
Bruant à face noire (Zonotrichia querula) : évaluation et rapport de situation du COSEPAC 2017
Bruant à face noire

Préoccupante
2017
Table des matières
- Table des matières
- COSEPAC sommaire de l’évaluation
- COSEPAC résumé
- Résumé technique
- Description et importance de l’espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Catégorie 9.3 : Effluents agricoles et sylvicoles (impact faible à élevé)
- Catégorie 1.1 : Zones résidentielles et urbaines (impact faible)
- Catégorie 4 : Corridors de transport et de service (impact faible)
- Catégorie 8.1 : Espèces exotiques (non indigènes) envahissantes (impact faible)
- Menaces négligeables et inconnues
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d’information
- Sommaire biographique du rédacteur du rapport
- Collections examinées
Liste des figures
- Figure 1. Répartition mondiale du Bruant à face noire en période de reproduction (gris pâle; adaptation à partir de James et al., 1976; Cadman, 2007) et d’hivernage (gris foncé; adaptation à partir de National Audubon Society, 2015; eBird, 2016; Norment et al., 2016), y compris les occurrences irrégulières (lignes tiretées).
- Figure 2. Indices de nidification du Bruant à face noire relevés durant l’élaboration de l’Atlas des oiseaux nicheurs du Manitoba, 2010-2014 (MMBA, 2015).
- Figure 3. Indices de nidification du Bruant à face noire relevés durant l’élaboration du deuxième (2001-2005) Atlas des oiseaux nicheurs de l’Ontario, 2010-2014 (Cadman et al., 1987 et 2007).
- Figure 4. Tendances annuelles des observations de Bruants à face noire (de 1980 à 2014) d’après les données du Recensement des oiseaux de Noël (National Audubon Society, 2015) aux États-Unis, au Kansas, au Nebraska, en Oklahoma et au Texas.
- Figure 5. Mentions de Bruants à face noire dans eBird (eBird, 2016) de décembre à février (de 2004 à 2014) dans les États américains faisant partie de l’aire de répartition principale en dehors de la période de reproduction.
- Figure 6. Tendances des effectifs de Bruants à face noire au printemps (de 1994 à 2014) à l’observatoire du lac de la Dernière-Montagne.
- Figure 7. Tendances des effectifs de Bruants à face noire à l’automne (de 1993 à 2014) à l’observatoire d’oiseaux du lac des Deux-Montagnes.
- Figure 8. Tendances des effectifs de Bruants à face noire au printemps (de 2006 à 2010) à l’observatoire d’oiseaux du marais Delta.
- Figure 9. Tendances des effectifs de Bruants à face noire à l’automne (de 1993 à 2010) à l’observatoire d’oiseaux du marais Delta.
Liste des tableaux
- Tableau 1. Résumé des tendances de la population de Bruants à face noire sur les 10 et les 35 dernières années, d’après le Recensement des oiseaux de Noël (National Audubon Society, 2015).
- Tableau 2. Résumé des tendances de la population de Bruants à face noire dans les 10 dernières années d’après les données tirées de eBird (2016), rajustées selon la fréquence d’observation, entre le 1er décembre et le 28 février de 2004 à 2014.
Liste des annexes
- Annexe 1. Tableau d’évaluation des menaces pour le Bruant à face noire
Information sur le document
Évaluation et rapport de situation du COSEPAC sur le Bruant à face noire (Zonotrichia querula), 2017

COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les sommaires du statut de l’espèce du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages au Canada que l’on croit en péril. On peut citer le présent document de la façon suivante :
COSEPAC. 2017. Évaluation et Rapport de situation du COSEPAC sur leBruant à face noire (Zonotrichia querula) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xi + 43 p. (Registre public des espèces en péril site Web).
Note de production :
Le COSEPAC remercie Natural Resource Solutions Inc. (Kenneth Burrell) d’avoir rédigé le rapport de situation sur le Bruant à face noire (Zonotrichia querula) au Canada, aux termes d’un marché conclu avec Environnement et Changement climatique Canada. La supervision et la révision du rapport ont été assurées par Marcel Gahbauer, coprésident du Sous-comité de spécialistes des oiseaux du COSEPAC.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site Web du COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Harris's Sparrow Zonotrichia querula in Canada.
Illustration/photo de la couverture :
Bruant à face noire, photo de Ron Ridout (reproduction autorisée).
COSEPAC sommaire de l’évaluation
Sommaire de l’évaluation - avril 2017
- Nom commun
- Bruant à face noire
- Nom scientifique
- Zonotrichia querula
- Statut
- Préoccupante
- Justification de la désignation
- Cet oiseau septentrional qui niche au sol est le seul oiseau chanteur à se reproduire exclusivement au Canada. Les données des Recensements des oiseaux de Noël dans les lieux d’hivernage du Midwest américain montrent un déclin significatif à long terme de 59 % au cours des 35 dernières années, dont 16 % au cours de la dernière décennie. L’espèce pourrait être touchée par les changements climatiques sur les lieux de reproduction, tandis que les menaces qui pèsent sur les lieux d’hivernage sont entre autres la perte d’habitat, l’utilisation de pesticides, la mortalité routière et la prédation par les chats féraux.
- Répartition
- Territoires du Nord-Ouest, Nunavut, Alberta, Saskatchewan, Manitoba, Ontario
- Historique du statut
- Espèce désignée « préoccupante » en avril 2017.
COSEPAC résumé
Bruant à face noire
Zonotrichia querula
Description et importance de l’espèce sauvage
Le Bruant à face noire est un bruant de grande taille doté d’un capuchon et d’une bavette noirs distinctifs. Les individus des deux sexes ont un plumage semblable. Les oiseaux non nicheurs et ceux se trouvant dans leur première année ont un plumage similaire, une bonne partie de la bavette et des motifs faciaux noirs présents chez les individus nicheurs étant absents chez eux. Cet oiseau est le seul passereau qui ne niche qu’au Canada.
Répartition
Le Bruant à face noire, présent seulement en Amérique du Nord, est un oiseau qui migre en zone tempérée sur de longues distances. Il niche le long de la limite forestière dans le nord du Canada (Territoires du Nord-Ouest, Nunavut, Saskatchewan, Manitoba et, irrégulièrement, Ontario). Il hiverne dans le centre du Midwest américain (régulièrement au Nebraska, au Kansas, en Oklahoma et au Texas, et irrégulièrement en Arkansas, au Missouri, en Iowa et au Dakota du Sud). Comme l’accès à son aire de reproduction est limité, on en sait relativement peu sur l’espèce au Canada.
Habitat
Le Bruant à face noire préfère une mosaïque de terrain élevé et de toundra parsemée de lacs. Les territoires de reproduction incluent habituellement des conifères; les densités les plus élevées sont observées dans les peuplements forestiers dominés par l’épinette ou le mélèze, émaillés d’arbustes ayant généralement moins de un mètre de hauteur. En hiver et durant la migration, l’espèce fréquente divers habitats, notamment des fourrés riverains, des prairies, des lisières de forêts, des haies et des fourrés de saules.
Biologie
Le Bruant à face noire est une espèce socialement monogame qui se nourrit de fruits, de graines et d’insectes. Durant la saison de reproduction, il dépend d’abord principalement des fruits, puis il modifie son régime alimentaire pour y inclure davantage d’insectes et de graines au fil de la saison, au fur et à mesure que la couverture de neige disparaît. Les nids sont construits par les femelles, qui couvent aussi les œufs; ils sont placés sur le sol là où la végétation dense peut les dissimuler. Les couvées comptent en moyenne 4,07 œufs, la fourchette étant de 3 à 5. L’incubation dure de 12 à 13,5 jours, et les oisillons quittent le nid après 8,5 à 10 jours. Des études effectuées dans le secteur de la rivière Thelon (Territoires du Nord-Ouest) ont montré un taux d’éclosion de 76 %, un pourcentage d’oisillons ayant quitté le nid de 62,5 % et un taux global de succès de nidification de 47,5 %, la moyenne étant de 2,07 oisillons ayant quitté le nid par couple.
Taille et tendances des populations
La population mondiale, qui se reproduit exclusivement au Canada, est estimée à entre 500 000 et 5 000 000 d’individus, les plus récentes estimations se situant autour de 2 000 000 d’individus.
D’après les données obtenues du Recensement des oiseaux de Noël, le Bruant à face noire a connu un déclin annuel à long terme significatif de 2,58 % entre 1980 et 2014. Cela représente une baisse totale des effectifs de 59 % sur les 35 dernières années. Pour la plus récente période décennale (2004 à 2014), les données du Recensement montrent un déclin de 1,77 % par année, pour une perte cumulative de 16 %.
Menaces et facteurs limitatifs
À la grandeur des lieux d’hivernage du Midwest américain, la conversion de prairies et de terres périphériques à des fins agricoles pourrait être un facteur du déclin du Bruant à face noire. L’épandage de pesticides un peu partout dans l’aire d’hivernage a été lié au déclin de l’espèce; l’incidence relative de ce facteur est inconnue, mais elle est probablement négative et pourrait être grave.
Au sein de l’aire de reproduction, les préoccupations incluent la perte d’habitat liée au déboisement près de la limite septentrionale de l’aire de répartition de l’espèce. Ce déboisement est dû aux feux de forêt, à l’exploitation de mines et de carrières, et aux changements climatiques, qui pourraient réduire l’habitat de reproduction convenable tout en permettant aux ectoparasites et aux mammifères prédateurs, comme le renard roux, d’étendre leur répartition vers le nord.
Des études supplémentaires doivent être effectuées pour évaluer l’espèce tout au long de son cycle de vie annuel et pour étudier l’importance relative des différentes menaces qui la guettent dans ses lieux de reproduction et d’hivernage.
Protection, statuts et classements
Le Bruant à face noire, ses nids et ses œufs sont protégés au Canada par la Loi sur la convention concernant les oiseaux migrateurs. Cette loiinterdit la vente et la possession d’oiseaux migrateurs et de leurs nids, et toute activité qui nuit aux oiseaux migrateurs, à leurs œufs ou à leurs nids, sauf exceptions autorisées par le Règlement sur les oiseaux migrateurs. L’espèce est également protégée aux États-Unis aux termes de la Migratory Birds Treaty Act.
Le Bruant à face noire est classé « non en péril à l’échelle mondiale » par NatureServe. Au Canada, l’espèce est classée « non en péril » à l’échelle nationale, « non en péril » en Alberta et en Saskatchewan, et « vulnérable » au Manitoba et dans les Territoires du Nord-Ouest. Le COSEPAC a évalué l’espèce comme étant préoccupante en avril 2017.
