
Évaluation et Rapport de situation du COSEPAC sur la Pie-grièche migratrice Lanius ludovicianus au Canada - 2014
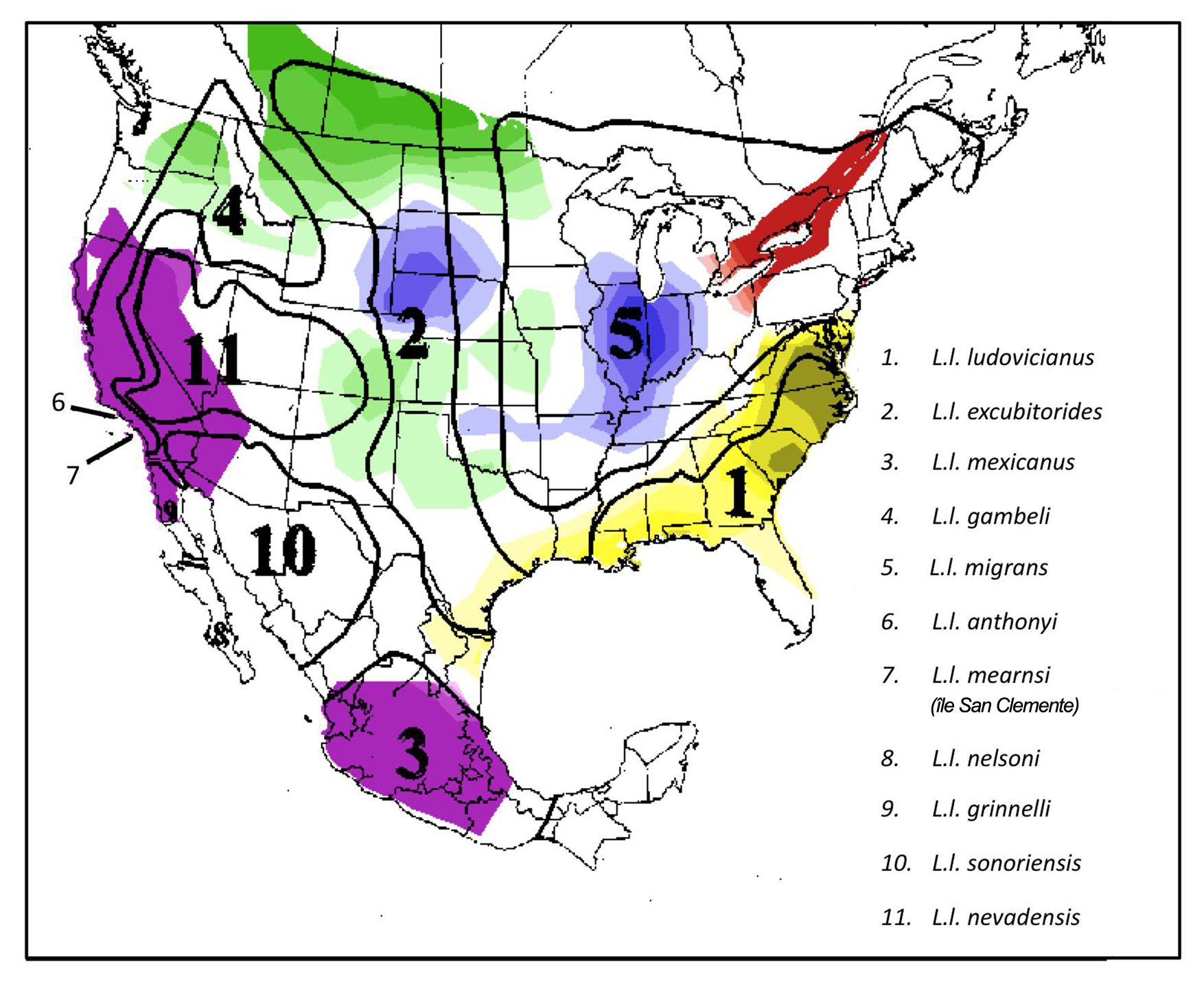
- Figure 1. Aires de reproduction en Amérique du Nord des différentes sous-espèces de Pies-grièches migratrices. Les aires numérotées délimitées par des lignes noires sont fondées sur les différences morphométriques relevées par Miller (1931). Ces aires sont comparées aux groupes génétiques identifiés par des couleurs différentes distingués d’après le coefficient d’admixture génétique des oiseaux échantillonnés par Chabot (2011a). Figure reproduite avec l’autorisation d’A. Chabot.
- Figure 2. Structure génétique estimée des populations chez la Pie-grièche migratrice selon Chabot (2011a). Chaque individu est représenté par une ligne verticale. Sont ici illustrés k = 5 groupes, chacun représenté par une couleur différente. Au sein de chaque groupe, les individus sont séparés par leur coefficient d’admixture génétique (Q). Les groupes génétiques sont identifiés sous la figure par les noms trinomiaux proposés par Miller (1931) et le groupe génétique présent dans l’est de l’Ontario. Figure reproduite avec l’autorisation d’A. Chabot.
- Figure 3. Aire de répartition nord-américaine de la Pie-grièche migratrice durant les saisons de reproduction et d’hivernage, d’après Yosef (1996). L’aire de répartition canadienne de la sous-espèce des Prairies englobe l’Alberta, la Saskatchewan et le Manitoba. L’aire de répartition canadienne de la sous-espèce de l’Est (sud de l’Ontario et extrême sud-ouest du Québec) consiste essentiellement en deux zones de nidification principales.
- Figure 4. Aire de reproduction canadienne de la Pie-grièche migratrice de la sous-espèce de l’Est, montrant le polygone utilisé aux fins du calcul de la zone d’occurrence (carte créée par A. Filion).
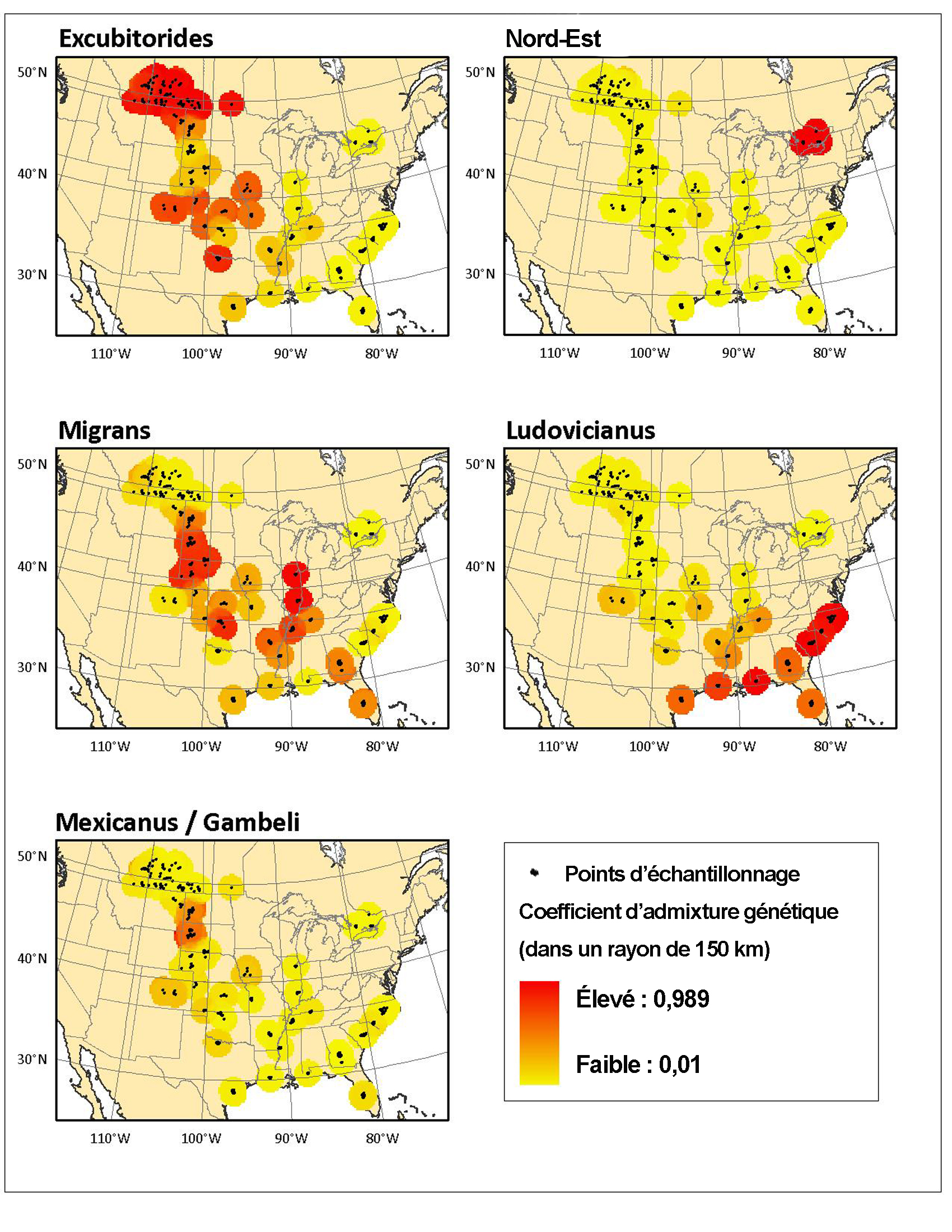
- Figure 5. Représentation géospatiale des estimations du degré de parenté génétique pour chaque groupe génétique de Pies-grièches migratrices échantillonné par Chabot (2011a). Les zones plus foncées indiquent des niveaux de flux génétique plus faibles parmi les zones échantillonnées voisines, et les zones plus claires, des niveaux de flux génétique plus élevés. Figure reproduite avec l’autorisation d’A. Chabot.
- Figure 6. Indices annuels des fluctuations de la population de la sous-espèce des Prairies de la Pie-grièche migratrice au Canada, d’après les résultats du Recensement des oiseaux nicheurs, 1970-2011. Graphique reproduit avec l’autorisation d’Environnement Canada (2013).
- Figure 7. Fluctuations du nombre de couples nicheurs de Pies-grièches migratrices dans le sud-ouest du Manitoba entre 1987 et 2013. La valeur de 2013 reflète une réduction de l’intensité des recherches et a donc été exclue de l’évaluation des tendances de la population. Données gracieusement fournies par K. DeSmet, ministère de la Conservation du Manitoba.
- Figure 8. Nombre de Pies-grièches migratrices matures recensées durant la saison de nidification lors des relevés ciblés réalisés en Ontario entre 1992 et 2013. Les relevés indiquent les nombres d’adultes appariés et d’adultes non appariés. Les résultats obtenus lors de l’année du premier relevé (1991) sont des sous-estimations et ont donc été omis. Données gracieusement fournies par le Service canadien de la faune, région de l’Ontario.
- Tableau 1. Estimations appariées de la différenciation des populations dérivées de données de microsatellites nucléaires entre les sous-espèces L. l. excubitorides, L. l. ludovicianus, L. l. migrans et la nouvelle sous-espèce présente en Ontario. PhiPT, un équivalent de FST; au-dessus de la diagonale. Les nombres en caractères gras indiquent des différences statistiquement significatives (valeurs de P sous la diagonale). N = 703 avec 9 999 permutations effectuées au moyen de sous-programmes dans GENALEX (Peakall et Smouse, 2006). La variation génétique parmi les sous-groupes a d’abord été mesurée au moyen de PhiPT, un équivalent de FST, qui calcule la différenciation des populations d’après la variance génotypique, où les plus faibles niveaux de flux génétique entre les sous-groupes sont indiqués par des valeurs plus élevées. Voir Chabot (2011a) pour de plus amples renseignements sur les régions échantillonnées
- Tableau 2. Estimations du nombre de Pies-grièches migratrices (individus matures) entre 1987 et 2013 d’après les résultats des relevés quinquennaux réalisés en Alberta et en Saskatchewan et de relevés annuels effectués au Manitoba
- Tableau 3. Tendances à long terme et à court terme des populations de la Pie-grièche migratrice aux États-Unis et au Canada révélées par le Relevé des oiseaux nicheurs (Sauer et al., 2011; Environnement Canada, 2013). Les tendances correspondent au changement annuel moyen en pourcentage calculé par modélisation hiérarchique. Les intervalles de confiance à 95 % représentent les percentiles 2,5 % et 97,5 % de la distribution postérieure des estimations des tendances. Les valeurs statistiquement significatives sont indiquées en caractères gras.

Les rapports de situation du Comité sur la situation des espèces en péril au Canada (COSEPAC) sont des documents de travail servant à déterminer le statut des espèces sauvages que l’on croit en péril. On peut citer le présent rapport de la façon suivante :
Évaluation et Rapport de situation du COSEPAC sur la Pie-grièche migratrice de la sous-espèce de l'Est (Lanius ludovicianus ssp.) et la sous-espèce des Prairies (Lanius ludovicianus excubitorides) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xiv + 57 p. (Registre public des espèces en péril site Web).
Rapport(s) précédent(s) :
COSEWIC. 2004. COSEWIC assessment and update status report on the Loggerhead Shrike excubitorides subspecies Lanius ludovicianus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. vi + 24 pp. (Registre public des espèces en péril site Web).
COSEWIC. 2000. COSEWIC assessment and update status report on the Loggerhead Shrike migrans subspecies, Lanius ludovicianus migrans in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. viii + 13 pp. (Registre public des espèces en péril site Web).
James, R.D. 2000. Update COSEWIC status report on the Loggerhead Shrike migrans subspecies, Lanius ludovicianus migrans in Canada, in COSEWIC assessment and status report on the Loggerhead Shrike migrans subspecies, Lanius ludovicianus migrans in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 1-13
Cadman, M.D. 1991. Update COSEWIC status report on the Loggerhead Shrike (Eastern population) Lanius ludovicianus migrans in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 26 pp.
Cadman, M.D. 1986. COSEWIC status report on the Loggerhead Shrike Lanius ludovicianus in Canada. Committee on the Status of Endangered Wildlife in Canada. Ottawa. 100 pp.
Le COSEPAC remercie Amy Chabot d’avoir rédigé le rapport de situation sur la Pie-grièche migratrice (Lanius ludovicianus) des sous-espèces de l’Est et des Prairies, aux termes d’un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par Jon McCracken, coprésident du sous-comité de spécialistes des oiseaux du COSEPAC.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-953-3215
Téléc. : 819-994-3684
COSEPAC Courriel
COSEPAC site Web
Also available in English under the title COSEWIC assessment and status report on the Loggerhead Shrike Eastern subspecies Lanius ludovicianus ssp. and the Prairie subspecies Lanius ludovicianus excubitorides in Canada.
Illustration/photo de la couverture :
Pie-grièche migratrice -- Photo par Larry Kirtley (utilisé avec permission).
© Sa Majesté la Reine du chef du Canada, 2014.
No de catalogue CW69-14/390-2014F-PDF
ISBN 978-0-660-22201-1

Sommaire de l’évaluation - mai 2014
Sommaire de l’évaluation - mai 2014
La Pie-grièche migratrice est un oiseau chanteur de taille moyenne (environ 21 23 cm de longueur). Les deux sexes sont semblables. Le sommet de la tête, le dos et le croupion sont gris foncé, tandis que les parties inférieures sont blanches à grisâtres. Les ailes sont en grande partie noires, avec une tache blanche bien visible en vol. Les rectrices sont noires, certaines avec l’extrémité blanche. Un masque facial couvre les yeux et s’étend jusqu’au bec. Le bec, crochu, évoque celui d’un rapace. Oiseau prédateur, la pie-grièche migratrice se distingue par son habitude d’empaler ses proies. Elle peut également être utile à titre d’espèce bioindicatrice ou d’espèce phare pour les oiseaux de prairie à statut de conservation précaire.
La Pie-grièche migratrice se rencontre uniquement en Amérique du Nord. Dans l’Ouest canadien, elle est présente du sud-ouest de l’Alberta au sud du Manitoba, en passant par le sud de la Saskatchewan. Dans l’est du pays, elle est régulièrement observée dans seulement deux régions du sud de l’Ontario, et elle se rencontre seulement de façon sporadique dans le sud-ouest du Québec. L’espèce est migratrice saisonnière. L’aire d’hivernage des individus nichant au Canada chevauche celle des oiseaux qui résident en permanence aux États-Unis.
Deux unités désignables sont distinguées au Canada : la sous-espèce des « Prairies » (L. l. excubitorides), présente au Manitoba, en Saskatchewan et en Alberta, et la sous-espèce de l’Est, présente en Ontario et au Québec. Les individus de l’Ontario et du Québec étaient assignés jusqu’à maintenant à la sous-espèce L. l. migrans, mais de nouvelles données génétiques révèlent qu’ils forment un groupe génétique distinct appartenant à une sous-espèce encore non nommée.
La Pie-grièche migratrice niche dans des milieux ouverts dominés par les graminées et/ou les herbacées non graminoïdes et parsemés de quelques arbustes ou arbres et de zones de sol dénudé. Les pâturages, les friches, les prairies, les savanes, les forêts claires de pinyon-genévrier, les steppes arbustives et les alvars constituent des milieux propices à sa nidification. Les habitats d’hivernage et de migration présentent des caractéristiques similaires à celles de l’habitat de reproduction. La superficie des territoires oscille entre 2,7 et 47,0 ha et est inversement proportionnelle à la densité des arbres et des arbustes pouvant servir de perchoirs. Le nid est habituellement aménagé dans un arbre ou un arbuste à cime dense et très ramifiée.
La Pie-grièche migratrice peut arriver sur les lieux de nidification au Canada dès la fin de mars. Le nid contient en moyenne 5 ou 6 œufs. L’incubation dure entre 16 et 18 jours. Les jeunes prennent leur premier envol à l’âge de 16 à 20 jours. La Pie-grièche migratrice se montre fidèle au site de nidification, mais le taux de réutilisation des sites est variable et plus élevé chez les mâles que chez les femelles. La fidélité au site de nidification est plus élevée chez les adultes que chez les jeunes et semble liée au succès de la nidification au cours de la saison précédente.
La Pie-grièche migratrice subit un déclin soutenu depuis de nombreuses années à l’échelle de son aire de nidification nord-américaine et canadienne. Riche d’environ 55 000 individus, la sous-espèce des Prairies au Canada est en déclin depuis au moins 1970, et ses effectifs ont chuté d’environ 47 % au cours des dix dernières années. La sous-espèce de l’Est a également subi un important déclin, tant en ce qui a trait à la taille de sa population qu’à la superficie de son aire de reproduction. La population compte moins de 100 individus en Ontario, et moins de 10 individus au Québec. La sous-espèce est tenue pour disparue au Nouveau-Brunswick depuis les années 1970.
La disparition et la dégradation de l’habitat de reproduction et d’hivernage sont tenues pour responsables des déclins d’effectifs observés à l’échelle de l’aire de répartition de l’espèce. La mortalité routière, les pesticides, la prédation et les phénomènes météorologiques extrêmes auraient également contribué au déclin de l’espèce. Le virus du Nil occidental a également été incriminé dans la mort d’un certain nombre d’individus, mais la gravité de cette menace est actuellement inconnue.
La Pie-grièche migratrice bénéficie d’une protection internationale (Canada, Mexique et États-Unis) en vertu de la Loi sur la Convention concernant les oiseaux migrateurs. En vertu de la Loi sur les espèces en péril du gouvernement fédéral, la sous-espèce des Prairies (L. l. excubitorides) est actuellement désignée « menacée », et la sous-espèce de l’Est (anciennement appelée L. l. migrans), « en voie de disparition ». Des ébauches de programme de rétablissement ont été élaborées pour les deux unités désignables. À l’échelle provinciale, la Pie-grièche migratrice est cotée « espèce sensible » (Sensitive Species) et « espèce préoccupante » (Species of Special Concern) en Alberta. Elle est désignée « espèce en voie de disparition » au Manitoba et en Ontario, et « espèce menacée au Québec. Elle est considérée comme en voie de disparition, menacée ou préoccupante dans 26 États.
Comme la vaste majorité des parcelles d’habitat propice à la Pie-grièche migratrice se trouvent sur des terres privées, il y a lieu de s’interroger sur l’efficacité des mécanismes de protection juridique en place.
Lanius ludovicianus excubitorides
Pie-grièche migratrice de la sous-espèce des Prairies (Unité désignable 1)
Loggerhead Shrike Prairie Subspecies (Designatable Unit 1)
Données démographiques
Information sur la répartition
| Population | Nombre d’individus matures |
|---|---|
| Alberta | 15 000 |
| Saskatchewan | 39 600 |
| Manitoba | 100 à 200 |
| Total | 54 700 à 54 800 |
Analyse quantitative
Menaces (réelles ou imminentes pour les populations ou leur habitat)
- Disparition et dégradation de l’habitat de reproduction, de migration et d’hivernage.
- Pesticides
- Mortalité routière (adultes et jeunes ayant pris leur envol).
Immigration de source externe (immigration de l’extérieur du Canada)
Historique du statut
Espèce désignée « menacée » en mai 2014.
Statut et justification de la désignation
Applicabilité des critères
Lanius ludovicianus ssp.
Pie-grièche migratrice de la sous-espèce de l'Est (Unité désignable 2)
Loggerhead Shrike Eastern Subspecies (Designatable Unit 2)
Données démographiques
Information sur la répartition
| Population | Nombre d’individus matures |
|---|---|
| Ontario | 50 à 100 |
| Québec | 10 (maximum) |
| Total | 60 à 110 |
Analyse quantitative
Menaces (réelles ou imminentes pour les populations ou leur habitat)
- Disparition et dégradation de l’habitat de reproduction, de migration et d’hivernage.
- Pesticides
- Prédation entraînant une réduction du succès de la nidification.
- Mortalité routière (adultes et jeunes ayant pris leur envol).
Immigration de source externe (immigration de l’extérieur du Canada)
Historique du statut
L’espèce a été considérée comme une unité et a été désignée « menacée » en avril 1986. Division en sous-espèces (excubitorides et migrans) en avril 1991, et chacune a reçu une désignation spécifique. La sous-espèce migrans a été désactivée en mai 2014 en raison de la nouvelle information génétique indiquant que certains individus du sud-est du Manitoba n’auraient pas dû être inclus dans la sous-espèce migrans. Divisée par la suite en une nouvelle sous-espèce n’ayant pas de nom (sous-espèce de l’Est, Lanius ludovicianus ssp.) en mai 2014 et a été désignée « en voie de disparition ».
