
Évaluation et Rapport de situation du COSEPAC sur Bourdon de l'Ouest Bombus occidentalis au Canada - 2014
- Description et importance de l’espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d’information
- Sommaire biographique des rédacteurs du rapport
- Collections examinées
- Figure 1. Femelle (ouvrière) de la sous-espèce occidentalis du bourdon de l’Ouest (soulignons qu’il en existe plus d’une forme et que la photo présente la forme habituelle). Photo prise par Sheila Colla. Spécimen conservé dans la collection d’abeilles Packer de l’Université York, à Toronto.
- Figure 2. Mâle de la sous-espèce occidentalis du bourdon de l’Ouest (soulignons qu’il en existe plus d’une forme et que la photo présente la forme habituelle). Photo prise par Sheila Colla. Spécimen conservé dans la collection d’abeilles Packer de l’Université York, à Toronto.
- Figure 3. Femelle (ouvrière) de la sous-espèce mckayi du bourdon de l’Ouest (Bombus occidentalis mckayi). Photo prise par Cory Sheffield. Spécimen conservé au Royal Saskatchewan Museum, à Regina, en Saskatchewan.
- Figure 4. Carte de l’aire de répartition mondiale de Bombus occidentalis montrant la répartition des deux sous-espèces, B. o. occidentalis (sous le trait) et B. o. mckayi (au-dessus du trait). À remarquer que la limite sud de la répartition de B. o. mckayi et la limite nord de celle de B. o. occidentalis ne sont pas bien définies. Carte créée à partir de données de Sheffield et al. (2013 et en prép.).
- Figure 5. Lieux de prélèvement de bourdons (236 260 points au total) en Amérique du Nord entre 1892 et 2012. Points rouges = bourdon de l’Ouest (Bombus occidentalis (12 898 mentions), les deux sous-espèces); points bleus = espèce étroitement apparentée du bourdon terricole (B. terricola) (13 857 mentions) y compris des régions de chevauchement (voir sous « Description et importance de l’espèce sauvage »). Il est à noter que le statut taxinomique de B. occidentalis et de B. terricola a posé des problèmes, et qu’il n’est pas garanti que tous les spécimens cartographiés soient correctement identifiés. On pense que l’unique signalement d’un spécimen de B. occidentalis dans l’est constitue une erreur. Ces cartes situent l’aire de répartition de façon générale, et il convient d’analyser les observations aberrantes. On ne dispose d’aucune donnée pour les emplacements où il n’y a pas de points. Plus de 70 personnes et établissements ont contribué à constituer l’ensemble de données. Ils sont nommés sur le site www.leifrichardson.org/bbna.html. Carte établie par Leif Richardson. Spécimens réunis dans un ensemble de données pour Williams et al. (2014).
- Figure 6. Répartition spatiale des données d’échantillonnage du bourdon de l’Ouest au Canada de 1882 à 2010. Chaque cercle est proportionnel au nombre d’observations consignées à cet endroit entre 1882 et 2010, inclusivement (y compris les échantillons non indépendants et les sites où plusieurs observations ont été consignées au cours d’une année). Les données ont été cartographiées à l’aide de carrés représentant 50 km sur 50 km. Les carrés gris indiquent les mentions antérieures à 1996 (128 carrés); les carrés jaunes montrent les sites où des spécimens ont été recueillis avant et après 1996 (27 carrés); les carrés verts indiquent les prélèvements effectués après 1996 (13 carrés) (n = 1 706 spécimens). Les données ont été compilées à partie des informations fournies par le Système canadien d’information sur la biodiversité et les collaborateurs indiqués dans les « Remerciements ». Il est impossible de mettre cette carte à jour.
- Figure 7. Abondance relative (AR) du bourdon de l’Ouest dans le sud de l’Alberta au cours deux intervalles de temps (la taille combinée des échantillons est indiquée entre parenthèses). Toutes les données provenant du sud de l’Alberta sont indiquées (c.-à-d. les relevés de 2000 et de 2010, ainsi que les autres sites d’études où des échantillons ont été prélevés à d’autres moments). À l’exception de deux sites où le bourdon de l’Ouest n’a pas été repéré à aucune des deux périodes (Innisfail et route Trunk), on a observé un déclin de l’AR entre 2000 et 2010. Les habitats où des spécimens de l’espèce ont été recueillis sont Calgary (zone urbaine), Clarsehom (zone rurale), lac Barrier (zone naturelle), Drumheller (limite de l’aire de répartition de l’espèce), mont Fortress (altitude la plus élevée), Coleman (zone rurale) et Innisfail (à l’extérieur de l’aire de répartition de la sous-espèce) et route Trunk (à l’extérieur de l’aire de répartition de la sous-espèce).
- Figure 8. Probabilité de prélèvement prévue du bourdon de l’Ouest, selon les données obtenues de 14 sites du sud de l’Alberta échantillonnés entre 1985 et 2010. Les points indiquent les probabilités moyennes (avec un intervalle de confiance de 95 %) des sites échantillonnés au cours d’une année donnée. La ligne pointillée indique un ajustement de la régression quadratique; entre 2010 et 2018, la ligne représente les valeurs projetées de l’équation de régression.
- Figure 9. Abondance relative (AR) du bourdon de l’Ouest selon toutes les mentions de Bombus au Canada (de 1882 à 2011) versées dans des bases de données. L’axe des Y de gauche (partie ombragée de la barre) montre le nombre de spécimens du bourdon de l’Ouest et l’axe des Y de droite (triangles) représente la proportion des spécimens de bourdon de l’Ouest par intervalles de 10 ans. On a utilisé une régression linéaire pour examiner les tendances de l’AR au cours des intervalles de 10 ans : la ligne représente le meilleur ajustement des données. Voir également le tableau 3. Les graphiques ont été générés à l’aide du logiciel Minitab ®.
- Figure 10. Nombre total de spécimens de bourdons prélevés au Canada (de 1882 à 2011) dans chaque province et territoire; les triangles représentent le nombre de spécimens de bourdon de l’Ouest. Les valeurs indiquées au-dessus de chaque barre représentent le pourcentage de bourdons prélevés par rapport à l’ensemble de la collecte. Voir également le tableau 3. Les graphiques ont été générés à l’aide du logiciel Minitab ®.
- Tableau 1. Résumé des chiffres de superficie et d’altitude de la répartition du bourdon de l’Ouest recueillis pour quatre intervalles d’échantillonnage s’échelonnant de 1882 à 2010. Voir également la figure 6.
- Tableau 2. Relevés récents (depuis 2002) ciblant les bourdons dans l’aire de répartition du bourdon de l’Ouest.
- Tableau 3. Abondance relative (AR) du bourdon de l’Ouest comparativement aux données de prélèvement de Bombus saisies (de 1882 à 2011) au Canada. Plus de 70 personnes et institutions ont contribué à l’élaboration de l’ensemble de données. Les données des spécimens ont été réunies en un ensemble de données pour Williams et al., 2014. L’AR du bourdon de l’Ouest est indiquée en intervalles de 10 ans (voir la représentation graphique à la figure 9 et à la figure 10). Les spécimens observés au Manitoba ne sont pas considérés comme des populations naturelles. La plupart des bourdons signalés en C.-B. proviennent du tiers méridional de la province et sont considérés comme appartenant à la sous-espèce occidentalis.
- Tableau 4. Tableau de classification des menaces pesant sur la sous-espèce occidentalis du bourdon de l’Ouest (Bombus occidentalis occidentalis) dans l’ensemble de son aire de répartition au Canada. Tableau fondé sur le système unifié de classification des menaces proposé par l’IUCN–CMP (Partenariat pour les mesures de conservation de l’Union internationale pour la conservation de la nature). Pour une description détaillée du système de classification des menaces, consulter le site Web du Partenariat pour les mesures de conservation (CMP, 2006). Pour de plus amples renseignements sur la façon dont les valeurs sont attribuées, voir Master et al. (2009). Le calculateur de menaces a été rempli par J. Heron et C. Sheffield avec la collaboration de D. Fraser, S. Colla et L. Richardson.

Les rapports de situation du Comité sur la situation des espèces en péril au Canada (COSEPAC) sont des documents de travail servant à déterminer le statut des espèces sauvages que l’on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2014. Évaluation et Rapport de situation du COSEPAC sur le bourdon de l’Ouest (Bombus occidentalis) de la sous-espèce occidentalis (Bombus occidentalis occidentalis) et la sous-espèce mckayi (Bombus occidentalis mckayi) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xii + 58 p. (Registre public des espèces en péril site Web).
Le COSEPAC remercie Sheila Colla, Cory Sheffield et Leif Richardson d’avoir rédigé le rapport de situation sur le bourdon de l’Ouest (Bombus occidentalis) au Canada, en vertu d’un contrat avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Jennifer Heron, coprésidente du Sous-comité de spécialistes des arthropodes.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-953-3215
Téléc. : 819-994-3684
COSEPAC Courriel
COSEPAC site Web
Also available in English under the title COSEWIC Assessment and Status Report on the Western Bumble Bee Bombus occidentalis, Bombus occidentalis, occidentalis subspecies (Bombus occidentalis occidentalis) and the mckayi subspecies (Bombus occidentalis mckayi) in Canada.
Illustration/photo de la couverture :
Bourdon de l'Ouest -- Photographie de la page couverture : David Inouye. Ouvrière de bourdon de l’Ouest en train de butiner le nectar d’une fleur d’Ipomopsis.
© Sa Majesté la Reine du chef du Canada, 2014.
No de catalogue CW69-14/694-2014F-PDF
ISBN 978-0-660-22200-4

Sommaire de l’évaluation - mai 2014
Sommaire de l’évaluation - mai 2014
Le bourdon de l’Ouest (Western Bumble Bee) (Bombus occidentalis Greene) est une des cinq espèces nord-américaines classées dans le sous-genre Bombus sensu stricto. De taille moyenne (entre 1 et 2 cm), il a une tête courte. La coloration du pelage de l’abdomen varie, mais il a toujours une bande transversale jaune sur le dessus du thorax devant la base des ailes, et l’extrémité de l’abdomen est presque toujours blanche.
La taxinomie des bourdons fait l’objet d’un vaste débat, y compris l’historique taxinomique du bourdon de l’Ouest. L’espèce a déjà été placée en synonymie du bourdon terricole; toutefois, de récents travaux de génétique ont confirmé qu’il s’agissait de deux espèces distinctes. Des travaux de taxinomie récents divisent l’espèce en deux sous-espèces, Bombus occidentalis occidentalis et Bombus occidentalis mckayi, d’après les informations génétiques et morphologiques et les données sur la répartition.
Le bourdon de l’Ouest se répartit dans presque tout l’ouest des États-Unis. La sous-espèce occidentalis est présente, en direction nord, du centre de la Californie au nord de la Colombie-Britannique et, vers l’est, jusque dans le sud de la Saskatchewan et dans le Dakota du Sud. La sous-espèce mckayi est présente, vers le nord, du centre-nord de la Colombie-Britannique au Yukon, aux Territoires du Nord-Ouest et à l’Alaska.
Le bourdon de l’Ouest vit dans des milieux variés, notamment les boisés mixtes, les terres agricoles, les zones urbaines, les prairies montagnardes et la limite ouest des prairies. La sous-espèce mckayi semble se cantonner à la Cordillère et à l’aire écologique boréale. Le bourdon de l’Ouest exploite de nombreux genres de plantes comme sources de pollen et de nectar. Comme de nombreux bourdons, Il fait habituellement son nid dans des terriers abandonnés de rongeurs ou dans des cavités du bois en décomposition.
Le bourdon de l’Ouest a un cycle vital annuel. Les reines fécondées émergent de leur cachette hivernale au printemps et partent à la recherche d’un site de nidification où fonder une colonie. Une fois qu’elle a trouvé le nid, la reine va chercher du pollen et du nectar pour s’alimenter, puis elle y retourne pondre ses œufs, qui produiront des ouvrières. Celles-ci se chargent de l’entretien de la colonie et de son approvisionnement en nourriture. À la fin de l’été, la reine pond des mâles et des femelles fertiles. Ces reproducteurs quittent la colonie et s’accouplent, et seules les reines fécondées hibernent. Le reste de la colonie, y compris la vieille reine, meurt à l’arrivée du froid.
La sous-espèce occidentalis continue d’habiter l’essentiel de son aire historique au Canada, mais à moins d’endroits et dans une moindre abondance : les données d’abondance relative des dix dernières années pointent vers une baisse probable des effectifs de plus de 30 %. Là où cette sous-espèce a été le plus étudiée au pays (soit dans le sud de la Colombie-Britannique et de l’Alberta), l’abondance relative a décliné considérablement à tous les sites recensés depuis une trentaine d’années. On observe plus souvent la sous-espèce mckayi, dont l’abondance semble constante, même s’il existe peu de données historiques à partir desquelles dégager des tendances.
Parmi les menaces qui pourraient peser sur la sous-espèce occidentalis, il y a la transmission d’agents pathogènes par des bourdons échappés d’élevages qui sont utilisés pour la pollinisation des cultures sous serre. L’emploi de produits chimiques et de pesticides agricoles et la perte d’habitat constituent d’autres facteurs de menace régionaux.
À l’heure actuelle, les deux sous-espèces du bourdon de l’Ouest ne bénéficient d’aucune protection juridique au Canada. Toutes les espèces du sous-genre Bombus sensu stricto semblent décliner à l’échelle mondiale.
Bombus occidentalis occidentalis
Bourdon de l'Ouest de la sous-espèce occidentalis
Western Bumble Bee occidentalis subspecies
Données démographiques
Information sur la répartition
| Population | Nombre d’individus matures |
|---|---|
| sous-espèce occidentalis | Inconnu. |
| Total | Inconnu. |
Analyse quantitative
Menaces (réelles ou imminentes pour les populations ou leur habitat)
La sous espèce occidentalis présente l’une des charges les plus lourdes de parasites (notamment de la microsporidie Nosema bombi) parmi les espèces de bourdons de l’Amérique du Nord. Les menaces actuelles pesant sur l’espèce, en particulier dans les portions sud de son aire de répartition, incluent la propagation de pathogènes et la transmission de maladies provenant de colonies de bourdons exotiques et de colonies de bourdons gérées à des fins commerciales pour la pollinisation des cultures en serre, l’utilisation de pesticides (dont les néonicotinoïdes), l’agriculture et d’autres pratiques d’utilisation des terres de plus en plus intensives et les modifications de l’habitat.
Immigration de source externe (immigration de l’extérieur du Canada)
Nature délicate de l’information sur l’espèce
Historique du statut
COSEPAC : Espèce désignée « menacée » en mai 2014.
Statut et justification de la désignation
Applicabilité des critères
Bombus occidentalis mckayi
Bourdon de l'Ouest de la sous-espèce mckayi
Western Bumble Bee mckayi subspecies
Données démographiques
Information sur la répartition
| Population | Nombre d’individus matures |
|---|---|
| sous-espèce mckayi | Inconnu. |
| Total | Inconnu. |
Analyse quantitative
Menaces (réelles ou imminentes pour les populations ou leur habitat)
Selon de récentes études menées en Alaska, cette sous-espèce présente l’une des charges les plus lourdes de parasites (notamment de la microsporidie Nosema bombi) parmi les espèces de bourdons de l’Amérique du Nord. Parmi les autres menaces possibles, il y a la transmission non connue de maladies par des bourdons exotiques introduits comme agents de pollinisation dans les serres (en cours au Yukon), l’emploi de pesticides (dont les néonicotinoïdes) et les modifications de l’habitat.
Effet d'une immigration de source externe (immigration de l’extérieur du Canada)
Historique du statut
COSEPAC : Espèce désignée « préoccupante » en mai 2014.
Statut et justification de la désignation
Applicabilité des critères

Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsable des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d’animal, de plante ou d’un autre organisme d’origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s’est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
Le Service canadien de la faune d’Environnement Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Le genre Bombus comprend environ 250 espèces réparties principalement dans les régions tempérées de l’Amérique du Nord, de l’Amérique centrale, de l’Amérique du Sud, de l’Europe et de l’Asie. Le bourdon de l’Ouest (Bombus occidentalis) appartient au sous-genre Bombus sensu stricto, un des 15 sous-genres de bourdons reconnus dans le monde. En Amérique du Nord, le sous-genre Bombus s. str. réunit quatre autres espèces : Bombus affinis Cresson, B. cryptarum (Fabricius), B. franklini Frison et B. terricola Kirby.
On identifie les bourdons principalement à leur coloration à l’âge adulte, quoique celle-ci varie chez de nombreuses espèces. Cette variation explique en partie les difficultés taxinomiques passées et récentes soulevées par de nombreuses espèces, y compris le bourdon de l’Ouest.
Bombus occidentalis a été décrit comme espèce distincte par Greene (1858), puis reconnu comme appartenant à la même espèce que B. terricola par certains auteurs (voir par exemple Milliron, 1971; Cameron et al., 2007 [mais Milliron en faisait une sous-espèce]) et encore comme espèce distincte par d’autres (voir par exemple Stephen, 1957; Thorp et al., 1983).
Cameron et al. (2007) ont récemment comparé des séquences d’ADN du gène mitochondrial 16S et conclu que les deux taxons (B. occidentalis et B. terricola) sont conspécifiques. Toutefois, Williams et al. (2012) ont indiqué que les séquences de la sous-unité I de la cytochrome c oxydase (COI) de l’ADN mitochondrial (code à barres de l’ADN) différaient suffisamment pour qu’on considère B. occidentalis comme une espèce distincte. Ces résultats confortent ceux de Bertsch et al. (2010), une divergence globale des séquences de COI d’environ 5 % étant constatée entre les deux bourdons. De plus, Owen et Whidden (2013) ont trouvé des caractères morphologiques et moléculaires à l’appui de la distinction de deux taxons. Par conséquent, B. occidentalis est considéré comme une espèce distincte de B. terricola.
Williams et al. (2012) ont aussi découvert des divergences dans les séquences de COI d’échantillons du bourdon de l’Ouest qui étaient corrélées avec la géographie, et ils ont reconnu deux sous-espèces : B. o. occidentalis et B. o. mckayi. Pour obtenir plus de renseignements au sujet de la phylogénie moléculaire, voir Williams et al. (2012), qui ont vérifié la division du bourdon de l’Ouest en deux génotypes distincts. Des travaux en cours fondés sur des données morphologiques et moléculaires et des données sur la répartition appuient cette conclusion (Sheffield et al., 2013). C’est pourquoi B. occidentalis est présenté dans ce rapport de situation comme deux sous-espèces : la sous-espèce occidentalis et la sous-espèce mckayi.
