagoraraki, I., Battigelli, D.A. et Standridge, J.H. (2001). Removal of emerging waterborne pathogens. AWWA Research Foundation and American Water Works Association, Denver, Colorado.
Harrington, G.W., Xagoraraki, I., Assavasilavasukul, P. et Standridge, J.H. (2003). Effect of filtration conditions on removal of emerging waterborne pathogens. J. Am. Water Works Assoc., 95(12): 95–104.
Hartmann, N.M., Dartscht, M., Szewzyk, R. et Selinka, H-C. (2013). Monitoring of adenovirus serotypes in environmental samples by combined PCR and melting point analyses. Virol. J., 10 : 190.
Harvey, R.W., Metge, D.W., LeBlanc, D.R., Underwood, J., Aiken, G.R., Butler, K., McCobb, T.D. et Jasperse, J. (2015). Importance of the colmation layer in the transport and removal of cyanobacteria, viruses, and dissolved organic carbon during natural lake-bank filtration. J. Environ. Qual., 44(5) : 1413–1423.
Hata, A., Katayama, H., Kojima, K., Sano, S., Kasuga, I., Kitajima, M. et Furumai, H. (2014). Effects of rainfall events on the occurrence and detection efficiency of viruses in river water impacted by combined sewer overflows. Sci. Total Environ., 468–469 : 757–763.
Hata, A., Kitajima, M. et Katayama, H. (2013). Occurrence and reduction of human viruses, F-specific RNA coliphage genogroups and microbial indicators at a full-scale wastewater treatment plant in Japan. J. Appl. Microbiol., 114(2) : 545–554.
Hauri, A.M., Schimmelpfennig, M., Walter-Domes, M., Letz, A., Diedrich, S., Lopez-Pila, J. et Schreier, E. (2005). An outbreak of viral meningitis associated with a public swimming pond. Epidemiol. Infect., 133(2) : 291–298.
Havelaar, A. et Melse, J.M. (2003). Quantifying public health risk in the WHO guidelines for drinking-water quality: A burden of disease approach. Rijkinstituut voor Volskgezondheid en Milieu, RIVM Report 734301022, Bilthoven, Pays-Bas.
Havelaar, A.H. (1987). Bacteriophages as model organisms in water treatment. Microbiol. Sci., 4(12) : 362–364.
Havelaar, A.H., Van Olphen, M. et Drost, Y.C. (1993). F-specific RNA bacteriophages are adequate model organisms for enteric viruses in fresh water. Appl. Environ. Microbiol., 59(9) : 2956–2962.
Havelaar, A.H., Van Olphen, M. et Schijven, J.F. (1995). Removal and inactivation of viruses by drinking water treatment processes under full scale conditions. Water Sci. Technol., 31(5–6) : 55–62.
He, X.Q., Cheng, L., Zhang, D.Y., Xie, X.M., Wang, D.H. et Wang, Z. (2011). One-year monthly survey of rotavirus, astrovirus and norovirus in three sewage treatment plants (STPs) in Beijing, China and associated health risk assessment. Water Sci. Technol., 64(6) : 1202–1210.
Hellard, M.E., Sinclair, M.I., Forbes, A.B. et Fairley, C.K. (2001). A randomized, blinded, controlled trial investigating the gastrointestinal health effects of drinking water quality. Environ. Health Perspect., 109(8): 773-778
Hellmer, M., Paxeus, N., Magnius, L., Enache, L., Arnholm, B., Johansson, A., Bergstrom, T. et Norder, H. (2014). Detection of pathogenic viruses in sewage provided early warnings of hepatitis A virus and norovirus outbreaks. Appl. Environ. Microbiol., 80(21) : 6771–6781.
Hendricks, D.W., Clunie, W.F., Sturbaum, G.D., Klein, D.A., Champlin, T.L., Kugrens, P., Hirsch, J., McCourt, B., Nordby, G.R., Sobsey, M.D., Hunt, D.J. et Allen, M.J. (2005). Filtration removals of microorganisms and particles. J. Environ. Eng., 131(12) : 1621–1632.
Herrmann, J.E., Kostenbader, K.D.,Jr et CLIVER, D.O. (1974). Persistence of enteroviruses in lake water. Appl. Microbiol., 28(5) : 895–896.
Hewitt, J., Bell, D., Simmons, G.C., Rivera-Aban, M., Wolf, S. et Greening, G.E. (2007). Gastroenteritis outbreak caused by waterborne norovirus at a New Zealand ski resort. Appl. Environ. Microbiol., 73(24) : 7853–7857.
Hijnen, W.A. et Medema, G.J. (2010). Elimination of micro-organisms by drinking water treatment processes—A review. KWR Watercycle Research Institute. IWA Publishing, Londres, Royaume-Uni.
Hijnen, W.A., Beerendonk, E.F. et Medema, G.J. (2006). Inactivation credit of UV radiation for viruses, bacteria and protozoan (oo)cysts in water: A review. Water Res., 40(1) : 3–22.
Hijnen, W.A., Schijven, J.F., Bonne, P., Visser, A. et Medema, G.J. (2004a). Elimination of viruses, bacteria and protozoan oocysts by slow sand filtration. Water Sci. Technol., 50(1) : 147–154.
Hijnen, W.A., Suylen, G.M., Bahlman, J.A., Brouwer-Hanzens, A. et Medema, G.J. (2010). GAC adsorption filters as barriers for viruses, bacteria and protozoan (oo)cysts in water treatment. Water Res., 44(4) : 1224–1234.
Hijnen, W.A.M. (2011). Chapter 12: Quantitative methods to assess capacity of water treatment processes to eliminate micro-organisms. Dans : Quantitative methods to assess capacity of water treatment to eliminate micro-organisms. IWA Publishing, Londres, Royaume-Uni, p. 162–166.
Hijnen, W.A.M., Van Der Veer, A.J., Beerendonk, E.F. et Medema, G.J. (2004b). Increased resistance of environmental anaerobic spores to inactivation by uv. Water Science and Technology: Water Supply, 4(2) : 55–61.
Hill, V.R., Kahler, A.M., Jothikumar, N., Johnson, T.B., Hahn, D. et Cromeans, T.L. (2007). Multistate evaluation of an ultrafiltration-based procedure for simultaneous recovery of enteric microbes in 100-liter tap water samples. Appl. Environ. Microbiol., 73(13) : 4218–4225.
Hill, V.R., Polaczyk, A.L., Hahn, D., Narayanan, J., Cromeans, T.L., Roberts, J.M. et Amburgey, J.E. (2005). Development of a rapid method for simultaneous recovery of diverse microbes in drinking water by ultrafiltration with sodium polyphosphate and surfactants. Appl. Environ. Microbiol., 71(11) : 6878–6884.
Hilt, E.E., McKinley, K., Pearce, M.M., Rosenfeld, A.B., Zilliox, M.J., Mueller, E.R., Brubaker, L., Gai, X., Wolfe , A.J. and Schreckenberger, P.C. (2014). Urine is not sterile: Use of enhanced urine culture techniques to detect resident bacterial flora in the adult female bladder. J. Clin. Microbiol., 52(3): 871-876.
Hoa Tran, T.N., Trainor, E., Nakagomi, T., Cunliffe, N.A. et Nakagomi, O. (2013). Molecular epidemiology of noroviruses associated with acute sporadic gastroenteritis in children: global distribution of genogroups, genotypes and GII.4 variants. J. Clin. Virol., 56(3) : 185–193.
Hoebe, C.J., Vennema, H., de Roda Husman, A.M. et van Duynhoven, Y.T. (2004). Norovirus outbreak among primary schoolchildren who had played in a recreational water fountain. J. Infect. Dis., 189(4) : 699–705.
Hoff, J.C. (1986). Inactivation of microbial agents by chemical disinfectants. U.S. Environmental Protection Agency des É.‑U., EPA/600/S2-86/067, Cincinnati, Ohio.
Hofmann, R. (2015). UV dose requirements for enteric viruses. Report for Water, Air and Climate Change Bureau, Healthy Environments and Consumer Safety Branch, Health Canada, Ottawa, Ontario.
Hollinger, F.B. et Emerson, S.U. (2007). Hepatitis A virus. Knipe, D.M. and Howley, P.M. (éditeurs). 5th edition. Lippincott Williams and Wilkins, Philadelphie, Pennsylvanie.
Hörman, A., Rimhanen-Finne, R., Maunula, L., Von Bonsdorff, C., Torvela, N., Heikinheimo, A. et Hänninen, M. (2004). Campylobacter spp., giardia spp., noroviruses, and indicator organisms in surface waters in southwestern Findland, 2000–2001. Appl. Environ. Microbiol., 71 : 87–95.
Hot, D., Legeay, O., Jacques, J., Gantzer, C., Caudrelier, Y., Guyard, K., Lange, M. et Andreoletti, L. (2003). Detection of somatic phages, infectious enteroviruses and enterovirus genomes as indicators of human enteric viral pollution in surface water. Water Res., 37(19) : 4703–4710.
Hsu, B.M., Chen, C.H., Kung, C.M., Wan, M.T. et Shen, S.M. (2007). Evaluation of enterovirus recovery in surface water by different adsorption and elution procedures. Chemosphere, 66(5) : 964–969.
Huck, P. et Sozański, M. (2011). 3.16 – chemical basis for water technology. Dans : Treatise on water science. Wilderer, P. (éditeurs). Oxford : Elsevier, p. 429–469.
Huck, P.M. et Coffey, B.M. (2004). The importance of robustness in drinking-water systems. J. Toxicol. Environ. Health A, 67(20–22) : 1581–1590.
Hunt et Johnson (2017). Pathogen transport in groundwater systems: contrasts with traditional solute transport. Hydrogeol J., 25:921-930.
Hunt, R.J., Borchardt, M.A. et Bradbury, K.R. (2014). Viruses as groundwater tracers: Using ecohydrology to characterize short travel times in aquifers. Ground Water, 52(2) : 187–193.
Hunt, R.J., Borchardt, M.A., Richards, K.D. et Spencer, S.K. (2010). Assessment of sewer source contamination of drinking water wells using tracers and human enteric viruses. Environ. Sci. Technol., 44(20) : 7956–7963.
Hunter, P.R., Chalmers, R.M., Hughes, S. et Syed, Q. (2005). Self-reported diarrhea in a control group: A strong association with reporting of low-pressure events in tap water. Clin. Infect. Dis., 40(4) : e32–34.
Hurst, C.J., Knudsen, G.R., McInerney, M.J., Stetzenbach, L.D. et Walter, M.V. (2001). Manual of environmental microbiology. American Society for Microbiology Press, Washington, DC.
Hutson, A.M., Atmar, R.L., Marcus, D.M. et Estes, M.K. (2003). Norwalk virus-like particle hemagglutination by binding to H histoblood group antigens. J. Virol, 77 : 405–415.
Hynds, P., Misstear, B.D., Gill, L.W. et Murphy, H.M. (2014a). Groundwater source contamination mechanisms: physicochemical profile clustering, risk factor analysis and multivariate modelling. J. Contam. Hydrol., 159 : 47–56.
Hynds, P.D., Misstear, B.D. et Gill, L.W. (2012). Development of a microbial contamination susceptibility model for private domestic groundwater sources. Water Resour. Res., 48(12).
Hynds, P.D., Thomas, M.K. et Pintar, K.D. (2014b). Contamination of groundwater systems in the US and Canada by enteric pathogens, 1990-2013: a review and pooled-analysis. PLoS One, 9(5) : e93301.
INSPQ (1994). Bilan des maladies d'origine hydrique signalées dans les directions régionales de la santé publique du Québec en 1991 et 1992. Institut national de santé publique du Québec, Québec (Québec).
INSPQ (1998). Bilan des éclosions de maladies d'origine hydrique signalées dans les directions régionales de la santé publique du Québec en 1993, 1994 et 1995. Institut national de santé publique du Québec, Québec, Québec.
INSPQ (2001). Bilan des éclosions de maladies d'origine hydrique signalées dans les directions régionales de la santé publique du Québec en 1998 et en 1999. Institut national de santé publique du Québec, Québec, Québec.
Irving, L.G. et Smith, F.A. (1981). One-year survey of enteroviruses, adenoviruses, and reoviruses isolated from effluent at an activated-sludge purification plant. Appl. Environ. Microbiol., 41(1) : 51–59.
Jacangelo, J.G. (1991). The development of membrane technology. International report (IR3). Water Supply: Rev. J. Int. Water Supply Assoc., 9(3/4).
Jacangelo, J.G., Adham, S. et Laine, J-M. (1997). Membrane filtration for microbial removal. American Water Works Association Research Foundation and American Water Works Association, Denver, Colorado.
Jacangelo, J.G., Adham, S.S. et Laine, J-M. (1995). Mechanism of Cryptosporidium, Giardia and MS2 virus removal by MF and UF. J. Am. Water Works Assoc, 87(9) : 107–121.
Jacangelo, J.G., Laine, J., Carns, K.E., Cummings, E.W. et Mallevialle, J. (1991). Low-pressure membrane filtration for removing giardia and microbial indicators. J. Am. Water Works Assoc., 83(9) : 97–106.
Jacangelo, J.G., Patania Brown, N.L., Madec, A., Schwab, K., Huffman, D., Amy, G. et Prescott, A. (2006). Micro- and ultrafiltration performance specifications based on microbial removal. AWWA Research Foundation and American Water Works Association, Denver, Colorado.
Jackson, R.E et Heagle, D.J. (2016). Sampling domestic/farm wells for baseline groundwater quality and fugitive gas. Hydrogeol J., 24:269-272.
Jacobsen, K.H. et Wiersma, S.T. (2010). Hepatitis A virus seroprevalence by age and world region, 1990 and 2005. Vaccine, 28(41) : 6653–6657.
Jiang, B., Dennehy, P.H., Spangenberger, S., Gentsch, J.R. et Glass, R.I. (1995). First detection of group C rotavirus in faecal specimens of children with diarrhea in the united states. J. Infect. Dis., 172(1) : 45–50.
Jiang, H., Holtz, L.R., Bauer, I., Franz, C.J., Zhao, G., Bodhidatta, L., Shrestha, S.K., Kang, G. et Wang, D. (2013). Comparison of novel MLB-clade, VA-clade and classic human astroviruses highlights constrained evolution of the classic human astrovirus nonstructural genes. Virology, 436(1) : 8–14.
Jiang, S., Noble, R. et Chu, W. (2001). Human adenoviruses and coliphages in urban runoff-impacted coastal waters of southern California. Appl. Environ. Microbiol., 67(1) : 179–184.
Jiang, S.C. (2006). Human adenoviruses in water: Occurrence and health implications: A critical review. Environ. Sci. Technol., 40(23) : 7132–7140.
Johansson, P.J.H., Bergentoft, K., Larsson, P.A., Magnusson, G., Widell, A., Thorhagen, M. et Hedlund, K.-. (2005). A nosocomial sapovirus-associated outbreak of gastroenteritis in adults. Scand. J. Infect. Dis., 37(3) : 200–204.
John, D.E. et Rose, J.B. (2005). Review of factors affecting microbial survival in groundwater. Environ. Sci. Technol., 39(19) : 7345–7356.
Johnson, T.B., McKay, L.D., Layton, A.C., Jones, S.W., Johnson, G.C., Cashdollar, J.L., Dahling, D.R., Villegas, L.F., Fout, G.S., Williams, D.E. et Sayler, G. (2011). Viruses and bacteria in karst and fractured rock aquifers in East Tennessee, USA. Ground Water, 49(1) : 98–110.
Johnson, W.P., Ma, H. et Pazmino, E. (2011). Straining credibility: A general comment regarding common arguments used to infer straining as the mechanism of colloid retention in porous media. Environ. Sci. Technol., 45(9) : 3831–3832.
Joshi, M.S., Bhalla, S., Kalrao, V.R., Dhongade, R.K. et Chitambar, S.D. (2014). Exploring the concurrent presence of hepatitis A virus genome in serum, stool, saliva, and urine samples of hepatitis A patients. Diagn. Microbiol. Infect. Dis., 78(4) : 379–382.
Jothikumar, N., Paulmurugan, R., Padmanabhan, P., Sundari, R.B., Kamatchiammal, S. et Rao, K.S. (2000). Duplex RT-PCR for simultaneous detection of hepatitis A and hepatitis E virus isolated from drinking water samples. J. Environ. Monit., 2(6) : 587–590.
Kahler, A.M., Johnson, T.B., Hahn, D., Narayanan, J., Derado, G. et Hill, V.R. (2015). Evaluation of an ultrafiltration-based procedure for simultaneous recovery of diverse microbes in source waters. Water. (Basel), 7(3) : 1202–1216.
Kang, G., Kelkar, S.D., Chitambar, S.D., Ray, P. et Naik, T. (2005). Epidemiological profile of rotaviral infection in India: Challenges for the 21st century. J. Infect. Dis., 192 Suppl 1 : S120-6.
Kapikian, A.Z. et Chanock, R.M. (1996). Rotaviruses. Fields, B.N., Knipe, D.M. and Howley, P.M. (éditeurs). 3rd. Lippincott-Raven Publishers, Philadelphie, Pennsylvanie.
Kapoor, A., Li, L., Victoria, J., Oderinde, B., Mason, C., Pandey, P., Zaidi, S.Z. et Delwart, E. (2009). Multiple novel astrovirus species in human stool. J. Gen. Virol., 90(Pt 12) : 2965–2972.
Karim, M.R., Abbaszadegan, M. et Lechevallier, M. (2003). Potential for pathogen intrusion during pressure transients. J. Am. Water Works Assoc., 95(5) : 134–146.
Karim, M.R., Rhodes, E.R., Brinkman, N., Wymer, L. et Fout, G.S. (2009). New electropositive filter for concentrating enteroviruses and noroviruses from large volumes of water. Appl. Environ. Microbiol., 75(8) : 2393–2399.
Karst, S.M., Wobus, C.E., Lay, M., Davidson, J. et Virgin IV, H.W. (2003). STAT1-dependent innate immunity to a norwalk-like virus. Science, 299(5612) : 1575–1578.
