
Salmonella enterica au Canada, 2006 à 2019

 Téléchargez cet article en format PDF
Téléchargez cet article en format PDFPublié par : L'Agence de la santé publique du Canada
Numéro : Volume 47-5/6, mai/juin 2021 : Répondre aux nouveaux agrégats de COVID-19
Date de publication : mai/juin 2021
ISSN : 1719-3109
Soumettre un article
À propos du RMTC
Naviguer
RMTC : Volume 47-5/6, mai/juin 2021 : Répondre aux nouveaux agrégats de COVID-19
Surveillance
Sérovars de Salmonella enterica associés à des bactériémies au Canada, 2006 à 2019
Sandeep Tamber1, Brendan Dougherty2, Kimberly Nguy2
Affiliations
1 Direction des aliments, Santé Canada, Ottawa, ON
2 Centre des maladies infectieuses d’origine alimentaire, environnementale et zoonotique, Agence de la santé publique du Canada, Ottawa, ON
Correspondance
Citation proposée
Tamber S, Dougherty B, Nguy K. Sérovars de Salmonella enterica associés à des bactériémies au Canada, 2006 à 2019. Relevé des maladies transmissibles au Canada 2021;47(5/6):284–93. https://doi.org/10.14745/ccdr.v47i56a03f
Mots-clés : typhique, salmonellose non typhique, Programme national de surveillance des maladies entériques, PNSME
Résumé
Contexte : Les membres du genre bactérien Salmonella sont à l’origine de la salmonellose, une maladie dont les manifestations cliniques varient d’une gastro-entérite autolimitée à une bactériémie plus grave, une défaillance organique et une septicémie. Le genre se compose de plus de 2 600 variantes sérologiques (sérovars). On a remarqué des différences importantes dans la pathogenèse des sérovars de Salmonella.
Objectif : Le but de cette étude était de déterminer quels sérovars de Salmonella étaient plus susceptibles d’être associés à des bactériémies au Canada.
Méthodes : Pour chaque sérovar, on a extrait les informations sur le nombre total d’infections à Salmonella et d’isolats sanguins signalés au Programme national de surveillance des maladies entériques de 2006 à 2019. Le risque (proportion) et la probabilité (cote) de bactériémie ont été calculés pour tous les sérovars.
Résultats : Sur les 96 082 cas de salmonellose signalés au Programme national de surveillance des maladies entériques pendant la période d’étude de 14 ans, 4,4 % (IC 95 % : 4,3 %–4,6 %) étaient bactériémiques. Vingt sérovars de Salmonella non typhique ont été associés à des taux plus faibles de bactériémie par rapport à tous les sérovars de Salmonella non typhique, et 19 sérovars de Salmonella non typhique ont été identifiés comme ayant des taux plus élevés, Heidelberg, Oranienburg, Schwarzengrund, Virchow, Panama et Poona comptant parmi les 25 sérovars les plus fréquemment signalés au Canada pendant la période d’étude.
Conclusion : L’identification des sérovars associés aux bactériémies à Salmonella au Canada est un premier pas vers la compréhension des différences dans la pathogenèse et la présentation de la maladie.
Introduction
Les bactéries de Salmonella sont une cause importante de maladie humaine et comptent une variété de présentations cliniquesNote de bas de page 1. Le genre comprend deux espèces, six sous-espèces et plus de 2 600 variantes sérologiques (sérovars). Bien que toutes les espèces et sous-espèces aient été rapportées comme pouvant causer des maladies, la plupart des infections humaines sont causées par S. enterica spp. enterica (sous-espèce I)Note de bas de page 2. La sous-espèce I comprend des sérovars typhiques et non typhiques. Les sérovars typhiques provoquent la fièvre typhique (entérique), une infection invasive grave qui peut entraîner une défaillance de plusieurs organes, une bactériémie et une septicémieNote de bas de page 3. La salmonellose non typhique se traduit principalement par une gastro-entérite aiguë autolimitée, bien que des infections plus graves puissent survenirNote de bas de page 3. Au Canada, la majorité des cas de salmonellose signalés annuellement, y compris environ 925 hospitalisations et 17 décès, sont dus à la Salmonella non typhiqueNote de bas de page 4.
L’une des caractéristiques de Salmonella est la présence de gènes de virulence codés sur des sections du génome appelées îlots de pathogénicité de Salmonella. Les 2 659 sérovars de Salmonella possèdent tous des îlots de pathogénicité de Salmonella et codent donc la capacité à provoquer des infectionsNote de bas de page 5. Néanmoins, seule une petite proportion de sérovars est responsable de la majorité des infections humaines signalées, plus de 80 % des cas de salmonellose signalés étant attribués à 20 sérovars alors que plus de 200 sérovars sont signalés chaque année au CanadaNote de bas de page 6. Bien que certains sérovars fréquemment signalés sont associés à des risques d’exposition plus importants parce qu’ils sont plus répandus dans l’environnement, la présence de plusieurs sérovars très répandus, mais rarement signalés suggère des différences de virulence entre les sérovarsNote de bas de page 7. Cette hypothèse est soutenue par des études sur la pathogenèse des sérovars typhiques et des sérovars de Salmonella non typhique tels que Choleraesuis, Dublin, Typhimurium et EnteritidisNote de bas de page 1Note de bas de page 7. La pathogenèse de la plupart des sérovars de la Salmonella non typhique est cependant peu étudiée et donc mal comprise.
L’examen des taux de bactériémie à Salmonella chez les humains est un moyen de comparer la virulence relative des sérovars. Mondialement, environ 2 % à 8 % des infections signalées de la salmonellose non typhique conduisent à une bactériémie, et dans certains cas, la bactériémie n’est pas précédée d’une gastro-entérite Note de bas de page 8Note de bas de page 9Note de bas de page 10Note de bas de page 11. Les patients atteints de bactériémie sont plus susceptibles de connaître des issues graves, notamment l’hospitalisation et le décès, s’ils ne sont pas traités rapidement par des antibiotiquesNote de bas de page 12Note de bas de page 13. Par conséquent, l’identification précoce des patients à haut risque peut améliorer le pronostic des bactériémies à SalmonellaNote de bas de page 9Note de bas de page 14. Les facteurs de risque humains pour la dissémination bactérienne à partir de l’intestin comprennent une immunité compromise, des conditions médicales sous-jacentes et des âges extrêmes. Les facteurs de virulence bactérienne impliqués dans ce processus comprennent des protéines d’adhésion, des protéines d’évasion immunitaire et d’autres effecteurs sécrétésNote de bas de page 3Note de bas de page 10Note de bas de page 15Note de bas de page 16.
L’objectif de cette étude était de comparer les taux de bactériémie de différentes sous-espèces et sérovars de Salmonella afin de mieux comprendre les différences de virulence. L’identification des sérovars de Salmonella qui sont plus ou moins susceptibles d’être associés à une bactériémie est une étape clé vers le développement de meilleurs prédicteurs de la bactériémie à Salmonella et, à terme, de meilleures normes de soins.
