
Évaluation et Rapport de situation du COSEPAC sur le Fissident pygmée Fissidens exilis au Canada - 2016

- Table des matières
- Sommaire de l'évaluation
- Résumé
- Résumé technique
- Préface
- Description et importance de l'espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Activités récréatives
- Barrages et gestion de l'eau
- Agriculture et effluents agricoles
- Exploitation forestière et récolte du bois
- Corridors terrestres de transport et de service
- Développement résidentiel et commercial
- Production d'énergie et exploitation minière
- Changements climatiques
- Nombre de localités
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d'information
- Sommaire biographique de la rédactrice du rapport
- Collections examinées
- Figure 1. Photographie de fissident pygmée poussant sur une berge argileuse à Franquelin, au Québec, prise par le bryologue Stéphane Leclerc. Les plantes (feuilles et capsules immatures) mesurent moins de 1 cm de hauteur.
- Figure 2. Aire de répartition approximative connue du fissident pygmée en Amérique du Nord, selon toutes les sources disponibles pour le Canada et d'après des mentions d'herbier pour les États Unis. Il existe probablement un plus grand nombre de mentions d'herbier pour les États Unis que celles qui ont été utilisées dans le cadre du présent rapport. Cependant, des spécimens ont pu être trouvés pour tous les États pour lesquels des mentions de l'espèce ont été publiées, sauf l'Illinois. La mention provenant de l'Illinois est crédible (Pursell, 2007), mais aucun point correspondant n'a été inclus dans la figure ci dessus.
- Figure 3. Aire de répartition connue du fissident pygmée (Fissidens exilis) A) dans l'est du Canada et B) dans l'ouest du Canada, fondée sur 20 spécimens canadiens connus et/ou mentions publiées.
- Tableau 1. Sommaire des occurrences de fissident pygmée in Canada, avec indication des spécimens ou mentions examinés. Les nouvelles mentions signalées depuis la publication du premier rapport de situation apparaissent en caractères gras. Les mentions ont été obtenues auprès des herborisateurs dont le nom figure dans la section Experts contactés du présent rapport, et auprès d'herbiers (Herbier national du Canada, Musée canadien de la nature (CANM), Herbier Marie-Victorin, Jardin botanique de Montréal (MT), Musée du Nouveau Brunswick (NBM), University of Guelph (OAC), Herbier Louis-Marie, Université du Québec (QFA), Musée royal de l'Ontario (TRT), Beaty Biodiversity Museum, University of British Columbia (UBC), University of Western Ontario (UWO) et Devonian Botanic Garden, University of Alberta (ALTA-DBG)), de bases de données sur la conservation (Centre d'information sur le patrimoine naturel de l'Ontario, Centre de données sur la conservation du Canada Atlantique) et au moyen de recherches effectuées dans les bases de données en ligne de Canadensys, du Prairie and Northern Plant Diversity Centre, de l'herbier de l'Acadia University (ACAD) et de l'herbier du New York Botanical Garden (NY). Les spécimens marqués d'une croix (⁺) ont été examinés par la rédactrice du présent rapport. Tous les spécimens figurant dans le tableau (y compris le spécimen de référence manquant de North Dumfries, Ontario) ont été vérifiés de manière fiable par Jennifer Doubt, Steve Joya, Ron Pursell, Linda Ley, Tom Neily, Wilf Schofield et/ou Jean Faubert. Aucun spécimen n'a été trouvé pour l'occurrence de Montréal.
- Annexe 1. Calculateur des menaces pour le Fissidens exilis
COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l'on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2016. Évaluation et Rapport de situation du COSEPAC sur lefissident pygmée (Fissidens exilis) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xii + 31 p.
COSEPAC. 2005. Évaluation et Rapport de situation du COSEPAC sur le fissident pygmée (Fissidens exiilis) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. vii + 20 p.
Le COSEPAC remercie Jennifer Doubt d'avoir rédigé le rapport de situation du fissident pygmée (Fissidens exilis) au Canada, aux termes d'un contrat conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par René Belland, coprésident du Sous-comité de spécialistes des mousses et lichens du COSEPAC.
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement et Changement climatique Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Species Name Fissidens exilis in Canada.
Fissident pygmée - Photographie de fissident pygmée poussant sur une berge argileuse à Franquelin, au Québec, prise par le bryologue Stéphane Leclerc. Les plantes (feuilles et capsules immatures) mesurent moins de 1 cm de hauteur.
Le fissident pygmée (Fissidens exilis) est une mousse éphémère qui produit périodiquement des plantes minuscules (jusqu'à 2 mm de hauteur) portant 4 à 8 feuilles. Les plantes prennent naissance sur un tapis constitué de filaments verts indifférenciés, ou « protonéma », qui persiste à la surface et dans la couche superficielle du sol entre les périodes d'activité reproductrice. L'espèce peut être identifiée par l'examen de certaines caractéristiques microscopiques de ses individus feuillés (gamétophores), mais il est impossible d'identifier visuellement le protonéma. Des capsules remplies de spores, portées sur des soies mesurant 2 à 9 mm, sont fixées au sommet de chaque plante mature fécondée. La probabilité de détection du fissident pygmée est plus élevée lorsque des capsules sont présentes, surtout dans les grandes colonies de l'espèce.
Le fissident pygmée est présent en Europe, en Asie, en Afrique, dans les Antilles, en Nouvelle-Zélande et en Amérique du Nord. Certains auteurs avancent que l'espèce pourrait avoir été introduite dans les trois dernières régions, mais aucune donnée concluante ne le prouve. En Amérique du Nord, le fissident pygmée a été découvert en 1947, à Cleveland, en Ohio, et l'espèce a été signalée dans au moins quinze États de l'est des États-Unis ainsi que dans les provinces canadiennes suivantes : Nouvelle-Écosse, Québec, Ontario et Colombie-Britannique. Certains experts croient que l'espèce pourrait avoir été introduite en Colombie-Britannique.
Les activités de recherche ciblant le fissident pygmée exigent l'application de méthodes exhaustives particulières tenant compte des difficultés propres à la détection des mousses éphémères, dont l'identification visuelle est possible uniquement dans certaines conditions sporadiques. De telles méthodes n'ont pas été appliquées, et la plupart des sous-populations connues de l'espèce ont été découvertes de manière opportuniste.
En Amérique du Nord, la plupart des individus de l'espèce ont été trouvés principalement sur des sols ou des loams de type argileux dénudés, humides et au moins partiellement ombragés. L'espèce a été récoltée dans des zones boisées en bordure de cours d'eau et de ravins, dans des plaines inondables, sur des falaises, des plages, en bordure de routes, sur des sentiers et dans d'autres milieux où du sol dénudé est exposé. L'emplacement des parcelles d'habitat est transitoire et peut être imprévisible, en raison de divers phénomènes naturels ou liés aux activités humaines. Aucune tendance générale ne se dégage en ce qui concerne l'habitat de prédilection du fissident pygmée.
Le fissident pygmée est une espèce éphémère dont la stratégie de survie est « fugitive » : le cycle vital et le cycle de reproduction des plantes feuillées de l'espèce sont courts (moins de un an), ne sont pas liés aux saisons et sont largement tributaires de facteurs abiotiques. La petite taille du fissident pygmée lui permet de parvenir à maturité plus rapidement que les grandes mousses, dont les processus de développement prennent plus de temps. L'effort de reproduction est relativement élevé chez le fissident pygmée, puisque chaque plante minuscule ou presque produit une capsule remplie de spores. Ces petites spores (mesurant moins de 20 μm) sont caractéristiques des espèces dont les spores persistent longtemps dans le réservoir de spores. Ces caractéristiques permettent aux individus feuillés du fissident pygmée d'accomplir leur cycle vital dans les milieux transitoires de début de succession et d'éviter les facteurs de stress durant les périodes où l'habitat n'est pas propice en persistant sous des formes (spores et filaments souterrains) qui sont moins vulnérables aux conditions défavorables.
Les spores sont dispersées d'une hauteur de moins de 1 cm au-dessus du substrat, et la plupart des individus de l'espèce ont été trouvés dans des milieux au moins partiellement abrités. La dispersion des spores sur de grandes distances pourrait donc être très peu fréquente. La dispersion de sol comportant des spores ou des individus de l'espèce par l'intermédiaire de divers vecteurs biotiques et abiotiques pourrait être importante.
On ignore la taille et les tendances des populations de fissident pygmée; les efforts déployés pour établir ces données devront tenir compte des difficultés liées à la nature éphémère et à la taille minuscule de l'espèce.
Il est possible d'inférer certaines menaces en se fondant sur la biologie générale des mousses et sur les différents milieux où le fissident pygmée a été trouvé, mais aucune recherche n'a permis de déterminer les menaces particulières pesant sur cette espèce. Certaines activités humaines qui menacent habituellement d'autres espèces végétales pourraient avoir un effet neutre ou bénéfique sur le fissident pygmée, qui dépend de perturbations localisées du sol.