Kasorndorkbua, C., Opriessnig, T., Huang, F.F., Guenette, D.K., Thomas, P.J., Meng, X.J. et Halbur, P.G. (2005). Infectious swine hepatitis E virus is present in pig manure storage facilities on United States farms, but evidence of water contamination is lacking. Appl. Environ. Microbiol., 71(12) : 7831–7837.
Katayama, H., Shimasaki, A. et Ohgaki, S. (2002). Development of a virus concentration method and its application to detection of enterovirus and norwalk virus from coastal seawater. Appl. Environ. Microbiol., 68(3) : 1033–1039.
Katz, B.G., Eberts, S.M., et Kauffman, L.J. (2011). Using Cl/Br ratios and other indicators to assess potential impacts on groundwater quality from septic systems: a review and examples from principal aquifers in the United States. J. Hydrol., 397: 151-166.
Kauppinen, A., Ikonen, J., Pursiainen, A., Pitkanen, T. et Miettinen, I.T. (2012). Decontamination of a drinking water pipeline system contaminated with adenovirus and escherichia coli utilizing peracetic acid and chlorine. J. Water. Health., 10(3) : 406–418.
Kawamura, S. (2000). Integrated design and operation of water treatment facilities. 2nd edition. John Wiley & Sons, New York, New York.
Keswick, B.H. et Gerba, C.P. (1980). Viruses in groundwater. Environ. Sci. Technol., 14(11) : 1290–1297.
Keswick, B.H., Secor, S.L., Gerba, C.P. et Cech, I. (1982). Survival of enteric viruses and indicator bacteria in groundwater. J. Environ. Sci. Health Part A Environ. Sci. Eng., 17(6) : 903–912.
Kim, S.H., Cheon, D.S., Kim, J.H., Lee, D.H., Jheong, W.H., Heo, Y.J., Chung, H.M., Jee, Y. et Lee, J.S. (2005). Outbreaks of gastroenteritis that occurred during school excursions in Korea were associated with several waterborne strains of norovirus. J. Clin. Microbiol., 43(9) : 4836–4839.
Kirmeyer, G.J., Friedman, M., Martel, K., Howie, D., LeChevallier, M., Abbaszadegan, M., Karim, M., Funk, J. et Harbour, J. (2001). Pathogen intrusion into the distribution system. AWWA Research Foundation and American Water Works Association, Denver, Colorado.
Kirmeyer, G.J., Thomure, T.M., Rahman, R., Marie, J.L., LeChevallier, M.W., Yang, J., Hughes, D.M. et Schneider, O. (2014). Effective microbial control strategies for main breaks and depressurization. Water Research Foundation, Denver, Colorado.
Kitajima, M., Haramoto, E., Phanuwan, C., Katayama, H. et Furumai, H. (2012). Molecular detection and genotyping of human noroviruses in influent and effluent water at a wastewater treatment plant in Japan. J. Appl. Microbiol., 112(3) : 605–613.
Kitajima, M., Iker, B.C., Pepper, I.L. et Gerba, C.P. (2014). Relative abundance and treatment reduction of viruses during wastewater treatment processes–identification of potential viral indicators. Sci. Total Environ., 488–489 : 290–296.
Kitajima, M., Oka, T., Haramoto, E., Katayama, H., Takeda, N., Katayama, K. et Ohgaki, S. (2010a). Detection and genetic analysis of human sapoviruses in river water in Japan. Appl. Environ. Microbiol., 76(8) : 2461–2467.
Kitajima, M., Tohya, Y., Matsubara, K., Haramoto, E., Utagawa, E. et Katayama, H. (2010b). Chlorine inactivation of human norovirus, murine norovirus and poliovirus in drinking water. Lett. Appl. Microbiol., 51(1) : 119–121.
Kitis, M., Lozier, J.C., Kim, J-H., Mi, B. et Mariñas, B.J. (2003). Microbial removal and integrity monitoring of RO and NF membranes. J. Am. Water Works Assoc., 95(12) : 105–119.
Kiulia, N.M., Hofstra, N., Vermeulen, L.C., Obara, M.A., Medema, G. et Rose, J.B. (2015). Global occurrence and emission of rotaviruses to surface waters. Pathogens, 4(2) : 229–255.
Ko, G., Cromeans, T.L. et Sobsey, M.D. (2003). Detection of infectious adenovirus in cell culture by mRNA reverse transcription-PCR. Appl. Environ. Microbiol., 69(12) : 7377–7384.
Kozuskanich, J.C., Novakowski, K.S., Anderson, B.C., Crowe, A.S. et Balakrishnan, V.K. (2014). Anthropogenic impacts on a bedrock aquifer at the village scale. Groundwater, 52(3) : 474–486.
Kramer, M.H., Herwaldt, B.L., Craun, G.F., Calderon, R.L. et Juranek, D.D. (1996). Surveillance for waterborne-disease outbreaks – United States, 1993–1994. MMWR CDC Surveill Summ, 45(1) : 1–33.
Krasner, S.W., Weinberg, H.S., Richardson, S.D., Pastor, S.J., Chinn, R., Sclimenti, M.J., Onstad, G.D. et Thruston Jr., A.D. (2006). Occurrence of a new generation of disinfection byproducts. Environ. Sci. Technol., 40(23) : 7175–7185.
Kruithof, J.C., Hiemstra, P., Kamp, P., van der Hoek, J. et Schippers, J.C. (1997). Integrated multi-objective membrane systems for control of microbials and DBP-precursors. Dans : Proceedings of the Membrane Technology Conference, American Water Works Association, New Orleans, Louisiane.
Kruithof, J.C., Kamp, P.C., Folmer, H.C., Nederlof, M.M. et van Hoof, S.C.J.M. (2001). Development of a membrane integrity monitoring strategy for the UF/RO Heemskerk drinking water treatment plant. Water Sci. Technol.: Water Supply, 1(5–6) : 261–271.
Kuehn, W. et Mueller, U. (2000). Riverbank filtration: an overview. J. Am. Water Works Assoc., 92(12) : 60–69.
Kukkula, M., Arstila, P., Klossner, M.L., Maunula, L., Bonsdorff, C-H.V. et Jaatinen, P. (1997). Waterborne outbreak of viral gastroenteritis. Scand. J. Infect. Dis., 29(4) : 415–418.
Kukkula, M., Maunula, L., Silvennoinen, E. et Von Bonsdorff, C-H. (1999). Outbreak of viral gastroenteritis due to drinking water contaminated by norwalk-like viruses. J. Infect. Dis., 180(6) : 1771–1776.
Kumar, M., Adham. S., et DeCarolis, J. (2007). Reverse osmosis integrity monitoring. Desalination, 214 : 138–149.
Kundert, K. (2014). Alberta flood 2013 – city of Calgary water treatment system resiliency. Conférence nationale canadienne sur l'eau potable, Ottawa (Ontario).
Laine, J., Huovinen, E., Virtanen, M.J., Snellman, M., Lumio, J., Ruutu, P., Kujansuu, E., Vuento, R., Pitkänen, T., Miettinen, I., Herrala, J., Lepistö, O., Antonen, J., Helenius, J., Hänninen, M.-., Maunula, L., Mustonen, J., Kuusi, M., Collin, P., Korpela, M., Kuusela, A.-., Mustajoki, S., Oksa, H., Räsänen, S., Uotila, T. et Katto, T. (2011). An extensive gastroenteritis outbreak after drinking-water contamination by sewage effluent, Finland. Epidemiol. Infect., 139(7) : 1105–1113.
Laitinen, O.H., Honkanen, H., Pakkanen, O., Oikarinen, S., Hankaniemi, M.M., Huhtala, H., Ruokoranta, T., Lecouturier, V., André, P., Harju, R., Virtanen, S.M., Lehtonen, J., Almond, J.W., Simell, T., Simell, O., Ilonen, J., Veijola, R., Knip, M. et Hyöty, H. (2014). Coxsackievirus B1 is associated with induction of b-cell autoimmunity that portends type 1 diabetes. Diabetes, 63 : 446–455.
Lambertini, E., Borchardt, M.A., Kieke, B.A., Spencer, S.K. et Loge, F.J. (2012). Risk of viral acute gastrointestinal illness from nondisinfected drinking water distribution systems. Environ. Sci. Technol., 46(17) : 9299–9307.
Lambertini, E., Spencer, S.K., Bertz, P.D., Loge, F.J., Kieke, B.A. et Borchardt, M.A. (2008). Concentration of enteroviruses, adenoviruses, and noroviruses from drinking water by use of glass wool filters. Appl. Environ. Microbiol., 74(10) : 2990–2996.
Lambertini, E., Spencer, S.K., Kieke Jr., B.A., Loge, F.J. et Borchardt, M.A. (2011). Virus contamination from operation and maintenance events in small drinking water distribution systems. J. Water Health, 9(4) : 799–812.
Lapointe, B.E., Herren, L.W. et Bedford, B.J. (2012). Effects of hurricanes, land use, and water management on nutrient and microbial pollution: St. lucie estuary, southeast florida. J. Coast. Res., 28(6): 1345-1361.
Larsson, C., Andersson, Y., Allestam, G., Lindqvist, A., Nenonen, N. et Bergstedt, O. (2014). Epidemiology and estimated costs of a large waterborne outbreak of norovirus infection in Sweden. Epidemiol. Infect., 142(3) : 592–600.
LeChevallier, M.W. et Au, K.K. (2004). Water treatment and pathogen control: Process efficiency in achieving safe drinking water. IWA Publishing, Londres, Royaume-Uni.
LeChevallier, M.W., Evans, T.M. et Seidler, R.J. (1981). Effect of turbidity on chlorination efficiency and bacterial persistence in drinking water. Appl. Environ. Microbiol., 42: 159-167.
LeChevallier, M.W., Gullick, R.W., Karim, M.R., Friedman, M. et Funk, J.E. (2003). The potential for health risks from intrusion of contaminants into the distribution system from pressure transients. J. Water Health, 1(1) : 3–14.
LeChevallier, M.W., Xu, M., Yang, J., Teunis, P. et Fleming, K.K. (2011). Managing distribution system low transient pressures for water quality. Water Research Foundation, Denver, Colorado
Leclerc, H., Edberg, S., Pierzo, V. et Delattre, J.M. (2000). Bacteriophages as indicators of enteric viruses and public health risk in groundwaters. J. Appl. Microbiol., 88(1) : 5–21.
Lee, G., Lee, C., Park, C. et Jeong, S. (2008). Detection and molecular characterization of enteroviruses in Korean surface water by using integrated cell culture multiplex RT-PCR. Biomed. Environ. Sci., 21(5) : 425–431.
Lee, R.M., Lessler, J., Lee, R.A., Rudolph, K.E., Reich, N.G., Perl, T.M. et Cummings, D.A.T. (2013). Incubation periods of viral gastroenteritis: A systematic review. BMC Infect. Dis., 13(1).
Lee, S.H., Levy, D.A., Craun, G.F., Beach, M.J. et Calderon, R.L. (2002). Surveillance for waterborne-disease outbreaks – United States, 1999–2000. MMWR Surveill Summ, 51(8) : 1–47.
Lennon, G., Cashman, O., Lane, K., Cryan, B. et O'Shea, H. (2007). Prevalence and characterization of enteric adenoviruses in the south of Ireland. J. Med. Virol., 79(10) : 1518–1526.
Levine, W.C., Stephenson, W.T. et Craun, G.F. (1990). Waterborne disease outbreaks, 1986–1988. MMWR CDC Surveill Summ, 39(1) : 1–13.
Levison, J.K. et Novakowski, K.S. (2012). Rapid transport from the surface to wells in fractured rock: a unique infiltration tracer experiment. J. Contam. Hydrol., 131(1–4) : 29–38.
Li, D., Gu, A.Z., He, M., Shi, H. et Yang, W. (2009). UV inactivation and resistance of rotavirus evaluated by integrated cell culture and real-time RT-PCR assay. Water Res., 43(13) : 3261–3269.
Li, D., Gu, A.Z., Zeng, S., Yang, W., He, M. et Shi, H. (2011). Monitoring and evaluation of infectious rotaviruses in various wastewater effluents and receiving waters revealed correlation and seasonal pattern of occurrences. J. Appl. Microbiol., 110(5) : 1129–1137.
Li, D., He, M. et Jiang, S.C. (2010). Detection of infectious adenoviruses in environmental waters by fluorescence-activated cell sorting assay. Appl. Environ. Microbiol., 76(5) : 1442–1448.
Liang, J.L., Dziuban, E.J., Craun, G.F., Hill, V., Moore, M.R., Gelting, R.J., Calderon, R.L., Beach, M.J. et Roy, S.L. (2006). Surveillance for waterborne disease and outbreaks associated with drinking water and water not intended for drinking–United States, 2003–2004. MMWR Surveill Summ, 55(12) : 31–65.
Lieberman, R.J., Shadix, L.C., Newport, B.S., Frebis, C.P., Moyer, M.W.N., Safferman, R.S., Stetler, R.E., Lye, D., Fout, G.S., et Dahling, D. R. (2002). Microbial monitoring of vulnerable public groundwater supplies. American Water Works Association Research Foundation, Denver, Colorado.
Lim, M.Y., Kim, J.-. et Ko, G. (2010). Disinfection kinetics of murine norovirus using chlorine and chlorine dioxide. Water Res., 44(10) : 3243–3251.
Lin, J. et Ganesh, A. (2013). Water quality indicators: Bacteria, coliphages, enteric viruses. Int. J. Environ. Health Res., 23(6) : 484–506.
Linden, K.G., Shin, G.A., Lee, J.K., Scheible, K., Shen, C. et Posy, P. (2009). Demonstration 4-log adenovirus inactivation in a medium-pressure UV disinfection reactor. Journal of the American Water Works Association, 104(4) : 90–99.
Linden, K.G., Thurston, J., Schaefer, R. et Malley Jr., J.P. (2007). Enhanced UV inactivation of adenoviruses under polychromatic UV lamps. Appl. Environ. Microbiol., 73(23) : 7571–7574.
Linden, K.G., Wrights, H.B., Collins, J., Cotton, C. et Beck, S.E. (2015). Guidance for implementing action spectra correction with medium pressure UV disinfection. Water Research Foundation, Denver, Colorado.
Lindesmith, L., Moe, C., Marionneau, S., Ruvoen, N., Jiang, X., Lindblad, L., Stewart, P., Lependu, J. et Baric, R. (2003). Human susceptibility and resistance to norwalk virus infection. Nat. Med., 9(5) : 548–553.
Lindsey, B.D., Rasberry, J.S. et Zimmerman, T.M. (2002). Microbiological quality of water from noncommunity supply wells in carbonate and crystalline aquifers of pennsylvania. U.S.G.S, 01-4268, .
Liu, P., Hill, V.R., Hahn, D., Johnson, T.B., Pan, Y., Jothikumar, N. et Moe, C.L. (2012). Hollow-fiber ultrafiltration for simultaneous recovery of viruses, bacteria and parasites from reclaimed water. J. Microbiol. Methods, 88(1) : 155–161.
Locas, A., Barthe, C., Barbeau, B., Carrière, A. et Payment, P. (2007). Virus occurrence in municipal groundwater sources in Quebec, Canada. R. can. microbiol., 53(6) : 688–694.
Locas, A., Barthe, C., Margolin, A.B. et Payment, P. (2008). Groundwater microbiological quality in Canadian drinking water municipal wells. R. Can. microbiol., 54(6) : 472–478.
Lodder, W.J., Schijven, J.F., Rutjes, S.A., de Roda Husman, A.M. et Teunis, P.F.M. (2015). Entero- and parechovirus distributions in surface water and probabilities of exposure to these viruses during water recreation. Water Res., 75 : 25–32.
Lodder, w.J., van den Berg, H. H. J. L., Rutjes, S.A., et de Roda Husman, A. M. (2010). Presence of enteric viruses in source waters for drinking water production in the Netherlands. Appl. Environ. Microbiol. 76(17); 5965-5971.
Logsdon, G.S., Kohne, R., Abel, S. et LaBonde, S. (2002). Slow sand filtration for small water systems. J. Environ. Eng. Sci., 1(5): 339–348.
Lönnrot, M., Korpela, K., Knip, M., Ilonen, J., Simell, O., Korhonen, S., Savola, K., Muona, P., Simell, T., Koskela, P. et Hyoty, H. (2000). Enterovirus infections as a risk factor for ß-cell autoimmunity in a prospectively observed birth cohort: The Finnish diabetes prediction and prevention study. Diabetes, 49(8) : 1314–1318.
Lopman, B., Gastañaduy, P., Park, G.W., Hall, A.J., Parashar, U.D. et Vinjé, J. (2012). Environmental transmission of norovirus gastroenteritis. Curr. Opin. Virol., 2(1) : 96–102.
Loveland, J.P., Ryan, J.N., Amy, G.L. et Harvey, R.W. (1996). The reversibility of virus attachment to mineral surfaces. Colloids Surf. a Physicochem. Eng. Asp., 107 : 205–221.
Lovins, W.A., Taylor, J.S., et Hong, S.K. (2002). Micro-organism rejection by membrane systems. Environ. Eng. Sci., 19(6) : 453–465.
Lozier, J., Kitis, M., Colvin, C., Kim, J., Mi, B. et Marinas, B. (2003). Microbial removal and integrity monitoring of high-pressure membranes. AWWA Research Foundation, Denver, Colorado.
MacDougall, L., Majowicz, S., Doré, K., Flint, J., Thomas, K., Kovacs, S. et Sockett, P. (2008). Under-reporting of infectious gastrointestinal illness in british columbia, canada: Who is counted in provincial communicable disease statistics? Epidemiol. Infect., 136(2): 248-256.
Macler, B.A. et Regli, S. (1993). Use of microbial risk assessment in setting US drinking water standards. Int. J. Food Microbiol., 18(4) : 245–256.