Résumé technique
- Nom scientifique:
- Zonotrichia querula
- Nom français:
- Bruant à face noire
- Nom anglais:
- Harris’s Sparrow
- Répartition au Canada:
- Territoires du Nord-Ouest, Nunavut, Alberta, Saskatchewan, Manitoba, Ontario
Information démographique
| Sujet | Information |
|---|---|
| Durée d’une génération | De deux à trois ans |
| Y a-t-il un déclin continu [observe, inféré ou prévu] du nombre total d’individus matures? | Oui, observé |
| Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations]. | Inconnu |
| Pourcentage [observe, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations]. | Réduction observée de 16 % d’après les données du Recensement des oiseaux de Noël pour la plus récente période de 10 ans. |
| Pourcentage [prévu ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu, mais on s’attend à ce que le déclin à long terme continue à moins que les menaces soient relevées et atténuées. |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu, mais on s’attend à ce que le déclin à long terme continue à moins que les menaces soient relevées et atténuées. |
| Est-ce que les causes du déclin sont a) clairement réversibles et b) comprises et c) ont effectivement cessé? | a. On ne sait pas. b. Non c. Non |
| Y a-t-il des fluctuations extrêmes du nombre d’individus matures? | Non |
Information sur la répartition
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d’occurrence Selon la méthode du plus petit polygone convexe (voir la figure 1). | 1 368 784 km2 (aire de reproduction) 1 377 973 km2 (aire d’hivernage) |
| Indice de zone d’occupation (IZO) [Fournissez toujours une valeur établie à partir d’une grille à carrés de 2 km de côté.] |
Plus de 2 000 km2 |
| La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d’occupation totale se trouvent dans des parcelles d’habitat qui sont a) plus petites que la superficie nécessaire au maintien d’une population viable et b) séparées d’autres parcelles d’habitat par une distance supérieure à la distance de dispersion maximale présumée pour l’espèce? | a) Non b) Non |
| Nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme. (utilisez une fourchette plausible pour refléter l’incertitude, le cas échéant) |
Inconnu, mais plus de 10 |
| Y a-t-il un déclin [observé, inféré ou prévu] de la zone d’occurrence? | On ne sait pas. |
| Y a-t-il un déclin [observé, inféré ou prévu] de l’indice de zone d’occupation? | On ne sait pas. |
| Y a-t-il un déclin [observé, inféré ou prévu] du nombre de sous-populations? | S.O. |
| Y a-t-il un déclin [observé, inféré ou prévu] du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
On ne sait pas. |
| Y a-t-il un déclin [observé, inféré ou prévu] de [la superficie, l’étendue ou la qualité] de l’habitat? | On ne sait pas. |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | S.O. |
| Y a-t-il des fluctuations extrêmes du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d’occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation? | Non |
Nombre d’individus matures (dans chaque sous-population)
| Population | Nombre d’individus matures |
|---|---|
| cellule vide | Selon une estimation grossière, entre 500 000 et 5 000 000 (Blancher et al., 2007); la plus récente estimation est de 2 000 000 (Rosenberg et al., 2016). |
| Total | 2 000 000 |
Analyse quantitative
| Sujet | Information |
|---|---|
| Probabilité de disparition de l’espèce à l’état sauvage | Analyse quantitative non réalisée |
Menaces (réelles ou imminentes pour les populations ou leur habitat, de l’impact le plus élevé à l’impact le plus faible)
| Sujet | Information |
|---|---|
| Un calculateur des menaces a-t-il été rempli pour l’espèce, et dans l’affirmative, par qui? | Oui, le 26 septembre 2016, par Kenneth Burrell, Dave Fraser, Marcel Gahbauer, Suzanne Carrière, Richard Elliot, Pam Sinclair, Myles Lamont, Chris Norment, Rudolf Koes, Jim Rising, Samuel Hache, Amy Ganton, Joanna James. Impact global des menaces évalué à élevé-moyen, sur la base des évaluations suivantes :
La conversion de l’habitat pour l’agriculture, la production d’énergie renouvelable, les incendies et la suppression des incendies de même que les changements climatiques comptent parmi les autres menaces potentielles pour le Bruant à face noire, mais on n’en sait pas suffisamment, pour l’instant, pour évaluer leurs effets. |
Immigration de source externe (immigration de l’extérieur du Canada)
| Sujet | Information |
|---|---|
| Situation des populations de l’extérieur les plus susceptibles de fournir des individus immigrants au Canada | S.O. L’aire de reproduction de l’espèce se trouve exclusivement au Canada. |
| Une immigration a-t-elle été constatée ou est-elle possible? | S.O. |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | S.O. |
| Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants? | S.O. |
| Les conditions se détériorent-elles au Canada Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) ? |
S.O. |
| Les conditions de la population source se détériorent-elles Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) ? |
S.O. |
| La population canadienne est-elle considérée comme un puits Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe) ? |
S.O. |
| La possibilité d’une immigration depuis des populations externes existe-t-elle? | S.O. |
Nature délicate de l’information sur l’espèce
| Sujet | Information |
|---|---|
| L’information concernant l’espèce est-elle de nature délicate? | Non; l’espèce est largement répandue et demeure relativement commune à la grandeur de l’aire de reproduction au Canada. |
Statut historique
| Sujet | Information |
|---|---|
| COSEPAC : | Espèce désignée « préocupante » en avril 2017. |
Statut et justification de la désignation
| Sujet | Information |
|---|---|
| Statut | Préoccupante |
| Code alphanumérique | Sans objet |
| Justification de la désignation | Cet oiseau septentrional qui niche au sol est le seul oiseau chanteur à se reproduire exclusivement au Canada. Les données des Recensements des oiseaux de Noël dans les lieux d’hivernage du Midwest américain montrent un déclin significatif à long terme de 59 % au cours des 35 dernières années, dont 16 % au cours de la dernière décennie. L’espèce pourrait être touchée par les changements climatiques sur les lieux de reproduction, tandis que les menaces qui pèsent sur les lieux d’hivernage sont entre autres la perte d’habitat, l’utilisation de pesticides, la mortalité routière et la prédation par les chats féraux. |
Applicabilité des critères
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d’individus matures) | Sans objet puisque le déclin dans les 10 dernières années est d’environ 16 %. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Sans objet étant donné la vaste répartition de l’espèce. |
| Critère C (nombre d’individus matures peu élevé et en déclin) | Sans objet étant donné la taille de la population, estimée à environ deux millions. |
| Critère D (très petite population totale ou répartition restreinte) | Sans objet étant donné la taille de la population, estimée à environ deux millions. |
| Critère E (analyse quantitative) | Analyse non réalisée. |
Historique du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Mandat du COSEPAC
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Composition du COSEPAC
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Définitions (2016)
- Espèce sauvage
- Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d’animal, de plante ou d’un autre organisme d’origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s’est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
- Disparue (D)
- Espèce sauvage qui n’existe plus.
- Disparue du pays (DP)
- Espèce sauvage qui n’existe plus à l’état sauvage au Canada, mais qui est présente ailleurs.
-
En voie de disparition (VD)
(Remarque : Appelée « espèce disparue du Canada » jusqu’en 2003.) - Espèce sauvage exposée à une disparition de la planète ou à une disparition du pays imminente.
- Menacée (M)
- Espèce sauvage susceptible de devenir en voie de disparition si les facteurs limitants ne sont pas renversés.
-
Préoccupante (P)
(Remarque : Appelée « espèce en danger de disparition » jusqu’en 2000.) - Espèce sauvage qui peut devenir une espèce menacée ou en voie de disparition en raison de l'effet cumulatif de ses caractéristiques biologiques et des menaces reconnues qui pèsent sur elle.
-
Non en péril (NEP)
(Remarque : Appelée « espèce rare » jusqu’en 1990, puis « espèce vulnérable » de 1990 à 1999.) - Espèce sauvage qui a été évaluée et jugée comme ne risquant pas de disparaître étant donné les circonstances actuelles.
-
Données insuffisantes (DI)
(Remarque :Autrefois « aucune catégorie » ou « aucune désignation nécessaire ».) - Une catégorie qui s’applique lorsque l’information disponible est insuffisante (a) pour déterminer l’admissibilité d’une espèce à l’évaluation ou (b) pour permettre une évaluation du risque de disparition de l’espèce.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu’en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d’Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Description et importance de l’espèce sauvage
Nom et classification
Nom scientifique : Zonotrichia querula (Nuttall, 1840)
Nom français : Bruant à face noire
Nom anglais : Harris’s Sparrow
Classe : Oiseaux
Ordre : Passereaux
Famille : Embérizidés
Genre : Zonotrichia
Espèce : Zonotrichia querula
La classification retenue est celle proposée par l’American Ornithologists’ Union (AOU, 1998; Chesser et al., 2015). Aucune variation géographique ni aucune sous-espèce n’est reconnue (AOU, 1998; Norment et al., 2016). Il existe actuellement cinq espèces au sein du genre Zonotrichia (AOU, 1998).
Description morphologique
Le Bruant à face noire est un bruant de grande taille (longueur corporelle : de 17 à 20 cm, masse corporelle : de 30 à 45 g) portant des marques distinctives sur la face et la poitrine. Les individus des deux sexes ont un plumage semblable. Les adultes nicheurs sont dotés d’un capuchon et d’une bavette noirs bien définis qui couvrent le devant de la face et le dessus de la tête, jusqu’à la nuque et au haut de la gorge. La poitrine et les flancs sont striés de noir. Le bec et les pattes sont d’un rose chair. Les caractéristiques principales des ailes sont deux barres alaires blanches; le reste des ailes et le dos sont tachetés de brun et de gris. Les oiseaux non nicheurs et ceux se trouvant dans leur première année ont un plumage semblable, une bonne partie de la bavette et des motifs faciaux noirs présents chez les individus nicheurs étant absents chez eux. La quantité de noir sur la face et la bavette varie, les oiseaux les plus âgés et les mâles en ayant généralement davantage, la face demeurant principalement brune et grise. Les vocalisations de l’espèce ressemblent à celles d’autres espèces du même genre, en particulier à la première partie du chant du Bruant à gorge blanche (Zonotrichia albicollis). Son chant est décrit comme un sifflement plaintif par Semple et Sutton (1932), et comme un « siiiiiii siiii siiii » à une seule tonalité par Sibley (2000). On peut établir l’âge des individus, plus particulièrement des oiseaux se trouvant dans leur première année, en vérifiant la présence éventuelle de rémiges juvéniles (Pyle, 1997; Norment et al., 2016).
Structure spatiale et variabilité de la population
On ne possède aucune preuve de structuration au sein de la population de l’espèce. La population est répartie de façon continue dans toute l’aire de reproduction; aucun obstacle important à la dispersion et aux déplacements n’a été relevé. On remarque une hausse clinale de la taille des couvées avec l’accroissement de la latitude (Norment 1992a, b).
Unités désignables
Le Bruant à face noire ne compte aucune sous-espèce (AOU, 1998; Norment et al., 2016); tous les individus qui se reproduisent au Canada font partie d’une même population, et le présent rapport ne considère donc qu’une seule unité désignable.
Importance de l’espèce
Le Bruant à face noire est le seul passereau qui ne niche qu’au Canada (Norment et al., 2016). Il s’agit d’une des dernières espèces d’oiseaux chanteurs nord-américains dont les nids et les œufs ont été découverts, en 1931 dans son cas, de sorte que les éléments de base de l’écologie de sa reproduction étaient relativement peu connus jusqu’à récemment (Norment et al., 2016).
Aucune connaissance traditionnelle autochtone sur cette espèce n’est disponible actuellement.
Répartition
Aire de répartition mondiale
Le Bruant à face noire, présent seulement en Amérique du Nord, est un oiseau qui migre en zone tempérée sur des distances moyennes. Il niche le long de la limite forestière dans le nord du Canada (Territoires du Nord-Ouest, Nunavut, Saskatchewan, Manitoba et, irrégulièrement, Ontario); il hiverne dans le centre du Midwest américain (Dakota du Sud, Nebraska, Kansas, Oklahoma, Texas, Arkansas, Missouri et Iowa). Son aire d’hivernage principale inclut le centre-nord du Texas, le Kansas, l’Oklahoma et le Nebraska (figure 1). Ses routes migratoires ne sont pas bien connues, mais il suit sans doute une bande étroite de terres entre les aires de nidification et d’hivernage. De plus, l’espèce passe l’hiver, à titre de visiteur occasionnel, dans des emplacements périphériques éparpillés des 48 États contigus des États-Unis ainsi que, dans une moindre mesure, dans le sud du Canada et l’extrême nord du Mexique (National Audubon Society, 2015; eBird, 2016).
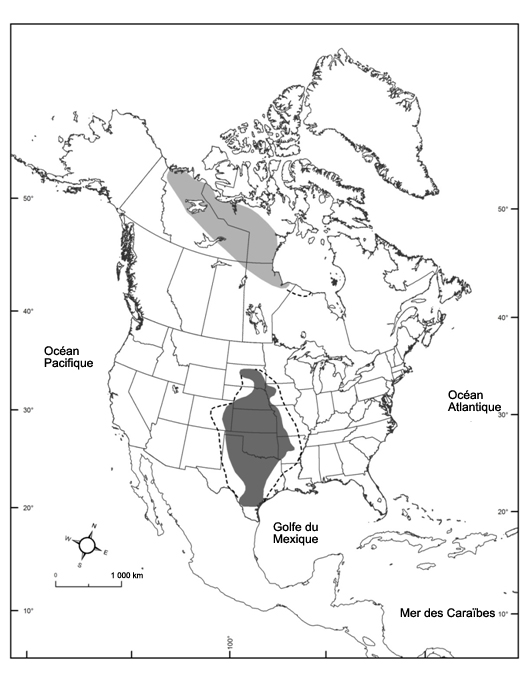
Description longue de la figure 1
Carte montrant l’aire de répartition mondiale du Bruant à face noire en Amérique du Nord. L’espèce se reproduit le long de la limite des arbres dans le nord du Canada (dans les Territoires du Nord Ouest, au Nunavut, en Saskatchewan, au Manitoba et, de façon irrégulière, en Ontario) et passe l’hiver dans le centre de la région du Midwest des États Unis (dans les États du Dakota du Sud, du Nebraska, du Kansas, de l’Oklahoma, du Texas, de l’Arkansas, du Missouri et de l’Iowa).
Aire de répartition canadienne
Le Bruant à face noire niche principalement près de la limite forestière dans certains secteurs des écozones du Bas-Arctique, de la taïga des plaines, de la taïga du Bouclier et des plaines hudsonniennes, à l’ouest de la baie d’Hudson (Government of Canada, 2013).
Au Manitoba, l’espèce niche à l’extrémité septentrionale de la province, où on la voit régulièrement le long du fleuve Nelson et à l’ouest de celui-ci, jusque dans le coin nord-ouest de la province et dans le nord-est de la Saskatchewan (eBird, 2016; Norment et al., 2016; figure 2). Au Nunavut, l’espèce niche dans les régions du Kivalliq et du Kitikmeot. Au sein de la région du Kivalliq, on la trouve principalement dans les deux tiers méridionaux, jusqu’à Rankin Inlet au nord; après cela, l’aire de reproduction se continue vers le nord-ouest depuis la côte jusqu’à un secteur situé à l’est d’Umingmaktok, dans la région du Kitikmeok (eBird, 2016; Norment et al., 2016). Dans le Kitikmeot, le Bruant à face noire vit le long de la côte de l’océan Arctique, jusqu’à Kugluktuk et Clifton Point à l’ouest (eBird, 2016; Norment et al., 2016). Dans les Territoires du Nord-Ouest, l’espèce niche jusqu’au delta du fleuve Mackenzie à l’ouest, et même peut-être jusque dans l’extrême nord-est du Yukon (Eckert, comm. pers., 2015). La limite sud-ouest de l’aire de reproduction part du nord-ouest des Territoires du Nord-Ouest, passe au sud du Grand lac de l’Ours et se rend jusque dans le nord-est de la Saskatchewan (eBird, 2016; Norment et al., 2016).
D’après les données tirées de l’Atlas des oiseaux nicheurs de l’Ontario (1981-1985 et 2001-2005; Cadman et al.,1987, 2007) et de James et al. (1976), l’oiseau niche irrégulièrement dans des endroits épars près de la côte de la baie d’Hudson (figure 3).