Méthodes
Sources de données
L’équipe de recherche a obtenu des données du Programme national de surveillance des maladies entériques, qui est administré par l’Agence de la santé publique du CanadaNote de bas de page 6. Les données sont un résumé des données de laboratoire soumises chaque semaine par les laboratoires de microbiologie de la santé publique provinciaux et territoriaux au Programme national de surveillance des maladies entériques. Aux fins de cette analyse, nous avons extrait les données de 2006 à 2019, y compris le nombre d’infections à Salmonella par sous-espèce, sérovar et source de spécimen pour les isolations extra-intestinales. Chaque isolation signalée représente un cas clinique. Les isolats sanguins étant récupérés dans le sang et utilisé à titre d'approximation du nombre de cas de bactériémie causés par Salmonella (bactériémie à Salmonella). Les isolats totaux sont ceux qui ont été récupéré à partir de l’ensemble des sources d’échantillons prélevées.
Analyse des données
Les informations sur la fréquence de signalement et la distribution des sous-espèces et des sérovars de Salmonella ont été mises sous forme de tableaux à l’aide de Microsoft Excel 2016 et visualisées à l’aide de GraphPad Prism 8.0.2.
Nous avons calculé les taux de bactériémie à Salmonella au fil du temps en divisant le nombre d’isolats sanguins déclarés chaque année par le nombre total de signalements de Salmonella pour cette année. Le test de tendance du chi carré/Cochran-Armitage a été utilisé pour déterminer s’il existait une tendance linéaire dans les taux de bactériémie au fil du temps. Cette analyse temporelle a été menée au niveau du genre ainsi qu’au niveau des sérovars de Salmonella non typhique individuels.
Les proportions de bactériémie (PB) des différents sérovars ou groupes de Salmonella ont été calculées en divisant le nombre d’isolats sanguins d’un sérovar/groupe donné par le nombre total d’isolats du même sérovar/groupe pour toutes les années combinées. Les intervalles de confiance (IC) à 95 % pour la proportion de bactériémie ont été calculés à l’aide de la formule suivante :
BP ±(1.96√(BP(1-BP)))/n
Pour certaines analyses, nous avons regroupé les sérovars selon la désignation de leur sous-espèce (I-VI) et la présentation de la maladie (typhique ou non typhique). Les taux de bactériémie significativement plus élevés ou plus faibles dans les sérovars ou groupes de Salmonella individuels ont été évalués à l’aide du test exact de Fisher. Pour identifier les sous-espèces et les sérovars présentant des taux plus élevés d’isolations sanguines, nous avons comparé les groupes suivants : la sous-espèce I par rapport à toutes les autres sous-espèces (II, IIIa, IIIb, IV et II-VI); les sérovars typhiques par rapport aux sérovars de Salmonella non typhique; et un seul sérovar de Salmonella non typhique par rapport à tous les sérovars de Salmonella non typhique.
Les rapports de cotes ont été calculés en comparant la probabilité qu’un sérovar donné infecte le sang(a/b) à la probabilité que toute souche de Salmonella non typhique infecte le sang(c/d); où a et b sont les nombres respectifs d’isolats sanguins et non sanguins pour chaque sérovar et c et d sont les nombres correspondants pour toutes les souches de Salmonella non typhique. Nous avons utilisé la formule suivante pour calculer l’IC à 95 % du rapport des cotes :
e^([ln(OR)±1.96√(1/a+1/b+1/c+1/d]))
Nous avons effectué tous les calculs et tests statistiques en utilisant Microsoft Excel 2016 et GraphPad Prism 8.0.2. Une valeur-p inférieure à 0,05 a été considérée comme significative pour tous les tests statistiques.