Le fissident pygmée figure actuellement sur la liste des espèces préoccupantes de la Loi sur les espèces en péril du Canada. L'espèce est également protégée aux termes de la Loi de 2007 sur les espèces en voie de disparition de l'Ontario, et au moins la moitié des sites où elle a été trouvée sont gérés par le gouvernement fédéral ou par le gouvernement de l'Ontario, ou encore par des organismes de conservation. Certaines instances en Amérique du Nord, y compris la Colombie-Britannique, ont attribué la cote SE (exotique) au fissident pygmée.
| Sujet | Information |
|---|---|
| Durée d'une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN (2011) est utilisée) Espèce éphémère dont les sporophytes subsistent peu longtemps (< 1 an), mais dont lre protonéma persistant a une longévité inconnue. |
Inconnue |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matues? | Non |
| Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] | Sans objet |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations]. | Inconnu |
| Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
Est-ce que les causes du déclin sont
|
Sans objet |
| Y a-t-il des fluctuations extrêmes du nombre d'individus matures? Les fluctuations extrêmes sont caractéristiques des mousses éphémères, qui réagissent de façon opportuniste au microclimat. |
Présumées, mais seulement à l'échelle locale |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d'occurrence | 2 030 000 km2 |
| Indice de zone d'occupation (IZO) (Fournissez toujours une valeur établie à partir d'une grille à carrés de 2 km de côté) |
84 km2 (constitue probablement une grossière sous-estimation de l'IZO réel) |
La population totale est-elle gravement fragmentée, c.-à-d. que plus de 50 % de sa zone d'occupation totale se trouvent dans des parcelles d'habitat qui sont
|
Probablement pas |
| Nombre de localités? (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.) (utilisez une fourchette plausible pour refléter l'incertitude, le cas échéant) En l'absence de menaces imminentes connues, on ne s'attend pas à ce qu'une menace en particulier touche plus d'une occurrence connue de fissident pygmée à la fois. Le nombre de localités équivaut donc au nombre d'occurrences connues. On s'attend à ce que la réalisation d'activités de recherche supplémentaires/ciblées entraîne une augmentation du nombre d'occurrences (voir les sections Répartition et Taille et tendances des populations) |
21, mais un nombre plus élevé est prévu |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de sous-populations? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.)? |
Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue et/ou la qualité] de l'habitat? | Inconnu |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités (Voir « Définitions et abréviations » sur le site Web du COSEPAC et IUCN 2010 (en anglais seulement) pour obtenir des précisions sur ce terme.)? |
Inconnu |
| Y a-t-il des fluctuations extrêmes de la zone d'occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation? | Inconnu |
| Sous-populations (indiquez une fourchette plausible) | Nombre d'individus matures |
|---|---|
| Total | Inconnu |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans]. | Non calculée |
| Sujet | Information |
|---|---|
| Les impacts séparés et combinés de toutes les menaces plausibles considérées pour cette espèce sont inconnus, soit parce qu'aucun exemple précis d'événement menaçant n'est connu, soit parce que la réponse du fissident pygmée à la menace potentielle n'est pas bien comprise. Un calculateur des menaces a-t-il été rempli pour l'espèce, et dans l'affirmative, par qui? |
Oui D. Fraser (modérateur de la discussion sur les menaces), R. Belland (coprésident du Sous-comité de spécialistes des mousses et lichens), J. Doubt (rédactrice du rapport), R. Boles (Service canadien de la faune), J. McKnight (Service canadien de la faune), E. Snyder (Ontario), S. Bureau (Service canadien de la faune), K. Golinski (Sous-comité de spécialistes des mousses et lichens), N. Fenton (Sous-comité de spécialistes des mousses et lichens) |
| Sujet | Information |
|---|---|
| Situation des populations de l'extérieur les plus susceptibles de fournir des individus immigrants au Canada Allen et al. (2004) présument que la population nord-américaine est en expansion, mais la faible détectabilité de l'espèce fait en sorte qu'il est difficile de déterminer lesquelles parmi les populations nouvellement répertoriées sont des populations plus récentes. |
Stable ou en expansion |
| Une immigration a-t-elle été constatée ou est-elle possible? | Non démontré, mais probable |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui |
| Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants? | Oui |
| Les conditions se détériorent-elles au Canada? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Peu probable |
| Les conditions de la population source se détériorent elles? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Inconnu |
| La population canadienne est-elle considérée comme un puits? Voir le tableau 3 (Lignes directrices pour la modification de l'évaluation de la situation d'après une immigration de source externe) |
Non |
| La possibilité d'une immigration depuis des populations externes existe-t-elle? | Oui |
| Sujet | Information |
|---|---|
| L'information concernant l'espèce est-elle de nature délicate? | Non |
| Sujet | Information |
|---|---|
| COSEPAC : Espèce désignée « préoccupante » en mai 2005. | Réexamen du statut et inscription à la catégorie « non en péril » en avril 2016. |
| Sujet | Information |
|---|---|
| Statut | Non en péril |
| Code alphanumérique | Sans objet |
| Justification de la désignation | Cette espèce a une zone d'occurrence canadienne très grande, présente sur les côtes du Pacifique et de l'Atlantique ainsi que dans le centre du Canada. Malgré une faible détectabilité qui complique les tentatives de quantifier la taille et les tendances de la population, le nombre d'occurrences connues a augmenté de 7 à 21 depuis 2005, et il est attendu que davantage d'occurrences seront documentées grâce aux efforts de recherche qui se poursuivent. Bien que l'espèce soit présente dans certaines régions densément peuplées du Canada, incluant le sud de l'Ontario, aucun déclin ou menaces imminentes directes ne sont connus pour cette espèce. Une perturbation localisée du sol est requise pour un habitat convenable, de sorte que certaines perturbations humaines pourraient en fait profiter à l'espèce. Bien que les données soient insuffisantes dans de multiples aspects de sa biologie, de son écologie, de sa répartition et de son abondance, rien n'indique que cette espèce est en péril au Canada. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d'individus matures) | Sans objet. Rien n'indique qu'il y ait un déclin du nombre d'individus matures. |
| Critère B (petite aire de répartition et déclin ou fluctuation) | Sans objet. La zone d'occurrence dépasse les seuils établis pour tous les critères. L'IZO est inférieur au seuil établi pour les espèces en voie de disparition, mais le nombre de localités dépasse les seuils établis pour les espèces en péril. Cette espèce ne subit pas de fluctuations extrêmes, sa population n'est pas gravement fragmentée et rien n'indique qu'il y ait un déclin de la population. |
| Critère C (nombre d'individus matures peu élevé et en déclin) | Sans objet. Le nombre d'individus matures n'est pas connu; dans le cas des occurrences connues, ce nombre ne peut être établi sans la réalisation d'une étude exhaustive à long terme. |
| Critère D (très petite population totale ou répartition restreinte) | Sans objet. Le nombre d'individus matures n'est pas connu, et ni l'IZO ni le nombre de localités connues ne correspondent aux seuils établis pour les espèces en péril. |
| Critère E (analyse quantitative) | Sans objet. Aucune analyse quantitative n'a été réalisée. |
Le fissident pygmée a été évalué pour la première fois en 2005 (COSEWIC, 2005). Depuis ce temps, les mentions préexistantes de l'espèce en Nouvelle-Écosse et au Québec ont été confirmées et de nouvelles occurrences ont été signalées en Nouvelle-Écosse (Anderson et Neily, 2010), au Québec, en Ontario et en Colombie-Britannique (SQB, 2014, mentions d'herbier), ce qui a fait passer le nombre total d'occurrences au Canada de 7 à 21 jusqu'à présent.
L'aire de répartition connue du fissident pygmée en Amérique du Nord a également augmenté (bien que dans une moindre mesure) depuis l'évaluation de 2005, et l'espèce s'est vu attribuer de nouvelles désignations dans plusieurs territoires à l'échelle nationale et infranationale.
Après avoir envisagé l'ébauche d'un sommaire du statut de l'espèce en 2013, le Sous-comité de spécialistes des mousses et lichens du COSEPAC a déterminé que les nouvelles connaissances acquises sur l'aire de répartition et l'abondance potentielle de l'espèce justifiaient la publication d'un rapport de situation mis à jour pouvant appuyer l'étude de diverses options concernant le statut de l'espèce.
Compte tenu des limites naturelles qui rendent difficile la détection du fissident pygmée, le Sous-comité a reconnu que la réalisation de travaux de terrain à une échelle réalisable dans le cadre de la préparation du rapport de situation ne permettrait pas de déterminer avec confiance la zone d'occurrence, l'indice de zone d'occupation ou le nombre de localités de l'espèce. L'information disponible est toutefois suffisante pour permettre la recommandation d'un statut pour l'espèce.
Un plan de gestion du fissident pygmée a été publié en vue de l'obtention de commentaires en juin 2015 (Environment Canada, 2015).
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d'une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d'une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d'être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d'autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l'Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d'information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu'en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d'Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Le fissident pygmée (Fissidens exilis Hedw.) est une mousse de la famille des Fissidentacées qui appartient au sous-genre Aloma Müll. Hal. (Pursell, 2007; Beever, 1999).
Le nom du genre, Fissidens, signifie « dent fendue » et fait allusion aux dents qui entourent l'orifice de la capsule renfermant les spores. Le nom anglais de l'espèce – Pocket Moss – fait allusion au dédoublement caractéristique, plus facilement visible, de la moitié supérieure de chaque feuille, qui forme une gouttière embrassant la moitié inférieure de la feuille suivante. L'épithète exilis signifie « petit » ou « mince ».
Les noms suivants sont considérés comme des synonymes de F. exilis : F. bloxamii Wilson, Bryum viridulum Dicks., Dicranum exile (Hedw.) Muhl., Schistophyllum exile (Hedw.) Lindb., Skitophyllum exile (Hedw.) Bach. Pyl. et Hypnum minutum Wilson (Steere, 1950; Allen, 2005; Missouri Botanical Garden, 2014). L'espèce a déjà été considérée comme une variété du F. bryoides, du F. viridulus et du Dicranum palmatum.
Les descriptions détaillées de l'espèce rédigées par Steere (1950) ainsi que Crum et Anderson (1981) sont résumées dans le rapport de situation du COSEPAC publié en 2005 (COSEWIC, 2005). Des descriptions plus récentes sont fournies par Allen (2005), Pursell (2007) et Faubert (2013). La figure 1 présente une photographie de l'espèce.
Le fissident pygmée produit des plantes minuscules (jusqu'à 2 mm de hauteur) portant 4 à 8 feuilles. Les plantes prennent naissance sur un tapis constitué de filaments verts indifférenciés, ou « protonéma », qui se trouve à la surface et dans la couche superficielle du sol. Les structures sexuées se forment sur les individus feuillés.
On peut identifier les individus feuillés de fissident pygmée et les distinguer d'autres espèces de fissident par l'examen de certaines caractéristiques cellulaires microscopiques, mais leur couleur vert foncé ou brun n'offre qu'un faible contraste avec le substrat dénudé où pousse l'espèce et avec son protonéma. Entre les périodes d'activité reproductrice, le protonéma, qu'il est impossible d'identifier visuellement à l'espèce, persiste dans le sol.
Des capsules remplies de spores, portées sur des soies mesurant 2 à 9 mm, sont fixées au sommet de chaque plante mature fécondée. La probabilité de détection du fissident pygmée est plus élevée lorsque des capsules sont présentes, surtout dans les grandes colonies de l'espèce. Les spores sont considérées comme petites (11–14 μm) pour des spores de bryophytes, dont la taille varie entre 5 μm et 310 μm (Crum, 2001).