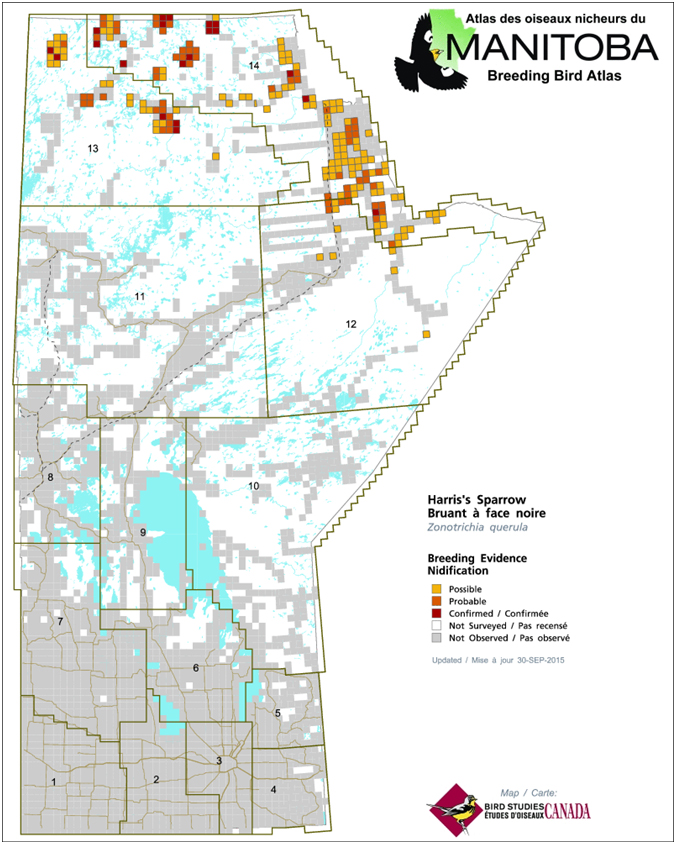
Description longue de la figure 2
Carte montrant l’aire de reproduction (à l’aide de carrés de grille) du Bruant à face noire au Manitoba, selon les données de l’Atlas des oiseaux nicheurs de 2010 à 2014. L’espèce est observée le long du fleuve Nelson et à l’ouest de celui ci, jusqu’au coin nord ouest de la province.
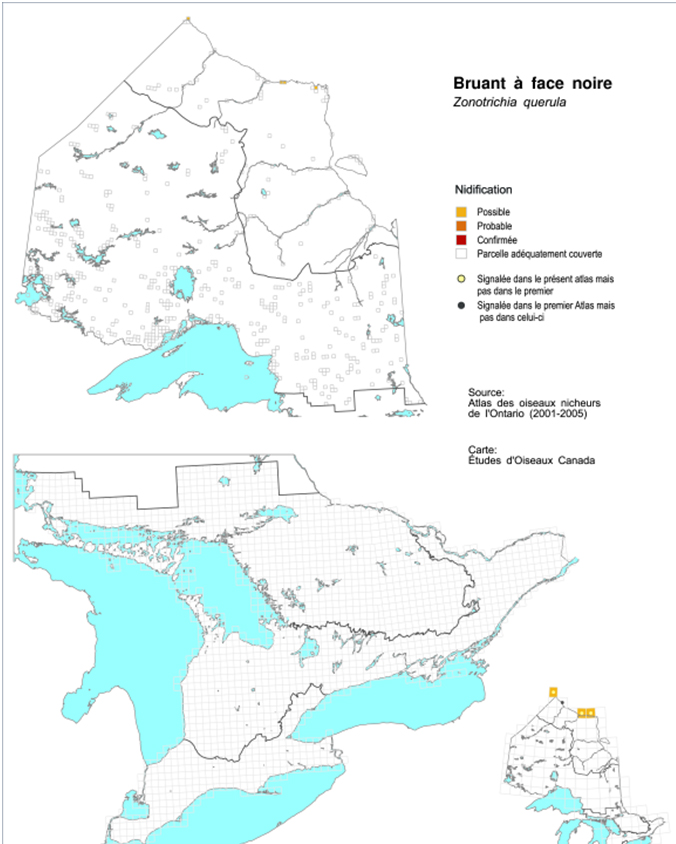
Description longue de la figure 3
Carte indiquant l’aire de reproduction (à l’aide de carrés de grille) du Bruant à face noire en Ontario, selon les données de l’Atlas des oiseaux nicheurs. La carte montre une reproduction irrégulière dans des localités dispersées près de la côte de la baie d’Hudson.
Zone d’occurrence et zone d’occupation
On estime la superficie de la zone d’occurrence à 1 368 784 km2 pour l’aire de reproduction et à 1 377 973 km2 pour l’aire d’hivernage (figure 1). La répartition au sein de l’aire de reproduction n’est pas suffisamment documentée pour permettre le calcul de la zone d’occupation, mais celle-ci comporte certainement plus de 2 000 km2.
Activités de recherche
Les données de répartition du Bruant à face noire au Canada sont principalement tirées de l’Atlas des oiseaux nicheurs du Manitoba (2015), élaboré en 2010-2014, du Conservation Data Centre de la Saskatchewan et d’observations de particuliers avec lesquels on a communiqué lors de la préparation du présent rapport de situation (voir la section « Remerciements »).
Les cartes de répartition tirées de NatureServe Explorer (2015) et de The Birds of North America (Norment et al., 2016) sont considérées comme les cartes les plus fiables pour cette espèce; elles ont été corroborées par des données provenant du Relevé des oiseaux des Territoires du Nord-Ouest et du Nunavut (ECCC, 2016) et de eBird (2016) pour juin et juillet dans les dix dernières années.
Habitat
Besoins en matière d’habitat
Habitat de reproduction
Dans toute son aire de reproduction, le Bruant à face noire préfère la zone de transition entre la forêt et la toundra de terrain élevé. Les territoires de reproduction incluent habituellement des conifères; les densités les plus élevées sont observées dans les peuplements forestiers dominés par l’épinette (Picea sp.) et, dans une moindre mesure, par le mélèze (Larix laricina), où le sous-étage comporte de la végétation arbustive (Harper, 1953; Gillespie et Kendeigh, 1982; Norment, 1992a; Norment et al., 2016). On a observé une corrélation positive entre la densité de Bruants à face noire et l’abondance d’arbres et de végétation de sous-étage (Norment et al., 2016). L’espèce niche à l’occasion dans des secteurs exempts d’arbres; dans ces cas, le couvert arbustif est particulièrement important pour l’établissement et la dissimulation des nids (Clarke, 1944; Sealy, 1967; Norment et al., 2016). L’habitat de reproduction favorisé consiste généralement en de petits peuplements forestiers isolés (allant d’un petit bouquet d’arbres à une superficie de 12 ha) disséminés un peu partout sur le territoire de reproduction (Norment, 1992a; Norment et al., 2016).
Les densités les plus élevées sont observées là où la composition de la végétation est diversifiée et où les arbustes sont généralement grands (plus de un mètre de hauteur) (Obst, comm. pers., 2015). Les abondances les plus élevées sont signalées là où le territoire comporte 10 % de forêt d’épinettes noires (P. mariana) et d’épinettes blanches (P. glauca), 27 % d’arbustaie de bouleaux glanduleux (Betula glandulosa) et de saules (Salix sp.), et 63 % de toundra (Norment,1992b; Norment et al., 2016). D’après les estimations, les populations d’oiseaux nicheurs varient d’aussi peu que 0,025 mâle territorial par hectare dans les habitats de faible densité (Harris et al., 1974) à 0,125–0,82 couple nicheur par hectare dans les habitats de haute densité (Gillespie et Kendeigh, 1982; Norment, 1992b).
Des variations clinales subtiles sont observées à la grandeur de l’aire de reproduction de l’espèce en ce qui a trait à la végétation présente sur le territoire de reproduction (Norment et al., 2016). Dans les Territoires du Nord-Ouest, l’espèce niche généralement dans de la végétation arbustive dominée par le bouleau glanduleux, l’aulne (Alnus spp.) et le saule (Semple et Sutton, 1932; Harper, 1953; Norment, 1992b). Par contre, à Churchill, au Manitoba, on observe plus souvent l’espèce à la lisière de forêts d’épinettes et de mélèzes (Gillespie et Kendeigh, 1982), dans des clairières et en bordure de secteurs incendiés (Semple et Sutton, 1932; Norment et al., 2016).
Habitat d’hivernage et de migration
L’espèce fréquente divers habitats tout au long de l’hiver et des périodes de migration, et est fréquemment observée dans des fourrés riverains, des lisières de forêts, des haies et des fourrés de saules (NatureServe, 2015). Des études menées au Kansas (Graul, 1967) et en Oklahoma (Nice, 1929; Bridgwater, 1966) montrent que l’espèce préfère les secteurs perturbés, par exemple les haies, les tas de broussailles et les terres agricoles, de même que l’habitat naturel comme la succession secondaire et les corridors riverains (Norment et al., 2016).
Tendances en matière d’habitat
Habitat de reproduction
Des activités humaines, par exemple l’exploitation minière et l’aménagement de routes, ont lieu dans l’aire de reproduction; la portée de leur incidence sur l’habitat de reproduction demeure incertaine, mais le taux global de changement de l’habitat de reproduction est probablement faible.
Les effets quantitatifs des changements climatiques ne sont pas connus, mais des témoignages anecdotiques laissent supposer que le déplacement vers le nord de la limite forestière pourrait avoir sur l’espèce une incidence (Obst, comm. pers., 2015) à la fois positive (p. ex. hausse de l’abondance d’espèces d’arbustes utiles au Bruant à face noire pour la recherche de nourriture et la nidification) et négative (p. ex. diminution de l’habitat convenable à la limite sud de l’aire de répartition, c.-à-d. dans le nord du Manitoba). La quantité et la qualité de l’habitat de reproduction de l’espèce ne font pas l’objet d’une surveillance (aucune tendance n’est connue).
Habitat d’hivernage
L’aire d’hivernage du Bruant à face noire inclut le centre et le sud de la région des Grandes Plaines des États-Unis, du Dakota du Sud au Texas (Norment et al., 2016), où l’espèce utilise des habitats ouverts, y compris les fourrés riverains, les lisières de forêts, les haies et les fourrés de saules. La conversion de ce type d’habitat en terres agricoles et, dans une moindre mesure, l’étalement urbain constituent des menaces pour l’espèce, mais on ne connaît pas la portée de ces menaces (NatureServe, 2015).
Biologie
Peu d’études récentes ont été faites sur le Bruant à face noire au Canada. Une bonne partie des connaissances que nous possédons sur la biologie de reproduction de l’espèce sont tirées d’études antérieures menées par C. Norment dans le secteur de la rivière Thelon, dans les Territoires du Nord-Ouest (p. ex. Norment, 1992a, b, 1993, 1994, 1995a, b). Ainsi, une bonne partie de l’information donnée ci-dessous est tirée de renseignements résumés dans la portion de l’ouvrage révisé The Birds of North America consacrée à l’espèce (Norment et al., 2016).
Cycle vital et reproduction
Le Bruant à face noire est une espèce socialement monogame (Norment et al., 2016). Les oiseaux en couple cherchent leur nourriture ensemble, le mâle suivant la femelle, de près juste avant l’incubation (Norment et al., 2016). Les individus forment des groupes lâches à la fin de l’été, juste avant la migration automnale (Norment et al., 2016).
Le Bruant à face noire arrive généralement dans son territoire de reproduction entre la fin de mai et le début de juin (Norment et al., 2016). Il commence habituellement à construire son nid entre la deuxième et la troisième semaine de juin (c.-à-d. environ 14 jours après la date moyenne d’arrivée des oiseaux) ou lorsque la couverture de neige ne dépasse pas 60 % (NatureServe Explorer, 2015; Norment et al., 2016). Les nids sont composés de deux couches : la couche intérieure est principalement faite de mousses, de cypéracées et de graminées sèches, et la couche extérieure est faite de lichens, de mousses et de brindilles (Rees, 1973). Les femelles construisent les nids, couvent les œufs et élèvent les petits (Norment et al., 2016). Les deux parents nourrissent les oisillons, mais les mâles ont tendance à le faire moins fréquemment, surtout au début du stade de l’éclosion (Norment et al., 2016). Les couples n’ont une deuxième couvée que si la première échoue avant l’éclosion (Norment, 1992b; Norment et al., 2016).
Norment (1992b) a établi que la période d’incubation dure de 12 à 13,5 jours, et que les oisillons quittent le nid après 8,5 à 10 jours. Les couvées comportent en moyenne (n = 155) 4,07 œufs, la fourchette étant de 3 à 5; le moment de l’éclosion est synchronisé entre les nids (Norment, 1992b). Le taux d’éclosion, dans les Territoires du Nord-Ouest, était de 76 %, le pourcentage d’oisillons ayant quitté le nid, de 62,5 %, et le taux global de succès de nidification (au moins un petit ayant quitté le nid), de 47,5 %, 2,07 oisillons ayant quitté le nid par couple en moyenne (Norment, 1992b; NatureServe Explorer, 2015; Norment et al., 2016). La mortalité observée des jeunes à l’envol était surtout due à la prédation, les principaux prédateurs étant le spermophile arctique (Spermophilus parryii) et, dans une moindre mesure, l’hermine (Mustela erminea) (NatureServe Explorer, 2015).
Les nids sont toujours placés au sol, habituellement au sein d’une couverture de végétation dense (Norment, 1993). La probabilité de succès du nid augmente en fonction de la dissimulation de celui-ci (Norment, 1993). Les nids se trouvent souvent sous des arbustes de moins de un mètre de hauteur (NatureServe Explorer, 2015; Norment et al., 2016). Les espèces de végétaux servant de couverture dépendent selon la latitude, mais la plupart sont ligneuses (Norment, 1993). Au Nunavut, le Bruant à face noire choisit le plus souvent le bouleau nain (Betula pumila) (68 %; n = 65), tandis qu’au Manitoba, il choisit plutôt le petit thé du Labrador (Rhododendron tomentosum) (32 %; n = 26) (Norment 1993). Dans les Territoires du Nord-Ouest, l’orientation moyenne de l’entrée du nid est de 140,5°, soit éloignée à 170° de l’orientation des vents de tempête dominants (Norment, 1993).
L’âge à la première reproduction n’est pas connu, mais on croit qu’il s’agirait d’un an, comme pour d’autres bruants du genre Zonotrichia (Norment, 1992a; Chilton et al., 1995). Le rapport mâle-femelle est de 1:1 dans tous les territoires de reproduction établis (Norment et al., 2016).