Résultats
De 2006 à 2019, 96 082 isolations de Salmonella ont été signalées au Programme national de surveillance des maladies entériques. Au cours de cette période, 4 252 isolations du système sanguin ont été signalées, soit un taux global de bactériémie de 4,4 % (IC 95 % : 4.3 %–4.6 %). L’étude des taux annuels de bactériémies a montré une tendance à la hausse sur la période de 14 ans, avec un taux d’augmentation annuel moyen de 0,2 % (IC 95 % : -0,4 % à 0,7 %) et une fourchette de 2,7 % en 2007 (IC 95 % : 2,4 %–3,2 %) à 5,9 % (IC 95 % : 5,4 %–6,5 %) en 2017(P < 0,0001, figure 1 et figure 2).
Figure 1 : Nombre total de signalements et d’isolations sanguines de bactériémies à Salmonella signalées au PNSME, 2006 à 2019Footnote a
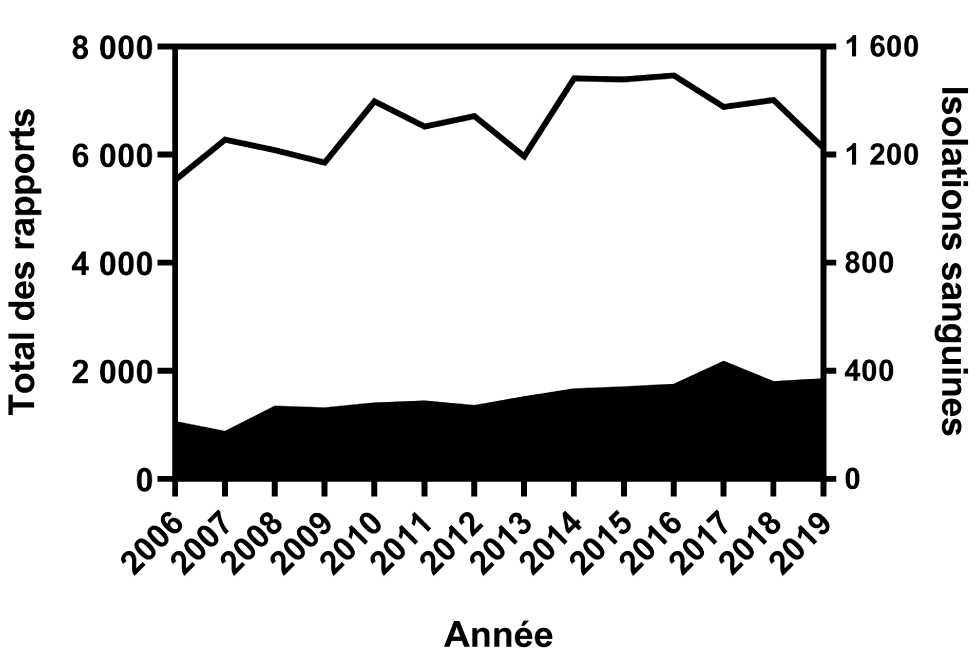
Description textuelle : Figure 1
| Année | Total des rapports | Isolations sanguines |
|---|---|---|
| 2006 | 5 745 | 211 |
| 2007 | 6 455 | 175 |
| 2008 | 6 358 | 269 |
| 2009 | 6 119 | 263 |
| 2010 | 7 268 | 280 |
| 2011 | 6 813 | 288 |
| 2012 | 6 991 | 271 |
| 2013 | 6 278 | 304 |
| 2014 | 7 748 | 333 |
| 2015 | 7 738 | 340 |
| 2016 | 7 820 | 350 |
| 2017 | 7 323 | 434 |
| 2018 | 7 375 | 359 |
| 2019 | 6 492 | 370 |
Figure 2 : Évolution dans le temps des proportions de bactériémies à Salmonella, d’après les rapports adressés au PNSME, 2006 à 2019Footnote a
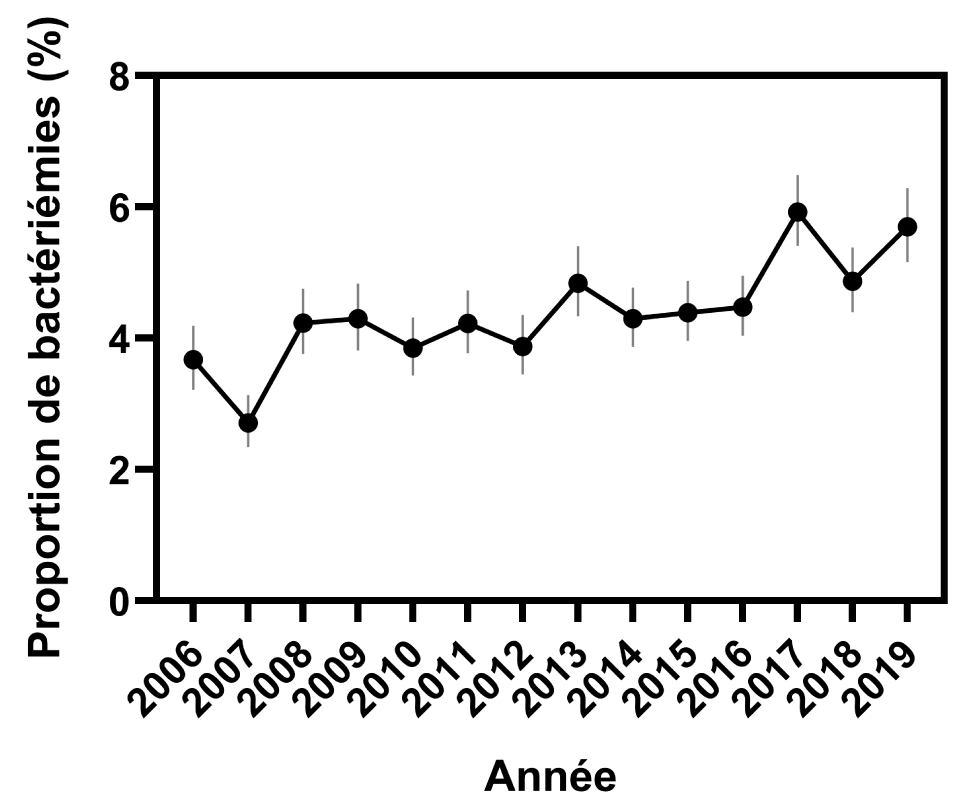
Description textuelle : Figure 2
| Année | Proportion de bactériémies (%) | Intervalle de confiance à 95 % |
|---|---|---|
| 2006 | 3,7 | 3,2–4,2 |
| 2007 | 2,7 | 2,3–3,1 |
| 2008 | 4,2 | 3,8–4,8 |
| 2009 | 4,3 | 3,8–4,8 |
| 2010 | 3,9 | 3,4–4,3 |
| 2011 | 4,2 | 3,8–4,7 |
| 2012 | 3,9 | 3,4–4,4 |
| 2013 | 4,8 | 4,3–5,4 |
| 2014 | 4,3 | 3,9–4,8 |
| 2015 | 4,4 | 4,0–4,9 |
| 2016 | 4,5 | 4,0–5,0 |
| 2017 | 5,9 | 5,4–6,5 |
| 2018 | 4,9 | 4,4–5,4 |
| 2019 | 5,7 | 5,2–6,3 |
Sur les 96 082 isolats de Salmonella signalés, 95 385 (99,3 %) ont été typés au niveau du sérovar, 258 (0,3 %) au niveau de la sous-espèce et 439 (0,5 %) au niveau du genre. Le groupe du genre a été exclu de l’analyse ultérieure. Parmi les 4 252 isolats sanguins, sept ont été exclus de l’analyse parce qu’ils n’ont été typés qu’au niveau du genre.
Le tableau 1 présente la distribution des sous-espèces des isolats typés qui ont été rapportés. Il n’y a eu que deux signalements de S. bongori pendant la période de référence; tous deux ont été isolés à partir de selles. Les autres isolats étaient des S. enterica, 99,3 % des isolats totaux et 99,5 % des isolats sanguins étant typés comme étant de la sous-espèce I.
| Espèce | Sous-espèces | Total des rapports | Rapports sanguins | ||
|---|---|---|---|---|---|
| Nombre de sérovars | Nombre d'isolats | Nombre de sérovars | Nombre d'isolats | ||
| Salmonella enterica | enterica (I) | 574 | 94 972 | 139 | 4 218 |
| salamae (II) | 40 | 80 | 2 | 2 | |
| arizoniae (IIIa) | 19 | 104 | 4 | 5 | |
| diarizonae (IIIb) | 95 | 285 | 4 | 4 | |
| houtenae (IV) | 24 | 199 | 4 | 9 | |
| indica (VI) | 1 | 1 | 0 | 0 | |
| Salmonella bongori | s.o. | 1 | 1 | 0 | 0 |
Le taux de bactériémie de la sous-espèce I était de 4,4 % (IC 95 % : 4.3 %–4.6 %). À l’exception de la sous-espèce IIIb, ce taux ne différait pas significativement de ceux des autres sous-espèces, même lorsqu’elles étaient analysées en tant que groupe agrégé pour réduire l’imprécision associée au faible nombre d’isolats (P = 0,0728, figure 3). Le taux de bactériémie de la sous-espèce IIIb, 1,4 % (IC 95 % : 0.5 %–3,5 %), était significativement plus faible que celle de la sous-espèce I (P = 0,0087).