Majowicz, S.E., Doré, K., Flint, J.A., Edge, V.L., Read, S., Buffett, M.C., McEwen, S., McNab, W.B., Stacey, D., Sockett, P. et Wilson, J.B. (2004). Magnitude and distribution of acute, self-reported gastrointestinal illness in a canadian community. Epidemiol. Infect., 132(4): 607-617.
Malley, J.P., Ballester, N.A., Margolin, A.B., Linden, K.G., Mofidi, A., Bolton, J.R., Crozes, G., Cushing, B., Mackey, E., Laine, J.M. et Janex, M. (2004). Inactivation of pathogens with innovative UV technologies. AWWA Research Foundation and American Water Works Association, Denver, Colorado.
Mamane-Gravetz, H. et Linden, K.G. (2004). Impact of particle aggregated microbes on UV disinfection. Proceedings of the American Water Works Association Water Quality Technology Conference. San Antonio, Texas, November 14–18.
Marion, J.W., Lee, C., Lee, C.S., Wang, Q., Lemeshow, S., Buckley, T.J., Saif, L.J. et Lee, J. (2014). Integrating bacterial and viral water quality assessment to predict swimming-associated illness at a freshwater beach: A cohort study. Plos One, 9(11).
Marks, P.J., Vipond, I.B., Carlisle, D., Deakin, D., Fey, R.E. et Caul, E.O. (2000). Evidence for airborne transmission of norwalk-like virus (nlv) in a hotel restaurant. Epidemiol. Infect., 124(3) : 481–487.
Marks, P.J., Vipond, I.B., Regan, F.M., Wedgwood, K., Fey, R.E. et Caul, E.O. (2003). A school outbreak of norwalk-like virus: Evidence for airborne transmission. Epidemiol. Infect., 131(1) : 727–736.
Marthaler, D., Rossow, K., Gramer, M., Collins, J., Goyal, S., Tsunemitsu, H., Kuga, K., Suzuki, T., Ciarlet, M. et Matthijnssens, J. (2012). Detection of substantial porcine group B rotavirus genetic diversity in the United States, resulting in a modified classification proposal for G genotypes. Virology, 433(1) : 85–96.
Maslova, L. (2017). Water quality responses from major hurricanes in Robertson County: a review and historic data analysis. Thèse de doctorat, Université de la Caroline du Nord, Raleigh, Caroline du Nord.
Matson, D.O. (2004). Caliciviruses and hepatitis E virus. Dans : Textbook of pediatric infectious diseases. Feigin, R.D., Cherry, J.D., Demmler, G.J. and Kaplan, S.L. (éditeurs). 5th. Saunders Co., Philadelphie, Pennsylvanie.
Matsushita, T., Matsui, Y., Shirasaki, N. et Kato, Y. (2005). Effect of membrane pore size, coagulation time, and coagulant dose on virus removal by a coagulation-ceramic microfiltration hybrid system. Desalination, 178(1–3 SPEC. ISS.) : 21–26.
Matsushita, T., Shirasaki, N., Matsui, Y. et Ohno, K. (2011). Virus inactivation during coagulation with aluminum coagulants. Chemosphere, 85(4) : 571–576.
Matsushita, T., Shirasaki, N., Tatsuki, Y. et Matsui, Y. (2013). Investigating norovirus removal by microfiltration, ultrafiltration, and precoagulation-microfiltration processes using recombinant norovirus virus-like particles and real-time immuno-PCR. Water Res., 47(15) : 5819–5827.
Mattison, K., Shukla, A., Cook, A., Pollari, F., Friendship, R., Kelton, D., Bidawid, S. et Farber, J.M. (2007). Human noroviruses in swine and cattle. Emerg. Infect. Dis., 13(8) : 1184–1188.
Maunula, L., Miettinen, I.T. et Von Bonsdorff, C.-. (2005). Norovirus outbreaks from drinking water. Emerg. Infect. Dis., 11(11) : 1716–1721.
Maunula, L.P., Klemola, A., Kauppinen, K., Söderberg, T., Nguyen, T., Pitkänen, T., Kaijalainen, S., Simonen, M.L., Miettinen, I.T., Lappalainen, M., Laine, J., Vuento, R., Kuusi, M. et Roivainen, M. (2009). Enteric viruses in a large waterborne outbreak of acute gastroenteritis in Finland. Food Environ Virol, 1 : 31–36.
Maurer, A.M. et Stürchler, D. (2000). A waterborne outbreak of small round structured virus, campylobacter and shigella co-infections in La Neuveville, Switzerland, 1998. Epidemiol. Infect., 125(2) : 325–332.
Mayer, B.K., Ryu, H. et Abbaszadegan, M. (2008). Treatability of U.S. Environmental Protection Agency contaminant candidate list viruses: removal of coxsackievirus and echovirus using enhanced coagulation. Environ. Sci. Technol., 42(18) : 6890–6896.
Mayer, B.K., Ryu, H., Gerrity, D. et Abbaszadegan, M. (2010). Development and validation of an integrated cell culture-qRTPCR assay for simultaneous quantification of coxsackieviruses, echoviruses, and polioviruses in disinfection studies. Water Sci. Technol., 61(2) : 375–387.
Mayer, B.K., Yang, Y., Gerrity, D.W. et Abbaszadegan, M. (2015). The impact of capsid proteins on virus removal and inactivation during water treatment processes. Microbiol. Insights, 8(Suppl 2) : 15–28.
McConnell, L.K., Sims, R.C. et Barnett, B.B. (1984). Reovirus removal and inactivation by slow-rate sand filtration. Appl. Environ. Microbiol., 48(4) : 818–825.
McVicar, M., Bickerton, B., Chaulk, M. et Walsh, M. (2015). UV254 and streaming current monitors can improve coagulation control in challenging conditions. Opflow, 41(7) : 26–28.
Medema, G., Loret, J., Stenström, T. et Ashbolt, N. (2006). Quantitative microbial risk assessment in the water safety plan. European Union MicroRisk Project, Report for the European Commission under the Fifth Framework Programme, Theme 4: Energy, environment and sustainable development.
Mena, K.D. et Gerba, C.P. (2009). Waterborne adenovirus. Rev. Environ. Contam. Toxicol., 198 : 133–167.
Mena, K.D., Gerba, C.P., Haas, C.N. et Rose, J.B. (2003). Risk assessment of waterborne coxsackievirus. J. Am. Water Works Assoc., 95(7) : 122–131.
Méndez, E. et Arias, C.F. (2007). Astroviruses. Dans : Fields virology. Knipe, D.M. and Howley, P.M. (éditeurs). 5th edition. Lippincott Williams and Wilkins, Philadelphie, Pennsylvanie.
Meng, Q.S. et Gerba, C.P. (1996). Comparative inactivation of enteric adenoviruses, poliovirus and coliphages by ultraviolet irradiation. Water Res., 30(11) : 2665–2668.
Meng, X., Dea, S., Engle, R.E., Friendship, R., Lyoo, Y.S., Sirinarumitr, T., Urairong, K., Wang, D., Wong, D., Yoo, D., Zhang, Y., Purcell, R.H. et Emerson, S.U. (1999). Prevalence of antibodies to the hepatitis E virus in pigs from countries where hepatitis E is common or is rare in the human population. J. Med. Virol., 59(3) : 297–302.
Mesquita, J.R., Barclay, L., Nascimento, M.S.J. et Vinjé, J. (2010). Novel norovirus in dogs with diarrhea. Emerg. Infect. Dis., 16(6) : 980–982.
Mesquita, M.F. et Emelko, M.B. (2012). Bacteriophages as surrogates for the fate and transport of pathogens in source water and in drinking water treatment processes. Dans : Bacteriophages. Kurtboke, I. (éditeurs). InTech, Rijeka, Croatie.
Mesquita, M.M.F. et Emelko, M.B. (2015). New guidance for groundwater under the direct influence of surface water: the how and why of F-specific coliphage. OWWA Annual Conference and Trade Show, Toronto Ontario.
Michen, B. et Graule, T. (2010). Isoelectric points of viruses. J. Appl. Microbiol., 109(2) : 388–397.
Mijatovic-Rustempasic, S., Roy, S., Sturgeon, M., Rungsrisuriyachai, K., Reisdorf, E., Cortese, M.M. et Bowen, M.D. (2015). Full-genome sequence of the first G8P [14] rotavirus strain detected in the United States. Genome Announc, 3(3) : e00677–15.
Mijatovic-Rustempasic, S., Roy, S., Teel, E.N., Weinberg, G.A., Payne, D.C., Parashar, U.D. et Bowen, M.D. (2016). Full genome characterization of the first G3P [24] rotavirus strain detected in humans provides evidence of interspecies reassortment and mutational saturation in the VP7 gene. J. Gen. Virol., 97(2) : 389–402.
Ming, H., Zhu, L. et Zhang, Y. (2011). Rapid quantification of infectious enterovirus from surface water in Bohai Bay, China using an integrated cell culture-qPCR assay. Mar. Pollut. Bull., 62(10) : 2047–2054.
Ministère des Finances de l'Ontario (2003a). Faits saillants du recensement 2001 : Feuille de renseignements 1, Bureau des politiques économiques, Direction de l'analyse en matière démographie et de travail, Toronto, Ontario.
Ministère des Finances de l'Ontario (2003b). Faits saillants du recensement 2001 : Feuille de renseignements 2, Bureau des politiques économiques, Direction de l'analyse en matière démographie et de travail, Toronto, Ontario.
Moe, K. et Shirley, J.A. (1982). The effects of relative humidity and temperature on the survival of human rotavirus in faeces. Arch. Virol., 72(3) : 179–186.
Mons, M.N., van der Wielen, J.M.L., Blokker, E.J.M., Sinclair, M.I., Hulshof, K.F.A.M., Dangendorf, F., Hunter, P.R. et Medema, G.J. (2007). Estimation of the consumption of cold tap water for microbiological risk assessment: An overview of studies and statistical analysis of data. J. Water Health, 5(SUPPL. 1) : 151–170.
Mooijman, K.A., Bahar, M., Contreras, N. et Havelaar, A.H. (2001). Optimisation of the ISO-method on enumeration of somatic coliphages (draft ISO 10705-2). Water Sci. Technol., 43(12) : 205–208.
Mooijman, K.A., Ghameshlou, Z., Bahar, M., Jofre, J. et Havelaar, A.H. (2005). Enumeration of bacteriophages in water by different laboratories of the European Union in two interlaboratory comparison studies. J. Virol. Methods, 127(1) : 60–68.
Morales-Morales, H.A., Vidal, G., Olszewski, J., Rock, C.M., Dasgupta, D., Oshima, K.H. et Smith, G.B. (2003). Optimization of a reusable hollow-fiber ultrafilter for simultaneous concentration of enteric bacteria, protozoa, and viruses from water. Appl. Environ. Microbiol., 69(7) : 4098–4102.
Morrissey, P.J., Johnston, P.M. et Gill, L.W. (2015). The impact of on-site wastewater from high density cluster developments on groundwater quality. J. Contam. Hydrol., 182 : 36–50.
Murphy, H.M., Prioleau, M.D., Borchardt, M.A. and Hynds, P.D. (2017). Review: Epidemiological evidence of groundwater contribution to global enteric disease, 1948–2015. Hydrogeol. J., 25(4): 981-1001.
Murphy, H.M., Thomas, M.K., Medeiros, D.T., McFadyen, S. et Pintar, K.D. (2016a). Estimating the number of cases of acute gastrointestinal illness (AGI) associated with Canadian municipal drinking water systems. Epidemiol. Infect., 144(7) : 1371–1385.
Murphy, H.M., Thomas, M.K., Schmidt, P.J., Medeiros, D.T., McFadyen, S. et Pintar, K.D. (2016b). Estimating the burden of acute gastrointestinal illness due to Giardia, Cryptosporidium, Campylobacter, E. coli O157 and norovirus associated with private wells and small water systems in Canada. Epidemiol. Infect., 144(7) : 1355–1370.
Murray, C. et Lopez, A. (1996a). Global health statistics. Harvard School of Public Health, Cambridge, Massachusetts.
Murray, C.J. et Lopez, A.D. (1996b). The global burden of disease: A comprehensive assessment of mortality and disability from disease, injury and risk factors in 1990 and projected to 2020. Harvard School of Public Health, Cambridge, Massachusetts.
MWH (2012). Water treatment principles and design. 3rd edition. John Wiley & Sons, New York, New York.
Nairn, C., Galbraith, D.N., Taylor, K.W. et Clements, G.B. (1999). Enterovirus variants in the serum of children at the onset of type 1 diabetes mellitus. Diabetic Med., 16(6) : 509–513.
Nakajima, H., Nakagomi, T., Kamisawa, T., Sakaki, N., Muramoto, K., Mikami, T., Nara, H. et Nakagomi, O. (2001). Winter seasonality and rotavirus diarrhoea in adults. Lancet, 357(9272) : 1950.
Nasser, A., Weinberg, D., Dinoor, N., Fattal, B. et Adin, A. (1995). Removal of hepatitis A virus (HAV), poliovirus and MS2 coliphage by coagulation and high rate filtration. Water Sci. Technol., 31(5–6) : 63–68.
Newcombe, G. et Dixon, D. (2006). Introduction. Dans : Interface science in drinking water treatment – theory and applications. Newcombe, G. et Dixon, D. (éditeurs). Academic Press, Londres, Royaume-Uni, p. 1–4.
New Zealand Department of Internal Affairs (2017).Report of the Havelock North Drinking Water Inquiry: Stage 2. Auckland, Nouvelle-Zélande. Disponible à l'adresse : www.dia.govt.nz/Government-Inquiry-into-Havelock-North-Drinking-Water
Nichols, G., Lane, C., Asgari, N., Verlander, N.Q. et Charlett, A. (2009). Rainfall and outbreaks of drinking water related disease and in England and Wales. J. Water Health, 7(1) : 1–8.
Nishizawa, T., Okamoto, H., Konishi, K., Yoshizawa, H., Miyakawa, Y. et Mayumi, M. (1997). A novel DNA virus (TTV) associated with elevated transaminase levels in post transfusion hepatitis of unknown etiology. Biochem. Biophys. Res. Commun., 241(1) : 92–97.
Noss, C.I. et Olivieri, V.P. (1985). Disinfecting capabilities of oxychlorine compounds. Appl. Environ. Microbiol., 50(5) : 1162–1164.
Novakowski, K. (2015). Pathogens in bedrock aquifers. A detailed study of the presence and transport of pathogens in urbanized settings with private bedrock wells. Canadian Water Network.
Novakowski, K., Bickerton, G., Lapcevic, P., Voralek, J., et Ross, N. (2006a). Measurements of groundwater velocity in discrete rock fractures. J. Contam. Hyrdol., 82:44-60.
Novakowski, K., Beatty, B., Conboy, M.J., et Lebedin, J. (2006b). Water Well Sustainability in Ontario: Expert Panel Report. Préparé à l'intention du ministère de l'Environnement de l'Ontario, Sustainable Water Well Initiative.
NRMMC-EPHC (2006). Australian guidelines for water recycling: Managing health and environmental risks (Phase 1). Adelaide, Australie. Natural Resource Management Ministerial Council, Environment Protection and Heritage Council.
NSF/ANSI (2015). Standard 419—Public drinking water equipment performance - filtration. NSF International, Ann Arbor, Michigan.
NSF/ANSI (2016a). Standard 55—Ultraviolet microbiological water treatment systems. NSF International, Ann Arbor, Michigan.
NSF/ANSI (2016b). Standard 58—Reverse osmosis drinking water treatment systems. NSF International, Ann Arbor, Michigan.
NSF/ANSI (2017). Standard 60— Drinking water treatment chemicals health effects. NSF International, Ann Arbor, Michigan.
Nwachuku, N. et Gerba, C.P. (2006). Health risks of enteric viral infections in children. Rev. Environ. Contam. Toxicol., 186 : 1–56.
Nwachuku, N., Gerba, C.P., Oswald, A. et Mashadi, F.D. (2005). Comparative inactivation of adenovirus serotypes by UV light disinfection. Appl. Environ. Microbiol., 71(9) : 5633–5636.
Nygård, K., Vold, L., Halvorsen, E., Bringeland, E., Røttingen, J.A. and Aavitsland, P. (2004). Waterborne outbreak of gastroenteritis in a religious summer camp in Norway, 2002. Epidemiol. Infect., 132(2) : 223–229.
Nygård, K., Wahl, E., Krogh, T., Tveit, O.A., Bøhleng, E., Tverdal, A. et Aavitsland, P. (2007). Breaks and maintenance work in the water distribution systems and gastrointestinal illness: A cohort study. Int. J. Epidemiol., 36(4) : 873–880.
O'Connor, D.R. (2002). Deuxième partie : Rapport de la Commission d'enquête sur Walkerton : stratégie relative à la salubrité de l'eau potable, ministère du Procureur général de l'Ontario.
Ogorzaly, L., Bertrand, I., Paris, M., Maul, A. et Gantzer, C. (2010). Occurrence, survival, and persistence of human adenoviruses and F-specific RNA phages in raw groundwater. Appl. Environ. Microbiol., 76(24) : 8019–8025.
Ogorzaly, L., Bonot, S., Moualij, B.E., Zorzi, W. et Cauchie, H. (2013a). Development of a quantitative immunocapture real-time PCR assay for detecting structurally intact adenoviral particles in water. J. Virol. Methods, 194(1–2) : 235–241.
Ogorzaly, L., Cauchie, H., Penny, C., Perrin, A., Gantzer, C. et Bertrand, I. (2013b). Two-day detection of infectious enteric and non-enteric adenoviruses by improved ICC-qPCR. Appl. Microbiol. Biotechnol., 97(9) : 4159–4166.
Oikarinen, S., Tauriainen, S., Hober, D., Lucas, B., Vazeou, A., Sioofy-Khojine, A., Bozas, E., Muir, P., Honkanen, H., Ilonen, J., Knip, M., Keskinen, P., Saha, M., Huhtala, H., Stanway, G., Bartsocas, C., Ludvigsson, J., Taylor, K. et Hyöty, H. (2014). Virus antibody survey in different European populations indicates risk association between coxsackievirus B1 and type 1 diabetes. Diabetes, 63(2) : 655–662.