Le record de longévité pour cette espèce est de 11 ans et 8 mois (Norment et al., 2016). Le taux moyen de survie des œufs jusqu’à l’éclosion a varié entre 63,8 % et 90,5 % (Norment, 1992b). D’après les données sur le taux de survie annuel, celui-ci, pour les adultes, a été de 38 % en moyenne sur 2 années d’étude au Nunavut (Norment, 1992b); ce faible taux estimé reflète probablement le fait que les individus changent de site de nidification. Si la durée d’une génération n’a pas été calculée pour l’espèce, on suppose qu’elle est de 2 ou 3 ans comme pour la plupart des autres petits passereaux. Un individu hybride, issu d’un croisement avec le Bruant à couronne blanche (Zonotrichia leucophrys), a été pris à la pointe Long, en Ontario (Payne, 1979).
Alimentation
Le Bruant à face noire se nourrit de fruits, de graines et d’insectes; au sol, il obtient ces aliments d’une manière semblable à celle employée par d’autres bruants du genre Zonotrichia, en grattant le sol de ses pattes pour les déloger (Norment,1992a). Il se nourrit moins souvent dans les arbustes et les arbres qu’au sol (Nice, 1929; Semple et Sutton, 1932; Norment et al., 2016).
Le Bruant à face noire se nourrit d’une grande diversité d’espèces végétales : cypéracées (Carex spp. et Cyperus spp.),scirpe (Scirpus spp.), bouleau (Betula spp.), amarante (Amaranthus spp.), chénopode (Chenopodium spp.), airelle (Vaccinium spp.), camarine noire (Empetrum nigrum) et raisin d’ours (Arctostaphylos uva-ursi) (Semple et Sutton, 1932). Les baies sont particulièrement importantes pour cet oiseau lorsqu’il arrive sur son territoire de reproduction avant l’émergence des insectes (Norment et Fuller, 1997). Norment et Fuller (1997) ont constaté que le contenu stomacal des oiseaux avant la nidification était composé à 82 % de fruits comme l’airelle (Vaccinium spp.), la camarine noire (Empetrum nigrum) et le raisin d’ours (Arctostaphylos spp.) La même étude a permis à Norment et Fuller (1997) de conclure que les arthropodes représentaient 72,5 % du régime alimentaire des Bruants à face noire adultes durant la période de nidification et 81,5 % de celui des oisillons au nid.
Physiologie et adaptabilité
On en sait relativement peu sur la physiologie de l’espèce; Norment (1995a) a établi que la température optimale pour l’incubation des œufs du Bruant à face noire se situe entre +10 °C et +20 °C, ce qui permet aux femelles de quitter le nid pour une plus longue période qu’autrement et d’y rentrer moins souvent pour couver. La même étude (Norment, 1995a) a conclu qu’à des températures supérieures à +20 °C, les adultes passaient plus de temps au nid pour faire de l’ombre aux œufs et tenter d’aider les jeunes à l’envol à réguler leur température corporelle (Norment et al., 2016). Il a également été établi que le Bruant à face noire est mal adapté aux températures extrêmes durant la nidification, surtout lorsque le mercure monte au-dessus de +30 °C ou descend sous -10 °C (Norment et al., 2016).
En périodes de mauvais temps, les oiseaux abandonnent des territoires de reproduction nouvellement acquis et y reviennent pour reprendre leur nidification une fois les intempéries terminées (Norment et al., 2016). Cette stratégie profite à l’espèce lorsque les oiseaux n’ont pas commencé la ponte, mais elle est nuisible aux oisillons et aux œufs fraîchement pondus, et peut même parfois leur être mortelle.
Dispersion et migration
Après l’envol, les juvéniles se dispersent à un maximum de 500 m de leur lieu de naissance; les jeunes et certains adultes ont peut-être une distance de dispersion plus élevée au sein de l’aire de reproduction, mais cela n’a pas été documenté (c.-à-d. qu’aucune des observations d’oiseaux bagués n’a confirmé le déplacement sur une distance plus grande que celle-ci d’oiseaux capturés ou bagués au Canada; Brewer et al., 2006; Norment et al., 2016).
Norment (1994) a documenté un taux de retour de 38 % des oiseaux bagués dans les Territoires du Nord-Ouest, sans différence notable selon le sexe. Les adultes n’ayant pas réussi leur nidification sont beaucoup moins nombreux à retourner au même site les années suivantes que ceux l’ayant réussi (Norment, 1994).
D’après les données de baguage et d’observation (Brewer et al., 2006; eBird, 2016), la plupart des oiseaux migrent vers le nord et le sud en empruntant un couloir étroit reliant l’aire de reproduction à l’aire d’hivernage. Des individus seuls sont fréquemment observés hors de l’aire d’hivernage et du couloir de migration habituels.
Relations interspécifiques
À la grandeur de l’aire de reproduction, l’espèce est socialement monogame et forme des groupes lâches avant le début de la migration automnale (Norment et al., 2016). Lors des migrations printanière et automnale de même que durant l’hiver, le Bruant à face noire se joint à des groupes comportant diverses espèces, par exemple le Bruant à couronne blanche, le Bruant hudsonien (Spizelloides arborea)et le Cardinal rouge (Cardinalis cardinalis) (Shuman et al., 1992; Thompson, 1994; Robel et al., 1997).
Taille et tendances des populations
Activités et méthodes d’échantillonnage
Le Bruant à face noire est un des passereaux nord-américains les moins étudiés; seuls quelques programmes ont été conçus correctement afin d’estimer la taille et les tendances de la population. Étant donné que l’oiseau se reproduit dans des secteurs nordiques isolés, il n’est pas vraiment visé par le Relevé des oiseaux nicheurs d’Amérique du Nord, sur lequel on se fie habituellement pour connaître les données sur les tendances des populations d’oiseaux chanteurs. Les relevés fournissant des données pertinentes sont expliqués ci-dessous.
Recensement des oiseaux de Noël
Le Recensement des oiseaux de Noël (RON) remonte à 1900; il permet de relever les populations hivernales d’oiseaux grâce à des recensements annuels à l’intérieur de cercles de dénombrement de 24 km de diamètre (National Audubon Society, 2015). Ce programme fournit des estimations des populations et de l’abondance pour la plupart des oiseaux terrestres qui hivernent au Canada ou aux États-Unis, y compris le Bruant à face noire. Au sein de chaque cercle de dénombrement, les bénévoles participant au RON consignent toutes les espèces d’oiseaux et tous les individus observés en une seule journée, entre le 14 décembre et le 5 janvier d’une année donnée. L’aire de reproduction du Bruant à face noire étant presque entièrement non couverte, le RON constitue la source de données la plus fiable pour l’analyse des tendances de la population.
La couverture du RON dans l’aire de répartition principale du Bruant à face noire en dehors de la période de reproduction (Texas, Oklahoma, Kansas et Nebraska) a été relativement limitée jusqu’en 1980. Les analyses des tendances ont été faites à partir de données recueillies de 1980 à 2014 qui ont été converties en fonction de l’effort d’observation (oiseaux par heure-équipe). L’analyse a également porté sur l’ensemble de l’aire d’hivernage durant la même période pour relever la tendance globale et tenir compte du nombre, peut-être important, d’oiseaux observés à l’extérieur de l’aire de répartition principale en dehors de la période de reproduction. Des analyses par État ont également été menées pour cinq États situés en bordure de l’aire de répartition principale en dehors de la période de reproduction, mais ce n’est qu’au Missouri et en Iowa que la taille des échantillons était suffisante pour que l’on puisse dégager des tendances. Dans les autres États étudiés (Colorado, Dakota du Sud et Arkansas) où l’on observe des Bruants à face noire assez régulièrement en hiver, la taille des échantillons était trop petite pour que les résultats soient significatifs ou pour que l’on dégage des tendances. Ces États ne seront donc pas inclus dans les discussions qui suivent. Aucune analyse n’a été effectuée pour le Canada ou le Mexique étant donné la petite taille des échantillons dans ces régions.
L’analyse des tendances pour le RON a consisté à ajuster, par régression linéaire simple, les courbes de tendance aux indices annuels corrigés pour l’effort (oiseaux par heure-équipe), les tendances étant exprimées sous la forme du taux de changement moyen géométrique entre la valeur de la première année et la valeur de la dernière année (Smith et al., 2014). Les indices annuels étaient des points de donnée uniques pour chaque région et année, calculés en tant que moyennes de tous les cercles de dénombrement du RON par année, données pouvant être téléchargées auprès de la National Audubon Society (2015) en se servant des filtres par espèce pour la consultation des résultats historiques. La signification statistique (p < 0,05) a été établie d’après la valeur de p associée à la pente des droites de régression. Les données ont été analysées au moyen de R v.3.2.3 (R Core Team 2015).
Atlas des oiseaux nicheurs du Manitoba
On a effectué pour l’Atlas des oiseaux nicheurs du Manitoba la première cartographie systématique des oiseaux nicheurs de la province. Des données ont été recueillies sur le terrain de 2010 à 2014 (MBBA, 2015). La couverture n’était pas uniforme, en particulier à l’extrémité nord de la province, où se trouve l’aire de reproduction du Bruant à face noire; dans l’ensemble, 30 % des parcelles de l’Atlas (10 km x 10 km) ont été couvertes au moins en partie, et un effort d’au moins 10 heures a été considéré comme suffisant pour relever la majorité des espèces présentes dans une parcelle (Artuso, comm. pers., 2016).
Comme il s’agit du premier atlas des oiseaux nicheurs de la province, les données recueillies ont uniquement été utilisées pour établir la présence et l’abondance de l’espèce, mais pas les tendances.
eBird
Le programme interactif eBird permet aux utilisateurs de soumettre leurs observations en temps réel sous forme de listes (eBird, 2016). Les données peuvent être compilées aux fins d’établissement de tendances en matière de répartition et d’abondance, puis elles peuvent être utilisées à des fins de conservation et d’éducation (eBird, 2016). Ce programme est relativement nouveau (créé en 2002), et le nombre d’utilisateurs augmente régulièrement depuis sa création (eBird, 2016). Bien que les données contenues dans eBird comportent un biais important découlant de faux positifs, il a été décidé qu’il valait la peine de les utiliser étant donné le manque de données autres sur le Bruant à face noire et dans le but de pouvoir les comparer avec les tendances tirées du Recensement des oiseaux de Noël.
Aux fins du présent rapport de situation, les données ont été analysées pour l’aire de répartition principale de l’espèce en dehors de la période de reproduction aux États-Unis (Texas, Oklahoma, Kansas et Nebraska) ainsi que pour chacun de ces États individuellement et pour deux États périphériques (Iowa et Missouri), où l’on disposait de données suffisantes. Celles-ci ont été compilées pour les mois de décembre à février, ce qui a permis d’obtenir la fréquence moyenne d’observation de l’espèce dans les listes d’observation afin de procéder à l’analyse des tendances entre 2004 et 2014.
Une modélisation statistique a été effectuée de la même manière que pour l’analyse des données du Recensement des oiseaux de Noël, décrite ci-dessus, en utilisant la fréquence d’observation de l’espèce dans les listes pour calculer les tendances rajustées à l’échelle des années.
Réseau canadien de surveillance des migrations
Le Réseau canadien de surveillance des migrations (RCSM) est un programme conjoint, mené par différents observatoires en collaboration avec Études d’Oiseaux Canada et le Service canadien de la faune afin d’effectuer le suivi des populations d’oiseaux terrestres dans tout le pays (Bird Studies Canada, 2016). Le Réseau comporte plus de 25 stations de surveillance un peu partout au pays, où l’on suit les populations d’oiseaux terrestres grâce à des méthodes de suivi normalisées, comme la capture au filet japonais et des relevés quotidiens (Bird Studies Canada, 2016).
Les données ont été analysées aux seuls sites du Réseau se trouvant dans l’étroit couloir de migration du Bruant à face noire : l’observatoire d’oiseaux du marais Delta, au Manitoba, et l’observatoire d’oiseaux du lac de la Dernière-Montagne, en Saskatchewan. On a analysé les données recueillies aux printemps 2006 à 2010 et aux automnes 1993 à 2010 au marais Delta, et les données des printemps 1994 à 2014 et des automnes 1993 à 2014 au lac de la Dernière-Montagne.
Les analyses des tendances ont été produites par NatureCounts (2016) au moyen de l’outil « Population Trends ». La modélisation statistique a été faite au moyen des totaux des recensements et des données de baguage pour calculer les tendances rajustées à l’échelle des années.
Abondance
En se basant principalement sur les données extrapolées à partir du Recensement des oiseaux de Noël, Blancher et al. (2007) avaient estimé que la population de Bruants à face noire comptait entre 500 000 et 5 000 000 d’individus, mais il est à noter que les données sur l’espèce étaient alors limitées. En se fondant sur des données plus récentes, Rosenberg et al. (2016) ont revu l’estimation à environ 2 000 000 d’individus. Puisque l’ensemble de la population niche au Canada, cette estimation se trouve à être aussi celle de la population canadienne.
D’après les résultats préliminaires tirés de l’Atlas des oiseaux nicheurs du Manitoba, des indices de nidification du Bruant à face noire ont été enregistrés dans 200 parcelles d’atlas (10 km x 10 km). La plupart des mentions pour l’espèce avaient trait à une nidification possible (118 carrés), les indices de nidification probable (62) ou confirmée (20) étant moins communs (MMBA, 2015). Dans les parcelles d’atlas où on a relevé la présence du Bruant à face noire, l’espèce a été observée à 44,4 % de tous les points d’écoute (n = 1 096), l’abondance moyenne étant de 0,71 oiseau par dénombrement (n = 2 469). Des données supplémentaires publiées dans l’Atlas des oiseaux nicheurs du Manitoba montrent que la probabilité de détection est élevée (plus de 70 %) à l’extrémité nord-ouest du Manitoba, même le long du littoral de la baie d’Hudson à l’est du fleuve Nelson jusqu’à la limite de l’Ontario, et ce malgré la rareté des observations précédentes à cet endroit (Artuso, comm. pers., 2016, inédit).