Figure 3 : Comparaison des proportions de bactériémies des sous-espèces de Salmonella enterica, sur la base des rapports au PNSME, 2006 à 2019Footnote aFootnote bFootnote c
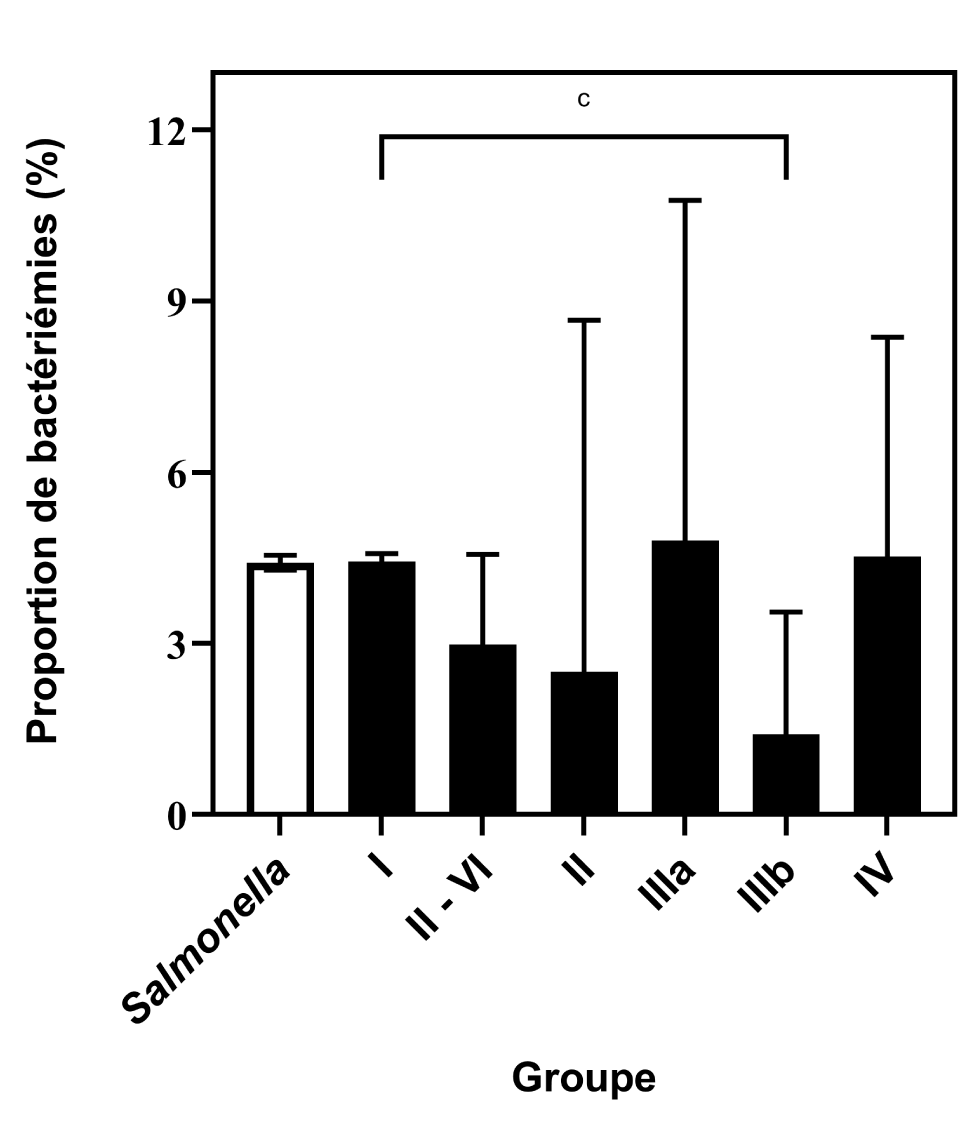
Description textuelle : Figure 3
| Sous-espèces de Salmonella | Proportion de bactériémies (%) | Intervalle de confiance à 95 % |
|---|---|---|
| Salmonella (toutes les sous-espèces, I-VI) | 4,4 | 4,3–4,6 |
| Sous-espèce I | 4,4 | 4,3–4,6 |
| Sous-espèces II-VI | 3,0 | 1,9–4,6 |
| Sous-espèce II | 2,5 | 0,4–8,7 |
| Sous-espèce IIIa | 4,8 | 2,1–10,8 |
| Sous-espèce IIIb | 1,4 | 0,5–3,6 |
| Sous-espèce IV | 4,5 | 2,4–8,4 |
Parmi les isolats de la sous-espèce I, il y avait 3 678 isolations totales de souches typhiques (2 350 Typhi, 1 198 Paratyphi A, 124 Paratyphi B et 6 Paratyphi C) et 1 295 isolations sanguines (794 Typhi, 485 Paratyphi A, 16 Paratyphi B et 0 Paratyphi C). Le taux de bactériémie des souches typhiques était de 35,2 % (IC 95 % : 33.7 %–36,8 %), significativement plus élevé que le taux calculé pour les souches non typhiques (3,2 %, IC 95 % : 3.1 %–3,3 %, P < 0,0001, figure 4).
Figure 4 : Comparaison des proportions de bactériémies dues à des souches typhiques et non typhiques des sous-espèces de Salmonella enterica, d’après les rapports transmis au PNSME, 2006 à 2019Footnote aFootnote bFootnote c
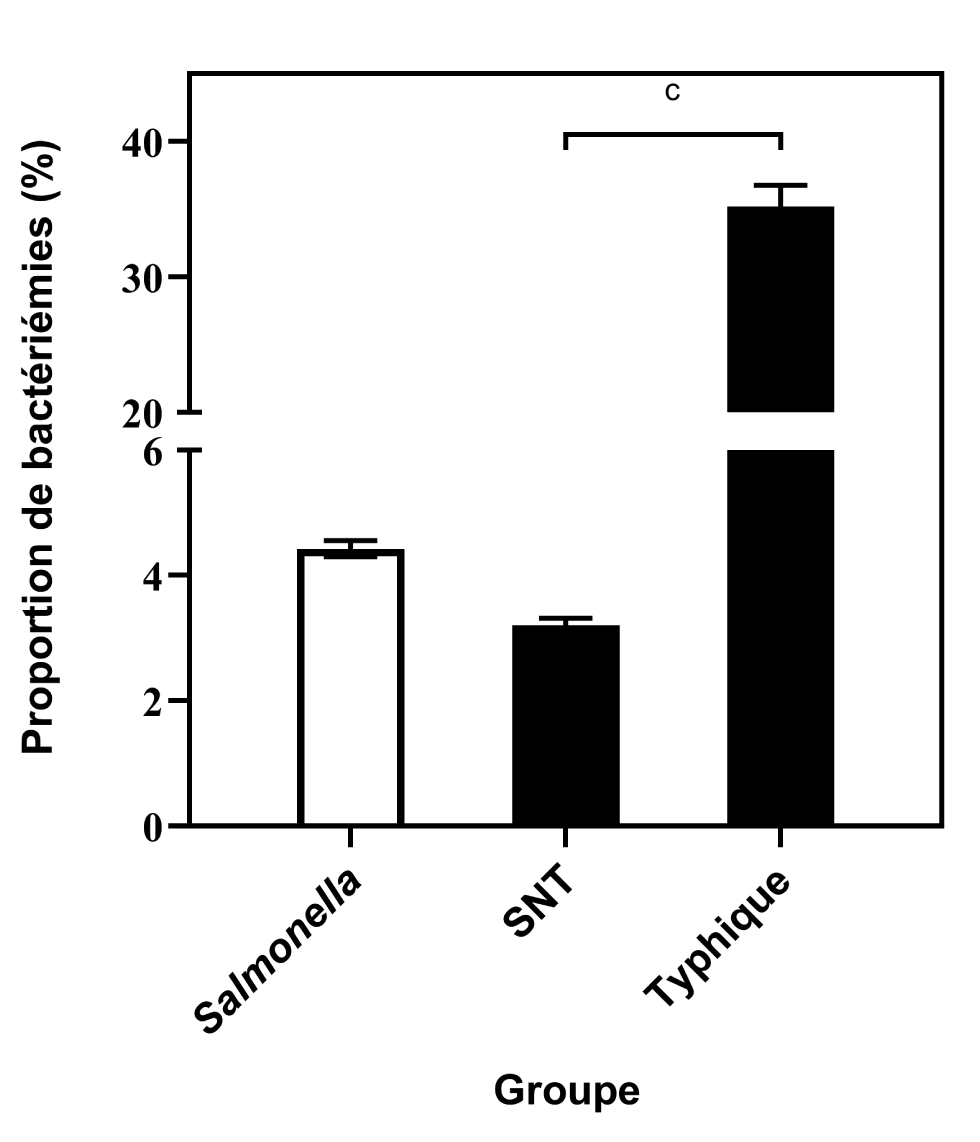
Description textuelle : Figure 4
| Groupe de Salmonella | Proportion de bactériémies (%) | Intervalle de confiance à 95 % |
|---|---|---|
| Salmonella (tous les groupes) | 4,4 | 4,3–4,6 |
| Non typhique (SNT) | 3,2 | 3,1–3,3 |
| Typhique | 35,2 | 33,7–36,8 |
Sur les 570 sérovars de la Salmonella non typhique signalés au Programme national de surveillance des maladies entériques de 2006 à 2019, 136 ont été associés à des bactériémies. Enteritidis était le plus fréquemment associé à la bactériémie, avec plus de 1 000 rapports. Avec Heidelberg, ces deux sérovars représentaient 60 % des rapports sanguins (figure 5). Typhimurium, Dublin et Oranienburg étaient, respectivement, les troisième, quatrième et cinquième sérovars les plus fréquemment signalés dans le sang (figure 5).