Oishi, I., Yamazaki, K., Kimoto, T., Minekawa, Y., Utagawa, E., Yamazaki, S., Inouye, S., Grohmann, G.S., Monroe, S.S., Stine, S.E., Carcamo, C., Ando, T. et Glass, R.I. (1994). A large outbreak of acute gastroenteritis associated with astrovirus among students and teachers in Osaka, Japan. J. Infect. Dis., 170(2) : 439–443.
Olszewski, J., Winona, L. et Oshima, K.H. (2005). Comparison of 2 ultrafiltration systems for the concentration of seeded viruses from environmental waters. R. Can. microbiol., 51(4) : 295–303.
OMS (2004). Directives de qualité pour l'eau de boisson. 3e édition. Organisation mondiale de la Santé, Genève, Suisse.
OMS (2012). Planifier la gestion de la sécurité sanitaire de l'eau pour l'approvisionnement en eau des petites communautés. Organisation mondiale de la Santé, Genève, Suisse.
OMS (2017). Directives pour la qualité de l'eau de boisson. 4ème édition. Organisation mondiale de la Santé. Genève, Suisse. Disponible à : https://www.who.int/water_sanitation_health/water-quality/guidelines/fr/
O'Neil, A.E., Richen, D. et Lundrie, P. (1985). A waterborne epidemic of acute infectious non-bacterial gastroenteritis in Alberta, Canada. R. Can. santé publique, 76(3) : 199–203.
ÖNORM (2001). Austrian national standard: ÖNORM M 5873-1E, plants for disinfection of water using ultraviolet radiation: requirements and testing, part 1: low pressure mercury lamp plants . Austrian Standards Institute, Vienne, Autriche.
ÖNORM (2003). Austrian national standard: ÖNORM 5873-2 E, plants for disinfection of water using ultraviolet radiation: requirements and testing, part 2: medium pressure mercury lamp plants. Austrian Standards Institute, Vienne, Autriche.
Oosterveld, A. (2017). Personal communication. Viqua, Guelph, Ontario.
O'Reilly, C.E., Bowen, A.B., Perez, N.E., Sarisky, J.P., Shepherd, C.A., Miller, M.D., Hubbard, B.C., Herring, M., Buchanan, S.D., Fitzgerald, C.C., Hill, V., Arrowood, M.J., Xiao, L.X., Hoekstra, R.M., Mintz, E.D., Lynch, M.F., Bannerman, T., Bradley, B., Bratka, L., Carmean, J., Fugitt, R., Golden, S., Koch, E., Murphy, M., Salehi, E., Smith, F., Osborn, N.C., Wertenbach, J.M., Young, S.M., Baker, M., Bowman, C., Kolaz, H., Messer, B., Nabors, S., White, D., Achen, M., Agin, J., Brisker, T., Byrum, B., Flynn, T., Gerhardt, T., Gosnell, C., Kallis, T., Kirchner, C., McDonnell, D., Ott, M., Panico, P., Utter, C., Bopp, C.A. et Browne, L.H. (2007). A waterborne outbreak of gastroenteritis with multiple etiologies among resort island visitors and residents: Ohio, 2004. Clin. Infect. Dis., 44(4) : 506–512.
Owen, C. (1999). Microbial challenge of integrated membrane system large-scale pilot plants treating a highly organic surface water. Dans : Proceedings of the 1999 AWWA Water Quality Technology Conference, Tampa, FL. American Water Works Association, Denver, Colorado..
Page, D., Dillon, P., Toze, S., Bixio, D., Genthe, B., Jiménez Cisneros, B.E. et Wintgens, T. (2010). Valuing the subsurface pathogen treatment barrier in water recycling via aquifers for drinking supplies. Water Res., 44(6) : 1841–1852.
Pallansch, M. et Roos, R. (2007). Enteroviruses: Polioviruses, coxsackieviruses, echoviruses, and newer enteroviruses. Dans : Fields virology. Knipe, D.M. et Howley, P.M. (éditeurs). 5ème édition. Lippincott Williams and Wilkins, Philadelphie, Pennsylvanie.
Palombo, E.A. et Bishop, R.F. (1996). Annual incidence, serotype distribution, and genetic diversity of human astrovirus isolates from hospitalized children in Melbourne, Australia. J. Clin. Microbiol., 34(7) : 1750–1753.
Pang, L. (2009). Microbial removal rates in subsurface media estimated from published studies of field experiments and large intact soil cores. J. Environ. Qual., 38(4): 1531–1559.
Pang, L., Close, M., Goltz, M., Noonan, M. et Sinton, L. (2005). Filtration and transport of bacillus subtilis spores and the F-RNA phage MS2 in a coarse alluvial gravel aquifer: Implications in the estimation of setback distances. J. Contam. Hydrol., 77(3) : 165–194.
Pang, X.L., Bonita, E.L., Mooney, D., Gabos, S., Craik, S., Zurawell, R. et Neumann, N. (2014). Détection des entérovirus dans les eaux de surface et les eaux souterraines en Alberta au Canada. Communication présentée à la 16e Conférence nationale canadienne sur l'eau potable, Gatineau, Québec.
Pang, X.L., Lee, B.E., Pabbaraju, K., Gabos, S., Craik, S., Payment, P. et Neumann, N. (2012). Procédures préanalytiques et analytiques pour la détection d'entérovirus et d'entérovirus dans des échantillons d'eau. J. Virol. Methods, 184(1-2) : 77-83.
Parshionikar, S.U., Willian-True, S., Fout, G.S., Robbins, D.E., Seys, S.A., Cassady, J.D. et Harris, R. (2003). Waterborne outbreak of gastroenteritis associated with a norovirus. Appl. Environ. Microbiol., 69(9) : 5263–5268.
Passantino, L., Malley Jr., J., Knudson, M., Ward, R. et Kim, J. (2004). Effect of low turbidity and algae on UV disinfection performance. J. Am. Water Works Assoc., 96(6) : 128–137.
Payment, P. (1989). Elimination of viruses and bacteria during drinking water treatment: Review of 10 years of data from the Montréal metropolitan area. Dans : Biohazards of drinking water treatment. Larson, R.A. (éditeur). Lewis Publishers, Chelsea, Michigan, p. 59–65.
Payment, P. (1991). Fate of human enteric viruses, coliphages, and clostridium perfringens during drinking-water treatment. R. Can. microbiol., 37(2) : 154–157.
Payment, P. (1993). Viruses in water: An underestimated health risk for a variety of diseases. Dans : Disinfection dilemma: Microbiological control versus by-products. Robertson, W., Tobin, R. et Kjartanson, K. (éditeurs). American Water Works Association, Denver, Colorado, p. 157–164.
Payment, P. (1999). Poor efficacy of residual chlorine disinfectant in drinking water to inactivate waterborne pathogens in distribution systems. R. Can. microbiol., 45(8) : 709–715.
Payment, P. (2001). Cultivation of viruses from environmental samples. Dans : Manual of environmental microbiology. Hurst, C.J., Crawford, R.L., Knudsen, G.R., McInerney, M.J. et Stenzenbach, L.D. (éditeurs). 2ème édition. American Society for Microbiology, Washington, DC.
Payment, P. (2007). Cultivation and assay of animal viruses. Dans : Manual of environmental microbiology. Hurst, C., Crawford, R., Garland, J., Lipson, D., Mills, A. et Stetzenbach, L. (éditeurs). 3ème edition. American Society for Microbiology, Washington, DC.
Payment, P. et Armon, R. (1989). Virus removal by drinking water treatment processes. Crit. Rev. Environ. Control, 19(1) : 15–31.
Payment, P. et Franco, E. (1993). Clostridium perfringens and somatic coliphages as indicators of the efficiency of drinking water treatment for viruses and protozoan cysts. Appl. Environ. Microbiol., 59(8) : 2418–2424.
Payment, P. et Locas, A. (2011). Pathogens in water: Value and limits of correlation with microbial indicators. Ground Water, 49(1) : 4–11.
Payment, P. et Morin, E. (1990). Minimal infective dose of the OSU strain of porcine rotavirus. Arch. Virol., 112(3‑4) : 277–282.
Payment, P. et Pintar, K. (2006). Waterborne pathogens: A critical assessment of methods, results and data analysis. Rev. Sci. Eau, 19(3) : 233–245.
Payment, P. et Trudel, M. (1993). Methods and techniques in virology. Marcel Dekker, New York, New York.
Payment, P., Trudel, M., Sattar, S.A., Springthorpe, V.S., Subrahmanyan, T.P., Gregory, B.E., Vajdic, A.H., Blaskovic, P., Guglielmi, I.J. et Kudrewko, O. (1984). Virological examination of drinking water: a Canadian collaborative study. R. Can. microbiol., 30(1) : 105–112.
Payment, P., Trudel, M. et Plante, R. (1985). Elimination of viruses and indicator bacteria at each step of treatment during preparation of drinking water at seven water treatment plants. Appl. Environ. Microbiol., 49(6) : 1418–1428.
Payment, P., Richardson, L., Siemiatycki, J., Dewar, R., Edwardes, M. et Franco, E. (1991). A randomized trial to evaluate the risk of gastrointestinal disease due to consumption of drinking water meeting current microbiological standards. Am. J. Public Health, 81(6) : 703–708.
Payment, P., Siemiatycki, J., Richardson, L., Renaud, G., Franco, E. et Prévost, M. (1997). A prospective epidemiological study of gastrointestinal health effects due to the consumption of drinking water. Int. J. Environ. Health Res., 7(1) : 5–31.
Payment, P., Berte, A., Prévost, M., Ménard, B. et Barbeau, B. (2000). Occurrence of pathogenic microorganisms in the Saint Lawrence River (Canada) and comparison of health risks for populations using it as their source of drinking water. R. Can. microbiol., 46(6) : 565–576.
Peldszus, S., Hallé, C., Peiris, R.H., Hamouda, M., Jin, X., Legge, R.L., Budman, H., Moresoli, C. et Huck, P.M. (2011). Reversible and irreversible low-pressure membrane foulants in drinking water treatment: Identification by principal component analysis of fluorescence EEM and mitigation by biofiltration pretreatment. Water Res., 45(16) : 5161–5170.
Peldszus, S., Souza, R., Bolton, J.R., Dussert, B.W., Smith, E.F. et Andrews, S.A. (2000). Impact of medium pressure uv disinfection on the formation of low molecular weight organic by-products and nitrite, and the reduction of bromate. Proceedings, AWWA Water Quality Technology Conference, Salt Lake City, Utah.
Percival, S., Chalmers, R., Embrey, M., Hunter, P., Sellwood, J. et Wyn-Jones, P. (2004). Hepatitis E virus (HEV). Dans : Microbiology of waterborne diseases. Elsevier Academic Press, Amsterdam, Pays-Bas, p. 427–431.
Persaud, E., Levison, J., Pehme, P., Novakowski, K. and Parker, B. (2018). Cross-hole fracture connectivity assessed using hydraulic responses during liner installations in crystalline bedrock boreholes. J. Hydrol., 556:233-246.
Petterson, S., Roser, D. et Deere, D. (2015). Characterizing the concentration of cryptosporidium in Australian surface waters for setting health-based targets for drinking water treatment. J. Water Health, 13(3) : 879–896.
Pham, B., Duval, B., De Serres, G., Gilca, V., Tricco, A.C., Ochnio, J. et Scheifele, D.W. (2005). Seroprevalence of hepatitis A infection in a low endemicity country: a systematic review. BMC Infect. Dis., 5 : 56.
Phan, T.G., Trinh, Q.D., Yagyu, F., Sugita, K., Okitsu, S., Müller, W.E.G. et Ushijima, H. (2006). Outbreak of sapovirus infection among infants and children with acute gastroenteritis in Osaka City, Japan during 2004–2005. J. Med. Virol., 78(6) : 839–846.
Pieper, A.P., Ryan, J.N., Harvey, R.W., Amy, G.L., Illangasekare, T.H. et Metge, D.W. (1997). Transport and recovery of bacteriophage PRD1 in a sand and gravel aquifer: Effect of sewage-derived organic matter. Environ. Sci. Technol., 31(4) : 1163–1170.
Pina, S., Buti, M., Jardí, R., Clemente-Casares, P., Jofre, J. et Girones, R. (2001). Genetic analysis of hepatitis A virus strains recovered from the environment and from patients with acute hepatitis. J. Gen. Virol., 82(12) : 2955–2963.
Pina, S., Puig, M., Lucena, F., Jofre, J. et Girones, R. (1998). Viral pollution in the environment and in shellfish: Human adenovirus detection by PCR as an index of human viruses. Appl. Environ. Microbiol., 64(9) : 3376–3382.
Plummer, J.D. et Long, S.C. (2013). Assessment of torque teno virus as a viral pathogen indicator in waters. Water Research Foundation, 4288.
Plummer, J.D., Long, S.C., Liu, Z. et Charest, A.A. (2014). Torque teno virus occurrence and relationship to bacterial and viral indicators in feces, wastewaters, and waters in the United States. Environ. Eng. Sci., 31(12) : 671–680.
Polo, C., Perez, J.L., Mielnichuck, A.., Fedele, C., Niubo, J.et Tenorio, A. (2004). Prevalence and patterns of polyomavirus urinary excretion in immunocompetent adults and children. Clin. Microbiol. Infect., 10(7): 640-644.
Pons, W., Young, I., Truong, J., Jones-Bitton, A., McEwen, S., Pintar, K. et Papadopoulos, A. (2015). A systematic review of waterborne disease outbreaks associated with small non-community drinking water systems in Canada and the United States. Plos One, 10(10).
Powell, K.L., Taylor, R.G., Cronin, A.A., Barrett, M.H., Pedley, S., Sellwood, J., Trowsdale, S.A. et Lerner, D.N. (2003). Microbial contamination of two urban sandstone aquifers in the UK. Water Res., 37(2) : 339–352.
Powelson, D.K., Simpson, J.R. et Gerba, C.P. (1991). Effects of organic matter on virus transport in unsaturated flow. Appl. Environ. Microbiol., 57(8) : 2192–2196.
Poytner, S.F.B. et Slade, J.S. (1977). The removal of viruses by slow sand filtration. Prog. Wat. Tech., 9 : 75–88.
Praamsma, T. (2017). Rock outcrops in the Canadian Shield: an investigation of contaminant transport from surface sources in fractured rock aquifers. Thèse de doctorat, Université Queen's, Kingston, Ontario.
Purcell, R.H. (1996). Hepatitis E virus. Dans : Fields virology. Fields, B.N., Knipe, D.M. et Howley, P.M. (éditeurs). 3rd edition. Lippincott-Raven Publishers, Philadelphie, Pennsylvanie.
Pype, M-L, Lawrence, M.G., Keller, J. et Gernjak, W. (2016). Reverse osmosis integrity monitoring in water reuse: The challenge to verify virus removal – A review. Water Res., 98 : 384–395
Qiu, Y., Lee, B.E., Neumann, N., Ashbolt, N., Craik, S., Maal-Bared, R. et Pang, X.L. (2015). Assessment of human virus removal during municipal wastewater treatment in Edmonton, Canada. J. Appl. Microbiol., 119(6) : 1729–1739.
Quignon, F., Sardin, M., Kiene, L. et Schwartzbrod, L. (1997). Poliovirus-1 inactivation and interaction with biofilm: a pilot-scale study. Appl. Environ. Microbiol., 63(3) : 978–982.
Ramachandran, M., Gentsch, J.R., Parashar, U.D., Jin, S., Woods, P.A., Holmes, J.L., Kirkwood, C.D., Bishop, R.F., Greenberg, H.B., Urasawa, S., Gerna, G., Coulson, B.S., Taniguchi, K., Bresee, J.S., Glass, R.I., Nelson, R., Hartin, M., Robinson, C.C., Arcilla, Y., Mason, T.J., Hamilton, M., Yam, P.A., Aiazzi, D., Clark, H.F. et Johnson, T. (1998). Detection and characterization of novel rotavirus strains in the United States. J. Clin. Microbiol., 36(11) : 3223–3229.
Rao, V. C., Symons, J. M., Ling, A., Wang, P., Metcalf, T.G., Hoff, J. C. et Melnick J.L. (1988). Removal of hepatitis A virus and rotavirus by drinking water treatment. J. Am. Water Works Assoc., 80(2) : 59–67.
Raphael, R.A., Sattar, S.A. et Springthorpe, V.S. (1985a). Long-term survival of human rotavirus in raw and treated river water. R. Can. microbiol., 31(2) : 124–128.
Raphael, R.A., Sattar, S.A. et Springthorpe, V.S. (1985b). Rota virus concentration from raw water using positively charged filters. J. Virol. Methods, 11(2) : 131–140.
Rattanakul, S., Oguma, K. et Takizawa, S. (2015). Sequential and simultaneous applications of UV and chlorine for adenovirus inactivation. Food Environ. Virol., 7(3) : 295–304.
Ray, C., Grischek, T., Schubert, J., Wang, J.Z. et Speth, T.F. (2002b). A perspective of riverbank filtration. J. Am. Water Works Assoc., 94(4) : 149–160.
Ray, C., Schubert, J., Linsky, R.B. et Melin, G. (2002a). Introduction. Dans : Riverbank filtration: Improving source-water quality. Ray, C., Melin, G. et Linsky, R.B. (éditeurs). Kluwer Academic Publishers, Dordrecht, Pays-Bas, p. 1–15.
Regli, S., Rose, J.B., Haas, C.N. et Gerba, C.P. (1991). Modeling the risk from giardia and viruses in drinking water. J. Am. Water Works Assoc., 83(11) : 76–84.
Repp, K.K. et Keene, W.E. (2012). A point-source norovirus outbreak caused by exposure to fomites. J. Infect. Dis., 205(11) : 1639–1641.
Ressources naturelles Canada (RNCan) (2005). How well do we understand groundwater in Canada? A science case study. Sciences de la Terre, rapport no 2005030.
