
Évaluation et Rapport de situation du COSEPAC sur la Raie tachetée Leucoraja ocellata au Canada - 2015
Population du golfe du Saint-Laurent
Population de l'est du plateau néo-écossais et de Terre-Neuve
Population de l'ouest du plateau néo-écossais et du banc Georges

Photo © C. Miri
Description longue pour la figure 1
Photo d’une raie tachetée (Leucoraja ocellata), vue dorsale. La raie tachetée est un poisson en forme de disque dont le museau très arrondi se termine par un angle obtus et porte deux taches opaques de cartilage. La queue est mince et arrondie et comprend deux petites nageoires à l’extrémité. La surface dorsale (disque supérieur) est brune et présente des taches arrondies foncées et, généralement, un ocelle ou plus près du bord postérieur de chaque nageoire pectorale. Les yeux se trouvent sur la surface dorsale, tandis que les branchies et la bouche sont sur la surface ventrale.
- Table des matières
- Sommaire de l’évaluation
- Résumé
- Résumé technique – Population du golfe du Saint Laurent
- Résumé technique – Population de l'est du plateau néo écossais et de Terre Neuve
- Résumé technique – Population de l'ouest du plateau néo écossais et du banc Georges
- Préface
- Description et importance de l’espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances de la population
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d’information
- Sommaire biographique des rédactrices du rapport
- Figure 1. Dessin de la raie tachetée (Leucoraja ocellata) tiré de Scott et Scott (1988)
- Figure 2. Carte de référence montrant la bathymétrie (100, 200, 500 m de contour), les limites de l’OPANO et la toponymie mentionnées dans le présent rapport. La ligne foncée dans le coin inférieur droit représente la frontière entre le Canada et les États-Unis
- Figure 3. Lieux de capture de raies tachetées lors de relevés au chalut de fond du MPO dans les divisions 4VWX (juillet), 4T (septembre), 4RS (août), 3LNOP (printemps) et 5Z (printemps). Les données datent de 1970 à 2013, et les raies de toutes tailles sont représentées. Les données sur les captures ont été normalisées en fonction du nombre de prises par 1,75 mille marin, et les valeurs moyennes de la série chronologique ont été calculées selon une grille de 10’ de latitude sur 10’ de longitude. Les lignes foncées représentent les limites proposées de l’unité désignable (UD). La région sud-ouest constitue l’UD de l’ouest du plateau néo-écossais et du banc Georges. La ligne traversant le banc Georges montre la frontière entre le Canada et les États-Unis. La région sud-est représente l’UD de l’est du plateau néo-écossais et de Terre-Neuve, et la région nord, l’UD du golfe du Saint-Laurent
- Figure 4. Lieux de capture de raies tachetées dans les environs du chenal Laurentien, dans l’UD de l’est du plateau néo-écossais et de Terre-Neuve, lors du relevé du MPO sur le plateau néo-écossais (en juillet) et du relevé des divisions 3LNOP (au printemps). Les données datent de 1970 à 2013, et les raies de toutes tailles sont représentées. Le secteur non recensé dans le chenal Laurentien sert de démarcation entre les zones couvertes par les deux relevés
- Figure 5. Distribution des longueurs de raies tachetées mesurées lors A) du relevé printanier à Terre-Neuve (partie de l’UD de l’est du plateau néo-écossais et de Terre-Neuve) et B) du relevé dans le nord du golfe du Saint-Laurent en août (partie de l’UD du golfe du Saint-Laurent). Un intervalle de longueur de 5 cm a été utilisé, et les valeurs sur l’axe des x correspondant à la valeur médiane de l’intervalle
- Figure 6. Distribution des longueurs de raies tachetées mesurées lors A) du relevé printanier à Terre-Neuve (partie de l’UD de l’est du plateau néo-écossais et de Terre-Neuve) et B) du relevé dans le nord du golfe du Saint-Laurent en août (partie de l’UD du golfe du Saint-Laurent). Un intervalle de longueur de 5 cm a été utilisé, et les valeurs sur l’axe des x correspondant à la valeur médiane de l’intervalle
- Figure 7. Tendances des estimations réalisées lors de relevés au chalut de fond de l’abondance des raies tachetées matures dans le sud du golfe du Saint-Laurent (1971-2013). Le graphique du haut présente les résultats sur une échelle arithmétique. La ligne continue représente la régression log-linéaire ajustée. Dans le graphique du bas, les estimations de l’abondance sont représentées à l’échelle logarithmique naturelle, et la ligne continue est la régression ajustée. Les points de données pleins proviennent des 3 générations les plus récentes (30 ans). Aucune raie tachetée mature n’a été capturée en 2013; aux fins de l’analyse de régression, on a utilisé la valeur de l’estimation la plus faible qui n’est pas 0 (2011)
- Figure 8. Tendances de la zone du relevé occupée par la raie tachetée (km²) dans le sud du golfe du Saint-Laurent (1971-2013). La ligne ajustée, qui résulte d’un lissage localement pondéré (Lowess), est utilisée ici pour montrer la tendance générale
- Figure 9. Tendances des estimations réalisées lors de relevés au chalut de fond de l’effectif des raies tachetées matures dans l’est du plateau néo-écossais, dans 4VW de l’OPANO (1970-2013). Ce secteur fait partie de l’UD de l’est du plateau néo-écossais et de Terre-Neuve. Le graphique du haut présente les résultats sur une échelle arithmétique. La ligne continue représente la régression log-linéaire ajustée. Dans le graphique du bas, les estimations de l’effectif sont représentées à l’échelle logarithmique naturelle, et la ligne continue est la régression ajustée. Aucune raie tachetée mature n’a été capturée en 2001, 2002, 2008 et 2013. Aux fins de l’analyse de régression, on a utilisé la valeur de l’estimation la plus faible qui n’est pas 0 (2012)
- Figure 10. Tendances de la zone du relevé occupée par la raie tachetée (km²) dans l’est du plateau néo-écossais (1970-2013), qui fait partie de l’UD de l’est du plateau néo-écossais et de Terre-Neuve. La ligne ajustée, qui résulte d’un lissage localement pondéré (Lowess), est utilisée ici pour montrer la tendance générale.
- Figure 11. Tendances des estimations réalisées lors de relevés au chalut de fond de l’effectif des raies tachetées au sud de Terre-Neuve, dans 3LNOP de l’OPANO (1971-2013). Ce secteur fait partie de l’UD de l’est du plateau néo-écossais et de Terre-Neuve. La série chronologique du printemps est divisée en trois parties, représentées par des symboles différents. Durant la première partie, on a utilisé un chalut Yankee, durant la deuxième (1983-1995), un chalut Engel, et durant la troisième (1996-2013), un chalut Campelen. Le graphique du haut présente les résultats sur une échelle arithmétique. La ligne continue représente la régression log-linéaire ajustée pour la période où on utilisait le chalut Campelen. Dans le graphique du bas, les estimations de l’effectif sont représentées à l’échelle logarithmique naturelle, et la ligne continue est la régression ajustée. Aucune raie tachetée n’a été capturée en 2011 et en 2013. Aux fins de l’analyse de régression, on a utilisé la valeur de l’estimation la plus faible qui n’est pas 0 (2012)
- Figure 12. Tendances des estimations réalisées lors de relevés au chalut de fond de l’effectif des raies tachetées matures dans l’ouest du plateau néo-écossais et la baie de Fundy, dans 4X de l’OPANO (1970-2013). Ce secteur zone fait partie de l’UD de l’ouest du plateau néo-écossais et du banc Georges. Le graphique du haut présente les résultats sur une échelle arithmétique. La ligne continue représente la régression log-linéaire ajustée. Dans le graphique du bas, les estimations de l’effectif sont représentées à l’échelle logarithmique naturelle, et la ligne continue est la régression ajustée. Aucune raie tachetée mature n’a été capturée en 1971. Aux fins de l’analyse de régression, on a utilisé la valeur de l’estimation la plus faible qui n’est pas 0 (2002)
- Figure 13. Tendances de la zone du relevé occupée par la raie tachetée (km²) dans l’ouest du plateau néo-écossais et la baie de Fundy (1970-2013), qui fait partie de l’UD de l’ouest du plateau néo-écossais et du banc Georges. La ligne ajustée, qui résulte d’un lissage localement pondéré (Lowess), est utilisée ici pour montrer la tendance générale.
- Figure 14. Tendances des estimations réalisées lors de relevés canadiens au chalut de fond de l’effectif des raies tachetées matures dans la portion canadienne du banc Georges (1987-2012). Ce secteur zone fait partie de l’UD de l’ouest du plateau néo-écossais et du banc Georges. Le graphique du haut présente les résultats sur une échelle arithmétique. La ligne continue représente la régression log-linéaire ajustée. Dans le graphique du bas, les estimations de l’effectif sont représentées à l’échelle logarithmique naturelle, et la ligne continue est la régression ajustée
- Figure 15. Tendances de la zone du relevé occupée par la raie tachetée (km²) sur le banc Georges (1987-2012), qui fait partie de l’UD de l’ouest du plateau néo-écossais et du banc Georges. La ligne ajustée, qui résulte d’un lissage localement pondéré (Lowess), est utilisée ici pour montrer la tendance générale
- Figure 16. Indices de biomasse (kg/trait) de la raie tachetée (1967-2013) dans les eaux états-uniennes, selon les relevés automnaux effectués par le Northeast Fisheries Science Center (d’après la figure 31 de Sosebee, 2014)
- Figure 17. Rejets estimés de raies tachetées par les pêches des poissons de fond dans le sud du golfe du Saint-Laurent par classes de taille (juvéniles : 21-41 cm; adultes : 42 cm et plus) (source : figure 22 de Swain et al., 2006)
- Figure 18. Prises estimées de raies tachetées dans l’est du plateau néo-écossais, 1970-2004 (source : annexe D1 de Swain et al., 2006)
COSEPAC
Comité sur la situation
des espèces en péril
au Cananda

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages que l’on croit en péril. On peut citer le présent rapport de la façon suivante :
COSEPAC. 2015. Évaluation et Rapport de situation du COSEPAC sur la raie tachetée (Leucoraja ocellata), population du golfe du Saint-Laurent, population de l'est du plateau néo-écossais et de Terre-Neuve et population de l'ouest du plateau néo-écossais et du banc Georges au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xviii + 49 p. Visitez le site Registre public des espèces en péril.
COSEPAC 2005. Évaluation et Rapport de situation du COSEPAC sur la raie tachetée (Leucoraja ocellata) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. viii + 47 p. Visitez le site Registre public des espèces en péril.
Le COSEPAC remercie Alan Sinclair d’avoir rédigé le rapport de situation sur la raie tachetée (Leucoraja ocellata), population du sud du golfe Saint Laurent, population de l’est du plateau néo-écossais et population de l’ouest du plateau néo-écossais et du banc Georges au Canada, aux termes d’un marché conclu avec Environnement Canada. La supervision et la révision du rapport ont été assurées par John Reynolds, coprésident du Sous-comité de spécialistes des poissons marins.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site web : COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Winter Skate Leucoraja ocellata, Gulf of St. Lawrence population, Eastern Scotian Shelf - Newfoundland population and Western Scotian Shelf - Georges Bank population in Canada. Committee on the Status of Endangered Wildlifein Canada.
Raie tachetée -- Photo : C. Miri, Pêches et Océans Canada, Région de Terre-Neuve-et-Labrador.
Population du sud du golfe Saint-Laurent
Population de l'est du plateau néo-écossais
Population de l'ouest du plateau néo-écossais et du banc Georges
La raie tachetée (Leucoraja ocellata Mitchill 1815) se distingue de toutes les autres espèces de raies, à l’exception de la raie hérisson (Leucoraja erinacea), espèce qui lui est sympatrique, par les caractéristiques suivantes : museau très arrondi se terminant par un angle obtus et portant deux taches opaques de cartilage; présence de trois rangées ou plus d’épines sur la queue et le haut du corps (disque), depuis la région scapulaire jusqu’à la naissance de la première nageoire dorsale; taches arrondies foncées sur la partie postérieure du disque; généralement, un ocelle ou plus près du bord postérieur de chaque nageoire pectorale. Les spécimens dépourvus de ces taches ressemblent tellement à la raie hérisson lorsqu’ils sont petits (< 35 cm) qu’un simple examen visuel ne permet pas de les distinguer de cette espèce. Le nombre de rangées de dents sur la mâchoire supérieure, variable selon la taille de l’individu, est plus élevé chez la raie tachetée que chez la raie hérisson. Le nombre d’épines de la rangée médiocaudale est également plus élevé chez la raie tachetée (généralement > 21).
La raie tachetée est endémique dans l’Atlantique Nord-Ouest, où on la rencontre depuis le golfe du Saint-Laurent et les eaux méridionales de Terre-Neuve jusqu’au cap Hatteras, en Caroline du Nord. Dans les eaux canadiennes, l’espèce se concentre dans trois zones : le golfe du Saint-Laurent; l’est du plateau néo-écossais et le sud de Terre-Neuve; l’ouest du plateau néo-écossais, la baie de Fundy et la portion canadienne du banc Georges. Ces populations sont séparées par des distances considérables, ce qui laisse croire qu’elles sont distinctes. Elles montrent également des variations des caractères héréditaires qui sont indicatrices d’adaptations importantes. Ainsi, on propose trois unités désignables (UD) distinctes dans le présent rapport :
- UD du golfe du Saint-Laurent;
- UD de l’est du plateau néo-écossais;
- UD de l’ouest du plateau néo-écossais et du banc Georges.
La raie tachetée est une espèce benthique vivant généralement sur les fonds de sable ou de gravier. On la retrouve à des profondeurs pouvant atteindre 371 m, mais principalement dans des eaux de moins de 111 m. La plupart des raies tachetées sont capturées dans des eaux variant de 5 à 16 °C (plage de -1,2 à 19 °C).
Hors du golfe du Saint-Laurent, les raies tachetées atteignent la maturité à une longueur totale de 75 cm ou à 13 ans. Dans le golfe, la maturité est atteinte à 42 cm ou à 5 ans. On estime la durée d’une génération à 18 ans hors du golfe et à 10 ans dans le golfe.
La taille et les tendances des populations ont été estimées d’après des relevés indépendants des pêches. Au cours des 3 dernières générations, les raies tachetées matures du golfe du Saint-Laurent auraient subi un déclin de 99 %. Les données disponibles provenant des relevés réalisées dans l’est du plateau néo-écossais couvrent les 2,4 dernières générations, durant lesquelles le nombre d’individus matures aurait diminué de 98 %. On a noté une tendance à la baisse peu importante dans l’ouest du plateau néo-écossais au cours des 2,4 dernières générations ainsi qu’une tendance à la hausse, encore une fois peu importante, dans la portion canadienne du banc Georges, où se trouve la plus grande partie de la population. Dans l’ensemble, on n’observe aucune tendance discernable au sein de la population de l’ouest du plateau néo-écossais et du banc Georges.
Certaines caractéristiques du cycle vital des raies tachetées, comme la maturité tardive et les faibles taux de reproduction, les rendent vulnérables à la surexploitation, réduisent le taux de rétablissement et augmentent le risque de disparition. La surpêche explique probablement une partie du déclin de l’espèce dans le golfe du Saint-Laurent et l’est du plateau néo-écossais, mais ce sont les taux anormalement élevés de mortalité due à des causes autres que les pêches, possiblement le nombre de plus en plus élevé de phoques gris (Halichoerus grypus), qui menacent la viabilité de ces deux populations de raies tachetées. Les prises accessoires d’autres pêches ont été grandement réduites, mais leurs conséquences actuelles sur les populations très diminuées du golfe du Saint-Laurent et de l’est du plateau néo-écossais sont inconnues.
La raie tachetée est endémique dans l’Atlantique Nord-Ouest, et une portion considérable de l’aire de répartition se trouve dans les eaux canadiennes. L’âge à maturité et la répartition par taille uniques des raies tachetées du golfe du Saint-Laurent donnent à penser que cette population forme en réalité une espèce distincte, mais cela reste à démontrer.
En 2005, le COSEPAC a évalué la raie tachetée et défini quatre UD séparées : 1. UD du sud du golfe du Saint-Laurent (« en voie de disparition »); UD de l’est du plateau néo-écossais (« menacée »); 3. UD de l’ouest du plateau néo-écossais, de la baie de Fundy et du banc Georges (« préoccupante »); UD du nord du golfe du Saint-Laurent et de Terre-Neuve (« données insuffisantes »). Le gouverneur en conseil a décidé de ne pas inscrire la raie tachetée à l’annexe de la Loi sur les espèces en péril (LEP). La mortalité des raies tachetées due à des causes anthropiques est plutôt réglementée par Pêches et Océans Canada aux termes de la Loi sur les pêches. La pêche dirigée des raies tachetées est interdite au Canada, et toutes les raies faisant partie des prises accessoires doivent être remises à l’eau. Le taux de survie des individus rejetés est inconnu. L’Union internationale pour la conservation de la nature (UICN) évalue que la raie tachetée est en voie de disparition dans toute son aire de répartition. En 2015, le COSEPAC a évalué la raie tachetée selon trois UD séparées, soit la population du golfe du Saint-Laurent (« en voie de disparition »), la population de l’est du plateau néo-écossais et de Terre-Neuve (« en voie de disparition ») et la population de l’ouest du plateau néo-écossais et du banc Georges (« non en péril »).