Figure 5 : Distribution de fréquence des cinq principaux sérovars non typhiques de Salmonella enterica isolés dans le sang, signalés au PNSME, 2006 à 2019Footnote a
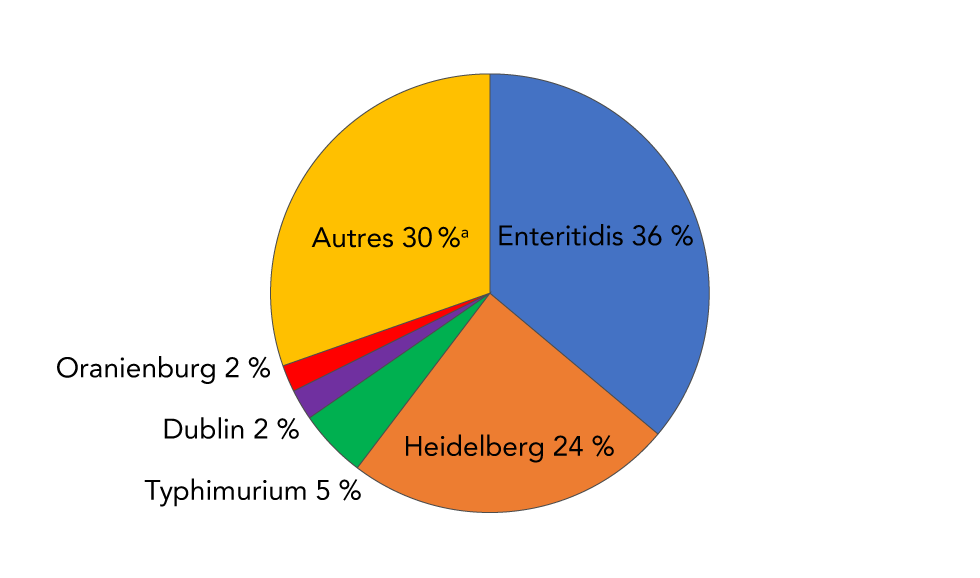
Description textuelle : Figure 5
|
Sérovars de Salmonella |
Pourcentage d'isolations sanguines |
|---|---|
| Enteritidis (bleu) | 36,1 |
| Heidelberg (orange) | 24,2 |
| Typhimurium (vert) | 5,0 |
| Dublin (violet) | 2,3 |
| Oranienburg (rouge) | 2,0 |
| Tous les autres sérovars (n = 131) (jaune) | 30,4 |
Soixante-trois sérovars n’ont été associés qu’à un seul rapport sanguin au cours de la période de signalement de 14 ans. À titre de comparaison, les deux principaux sérovars de la Salmonella non typhique, toutes sources de spécimens confondues, étaient Enteritidis et Typhimurium, représentant 50 % du total des rapports. Les sérovars Heidelberg, I 4, [5],12:i:- et Infantis complètent le top 5 (figure 6).
Figure 6 : Distribution de fréquence des cinq principales Salmonella enterica non typhiques isolées de toutes les sources d’échantillons déclarées au PNSME, 2006 à 2019Footnote a
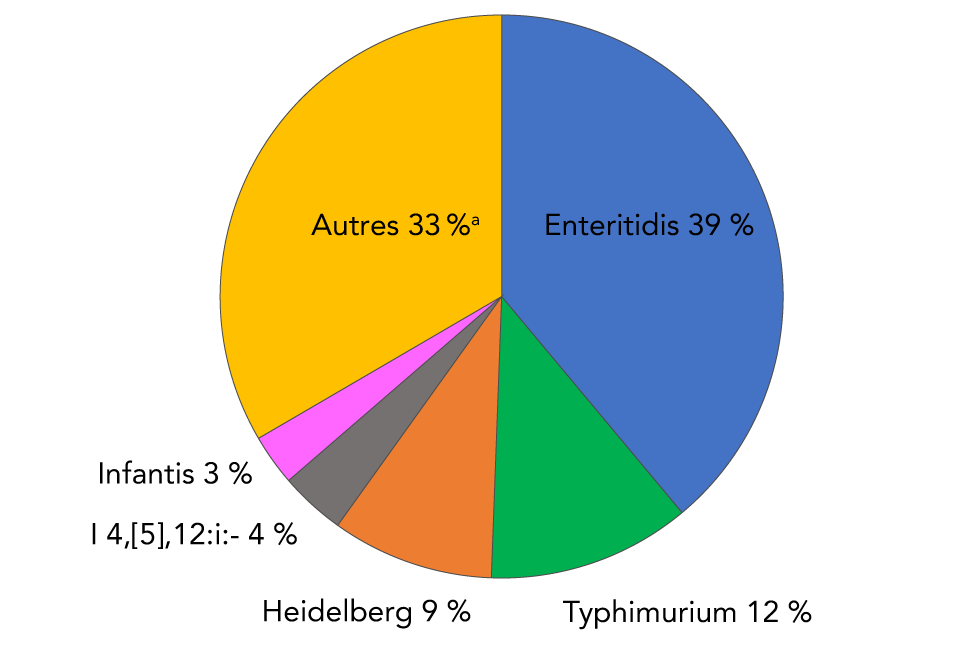
Description textuelle : Figure 6
| Sérovars de Salmonella | Pourcentage du total des isolations |
|---|---|
| Enteritidis (bleu) | 38,9 |
| Typhimurium (vert) | 11,7 |
| Heidelberg (orange) | 9,3 |
| I 4, [5],12:i:- (gris) | 3,8 |
| Infantis (rose) | 2,9 |
| Tous les autres sérovars (n = 565) (jaune) | 33,4 |
Les proportions de bactériémie pour les sérovars individuels de la Salmonella non typhique associés à au moins 10 infections sont énumérées dans le tableau 2 par ordre décroissant de fréquence.