Description longue de la figure 1
Photo de fissident pygmée poussant sur une berge argileuse. Cette mousse produit des plantes minuscules (jusqu'à deux millimètres) portant quatre à huit feuilles. Les plantes prennent naissance sur un tapis constitué de filaments verts indifférenciés qui se trouve à la surface et dans la couche superficielle du sol. Des capsules remplies de spores, portées sur des soies (mesurant deux à neuf millimètres), sont fixées au sommet de chaque plante mature fécondée
La population canadienne n'a pas été suffisamment étudiée pour qu'on dispose d'information fiable sur la structure et la variabilité de la population.
En raison de l'insuffisance de l'information sur la structure génétique, la répartition, la dispersion ou l'écologie de l'espèce, il n'est pas possible d'évaluer adéquatement le caractère distinct ou l'importance d'une partie quelconque de la population canadienne. Cette population est donc considérée comme une seule unité désignable.
Les mousses éphémères comme le fissident pygmée, plus particulièrement celles qui poussent dans les milieux boisés, sont peu communes au Canada. Les sous-populations canadiennes de fissident pygmée se trouvent à la limite nord de l'aire de répartition nord-américaine de l'espèce.
Le fissident pygmée est présent en Europe (îles Britanniques, centre et nord de l'Europe, Scandinavie (Steere, 1950; Pursell, 2007)), en Asie (Japon (Iwatsuki et Noguchi, 1973), Cachemire (NatureServe, 2014)) et en Afrique (Algérie (Pursell, 2007)). NatureServe (2014), citant Smith (1978), attribue également une cote à l'espèce en Amérique du Sud, mais Smith (2004) ne mentionne pas cette région. Le fissident pygmée est aussi présent en Nouvelle-Zélande (Beever, 1999) et dans les Antilles (Pursell, 2007), où l'on croit que l'espèce a été introduite. Dans plusieurs parties de son aire de répartition, l'espèce est considérée comme rare.
En Amérique du Nord, le fissident pygmée a été découvert en 1947, à Cleveland, en Ohio (Steere, 1950). Récemment, certains auteurs (Allen et al., 2004; Pursell, 2007; Faubert, 2013) ont avancé que l'espèce aurait été introduite d'Europe, vu sa découverte relativement récente dans des régions du continent qui sont densément peuplées, sujettes aux perturbations d'origine humaine et dont la flore a été assez bien étudiée. Jusqu'à présent, les cotes de conservation attribuées pour la Caroline du Nord (NatureServe, 2014) et la Colombie-Britannique (B.C. Conservation Data Centre, 2016) reflètent l'opinion selon laquelle le fissident pygmée aurait été introduit. Aucune donnée concluante ne montre toutefois que c'est le cas. Bien que les sites où le fissident pygmée a été trouvé soient situés dans des zones urbaines, l'espèce n'est pas associée aux milieux anthropiques, comme on pourrait s'y attendre pour une espèce introduite. De plus, le fissident pygmée est une espèce opportuniste qui pousse dans les sols perturbés et qui peut y prospérer, peu importe que le milieu soit anthropique ou non (voir l'analyse qui suit).
Cependant, certaines espèces de bryophytes indigènes de l'Amérique du Nord qui sont plus facilement détectables (plantes vivaces de plus grande taille) que le fissident pygmée ont été découvertes récemment, ce qui donne à penser qu'une espèce récemment découverte n'est pas nécessairement introduite. Ainsi, l'entodon de Schleicher (Entodon schleicheri) a été signalé pour la première fois en Amérique du Nord dans les années 1970 (Buck et Crum, 1978, selon du matériel récolté en 1938), mais était auparavant connu comme une espèce européenne. Dans le cadre d'une seule étude sur la Colombie-Britannique, McIntosh (1989) a plus récemment signalé pour la première fois la présence de quatre espèces nord-américaines ainsi que de deux genres et sept espèces jusque-là inconnus au Canada.
De plus, les occurrences de fissident pygmée ne sont pas concentrées autour des ports, dans les milieux urbains ou le long de perturbations linéaires anthropiques, comme c'est le cas de nombreuses espèces de bryophytes introduites (Schofield, 1988) ou qui prolifèrent rapidement (Hassel et Söderström, 1998). On trouve le fissident pygmée aussi bien dans des milieux naturels que dans des zones urbaines soumises à divers types de perturbations. Comme l'espèce a besoin de sol minéral exposé, elle est en mesure de coloniser les zones d'activité humaine, peu importe qu'elle ait été introduite sur ce continent par l'activité humaine ou non.
Aux États-Unis (figure 2), le fissident pygmée a été signalé dans au moins quinze États de l'est du pays (Alabama, Caroline du Nord, Illinois, Indiana, Kentucky, Maine, Maryland, Michigan, Missouri, New Jersey, New York, Ohio, Pennsylvanie, Tennessee, Vermont et Virginie-Occidentale (Allen et al., 2004; Pursell, 2007)). Contrairement au Canada, aucune population n'a été signalée sur la côte du Pacifique aux États-Unis.
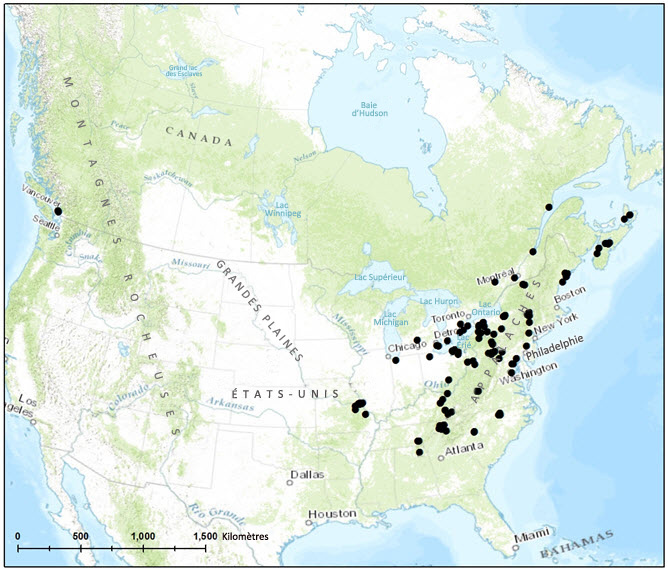
Description longue de la figure 2
Carte de l'aire de répartition du fissident pygmée en Amérique du Nord. Aux États Unis, le fissident pygmée a été signalé dans au moins quinze États de l'est du pays (Alabama, Caroline du Nord, Illinois, Indiana, Kentucky, Maine, Maryland, Michigan, Missouri, New Jersey, New York, Ohio, Pennsylvanie, Tennessee, Vermont et Virginie-Occidentale). Au Canada, l'espèce a été signalée en Nouvelle Écosse, au Québec, en Ontario et en Colombie-Britannique.
Des spécimens canadiens de fissident pygmée ont été récoltés en Nouvelle-Écosse, au Québec, en Ontario et en Colombie-Britannique (tableau 1, figure 3). L'espèce a été trouvée dans les écozones suivantes : plaines à forêts mixtes, partie sud du Bouclier boréal, maritime de l'Atlantique et maritime du Pacifique (Ecological Stratification Working Group, 1996). La Colombie-Britannique est la seule province où aucune population de l'espèce n'a encore été signalée en milieu naturel. Les deux sites connus se trouvent dans la région de Vancouver.