Durée d’une génération: 10 ans
Durée d’une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d’estimation de la durée d’une génération autre que celle qui est présentée dans les lignes directrices de l’UICN [2008] est utilisée)
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d’individus matures?
Oui, observé
Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations] (52-60 ans).
Déclin de 97 % sur 2 générations
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations] (78-90 ans).
Déclin de 99 % sur 3 générations
Pourcentage [prévu ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations] (78-90 ans).
Non estimé, mais on s’attend à ce que le déclin continue.
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur.
Déclin de 99 %
Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé?
Non. Pas clairement réversibles et comprises, et n’ont pas cessé
Y a-t-il des fluctuations extrêmes du nombre d’individus matures?
Non
Superficie estimée de la zone d’occurrence Tirée de COSEPAC (2004)
161 515 km²
Indice de zone d’occupation (IZO; valeurs d’une grille à carrés de 2 km de côté)
380 km² selon la zone d’occupation pondérée par l’échantillonnage obtenue au cours des 5 dernières années de relevés par navire de recherche
La population totale est-elle gravement fragmentée?
Non
Nombre de localités (selon le nombre de menaces)
Nombre de localités incertain. Possiblement 1 puisque la prédation par le phoque gris pourrait être responsable d'un déclin de l'effectif de 25 % par année
Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d’occurrence?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de l’indice de zone d’occupation?
Oui
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l’étendue ou la qualité] de l’habitat?
Non
Y a-t-il des fluctuations extrêmes du nombre de populations?
Non
Y a-t-il des fluctuations extrêmes du nombre de localités?
Non
Y a-t-il des fluctuations extrêmes de la zone d’occurrence?
Non
Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation?
Non
Nombre d’individus matures (mâles et femelles)
Total: 6 300 (estimation de la population minimale d’après les 5 dernières années de relevés par navire de recherche). La population réelle est vraisemblablement plus grande.
La probabilité de disparition de l’espèce à l’état sauvage est d’au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans].
L'analyse n'a pas été effectuée
La principale menace est le taux élevé de mortalité due à des causes autres que la pêche, possiblement causée par la prédation par les phoques gris de plus en plus nombreux dans cette zone. Il y a également une possibilité de mortalité attribuable aux rejets des prises accessoires de la pêche ciblant d’autres espèces.
Situation des populations de l’extérieur?
Aucune population états-unienne contiguë
Une immigration a-t-elle été constatée ou est-elle possible?
Non
Des individus immigrants seraient-ils adaptés pour survivre au Canada?
Oui
Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants dans l’estuaire du Saint-Laurent?
Oui
La possibilité d’une immigration depuis des populations externes existe-t-elle?
Non
L’information concernant l’espèce est-elle de nature délicate?
Non
COSEPAC : La population du sud du golfe du Saint-Laurent de la raie tachetée a été évaluée en « voie de disparition » en mai 2005, et la population du nord du Golfe et de Terre-Neuve a été évaluée comme faisant partie de la catégorie « données insuffisantes » en mai 2005. Les Lignes directrices pour reconnaître les unités désignables du COSEPAC (2013) ont été utilisées pour réviser la structure des populations pour l’évaluation de 2015, résultant ainsi en de nouvelles unités désignables. La nouvelle population du golfe du Saint-Laurent est composée de la population précédente du sud du golfe du Saint-Laurent et de parties de la population précédente du nord du Golfe et de Terre-Neuve. Les sous-populations restantes de la population du nord du Golfe et de Terre-Neuve ont été assignées à la nouvelle population de l’est du plateau néo-écossais et de Terre-Neuve. La population du golfe du Saint-Laurent a été désignée « en voie de disparition » en mai 2015.
Durée d'une génération: 18 ans
Durée d'une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN [2008] est utilisée)
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures?
Oui
Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] (52-60 ans).
Déclin de 97 % sur 2 générations
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations] (78-90 ans).
Déclin de 98 % sur 2,4 générations
Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations] (78-90 ans).
Non estimé
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur.
Déclin de 98 %. Le déclin observé au cours des 2,4 dernières générations est continu et devrait se poursuivre dans le futur
Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé?
Non. Pas clairement réversibles et comprises, et n’ont pas cessé.
Y a-t-il des fluctuations extrêmes du nombre d'individus matures?
Non
Superficie estimée de la zone d'occurrence Tirée de COSEPAC (2004)
215 436 km²
Indice de zone d'occupation (IZO; valeurs d'une grille à carrés de 2 km de côté)
5 200 km² selon la zone d’occupation pondérée par l’échantillonnage obtenue au cours des 5 dernières années de relevés par navire de recherche
La population totale est-elle gravement fragmentée?
Non
Nombre de localités (selon le nombre de menaces)
Probablement > 10 en raison des prédateurs et des prises accessoires de multiples types de pêches
Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation?
Oui
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat?
Non
Y a-t-il des fluctuations extrêmes du nombre de populations?
Non
Y a-t-il des fluctuations extrêmes du nombre de localités?
Non
Y a-t-il des fluctuations extrêmes de la zone d'occurrence?
Non
Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation?
Non
- Population: Est du plateau néo-écossais
27 000 (estimation de la population minimale d’après les dernières années de relevés par navire de recherche) - Population Sud de Terre-Neuve
77 000 (estimation de la population minimale d’après les 5 dernières années de relevés par navire de recherche) - Population Total : 104 000 (population réelle vraisemblablement plus grande)
La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans].
L'analyse n'a pas été effectuée
La principale menace est le taux élevé de mortalité due à des causes autres que les pêches, possiblement causée par la prédation par les phoques gris de plus en plus nombreux dans cette zone. Il y a également une possibilité de mortalité attribuable aux rejets des prises accessoires des pêches ciblant d’autres espèces.
Situation des populations de l'extérieur?
Aucune population états-unienne contiguë
Une immigration a-t-elle été constatée ou est-elle possible?
Non
Des individus immigrants seraient-ils adaptés pour survivre au Canada?
Oui
Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants dans l'estuaire du Saint-Laurent?
Oui
La possibilité d'une immigration depuis des populations externes existe-t-elle?
Non
L'information concernant l'espèce est-elle de nature délicate?
Non
COSEPAC : La population de l’est du plateau néo-écossais de la raie tachetée a été évaluée « menacée » en mai 2005, et la population du nord du Golfe et de Terre-Neuve a été évaluée comme faisant partie de la catégorie « données insuffisantes » en mai 2005. Les Lignes directrices pour reconnaître les unités désignables du COSEPAC (2013) ont été utilisées pour réviser la structure des populations pour l’évaluation de 2015, résultant ainsi en de nouvelles unités désignables. La nouvelle population de l’est du plateau néo-écossais et de Terre-Neuve est composée de la population précédente de l’est du plateau néo-écossais et de parties de la population précédente du nord du Golfe et de Terre-Neuve. Les sous-populations restantes de la population du nord du Golfe et de Terre-Neuve ont été assignées à la nouvelle population du golfe du Saint-Laurent. La population de l’est du plateau néo-écossais et de Terre-Neuve a été désignée « en voie de disparition » en mai 2015.
Durée d'une génération: 18 ans
Durée d'une génération (généralement, âge moyen des parents dans la population; indiquer si une méthode d'estimation de la durée d'une génération autre que celle qui est présentée dans les lignes directrices de l'UICN [2008] est utilisée)
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre total d'individus matures?
Non
Pourcentage estimé de déclin continu du nombre total d'individus matures sur [cinq ans ou deux générations] (52-60 ans).
Aucun déclin au cours des 2 dernières générations
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix dernières années ou trois dernières générations] (78-90 ans).
Aucun déclin au cours des 2 dernières générations
Pourcentage [prévu ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours des [dix prochaines années ou trois prochaines générations] (78-90 ans).
s.o.
Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d'augmentation] du nombre total d'individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur.
Aucun déclin observé au cours des 2,4 dernières générations, et il n’y a aucune indication que cela changera
Est-ce que les causes du déclin sont clairement réversibles et comprises et ont effectivement cessé?
s.o.
Y a-t-il des fluctuations extrêmes du nombre d'individus matures?
Non
Superficie estimée de la zone d'occurrence Tirée de COSEPAC (2004)
91 409 km²
Indice de zone d'occupation (IZO; valeurs d'une grille à carrés de 2 km de côté)
28 300 km² selon la zone d’occupation pondérée par l’échantillonnage obtenue au cours des 5 dernières années de relevés par navire de recherche
La population totale est-elle gravement fragmentée?
Non
Nombre de localités (selon le nombre de menaces)
s.o.
Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d'occurrence?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de l'indice de zone d'occupation?
Oui
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de populations?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités?
Non
Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l'étendue ou la qualité] de l'habitat?
Non
Y a-t-il des fluctuations extrêmes du nombre de populations?
Non
Y a-t-il des fluctuations extrêmes du nombre de localités?
Non
Y a-t-il des fluctuations extrêmes de la zone d'occurrence?
Non
Y a-t-il des fluctuations extrêmes de l'indice de zone d'occupation?
Non
Nombre d'individus matures
Total: 614 000 (estimation de la population minimale d’après les 5 dernières années de relevés par navire de recherche dans l’ouest du plateau néo-écossais et la portion canadienne du banc Georges) La population réelle est vraisemblablement plus grande.
La probabilité de disparition de l'espèce à l'état sauvage est d'au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans].
L'analyse n'a pas été effectuée
Les menaces pesant sur cette population sont limitées.
Situation des populations de l'extérieur?
La population des États-Unis est stable
Une immigration a-t-elle été constatée ou est-elle possible?
Oui
Des individus immigrants seraient-ils adaptés pour survivre au Canada?
Oui
Y a-t-il suffisamment d'habitat disponible au Canada pour les individus immigrants dans l'estuaire du Saint-Laurent?
Oui
La possibilité d'une immigration depuis des populations externes existe-t-elle?
Possible
L'information concernant l'espèce est-elle de nature délicate?
Non
COSEPAC : Espèce désignée « préoccupante » en mai 2005. Réexamen du statut et inscription à la catégorie « non en péril » en mai 2015.
En 2005, le COSEPAC a évalué la raie tachetée et défini quatre unités désignables (UD) séparées : 1. UD du sud du golfe du Saint-Laurent (« en voie de disparition »); UD de l’est du plateau néo-écossais (« menacée »); 3. UD de l’ouest du plateau néo-écossais, de la baie de Fundy et du banc Georges (« préoccupante »); UD du nord du golfe du Saint-Laurent et de Terre-Neuve (« données insuffisantes »). En 2010, le gouverneur en conseil a décidé de ne pas ajouter la raie tachetée à l’annexe de la Loi sur les espèces en péril (LEP) parce que l’inscription entraînerait la perte de millions de dollars de revenus liés aux pêches ainsi qu’une perte directe et indirecte importante d’emplois.
Nos connaissances sur la biologie de la raie tachetée se sont beaucoup améliorées depuis l’évaluation effectuée par le COSEPAC en 2005. Une analyse du potentiel de rétablissement de l’espèce a également été réalisée par Pêches et Océans Canada (MPO) en 2006, ce qui nous a permis d’obtenir davantage de données sur la répartition de l’espèce et la dynamique des populations. De nouvelles données sur la taille et l’âge à maturité dans l’aire de répartition au Canada ont mené à la révision de la durée d’une génération estimée. Un modèle d’analyse démographique a été élaboré et utilisé pour suivre les effectifs et les changements relatifs à la mortalité due aux pêches et à la mortalité due à des causes autres que les pêches. Les nouvelles lignes directrices du COSEPAC pour déterminer les unités désignables ont été suivies lors de la révision de la structure proposée des populations dans le cadre de la réévaluation. Des données additionnelles sur les tendances des effectifs couvrant une période de plus de dix ans sont disponibles grâce aux relevés au chalut de fond du MPO.
Dans l’ensemble, le présent rapport constitue une mise à jour importante de l’évaluation du COSEPAC réalisée en 2005.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsable des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu’en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d’Environnement Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
La raie tachetée (Leucoraja ocellata Mitchill, 1815) appartient à la classe des Chondrichthyens, à la sous-classe des Élasmobranches, à l’ordre des Rajiformes et à la famille des Rajidés (voir la figure 1). En anglais, on la désigne par plusieurs noms : winter skate, big skate, eyed skate et spotted skate.
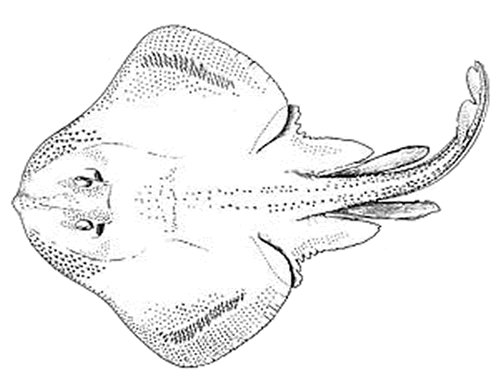
Description longue pour la figure 1
Dessin au trait de la raie tachetée, vue dorsale. La raie tachetée est un poisson en forme de disque dont le museau très arrondi se termine par un angle obtus et porte deux taches opaques de cartilage. La queue est mince et arrondie et comprend deux petites nageoires à l'extrémité. Les yeux se trouvent sur la surface dorsale, tandis que les branchies et la bouche sont sur la surface ventrale.
La raie tachetée se distingue des autres espèces de raies, à l’exception d’une espèce sympatrique, la raie hérisson (Leucoraja erinacea), par une combinaison des caractéristiques suivantes : la raie tachetée a le museau très arrondi, qui se termine par un angle obtus et porte 2 taches opaques de cartilage; le milieu de la partie postérieure du corps (disque) et la queue portent 3 rangées ou plus d’épines, depuis la région scapulaire jusqu’à la naissance de la première nageoire dorsale, la rangée médiane disparaissant durant la croissance; le disque supérieur présente des taches arrondies foncées et, généralement, un ocelle ou plus près du bord postérieur de chaque nageoire pectorale (Collette et Klein-MacPhee, 2002). Cependant, les spécimens dépourvus de ces taches ressemblent tellement à la raie hérisson lorsqu’ils sont petits (< 35 cm) qu’un simple examen visuel ne permet pas de les distinguer de cette espèce. Il est à noter que cette difficulté de distinguer la raie tachetée de la raie hérisson n’a pas d’incidence sur les estimations des tendances démographiques du présent rapport puisque celles-ci portent uniquement sur la population d’adultes de l’espèce et excluent donc les individus dont la taille est inférieure à 35 cm. Le nombre de rangées de dents sur la mâchoire supérieure, variable selon la taille de l’individu, est plus élevé chez la raie tachetée que chez la raie hérisson. Le nombre d’épines de la rangée médiocaudale est également plus élevé chez la raie tachetée (généralement > 21) (Collette et Klein-MacPhee, 2002). La raie tachetée se distingue en outre par son profil antérieur cunéiforme, l’absence d’épines bien développées derrière les épaules et la partie postérieure de la queue, et la présence de denticules densément distribués sur la face inférieure de la queue (sauf à l’extrémité) (Scott et Scott, 1988).