Les bourdons sont des insectes holométaboles. Ils ont quatre stades de développement (œuf, larve, nymphe et adulte) et forment des colonies primitivement eusociales dont les adultes se divisent en trois castes : la reine (femelle reproductrice), les ouvrières (stériles) et les mâles. Le bourdon de l’Ouest adulte a une coloration qui varie beaucoup, surtout sur le scutellum (pièce dorsale dure du thorax qui se situe habituellement entre les points d’insertion des ailes) et sur les deuxième et troisième segments abdominaux; elle va du noir au jaune (voir les précisions plus loin).
Femelles : les reines et les ouvrières présentent une coloration similaire, mais leur taille diffère (reine : entre 1,6 et 1,9 cm de longueur; ouvrière : entre 1,1 et 1,3 cm de longueur). La tête est entièrement noire (figure 1), et l’espace malaire (entre le bord inférieur de l’œil composé et la base de la mandibule) est court (à peu près aussi long que large). Toutes les femelles ont une bande transversale de poils jaunes devant la base des ailes. La coloration de l’abdomen varie, parfois noire sur les quatre premiers segments (figure 1) et parfois avec du jaune sur le troisième et occasionnellement aussi sur une partie du deuxième segment. La plupart des femelles ont du poil blanc (moins souvent jaunâtre) sur le tergum apical (l’extrémité de l’abdomen). La coloration varie davantage chez la sous-espèce occidentalis. Chez la sous-espèce mckayi, le poil est constamment jaune sur le troisième segment abdominal (figure 3).

Photo : S. Colla © Environnement Canada
Description longue pour la figure 1
Photo d’un spécimen d’ouvrière de bourdon de l’Ouest, de la sous espèce occidentalis. Les reines et les ouvrières de l’espèce présentent une coloration similaire. La tête est entièrement noire, et l’espace malaire est court (à peu près aussi long que large). Une bande transversale de poils jaunes est présente devant la base des ailes, et les quatre premiers segments de l’abdomen sont noirs. La plupart des femelles ont du poil blanc (moins souvent jaunâtre) sur l’extrémité de l’abdomen.

Photo : S. Colla © Environnement Canada
Description longue pour la figure 2
Photo d’un spécimen de bourdon de l’Ouest mâle, de la sous espèce occidentalis. Les mâles ont une apparence semblable à celle des femelles (voir le texte du rapport), avec une coloration variable. Le corps mesure de 1 à 2 centimètres de long. Chez le mâle, la pilosité de la face est constituée de poils jaune pâle entremêlés de poils noirs. L’espace malaire est court.

Photo : S. Colla © Environnement Canada
Description longue pour la figure 3
Photo d’un spécimen d’ouvrière de bourdon de l’Ouest, de la sous espèce mckayi. Les ouvrières et les reines présentent une coloration similaire : la tête est entièrement noire, et l’espace malaire est court (à peu près aussi long que large). Une bande transversale de poils jaunes est présente devant la base des ailes, et les quatre premiers segments de l’abdomen sont noirs. La plupart des individus ont du poil blanc (moins souvent jaunâtre) sur l’extrémité de l’abdomen. Chez la sous-espèce mckayi, le poil est toujours jaune sur le troisième segment abdominal.
Koch et al. (2012) et Williams et al. (2014) recensent les variantes de couleur chez le bourdon de l’Ouest adulte. Il est rare d’observer ou d’identifier les bourdons, comme la plupart des abeilles, aux stades immatures (œuf, larve ou nymphe). Ces stades restent pratiquement inaperçus, et on n’a pas étudié la biologie de nidification ni les stades immatures de presque toutes les espèces d’abeilles. Stephen et al. (1969) et Michener (2007) présentent des descriptions sommaires des stades vitaux des abeilles. Stephen et Koontz (1973a, 1973b) traitent plus précisément des stades larvaires des bourdons. Pour ce qui est du bourdon de l’Ouest, les stades immatures ne sont pas décrits dans la littérature, non plus que la plupart des caractères distinctifs des espèces du genre.
Cameron et al. (2011) ont récemment étudié la diversité génétique et la stabilité des populations américaines du bourdon de l’Ouest, précisément de la sous-espèce occidentalis, à l’aide de 8 à 11 locus microsatellites. D’après leurs résultats, cette sous-espèce montrerait une faible diversité génétique parmi les sous-populations (n = 93, locus = 8, HE totale = 0,584), et serait susceptible de plus grandes baisses d’effectifs que d’autres bourdons étudiés en raison du risque accru de consanguinité et de la dérive génétique dans les populations ayant un petit effectif efficace.
Le séquençage des codes à barres ADN (c.-à-d. de la COI) de nombreux spécimens du bourdon de l’Ouest, provenant de l’ensemble de l’aire de répartition naturelle de l’espèce, a été fait, et les résultats ont été versés dans la plateforme de Barcode of Life Data Systems (BOLD) (voir Williams et al., 2012). Les séquences ont servi à appuyer la classification des deux sous-espèces : B. o. mckayi et B. o. occidentalis (Williams et al., 2012). Sheffield et al. (2013) ont présenté d’autres données morphologiques et biogéographiques à l’appui de la distinction des deux sous-espèces du bourdon de l’Ouest reconnues par Williams et al. (2012); les auteurs préparent les données pour publication (Sheffield et al., en prép.).
Il est proposé de définir deux unités désignables selon les sous-espèces reconnues par Williams et al. (2012). Au Canada, la sous-espèce occidentalis est présente dans les aires écologiques nationales suivantes du COSEPAC : Pacifique, Montagnes du Sud, Prairie et l’extrémité sud de l’aire des Montagnes du Nord (COSEPAC, 2010). La sous-espèce mckayi est présente dans l’aire écologique nationale des Montagnes du Nord, mais il se peut qu’on la trouve jusque dans l’extrémité nord-ouest de l’aire écologique nationale boréale. Les deux sous-espèces semblent avoir une division sur les plans morphologique, moléculaire et écologique entre le 55° et le 60° de latitude nord (Sheffield, données pers.) (voir sous « Répartion »), ce qui plaide en faveur de l’établissement de deux unités désignables.
Avant son déclin, le bourdon de l’Ouest était parmi les bourdons les plus couramment observés dans l’ouest du Canada (Hobbs, 1968; Richards, 1978). Comme toutes les abeilles, le bourdon de l’Ouest est important sur le plan écologique, car il pollinise diverses plantes indigènes dans son aire de répartition (voir Ascher et Pickering, 2013, pour obtenir une liste des plantes). Les bourdons sont actifs pendant toute la période de végétation et volent quand le mauvais temps en empêche la plupart des autres insectes (Heinrich, 2004). Pollinisateurs, il facilitent la reproduction des plantes, aidant donc à abriter et à nourrir d’autres animaux et contribuant à la pérennité des écosystèmes indigènes (Goulson, 2010; Heinrich, 2004). Goulson (2010) et Heinrich (2004) donnent d’amples renseignements généraux sur les biens et services écosystémiques fournis par les bourdons.
Des colonies de bourdons d’élevage sont expédiées dans le monde entier pour être utilisées dans les serres afin de polliniser les cultures de concombres, de poivrons et de tomates, par exemple (Patten et al., 1993; Macfarlane et Patten, 1997). Ces plantes sont cultivées et fleurissent toute l’année. Elles exigent donc une pollinisation continue. Le bourdon de l’Ouest a déjà été élevé pour la pollinisation sous serre en Amérique du Nord, avant que l’élevage de l’espèce en captivité ne soulève des problèmes au début des années 2000 (voir par exemple Whittington et Winston, 2003). Jusqu’à ce moment-là, les producteurs commerciaux de bourdons (Koppert® et BioBest®) élevaient le bourdon fébrile (B. impatiens) comme pollinisateur dans l’est de l’Amérique du Nord et le bourdon de l’Ouest pour l’ouest. Maintenant, seul le bourdon fébrile est vendu pour la pollinisation sous serre en Amérique du Nord, et aucune restriction n’empêche de l’importer à cette fin dans l’ensemble du Canada; on l’utilise en Colombie-Britannique, où il a établi des populations férales (Ratti et Colla, 2010). On soupçonne que le bourdon fébrile est expédié dans les Territoires du Nord-Ouest pour usage dans des serres privées (Carriere, comm. pers., 2014). Certains États américains, comme l’Oregon, ont interdit l’importation d’espèces de Bombus hors de leurs aires de répartition (dont le bourdon fébrile).
Les abeilles fournissent des services de pollinisation pour la production de fruits sauvages et la pérennité des écosystèmes naturels. Ascher et Pickering (2013) donnent une liste partielle des genres et espèces de plantes que butine le bourdon de l’Ouest. Par ailleurs, les abeilles revêtent une grande importance culturelle pour les groupes autochtones. Parmi les plantes que butine le bourdon de l’Ouest qui sont importantes pour les Premières Nations, il y a des espèces de la famille des Astéracées (p. ex. du genre Helianthus), de la famille des Caprifoliacées, de la famille des Éricacées (p. ex. du genre Vaccinium), de la famille des Rosacées (p. ex. des genres Rosa et Rubus), et bien d’autres (Turner, 1975).
En Amérique du Nord,le sous-genre Bombus sensu stricto est représenté par cinq espèces (un peu plus de 10 % des 46 espèces; Williams et al., 2014). Pour des raisons qui demeurent obscures (bien qu’il y ait d’importants chevauchements de zones de grande urbanisation et d’agriculture), le sous-genre présente le plus grand risque inhérent de vulnérabilité de tous les sous-genres du continent – quatre espèces sont préoccupantes. Deux des espèces sont jugées en péril : le bourdon à taches rousses (Bombus affinis Cresson) a été classifié comme étant menacé par le COSEPAC (2010), et le bourdon de Franklin (B. franklini Frison; trouvé aux États-Unis seulement) pourrait avoir disparu, car on ne l’a pas vu depuis 2008. Le COSEPAC est en train d’établir un rapport au sujet du bourdon terricole (B. terricola Kirby). En outre, le bourdon de l’Ouest est l’hôte de deux bourdons parasites ou bourdons coucous : le B. bohemicus Cresson et le B. suckleyi Greene, dont le premier semble avoir subi d’importants déclins.
Le bourdon de l’Ouest est présent dans l’ouest de l’Amérique du Nord (figures 4 et 5). La sous-espèce occidentalis se répartit, vers le nord, du centre de la Californie au centre de la Colombie-Britannique, vers l’est, en Alberta et dans le sud de la Saskatchewan, et, vers le sud, dans le Dakota du Nord, le Dakota du Sud, l’Idaho, le Montana, le Wyoming, l’Utah, le Colorado, le Nouveau-Mexique, le nord de l’Arizona et le Nevada. La sous-espèce mckayi se répartit, vers le nord, du nord de la Colombie-Britannique au Yukon à l’ouest des Territoires du Nord-Ouest et en Alaska. L’espèce a été signalée à des altitudes variant du niveau de la mer à 1 350 mètres, mais l’altitude des sites de prélèvement change avec la latitude : l’espèce loge en hauteur dans les parties sud de son aire mondiale.
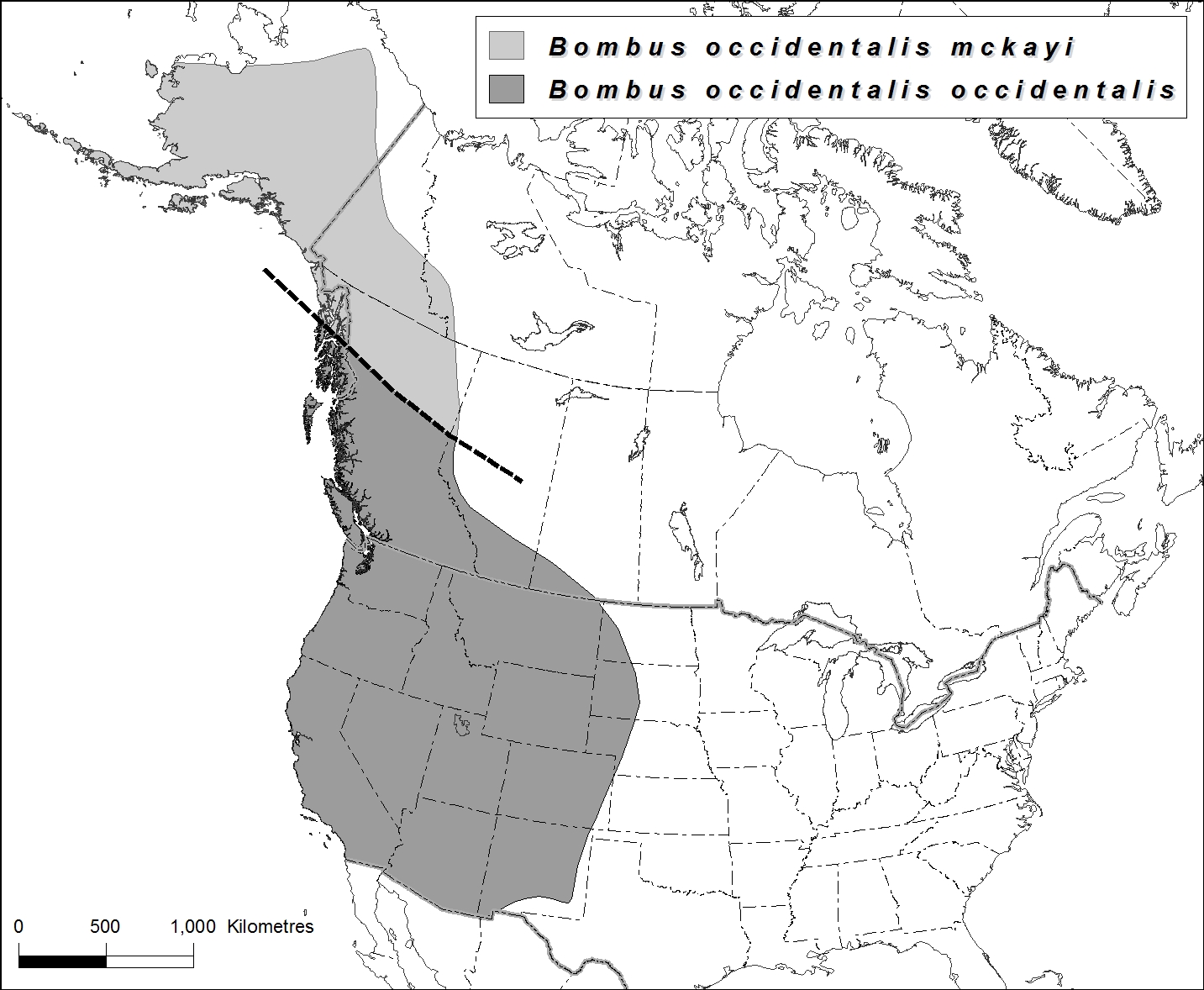
Image : © Environnement Canada
Description longue pour la figure 4
Carte de l’Amérique du Nord montrant l’aire de répartition des sous espèces occidentalis et mckayi du bourdon de l’Ouest. La sous-espèce occidentalis est présente, en direction nord, du centre de la Californie au centre de la Colombie-Britannique, jusque dans l’est de l’Alberta et dans le sud de la Saskatchewan et, vers le sud, dans le Dakota du Nord, le Dakota du Sud, l’Idaho, le Montana, le Wyoming, l’Utah, le Colorado, le Nouveau-Mexique, le nord de l’Arizona et le Nevada. La répartition de la sous espèce mckayi s’étend du nord de la Colombie Britannique à l’Alaska, en passant par le Yukon et l’ouest des Territoires du Nord Ouest.
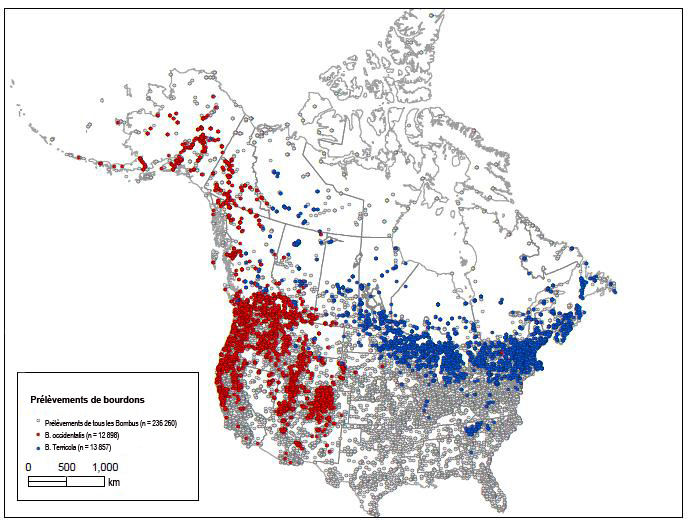
Image : © Environnement Canada
Description longue pour la figure 5
Carte de l’Amérique du Nord montrant l’aire de répartition (ensemble de points) des sous espèces occidentalis et mckayi du bourdon de l’Ouest, et du proche parent terricola. La sous-espèce occidentalis est présente dans l’ouest du Canada, dans tout le sud de la Colombie-Britannique jusqu’au 55° de latitude nord environ et, vers l’est, dans le sud de l’Alberta et de la Saskatchewan. La sous-espèce mckayi est présente en Colombie-Britannique au nord du 55° de latitude nord environ, au Yukon et vraisemblablement dans l’ouest des Territoires du Nord-Ouest.
La sous-espèce occidentalis est présente dans l’ouest du Canada, dans tout le sud de la Colombie-Britannique jusqu’au 55° de latitude nord environ et, vers l’est, dans le sud de l’Alberta et de la Saskatchewan. La sous-espèce mckayi est présente en Colombie-Britannique au nord du 55° de latitude nord environ, au Yukon et vraisemblablement dans l’ouest des Territoires du Nord-Ouest (Sheffield et al., en prép.) (figures 4 et 5).
La superficie de la zone d’occurrence des deux sous-espèces du bourdon de l’Ouest est approximative : de 720 170 km2 pour la sous-espèce occidentalis et de 623 837 km2 pour la sous-espèce mckayi. De légères incertitudes entourent la délimitation de l’interface des aires de répartition des deux sous-espèces en raison d’un échantillonnage insuffisant dans ce secteur crucial (Sheffield, comm. pers., 2013).
Il est impossible de calculer un indice de la zone d’occupation (IZO). La taille des populations et la dispersion des mentions sur un vaste territoire portent à établir la valeur de l’IZO pour les deux sous-espèces à plus de 2 000 km2.