| Numéro | Localité | Première observation | Recherche subséquente | Spécimen ou mention | Substrat, habitat | Propriété |
|---|---|---|---|---|---|---|
| 1 | Richmond, C.-B. île Sea | 17 mars 2012 | 18 mai 2015 : espèce absente | UBC B212546 | Sur la berge argileuse d'un marécage, sous des arbustes | Municipalité de Richmond? |
| 2 | Vancouver, C.-B. pointe Grey, plage Wreck | 24 mars 2010, | 9 mars 2012 : espèce présente; 18 mai 2015 : espèce absente | UBC B211597, B218058 | Sur une motte d'argile en bordure du sentier derrière la plage. | Parc régional Pacific Spirit, District régional du Grand Vancouver |
| 3 | Dunnville, Ont. zone de protection de la nature Ruigrok Tract | 18 novembre 2012 | CANM 335576⁺ | Sur sol argileux, dans une zone de transition entre bois décidu sec et fourré en terrain marécageux humique | Office de protection de la nature de la péninsule du Niagara | |
| 4 | Port Dover, Ont. | 23 novembre 2011 | CANM 331159⁺ | Sur la masse racinaire recouverte d'argile d'un pin blanc déraciné, près d'un champ labouré | Entreprise agricole | |
| 5 | Chatham-Kent, Ont. Zone de protection de la nature Sinclair's Bush | 16 août 2002 | ALTA-DBG B-14643⁺ | Sur boue dénudée, dans une forêt d'érable et de hêtre | Office de protection de la nature de la vallée de la Thames inférieure | |
| 6 | Canton de North Dumfries, Ont. Sudden Tract | 23 septembre 1995 | Spécimen manquant, probablement à NB ou ALTA-DBG (Bradley, comm. pers., 2015) | Dans une forêt décidue fraîche et humide dominée par l'érable à sucre | Ministère des Richesses naturelles de l'Ontario (forêt régionale) | |
| 7 | Canton de Walsingham, Ont. zone de protection de la nature du ruisseau Deer | 22 juin 1995 | ALTA-DBG B-6969 | Dans une forêt mixte de feuillus et de pruche, sèche à fraîche | Office de protection de la nature de la région de Long Point | |
| 8 | Canton d'Anderdon, Ont. « Canard River Kentucky Coffee Tree Woods » | 24 mars 1984 | Août 2002 : espèce non trouvée | CANM 290756⁺ | Sur des mottes d'argile, dans un bois en plaine inondable | Office de protection de la nature de la région d'Essex |
| 9 | Canton de Colchester Sud, Ont. | 26 mars 1981 | Août 2002 : espèce non trouvée | CANM 275055⁺, UBC B24863, NY 113173 | Dans un bois décidu mature dominé par le chêne | Ministère des Richesses naturelles de l'Ontario |
| 10 | Sainte-Foy, Qc | 3 juin 1987 | Herbier privé de Jean Faubert, spécimen no 7965 | Sur sol dénudé, dans une forêt décidue | Campus de l'Université Laval | |
| 11 | Gatineau, Qc | Juin 1982 | CANM 291533⁺ | Non consigné | Parc de la Gatineau (Commission de la capitale nationale) | |
| 12 | Montréal, Qc | Automne 1973 | Juillet 2002 : espèce non trouvée | Molnar, 1975 (mention publiée seulement) | Sur sol argileux, dans une plantation d'épinettes et de mélèzes | Arboretum Morgan, Université McGill |
| 13 | Franquelin, Qc | 12 octobre 2014 | 17 Juillet 2015 : espèce présente, mais moins abondante | SQB (2014); spécimen STL-0053 vérifié par Jean Faubert | Sur sol argileux en bordure d'un ruisseau s'écoulant dans une forêt boréale, avec roches granitiques sèches | Province de Québec; propriété gérée par la municipalité de Franquelin |
| 14 | Ruisseau Teare, N.-É. | 26 avril 2012 | CANM 331676⁺ | Sur sol exposé, sur la pente descendant vers le ruisseau | Propriété privée | |
| 15 | Cap White, N.-É. | 7 et 8 mai 2010 | CANM 331674⁺, NB BB-21566, ACAD ECS03857 | Sur du sol, derrière une falaise de gypse | Propriété privée | |
| 16 | Rivière Herbert, N.-É. | 3 avril 2012 | CANM 331675⁺ | Sur du sol, sur la berge de la rivière | Propriété privée | |
| 17 | Ruisseau Glen, N.-É. | 23 avril 2012 | ACAD ECS039057 | Sur du sol, sur le versant d'un ravin | Inconnu | |
| 18 | Glendyer, N.-É. | 10 octobre 2013 | ACAD ESC039909 | Sur un sol humide et ombragé, dans une doline | Propriété privée | |
| 19a | Big Harbour, N.-É. | 10 octobre 2013 | ACAD ECS039908 | Sur sol argileux, sous des feuillus | Couronne | |
| 19b | Big Harbour, N.-É. | 10 octobre 2013 | ACAD ECS039907 | Sur sol argileux, sous des feuillus | Couronne | |
| 20 | Ruisseau Bishop, N.-É. | 15 janvier 2014 | ACAD ECS039911, CANM 335560⁺ | Sur des rochers, dans une pente sous des aulnes | Propriété privée | |
| 21 | Belle-Isle, N.-É. | 16 juillet 1987 | UBC B114963 | Dans une forêt d'épinettes et de sapins de seconde venue, sur sol argileux et humide | Inconnu |
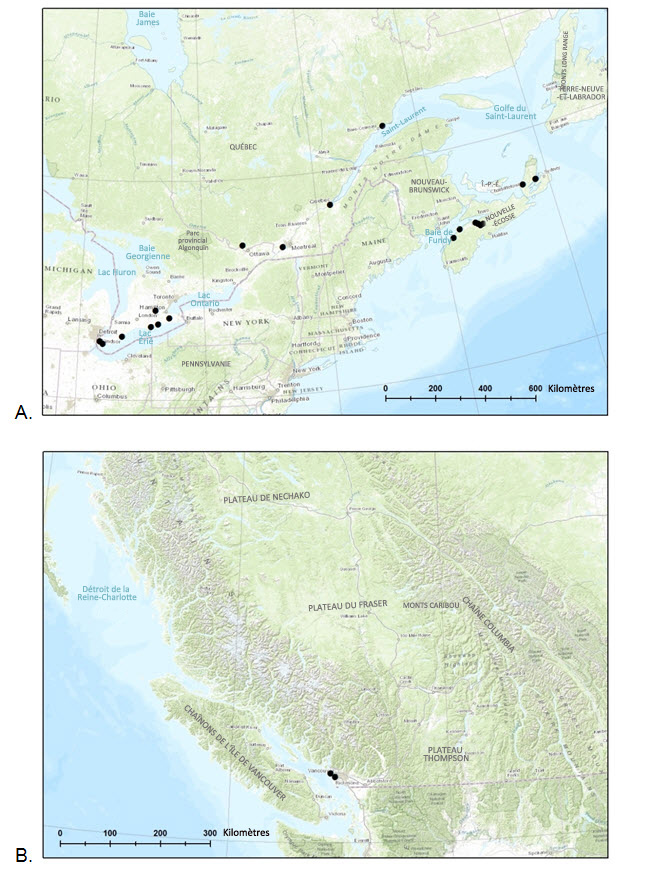
Description longue de la figure 3
Deux cartes illustrant l'aire de répartition connue du fissident pygmée dans l'est du Canada (carte A) et dans l'ouest du Canada (carte B).
Compte tenu des occurrences nouvellement répertoriées qui n'étaient pas connues au moment de la préparation du rapport de situation précédent (COSEWIC, 2005), la zone d'occurrence connue du fissident pygmée a augmenté, passant de 30 000 km2 à 2,03 millions de km2. L'IZO, calculé selon une grille à carrés de 2 km de côté, s'établit à 84 km2. Cette valeur constitue probablement une sous-estimation de la superficie occupée par l'espèce, étant donné que celle-ci est moins susceptible d'être détectée que la plupart des autres mousses et que de nombreux secteurs d'habitat potentiellement propice à l'espèce n'ont pas encore été explorés.
Des recherches ciblées ont été effectuées en 2002 pour la préparation du premier rapport de situation sur le fissident pygmée. Les recherches ont porté sur un site en Ontario et deux sites au Québec qui étaient décrits sur les étiquettes des spécimens d'herbier et dans les mentions publiées, et ont également ciblé des sites explorés à la recherche de deux autres espèces de mousses (le Bryoandersonia illecebra, dans le sud de l'Ontario, et l'Helodium paludosum, dans le sud de l'Ontario et du Québec) pour lesquelles des rapports de situation étaient préparés au même moment. De minuscules spécimens de mousses du genre Fissidens ont été récoltés dans 11 des 36 sites visités, et un total de 46 échantillons (COSEWIC, 2005) ont été prélevés aux fins de l'examen au microscope. Le fissident pygmée a été trouvé une seule fois, dans un site qui n'avait jamais été répertorié (tableau 1). Par ailleurs, les 19 autres spécimens canadiens connus de l'espèce ont été répertoriés par des botanistes qui réalisaient des études générales sur la biodiversité ou des projets ciblant d'autres espèces que le fissident pygmée.
Aucune activité de recherche ciblée n'est associée au présent rapport de situation. La réalisation de ce type de recherche soulève de nombreuses difficultés dans le cas du fissident pygmée :
- La petite taille du fissident pygmée fait en sorte que l'espèce est moins visible que de nombreuses autres espèces de bryophytes. La probabilité de détection du fissident pygmée est plus élevée lorsque ses capsules remplies de spores, qui subsistent peu de temps, sont pleinement formées; l'espèce risque de passer inaperçue aux autres stades de son développement (p. ex. Allen et al., 2004; Allen, 2005; Faubert, 2013).
- En l'absence d'individus feuillés et de capsules, le fissident pygmée persiste sous forme de protonéma (et probablement aussi sous forme de spores) et peut donc souvent passer inaperçu pendant des périodes de durée inconnue, pouvant être longues. En présence d'habitat apparemment propice, on ne peut, en une seule visite, déterminer si le fissident pygmée est présent ou non, ou établir une tendance apparente de l'abondance de l'espèce.
- Contrairement aux périodes de détectabilité qui sont raisonnablement bien documentées chez certaines plantes vasculaires, le moment le plus propice pour détecter le fissident pygmée n'est pas clairement établi et varie probablement (comme chez d'autres espèces éphémères) sur le plan géographique et temporel en fonction du climat local, du microclimat et des perturbations, sans qu'on puisse établir de lien fiable par rapport au cycle annuel des saisons. Les capsules parviennent à maturité en hiver (p. ex. Steere, 1950; Smith, 2004; Atherton et al., 2010), lorsque l'humidité est élevée, ce qui rend plus probable la détection de l'espèce au printemps et à l'automne. Les spécimens canadiens ont été récoltés durant tous les mois de l'année sauf février et décembre, un quart des spécimens ayant été prélevés entre les mois de mai et août (tableau 1).
- Dans les régions boisées où se rencontre le fissident pygmée, les conditions humides prolongées peuvent être plus courantes au printemps et à l'automne, mais peuvent survenir à n'importe quel moment de l'année; de telles conditions peuvent être plus fréquentes dans la partie nord de l'aire de répartition de l'espèce que dans la partie sud. De même, les perturbations localisées (p. ex. chute d'un arbre, crue d'un cours d'eau) qui exposent des parcelles de sol propices à la croissance du fissident pygmée surviennent de manière imprévisible au cours d'une année et d'une année à l'autre.
Selon Allen et al. (2004), le fissident pygmée serait plus commun que ne le laissent prévoir les mentions d'occurrence, vu sa petite taille et la courte période au cours de laquelle il peut être récolté. Ce fait est souvent évoqué pour d'autres mousses éphémères (Allen, 1979; Risk, 2002; Glime, 2007). Vanderpoorten et Engels (2002) ont constaté que les bryophytes éphémères ne pouvaient faire l'objet d'une modélisation prédictive de la répartition fondée sur des variables environnementales. Une observation attentive de l'habitat de prédilection de l'espèce ainsi que des visites ciblées effectuées à des moments soigneusement planifiés ont néanmoins permis à Risk (2002) de répertorier 50 nouvelles localités américaines abritant l'espèce apparentée Fissidens hyalinus, ce qui a porté le nombre de sites connus de 15 à 65. Des recherches ciblées semblables pourraient s'avérer profitables pour le fissident pygmée.
Comme Beever (1999) l'indique, les mousses du genre Fissidens, à l'instar de nombreuses mousses, ont besoin de conditions microenvironnementales précises. En Amérique du Nord, le fissident pygmée a été trouvé sur des sols ou des loams de type argileux dénudés, humides et au moins partiellement ombragés (p. ex. Allen, 2005; Faubert, 2013, mentions d'herbier). L'espèce a été récoltée dans des zones boisées en bordure de cours d'eau et de ravins, dans des plaines inondables, sur des falaises, en bordure de routes, sur des sentiers et dans d'autres milieux où du sol dénudé est exposé (Crum et Anderson, 1981; Pursell, 2007; Anderson et Neily, 2010, mentions d'herbier, tableau 1). L'espèce peut être associée à d'autres mousses éphémères telles que celles des genres Ephemerum et Micromitrium (Crum et Anderson, 1981), dont on connaît les préférences pour les petites superficies de milieux temporaires. Tom Neily (comm. pers., 2015) a noté que les sites abritant le fissident pygmée en Nouvelle-Écosse se trouvaient dans des zones karstiques et basaltiques.