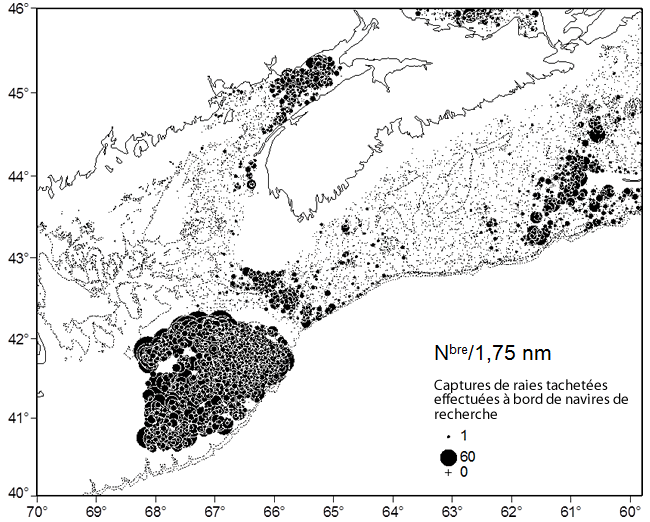
Les résultats de relevés au chalut de fond effectués par Pêches et Océans Canada (MPO) sont utilisés pour déterminer la répartition spatiale de la raie tachetée au Canada. Les relevés ont été réalisés par le personnel de quatre Régions du MPO dans des sections d’eaux côtières ne se chevauchant pas. La Région du Golfe du MPO a effectué des relevés dans le sud du golfe du Saint-Laurent (SGSL; div. 4T de l’Organisation des pêches de l’Atlantique Nord-Ouest [OPANO]; voir la figure 2) en septembre principalement. Des relevés printaniers de la Région de Terre-Neuve-et-Labrador ont permis de couvrir les divisions 3LNOP de février à avril. Les relevés de la Région du Québec, en août, ont couvert le nord du golfe du Saint-Laurent (NGSL; div. 4RS). La Région des Maritimes a mené des relevés dans l’est du plateau néo-écossais (EPNE; div. 4VW), dans l’ouest du plateau néo-écossais (OPNE) et dans la baie de Fundy (div. 4X) en juillet principalement, ainsi que sur le banc Georges (div. 5Z) en février et en mars. La répartition spatiale de la raie tachetée est montrée à la figure 3. Pour cette figure, la densité moyenne de l’espèce (nombre par trait de 1,75 nm) a été déterminée sur une grille de 10’ de latitude sur 10’ de longitude (1,82 x 1,29 km à une latitude de 46°).
Dans les eaux canadiennes, les raies tachetées forment trois principales concentrations (voir la figure 3), essentiellement à des profondeurs de moins de 110 m (McEachran et Martin, 1975; Swain et al., 2006) : dans le SGSL, dans l’EPNE et dans la baie de Fundy-OPNE (notamment le banc Browns) ainsi que dans la portion canadienne du banc Georges.
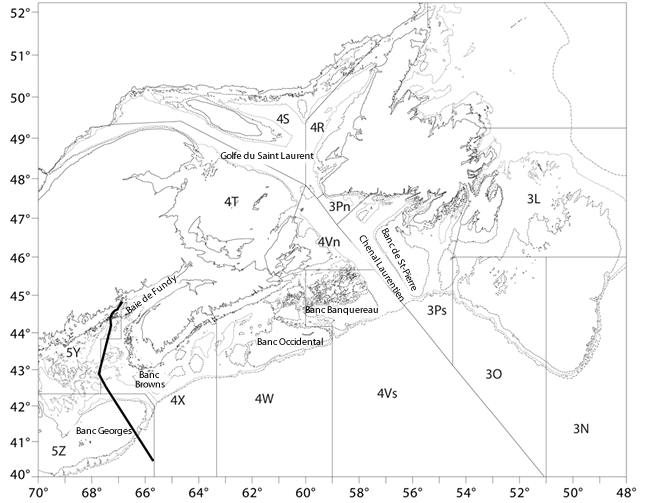
Description longue pour la figure 2
Carte de référence (40 52 ° nord; 48 70 ° ouest) montrant la bathymétrie (100, 200 et 500 m de contour), les limites de l'Organisation des pêches de l'Atlantique Nord Ouest (OPANO) et la toponymie mentionnées dans le présent rapport.
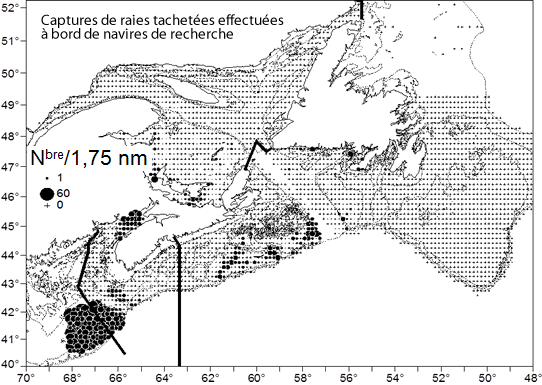
Description longue pour la figure 3
Carte montrant les lieux de capture de raies tachetées lors de relevés au chalut de fond de Pêches et Océans Canada dans les divisions 4VWX (juillet), 4T (septembre), 4RS (août), 3LNOP (printemps) et 5Z (printemps) de l'OPANO. Les données datent de 1970 à 2013, et les raies de toutes tailles sont représentées. Les limites des unités désignables (UD) suivantes sont illustrées : UD de l'ouest du plateau néo écossais et du banc Georges, UD de l'est du plateau néo écossais et de Terre Neuve et UD du golfe du Saint Laurent.
Dans la région du SGSL, en septembre, lorsque les principaux relevés aux fins de recherches sont effectués, les raies tachetées se trouvent essentiellement à l’est du Nouveau-Brunswick, dans le détroit de Northumberland, à l’est de l’Île-du-Prince-Édouard et à l’est des Îles-de-la-Madeleine. Très peu de prises ont été rapportées dans la portion centrale du SGSL et le long de la côte ouest de l’île du Cap-Breton. Les rares raies tachetées capturées dans des lieux dispersés (53 individus) de la région du NGSL se trouvaient dans des eaux relativement profondes (120-500 m). Il est possible que certains de ces spécimens aient en fait été des raies épineuses (Amblyraja radiata) mal identifiées (Dutil et al., 2006) ou des individus errants du SGSL (D. Swain, comm. pers., 2014).
Dans l’EPNE, des taies tachetées ont été observées dans les eaux relativement peu profondes (< 100 m) des bancs Western et Banquereau (voir la figure 3). En plus, des prises ont été réalisées le long de la bordure ouest du chenal Laurentien (à l’est du banc Banquereau), à la limite est de la zone de couverture du relevé de juillet, dans des eaux d’une profondeur de plus de 200 m (voir la figure 4). On note une zone non recensée dans le chenal Laurentien, entre le relevé de juillet réalisé par la Région des Maritimes du MPO et le relevé printanier effectué par la Région de Terre-Neuve-et-Labrador du MPO, et l’on n’est pas certain que des raies tachetées soient présentes dans la portion ouest du chenal (voir la figure 4). Des raies tachetées ont été observées dans la portion centrale du chenal Laurentien durant le relevé printanier, à des profondeurs de 200 à 400 m. On a également fait mention d’individus se trouvant à proximité de l’isobathe de 200 m et dans les eaux moins profondes du banc de St-Pierre. Il est donc possible que des raies tachetées se répartissent de l’EPNE au banc de St-Pierre en passant par le chenal Laurentien. L’espèce a également été observée dans les zones côtières du sud de Terre-Neuve (voir la figure 3).
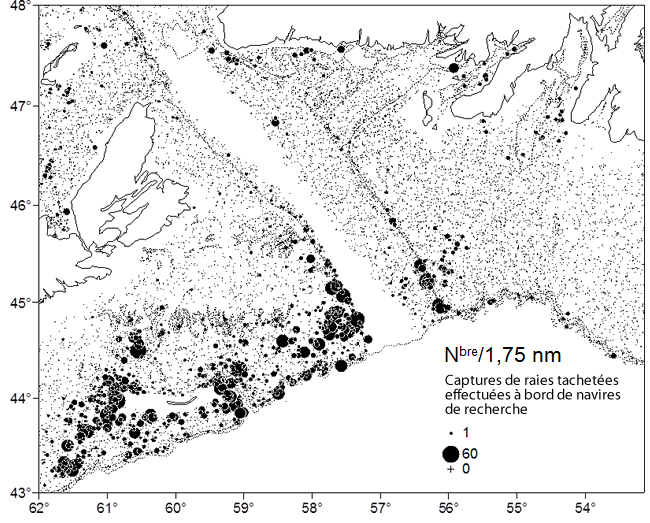
Description longue pour la figure 4
Carte illustrant les raies tachetées dans les environs du chenal Laurentien, dans l'UD de l'est du plateau néo écossais et de Terre Neuve, lors du relevé de Pêches et Océans Canada sur le plateau néo écossais (en juillet) et du relevé des divisions 3LNOP (au printemps). Les données datent de 1970 à 2013, et les raies de toutes tailles sont représentées.
Dans la zone de la troisième concentration de raies tachetées, des individus sont observés dans la baie de Fundy, dans l’OPNE (principalement sur le banc Browns et le banc Georges (voir la figure 3). L’absence visible de l’espèce au sud-ouest de la Nouvelle-Écosse, entre le banc Browns et la baie de Fundy, découle en fait de l’absence de relevés dans cette zone, à cause des fonds non chalutables. On sait cependant que la raie tachetée se trouve dans cette zone puisque des individus ont été capturés lors de relevés ciblant d’autres poissons de fond (Swain et al., 2006). Il est possible que le chenal de Fundy entre le banc Georges et le banc Browns agisse à titre de barrière bathymétrique nuisant aux déplacements entre ces zones puisque les profondeurs y atteignent plus de 200 m sur une distance de 40 km. Toutefois, on en sait peu à propos des déplacements des raies tachetées dans cette zone. Un faible nombre de prises ont été mentionnées dans le chenal de Fundy durant le relevé de juillet du MPO (voir la figure 5), et Scott (1982) a noté la présence de l’espèce dans la plage de profondeurs couverte par le relevé. De même, Frisk et al. (2008) a rapporté qu’un nombre restreint d’individus ont été capturés dans le chenal de Fundy au cours de relevés états-uniens au chalut de fond dans la zone. Selon ces mêmes auteurs, il se pourrait que les populations de raies tachetées sur le banc Georges et sur le plateau néo-écossais aient été liées et que la soudaine hausse des effectifs de l’espèce sur le banc Georges dans les années 1980 résulte d’une migration à partir du plateau néo-écossais.
Les 3 principales concentrations sont séparées par des distances considérables. Une centaine de kilomètres séparent les captures des relevés dans l’EPNE de celles des relevés dans l’OPNE au cours de la période de 1970 à 2013 (soit 43 ans; voir la figure 5). La distance entre les principales concentrations sur le banc Occidental et le banc Browns est de 300 km. Un total de 27 raies tachetées ont été capturées dans la sous-division 4Vn de l’OPANO (à l’est de l’île du Cap-Breton) au cours du relevé de juillet, tandis qu’on a fait état de 2 751 individus appartenant à la concentration principale dans l’OPNE. La distance entre les concentrations du SGSL et de l’EPNE est d’environ 240 km (voir la figure 4). On en sait peu à propos de la migration et du mélange entre les populations de raies tachetées. Cependant, ces longues distances dans des milieux apparemment propices laissent croire que les concentrations pourraient être distinctes.
On note des différences relatives aux possibles caractères héréditaires du cycle vital entre les raies tachetées du sud du golfe du Saint-Laurent et celles d’ailleurs. Dans les eaux du plateau néo-écossais, la longueur à laquelle 50 % des individus atteignent la maturité est de 75 à 78 cm (Simon et al., 2003; McPhie et Campana, 2009), et un grand nombre de spécimens mesurant plus de 80 cm a été signalé (McEachran et Martin, 1977; Simon et al., 2003). Les spécimens de plus de 80 cm étaient également abondants lors des relevés printaniers au sud de Terre-Neuve (voir la figure 6). La taille à laquelle 50 % des individus vivant dans le sud du golfe du Saint-Laurent atteignent la maturité est d’environ 40 cm, et aucun individu d’une longueur totale de plus de 80 cm n’a été observé dans cette région (Swain et al., 2006, 2009). Sur les 53 raies tachetées capturées lors de relevés dans le nord du golfe du Saint-Laurent, aucune ne dépassait 65 cm de longueur (voir la figure 6). Dans le sud du golfe, les raies tachetées possèdent également moins de rangées de dents et plus de vertèbres sur le pédoncule que les autres spécimens (McEachran et Martin, 1977). Les différences relatives au cycle vital entre la population du golfe Saint-Laurent et celles du plateau néo-écossais peuvent refléter un déplacement de caractère entre la raie tachetée et la raie hérisson (L. erinacea) sur le plateau néo-écossais, où ces espèces similaires sont sympatriques (McEachran et Martin, 1977).
Description longue pour la figure 5
Carte illustrant les lieux de capture de raies tachetées dans les zones de relevés de Pêches et Océans Canada dans l'ouest du plateau néo écossais (en juillet) et sur le banc Georges (au printemps).
Diagramme A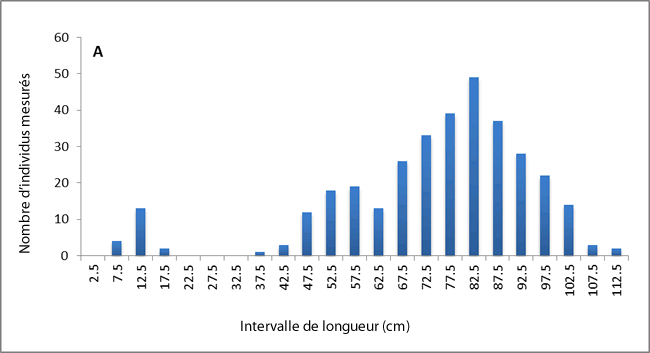
Diagramme B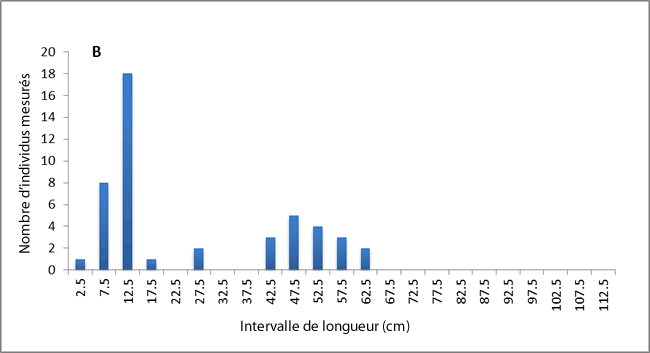
Description longue pour la figure 6
Deux diagrammes à barre illustrant la distribution des longueurs de raies tachetées mesurées lors du relevé printanier à Terre Neuve (partie de l'UD de l'est du plateau néo écossais et de Terre Neuve) (diagramme A) et du relevé dans le nord du golfe du Saint Laurent en août (partie de l'UD du golfe du Saint Laurent) (diagramme B). Les spécimens de plus de 80 cm étaient abondants lors des relevés printaniers dans le sud de Terre Neuve; cependant, aucune raie tachetée capturée lors des relevés dans le nord du golfe du Saint Laurent ne dépassait 65 cm.
Le précédent rapport de situation du COSEPAC sur la raie tachetée, qui date de 2005, a défini quatre UD selon les différences relatives à la répartition spatiale, aux caractéristiques du cycle vital, aux limites inférées et observées des déplacements des individus et aux diverses tendances des effectifs (COSEWIC, 2005). Ces zones ont été délimitées en fonction des divisions de l’OPANO (voir la figure 2) :
- population du sud du golfe du Saint-Laurent (4T de l’OPANO);
- population de l’est du plateau néo-écossais (4VW de l’OPANO);
- population du banc Georges, de l’ouest du plateau néo-écossais et de la baie de Fundy (4X5Ze de l’OPANO);
- population du nord du golfe Saint-Laurent et de Terre-Neuve (division 3LNOP4RS de l’OPANO).
La population de l’est du plateau néo-écossais a connu un grave déclin du nombre d’individus matures, tandis que la population du banc Georges, de l’ouest du plateau néo-écossais et de la baie de Fundy demeure relativement stable. Le précédent rapport de situation a été rédigé avant la mise en œuvre des Lignes directrices pour reconnaître les unités désignables du COSEPAC (COSEWIC, 2011), qui ne considèrent pas les différences relatives aux tendances des effectifs comme un critère. La quatrième UD précédemment désignée avait été séparée des autres parce qu’elle se trouvait dans les eaux à la limite septentrionale de l’aire de répartition de l’espèce, où les individus sont rares. Encore une fois, ce critère n’est pas reconnu dans les actuelles lignes directrices du COSEPAC.
Les lignes directrices incluent le caractère distinct et le caractère important dans l’évolution. Cependant, aucune étude génétique n’a été publiée sur la raie tachetée. La répartition spatiale de l’espèce laisse entrevoir trois populations distinctes. Une se trouve principalement dans le SGSL, et certains individus sont observés jusque dans le nord du golfe. Il existe des différences relatives aux caractéristiques du cycle vital entre les raies tachetées du golfe du Saint-Laurent et les raies d’ailleurs, qui pourraient être héréditaires (critère no 2 du caractère important). Chez les individus capturés à Terre-Neuve lors du relevé printanier, la distribution des longueurs était semblable à celle de la population de l’EPNE, ce qui indique que ces deux populations sont du même type.