Statut et justification de la désignation
Applicabilité des critères
Les recherches ciblant la Pie-grièche migratrice effectuées au cours de la dernière décennie comportaient trois grands objectifs :
- documenter la répartition et les tendances des populations;
- évaluer la structure génétique des populations;
- caractériser davantage l’aire d’hivernage de l’espèce.
D’autres travaux réalisés dans l’est du Canada ont permis d’évaluer la structure par âge des populations, le degré de fidélité au site de nidification et les taux d’immigration d’après les données amassées dans le cadre d’un programme de baguage couleur à long terme. Des analyses visant à établir un lien entre l’occurrence de l’espèce et divers attributs de l’habitat ont été effectuées en Ontario. Au Québec, un important projet de cartographie visant à répertorier les régions les plus propices à l’espèce a été réalisé.
D’après les résultats d’une analyse de microsatellites de l’ADN nucléaire provenant d’un échantillon d’oiseaux issus de diverses régions de l’Amérique du Nord, les individus présents en Ontario représenteraient un groupe génétique unique jusque-là non documenté et significativement différent du L. l. migrans auquel il était présumé appartenir. Deux unités désignables génétiquement distinctes et présentant peu d’échanges génétiques entre elles sont présentes au Canada. À une certaine époque, les sous-espèces L. l. excubitorides et L. l. migrans semblaient s’hybrider dans le sud-est du Manitoba. Tout individu rencontré dans cette portion de la province est vraisemblablement un représentant de la sous-espèce des Prairies. On estime aujourd’hui que les individus du sud-est du Manitoba n’auraient pas dû être considérés comme faisant partie de la sous-espèce L. l. migrans lors de la dernière évaluation de la situation de la sous-espèce par le COSEPAC.
Divers projets d’intendance de l’habitat misant sur la participation de propriétaires privés aux initiatives de rétablissement de l’espèce sont en cours dans l’ouest et l’est du Canada. En Ontario et au Québec, un programme d’élevage en captivité et de réintroduction en nature a connu un succès mitigé.

Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsable des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Espèce, sous-espèce, variété ou population géographiquement ou génétiquement distincte d’animal, de plante ou d’un autre organisme d’origine sauvage (sauf une bactérie ou un virus) qui est soit indigène du Canada ou qui s’est propagée au Canada sans intervention humaine et y est présente depuis au moins cinquante ans.
Le Service canadien de la faune d’Environnement Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
La Pie-grièche migratrice (Lanius ludovicianus) est un oiseau chanteur de taille moyenne qui mesure en moyenne 20 à 22 cm de longueur et pèse 47 à 48 g (Dunning, 1993; Yosef, 1996; Chabot, données inédites). Le sommet de la tête, le dos et le croupion sont gris foncé, tandis que les parties inférieures sont blanches à grisâtres. Les ailes sont en grande partie noires, mais la base des primaires dessine une tache blanche bien visible en vol. Les rectrices sont en grande partie noires, mais certaines ont l’extrémité blanche, la quantité de blanc augmentant progressivement du centre au bord externe de la queue. La Pie-grièche migratrice se différencie de la Pie-grièche grise (Lanius excubitor) par son masque facial noir qui se prolonge au-dessus des yeux en une fine ligne jusqu’à la base du bec; chez la Pie-grièche grise, le masque ne s’étend pas au-delà des yeux. Le bec est noir, mais peut présenter une teinte plus claire, particulièrement au niveau de la mandibule et principalement chez les femelles. Le bec est crochu comme celui des rapaces et armé de chaque côté d’une dent tomiale (Cade, 1995). La Pie-grièche migratrice doit son nom anglais de loggerhead à l’aspect de sa tête, relativement massive en raison des forts muscles qui confèrent au bec sa puissance.
Les deux sexes sont semblables. Durant la saison de reproduction, les mâles ont généralement la mandibule foncée et la poitrine blanche, tandis que les femelles ont souvent la mandibule pâle et la poitrine teintée de havane. Les mâles sont généralement plus grands que les femelles (Haas, 1987; Collister et Wicklum,1996; Yosef, 1996; Chabot, données inédites).
Les juvéniles ressemblent aux adultes, mais ils ont la poitrine finement barrée de lignes diffuses avant la première mue, qui survient à la fin de l’été ou au début de l’automne (Burnside, 2006). Ils ont également les ailes et la queue plus brunes que les adultes (Miller, 1928; Burnside, 2006; Chabot, 2011a).
Le chant printanier des mâles consiste en une série de trilles brefs ou d’une combinaison de notes claires répétées plusieurs fois et de rythme, tonalité et qualité variables. Le chant territorial, émis tant par les mâles que par les femelles, est semblable, mais contient des notes plus rauques qui ressemblent aux cris des oisillons quémandant leur nourriture. Les vocalises des mâles sont plus rapides et plus longues que celles des femelles et comportent des trilles, contrairement à celles des femelles (Soendjoto, 1995). Le cri d’alarme, émis lorsqu’un intrus ou un prédateur est repéré, consiste en une série de notes rauques.
À l’échelle de son aire de répartition, la Pie-grièche migratrice forme des colonies ou agrégations reproductrices lâches (Cade et Woods, 1997; Pruitt, 2000; Etterson, 2003). Woods (1994) mentionne avoir observé des regroupements spatiaux (« spatial clumping ») de Pies-grièches migratrices nichant dans des peuplements à armoise. La structure spatiale des populations reproductrices de la Pie-grièche migratrice a souvent été décrite comme irrégulière. Dans de nombreuses régions, des milieux apparemment propices demeurent inoccupés, alors qu’ailleurs des parcelles d’habitat similaire sont occupées par des groupes isolés. L’espèce est donc souvent répartie en populations reproductrices isolées spatialement et souvent de petite taille.
Chez la Pie-grièche migratrice, la coloration du plumage, en particulier la coloration du dos et la quantité de blanc sur la queue, le croupion, les sus-caudales et les scapulaires, varie d’une région à l’autre. Les dimensions du bec et la longueur de la queue et des ailes peuvent également différer d’une région à l’autre (Chabot, données inédites). Dans le cadre de la plus importante révision taxinomique fondée sur des caractères morphologiques réalisée à ce jour, Miller (1931) a scindé le taxon en 11 sous-espèces par classe d’âge et mue. Subséquemment, Rand (1960) et Phillips et Rea (Phillips, 1986) ont réduit le nombre de sous-espèces, mais proposé une nouvelle sous-espèce confinée au sud de la Floride, le L. l. miamensis.
Dans une étude utilisant un segment d’environ 200 paires de bases de la région de régulation de l’ADN mitochondrial et de la séquence du gène du « cytochrome b », Mundy et al. (1997) n’ont observé aucun haplotype unique parmi les populations des sous-espèces L. l. anthonyi, L. l. mearnsi, L. l. gambeli et L. l. excubitorides, même si un haplotype différent dominait chez chacune d’entre elles. Un test G étendu à toutes les populations a révélé l’existence d’une hétérogénéité significative dans les fréquences des haplotypes entre les différents sites d’échantillonnage et, dès lors, entre les sous-espèces (Mundy et al., 1997). La valeur de FST pour les cinq populations s’élevait à 0,78 (P <0,05), et toutes les paires intersubspécifiques de populations étaient hautement significatives (P <0,01). Les résultats de Mundy et al. (1997) sont étayés par des analyses récentes utilisant 10 nouveaux marqueurs microsatellites nucléaires (Coxon et al., 2011) et un fragment d’ADNmt d’environ 250 paires de bases (A. Coxon, comm. pers., 2012).
Vallianatos et al. (2002) ont examiné la structure génétique de populations de Pies-grièches migratrices du centre et de l’est de l’Amérique du Nord. Ils ont analysé un segment de 267 paires de bases de la séquence de la région de régulation de l’ADN mitochondrial provenant d’échantillons prélevés dans l’aire de répartition de trois sous-espèces présumées – L. l. migrans, L. l. ludovicianus et L. l. excubitorides – et leurs zones d’hybridation. L’analyse de variance moléculaire a révélé qu’une portion significative de la variation spatiale totale de la région régulatrice était répartie entre les trois sous-espèces (24,4 %; P <0,01). Les auteurs de cette étude ont proposé quatre unités de gestion correspondant aux désignations des sous-espèces, tout en scindant la sous-espèce L. l. migrans de manière telle que les populations de l’Ontario, du Québec et du nord-est des États-Unis (une unité de gestion de l’Est) étaient considérées génétiquement distinctes de celles du Manitoba, de l’Illinois, de l’Iowa, du Missouri, du Minnesota et d’autres États de l’Ouest compris dans l’aire de répartition occidentale présumée du L. l. migrans (faisant du L. l. migrans une unité de gestion occidentale; Vallianatos et al., 2002).
Plus récemment, Chabot (2011a) a réalisé une évaluation à grande échelle de la structure spatiale génétique de la population de Pie-grièche migratrice faisant appel à 15 marqueurs microsatellites, dont certains déjà utilisés par Mundy et al. (1997) et Coxon et al. (2011). Cette étude a une fois de plus validé la désignation des sous-espèces L. l. migrans, L. l. gambeli, L. l. excubitorides et L. l. ludovicianus par Miller (1931) et des unités de gestion proposées par Vallianatos et al. (2002) pour le L. l. migrans (Figure 1.). Toutefois, ces nouveaux résultats semblent indiquer que les oiseaux de l’Ontario (et du Québec par inférence) sont génétiquement distincts du L. l. migrans (figures 1 et 2). Les conclusions des études moléculaires ont été de façon générale étayées par des données morphométriques (Chabot, 2011a). Dans le cadre d’analyses additionnelles des données de microsatellites réalisées pour les besoins du présent rapport de situation et faisant appel à la statistique PhiPT (un équivalent de FST), les populations ont été désignées a priori d’après les résultats de l’analyse bayésienne par segmentation (Chabot, 2011a). Les résultats de ces analyses ont montré que les groupes génétiques reconnus par Chabot (2011a; figures 1 et 2) sont effectivement génétiquement distincts (Tableau 1; P <0,001).
Carte : © Environnement Canada
Description longue pour la figure 1
Carte illustrant l’aire de reproduction en Amérique du Nord de 11 sous espèces différentes de Pies grièches migratrices. La carte présente également deux types d’information : 1) les aires où les oiseaux sont morphométriquement différents (d’après les caractéristiques déterminées par Miller, 1931) et 2) les aires où les oiseaux sont groupés génétiquement d’après le coefficient d’admixture génétique échantillonnés par Chabot (2011a). Les oiseaux de l’Ontario (et du Québec par inférence) sont indiqués comme génétiquement distincts du L. l. migrans.
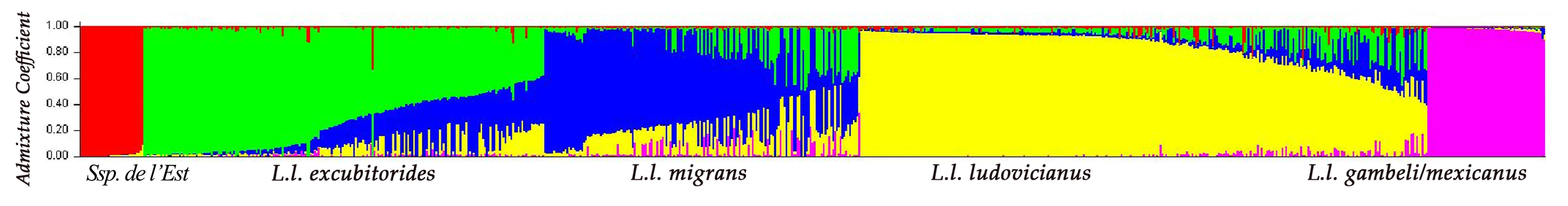
Figure : © A. Chabot, Environnement Canada
Description longue pour la figure 2
Graphique illustrant la structure génétique estimée des Pies grièches migratrices selon Chabot (2011a). L’axe y représente le coefficient d’admixture, et l’axe x, les groupes génétiques. Chaque oiseau correspond à une ligne verticale. Sont illustrés ici k = 5 groupes, chacun représenté par une couleur différente. Au sein de chaque groupe, les individus sont séparés par leur coefficient d’admixture génétique (Q). Les groupes génétiques sont identifiés sous la figure par les noms trinomiaux (L. l. excubitorides, L. l. migrans, L. l. ludovicianus, L. l. gambeli/mexicanus) proposés par Miller (1931) et le groupe génétique présent dans l’est de l’Ontario.
| Groupe génétique | L. l. excubitorides | L. l. ludovicianus | L. l. migrans | Ontario |
|---|---|---|---|---|
| L. l. excubitorides | - | 0,046 | 0,039 | 0,089 |
| L. l. ludovicianus | 0,001 | - | 0,047 | 0,124 |
| L. l. migrans | 0,001 | 0,001 | - | 0,121 |
| Ontario | 0,001 | 0,001 | 0,001 | - |
À la lumière des conclusions de Miller (1931), de Vallianatos et al. (2002) et de Chabot (2011a), deux unités désignables de Pies-grièches migratrices sont tenues pour présentes au Canada, à savoir la sous-espèce des Prairies (L. l. excubitorides) et une sous-espèce encore à nommer présente dans le sud de l’Ontario et du Québec. Cette dernière unité était traitée encore tout récemment comme une sous-population du L. l. migrans (Miller, 1931), mais elle est aujourd’hui considérée comme un vestige d’un groupe génétique unique qui demeure à nommer (Chabot, 2011a). Pour les besoins du présent rapport, nous appliquerons l’expression « Pie-grièche migratrice de l’Est » à cette unité désignable, désormais considérée comme distincte du L. l. migrans.
Vallianatos et al. (2002) considérait le centre-sud et le sud-est du Manitoba comme une zone d’hybridation entre le L. l. excubitorides et le L. l. migrans. Les résultats de Chabot (2011a) semblent indiquer que les individus rencontrés dans le sud-est du Manitoba au cours des dix dernières années correspondent génétiquement à la sous-espèce L. l. excubitorides, mais cette hypothèse est peut-être un artéfact de la disparition récente de l’espèce dans le centre-nord des États-Unis (Pruitt, 2000; Chabot, données inédites). Il est possible que le sud-est du Manitoba ait abrité autrefois une troisième unité désignable formée d’individus de la sous-espèce L. l. migrans, mais la chose est impossible à démontrer.
Outre leurs différences génétiques et morphométriques, les deux unités désignables susmentionnées occupent deux écozones distinctes au Canada. La sous-espèce des Prairies se rencontre principalement dans l’écozone des Prairies, même si son aire de répartition s’étendait anciennement jusqu’à l’écozone des Plaines boréales vers le nord et, vers l’ouest, jusqu’à l’écozone de la Cordillère montagnarde en Alberta (Cadman, 1985, 1990). La sous-espèce de l’Est est confinée à l’écozone des Plaines à forêts mixtes. L’écart important entre les aires de répartition actuelles des deux unités désignables est suffisant pour empêcher tout déplacement et, du coup, tout échange génétique entre les deux unités désignables.
La Pie-grièche migratrice est la seule espèce du genre Lanius endémique à l’Amérique du Nord (Lefranc, 1997). À la fois des passereaux et des prédateurs de niveau trophique supérieur, les pies-grièches occupent une position unique dans la chaîne alimentaire. Leur bec crochu pourvu d’une dent tomiale est fonctionnellement similaire au bec entaillé des faucons et leur permet d’être des prédateurs de vertébrés, ce qui les distingue des autres oiseaux chanteurs. Les proies de plus grande taille sont souvent empalées sur des objets pointus tels que des épines ou des fils barbelés. Ce comportement représente une adaptation unique qui permet aux pies-grièches de consommer des proies de forte taille même si elles n’ont pas de fortes serres et des pattes robustes comme les rapaces. Le nom générique Lanius (= boucher en français) et le nom familier anglais butcher bird reflètent le sentiment de répulsion que ce comportement inspire à certains observateurs plus sensibles.
Des travaux visant à préciser les besoins en matière d’habitat de l’espèce et à caractériser les milieux avec lesquels elle est associée en Ontario (Cuddy et Leviton, 1996; Glynn-Morris, 2010; Chabot et Lagios, 2012) semblent indiquer que la Pie-grièche migratrice pourrait être une espèce phare appropriée, dans la mesure où les initiatives visant à protéger son habitat se révéleront bénéfiques pour les autres espèces d’oiseaux de prairie qui connaissent également un déclin généralisé (Berlanga et al., 2010). La Commission de coopération environnementale (CCE) de l’Amérique du Nord a identifié la Pie-grièche migratrice comme l’une des onze espèces d’oiseaux qui suscitent des préoccupations communes en matière de conservation au Canada, aux États-Unis et au Mexique et qui sont considérées à ce titre comme des espèces porte-étendard (CCE, 2008).
La Pie-grièche migratrice a disparu de la majorité des régions de l’est du Canada et du nord-est des États-Unis comprises dans son aire de répartition historique. La population ontarienne est la plus grande et potentiellement la seule représentante restante de l’Unité désignable de l’Est (Chabot, 2011a) et présente à ce titre une importance particulière en ce qui a trait à la préservation de la diversité génétique au sein de l’espèce.
Aucune connaissance traditionnelle autochtone se rattachant à cette espèce n’a été répertoriée.
La Pie-grièche migratrice se rencontre uniquement en Amérique du Nord. À l’échelle du continent, elle présente une vaste aire de répartition qui englobe la majeure partie du Mexique et des États-Unis et le sud du Canada (Figure 3). Il n’est toutefois pas certain qu’elle soit présente dans la région de la côte du golfe au Mexique durant la saison de nidification (G. Perez, comm. pers., 2006). Dans les portions septentrionales de son aire de répartition, elle doit obligatoirement migrer vers le sud à l’approche de l’hiver. Plus au sud, de façon générale au sud du 40° de latitude et à l’extérieur du Canada, l’espèce est migratrice facultative ou comporte un certain pourcentage de résidents permanents (Figure 3). Les aires d’hivernage des individus migrateurs et des résidents permanents se chevauchent partiellement.
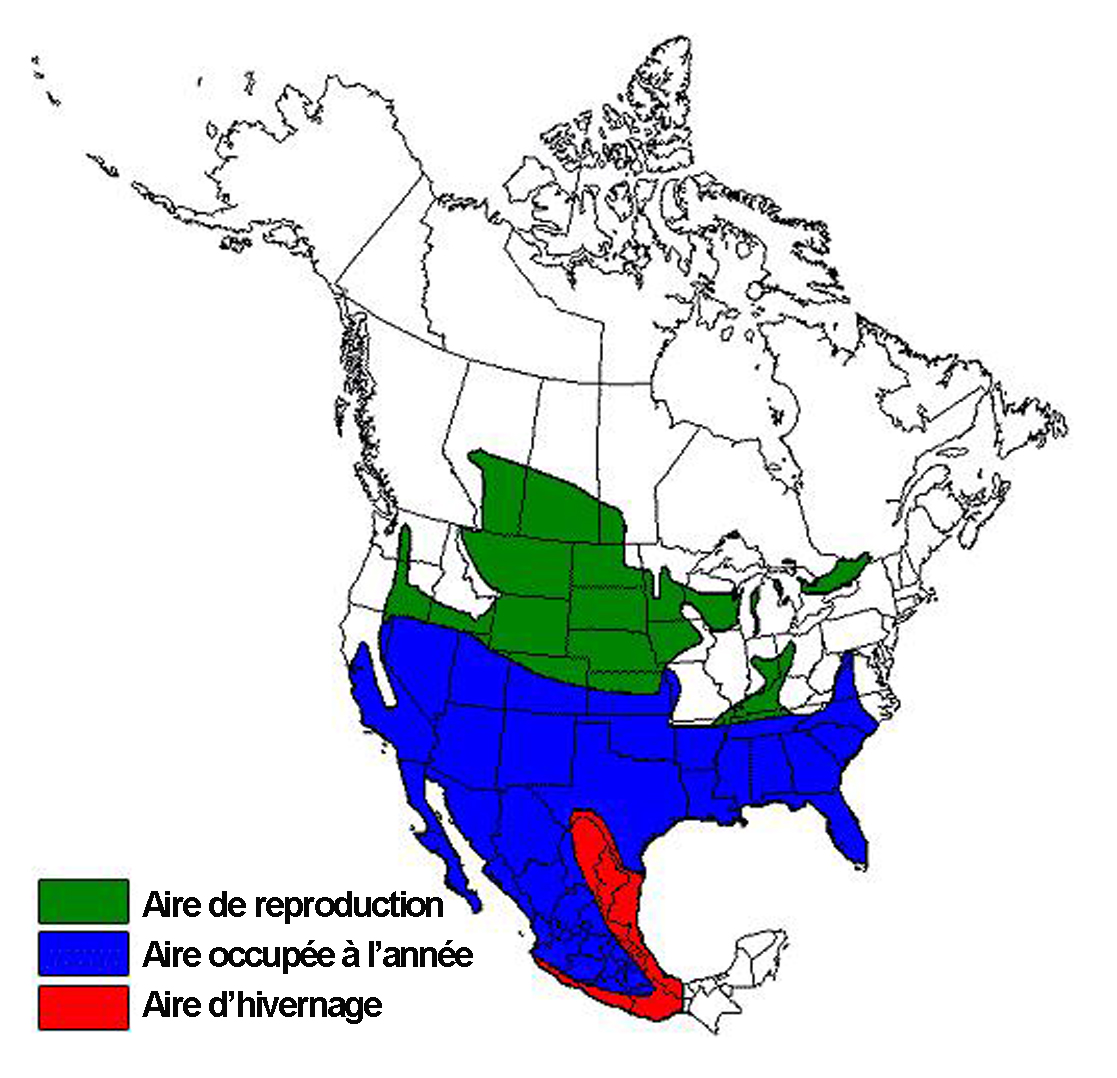
Carte : © Environnement Canada
Description longue pour la figure 3
Carte montrant l’aire de répartition nord américaine de la Pie grièche migratrice; on l’observe dans la plus grande partie du Mexique ainsi qu’aux États Unis et dans le sud du Canada. L’aire de reproduction, l’aire occupée à l’année et l’aire d’hivernage sont illustrées. L’aire de répartition canadienne de la sous espèce des Praires (L. l. excubitorides) englobe l’Alberta, la Saskatchewan et le Manitoba. L’aire de répartition canadienne de la sous espèce de l’Est (L. ludovicianus spp.) s’étend dans le sud de l’Ontario et l’extrême sud ouest du Québec.