Reuter, G., Boros, A. et Pankovics, P. (2011). Kobuviruses - a comprehensive review. Rev. Med. Virol., 21(1) : 32‑41.
Reynolds, K.A. (2004). Integrated cell culture/PCR for detection of enteric viruses in environmental samples. Methods Mol. Biol., 268 : 69–78.
Reynolds, K.A., Gerba, C.P., Abbaszadegan, M. et Pepper, I.L. (2001). ICC/PCR detection of enteroviruses and hepatitis A virus in environmental samples. R. Can. microbiol., 47(2) : 153–157.
Rigotto, C., Victoria, M., Moresco, V., Kolesnikovas, C.K., Corrêa, A., Souza, D.S.M., Miagostovich, M.P., Simões, C.M.O. et Barardi, C.R.M. (2010). Assessment of adenovirus, hepatitis A virus and rotavirus presence in environmental samples in Florianopolis, South Brazil. J. Appl. Microbiol., 109(6) : 1979–1987.
Rijal, G., Petropoulou, C., Tolson, J.K., DeFlaun, M., Gerba, C., Gore, R., Glymph, T., Granato, T., O'Connor, C., Kollias, L. et Lanyon, R. (2009). Dry and wet weather microbial characterization of the Chicago area waterway system. Water Sci. Technol., 60(7) : 1847–1855.
Rimhanen-Finne, R., Hänninen, M., Vuento, R., Laine, J., Jokiranta, T.S., Snellman, M., Pitkänen, T., Miettinen, I. et Kuusi, M. (2010). Contaminated water caused the first outbreak of giardiasis in Finland, 2007: a descriptive study. Scand. J. Infect. Dis., 42(8) : 613–619.
Robertson, J.B. (1997). Natural protection of spring and well drinking water against surface microbial contamination. I. hydrogeological parameters. Crit. Rev. Microbiol., 23(2) : 143–178.
Robertson, W.D., Sudicky, E.A., Cherry, J.A., Rapaport, R.A. et Shimp, R.J. (1989). Impact of a domestic septic system on an unconfined sand aquifer. Contaminant Transport in Groundwater. Proc. Symposium, Stuttgart, 1989 : 105–112.
Robinson, C. et Echavarria, M. (2007). Adenoviruses. Dans : Manual of clinical microbiology. Murray, P.R., Baron, E.J., Jorgensen, J., Pfaller, M. et Landry, M.L. (éditeurs). 9th. ASM Press, p. 1589.
Robinson, C.M., Singh, G., Lee, J.Y., Dehghan, S., Rajaiya, J., Liu, E.B., Yousuf, M.A., Betensky, R.A., Jones, M.S., Dyer, D.W., Seto, D. et Chodosh, J. (2013). Molecular evolution of human adenoviruses. Sci. Rep., 3.
Rochelle, P.A., Blatchley III, E.R., Chan, P.S., Scheible,O.K. et Chengyue, S.(2011) Challenge organisms for inactivation of viruses by ultraviolet treatment. Water Research Founation et Environmental Protection Agency des É.-U., Denver, Colorado.
Roivainen, M., Alfthan, G., Jousilahti, P., Kimpimäki, M., Hovi, T. et Tuomilehto, J. (1998). Enterovirus infections as a possible risk factor for myocardial infarction. Circulation, 98(23) : 2534–2537.
Rook, J.J. (1976). Haloforms in drinking water. J. Amer. Water Wks Ass., 68(3) : 168–172.
Rosario, K., Nilsson, C., Lim, Y.W., Ruan, Y. et Breitbart, M. (2009). Metagenomic analysis of viruses in reclaimed water. Environ. Microbiol., 11(11) : 2806–2820.
Rossi, P., De Carvalho-Dill, A., Müller, I. et Aragno, M. (1994). Comparative tracing experiments in a porous aquifer using bacteriophages and fluorescent dye on a test field located at Wilerwald (Switzerland) and simultaneously surveyed in detail on a local scale by radio-magneto-tellury (12-240 kHz). Environmental Geology, 23(3) : 192–200.
Rotbart, H.A. (1995). Human enterovirus infections. Human enterovirus infections. ASM Press, Washington, DC.
Rutjes, S.A., Lodder, W.J., Van Leeuwen, A.D., et de Roda Husman, A.M. (2009). Detection of infectious rotavirus in naturally contaminated source waters for drinking water production. Appl. Environ. Microbiol., 107(1): 97–105.
Ryan, J.N., Elimelech, M., Ard, R.A., Harvey, R.W. et Johnson, P.R. (1999). Bacteriophage PRD1 and silica colloid transport and recovery in an iron oxide-coated sand aquifer. Environ. Sci. Technol., 33(1) : 63–73.
Ryu, H., Cashdollar, J.L., Fout, G.S., Schrantz, K.A. et Hayes, S. (2015). Applicability of integrated cell culture quantitative PCR (ICC-qPCR) for the detection of infectious adenovirus type 2 in UV disinfection studies. J. Environ. Sci. Health Part A Toxic Hazard. Subst. Environ. Eng., 50(8) : 777–787.
Sangsanont, J., Katayama, H., Kurisu, F. et Furumai, H. (2014). Capsid-damaging effects of UV irradiation as measured by quantitative PCR coupled with ethidium monoazide treatment. Food Environ. Virol., 6(4) : 269–275.
Sano, D., Pérez-Sautu, U., Guix, S., Pintó, R.M., Miura, T., Okabe, S. et Bosch, A. (2011). Quantification and genotyping of human sapoviruses in the llobregat River catchment, Spain. Appl. Environ. Microbiol., 77(3) : 1111‑1114.
Santé et Bien-être social Canada (1990). Intoxications alimentaires au Canada : sommaire décennal, 1975-1984, Direction générale de la protection de la santé, Ottawa, Ontario.
Santé Canada (1994). Intoxications alimentaires et maladies d'origine hydrique au Canada : sommaire annuel, 1987, Direction générale de la protection de la santé, Ottawa, Ontario.
Santé Canada (1996). Intoxications alimentaires et maladies d'origine hydrique au Canada : sommaire annuel, 1988–1989, Direction générale de la protection de la santé, Ottawa, Ontario.
Santé Canada (2006). Recommandations pour la qualité de l'eau potable au Canada : document technique – Les trihalométhanes, Bureau de la qualité de l'eau et de la santé, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2008a). Recommandations pour la qualité de l'eau potable au Canada : document technique – Le chlorite et le chlorate, Bureau de l'eau, de l'air et des changements climatiques, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2008b). Recommandations pour la qualité de l'eau potable au Canada : document technique – Les acides haloacétiques, Bureau de l'eau, de l'air et des changements climatiques, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2011). Recommandations pour la qualité de l'eau potable au Canada : document technique – N-nitrosodiméthylamine (NDMA), Bureau de l'eau, de l'air et des changements climatiques, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2012a). Recommandations pour la qualité de l'eau potable au Canada : document technique – Escherichia coli, Bureau de l'eau, de l'air et des changements climatiques, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2012b). Recommandations pour la qualité de l'eau potable au Canada : document technique – Les coliformes totaux, Bureau de l'eau, de l'air et des changements climatiques, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2012c). Recommandations pour la qualité de l'eau potable au Canada : Document technique – Protozoaires entériques : Giardia et Cryptosporidium, Bureau de l'eau, de l'air et des changements climatiques, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2012d). Recommandations pour la qualité de l'eau potable au Canada : document technique – La turbidité, Bureau de la qualité de l'eau et de l'air, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2016). Recommandations pour la qualité de l'eau potable au Canada : Document technique – Le bromate, Bureau de la qualité de l'eau et de l'air, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Santé Canada (2018). Recommandations pour la qualité de l'eau potable au Canada : Conseils sur l'utilisation de l'évaluation quantitative du risque microbien dans l'eau potable – Document de consultation public. Bureau de la qualité de l'eau et de l'air, Direction générale de la santé environnementale et de la sécurité des consommateurs, Santé Canada, Ottawa, Ontario.
Sattar, S.A. (1978). Viruses. Dans : Water and health. Viruses. Pressses de l'Université d'Ottawa, Ottawa, Ontario.
Scandura, J.E. et Sobsey, M.D. (1997). Viral and bacterial contamination of groundwater from on-site sewage treatment systems. Water Sci. Technol., 35(11–12) : 141–146.
Schaider, L.A., Ackerman, J.M. et Rudel, R.A. (2016). Septic systems as sources of organic wastewater compounds in domestic drinking water wells in a shallow sand and gravel aquifer. Sci. Total Environ., 547 : 470–481.
Schijven, J., Berger, P. et Miettinen, I. (2002). Removal of pathogens, surrogates, indicators, and toxins using riverbank filtration. Dans : Riverbank filtration: improving source-water quality. Ray, C., Melin, G. et Linsky, R.B. (éditeurs). Kluwer Academic Publishers, Dordrecht, Pays-Bas, p. 73–116.
Schijven, J.F. et Hassanizadeh, S.M. (2000). Removal of viruses by soil passage: Overview of modeling, processes, and parameters. Crit. Rev. Environ. Sci. Technol., 30(1) : 49–127.
Schijven, J.F. et Hassanizadeh, S.M. (2002). Virus removal by soil passage at field scale and groundwater protection of sandy aquifers. Water Sci. Technol., 46(3) : 123–129.
Schijven, J.F., Hassanizadeh, S.M. et de Roda Husman, A.M. (2010). Vulnerability of unconfined aquifers to virus contamination. Water Res., 44(4): 1170-1181.
Schijven, J.F., Hoogenboezem, W., Hassanizadeh, S.M. et Peters, J.H. (1999). Modeling removal of bacteriophages MS2 and PRD1 by dune recharge at Castricum, Netherlands. Water Resour. Res., 35(4) : 1101–1111.
Schijven, J.F., Hoogenboezem, W., Nobel, P.J., Medema, G.J. et Stakelbeek, A. (1998). Reduction of FRNA-bacteriophages and faecal indicator bacteria by dune infiltration and estimation of sticking efficiencies. Water Sci. Technol., 38(12) : 127–131.
Schijven, J.F., Mülschlegel, J.H.C., Hassanizadeh, S.M., Teunis, P.F.M. et de Roda Husman, A.M. (2006). Determination of protection zones for dutch groundwater wells against virus contamination - uncertainty and sensitivity analysis. J. Water Health, 4(3) : 297–312.
Schijven, J.F., Van den Berg, H.H.J.L., Colin, M., Dullemont, Y., Hijnen, W.A.M., Magic-Knezev, A., Oorthuizen, W.A. et Wubbels, G. (2013). A mathematical model for removal of human pathogenic viruses and bacteria by slow sand filtration under variable operational conditions. Water Res., 47(7) : 2592–2602.
Schmidt, P.J. (2015). Norovirus dose-response: Are currently available data informative enough to determine how susceptible humans are to infection from a single virus? Risk Anal., 35(7) : 1364–1383.
Schmidt, P.J. et Emelko, M.B. (2010). QMRA and decision-making: Are we handling measurement errors associated with pathogen concentration data correctly? Water Research, 45(2) : 461–472.
Schmidt, P.J., Emelko, M.B. et Reilly, P.M. (2010). Quantification of analytical recovery in particle and microorganism enumeration methods. Environ. Sci. Technol., 44(5) : 1705–1712.
Schmidt, P.J., Pintar, K.D.M., Fazil, A.M. et Topp, E. (2013). Harnessing the theoretical foundations of the exponential and beta-poisson dose-response models to quantify parameter uncertainty using markov chain monte carlo. Risk Anal., 33(9) : 1677–1693.
Schuster, C.J., Ellis, A.G., Robertson, W.J., Charron, D.F., Aramini, J.J., Marshall, B.J. et Medeiros, D.T. (2005). Infectious disease outbreaks related to drinking water in Canada, 1974–2001. R. Can. santé publique, 96(4) : 254–255.
Sdiri-Loulizi, K., Hassine, M., Aouni, Z., Gharbi-Khelifi, H., Chouchane, S., Sakly, N., Neji-Guédiche, M., Pothier, P., Aouni, M. et Ambert-Balay, K. (2010). Detection and molecular characterization of enteric viruses in environmental samples in Monastir, Tunisia between January 2003 and April 2007. J. Appl. Microbiol., 109(3) : 1093–1104.
Sedmak, G., Bina, D., MacDonald, J. et Couillard, L. (2005). Nine-year study of the occurrence of culturable viruses in source water for two drinking water treatment plants and the influent and effluent of a wastewater treatment plant in Milwaukee, Wisconsin (August 1994 through July 2003). Appl. Environ. Microbiol., 71(2) : 1042–1050.
Seitz, S.R., Leon, J.S., Schwab, K.J., Lyon, G.M., Dowd, M., McDaniels, M., Abdulhafid, G., Fernandez, M.L., Lindesmith, L.C., Baric, R.S. et Moe, C.L. (2011). Norovirus infectivity in humans and persistence in water. Appl. Environ. Microbiol., 77(19) : 6884–6888.
Sekla, L., Stackiw, W., Kay, C. et VanBuckenhout, L. (1980). Enteric viruses in renovated water in Manitoba. R. Can. microbiol., 26(4) : 518–523.
Sharma, S.K. et Amy, G. (2011). Chapter 15: Natural treatment systems. Dans : Water quality and treatment: A handbook on drinking water. Edzwald, J.K. (éditeurs). McGraw Hill, New York, New York, p. 15.1–15.33.
Sharp, E. (2015). Using online zeta potential measurements for coagulation control: A first for the UK water industry. Dans : Proceedings of the 6th IWA Specialist Conference on Natural Organic Matter in Drinking Water, 7–10 September, Malmö, Suède.
Sharpless, C.M. et Linden, K.G. (2001). UV photolysis of nitrate: Effects of natural organic matter and dissolved inorganic carbon and implications for UV water disinfection. Environ. Sci. Technol., 35(14) : 2949–2955.
Shin, G. et Sobsey, M.D. (2015). Removal of norovirus from water by coagulation, flocculation and sedimentation processes. Water Sci. Technol. Water Supply, 15(1) : 158–163.
Shin, G., Linden, K.G. et Faubert, G. (2009). Inactivation of giardia lamblia cysts by polychromatic UV. Lett. Appl. Microbiol., 48(6) : 790–792.
Shin, G-A., Sobsey, M.D. (2008). Inactivation of norovirus by chlorine disinfection of water. Water Res., 42(17) : 4562–4568.
Sidhu, J.P.S., Hodgers, L., Ahmed, W., Chong, M.N. et Toze, S. (2012). Prevalence of human pathogens and indicators in stormwater runoff in Brisbane, Australia. Water Res., 46(20) : 6652–6660.
Siembida-Lösch, B., Anderson, W.B., Bonsteel, J. et Huck, P.M. (2014). Pretreatment impacts on biopolymers in adjacent ultrafiltration plants. J. Am. Water Works Assoc., 106(9) : E372–E382.
Simmons, F.J., Kuo, D.H. et Xagoraraki, I. (2011). Removal of human enteric viruses by a full-scale membrane bioreactor during municipal wastewater processing. Water Res., 45(9) : 2739–2750.
Simmons, K., Gambhir, M., Leon, J. et Lopman, B. (2013). Duration of immunity to norovirus gastroenteritis. Emerg. Infect. Dis., 19(8) : 1260–1267.
Sinclair, R.G., Jones, E.L. et Gerba, C.P. (2009). Viruses in recreational water-borne disease outbreaks: A review. J. Appl. Microbiol., 107(6) : 1769–1780.
Sinclair, R.G., Rose, J.B., Hashsham, S.A., Gerba, C.P. et Haase, C.N. (2012). Criteria for selection of surrogates used to study the fate and control of pathogens in the environment. Appl. Environ. Microbiol., 78(6) : 1969–1977.
Sinigalliano, C.D., Gidley, M.L., Shibata, T., Whitman, D., Dixon, T.H., Laws, E., Hou, A., Bachoon, D., Brand, L., Amaral-Zettler, L., Gast, R.J., Steward, G.F., Nigro, O.D., Fujioka, R., Betancourf, W.Q., Vithanage, G., Mathews, J., Fleming, L.E. et Solo-Gabriele, H.M. (2007). Impacts of hurricanes Katrina and Rita on the microbial landscape of the New Orleans area. Proc. Natl. Acad. Sci. U. S. A., 104(21): 9029–9034.
Sinton, L.W., Finlay, R.K., Pang, L. et Scott, D.M. (1997). Transport of bacteria and bacteriophages in irrigated effluent into and through an alluvial gravel aquifer. Water Air Soil Pollut., 98(1–2) : 17–42.
Sinton, L.W., Noonan, M.J., Finlay, R.K., Pang, L. et Close, M.E. (2000). Transport and attenuation of bacteria and bacteriophages in an alluvial gravel aquifer. New Zealand J. Mar. Freshw. Res., 34(1) : 175–186.
Skraber, S., Gassilloud, B. et Gantzer, C. (2004). Comparison of coliforms and coliphages as tools for assessment of viral contamination in river water. Appl. Environ. Microbiol., 70(6) : 3644–3649.
Skraber, S., Schijven, J., Gantzer, C. et De Roda Husman, A.M. (2005). Pathogenic viruses in drinking-water biofilms: A public health risk? Biofilms, 2(2) : 105–117.
Slade, J.S. (1978). Enteroviruses in slow sand filtered water. J. Inst.Water Eng. Sci., 32 : 530–536.
Smeets, P.W.M.H. (2010). Chapter 2: A stochastic pathogen reduction model for full-scale treatment. Dans : Stochastic modelling of drinking water treatment in quantitative microbial risk assessment. IWA Publishing. Londres, Royaume-Uni.
Smeets, P.W.M.H., Medema, G.J. et Van Dijk, J.C. (2009). The Dutch secret: How to provide safe drinking water without chlorine in the Netherlands. Drink. Water Eng. Sci., 2(1) : 1–14.
Smith, C.J. et Osborn, A.M. (2009). Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol. Ecol., 67(1) : 6–20.