Des études effectuées par Gillespie et Kendleigh (1982) et par Norment (1992b) au Manitoba et dans les Territoires du Nord-Ouest laissent supposer que les densités varient entre 0,125 et 0,82 couple par hectare dans l’habitat convenable. Toutefois, la densité moyenne à la grandeur de l’aire de répartition de l’espèce demeure inconnue.
Fluctuations et tendances
Recensement des oiseaux de Noël
D’après les données du Recensement des oiseaux de Noël (tableau 1; figure 4), sur une période de 35 ans (de 1980 à 2014), le Bruant à face noire a connu un déclin à long terme significatif sur le plan statistique, soit de 2,58 % par année (p < 0,001). Cela représente une baisse totale de l’effectif de 59 % sur cette période. Le taux de déclin semble avoir diminué dans les 10 dernières années (-1,77 % par année, p = 0,211), la baisse estimée totale de l’effectif, statistiquement non significative, étant de 15 % pour cette période (2004-2014).
| Région | Période | Taux annuel de changement (%/année) | Valeur de p |
|---|---|---|---|
| États-Unis | 1980 – 2014 | -2,58 | ** |
| États-Unis | 2004 – 2014 | -1,77 | ns |
| Texas | 1980 – 2014 | -3,18 | ** |
| Texas | 2004 – 2014 | +14,17 | ns |
| Oklahoma | 1980 – 2014 | -0,94 | * |
| Oklahoma | 2004 – 2014 | -1,55 | ns |
| Nebraska | 1980 – 2014 | -7,03 | ** |
| Nebraska | 2004 – 2014 | +2,96 | ns |
| Kansas | 1980 – 2014 | -0,78 | ns |
| Kansas | 2004 – 2014 | -9,48 | * |
| Missouri | 1980 – 2014 | +0,33 | ns |
| Missouri | 2004 – 2014 | +11,34 | ns |
| Iowa | 1980 – 2014 | -1,01 | ns |
| Iowa | 2004 – 2014 | +10,66 | ns |
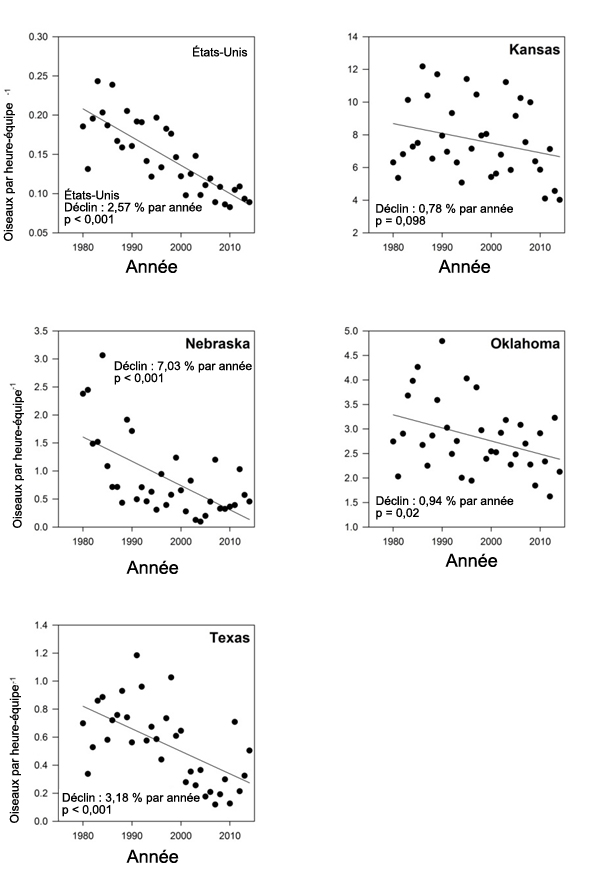
Description longue de la figure 4
Graphiques montrant les tendances annuelles en ce qui concerne les observations de Bruants à face noire (oiseaux par heure équipe) de 1980 à 2014 aux États Unis (dans les États du Kansas, du Nebraska, de l’Oklahoma et du Texas), selon le Recensement des oiseaux de Noël.
Les données du Recensement des oiseaux de Noël pour trois des quatre États de l’aire de répartition principale en dehors de la période de reproduction (Texas, Oklahoma et Nebraska) montrent un taux annuel de déclin à long terme significatif (-3,18 %, -0,94 % et -7,03 %, respectivement), tandis que dans l’État restant (Kansas), un déclin significatif sur le plan statistique (-9,48 % par année) a été enregistré à court terme (10 ans).
Les tendances pour les États périphériques étudiés (Missouri et Iowa) étaient principalement positives, mais non significatives. Ces États abritent une faible proportion des Bruants à face noire en hivernage, et les données les concernant ne reflètent pas la tendance globale pour l’espèce.
La seule tendance significative à court terme a été enregistrée au Kansas (-9,48 %; p < 0,05). Au Texas, une hausse importante de 14,2 % par année a été observée, mais elle n’est pas statistiquement significative au seuil de signification de 0,05 (p = 0,099). Cette situation pourrait s’expliquer par la concentration de l’espèce dans l’habitat convenable à la suite de la perte d’habitat (due à la conversion en terres agricoles) dans la partie nord de l’aire d’hivernage, ou encore par des hivers particulièrement rudes ayant force l’espèce à se rendre plus au sud récemment. S’il semble y avoir une certaine plasticité dans la répartition hivernale d’une année à l’autre, on ne constate aucun changement précis de la répartition de l’espèce dans son aire d’hivernage.
eBird
Aux fins du présent rapport de situation, les données tirées de eBird ont été comparées aux données du Recensement des oiseaux de Noël. D’après les analyses des tendances faites pour la période allant de 2004 à 2014, des tendances significatives ont été relevées pour l’ensemble des États faisant partie de l’aire de répartition principale en dehors de la période de reproduction (-13,47 %, p = 0,031; figure 5) de même que pour le Missouri (+11,09 %, p = 0,035) (tableau 2). Bien que la tendance à la baisse pour l’ensemble des États faisant partie de l’aire de répartition en dehors de la période de reproduction soit plus importante que celle relevée grâce au Recensement des oiseaux de Noël, les deux programmes permettent de constater des baisses globales des effectifs à court ou à long terme. Des tendances à la baisse non significatives ont été relevées au Texas, en Oklahoma, au Kansas et en Iowa, tandis que les observations au Nebraska ont augmenté.

Description longue de la figure 5
Graphique montrant les registres d’eBird (eBird, 2016) relatifs au Bruant à face noire de 2004 à 2014 (période de décembre à février) dans les principaux États où l’espèce est présente en dehors de sa période de reproduction.
| Région | Période | Taux annuel de changement (%/année) | Valeur de p |
|---|---|---|---|
| États de l’aire de répartition principale en dehors de la période de reproduction | 2004 – 2014 | -13,47 | * |
| Texas | 2004 – 2014 | -1,91 | ns |
| Oklahoma | 2004 – 2014 | -2,39 | ns |
| Nebraska | 2004 – 2014 | +8,16 | ns |
| Kansas | 2004 – 2014 | -1,54 | ns |
| Missouri | 2004 – 2014 | +11,09 | * |
| Iowa | 2004 – 2014 | -4,38 | ns |
Réseau canadien de surveillance des migrations
Les données du Réseau canadien de surveillance des migrations montrent que des tendances significatives ont été relevées à l’observatoire d’oiseaux du lac de la Dernière-Montagne (Saskatchewan), mais pas à l’observatoire d’oiseaux du marais Delta (Manitoba). Au lac de la Dernière-Montagne, le Bruant à face noire a subi un déclin significatif d’après le jeu de données à long terme pour l’automne (de 1993 à 2014); toutefois, les jeux de données pour le printemps (figure 6) et l’automne (figure 7) témoignent tous deux d’une hausse (24,23 % et 1,41 %, respectivement) à court terme (de 2004 à 2014), la tendance étant significative pour les données du printemps.
Les données enregistrées à l’observatoire d’oiseaux du marais Delta indiquent une tendance non significative dans les jeux de données du printemps (court terme; figure 8) et de l’automne (long terme; figure 9). Les données propres au printemps (de 2004 à 2010) montrent une hausse de 12,56 % par année, tandis que celles de l’automne (1993 à 2010) montrent une baisse de 0,02 % par année.

Description longue de la figure 6
Graphique illustrant la tendance de la population de Bruants à face noire observée à l’Observatoire d’oiseaux du lac Last Mountain au printemps, de 1994 à 2014.
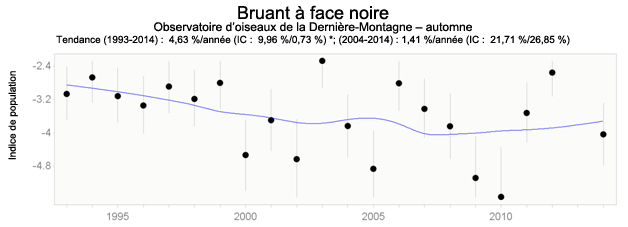
Description longue de la figure 7
Graphique illustrant la tendance de la population de Bruants à face noire observée à l’Observatoire d’oiseaux du lac Last Mountain à l’automne, de 1993 à 2014.

Description longue de la figure 8
Graphique illustrant la tendance de la population de Bruants à face noire observée à l’Observatoire d’oiseaux du marais Delta au printemps, de 2006 à 2010.

Description longue de la figure 9
Graphique illustrant la tendance de la population de Bruants à face noire observée à l’Observatoire d’oiseaux du marais Delta à l’automne, de 1993 à 2010.
Immigration de source externe
Sans objet. Comme le Bruant à face noire ne se reproduit qu’au Canada, la possibilité d’un sauvetage par immigration d’individus de populations de l’extérieur du Canada est exclue.
Menaces et facteurs limitatifs
Les menaces sont décrites ci-dessous et résumés à l’annexe 1 d’après une version modifiée du système unifié de classification des menaces proposé par l’Union international pour la conservation de la nature et le Partenariat pour les mesures de conservation (IUCN-CMP) (COSEWIC, 2014), qui donne pour les menaces pesant sur le Bruant à face noire un impact global élevé à moyen.
Catégorie 9.3 : Effluents agricoles et sylvicoles (impact faible à élevé)
L’épandage de pesticides un peu partout dans les lieux d’hivernage a été lié au déclin d’oiseaux de prairie (Norment, comm. pers., 2016). Les néonicotinoïdes, en particulier, ont été mis en cause dans le vaste déclin de divers taxons dans les dix dernières années (Hallmann et al., 2014; Gibbons et al., 2015). Le degré d’incidence passée ou actuelle des pesticides sur le Bruant à face noire n’est pas bien connu (Norment et al., 2016), mais la gravité est probablement au moins légère, et pourrait être modérée à élevée.
Catégorie 1.1 : Zones résidentielles et urbaines (impact faible)
Le développement résidentiel et, dans une moindre mesure, commercial pourrait éliminer une partie de l’habitat d’hivernage convenant au Bruant à face noire (Norment et al., 2016), mais ce développement est généralement confiné aux périphéries de centres urbains existants. Il cause principalement des préoccupations en raison de la mortalité liée aux collisions avec les fenêtres (Bracey et al., 2016), mais aucune étude n’a été faite pour documenter le nombre de Bruants à face noire qui meurent ainsi.
Catégorie 4 : Corridors de transport et de service (impact faible)
La plupart des Bruants à face noire sont exposés à des routes dans leur aire d’hivernage et, puisqu’il s’agit d’oiseaux qui cherchent leur nourriture au sol, ils pourraient être menacés par des collisions avec des véhicules (Molhoff, 1979). Toutefois, seul un faible pourcentage des individus sera probablement touché, et l’effet global est donc sans doute faible. Les lignes de transport et de distribution d’électricité sont peu susceptibles de causer plus qu’un risque négligeable de collision, et leur effet pourrait en fait être neutre ou bénéfique pour le Bruant à face noire dans son aire d’hivernage puisqu’elles offrent un habitat convenable (Norment et al., 2016).
Catégorie 8.1 : Espèces exotiques (non indigènes) envahissantes (impact faible)
La prédation par les chats constitue une menace possible dans les lieux d’hivernage étant donné la grande portée de ce problème et la vulnérabilité toute particulière des oiseaux qui cherchent leur nourriture au sol (Blancher. 2013; Loss et al., 2013). Si, à ce jour, aucun rapport n’a été rédigé qui porte précisément sur la prédation du Bruant à face noire par les chats, cette préoccupation demeure crédible.
Menaces négligeables et inconnues
La hausse de la population de divers oiseaux de proie (p. ex. Épervier brun [Accipiter striatus], Épervier de Cooper [A. cooperii] et Faucon pèlerin [Falco peregrinus]) pourrait entraîner une hausse du risque de prédation pour le Bruant à face noire tout au long de son cycle annuel (Sauer et al., 2014), mais on ne sait pas vraiment à quel point, et cette menace est donc actuellement évaluée comme étant inconnue.