Les limites exactes de l’aire de répartition de chaque sous-espèce ne peuvent être déterminées avec précision en raison de l’hybridation qui semble se produire dans les zones de contact entre les diverses sous-espèces. Avant le déclin de l’espèce, qui a eu pour effet de limiter son occurrence dans certaines anciennes zones de contact, les populations de la sous-espèce L. l. excubitorides s’hybridaient avec les sous-espèces présumées L. l. gambeli et L. l. nevadensis dans la région des Rocheuses, avec le L. l. mexicanus dans le nord du Mexique (Miller, 1931; Chabot, 2011a) et avec le L. l. migrans dans les Grandes Plaines et au Manitoba (Vallianatos et al., 2001; Chabot, 2011a).
L’aire de reproduction de la sous-espèce des Prairies, le L. l. excubitorides, s’étend du sud-est de l’Alberta jusque dans le sud-ouest du Manitoba et, vers le sud, à travers les Grandes Plaines jusqu’au centre du Texas et, à l’ouest, du nord-est de l’Idaho jusque dans le sud-est de la Californie et l’ouest du Texas, le désert Sonora et le nord de l’État de Durango, au Mexique (Figure 1.).
Anciennement, le L. l. migrans nichait probablement depuis le sud-est du Manitoba jusque dans l’est du Texas, le centre de la Louisiane et l’ouest de la Caroline du Nord et de la Virginie. Vers l’est, la sous-espèce était tenue pour présente jusque dans les Maritimes, mais les populations du nord-est des États-Unis étaient probablement plus étroitement apparentées génétiquement aux populations de l’Ontario, assignées ici à l’unité désignable de l’Est. La « véritable » sous-espèce L. l. migrans ne se rencontre plus aujourd’hui qu’en populations isolées en Illinois, en Indiana, en Ohio, en Iowa et plus au sud (Figure 1.).
En raison du mélange des sous-espèces dans l’aire d’hivernage et de la faible connectivité migratoire (Chabot, 2011a), il est difficile de préciser les limites des aires de reproduction et d’hivernage de populations individuelles. Il semble toutefois que les populations qui se reproduisent dans l’Ouest canadien migrent vers le sud-est, tandis que les individus qui nichent dans l’est du Canada migrent soit vers le sud, en se déplaçant à l’est des Adirondack jusqu’à la côte atlantique, soit vers le sud-ouest (Burnside, 1987; Hobson et Wassenaar, 1997; Perez, 2006; Perez et Hobson, 2007; Chabot, 2011a).
Au Canada, la sous-espèce des Prairies (L. l. excubitorides) est présente en Alberta, en Saskatchewan et au Manitoba (Figure 3). En Alberta, elle se rencontrait anciennement dans la portion centrale de la tremblaie-parc (Kiliaan et Prescott, 2002) et dans la région des Prairies (Salt et Wilk, 1958). Elle était également présente vers le nord dans la région de Peace River, mais elle y a disparu au cours des années 1950 (Salt et Wilk, 1958; Prescott et Bjorge, 1999). À l’heure actuelle, le cœur de son aire de répartition en Alberta correspond à la moitié septentrionale des prairies vers l’est, de Hanna et de Brooks (Bjorge et Kiliaan, 1997) jusque dans le sud de la région de la tremblaie-parc à l’est de Stettler (Kiliaan et Prescott, 2002; Prescott, 2013). Depuis 1993, les populations sont en déclin, et l’aire de reproduction de la sous-espèce s’est contractée (Prescott, 2009, 2013).
Les mêmes tendances ont été observées en Saskatchewan (voir Fluctuations et tendances). Les populations ont chuté, et l’aire de reproduction s’est contractée vers le sud de la province (A. Didiuk, comm. pers., 2012). La sous-espèce est actuellement largement répartie dans les régions de la tremblaie-parc et des Prairies, mais elle ne niche plus dans la plupart des régions du centre de la Saskatchewan (régions de Meadow Lake, Nipawin et Somme; Smith, 1996). Par suite de la disparition de nombreuses populations locales dans le sud-est de la province au cours des dix dernières années, la sous-espèce est aujourd’hui confinée au sud de la Saskatchewan, où elle présente une répartition irrégulière (A. Didiuk, comm. pers., 2012).
Historiquement, les aires de reproduction des sous-espèces L. l. excubitorides et L. l. migrans se chevauchaient dans certains secteurs de l’est du Manitoba, et les deux sous-espèces s’hybridaient (Miller, 1931; Vallianatos et al., 2001). Aujourd’hui, la Pie-grièche migratrice ne s’observe plus régulièrement que dans le sud-ouest de la province et n’y est plus représentée que par la sous-espèce L. l. excubitorides. Des relevés annuels ciblés menés au cours des dix dernières années dans d’anciens sites de nidification indiquent que l’aire de répartition de l’espèce s’est contractée vers le sud et l’ouest de la province (K. DeSmet, comm. pers., 2012). L’espèce ne se reproduit pour ainsi dire plus dans le sud-est du Manitoba.
Anciennement, la Pie-grièche migratrice se reproduisait sporadiquement dans les Maritimes, mais aucune activité reproductrice n’y a été signalée depuis 1972 (Erskine, 1992). La sous-espèce de l’Est est aujourd’hui confinée au sud de l’Ontario et à l’extrême sud-ouest du Québec (Figure 4).
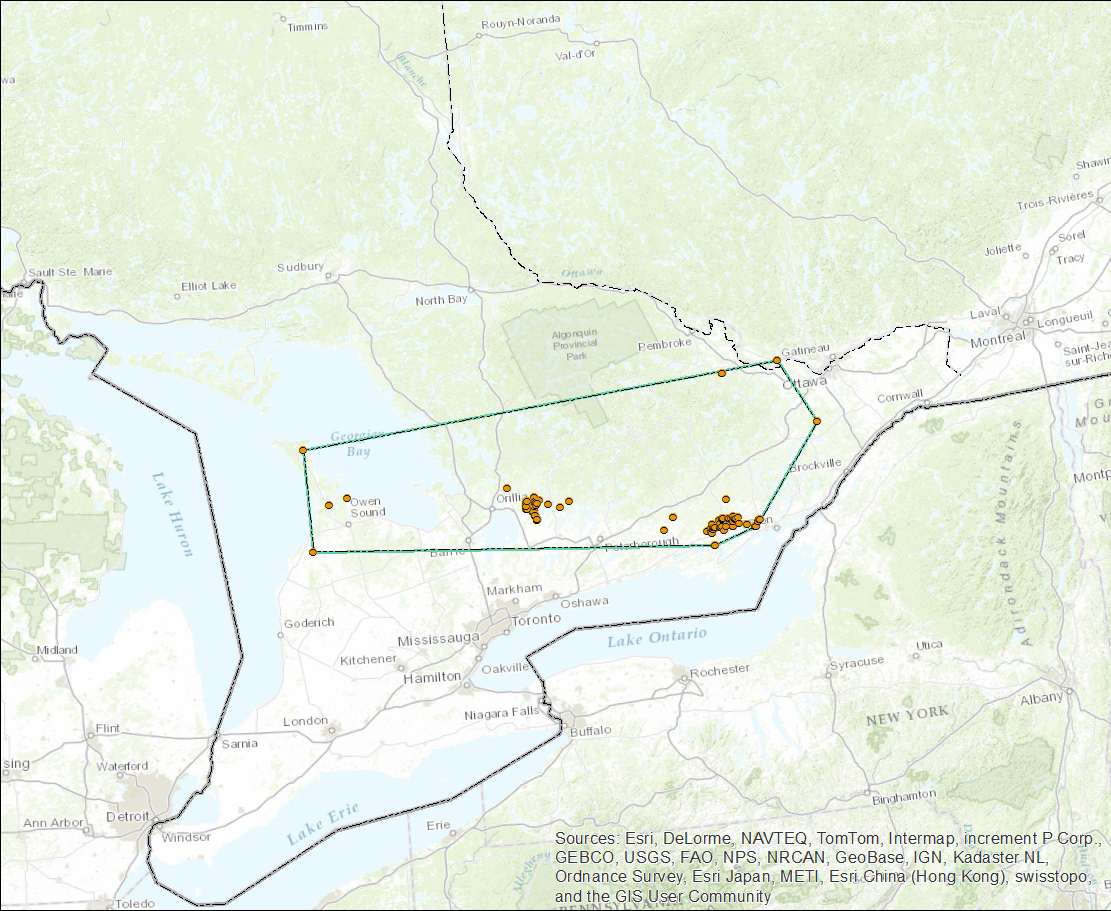
Carte : © Environnement Canada
Description longue pour la figure 4
Carte de la région des Grands Lacs indiquant l’aire de reproduction canadienne de la Pie grièche de la sous espèce de l’Est (L. ludovicianus spp.) en Ontario et dans l’extrême sud ouest du Québec (voir le texte pour de plus amples renseignements). L’aire de reproduction est estimée selon la méthode du polygone convexe minimum, qui englobe les sites où la sous espèce a été observée.
Entre 1981 et 1985, durant les travaux du premier Atlas des oiseaux nicheurs de l’Ontario, la Pie-grièche migratrice a été observée dans 145 carrés de 10 km de côté (Cadman et al., 1987). Plusieurs de ces carrés auraient abrité entre 2 et 10 couples nicheurs. À la fin des années 1980, les données amassées dans le cadre de l’Ontario Rare Breeding Bird Program et, subséquemment, de l’Ontario Bird’s at Risk Program, ont révélé que l’espèce avait pratiquement disparu de la province et était désormais confinée aux trois principales zones de reproduction dans les régions des plaines calcaires de Carden, Napanee et Smiths Falls. Durant les travaux du deuxième Atlas des oiseaux nicheurs (2001-2005), l’espèce a été observée dans seulement 38 carrés de 10 km de côté (Cadman et al., 2007). L’espèce est aujourd’hui observée uniquement de façon sporadique dans la région de Smiths Falls, et moins de 10 couples nicheurs fréquentent encore annuellement la région de Napanee (Imlay et Lapierre, 2012). Même si ses effectifs y sont restreints (au plus 20 couples nicheurs), l’espèce est encore observée régulièrement dans la plaine calcaire de Carden (Imlay et Lapierre, 2012).
La Pie-grièche migratrice était autrefois un nicheur relativement commun au Québec au début des années 1900, mais elle était devenue rare à la fin des années 1970. Durant les travaux du premier Atlas des oiseaux nicheurs au Québec, entre 1984 et 1989, l’espèce a été observée dans 30 carrés de 10 km de côté dans la portion sud-ouest de la province. Dans le cadre des travaux du deuxième atlas, qui se poursuivent actuellement, elle a été observée à un seul site. Avant le début de ces travaux, deux couples nidifiants avaient été observés en 1992, et un autre en 1993 (Grenier et al., 1999). En 1995, des preuves de nidification ont été décelées à trois sites, un à proximité de Montréal, les deux autres dans la région de l’Outaouais, dans le sud-ouest de la province (SOS-POP, 2011; F. Shaffer, comm. pers., 2014). Aucune autre preuve de nidification n’a été découverte entre 1996 et 2009, malgré les relevés effectués aux anciens sites de nidification et la remise en liberté de 101 jeunes élevés en captivité dans la région de l’Outaouais entre 2004 et 2009 (F. Shaffer, comm. pers., 2012). En 2009, un individu élevé en captivité et relâché au Québec en 2008 a été observé nidifiant dans la région de Carden en Ontario. En 2010, un couple (dont un des membres avait été élevé en captivité et relâché en Ontario) nidifiant a été observé dans la région de l’Outaouais.
La zone d’occurrence de la sous-espèce des Prairies, calculée selon la méthode du polygone convexe minimum, est estimée à 375 100 km2. Il est actuellement impossible de calculer un indice de zone d’occupation (IZO) pour cette population selon une grille à carrés de 2 km de côté, car on ignore l’emplacement exact des sites où l’espèce niche encore. Toutefois, on peut supposer que l’indice de zone d’occupation est supérieur à 2000 km2, compte tenu de la taille de la population et du fait que les sites de nidification sont disséminés sur un vaste territoire.
La zone d’occurrence de la sous-espèce de l’Est, également calculée selon la méthode du polygone convexe minimum, est estimée à 49 310 km2. L’IZO, établi selon une grille à carrés de 2 km, est estimé à moins de 200 km2, si l’on suppose que la population compte au plus 55 couples nicheurs et que ces derniers n’occupent pas tous un carré différent, ce qui est effectivement le cas.
L’ampleur des recherches ciblant la Pie-grièche migratrice et les méthodes utilisées pour les relevés diffèrent selon les provinces. À l’échelle de l’Amérique du Nord, l’espèce fait l’objet d’un suivi dans le cadre du Relevé des oiseaux nicheurs (BBS), lequel fournit la seule mesure normalisée des tendances des populations sur un vaste territoire (voir par exemple Robbins et al., 1986; Peterjohn et Sauer, 1995). Toutefois, l’exactitude des estimations de l’abondance de la Pie-grièche migratrice fournies par le BBS est sujette à caution (Smith, 1990; Peterjohn et Sauer, 1995). La répartition généralement agrégée de l’espèce (Etterson, 2003; Chabot, données inédites; SCF, données inédites) et la faible densité des populations rendent difficile l’estimation de la taille de la population selon la méthodologie du BBS.
Divers relevés ciblant la Pie-grièche migratrice ont été menés au fil des ans en Alberta (Telfer et al., 1989; Bjorge et Prescott, 1996; Collins, 1996; Bjorge et Kiliaan, 1997; Kiliaan et Prescott, 2002; Prescott, 2003, 2004). Le relevé le plus utile pour suivre l’évolution des populations sur un vaste territoire dans l’Ouest canadien est le relevé quinquennal réalisé en bordure de route depuis 1987 en Alberta et en Saskatchewan. Des milliers de kilomètres d’habitat de bord de route sont inventoriés annuellement dans le cadre de ces relevés. Le plus récent a été effectué en 2013 (Prescott, 2013; Didiuk et al., 2014). Comme le premier relevé réalisé en 1987 a donné des estimations erronément faibles (Prescott, 2013), ses résultats ne doivent pas être inclus dans le calcul des tendances des populations en Alberta ou en Saskatchewan. En outre, ce relevé quinquennal n’a pas été réalisé en Alberta en 1993, mais il l’a été en Saskatchewan. Dès lors, on dispose de résultats comparables pour ces deux provinces seulement pour les années 1998 à 2013.
Au Manitoba, les relevés ciblés sont effectués tous les ans, et non tous les cinq ans comme en Alberta et en Saskatchewan. Ces relevés sont effectués annuellement dans le sud-ouest du Manitoba par Conservation Manitoba depuis 1987 dans le cadre d’un projet de surveillance à grande échelle des oiseaux de prairie (K. DeSmet, comm. pers., 2014). Des changements ont été apportés au début des années 1990 à la méthodologie utilisée, mais de façon générale, la façon de faire a été normalisée, les relevés visant principalement à vérifier le statut d’occupation aux sites de nidification récemment et anciennement occupés (K. DeSmet, comm. pers., 2012). L’occurrence de la Pie-grièche migratrice est également documentée dans le cadre du présent projet d’atlas des oiseaux nicheurs du Manitoba, qui couvre la période 2010 à 2014. Les recherches effectuées dans le cadre des travaux de l’atlas diffèrent des recherches ciblant l’espèce menées en 2013 (K. DeSmet, comm. pers., 2014). Cette année a donc été exclue du calcul des estimations des tendances des populations dans le présent rapport.
Le deuxième Atlas des oiseaux nicheurs de l’Ontario couvre la période 2001 à 2005 et l’ensemble du territoire ontarien. Les données obtenues permettent de comparer la probabilité d’occurrence de l’espèce au cours de la période de vingt ans écoulée depuis les travaux du premier atlas (1981 à 1985) (Cadman et al., 2007). En Ontario, le Service canadien de la faune a également supervisé la réalisation de relevés annuels ciblés au cours des vingt dernières années dans la plupart des principales zones de nidification (Imlay et Lapierre, 2012). Des recherches ciblées de moindre ampleur ont été menées dans la région de Smiths Falls et les comtés de Grey et de Bruce et sur l’île Manitoulin. La méthodologie a été normalisée au cours des dix dernières années, et les recherches visent principalement les sites récemment ou anciennement occupés (Imlay et Lapierre, 2012). Les zones d’habitat propice ont également été cartographiées et sont inventoriées dans la mesure du possible dans les principales zones de nidification.
Le Service canadien de la faune a cartographié les zones d’habitat propice au Québec à la fin des années 1990. Cette analyse a montré que la plus grande partie de l’habitat propice se trouvait dans la région de l’Outaouais, adjacente aux régions de Pembroke et de Renfrew encore périodiquement occupées par l’espèce dans l’est de l’Ontario (Jobin et al., 2005). Des relevés ont été réalisés dans la région de l’Outaouais de 2004 à 2010, mais aucun relevé ciblant l’espèce n’y a été effectué depuis. Les travaux du deuxième Atlas des oiseaux nicheurs du Québec ont débuté en 2010. Cette année-là, un couple a été observé (Chabot, 2011b), et un individu isolé a été aperçu au même endroit l’année suivante (F. Shaffer, comm. pers., 2012).
La Pie-grièche migratrice se rencontre dans divers types de prairies et d’arbustaies. Les territoires de nidification contiennent normalement les éléments suivants :
- des supports de nidification (petits arbres ou arbustes);
- des perchoirs dominants pour la chasse, le maintien du couple et la proclamation territoriale (poteaux de clôture, arbustes et arbres, fils électriques);
- des sites propices à l’aménagement d’une cache de nourriture (buissons épineux, fils barbelés ou arbres densément ramifiés);
- des zones propices à la quête de nourriture (espaces ouverts à couvert de graminées courtes parsemés de perchoirs et de zones dénudées).
De nombreux milieux offrent de telles caractéristiques. Les Pies-grièches se rencontrent donc dans divers types de milieux comme des pâturages (prairies cultivées ou indigènes), des champs abandonnés, des prairies, des savanes, des forêts claires de pinyon-genévrier, des steppes arbustives et des alvars (Brownell et Riley, 2000; Pruitt, 2000; Prescott, 2013).
Dans de nombreuses régions, les caractéristiques particulières des microhabitats fréquentés par l’espèce ont changé au fil des ans. Ainsi, dans la plaine de Napanee, en Ontario, l’espèce se rencontrait autrefois des milieux dominés par l’aubépine (Cratageus sp.), mais elle semble préférer aujourd’hui les milieux dominés par le genévrier de Virginie (Juniperus virginiana). Dans d’autres régions, l’espèce occupe des milieux aménagés par les humains comme des aéroports et des cimetières (Temple, 1995; K. DeSmet, comm. pers., 2012; Chabot, données inédites).
Bien que la Pie-grièche migratrice aménage souvent son nid dans des haies-clôtures, en particulier là où les autres types de milieux sont en quantité restreinte, les tentatives de nidification le long de clôtures bordées d’arbres ou d’arbustes et dans des haies-clôtures sont rarement fructueuses, les pressions de prédation par les mammifères étant plus élevées le long de ces corridors de déplacement (Yosef, 1994; Esley et Bollinger, 2001).
En Alberta, des valeurs de superficie du territoire oscillant entre 8,5 et 13,4 ha ont été enregistrées dans le cadre de deux études ciblant la sous-espèce L. l. excubitorides (Collister, 1994; Collister et Wilson, 2007a). En 2009, des valeurs de 2,7 à 47,0 ha et de 2,9 à 11,7 ha ont été observées en Ontario à Carden et à Napanee, respectivement (Glynn-Morris, 2010). Aucune différence statistiquement significative n’a été relevée entre la superficie moyenne des territoires à Carden (15,45 ± 12,99 ha, n = 12) et à Napanee (6,86 ± 3,52 ha, n = 6). La superficie des territoires était significativement plus élevée au moment du premier envol des jeunes (12,97 ± 13,90 ha) qu’à tous les autres stades de développement (1,55 ± 1,43 ha à 3,33 ± 2,49 ha) et ne semblait pas liée à la superficie des parcelles d’habitat disponibles.
Ailleurs en Amérique du Nord, la superficie des territoires varie entre 0,8 à 17,6 ha (valeurs tirées de Yosef, 1996, et de Pruitt, 2000). Il a été démontré que la superficie des territoires varie en fonction inverse de l’abondance des arbres et arbustes pouvant servir de perchoir (Miller, 1951; Yosef, 1996). Ce facteur pourrait expliquer les différences liées à la superficie moyenne des territoires relevées entre les régions. La superficie des territoires semble en outre varier durant la saison de reproduction, atteignant sa valeur maximale après que les jeunes ont quitté le nid, mais dépendent encore de leurs parents (Glynn-Morris, 2010).
La Pie-grièche migratrice aménage normalement son nid dans un petit arbre ou un arbuste (Peck et James, 1987; Yosef, 1996; Pruitt, 2000; Chabot et al., 2001a). À l’échelle de son aire de répartition, elle utilise comme support de nombreuses espèces d’arbres ou d’arbustes, mais elle montre des préférences localement. Les essences au port relativement dense lui confèrent une meilleure protection et sont donc préférées (Porter et al., 1975; Chabot et al., 2001 b; Glynn-Morris, 2010). En général, la Pie-grièche migratrice utilise l’espèce d’arbre ou d’arbuste qui, à l’échelle locale, satisfait le plus à ses exigences. La sous-espèce des Prairies niche dans des haies brise-vent de Caragena et dans d’autres arbustes tels que le cerisier de Virginie (Prunus virginiana), les saules et la shépherdie argentée (Shepherdia argentea; Collister et Wilson, 2007a; A. Didiuk, comm. pers., 2014). En Ontario, l’aubépine (Cratageus sp.) est préférée (Peck et James, 1987), quoique le genévrier de Virginie, qui a supplanté l’aubépine dans certaines régions, est maintenant plus souvent utilisé comme support de nidification (Chabot et al., 2001a).