On dispose de spécimens du bourdon de l’Ouest dans les musées et autres collections qui datent d’entre 1882 et 2013. Les prélèvements en Amérique du Nord ont été représentés sur une carte. Ils illustrent les activités de recherche visant directement ou indirectement le bourdon de l’Ouest (figure 5, 236 260 points au total; tous les Bombus). Les relevés n’ont pas toujours été systématiques ou complets sur l’ensemble de l’aire de répartition du bourdon de l’Ouest, mais on présume que si l’espèce avait été présente, des spécimens auraient été recueillis durant les campagnes de prélèvement. Il y a de grandes régions et de longues périodes de temps pour lesquelles on a peu d’informations.
Depuis une dizaine d’années, les activités de recherche axées sur les bourdons (collectivement) dans certaines parties de l’Amérique du Nord ont été considérables. Celles menées dans l’aire du bourdon de l’Ouest sont résumées en partie par province ci-dessous et énumérées au tableau 2.
Dans l’aire de répartition de la sous-espèce occidentalis, les relevés entre 2009 et 2013 ont recensé des spécimens dans le sud de l’île de Vancouver, les basses-terres continentales, la vallée du bas Fraser, la région de l’Okanagan et le Kootenay (tableau 2). En 2010, des relevés de grande envergure n’ont livré que 17 observations (> 575 heures, > 115 sites et > 800 km) (tableau 2).
Au cours de la dernière décennie, il y a eu très peu de relevés ou de prélèvements de bourdons dans le centre et le nord de la Colombie-Britannique. En 2013, des relevés (au moins 281 heures cumulées sur environ 104 sites; d’autres sites et échantillons sont encore en traitement) ont été réalisés en Colombie-Britannique (Sheffield et al., 2013; données utilisées dans un manuscrit en prép.). Ces relevés ont produit au moins 6 447 spécimens de Bombus (d’autres échantillons sont encore en traitement), dont 115 (soit 1,7 % de tous les spécimens examinés) étaient du bourdon de l’Ouest (Sheffield et al., en prép.), trouvés à seulement 36 des 104 sites et plus (le compte des sites n’est pas terminé) (Sheffield, données pers., 2013).
Dans les années 1960 et 1970, les chercheurs qui étudiaient l’écologie des bourdons ont disposé des milliers de nids artificiels dans des zones naturelles du sud de l’Alberta, attirant ainsi des centaines de reines du bourdon de l’Ouest (Hobbs, 1968; Richards, 1978). Au printemps de 1985 et de 1986, les chercheurs ont recueilli quotidiennement toutes les reines du genre Bombus dans les régions de Calgary et de la vallée de la Kananaskis dans le sud de l’Alberta, prélevant bien plus de 200 reines chaque année et produisant un total de 126 mentions du bourdon de l’Ouest (Owen, 1988). Au cours des étés de 1997 à 2000, les chercheurs ont échantillonné périodiquement les ouvrières et les mâles des bourdons dans la vallée de la Kananaskis et le sud de l’Alberta, réunissant plus de 700 mentions du bourdon de l’Ouest (Otterstatter et Whidden, 2004). Les collectes dans la vallée de la Kananaskis entre 1985 et 2000 ont exigé au moins 1500 heures-personnes.
Au cours de la dernière décennie, il s’est fait très peu de recherches. En 2007, des relevés dans le sud-est de l’Alberta ont repéré le bourdon de l’Ouest dans les collines du Cyprès (environ 60 reines; Sheffield, données pers.). En 2013, des relevés ont été effectués dans quatre secteurs de la province. On a recueilli des spécimens, peu fréquemment, dans les régions du parc provincial des Dinosaures, de Red Cliff (au sud de Medicine Hat) et des collines du Cyprès (Sheffield, données pers.), mais aucun dans la région d’Edmonton (Sheffield, données pers.). Ce secteur, croit-on, se trouve hors de l’aire de l’espèce.
Il y a peu de relevés historiques ou de collections muséales de la Saskatchewan. Récemment (2012-2013), il a été fait mention de la sous-espèce occidentalis dans le tiers inférieur de la province. Le Royal Saskatchewan Museum a mené de vastes relevés d’insectes, se prolongeant sur tout l’été, dans quatre régions géographiques du sud-ouest de la province (parc national des Prairies, parc provincial Saskatchewan Landing, Great Sandhills, vallée de la Big Muddy et parc interprovincial des Collines-Cypress). On a repéré la sous-espèce occidentalis sur plusieurs des sites visés et ailleurs dans la province, mais les échantillons sont encore en traitement (Sheffield, comm. pers.). La sous-espèce semble peu commune par comparaison à d’autres bourdons (Sheffield, comm. pers., 2013). Aucune mention de la sous-espèce dans ces secteurs n’a été versée dans les bases de données avant ces relevés. Il est possible que la sous-espèce n’ait pas été aussi présente dans les Prairies que dans les parties ouest de son aire.
Depuis trois ans, des relevés de bourdons le long des routes principales sont effectués au moins quatre jours par an. La sous-espèce mckayi était présente à de nombreux sites inventoriés en 2009, en 2010 et en 2013 (Cannings, comm. pers., 2013; Sheffield, comm. pers., 2013). La sous-espèce mckayi est encore considérée commune dans l’Alaska voisin, où ses représentants constituaient plus de 30 % de tous les bourdons observés (Koch et Strange, 2012). Avant ces relevés, les mentions sont très rares au Yukon.
Il existe très peu de mentions de la sous-espèce mckayi dans les Territoires du Nord-Ouest, et elles proviennent de la limite ouest. Il n’y a qu’une mention antérieure à 2011 (elle date du 4 août 1944, et l’emplacement exact n’est pas indiqué). Les huit autres spécimens proviennent de divers sites le long de la rivière Nahanni-Sud, et ils ont été recueillis en août 2011 (Stotyn et Tate, 2012).
Le bourdon de l’Ouest a besoin d’un habitat qui contient des ressources florales abondantes et des sites de nidification propices. Le bourdon est une espèce qui n’a pas d’exigences précises du point de vue de l’habitat. On le retrouve dans les forêts ouvertes de conifères et de feuillus et les forêts mixtes, les prés humides ou secs, les prairies montagnardes ou herbeuses, les prés bordant les zones riveraines, les bordures de route de la taïga adjacente à des zones boisées, les parcs urbains, les jardins et les zones agricoles, les milieux subalpins et les milieux naturels plus isolés.
Il n’existe que quelques études sur les préférences du bourdon de l’Ouest en matière de nidification naturelle. En Alberta, Hobbs (1968) a attiré 37 reines pour les faire nicher dans des nids artificiels souterrains. Trois autres reines ont établi des colonies dans des nids artificiels situés au-dessus du sol, ce qui laisse croire que l’insecte aurait une préférence pour les nids souterrains (une hypothèse soutenue par Kearns et Thomson, 2001), mais qui fait aussi penser qu’il possède une certaine capacité d’adaptation du comportement (Hobbs, 1968). Dans le même ordre d’idée, Richards (1978) a placé des nids artificiels dans divers habitats du sud-est de l’Alberta et a constaté que 88 reines du bourdon de l’Ouest (12 % de 709 reines parmi 15 espèces de Bombus) avaient établi des nids. Elles semblaient privilégier les nids souterrains reliés à la surface par des tunnels à pente descendante avec une ouverture orientée ouest-sud-ouest (Hobbs, 1968). Les reines préféraient nettement établir leur nid dans les zones boisées et transitionnelles (de boisées à prairiales) plutôt que dans les prairies ouvertes (Richards, 1978).
Chez tous les bourdons, les reines fécondées hivernent en solitaire et passent généralement l’hiver en s’enfouissant dans un sol meuble ou en s’introduisant dans des arbres en décomposition (Benton, 2006). Hobbs (1968) décrit la présence d’un hibernacle de bourdons de l’Ouest à deux pouces sous la surface du versant ouest abrupt d’un monticule de terre.
En général, les bourdons ont besoin de ressources florales et de sites de nidification propices qui permettent à une colonie de subsister pendant toute la saison, ainsi que de sites d’hivernage protégés. Aucune étude n’a permis d’associer des tendances particulières en matière d’habitat aux populations de bourdons de l’Ouest. Cependant, la transformation considérable et cumulative de l’habitat a vraisemblablement causé un déclin dans certaines portions de leur aire de répartition. Les principaux centres urbains du Lower Mainland, de la région du Grand Victoria et de Calgary, conjugués à l’agriculture à grande échelle pratiquée dans ces régions, ont mené à un déclin cumulatif de la qualité de l’habitat.
La fragmentation de l’habitat, les nouveaux développements agricoles, y compris la conversion des cultures pollinisées par des insectes à des cultures pollinisées par le vent ou à des cultures sous serre, et l’intensification de l’agriculture, éventuellement conjugués à une augmentation de la présence d’agents pathogènes, ont vraisemblablement contribué au déclin de la sous-espèce dans une bonne partie de son aire du sud-ouest du Canada (principalement en Colombie-Britannique et dans l’ouest de l’Alberta).
Au cours des dernières décennies, le développement de l’agriculture dans le sud de la Colombie-Britannique et de l’Alberta a rendu moins propice l’environnement de la faune (y compris des pollinisateurs) (Javorek et Grant, 2011). L’exploitation forestière, le broutage par les animaux et l’assèchement des terres humides pourraient avoir altéré la qualité des habitats. En Alberta, par exemple, les contreforts où habite le bourdon de l’Ouest changent rapidement en raison du développement considérable de l’industrie de l’énergie et de l’exploitation forestière et en raison de l’intensification des activités récréatives, de l’agriculture et de la superficie consacrée à celle-ci. Ces différents usages provoquent des perturbations importantes des terres et de l’eau et entraînent une fragmentation de l’habitat (Gardner, 2007); ils sont susceptibles d’avoir des répercussions négatives sur le bourdon de l’Ouest.
Cette sous-espèce pourrait ne pas être aussi courante dans les prairies que dans les parties occidentales de son aire. La conversion de la prairie naturelle s’est produite en bonne partie il y a plusieurs décennies. Par ailleurs, on possède peu de données historiques du bourdon de l’Ouest. C’est pourquoi il est difficile d’en évaluer les tendances.
L’habitat de la sous-espèce ne semble pas avoir changé considérablement au fil du temps, bien que les effets cumulatifs du développement des ressources et des changements climatiques puissent avoir une incidence sur la période de croissance des ressources florales nécessaires au maintien des populations durant toute la saison.
Les données sont réunies à partir de références générales sur les bourdons (Alford, 1975; Goulson, 2003; Benton, 2006) et des références sont fournies, le cas échéant, se rapportant précisément au bourdon de l’Ouest.
Le bourdon de l’Ouest est une espèce au comportement eusocial primitif qui vit en colonie comportant une reine et des ouvrières issues de cette reine. Les colonies ne durent qu’un an et ne produisent qu’une génération par année. L’accouplement se fait à l’automne; les mâles meurent et seules les reines survivent à l’hiver et émergent au printemps pour fonder une nouvelle colonie.
Normalement, les reines émergent d’avril à mai et se mettent immédiatement à la recherche d’un site de nidification propice. Elles établissent leur nid dans des terriers de rongeurs abandonnés, des tertres herbeux, des billes de bois pourries ou des trous dans du bois mort. Les reines qui hivernent dans les zones méridionales de leur aire de répartition émergent plus rapidement que celles qui hivernent dans les zones situées plus au nord. De plus, la période d’émergence varie en fonction de la température et du climat. Les reines de l’Alberta établissent leurs nids de la mi-mai à la fin mai (Hobbs, 1968).
Quelques semaines après la première période de ponte, les ouvrières émergent et se chargent d’approvisionner la colonie en nourriture et de prodiguer les soins au couvain. À mesure que l’été progresse, la production d’ouvrières culmine, et la colonie commence à produire des mâles et de futures reines. Ces individus reproducteurs quittent la colonie et s’accouplent. Les jeunes reines fécondées entrent en diapause et hibernent. Le déclin de la colonie s’amorce à l’approche de l’automne et la vieille reine, les mâles et les ouvrières finissent par mourir au premier gel. On en sait relativement peu sur le comportement d’accouplement et la dynamique des colonies du bourdon de l’Ouest. Chez une espèce étroitement apparentée, le bourdon fébrile, les femelles s’accouplent une seule fois avec un seul mâle et, comme c’est le cas pour toutes les espèces de bourdon, elles conservent le sperme ainsi obtenu dans une spermathèque jusqu’à son utilisation pour la fertilisation (Greeff et Schmid-Hempel, 2008).
L’éclosion des œufs survient environ quatre jours après la ponte. Les larves néonates commencent aussitôt à se nourrir du pollen et du nectar apportés à la ruche par les ouvrières. La vie larvaire des bourdons comporte quatre stades. Après environ deux semaines, les larves se tissent un cocon où elles se transforment en nymphes. Deux semaines plus tard, les adultes émergent. Au total, le développement de l’œuf à l’adulte dure environ cinq semaines, mais la durée de leur développement varie selon la température et la quantité de nourriture disponible (Alford, 1975). Le bourdon de l’Ouest est une espèce qui emmagasine du pollen (« pollen-storer »). Les larves vivent dans des alvéoles et sont nourries individuellement par les adultes, qui ouvrent régulièrement les cellules où elles se trouvent pendant leur développement. Les adultes des espèces qui emmagasinent du pollen sont tous approximativement de la même taille, contrairement à ceux des espèces qui fabriquent des poches (« pocket-making »), chez qui la taille des ouvrières varie considérablement selon la quantité de nourriture à laquelle elles ont eu accès durant leur développement larvaire. Les reines de bourdon de l’Ouest pourraient avoir besoin de plus de pollen que celles d’autres espèces de bourdon pour amorcer la production d’ouvrières (Hobbs, 1968).
Une grande variété d’invertébrés parasitent le bourdon de l’Ouest à tous les stades du cycle de la colonie (Schmid-Hempel, 1998). Les reines qui émergent au printemps peuvent être infectées par des nématodes (Sphaerularia bombi) ou des protozoaires (Apicystis bombi), ce qui les rend incapables de fonder des colonies. Au cours de l’été, les ouvrières peuvent attraper des parasites (p. ex. Crithidia bombi) lorsqu’elles butinent des fleurs contaminées par des abeilles infectées.
Les bourdons « coucous » (sous-genre Psithyrus) se spécialisent dans l’usurpation de reines. Les femelles adultes entrent dans la colonie, tuant parfois la reine, et pondent leurs propres œufs qui sont soignés par les ouvrières restantes de la colonie hôte. Le bourdon de l’Ouest est l’hôte de deux espèces de bourdon coucou, le B. bohemicus Cresson et le B. suckleyi Greene.
L’acarien interne Locustacarus buchneri est un parasite commun qui vit dans les voies respiratoires de la plupart des espèces de Bombus (voire toutes). Une étude menée dans le sud de l’Alberta a montré que cet acarien se retrouvait principalement chez le bourdon de l’Ouest, dont jusqu’à 50 % des reines et des ouvrières étaient infectées (Otterstatter et Whidden, 2004). On n’a pu établir clairement la ou les raisons de cette spécificité; cependant, ce parasite représente une menace pour les populations de bourdon de l’Ouest.
Nosema bombi est une microsporidie qui parasite l’intestin et les tissus des bourdons. Bien que le taux d’infection par Nosema bombi soit considéré faible chez les bourdons sauvages au Canada (taux d’infection moyens entre 5 et 10 %), des études récentes sur le terrain aux États-Unis (Cameron et al., 2011) ont permis de constater les taux les plus élevés d’infection à N. bombi (c.-à-d. plus de 35 %) chez les espèces de bourdon en déclin, particulièrement le bourdon de l’Ouest, ce qui vient étayer l’hypothèse que ce parasite constitue une menace grave. Cependant, Koch et Strange (2012) ont enregistré des taux élevés semblables (c.-à-d, 44 %) chez la sous-espèce mckayi en Alaska, où celle-ci demeure le bourdon le plus couramment observé. Ainsi, des taux élevés d’infection à Nosema pourraient faire partie de la dynamique normale hôte-agent pathogène du bourdon de l’Ouest (Koch et Strange, 2012) et le déclin de la sous-espèce occidentalis pourrait être attribué à la cumulation de plusieurs menaces (voir plus haut). Cependant, on croit que cet agent pathogène serait responsable de l’effondrement des élevages commerciaux du bourdon de l’Ouest en Amérique du Nord, au milieu des années 1990 (Thorp et Shepherd, 2005).
Les prédateurs du bourdon de l’Ouest adulte comprennent divers asiles (famille des Asilidés) et araignées-crabes (famille des Thomisidés). Les mouches conopides (famille des Conopidés) sont des parasitoïdes des bourdons adultes. Le raton-laveur, la moufette, les ours et d’autres mammifères attaquent également les colonies de bourdons (Breed et al., 2004).
Les bourdons hivernent sous forme de reines adultes qui émergent tôt au printemps et doivent compter sur des espèces de plantes qui fleurissent tôt pour fonder la colonie et sur d’autres ressources florales pour assurer le développement de la colonie tout au long du printemps et de l’été. Comme les bourdons ont un comportement obligatoirement social, ils dépendent de groupements végétaux variés et nécessitent des ressources importantes de pollen et de nectar durant toute la période d’activité de la colonie. Par conséquent, seuls les habitats offrant des communautés végétales riches fournissent suffisamment de nourriture pour soutenir les colonies.
On trouve des bourdons presque partout au Canada et de nombreuses espèces (dont le bourdon de l’Ouest) semblent tolérer relativement bien le froid durant leur période active d’adulte. En effet, on a trouvé des spécimens à des altitudes supérieures à 3 800 m dans les parties méridionales de leur aire de répartition globale (USDA, 2010). Les bourdons sont des insectes qui possèdent la capacité physiologique de thermorégulation (Heinrich, 2004). En vibrant très rapidement, ils génèrent de la chaleur dans leurs muscles thoraciques et parviennent ainsi à atteindre la température minimale requise (approximativement 30 °C) lorsque la température ambiante est basse (Heinrich, 2004). Comme ils volent au printemps et en automne dans les régions tempérées et arctiques, cette vibration leur permet d’élever leur température interne bien au-dessus de la température ambiante. Puisque le bourdon de l’Ouest est une espèce qui émerge tôt à des latitudes et à des altitudes élevées, la thermorégulation est un mécanisme d’adaptation extrêmement important.