Dix-neuf sérovars présentaient des taux de bactériémie significativement plus élevés que ceux du groupe de Salmonella non typhique. En termes de fréquence, ces sérovars varient du troisième le plus signalé (Heidelberg) au 85e (I 6.7:c:-). Les sérovars présentant les taux de bactériémie les plus élevés étaient Choleraesuis (33 signalements, taux de bactériémie 36,4 %, IC 95 % : 22.2 %–53,3 %) ; Dublin (187 rapports, taux de bactériémie 35,3 %, IC 95 % : 28.8 %–42,4 %) ; et I 6,7:c:- (30 signalements, taux de bactériémie 30,0 %, IC 95 % : 16.7 %–47.9 %).
Le tableau 3 énumère les sérovars de la Salmonella non typhique ayant fait l’objet d’au moins 10 signalements et n’ayant pas fait l’objet d’une isolation sanguine. Parmi ceux-ci, 20 présentaient des taux de bactériémie significativement plus faibles que le groupe de tous les sérovars de la Salmonella non typhique. Dans ce groupe se trouvaient Enteritidis et Typhimurium. En outre, 48 sérovars signalés au moins 10 fois entre 2006 et 2019 n’étaient associés à aucune isolation sanguine.
| Sérovar | Total des isolats | valeur-pTableau 1 footnote a |
|---|---|---|
| Weltevreden | 382 | < 0,0001 |
| Senftenberg | 313 | < 0,0001 |
| Mississippi | 170 | 0,0478 |
| Blockley | 117 | 0,0568 |
| Miami | 105 | 0,0847 |
| I 9,12:-:1,5 | 100 | 0,0804 |
| Adélaïde | 86 | 0,1188 |
| Albany | 86 | 0,1188 |
| Alachua | 54 | 0,4216 |
| Concord | 53 | 0,4187 |
| Cotham | 46 | 0,4059 |
| Norwich | 46 | 0,4059 |
| Ealing | 45 | 0,4053 |
| Singapour | 44 | 0,4050 |
| Richmond | 38 | 0,6349 |
| I 4,[5],12:-:1,2 | 36 | 0,6309 |
| I 6,7:-:- | 31 | 0,6260 |
| Altona | 27 | > 0,9999 |
| I 4, [5],12:d:- | 27 | > 0,9999 |
| Potsdam | 25 | > 0,9999 |
| I Rough-O:HNM | 25 | > 0,9999 |
| Ago | 24 | > 0,9999 |
| Bardo | 24 | > 0,9999 |
| Irumu | 23 | > 0,9999 |
| Liverpool | 23 | > 0,9999 |
| Othmarschen | 23 | > 0,9999 |
| Istanbul | 21 | > 0,9999 |
| Fluntern | 20 | > 0,9999 |
| Putten | 20 | > 0,9999 |
| I 6,8:e,h:- | 20 | > 0,9999 |
| Kedougou | 19 | > 0,9999 |
| Larochelle | 19 | > 0,9999 |
| I 6,7:e,h:- | 19 | > 0,9999 |
| Isangi | 16 | > 0,9999 |
| Goettingen | 15 | > 0,9999 |
| Lexington | 15 | > 0,9999 |
| Amsterdam | 14 | > 0,9999 |
| Apapa | 14 | > 0,9999 |
| Kingabwa | 13 | > 0,9999 |
| I 13,23:b:- | 13 | > 0,9999 |
| Abony | 12 | > 0,9999 |
| I Rough-O:i:1,2 | 12 | > 0,9999 |
| Kisarawe | 11 | > 0,9999 |
| Saphra | 10 | > 0,9999 |
| I 6,8:-:- | 10 | > 0,9999 |
Les tendances de signalement des trois sérovars les plus fréquemment signalés, Enteritidis, Typhimurium et Heidelberg, sont présentées dans la figure 7. De 2006 à 2019, le nombre de rapport sanguins pour le sérovar Enteritidis est passé de 15 à 97 et le nombre total de signalements est passé de 1 338 à 2 254, soit un taux de bactériémie qui est passé de 1,1 % (IC 95 % : 0,7 %–1,8 %) à 4,1 % (IC 95 % : 3,4 %–5,0 %, P < 0.0001). Le taux de bactériémie de Typhimurium a également semblé augmenter au cours de la période de référence, passant de 0,4 % (IC 95 % : 0,2 %–1,0 %) à 2,3 % (IC 95 % : 1,3 %–3.9 %, P = 0.0031). Le nombre de rapports sanguin est resté relativement stable, avec quatre en 2006 et 13 en 2019, mais le nombre total de rapports a diminué, passant de 998 en 2006 à 557 en 2019.
Figure 7 : Taux de bactériémie des différents sérovars de Salmonella non typhique, 2006 à 2019Footnote aFootnote bFootnote c

Description textuelle : Figure 7
Sérovars de Salmonella |
Année | Total des rapports | Isolations sanguines | Proportion de bactériémies (%) | Intervalle de confiance à 95 % |
|---|---|---|---|---|---|
| Enteritidis | 2006 | 1 338 | 15 | 1,1 | 0,7–1,8 |
| Enteritidis | 2007 | 1 661 | 26 | 1,6 | 1,1–2,2 |
| Enteritidis | 2008 | 2 225 | 35 | 1,6 | 1,1–2,1 |
| Enteritidis | 2009 | 1 955 | 53 | 2,7 | 2,0–3,4 |
| Enteritidis | 2010 | 2 828 | 47 | 1,7 | 1,2–2,2 |
| Enteritidis | 2011 | 2 763 | 66 | 2,4 | 1,8–3,0 |
| Enteritidis | 2012 | 2 117 | 49 | 2,3 | 1,7–3,0 |
| Enteritidis | 2013 | 2 019 | 57 | 2,8 | 2,1–3,5 |
| Enteritidis | 2014 | 3 296 | 100 | 3,0 | 2,4–3,6 |
| Enteritidis | 2015 | 3 209 | 104 | 3,2 | 2,6–3,8 |
| Enteritidis | 2016 | 3 433 | 142 | 4,1 | 3,4–4,7 |
| Enteritidis | 2017 | 3 278 | 143 | 4,4 | 3,6–4,9 |
| Enteritidis | 2018 | 3 083 | 119 | 3,9 | 3,1–4,4 |
| Enteritidis | 2019 | 2 254 | 97 | 4,3 | 3,4–5,0 |
Les fréquences de signalement du sérovar Heidelberg ont fluctué de manière non linéaire au cours de la période de signalement, passant d’un minimum de 19 rapports sanguins en 2019 à un maximum de 99 en 2017. Le nombre total de rapports a varié entre un minimum de 267 en 2019 et un maximum de 1 071 en 2012. Le taux de bactériémie correspondant a également fluctué, passant d’un minimum de 5,4 % (IC 95 % : 3,9 %–7,5 %) en 2007 à un maximum de 18,2 % (IC 95 % : 15,2 %–21,7 %) en 2017.