Les préférences de la Pie-grièche migratrice en ce qui a trait à la hauteur du couvert herbacé dans les territoires de chasse sont incertaines. Elles semblent varier à l’échelle de l’aire de répartition de l’espèce et pourraient être liées à d’autres facteurs orientant le choix de l’habitat. Dans l’est des États-Unis et en Ontario, l’espèce semble préférer les milieux à graminées relativement courtes, peut-être parce qu’elle y trouve plus facilement sa nourriture (Gawlik et Bildstein, 1993; Chabot et al., 2001 b; Glynn-Morris, 2010) ou dépense moins d’énergie pour ce faire (Yosef et Grubb, 1993). Dans l’ouest du Canada, Prescott et Collister (1993) ont constaté que les individus qui nichent en Alberta préféraient les terrains arides à graminées moyennes (15 – 35 cm) à hautes (> 35 cm). La structure du couvert herbacé (homogène ou hétérogène) est importante, la préférence étant accordée aux terrains à couvert herbacé hétérogène parsemés de zones dénudées (Michaels et Cully, 1998). Anciennement, le broutage ou les feux de friche contribuaient au maintien des zones d’habitat dégagé et de la structure hétérogène préférées par l’espèce (Cuddy, 1995).
Dans de nombreuses régions, l’occurrence de l’espèce est liée à la proportion d’habitat propice disponible à l’échelle du paysage (Brooks et Temple, 1990a; Bjorge et Prescott, 1996; Cuddy et Leviton, 1996; Yosef, 1996; Chabot et al., 2001a; Glynn-Morris, 2010), les domaines vitaux ou territoires représentant habituellement seulement une proportion de l’habitat propice disponible (Glynn-Morris, 2010), ce qui pourrait expliquer la répartition de l’espèce en colonies lâches. Selon Cuddy et Leviton (1996), une proportion minimale d’habitat propice à l’échelle du paysage pourrait être requise pour que l’espèce occupe une région en particulier.
Bjorge et Prescott (1996) ont noté que la densité des individus reproducteurs dans le sud-est de l’Alberta était corrélée positivement à la densité des arbres et arbustes et à la présence de cours de ferme, de haies brise-vent et d’emprises. Les principales zones de nidification en Ontario se trouvent dans des plaines calcaires où le broutage par le bétail des pâturages non cultivés constitue la principale utilisation des terres.
En Ontario, une analyse à l’échelle du paysage de divers attributs des parcelles d’habitat a été réalisée dans chaque principale zone de nidification dans les régions de Carden, Napanee et Smiths Falls à partir des données d’occurrence amassées de 1991 à 2010 (Chabot et Lagois, 2012). L’analyse a permis d’évaluer l’impact des attributs dans un rayon de 0,5, 5 et 15 km des sites, valeurs qui correspondent de façon générale à la taille des territoires et à la distance de dispersion des individus nicheurs retournant sur les sites de nidification et des jeunes, respectivement. Une analyse de variables liées aux parcelles, aux classes et au paysage a mis en évidence des différences statistiquement significatives pour la plupart des variables entre les milieux occupés, les milieux propices mais inoccupés, et les parcelles d’habitat occupées choisies au hasard (Chabot et Lagois, 2012). Ces résultats donnent à croire que les effets à l’échelle du paysage ont une incidence déterminante sur la qualité de l’habitat pour l’espèce.
En général, les besoins de l’espèce en matière d’habitat d’hivernage ne semblent pas différer de façon notable des besoins liés à l’habitat de reproduction (Bartgis, 1992; Collins, 1996; Yosef, 1996; Chabot, données inédites). Dans la portion méridionale de l’aire de répartition de l’espèce en Amérique du Nord, les mêmes individus peuvent demeurer sur leur territoire à l’année (Miller, 1931; Chabot, données inédites), mais l’utilisation de l’habitat peut changer à l’intérieur du territoire. Ainsi, la superficie du territoire peut augmenter en hiver (Blumton, 1989; Collins, 1996), et certains types de milieux, notamment les terrains boisés, peuvent être utilisés plus fréquemment en hiver que durant la saison de reproduction (Blumton, 1989; Bartgis, 1992; Gawlik et Bildstein, 1993; Chabot, données inédites).
En raison des difficultés que soulève l’identification des différentes sous-espèces sur le terrain, on sait peu de choses sur la compétition qui oppose les individus migrateurs et les individus résidents dans les aires d’hivernage. Selon certains, les résidents pourraient surclasser les individus migrateurs en hiver et les refouler vers des milieux de qualité suboptimale. Perez et Hobson (2009) ont toutefois constaté que les individus migrateurs et les résidents permanents exploitent des milieux différents au Mexique. Le long de la côte du golfe du Mexique, au Texas, cette partition semble fondée sur les classes d’âge, les individus plus âgés occupant les milieux côtiers et les individus plus jeunes, les milieux plus à l’intérieur des terres (Craig et Chabot, 2012).
On en sait très peu sur les besoins en matière d’habitat de l’espèce en période de migration (Yosef, 1996). Toutefois, si l’on considère la plasticité dont l’espèce fait preuve à l’égard de ses habitats de reproduction et d’hivernage, on peut supposer qu’elle se montre tout aussi polyvalente à l’égard de son habitat de migration (Yosef, 1996).
L’habitat de reproduction a décliné et continue de décliner à l’échelle de l’aire de répartition de l’espèce (voir par exemple Telfer, 1992; Yosef, 1996; Cade et Woods, 1997). La plupart des spécialistes conviennent toutefois qu’une bonne partie de l’habitat apparemment propice est inoccupée (Brooks et Temple, 1990a; K. DeSmet, comm. pers., 2012; A. Didiuk, comm. pers., 2012; A. Chabot, obs. pers.; SCF, données inédites). Bien qu’ils se soient dits d’avis que la population de Pies-grièches migratrices en Alberta est limitée par la disponibilité de l’habitat de qualité optimale, Prescott et Collister (1993) n’ont pas écarté la possibilité que d’autres facteurs aient également un rôle limitatif. En Ontario, une analyse de la disponibilité de l’habitat a indiqué que la quantité d’habitat était suffisante pour supporter une population de 500 couples (Cuddy et Leviton, 1996), alors que le nombre de couples nicheurs était estimé à moins de 50 couples à l’époque. Selon une analyse similaire effectuée au Québec (Jobin et al. 2005), la quantité d’habitat de qualité dans la région de l’Outaouais devrait être suffisante pour assurer le maintien d’une population viable, mais la réalité est toute autre. La majorité des études portant sur la disponibilité de l’habitat menées à ce jour n’ont pas tenu compte de l’impact de la fragmentation de l’habitat ou d’autres effets influant à l’échelle du paysage sur la disponibilité de l’habitat.
Dans l’ouest du Canada, la perte d’habitat de reproduction semble causée principalement par la conversion des prairies en terres agricoles et, vers le nord, dans les portions périphériques de l’aire de reproduction de l’espèce, par la régénération forestière (Cadman, 1985). Dans les Prairies canadiennes, la superficie des pâturages non cultivés a décliné de 39 % entre 1946 et 1986 dans les régions où les déclins de populations les plus abrupts ont été observés, mais de seulement 12 % dans les régions où un nombre substantiel d’individus étaient encore présents (Telfer, 1992). Bien que les pâturages occupent encore une vaste superficie au Manitoba, une bonne partie de ces pâturages sont cultivés et sont donc dépourvus d’arbres susceptibles d’être utilisés comme supports de nidification ou perchoirs.
Dans l’est du Canada, la succession naturelle résultant de l’abandon des pâturages a été la principale cause de perte d’habitat au cours des dernières décennies (Cadman et al., 2007). La superficie des pâturages en Ontario a chuté d’environ 2,7 millions d’hectares en 1921 à environ 660 000 hectares en 2011, soit un déclin de 75 %. Cette tendance se poursuit, puisqu’un déclin de 12,3 % a été enregistré de 2006 à 2011. Dans la région de Smiths Falls, les pertes d’habitat ont souvent été causées par des projets domiciliaires en milieu rural, mais une partie des pertes est attribuable à des projets de reboisement (D. Cuddy, comm. pers., 2012). L’extraction d’agrégats a également eu un impact local en Ontario, en particulier dans la région de Carden (Cadman, 1990). L’aménagement de « fermes solaires » menace de plus en plus l’habitat de l’espèce dans les régions de Napanee et de Carden.
Avant l’arrivée des premiers colons européens dans l’est de l’Amérique du Nord, les oiseaux de prairie, y compris la Pie-grièche migratrice, nichaient dans des milieux prairiaux indigènes tels que des prairies, des savanes, des prairies de castor, des alvars et des zones balayées par le feu ou déboisées à des fins agricoles par les Premières Nations (Askins et al., 2007; Catling, 2008). La colonisation européenne a occasionné la disparition de bon nombre de ces milieux (Askins et al., 2007). À l’échelle de l’Amérique du Nord, la superficie actuelle des prairies nordiques à grandes graminées est estimée à seulement 2,4 % de la superficie originale (Samson et al., 2004); en Ontario, ce pourcentage est inférieur à 1 % (Bakowsky et Riley, 1994; Catling et Brownell, 1999; Catling, 2008). Bien qu’il soit difficile d’estimer de façon précise les pertes d’habitat d’alvar et de savane en Ontario, il est clairement établi qu’une importante portion de ces types d’habitat a disparu et que l’agriculture et d’autres activités humaines ont entraîné la dégradation de la majeure partie des parcelles existantes occupées par ces types de milieux (Reschke et al., 1999; Brownell et Riley, 2000).
Au Québec, la superficie des pâturages a chuté de 85 % entre 1941 et 1990 (Cadman, 1990). Une analyse comparative de photographies aériennes prises entre les années 1960 et 1980 dans le sud du Québec a permis d’évaluer les changements survenus à l’échelle du paysage rural dans les basses-terres du Saint-Laurent (Jobin et al., 1996). Ces changements résultent principalement de l’abandon des fermes peu productives et de la régénération forestière qui s’est ensuivie, de plantations, de l’étalement urbain et de l’essor de l’agriculture intensive. Dans le cadre d’une étude plus récente, Jobin et al. (2010) ont examiné des images Landsat couvrant les écorégions des basses-terres du Saint-Laurent et des Appalaches du sud du Québec prises en 1993 et en 2001. Un changement des principales classes agricoles a été noté dans les basses-terres du Saint-Laurent, les cultures fourragères pérennes étant converties en cultures annuelles et la superficie des paysages dominés par l’agriculture intensive augmentant (Jobin et al., 2010). Ces changements se sont vraisemblablement traduits par une perte d’habitat propice à la reproduction de la Pie-grièche migratrice.
La perte d’habitat a également contribué au déclin des populations dans le nord des États-Unis. Par exemple, 70 % de l’habitat de steppe à armoise a été converti en terres agricoles en Idaho, l’exploitation de mines à ciel ouvert a détruit une bonne partie de l’habitat propice à l’espèce en Indiana, et l’abandon des exploitations agricoles moins productives et la régénération forestière qui s’en est ensuivie ont entraîné un déclin de 74 % à 30 % de la superficie des terres agricoles dans l’État de New York entre 1900 et 1982 (Yosef, 1996).
Les habitats de migration et d’hivernage ont également subi un déclin (Telfer, 1992; Yosef, 1996; Cade et Woods, 1997). Étant donné l’ampleur des pertes d’habitat de prairie survenues à l’échelle de l’Amérique du Nord, on peut supposer que ces pertes ont eu un impact négatif important sur les populations des sous-espèces des Prairies et de l’Est dans leur aire d’hivernage.
Les Pies-grièches migratrices commencent à arriver sur les lieux de reproduction à la fin de mars ou au début d’avril et entreprennent leur migration vers le sud en septembre. Elles passent donc environ cinq mois de l’année au Canada.
Selon la plupart des études, la Pie-grièche migratrice serait une espèce monogame (Yosef, 1996). Des analyses de l’ADN nucléaire d’oisillons effectuées en Ontario font toutefois état d’une faible occurrence d’accouplements extra-conjugaux et, plus fréquemment, de cas de maternité multiple (p. ex. parasitisme de couvée) dans des nids individuels (Chabot et Lougheed, 2005). Dans le cadre de travaux de baguage, plusieurs femelles ont été observées à un même site de nidification, et des aides parentaux des deux sexes ont été observés autour de nids en Ontario (Chabot, 2009, 2010, 2011 b). Etterson (2004) a toutefois noté que seulement 4 % des jeunes ou 14 % des couvées étaient issues d’accouplements extra-conjugaux en Oklahoma, et il n’y a observé aucun cas de maternité multiple. La polygynie (situation où un mâle s’accouple avec plusieurs femelles) a cependant déjà été observée chez la Pie-grièche migratrice (Yosef, 1992).
En général, les Pies-grièches migratrices se reproduisent pour la première fois à l’âge de un an, au cours du premier printemps suivant l’année de leur naissance (Miller, 1931). L’espèce ne produit généralement qu’une seule couvée, mais elle peut en produire une deuxième si la première tentative de nidification a échoué (Miller, 1931; Yosef, 1996). Les doubles couvées sont occasionnelles sous nos latitudes, mais plus fréquentes plus au sud (Yosef, 1996).
Les deux sexes participent à la sélection du site de nidification et à la construction du nid (A. Chabot, obs. pers.). Le nid est une coupe ouverte faite de brindilles et habituellement tapissée de plumes ou de poils. La taille des couvées augmente en fonction de la latitude et est généralement plus élevée chez les populations de l’ouest du continent (Yosef, 1996). Les œufs, en moyenne au nombre de 5 ou 6 (Yosef, 1996; Chabot et al., 2001a; Collister et Wilson, 2007 b), sont couvés durant 15 à 17 jours (Lohrer, 1974; Collister et Wilson, 2007 b). Les jeunes demeurent au nid durant 16 à 20 jours (Yosef, 1996; Collister et Wilson, 2007 b). Les deux parents continuent de les nourrir durant un mois une fois qu’ils ont quitté le nid, mais ils utilisent souvent des portions différentes de leur territoire. Les jeunes s’initient à l’empalage à l’âge de 20 à 25 jours et maîtrisent déjà la technique à l’âge de 35 jours (Smith, 1972).
Le succès de reproduction (au moins un jeune atteignant l’âge de l’envol) fluctue considérablement d’une année et d’une région à l’autre (Pruitt, 2000). Un taux moyen de 56 % a été calculé d’après les résultats d’un certain nombre d’études (passées en revue par Yosef, 1996), mais des taux plus faibles ont été enregistrés dans le cadre d’études plus récentes. Par exemple, dans le sud-est de l’Alberta, Collister et Wilson (2007b) ont observé un taux de survie journalier moyen des nids de 0,973 (IC 95 % : 0,967, 0,978) qui, étendu à toute la durée du cycle de nidification, donne un taux de reproduction estimé de 35 % (IC 95 % : 28 – 43 %; S. Wilson, comm. pers., 2014). D’autres estimations, calculées à l’aide de méthodes d’analyse fondées sur les jours d’exposition, s’établissent à 40 % en Oregon (Nur et al., 2004), à 43 % en Oklahoma (Etterson et al., 2007) et à seulement 26 % en Illinois (Walk et al., 2006).
Les nids sont très rarement parasités par le Vacher à tête brune (DeGues et Best, 1991), mais la prédation peut réduire considérablement la productivité des couvées, notamment au Manitoba (K. DeSmet, comm. pers., 2012), en Ontario (SCF, données inédites) et en Illinois (voir par exemple Walk et al., 2006), trois régions qui abritent de petites populations de Pies-grièches migratrices.
La mortalité post-envol des jeunes, généralement attribuable à la prédation, est apparemment élevée et oscille entre 33 et 46 % au cours des premiers 7 à 10 jours suivant le premier envol (Yosef, 1996; Chabot et al., 2001a). D’autres études (voir par exemple Blumton, 1989) font toutefois état de taux de survie post-envol relativement élevés. En Ontario, un taux de survie jusqu’à la migration automnale relativement élevé a été observé dans le cadre d’un suivi radio-télémétrique de jeunes plus âgés ayant acquis leur indépendance (Imlay et al., 2010).
En nature, le record de longévité pour l’espèce est de 12 ans, mais en captivité, les oiseaux peuvent vivre jusqu’à 15 ans (T. Imlay, comm. pers., 2012). D’après des données de baguage amassées en Ontario, la durée de vie moyenne de l’espèce en nature oscille entre 2 et 4 ans. Une estimation de l’intervalle de survie des adultes de 0,5 à 0,7, calculée à l’aide de l’équation α + [S/(1-S)], où α correspond à l’âge moyen des femelles au moment de la première reproduction et S est le taux de survie adulte, donne une durée de génération de 2 à 3,3 ans. Une durée de génération de 3 ans est utilisée aux fins de la présente évaluation.
La structure par âge des populations n’a pas été étudiée récemment dans les Prairies, mais elle l’a été en Ontario et en Illinois dans le cadre d’études de baguage (Chabot, 2009, 2010, 2011 b). En Ontario, la majorité des individus reproducteurs sont des oiseaux qui ont plus de deux années civiles (After Second Year ou ASY en anglais). Bien que le rapport du nombre d’oiseaux qui en sont à leur deuxième année civile (Second Year ou SY en anglais) au nombre d’ASY fluctue considérablement d’une année à l’autre, le nombre de femelles SY observées annuellement en Illinois et en Ontario est habituellement supérieur au nombre de mâles. Comme le succès de reproduction est comparable d’une région à l’autre (Chabot, 2011b), la mortalité hivernale chez les oiseaux écoulant leur première année de vie est probablement un facteur déterminant de la structure par âge des populations.
La Pie-grièche migratrice est un prédateur opportuniste qui s’adapte à la base de proies disponibles (Miller, 1931; Craig, 1978; Scott et Morrison, 1995). Les insectes forment la principale composante de son alimentation durant la saison de reproduction, mais la contribution des vertébrés augmente en hiver. Le comportement de l’espèce d’aménager des caches de nourriture est considéré comme un système d’entreposage de la nourriture et contribue probablement au succès de la reproduction, en particulier lorsque les conditions météorologiques sont défavorables (Yosef, 1996). Toutefois, une réduction des couvées vraisemblablement imputable à une pénurie de nourriture a été observée durant des épisodes de conditions météorologiques défavorables (A. Chabot, obs. pers.; K. DeSmet, comm. pers., 2012).
Les Pies-grièches migratrices tolèrent relativement bien les activités humaines autour des sites de nidification (Luukkonen, 1987; Brooks, 1988; Bartgis, 1989), mais les données sont contradictoires en ce qui a trait à la fréquence à laquelle elles abandonnent leur nid en réaction à des perturbations anthropiques (Porter et al., 1975; Siegel, 1980). Elles semblent toutefois moins dérangées par les perturbations mécaniques telles que celles occasionnées par les tracteurs que par la présence d’humains (A. Chabot, obs. pers.).
Les pies-grièches qui nichaient dans l’est du Manitoba au début des années 1990 étaient souvent associées à des milieux anthropiques (p. ex. cimetières, aéroports, quartiers résidentiels en milieu rural). L’espèce niche d’ailleurs dans des milieux similaires dans d’autres portions de son aire de reproduction (Chabot, données inédites). Même si cela donne à croire que l’espèce est capable de s’adapter avec succès à ces types de milieux, ça ne semble pas être le cas à l’échelle de son aire de répartition (Jones et Bock, 2002). Une seule étude a été consacrée à l’évaluation du succès reproducteur de l’espèce en milieu urbain (Boal et al., 2003). Cette étude révèle que, même si le succès reproducteur en milieu urbain était similaire à celui observé en milieu rural, les proportions de zones résidentielles et commerciales et de zones dégagées à couvert herbacé bas dans les territoires de nidification étaient respectivement moins élevées et plus élevées que les proportions de ces mêmes milieux à l’échelle du paysage (Boal et al., 2003). En outre, l’utilisation du milieu urbain est peut-être associée à une perte concurrente de l’habitat naturel (Boal et al., 2003). En conséquence, on ignore si la capacité de l’espèce de s’adapter au changement de milieu et à la perte d’habitat contribue à la viabilité de l’espèce à long terme.
L’espèce est migratrice dans la moitié septentrionale de son aire de répartition, qui inclut la totalité de l’aire de répartition canadienne. Plus au sud, les déplacements sont irréguliers et variables au sein de l’espèce (Miller, 1931). Aux États-Unis, certains individus appartenant à des populations essentiellement migratrices demeurent tout l’hiver sur leur site de reproduction (Figure 3).
Les résultats d’une analyse d’autocorrélation spatiale de la distance génétique versus géographique entre individus donnent à croire qu’un flux génétique (résultat d’une dispersion fructueuse menant à la reproduction) se produit sur de grandes distances. Une structure génétique spatiale positive (similarité génétique entre individus) s’observe sur une distance pouvant atteindre 500 km chez cette espèce (Chabot, 2011a). Les résultats de l’analyse d’autocorrélation spatiale semblent témoigner de l’existence d’un isolement par la distance (Wright, 1943), où le flux génétique (ou dispersion) est plus élevé sur de courtes distances et s’atténue graduellement en fonction de la distance (Figure 5). Les femelles et les jeunes qui ne se sont pas encore reproduits se dispersent sur des distances plus grandes que les mâles ou les reproducteurs ASY (Chabot, 2011a). Les individus migrateurs de toutes les cohortes se dispersent sur des distances significativement plus grandes que les individus résidents non migrateurs. Des estimations du flux génétique indiquent que la dispersion s’effectue plus souvent dans l’axe nord-sud plutôt qu’est-ouest (Figure 5; Chabot, 2011a). Le flux génétique est toutefois réduit dans les régions où l’espèce réside à l’année et à la limite nord de l’aire de répartition de l’espèce, c’est-à-dire dans l’Ouest canadien et en Ontario.
Cartes et Graphique : © Environnement Canada
Description longue pour la figure 5
Cinq cartes représentant de façon géographique les estimations du degré de parenté génétique pour chaque groupe génétique de Pies grièches migratrices échantillonné par Chabot (2011a). Ces graphiques sont intitulés « sous espèce excubitorides », « sous espèce du nord est », « sous espèce migrans », « sous espèce ludovicianus » et « sous espèce mexicanus/gambeli ». Les zones échantillonnées sont indiquées. Le taux d’admixture génétique est mentionné dans un rayon de 150 km de chaque zone. Le taux d’admixture génétique semble diminuer en fonction de la distance entre les zones d’échantillonnage et être plus élevé sur l’axe nord sud que l’axe est ouest.