Smith, D.B., Purdy, M.A. et Simmonds, P. (2013). Genetic variability and the classification of hepatitis E virus. J. Virol., 87(8) : 4161–4169.
Smith, D.B., Simmonds, P., Jameel, S., Emerson, S.U., Harrison, T.J., Meng, X., Okamoto, H., Van der Poel, W.H.M. et Purdy, M.A. (2014). Consensus proposals for classification of the family hepeviridae. J. Gen. Virol., 95 : 2223–2232.
Smith, H.M., Reporter, R., Rood, M.P., Linscott, A.J., Mascola, L.M., Hogrefe, W. et Purcell, R.H. (2002). Prevalence study of antibody to ratborne pathogens and other agents among patients using a free clinic in downtown Los Angeles. J. Infect. Dis., 186(11) : 1673–1676.
Sobsey, M.D. (1989). Inactivation of health-related microorganisms in water by disinfection processes. Water Sci. Technol., 21(3) : 179–195.
Sobsey, M.D., Fuji, T. et Shields, P.A. (1988). Inactivation of hepatitis A virus and model viruses in water by free choline and monochloramine. Water Sci. Technol., 20(11–12) : 385–391.
Sobsey, M.D., Shields, P.A., Hauchman, F.H., Hazard, R.L. et Caton III, L.W. (1986). Survival and transport of hepatitis A virus in soils, groundwater and wastewater. Water Sci. Technol., 18(10) : 97–106.
Song, K., Mohseni, M. et Taghipour, F. (2016). Application of ultraviolet light-emitting diodes (UV-LEDs) for water disinfection: A review. Water Res., 94 : 341–349.
Song, L. et Elimelech, M. (1994). Transient deposition of colloidal particles in heterogeneous porous media. J. Colloid Interface Sci., 167(2) : 301–313.
Soucie, W.J. et Sheen, B.J. (2007). Filter-to-waste optimization. J. Am. Water Works Assoc., 99(5) : 148–157.
Staatscourant (2001). Dutch Drinking Water Decree.
Statistique Canada (2004). Enquête sur la santé dans les collectivités canadiennes : cycle 2.2 – Nutrition (vague 3) : santé générale et rappel alimentaire de 24 heures, Ottawa (Ontario).
Statistique Canada (2008). Enquête sur la santé dans les collectivités canadiennes (ESCC) : cycle 2.2 (2004) – Nutrition : Volet général sur la santé (y compris les suppléments de vitamines et de minéraux) et du rappel alimentaire de 24 heures – Guide de l'utilisateur, Ottawa, Ontario.
Statistique Canada (2012). Tables de mortalité, Canada, provinces et territoires, 2007 à 2009, ministère de l'Industrie.
Statistique Canada (2013a). Enquête sur les ménages et l'environnement (EME), 2011.
Statistique Canada (2013b). Enquête sur les usines de traitement de l'eau potable, 2011.
Stetler, R.E. (1984). Coliphages as indicators of enteroviruses. Appl. Environ. Microbiol., 48(3) : 668-670.
Stetler, R.E., Ward, R.L. et Waltrip, S.C. (1984). Enteric virus and indicator bacteria levels in a water treatment system modified to reduce trihalomethane production. Appl. Environ. Microbiol., 47(2) : 319–324.
Steyer, A., Poljšak-Prijatelj, M., Barlic-Maganja, D. et Marin, J. (2008). Human, porcine and bovine rotaviruses in Slovenia: Evidence of interspecies transmission and genome reassortment. J. Gen. Virol., 89(7) : 1690–1698.
Storey, M.V. et Ashbolt, N.J. (2003). Enteric virions and microbial biofilms - a secondary source of public health concern? Water Sci. Technol., 48(3) : 97–104.
Summa, M., von Bonsdorff, C.. et Maunula, L. (2012). Pet dogs - a transmission route for human noroviruses? J. Clin. Virol., 53(3) : 244–247.
Symonds, E.M. et Breitbart, M. (2015). Affordable enteric virus detection techniques are needed to support changing paradigms in water quality management. Clean Soil Air Water, 43(1) : 8–12.
Talbot, C., Heath, M., Wright, H. and Peters, D. (2011). 12-month UV fouling study on unfiltered source water. In: Proceedings of the AWWA Water Quality Technology Conference. American Water Works Association, Denver, Colorado.
Tapparel, C., Siegrist, F., Petty, T.J. et Kaiser, L. (2013). Picornavirus and enterovirus diversity with associated human diseases. Infec. Genet. Evol., 14(1) : 282–293.
Taylor, L.H., Latham, S.M. et Woolhouse, M.E.J. (2001). Risk factors for human disease emergence. Philos. Trans. R. Soc. B Biol. Sci., 356(1411) : 983–989.
Teixeira, D.M., Hernandez, J.M., Silva, L.D., Oliveira, D.S., Spada, P.K.P., Gurjão, T.C.M., Mascarenhas, J.D.'.P., Linhares, A.C., Morais, L.L.C.S. et Gabbay, Y.B. (2016). Occurrence of norovirus GIV in environmental water samples from Belém City, Amazon Region, Brazil. Food Environ. Virol., 8(1) : 101–104.
Templeton, M.R., Andrews, R.C. et Hofmann, R. (2005). Inactivation of particle-associated viral surrogates by ultraviolet light. Water Res., 39(15) : 3487–3500.
Templeton, M.R., Andrews, R.C. et Hofmann, R. (2006). Impact of iron particles in groundwater on the UV inactivation of bacteriophages MS2 and T4. J. Appl. Microbiol., 101(3) : 732–741.
Templeton, M.R., Andrews, R.C. et Hofmann, R. (2007). Removal of particle-associated bacteriophages by dual-media filtration at different filter cycle stages and impacts on subsequent UV disinfection. Water Res., 41(11) : 2393–2406.
Templeton, M.R., Andrews, R.C. et Hofmann, R. (2008). Particle-associated viruses in water: Impacts on disinfection processes. Crit. Rev. Environ. Sci. Technol., 38(3) : 137–164.
Teunis, P.F.M., Lodder, W.J., Heisterkamp, S.H. et De Roda Husman, A.M. (2005). Mixed plaques: Statistical evidence how plaque assays may underestimate virus concentrations. Water Res., 39(17) : 4240–4250.
Teunis, P.F.M., Moe, C.L., Liu, P., Miller, S.E., Lindesmith, L., Baric, R.S., Le Pendu, J. et Calderon, R.L. (2008). Norwalk virus: How infectious is it? J. Med. Virol., 80(8) : 1468–1476.
Teunis, P.F.M., Xu, M., Fleming, K.K., Yang, J., Moe, C.L. et Lechevallier, M.W. (2010). Enteric virus infection risk from intrusion of sewage into a drinking water distribution network. Environ. Sci. Technol., 44(22) : 8561–8566.
Tfaily, R., Papineau, I., Andrews, R.C. et Barbeau, B. (2015). Application of quantitative microbial risk assessment at 17 Canadian water treatment facilities. J. Am. Water Works Assoc., 107(10) : E497–E508.
The Pirbright Institute (2016). The picornavirus pages. Disponible à : www.picornaviridae.com.
Thomas, M.K., Charron, D.F., Waltner-Toews, D., Schuster, C., Maarouf, A.R. et Holt, J.D. (2006). A role of high impact weather events in waterborne disease outbreaks in Canada, 1975-2001. Int. J. Environ. Health Res., 16(3) : 167–180.
Thomas, M.K., Murray, R., Flockhart, L., Pintar, K., Pollari, F., Fazil, A., Nesbitt, A. et Marshall, B. (2013). Estimates of the burden of foodborne illness in canada for 30 specified pathogens and unspecified agents, circa 2006. Foodborne Pathog. Dis., 10(7) : 639–648.
Thurston-Enriquez, J.A., Haas, C.N., Jacangelo, J. et Gerba, C.P. (2003a). Chlorine inactivation of adenovirus type 40 and feline calicivirus. Appl. Environ. Microbiol., 69(7) : 3979–3985.
Thurston-Enriquez, J.A., Haas, C.N., Jacangelo, J., Riley, K. et Gerba, C.P. (2003b). Inactivation of feline calicivirus and adenovirus type 40 by UV radiation. Appl. Environ. Microbiol., 69(1) : 577–582.
Thurston-Enriquez, J.A., Haas, C.N., Jacangelo, J. et Gerba, C.P. (2005a). Inactivation of enteric adenovirus and feline calicivirus by chlorine dioxide. Appl. Environ. Microbiol., 71(6) : 3100–3105.
Thurston-Enriquez, J.A., Haas, C.N., Jacangelo, J. et Gerba, C.P. (2005b). Inactivation of enteric adenovirus and feline calicivirus by ozone. Water Res., 39(15) : 3650–3656.
Todd, E.C.D. (1974-2001). Intoxications alimentaires et maladies d'origine hydrique au Canada : sommaires annuels, Direction générale des produits de santé et des aliments, Santé Canada, Polyscience Publications Inc., Laval, Québec.
Tu, E.T., Bull, R.A., Kim, M., McIver, C.J., Heron, L., Rawlinson, W.D. et White, P.A. (2008). Norovirus excretion in an aged-care setting. J. Clin. Microbiol., 46(6) : 2119–2121.
Tufenkji, N. et Emelko, M.B. (2011). Fate and transport of microbial contaminants. Dans : Encyclopedia of Environmental Health. J.O. Nriagu (éditeurs). Elsevier B.V. p. 715–726.
Tung-Thompson, G., Libera, D.A., Koch, K.L., De Los Reyes, F.L. et Jaykus, L-A. (2015). Aerosolization of a human norovirus surrogate, bacteriophage MS2, during simulated vomiting. Plos One, 10(8).
Uejio, C.K., Yale, S.H., Malecki, K., Borchardt, M.A., Anderson, H.A. et Patz, J.A. (2014). Drinking water systems, hydrology, and childhood gastrointestinal illness in Central and Northern Wisconsin. Am. J. Public Health, 104(4) : 639–646.
U.S. EPA (1989). Part V, environmental protection agency, 40 CFR parts 141 and 142. national primary drinking water regulations: Filtration; disinfection; turbidity; Giardia lamblia; viruses; legionella, and heterotrophic bacteria. final rule. Fed. Regist., 54 : 27485–27541.
U.S. EPA (1991). Guidance manual for compliance with the filtration and disinfection requirements for public water systems using surface water sources. U.S. Environmental Protection Agency, Washington, DC.
U.S. EPA (1996). ICR microbial laboratory manual. Office of Research and Development, U.S. Environmental Protection Agency, EPA/600/R-95/178, Washington, DC.
U.S. EPA (1999). Disinfection profiling and benchmarking guidance manual. U.S. Environmental Protection Agency, EPA/815/R-99/013, Washington, DC.
U.S. EPA (2000). Part II, environmental protection agency, 40 CFR parts 141 and 142. national primary drinking water regulations: Ground water rule: Proposed rule. Fed. Regist., 65(91) : 30193–30274.
U.S. EPA (2001a). Method 1601: Male-specific (F+) and somatic coliphage in water by two-step enrichment procedure. Office of Water, U.S. Environmental Protection Agency, EPA/821-R-01-030, Washington, DC.
U.S. EPA (2001b). Method 1602: Male-specific (F+) and somatic coliphage in water by single agar layer (SAL) procedure. Office of Water, U.S. Environmental Protection Agency, EPA/821-R-01-029, Washington, DC.
U.S. EPA (2001c). Manual of methods for virology. Office of Research and Development, U.S. Environmental Protection Agency, EPA-600/4-84/013, Washington, DC.
U.S. EPA (2001d). Low-pressure membrane filtration for pathogen removal: application, implementation and regulatory issues. U.S. Environmental Protection Agency, EPA 815-C-01-001, Cincinnati, Ohio.
U.S. EPA (2004). Long term 1 enhanced surface water treatment rule - turbidity provisions technical guidance manual. Office of Water, U.S. Environmental Protection Agency, EPA 816-R-04-007, Washington, DC.
U.S. EPA (2005). Membrane filtration guidance manual. U.S. Environmental Protection Agency, Cincinnati, Ohio.
U.S. EPA (2006a). National primary drinking water regulations: Long term 2 enhanced surface water treatment rule. final rule. Fed. Regist., 71(3): 653–702.
U.S. EPA (2006b). Ultraviolet disinfection guidance manual. Office of Water, U.S. Environmental Protection Agency, EPA 815-R-06-007, Washington, DC.
U.S. EPA (2006c). 40 CFR parts 9, 141 and 142. national primary drinking water regulations: Ground water rule. final rule. Fed. Regist., 71(216) : 65573–65660.
U.S. EPA (2006d). Occurrence and monitoring document for the final ground water rule. Office of Water (4606-M), U.S. Environmental Protection Agency, EPA 815-R-06-012, Washington, DC.
U.S. EPA (2012). Method 1615.1: Measurement of enterovirus and norovirus occurrence in water by culture and RT-qPCR. Office of Research and Development, National Exposure Research Laboratory, U.S. Environmental Protection Agency, EPA 600/R-10/181, Cincinnati, Ohio.
U.S. EPA (2014). Microbiological risk assessment (MRA) tools, methods, and approaches for water media. Office of Water, Office of Science and Technology. U.S. Environmental Protection Agency, EPA-820-R-14-009, Washington, D.C.
Vaidya, S.R., Chitambar, S.D. et Arankalle, V.A. (2002). Polymerase chain reaction-based prevalence of hepatitis A, hepatitis E and TT viruses in sewage from an endemic area. J. Hepatol., 37(1) : 131–136.
Valade, M.T., Becker, W.C. et Edzwald, J.K. (2009). Treatment selection guidelines for particle and NOM removal. J. Water Supply Res. Technol. Aqua, 58(6) : 424–432.
Van Heerden, J., Ehlers, M.M., Heim, A. et Grabow, W.O.K. (2005). Prevalence, quantification and typing of adenoviruses detected in river and treated drinking water in South Africa. J. Appl. Microbiol., 99(2) : 234–242.
Vaughn, J.M., Landry, E.F. et Thomas, M.Z. (1983). Entrainment of viruses from septic tank leach fields through a shallow, sandy soil aquifer. Appl. Environ. Microbiol., 45(5) : 1474–1480.
Verhoef, L., Hewitt, J., Barclay, L., Ahmed, S.M., Lake, R., Hall, A.J., Lopman, B., Kroneman, A., Vennema, H., Vinje, J. et Koopmans, M. (2015). Norovirus genotype profiles associated with foodborne transmission, 1999-2012. Emerg. Infect. Dis., 21(4) : 592–599.
Villar, L.M., de Paula, V.S., Diniz-Mendes, L., Lampe, E. et Gaspar, A.M.C. (2006). Evaluation of methods used to concentrate and detect hepatitis A virus in water samples. J. Virol. Methods, 137(2) : 169–176.
Vinje, J. (2015). Advances in laboratory methods for detection and typing of norovirus. J. Clin. Microbiol., 53(2) : 373–381.
VROM (2005). Inspectorate guideline: Assessment of the microbial safety of drinking water. VROM Inspectorate, Pays-Bas.
Wait, I.W. et Blatchley III, E.R. (2010). Model of radiation transmittance by inorganic fouling on UV reactor lamp sleeves. Water Environ. Res., 82(11): 2272-2278
Wait, I.W., Johnston, C.T., Schwab, A.P. et Blatchley III, E.R. (2005). The influence of oxidation reduction potential on inorganic fouling of quartz surfaces in UV disinfection systems. In: Proceedings of the AWWA Water Quality Technology Conference. American Water Works Association, Denver, Colorado.
Wait, I.W., Johnston, C.T. et Blatchley III, E.R. (2007). The influence of oxidation reduction potential and water treatment processes on quartz lamp sleeve fouling in ultraviolet disinfection reactors. Water Res., 41(11): 2427-2436.
Wallender, E.K., Ailes, E.C., Yoder, J.S., Roberts, V.A. et Brunkard, J.M. (2014). Contributing factors to disease outbreaks associated with untreated groundwater. Groundwater, 52(6) : 886–897.
Wang, D., Bolton, J.R., Andrews, S.A. et Hofmann, R. (2015). Formation of disinfection by-products in the ultraviolet/chlorine advanced oxidation process. Sci. Total Environ., 518–519 : 49–57.
Ward, R.L., Bernstein, D.I., Young, E.C., Sherwood, J.R., Knowlton, D.R. et Schiff, G.M. (1986). Human rotavirus studies in volunteers: Determination of infectious dose and serological response to infection. J. Infect. Dis., 154(5) : 871–880.
Werber, D., Lauševic, D., Mugoša, B., Vratnica, Z., Ivanovic-Nikolic, L., Žižic, L., Alexandre-Bird, A., Fiore, L., Ruggeri, F.M., Di Bartolo, I., Battistone, A., Gassilloud, B., Perelle, S., Nitzan Kaluski, D., Kivi, M., Andraghetti, R. et Pollock, K.G.J. (2009). Massive outbreak of viral gastroenteritis associated with consumption of municipal drinking water in a european capital city. Epidemiol. Infect., 137(12) : 1713–1720.
Westrell, T., Anderson, Y. et Stenström, T.A. (2006b). Drinking water consumption patterns in sweden. J. Water Health, 4(4) : 511–522.
Westrell, T., Teunis, P., van den Berg, H., Lodder, W., Ketelaars, H., Stenström, T.A. et de Roda Husman, A.M. (2006a). Short- and long-term variations of norovirus concentrations in the Meuse River during a 2-year study period. Water Res., 40(14) : 2613–2620.
Whelan, K. (2017). Assessing aquifer vulnerability of private wells through geological and geochemical analysis near the Clarington Transformer Station. Master's Thesis, University of Guelph, Guelph, Ontario.
Wilkes, G., Edge, T., Gannon, V., Jokinen, C., Lyautey, E., Medeiros, D., Neumann, N., Ruecker, N., Topp, E. et Lapen, D.R. (2009). Seasonal relationships among indicator bacteria, pathogenic bacteria, cryptosporidium oocysts, giardia cysts, and hydrological indices for surface waters within an agricultural landscape. Water Res., 43(8) : 2209–2223.