La deuxième UD se trouve principalement dans l’EPNE, mais aussi sur le banc de St-Pierre. Des raies tachetées sont également capturées dans les eaux au sud de Terre-Neuve. La troisième UD est dans l’OPNE, la baie de Fundy et le banc Georges. La séparation spatiale entre ces régions est telle que les déplacements des individus d’une région à l’autre sont très limités. On note également des différences biogéographiques entre l’ouest et l’est du plateau néo-écossais. L’ouest du plateau (et le banc Georges) subit l’influence du Gulf Stream; les températures et la salinité y sont plus élevées que dans l’est du plateau, qui se trouve dans le courant du Labrador (Mahon et al., 1984). Plusieurs espèces de poissons de fond se divisent en deux stocks, entre l’ouest et l’est du plateau néo-écossais. C’est notamment le cas de la morue (Gadus morhua), de l’aiglefin (Melanogrammus aeglefinus) et de la goberge (Pollachius virens). Ainsi, les raies tachetées se trouvant de part et d’autre du plateau néo-écossais pourraient présenter des adaptations locales (critère no 2 du caractère distinct). Si la population de l’EPNE disparaissait, il y aurait une disjonction considérable dans l’aire de répartition de l’espèce au Canada (critère no 4 du caractère important). Compte tenu de ces facteurs, on reconnaît trois UD de raies tachetées au pays (voir la figure 2) :
- UD du golfe du Saint-Laurent (4RST de l’OPANO)
- UD de l’est du plateau néo-écossais et de Terre-Neuve (3LNOP4VW de l’OPANO)
- UD de l’ouest du plateau néo-écossais et du banc Georges (inclut la baie de Fundy, la portion canadienne du banc Georges et la division 5Y de l’OPANO [4X5Ze5Yb]. Cette UD chevauche la frontière avec les États-Unis, et les poissons traversent librement cette frontière.)
La raie tachetée est endémique dans l’Atlantique Nord-Ouest, où elle vit en sympatrie avec la raie hérisson dans la plus grande partie de son aire de répartition. Dans les zones de sympatrie, la taille à maturité et la taille finale sont plus grandes chez la raie tachetée que chez la raie hérisson. Cependant, les individus de la raie tachetée sont allopatriques dans le sud du golfe du Saint-Laurent, où ils présentent des caractères du cycle vital intermédiaires entre la raie hérisson et la raie tachetée. Les différences entre les individus du sud du golfe du Saint-Laurent et ceux d’ailleurs pourraient être suffisantes pour justifier une réévaluation taxinomique de la population et une désignation possible à titre d’espèce distincte (Kelly et Hanson, 2013a).
La raie tachetée est endémique dans l’Atlantique Nord-Ouest (Scott et Scott, 1988), où on la rencontre depuis le golfe du Saint-Laurent et les eaux méridionales de Terre-Neuve jusqu’au cap Hatteras, en Caroline du Nord (Robins et Ray, 1986).
Dans les eaux canadiennes, la raie tachetée se rencontre depuis la zone au sud de Terre-Neuve et le golfe du Saint-Laurent jusqu’à la frontière entre le Canada et les États-Unis. La zone d’occurrence fondée sur la distribution des captures réalisées lors de relevés scientifiques (montrées à la figure 3) a été estimée à 161 515 km² pour l’UD du golfe du Saint-Laurent, à 215 436 km² pour l’UD de l’est du plateau néo-écossais et de Terre-Neuve et à 91 409 km² pour l’UD de l’ouest du plateau néo-écossais et du banc Georges (J. WU, Secrétariat du COSEPAC, comm. pers., février 2015).
La raie tachetée est une espèce benthique vivant sur les fonds de sable ou de gravier. Lors des relevés réalisés à bord de navires de recherche, plus de 90 % des individus sont capturés dans des eaux de moins de 150 m de profondeur, mais certains le sont à presque 400 m. Dans le sud du golfe, les raies tachetées occupent les eaux côtières très peu profondes à la fin de l’été et au début de l’automne, et se dispersent sur le plateau madelinien en hiver (Darbyson et Benoît, 2003). Dans les eaux du plateau néo-écossais, Scott et Scott (1988) ont indiqué que les raies préféraient une profondeur de 37 à 90 m. Des jeunes de l’année ont également été observés dans les eaux côtières très peu profondes du sud de Terre-Neuve et le long des plages de la baie de la Conception (M. Simpson, Région de Terre-Neuve-et-Labrador du MPO, comm. pers., avril 2015).
Dans les eaux canadiennes, les raies tachetées se trouvent principalement dans les zones où la température est la plus élevée et évitent les températures de moins de 2 ºC. Dans le sud du golfe, la température médiane de la zone occupée lors du relevé de septembre était de 8,7 °C (Swain et al., 2006). Le détroit de Northumberland a été recensé au chalut de fond en juillet et en août de 2001 à 2009, et les raies tachetées capturées se trouvaient alors dans des eaux peu profondes et chaudes, à une profondeur médiane de 12 m et à une température médiane de 16,3 °C (Kelly et Hanson, 2013a). En hiver, les températures du fond des lieux de capture sur le plateau madelinien étaient considérablement plus basses, soit près de 0 °C (Galbraith et al., 2013). Ailleurs, l’espèce a été capturée dans des eaux de 1,1 à 12,7 °C à l’est de la Nouvelle-Écosse et de 2 à 15 °C entre le sud de la Nouvelle-Écosse et le cap Hatteras. Dans les eaux du plateau néo-écossais, l’espèce préfère les profondeurs où la température est de 5 à 9 °C (Collette et Klein-MacPhee, 2002). On note peu de changements saisonniers de la répartition spatiale sur tout le plateau néo-écossais et dans la baie de Fundy (Simon et al., 2003).
En Nouvelle-Angleterre, l’espèce quitte généralement la côte méridionale et ses eaux très peu profondes au début de l’été, lorsque la température atteint 18 ou 19 °C, pour y revenir, jusqu’à la hauteur de l’État de New York, au début de l’automne (Bigelow et Schroeder, 1953). En été, elle est régulièrement observée près des côtes de la baie Passamaquoddy, à l’entrée de la baie de Fundy, en Nouvelle-Écosse et autour de l’Île-du-Prince-Édouard.
On a noté une forte tendance de réchauffement dans la région où vit la raie tachetée. On ne sait pas comment les variations des conditions océaniques touchent cette espèce, mais les effets pourraient être à la fois directs et indirects (changements dans le réseau trophique). En 2013, les températures à la surface de la mer ont atteint des sommets dans la zone de l’Atlantique durant l’été, et des températures record ou proches des records durant les mois où les eaux sont libres de glace (DFO, 2013). La couverture de glace de mer a été mince durant trois hivers consécutifs (de 2011 à 2013), et les températures estivales de la couche intermédiaire froide sont les plus élevées depuis le début des années 1980 (voire avant) dans le golfe du Saint-Laurent et sur le plateau néo-écossais. Au cours de cette période, les températures de fond étaient supérieures à la normale dans presque toute la zone, et des records ont été enregistrés dans le nord du golfe, des régions du plateau néo-écossais (divisions 4X et 4W de l’OPANO) et le sud de Terre-Neuve (division 3P).
On pense que la raie tachetée pond de 40 à 70 sacs ovigères annuellement (Kelly et Hanson, 2013b), mais le nombre exact est inconnu (Collette et Klein-MacPhee, 2002). L’incubation pourrait durer jusqu’à 22 mois (Simon et Frank, 1998). Dans les eaux du sud de la Nouvelle-Angleterre, la ponte s’étale sur toute l’année, tandis que, dans les eaux de la Nouvelle-Écosse, elle a lieu seulement à l’été et à l’automne (Vladykov, 1936; Collette et Klein-MacPhee, 2002). À l’ouest de l’île de Sable, des pêcheurs ont noté que les femelles pondaient des sacs ovigères complets uniquement à la fin de l’été et au début de l’automne, ce qui laisse croire que ce secteur est une frayère (Simon et Frank, 2000). Chaque embryon est enfermé dans un sac ovigère dur de couleur brune mesurant de 5,5 à 8,6 cm de longueur et de 3,5 à 5,2 cm de largeur, cornes non comprises (Scott et Scott, 1988). Le sac ovigère est muni de 4 longs filaments (« cornes »), qui font probablement office d’ancres (Collette et Klein-MacPhee, 2002). Les spécimens récemment éclos (c’est-à-dire les spécimens à l’abdomen encore rempli de vitellus) mesuraient environ de 112 à 127 mm de longueur.
Les raies tachetées de l’UD de l’est du plateau néo-écossais et de Terre-Neuve et de l’UD de l’ouest du plateau néo-écossais et du banc Georges atteignent la maturité à une taille nettement plus grande que celles du golfe du Saint-Laurent (McEachran et Martin, 1977). Dans l’est du plateau néo-écossais, la longueur totale à laquelle 50 % des individus atteignent la maturité est de quelque 77 cm. De nouvelles données sur la maturité indiquent que l’âge auquel 50 % des individus atteignent la maturité est de 11 ans chez les mâles et de 13 ans chez les femelles (McPhie et Campana, 2009). Ces valeurs sont plus élevées que celles utilisées lors de l’évaluation originale du COSEPAC, qui avaient été inférées d’après des données préliminaires sur l’âge lié à la longueur provenant d’un ensemble de données limité (COSEWIC, 2005). Ces nouvelles données sur la maturité sont similaires à celles rapportées pour la raie tachetée du golfe du Maine (Sulikowski et al., 2003). La longueur à laquelle 50 % des individus atteignent la maturité dans le sud du golfe est de 42 cm pour les femelles et de 40 cm pour les mâles (Kelly et Hanson, 2013b), tandis que l’âge auquel 50 % des individus sont matures est de 5 ans (chez les 2 sexes). Il est à noter que ces nouvelles estimations de l’âge à maturité pour les 2 populations ont été obtenues de populations grandement réduites. Il est donc possible que les données sous-estiment l’âge à maturité des individus non reproducteurs. Les estimations de l’âge à maturité n’étaient pas disponibles pour les raies tachetées de l’ouest du plateau néo-écossais et de la baie de Fundy. Cependant, étant donné que ces dernières ont une taille semblable à celle des raies tachetées de l’est du plateau néo-écossais, on a présumé qu’elles atteignaient la maturité à un âge semblable.
Il existe plusieurs estimations du coefficient instantané de mortalité due à des causes autres que les pêches (M), composante importante de l’estimation de la durée d’une génération. Simon et Frank (1996) ont estimé M à 0,21 an-1 d’après les estimations de la longévité. McPhie et Campana (2009) ont présenté 4 estimations empiriques de M pour la population de l’est du plateau néo-écossais d’après divers paramètres du cycle vital (p. ex. coefficient de croissance de von Bertalanffy, température de la zone occupée, âge à maturité). Les valeurs estimées variaient de 0,13 à 0,44 an-1 (moyenne de 0,26 an-1). Swain et al. (2009) ont utilisé un modèle de population basé sur la longueur des individus pour estimer les indices vitaux à valeur temporelle variable pour les raies tachetées du sud du golfe du Saint-Laurent et de l’est du plateau néo-écossais, où M est estimé sur une échelle décennale. Les auteurs ont noté des signes d’augmentation de M chez les adultes. Durant les années 1970, M était estimé à 0,05 an-1 pour les 2 populations. Il a augmenté, pour atteindre environ 0,6 an-1 dans le sud du golfe et 0,35 an-1 sur le plateau néo-écossais au cours de la période la plus récente. Ces valeurs estimées élevées de M concordent avec l’absence de rétablissement des 2 populations, et ce, malgré une période prolongée où il y a eu très peu de pêche. Cependant, les estimations ne représentent pas les conditions normales puisque, comme il sera abordé plus en détail à la section Menaces et facteurs limitatifs, les populations ayant un M aussi élevé pourraient ne pas être viables. En revanche, les valeurs très faibles de M lors de la période initiale semblent trop basses pour permettre l’estimation de la durée d’une génération, car elles laissent croire à un âge moyen des reproducteurs trop élevé de 25 ans et à une longévité de 60 ans. Or, des raies tachetées de cet âge n’ont jamais été observées. Lors de l’évaluation précédente du COSEPAC, on a utilisé une valeur de M de 0,1 an-1, et ce, malgré le peu de données probantes dans la littérature (COSEWIC, 2005).
L’Union internationale pour la conservation de la nature (UICN), dans ses lignes directrices (IUCN, 2013), recommande d’utiliser l’âge auquel 50 % des individus atteignent la maturité + 1/M pour estimer la durée de génération. Ces lignes directrices mentionnent également que la durée d’une génération devrait refléter les conditions présentes avant la réduction des populations due à l’exploitation. D’après l’information disponible et compte tenu de la distribution des âges observée des populations, on présume que M est de 0,2 an-1 pour les 3 populations de raies tachetées, ce qui permet d’estimer des durées de génération de 18 ans pour les individus de l’UD de l’est du plateau néo-écossais et de Terre-Neuve et de l’UD de l’ouest du plateau néo-écossais et du banc Georges, et de 10 ans, pour les plus petites raies de l’UD du golfe du Saint-Laurent. La durée d’une génération de l’UD du golfe du Saint-Laurent est considérablement plus courte que celle qui avait été utilisée dans le cadre de la dernière évaluation (22 ans), alors que l’on ne possédait pas de données propres à la population en ce qui concerne l’âge de maturité. La nouvelle estimation repose sur des bases plus solides.
On en sait très peu à propos de la physiologie et de l’adaptabilité de la raie tachetée.
Au Canada, les raies tachetées se rencontrent dans 3 principales concentrations (voir la figure 3), séparées les unes des autres par des distances de 200 à 250 km. Des relevés saisonniers réalisés sur le plateau néo-écossais et dans la baie de Fundy (printemps 1979-1984, automne 1978-1994, été 1971-2013) ont indiqué peu de variations de la répartition spatiale et bathymétrique de la raie tachetée (Simon et al., 2003). Dans le sud du golfe du Saint-Laurent, les raies tachetées favorisent les eaux côtières chaudes et peu profondes en été. Elles quittent ces eaux en hiver, possiblement pour éviter l’affouillement glaciel, et se répartissent dans les eaux plus profondes et beaucoup plus froides du plateau madelinien et du chenal Laurentien (Clay, 1991; Darbyson et Benoît, 2003). L’espèce se déplace probablement de part et d’autre de la frontière entre le Canada et les États-Unis sur le banc Georges.
La raie tachetée préfère le tourteau commun (Cancer irroratus) et le calmar, mais elle se nourrit également d’annélides, d’amphipodes, de crevettes, de couteaux et de n’importe quels petits poissons facilement accessibles (Collette et Klein-MacPhee, 2002). Selon les données historiques, la raie tachetée chasse également des raies de petite taille, et le lançon d’Amérique (Ammodytes americanus) serait parmi ses poissons préférés (Simon et Frank, 2000). Les juvéniles se nourrissent principalement de crustacés, tandis que les adultes chassent les poissons et les tourteaux communs de grande taille (Kelly et Hanson, 2013b).
Dans les zones où la raie tachetée et la raie hérisson sont sympatriques, McEachran et al. (1976) ont laissé entendre que les deux espèces évitent une trop forte compétition en consommant différentes espèces de proies. Ainsi, la raie tachetée exploite principalement l’endofaune, et la raie hérisson, l’épifaune. La portion nord de l’aire de répartition canadienne de la raie tachetée chevauche également l’aire de répartition de la raie épineuse (Amblyraja radiata), mais on en sait peu sur les interactions entre les deux espèces.
On en sait peu sur la prédation exercée sur la raie tachetée, mais cette dernière semble être la proie de nombreuses espèces, y compris des requins, d’autres raies (comme la raie hérisson) et le phoque gris (Scott et Scott, 1988; Beck et al., 2007). Des études réalisées sur des échantillons de matières fécales et de contenus stomacaux de phoques gris donnent à penser que la raie tachetée constitue une faible partie de leur alimentation (voir par exemple Bowen et Harrison, 1994; Hammill et Stenson, 2000). Cependant, l’examen d’échantillons fécaux sous-estime la présence des raies dans l’alimentation des phoques, les raies n’ayant pas de structures dures comme des os et des otolithes. Lors d’une récente étude se basant sur les profils des acides gras, Beck et al. (2007) ont affirmé que la raie tachetée forme moins de 1 % de l’alimentation du phoque gris de l’île de Sable, au sud de la Nouvelle-Écosse. De l’information additionnelle sur la prédation par le phoque gris se trouve à la section Menaces et facteurs limitatifs.
L’espèce est exposée à plusieurs parasites, notamment des protozoaires, des myxosporidies, des hématozoaires, des trématodes et des nématodes (Scott et Scott, 1988).