Le taux de retour apparent des jeunes au lieu de naissance est faible. Collister et DeSmet (1997) ont observé 1,2 % des juvéniles en Alberta et 0,8 % des juvéniles au Manitoba au cours de l’année suivant leur baguage. En Ontario, le taux de retour des jeunes a oscillé entre 3,1 % et 12 % (Okines et McCracken, 2003, 2005). Aux États-Unis, les taux de retour des jeunes sont également faibles : 3,6 % en Virginie (Luukkonen, 1987), 2,4 % en Indiana (Burton et Whitehead, 1990), 1,7 % en Virginie (Blumton, 1989), 1,1 % au Missouri (Kridelbaugh, 1982), 0,8 % au Dakota du Nord, et 0,0 % au Minnesota (Brooks et Temple, 1990 b).
En Alberta, des juvéniles se sont dispersés sur une distance moyenne de 12,4 km et jusqu’à 70 km de leur lieu de naissance (Collister, 1994). Au Manitoba, la dispersion natale moyenne s’établissait à 15,4 km. En Ontario, les jeunes sont retournés de façon générale à moins de 44 km de leur lieu de naissance, mais certains se sont dispersés sur des distances plus grandes (150 km et plus) (Okines et McCracken, 2003; Chabot, 2011 b). Les jeunes se sont dispersés en moyenne sur des distances de 9,9 à 15,0 km en Ontario (Okines et McCracken, 2003). Les mesures des distances de dispersion fondées sur des études de baguage varient en fonction de l’intensité des recherches et des distances sur lesquelles elles ont été effectuées et ne devraient donc pas être interprétées au pied de la lettre.
Dans le cadre d’une étude réalisée en Ontario, les taux de retour des adultes bagués ont oscillé entre 11 % et 28 % (Okines et McCracken, 2003, 2005). En Alberta, 32 % des adultes bagués ont été observés de nouveau aux mêmes sites l’année suivante; au Manitoba, ce pourcentage s’établissait à seulement 16 % (Collister et DeSmet, 1997; Collister, 2013). Ailleurs, les taux de retour diffèrent selon les régions : 47 % au Missouri (Kridelbaugh, 1982), 41 % en Indiana (Burton et Whitehead, 2002), 30 % en Idaho (Woods, 1994) et 14 % au Dakota du Nord (Haas et Sloane, 1989). Comme les taux de retour reflètent les taux de survie apparente des adultes, Collister (2013) s’est dit d’avis que les taux susmentionnés chez la Pie-grièche migratrice étaient généralement plus faibles que ceux observés chez d’autres espèces d’oiseaux chanteurs et, dès lors, que la survie des adultes est relativement faible dans de nombreuses régions.
Au Dakota du Nord, des taux de fidélité au site natal de 28 % ont été observés chez les mâles adultes, et de 5 % chez les femelles (Haas et Sloane, 1989). Dans le sud de l’Idaho, 30 % de tous les adultes bagués ont regagné le territoire qu’ils occupaient l’année précédente (Woods, 1994). Au Missouri, 47 % des mâles et 0 % des femelles ont regagné le site de nidification qu’ils occupaient l’année précédente.
Collister et DeSmet (1997) ont noté que les adultes revenaient en moyenne à 1,9 km du site de nidification occupé l’année précédente en Alberta et à 3,1 km au Manitoba. Selon ces auteurs, 95 % des adultes reviendraient en moyenne à 4,7 km du site de nidification occupé l’année précédente. La distance de dispersion était plus élevée chez les femelles, tant en Alberta qu’au Manitoba. En Ontario, les adultes sont retournés à 47 km ou moins de l’endroit où ils avaient été bagués (moyenne de 18,6 km; Okines et McCracken, 2003).
À l’échelle de l’aire de répartition de l’espèce, les taux de réutilisation des territoires et des parcelles d’habitat sont plus élevés que les taux de fidélité au site de nidification (Yosef, 1996; Pruitt, 2000). En Ontario, les taux de réutilisation du site et de fidélité au site semblent corrélés positivement au succès de la reproduction (Glynn-Morris, 2010). La fidélité au site de nidification semblait également corrélée positivement au succès de reproduction au Manitoba (Collister et DeSmet, 1997).
Les pies-grièches interagissent avec de nombreuses autres espèces d’oiseaux, probablement pour défendre leur aire d’alimentation (Cadman, 1985; Smith, 1991; Collister, 1994; Woods, 1994; Yosef, 1996). Des Moqueurs chats (Mimus polyglottos), des Chevêches des terriers (Athene cunicularia; Yosef 1996) et des Crécerelles d’Amérique (Falco sparverius) ont été observés en train de dérober de la nourriture dans des caches de pies-grièches (cleptoparasitisme) (A. Chabot, obs. pers.). Les Moqueurs chats et les Moqueurs roux (Toxostoma rufum) semblent capables de forcer les pies-grièches à abandonner leur territoire et leur site de nidification, même durant la période d’incubation (Chabot, données inédites). Des jeunes pies-grièches ayant atteint l’âge de l’envol ont été attaquées ou harcelées par un certain nombre d’espèces de passereaux (Smith 1991). Bien que la compétition interspécifique avec des espèces telles que la Crécerelle d’Amérique, l’Étourneau sansonnet (Sturnus vulgaris) et la fourmi de feu rouge (Solenipsis invicta) semble influer sur la démographie des populations locales (Cadman, 1985; Lymn et Temple, 1991; A. Chabot, obs. pers.), rien n’indique qu’elle ait contribué aux déclins des populations de la Pie-grièche migratrice à l’échelle de l’aire de répartition de l’espèce.
Une description détaillée des activités et méthodes d’échantillonnage est présentée à la section Activités de recherche. De tous les relevés fournissant des données sur l’espèce à l’échelle de son aire de répartition, c’est le Relevé des oiseaux nicheurs (BBS) qui couvre la période la plus longue. Le BBS comporte toutefois plusieurs limites, et la fiabilité des données régionales sur l’espèce est jugée faible à modérée au Canada (Environnement Canada, 2013; A. Didiuk, comm. pers., 2014). Les taux de détection de l’espèce dans le cadre du BBS sont faibles, car l’espèce est répartie en populations très localisées et est difficile à détecter dans le cadre de brèves observations en bordure de route en raison de son comportement. Les relevés ciblés en bordure de route visant à optimiser la détection de l’espèce fournissent des estimations plus fiables de la taille et des tendances des populations régionales que le BBS (A. Didiuk, comm. pers., 2014). La méthodologie des relevés est normalisée en Alberta et Saskatchewan. Les relevés ciblant l’espèce y sont réalisés selon la même méthodologie tous les cinq ans depuis 1987, le plus récent ayant été effectué en 2013. Les résultats des premiers relevés effectués en Alberta et en Saskatchewan en 1987 ne devraient toutefois pas être inclus dans l’analyse des tendances en raison de l’intensité relativement faible des échantillonnages réalisés cette année-là et de l’inexpérience des observateurs (Prescott, 2013; A. Didiuk, comm. pers., 2014). Au Manitoba, en Ontario et au Québec, l’objectif principal des relevés ciblés annuels consistait à vérifier la présence de l’espèce dans les sites de reproduction historiques et, dans une moindre mesure, dans les parcelles d’habitat apparemment propice.
D’après les données du BBS, on estime que la population mondiale de pie-grièche compte entre 3,7 et 4,2 millions d’individus.
La population de la sous-espèce des Prairies compte environ 54 700 individus matures répartis comme suit : près de 15 000 en Alberta, 39 600 en Saskatchewan et entre 100 et 200 au Manitoba.
Par extrapolation des observations effectuées en bordure de route et en tenant compte de la détectabilité de l’espèce, on estime à 7 508 couples reproducteurs (environ 15 000 individus matures) la taille de la population totale en Alberta en 2013 (Prescott, 2013). L’abondance de l’espèce varie d’une région à l’autre, les effectifs estimés les plus élevés étant observés, par ordre décroissant d’abondance, dans le centre-est, puis dans le sud-est de la province. Aucun individu n’a été observé dans le quadrant sud-ouest de la province.
La Pie-grièche migratrice est plus abondante en Saskatchewan qu’en Alberta. Si l’on se fonde sur la plus récente estimation de la taille de la population réalisée selon les mêmes techniques que celles utilisées en Alberta, la population de la Saskatchewan comptait environ 39 600 individus en 2013 (Didiuk et al., 2014).
Au Manitoba, des dénombrements ciblant l’espèce sont effectués annuellement depuis 1987. En date de 2012, la population y était estimée à environ 50 couples (100 individus matures), mais tous les individus n’ont probablement pas été détectés. Dans le cadre des travaux en cours du projet d’atlas provincial des oiseaux nicheurs, des preuves de nidification ont été observées dans 36 carrés à côté de 10 km jusqu’à la fin de la saison de nidification de 2013.
En Ontario, un programme annuel de « réintroduction » d’oiseaux élevés en captivité dans la province est en cours depuis 2001. Ce programme a connu un certain succès, si l’on se fie au nombre d’oiseaux élevés en captivité qui se sont accouplés avec des individus sauvages (Imlay et Lapierre, 2012). Entre 2004 et 2013, 698 oiseaux élevés en captivité ont été remis en liberté en Ontario, et 35 (5 %) d’entre eux sont retournés sur les lieux où ils avaient été relâchés (Wildlife Preservation Canada, comm. pers., 2014). Parmi ces 35 individus, 14 se sont reproduits avec succès (12 à Carden, 1 à Grey-Bruce et 1 au Québec), 10 n’ont pas formé de couples et 11 ont formé un couple, mais ne se sont pas reproduits avec succès.
Bien que les individus sauvages dominent largement chez la sous-espèce de l’Est (Chabot, 2009, 2010, 2011 b), le programme de réintroduction est considéré comme une mesure importante pour assurer la pérennité de la population sauvage (Tischendorf, 2009). Les individus élevés en captivité et relâchés sont inclus dans l’estimation de la taille de la population, mais selon les lignes directrices du COSEPAC applicables aux populations manipulées, ces individus ne contribuent pas de façon efficace au maintien de la population et devraient être exclus de l’estimation de la taille des populations.
En Ontario, les relevés ciblant la Pie-grièche migratrice ont débuté en 1992. Les recherches ont surtout été effectuées dans les principales zones de nidification de Smiths Falls, de Napanee et de Carden, mais aussi, dans une moindre mesure, dans la péninsule du Niagara et sur l’île Manitoulin. Environ 50 couples reproducteurs ont été recensés en 1992, mais la population a subi un déclin relativement soutenu depuis. Au cours de la période de dix ans la plus récente (2003 à 2013), le nombre connu de couples reproducteurs dans la province a oscillé entre 18 et 31 couples (SCF, données inédites, 2014). Si l’on inclut les individus non appariés, la population de l’Ontario compte probablement entre 50 et 100 adultes. Le nombre réel est probablement plus près de la borne inférieure de cette estimation (voir ci-dessous).
Jusqu’en 2003, la majorité des couples nicheurs en Ontario se reproduisaient dans la principale zone de nidification de Napanee. Le nombre de couples nicheurs a cependant décliné de façon soutenue dans cette région, pour s’établir à seulement 5 en 2012. À l’inverse, le nombre de couples nicheurs dans la plaine de Carden est passé d’environ 5 au début des années 1990 à un sommet de 18 en 2009. Cette augmentation est peut-être attribuable à la réintroduction à compter de 2003 d’oiseaux élevés en captivité dans cette région (Tischendorf, 2009). Des individus élevés en captivité sont observés chaque année parmi la population se reproduisant dans cette région, mais le gros des effectifs est encore formé d’oiseaux sauvages (Chabot, 2009, 2010, 2011 b).
Aucun individu n’a niché dans la plaine calcaire de Smiths Falls entre 2000 et 2008, mais jusqu’à trois couples y ont été observés chaque année depuis 2009 (SCF, données inédites). En dépit des lâchers d’oiseaux élevés en captivité effectués près de la baie Dyer’s sur la péninsule Bruce, l’espèce n’a été observée que sporadiquement au cours de la dernière décennie sur l’île Manitoulin et dans la principale zone de nidification de Grey-Bruce.
Entre 1980 et 1990, seulement 14 mentions de nidification ont été enregistrées au Québec. Durant la période couverte par le premier atlas des oiseaux nicheurs (1984 à 1989), seulement 30 observations avaient été enregistrées, dont sept mentions confirmées se rapportant à des couples nicheurs (Gauthier et Aubry, 1996). Lors d’un relevé étendu à toutes les zones de nidification anciennement occupées mené en 1990, un seul couple nicheur a été observé (Cadman, 1990), pour une estimation optimiste de 10 couples nicheurs (Robert et Laporte, 1991). Deux couples nicheurs ont niché au Québec en 1992, et un seul l’année suivante (Grenier et al., 1999). Des preuves de nidification ont été relevées à trois sites en 1995 (SOS-POP, 2011). Aucune autre preuve de nidification n’a été enregistrée entre 1996 et 2009, malgré les relevés effectués dans les sites de nidification anciennement occupés et en dépit du lâcher de 101 jeunes élevés en captivité dans la région de l’Outaouais entre 2004 et 2009. Un oiseau élevé en captivité relâché au Québec a été observé dans la zone de nidification principale de Carden, en Ontario. En 2010, un couple nicheur composé d’un mâle sauvage et d’une femelle élevée en captivité et relâchée dans la région de Carden en 2009 a été observé à proximité du point de lâcher. Ce couple est parvenu à élever au moins un jeune jusqu’à l’âge de l’envol, mais en 2011, un seul des deux oiseaux a été revu à ce site (F. Shaffer, comm. pers., 2012).
En bref, la taille de la population de la sous-espèce de l’Est (excluant les individus élevés en captivité) est actuellement estimée à tout au plus 110 individus matures, soit moins de 100 individus en Ontario et moins de 10 au Québec.
Le Relevé des oiseaux nicheurs (BBS) fournit des informations sur la tendance des populations à l’échelle de l’aire de répartition de l’espèce. Le déclin généralisé de l’espèce était déjà amorcé au moment du premier BBS (Cade et Woods, 1997). Les tendances estimées des populations par le BBS donnent vraisemblablement un portrait prudent des déclins survenus au cours du siècle dernier. À l’échelle de la région couverte par le BBS (États-Unis et Canada combinés), les populations de l’espèce ont décliné à un taux moyen de 3,2 % par année entre 1966 et 2011 et de 1,7 % par année entre 2001 et 2011 (voir le tableau 3, qui inclut également les intervalles de confiance associés à ces estimations).
| Région | 1987 Tableau note3 | 1993 | 1998 | 2003 | 2008 | 2013 |
|---|---|---|---|---|---|---|
| Saskatchewan | 67 048 | 116 135 | 93 617 | 86 697 | 35 582 | 39 579 |
| Alberta Tableau note2 | 5 650 | s/o | 23 428 | 16 654 | 15 442 | 15 016 |
| Manitoba | 530 | 654 | 332 | 162 | 98 | 70 Tableau note4 |
| Région | Nombre de parcours | 1970 Tableau note1.3-2011 Tendance |
1970-2011 IC à 95 % |
2001-2011 Tendance |
2001-2011 IC à 95 % |
|---|---|---|---|---|---|
| Amérique du Nord | 2011 | -3,2 | -3,6, -2,9 | -1,7 | -2,5, -0,9 |
| États-Unis | 1870 | -3,2 | -3,6, -2,9 | -1,7 | -2,5, -0,9 |
| Canada | 123 | -2,9 | -4,9, -1,3 | -1,4 | -4,6, 3,07 |
| Alberta | 40 | 0,3 | -1,7, 2,4 | 1,3 | -1,8, 8,1 |
| Saskatchewan | 49 | -3,6 | -5,5, -1,6 | -3,0 | -7,3, 4,0 |
| Manitoba | 16 | -5,1 | -9,5, -1,1 | -5,2 | -11,3, 0,2 |
| Fondrières des Prairies (Alb., Sask., Man.) | 105 | -2,8 | -4,8, -1,2 | -1,3 | -4,6, 3,1 |
| Ontario | 18 | -12,4 | -17,6, -7,9 | -12,2 | -20,8, -3,3 |
Les résultats du BBS indiquent que les populations de l’espèce aux États-Unis ont décliné en moyenne de 3,2 % par année entre 1966 et 2011 et de 1,7 % au cours de la période de 10 ans écoulée entre 2001 et 2011 (Tableau 3). À l’échelle de l’aire de répartition de l’espèce au Canada, le déclin annuel moyen de l’espèce (principalement de la sous-espèce des Prairies) s’établit à 2,9 % entre 1970 et 2011 et à 1,3 % entre 2001 et 2011. Cette dernière valeur n’est toutefois pas statistiquement significative (voir le Tableau 3). Des précisions pour chacune des unités désignables sont présentées ci-dessous.
Il convient de rappeler que le BBS fournit une estimation de la tendance de la population de la sous-espèce des Prairies et qu’en raison des limites du relevé, la fiabilité de cette estimation est faible à modérée (Environnement Canada, 2013). Selon les résultats du BBS, la population a décliné de manière statistiquement significative de 2,8 % par année entre 1970 et 2011 (soit l’équivalent d’une perte de 69 % pour l’ensemble de cette période) et de manière non significative de 1,3 % par année au cours de la période de dix ans écoulée entre 2001 et 2011 (soit l’équivalent d’un déclin de 13 % pour l’ensemble de cette période (Tableau 3; Figure 6). Si l’on se fonde sur l’estimation du déclin à long terme du BBS (2,8 % par année) pour inférer la tendance au cours de la période de 10 ans la plus récente, qui fournit probablement une meilleure représentation de la tendance à court terme, le déclin global de la population de la sous-espèce des Prairies sur 10 ans s’élève à 24,7 %.
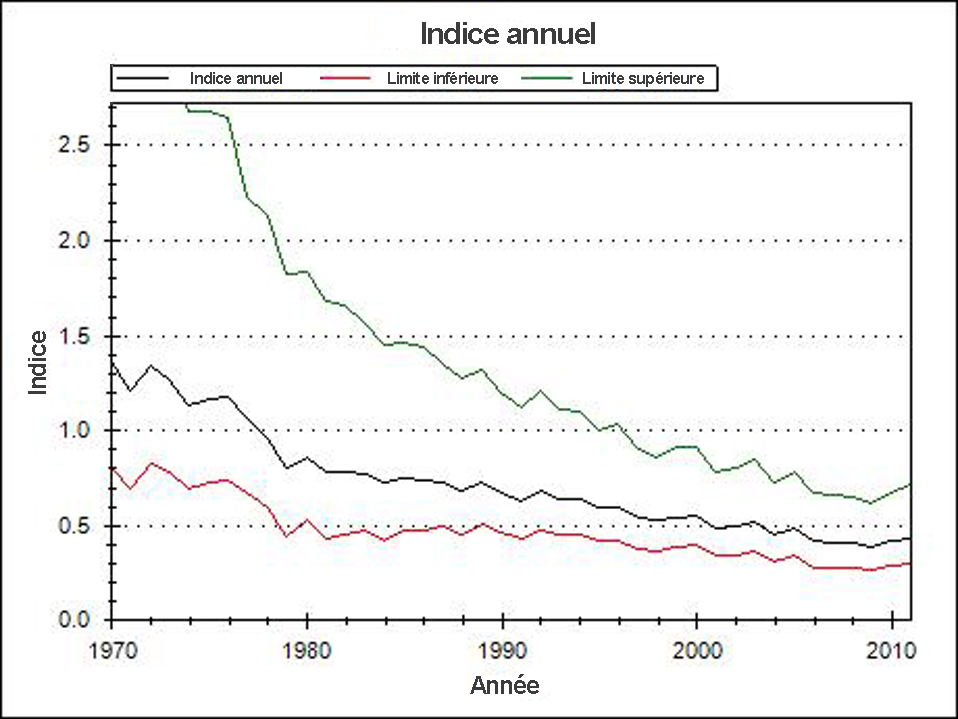
Graphique : © Environnement Canada
Description longue pour la figure 6
Graphique présentant les indices annuels des fluctuations des changements au sein de la population de la sous espèce des Prairies de la Pie grièche migratrice (L. l. excubitorides) au Canada d’après les résultats du Recensement des oiseaux nicheurs de 1970 à 2011. L’indice annuel montre un déclin, passant d’environ 1,4 en 1970 à moins de 0,5 en 2011. Le déclin est plus prononcé avant 2000.
D’après les données du BBS, la population albertaine semble relativement stable, tant à long terme qu’à court terme (hausses de 0,3 % par année de 1970 à 2011 et de 1,3 % par année de 2001 à 2011; Tableau 3), mais il convient de rappeler que la fiabilité du relevé est faible. Des relevés ciblés réalisés en 1993 et en 1996 dans la principale zone de nidification de l’espèce en Alberta ont également révélé que la population était relativement stable (Bjorge et Kiliaan, 1997). En revanche, selon des relevés effectués en bordure de route, l’indice de population aurait décliné de 19,1 % au cours de la période de 15 ans écoulée entre 1998 et 2013 et de 11,0 % au cours de la plus récente période de dix ans (Prescott, 2013; voir également le Tableau 2).
En Saskatchewan, les données du BBS indiquent un déclin statistiquement significatif de 3,6 % par année entre 1970 et 2011 et un déclin non significatif de 3,0 % par année au cours de la période de dix ans écoulée entre 2001 et 2011 (Tableau 3). D’autres relevés ciblés font cependant état de déclins plus abrupts, les pertes étant évaluées à 54,3 % au cours de la période de dix ans écoulée entre 2003 et 2013 (Didiuk et al., 2014; Tableau 2).
Au Manitoba, les données du BBS font état d’un déclin statistiquement significatif de l’effectif de 5,1 % par année entre 1970 et 2011 et de 5,2 % par année entre 2001 et 2011 (soit l’équivalent d’un déclin de 52 % sur dix ans; Tableau 3). Les relevés ciblés réalisés au Manitoba depuis 1987 confirment ces tendances (Figure 7), les pertes sur dix ans oscillant entre 40,1 % (calcul fondé uniquement sur les données amassées entre 2002 et 2012) et 61,9 % (calcul fondé sur le déclin annuel moyen à long terme de 9,2 % par année enregistré entre 1993 et 2012). Les occurrences de nidification sont aujourd’hui presque entièrement confinées au coin sud-ouest de la province, ce qui reflète le retrait global de l’espèce du sud-est de la province (K. DeSmet, comm. pers., 2012). Dans le sud-est du Manitoba, le nombre de couples nicheurs a chuté d’environ 60 en 1987 à 20 en 2004, alors que la population restante était concentrée dans la région de Winnipeg (K. DeSmet, comm. pers., 2012). Aucune preuve concluante de nidification n’a été décelée dans le sud-est du Manitoba depuis 2009 (K. DeSmet, comm. pers., 2012), mais des signes de nidification potentielle ont été observés dans trois carrés de l’atlas dans le cadre des travaux du présent atlas des oiseaux nicheurs. Les relevés intensifs réalisés depuis 2010 indiquent que moins de 50 couples nichent encore dans la province, la presque totalité d’entre eux occupant quelques enclaves de prairies dans l’extrême sud-ouest de la province (K. DeSmet, comm. pers., 2014).