Wilson, R., Anderson, L.J., Holman, R.C., Gary, G.W. et Greenberg, H.B. (1982). Waterborne gastroenteritis due to the norwalk agent: Clinical and epidemiologic investigation. Am. J. Public Health, 72(1) : 72–74.
Wingender, J. et Flemming, H. (2011). Biofilms in drinking water and their role as reservoir for pathogens. Int. J. Hyg. Environ. Health, 214(6) : 417–423.
Woessner, W.W., Ball, P.N., DeBorde, D.C. et Troy, T.L. (2001). Viral transport in a sand and gravel aquifer under field pumping conditions. Ground Water, 39(6) : 886–894.
Wolf, S., Williamson, W., Hewitt, J., Lin, S., Rivera-Aban, M., Ball, A., Scholes, P., Savill, M. et Greening, G.E. (2009). Molecular detection of norovirus in sheep and pigs in New Zealand farms. Vet. Microbiol., 133(1–2) : 184‑189.
Wright, H., Gaithuma, D., Heath, M., Schulz, C. Bogan, T., Cabaj, A. et Schmalweiser, A. (2012). UV Disinfection Knowledge Base. Water Research Foundation, Denver, CO.
Wu, J., Chen, C., Chiang, T., Sheen, I., Chen, J., Tsai, W., Huang, Y. et Lee, S. (2000). Clinical and epidemiological implications of swine hepatitis E virus infection. J. Med. Virol., 60(2) : 166–171.
Wu, J., Long, S.C., Das, D. et Dorner, S.M. (2011). Are microbial indicators and pathogens correlated? A statistical analysis of 40 years of research. J. Water Health, 9(2) : 265–278.
Xagoraraki, I., Harrington, G.W., Assavasilavasukul, P. et Standridge, J.H. (2004). Removal of emerging waterborne pathogens and pathogen indicators pilot-scale conventional treatment. J. Am. Water Works Assoc., 96(5) : 102–113+12.
Xagoraraki, I., Kuo, D.H., Wong, K., Wong, M. et Rose, J.B. (2007). Occurrence of human adenoviruses at two recreational beaches of the Great Lakes. Appl. Environ. Microbiol., 73(24) : 7874–7881.
Yamashita, T., Kobayashi, S., Sakae, K., Nakata, S., Chiba, S., Ishihara, Y. et Isomura, S. (1991). Isolation of cytopathic small roundviruses with BS-C-1 cells from patients with gastroenteritis. J. Infect. Dis., 164 : 954–957.
Yang, J., LeChevallier, M.W., Teunis, P.F.M. et Xu, M. (2011). Managing risks from virus intrusion into water distribution systems due to pressure transients. J. Water and Health, 9(2): 291–305.
Yates, M.V. et Yates, S.R. (1988). Modeling microbial fate in the subsurface environment. Crit. Rev. Environ. Control, 17(4) : 307–344.
Yates, M.V. et Yates, S.R. (1989). Septic tank setback distances: A way to minimize virus contamination of drinking water. Ground Water, 27(2) : 202–208.
Yates, M.V., Gerba, C.P. et Kelley, L.M. (1985). Virus persistence in groundwater. Appl. Environ. Microbiol., 49(4) : 778–781.
Yates, M.V., Stetzenbach, L.D., Gerba, C.P. et Sinclair, N.A. (1990). The effect of indigenous bacteria on virus survival in ground water. J. Environ. Sci. Health Part A Environ. Sci. Eng. Toxic Hazard. Subst. Control, 25(1) : 81–100.
Yates, M.V., Yates, S.R., Wagner, J. et Gerba, C.P. (1987). Modeling virus survival and transport in the subsurface. J. Contam. Hydrol., 1(3) : 329–345.
Yayli, G., Kiliç, S. et Örmeci, A.R. (2002). Hepatitis agents with enteric transmission - an epidemiological analysis. Infection, 30(6) : 334–337.
Yoder, J., Roberts, V., Craun, G.F., Hill, V., Hicks, L.A., Alexander, N.T., Radke, V., Calderon, R.L., Hlavsa, M.C., Beach, M.J. et Roy, S.L. (2008). Surveillance for waterborne disease and outbreaks associated with drinking water and water not intended for drinking-United States, 2005-2006. MMWR Surveill Summ, 57(9) : 39–62.
Zhang, M., Zeng, C.Q., Morris, A.P. et Estes, M.K. (2000). A functional NSP4 enterotoxin peptide secreted from rotavirus-infected cells. J. Virol., 74(24) : 11663–11670.
Zhu, B., Clifford, D.A. et Chellam, S. (2005a). Virus removal by iron coagulation-microfiltration. Water Res., 39(20) : 5153–5161.
Zhu, B., Clifford, D.A. et Chellam, S. (2005b). Comparison of electrocoagulation and chemical coagulation pretreatment for enhanced virus removal using microfiltration membranes. Water Res., 39(13) : 3098–3108.
Annexe A : Liste d'acronymes
- ADN
- acide désoxyribonucléique
- ANSI
- American National Standards Institute
- ARN
- acide ribonucléique
- AVCI
- années de vie corrigées du facteur invalidité
- AVI
- années vécues avec l'invalidité
- AVP
- années de vie perdues
- C
- concentration résiduelle de désinfectant
- CDC
- Centers for Disease Control and Prevention (États-Unis)
- CG
- copies du génome
- CT
- concentration de désinfectant (C) × temps de contact avec le désinfectant (T)
- ECP
- effet cytopathogénique
- EPA
- Environmental Protection Agency (É.‑U.)
- ÉQRM
- évaluation quantitative du risque microbien
- ESSIDES
- eaux souterraines subissant l'influence directe des eaux de surface
- FB
- filtration sur berge
- ICC-PCR
- culture cellulaire intégrée et réaction en chaîne de la polymérase
- ISO
- organisation internationale de normalisation
- kD
- kilodaltons
- MF
- microfiltration
- MGA
- maladie gastro-intestinale aiguë
- NF
- nanofiltration
- NPP-UI
- nombre le plus probable d'unités infectieuses
- NSF
- NSF International
- OI
- osmose inverse
- OMS
- Organisation mondiale de la Santé
- PCR
- réaction en chaîne de la polymérase
- PE
- point d'entrée
- PU
- point d'utilisation
- qPCR
- réaction en chaîne de la polymérase quantitative
- RT-PCR
- réaction en chaîne de la polymérase avec transcriptase inverse
- SPD
- sous-produit de désinfection
- SRM
- seuil de rétention des molécules
- T
- temps de contact avec le désinfectant
- UF
- ultrafiltration
- UV
- ultraviolet
- VHA
- virus de l'hépatite A
- VHE
- virus de l'hépatite E
Annexe B : Tableaux
| Virus ou groupe de virus | Groupe (c.-à-d. nature de l'acide nucléique) | Enveloppe, forme et diamètre | Taxonomie | Stéréotypes/génotypes | Transmission | Dose infectieuse | Période d'incubation | Effets connexes sur la santé, complications et immunité |
|---|---|---|---|---|---|---|---|---|
| Généralement associés à des maladies humaines | ||||||||
Entérovirus |
Groupe IV : ARNmc (+) linéaire |
Non enveloppés 20 à 30 nm |
Famille : Picornaviridae |
12 espèces; sept associés à la maladie humaine : EV-A à EV-D et rhinovirus (RV)-A, B et C |
Voie fécale-oralepar contact direct entre personnes; ingestion d'un aliment ou d'eau contaminés (peu d'information); contact avec des vecteurs passifs; muqueuse respiratoire ou oculaire |
Inconnue |
2 à 35 jours; médiane : 7 à 14 jours |
Méningite, encéphalite, poliomyélite, myocardite; gastroentérite; affections respiratoires; infection oculaire; complications possibles : diabète type 1, myalgie, syndrome de la fatigue chronique |
Hépatovirus (hépatite A, VHA) |
Groupe IV : ARNmc (+) linéaire |
Non-enveloppés, icosahédrique 27 à 32 nm |
Famille : Picornaviridae |
S.O. |
Voie fécale-oralepar contact direct entre personnes ou ingestion d'un aliment ou d'eau contaminés peut y avoir excrétion de 3 à 10 jours avant l'apparition des symptômes |
Inconnue, mais on suppose qu'elle est faible (10 à 100 particules virales) |
15 à 50 jours; moyenne : 28 jours |
Hépatite bénigne, habituellement < 2 mois; dans un faible pourcentage de cas, maladie prolongée ou récidivante pendant jusqu'à 6 mois |
Hepevirus (hépatite E, VHE) |
Groupe IV : ARNmc (+) linéaire |
Non enveloppés 27 à 34 nm |
Famille : Hepeviridae |
4 génotypes : 1 à 4, et 24 sous-types |
Voie fécale-oralepar ingestion d'eau ou d'aliments contaminés (moins courante); zoonotique; |
Inconnue |
15 à 60 jours; moyenne : 40 jours |
hépatite; taux élevé de mortalité au cours de la grossesse aucun vaccin |
Norovirus |
Groupe IV : ARNmc (+) linéaire
|
Non enveloppés, icosa-hydrique 35 à 40 nm |
Famille : Caliciviridae |
6 géno-groupes : GI, GII, GIII, GIV, GV, GVI) 25 génotypes distincts |
Voie fécale-oralepar contact direct entre personnes ou ingestion d'aliments, d'eau ou de vomissure aérosolisée Il peut y avoir excrétion chez les sujets asymptomatiques (avant l'apparition des symptômes et 2 semaines ou plus après le rétablissement) |
Beaucoup d'incertitude au sujet de la dose infectieuse, mais considéré comme très contagieux |
12 à 48 heures |
Gastroentérite (y compris vomissements d'apparition aiguë, diarrhée aqueuse non sanglante accompagnée de crampes abdominales et de nausées), durant habituellement de 24 à 48 heures |
Rotavirus |
Groupe III : ARN bicaténaire linéaire |
Non enveloppés 80 nm |
Famille : Reoviridae |
8 groupes sérologiques : A, B, C, D, E, F, G et H Groupe A subdivisé en 28 G types et 39 P types |
Voie fécale-oralepar contact direct entre personnes, contact avec vecteurs passifs; ingestion d'un aliment ou d'eau contaminés (peu courant) |
Dose infectieuse médiane : ~ 6 particules virales |
< 48 h |
Gastroentérite; diarrhée aqueuse grave causant déshydratation, plus particulièrement chez les jeunes enfants |
| Association possible ou rare avec des maladies humaines | ||||||||
Adénovirus |
Groupe I : ADN bicaténaire linéaire |
Non enveloppés 70 à 100 nm |
Famille : Adenoviridae |
7 espèces : A, B, C, D, E, F et G plus de 60 types reconnus d'adénovirus humain |
Voie orofécale par contact de personne à personne; eau contaminée (moins courant) ou solutions ophtalmiques; contact avec des vecteurs passifs; muqueuse respiratoire ou oculaire |
Varie selon le type : |
2 à 14 jours |
Gastroentérite; affections respiratoires; infections oculaires vaccin pour l'adénovirus humain E de sérotype 4 et l'adénovirus humain B de sérotype 7 |
Aichivirus |
ARNmc(+) |
Non enveloppés, icosahédrique |
Famille : Picornaviridae |
|
Voie fécale-orale; produits de la mer excrété dans les matières fécales; détecté dans les eaux usées non traitées et traitées |
|
|
diarrhée, nausées, vomissements, douleurs abdominales et fièvre |
Astrovirus |
Groupe IV : ARN monocaténaire linéaire à brin positif [ARNmc (+)] |
Non enveloppés |
Famille : Astroviridae |
8 sérotypes |
Voie fécale-orale |
|
1 à 5 jours |
gastroentérite aucun vaccin |
Polyomavirus |
Groupe II : |
Non enveloppés 40 à 45 nm |
Famille : Polyomaviridae |
|
Incertaine; peut-être respiratoire, par les muqueuses; nourriture ou eau contaminées; |
|
|
La plupart des personnes infectées sont asymptomatiques; leucoencéphalopathie multifocale progressive |
Sapovirus |
Groupe IV : ARNmc (+) linéaire |
Non enveloppés, icosahédrique |
Famille : Caliciviridae |
5 génogroupes (GI à GV) |
Voie fécale-orale par contact direct entre personnes ou ingestion d'un aliment ou d'eau contaminés |
Faible |
1 à 2 jours |
Gastroentérite, mais asymptomatique dans de nombreux cas |
| Lieu et date | Méthodes d'échantillonnage et de détection | Virus | Fréquence des échantillons positifs | Concentration | Autres constatations | Référence |
|---|---|---|---|---|---|---|
Milwaukee, WI (É.-U.) |
Prélevé au cours de périodes de faible débit et de périodes de ruissellement accru (précipitation et fonte des neiges) |
Adénovirus |
Virus (tous) : 31/63 (49 %) |
Moyenne (tous les virus) = 56 cg/L |
Concentrations les plus élevées au cours du ruissellement du printemps, adénovirus et entérovirus infectieux détectés; observées seulement au cours d'événements de ruissellement |
Corsi et coll., 2014 |
Madison, WI (É.-U.) |
Échantillonnage aux 2 à 4 semaines, y compris 26 événements |
Adénovirus |
Virus (tous) : 82 % |
Plage = non-détect. – 530 cg/L |
S.O. |
Bradbury et coll., 2013 |
Credit River, lac Ontario (CAN) |
Échantillonnage aux 2 semaines, ainsi que lors d'événements de précipitation |
virus entériques cultivables |
STE1 = 28 % (n=25) |
Conc. max. = 0.33 NPP-UI2/L (à STE1 le 15 mars 2010)
|
Aucun virus détecté (n = 15) d'avril à sept.; virus seulement détecté (44 %, n = 41) dans les influents aux STE au cours des mois plus froids (oct. à mars) |
Edge et coll., 2013 |
Lower Yakima Valley, WA (É.-U.) |
21 échantillons |
Norovirus |
Norovirus : 2/11 (18 %) |
S.O. |
Inhibition détectée dans tous les échantillons, peut donc constituer une sous-estimation de l'exposition au risque |
Gibson et Schwab, 2011 |
Edmonton AB (CAN) |
Échantillonnage mensuel pendant 5 mois (n = 5) à chaque station |
Virus entériques humains |
S.O. |
Rossdale : |
S.O. |
EPCOR, 2011 |
Edmonton AB (CAN) |
Échantillonnage mensuel (n=12) à chaque station |
Virus entériques humains |
|
Rossdale : |
|
EPCOR, 2010 |
Milwaukee, WI (É.-U.) |
Méthode modifiée de l'U.S. EPA de la culture cellulaire par floculation organique ICR |
Virus cultivables |
STE1 : |
STE1 : max. calculé = 59,0 NPP4/100 L (févr. 2001) |
Les détections se sont produites en majorité au cours des mois plus froids |
Sedmak et coll., 2005 |
| Lieu et date | Méthode d'échantillonnage et de détection | Virus | Fréquence des puits/échantillons positifs | Concentra-tion | Autres constatations | Référence |
|---|---|---|---|---|---|---|
Nord de l'AB (CAN), 5 puits juin – oct. 2013
|
54 échantillons qPCR multiple en temps réel (rtMP)-qPCR intégrée à la culture de cellules |
Norovirus |
6/54 (11 %) provenant de deux puits
|
S.O. |
Le rotavirus est le virus détecté le plus souvent; |
Pang et coll., 2014 |
Sud du comté de Wellington, ON (CAN) |
118 échantillons pour 22 puits Échantillonnage mensuel pendant six mois RT-qPCR |
Virus entériques humains |
10/118 (8,5 %) échantillons : |
Puits privés = 1,16 –15,16 CG/L Puits municipaux = 0,09 – 15,63 CG/L |
Chaque résultat positif a montré la présence de virus une fois seulement durant toute la période d'échantillonnage |
Allen et coll., 2013 |
Madison, WI (É.-U.), 6 puits municipaux profonds (220 à 300 m) : 3 provenant d'un aquifère de calcaire/dolomite au-dessous de la couche semi-perméable régionale (confinée); |
Échantillonnage aux 2 à 4 semaines, comportant un total de 26 événements sur une période de 8 mois n = 147 RT-qPCRnote 1 Culture cellulaire (infectiosité) |
Adénovirus |
67/147 (46 %) 6/6 (100 %) puits positifs de façon répétée pendant la période d'échantillonnage |
Plage = non détectable jusqu'à 6,3 cg/L Moyenne = 0,7 cg/L Médiane = 0,2 cg/L |
L'adénovirus 41 est le sérotype identifié le plus souvent |
Bradbury et coll., 2013 |
Lower Yakima Valley, WA (É-U.), puits de profondeurs variables (10 à 250 pieds de profondeur) |
n = 10 RT-PCR |
Entérovirus |
4/10 (40 %) Aucun virus entériques détecté |
S.O. |
Inhibition détectée dans 4/10 échantillons ES : peut représenter une sous-estimation de l'exposition au risque |
Gibson et Schwab, 2011 |
WI (É.-U.), multiples communautés 2005–2007 |
Un seul échantillonnage dans 33 puits de 14 collectivités Échantillonnage mensuel dans trois puits municipaux pendant un an RT-qPCR en deux temps Test d'infectiosité par ICC-PCRnote 2 Sérotypage (séquençage) |
Entérovirus |
5/33 (15 %) puits, y compris 2/33 (6 %) pour adénovirus, 2/33 (6 %) pour entérovirus et 1/33 (3 %) pour norovirus GII; l'infectiosité (+) des deux entérovirus |
Adénovirus : 0,8 et 5,4 cg/L Entérovirus : 1,7 et 4,8 cg/L Norovirus : 77 cg/L |
Il faut de multiples échantillons de virus étalés sur plusieurs saisons pour bien décrire la vulnérabilité des puits à la contamination virale |
Hunt et coll., 2010 |
Provinces canadiennes : AB, ON et QC Puits municipaux à 25 endroits QC = oct. 2005 - nov. 2006 ON = mars - déc. 2006 AB = nov. - déc. 2006 |
167 échantillons (129 de sites propres; 38 de sites contaminés); 130 analysés pour déceler la présence de virus |
Virus cultivables totaux et : norovirus, adénovirus, entérovirus, réovirus 1, 2 et 3 |