Comme le fissident pygmée a besoin de sol exposé pour pousser, les tendances générales en matière d'habitat pourraient être moins importantes que les tendances à l'échelle locale. En règle générale, les perturbations naturelles résultant de la chute d'arbres ou d'inondations sont cependant moins courantes dans les régions très peuplées et aménagées, y compris dans les villes canadiennes où le fissident pygmée a été répertorié. Parallèlement, les perturbations d'origine humaine peuvent entraîner l'exposition de sol, que ce soit lors de l'aménagement et de l'utilisation de corridors terrestres de transport et de service ou lors d'activités liées à l'agriculture et à la foresterie. Les sites où le fissident pygmée a été récolté sont soumis à des perturbations naturelles et/ou humaines (tableau 1).
Très peu de publications traitent de la biologie du fissident pygmée. Le rapport de situation du COSEPAC de 2005 (COSEWIC, 2005) énonce quelques-unes des caractéristiques propres à toutes les mousses acrocarpes autoïques poussant sur le sol.
La plupart des auteurs (p. ex. Allen et al., 2004; Allen, 2005; Pursell, 2007) décrivent le fissident pygmée comme une espèce éphémère, ce qui signifie que le cycle vital et le cycle de reproduction des plantes feuillées sont courts (moins de un an), ne sont pas liés aux saisons et sont largement tributaires de facteurs abiotiques (During, 1979). L'espèce pourrait donc avoir certaines caractéristiques biologiques en commun avec des mousses appartenant à d'autres familles – Ephemeraceae, Micromitraceae, Funariaceae, Buxbaumiaceae et Polytrichaceae – dont les plantes sont aussi éphémères (avec ou sans protonéma persistant). La petite taille de ces mousses leur permet de parvenir à maturité plus rapidement que les grandes mousses, dont les processus de développement prennent plus de temps (p. ex. Goffinet et al., 2011).
Selon la classification des stratégies de cycle vital des bryophytes établie par During (During, 1979), le fissident pygmée est une espèce « fugitive » : elle ne vit pas longtemps (cycles de moins de un an) et se reproduit par voie sexuée très peu de temps après l'émergence des plantes feuillées. L'effort de reproduction est relativement élevé chez le fissident pygmée, puisque chaque plante minuscule ou presque produit une capsule remplie de spores. Les spores sont petites (toute spore de moins de 20 μm est considérée comme petite dans la classification de During). Ces caractéristiques permettent aux individus feuillés du fissident pygmée d'accomplir leur cycle vital dans les milieux transitoires de début de succession et d'éviter les facteurs de stress durant les périodes où l'habitat n'est pas propice en persistant sous des formes – spores et protonéma – qui sont moins vulnérables.
Étant donné que chaque individu feuillé produit des gamétanges mâles et femelles, il est probable que le fissident pygmée s'autoféconde. La fécondation entraîne la formation d'une soie terminée par une capsule remplie de spores. Presque toutes les plantes produisent une capsule.
Chez les espèces qui vivent peu longtemps, comme les mousses éphémères, dont le cycle vital est déclenché par des conditions transitoires survenant dans le microclimat, il est possible que plus d'une génération soit produite par année (p. ex. Gray, 1935 in Glime, 2007; Furness et Hall, 1981). Lorsque les conditions sont optimales, chaque stade de développement peut être très bref. Allen et al. (2004), par exemple, ont observé une réduction marquée de l'abondance le long d'un sentier dans un parc urbain entre le 20 et le 29 mars 2003.
Durant les périodes qui ne sont pas propices à l'activité reproductrice, le fissident pygmée persiste dans le sol sous forme de protonéma indifférencié (tissu gamétophytique juvénile dont l'aspect est semblable à celui d'algues filamenteuses) qui ne peut être identifié à l'espèce, et sous forme de spores, qui tombent près des plantes mères (en raison de la petite taille de celles-ci). Bien qu'aucune expérience n'ait été menée sur le fissident pygmée, on croit que les espèces éphémères dont les spores sont petites (< 20 μm) persistent beaucoup plus longtemps dans le sol que les espèces dont les spores sont grosses (> 25 μm). Ces dernières sont davantage caractéristiques des milieux où les perturbations surviennent de manière prévisible (During, 1979).
Furness et Hall (1981) ont étudié l'abondance du Physcomitrium sphaericum, une mousse éphémère produisant de grosses spores (et dont le protonéma n'est pas persistant), sur une période de plus de quarante ans dans deux sites de la Grande-Bretagne. Ils ont constaté que les spores présentes dans les sédiments d'étangs persistaient pendant plus d'une décennie et pouvaient donner naissance à des populations prospères dès que le niveau d'eau des lacs baissait suffisamment pour que le substrat boueux privilégié par l'espèce soit exposé. Aucune étude n'a encore été réalisée sur la longévité des spores du fissident pygmée.
Le tapis de protonéma joue probablement un rôle très important dans la multiplication végétative des colonies de fissident pygmée à l'échelle locale. Les besoins et la longévité du protonéma sont inconnus. Enfoui dans le substrat, le protonéma est probablement davantage à l'abri des pertes d'eau et des perturbations mineures de la surface du sol que les plantes feuillées. Dans les sols humides, le protonéma de certaines espèces peut survivre pendant 50 ans (Bristol, 1916, in Schofield, 1981).
La durée d'une génération est inconnue chez le fissident pygmée. Cependant, les espèces éphémères, comme le fissident pygmée, produisent habituellement des sporophytes qui subsistent peu longtemps (< 1 an), mais possèdent un protonéma persistant dont la longévité est inconnue.
Le fissident pygmée a besoin de sol minéral exposé, ce qui le rend vulnérable à la succession végétale dans son habitat. La stratégie de cycle vital fugitive de l'espèce concernant ses gamétophores lui permet d'éviter ce facteur de stress plutôt que d'avoir à le tolérer. Dans les endroits où les conditions d'humidité et les perturbations surviennent de façon périodique (p. ex. berges de cours d'eau, plaines inondables), le sol peut demeurer dénudé, ou de nouvelles parcelles de sol dénudé peuvent apparaître à mesure que d'autres sont recouvertes par la végétation, ce qui permet au protonéma persistant dans le sol de tolérer les périodes où l'habitat n'est pas propice. Lorsque les perturbations ne se répètent pas, les individus de l'espèce ayant colonisé le sol dénudé peuvent être éliminés avec le temps.
La production de spores est très importante pour les espèces fugitives, qui dépendent de substrats dispersés, temporaires et dont la récurrence est imprévisible (p. ex. During, 1979). Cependant, il est peu probable que les spores du fissident pygmée, qui sont libérées à moins de 1 cm au-dessus du sol dans des milieux abrités, puissent être dispersées sur de grandes distances. Il est également possible que les perturbations répétées du sol fragmentent les plantes (presque n'importe quelle cellule de mousse peut générer un clone) ou le protonéma enfoui. La dispersion de spores ou de fragments dans le sol par l'eau ou par des animaux ou des machines est probable. Bien qu'aucun vecteur animal particulier n'ait été répertorié, de nombreuses espèces animales pourraient sans doute prélever des spores ou des fragments présents dans le sol, plus particulièrement sur les berges de ruisseaux. Le fissident pygmée pourrait aussi être dispersé à l'occasion de projets de reboisement ou d'aménagement des terres, pour lesquels la machinerie utilisée afin de préparer le sol est transportée d'un site à l'autre.
Les mousses éphémères évitent la compétition en colonisant de nouveaux substrats et en accomplissant leur cycle vital très rapidement (Slack, 1990). L'établissement d'autres mousses et plantes vasculaires rend ces parcelles d'habitat non propices à ces espèces, jusqu'à ce qu'une nouvelle perturbation vienne « réamorcer » le processus de succession.
Le fissident pygmée peut être associé à n'importe quelle perturbation localisée du sol pouvant résulter, par exemple, d'une activité animale directe (creusage, piétinement par des dindons ou des cerfs) ou indirecte (inondation causée par des castors). De même, la perturbation de substrat naturel par des humains (p. ex. utilisation de sentiers) pourrait créer ou maintenir des parcelles de sol minéral dénudées propices à l'établissement du fissident pygmée. La prolifération de vers de terre non indigènes dans de nombreuses régions du Canada contribue aussi probablement à la disponibilité accrue de substrat de sol minéral (p. ex. Sackett et al., 2013). Toutefois, comme le sol peut également être perturbé de façon naturelle par d'autres facteurs (p. ex. inondations, érosion, arbres renversés par le vent), le fissident pygmée ne dépend vraisemblablement pas d'autres espèces.
Hallingbäck et Hodgetts (2000) recommandent que les localités abritant des bryophytes éphémères soient explorées de façon répétée, à des moments propices de l'année et durant plusieurs années, pour tenir compte des fluctuations de population et de la persistance invisible des espèces dans le réservoir de diaspores (dans le cas du fissident pygmée, le protonéma persistant fait également partie du réservoir de propagules de l'espèce). On ne dispose pas de telles données de relevé pour le fissident pygmée. Certains des facteurs pouvant compliquer les tentatives de quantification de la taille et des tendances des populations ont déjà été énoncés dans le présent rapport (voir la section Activités de recherche). La plupart des occurrences de fissident pygmée répertoriées ont été découvertes de façon fortuite par des botanistes travaillant à d'autres projets. En outre, l'impossibilité d'identifier l'espèce sans microscope entraîne un délai entre la récolte des spécimens et la confirmation de leur identification. Pour ces raisons, l'abondance de l'espèce n'a pas été évaluée, même au moment où chaque sous-population a été observée pour la première fois. Les visites subséquentes des sites – même celles qui sont réalisées relativement peu de temps après la première observation – sont susceptibles de donner des résultats différents (p. ex. Allen et al., 2004) et de favoriser une compréhension dynamique de l'abondance et des conditions optimales de reproduction de l'espèce.