On en sait peu sur les taux de dispersion naturelle du bourdon. La dispersion se fait principalement au printemps lorsque les reines se mettent à la recherche de sites de nidifications propices (Goulson, 2003). Certaines données indiquent que les bourdons peuvent se disperser sur de grandes distances. Ainsi, des mâles d’une espèce étroitement apparentée, B. terrestris, ont été trouvés dans un rayon variant de 2,6 à 9,9 km de leur colonie d’origine (Kraus et al., 2008). En Tasmanie, où il a été introduit au début des années 1990, B. terrestris s’est dispersé au rythme moyen d’une dizaine de kilomètres par année (Stout et Goulson, 2000). Selon des études axées sur la répartition irrégulière des habitats propices pour les bourdons (voir par exemple Hatfield et LeBuhn, 2007) et la difficulté accrue pour les petites populations d’insectes haplodiploïdes d’atteindre un nombre d’individus efficace (Zayed et Packer, 2005), la dispersion est un atout important pour la survie (voir les « Facteurs limitatifs »).
Le bourdon de l’Ouest est une espèce généraliste sur le plan de l’alimentation; il doit entrer en compétition avec de nombreuses autres espèces de bourdons pour butiner le pollen et le nectar. Cette espèce peut également « voler » du nectar (c.-à-d. qu’elle peut visiter les fleurs sans effectuer de pollinisation : elle perce un trou à la base de la fleur pour atteindre le nectar sans toucher aux anthères ou au stigmate), ce qui permet aux ouvrières de butiner le nectar des fleurs à longue corolle tubulaire. Ainsi, l’espèce arrive à s’alimenter du nectar d’une grande variété d’espèces de plantes à fleurs.
Le bourdon de l’Ouest entretient vraisemblablement d’importantes relations mutualistes avec de nombreuses espèces de plantes qui fleurissent tôt au printemps et d’espèces montagnardes qui pourraient dépendre de lui et d’autres bourdons pour la pollinisation. Cependant, on ne connaît pas l’intensité de cette interdépendance. Lorsque l’aire de répartition du bourdon de l’Ouest chevauche celle d’une autre espèce étroitement apparentée, le bourdon terricole, on le rencontre plus fréquemment dans les habitats montagnards, alors qu’on trouve davantage le bourdon terricole à moindre altitude (Hobbs, 1968), même si les deux espèces vivent en sympatrie dans certaines parties des prairies.
Le bourdon de l’Ouest est l’hôte des bourdons coucous B. bohemicus Cresson et B. suckleyi Greene (voir « Prédation et parasitisme »).
Quatre ensembles de données sont utilisés pour montrer le déclin de l’abondance relative du bourdon de l’Ouest. L’abondance relative (AR) est le nombre d’individus d’une seule espèce (p. ex. le bourdon de l’Ouest) divisé par le nombre total d’individus (p. ex, Bombus) recueillis; cette valeur sert souvent comme mesure substitutive de l’abondance lorsque les données ne se prêtent pas à d’autres analyses. Pour le bourdon de l’Ouest, l’AR est aussi utilisée comme indicateur des activités de recherche, et l’on présume que si l’espèce était présente dans une région durant une activité de prélèvement, des spécimens en auraient été vraisemblablement recueillis. Il convient de noter qu’il est possible que la mesure de l’AR d’une espèce puisse ne pas correspondre à l’abondance relative réelle de la population. Dans les prochaines sections, les études sont numérotées pour faciliter la référence d’une section à l’autre.
- Dans la première étude, on a utilisé un ensemble de données concernant les bourdons du Canada; cet ensemble était composé de 44 706 enregistrements provenant de musées et d’observations recueillies sur une période s’échelonnant de 1882 à 2011 (l’ensemble de données n’englobait pas encore les années 2012 et 2013 [p. ex. Sheffield et al., en prép.]). L’AR du bourdon de l’Ouest a été calculée par tranches de 10 ans, de même que par territoire où le bourdon a été observé au Canada, et représentée graphiquement (tableau 3; figure 9 et figure 10).
- Dans la seconde étude (en utilisant une version plus ancienne et moins complète de l’ensemble de données sur les bourdons précité), la répartition du bourdon au Canada a été portée sur une grille dont les cellules représentent une superficie de 50 km sur 50 km, de 1882 à 1995 et de 1996 à 2010 (incluant des échantillons non indépendants et des emplacements où plusieurs observations ont été enregistrées au cours d’une année). La figure 6 donne la représentation graphique de ces données.
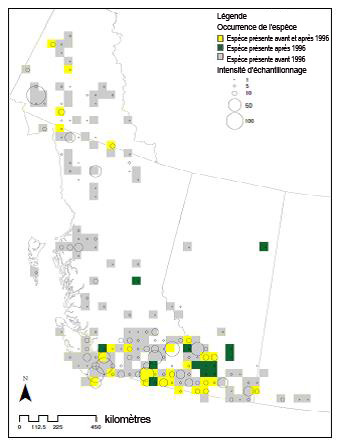
Image : © Environnement Canada
Description longue pour la figure 6
Carte montrant la répartition spatiale des données d’échantillonnage du bourdon de l’Ouest au Canada de 1882 à 2010. La carte est basée sur la seconde étude dont il est question dans le texte. Dans cette étude, la répartition du bourdon de l’Ouest au Canada de 1882 à 1995 et de 1996 à 2010 a été portée sur une grille dont les cellules représentent une superficie de 50 km sur 50 km (y compris les échantillons non indépendants et les sites où plusieurs observations ont été consignées au cours d’une année). Les symboles indiquent 128 mentions avant 1996, 27 sites où des individus ont été prélevés avant et après 1996, et 13 prélèvements après 1996.

Image : © Environnement Canada
Description longue pour la figure 7
Diagramme à barres montrant l’abondance relative (AR) du bourdon de l’Ouest dans le sud de l’Alberta au cours de deux intervalles de temps : 1985 à 2000 et 2010. Les données montrent que, à l’exception de deux sites où le bourdon de l’Ouest n’a été repéré durant aucune des deux périodes (Innisfail et route Trunk), on a observé une baisse de l’AR entre 2000 et 2010. Les baisses sont les suivantes : Calgary (zone urbaine), 59 à 4 %; Clarsehom (zone rurale), 21 à 3 %; lac Barrier (zone naturelle), 15 à 0 %; Drumheller (limite de l’aire de répartition de l’espèce), 5 à 3 %; mont Fortress, 5 à 4 %; Coleman (zone rurale) 4 à 0 %; Innisfail (à l’extérieur de l’aire de répartition de la sous-espèce) et route Trunk, 0 % au cours des deux périodes.
Deux autres études, de portée régionale, ont été menées, l’une dans le sud de l’Alberta et l’autre dans la vallée du Fraser (en Colombie-Britannique), et ont servi à montrer le déclin de l’AR dans chacune de ces régions. Il est important de noter que, vu l’emplacement géographique des sites d’échantillonnage, on présume que les spécimens prélevés appartiennent à la sous-espèce occidentalis.
3) Dans l’étude menée dans le sud de l’Alberta, on a comparé des données recueillies de 1985 à 2000 et l’on a répété, en 2010, la collecte de données aux mêmes sites en utilisant des méthodes semblables. L’étude menée dans la vallée du Fraser compare les populations de 1981-1982 avec celles de 2003-2004. Pour établir le présent rapport de situation, ces études ont été analysées afin de déterminer l’évolution de l’AR du bourdon de l’Ouest. La proportion de bourdons de l’Ouest par rapport au total de bourdons capturés a été évaluée au moyen d’une régression logistique; l’analyse tenait compte de la période (années au début ou à la fin de l’étude) et du site de l’étude (dans le cas des relevés effectués dans le sud de l’Alberta seulement). Cette démarche permet d’estimer la variation, au fil du temps, de la probabilité de prélever des bourdons de l’Ouest au site de l’étude.
Dans l’étude sur les bourdons menée en Alberta, des ouvrières et des mâles ont été relevés à huit endroits du sud de la province au cours de l’été 2000 (n = 1 672, toutes espèces confondues); la démarche a été répétée aux mêmes endroits en 2010 (n = 775). On peut trouver des détails sur l’étude de 2000 dans d’autres publications (Otterstatter, 2001), tandis que la deuxième étude a été effectuée particulièrement pour recueillir des données aux fins du présent rapport (études menées par R. Owen [données pers.]; R. Longair [données pers.]). Les lieux et la période de prélèvement ainsi que les activités de recherche ont été les mêmes en 2000 et en 2010, et les deux études sont jugées hautement comparables.
Les résultats d’autres relevés effectués dans la même région géographique, mais à des périodes différentes ou à des emplacements différents sont venus compléter ces données. Parmi les autres études ayant permis de recueillir des données, mentionnons les suivantes : une étude en 1985-1986 sur les reines de bourdon (n = 442) dans la vallée de la Kananaskis et la région de Calgary (Owen, 1988); des relevés intensifs de bourdons près du lac Barrier, dans la région de Kananaskis, durant la période de 1997 à 2000 (n = 4 376) et à Calgary au cours de l’année 1998 (n = 367) (Otterstatter et al., 2002; Otterstatter, 2004; Otterstatter et Whidden, 2004); une étude sur le butinage du pollen par les reines et les ouvrières (n = 99) à trois emplacements sur les contreforts du sud-ouest de l’Alberta durant l’année 1991-1992 (Rasheed et Harder, 1997); des relevés d’ouvrières et de mâles (n = 109), réalisés selon les opportunités à cinq sites dans le sud de l’Alberta au cours de l’année 2000 (R. Owen, données pers.).
4) Dans la vallée du Fraser, des bourdons sauvages ont été prélevés dans des cultures commerciales de petits fruits en 1981-1982, puis encore en 2003-2004 (voir les détails dans Winston et Graf, 1982; MacKenzie et Winston, 1984; Ratti, 2006; Ratti et al., 2008; Colla et Ratti, 2010). Au cours de l’étude la plus récente, des prélèvements ont été effectués dans douze sites au moyen de filets fauchoirs et de pièges à eau. Les deux études (1981-1982 et 2003-2004) ont été menées dans la même région, au moyen de méthodes semblables, et sont considérées hautement comparables.
- L’AR du bourdon de l’Ouest semble décliner dans chaque territoire (tableau 3, figure 9) et dans l’ensemble de l’aire de répartition de l’espèce au Canada lorsqu’on combine toutes les mentions (tableau 3; figure 10). En Colombie-Britannique, l’AR a chuté pour passer d’environ 43 % (1992-2001) à 3 % au cours de la dernière période de 10 ans (2002-2011), ce qui donne un déclin total de plus de 85 %. En Alberta, l’AR du bourdon de l’Ouest est passé de 83 % (1992-2001) à moins de 10 % (2002-2011). En ce qui concerne la Saskatchewan, dans l’ensemble, on possède peu de données historiques sur le bourdon; cependant, on observe un déclin de l’AR, qui est passée de 9 % à 3 % au cours de la dernière période de 10 ans.
- L’étude montre la répartition spatiale des données d’échantillonnage du bourdon de l’Ouest au Canada de 1882 à 2010 (figure 6). Chaque cercle est proportionnel au nombre d’observations consignées à cet endroit entre 1882 et 2010, inclusivement (y compris les échantillons non indépendants et les sites où plusieurs observations ont été enregistrées au cours d’une année). Les données ont été cartographiées à l’aide de carrés représentant 50 km sur 50 km. Les carrés gris indiquent les mentions antérieures à 1996 (128 carrés); les carrés jaunes montrent les sites où des spécimens ont été recueillis avant et après 1996 (27 carrés); les carrés verts indiquent les prélèvements effectués après 1996 (13 carrés) (n = 1 706 spécimens).
Deux études ciblant des régions particulières montrent un déclin de l’AR de la sous-espèce occidentalis. - En Alberta, on observe un déclin de l’AR, qui passe de 16,9 % (n = 1 017 [sous-espèce occidentalis] sur 6 006 [Bombus prélevés]) au cours de la période 1985 à 2000 à 3,2 % (n = 25 [sous-espèce occidentalis] sur 775 [Bombus prélevés]) en 2010 (figures 8 et 9). Le nombre le plus élevé de bourdons de l’Ouest dans cette étude a été consigné au lac Barrier (n = 4 924), et l’un des changements les plus marquants observés est la disparition totale (ou presque) de cette espèce de bourdon d’un des sites du lac Barrier (Kananaskis) alors qu’elle se situait auparavant au 3e et au 4e rang des espèces de bourdons les plus couramment prélevées. Dans six emplacements échantillonnés en 2000, puis à nouveau en 2010 (huit sites au total, mais deux sites ont été exclus parce que le bourdon de l’Ouest n’avait pas été signalé à aucune de ces périodes), l’AR a chuté de 80 %, passant de 14 % à 0,7 % en 10 ans.
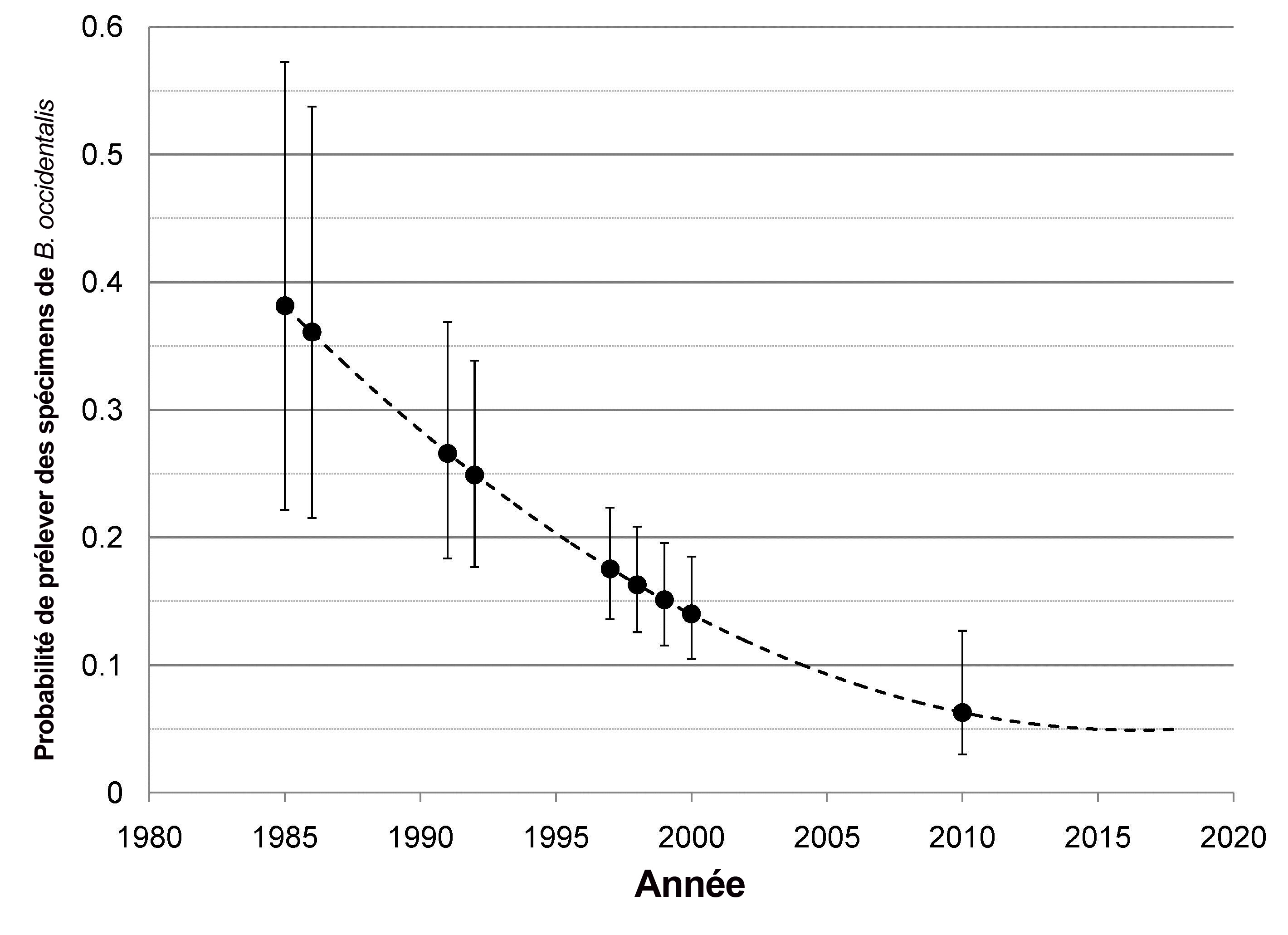
Image : © Environnement Canada
Description longue pour la figure 8
Graphique illustrant la probabilité moyenne de prélever un bourdon de l’Ouest (axe des Y) durant une année donnée (axe des X), selon les données obtenues de 14 sites du sud de l’Alberta échantillonnés entre 1985 et 2010. Les points indiquent les probabilités moyennes des sites échantillonnés au cours d’une année donnée. Une ligne relie les points de 1985 à 2010, et elle est étendue jusqu’à 2018 en fonction des valeurs prévues d’après une équation de régression. Le graphique montre une baisse abrupte de la probabilité de prélever un bourdon de l’Ouest de 1985 à 2010 en Alberta, avec une probabilité moyenne chutant de presque 0,4 à tout juste plus de 0,05.
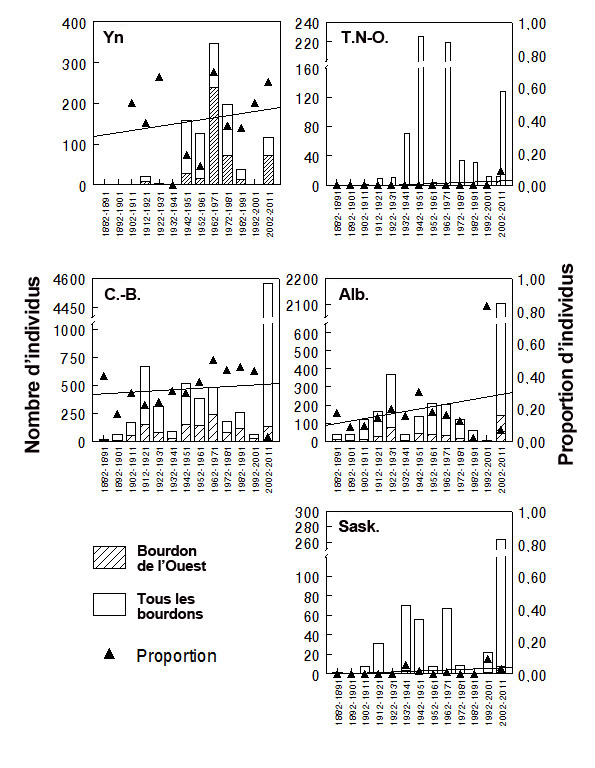
Image : © Environnement Canada
Description longue pour la figure 9
Graphique à cinq volets illustrant l’abondance relative (AR) du bourdon de l’Ouest d’après toutes les mentions de Bombus compilées au Canada (de 1882 à 2011; les données sont présentées à des intervalles de 10 ans) (voir aussi le tableau 3). Un axe des Y indique le nombre de spécimens de bourdon de l’Ouest, et l’autre axe des Y représente la proportion de spécimens de bourdon de l’Ouest par rapport à l’ensemble. Les volets présentent les données pour le Yukon, les Territoires du Nord Ouest, la Colombie-Britannique, l’Alberta et la Saskatchewan. L’abondance relative du bourdon de l’Ouest semble baisser dans chaque territoire (voir le texte du rapport, à la section Abondance).