La conversion de prairies et de terres périphériques à des fins agricoles dans le Midwest des États-Unis pourrait être l’un des facteurs les plus importants du déclin du Bruant à face noire (Norment, comm. pers., 2016; Norment et al., 2016). À l’extrémité septentrionale de l’aire d’hivernage de l’espèce (c.-à-d. Nebraska, Dakota du Sud, Iowa, Missouri, et Kansas), quelque 530 000 ha de terres dominées par les graminées ont été converties en terres cultivées entre 2006 et 2011 (Wright et Wimberly, 2013). Des changements semblables ont été documentés dans l’aire de répartition principale en dehors de la période de reproduction, soit au Texas, au Nebraska, au Kansas et en Oklahoma; ils ont une incidence négative sur toute une série d’espèces de prairie dans cette région (Coppedge et al.,2001; Evans et Potts, 2015). L’enlèvement de haies et d’autres brise-vent lié à l’intensification de l’agriculture, et la réduction de la superficie de terres périphériques, pourraient tout particulièrement nuire à l’espèce (Norment et al., 2016). Toutefois, comme le Bruant à face noire utilise de l’habitat ouvert et pourrait se nourrir de résidus de cultures de céréales, l’incidence nette de l’agriculture (catégorie de menace 2) est inconnue pour l’instant.
L’exploitation du pétrole et du gaz ainsi que de l’énergie renouvelable (catégorie de menace 3) pourrait constituer une menace potentielle pour une vaste gamme d’espèces, y compris le Bruant à face noire, étant donné les vastes superficies nécessaires à l’aménagement de l’infrastructure (Preston et Kim, 2016). Elle peut également être une cause de mortalité, en particulier aux installations d’énergie éolienne (Zimmerling et al., 2013; Bird Studies Canada et al.,2016). Ces impacts potentiels n’ont toutefois pas été étudiés pour le Bruant à face noire en particulier, et on a par ailleurs remarqué que les emprises pourraient pour leur part profiter à l’espèce (Norment, comm. pers., 2016). Ces effets sont donc pour l’instant considérés comme négligeables à inconnus. Les effets de l’exploitation minière sont pour leur part évalués comme négligeables étant donné que jusqu’à 2,8 % de l’aire de reproduction du Bruant à face noire se trouve dans un rayon de 10 km de carrières au stade de l’exploration ou de mines (Fournier et Carrière, comm. pers., 2016). Toutefois, la principale incidence, soit l’enlèvement de la végétation aux fins d’exploitation minière, se ferait sentir sur une petite superficie, et les incidences connexes, notamment la hausse de la circulation routière, augmenteraient.
Au sein de l’aire de reproduction, le feu peut avoir une incidence négative sur le succès de nidification dans une année donnée (Black et Bliss, 1980; Arsenault et Payette, 1992; Flannigan et al., 2001), mais il peut aussi avoir un effet positif sur l’espèce en créant, dans l’habitat, des ouvertures convenant à la nidification. L’effet net des incendies et de la suppression des incendies (catégorie de menace 7.1) demeure donc inconnu.
Les changements climatiques (catégorie de menace 11) pourraient réduire la zone de toundra forestière située le long de la limite sud de l’aire de reproduction de l’espèce et ainsi, peut-être, entraîner un déplacement géographique vers le nord (Rizzo, 1988; Zoltai, 1988; Payette et al., 2001). La réaction de l’espèce au déplacement de la limite forestière vers le nord est documentée par les changements relevés dans les observations de l’espèce aux environs de Churchill, au Manitoba, depuis les années 1930, lorsque l’espèce y était plus commune qu’aujourd’hui (Jehl et Smith, 1970; Jehl, 2004; Di Labio, comm. pers., 2015). De plus, les changements climatiques pourraient augmenter la violence des phénomènes météorologiques, ce qui réduirait le taux de survie des oisillons (Norment et al., 2016). L’élargissement de la répartition des mammifères prédateurs, par exemple le renard roux (Vulpes vulpes) (Naughton, 2012), et des ectoparasites volants (p. ex. brûlots [diptères de la famille des Cératopogonidés] et moustiques [diptères de la famille des Culicidés]) (Tomás et al., 2008) que risquent d’entraîner les changements climatiques pourrait avoir une incidence négative sur les oisillons du Bruant à face noire.
Protection, statuts et classements
Statuts et protection juridique
Au Canada, le Bruant à face noire, ses nids et ses œufs sont protégés par la Loi sur la convention concernant les oiseaux migrateurs (1994). Cette loi interdit la vente et la possession d’oiseaux migrateurs et de leurs nids de même que toute activité nuisant aux oiseaux migrateurs, à leurs œufs ou à leurs nids, sauf exceptions autorisées par le Règlement sur les oiseaux migrateurs. L’espèce est protégée de façon semblable aux États-Unis aux termes de la Migratory Birds Treaty Act (1918).
Statuts et classement non juridiques
Le Bruant à face noire est classé « préoccupation mineure » (Least Concern) par BirdLife International (2015) dans la liste rouge de l’UICN, et « non en péril à l’échelle mondiale » (G5; revu en 1996) par NatureServe Explorer (2015). Au Canada, l’espèce est désignée « non en péril, individus nicheurs et non nicheurs » (N5B et N5N); au Nunavut, l’espèce est « non classée » (SNR), en Saskatchewan, elle est classée « non en péril » (S5), tandis qu’au Manitoba et dans les Territoires du Nord-Ouest, l’espèce est classée « vulnérable » (S3; NatureServe Explorer 2015). Au Texas, l’espèce est classée « apparemment non en péril » (S4; NatureServe Explorer, 2015). L’espèce n’a pas été classée ailleurs.
Protection et propriété de l’habitat
On signale que le Bruant à face noire se reproduit dans le parc national Wapusk, au Manitoba, et c’est probablement aussi le cas dans le parc national Tuktut Nogait, dans les Territoires du Nord-Ouest. Des individus pourraient parfois passer par d’autres aires protégées durant leur migration ou en fréquenter durant l’hivernage, mais de manière générale, la protection juridique de l’habitat de l’espèce est limitée dans toute son aire de répartition annuelle.
Remerciements et experts contactés
Beaucoup de personnes ont répondu à notre demande générale de renseignements sur le Bruant à face noire au Canada et aux États-Unis : Ken Abraham, Christian Artuso, Bruce Bennett, David Britton, Dick Cannings, Syd Cannings, Suzanne Carrière, Vanessa Charlwood, Mike Chutter, Bruce Di Labio, Kiel Drake, Cameron Eckert, Bonnie Fournier, Marcel Gahbauer, Samuel Hache, Audrey Heagy, Reid Hildebrandt, Rudolf Koes, Tom Jung, Bruce MacDonald, Scott MacDougall-Shackleton, Stuart Mackenzie, Jon McCracken, Patrick Nantel, Chris Norment, Joachim Obst, Rhiannon Pankratz, Lisa Pirie-Dominix, Jennie Rausch, James Rising, Rich Russell, Pamela Sinclair, Sonia Schnobb, Don Sutherland, Doug Tate, Phil Taylor, Doug Tozer et Ken Tuininga. Le rédacteur du rapport tient à remercier en plus de ces personnes, qui ont toutes fourni de l’information, les centres de données sur la conservation et les centres d’information sur le patrimoine naturel concernés, de même que l’Agence Parcs Canada, pour avoir donné des renseignements sur l’espèce.
Le présent rapport a grandement profité de commentaires reçus des personnes suivantes : Mike Burrell, Richard Elliot, Marcel Gahbauer, Elaine Gosnell, Lillian Knopf, Nathan Miller et David Stephenson.
Doug Tozer a aimablement aidé à l’analyse statistique des données du Recensement des oiseaux de Noël, élément essentiel du présent rapport de situation. Lillian Knopf a créé les figures 4 et 5. Gerry Schaus a créé la figure 1 et effectué le calcul de la zone d’occurrence avec les conseils de Jenny Wu.
Sources d’information
American Ornithologists’ Union (AOU). 1998. Check-list of North American birds: the species of birds of North America from the Arctic through Panama, including the West Indies and Hawaiian Islands. Seventh edition, American Ornithologists’ Union, Washington, D.C.
Arsenault, D., et S. Payette. 1992. A post-fire shift from lichen-spruce to lichen-tundra vegetation at tree line. Ecology 73:1007-1018.
Artuso, C., comm. pers. 2016. Correspondance par courriel adressée à K.G.D. Burrell. Janvier 2016. Gestionnaire de projets du Manitoba, Études d’Oiseaux Canada, Winnipeg, Manitoba.
BirdLife International. 2015. Species factsheet: Zonotrichia querula. Site Web : http://www.birdlife.org [consulté en décembre 2015].
Bird Studies Canada. 2016. About the Canadian Migration Monitoring Network. Site Web : http://www.birdscanada.org/volunteer/cmmn/index.jsp?targetpg=about [consulté en octobre 2016]. (Également disponible en français : Études d’oiseaux Canada. 2016. [Réseau canadien de surveillance des migrations] : À propos de nous http://www.birdscanada.org/volunteer/cmmn/index.jsp?lang=FR&targetpg=about)
Bird Studies Canada, Canadian Wind Energy Association, Environment Canada, and Ontario Ministry of Natural Resources and Forestry. 2016. Wind energy bird and bat monitoring database summary of the findings from post-construction monitoring reports. July 2016. 47pp. Site Web : http://www.bsc-eoc.org/birdmon/wind/resources.jsp?dir=reports [consulté en octobre 2016]. (Également disponible en français : Études d’Oiseaux Canada, Environnement et Changement climatique Canada, l’Association canadienne de l’énergie éolienne, et du ministère des Richesses naturelles et des Forêts de l’Ontario. 2016. Base de données du Suivi des populations d’oiseaux et de chauves-souris relié à l’énergie éolienne : Sommaire de l’information présentée dans les rapports de suivi postérieur à la construction. Juillet 2016. 48 p. https://www.bsc-eoc.org/resources/wind/Jul2016_sommaire_base_de_donnees.pdf.)
Black, R.A., et L.C. Bliss. 1980. Reproductive ecology of Picea mariana (Mill.) BSP, at tree line near Inuvik, Northwest Territories, Canada. Ecological Monographs 50:331-354.
Blancher, P.J., K.V. Rosenberg, A.O. Panjabi, B. Altman, J. Bart, C.J. Beardmore, G.S. Butcher, D. Demarest, R. Dettmers, E.H. Dunn, W. Easton, W.C. Hunter, E.E. Iñigo-Elias, D.N. Pashley, C.J. Ralph, T.D. Rich, C.M. Rustay, J.M. Ruth et T.C. Will. 2007. Guide to the Partners in Flight Population Estimates Database. Version: North American Landbird Conservation Plan 2004. Partners in Flight Technical Series No 5.
Blancher, P.J. 2013. Estimated number of birds killed by house cats (Felis catus) in Canada. Avian Conservation and Ecology 8(2): 3.
Bracey, A.M., M.A. Etterson, G.J. Niemi et R.F. Green. 2016. Variation in bird-window collision mortality and scavenging rates within an urban landscape. The Wilson Journal of Ornithology 128: 355-367.
Brewer, D., A. Diamond, E.J. Woodsworth, B.T. Collins et E.H. Dunn. 2006. Canadian Atlas of Bird Banding. Volume 1: Doves, Cuckoos, and Hummingbirds through Passerines, 1921-1995, 2nd edition. Site Web : http://www.ec.gc.ca/aobc-cabb/index.aspx?lang=En [consulté en novembre 2015]. (Également disponible en français : Brewer, D., A. Diamond, E.J. Woodsworth, B.T. Collins et E.H. Dunn. 2006. Atlas des oiseaux bagués ou repris au Canada. Volume 1 : Pigeons et tourterelles, coulicous, colibris et passereaux, 1921-1995, 2e éd. http://www.ec.gc.ca/aobc-cabb/index.aspx?lang=fr.)
Bridgwater, D.D. 1966. Winter movements and habitat use by Harris’ sparrow, Zonotrichia querula (Nuttall). Proceedings of the Oklahoma Academy of Science 46:53-59.
Cadman, M.D., P.F.J. Eagles et F.M. Helleiner (eds.). 1987. Atlas of the Breeding Birds of Ontario. University of Waterloo Press, Waterloo, Ontario. 617 pp.
Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage et A.R. Couturier, eds. 2007. Atlas of the Breeding Birds Of Ontario, 2001-2005. Bird Studies Canada, Environment Canada, Ontario Field Ornithologists’, Ontario Ministry of Natural Resources, and Ontario Nature, Toronto, Ontario, xxii + 706 pp. (Également disponible en français : Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage et A.R. Couturier [éd.]. 2007. Atlas des oiseaux nicheurs de l’Ontario, 2001-2005. Environnement Canada, Études d’Oiseaux Canada, le ministère des Richesses naturelles de l’Ontario, Ontario Field Ornithologists et Ontario Nature, Toronto, Ontario, xxii + 728 p.)
Cadman, M.D. 2007. Harris’s Sparrow, pp. 568-569 in Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage et A.R. Couturier, eds. Atlas of the Breeding Birds Of Ontario, 2001-2005. Bird Studies Canada, Environment Canada, Ontario Field Ornithologists’, Ontario Ministry of Natural Resources, and Ontario Nature, Toronto, Ontario, xxii + 706 pp. (Également disponible en français : Cadman, M.D. 2007. Bruant à face noire, p. 568-569 in Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage et A.R. Couturier [éd.]. Atlas des oiseaux nicheurs de l’Ontario, 2001-2005. Environnement Canada, Études d’Oiseaux Canada, le ministère des Richesses naturelles de l’Ontario, Ontario Field Ornithologists et Ontario Nature, Toronto, Ontario, xxii + 728 p.)
Carrière, S., comm. pers. 2016. Correspondance par courriel adressée à K.G.D. Burrell. Septembre 2016. Biologiste de la faune, gouvernement des Territoires du Nord-Ouest, Yellowknife (Territoires du Nord-Ouest).
Chesser, R.T., R.C. Banks, K.J. Burns, C. Cicero, J.L. Dunn, A.W. Kratter, I.J. Lovette, A.G. Navarro-Sigüenza, P.C. Rasmussen, J.V. Remsen, Jr., J.D. Rising, D.F. Stotze et K. Winker. 2015. Fifty-sixth Supplement to the American Ornithologists’ Union Check-list of North American Birds. Auk 132:748-764.