La plupart des auteurs considèrent le fissident pygmée comme une espèce rare ou dont la collecte est sous-optimale (p. ex. Crum et Anderson, 1981; Allen et al., 2004; Allen, 2005; NatureServe, 2014; Faubert, 2013). La découverte récente de populations qui n'avaient jamais été répertoriées porte à croire que la réalisation d'activités de recherche supplémentaires importantes et soigneusement planifiées dans les zones se trouvant entre les sites (Environment Canada, 2015) est nécessaire pour estimer adéquatement la zone d'occurrence, l'IZO ou le nombre d'individus matures de l'espèce (voir aussi Activités et méthodes d'échantillonnage).
Les fluctuations de population à l'échelle locale sont caractéristiques des espèces éphémères (p. ex. Furness et Hall, 1981; Allen et al., 2004). Allen et al. (2004) présument que la population de fissident pygmée de l'est de l'Amérique du Nord est en expansion, compte tenu de l'augmentation de l'aire de répartition et du nombre de spécimens récoltés, mais aucune étude n'a été menée à ce sujet.
La population de fissident pygmée du nord des États-Unis, qui constitue la source potentielle d'individus de l'espèce la plus proche à l'extérieur du Canada, pourrait probablement constituer une source d'immigration pour la population canadienne, si cette dernière devait disparaître. Malgré la très petite taille des individus feuillés et des spores du fissident pygmée, la propagation de l'espèce (voir la section Dispersion) aux États-Unis et dans les régions limitrophes du Canada semble tout aussi probable, surtout du fait que les aires de répartition canadienne et américaine se chevauchent sur le plan latitudinal (figure 2). La dispersion naturelle pourrait être ralentie par les Grands Lacs, dans le centre du Canada. Toutefois, comme les mécanismes de dispersion de l'espèce sont mal compris, l'importance de cette barrière possible demeure inconnue. Les individus de l'espèce aux États-Unis seraient probablement adaptés pour survivre au Canada.
La dispersion du fissident pygmée entre les États-Unis et le Canada, que ce soit par ses spores ou ses fragments végétatifs transportés par l'eau ou par des vecteurs animaux, est probable. Aujourd'hui plus que par le passé, les restrictions imposées sur le déplacement de sol de part et d'autre de la frontière canadienne limitent probablement le transport de l'espèce par des moyens artificiels.
Comme tous les végétaux, le fissident pygmée présente une plage de tolérance caractéristique pour ce qui est de l'humidité, des éléments nutritifs et de l'éclairement. Mis à part les milieux et les substrats auxquels l'espèce a été associée (voir la section Habitat), les conditions particulières exigées par le fissident pygmée n'ont cependant pas été étudiées. Par conséquent, on s'attend à ce que le fissident pygmée soit menacé par les activités qui modifient l'humidité, les éléments nutritifs, l'éclairement ou le régime de perturbation de son habitat à long terme. On présume que l'enlèvement de sol à grande échelle ou la destruction d'habitat due au développement, par exemple, aura une incidence négative sur les populations locales. De même, l'interruption du régime de perturbation entraînant l'exposition de nouveau sol minéral non colonisé rendrait l'habitat moins propice pour l'espèce.
Le système de classification des menaces de l'IUCN-CMP (Union internationale pour la conservation de la nature-Partenariat pour les mesures de conservation) (IUCN-CMP, 2006) permet de considérer de manière systématique les impacts d'une grande variété de menaces qui pèsent sur une espèce donnée. Les rubriques qui suivent correspondent aux catégories de menaces de l'UICN pouvant s'appliquer au fissident pygmée (annexe 1). Aucune de ces menaces ne présente cependant un caractère imminent certain. Au Canada, on trouve le fissident pygmée aussi bien dans des zones naturelles que dans des zones aménagées urbaines ou récréatives. Rien n'indique qu'il y ait un déclin de la population, et Allen et al. (2004) présument qu'il y a une augmentation continue de la population en Amérique du Nord. Certaines activités humaines, telles que l'aménagement de sentiers et de routes, de même que certains types d'activités forestières, minières et agricoles qui perturbent le sol tout en le laissant disponible pour la colonisation par les végétaux, pourraient favoriser la dispersion et la propagation du fissident pygmée. L'information disponible n'a pas permis de calculer l'impact global des menaces qui pèsent sur cette espèce, mais cet impact semble minime (tableau 1).
Au moins sept occurrences canadiennes de fissident pygmée se trouvent dans des parcs accessibles au public et/ou des milieux naturels aménagés. L'aménagement et l'utilisation de sentiers sur le sol indigène favorisent l'espèce en maintenant le sol dénudé. L'espèce a cependant été trouvée aussi bien sur des sentiers que loin de ceux-ci. Par ailleurs, l'application de pesticides chimiques, l'élimination de substrat approprié due à la construction de bâtiments et d'autres infrastructures et le piétinement excessif auraient une incidence négative sur l'espèce.
Au moins trois spécimens canadiens de fissident pygmée ont été récoltés sur des berges de cours d'eau ou dans des plaines inondables. La régulation de l'eau dans ce type d'habitat peut réduire la disponibilité de substrat de sol perturbé à des périodes propices (plus humides) de l'année. Aucun nouveau projet de gestion de l'eau n'a été proposé pour les sites où le fissident pygmée a été observé.
Une occurrence canadienne de fissident pygmée se trouve près d'un champ labouré (tableau 1). Aucune occurrence n'a encore été observée dans des champs agricoles, peut-être à cause de l'éclairement plus élevé et/ou du taux d'humidité plus faible qu'on y trouve comparativement aux milieux où l'espèce est habituellement observée, mais les milieux agricoles sont souvent peu explorés. Certaines bryophytes éphémères sont étroitement associées à l'activité agricole (p. ex. Porley, 2008). Selon des études européennes, la culture des champs à l'automne peut perturber le développement des bryophytes éphémères, tandis que les champs recouverts de chaumes pour l'hiver peuvent favoriser le déclenchement du cycle de reproduction des plantes lorsque l'humidité augmente à l'automne, et la conclusion de ce cycle avant la réactivation du champ au printemps (Porley, 2008; Hallingbäck et Hodgetts, 2000). Comme les bryophytes sont généralement très sensibles aux changements chimiques qui se produisent dans leur environnement, on ne s'attend pas à ce que l'utilisation d'engrais ou d'herbicides favorise le fissident pygmée.
Une occurrence canadienne de fissident pygmée se trouve dans une forêt régionale (tableau 1). Il est possible que la machinerie utilisée pour les activités forestières (récolte, reboisement) puisse favoriser la dispersion du fissident pygmée entre deux sites situés à proximité. La mesure dans laquelle les activités forestières aident le fissident pygmée ou lui nuisent en créant un nouvel habitat de sol perturbé dépend probablement de la mesure dans laquelle les conditions globales du site – humidité, éléments nutritifs et éclairement – sont modifiées par la même occasion.
En général, la construction et l'entretien de corridors terrestres de transport (voies ferrées, routes) qui entraînent la perturbation du sol en périphérie sont susceptibles de favoriser le fissident pygmée lorsque ces corridors traversent un milieu propice, à la condition que l'apport de produits chimiques, tels que le sel de voirie et les herbicides, soit minime.
L'enlèvement ou le recouvrement de substrat ou l'instauration de mesures de gestion intensive (fauchage, application d'engrais ou de pesticides/herbicides) pourraient menacer le fissident pygmée, mais de tels travaux ne sont pas prévus dans les sites canadiens où l'espèce a été répertoriée. De plus, certains sites de la Colombie-Britannique et du Québec où l'espèce a été récoltée ainsi que de nombreux sites américains se trouvent en milieu urbain, ce qui indique un certain degré de tolérance ou d'affinité de l'espèce pour l'activité humaine.
On s'attend à ce que les activités qui nécessitent l'enlèvement de sol à grande échelle et/ou des concentrations de substances chimiques non naturelles aient des effets négatifs sur le fissident pygmée. Aucun projet d'exploitation minière ou de production d'énergie n'est prévu dans les sites où l'espèce a été observée.
Dans le rapport de situation de 2005 (COSEWIC, 2005), le COSEPAC a indiqué que le climat comptait parmi les facteurs limitatifs potentiels de l'espèce, mais la plage de tolérance de celle-ci aux conditions climatiques semble large, compte tenu de son aire de répartition mondiale. L'impact du réchauffement climatique sur le fissident pygmée, qui pourrait modifier certains paramètres de l'habitat tels que l'humidité, la température, les perturbations naturelles et l'activité de vecteurs de dispersion possibles, est inconnu.
Il est peu probable qu'un même phénomène constituant une menace plausible puisse affecter un site où le fissident pygmée a été observé et l'occurrence la plus proche de ce site. Vingt-et-une occurrences de fissident pygmée ont été répertoriées depuis 1973 (tableau 1); le nombre de localités répertoriées est donc le même. Cinq de ces occurrences ont été visitées de nouveau depuis leur découverte; l'espèce était encore présente dans seulement deux d'entre elles (tableau 1). Toutefois, compte tenu de la présence continue de substrat apparemment approprié, de la visibilité intermittente de l'espèce et de l'absence de données très précises sur l'emplacement des localités, on suppose que tous les sites abritent encore le fissident pygmée.
Le fissident pygmée figure actuellement sur la liste des espèces préoccupantes de l'annexe 1 de la Loi sur les espèces en péril du Canada et sur la liste officielle des espèces en péril en Ontario. Un plan de gestion visant à caractériser avec une plus grande certitude la répartition et l'abondance de cette espèce grâce à l'élaboration et à la mise en œuvre de protocoles d'inventaire et de suivi précis a été ébauché (Environment Canada, 2015).
Le fissident pygmée est coté G3/G4 (vulnérable) à l'échelle mondiale et N1 au Canada (NatureServe, 2014). À l'échelle infranationale, l'espèce est cotée S1 (gravement en péril) au Tennessee et au Vermont (NatureServe, 2014), S1? en Nouvelle-Écosse (ACCDC, 2014), S1S2 au Québec (CDPNQ, 2015) et S2 (en péril) en Ontario (NHIC, 2014) et dans l'État de New York (NatureServe, 2014). En Colombie-Britannique (Penny, comm. pers., 2015) et en Caroline du Nord (NatureServe, 2014), le fissident pygmée est considéré comme une espèce introduite. Ailleurs, l'espèce est jugé inclassable (Missouri) ou n'est simplement pas classée (NatureServe, 2014).