Les analyses de la taille et des tendances des populations dans le présent rapport sont fondées sur des relevés annuels, indépendants des pêches, que le MPO effectue à bord de navires de recherche depuis plusieurs décennies en suivant les protocoles normalisés décrits par Chadwick et al. (2007). Ces relevés ont une vaste couverture spatiale et temporelle et fournissent des indices d’abondance relative fiables sur une période pertinente aux fins de l’évaluation. Les relevés spécifiques utilisés sont mentionnés à la section Structure spatiale et variabilité des populations. D’autres sources de données ont été utilisées lors de la précédente évaluation du COSEPAC, dont les suivantes : 1) des relevés effectués à bord de navires de recherche avant les années 1970, alors que les protocoles n’étaient pas encore normalisés; 2) des relevés couvrant de courtes périodes effectués par des pêcheurs ou des scientifiques à partir du milieu des années 1990 au moyen d’engins fixes ou mobiles; 3) les données recueillies par les pêches commerciales dirigées de la raie tachetée sur le plateau néo-écossais. Ces deux dernières sources de données complétaient et confirmaient l’information obtenue lors des principaux relevés, elles ne seront donc pas répétées ici.
Le présent rapport met l’accent sur les tendances de l’effectif des individus matures. Les estimations des relevés sont considérées comme fiables pour déterminer approximativement les tendances, mais les valeurs obtenues risquent d’être inférieures à l’effectif réel, car le filet utilisé ne capture pas toutes les raies sur son passage. Les variations en pourcentage de l’effectif ont été estimées à 100*(exp(t*b)-1), où t est le nombre d’années couvertes par l’indice et b est la pente de la régression linéaire du logarithme naturel de l’indice de relevé par rapport à l’année (log-linéaire). Certaines années (2013 dans le sud du golfe; 2001, 2002, 2008 et 2013 dans l’est du plateau néo-écossais; 1971 dans l’ouest du plateau néo-écossais), des relevés ne capturaient aucune raie tachetée mature. Dans les analyses de régression, les valeurs 0 ont été remplacées par la valeur la plus faible n’étant pas 0 dans la série chronologique. Les séries chronologiques des estimations de l’effectif des relevés utilisées pour les analyses sont présentées à l’annexe 1.
La zone d’occupation annuelle (At) de l’espèce a été estimée d’après les résultats des relevés au moyen de la zone d’occupation pondérée par l’échantillonnage :

où n est le nombre de traits de chalut du relevé de l’année t, Yi est le nombre d’individus capturés au trait i, et ai est la superficie de la strate échantillonnée au trait i divisée par le nombre de traits réalisés dans la strate (Smedbol et al., 2002).
Les relevés dans le sud du golfe Saint-Laurent ont commencé en 1971. Au fil du temps, le navire, l’engin de pêche et les périodes quotidiennes d’échantillonnage ont varié (Chadwick et al., 2007). Les pêches ont été effectuées le jour seulement durant la première partie de la série chronologique (1971-1984), puis on a adopté des pêches sur 24 h. Les résultats de la comparaison des prises ont été utilisés pour ajuster les résultats des relevés pour tenir compte des différences relatives à l’engin de pêche, au navire et aux heures d’opération (Benoît et Swain, 2003a, b; Benoît, 2006). La variation la plus marquée de l’efficacité de capture a été observée lorsqu’on est passé d’un échantillonnage de jour seulement à un échantillonnage sur 24 h.
Les estimations de l’effectif des raies tachetées matures (42 cm et plus LT) dans le sud du golfe variaient autour d’une moyenne d’environ 580 000 individus (sans tendance) dans les années 1970 (voir la figure 7). Les estimations ont commencé à diminuer dans les années 1980, où la moyenne décennale était de 318 000 individus. Le déclin s’est poursuivi, l’effectif moyen étant de 87 000, de 29 000 et de 7 000 dans les années 1990, 2000 et 2010, respectivement. Aucune raie tachetée mature n’a été capturée lors du relevé de 2013. L’indice du relevé a été utilisé à titre d’indice d’abondance principal de l’espèce dans l’UD du golfe du Saint-Laurent puisque très peu de raies ont été capturées lors des relevés dans le nord du golfe (voir ci-dessous).
La pente de régression log-linéaire de l’effectif au cours des 3 générations précédentes (30 ans) était négative (-0,15 an-1, p < 0,0001; voir le tableau 1). On estime que l’effectif durant cette période a connu un déclin de 99 %. D’après les données des 2 dernières générations, la pente estimée était également négative (-0,17 an-1, p = 0,0001), et on estime le déclin à 97 %. La régression de la plus récente génération était également significative (-0,29 an-1, p = 0,0351), et l’effectif a diminué de 95 %. La durée de la plus récente génération correspond environ au temps écoulé depuis la première évaluation du COSEPAC, et les données des tendances indiquent que la population a connu un déclin considérable depuis.
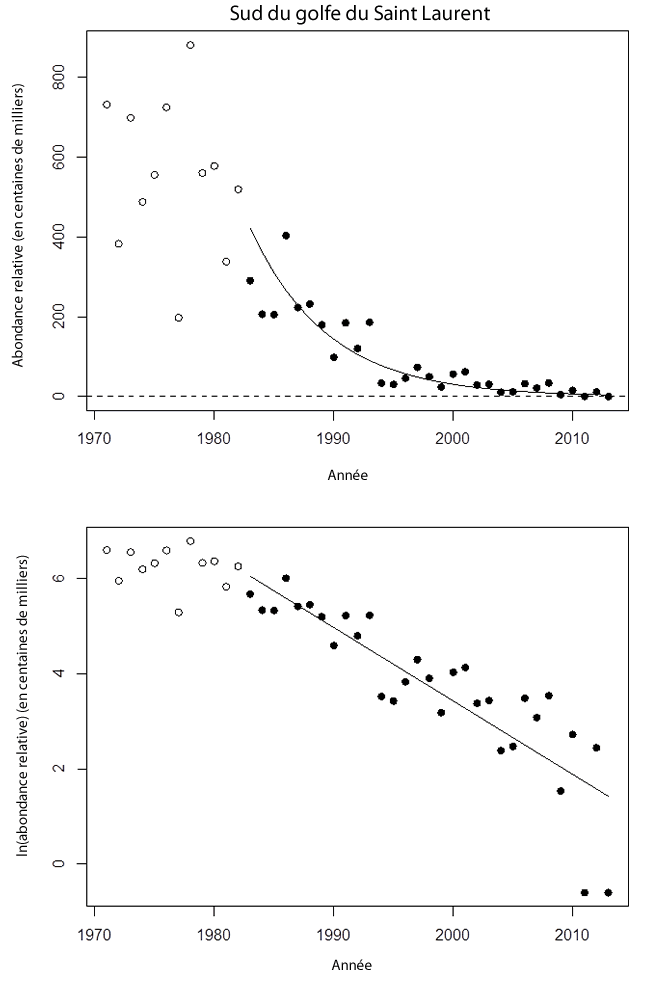
Description longue pour la figure 7
Deux graphiques illustrant les tendances des estimations réalisées lors de relevés au chalut de fond de l’abondance des raies tachetées matures dans le sud du golfe Saint-Laurent (1971-2013). Un graphique présente les résultats sur une échelle arithmétique, tandis que l’autre présente les estimations de l’abondance à l’échelle logarithmique naturelle. Voir Annexe 1a [ou 1b] pour plus d'information.
Description longue pour la figure 8
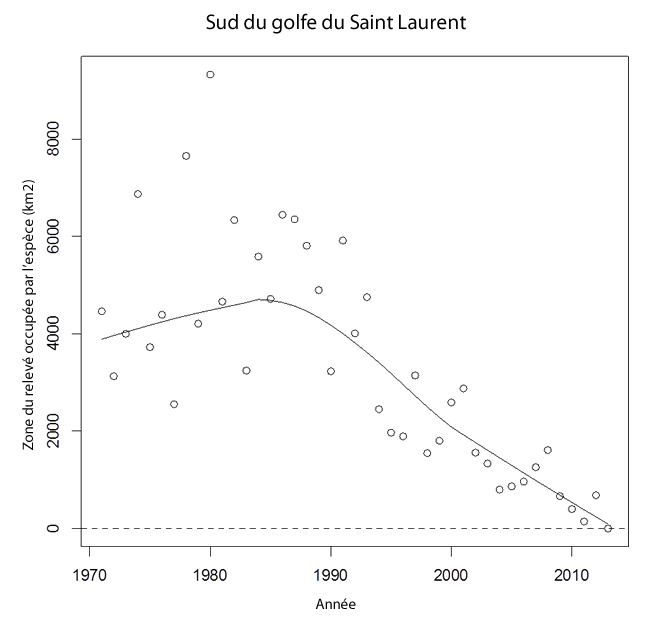
Graphique illustrant les tendances de la zone du relevé occupée par la raie tachetée dans le sud du golfe du Saint Laurent (1971 2013). Durant les années 1970 et 1980, la raie tachetée occupait environ 5 000 km² de cette zone du relevé. La superficie a passé à environ 1 400 km² dans les années 2000 et à 300 km² dans les années 2010. Voir Annexe 1a [ou 1b] pour plus d'information.
Le déclin des effectifs de raies tachetées dans le sud du golfe a été accompagné d’une contraction de la zone d’occupation (voir la figure 8). Au cours des années 1970 et 1980, l’espèce occupait environ 5 000 km² de la superficie couverte annuellement par le relevé. La zone d’occupation est ensuite passée à environ 1 400 km² dans les années 2000 et à quelque 300 km² dans les années 2010.
Au cours des relevés au chalut de fond dans le nord du golfe du Saint-Laurent de 1984 à 2013, 35 traits de pêche sur un total de 6 134 auraient permis de capturer 52 raies tachetées, et ce, 14 années sur 30 seulement. À cause de la trop faible fréquence de ces captures, il n’a pas été possible de générer un indice d’abondance fiable dans ce secteur.
Les relevés au chalut de fond de juillet dans l’est du plateau néo-écossais ont commencé en 1970. Le navire et l’engin de pêche ont été changés en 1982, lorsque le A.T. Cameron, qui utilisait un chalut Yankee 36, a été remplacé par le Lady Hammond et son chalut Western IIA (Chadwick et al., 2007). Une comparaison des prises a été réalisée entre le A.T. Cameron et le Lady Hammond, mais aucun facteur de correction n’a été recommandé pour la raie tachetée (Fanning, 1985). Le relevé a été effectué par le Lady Hammond et deux autres navires de recherche au cours d’années subséquentes au moyen du même engin de pêche et du même protocole d’échantillonnage. Aucune comparaison des prises n’a été réalisée entre ces navires, et les changements relatifs aux opérations étaient relativement mineurs (navire seulement); la période non ajustée a donc été utilisée pour l’analyse des tendances.
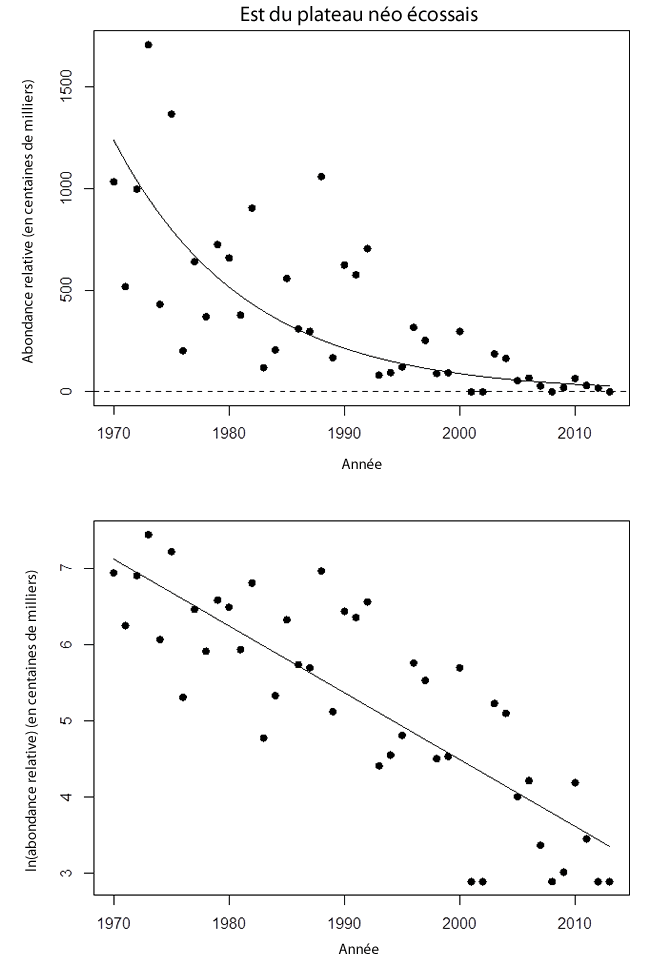
Les estimations annuelles de l’effectif de raies tachetées matures (75 cm et plus LT) dans le secteur ont baissé au cours de la période (voir la figure 9). Les moyennes décennales étaient de 799 000 individus dans les années 1970, de 466 000 dans les années 1980, de 296 000 dans les années 1990, de 82 000 dans les années 2000 et de 38 000 dans les années 2010. Aucune raie tachetée mature n’a été capturée lors des relevés de 2001, 2002, 2008 et 2013. Aux fins de la régression log-linéaire, l’estimation la plus basse de la série chronologique (2012) a été utilisée pour ces 4 années.
La série chronologique couvre seulement les 2,4 dernières générations. La pente de régression log-linéaire de l’effectif au cours de la série complète (43 ans) était négative (-0,088 an-1, p < 0,0001; voir le tableau 1). L’effectif au cours de cette période a diminué de 98 %. D’après les données des 2 dernières générations, la pente estimée était également négative (-0,097 an-1, p < 0,0001), et le déclin aurait été de 97 %. La régression de la plus récente génération était également significative (-0,127 an-1, p = 0,0013), et on a noté un déclin de 90 % de l’effectif.
La zone d’occupation de la raie tachetée dans l’est du plateau néo-écossais a augmenté dans les années 1970 et a atteint un sommet à environ 16 000 km² au milieu des années 1980 (voir la figure 10), malgré une variabilité interannuelle considérable dans les estimations. Elle a diminué au cours du reste de la série chronologique, s’établissant à environ 5 000 km² depuis 2010.
Description longue pour la figure 9
Deux graphiques illustrant les tendances (1970 2013) des estimations réalisées lors de relevés au chalut de fond de l'effectif des raies tachetées matures dans l'est du plateau néo écossais, dans 4VW de l'OPANO. Un graphique présente les résultats sur une échelle arithmétique, tandis que l'autre présente les estimations de l'abondance à l'échelle logarithmique naturelle. Voir Annexe 1a [ou 1b] pour plus d'information.
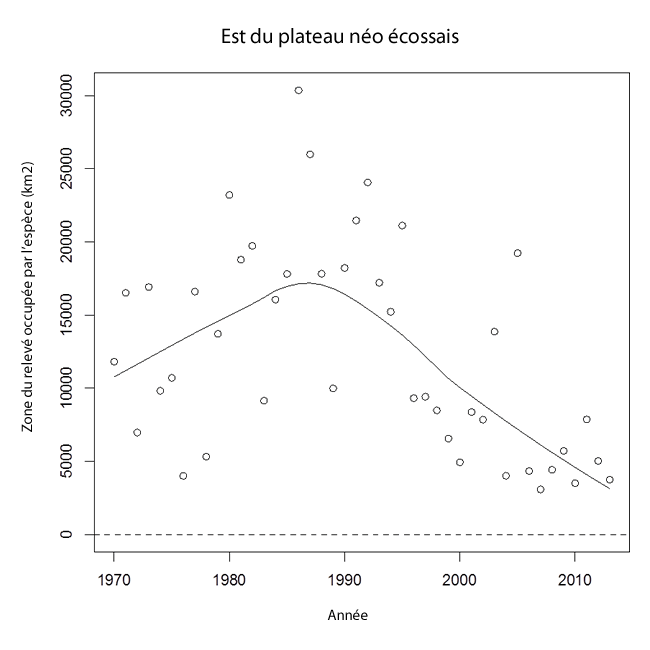
Description longue pour la figure 10
Graphique illustrant les tendances de la zone du relevé occupée par la raie tachetée dans l'est du plateau néo écossais (1970 2013). La zone occupée par la raie tachetée a augmenté durant les années 1970 et a atteint un sommet à environ 16 000 km² au milieu des années 1980. La superficie a ensuite décliné pour le reste de la période jusqu'à une moyenne d'environ 5 000 km² depuis 2010. Voir Annexe 1a [ou 1b] pour plus d'information.