Chilton, G., M.C. Baker, C.D. Barrentine et M.A. Cunningham. 1995. White-crowned Sparrow (Zontotrichia leucophrys), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology. Site Web : http://bna.birds.cornell.edu/bna/species/183 [consulté en octobre 2016].
Clarke, C.H.D. 1944. Notes on the status and distribution of certain mammals and birds in the Mackenzie River and western arctic area in 1942 and 1943. Canadian Field-Naturalist 58:97-103.
Coppedge, B.R., D.M. Engle, R.E. Masters et M.S. Gregory 2001. Avian response to landscape change in fragmented southern Great Plains grasslands. Ecological Applications 11:47-59.
Di Labio, B., comm. pers. 2015. Correspondance par courriel adressée à K.G.D. Burrell. Juillet 2015. Guide d’ornithologie, Di Labio Birding, Ottawa (Ontario).
eBird. 2016. eBird: An online database of bird distribution and abundance. Cornell Lab of Ornithology, Ithaca, New York. Site Web : http://www.eBird.org [consulté en août 2016].
Eckert, C., comm. pers. 2015. Correspondance par courriel adressée à K.G.D. Burrell. Juillet 2015. Biologiste de la conservation, gouvernement du Yukon, Whitehorse (Yukon).
Environment and Climate Change Canada (ECCC). 2016. eBird Canada (Northwest Territories/Nunavut). Site Web : https://www.ec.gc.ca/reom-mbs/default.asp?lang=En&n=60E48D07-1 [consulté en octobre 2016]. (Également disponible en français : Environnement et Changement climatique Canada [ECCC]. 2016. eBird Canada [Territoires du Nord-Ouest/Nunavut] https://www.ec.gc.ca/reom-mbs/default.asp?lang=Fr&n=60E48D07-1.)
Evans, S.G., et M.D. Potts. 2015. Effect of agricultural commodity prices on species abundance of US grassland birds. Environmental and Resource Economics 62:549-565.
Flannigan, M., I. Campbell, M. Wotton et C. Carcaillet. 2001. Future fire in Canada’s boreal forest: paleoecology results and general circulation model - regional climate model simulations. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestière 31:854-864.
Gibbons, D., C. Morrissey et P. Mineau. 2015. A review of the direct and indirect effects of neonicotinoids and fipronil on vertebrate wildlife. Environmental Science and Pollution Resources 22:103-118.
Gillespie, W.L., et S.C. Kendeigh. 1982. Breeding bird populations of northern Manitoba. Canadian Field-Naturalist 96:272-281.
Government of Canada. 2013. EcoZones, EcoRegions, and EcoDistricts. Site Web : http://sis.agr.gc.ca/cansis/nsdb/ecostrat/hierarchy.html [consulté en décembre 2015]. (Également disponible en français : Gouvernement du Canada. 2013. Écozones, écorégions et Écodistricts http://sis.agr.gc.ca/siscan/nsdb/ecostrat/hierarchy.html.)
Graul, W.D. 1967. Population movements of the Harris’s Sparrow and tree sparrow in eastern Kansas. Southwestern Naturalist 12:303-310.
Hallmann, C.A., R.P.B. Foppen, C.A.M. van Turnhout, H. de Kroon et E. Jongejans. 2014. Declines in insectivorous birds are associated with high neonicotinoid concentrations. Nature 511:341-343.
Harris, W.C., E.A. Johnson, S.J.B. Johnson et K.M. Traynor. 1974. Upland lichen woodland. American Birds 28:1048-1049.
Harper, F. 1953. Birds of the Nueltin Lake expedition. American Midland Naturalist 49:1-116.
James, R.D., P.L. McLaren et J.C. Barlow. 1976. Annotated checklist of the birds of Ontario. Life Science Miscellaneous Publications, Royal Ontario Museum, Toronto, Ontario.
Jehl, J.R., et B.A. Smith. 1970. Birds of the Churchill Region, Manitoba. Manitoba Museum of Nature, Winnipeg, Manitoba.
Jehl, J. 2004. Birdlife of the Churchill Region: status, history, biology. Manitoba Special Conservation Fund and Churchill Northern Studies Center, Churchill, Manitoba.
Loss, S.R., T. Will et P.P. Marra. 2013. The impact of free-ranging domestic cats on wildlife in the United States. Nature Communications 4(1396) (doi:10.1038/ncomms2380).
Manitoba Breeding Bird Atlas (MBBA). 2015. Welcome to the Manitoba Breeding Bird Atlas. Site Web : http://www.birdatlas.mb.ca/ [consulté en décembre 2015] (Également disponible en français : Atlas des oiseaux nicheurs du Manitoba [AONM]. 2015. Bienvenue sur le site de l’Atlas des oiseaux nicheurs du Manitoba! http://www.birdatlas.mb.ca/index_fr.jsp.)
Migratory Birds Convention Act (MBCA). 1994. Migratory Birds Convention Act, 1994. Site Web : http://laws.justice.gc.ca/en/M-8.01/ [consulté en décembre 2015]. (Également disponible en français : Loi sur la convention concernant les oiseaux migrateurs [LCCOM]. 1994. Loi de 1994 sur la convention concernant les oiseaux migrateurs http://laws-lois.justice.gc.ca/fra/lois/m-7.01/.)
Mollhoff, W. 1979. Tower kills. Nebraska Bird Review 47:58.
National Audubon Society. 2015. The Christmas Bird Count Historical Results. Site Web : http://www.christmasbirdcount.org [consulté en novembre 2015].
NatureCounts. 2016. Canadian Migration Monitoring Network: Population trends and seasonal abundance. Site Web : http://www.bsc-eoc.org/birdmon/cmmn/popindices.jsp [consulté en octobre 2015]. (Également disponible en français : NatureCounts. 2016. Réseau canadien de surveillance des migrations - [Indices de population] (https://www.bsc-eoc.org/birdmon/cmmn/popindices.jsp?switchlang=FR.)
NatureServe Explorer. 2015. Harris’s Sparrow Zonotrichia querula. Site Web : http://explorer.natureserve.org/servlet/NatureServe?searchName=Zonotrichia%20querula [consulté en décembre 2015].
Naughton, D. 2012. The natural history of Canadian mammals. Canadian Museum of Nature. Ottawa, Ontario. 824pp. (Également disponible en français : Naughton, D. 2016. Histoire naturelle des mammifères du Canada. Musée canadien de la nature, Ottawa, Ontario; Éditions Michel Quintin, Waterloo, Québec. 858 p.)
Nice, M.M. 1929. The Harris’ sparrow in central Oklahoma. Condor 31:57-61.
Norment, C.J. 1992a. The comparative breeding ecology of the Harris’s Sparrow (Zonotrichia querula) and White-crowned Sparrow (Zonotrichia leucophrys) in the Northwest Territtories, Canada. Thèse de doctorat. University of Kansas, Lawrence, Kansas.
Norment, C.J. 1992b. Comparative breeding biology of Harris’s Sparrows (Zonotrichia querula) and Gambel’s White-crowned Sparrows (Zonotrichia leucophrys gambelii) in the Northwest Territories, Canada. Condor 94:955-975.
Norment, C.J. 1993. Nest-site characteristics and nest predation in Harris’s Sparrows and White-crowned Sparrows in the Northwest Territories, Canada. Auk 110:769-777.
Norment, C.J. 1994. Breeding site fidelity in Harris’ Sparrows, Zonotrichia querula, in the Northwest Territories. Canadian Field-Naturalist 108:234-236.
Norment, C.J. 1995a. Incubation patterns in Harris’ Sparrows and White-Crowned Sparrows in the Northwest Territories, Canada. Journal of Field Ornithology 66:553-563.
Norment, C.J. 1995b. Prebasic (postnuptial) molt in free-ranging Harris’s Sparrows, Zonotrichia querula, in the Northwest Territories, Canada. The Canadian Field-Naturalist 109:470-475.
Norment, C.J. 2003. Patterns of nestling feeding in Harris’s Sparrows, Zonotrichia querula and White-crowned Sparrows, Z. leucophrys, in the Northwest Territories, Canada. Canadian Field-Naturalist 117:203-208.
Norment, C., comm. pers. 2016. Correspondance par courriel adressée à K.G.D. Burrell. Septembre 2016. Professeur et président, Dept. of Environmental Science and Biology, The College at Brockport, State University of New York, Brockport, New York.
Norment, C.J., et M.E. Fuller. 1997. Breeding-season frugivory by Harris’s sparrows (Zonotrichia querula) and white-crowned sparrows (Zonotrichia leucophrys) in a low-arctic ecosystem. Canadian Journal of Zoology 75:670-679.
Norment, C.J., S.A. Shackleton, D.J. Watt, P. Pyle et M.A. Patten. 2016. Harris’s Sparrow (Zonotrichiaquerula), The Birds of North America (P.G. Rodewald, Ed.). Cornell Lab of Ornithology, Ithaca, New York. Site Web : https://birdsna.org/Species-Account/bna/species/harspa [consulté en octobre 2016].
Obst, J., comm. pers. 2015. Correspondance par courriel adressée à K.G.D. Burrell. Septembre 2015. Technicien de terrain, Daring Lake Natural Heritage Study, Yellowknife (Territoires du Nord-Ouest).
Payette, S., M.J. Fortin et I. Gamache. 2001. The subarctic forest-tundra: the structure of a biome in a changing climate. Bioscience 51:709-718.
Payne, R.B. 1979. Two apparent hybrid Zonotrichia sparrows. Auk 96:595-599.
Preston, T.M., et K. Kim. 2016. Land cover changes associated with recent energy development in the Williston Basin; Northern Great Plains, USA. Science of the Total Environment 566-567: 1511-1518.
Pyle, P. 1997. Identification guide to North American birds: Columbidae to Ploceidae. Part I. Slate Creek Press, Bolinas, California.
Rees, W.R. 1973. Comparative ecology of three sympatric species of Zonotrichia. Thèse de doctorat. University of Toronto, Toronto, Ontario.
Rizzo, B. 1988. The sensitivity of Canada’s ecosystems to climatic change. Canadian Community Ecological Land Classification Newsletter 17:10-12.
Robel, R.J., J.F. Keating, J.L. Zimmerman, K.C. Behnke et K.E. Kemp. 1997. Consumption of coloured and flavored food morsels by Harris’s and American Tree Sparrows. Wilson Bulletin 109:218-225.
Rosenberg, K.V., J.A. Kennedy, R. Dettmers, R.P. Ford, D. Reynolds, J.D. Alexander, C.J. Beardmore, P.J. Blancher, R.E. Bogart, G.S. Butcher, A. F. Camfield, A. Couturier, D.W. Demarest, W.E. Easton, J.J. Giocomo, R.H. Keller, A.E. Mini, A.O. Panjabi, D.N. Pashley, T.D. Rich, J.M. Ruth, H. Stabins, J. Stanton et T. Will. 2016. Partners in Flight Landbird Conservation Plan: 2016 Revision for Canada and Continental United States. Partners in Flight Science Committee.
R Core Team. 2015. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Site Web : http://www.R-project.org/ [consulté en décembre 2015].
Sauer, J.R., J.E. Hines, J.E. Fallon, K.L. Pardieck, D.J. Ziolkowski, Jr. et W.A. Link. 2014. The North American Breeding Bird Survey, Results and Analysis 1966-2013. Version 01. 30.2015. USGS Patuxent Wildlife Research Center, Laurel, Maryland.
Sealy, S.G. 1967. Range extension of the Harris’s Sparrow. Canadian Field-Naturalist 81:152-153.
Semple, J.B., et G.M. Sutton. 1932. Nesting of Harris’s Sparrow Zonotrichia querula at Churchill, Manitoba. Auk 49:166-183.
Shuman, T.W., R.J. Robel, J.L. Zimmerman et K.E. Kemp. 1992. Time budgets of confined Northern Cardinals and Harris’s Sparrows in flocks of different size and composition. Journal of Field Ornithology 63:129-137.
Sibley, D.A. 2000. The Sibley Guide to Birds. National Audubon Society, NY.
Smith, A.C., M.R. Hudson, C. Downes et C.M. Francis. 2014. Estimating breeding bird survey trends and annual indices for Canada: how do the new hierarchical Bayesian estimates differ from previous estimates? Canadian Field-Naturalist 128:119-134.
Thompson, F.R., III. 1994. Temporal and spatial patterns of breeding brown-headed cowbirds in the midwestern United States. Auk 111:979-990.
Tomás, G., S. Merino, J. Martínez-de la Puente, J. Moreno, J. Morales et E. Lobato. 2008. Determinants of abundance and effects of blood-sucking flying insects in the nest of a hole-nesting bird. Oecologia 156: 305-312.
Wright, C.K., et M.C. Wimberly. 2013. Recent land use change in the Western Corn Belt threatens grasslands and wetlands. Proceedings of the National Academy of Sciences 110: 4134-4139.
Zimmerling, J.R., A.C. Pomeroy, M.V. d’Entremont et C.M. Francis. 2013. Canadian estimate of bird mortality due to collisions and direct habitat loss associated with wind turbine developments. Avian Conservation and Ecology, 8(2):10.
Zoltai, S.C. 1988. Ecoclimatic provinces of Canada and man-induced climatic change. Canadian Community Ecological Land Classification Newsletter 17:12-15.
Sommaire biographique du rédacteur du rapport
Kenneth Burrell est un biologiste travaillant pour Natural Resource Solutions Inc. (NRSI), société d’experts-conseils en environnement située à Waterloo, en Ontario. Il est le principal expert-conseil en ornithologie de cette société, et il se spécialise dans l’inventaire et l’évaluation des ressources naturelles, la recherche et les études d’impact. M. Burrell est membre actif de la communauté des ornithologues de l’Ontario, notamment de eBird et de l’organsime Ontario Field Ornithologists, où il siège au conseil d’administration depuis 2014, et de l’Ontario Bird Records Committee depuis 2011. Depuis qu’il a décroché une maîtrise ès sciences en 2013, M. Burrell a publié des articles sur une vaste gamme de sujets liés à l’ornithologie de terrain, notamment les vols de réorientation printaniers dans la région de Pelee, les parulines hybrides, eBird et ses utilités, et plus récemment l’assignation des lieux de provenance des oiseaux au moyen des isotopes stables.