En bref, la sous-espèce des Prairies a subi d’importants déclins répartis sur de nombreuses années, et cette tendance se maintient. Les résultats du BBS font état d’un déclin de seulement environ 13 % entre 2001 et 2011, mais une fois de plus, tel qu’il a déjà été mentionné, le BBS comporte plusieurs limites, et les relevés ciblés fournissent une meilleure estimation de l’évolution de la population régionale de l’espèce. Selon ces relevés ciblés, le déclin survenu sur une période de dix ans est estimé à 11,0 % en Alberta, à 54,3 % en Saskatchewan et à 40,1 % au Manitoba. Une fois les résultats ajustés en fonction de la taille des populations dans les trois provinces, on obtient un déclin global d’environ 47 % pour la sous-espèce des Prairies au cours de la plus récente période de dix ans (2003-2013).
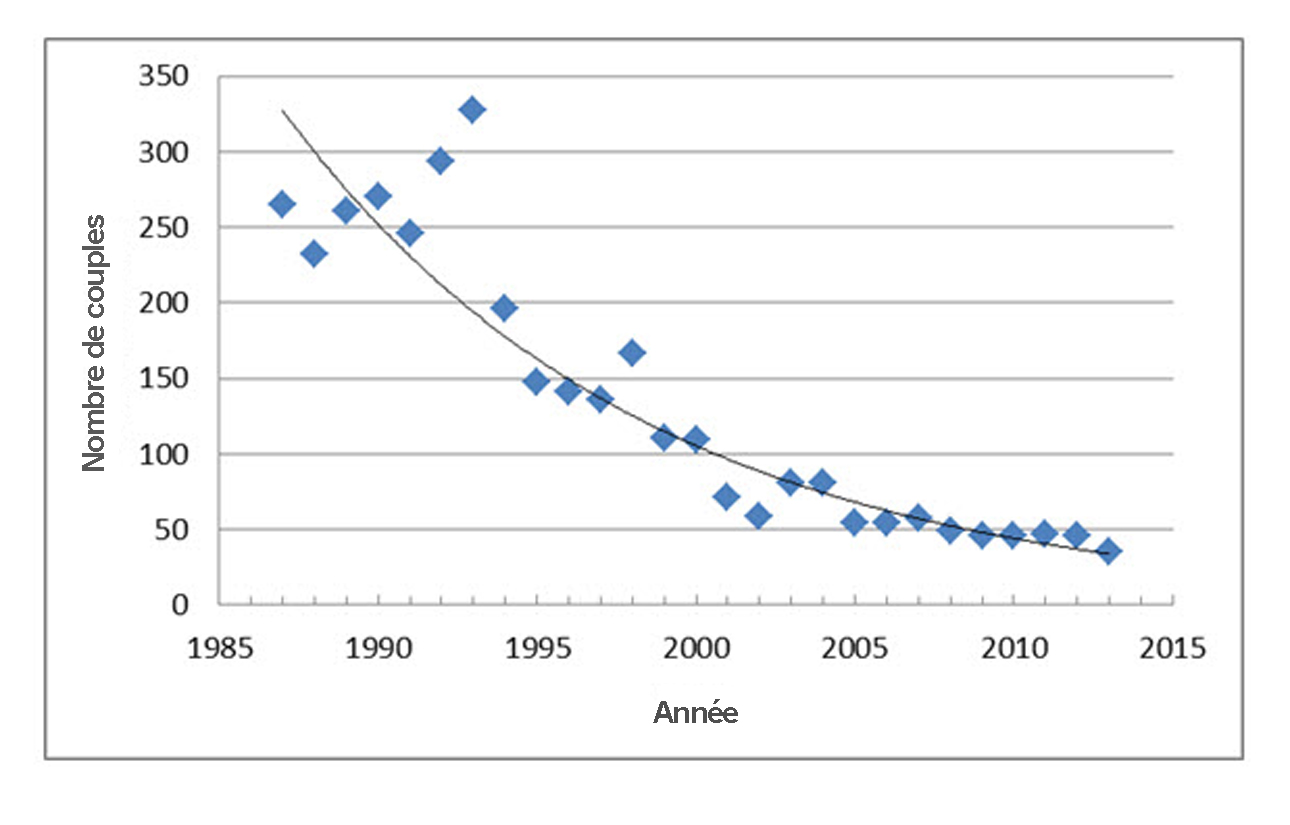
Graphique : © Environnement Canada
Description longue pour la figure 7
Graphique indiquant les tendances du nombre de couples nicheurs de Pies grièches dans le sud ouest du Manitoba entre 1987 et 2013, d’après des relevés ciblant l’espèce effectués depuis 1987. Le nombre de couples a passé de plus de 250 en 1987 à moins de 50 en 2013, et on a noté une pointe de plus de 325 couples au début des années 1990.
En Ontario, les données du BBS semblent indiquer que la population a décliné de manière statistiquement significative de 12,4 % par année entre 1970 et 2011 et de 12,2 % entre 2001 et 2011 (Tableau 3). Ces résultats doivent être interprétés avec prudence en raison de la faible taille des échantillons et de l’occurrence restreinte de l’espèce. Les résultats du deuxième Atlas des oiseaux nicheurs de l’Ontario font état d’un déclin de 63 % de la probabilité d’observer l’espèce durant les 20 années qui se sont écoulées entre les travaux du premier et du deuxième atlas (1981-1985 et 2001-2005) (Cadman et al., 2007), soit l’équivalent d’une perte de plus de 30 % sur dix ans. Les relevés ciblés réalisés par le Service canadien de la faune entre 1992 et 2013 indiquent également un déclin équivalent à une réduction d’effectif moyenne de 2,9 % par année, soit l’équivalent d’une perte de 46 % sur les 22 ans écoulés durant cette période (Figure 8). Au cours de la plus récente période de dix ans écoulée entre 2003 et 2013, le nombre d’individus a chuté de 59 à 51 adultes, soit un déclin de 13 %. Pour cette espèce, les tendances calculées à partir de séries temporelles plus longues sont probablement plus révélatrices. Si l’on utilise la tendance établie sur la série temporelle plus longue (déclin de 2,9 % par année), le déclin s’établit à environ 26 % sur une période de dix ans. C’est cette valeur qui est utilisée dans le présent rapport.
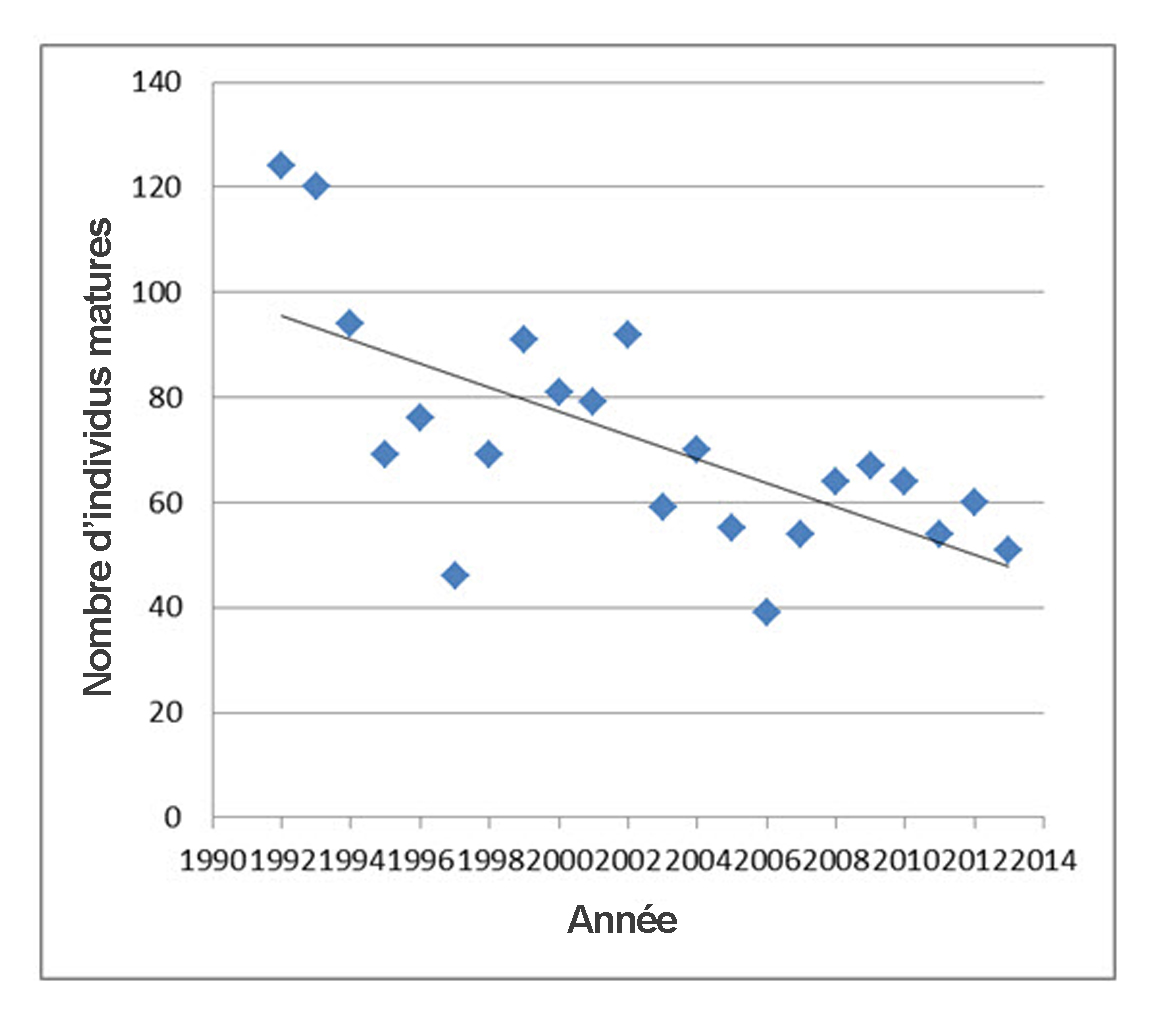
Graphique : © Environnement Canada
Description longue pour la figure 8
Graphique montrant les tendances du nombre de Pies grièches migratrices matures recensées durant la saison de nidification lors des relevés ciblés réalisés en Ontario entre 1992 et 2013. Le nombre est passé de plus de 125 en 1992 à environ 50 en 2013.
La situation de l’espèce au Québec est encore plus catastrophique qu’en Ontario. Durant les travaux du premier atlas des oiseaux nicheurs, qui se sont échelonnés de 1984 à 1989, l’espèce a été observée dans 30 carrés. Toutefois, sa présence n’a été relevée que dans un seul carré durant les travaux en cours du présent atlas. L’espèce a pratiquement disparu du Québec.
La Pie-grièche migratrice peut se disperser sur des distances pouvant atteindre 500 km (Chabot, 2011a). Une immigration de source externe est donc possible. Les estimations du flux génétique indiquent que la dispersion se produit plus souvent de la limite nord de l’aire de répartition de l’espèce vers le sud (Chabot, 2011a; Figure 5). Bien que les études de baguage aient fait état de déplacements entre les principales zones de nidification de l’espèce en Ontario et entre l’Ontario et la région de l’Outaouais au Québec, aucun échange d’individus bagués en provenance d’autres régions de l’Amérique du Nord n’a été signalé à ce jour durant la période de reproduction (Burnside, 1987; Chabot, 2011a).
Au moins quelques parcelles d’habitat propice semblent subsister dans les régions anciennement occupées par l’espèce. Bien que l’impact de la fragmentation de l’habitat sur l’espèce ne soit pas entièrement compris, il semble que la taille des parcelles d’habitat et divers facteurs agissant à l’échelle du paysage aient un impact déterminant sur la qualité de l’habitat pour l’espèce (Cuddy et Leviton, 1996; Glynn-Morris, 2010; Chabot et Lagois, 2012), qui pourrait être liée à la répartition irrégulière en colonies lâches de l’espèce. Quoi qu’il en soit, des parcelles d’habitat inoccupées, mais apparemment propices existent encore dans des régions qui abritaient autrefois l’espèce, et leur présence accroît la probabilité d’une immigration de source externe.
Dans le cas de la population des Prairies, une immigration de source externe en provenance des États-Unis paraît peu probable, car les populations au sud de la frontière canado-états-unienne sont en déclin. De la même façon, dans le nord-est des États-Unis, l’espèce est soit disparue, soit présente en très faible nombre, et elle y connaît un déclin généralisé. En conséquence, dans le cas de la sous-espèce de l’Est, une immigration de source externe est jugée impossible.
Un lien a été clairement établi à l’échelle de l’Amérique du Nord entre le déclin des populations de la Pie-grièche migratrice et la destruction et la dégradation de l’habitat de l’espèce (Kridelbaugh, 1982; Luukkonen, 1987; Woods, 1994; Yosef, 1996). La superficie des pâturages non cultivés a chuté de 39 % entre 1946 et 1986 dans les régions des provinces des Prairies où les déclins des populations de l’espèce ont été les plus abrupts (Telfer, 1992). Samson et Knopf (1994) ont quantifié l’ampleur des réductions de superficie des prairies indigènes en Alberta (prairies mixtes : 61 %), en Saskatchewan (prairies à graminées hautes : 81 %; prairies à graminées courtes : 85 %), au Manitoba (prairies à graminées hautes et prairies mixtes : 99 %) et plus au sud, le long des portions occidentale et centrale des Grandes Plaines. Dans le nord-est des États-Unis et l’est du Canada, les pertes d’habitat ont été causées par l’abandon des terres agricoles, le développement, les changements généralisés touchant les pratiques agricoles, la reforestation et la succession végétale (Novak, 1989; Cadman, 1985, 1990; Cuddy, 1995; Jobin et al., 1996, 2010). Bien que la perte d’habitat de reproduction ait eu un impact négatif important sur les populations de Pies-grièches migratrices et ait vraisemblablement entraîné une contraction de l’aire de reproduction à la limite nord de l’aire de répartition de l’espèce, des parcelles d’habitat apparemment propice demeurent inoccupées dans de nombreuses régions des États-Unis (voir par exemple Brooks et Temple, 1990a; Robbins, 1991; Cuddy et Leviton, 1996) et dans l’est et l’ouest du Canada (SCF, données inédites). D’autres facteurs pourraient donc être en cause.
La destruction et la dégradation de l’habitat d’hivernage ont vraisemblablement contribué au déclin des populations de l’espèce (Brooks, 1988; Lymn et Temple, 1991). Dans l’aire d’hivernage de l’espèce, de très nombreux pâturages et vieux champs abandonnés ont été convertis en terres agricoles. De vastes étendues de prairie ont été détruites le long de la côte du golfe du Mexique (Lymn et Temple, 1991), importante aire d’hivernage de l’espèce (Chabot, 2011a). Telfer (1992) a étudié l’utilisation de l’habitat par la Pie-grièche migratrice au Texas et a constaté que seulement 17 % des prairies indigènes originales étaient encore présentes.
La conversion des prairies en quartiers résidentiels le long de la côte du golfe du Mexique a probablement un impact sur les quantités de nourriture disponibles et d’autres besoins de l’espèce liés à l’habitat. Les individus résidents non migrateurs défendent leur territoire en hiver, et dans les régions où la quantité d’habitat disponible est limitée, les individus migrateurs sont peut-être forcés d’utiliser des parcelles d’habitat d’hivernage de qualité marginale, ce qui peut compromettre leur survie en hiver (Brooks et Temple, 1990a). Craig et Chabot (2012) ont noté que, dans la région de la côte du golfe du Mexique, au Texas, la répartition des parcelles d’habitat disponibles est fondée sur l’âge, les individus plus âgés étant plus nombreux le long de la côte dans les secteurs résidentiels densément peuplés et les individus plus jeunes, plus abondants dans les régions situées plus à l’intérieur des terres. L’espèce n’est pas communément rencontrée dans ces régions côtières durant la saison de reproduction (F. Chavez-Ramirez, comm. pers., 2012), ce qui donne à croire que ces régions sont utilisées exclusivement par les individus migrateurs et non par les résidents permanents.
La mortalité résultant de collisions avec des véhicules a peut-être un impact négatif important sur les populations de l’espèce (Miller, 1931; Campbell, 1975; Novak, 1989; Luukkonen, 1987; Flickinger, 1995). Les pies-grièches volent souvent près du sol lorsqu’elles traversent des routes, en particulier en bordure de routes où la végétation herbacée a été tondue et est plus courte que la végétation environnante. Les haies, les clôtures, les fils électriques et autres éléments de l’habitat fréquemment utilisés par les pies-grièches sont souvent plus nombreux le long des routes, ce qui accroît le risque de collisions avec des véhicules. La menace globale posée par la mortalité routière augmente avec l’expansion et l’amélioration du réseau routier et l’intensification de la circulation routière à l’échelle de l’aire de répartition de l’espèce.
Les pesticides ont probablement contribué au déclin des populations de l’espèce, car les pies-grièches sont des prédateurs de niveau trophique supérieur (Fraser et Luukkonen, 1986). Il y a quelques décennies, deux études ont révélé la présence de concentrations relativement élevées de DDE ou de DDT dans les tissus de pies-grièches nichant en Illinois et en Virginie (Anderson et Duzan, 1978) et en Californie (Rudd et al., 1981). La présence de DDE a également été détectée dans des œufs non éclos en Ontario en 2010 (SCF, données inédites). Des signes de réduction de l’épaisseur de la coquille ont été observés en Illinois, mais aucun signe apparent n’a été détecté en Californie ou en Floride (Hands et al., 1989). Busbee (1977) a noté que les jeunes exposés à la dieldrine étaient plus lents à attaquer et à tuer leurs proies que les individus qui n’y avaient pas été exposés. Blumton (1989) a indiqué que des traces de contamination par des pesticides avaient été décelées en Virginie chez des pies-grièches autopsiées, mais il n’a établi aucun lien entre la contamination et la mortalité des oiseaux.
Les pesticides peuvent également avoir un impact indirect sur l’espèce en réduisant la quantité de nourriture disponible. Un lien a été établi entre le déclin des populations de pies-grièches dans les Prairies canadiennes et l’épandage de dieldrine contre les criquets, qui constituent 30 à 75 % du régime alimentaire de l’espèce (C. Campbell, in Cadman, 1985; Yosef, 1994, 1996). Récemment, pour freiner la propagation des fourmis de feu dans les quartiers d’hiver, on a fait appel au mirex, produit qui peut s’accumuler et atteindre des concentrations relativement élevées dans les tissus des pies-grièches (Collins et al., 1974; Lymn et Temple, 1991). De façon générale, si l’on se fonde sur les preuves amassées à ce jour (Herkert, 2004; SCF, données inédites), les pesticides ne représentent vraisemblablement pas la principale cause du déclin de l’espèce, en particulier dans les régions où le pacage constitue la principale utilisation des terres et où les applications de pesticides sont limitées. Les pesticides doivent à tout le moins être considérés comme une menace de faible niveau.
La Pie-grièche migratrice compte plusieurs prédateurs d’œufs et d’oisillons, dont la Pie d’Amérique (Pica hudsonia) et la couleuvre à nez mince (Pituophis melanoleucus) dans l’Ouest, de même que la belette à longue queue (Mustela frenata), les chats féraux (Felis domesticus), le renard (Vulpes sp.) et le raton laveur (Procyon lotor) (Bent, 1950; DeGues, 1990; Gawlik et Bildstein, 1990; Scott et Morrison, 1990; Collister, 1994; Collister et Wilson, 2007 b). Durant la saison de reproduction, les taux de prédation des adultes, des œufs et des oisillons au nid semblent plus élevés près des routes et des haies (Yosef, 1994). La prédation des nids peut être une cause de mortalité importante dans certaines régions (Collister et Wilson, 2007b; K. DeSmet, comm. pers.; SCF, données inédites). Le succès de nidification peut être aussi bas que 35 % (et même plus faible que cette valeur) dans les régions où la pression de prédation est élevée (voir par exemple DeGeus et Best, 1991). La prédation peut donc être un facteur limitatif important, en particulier dans les régions où les populations sont de petite taille. Loin d’agir comme un facteur naturel de régulation des populations, l’augmentation des effectifs des prédateurs non indigènes (p. ex, chats féraux) et d’autres espèces qui tirent profit de la présence des humains (p. ex. raton laveur, corvidés) peut constituer une menace plus directe au succès de reproduction des pies-grièches.
La mortalité post-envol est apparemment élevée et pourrait osciller selon certaines sources entre 33 à 53 % dans les dix jours suivant l’envol (Burton, 1990; Collister, 1994; Yosef, 1996; Chabot et al., 2001 b). Toutefois, d’autres études (voir par exemple Blumton, 1989) font état de taux de survie post-envol relativement élevés. On ignore dans quelle mesure la survie post-envol est un facteur limitatif, mais elle a probablement un impact important dans certaines régions.
Les conditions météorologiques peuvent avoir un impact négatif important sur le succès de reproduction de l’espèce. Les épisodes de temps défavorable, dont la fréquence risque d’augmenter sous l’effet des changements climatiques, peuvent provoquer la perte de nids et de couvées. Porter et al. (1975) rapporte que de graves orages ont entraîné la perte de 9 nids sur 12 dans l’est du Colorado. De tels orages causent fréquemment la perte de nids dans les Grandes Plaines de l’est du Colorado (S. Craig, comm. pers., 2012). Des périodes de temps froid et/ou pluvieux peuvent également causer la perte de nids et compromettre le succès des couvées au Canada (Collister et Wilson, 2007; A. Chabot, obs. pers.). Bien que les pies-grièches produisent souvent une deuxième couvée si la première tentative de nidification a échoué (Miller, 1931; Collister et Wilson, 2007 b), l’ampleur de la menace posée par les conditions météorologiques défavorables augmente en fonction de la fréquence avec laquelle ces épisodes se répètent.
Le virus du Nil occidental a été incriminé dans la mort de Pies-grièches migratrices en Ontario (Bertelsen et al., 2004) et au Manitoba (Lindgren et al., 2009). Toutefois, la sensibilité de l’espèce à cette maladie ou à toute autre maladie n’a pas été documentée. En conséquence, la gravité de cette menace est présentement inconnue.