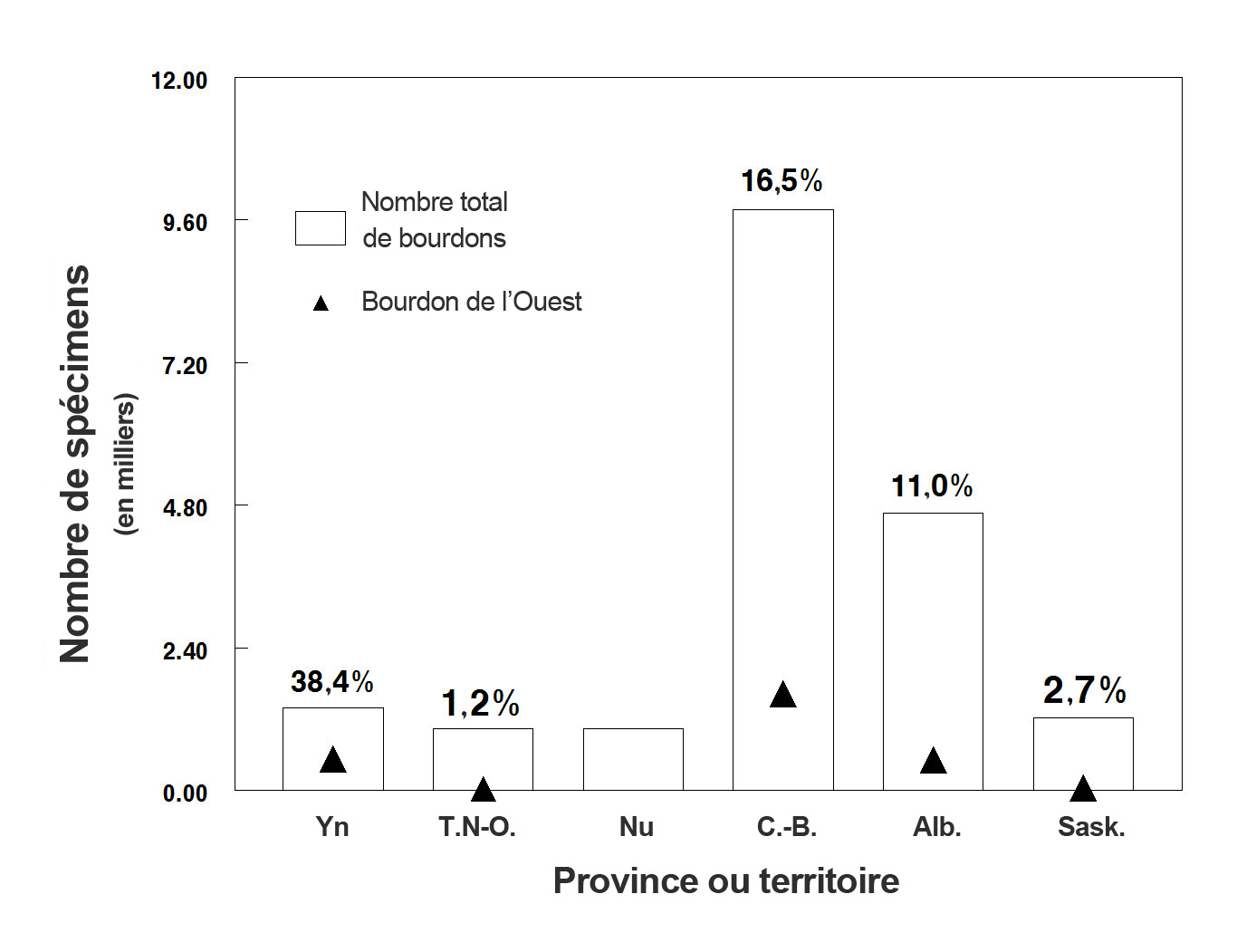
Image : © Environnement Canada
Description longue pour la figure 10
Diagramme à barres indiquant le nombre total de spécimens de bourdons capturés au Canada et répertoriés dans la base de données, de 1882 à 2011, dans chaque province et territoire. Les symboles indiquent le nombre de spécimens de bourdon de l’Ouest, et le pourcentage de bourdons de l’Ouest prélevés par rapport au reste de l’ensemble est indiqué au dessus de chaque barre. (Voir aussi le tableau 3.) Les bourdons de l’Ouest représentent 38,4 % des spécimens au Yukon, 1,2 % dans les Territoires du Nord Ouest, 0 % au Nunavut, 16,5 % en Colombie-Britannique, 11 % en Alberta et 2,7 % en Saskatchewan.
L’aire de répartition du bourdon de l’Ouest ne semble pas s’être déplacée vers le nord : aucun bourdon n’a été signalé dans les deux sites le plus aux nord figurant dans l’ensemble de données de l’Alberta (c.-à-d. le chemin Forestry Trunk et Innisfail) au cours de la période la plus récente. Le classement des sites en fonction de l’écorégion (selon la définition de Strong, 1992) laisse entendre que les déclins les plus importants ont été enregistrés dans les prairies-parcs (régions autour de Calgary et au sud de celle-ci; l’AR a chuté, passant de 58,6 % à 6,3 %). Des changements importants ont également été observés dans les régions montagneuses (vallée de la Kananaskis et régions situées au sud entre les prairies herbeuses et les Rocheuses; on a observé un déclin de l’AR, qui est passé de 14,8 % à 0 %). Les changements observés dans les prairies herbeuses étaient moins prononcés (de 12,7 à 3,1 %). Pour ce qui est des habitats subalpins, on n’a constaté aucun changement important (de 5,0 % à 5,2 %; cependant, l’abondance du bourdon de l’Ouest est généralement faible dans les régions subalpines.
La variation de la probabilité de prélever des bourdons de l’Ouest durant la période 1985-2010 en Alberta montre une forte décroissance au cours des 25 dernières années; cette estimation a été obtenue à partir de la régression logistique de l’AR du bourdon de l’Ouest dans l’ensemble des 14 sites (figure 8). Étant donné que ces prévisions sont fondées sur les données cumulatives des diverses régions, la marge d’erreur est importante (comme l’indiquent les grands intervalles de confiance des premières années) et tient compte des activités et des méthodes de recherche. Par exemple, les activités de recherche en Alberta ont été plus importantes au cours des années 1985 à 2000 (n = 6 006, en raison des nombreuses heures-personnes sollicitées par plusieurs études) qu’en 2010 (n = 775, résultat de l’utilisation de deux préleveurs d’échantillons dans une seule étude).
4) L’étude menée dans la vallée du Fraser a montré un déclin de l’AR dans les champs de petits fruits, de 33,3 % (n = 608/1 828) en 1981-1982 à 0,7 % (30/4 221) en 2003-2004 (pour plus de précisions, voir Winston et Graf, 1982; MacKenzie et Winston, 1984; Ratti, 2006; Ratti et al., 2008; Colla et Ratti, 2010).
Aux États-Unis, on a observé une décroissance des populations chez certaines des espèces de bourdons les plus abondantes historiquement (dont le bourdon de l’Ouest) qui occupaient autrefois de grandes aires de répartition (Cameron et al., 2011). En tout, 16 788 bourdons ont été prélevés sur tout le territoire des États-Unis sauf en Alaska (p. ex. à l’intérieur de l’aire de répartition de la sous-espèce occidentalis) entre 2007 et 2009 et, de ce nombre, seulement 129 spécimens de bourdon de l’Ouest ont été prélevés (Cameron et al., 2011). Toutes les détections se sont produites dans les montagnes Rocheuses et dans la zone intramontagneuse de l’ouest (soit la région d’Amérique du Nord située entre les montagnes Rocheuses à l’est et la chaîne des Cascades et la Sierra Nevada à l’ouest). L’espèce est largement absente de la partie occidentale de son aire de répartition (historiquement la côte Ouest du Pacifique, la Californie, l’Orégon et l’État de Washington) (Cameron et al., 2011). On estime la diminution de l’aire de répartition où des bourdons ont été détectés dans le cadre de cette étude à 28 % sur plus de 100 ans.
Au Yukon et dans les Territoires du Nord-Ouest, l’AR semble stable (tableau 3; figure 10). Il y a peu de spécimens historiques et, par conséquent, on possède peu de données comparables à partir desquelles on pourrait dégager des tendances. En se basant sur les relevés de 2013 (au moins 20 sites d’échantillonnage, durée de prélèvement d’au moins 30 minutes; les données n’ont pas encore été versées dans l’ensemble de données sur les bourdons au Canada), ce bourdon a été observé dans de nombreux sites de l’aire de répartition au Yukon (Sheffield, données pers., 2013). Bien que l’on connaisse les données visant la partie ouest des Territoires du Nord-Ouest, aucune tendance n’est dégagée, car la plupart des observations de l’espèce datent de 2011 (voir également Stotyn, 2012).
En Alaska, Koch et Strange (2012) indiquent que le bourdon de l’Ouest est l’espèce la plus courante. On le trouve fréquemment en bordure des routes, de la taïga aux forêts boréales, et il était plus abondant en août 2010 dans cet État que dans les États de l’ouest situés plus bas (c.-à-d. dans l’aire de répartition de la sous-espèce occidentalis) où des relevés ont été effectués la même année (Strange, comm. pers., 2010).
On en sait peu sur les variations naturelles et les tendances des populations de bourdons. Le bourdon de l’Ouest était autrefois l’une des espèces les plus fréquentes dans l’ouest de l’Amérique du Nord, et la décroissance récente de la sous-espèce occidentalis sur la majeure partie de son aire de répartition mondiale laisse croire que cette tendance n’est pas due à une variation naturelle.
Si l’on compare les données recueillies avant 1996 et après 1995 (voir la figure 6), l’aire de répartition de la sous-espèce occidentalis semble avoir rétréci. En combinant les statistiques sur la distribution des fréquences aux données sur la répartition spatiale des observations, il semble que le déclin des populations du bourdon de l’Ouest se soit amorcé après les années 1970 (figure 4). Cependant, les mesures servant à estimer les activités d’échantillonnage ne sont pas disponibles; ainsi, les tendances au chapitre de la répartition spatiale pourraient également refléter des écarts en ce qui concerne les activités d’échantillonnage ainsi que des inexactitudes spatiales liées au géoréférencement des données sur les spécimens et les observations. Les tendances en matière de déclin établies à partir des sites où des échantillons ont été prélevés au cours des trois intervalles laissent croire qu’une décroissance d’environ 60 % s’est produite dans les aires occupées (tableau 1). Les sites représentés sous forme de grille dont les cellules équivalent à 50 km de côté ont été choisis de façon à correspondre aux travaux précédents sur le Bombus (p. ex. Fitzpatrick et al., 2007). De plus, l’altitude moyenne où des bourdons de l’Ouest ont été observés a été la plus basse dans la période précédant 1996 et la plus élevée dans la période suivant 1996 (tableau 1). Cependant, la différence d’altitude moyenne entre ces deux périodes n’était que d’environ 100 m. L’intensité d’échantillonnage relative n’était pas uniforme d’une période à l’autre (n = 128, 27 et 13); il est donc difficile d’établir une tendance uniforme en faveur d’altitudes plus élevées.
| Avant 1996 | Avant 1996 et ultérieurement | Après 1995 | Répartition actuelle (toutes les observations depuis 1995) | |
|---|---|---|---|---|
| Superficie (km2) | 320 000 | 67 500 | 32 500 | 100 000 |
| Nbre de sites (chaque site = 50 km2) | 128 | 27 | 13 | 40 |
| Altitude moyenne (m) | 1 114 | 1 210 | 1 218 | |
| Écart-type de l’altitude (m) | 520 | 600 | 441 | |
| Variation de l’altitude (m) | 0 – 2 615 | 20 – 2 177 | 396 – 1 920 |
Des zones étendues de l’aire de répartition du bourdon de l’Ouest au Canada sont sous-échantillonnées. À l’intérieur des aires de répartition géographique des deux sous espèces, les populations se trouvant dans un milieu naturel propice pourraient éventuellement se disperser et recoloniser les secteurs où les effectifs ont accusé une décroissance. Cependant, on ne connaît pas les effets de cette dynamique source-puits de l’effet de l’immigration de source externe. L’immigration de source externe entre les sous-espèces occidentalis et mckayi n’est pas applicable.
La majeure partie de la zone nordique de l’aire de répartition de la sous-espèce occidentalis au Canada demeure propice et n’a presque pas subi de modification, mais elle demeure toutefois sous-échantillonnée. Les travaux récents (2013) montrent que la sous-espèce est toujours présente dans la région centrale intérieure de la Colombie-Britannique (voir les « Activités de recherche »), bien qu’un faible pourcentage, soit 1,7 % du total examiné lors de l’étude menée en 2013 (des données sur certains spécimens sont encore en cours de traitement) soient des bourdons de l’Ouest (Sheffield et al., en prép; Sheffield et al., 2013). L’aire de répartition de la sous-espèce occidentalis s’étend jusque dans l’ouest des États-Unis où sa population a également décliné (Cameron et al., 2011). Ainsi, bien qu’il existe des milieux propices, les populations des États-Unis pourraient ne pas être suffisamment abondantes pour contribuer à l’effet de sauvetage des habitats canadiens par immigration de source externe.
La sous-espèce mckayi est plus souvent prélevée dans l’ensemble de son aire de répartition du nord de la Colombie-Britannique, du Yukon et de l’ouest des Territoires du Nord-Ouest. Le sauvetage par immigration de source externe de la sous-espèce mckayi est possible.
On s’est servi du calculateur des menaces (Salafsky et al., 2008; Master et al., 2009) du Partenariat pour les mesures de conservation de l’Union internationale pour la conservation de la nature (IUCN-CMP) pour classer et énumérer les menaces qui pèsent sur la sous-espèce occidentalis, et on a obtenu un faible taux d’incidence générale des menaces (tableau 3). Malgré la faible incidence apparente, les populations de la sous-espèce occidentalis semblent décroître, si l’on se fie à l’abondance observée lors des activités de prélèvement récentes (voir sous « Activités de recherche » et « Taille et tendances des populations »). On croit que les répercussions cumulatives des nombreuses menaces ont contribué au déclin de cette sous-espèce. L’évaluation des menaces pour la sous-espèce mckayi n’est pas terminée.
En raison d’éléments génétiques inhérents à l’haplodiploïdie, les bourdons sont plus vulnérables que d’autres espèces animales à la fragmentation de leur habitat (Packer et Owen, 2001) (voir les « Facteurs limitatifs »). De plus, ils ont besoin de quantités importantes de ressources florales (c.-à-d. le pollen et le nectar) pendant toute la saison de croissance, car les reines qui produiront les nouvelles générations ne se développeront qu’à la fin du cycle de la colonie. Les menaces qui pèsent sur les ressources florales, les sites de nidification (durant la saison de croissance) et les sites d’hivernage peuvent avoir des répercussions immenses sur les populations de bourdon locales.
La perte d’habitat attribuable au développement résidentiel et commercial intensif dans les zones urbaines peut contribuer au déclin des populations locales de la sous-espèce occidentalis. Cependant, il y a eu quelques observations récentes (au cours des cinq dernières années) de la sous-espèce à Victoria (2012), à Delta (2010) et dans d’autres zones urbaines (voir le tableau 2). Cette menace s’applique uniquement aux zones fortement urbanisées et peuplées par des humains : Colombie-Britannique – les basses-terres continentales, la vallée du bas Fraser et la région de Victoria; Alberta – Calgary et les régions environnantes. Cette menace ne semble pas s’appliquer à la sous-espèce mckayi.