De 1971 à 2013, 17 881 traits ont été effectués dans le cadre de relevés printaniers à bord d’un navire de recherche au sud de Terre-Neuve. Un total de 288 raies tachetées ont été capturées par 137 de ces traits. Aucune raie tachetée n’a été capturée lors des relevés de 1971-1976, 1978-1982, 1984, 2011 et 2013 (voir la figure 11). Le relevé n’a pas été réalisé en 2006, et il n’y a donc pas d’indice pour cette année. Les prises n’ont pas été divisées en classes de taille, et l’indice est fondé sur toutes les raies capturées. Les estimations des années où des raies tachetées ont été capturées étaient très variables, et on recommande donc de faire preuve de prudence lors de l’interprétation des résultats. L’indice d’abondance a augmenté au début des années 1980 et est demeuré relativement élevé (sans tendance) jusqu’en 1995. L’estimation de l’effectif moyen pour la période de 1985 à 1995 est de 64 000 individus. On a changé l’engin de pêche en 1996 en faveur d’un chalut Campelen, réputé être plus efficace pour capturer les poissons, notamment eux de petite taille. Ainsi, comme cela a déjà été fait dans d’autres rapports de situation du COSEPAC (p. ex. merluche blanche [Urophycis tenuis]), les estimations subséquentes (1996-2013) ont été considérées comme faisant partie d’une série chronologique distincte. Durant cette série, on a obtenu des estimations très élevées en 2003 (556 000 individus) et en 2010 (305 000 individus), mais aucune raie tachetée n’a été capturée en 2011 et en 2013. La pente de régression log-linéaire pour la période de 1996 à 2013 était négative (-0,084 an-1, p = 0,0313), et l’effectif a diminué de 76 %. À la limite septentrionale de l’aire de répartition de ce secteur, on a observé des raies tachetées seulement dans une très petite portion de la zone de relevé, et les prises étaient sporadiques, ce qui rend les indices du relevé peu fiables.
Pour résumer les indices d’abondance de cette UD, la plus grande partie de la population a été observée dans l’est du plateau néo-écossais lors des premières années des relevés (1970-1990). Cette portion de la population a subi un très fort déclin de 98 % au cours des 2,4 dernières générations. Il semble y avoir eu un déclin de l’effectif au sud de Terre-Neuve depuis l’utilisation d’un nouvel engin de pêche, soit en 1996, même si cet indice a varié considérablement d’une année à l’autre. L’indice du relevé dans l’est du plateau néo-écossais a couvert la plus longue période, et les raies tachetées étaient plus abondantes à cet endroit dans les années 1970 que dans le sud de Terre-Neuve. Par conséquent, l’indice de l’est du plateau néo-écossais a été utilisé comme principal indicateur pour cette UD.
Description longue pour la figure 11
Deux graphiques illustrant les tendances des estimations réalisées lors de relevés au chalut de fond de l'effectif des raies tachetées matures dans le sud de Terre Neuve, dans 3LNOP de l'OPANO (1971 2013). Un graphique présente les résultats sur une échelle arithmétique, tandis que l'autre présente les estimations de l'abondance à l'échelle logarithmique naturelle. Différents symboles montrent les types de chaluts utilisés (Yankee, Engel, Campelen). Voir Annexe 1a [ou 1b] pour plus d'information.
Deux relevés au chalut de fond ont servi à estimer les tendances de l’effectif des raies tachetées adultes dans cette UD, c’est-à-dire le relevé de juillet dans l’ouest du plateau néo-écossais et la baie de Fundy (1970-2012) et le relevé de mars du MPO sur le banc Georges (1987-2012). Le relevé de l’ouest du plateau néo-écossais et de la baie de Fundy a été réalisé à bord du même navire et au moyen du même engin de pêche et des mêmes protocoles décrits plus haut pour le relevé de l’est du plateau néo-écossais, et aucun ajustement n’a été fait aux résultats du relevé. Seuls les résultats de la portion canadienne du banc Georges ont été utilisés.
Les estimations annuelles de l’effectif des individus matures (75 cm et plus LT) dans le cadre du relevé de l’ouest du plateau néo-écossais et de la baie de Fundy étaient très variables, et on n’a discerné aucune tendance apparente. Les estimations décennales moyennes étaient de 107 000 individus dans les années 1970, de 91 000 dans les années 1980, de 81 000 dans les années 1990, de 47 000 dans les années 2000 et de 57 000 dans les années 2010 (voir la figure 12). Aucune raie tachetée mature n’a été observée lors du relevé du 1971. Dans ce cas, la valeur nulle (0) a été remplacée par la valeur la plus faible de la série chronologique (2002) aux fins de la régression log-linéaire. Cette série couvre seulement 2,4 générations. L’estimation de la pente log-linéaire n’était pas statistiquement significative (-0,015 an-1, p = 0,15), et on estime que l’effectif a diminué de 53 %.
Les estimations annuelles de la zone d’occupation de la raie tachetée de l’ouest du plateau néo-écossais étaient très variables (sans tendance nette) (voir la figure 13). Celle-ci a légèrement augmenté au début des années 1990 et était en moyenne, au cours des 5 dernières années (2009-2013) du relevé, de 9 100 km².
Les estimations initiales du relevé du banc Georges à la fin des années 1980 étaient de 400 000 individus matures (voir la figure 14). Ce nombre a diminué, s’établissant à moins de 100 000 individus au milieu des années 1990, avant d’augmenter et d’atteindre environ 200 000 individus. Trois estimations très élevées ont été réalisées de 2008 à 2010, la plus élevée dépassant 1,2 million d’individus. On ne sait pas pourquoi les valeurs étaient aussi élevées, mais celles-ci pourraient être indicatrices d’un changement dans l’aire de répartition de l’espèce, dont certains individus des États-Unis se déplacent vers d’autres zones (Frisk et al., 2008). Des raies matures ont été capturées lors de tous les relevés menés sur le banc Georges.
La série chronologique du relevé sur le banc Georges couvre seulement 1,4 génération. La régression log-linéaire n’a pas donné une estimation de la pente significative (0,013 an-1, p = 0,52). On a estimé une hausse de 39 % de l’effectif au cours de cette période.
La zone d’occupation de la raie tachetée sur le banc Georges n’a affiché aucune tendance depuis le début des relevés, en 1987 (voir la figure 15) et était en moyenne, au cours des 5 dernières années du relevé (2009-2013), de 19 200 km².
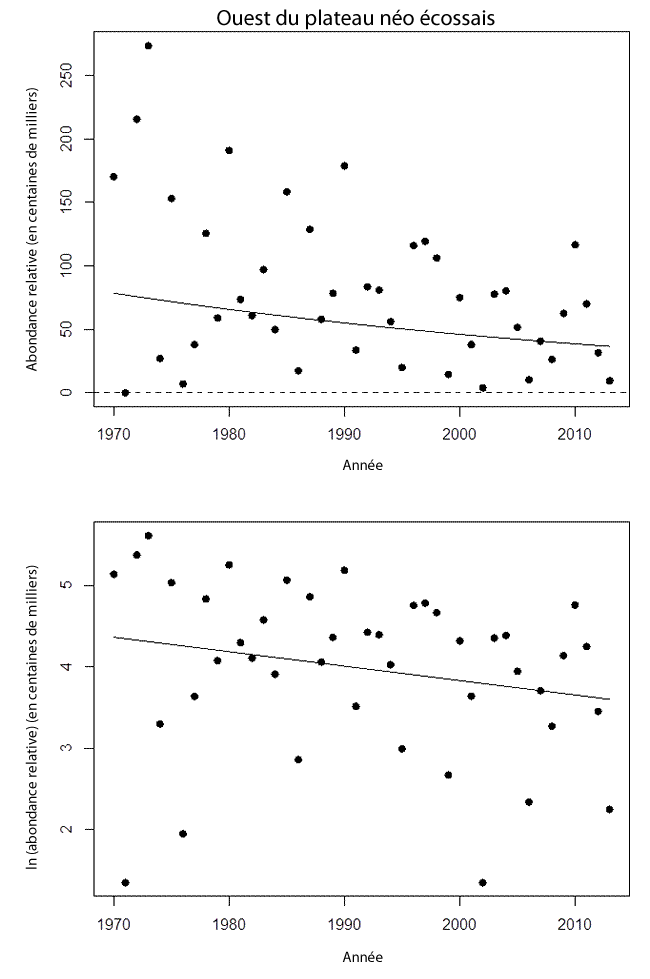
Description longue pour la figure 12
Deux graphiques illustrant les tendances des estimations réalisées lors de relevés au chalut de fond de l'effectif des raies tachetées matures dans l'ouest du plateau néo écossais et la baie de Fundy, dans 4X de l'OPANO (1970 2012). Un graphique présente les résultats sur une échelle arithmétique, tandis que l'autre présente les estimations de l'abondance à l'échelle logarithmique naturelle. Voir Annexe 1a [ou 1b] pour plus d'information.
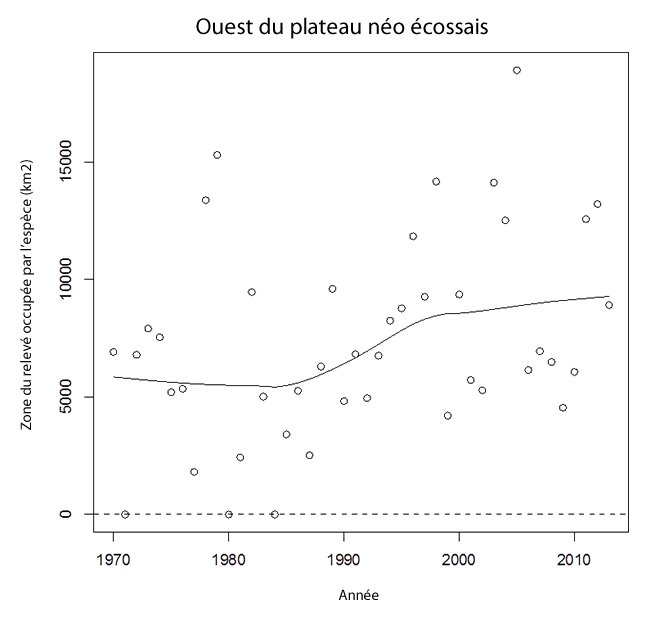
Description longue pour la figure 13
Graphique illustrant les tendances de la zone du relevé occupée par la raie tachetée dans l'ouest du plateau néo écossais et la baie de Fundy (1970 2013). Les estimations annuelles de la zone occupée par la raie tachetée dans l'ouest du plateau néo écossais étaient très variables. On n'a noté aucune tendance particulière, sauf une légère augmentation de la zone occupée au début des années 1990. La moyenne de la zone occupée de 2009 à 2013 était de 9 100 km². Voir Annexe 1a [ou 1b] pour plus d'information.
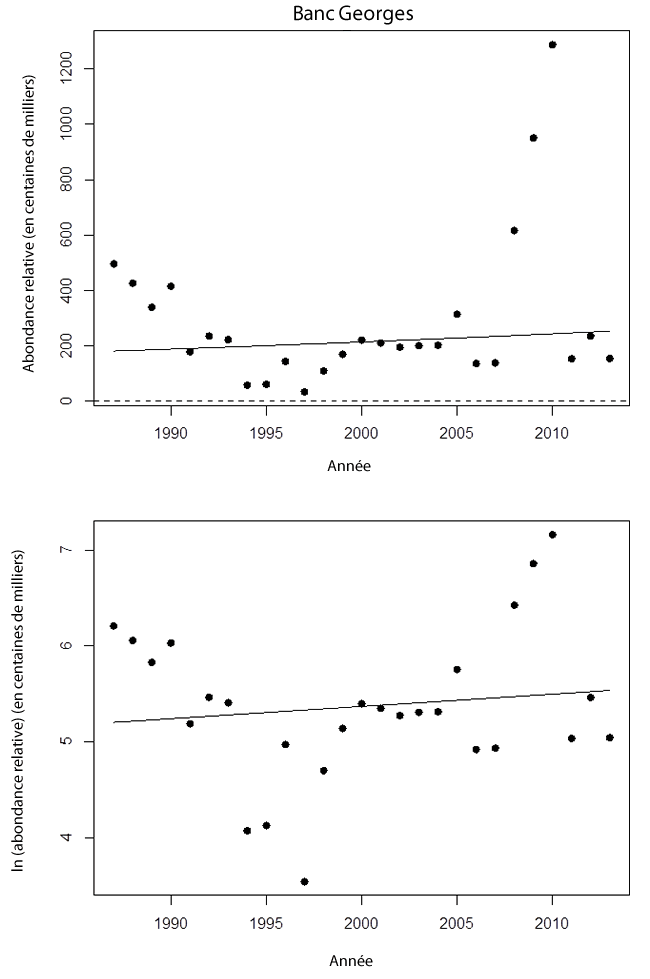
Description longue pour la figure 14
Deux graphiques illustrant les tendances des estimations réalisées lors de relevés canadiens au chalut de fond de l'effectif des raies tachetées matures dans la portion canadienne du banc Georges (1987 2012). Un graphique présente les résultats sur une échelle arithmétique, tandis que l'autre présente les estimations de l'abondance à l'échelle logarithmique naturelle. Voir Annexe 1a [ou 1b] pour plus d'information.
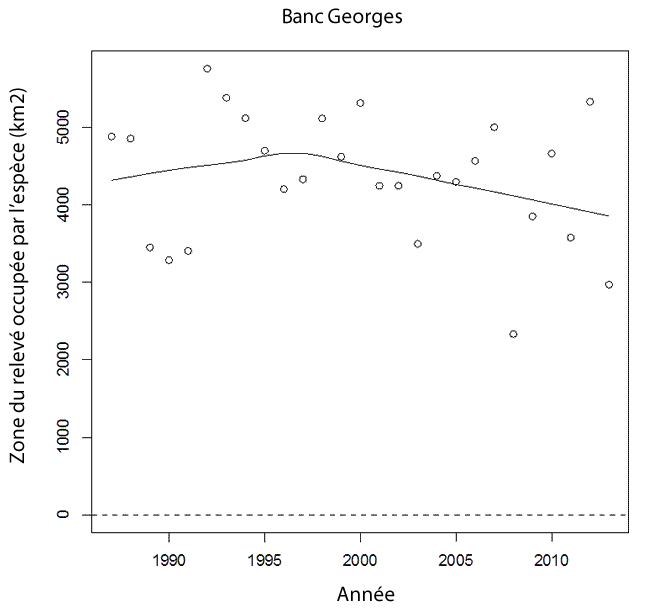
Description longue pour la figure 15
Graphique illustrant les tendances de la zone du relevé occupée par la raie tachetée sur le banc Georges (1987 2012). Aucune tendance n'a été observée dans la zone occupée. Voir Annexe 1a [ou 1b] pour plus d'information.
Pour résumer ces 2 indices, le nombre d’individus matures dans l’UD de l’ouest du plateau néo-écossais et du banc Georges semble être relativement stable, et on ne discerne pas de tendance. L’indice du relevé au chalut de fond dans l’ouest du plateau néo-écossais est très variable et, tandis qu’on estime une tendance au déclin (53 %) au cours des 2,4 dernières générations, la pente de régression n’est pas significative sur le plan statistique. Sur le banc Georges, le relevé révèle une tendance globale à la hausse (39 %) mais, encore une fois, la pente de régression est statistiquement non significative. Les estimations tirées du relevé sur le banc Georges sont environ 4 fois supérieures à celles tirées du relevé dans l’ouest du plateau néo-écossais au cours de la même période (1987-2013), et la tendance dans ce secteur zone refléterait mieux la tendance de l’UD entière. Cependant, contrairement aux autres UD, la durée de la série chronologique considérée dans la régression influe sur les inférences visant à déterminer si la tendance du nombre d’individus matures dans l’ouest du plateau néo-écossais et sur le banc Georges est significative ou non (voir le tableau 1).
Les indices de biomasse de la raie tachetée déterminés lors des relevés d’automne du Northeast Fisheries Science Center (NEFSC) montrent deux fortes augmentations de l’effectif, lesquelles sont suivies de déclins (Sosebee, 2014; voir la figure 16). La première augmentation a commencé au milieu des années 1970 et a atteint un sommet au milieu des années 1980. L’indice a ensuite diminué considérablement jusqu’au milieu des années 1990. La deuxième augmentation notable a eu lieu de 2006 à 2009. Les estimations subséquentes ont diminué de façon importante. Cette grande variabilité est difficile à expliquer pour une espèce caractérisée par une productivité aussi faible; c’est pourquoi elle pourrait refléter des changements dans la répartition et/ou l’efficacité des relevés (Frisk et al., 2008).