Discussion
Selon les données soumises au Programme national de surveillance des maladies entériques, le taux global de bactériémie à Salmonella au Canada était de 4,4 % (IC 95 % : 4,3 %–4,6 %) de 2006 à 2019. Les taux annuels ont varié de 2,7 % à 5,9 % et ont augmenté en moyenne de 0,2 % par an au cours de la période en question. L’analyse du taux de bactériémie pour les sérovars individuels a montré qu’il évoluait dans le temps en fonction du nombre de rapports totaux et sanguins. Le taux global pour le genre devrait donc refléter les changements dans les habitudes de signalement de tous les sérovars de Salmonella au Canada. Les raisons sous-jacentes de ces changements méritent d’être étudiées plus avant, mais pourraient refléter des changements dans les voies d’exposition, des changements dans la prévalence des sérovars, des changements démographiques ou autres dans la population des patients, ou une combinaison de ces facteursNote de bas de page 14Note de bas de page 15.
Le taux de bactériémie pour les isolats de typhoïde était de 35,2 % (IC 95 % : 33,7 %–36,8 %), au cours de la période de référence de 14 ans. La bactériémie est souvent une conséquence de la fièvre typhoïde et le taux rapporté ici est conforme aux valeurs publiéesNote de bas de page 17. Le taux de bactériémie des isolats de la Salmonella non typhique était de 3,2 % (IC 95 % : 3,1 %–3,3 %). Ce taux était légèrement inférieur aux estimations des États-Unis (5 %)Note de bas de page 18Note de bas de page 19 et supérieur aux 2,1 % rapportés pour l’AngleterreNote de bas de page 14.
Ces différences pourraient être dues à des différences dans les périodes de signalement des trois études (États-Unis : 1996 à 2006, Angleterre : 2004 à 2015) et des différences dans la prévalence des sérovarsNote de bas de page 14Note de bas de page 19. Les cinq principaux sérovars signalés dans l’étude américaine étaient Typhimurium, Enteritidis, Newport, Heidelberg et JavianaNote de bas de page 19. En Angleterre, les cinq principaux sérovars étaient Enteritidis, Typhimurium, Virchow, Newport et InfantisNote de bas de page 14. On a constaté un chevauchement considérable des sérovars les plus fréquemment associés aux bactériémies au Canada, aux États-Unis et en AngleterreNote de bas de page 14Note de bas de page 19. Parmi les 19 sérovars identifiés comme ayant des taux de bactériémie élevés dans cette étude, huit ont également été identifiés comme ayant des taux de bactériémie élevés en Angleterre et aux États-Unis : Heidelberg, Oranienburg, Virchow, Schwarzengrund, Panama, Poona, Brandenburg et Dublin. Le Canada partageait quatre sérovars supplémentaires avec les États-Unis (Reading, Sandiego, Urbana et Choleraesuis) et deux avec l’Angleterre (Chester et Paratyphi B var. Java)Note de bas de page 14Note de bas de page 19.
Des variations géographiques des sérovars ont également été remarquées entre les trois études. Les sérovars Durban, Telelkebir, Lomalinda, Stanleyville et I 6,7:c:- ont tous été identifiés dans cette étude comme ayant des taux de bactériémie plus élevés. Collectivement, ces cinq sérovars sont rares et ont été à l’origine de 257 cas confirmés en laboratoire entre 2006 et 2019 (soit 0,3 % de l’ensemble des signalements de Salmonella non typhique). Ces sérovars n’étaient pas répertoriés dans les rapports anglais et américains, ce qui suggère que les expositions sont liées à des voyages ou sont uniques au Canada.
La poursuite de l’étude de la pathogenèse des sérovars de la Salmonella non typhique associés et non associés à la bactériémie permettra d’acquérir des connaissances sur les risques relatifs des sérovars de Salmonella. La compréhension de ces différences permettra d’élaborer des méthodes pour améliorer les soins aux patients grâce à l’identification et au traitement précoces des cas de salmonellose qui présentent un risque élevé de développer une bactériémie.
Forces et faiblesses
Les taux de bactériémies présentés ci-dessus sont des estimations du taux réel fondées sur les données soumises au Programme national de surveillance des maladies entériques. La soumission des données au Programme national de surveillance des maladies entériques est volontaire. Les valeurs indiquées dans ce rapport ne représentent que les isolats soumis par les laboratoires provinciaux et territoriaux et sous-estiment le nombre de cas de salmonellose au CanadaNote de bas de page 20.
Les données sur la source de l’isolat sont saisies par le Programme national de surveillance des maladies entériques sur la base des détails recueillis par les laboratoires soumissionnaires. Comme cette information n’est pas toujours disponible, les données sur les infections extra-intestinales à Salmonella peuvent également être sous-déclarées. Bien que tous les efforts soient faits pour éviter les doublons, les données sources limitées ne permettent pas d’identifier les isolations en double provenant de différentes sources et provenant du même patient. De plus, les données incluent les cas liés aux voyages, ce qui ne reflète pas la véritable représentation des souches endémiques au Canada.
La solidité d’études comme celle-ci repose sur la qualité et la quantité des données de surveillance disponibles. Un nombre plus faible d’isolats rapportés conduit à un plus grand degré d’imprécision. Les chiffres présentés ici sont probablement biaisés en faveur de la bactériémie, car les patients présentant des symptômes plus graves sont plus susceptibles de consulter un médecin que ceux qui présentent une forme autolimitée de la maladie. En l’absence de données cliniques, on suppose que les cas dans lesquels des isolats sanguins de Salmonella ont été récupérés étaient plus graves que ceux qui avaient des cultures de selles positives.