| Province | Région générale | Année | Activités de recherche | Observations | Référence |
|---|---|---|---|---|---|
| C.-B. | Sud-est de l’île de Vancouver | 2010 | 332 km 106 heures | 0 | Page, Lilley et Heron, 2010 |
| Basses-terres continentales | 2010 | 64 sites 271 heures 355 km | 1 (sous-espèce occidentalis) | Parkinson et Heron, 2010 | |
| Vallée du bas Fraser | 2010 | 46 sites 18 jours | 6 (sous-espèce occidentalis) | Knopp, Larkin et Heron, 2010 | |
| Okanagan | 2010 | 40 sites 158 heures 147 km | 4 (sous-espèce occidentalis) | Marks et Heron, 2010 | |
| Kootenay-Ouest | 2010 | 11 sites 19 km 40 heures | 6 (sous-espèce occidentalis) | Westcott et Heron, 2010 | |
| Ensemble des parcs du sud de l’aire de répartition de la C.-B., de l’Alb. et de la Sask. | 2010 | Inconnu | Oui (sous-espèce occidentalis) | Best, données pers., 2010 | |
| Région de Victoria, sud de l’île de Vancouver | 2012 | 7 sites; tous des parcs municipaux en milieu urbain | 9 (sous-espèce occidentalis) | Wray, comm. pers., 2013 | |
| Région centrale intérieure de la C.-B. | 2013 | 281 heures | En cours, les deux sous-espèces | Sheffield, données pers., 2013; Heron, données pers., 2013 | |
| Parties nord le long de l’autoroute de l’Alaska de Fort St. John à Atlin | 2013 | Du 28 mai au 2 août; 55 sites; activités de recherche pendant au moins 30 minutes par site. | Oui. De nombreux spécimens au Yn et dans le nord de la C.-B. | Cannings, données pers., 2013 | |
| Alb. | Edmonton | 2013 | En cours | Oui, en cours (sous-espèce occidentalis) | C. Sheffield, données pers., 2013 |
| Sask. | Les collines du Cyprès et loin dans les prairies (Leader, Eastend, Shaunovan, Swift Current et vers l’est jusqu’à Regina) | 2013 | En cours | Oui, en cours (sous-espèce occidentalis) | C. Sheffield, données pers., 2013 |
| T.N-O. | Rivière Nahanni-Sud de Moose Ponds à Blackstone Landing sur la rivière Liard, incluant la réserve du parc national Nahanni | 2011 | Prélèvement opportuniste de bourdons à 19 sites du 5 au 26 juillet. | Oui. Trois Bombus occidentalis mckayi (des 78 bourdons prélevés). | Stotyn et Tate, 2012 |
| Yn | Toutes les parties sud | 2013 | Du 28 mai au 2 août; 16 sites; activités de recherche pendant au moins 30 minutes par site. | Oui. De nombreux spécimens de Bombus occidentalis mckayi au Yn et dans le nord de la C.-B. | Cannings, données pers., 2013 |
| Yn (sous-espèce mckayi) | T.N.-O. (sous-espèce mckayi) | T.N.-O. (sous-espèce mckayi) | C.-B. (sous-espèce occidentalis et sous-espèce mckayi) | Alb. (sous-espèce occidentalis) | Sask. (sous-espèce occidentalis) | Sask. (sous-espèce occidentalis) | Variation globale de l’AR par rapport à la décennie précédente | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1882-1891 | 0 | 0 | 1 | 15 | 39 | 2 | 57 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1882-1891 | 0 | 0 | 0 | 6 | 7 | 0 | 13 | |
| Nombre de spécimens dans la base de données | AR | 1882-1891 | - | - | - | 0,4 | 0,179487 | - | 0,22807 | |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1892-1901 | 0 | 0 | 0 | 59 | 34 | 0 | 93 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1892-1901 | 0 | 0 | 0 | 10 | 3 | 0 | 13 | |
| Nombre de spécimens dans la base de données | AR | 1892-1901 | 0 | - | - | 0,169492 | 0,088235 | - | 0,139785 | 39 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1902-1911 | 2 | 1 | 3 | 166 | 119 | 8 | 299 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1902-1911 | 1 | 0 | 0 | 49 | 11 | 0 | 61 | |
| Nombre de spécimens dans la base de données | AR | 1902-1911 | 0,5 | - | - | 0,295181 | 0,092437 | - | 0,204013 | 46 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1912-1921 | 21 | 9 | 44 | 668 | 166 | 31 | 939 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1912-1921 | 8 | 0 | 0 | 149 | 24 | 0 | 181 | |
| Nombre de spécimens dans la base de données | AR | 1912-1921 | 0,380952 | - | - | 0,223054 | 0,144578 | - | 0,192758 | 0,055169 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1922-1931 | 3 | 10 | 13 | 313 | 372 | 0 | 711 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1922-1931 | 2 | 0 | 0 | 76 | 74 | 0 | 152 | |
| Nombre de spécimens dans la base de données | AR | 1922-1931 | 0,666667 | - | - | 0,242812 | 0,198925 | - | 0,213783 | -11 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1932-1941 | 1 | 70 | 15 | 88 | 39 | 70 | 283 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1932-1941 | 0 | 0 | 0 | 27 | 6 | 4 | 37 | |
| Nombre de spécimens dans la base de données | AR | 1932-1941 | - | - | - | 0,306818 | 0,153846 | 0,057143 | 0,130742 | -38 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1942-1951 | 157 | 226 | 92 | 513 | 135 | 56 | 1179 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1942-1951 | 29 | 1 | 0 | 152 | 41 | 1 | 224 | |
| Nombre de spécimens dans la base de données | AR | 1942-1951 | 0,184713 | - | - | 0,296296 | 0,303704 | 0,017857 | 0,189992 | -0,45318 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1952-1961 | 126 | 4 | 33 | 377 | 211 | 8 | 759 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1952-1961 | 15 | 0 | 0 | 137 | 38 | 0 | 190 | |
| Nombre de spécimens dans la base de données | AR | 1952-1961 | 0,119048 | - | - | 0,363395 | 0,180095 | - | 0,250329 | 32 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1962-1971 | 348 | 219 | 135 | 478 | 201 | 67 | 1448 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1962-1971 | 239 | 0 | 0 | 239 | 32 | 1 | 511 | |
| Nombre de spécimens dans la base de données | AR | 1962-1971 | 0,686782 | - | - | 0,5 | 0,159204 | 0,014925 | 0,352901 | 41 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1972-1981 | 198 | 33 | 1 | 174 | 120 | 9 | 535 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1972-1981 | 71 | 0 | 0 | 76 | 15 | 0 | 162 | |
| Nombre de spécimens dans la base de données | AR | 1972-1981 | 0,358586 | - | - | 0,436782 | 0,125 | - | 0,302804 | -14 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1982-1991 | 37 | 31 | 10 | 253 | 59 | 1 | 391 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1982-1991 | 13 | 0 | 0 | 115 | 1 | 0 | 129 | |
| Nombre de spécimens dans la base de données | AR | 1982-1991 | 0,351351 | - | - | 0,454545 | 0,016949 | - | 0,329923 | 9 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 1992-2001 | 2 | 12 | 0 | 58 | 6 | 22 | 100 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 1992-2001 | 1 | 0 | 0 | 25 | 5 | 2 | 33 | |
| Nombre de spécimens dans la base de données | AR | 1992-2001 | 0,5 | - | - | 0,431034 | 0,833333 | 0,090909 | 0,33 | 0 |
| Nombre de spécimens dans la base de données | Toutes les espèces de bourdons | 2002-2011 | 116 | 140 | 59 | 4573 | 2103 | 263 | 7254 | |
| Nombre de spécimens dans la base de données | Bourdon de l’Ouest | 2002-2011 | 73 | 12 | 0 | 128 | 142 | 8 | 363 | |
| Nombre de spécimens dans la base de données | AR | 2002-2011 | 0,62931 | 0,085714 | - | 0,02799 | 0,067523 | 0,030418 | 0,050041 | -85 |
| Impact des menaces | Impact des menaces (descriptions) | Maximum | Minimum |
|---|---|---|---|
| Explication de l’impact des menaces La sous-espèce occidentalis est principalement menacée par les effets cumulatifs (en ordre d’importance) : 8.1) Espèces exotiques et non indigènes envahissantes : Propagation d’agents pathogènes (l’utilisation pour la pollinisation des cultures sous serre d’espèces de bourdons commerciaux infectés [p. ex. l’utilisation du bourdon fébrile dans l’ouest du Canada] pourrait contribuer à la propagation d’agents pathogènes chez les populations sauvages de bourdons butinant à proximité). Des études en laboratoire montrent que les espèces parasites Crithidia bombi et Nosema bombi (soupçonnée) ont des effets indésirables sur les reines qui fondent les colonies de bourdons, les ouvrières participant à la quête de nourriture et sur les colonies tout entières. 9.3) Eaux résiduaires de l’agriculture et de la foresterie : Le pesticide imidaclopride (un néonicotinoïde) est nuisible à des concentrations de l’ordre de quelques parties par milliard (ppb). Les néonicotinoïdes sont des pesticides systémiques qui s’accumulent et circulent dans toute la plante et qui atteignent le pollen et le nectar. Ils sont fréquemment utilisés sur les parcours de golf et les terres agricoles. 2.1) Cultures non ligneuses annuelles et pluriannuelles : Réductions cumulatives des ressources florales pour les bourdons sauvages dans les aménagements paysagers dominés par des monocultures, en particulier celles qui n’ont pas besoin des insectes pour la pollinisation. |
|||
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 0 | 0 |
| D | Faible | 2 | 2 |
| Valeur d’impact global des menaces calculée : | Faible | Faible | |
| # | Menace | Impact (calculé) Critère |
Impact (calculé) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | La portée est négligeable, parce qu’il y a de vastes étendues de milieux naturels où il n’y a pas de développement. La gravité est légère parce que les effets cumulatifs du développement résidentiel et commercial autour des centres urbains de l’ouest du Canada, en particulier dans les régions du sud situées à environ 200 km de la frontière des États-Unis entraînent souvent une perte complète de l’habitat. Cependant, on observe encore la sous-espèce occidentalis (p. ex., à Victoria, Delta, Edmonton, Regina). L’immédiateté est élevée, parce que la pratique est continue. | |
| 1.1 | Habitations et zones urbaines | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | Perte d’habitat résultant de l’urbanisation accrue, bien que cela ne touche qu’un faible pourcentage de l’aire de répartition de la sous-espèce. | |
| 1.2 | Zones commerciales et industrielles | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | Perte d’habitat résultant de l’urbanisation accrue, bien que cela ne touche qu’un faible pourcentage de l’aire de répartition de la sous-espèce. | |
| 1.3 | S.O. Le développement récréatif effectué dans une certaine mesure peut entraîner la perte d’habitat pour les bourdons, mais, dans l’ensemble, d’autres effets tangentiels peuvent influer sur l’habitat des bourdons (p. ex. l’utilisation de pesticides sur les terrains de golf, la dérivation des cours d’eau, la diminution des ressources florales, etc.). Ces menaces sont traitées ailleurs dans le tableau. | ||||||
| 2 | Agriculture et aquaculture (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | La portée est négligeable parce qu’il y a de vastes étendues de milieux naturels où l’on ne pratique pas l’agriculture. La gravité est légère, car dans certaines régions agricoles, les bourdons sont abondants et répandus. L’immédiateté est élevée, parce que les activités se poursuivent. | |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | L’intensification de l’agriculture dans les basses altitudes, en particulier dans les parties méridionales de l’aire de répartition (p. ex. C.-B. – Okanagan, Kootenay, île de Vancouver, basses-terres continentales; Alb. – Calgary et sud de l’Alb.). Bien que cette menace existe de tout temps, on a assisté à une intensification de l’agriculture au cours des 25 dernières années et celle-ci a entraîné une diminution de la qualité de l’habitat (p. ex. voir Javorek et Grant, 2011). Des habitats agricoles favorisant en quelque sorte les bourdons (p. ex. prairies de fauche et légumes-feuilles) sont remplacés par des cultures en serres fermées, des vignobles et autres systèmes agricoles qui pourraient ne pas avoir besoin de pollinisation par les insectes. | |
| 4 | Corridors de transport et de service (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | La portée est négligeable, parce qu’il y a de vastes étendues de milieux naturels où la construction de routes et de lignes de services publics n’est pas prévue. La gravité est négligeable parce que dans de nombreux cas, les corridors de transport peuvent laisser un habitat plus ouvert pour les bourdons (à condition que ces corridors ne soient pas pavés). L’immédiateté est élevée, parce que la pratique est continue. | |
| 4.1 | Routes et voies ferrées | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | La menace est jugée négligeable. Elle pourrait agrandir temporairement les habitats situés le long des routes où des zones doivent être nettoyées. Il pourrait cependant y avoir des impacts cumulatifs dus aux herbicides (cette menace est traitée ailleurs). | |
| 4.2 | Lignes de services publics | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | La menace est jugée négligeable. Elle pourrait agrandir temporairement les habitats situés le long des routes où des zones doivent être nettoyées. Il pourrait cependant y avoir des impacts cumulatifs dus aux herbicides (cette menace est traitée ailleurs). | |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | S.O. | |
| 5.3 | Exploitation forestière et récolte du bois | Négligeable | Négligeable (< 1 %) | Négligeable (< 1 %) | Élevée (continue) | La menace est jugée négligeable. L’exploitation forestière pourrait augmenter temporairement les habitats disponibles s’ils sont reliés entre eux. | |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | La portée est négligeable, parce qu’il y a de vastes étendues de milieux naturels où il n’y a pas d’activités récréatives en cours; la gravité est légère, parce des nids pourraient être piétinés lors d’activités récréatives ou celles-ci pourraient contribuer à dégrade les sites de nidification. L’immédiateté est élevée, parce que la pratique est continue. | |
| 6.1 | Activités récréatives | Négligeable | Négligeable (< 1 %) | Légère (1-10 %) | Élevée (continue) | La menace est jugée négligeable. Certaines activités récréatives pourraient nécessiter un enlèvement des nids à l’échelle locale, bien que dans toute l’aire de répartition de la sous-espèce, cette menace est vraisemblablement mineure dans l’ensemble. | |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | Faible | Faible (1-10 %) | Extrême (71-100 %) | Élevée (continue) | La portée est faible, car la propagation des espèces envahissantes se fait principalement dans les zones urbaines et agricoles du Canada. Les habitats naturels ne semblent pas contenir de bourdons non indigènes. La propagation et l’impact des agents pathogènes demeurent inconnus dans la majeure partie de l’aire de répartition. La gravité est extrême, parce que ces pratiques ont une incidence sur les populations de bourdons. L’immédiateté est élevée, parce que la pratique est continue. | |
| 8.1 | Espèces exotiques/non indigènes envahissantes | Faible | Faible (1-10 %) | Extrême (71-100 %) | Élevée (continue) | L’introduction du bourdon fébrile et son utilisation pour la pollinisation des cultures sous serre dans l’ouest du Canada pourraient avoir des répercussions sur les populations sauvages du bourdon de l’Ouest. Les populations de bourdons fébriles qui se sont échappées et établies pourraient déloger les bourdons indigènes des sites de nidification et leur faire concurrence pour les ressources florales. Les colonies introduites pourraient être un vecteur de la propagation d’agents pathogènes ou de maladies. On ne connaît pas la situation quant à l’établissement des populations sauvages de bourdons fébriles dans l’Ouest du Canada, mais cet établissement aurait vraisemblablement un impact négatif sur les espèces de bourdons indigènes, comme cela a été documenté ailleurs dans le monde pour d’autres espèces domestiquées (Williams et Osborne, 2009). Des études en laboratoire montrent que les espèces parasites Crithidia bombi et Nosema bombi (soupçonnée) ont des effets dévastateurs sur les reines qui fondent les colonies de bourdons, les ouvrières participant à la quête de nourriture et les colonies tout entières. (Brown et al., 2000, 2003; Otterstatter et al., 2005). | |
| 9 | Pollution (en anglais seulement) | Faible | Faible (1-10 %) | Élevée (31-70 %) | Élevée (continue) | La portée est faible, parce que l’habitat naturel comporte de vastes étendues où les pesticides ne sont pas utilisés. La gravité est élevée, parce que les répercussions des pesticides sont connues. L’immédiateté est élevée, parce que la pratique est continue. | |
| 9.3 | Effluents agricoles et forestiers | Faible | Faible (1-10 %) | Élevée (1-100 %) | Élevée (continue) | Les néonicotinoïdes représentent une menace particulière pour les bourdons (comparativement à d’autres pesticides), parce qu’ils sont dangereux même à des concentrations de quelques parties par milliard (ppb) (Agence des États-Unis pour la protection de l’environnement [EPA] 1994; Marletto et al., 2003). Les néonicotinoïdes sont des pesticides systémiques qui circulent dans toute la plante et qui atteignent le pollen et le nectar. | |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | Non calcule | Généralisée (71-10 %) | Inconnu | Élevée (continue) | La portée est généralisée, car les changements climatiques touchent l’ensemble de l’aire de répartition de l’espèce. On ne connaît pas la gravité, parce les répercussions ne sont pas étudiées à une grande échelle. L’immédiateté est élevée, parce que la menace est continue. | |
| 11.2 | Sécheresses | Non calcule | Généralisée (71-10 %) | Inconnu | Élevée (continue) | Les changements climatiques constituent une autre menace possible (Williams et Osborne, 2009). On sait que les espèces de bourdons ont une niche climatique étroite et sont plus vulnérables aux menaces extrinsèques (Williams et al., 2009). On ne connaît pas actuellement les tolérances climatiques du bourdon de l’Ouest. |
Classification des menaces d’après l’IUCN-CMP (International Union for Conservation of Nature - Conservation Measures Partnership), Salafsky et al. (2008).
La perte d’habitat attribuable à l’intensification de l’agriculture est continue dans toutes les parties méridionales de l’aire de répartition de la sous-espèce occidentalis en Colombie-Britannique, en Alberta et en Saskatchewan, où se trouvent certaines des régions les plus fortement urbanisées ou agricoles du Canada. L’essor de l’agriculture intensive au cours des dernières décennies a entraîné une réduction de la qualité de l’habitat d’alimentation des bourdons à l’échelle mondiale (voir par exemple Williams, 1989; Kosior et al., 2007). L’intensification de l’agriculture est jugée moins menaçante pour la sous-espèce mckayi. L’utilisation de produits chimiques (p. ex. de pesticides) dans les régions agricoles ajoute un facteur supplémentaire de menace pouvant être grave pour tous les bourdons. Ce point est traité plus loin dans la section sur la pollution (menace no 9).
La propagation d’agents pathogènes a provoqué des déclins prononcés chez de nombreuses espèces animales (Morton et al., 2004; Power et Mitchell, 2004), mais l’ampleur de cette menace pour les bourdons demeure à préciser. Le phénomène survient lorsqu’un agent pathogène se transmet d’une population-hôte réservoir gravement infectée à une population-hôte sympatrique non-réservoir (Power et Mitchell, 2004). L’utilisation d’espèces de bourdons commerciaux infectés (le bourdon fébrile au Canada) pour la pollinisation des cultures sous serre a été incriminée dans la propagation de pathogènes aux populations de bourdons sauvages se nourrissant à proximité des serres (Colla et al., 2006; Otterstatter et Thomson, 2008). Dans l’ouest du Canada, les serres qui utilisent des bourdons d’élevage pour les cultures en serre sont présentes principalement dans le sud de la Colombie-Britannique et, dans une moindre mesure, dans les régions Lacombe et Redcliff de l’Alberta. Les espèces de parasites en cause (Crithidia bombi) dans la propagation d’agents pathogènes aux bourdons sauvages, ou soupçonnées de l’être (Nosema bombi), ont des effets nuisibles sur les reines qui fondent les colonies, les ouvrières participant à la quête de nourriture et sur les colonies tout entières (Brown et al., 2000, 2003; Otterstatter et al., 2005). On trouve ces parasites chez plusieurs espèces de bourdons (Macfarlane, 1974; Macfarlane et al., 1995; Colla et al., 2006), mais on ignore toujours leur virulence chez le bourdon de l’Ouest sauvage. Quoi qu’il en soit, un lien a été établi entre l’utilisation accrue de bourdons par les serriculteurs au cours des dernières décennies et le déclin de certaines espèces du sous-genre Bombus, dont le B. affinis et le bourdon de l’Ouest (Thorpe et Shepherd, 2005; RNC, 2007; Evans et al., 2008).
Des relevés récents effectués en Alaska (Koch et Strange, 2012) ont révélé que les taux d’infection à Nosema chez la sous-espèce mckayi étaient équivalents, sinon légèrement plus élevés, que chez les populations de la sous-espèce occidentalis aux États-Unis. Les colonies de bourdons d’élevage sont moins utilisées dans le nord et la présence de taux plus élevés de Nosema pourrait correspondre aux niveaux naturels hôte-parasite (Koch et Strange, 2012) et pourrait ne pas être due à une propagation d’agents pathogènes provenant de serres.