Dans les prairies à graminées courtes, le bétail peut endommager ou tuer les quelques arbres susceptibles d’être utilisés comme support de nidification. Dans le sud est du Colorado, les couples nicheurs de la sous-espèce L. l. excubitorides aménagent leur nid plus fréquemment dans des arbustes protégés du bétail par des exclos que dans des prairies broutées par le bétail (S. Craig, comm. pers., 2012) où la végétation ligneuse est exposée aux effets négatifs du broutage. En se frottant contre de petits arbres utilisés comme support de nidification, le bétail peut provoquer la chute du contenu des nids sur le sol et la mort d’œufs et d’oisillons (A. Chabot, obs. pers.).
Pour les deux populations de la Pie-grièche migratrice, le nombre de localités dépend du nombre de propriétaires privés susceptibles d’avoir un impact sur les populations reproductrices en modifiant l’habitat. Le nombre de propriétaires n’a pas été déterminé, mais d’après les nombres estimés de couples nicheurs, le nombre de localités est tenu pour largement supérieur à 1 000 dans le cas de la sous-espèce des Prairies, mais ne dépasse pas 25 à 50 dans celui de la sous-espèce de l’Est.
La Pie-grièche migratrice bénéficie d’une protection internationale (Canada, Mexique et États-Unis) aux termes de la Loi sur la convention concernant les oiseaux migrateurs, en vertu de laquelle nul ne peut avoir en sa possession des œufs, des nids, des plumes ou des individus de cette espèce.
En vertu de la Loi sur les espèces en péril, la Pie-grièche de la sous-espèce L. l. excubitorides a été désignée à l’échelle nationale espèce menacée, et la sous-espèce L. l. migrans (appelée Pie-grièche de l’Est dans le présent rapport), espèce en voie de disparition. Une ébauche de programme de rétablissement a été élaborée pour la Pie-grièche de la sous-espèce des Prairies. L’habitat essentiel de la sous-espèce a été désigné dans la réserve nationale de faune de la base des Forces canadiennes Suffield, dans le sud-est de l’Alberta (A. Didiuk, comm. pers., 2013). D’autres parcelles d’habitat essentiel ont été désignées dans le sud-ouest de la Saskatchewan et ont été incluses dans un plan d’action en préparation applicable aux espèces en péril dans les régions situées au sud de la ligne de partage des eaux (South of the Divide species at risk action plan) (A. Didiuk, comm. pers., 2013).
Dans l’est du Canada, une ébauche de programme de rétablissement (Environnement Canada, 2010a) a été élaborée pour la Pie-grièche de la sous-espèce de l’Est. L’habitat essentiel de la sous-espèce en Ontario a été désigné dans le cadre de cette initiative.
La Pie-grièche migratrice est désignée espèce sensible (Sensitive Species) et espèce préoccupante (Species of special concern) en Alberta (Anonyme, 2001). Elle n’a aucun statut juridique en Saskatchewan et est classée espèce en voie de disparition au Manitoba et en Ontario (Loi de 2007 sur les espèces en voie de disparition). Elle est cotée espèce menacée au Québec (Gouvernement du Québec, 2000), statut qui correspond à celui d’espèce en voie de disparition dans d’autres provinces.
Aux États-Unis, l’espèce est cotée en voie de disparition, menacée ou préoccupante dans 26 États (Pruitt, 2000) ou fait l’objet d’une évaluation en vue d’un éventuel classement dans d’autres États (p. ex. Tennessee).
La Pie-grièche migratrice n’est pas classée au Mexique (Commission de coopération environnementale, 2000) et ne jouit d’aucune protection juridique spéciale autre que celle que lui confère la Loi sur la convention concernant les oiseaux migrateurs.
À l’échelle mondiale, la Pie-grièche migratrice est cotée G4 (apparemment non en péril) depuis le dernier réexamen de son statut en février 2001. En dépit du déclin de ses populations à l’échelle de l’Amérique du Nord, l’espèce était considérée comme encore largement répartie et commune dans certaines régions. À l’échelle nationale, elle est cotée N4 aux États-Unis et N4B au Canada. L’espèce est inscrite sous la catégorie « préoccupation mineure » (Least Concern) sur la liste rouge de l’UICN.
Les cotes suivantes ont été attribuées à la Pie-grièche migratrice dans les États situés immédiatement au sud des Prairies canadiennes : Montana (S3B), Dakota du Nord (SU), Dakota du Sud (S3S4B), Wyoming (S3), Minnesota (S2B), Wisconsin (S1B), Michigan (SNR). Les cotes suivantes lui ont été attribuées dans les États situés immédiatement au sud de l’aire de répartition canadienne de la sous-espèce de l’Est : Maine (SHB S1?N), New Hampshire (SHB), Vermont (SHB), New York (S1B), Pennsylvanie (SNRB), Ohio (S1), Indiana (S3B).
Bien que ses populations aient décliné considérablement au cours du siècle dernier et que les baisses d’effectif semblent se poursuivre dans certaines régions, la sous-espèce L. l. excubitorides a été classée mondialement G4T4 (apparemment non en péril) lors du dernier réexamen de son statut en janvier 2003. Sa cote mondiale arrondie est T4 (apparemment non en péril). À l’échelle nationale, la sous-espèce est cotée NNR au Canada. Elle est classée S2B au Manitoba et S3B en Saskatchewan.
De la même façon, même si elle est passablement rare dans certaines portions de son aire de répartition, la sous-espèce L. l. migrans a été désignée G4T3Q à l’échelle mondiale (apparemment non en péril mondialement) lors du dernier réexamen de son statut en octobre 1999. Sa cote mondiale arrondie, T3 (vulnérable), lui a été attribuée en considération de sa répartition irrégulière et des déclins spectaculaires survenus à l’échelle de son aire de répartition. À l’échelle nationale, la sous-espèce est cotée N3B, N3N aux États-Unis et NNR au Canada. Elle est actuellement classée S1B au Manitoba et au Québec, SNR au Nouveau-Brunswick, en Nouvelle-Écosse et à l’Île-du-Prince-Édouard, et S2B en Ontario. Toutes les cotes susmentionnées ont été attribuées à la sous-espèce de l’Est avant que celle-ci soit reconnue au Canada comme une sous-espèce distincte encore à nommer.
Tant dans l’est que dans l’ouest du Canada, la vaste majorité des parcelles d’habitat propice à l’espèce se trouve sur des terres privées. Dans l’ouest du pays, le bloc Ouest du parc national des Prairies et des portions de certains pâturages communautaires fédéraux en Saskatchewan abritent des nombres relativement élevés d’individus de la sous-espèce des Prairies (A. Didiuk, comm. pers., 2012). Toutefois, l’avenir du programme fédéral des pâturages communautaires est actuellement incertain et pourrait même être compromis (Stewart, 2013).
Dans les Prairies, les mesures de conservation visent principalement à répertorier et à protéger les parcelles d’habitat propice à l’espèce en encourageant les propriétaires privés à conclure des accords d’intendance et à clôturer et à protéger les secteurs reconnus comme des sites de nidification et d’alimentation importants. Dans le sud-est de l’Alberta, on a planté un grand nombre de shépherdies argentées le long de la ligne de chemin de fer du Canadien Pacifique. En Alberta, divers projets en cours ont pour but de protéger les secteurs abritant de fortes densités de pies-grièches (voir Prescott et Bjorge, 1999). Les initiatives Operation Grassland Community et Shrubs for Shrikes sensibilisent les propriétaires à la situation de la Pie-grièche migratrice dans les Prairies.
En Ontario, la Loi de 2007 sur les espèces en voie de disparition interdit à quiconque d’endommager ou de détruire l’habitat de nidification de la Pie-grièche migratrice. Parcs Ontario a pris des mesures dans le but de protéger l’habitat de l’espèce aux ranchs Cameron et Windmill, dans la plaine de Carden. Une proposition vise à inclure ces deux grandes propriétés dans un nouveau parc provincial. Dans la plaine Napanee, Conservation de la nature Canada est propriétaire de la réserve naturelle Scheck, où la Pie-grièche migratrice a niché régulièrement au cours de la dernière décennie et où des jeunes pies-grièches élevées en captivité ont été relâchées en 2012. En Ontario, les gouvernements fédéral et provincial ont tous deux financé diverses initiatives d’intendance de l’habitat bénéfiques pour la Pie-grièche migratrice de la sous-espèce de l’Est. Les mesures de conservation mises en place consistent principalement à éclaircir la strate arbustive, à installer des clôtures et à aménager des puits pour favoriser le broutage par le bétail. La plupart des projets se déroulent dans les principales zones de nidification des plaines de Carden et Napanee.
Au Québec, Conservation de la nature Canada et Protection des oiseaux du Québec sont copropriétaires de la ferme de Ghost Hill, riche en habitat propice et où des jeunes pies-grièches élevées en captivité ont été relâchées en 2004 et 2009.
La rédactrice remercie les nombreuses personnes qui lui ont fourni des données inédites sur l’espèce ou qui ont examiné et commenté les différentes versions du manuscrit, en particulier Anna Calvert, Nikki Boucher, Vivian Brownell, Rhonda Donley, Gilles Falardeau, Vicki Friesen, Marcel Gahbauer, Hazel Wheeler, Darren Irwin, Marty Leonard, Marie-France Noel, Kate Pitt, Megan Rasmussen, Chris Risley, Ben Sawa et Scott Wilson. Ont également fourni de précieuses informations et participé à l’examen du manuscrit Susan Craig, Ken DeSmet (Conservation Manitoba), Andrew Didiuk (Service canadien de la faune), David Fraser (Ministry of Environment de la Colombie-Britannique), Todd Norris (ministère des Richesses naturelles de l’Ontario), David Prescott (Alberta’s Environment and Sustainable Resource Development), François Shaffer (Service canadien de la faune) et Ken Tuininga (Service canadien de la faune). Alain Filion (Environnement Canada) a fourni des services SIG. Le financement nécessaire à la préparation de ce rapport a été fourni par Environnement Canada.
- Luc Bélanger, Environnement Canada
- Bill Bristol, Agriculture et Agroalimentaire Canada
- Vivian Brownell, ministère des Richesses naturelles de l’Ontario
- Ken DeSmet, Conservation Manitoba
- Andrew Didiuk, Service canadien de la faune, président de l’équipe de rétablissement de la Pie-grièche migratrice (population des Prairies)
- Dave Duncan, Service canadien de la faune
- Gilles Falardeau, Environnement Canada
- David Fraser, Ministry of the Environment de la Colombie-Britannique
- Nicole Fergus, Nature Saskatchewan
- Chris Friesen, Centre de données sur la conservation du Manitoba, Conservation Manitoba
- Isabelle Gauthier, ministère du Développement durable, de l’Environnement, de la Faune et des Parcs du Québec
- Kerry Grisley, Operation Grassland Community, Alberta Fish and Game Association
- Tara Imlay, Wildlife Preservation Canada, Ontario
- David Ingstrup, Service canadien de la faune
- Neil Jones, Secrétariat du COSEPAC, Environnement Canada
- Angela McConnell, Service canadien de la faune
- Patrick Nantel, Parcs Canada
- Todd Norris, ministère des Richesses naturelles de l’Ontario
- Annie Paquet, ministère du Développement durable, de l’Environnement, de la Faune et des Parcs du Québec
- Jeanette Pepper, Ministry of the Environment de la Saskatchewan
- Gigi Pittoello, Ministry of the Environment de la Saskatchewan
- Dave Prescott, Alberta Environment and Sustainable Resource Development
- Richard Quinlan, Alberta Environment and Sustainable Resource Development
- Agnes Richards, Service canadien de la faune
- Jean-Pierre Savard, Environnement Canada
- Ben Sawa, Ministry of the Environment de la Saskatchewan
- Sonia Schnobb, Secrétariat du COSEPAC, Environnement Canada
- François Shaffer, Service canadien de la faune
- Don Sutherland, ministère des Richesses naturelles de l’Ontario
- Katrina Stipec, Ministry of the Environment de la Colombie-Britannique
- Benjamin Taylor, ministère de la Défense nationale
- Junior A. Tremblay, ministère du Développement durable, de l’Environnement, de la Faune et des Parcs du Québec
- Ken Tuininga, Service canadien de la faune, Président, Équipe de rétablissement de la Pie-grièche migratrice (population de l’Est)
- Drogomir Drajs Vujnovic, Alberta Tourism Parks and Recreation
Mme Amy Chabot (Ph.D.) est titulaire d’un diplôme de baccalauréat ès sciences (1991) et de maîtrise ès sciences (1994) de l’Université McGill. Son mémoire de maîtrise portait sur la biologie reproductive de la Pie-grièche migratrice en Ontario et au Québec et sur la sélection de l’habitat par l’espèce. En 2010, elle a obtenu un diplôme de doctorat de la Queen’s University. Ses recherches visaient à évaluer l’impact de la migration sur l’évolution et la conservation des passereaux en Amérique du Nord et, plus précisément, à documenter la structure génétique des populations et le comportement migrateur de la Pie-grièche migratrice. Elle continue d’étudier l’écologie des pies-grièches au Canada et aux États-Unis et s’intéresse tout particulièrement aux interactions saisonnières et à la dynamique des populations dans les aires d’hivernage. Elle agit à tire de conseillère scientifique au sein de l’équipe de rétablissement de la Pie-grièche de la sous-espèce de l’Est (Eastern Loggerhead Shrike Recovery Team) depuis 1994. Mme Chabot a assumé diverses fonctions à titre de biologiste, notamment en qualité de biologiste-conseil indépendante pour divers organismes gouvernementaux et non gouvernementaux et de membre du personnel d’Études Oiseaux Canada, de la New Hampshire Audubon Society, du North American Loon Fund et de la Society for Protection of New Hampshire Forests.
- Anderson, W.L., et R.E. Duzan. 1978. DDE residues and eggshell thinning in Loggerhead Shrikes, Wilson Bulletin 90:215 220.
- Anonyme. 2001. The general status of Alberta wild species in 2000, Alberta Sustainable Resource Development, Edmonton (Alberta).
- Askins, R.A., F. Chavez-Ramirez, B.C. Dale, C.A. Haas, J.R. Herkert, F.L. Knopf et P.D. Vickery. 2007. Conservation of grassland birds in North America: understanding ecological processes in different regions, Ornithological Monographs 64:1 46.
- Bakowsky, W., et J.L. Riley. 1994. A survey of the prairies and savannas of southern Ontario, p. 7 16, in R. Wickett, P. Dolan-Lewis, A. Woodliffe et P. Pratt (éd.), Proceedings of the Thirteenth North American Prairie Conference: Spirit of the Land, Our Prairie Legacy, 6 9 août 1992, Windsor: Corporation of the City of Windsor.
- Bartgis, R. 1992. Loggerhead Shrike, Lanius ludovicianus, p. 281 297 in K.J. Schneider et D.M. Pence (éd.), Migratory nongame birds of management concern in the Northeast, U.S. Dept. Interior, Fish and Wildlife Service, Newton Corner (Massachusetts).
- Bent, A.C. 1950. Life histories of North American Wagtails, Shrikes, Vireos and Their Allies, U.S. National Museum Bulletin 197.
- Berlanga, H., J.A. Kennedy, T.D. Rich, M.C. Arizmendi, C.J. Beardmore, P.J. Blancher, G.S. Butcher, A.R. Couturier, A.A. Dayer, D.W. Demarest, W.E. Eason, M. Gustafson, E. Iñigo-Elias, E.A. Krebs, A.O. Panjabi, V. Rodriguez Contreras, K.V. Rosenberg, J.M. Ruth, E. Santana Castellón, R. Ma. Vidal et T. Will. 2010. Sauvegardons nos oiseaux en commun : vision tri-nationale de Partenaires d’envol pour la conservation des oiseaux terrestres, Cornell Lab of Ornithology, Ithaca (New York).
- Bertelsen, M.F, R.A. Olberg, G.J. Crawshaw, A. Dibernardo, L.R. Lindsay, M. Drebot et I.K. Barker. 2004. West Nile virus infection in the eastern Loggerhead Shrike (Lanius ludovicianus migrans): Pathology, epidemiology and immunization, Journal of Wildlife Diseases 40:538 542.
- Bjorge, R.R., et H. Kiliaan. 1997. Densities of Loggerhead Shrikes on study blocks in southeastern Alberta inventoried in both 1993 and 1996, Blue Jay 55:217 219.
- Bjorge, R.R., et D.R.C. Prescott. 1996. Population estimate and habitat associations of the Loggerhead Shrike, Lanius ludovicianus, in southeastern Alberta, Canadian Field Naturalist 110:445 449.
- Blumton, A.K. 1989. Factors affecting Loggerhead Shrike mortality in Virginia, Master’s thesis, Virginia Polytechnique Institute and State University, Blacksburg (Virginie).
- Boal, C.W., T.S. Estabrook et A.E. Duerr. 2003. Productivity and breeding habitat of Loggerhead Shrikes in a southwestern urban environment, Southwestern Naturalist 48:557 52.
- Brooks, B.L. 1988. The breeding distribution, population dynamics, and habitat availability and suitability of an upper Midwest Loggerhead Shrike population, mémoire de maîtrise ès sciences, University of Wisconsin, Madison (Wisconsin).
- Brooks, B.L., et S.A. Temple. 1990a. Habitat availability and sustainability for Loggerhead Shrikes in the Upper Midwest, American Midland Naturalist 123:75 83.
- Brooks, B.L., et S.A. Temple. 1990 b. Dynamics of a Loggerhead Shrike population in Minnesota, Wilson Bulletin 102:441 450.
- Brownell, V.R., et J.L. Riley. 2000. The Alvars of Ontario: Significant Natural Areas in the Ontario Great Lakes Region, Federation of Ontario Naturalists.
- Burnside, F. 1987. Long-distance movements in Loggerhead Shrikes, Journal of Field Ornithology 58:62 65.
- Burnside, K.M. 2006. Molts, plumages, and age classes of passerines and “near-passerines”: a bander’s overview, North American Bird Bander 31:175 193.
- Burton, K.M. 1990. An investigation of population status and breeding biology of the Loggerhead Shrike (Lanius ludovicianus) in Indiana, mémoire de maîtrise ès sciences, Indiana Univ. Bloomington.
- Burton, K.M., et D.R. Whitehead. 1990. An investigation of population status and breeding biology of the Loggerhead Shrike in Indiana, rapport inédit to the Indiana Department of Natural Resources, Nongame and Endangered Wildlife Program.
- Busbee, E.L. 1977. The effects of dieldrin on the behavior of young Loggerhead Shrikes, The Auk 94:28 35.
- Cade, T.J. 1995. Shrikes as predators, p. 1 5 in R. Yosef et F.E. Lohrer (éd.), Shrikes (Laniidae) of the World: Biology and Conservation, Proceedings of the Western Foundation of Vertebrate Zoology 6.
- Cade, T.J., et C.P. Woods. 1997. Changes in distribution and abundance of Loggerhead Shrike, Conservation Biology 11:21 31.
- Cadman, M.D. 1985. Status report on the Loggerhead Shrike (Lanius ludovicianus) in Canada, Committee on the Status of Endangered Wildlife in Canada, Ottawa (Ontario).
- Cadman, M.D., P.F.J. Eagles et F.M. Helleiner (éd.). 1987. Atlas of Breeding Birds of Ontario, University of Waterloo Press, Waterloo (Ontario).
- Cadman, M.D. 1990. Update status report on the Loggerhead Shrike (Lanius ludovicianus) in Canada, COSEWIC, Ottawa.
- Cadman, M.D., D.A. Sutherland, G.G. Beck, D. Lepage et A.R. Couturier (éd.). 2010. Atlas des oiseaux nicheurs de l’Ontario, 2001-2005, Environnement Canada, Études d’Oiseaux Canada, ministère des Richesses naturelles de l’Ontario, Ontario Field Ornithologists, et Ontario Nature, Toronto.
- Campbell, C. 1975. Distribution and breeding success of the Loggerhead Shrike in southern Ontario, Sefvice canadien de la faune, rapport no 6065.
- Catling, P.M. 2008. The extent and floristic composition of the Rice Lake Plains based on remnants, Canadian Field-Naturalist 122:1 20.
- Catling, P.M., et V.R. Brownell. 1999. Additional notes on the vegetation of dry openings along the Trent River, Ontario, Canadian Field-Naturalist 113:506 509.
- Chabot, A. 2009. Loggerhead Shrike Ontario 2009 Adult Banding Summary, rapport inédit pour le Canadian Wildlife Service – Ontario Region.
- Chabot, A. 2010. 2010 Loggerhead Shrike Banding Summary, rapport inédit pour le Service canadien de la faune, Région de l’Ontario Region.
- Chabot, A.A. 2011a. The impact of migration on the evolutionary and conservation of an endemic North America passerine: Loggerhead Shrike (Lanius ludovicianus), Dissertation, Queen’s University.
- Chabot, A. 2011b. 2011 Ontario Loggerhead Shrike Banding Summary, rapport inédit pour le Canadian Wildlife Service – Ontario Region.
- Chabot, A.A., D.M. Bird et R.D. Titman. 2001a. Breeding biology and nesting success of Loggerhead Shrikes in Ontario, Wilson Bulletin 113:285 289.
- Chabot, A.A., R.D. Titman et D.M. Bird. 2001 b. Habitat use by Loggerhead Shrikes in Ontario and Quebec, Canadian Journal of Zoology 79:916 925.
- Chabot, A.A., et E. Lagois. 2012. Analysis of Loggerhead Shrike habitat associations and grassland bird species assemblages in Ontario, rapport inédit pour Wildlife Preservation Canada.
- Chabot, A.A., et S.C. Lougheed. 2005. Conservation genetics of the eastern Loggerhead Shrike in Ontario, rapport inédit pour le Endangered Species Recovery Fund.
- Collins, J.A. 1996. Breeding and wintering ecology of the Loggerhead Shrike in southern Illinois, mémoire de maîtrise ès sciences, Southern Illinois University, Carbondale (Illinois).
- Collins, H.L., G.P. Markin et J. Davis. 1974. Residue accumulation in selected vertebrates following a single aerial application of mirex bait, Louisiana – 1971 72, Pesticides Monitoring Journal 8:125 130.
- Collister, D.M. 1994. Breeding ecology and habitat preservation of the Loggerhead Shrike in southeastern Alberta, Master’s thesis, Univ. of Calgary, Calgary (Alberta).