Dans la liste des mousses du programme sur la situation générale des espèces, qui devait être mise à jour en 2015, le fissident pygmée est coté S2 en Ontario et S1? au Québec (Belland, comm. pers., 2015). Les cotes de situation générale n'étaient pas disponibles pour la Colombie-Britannique et la Nouvelle-Écosse au moment de la rédaction du présent rapport.
Les données disponibles sur la propriété des terres où se trouvent les occurrences canadiennes de fissident pygmée sont résumées au tableau 1. Les trois quarts des sites environ sont accessibles au public, et la moitié sont gérés par des organismes de conservation ou par des gouvernements qui administrent les protections juridiques applicables en vertu des lois sur les espèces en péril du gouvernement fédéral et de l'Ontario.
Nous exprimons notre reconnaissance à tous les experts contactés pour la compilation des données du présent rapport ainsi qu'aux membres travaillants et enthousiasmes du Sous-comité de spécialistes des mousses et lichens du COSEPAC, qui ont transmis des données, de sages conseils et des avis d'experts au sujet du fissident pygmée au Canada. Alain Filion, Julie Perrault et Shirley Sheppard, du Secrétariat du COSEPAC, ont fourni des cartes, des encouragements et un soutien administratif exemplaire.
Allan Aubin, bryologue, Simcoe (Ontario).
Annie St-Louis, technicienne en travaux d'enseignement et de recherche, Herbier Louis-Marie, Université Laval, Québec (Québec).
Bruce Bagnell, bryologue (Nouveau-Brunswick).
Carole Ann Lacroix, conservatrice adjointe, Herbarium, University of Guelph (OAC), Guelph (Ontario).
Deb Metsger, conservatrice adjointe, Herbarium, Musée royal de l'Ontario (TRT), Toronto (Ontario).
David Bradley, botaniste et bryologue, Centre d'information sur le patrimoine naturel de l'Ontario, ministère des Richesses naturelles et des Forêts de l'Ontario, Peterborough (Ontario).
Emmanuelle Fay, coordonnatrice – Rétablissement des espèces en péril, Service canadien de la faune, Direction générale de l'intendance environnementale, Environnement Canada, Québec (Québec).
Geoffrey Hall, coordonnateur de la collection, Herbier Marie-Victorin (MT), Montréal (Québec).
Jean Faubert, bryologue et président, Société québécoise de bryologie, Rimouski (Québec).
Jenifer Penny, Program Botanist, British Columbia Conservation Data Centre, Victoria (Colombie-Britannique).
Linda Ley, bryologue, Ottawa (Ontario).
Michael Oldham, botaniste et herpétologiste, Centre d'information sur le patrimoine naturel de l'Ontario, ministère des Richesses naturelles et des Forêts de l'Ontario, Peterborough (Ontario)
Olivia Lee, Bryophyte, Fungi and Lichen Collections Manager, University of British Columbia Herbarium, Beaty Biodiversity Museum (UBC), Vancouver (Colombie-Britannique).
René Belland, Bryologist and Curator, Plant Herbarium, Devonian Botanic Garden (ALTA-DBG); Faculty Service Officer, Department of Renewable Resources, University of Alberta, Edmonton (Alberta).
Ruth Newell, Curator, Acadia University Herbarium, Wolfville (Nouvelle-Écosse).
Sandra Mackin, technicienne d'herbier, University of Western Ontario Herbarium (UWO), London (Ontario).
Scott Schuette, Treasurer and Copyright Licences, American Bryological and Lichenological Association
Sean Blaney, botaniste et directeur adjoint, Centre de données sur la conservation du Canada Atlantique, Sackville (Nouveau-Brunswick).
Stéphane Leclerc, bryologue, Sainte-Anne-de-Beaupré (Québec).
Steve Joya, Bryologist and Herbarium Technician, Beaty Biodiversity Museum, University of British Columbia (UBC), Vancouver (Colombie-Britannique).
Stephen Clayden, conservateur, Herbier du Musée du Nouveau-Brunswick, Saint John (Nouveau-Brunswick).
Terry McIntosh, Bryologist and Consultant, Vancouver (Colombie-Britannique).
Tom Neily, Bryologist/lichenologist and Consultant, Middleton (Nouvelle-Écosse).
ACCDC (Atlantic Canada Conservation Data Centre). 2014. Rarity ranks for non-vascular plants. (Disponible en anglais seulement, consulté le 29 décembre 2014)
Allen, B.H. 1979. Additional distributional records of mosses in Pennsylvania. The Bryologist 82:289–291.
Allen, B.H., R.A. Pursell et C. Darigo. 2004. Fissidens exilis and a key to the species of Fissidens in Missouri. Evansia 21:111–115
Allen, B.H. 2005. Maine Mosses: Sphagnaceae – Timmiaceae. Memoirs of the New York Botanical Garden 93. The New York Botanical Garden Press, New York. 419 pp.
Atherton, D.M., S.D.S. Bosanquet et M. Llawley. 2010. Mosses and Liverworts of Britain and Ireland: A Field Guide. British Bryological Society, England. 856 pp.
ACCDC (Atlantic Canada Conservation Data Centre). 2014. List of species ranks. (Disponible en anglais seulement, consulté le 31 décembre 2014).
Anderson, F. et T. Neily. 2010. A reconnaissance level survey of calciphilous lichens in selected karst topography in Nova Scotia with nots on incidental bryophytes. Report to Nova Scotia Department of Natural Resources. Mersey Tobeatic Research Institute, August 2010.16 pp.
B.C. Conservation Data Centre. 2016. BC Species and Ecosystems Explorer. B.C. Ministry of Environment, Victoria B.C. (Disponible en anglais seulement, dernière évaluation en avril 2016)
Beever, J.E. 1999. Studies of Fissidens (Bryophyta: Musci) in New Zealand: a synopsis and key to taxa. New Zealand Journal of Botany 37: 659–670.
Bryoquel (PDF; 136 Ko). 2015. Liste des taxons rares au Qc-Labr. (liste à jour des rangs des espèces résultant de la collecte et de l'enrichissement de données effectués en continu et de façon consultative; les rangs du Centre de données sur le patrimoine naturel du Québec, qui sont officiels, sont coordonnés avec ceux de Bryoquel). (consulté en août 2015)
Buck, W.R. et H.A. Crum. 1978. Entodon schleicheri new to North America. The Bryologist 81:429–432.
COSEWIC (Committee on the Status of Endangered Wildlife in Canada). 2005. COSEWIC assessment and status report on the pygmy pocket moss Fissidens exilis in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 18 pp. (Également disponible en français : COSEPAC (Comité sur la situation des espèces en péril au Canada). 2005. Évaluation et Rapport de situation du COSEPAC sur le fissident pygmée (Fissidens exilis) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. vii + 20 p.)
Crum, H.A. 2001. Structural Diversity of Bryophytes. University of Michigan Herbarium, Ann Arbor, 379 pp.
Crum, H.A. et L.E. Anderson. 1981. Mosses of Eastern North America. Columbia University Press, New York. 1328 pp.
During, H.J. 1979. Life strategies in bryophytes : a preliminary review. Lindbergia 5 :2–18.
Ecological Stratification Working Group. 1996. A National Ecological Framework for Canada. Agriculture and Agri-Food Canada, Research Branch, Centre for Land and Biological Resources Research, and Environment Canada, State of the Environment Directorate, Ecozone Analysis Branch, Ottawa/Hull. Report and national map at 1:7,500,000 scale. (Également disponible en français : Groupe de travail sur la stratification écologique. 1996. Cadre écologique national pour le Canada. Agriculture et Agroalimentaire Canada, Direction de la recherche, Centre de recherches sur les terres et les ressources biologiques, et Environnement Canada, Direction générale de l'état de l'environnement, Direction de l'analyse des écozones, Ottawa/Hull. Rapport et carte nationale 1/7 500 000.)
Environment Canada. 2015. Management Plan for the Pygmy Pocket Moss (Fissidens exilis) in Canada [Draft]. Species at Risk Act Management Plan Series. Environment Canada, Ottawa. iv + 16 pp. (consulté en août 2015) (Également disponible en français : Environnement Canada. 2015. Plan de gestion du fissident pygmée (Fissidens exilis) au Canada [Proposition]. Série de Plans de gestion de la Loi sur les espèces en péril. Environnement Canada, Ottawa. iv + 19 p. )
Faubert J. 2007. Catalogue des bryophytes du Québec et du Labrador. Provancheria N°30, Mémoire de l'Herbier Louis-Marie, Université Laval, 138 p.
Faubert, J. 2013. Flore des bryophytes du Québec-Labrador. Volume 2 : Mousses, première partie. Société québécoise de bryologie, Saint-Valérien, Québec, xiv + 402 p.
Furness, S.B. et R.H. Hall. 1981. An explanation of the intermittent occurrence of Phycomitrium sphaericum (Hedw.) Brid. Journal of Bryology 11:733–742.
Glime, Janice M. 2007. Bryophyte Ecology. Volume 1. Physiological Ecology. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. (Disponible en anglais seulement, consulté le 14 janvier 2015)
Goffinet, B., J.M. Budke et L.C. Newman. 2011. Micromitraceae: A new family of highly reduced mosses. Taxon 60:1245–1254.
Hallingbäck, T. et N. Hodgetts (compilers). 2000. Mosses, Liverworts and Hornworts. Status Survey and Conservation Action Plan for Bryophytes. International Union for Conservation of Nature and Natural Resources (IUCN) Bryophyte Specialist Group, Gland, Switzerland and Cambridge, UK. x + 106 pp.
Hassel, K. et L. Söderström. 1998. The presence of Pogonatum dentatum (Brid.) Brid. In roadside diaspore banks in Sweden. Lindbergia 23:113–118.
IUCN-CMP (International Union for Conservation of Nature and Conservation Measures Partnership). 2006. IUCN – CMP unified classification of direct threats, ver. 1.0 – June 2006. Gland, Switzerland. 17 pp. (Disponible en anglais seulement, consulté le 14 janvier 2015)
Iwatsuki, Z. et A. Noguchi. 1973. Index muscorum Japonicarum. Journal of the Hattori Botanical Laboratory 37:299–418.
McIntosh, T.T. 1989. Bryophyte records from the semiarid steppe of northwestern North America, including four species new to North America. The Bryologist 92:356–362.