L’introduction et l’usage dans l’ouest du Canada du bourdon fébrile (B. impatiens), qui est très efficace pour la pollinisation, pourraient avoir une incidence accrue sur les populations de bourdon de l’Ouest. Les bourdons fébriles importés pour usage dans des serres de la Colombie-Britannique sont renommés pour s’échapper des serres et se retrouver dans la nature, où ils pourraient s’avérer être des compétiteurs et une menace pour les populations sauvages en déclin (Ratti et Colla, 2010). Cette espèce pourrait déloger le bourdon de l’Ouest des habitats de nidification ou lui faire concurrence pour les ressources alimentaires. Elle peut également servir de vecteur aux agents pathogènes et aux maladies. On ne connaît pas la situation des populations sauvages de cette espèce de l’est introduite dans l’ouest du Canada, mais elles pourraient avoir une incidence nuisible sur les espèces indigènes, comme il a été observé dans le cadre d’autres travaux (Williams et Osborne, 2009).
Dans les paysages fortement agricoles, le bourdon fébrile est susceptible de livrer une concurrence pour le nectar à l’abeille européenne domestiquée (Apis mellifera L.) introduite au Canada. Comme il est extrêmement difficile d’étudier la compétition en nature (Thomson, 2006), on ne connaît pas les impacts sur le paysage agricole et, vu que l’abeille domestique est présente en Amérique du Nord depuis des centaines d’années, on voit difficilement comment elle pourrait être directement responsable des déclins récents observés chez le bourdon de l’Ouest.
On sait que la pollution causée par les produits agrochimiques (pesticides) a des impacts nuisibles sur les abeilles et d’autres insectes utiles (voir par exemple Whitehorn et al., 2012; Baron et al., 2014). De nombreux produits agrochimiques sont reconnus pour avoir des répercussions sur les abeilles, mais les néonicotinoïdes ont suscité beaucoup d’attention au cours des dernières décennies et ont été reconnus comme risque majeur pour les insectes pollinisateurs (Morandin et Winston 2003; Laycock et al., 2012; Whitehorn et al., 2012). À peu près au moment où l’on a commencé à remarquer un déclin du bourdon de l’Ouest et d’autres bourdons de ce sous-genre, le nouveau pesticide imidaclopride (un néonicotinoïde) était homologué pour usage aux États-Unis (1994) et au Canada (1995) (Cox, 2001; ARLA, 2001). Les néonicotinoïdes sont des pesticides systémiques qui circulent et s’accumulent dans la plante, y compris dans le pollen et le nectar. Ces pesticides sont nuisibles pour les abeilles (comparativement à d’autres pesticides) à des concentrations de l’ordre de quelques parties par milliard (ppb) (EPA, 1994; Marletto et al., 2003). L’imidaclopride est non létal pour les bourdons s’il est utilisé conformément au mode d’emploi (voir par exemple Tasei et al., 2001). Ses effets sur les bourdons n’ont toutefois été évalués que chez une espèce, B. impatiens, jugée représentative de toutes les espèces habitant l’Amérique du Nord. (Gels et al., 2002; Morandin et Winston, 2003). Une autre étude a démontré que les néonicotinoïdes avaient des effets nuisibles sur une espèce de bourdon européenne appartenant au même sous-genre que le bourdon de l’Ouest (Tasei et al., 2001). Diverses particularités du cycle vital du bourdon de l’Ouest (forte taille corporelle, émergence hâtive, longue durée du cycle de la colonie, etc.) rendent peut-être l’espèce plus vulnérable à l’accumulation de pesticides dans la colonie.
L’usage répandu des pesticides de la catégorie des néonicotinoïdes dans l’aire de répartition des deux sous-espèces n’a pas été quantifié et, par conséquent, on ne peut que spéculer sur leurs effets. Les néonicotinoïdes sont fréquemment utilisés sur les parcours de golf et les terres agricoles (Sur et Stork, 2003). Le traitement de zones étendues servant à la pratique du golf pourrait exposer les bourdons à de grandes quantités de pesticides dans des milieux qui autrement offriraient un habitat propice à ces insectes (Tanner et Gange, 2004).
Les changements climatiques sont une autre menace possible pour les bourdons (Williams et Osborne, 2009). Les espèces de bourdons qui possèdent une niche climatique étroite sont plus vulnérables aux menaces extrinsèques (Williams et al., 2009).
Les bourdons ont un besoin constant de ressources florales durant la saison de végétation pour soutenir la croissance de la colonie. Sans ces ressources, l’émergence des reines, la production d’ouvrières et la croissance de la colonie pourraient être entravées. La présence de ressources alimentaires abondantes durant toute la saison permet aux populations locales de subsister. Seules les reines fécondées hivernent, alors un manque de ressources florales tôt dans la saison entraînera la mort de colonies ou la dispersion des reines nouvellement émergées.
Les bourdons sont des organismes haplodiploïdes chez qui le déterminisme complémentaire du sexe accroît considérablement le risque d’extinction lorsque la taille efficace de la population diminue (Zayed et Packer, 2005). Cette situation est le fait d’une spirale d’extinction induite par les mâles diploïdes (Zayed et Packer, 2005). Chez les bourdons et la plupart des autres organismes haplodiploïdes, le sexe est déterminé par le génotype à locus unique : les hémizygotes (haploïdes) sont des mâles, les hétérozygotes sont des femelles, et les homozygotes sont des mâles diploïdes. Les mâles diploïdes sont habituellement stériles. Le nombre d’allèles codeurs du sexe dans une population détermine la proportion d’individus diploïdes qui sont des mâles et dépend lui-même principalement de la taille efficace de la population. En d’autres mots, lorsque la taille d’une population de bourdons s’amenuise, la fréquence des mâles diploïdes augmente. Comme les mâles diploïdes sont stériles ou inviables, leur production accrue dans une population de taille réduite a pour effet d’accélérer le déclin de la population et détermine du coup une forme particulière de spirale d’extinction induite par les mâles diploïdes. Ce type particulier de fardeau génétique est le plus lourd connu (Hedrick et al., 2006). En pratique, lorsqu’une population de bourdons décline au point de ne plus compter que quelques individus reproducteurs, elle est inévitablement vouée à disparaître même si les conditions environnementales demeurent stables, à moins qu’elle augmente subitement en l’espace de quelques générations.
Aucune loi fédérale ou provinciale ne permet de protéger le bourdon de l’Ouest, d’atténuer les menaces qui pèsent sur ce groupe ou de protéger les sites de nidification ou l’habitat de l’espèce.
Certaines provinces canadiennes envisagent d’imposer par la loi une interdiction d’utiliser les pesticides de la catégorie des néonicotinoïdes, ce qui permettrait d’en atténuer la menace pour les bourdons (voir la section sur la pollution parmi les menaces). À l’heure actuelle, aucun des gouvernements des territoires dans l’aire de répartition du bourdon de l’Ouest n’a proposé de mesures législatives.
Les cotes de conservation provinciales et territoriales fondées sur les critères établis par NatureServe seront incluses dans le rapport Espèces sauvages 2015 (Hebert, comm. pers., 2013). Les cotes de conservation nationales pour le Canada (Conseil canadien pour la conservation des espèces en péril [CCSEP, 2011]) sont les suivantes : Colombie-Britannique et Alberta – N3 (sensible); Yukon et Saskatchewab – N5 (indéterminée). La cote de conservation à l’échelle mondiale est GU (non cotée en raison du manque d’information) (NatureServe 2013). Dans son évaluation aux fins de sa liste rouge, la Xerces Society for Invertebrate Conservation (2013) a attribué à l’espèce la cote « en péril ». Dans la Liste rouge de l’UICN (2013), l’espèce est désignée n’avoir aucune protection. L’espèce n’a pas été évaluée ni classée sous le régime de la Federal Endangered Species Act des États-Unis.
Dans leur étude sur le déclin général du bourdon à l’échelle mondiale, Williams et Osborne (2009) évaluent la situation du bourdon fébrile (y compris son espèce occidentale apparentée, le bourdon de l’Ouest) comme espèce en danger selon les critères suivants de l’IUCN : A2 : réduction de la population depuis 1995 > 50 % (présumée); les causes de la réduction ne sont peut-être pas réversibles ou n’ont peut-être pas cessé, en se basant sur le faible taux d’observations de spécimens au cours des quatre dernières années, au moins dans le quart sud-ouest de son aire de répartition.
Le déclin rapide du bourdon de l’Ouest et d’autres espèces du sous-genre Bombus s. str. semble s’être amorcé au milieu des années 1990 (RNC, 2007). Le B. franklini a disparu de son aire dans l’ouest des États-Unis et figure sur la liste des espèces gravement menacées d’extinction de l’UICN (Evan et al., 2008). Bombus affinis a été récemment désigné comme espèce en voie de disparition par le COSEPAC au Canada. Bombus terricola semble également sur le déclin (RNC, 2007; Evans et al., 2008). Les facteurs responsables de ces déclins demeurent inconnus (RNC, 2007; Evans et al., 2008).
Aucune analyse n’a été effectuée pour déterminer le statut de propriété de l’aire de répartition de l’espèce. La majeure partie des terres situées dans les régions peuplées adjacentes à la frontière internationale appartiennent à des intérêts privés. Ce sont soit des zones urbaines, soit des terres consacrées à une agriculture intensive.
L’aire de répartition du bourdon de l’Ouest au Canada s’étend sur plusieurs parcs provinciaux et nationaux et zones protégées. Les observations récentes (depuis 2013) ont été effectuées notamment dans les zones protégées suivantes :
- Colombie-Britannique : La sous-espèce occidentalis a été enregistrée dans des parcs gouvernementaux locaux : au parc régional de la baie Boundary (2010, agglomération de Vancouver), parc du mont Douglas (2012, Saanich), parc du mont Tolmie (2012, Saanich), sanctuaire de la nature de Christmas Hill (2012, Saanich), Parc Highrock (Cairn) (2012, Esquimalt), (jardins) de Brentwood Bay (2012, Central Saanich). Parcs provinciaux : le parc provincial Manning. Parcs fédéraux : parc national du Mont-Revelstoke et parc national Yoho. En 2013, des relevés ont été faits aux parcs provinciaux situés dans l’aire de répartition de la sous-espèce mckayi; les données n’ont toutefois pas encore été analysées (Sheffield, comm. pers., 2013).
- Alberta : La sous-espèce occidentalis a été signalée au parc interprovincial des collines-Cypress (Alberta et Saskatchewan), au parc national Banff et au parc provincial des Dinosaures.
- Saskatchewan : La sous-espèce occidentalis a été observée au parc interprovincial des collines-Cypress (Alberta et Saskatchewan.) et au parc national des Prairies.
- Territoires du Nord-Ouest : La sous-espèce mckayi a été observée à la Réserve du parc national Nahanni et à la réserve du parc national Nááts’ihch’oh.
- Yukon : Aucun signalement dans les parcs ou les aires protégées, bien que la sous-espèce mckayi y vive vraisemblablement.
Nous remercions les personnes et les établissements suivants qui nous ont fourni des données et des spécimens aux fins de la présente évaluation : Elizabeth Elle (Université Simon Fraser), Xerces Society for Invertebrate Conservation, Jamie Strange (USDA), Jonathan Koch (Utah State University), John Ascher (American Museum of Natural History), Ralph Cartar (Université de Calgary), Robin Owen (Collège Mont-Royal), Lincoln Best (Université York), Cory Sheffield (Université York), Jason Gibbs (Université York), Jennifer Heron (BC Ministry of Environment), Syd Cannings (Environnement Canada), Rob Longair (Université de Calgary), David Larson, Katrina Stipec (BC Ministry of Environment), Alison Leslie, Claudia Ratti (York University), Dave Fraser (BC Ministry of Environment), Julie Wray (Université Simon Fraser), Leif Richardson (Dartmouth College). Nous voulons remercier tout spécialement Ilona Naujokaitis-Lewis (Université de Toronto) qui a analysé les données spatiales.
Photographie de la page couverture : David Inouye. Ouvrière de bourdon de l’Ouest en train de butiner le nectar d’une fleur d’Ipomopsis.
Une bonne partie des données récentes (2013) proviennent de projets financés par Earth Rangers; d’autres fonds ont été accordés à C. Sheffield (Royal Saskatchewan Museum). Le Service canadien de la faune et le ministre de l’Environnement de la Colombie-Britannique ont fourni des ressources et du personnel.
- Alford, D.V. 1975. Bumble bees. London: Davis-Poynter.
- Agence de réglementation de la lutte antiparasitaire (ARLA). 2001. Imidacloprid, Note réglementaire REG2001-11, Ottawa, Santé Canada, Agence de réglementation de la lutte antiparasitaire, (Format PDF, 65 Ko.)
- Ascher, J., et J. Pickering. 2013. Discover Life’s bee species guide and world checklist, Species account for Bombus occidentalis. (consulté le 12 novembre 2013; en anglais seulement).
- Baron, G.L., N.E. Raine, M.J.F. Brown. 2014. Impact of chronic exposure to a pyrethroid pesticide on bumblebees and interactions with a trypanosome parasite, Journal of Applied Ecology, doi: 10.1111/1365-2664.12205.
- Benton, T. 2006. Bumblebees, Harper-Collins, Royaume Uni.
- Bertsch, A., M. Hrabé de Angelis et G.K.H. Przemeck. 2010. A phylogenetic framework for the North American bumblebee species of the subgenus Bombus sensu stricto (Bombus affinis, B. franklini, B. monderatus, B. occidentalis and B. terricola) based on mitochondrial DNA markers (Hymenoptera: Apidae: Bombus), Beiträge zur Entomologie 60:229 242.
- Best, L. 2010. Données personnelles. York University, Toronto (Ontario).
- Breed, M.D., E. Guzman-Novoa et G.J. Hunt. 2004. Defensive Behavior of honey bees: Organization, Genetics, and Comparisons with Other Bees, Annual Review of Entomology 49:271 298.
- Brown M.J.F., R. Loosli et P. Schmid-Hempel. 2000. Condition-dependent expression of virulence in a trypanosome infecting bumble bees, Oikos 91:421 427.
- Brown M., R. Schmid-Hempel et P. Schmid-Hempel. 2003. Strong context-dependent virulence in a host-parasite system: reconciling genetic evidence with theory, Journal of Animal Ecology 72: 994 1002.
- Cameron, S.A., J.D. Lozier, J.P. Strange, J.B. Koch, N. Cordes, L.F. Solter et T. Griswold. 2011. Patterns of widespread decline in North America bumble bees, Proceedings of the National Academy of Science 108:662 667.
- Cameron, S.A., H.M. Hines et P. Williams. 2007: A comprehensive phylogeny of the bumble bees (Bombus), Biological Journal of the Linnean Society 91:161 188.
- Carriere, S. 2014. Ministère de l’Environnement et des Ressources naturelles (ERN) Gouvernement des Territoires du Nord-Ouest, Yellowknife (Territoires du Nord Ouest), communication personnelle adressée à J. Heron.
- Colla, S.R., M.C. Otterstatter, R.J. Gegear et J.D. Thomson. 2006. Plight of the bumble bee: Pathogen spillover from commercial to wild populations, Biological Conservation 129:461 467.
- Colla, S.R., et L. Packer. 2008: Evidence for decline in eastern North American bumble bees (Hymenoptera: Apidae), with special focus on Bombus affinis Cresson, Biodiversity and Conservation 17:1379 1391.
- Colla, S.R., et C.M. Ratti. 2010. Evidence for the decline of Western Bumble Bee (Bombus occidentalis Greene) in British Columbia, Pan-Pacific Entomologist 86:32-34.
- COSEPAC. 2010. Évaluation et Rapport de situation du COSEPAC sur le bourdon à tache rousse (Bombus affinis) au Canada, Comité sur la situation des espèces en péril au Canada, Ottawa.
- Cox, C. 2001. Insecticide factsheet: Imidacloprid, Journal of Pesticide Reform 21:15 22.
- Environmental Protection Agency (EPA). 1994. Pesticide fact sheet: Imidacloprid, Washington D.C., 18 mars.
- Evans, E., R. Thorp, S. Jepson et S.H. Black. 2008. Status review of three formerly common species of bumble bee in the subgenus Bombus, (Format PDF, 830 Ko.), établi pour la Xerces Society for Invertebrate Conservation.
- Fitzpatrick, U., T.E. Murray, R.J. Paxton, J. Breen, D. Cotton, V. Santorum et M.J.F. Brown. 2007. Rarity and decline in bumble bees – A test of causes and correlates in the Irish fauna, Biological Conservation 136:185 194.
- Gardner, A. 2007. The changing landscape of the southern Alberta foothills, Southern Foothills Study.
- Gels, J.A., D.W. Held et D.A. Potter. 2002. Hazards of insecticides to the bumble bees Bombus impatiens (Hymenoptera: Apidae) foraging on flowering white clover in turf, Journal of Economic Entomology 95:722 728.
- Gillespie, S. 2010. Factors affecting parasite prevalence among wild bumble bees, Ecological Entomology 35:737 747.
- Goulson, D. 2003. Bumblebees, Their Behaviour and Ecology, Oxford University Press, Oxford, 235 p.
- Greef, M., et P. Schmid-Hempel. 2008. Sperm reduces female longevity and increases melanization of the spermatheca in the bumble bee Bombus terrestris, Insectes Sociaux 55:313 319.
- Greene, J.W. 1858. Descriptions of several new hymenopterous insects from the northwest coast of America, Annals of Lyceum of Natural History of New York 7:11 12.
- Hatfield, R.G., et G. LeBuhn. 2007. Patch and landscape factors shape community assemblages of bumble bees, Bombus spp. (Hymenoptera:Apidae), in montane meadows, Biological Conservation 139:150 158.
- Heinrich, B. 2004. Bumblebee Economics, Harvard University Press, États Unis, 245 p.
- Heron, J. 2013. BC Ministry of Environment, Vancouver (Colombie Britannique), communication personnelle adressée à Sheila Colla.
- Hobbs, G.A. 1968. Ecology of species of Bombus Latr. (Hymenoptera: Apidae) in southern Alberta, VI. Subgenus Bombus, Canadian Entomologist 100:156 164.
- Javorek, S.K., et M.C. Grant. 2011. Tendances de la capacité d’habitat faunique des terres agricoles du Canada, de 1986 à 2006, Biodiversité canadienne : état et tendances des écosystèmes en 2010, Rapport technique thématique nº 14, Conseils canadiens des ministres des ressources, Ottawa (Ontario), vi + 51 p.
- Knopp, D., L. Larkin et J. Heron. 2010. Surveys for Dun Skipper (Euphyes vestris) and Western Bumble Bee (Bombus occidentalis) in the Lower Fraser Valley, B.C., B.C. Ministry of Environment, Terrestrial Conservation Science Section, UBC Office, 315 – 2202 Main Mall, Vancouver (Colombie Britannique), 53 p.