- Collister, D.M. 1999. Update Status Report, Loggerhead Shrike Western Canadian Population, Unpubl. COSEWIC report, Ottawa.
- Collister, D.M. 2013. Annual adult survival of Loggerhead Shrikes in southeastern Alberta, North American Bird Bander 38:97 105.
- Collister, D.M., et D. Wicklum. 1996. Intraspecific variation in Loggerhead Shrikes: sexual dimorphism and implication for subspecies classification, The Auk 113:221 223.
- Collister, D.M., et K. DeSmet. 1997. Breeding and natal dispersal in the Loggerhead Shrike, Journal of Field Ornithology 68:273 282.
- Collister, D.M., et S. Wilson. 2007a. Territory size and foraging habitat of Loggerhead Shrikes (Lanius ludovicianus) in southeastern Alberta, Journal of Raptor Research 41:130 138.
- Collister, D.M., et S. Wilson. 2007 b. Contributions of weather and predation to reduced breeding success in a threatened northern Loggerhead Shrike population, Avian Conservation and Ecology 2(2):11.
- Commission de coopération environnementale. 2008. La mosaïque nord-américaine : aperçu des principaux enjeux environnementaux, Secrétariat de la Commission de coopération environnementale.
- Coxon A., A.A. Chabot, S.C. Lougheed, J.A. Dávila et B.W. White. 2011. Characterization of 17 microsatellite loci from the Loggerhead Shrike (Lanius ludovicianus), Conservation Genetics Resources, DOI 10.1007/s12686-011-9585 2.
- Craig, R.B. 1978. An analysis of the predatory behavior of the Loggerhead Shrike, The Auk 95:221 234.
- Craig, S., et A. Chabot. 2012. A study of wintering Loggerhead Shrikes in the Texas Gulf Coastal Plain, Colorado Birds 46:121 126.
- Cuddy, D. 1995. Protection and restoration of breeding habitat for the Loggerhead Shrike (Lanius ludovicianus) in Ontario, Canada, p. 283 286 in R. Yosef et F.E. Lohrer (éd.), Shrikes (Laniidae) of the world: biology and conservation, Proceedings of the Western Foundation of Vertebrate Zoology 6.
- Cuddy, D., et B. Leviton. 1996. Habitat availability and supply analysis for Loggerhead Shrike in Ontario, Draft, rapport inédit pour le Ontario Ministry of Natural Resources.
- DeGeus, D.W. 1990. Productivity and habitat preferences of Loggerhead Shrikes inhabiting roadsides in a midwestern agro-environment, Master’s thesis, Iowa State University, Ames (Iowa).
- DeGeus, D.W., et L.B. Best. 1991. Brown-headed Cowbirds parasitize Loggerhead Shrikes: first records for family Laniidae, Wilson Bulletin 103:504 506.
- Didiuk, A., J. Conkin, K. Majewski, R. Greuel et D. Prescott. 2014. The 2013 Prairie Loggerhead Shrike Survey in Saskatchewan, Série de rapports techniques, Canadian Wildlife Service, Saskatoon (Saskatchewan).
- Dunning, J.B. Jr. 1993. CRC Handbook of Avian Body Masses, CRC Press, Boca Raton (Floride).
- Environnement Canada. 2010a. Programme de rétablissement de la Pie-grièche migratrice de la sous-espèce migrans (Lanius ludovicianus migrans) au Canada [Proposition], Loi sur les espèces en péril – Série de Programmes de rétablissement, Environnement Canada, Ottawa.
- Environnement Canada, 2010b. Site Internet du Relevé nord américain des oiseaux nicheurs – Résultats et analyses canadiens, version 3.00, Environnement Canada, Gatineau (Québec).
- Environnement Canada. 2013. Site Web du Relevé des oiseaux nicheurs de l’Amérique du Nord – Tendances démographiques au Canada, version des données de 2011, Environnement Canada, Gatineau (Québec) K1A 0H3.
- Erskine, A.J. 1992. Atlas of the Breeding Birds of the Maritime Provinces, Nimbus Publ., and the Nova Scotia Museum, Halifax (Nouvelle-Écosse).
- Esley, J.D., et E.K. Bollinger. 2001. Habitat selection and reproductive success of Loggerhead Shrikes in northwestern Missouri: A hierarchical approach, Wilson Bulletin 113:290 296.
- Etterson, M.A. 2003. Conspecific attraction in Loggerhead Shrikes: implications for habitat conservation and reintroduction, Biological Conservation 114:199 205.
- Etterson, M.A. 2004. Parentage in an Oklahoma population of Loggerhead Shrike assessed using nuclear microsatellites, Condor 106:401 404.
- Etterson, M.A., L.R. Nagy et T.R. Robinson. 2007. Partitioning risk among different causes of nest failure, The Auk 124:432 443.
- Flickinger, E.L. 1995. Loggerhead fatalities on a highway in Texas, p. 67-69 in R. Yosef et F.E. Lohrer (éd.), Shrikes (Laniidae) of the World: Biology and Conservation, Proceedings of the Western Foundation of Vertebrate Zoology 6.
- Fraser, J.D., et D.R. Luukkonen. 1986. The Loggerhead Shrike, p. 933 941 in R.L. DeSilvestro (éd.), Audubon Wildlife Report – 1986, National Audubon Society, New York (New York).
- Gauthier, J., et Y. Aubry (éd.). 1995. Les oiseaux nicheurs du Québec : atlas des oiseaux nicheurs du Québec méridional. Assoc. québécoise des groupes d’ornithologues, Société québécoise de protection des oiseaux, Service can. de la faune, Environnement Canada, Région du Québec, Montréal.
- Gawlik, D.E., et K.L. Bildstein. 1990. Reproductive success and nesting habitat of Loggerhead Shrikes in north-central South Carolina, Wilson Bulletin 102:37 48.
- Gawlik, D., et K.L. Bildstein. 1993. Seasonal habitat use and abundance of Loggerhead Shrikes in South Carolina, Journal Wildlife Management 57:352 357.
- Glynn-Morris, M. 2010. Breeding habitat selection by the Loggerhead Shrike in Ontario: a hierarchical analysis, B.Sc. Thesis, Department of Biology, Queen’s University, Ontario.
- Gouvernement de l’Ontario. 2007. Loi de 2007 sur les espèces en voie de disparition, disponible à Lois-en-ligne.
- Gouvernement du Québec. 2000. Décret 377-2000, 29 mars 2000, Loi sur les espèces menacées ou vulnérables (L.R.Q., c. E 12.01), Gazette officielle du Québec 12 avril 2000, 132e année, no 15.
- Grenier, M., B. Jobin et P. Laporte. 1999. A GIS approach to evaluating Loggerhead Shrike habitat availability in southern Québec, Canada, Picoides 12:6 7.
- Haas, C.A. 1987. Eastern subspecies of the Loggerhead Shrike: The need for measurements of live birds, North American Bird Bander 12:99 102.
- Haas, C.A., et S.L. Sloane. 1989. Low return rates of migratory Loggerhead Shrikes: winter mortality or low site fidelity? Wilson Bulletin 101:458 460.
- Hands, H.M., R.D. Drobney et M.R. Ryan. 1989. Status of the Loggerhead Shrike in the northcentral United States, Missouri Coop. Fish and Wildlife Research Unit, University of Missouri.
- Herkert, J. 2004. Organochlorine pesticides are not implicated in the decline of the Loggerhead Shrike, Condor 106:702 705.
- Hobson, K.A., et L.I. Wassenaar. 1997. Linking breeding and wintering grounds of Neotropical migrant songbirds using stable hydrogen isotopic analysis of feathers, Oecologia 109:142 148.
- Imlay, T.I, J.F. Crowley, A.M. Argue, J.C. Steiner, D.R. Norris et B.J.M. Stutchbury. 2010. Survival, dispersal and early migration movements of captive-bred juvenile eastern Loggerhead Shrikes (Lanius ludovicianus migrans), Biological Conservation 143:2578 2582.
- Imlay, T.I., et J. Lapierre. 2012. Population monitoring and field propagation and release programs, rapport inédit pour le Ontario Ministry of Natural Resources and Canadian Wildlife Service.
- Jobin, B., J.-L. DesGranges et C. Boutin. 1996. Population trends in selected species of farmland birds in relation to recent developments in agriculture in the St. Lawrence Valley, Agriculture, Ecosystems and Environment 57:103 116.
- Jobin, B, M. Grenier et P. Laporte. 2005. Using satellite imagery to assess breeding habitat availability of the endangered Loggerhead Shrike in Québec, Biodiversity and Conservation 14:81 95.
- Jobin, B., C. Latendresse, M. Grenier, C. Maisonneuve et A. Sebbane. 2010. Recent landscape change at the ecoregion scale in Southern Québec (Canada), 1993-2011, Environmental Monitoring and Assessment 164:631 647.
- Jones, Z.F., et C.E. Bock. 2002. Conservation of grassland birds in an urbanizing landscape: a historical perspective, Condor 104:643 651.
- Kiliaan, H., et D.R.C. Prescott. 2002. Survey of the Loggerhead Shrike in the Southern Aspen Parkland Region, 2000-2001, Alberta Sustainable Resource Development, Fish and Wildlife Division, Alberta Species at Risk Report No. 46, Edmonton (Alberta).
- Kridelbaugh, A.L. 1982. An ecological study of Loggerhead Shrikes in central Missouri, mémoire de maîtrise ès sciences, University of Missouri, Columbia.
- Lefranc, N. 1997. Shrikes: A Guide to Shrikes of the World, Yale University Press, New Haven (Connecticut).
- Lindgren, C.J., R. Postey, K. De Smet, C. Higgs et A.B. Thompon. 2009. West Nile virus as a cause of death among endangered eastern Loggerhead Shrikes, Lanius ludovicianus migrans, in West St. Paul, Manitoba, Canadian Field Naturalist 123:7 11.
- Lohrer, F.E. 1974. Post-hatching growth and development of the Loggerhead Shrike in Florida, mémoire de maîtrise ès sciences, University of South Florida, Tampa.
- Luukkonen, D.R. 1987. Status and breeding ecology of the Loggerhead Shrike in Virginia, Master’s thesis, Virginia Polytechnique Institute and State University, Blacksburg (Virginie).
- Lymn, N., et S.A. Temple. 1991. Land-use changes in the Gulf coast region: links to declines in midwestern Loggerhead Shrike populations, Passenger Pigeon 53:315 325.
- Michaels, H.L., et J.F. Cully. 1998. Landscape and fine scale habitat associations of the Loggerhead Shrike, Wilson Bulletin 110:474 482.
- Miller, A.H. 1928. The molts of the Loggerhead Shrike Lanius ludovicianus Linneaus, University of California Publications in Zoology 30:393 417.
- Miller, A.H. 1931. Systematic revision and natural history of the American shrikes (Lanius), University of California Publications in Zoology 38:11 242.
- Miller, A.H. 1951. A comparison of the avifaunas of Santa Cruz and Santa Rosa islands, California, Condor 53:117 123.
- Mundy, N.I., C.S. Winchell, T. Burr et D.S. Woodruff. 1997. Microsatellite variation and microevolution in the critically endangered San Clemente Island Loggerhead Shrike (Lanius ludovicianus mearnsi), Proceedings of the Royal Society of London Series B 264:869 875.
- Novak, P.G. 1989. Breeding ecology and status of the Loggerhead Shrike (Lanius ludovicianus) in New York state, mémoire de maîtrise ès sciences, Cornell University, Ithaca (New York).
- Nur, N., A.L. Holmes et G.R. Geupel. 2004. Use of survival time analysis to analyze nesting success in birds: an example using Loggerhead Shrikes, Condor 106:457 471.
- Okines, D., et J. McCracken. 2003. Loggerhead Shrike Banding in Ontario: Report on the 2003 Field Season, rapport inédit prepared for the Canadian Wildlife Service.
- Okines, D., et J. McCracken. 2005. Loggerhead Shrike Banding in Ontario: Report on the 2004 Field Season, rapport inédit prepared for the Canadian Wildlife Service.
- Peakall, R., et P.E. Smouse. 2006. GENALEX 6: Genetic Analysis in Excel, Population genetic software for teaching and research, Molecular Ecology Notes 6:288 295.
- Peck, G.K., et R.D. James. 1987. Breeding Birds of Ontario Nidiology and Distribution, Volume 2: Passerines, Royal Ontario Museum.
- Pérez, G.E. 2006. Migratory connectivity and wintering habitat structure of Loggerhead Shrikes: inferences from stable hydrogen isotope and microsatellite DNA analysis, mémoire de maîtrise ès sciences, Department of Biology, University of Saskatchewan (Saskatchewan).
- Pérez, G., et K.A. Hobson. 2006. Isotopic evaluation of interrupted molt of northern breeding populations of the Loggerhead Shrike, Condor 108:877 886.
- Pérez, G., et K.A. Hobson. 2007. Feather deuterium measurement reveal origins of migratory western Loggerhead Shrikes (Lanius ludovicianus excubitorides) wintering in Mexico, Diversity and Distributions 13:166 171.
- Pérez, G.E., et K.A. Hobson. 2009. Winter habitat use by Loggerhead Shrikes (Lanius ludovicianus) in Mexico: separating migrants from residents using stable isotopes, Journal of Field Ornithology 150:459 467.
- Peterjohn, B.G., et J.R. Sauer. 1995. Population trends of the Loggerhead Shrike from the North American Breeding Bird Survey, p. 117 121 in R. Yosef et F.E. Lohrer (éd.), Shrikes (Laniidae) of the World: Biology and Conservation, Proceedings of the Western Foundation of Vertebrate Zoology 6.
- Phillips, A.R. 1986. The Known Birds of North and Middle America, Part I, Privately published, Denver (Colorado).
- Porter, D.K., M.A. Strong, J.B. Giezentanner et R.A. Ryder. 1975. Nest ecology, productivity and growth of the Loggerhead Shrike on the shortgrass prairie, Southwestern Naturalist 19:429 436.
- Prescott, D.R.C. 2003. The use of call playbacks for censusing Loggerhead Shrikes in southern Alberta, Alberta Sustainable Resource Development, Fish and Wildlife Division, Alberta Species at Risk Report No. 67, Edmonton (Alberta).
- Prescott, D.R.C. 2013. The 2013 Loggerhead Shrike survey in Alberta, Alberta Environment and Sustainable Resource Development, Fish and Wildlife Policy Branch, Alberta Species at Risk Report No. 151, Edmonton (Alberta).
- Prescott, D.R.C., et R.R. Bjorge. 1999. Status of the Loggerhead Shrike (Lanius ludovicianus) in Alberta, Alberta Natural Resources Services, Edmonton (Alberta).
- Prescott, D.R.C., et D.M. Collister. 1993. Characteristics of occupied and unoccupied Loggerhead Shrike territories in southeastern Alberta, Journal of Wildlife Management 57:346 352.
- Pruitt, L. 2000. Loggerhead Shrike Status Assessment, United States Fish and Wildlife Service, Fort Snelling (Minnesota).
- Rand, A.L. 1960. Family Laniidae, p. 309 365 in E. Mayr et J.C. Greenway, Jr. (éd.), Check-list of Birds of the World, Vol. 9. Museum of Comparative Zoology, Cambridge (Massachusetts).
- Reschke, C., R. Reid, J. Jones, T. Feeney et H. Potter. 1999. Conserving Great Lakes Alvars, The Nature Conservancy, Great Lakes Program, Chicago (Illinois).
- Robbins, S.D. 1991. Wisconsin Birdlife, University of Wisconsin Press, Madison.
- Robbins, C.S., D. Bystrak et P.H. Geissler. 1986. The Breeding Bird Survey: Its First Fifteen Years, 1965-1979, U.S. Fish and Wildlife Service Resource Publication.
- Robert, M., et P. Laporte. 1991. Situations historique et actuelle de la Pie-grièche migratrice au Québec, Service canadien de la faune, Cahiers de biologie no 196.
- Rudd, P.L., R.B. Craig et W.S. Williams. 1981. Trophic accumulation of DDT in a terrestrial food web, Environmental Pollut. Serv. A Ecol. Biol. 25:219 228.
- Salt, W.R., et A.L. Wilk. 1958. The Birds of Alberta, Queen’s Printer, Edmonton (Alberta).
- Samson, F.B., and F.L. Knopf. 1994. Prairie conservation in North America, BioScience 44:418 421.
- Samson, F.B., F.L. Knopf et W.R. Ostlie. 2004. Great Plains ecosystems: past, present and future, Wildlife Society Bulletin 32:6 15.
- Sauer, J.R., J.E. Hines, J.E. Fallon, K.L. Pardieck, D.J. Ziolkowski, Jr. et W.A. Link. 2011. The North American Breeding Bird Survey, Results and Analysis 1966 – 2010, Version 12.07.2011 USGS Patuxent Wildllife Research Center, Laurel (Maryland).
- Scott, T.A., et M.L. Morrison. 1990. Natural history and management of the San Clemente Loggerhead Shrike, Proceedings of the Western Foundation of Vertebrate Zoology 4:23 57.
- Scott, T.A., et M.L. Morrison. 1995. Opportunistic foraging behavior of Loggerhead Shrikes, p. 186 193 in R. Yosef et F.E. Lohrer (éd.), Shrikes (Laniidae) of the World: Biology and Conservation, Proceedings of the Western Foundation of Vertebrate Zoology 6.
- Siegel, M.S. 1980. The nesting ecology and population dynamics of the Loggerhead Shrike in the blackbelt of Alabama, mémoire de maîtrise ès sciences, University of Alabama, Tuscaloosa.
- Smith, S.M. 1972. The ontogeny of impaling behaviour in the Loggerhead Shrike, Lanius ludovicianus, Behaviour 42:232 247.
- Smith, E.L. 1990. Factors influencing distribution and abundance of the Loggerhead Shrike (Lanius ludovicianus) in south-central Illinois, rapport inédit from Eastern Illinois University, Environmental Biology Program, Charlestown (Illinois).
- Smith, A.R. 1996. Atlas of Saskatchewan Birds, Environment Canada and Nature Saskatchewan, Regina.
- Smith, K. 1991. Brown Thrashers harass fledgling Loggerhead Shrikes, Bulletin of the Oklahoma Ornithological Society 24:34 35.
- Soendjoto, M.A. 1995. Vocalization behaviour of captive Loggerhead Shrikes (Lanius ludovicianus excubitorides), mémoire de maîtrise ès sciences, Macdonald Campus of McGill University, Sainte Anne-de-Bellevue (Québec).
- SOS-POP. 2011. Banque de données sur populations d’oiseaux en situation précaire au Québec [version du 30 novembre 2011], Regroupment Québec Oiseaux, Montréal (Québec).
- St. Laurent, K., A. Didiuk, L. Craig-Moore et B. Johns. 2013. Prairie Loggerhead Shrike (Lanius ludovicianus excubitorides) Saskatchewan five-year survey summary: 1987 2013, rapport inédit, Canadian Wildlife Service, Saskatoon (Saskatchewan).
- St. Louis, V., A.M. Pidgeon, M.K. Clayton, B.A. Locke, D. Bash et V.C. Radeloff. 2010. Habitat variables explain Loggerhead Shrike occurrence in the northern Chihuahuan Desert but are poor correlates of fitness measures, Landscape Ecology 25:643 654.
- Stewart, L. 2013. Nos prairies en péril, BirdWatch Canada 65:6 7.
- Telfer, E.S. 1992. Habitat change as a factor in the decline of the western Canadian Loggerhead Shrike, Lanius ludovicianus, population, Canadian Field Naturalist 106:321 326.
- Telfer, E.S, C. Adam, K. DeSmet et R. Wershler. 1989. Situation et distribution de la Pie grièche migratrice dans l’ouest du Canada, Service canadien de la faune, Cahiers de biologie no 184.
- Temple, S.A. 1995. Shrike research and conservation, p. 296 298 in R. Yosef et F.E. Lohrer (éd.), Shrikes (Laniidae) of the World: Biology and Conservation, Proceedings of the Western Foundation of Vertebrate Zoology 6.
- Tischendorf, L. 2009. Population viability analysis of the eastern Loggerhead Shrike (Lanius ludovicianus migrans), rapport inédit pour le Canadian Wildlife Service – Ontario, Environment Canada.
- United States Fish and Wildlife Service (USFWS). 1984. California Channel Islands Species Recovery Plan.
- United States Fish and Wildlife Service (USFWS). 1987. Migratory nongame birds of management concern in the U.S.: the 1987 list, U.S. Fish and Wildlife Service, Office of Migratory Bird Management, Washington D.C.
- United States Fish and Wildlife Service (USFWS). 1995. Migratory nongame birds of management concern in the U.S.: the 1995 list, U.S. Fish and Wildlife Service, Office of Migratory Bird Management, Washington D.C.
- Vallianatos, M., S.C. Lougheed et P.T. Boag. 2001. Phylogeography and genetic characteristics of a putative secondary-contact zone of the loggerhead shrike in central and eastern North America, Canadian Journal of Zoology 79:2221 2227.
- Vallianatos, M., S.C. Lougheed et P.T. Boag. 2002. Conservation genetics of the Loggerhead Shrike (Lanius ludovicianus) in central and eastern North America, Conservation Genetics 3:1 13.
- Walk, J.W., E.L. Kershner et R.E. Warner. Low nesting success of Loggerhead Shrikes in an agricultural landscape, Wilson Journal of Ornithology 118:70 74.
- Woods, C.P. 1994. The Loggerhead Shrike in southwest Idaho, Master’s thesis, Boise State University, Boise (Idaho).
- Wright, S. 1943. Isolation by distance, Genetics 28:114 138.
- Yosef, R. 1992. Behaviour of polygynous and monogamous Loggerhead Shrikes and a comparison with Northern Shrikes, Wilson Bulletin 104:747 749.
- Yosef, R. 1994. The effects of fencelines on the reproductive success of Loggerhead Shrikes, Conservation Biology 8:281 285.
- Yosef, R. 1996. Loggerhead Shrike (Lanius ludovicianus), in The Birds of North America, No. 231 (A. Poole et F. Gill, éd.), The Academy of Natural Science, Philadelphia (Pennsylvanie) et The American Ornithological Union, Washington D.C.
- Yosef, R., et T.C. Grubb, Jr. 1993. Effect of vegetation height on hunting behavior and diet of Loggerhead Shrikes, Condor 95:127 131.