1/130 (0,8 %) par culture cellulaire; provenait d'un site contaminé connu; analyse microscopique pour identifier des picornavirus 0/130 Tous (-) par méthodes moléculaires Aucun norovirus ni autre virus détecté |
10 NPP-UI /1 000 L |
S.O. |
Locas et coll., 2008 |
Québec (CAN), 12 puits municipaux alimentés par des aquifères différents (confinés et non confinés), divers types de sol, de profondeur et de contamination |
Échantillonnage mensuel plus 2X/mois printemps et automne 113 analysés pour détecter des virus entériques humains Culture cellulaire RT-PCR pour détecter des norovirus |
Virus totaux cultivables et norovirus |
9/113 (8 %) échantillons et 5/12 (42 %) des puits pour détecter des virus cultivables |
Plage = 3 à 589 NPP-UI/1 000 L |
S.O. |
Locas et coll., 2007 |
Sud-est du Michigan (É.-U.) |
169 échantillons et 32 paires reproductibles ont été prélevés de 38 puits. 31 puits ont fait l'objet de 5-6 échantillonnages. Les autres puits ont été analysés 1 à 2 fois. |
Virus cultivables |
- 2/93 (2 %) échantillons positifs pour virus cultivables représentant 2/34 puits (6 %) |
S.O. |
Entérovirus découverts dans 4 puits (10,5 %) par RT-PCR |
Francy et coll., 2004 |
É.-U., Îles vierges et Porto Rico 29 sites Étude d'un an |
Échantillonnage mensuel n = 321 Culture cellulaire RT-PCR multiplex |
Entérovirus |
- 50/321(16 %), représentant 21/29 (72 %) des sites |
S.O. |
S.O. |
Fout et coll., 2003 |
Wisconsin, É.-U. 50 puits résidentiels et privés dans 7 districts hydrogéologiques janvier 1999 à juin 2000 |
Échantillonnage quatre fois par année, une fois par saison RT-PCR (tous les virus) et culture cellulaire (virus entériques) |
Entérovirus, rotavirus, VHA, virus apparentés au virus de Norwalk |
5/194 (3 %) des échantillons, par Trois échantillons contenaient le VHA et l'autre, le rotavirus, un virus apparenté au virus de Norwalk et l'entérovirus Aucun virus cultivable détecté |
S.O. |
S.O. |
Borchardt et coll., 2003 |
448 sites dans 35 états américains Puits dont la profondeur varie de 15 à 152 m de zones hydrogéologiques différentes |
RT-PCR (tous les virus) et culture cellulaire (entérovirus) n= 539 |
Entérovirus rotavirus |
22/539 (4 %) des échantillons et 21/448 (4,6 %) des puits (+) pour virus infectieux |
Plage (culture cellulaire) = 0,09 à1,86 NPP-UI /100 L |
S.O. |
Abbaszadegan et coll., 2003 |
Comtés de Baltimore et de Hartford, MD (É.-U.) |
45 sites échantillonnés dans chaque comté, ainsi qu'un site au hasard RT-PCR (tous les virus) et culture cellulaire (virus entériques) |
Entérovirus |
Aucun virus cultivable détecté 1 échantillon (+) pour rotavirus déterminé par RT-PCR |
S.O. |
S.O. |
Banks et coll., 2002 |
PA (É.-U.) |
59 puits échantillonnés une seule fois pendant la période d'échantillonnage Culture cellulaire |
Virus cultivable |
5/59 (8,5 %) sur les quatre aquifères |
S.O. |
S.O. |
Lindsey et coll., 2002 |
Comtés de Worcester et de Wicomico, MD (É.-U.) Puits d'un petit réseau public de distribution d'eau dans un aquifère sablonneux semi-confiné |
n = 27 Culture cellulaire RT-PCR |
Virus cultivable |
3/27 (11 %) (+) pour virus : un déterminé par culture cellulaire et deux par RT-PCR |
S.O. |
S.O. |
Banks et coll., 2001 |
Eau souterraine provenant de différents lieux géographiques (É.-U.) |
n = 150 RT-PCR et culture cellulaire |
Entérovirus |
Virus cultivables détectés dans 13/150 (8,7 %). RT-PCR, 40/150 (26,7 %) pour entérovirus, 8/150 (5,3 % pour rotavirus et 12/150 (8 %) pour VHA |
S.O. |
Abbaszadegan et coll., 1999 |
|
|
||||||
| Lieu et date | Méthode d'échantillonnage et détection | Virus | Fréquence des échantillons positifs | Concentration | Autres constatations | Référence |
|---|---|---|---|---|---|---|
Wisconsin (É.-U.), eau potable provenant de multiples puits forés à des profondeurs différentes (23 avr. 2006– nov. 2007 (3 périodes d'échantillonnage) |
Échantillonnage mensuel Échantillonnage de l'eau de puits immédiatement après la désinfection par UV (avant le réseau de distribution) qPCR Stéréotypage (par séquençage) et culture cellulaire de l'adénovirus et des entérovirus (+) |
Adénovirus |
287/1 204 (24 %) étaient (+) pour au moins un type de virus et 3 % (41/1 204) étaient (+) pour 2 types de virus ou plus L'adénovirus était le virus le plus souvent détecté [157 (13 %)], suivi de l'entérovirus [109 (9 %)], puis du norovirus GI [51 (4 %)] VHA = 10 (1 %) Rotavirus = 1 (0,1 %) Norovirus GII = 0 (0 %) |
Maximal : Moyenne : Les cultures cellulaires d'adénovirus et les échantillons d'entérovirus qPCR (+) n'ont jamais montré d'effet cytopathique, MAIS la culture cellulaire intégrée (ICC)-qPCR d'adénovirus et d'entérovirus reproductible détecté dans 25 % et 28 % de ces échantillons, respectivement 5 sérotypes d'adénovirus identifiés dans des échantillons soumis à qPCR (+) |
1 843 épisodes d'affections MGA sur 48 semaines de surveillance Incidence des affections MGA (épisodes/année-personne) ÉQRM On a établi un lien entre des mesures qPCR et l'incidence d'affections MGA dans la population étudiée |
Borchardt et coll., 2012 |
Wisconsin (É.-U.) 8 collectivités avaient installé la désinfection UV au cours de la première année d'étude; les six autres n'avaient aucun traitement en place (changé au cours de la 2e année) ***Une collectivité a été exclue en raison de la présence récidivante de coliformes (difficile à chlorer) Périodes de surveillance :
|
Échantillonnage provenant de six à huit ménages (foyers choisis en fonction de leur emplacement dans le réseau de distribution) une fois par mois dans les puits avant et après la désinfection par UV et au robinet des ménages dans les réseaux de distribution Volume moyen de l'échantillon = 877 L (n = 902); filtré sur place Questionnaire rempli par les gestionnaires des services publics de distributions d'eau aux quatre mois (détails saisis au sujet de 12 types d'événements) RT-qPCR |
Virus entériques humains |
Adénovirus détecté le plus souvent, suivi de l'entérovirus et du norovirus GI Pour 18 échantillons post-UV qui étaient (+), 17 contenaient des adénovirus |
Concentration moyenne des virus la plus élevée dans les puits, réduite de 1 à 6 log par désinfection aux UV et augmentée d'un log dans le réseau de distribution (c.-à-d. virus entré directement dans le réseau de distribution)
|
L'étude démontre que des virus pathogènes pour l'homme peuvent entrer directement dans des réseaux de distribution et que le niveau de contamination virale était lié à des événements courants dans les réseaux de distribution On a établi le lien le plus significatif entre l'ajout (et non le remplacement) d'un tuyau et l'augmentation des concentrations de virus. Événement de chloration non associé à la prévalence des virus ou leur concentration moyenne |
Lambertini et coll., 2011, 2012 |
Région de Montréal, QC (CAN) Eau traitée provenant des 3 stations de traitement des eaux (STE) utilisant des traitements conventionnels ou plus efficaces |
Culture cellulaire |
Virus cultivables |
0/8 |
S.O. |
S.O. |
Payment et Franco, 1993 |
Montréal, Ottawa et Toronto (CAN) Trois stations de traitement de l'eau de chaque ville |
Culture cellulaire |
Virus cultivables |
0/16 |
S.O. |
S.O. |
Payment et coll., 1984 |
| Date | Lieu | Agent pathogène | Nombre est. de cas | Alimentation en eau | Causes premières | Références |
|---|---|---|---|---|---|---|
| i) Amérique du Nord | ||||||
1971 |
É.-U. (AR) |
VHAnote 2 |
98 |
petit réseau non communautaire |
contamination de l'approvisionnement en eau du puits par des effluents septiques |
AWWA, 1975 |
oct. 1972 |
É.-U. (AL) |
VHA |
50 |
petit réseau non communautaire |
contamination à la source par des effluents septiques conjuguée à un traitement inadéquat |
Baer et Walker, 1977 |
juil. 1978 |
É.-U. (PA) |
norovirus |
120 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Wilson et coll., 1982 |
juil. 1982 |
É.-U. (GA) |
VHA |
35 |
petit réseau non communautaire |
inconnue |
Bloch et coll., 1990 |
août 1986 |
É.-U. (SD) |
norovirus |
135 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Levine et coll., 1990 |
sept. 1987 |
É.-U. (PA, DE, NJ) |
norovirus |
5 000 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Levine et coll., 1990 |
juin 1988 |
É.-U. (ID) |
norovirus |
339 |
petit réseau non communautaire |
eau de puits non traitée |
Levine et coll., 1990 |
févr. 1989 |
CA (QC) |
norovirus |
26 |
petit réseau non communautaire |
inconnue |
Todd, 1971-2001 |
mars 1989 |
CA (ON) |
norovirus |
68 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Todd, 1971-2001 |
avr. 1989 |
É.-U. (AZ) |
norovirus |
900 |
petit réseau non communautaire |
eau de puits contaminée par des effluents septiques |
Herwaldt et coll., 1991 |
juil. 1989 |
CA (QC) |
norovirus |
159 |
petit réseau non communautaire |
inconnue |
Todd, 1971-2001 |
août 1989 |
CA (QC) |
norovirus |
57 |
petit réseau non communautaire |
inconnue |
Todd, 1971-2001 |
sept. 1989 |
CA (QC) |
VHA |
8 |
petit réseau non communautaire |
inconnue |
Todd, 1971-2001 |
avr. 1990 |
CA (QC) |
rotavirus |
67 |
petit réseau non communautaire |
contamination de l'eau de puits par des animaux |
Todd, 1971-2001 |
mai 1990 |
É.-U. (PA) |
VHA |
22 |
petit réseau non communautaire |
eau de puits non traitée |
Herwaldt et coll., 1991 |
avr. 1992 |
É.-U. (MO) |
VHA |
46 |
petit réseau non communautaire |
eau de puits non traitée |
Kramer et coll., 1996 |
mars 1993 |
CA (ON) |
rotavirus |
11 |
petit réseau non communautaire |
inconnue |
Todd, 1971-2001 |
mai 1994 |
CA (SK) |
VHA |
6 |
petit réseau non communautaire |
contamination par le réseau de distribution |
Todd, 1971-2001 |
juin –juil. 1995 |
CA (TY) /É.-U.(AK) |
multiples, y compris petit virus structuré rond |
126 |
restaurant (source d'approvisionnement en eau : puits) |
contamination du puits par une fosse septique |
Beller et coll., 1997 |
août 1995 |
CA (QC) |
VHA |
8 |
petit réseau non communautaire |
inconnue |
Todd, 1971-2001 |
sept. 1995 |
É.-U. (TN) |
VHA |
8 |
résidence privée (source d'approvisionnement en eau : puits) |
eau souterraine non traitée |
Yoder et coll., 2008 |
déc. 1997 |
É.-U. (NY) |
norovirus |
1 450 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Lee et coll., 2002 |
juil. 1999 |
É.-U. (NM) |
petit virus structuré rond |
70 |
petit réseau non communautaire |
eau de source insuffisamment traitée |
Lee et coll., 2002 |
juin 2000 |
É.-U. (KS) |
norovirus |
86 |
petit réseau non communautaire |
eau de puits non traitée |
Lee et coll., 2002; |
juin 2000 |
É.-U. (WV) |
norovirus |
123 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Lee et coll., 2002 |
juil. 2000 |
É.-U.(CA) |
norovirus |
147 |
petit réseau non communautaire |
eau de puits non traitée |
Lee et coll., 2002 |
janv. 2001 |
É.-U. (WY) |
norovirus |
230 |
petit réseau non communautaire |
pluies abondantes et contamination d'eau de puits non traitée par des effluents septiques |
Blackburn et coll., 2004; Gelting et coll., 2005 |
févr. 2001 |
É.-U. (WY) |
norovirus |
35 |
petit réseau non communautaire (chalet pour motoneigistes) |
contamination de l'eau de puits par des effluents septiques à cause d'un système d'élimination des eaux usées surchargé |
Anderson et coll., 2003 |
sept. 2001 |
É.-U.A. (WY) |
norovirus |
83 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Parshionikar et coll., 2003 |
juin 2002 |
É.-U. (CT) |
norovirus |
142 |
petit réseau non communautaire |
eau de puits non traitée |
Blackburn et coll., 2004 |
juil. 2002 |
É.-U. (NH) |
norovirus |
201 |
petit réseau non communautaire |
eau de puits non traitée |
Blackburn et coll., 2004 |
janv. 2004 |
É.-U. (PA) |
norovirus |
70 |
petit réseau non communautaire |
contamination qui a pénétré par le réseau de distribution |
Liang et coll., 2006 |
mai –sept. 2004 |
É.-U. (OH) |
multiple, y compris norovirus |
1 450 |
puits sur île |
contamination des eaux souterraines par des matières fécales (eaux usées) |
Fong et coll., 2007 |
juin 2006 |
É.-U. (WY) |
norovirus; Campylobacter |
139 |
petit réseau non communautaire |
eau de puits non traitée |
Yoder et coll., 2008 |
juil. 2006 |
É.-U. (MD) |
norovirus |
148 |
petit réseau non communautaire |
contamination qui a pénétré au point d'utilisation, eau de puits insuffisamment traitée |
Yoder et coll., 2008 |
déc. 2006 |
É.-U. (OR) |
norovirus |
48 |
petit réseau non communautaire |
eau de puits non traitée |
Yoder et coll., 2008 |
janv. 2007 |
É.-U. (WA) |
norovirus |
32 |
petit réseau non communautaire |
eau de puits non traitée |
Brunkard et coll., 2011 |
mai 2007 |
É.-U. (WI) |
norovirus |
229 |
petit réseau non communautaire |
eau de puits non traitée |
Borchardt et coll., 2011 |
juin 2007 |
É.-U. (CO) |
norovirus |
77 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Brunkard et coll., 2011 |
juin 2007 |
É.-U. (MD) |
norovirus |
94 |
petit réseau non communautaire |
eau de puits insuffisamment traitée |
Brunkard et coll., 2011 |
juin 2008 |
É.-U. (OK) |
norovirus |
62 |
communautaire (source d'approvisionnement en eau : puits) |
lacune du traitement et du réseau de distribution |
Brunkard et coll., 2011 |
mars 2008 |
É.-U. (TN) |
VHA |
9 |
individuel (source d'approvisionnement en eau: puits) |
eau souterraine non traitée |
Brunkard et coll., 2011 |
juil. 2009 |
É.-U. (ME) |
VHA |
2 |
résidence privée (source d'approvisionnement en eau: puits) |
inconnue |
CDC, 2013 |
juin 2010 |
É.-U. (CA) |
norovirus |
47 |
petit réseau non communautaire, restaurant (source d'approvisionnement en eau: puits) |
cause inconnue dans l'eau de puits |
CDC, 2013 |
juin 2011 |
É.-U. (NM) |
norovirus |
119 |
transitoire, non communautaire (source d'eau) |
eau souterraine non traitée |
CDC, 2015d |
août 2012 |
É.-U. (WI) |
norovirus |
19 |
transitoire non communautaire (source d'approvisionnement en eau: puits) |
eau souterraine non traitée |
CDC, 2015d |
| ii) International | ||||||
2004 |
Islande (lac Myvatn) |
norovirus |
> 100 |
petit approvisionnement rural |
source d'approvisionnement en eau non traitée |
Gunnarsdóttir et coll., 2013 |
2006 |
Nouvelle-Zélande |
norovirus |
218 |
centre de ski, approvisionnement en eau communautaire (source d'approvisionnement en eau: puits) |
source d'approvisionnement en eau contaminée par des eaux usées |
Hewitt et coll., 2007 |
2007 |
Finlande (Nokia) |
au moins sept agents pathogènes, y compris norovirus |
6 500 |
réseau municipal (source d'approvisionnement en eau: eau souterraine et eau souterraine artificielle); y compris filtration et désinfection au chlore |
contamination par des eaux usées |
Maunula et coll., 2009 |
2008 |
Montenegro (Podgorica) |
viral |
1 700 |
réseau municipal (source d'approvisionnement en eau: karstique et eau souterraine); chlorée, mais aucun résiduel |
contamination par des eaux usées |
Werber et coll., 2009 |
mars 2011 |
Italie (Sicile) |
norovirus |
156 |
réseau public (municipal) |
contamination du puits et des sources alimentant le réseau de distribution d'eau public |
Giammanco et coll., 2014 |
|
||||||
- Note de bas de page 1
-
L'espèce poliovirus n'existe plus. Ces virus appartiennent maintenant à l'espèce entérovirus C(voir la section 4.1.4).
- Note de bas de page 2
-
Pour les besoins du présent document, on entend par système d'approvisionnement en eau semi-public un système sans réseau de distribution ou doté d'un réseau minimal qui fournit de l'eau au public à partir d'une installation non reliée à une source d'approvisionnement en eau public.