Collections examinées
Aucune collection n’a été examinée.
Annexe 1. Tableau d’évaluation des menaces pour le Bruant à face noire
Annexe 1. tableau d’évaluation des menaces.
Tableau d’évaluation des menaces
- Nom scientifique de l’espèce ou de l’écosystème
- Bruant à face noire
- Code de l’élément
- ABPBXA4050
- Date
- 26/09/2016
- Évaluateur(s) :
- Kenneth Burrell, Dave Fraser, Marcel Gahbauer, Suzanne Carrière, Richard Elliot, Pam Sinclair, Myles Lamont, Chris Norment, Rudolf Koes, Jim Rising, Samuel Hache, Amy Ganton, Joanna James
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l’intensité de leur impact : Maximum de la plage d’intensité |
Comptes des menaces de niveau 1 selon l’intensité de leur impact : Minimum de la plage d’intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 1 | 0 |
| C | Moyen | 0 | 0 |
| D | Faible | 3 | 4 |
| - | Impact global des menaces calculé : | Élevé | Moyen |
- Impact global des menaces attribué :
- BC = Élevé-Moyen
- Commentaires sur l’impact global des menaces
- Les cotes dans plusieurs catégories sont « Inconnu », et les données sur les tendances sont plus limitées que pour certains autres oiseaux. Cependant, la plage moyen-élevé est probablement appropriée d’après la meilleure information accessible.
| Menace | Menace | Impact (calculé) | Impact (calculé) | Portée (10 prochaines années) | Gravité (10 ans ou 3 générations) | Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial | D | Faible | Grande (31-70 %) | Légère (1-10 %) | Élevée (menace toujours présente) | Dans l’ensemble, la population humaine est faible au sein de l’aire de reproduction et ne devrait pas avoir une incidence sur l’espèce. Au sein de l’aire d’hivernage, le développement pourrait détruire de l’habitat convenable, mais essentiellement à la périphérie des zones urbaines existantes. La plus grande préoccupation liée à cette menace pourrait être la mortalité due aux collisions avec les fenêtres, mais l’ampleur de ce problème n’a pas été documentée pour cette espèce. |
| 1.1 | Zones résidentielles et urbaines | D | Faible | Grande (31-70 %) | Légère (1-10 %) | Élevée (menace toujours présente) | Cette menace a principalement trait à la hausse du nombre de résidences dans l’aire d’hivernage, et plus précisément au risque de mortalité due aux collisions avec les fenêtres et aux effets des résidences en zones urbaines. Toutefois, l’expansion urbaine au sein de l’aire d’hivernage ne se fait généralement pas rapidement. Bien que cette espèce fréquente les mangeoires des résidences et pourrait donc tolérer une certaine suburbanisation, cette menace augmente également le risque de collisions avec des fenêtres. Cela peut aussi poser un risque dans certains secteurs du couloir migratoire, mais nous n’avons pas de preuves spécifiques pour l’affirmer. |
| 1.2 | Zones commerciales et industrielles | cellule vide | Négligeable | Négligeable (<1 %) | Inconnue | Élevée (menace toujours présente) | cellule vide |
| 1.3 | Zones touristiques et récréatives | cellule vide | Négligeable | Négligeable (<1 %) | Légère (1-10 %) | Modérée (peut-être dans le futur [court terme, < 10 ans]) | Il pourrait y avoir un impact faible dans toute l’aire de reproduction en raison de la présence de gîtes, mais on ne sait pas dans quelle mesure. La cote s’applique à toute l’aire de répartition (cycle de vie complet) de l’espèce. |
| 2 | Agriculture et aquaculture | cellule vide | Inconnu | Inconnue | Inconnue | Inconnue | |
| 2.1 | Cultures annuelles et pérennes de produits autres que le bois | cellule vide | Inconnu | Inconnue | Inconnue | Inconnue | La menace s’applique principalement à l’aire d’hivernage et, dans une moindre mesure, aux routes migratoires, mais pas à l’aire de reproduction. L’espèce préfère l’habitat ouvert adjacent à des abris (p. ex. haies), et la perte ou la conversion d’un tel habitat pourrait donc constituer une menace, mais il faudrait avoir davantage de données pour comprendre la portée et la gravité éventuelles de cette menace. |
| 2.2 | Plantations pour la production de bois et de pâte | cellule vide | Négligeable | Négligeable (<1 %) | Inconnue | Élevée (menace toujours présente) | Cette menace pourrait être minimalement présente dans l’aire d’hivernage. |
| 2.3 | Élevage de bétail | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | Les pâturages peuvent constituer un bon habitat comportant un mélange de zones herbeuses et arbustives. |
| 2.4 | Aquaculture en mer et en eau douce | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 3 | Production d’énergie et exploitation minière | cellule vide | Négligeable | Négligeable (<1 %) | Extrême – élevée (31-100 %) | Élevée (menace toujours présente) | |
| 3.1 | Forage pétrolier et gazier | cellule vide | Inconnu | Petite (1-10 %) | Inconnue | Modérée (peut-être dans le futur [court terme, < 10 ans]) | La portée est minime au sein de l’aire de reproduction, un peu plus importante dans l’aire d’hivernage, mais la gravité n’est pas bien connue puisque l’habitat que constituent les emprises des pipelines peut être attrayant pour l’espèce, et que la perturbation est limitée aux périodes d’aménagement. Des activités de fracturation ont lieu dans l’aire d’hivernage, mais leurs effets sur le Bruant à face noire n’ont pas été documentés. |
| 3.2 | Exploitation de mines et de carrières | cellule vide | Négligeable | Négligeable (<1 %) | Extrême – élevée (31-100 %) | Élevée (menace toujours présente) | Certaines carrières au stade de l’exploration et mines seront probablement aménagées au sein de l’aire de reproduction dans un proche avenir. Si l’on tient compte d’une zone tampon prudente de 10 km autour de chaque mine proposée, cela pourrait représenter 2,8 % de l’aire de reproduction de l’espèce, mais l’effet principal, soit la perte d’habitat directe, ne touchera qu’une fraction de cette superficie. Des effets moins graves liés au bruit et à la poussière pourraient cependant être ressentis à une plus grande distance. Cette menace ne touche pas les lieux d’hivernage. |
| 3.3 | Énergie renouvelable | cellule vide | Négligeable | Négligeable (<1 %) | Inconnue | Élevée (menace toujours présente) | Des projets d’énergie solaire et éolienne ont été aménagés dans l’aire d’hivernage, et d’autres suivront probablement, mais leur effet sur l’espèce n’est pas bien connu. |
| 4 | Corridors de transport et de service | D | Faible | Généralisée (71-100 %) | Légère (1-10 %) | Élevée (menace toujours présente) | cellule vide |
| 4.1 | Routes et voies ferrées | D | Faible | Généralisée (71-100 %) | Légère (1-10 %) | Élevée (menace toujours présente) | La plupart des individus sont probablement exposés à des routes et des voies ferrées dans l’aire d’hivernage; on trouve peu de ces structures dans l’aire de reproduction, mais leur nombre augmente un peu. Comme il n’existe aucune preuve montrant que l’espèce est particulièrement vulnérable aux collisions avec des véhicules, la gravité est considérée comme légère. |
| 4.2 | Lignes de services publics | cellule vide | Inconnu | Généralisée – grande (31-100 %) | Inconnue | Élevée (menace toujours présente) | Beaucoup d’individus, voire la plupart, sont sans doute exposés à des lignes de services publics durant leur migration et dans l’aire d’hivernage, mais le risque de collision est probablement minime, et dans certaines situations, les emprises ont peut-être un effet positif puisqu’elles offrent un habitat convenable à l’espèce; somme toute, la gravité demeure inconnue. |
| 4.3 | Voies de transport par eau | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 4.4 | Corridors aériens | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 5 | Utilisation des ressources biologiques | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 5.1 | Chasse et capture d’animaux terrestres | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | |
| 5.2 | Cueillette de plantes terrestres | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 5.3 | Exploitation forestière et récolte du bois | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 5.4 | Pêche et récolte de ressources aquatiques | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 6 | Intrusions et perturbations humaines | cellule vide | Négligeable | Négligeable (<1 %) | Inconnue | Élevée (menace toujours présente) | cellule vide |
| 6.1 | Activités récréatives | cellule vide | Négligeable | Restreinte (11-30 %) | Négligeable (<1 %) | Élevée (menace toujours présente) | Les populations humaines sont très peu nombreuses dans toute l’aire de reproduction, et même dans l’aire d’hivernage, les effets des activités récréatives sont sans doute négligeables. |
| 6.2 | Guerre, troubles civils et exercices militaires | cellule vide | Négligeable | Négligeable (<1 %) | Inconnue | Élevée (menace toujours présente) | cellule vide |
| 6.3 | Travail et autres activités | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 7 | Modifications des systèmes naturels | cellule vide | Inconnu | Restreinte (11-30 %) | Inconnue | Élevée (menace toujours présente) | cellule vide |
| 7.1 | Incendies et suppression des incendies | cellule vide | Inconnu | Restreinte (11-30 %) | Inconnue | Élevée (menace toujours présente) | On sait que des feux de forêt se produisent à l’extrémité sud de l’aire de répartition de l’espèce. Ceux-ci peuvent avoir une incidence négative (p. ex. perte de nids), mais aussi positive (p. ex. création de nouvel habitat); la gravité est donc considérée comme inconnue pour l’instant. |
| 7.2 | Gestion et utilisation de l’eau et exploitation de barrages | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 7.3 | Autres modifications de l’écosystème | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 8 | Espèces et gènes envahissants ou autrement problématiques | D | Faible | Grande (31-70 %) | Légère (1-10 %) | Élevée (menace toujours présente) | cellule vide |
| 8.1 | Espèces exotiques (non indigènes) envahissantes | D | Faible | Grande (31-70 %) | Légère (1-10 %) | Élevée (menace toujours présente) | La prédation par les chats est préoccupante pour la plupart des oiseaux terrestres, et elle cause sans doute une certaine mortalité dans l’aire d’hivernage et dans certaines parties des routes migratoires. |
| 8.2 | Espèces indigènes problématiques | cellule vide | Inconnu | Inconnue | Inconnue | Élevée (menace toujours présente) | La hausse de la population de divers oiseaux de proie (p. ex. Épervier brun, Épervier de Cooper et Faucon pèlerin) pourrait accroître les pressions de prédation tout au long du cycle annuel, mais on ne possède pas de données propres au Bruant à face noire. Dans l’aire de reproduction, l’expansion vers le nord de la répartition des renards roux et d’insectes hématophages et la hausse de leur abondance pourraient aussi poser un risque pour le Bruant à face noire, mais des recherches devront être menées pour en établir les effets possibles. |
| 8.3 | Matériel génétique introduit | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 9 | Pollution | BD | Élevé – faible | Généralisée – grande (31-100 %) | Légère (1-10 %) | Élevée (menace toujours présente) | cellule vide |
| 9.1 | Eaux usées domestiques et urbaines | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 9.2 | Effluents industriels et militaires | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 9.3 | Effluents agricoles et sylvicoles | BD | Élevé – faible | Généralisée - grande (31-100 %) | Légère (1-10 %) | Élevée (menace toujours présente) | Le Bruant à face noire pourrait être exposé à des pesticides (comme les néonicotinoïdes) dans son aire d’hivernage et durant la migration, surtout qu’il se nourrit de graines. Si la gravité de cet effet n’est pas bien connue, l’incidence est presque certainement négative et pourrait être considérable en raison d’effets autant directs (consommation de graines) qu’indirects (baisse du nombre d’insectes proies). |
| 9.4 | Déchets solides et ordures | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 9.5 | Polluants atmosphériques | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 9.6 | Apports excessifs d’énergie | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 10 | Phénomènes géologiques | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 10.1 | Volcans | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 10.2 | Tremblements de terre et tsunamis | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 10.3 | Avalanches et glissements de terrain | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 11 | Changements climatiques et phénomènes météorologiques violents | cellule vide | Non calculé (en dehors de la période d’évaluation) | Généralisée (71-100 %) | Inconnue | Faible (peut-être dans le futur [long terme, > 10 ans]) | cellule vide |
| 11.1 | Déplacement et altération de l’habitat | cellule vide | Inconnu | Petite (1-10 %) | Inconnue | Élevée (menace toujours présente) | Il existe certaines preuves de déplacement de la répartition du fait des changements climatiques, qu’on pense à l’accroissement de la couverture arbustive dans certains secteurs de l’aire de reproduction et à la réduction d’occurrence dans les environs de Churchill, au Manitoba, possiblement en raison de l’assèchement dû au rehaussement isostatique. Dans l’ensemble, on s’attend à voir un déplacement de la répartition vers le nord avec l’accroissement des températures, mais on ne sait pas si cela aura un effet net sur la population. |
| 11.2 | Sécheresses | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| 11.3 | Températures extrêmes | cellule vide | Non calculé (en dehors de la période d’évaluation) | Généralisée (71-100 %) | Inconnue | Faible (peut-être dans le futur [long terme, > 10 ans]) | L’espèce est vulnérable aux variations de température extrêmes durant la nidification, les oisillons connaissant alors des problèmes de thermorégulation. Il est possible que les changements climatiques donnent lieu à un accroissement des variations de température, mais probablement seulement à long terme, et la gravité est difficile à prédire pour l’instant. |
| 11.4 | Tempêtes et inondations | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
Classification des menaces d’après l’IUCN-CMP, Salafsky et al. (2008).