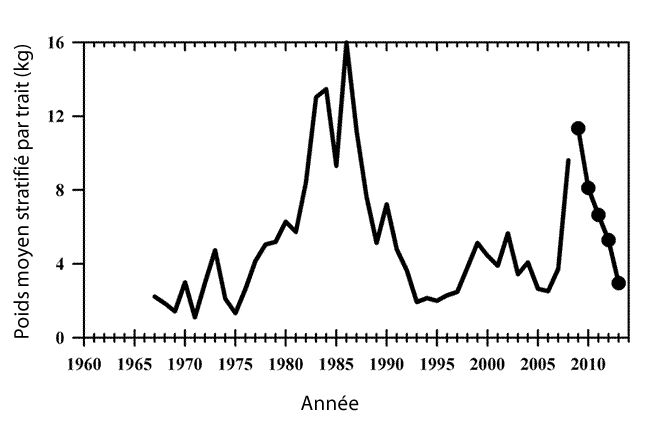
Description longue pour la figure 16
Graphique illustrant les indices de biomasse de la raie tachetée (1967 2013) dans les eaux états uniennes, selon les relevés automnaux effectués par le Northeast Fisheries Science Center. Le graphique montre une grande variabilité des tendances, avec des sommets importants au milieu des années 1980 et de 2006 à 2009. Les estimations subséquentes ont grandement diminué.
Les Élasmobranches, dont font partie les raies, seraient très vulnérables à l’exploitation à cause de leur grande taille et des caractéristiques connexes de leur cycle vital (p. ex. croissance lente, maturation tardive, faible fécondité par rapport aux poissons osseux) (Dulvy et Reynolds, 2002; Reynolds, 2003). McPhie et Campana (2009) ont comparé les caractéristiques démographiques et le taux de croissance intrinsèque de la raie tachetée dans l’est du plateau néo-écossais à ceux de trois espèces de raies sympatriques : la raie hérisson, la raie épineuse (Amblyraja radiata) et la raie à queue de velours (Malacoraja senta). La raie tachetée est la moins productive et la plus vulnérable à la surpêche, et c’est chez cette espèce que l’effectif des adultes a le plus baissé.
La raie tachetée n’est actuellement pas pêchée à des fins commerciales dans le sud du golfe du Saint-Laurent, et il est maintenant obligatoire que tous les individus capturés à titre de prises accessoires soient remis à l’eau. On ne connaît pas les taux de mortalité après la remise à l’eau. Des raies tachetées faisaient partie des prises accessoires des pêches des poissons de fond et de la pêche aux pétoncles, ce qui peut avoir contribué au déclin de l’espèce dans le sud du golfe durant les années 1970 et 1980. Swain et al. (2006) ont estimé les rejets à plus de 1 million de raies au début des années 1970. Ce nombre a baissé, s’établissant à 500 000-700 000 raies à la fin des années 1970 et dans les années 1980 (voir la figure 17). Les quantités de raies rejetées ont grandement diminué après la fermeture des pêches des poissons de fond au début des années 1990, et elles totalisent moins de 100 000 individus depuis 1992. En 2002-2004, les prises accessoires étaient inférieures à 10 000 raies. Les prises accessoires des pêches des poissons de fond estimées récemment demeurent très faibles, à moins de 2 tonnes annuellement pour la période de 2005 à 2011 (Benoît, 2013). Une récente analyse des prises accessoires dans le cadre de la pêche aux pétoncles dans la région indique aussi que très peu (moins de 1 tonne) de raies tachetées sont capturées (Benoît et al., 2010a).
Des raies sont observées dans les débarquements des pêches pratiquées dans l’est du plateau néo-écossais depuis la fin des années 1960; cependant, la composition en espèces n’est pas mentionnée dans les statistiques officielles. Une grande proportion des diverses espèces de raies capturées ont également été rejetées à la mer. Par conséquent, on a utilisé les données recueillies par les observateurs de pêches en mer pour estimer les prises et les rejets par espèce. Au cours des années 1970 et 1980, environ 2 500 tonnes de raies tachetées ont été capturées annuellement par les diverses pêches dirigées d’autres poissons de fond dans l’est du plateau néo-écossais (Swain et al., 2006; voir la figure 18). Après la fermeture de plusieurs pêches des poissons de fond au début des années 1990, les prises accessoires de raies tachetées ont quelque peu diminué. Cependant, une pêche dirigée des raies a commencé en 1994, et les prises étaient composées à environ 90 % de raies tachetées (Simon et al., 2003). Les débarquements de cette pêche a atteint un sommet en 1994, avec 2 200 tonnes. Ils ont diminué rapidement par la suite, et la pêche dirigée a été fermée en 2006. Les prises annuelles de raies tachetées étaient de 1 700 tonnes dans les années 1990, et de 300 tonnes, dans les années 2000. Il est possible que la surpêche dans les années 1980 et 1990 ait contribué au déclin de l’effectif des raies tachetées dans l’est du plateau néo-écossais. Il se peut également que, même si le nombre estimé de raies tachetées pêchées de 2000 à 2004 a été faible, ce nombre ait été suffisant pour décimer davantage la population (Swain et al., 2009).
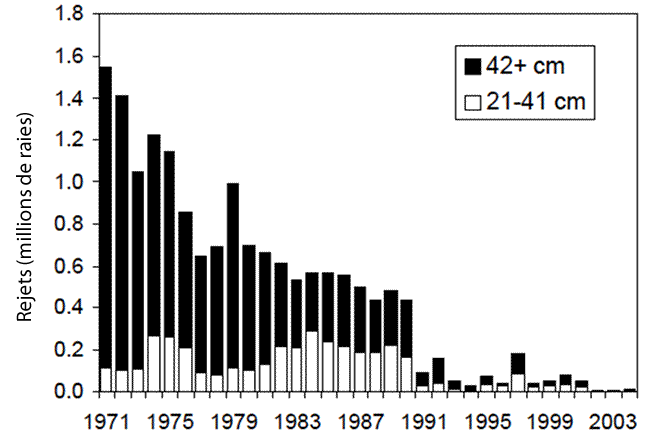
Description longue pour la figure 17
Graphique illustrant les tendances relatives aux rejets estimés de raies tachetées par les pêches des poissons de fond dans le sud du golfe du Saint Laurent par classes de taille (1971 2004). Les rejets ont diminué au cours de la période, passant de plus de 1 million au début des années 1970 à moins de 10 000 de 2002 à 2004.
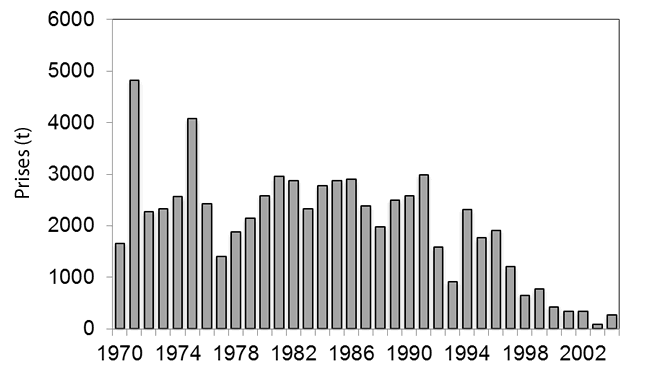
Description longue pour la figure 18
Graphique illustrant les tendances relatives aux prises estimées de raies tachetées dans l'est du plateau néo écossais (1970 2004), en tonnes.
Il n’y a aucune estimation des prises de raies tachetées dans le secteur de l’UD de l’ouest du plateau néo-écossais et du banc Georges. Les débarquements annuels rapportés pour toutes les espèces de raies dans la base de données de l’OPANO montrent un pic de 1973 à 1976, avec une moyenne annuelle de 964 tonnes. Depuis, les débarquements annuels rapportés pour toutes les raies sont de 40 tonnes en moyenne. Comme l’effectif dans ce secteur n’affiche aucune tendance, la mortalité attribuable aux pêches ne semble pas constituer une menace pour les raies tachetées de l’UD de l’ouest du plateau néo-écossais et du banc Georges.
Le déclin rapide de la raie tachetée dans le sud du golfe du Saint-Laurent et dans l’est du plateau néo-écossais a incité à analyser en détail les causes possibles (Swain et al., 2009). Un modèle d’espace d’états structuré en fonction du cycle vital a été utilisé pour examiner les variations du taux de mortalité (due aux pêches ou à d’autres causes que les pêches [M]) dépendantes du temps. L’analyse du secteur du sud du golfe a étudié 2 classes d’âge : les juvéniles (< 42 cm LT) et les adultes (42 cm et plus TL). Dans le cas de l’analyse de l’est du plateau néo-écossais, on a divisé les juvéniles en 2 classes (39-59 cm LT et 60-74 cm LT) et on a créé une classe pour les adultes (75 cm et plus LT).
On a estimé que la mortalité des juvéniles due aux pêches dans le sud du golfe était très faible au cours de la période (1971-2004). Pour ce qui est des adultes, la mortalité due aux pêches aurait été relativement élevée dans les années 1970, aurait baissé par la suite et, enfin, aurait été très faible au cours des 2 dernières décennies de l’analyse. Les estimations de la mortalité des adultes due à d’autres causes que les pêches (M) ont augmenté tout au long de la série chronologique. L’estimation médiane pour la période de 1990 à 2004 était de 0,6 an-1. Les causes possibles de cette augmentation ont été examinées, y compris les changements dans l’effectif des prédateurs (morue [Gadus morhua], phoque gris) et les changements dans les conditions environnementales reflétés par les températures du fond. De 1971 à 2004, la présence de phoques gris dans le sud du golfe a augmenté d’un facteur de près de 2,5 (Swain et al., 2009). L’augmentation de l’effectif des phoques gris serait la cause la plus probable de la hausse de la mortalité des raies tachetées adultes due à des causes autres que les pêches. On fonde cette hypothèse sur une corrélation qui est plus forte au fil du temps entre la variable M des raies adultes et les populations de phoques qu’entre la variable M et les températures à la hausse. Des projections démographiques de 2004 à 2014 ont indiqué que, si la récente hausse de la mortalité due à des causes autres que les pêches continuait, la population déclinerait même en l’absence de pêches. Ces projections sont appuyées par la tendance observée de l’indice tiré des relevés au chalut de fond réalisés ces 10 dernières années, et laissent croire à la disparition imminente de cette population (Kelly et Hanson, 2013a). Si la prédation par les phoques est réellement la principale cause de cette mortalité non liée aux pêches anormalement élevée et qu’elle est responsable du déclin continu et rapide de 25 % par année, il y a lieu de conclure qu’il y a 1 localité pour la population, d’après les critères de l’UICN, où le nombre de localités est lié au nombre de menaces.
Dans l’est du plateau néo-écossais, on a estimé que la mortalité due aux pêches était relativement faible au début et à la fin de la série chronologique période (1970-2004) pour les 3 classes de taille. La mortalité due aux pêches des 2 classes de taille les plus longues était la plus élevée durant les pêches dirigées du milieu des années 1990. La mortalité des adultes due à des causes autres que les pêches aurait augmenté tout au long de la série, et l’estimation médiane pour la période de 1990 à ce jour est de 0,35 an-1. Les causes possibles de cette augmentation ont été examinées, y compris la variation des effectifs de prédateurs (morue et phoque gris) et les changements environnementaux reflétés par les températures du fond. De 1971 à 2004, l’effectif des phoques gris dans le sud du golfe a plus que décuplé (Swain et al., 2009). Cette augmentation serait la cause la plus probable de la hausse de la mortalité des raies tachetées adultes due à des causes autres les pêches; on n’a en effet noté aucune corrélation entre la hausse de la mortalité des raies et la température de la mer. Des projections démographiques de 2004 à 2014 ont indiqué que l’effectif des adultes diminuerait un peu, mais se stabiliserait à un faible niveau en l’absence de pêches. Des prises annuelles continues de 300 tonnes, comme on a observé de 2002 à 2004, entraîneraient un déclin continu de l’effectif des adultes. L’ampleur du déclin de l’indice tiré des relevés au chalut de fond montre que les prises n’ont pas été réduites à zéro. Pour cette population, le maintien de la mortalité à la hausse due à la prédation, probablement par le phoque gris, et la mortalité due aux prises accessoires constituent les principales menaces pour cette population.
Il existe un grand nombre de localités (> 10) pour cette UD, d’après les critères de l’UICN, qui lient les localités aux menaces. La prédation par le phoque gris, à elle seule, n’est pas susceptible d’influer rapidement sur toutes les raies tachetées et ne peut donc pas mener à la désignation d’une seule localité pour cette UD. De plus, étant donné que les prises accessoires sont causées par un grand nombre de navires de pêche soumis à diverses mesures de gestion (p. ex. total admissible de captures, saisons, limites des prises accessoires), on compte plus de 10 localités pour cette population.
La propagation du crabe vert (Carcinus maenus), originaire d’Europe, pourrait avoir des conséquences sur la raie tachetée puisque l’aire de répartition de ce crabe chevauche celle d’autres crabes, du C. irrorus et d’autres proies de la raie tachetée, entraînant ainsi une compétition entre toutes ces espèces (question examinée par Klassen et Locke, 2007).
Le COSEPAC a évalué la raie tachetée et défini quatre UD en 2005 (COSEWIC, 2005). L’UD du sud du golfe du Saint-Laurent a été classée dans la catégorie « en voie de disparition »; l’UD de l’est du plateau néo-écossais, « menacée »; l’UD du banc Georges, de l’ouest du plateau néo-écossais et de la baie de Fundy, « préoccupante »; l’UD du nord du golfe Saint-Laurent et de Terre-Neuve, « données insuffisantes ». En 2010, le gouverneur en conseil a décidé de ne pas ajouter la raie tachetée à l’annexe de la Loi sur les espèces en péril (LEP) parce que l’inscription entraînerait la perte de millions de dollars en revenus ainsi qu’une perte importante directe et indirecte d’emplois (Canada Gazette, 2010). Il a plutôt été prévu de gérer la mortalité de la raie tachetée due aux pêches aux termes de la Loi sur les pêches. Dans ce contexte, on a fermé la pêche commerciale dirigée des raies; de plus, les raies tachetées doivent être rejetées à l’eau, et les taux de rejet font l’objet d’un suivi. Le taux de survie des raies tachetées rejetées à l’eau est inconnu. La décision n’a apporté aucune valeur intrinsèque à l’espèce.
En 2015, le COSEPAC a évalué la raie tachetée et défini trois UD distinctes, soit la population du golfe du Saint-Laurent (« en voie de disparition »), la population de l’est du plateau néo-écossais et de Terre-Neuve (« en voie de disparition ») et la population de l’ouest du plateau néo-écossais et du banc Georges (« non en péril »).
Au Québec, la raie tachetée n’est pas inscrite à titre d’espèce menacée ou vulnérable en vertu de la Loi sur les espèces menacées ou vulnérables (RLRQ, c E-12.01) (MRNF, 2014a). Cependant, l’espèce fait partie de la Liste des espèces susceptibles d’être désignées menacées ou vulnérables, qui est produite en vertu de cette même loi (MRNF, 2014b). Au Nouveau-Brunswick, la raie tachetée du sud du golfe du Saint-Laurent est inscrite comme « menacée », et celle de l’ouest du plateau néo-écossais et de la baie de Fundy, comme « préoccupante ».
L’UICN a évalué la raie tachetée comme étant « en voie de disparition » dans toute son aire de répartition (Kulka et al., 2009).
Jim Simon, Pêches et Océans Canada, Institut océanographique de Bedford, Dartmouth (Nouvelle-Écosse)
Isabelle Gauthier, ministère des Forêts, de la Faune et des Parcs, Québec
Doug Swain (Ph.D.), Pêches et Océans Canada, Centre des pêches du Golfe, Moncton (Nouveau-Brunswick)
Hugues Benoît, Pêches et Océans Canada, Centre des pêches du Golfe, Moncton (Nouveau-Brunswick)
Christie Whelan, Pêches et Océans Canada, Ottawa (Ontario)
Tara McIntyre, Pêches et Océans Canada, Institut océanographique de Bedford, Dartmouth (Nouvelle-Écosse)
Mark Simpson (Ph.D.), Pêches et Océans Canada, Centre des pêches de l’Atlantique Nord-Ouest, St. John’s (Terre-Neuve-et-Labrador)
Denis Bernier, Pêches et Océans Canada, Institut Maurice-Lamontagne, Mont-Joli (Québec)
Beck, C.A., S.J. Iverson, W.D. Bowen et W. Blanchard. 2007. Sex differences in grey seal diet reflect seasonal variation in foraging behaviour and reproductive expenditure: evidence from quantitative fatty acid signature analysis. Journal of Animal Ecology 76: 490-502.
Benoît, H.P. 2013. Two decades of annual landed and discarded catches of three southern Gulf of St Lawrence skate species estimated under multiple sources of uncertainty. ICES Journal of Marine Science 70: 554-563.
Benoît, H.P. 2006. Standardizing the southern Gulf of St. Lawrence bottom-trawl survey time series: results of the 2004-2005 comparative fishing experiments and other recommendations for the analysis of the survey data. DFO Can. Sci. Advis. Sec. Res. Doc. 2006/008. iv + 127 p. (Le titre ainsi qu’un résumé du document sont disponibles en français : Normalisation de la série chronologique des relevés au chalut de fond effectués dans le sud du golfe du Saint-Laurent : résultats d’études de pêche comparatives de 2004-2005 et autres recommandations pour l’analyse des données des relevés. Pêches et Océans Canada, Secrétariat canadien de consultation scientifique. Document de recherche 2006/008.)