Missouri Botanical Garden. 2014. Tropicos' details and synonyms for Fissidens exilis. (Disponible en anglais seulement, consulté le 31 décembre 2014).
Molnar, L. 1975. New distribution data on two mosses, Fissidens exilis and Thuidium pygmaeum, in Québec. The Canadian Field Naturalist 89: 324–325.
NatureServe. 2014. Arlington, VA. U.S.A. (Disponible en anglais seulement, consulté le 29 décembre 2014).
NHIC (Natural Heritage Information Centre). 2014. List of species ranks. (Disponible en anglais seulement, consulté le 29 décembre 2014).
Porley, R. 2008. Arable Bryophytes: A Field Guide to the Mosses, Liverworts and Hornworts of Cultivated Land in Britain and Ireland. WILDGuides Ltd., Hampshire. 140 pp.
Pursell, R.A. 2007. Fissidentaceae, pg. in Flora of North America Editorial Committee, Eds. Flora of North America North of Mexico. Vol 27. Oxford University Press, New York and Oxford. 713 pp.
Risk, A.C. 2002. The distribution, commonness and habitat characteristics of Fissidens hyalinus in the United States. The Bryologist 105:43–47.
Sackett, T.E., S.M. Smith et N. Basiliko. 2013. Indirect and direct effects of exotic earthworms on soil nutrient and carbon pools in North American temperate forests. Soil Biology & Biochemistry 57:459–467.
Schofield, W.B. 1981. Ecological significance of morphological characters in the moss gametophyte. The Bryologist 84:149–165.
Schofield, W.B. 1988. Bryophyte disjunctions in the Northern Hemisphere: Europe and North America. Botanical Journal of the Linnean Society (1988). 98:211–224.
Slack, N.G. 1990. Bryophytes and ecological niche theory. Botanical Journal of the Linnean Society 104:187–213.
Smith, A.J.E. 1978. The Moss Flora of Britain and Ireland. Cambridge University Press, Cambridge. 706 pp.
Smith, A.J.E. 2004. The Moss Flora of Britain and Ireland (2nd edition). Cambridge University Press, Cambridge.1012 pp.
SQB (Société québécoise de bryologie). 2014. Fil de nouvelles en ligne (consulté le 19 janvier 2015).
Steere, W.C. 1950. Notes on Fissidens. II. The discovery of Fissidens exilis in North America. The Bryologist 53: 131–136.
Vanderpoorten, A. et P. Engels. 2002. The effects of environmental variation on bryophtyes at a regional scale. Ecography 25:513–522.
Jennifer Doubt est bryologue et conservatrice en botanique à l'Herbier national du Canada du Musée canadien de la nature.
Voir le tableau 1.
| Impact des menaces | Impact des menaces (descriptions) | Comptes des menaces de niveau 1 selon l'intensité de leur impact : Maximum de la plage d'intensité |
Comptes des menaces de niveau 1 selon l'intensité de leur impact : Minimum de la plage d'intensité |
|---|---|---|---|
| A | Très élevé | 0 | 0 |
| B | Élevé | 0 | 0 |
| C | Moyen | 0 | 0 |
| D | Faible | 0 | 0 |
| - | Impact global des menaces calculé : | - | - |
| # | Menace | Impact (calculé) |
Impact (description) |
Portée (10 prochaines années) |
Gravité (10 années ou 3 générations) |
Immédiateté | Commentaires |
|---|---|---|---|---|---|---|---|
| 1 | Développement résidentiel et commercial (en anglais seulement) | - | - | - | - | - | - |
| 1.1 | Habitations et zones urbaines | - | - | - | - | - | Pas une menace. |
| 1.2 | Zones commerciales et industrielles | - | - | - | - | - | Pas une menace. (Ligne 542) : Le développement résidentiel et commercial pourrait constituer une menace, mais aucun projet de développement n'est prévu à l'heure actuelle. |
| 1.3 | Tourisme et espaces récréatifs | - | - | - | - | - | Pas une menace. Même chose qu'au point 1.2 (ci-dessus) : Pourrait constituer une menace si des projets étaient planifiés, mais rien n'indique que de tels projets sont prévus actuellement. - L'aménagement de sentiers pourrait être bénéfique pour cette espèce. |
| 2 | Agriculture et aquaculture (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Inconnue | - |
| 2.1 | Cultures annuelles et pluriannuelles de produits autres que le bois | - | Inconnu | Inconnue | Inconnue | Inconnue | Pas une menace. (Ligne 512) : Selon des études réalisées en Europe, les pratiques de gestion agricole pourraient à la fois être bénéfiques et constituer une menace pour l'espèce. Au Canada, l'espèce n'a pas été trouvée dans des champs cultivés (seulement en Europe). |
| 2.2 | Plantations pour la production de bois et de pâte | - | - | - | - | - | Pas une menace – pourrait être bénéfique. |
| 2.3 | Élevage et élevage à grande échelle | - | - | - | - | - | Pas une menace. |
| 2.4 | Aquaculture en mer et en eau douce | - | - | - | - | - | Pas une menace. |
| 3 | Production d'énergie et exploitation minière (en anglais seulement) | - | - | - | - | - | Pas une menace |
| 3.1 | Forage pétrolier et gazier | - | - | - | - | - | - |
| 3.2 | Exploitation de mines et de carrières | - | - | - | - | - | La plupart des sites de la N.-É. sont associés à des zones karstiques et basaltiques, qui sont généralement ciblées pour l'extraction de ressources. Cependant, aucun plan d'exploitation minière précis n'est connu concernant les sites où le fissident pygmée est présent. |
| 3.3 | Énergie renouvelable | - | - | - | - | - | - |
| 4 | Corridors de transport et de service (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Inconnue | - |
| 4.1 | Routes et voies ferrées | - | Inconnu | Inconnue | Inconnue | Inconnue | (Ligne 537) : La construction de routes pourrait avoir des effets néfastes (sels de voirie), mais aucune donnée ne le prouve. |
| 4.2 | Lignes de services publics | - | Inconnu | Inconnue | Inconnue | Inconnue | Même chose qu'au point 4.2 (ci-dessus) : Les lignes de services publics pourraient constituer une menace, mais celle-ci est inconnue. |
| 4.3 | Transport par eau | - | - | - | - | - | Pas une menace. |
| 4.4 | Trajectoires de vol | - | - | - | - | - | Pas une menace. |
| 5 | Utilisation des ressources biologiques (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Inconnue | - |
| 5.1 | Chasse et prélèvement d'animaux terrestres | - | - | - | - | - | Pas une menace. |
| 5.2 | Cueillette de plantes terrestres | - | - | - | - | - | Pas une menace. |
| 5.3 | Exploitation forestière et récolte du bois | - | Inconnu | Inconnue | Inconnue | Inconnue | (Ligne 528) : Effets positifs et négatifs. L'exploitation forestière perturberait le sol, mais pourrait aussi disperser les mousses. Les inconnues sont multiples; de l'habitat pourrait être créé, mais le site pourrait ne pas convenir à l'espèce pendant quelques années. |
| 5.4 | Pêche et récolte des ressources aquatiques | - | - | - | - | - | - |
| 6 | Intrusions et perturbations humaines (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | - |
| 6.1 | Activités récréatives | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | L'espèce a besoin de perturbations, mais pas trop. - Rien n'indique que le piétinement ait causé l'élimination de sous-populations. - L'espèce peut pousser en bordure de sentiers (C.-B. = 1 site; Ont. = 1 site; Qc = peut-être 2 sites) |
| 6.2 | Guerre, troubles civils et exercices militaires | - | - | - | - | - | - |
| 6.3 | Travaux et autres activités | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | - |
| 7 | Modification du système naturel (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | - |
| 7.1 | Incendies et suppression des incendies | - | - | - | - | - | Pas une menace. Les incendies sont supprimés dans de nombreux sites où pousse l'espèce. |
| 7.2 | Barrages, gestion et utilisation de l'eau | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | Les perturbations causées par les digues créent de l'habitat, mais le mode d'exploitation détermine si les digues constituent une menace ou non. |
| 7.3 | Autres modifications de l'écosystème | - | - | - | - | - | Le gravelage des sentiers pourrait constituer un problème; on ignore toutefois si des programmes importants de gravelage des sentiers sont prévus. |
| 8 | Espèces et gènes envahissants ou problématiques (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Inconnue | - |
| 8.1 | Espèces exotiques/non indigènes envahissantes | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | Rien n'a été signalé. |
| 8.2 | Espèces indigènes problématiques | - | Inconnu | Inconnue | Inconnue | Élevée (continue) | - |
| 8.3 | Introduction de matériel génétique | - | - | - | - | - | - |
| 9 | Pollution (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Modérée (peut-être à court terme, < 10 ans) | Préoccupations générales concernant la pollution près des champs agricoles et des routes qui entraînerait une modification de la composition du sol. |
| 9.1 | Eaux usées domestiques et urbaines | - | - | - | - | - | - |
| 9.2 | Effluents industriels et militaires | - | - | - | - | - | - |
| 9.3 | Effluents agricoles et forestiers | - | Inconnu | Inconnue | Inconnue | Modérée (peut-être à court terme, < 10 ans) | - |
| 9.4 | Détritus et déchets solides | - | - | - | - | - | - |
| 9.5 | Polluants atmosphériques | - | - | - | - | - | - |
| 9.6 | Énergie excessive | - | - | - | - | - | - |
| 10 | Phénomènes géologiques (en anglais seulement) | - | - | - | - | - | - |
| 10.1 | Volcans | - | - | - | - | - | - |
| 10.2 | Tremblements de terre et tsunamis | - | - | - | - | - | - |
| 10.3 | Avalanches et glissements de terrain | - | - | - | - | - | Pourrait être bénéfique. |
| 11 | Changement climatique et phénomènes météorologiques violents (en anglais seulement) | - | Inconnu | Inconnue | Inconnue | Inconnue | - |
| 11.1 | Déplacement et altération de l'habitat | - | - | - | - | - | - |
| 11.2 | Sécheresses | - | Inconnu | Inconnue | Inconnue | Inconnue | Pourrait être une menace. - Les sécheresses pourraient constituer un problème à long terme, mais pas au cours de la prochaine décennie. |
| 11.3 | Températures extrêmes | - | - | - | - | - | - |
| 11.4 | Tempêtes et inondations | - | Inconnu | Inconnue | Inconnue | Inconnue | - |