Conclusion
D’après les données soumises au Programme national de surveillance des maladies entériques, on estime que 4,4 % des cas de salmonellose survenus entre 2006 et 2019 ont entraîné une bactériémie. Des groupes et sérovars spécifiques de Salmonella associés à des taux de bactériémie plus ou moins élevés par rapport au groupe plus large de souches de Salmonella ont été identifiés. Les résultats de cette étude éclaireront les recherches futures visant à prédire et à améliorer l’issue des bactériémies à Salmonella au Canada.
Déclaration des auteurs
- S. T. — Conceptualisation, méthodologie, analyse formelle, rédaction–version originale, rédaction–révision et édition
- B. D. — Rédaction–révision et édition
- K. N. — Curation des données, rédaction–révision et édition
Intérêts concurrents
Aucun.
Remerciements
Les auteurs tiennent à remercier le personnel de l’Agence de la santé publique du Canada, du Programme des maladies entériques du Laboratoire national de microbiologie et du Centre des maladies infectieuses d’origine alimentaire, environnementale et zoonotique qui administre le Programme national de surveillance des maladies entériques. Les auteurs remercient également à A. Catford, E. Buenaventura, B. Dixon et J. Austin pour la révision critique du manuscrit.
Financement
Ce travail a été financé par Santé Canada (services votés) pour soutenir les programmes de sécurité alimentaire du Canada.
Références
- Référence 1
-
Jajere SM. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet World 2019;12(4):504-21. https://doi.org/10.14202/vetworld.2019.504-521
- Référence 2
-
Lamas A, Miranda JM, Regal P, Vázquez B, Franco CM, Cepeda A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol Res 2018;206:60-73. https://doi.org/10.1016/j.micres.2017.09.010
- Référence 3
-
Gal-Mor O, Boyle EC, Grassl GA. Same species, different diseases: how and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front Microbiol 2014;5:391. https://doi.org/10.3389/fmicb.2014.00391
- Référence 4
-
Thomas MK, Murray R, Flockhart L, Pintar K, Fazil A, Nesbitt A, Marshall B, Tataryn J, Pollari F. Estimates of foodborne illness-related hospitalizations and deaths in Canada for 30 specified pathogens and unspecified agents. Foodborne Pathog Dis 2015;12(10):820-7. https://doi.org/10.1089/fpd.2015.1966
- Référence 5
-
Ilyas B, Tsai CN, Coombes BK. Evolution of Salmonella-host cell interactions through a dynamic bacterial genome. Front Cell Infect Microbiol 2017;7:428. https://doi.org/10.3389/fcimb.2017.00428
- Référence 6
-
Agence de la santé publique du Canada. Programme national de surveillance des maladies entériques. 2018 (accédé 2020-12-04). https://www.canada.ca/fr/sante-publique/programmes/programme-national-surveillance-maladies-enteriques.html
- Référence 7
-
Cheng RA, Eade CR, Wiedmann M. Embracing diversity: differences in virulence mechanisms, disease severity, and host adaptations contribute to the success of nontyphoidal Salmonella as a foodborne pathogen. Front Microbiol 2019;10:1368. https://doi.org/10.3389/fmicb.2019.01368
- Référence 8
-
Gal-Mor O. Persistent infection and long-term carriage of typhoidal and nontyphoidal salmonellae. Clin Microbiol Rev 2018;32(1):e00088-18. https://doi.org/10.1128/CMR.00088-18
- Référence 9
-
Hohmann EL. Nontyphoidal salmonellosis. Clin Infect Dis 2001;32(2):263-9. https://doi.org/10.1086/318457
- Référence 10
-
Matheson N, Kingsley RA, Sturgess K, Aliyu SH, Wain J, Dougan G, Cooke FJ. Ten years experience of Salmonella infections in Cambridge, UK. J Infect 2010;60(1):21-5. https://doi.org/10.1016/j.jinf.2009.09.016
- Référence 11
-
Robinson JL. Salmonella infections in Canadian children. Paediatr Child Health 2019;24(1):50-1. https://doi.org/10.1093/pch/pxy199
- Référence 12
-
Galanakis E, Bitsori M, Maraki S, Giannakopoulou C, Samonis G, Tselentis Y. Invasive non-typhoidal salmonellosis in immunocompetent infants and children. Int J Infect Dis 2007;11(1):36-9. https://doi.org/10.1016/j.ijid.2005.09.004
- Référence 13
-
Hsu RB, Tsay YG, Chen RJ, Chu SH. Risk factors for primary bacteremia and endovascular infection in patients without acquired immunodeficiency syndrome who have nontyphoid salmonellosis. Clin Infect Dis 2003;36(7):829-34. https://doi.org/10.1086/367932
- Référence 14
-
Katiyo S, Muller-Pebody B, Minaji M, Powell D, Johnson AP, De Pinna E, Day M, Harris R, Godbole G. Epidemiology and outcomes of nontyphoidal Salmonella bacteremias from England, 2004 to 2015. J Clin Microbiol 2019;57(1):e01189-18. https://doi.org/10.1128/JCM.01189-18
- Référence 15
-
GBD 2017 Non-Typhoidal Salmonella Invasive Disease Collaborators. The global burden of non-typhoidal salmonella invasive disease: a systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect Dis 2019;19(12):1312-24. https://doi.org/10.1016/S1473-3099(19)30418-9
- Référence 16
-
Mandal BK, Brennand J. Bacteraemia in salmonellosis: a 15 year retrospective study from a regional infectious diseases unit. BMJ 1988;297(6658):1242-3. https://doi.org/10.1136/bmj.297.6658.1242
- Référence 17
-
Wain J, Diep TS, Ho VA, Walsh AM, Nguyen TT, Parry CM, White NJ. Quantitation of bacteria in blood of typhoid fever patients and relationship between counts and clinical features, transmissibility, and antibiotic resistance. J Clin Microbiol 1998;36(6):1683-7. https://doi.org/10.1128/JCM.36.6.1683-1687.1998
- Référence 18
-
Arshad MM, Wilkins MJ, Downes FP, Rahbar MH, Erskine RJ, Boulton ML, Younus M, Saeed AM. Epidemiologic attributes of invasive non-typhoidal Salmonella infections in Michigan, 1995--2001. Int J Infect Dis 2008;12(2):176-82. https://doi.org/10.1016/j.ijid.2007.06.006
- Référence 19
-
Jones TF, Ingram LA, Cieslak PR, Vugia DJ, Tobin-D'Angelo M, Hurd S, Medus C, Cronquist A, Angulo FJ. Salmonellosis outcomes differ substantially by serotype. J Infect Dis 2008;198(1):109-14. https://doi.org/10.1086/588823
- Référence 20
-
Thomas MK, Murray R, Flockhart L, Pintar K, Pollari F, Fazil A, Nesbitt A, Marshall B. Estimates of the burden of foodborne illness in Canada for 30 specified pathogens and unspecified agents, circa 2006. Foodborne Pathog Dis 2013;10(7):639-48. https://doi.org/10.1089/fpd.2012.1389

Cette œuvre est mise à disposition selon les termes de la Licence Creative Commons Attribution 4.0 International