Benoît, H.P. et Swain, D.P. 2003a. Accounting for length- and depth-dependent diel variation in catchability of fish and invertebrates in an annual bottom-trawl survey. ICES J. Mar. Sci. 60: 1298-1317.
Benoît, H.P. et Swain, D.P. 2003b. Standardizing the southern Gulf of St. Lawrence bottomtrawl survey time series: adjusting for changes in research vessel, gear and survey protocol. Can. Tech. Rep. Fish. Aquat. Sci. No. 2505: iv + 95 p. (Comprend un résumé en français.)
Benoît, H.P., D.P. Swain, M. Niles, S. LeBlanc, L.-A. Davidson. 2010. Incidental catch amounts and porential post-release survival of winter skate (Leucoraja ocellataI) captured in the scallop dredge fishery in the southern Guld of St. Lawrence (2006-2008). CSAS Res. Doc. 2010/043 (Le titre ainsi qu’un résumé du document sont disponibles en français : Quantité de prises accidentelles et survie éventuelle après le rejet à la mer des raies tachetées (Leucoraja ocellata) capturées lors de la pêche de pétoncles à la drague dans le sud du golfe du Saint-Laurent (2006-2008). Secrétariat canadien de consultation scientifique – Document de recherche 2010/043.)
Bigelow, H.B. et W.C. Schroeder. 1953. Fishes of the Western North Atlantic. Sawfishes, guitarfishes, skate and rays, chimaeroides, Memoir Sears Foundation Marine Research.
Canada Gazette. 2010. Order in Council giving notice of decisions not to add certain species to the list of endangered species, vol. 144, no 6, 17 mars 2010. (Également disponible en français : Décret donnant avis des décisions de ne pas inscrire certaines espèces sur la Liste des espèces en péril. Gazette du Canada, vol. 144, no 6 – 17 mars 2010.)
Chadwick, E.M.P., W. Brodie, D. Clark, D. Gascon et T.R. Hurlbut. 2007. History of annual multi-species trawl surveys on the Atlantic coast of Canada, Atlantic Zone Monitoring Program 6, Fisheries and Oceans Canada, Mont-Joli, Que. (Également disponible en français : Historique des relevés de chalut multi-spécifiques annuels sur la côte Atlantique du Canada. Bulletin PMZA 6: 25-41. Programme de monitorage de la zone Atlantique. Pêches et Océans Canada.)
Clay, D. 1991. Seasonal distribution of demersal fish (Osteichthyes) and skates (Chondrichthyes) in the southeastern Gulf of St. Lawrence. Canadian special publication of fisheries and aquatic sciences 113: 241-259. (Comprend un résumé en français : In J.-C. Therriault (éd.) Le Golfe du Saint-Laurent : petit océan ou grand estuaire? Compte rendu d’un atelier/symposium tenu à l’Institut Maurice-Lamontagne (Mont-Joli) du 14 au 17 mars 1989. Publication spéciale canadienne des sciences halieutiques et aquatiques 113.)
Collette, B.B. et G. Klein-MacPhee. 2002. Bigelow and Schroeder’s Fishes of the Gulf of Maine. Smithsonian Press. Washington.
COSEWIC. 2005. COSEWIC assessment and status report on the Winter Skate Leucoraja ocellata in Canada. Committee on the Status of Endangered Wildlife in Canada. vii + 41 pp. (Également disponible en français : Évaluation et Rapport de situation du COSEPAC sur la raie tachetée (Leucoraja ocellata) au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. viii + 47 p.)
COSEWIC. 2011. Guidelines for recognizing designatable units below the species level. (Également disponible en français : Lignes directrices pour reconnaître les unités désignables.
Darbyson, E. et Benoît, H.P. 2003. An atlas of the seasonal distribution of marine fish and invertebrates in the sGSL. Can. Data Rep. Fish. Aquat. Sci. 1113: 294 pp. (Comprend un résumé en français.)
DFO. 2013. Oceanographic conditions in the Atlantic zone in 2012. DFO Can. Sci. Advis. Sec. Sci. Advis. Rep. 2013/057. (Également disponible en français : Conditions océanographiques dans la zone Atlantique en 2012. Pêches et Océans Canada, Secrétariat canadien de consultation scientifique – Avis scientifique 2013/057.)
Dulvy, N.K. et J.D. Reynolds. 2002. Predicting extinction vulnerability in skates. Conservation Biology 16: 220-450.
Dutil, J.-D., R. Miller, C. Nozères, B. Bernier, D. Bernier et D. Gascon. 2006. Révision des identifications de poisson faites lors des relevés scientifique annuels de l’abondance des poissons de fonds et de la crevette nordique dans l’estuaire et le nord du golf du Saint-Laurent. Rapport manuscrit canadien des sciences halieutique et aquatiques 2760.
Fanning L.P. 1985. Intercalibration of Research Survey Results Obtained from Different Vessels. CAFSAC Rese Doc 85/03. (Comprend un résumé en français.)
Frisk, M.G., T.J. Miller, S.J.D. Martell et K. Sosebee. 2008. New hypothesis helps explain elasmobranch “outburst” on Georges Bank in the 1980s. Ecological Applications 18: 234-245.
Galbraith, P.S., J. Chassé, P. Larouche, D. Gilbert, D. Brickman, B. Pettigrew, L. Devine et C. Lafleur. 2013. Physical oceanographic conditions in the Gulf of St. Lawrence in 2012. Canadian Science Advisory Secretariat Research Document 2013/026.
Hammill, M.O. et G.B. Swenson. 2000. Estimated prey consumption by Harp seals (Phoca groenlandica), Hooded seals (Cystophora cristata), Grey seals (Halichoerus grypus) and Harbour seals (Phoca vitulina) in Atlantic Canada. J. Northw. Fish. Sci. 26: 1-23.
IUCN. 2013. IUCN Standards and Petitions Subcommittee. Guidelines for Using the IUCN Red List Categories and Criteria [PDF; 1.2 Mo] (en anglais seulement). Version 10. Prepared by the Standards and Petitions Subcommittee.
Kelly, J.T. et J.M. Hanson. 2013a. Abundance, distribution, and habitat characteristics of winter skate Leucoraja ocellata in the southern Gulf of St. Lawrence: a population on the brink of extinction? Journal of Fish Biology 82: 877-892.
Kelly, J.T. et J.M. Hanson. 2013b. Maturity, size at age and predator-prey relationships of winter skate Leucoraja ocellata in the southern Gulf of St. Lawrence: potentially an undescribed endemic facing extirpation. Fish Biology 82: 959-978.
Klassen, G. et A. Locke. 2007. A biological synopsis of the European Green Crab, Carcinus maenus. Canadian Manuscript Report of Fisheries and Aquatic Sciences No. 2818. (Comprend un résumé en français.)
Kulka, D. W., J. Sulikowski, et T. Gedamke. 2009. Leucoraja ocellata. In IUCN 2012. IUCN Red List of Threatened Species. Version 2012.2.
Mahon, R., R.W. Smith, B.B. Berstein et J.S. Scott. 1984. Spatial and temporal patterns of groundfish distribution on the Scotian Shelf and in the Bay of Fundy, 1970-1981. Can. Tech. Rep. Fish. Aqua. Sci. 1300, xi + 164 p. (Comprend un résumé en français.)
McEachran, J.D. et J.A. Martin. 1975. Distribution and relative abundance of seven species of skates (Pisces: Rajidea) which occur between Nova Scotia and Cape Hatteras. Fishery Bulletin 73: 110-136.
McEachran, J.D., D.F. Boesch et J.A. Musick. 1976. Food division within two sympatric species-pairs of skate (Pisces: Rajidae). Marine Biology. 35: 301-17.
McEachran, J.D. et C.O. Martin. 1977. Possible occurrence of character displacement in the sympatric skate Raja erinacea and Raja ocellata (Pisces: Rajidae). Environment and Biology of Fishes 2: 121-130.
McPhie, R.P. et S.E. Campana. 2009. Reproductive characteristics and population declines of four species of skate (Rajidae) off the eastern coast of Canada. Journal of Fish Biology 75: 223-246.
MRNF, 2014a. Liste des espèces désignées menacées ou vulnérables au Québec.
MRNF, 2014b. Liste des espèces désignées menacées ou vulnérables au Québec – Poissons.
Reynolds, J.D. 2003. Life histories and extinction risk. In Macroecology (eds K.J. Gaston et T.J. Blackburn), p. 195-217. Blackwell Publishing, Oxford.
Robins, C.R. et G.C. Ray. 1986. A field guide to Atlantic coast fishes of North America. Houghton Mifflin Company, Boston, U.S.A.
Scott, J.S. 1982. Depth, temperature and salinity preferences of common fishes of the Scotian Shelf. Journal of Northwest Atlantic Fishery Science. 3: 29-39.
Scott, W.B. et M.G. Scott. 1988. Atlantic Fishes of Canada. University of Toronto Press. Ontario, Canada.
Simon, J.E. et K.T. Frank. 1996. Assessment of the Division 4VsW skate fishery. DFO Atl. Fish. Res. Doc. 96/105. (Comprend un résumé en français.)
Simon, J.E. et K.T. Frank. 1998. Assessment of the Winter Skate fishery in Division 4VsW. DFO Atl. Fish. Res. Doc. 98/145. (Comprend un résumé en français.)
Simon, J.E. et K.T. Frank. 2000. Assessment of the Winter Skate fishery in Division 4VsW. DFO Canadian Stock Assessment Secretariat Res. Doc. 2000/140. (Comprend un résumé en français.)
Simon, J.E., L. Harris et T. Johnston. 2003. Distribution and abundance of Winter Skate, Leucoraja ocellata, in the Canadian Atlantic. Canadian Science Advisory Secretariat Research Document 2003/028. (Le titre ainsi qu’un résumé du document sont disponibles en français : Distribution et abondance de la raie tachetée, Leucoraja ocellatta, dans l’Atlantique canadien. Secrétariat canadien de consultation scientifique – Document de recherche 2003/028.)
Smedbol, R.K., P.A. Shelton, D.P. Swain, A. Fréchet et G.A. Chouinard. 2002. Review of the population structure of cod (Gadus morhua) in Atlantic Canada in a species-at-risk context. Canadian Science Advisory Secretariat Research Document 2002/082. (Le titre ainsi qu’un résumé du document sont disponibles en français : Survol de la structure de la population, de la distribution et de l’abondance de la morue (Gadus morhua) dans le Canada atlantique dans le contexte des espèces en péril. Secrétariat canadien de consultation scientifique – Document de recherche 2002/082.)
Sosebee, K.A. 2014. United States Research Report for 2013. NAFO SCS 14/11.
Sulikowski, J.A., M.D. Morin, S.H. Suk et W.H. Howell. 2003. Age and growth estimates of the Winter Skate (Leucoraja ocellata) in the western Gulf of Maine. Fishery Bulletin 101(2): 405-413.
Swain, D.P., J.E. Simon, L.E. Harris et H.P. Benoît, 2006. Recovery potential assessment of 4T and 4VW Winter Skate (Leucoraja ocellata): biology, current status and threats. Canadian Science Advisory Secretariat Research Document 2006/003. (Le titre ainsi qu’un résumé du document sont disponibles en français : Évaluation du potentiel de rétablissement de la raie tachetée (Leucoraja ocellata) de 4T et 4VW : biologie, situation actuelle et menaces. Secrétariat canadien de consultation scientifique – Document de recherche 2006/003.)
Swain, D.P., I.D. Jonson, J.E. Simon et R.A. Myers. 2009. Assessing threats to species at risk using stage-structured state-space models: mortality trends in skate populations. Ecological Applications 19: 1347-1364.
Vladykov, V.D. 1936. Capsules d’œufs de raies de l’Atlantique canadien appartenant au genre Raja. Nature Canadien 63: 211-231.
Alan Sinclair est titulaire d’un baccalauréat ès sciences (1976) et d’une maîtrise en sciences environnementales (1986) de l’Université de Dalhousie (Dalhousie University). Il a travaillé au MPO pendant 33 ans (1976-2009) à titre de technicien, de biologiste et de chercheur scientifique. Il a été membre du Sous-comité de spécialistes des poissons marins du COSEPAC de 2007 à 2009, et est coprésident de ce sous-comité depuis 2010.
| Année | Sud du golfe du Saint-Laurent (4T) ≤ 41 cm |
Sud du golfe du Saint-Laurent (4T) 42 cm et plus |
Est du plateau néo-écossais (4VW) 36-59 cm |
Est du plateau néo-écossais (4VW) 60-74 cm |
Est du plateau néo-écossais (4VW) 75 cm en plus |
|---|---|---|---|---|---|
| 1970 | - | - | 49,950 | 0,000 | 1 033,044 |
| 1971 | 15,949 | 732,121 | 213,379 | 17,703 | 518,039 |
| 1972 | 40,674 | 383,171 | 59,314 | 35,654 | 998,250 |
| 1973 | 76,846 | 698,632 | 485,666 | 231,644 | 1 708,245 |
| 1974 | 51,512 | 488,091 | 244,716 | 60,450 | 430,600 |
| 1975 | 373,176 | 555,708 | 631,235 | 347,241 | 1 366,806 |
| 1976 | 149,671 | 725,208 | 167,333 | 126,131 | 201,987 |
| 1977 | 0,000 | 197,599 | 1 374,273 | 501,510 | 640,852 |
| 1978 | 157,922 | 880,913 | 279,893 | 222,287 | 369,682 |
| 1979 | 102,402 | 560,570 | 3 269,690 | 1 089,737 | 724,458 |
| 1980 | 57,448 | 578,534 | 580,240 | 218,429 | 658,475 |
| 1981 | 120,732 | 338,378 | 1 439,324 | 913,716 | 377,610 |
| 1982 | 202,459 | 519,555 | 577,024 | 601,329 | 904,544 |
| 1983 | 349,743 | 290,809 | 592,827 | 224,059 | 118,185 |
| 1984 | 143,115 | 206,677 | 883,223 | 146,065 | 206,269 |
| 1985 | 357,681 | 205,259 | 960,011 | 448,612 | 557,778 |
| 1986 | 208,426 | 403,622 | 703,971 | 113,864 | 310,366 |
| 1987 | 133,641 | 223,451 | 359,047 | 89,057 | 297,332 |
| 1988 | 244,629 | 231,883 | 1 205,109 | 758,336 | 1 059,285 |
| 1989 | 166,360 | 180,039 | 126,468 | 147,099 | 167,334 |
| 1990 | 81,526 | 98,330 | 340,020 | 458,418 | 623,958 |
| 1991 | 69,927 | 184,716 | 2 051,950 | 673,013 | 575,751 |
| 1992 | 77,105 | 120,508 | 1 080,753 | 552,306 | 705,145 |
| 1993 | 48,562 | 186,225 | 2 386,211 | 155,920 | 82,207 |
| 1994 | 5,000 | 33,805 | 1 433,483 | 82,464 | 94,651 |
| 1995 | 70,971 | 30,649 | 329,766 | 192,180 | 122,721 |
| 1996 | 90,132 | 45,890 | 417,847 | 533,914 | 317,381 |
| 1997 | 85,757 | 73,219 | 958,292 | 517,888 | 252,389 |
| 1998 | 40,922 | 49,508 | 563,033 | 43,718 | 90,178 |
| 1999 | 35,820 | 24,018 | 304,053 | 368,330 | 93,091 |
| 2000 | 86,646 | 56,161 | 75,382 | 515,428 | 297,565 |
| 2001 | 30,992 | 61,931 | 195,941 | 85,893 | 0,000 |
| 2002 | 14,503 | 29,257 | 380,297 | 41,986 | 0,000 |
| 2003 | 14,827 | 31,009 | 994,035 | 170,237 | 186,282 |
| 2004 | 20,221 | 10,818 | 12,111 | 68,473 | 164,001 |
| 2005 | 1,154 | 11,813 | 367,424 | 13,978 | 54,862 |
| 2006 | 23,046 | 32,486 | 89,694 | 30,723 | 67,605 |
| 2007 | 7,764 | 21,693 | 12,723 | 43,415 | 28,986 |
| 2008 | 11,866 | 34,365 | 75,028 | 102,162 | 0,000 |
| 2009 | 35,947 | 4,625 | 220,514 | 117,971 | 20,361 |
| 2010 | 5,685 | 15,219 | 69,085 | 48,010 | 65,835 |
| 2011 | 1,018 | 0,548 | 150,687 | 68,185 | 31,496 |
| 2012 | 5,145 | 11,491 | 17,982 | 0,000 | 17,982 |
| 2013 | 0,000 | 0,000 | 465,549 | 0,000 | 0,000 |