- Koch, J.B., et J. Strange. 2009. Constructing a species database and historic range maps for North American bumble bees (Bombus sensu stricto Latrielle) to inform conservation decisions, Uludag Bee Journal 9:97 108.
- Koch, J.B., et J.P. Strange. 2012. The status of Bombus occidentalis and B. moderatus in Alaska with special focus on Nosema bombi incidence, Northwest Science 86:212-220.
- Kosoir, A., W. Celary, P. Olejniczak, J. Fijal, W. Krol, W. Solarz et P. Plonka. 2007. The decline of the bumble bees and cuckoo bees (Hymenoptera: Apidae: Bombini) of Western and Central Europe, Oryx 41:79 88.
- Kraus, F.B., S. Wolf et R.F.A. Moritz. 2008. Male flight distance and population substructure in the bumble bee, Bombus terrestris, Journal of Animal Ecology 78:247 252.
- Laycock, I., K.M. Lenthall, A.T. Barratt et J.E. Cresswell. 2012. Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris), Ecotoxicology, 21(7), 1937 1945.
- Lilley, P., N. Page et J. Heron. 2010. Surveys for butterfly species at risk on private and municipal lands on southeastern Vancouver Island and the Gulf Islands, British Columbia (2010), B.C. Ministry of Environment, Terrestrial Conservation Science Section, UBC Office, 315 – 2202 Main Mall, Vancouver (Colombie Britannique).
- Longair, R. 2010. University of Calgary, Calgary (Alberta), communication personnelle adressée à Sheila Colla.
- Macfarlane, R.P. 1974. Ecology of Bombinae (Hymenoptera: Apidae) of Southern Ontario, with emphasis on their natural enemies and relationships with flowers, thèse de doctorat, University of Guelph, Guelph (Ontario), Canada.
- Macfarlane, R.P., J.J. Lipa et H.J. Liu. 1995. Bumble bee pathogens and internal enemies, Bee World 76:130 148.
- Macfarlane, R.P., et K.D. Patten. 1997. Food sources in the management of bumble bee populations around cranberry marshes, in Proceedings of the 7th International Symposium on Pollination, K.W. Richards (éd.), Acta Horticulturae 239 244.
- MacKenzie K.E., et M.L. Winston. 1984. Diversity and abundance of native bee pollinators on berry crops and natural vegetation in the lower Fraser valley, British Columbia, Canadian Entomologist 116:965 974.
- Marks, D., et J. Heron. 2010. Surveys for Western Bumble Bee (Bombus occidentalis) and other arthropod species at risk on private and municipal lands in the south Okanagan – Similkameen River valleys, British Columbia 2010, B.C. Ministry of Environment, Terrestrial Conservation Science Section, UBC Office, 315 – 2202 Main Mall, Vancouver (Colombie Britannique).
- Marletto, F., A. Patetta et A. Manino. 2003. Laboratory assessment of pesticide toxicity to bumble bees, Bulletin of Insectology 56:155 158.
- Milliron, H.E. 1971: A monograph of the western hemisphere bumble bees (Hymenoptera: Apidae; Bombinae), I. The genera Bombus and Megabombus Subgenus Bombias, Mémoires de la Société entomologique du Canada / Memoirs of the Entomological Society of Canada 82:1 80.
- Morandin, L.A., et M.L. Winston. 2003. Effects of novel pesticides on bumble bee (Hymenoptera: Apidae) colony health and foraging ability, Community and Ecosystem Ecology 32:555 563.
- Morton, A., Routledge, R.C. Peet et A. Ladwig. 2004. Sea lice infection rates on juvenile pink and chum salmon in the nearshore environment of British Columbia, Canada, Journal canadien des sciences halieutiques et aquatiques / Canadian Journal of Fish and Aquatic Sciences 61:147 158.
- National Research Council (NRC). 2007. Status of Pollinators in North America, The National Academies Press, Washington DC.
- Otterstatter, M.C. 2001. The incidence of parasitism by conopid and phorid flies and its effects on the behavior of bumble bees (Hymenoptera: Apidae), mémoire de maîtrise ès sciences, University of Calgary.
- Otterstatter, M.C. 2004. Patterns of parasitism among conopid flies parasitizing bumble bees, Entomologia Experimentalis et Applicata 111:133 139.
- Otterstatter, M.C., R.J. Gegear, S.R. Colla et J.D. Thomson. 2005. Effects of parasitic mites and protozoa on the flower constancy and foraging rate of bumble bees, Behavioral Ecology and Sociobiology 58:383 389.
- Otterstatter, M.C., et T.L. Whidden. 2004. Patterns of parasitism by tracheal mites (Locustacarus buchneri) in natural bumble bee populations, Apidologie 35:351 357.
- Otterstatter, M.C., T.L. Whidden et R.E. Owen. 2002. Contrasting frequencies of parasitism and host mortality among phorid and conopid parasitoids of bumble-bees, Ecological Entomology 27:229 237.
- Otterstatter, M.C., et J.D. Thomson. 2008 Does Pathogen Spillover from Commercially Reared Bumble Bees Threaten Wild Pollinators? PLoS One 3: e2771.
- Owen, R.E. 1988. Body size variation and optimal body size of bumble bee queens (Hymenoptera: Apidae), Canadian Entomologist 120:19 27.
- Owen, R.E. 2010. Mount Royal College, Calgary (Alberta), communication personnelle adressée à Sheila Colla.
- Owen, R.E., T.L. Whidden. 2013. Discrimination of the bumble bee species Bombus occidentalis Greene and B. terricola Kirby by morphometric, colour and RAPD variation, Zootaxa 3608:328 344.
- Packer, L., et R. Owen. 2001. Population genetic aspects of pollinator decline, Ecology and Society 5:4.
- Parkinson, L., et J. Heron. 2010. Surveys for Western Bumble Bee (Bombus occidentalis) and other arthropod species at risk in the lower mainland, British Columbia (2010), B.C. Ministry of Environment, Terrestrial Conservation Science Section, UBC Office, 315 - 2202 Main Mall, Vancouver (Colombie Britannique).
- Patten, K.D., C.H. Shanks et D.F. Mayer. 1993. Evaluation of herbaceous plants for attractiveness to bumble bees for use near cranberry farms, Journal of Apicultural Research 32:73 79.
- Power, A.G., et C.E. Mitchell. 2004. Pathogen Spillover in Disease Epidemics, American Naturalist 164:S79 S89.
- Rasheed, S.A., et L.D. Harder. 1997. Economic motivation for plant species preferences of pollen-collecting bumble bees, Ecological Entomology 22:209 219.
- Ratti, C.M. 2006. Bee abundance and diversity in berry agriculture, mémoire de maîtrise ès sciences, Simon Fraser University.
- Ratti, C.M., H.A. Higo, T.L. Griswold et M.L. Winston. 2008. Bumble bees influence berry size in commercial Vaccinium spp. cultivation in British Columbia, The Canadian Entomologist 140:348 363.
- Ratti, C.M., et S.R. Colla. 2010. Discussion of the presence of an eastern bumble bee species (Bombus impatiens Cresson) in western Canada, Pan-Pacific Entomologist 86:29 31.
- Richards, K.W. 1978. Nest site selection by bumble bees (Hymenoptera: Apidae) in southern Alberta, The Canadian Entomologist 110:301 318.
- Schmid-Hempel, P. 1998. Parasites in social insects, Princeton University Press, Princeton (New Jersey).
- Sheffield, Cory. 2013. Royal Saskatchewan Museum, Regina (Saskatchewan), communication personnelle adressée à J. Heron.
- Stephen, W.P. 1957. Bumble Bees of Western America, Oregon State College, Agricultural Experiment Station, Technical Bulletin 40.
- Stephen, W.P., G.E. Bohart et P.F. Torchio. 1969. The Biology and External Morphology of Bees, with a Synopsis of the Genera of Northwestern America, Agricultural Experiment Station, Oregon State University, Corvallis (Oregon).
- Stephen, W.P., et T. Koontz. 1973a. larvae of the Bombini, I. Interspecific variation in larval head characteristics of Bombus (Humenoptera: Apoidea), Melanderia 13:1 12.
- Stephen, W.P., et T. Koontz. 1973b. Larvae of the Bombini, II. Developmental changes in the preadult stages of Bombus griseocollis (Degeer), Melanderia 13:13 29.
- Stephen, W.P., et T. Koontz. 1973. Larvae of the Bombini, I. Interspecific variation in larval head characteristics of Bombus (Humenoptera: Apoidea), Melanderia 13:1 12.
- Stephen, W.P., et T. Koontz. 1973. Larvae of the Bombini, II. Developmental changes in the preadult stages of Bombus griseocollis (Degeer), Melanderia 13:13 29.
- Stotyn, S., et D. Tate. 2012. Wild Bumble Bees of the Nahanni River Region, Northwest Territories, rapport présenté à l’Agence Parcs Canada, Réserve de parc national du Canada Nahanni, 9 p.
- Strange, J. 2010. United States Department of Agriculture, Bee Lab (Utah), communication personnelle adressée à Sheila Colla.
- Stout, J.C., et D. Goulson. 2000. Bumble bees in Tasmania: their distribution and potential impact on Australian flora and fauna, Bee World 81:80 86.
- Strong, W. 1992. Ecoregions of Alberta, Alberta Forestry, Lands and Wildlife, Edmonton (Alberta).
- Sur, R., et A. Stork. 2003. Uptake, translocation and metabolism of imidacloprid in plants, Bulletin of Insectology 1:35 40.
- Tanner, R.A., et A.C. Gange, 2004. Effects of golf courses on local biodiversity, Landscape and Urban Planning 71:137 146.
- Tasei, J.N., G. Ripault et E. Rivault. 2001. Hazards of Imidacloprid seed coating to Bombus terrestris (Hymenoptera: Apidae) when applied to Sunflower, Journal of Economic Entomology 94:623 627.
- Thomson, D.M. 2006. Detecting the effects of introduced species: a case study of competition between Apis and Bombus, Oikos 114:407 418.
- Thorp, R.W., D.S Horning et L.L. Dunning. 1983. Bumble bees and cuckoo bumble bees of California (Hymenoptera: Apidae), Bulletin of the California Insect Survey 23:viii.
- Thorp, R.W., et M.D. Shepherd. 2005. Subgenus Bombus Latreille 1802 (Apidae: Apinae: Bombini), in Shepherd, M.D., D.M. Vaughan et S.H. Black (éd.), Red List of Pollinator Insects of North America, CD-ROM, version 1 (mai 2005), Portland (Orégon), The Xerces Society for Invertebrate Conservation, (consulté le 12 novembre 2013; en anglais seulement).
- Turner, N. 1975. Food plants of British Columbia Indians Part 1 Coastal Peoples, Handbook 34, British Columbia Provincial Museum, Victoria (Colombie Britannique), 264 p.
- United States Department of Agriculture (USDA). 2010. U.S. National Pollinating Insects Database, Agriculture Research Service, Bee Biology and Systematics Laboratory, Logan (Utah) (base consultée le 11 novembre 2010).
- Westcott, L., et J. Heron. 2010. Surveys for Western Bumble Bee (Bombus occidentalis) and other arthropod species at risk on private and municipal lands in the West Kootenay region, British Columbia, August 2010, B.C. Ministry of Environment, Terrestrial Conservation Science Section, UBC Office, 315 – 2202 Main Mall, Vancouver (Colombie Britannique).
- Whidden, T., et R.E. Owen. 2011. Frequencies of diploid males in natural populations of three North American Bumble Bee (Bombus) species (Hymenoptera:Apidae), Annals of the Entomological Society of America 104:83 87.
- Whitehorn, P.R., S. O’Connor, F.L. Wackers et D. Goulson. 2012. Neonicotinoid pesticide reduces bumble bee colony growth and queen production, Science 336(6079), 351 352.
- Whittington, R., et M.L. Winston. 2003. Effects of Nosema bombi and its treatment fumagillin on bumble bee Bombus occidentalis colonies, Journal of Invertebrate Pathology 84:54 58.
- Whittington, R., et M.L. Winston. 2003. Are bumble bee colonies in tomato greenhourses obtaining adequate nutrition? The Canadian Entomologist 135:883 892.
- Williams, P.H. 2010. Bombus of the World, (en anglais seulement), consulté le 30 novembre 2010.
- Williams, P.H., et J.L. Osborne. 2009. Bumble bee vulnerability and conservation world-wide, Apidologie 40:367 387.
- Williams, P.H., S.R. Colla et Z. Xie. 2009. Bumble bee vulnerability: common correlates of winners and losers across three continents, Conservation Biology 23:931 940.
- Williams, P.H., R.W. Thorp, L.L. Richardson et S.R. Colla. 2014. Bumble Bees of North America: An Identification Guide, 208 p.
- Williams, P.H. 1989. Bumblebees and their decline in Britain, Ilford: Central Association of Bee-Keepers, 15 p.
- Williams, P.H., M.J.F. Brown, J.C. Carolan, D. Goulson, An, Jiandong, A.M. Aytekin, L.R. Best, A.M. Byvaltsev, B. Cederberg, R. Dawson, J. Huang, M. Ito, A. Monfared, R.H. Raina, P. Schmid-Hempel, C.S. Sheffield, P. Šima et Z. Xie. 2012. Unveiling cryptic species of the bumblebee subgenus Bombus s. str. worldwide with COI barcodes (Hymenoptera: Apidae), Systematics and Biodiversity 10:21 56, 10.1080/14772000.2012.664574.
- Winston, M.L., et L.H. Graf. 1982. Native bee pollinators of berry crops in the Fraser Valley of British Columbia, Journal of the Entomological Society of British Columbia 79:14 20.
- Wray, J. 2013. Simon Fraser University, Burnaby (Colombie Britannique), communication personnelle adressée à Jennifer Heron.
- Zayed, A., et L. Packer. 2005. Complementary sex determination substantially increases extinction proneness of haplodiploid populations, Proceedings of the National Academy of Sciences 102:10742 10746.
Sheila R. Colla a étudié divers aspects de l’écologie et du comportement des bourdons dans différentes régions de l’Amérique du Nord. À titre d’adjointe à la recherche pour James Thomson (Ph.D.), Michael Otterstatter (Ph.D.) et Robert Gegear (Ph.D.), de l’Université de Toronto, Campus St. George, elle a participé à un projet de recherche sur la propagation de pathogènes des populations de bourdons d’élevage aux populations de bourdons sauvages. Titulaire d’une bourse d’études supérieures du Canada Alexander-Graham-Bell, elle poursuit actuellement des études doctorales à l’Université York, à Toronto (Ontario), sous la supervision de Laurence Packer (Ph.D.). Ses travaux portent sur l’évolution de la communauté de bourdons au cours des cent dernières années et sur certaines des causes des déclins observés. Sheila R. Colla participe également à la Campagne pour la protection des pollinisateurs en Amérique du Nord. Les résultats de ses travaux ont fait l’objet d’articles dans les quotidiens et revues le Washington Post, Canadian Gardening, le Toronto Star et BioScience et ont été présentés aux émissions Quirks and Quarks de la CBC et The Daily Planet de Discovery Channel Canada.
Michael C. Otterstatter s’intéresse aux bourdons et à leurs parasites depuis 1997. Il travaille en collaboration avec des chercheurs renommés spécialisés dans les bourdons, dont Robin Owen, Lawrence Harder, James Thomson, Robert Gegear et Sheila Colla. Michael a effectué plusieurs études sur les bourdons de l’Alberta, ciblées entre autres sur les mouches et les acariens parasites (travaux effectués dans le cadre de sa maîtrise), et sur les bourdons du sud de l’Ontario, axées particulièrement sur les protozoaires parasites et la propagation de maladies des populations d’élevage aux populations sauvages (travaux effectués dans le cadre de son doctorat). Ces derniers travaux ont été publiés dans le New Scientist, Scientific American et le New York Times. D’autres travaux ont porté sur l’étude des ennemis naturels du bourdon dans le sud du Chili. Michael travaille actuellement comme épidémiologiste et collabore avec divers collaborateurs à des projets sur la conservation des bourdons.
Leif L. Richardson et un étudiant au doctorat au Dartmouth College, Hanover (New Hampshire). Dans le cadre de ses travaux, il examine les effets de la chimie du nectar des fleurs sur les abeilles et leurs parasites. De plus, il intègre les résultats des relevés avec les données sur les collections des musées pour étudier les changements dans la répartition des bourdons d’Amérique du Nord. En collaboration avec Sheila R. Colla et d’autres coauteurs, il a rédigé un guide des bourdons d’Amérique du Nord qui sera publié par la maison d’édition Princeton University Press en 2014.
Cory S. Sheffield étudie les abeilles et la pollinisation depuis 1993, alors qu’il était étudiant au baccalauréat spécialisé à l’Université Acadia, à Wolfville (Nouvelle-Écosse). Il a ensuite mené des études de deuxième cycle (M.Sc.) sur les interactions entre les insectes et les plantes à l’Université Acadia, ainsi qu’à Agriculture et Agroalimentaire Canada (AAC), à Kentville (Nouvelle-Écosse), de 1994 à 2006. Cory a fait son troisième cycle (Ph. D.) à l’Université de Guelph, en Ontario, tout en continuant à travailler à AAC. Il a étudié les abeilles de la Nouvelle-Écosse, notamment leur diversité et leur rôle dans la pollinisation des cultures. Durant cette période, il a publié, en collaboration avec plusieurs auteurs, des articles portant sur la redécouverte d’Epeoloides pilosulus en Nouvelle-Écosse, alors qu’on pensait que l’espèce avait disparu dans la province. Cory a ensuite entrepris des études postdoctorales à l’Université York (Ontario) sur la taxonomie des abeilles et le codage à barres de l’ADN, puis il a occupé un poste d’associé de recherche sur la taxonomie des abeilles auprès de l’Initiative de pollinisation canadienne (CANPOLIN). Il est actuellement chercheur scientifique et conservateur au service de la zoologie des invertébrés au Royal Saskatchewan Museum, à Regina (Saskatchewan). Ses travaux de recherche portent encore sur les abeilles; il a publié des articles sur la taxonomie des abeilles du Canada et de l’Amérique du Nord, l’utilité des codes à barres ADN pour les abeilles, la physiologie des abeilles, le rôle des abeilles dans la pollinisation et la diversité des abeilles du Canada.
Des données provenant de nombreuses collections nord-américaines ont servi à l’analyse présentée dans le présent rapport. Plus de 70 personnes et institutions ont contribué à rassembler les données. Vous en trouverez la liste sur le site suivant, www.leifrichardson.org/bbna.html.