
Évaluation et rapport de situation du COSEPAC sur le Requin-taupe bleu (Isurus oxyrinchus), population de l'Atlantique au Canada, 2017
Requin-taupe bleu

- Table des matières
- COSEPAC Sommaire de l’évaluation - Population de l'Atlantique
- COSEPAC Résumé
- Résumé technique - Population de l'Atlantique
- Préface
- Description et importance de l’espèce sauvage
- Répartition
- Habitat
- Biologie
- Taille et tendances des populations
- Menaces et facteurs limitatifs
- Protection, statuts et classements
- Remerciements et experts contactés
- Sources d’information
- Sommaire biographique du ou des rédacteurs du rapport
- Figure 1. Requin-taupe bleu (Isurus oxyrinchus). Figure de la Commission internationale pour la conservation des thonidés de l’Atlantique.
- Figure 2. Répartition des sites de marquage et de remise à l’eau de requins-taupes bleus dans l’océan Atlantique.
- Figure 3. Sites de remise à l’eau (n = 142) et de recapture (n = 7) de requins-taupes marqués au moyen de méthodes traditionnelles sur deux périodes.
- Figure 4. Sites de pose d’étiquettes de collecte de données détachables (ECDD) (n = 43) sur des requins-taupes bleus et sites de remise à l’eau (n = 34) au Canada.
- Figure 5. a) Aire de répartition mondiale approximative; b) aire de répartition de l’unité désignable dans l’Atlantique Nord; c) zone d’occupation canadienne de l’unité désignable du requin-taupe bleu.
- Figure 6. Observations combinées de requins-taupes bleus en eaux canadiennes provenant des bases de données des observateurs ZIFF et SIPMAR (1998-2014).
- Figure 7. Points chauds principaux montrant les probabilités de capture de requins-taupes bleus (débarquements + rejets) par les pêches canadiennes à la palangre pélagique de 2003 à 2013 : captures multipliées par a) deux fois (4 requins/mouillage); b) cinq fois (10 requins/mouillage) et c) dix fois (20 requins/mouillage) le nombre moyen de requins par mouillage.
- Figure 8. Distribution de la fréquence des longueurs des requins-taupes bleus capturés par des pêches japonaises à la palangre en eaux canadiennes de 1986 à 1996, ainsi que par des pêches canadiennes à la palangre de 1999 à 2014, répertoriées par les programmes canadiens d’observateurs en mer.
- Figure 9. a) Indices d’abondance du requin-taupe bleu dans l’Atlantique Nord et captures totales; b) indices de taux de capture normalisés de requins-taupes bleus produits par Showell et al. (2017).
- Figure 10. Taux de capture normalisés de requins-taupes bleus des pêches à la palangre pélagique dans la Région des Maritimes (1996-2014) sur le plateau néo-écossais.
- Figure 11. Médiane du rapport actuel entre la biomasse et la biomasse fondée sur le rendement équilibré maximal (BREM) du requin-taupe bleu, avec les intervalles de confiance à 80 % de 4 exécutions du modèle bayésien de production excédentaire (ICCAT, 2012).
- Figure 12. Indices d’appauvrissement de la biomasse relative des stocks reproducteurs du requin-taupe bleu dans l’Atlantique Nord selon deux exécutions du modèle « sans capture ».
- Figure 13. Débarquements canadiens de requins-taupes bleus (t) par a) type d’engin (« autre » inclut les débarquements dans le cadre de tournois de pêche, de pêches à la ligne à main et d’autres pêches) tirés du ZIFF et du SIPMAR; b) par région de gestion.
- Figure 14. Captures estimées de requins-taupes bleus (débarquements et rejets) dans l’Atlantique Nord par des palangriers (bleu) et d’autres engins (rouges).
- Figure 15. Mortalité des requins due à la capture ou à l’hameçonnage dans les pêches commerciales canadiennes à la palangre pélagique, répartie par espèce : a) proportion de requins qui meurent après la remise à l’eau, d’après les ECDD; b) proportion des captures totales qui meurent pendant l’hameçonnage (rayé) et après la remise à l’eau (gris plein).
- Figure 16. Prises accessoires totales annuelles estimées (t) de requins-taupes bleus par espèce ciblée et par engin (FM = filet maillant; P = palangre; CFP = chalut de fond à panneaux) dans la zone économique exclusive du Canada se trouvant dans les divisions 3LNOP, de 1998 à 2010.
- Tableau 1. Débarquements canadiens (t) de requins-taupes bleus par année, par engin de pêche et par région, calculés à partir du ZIFF et du SIPMAR. Showell et al. (2017).
- Tableau 2. Productivité (r, taux intrinsèque d’augmentation des populations par année) et durée d’une génération de 20 stocks de requins et de raies pélagiques, par ordre décroissant. Les estimations de la productivité sont des médianes; les limites de confiance à 80 % sont aussi indiquées. Source : ICCAT (2012).
Évaluation et rapport de situation du COSEPAC sur le Requin-taupe bleu Isurus oxyrinchus, population de l'Atlantique au Canada, 2017

COSEPAC
Comité sur la situation
des espèces en péril
au Canada

COSEWIC
Committee on the Status
of Endangered Wildlife
in Canada
Les rapports de situation du COSEPAC sont des documents de travail servant à déterminer le statut des espèces sauvages au Canada que l’on croit en péril. On peut citer le présent document de la façon suivante :
COSEPAC. 2017. Évaluation et Rapport de situation du COSEPAC sur le requin-taupe bleu (Isurus oxyrinchus), population de l'Atlantique, au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. xiii + 40 p. (Registre public des espèces en péril site Web).
Rapport(s) précédent(s) :
COSEPAC. 2006. Évaluation et Rapport de situation du COSEPAC sur le requin-taupe bleu (Isurus oxyrinchus)au Canada. Comité sur la situation des espèces en péril au Canada. Ottawa. vii + 30 p.
Le COSEPAC remercie Scott Wallace d’avoir rédigé le rapport de situation sur le requin-taupe bleu (Isurus oxyrinchus), aux termes d’un marché conclu entre Environnement et Changement climatique Canada. La supervision du rapport a été assurée par John Neilson, coprésident du Sous-comité de spécialistes des poissons marins.
Pour obtenir des exemplaires supplémentaires, s’adresser au :
Secrétariat du COSEPAC
a/s Service canadien de la faune
Environnement Canada
Ottawa (Ontario)
K1A 0H3
Tél. : 819-938-4125
Téléc. : 819-938-3984
Courriel : COSEPAC courriel
Site Web du COSEPAC
Also available in English under the title COSEWIC Assessment and Status Report on the Shortfin Mako Isurus oxyrinchus, Atlantic population, in Canada.
Requin-taupe bleu - Commission internationale pour la conservation des thonidés de l’Atlantique (A. López [‘Tokio’]).
Requin-taupe bleu
Isurus oxyrinchus
Population de l'Atlantique
Le requin-taupe bleu (Isurus oxyrinchus) est l’une des deux espèces du genre Isurus (l’autre étant le petit requin-taupe [I. paucus]) et l’une des cinq espèces de la famille des Lamnidés (requins-taupes). Parmi les autres lamnidés présents au Canada figurent le grand requin blanc (Carcharodon carcharias), la taupe du Pacifique (Lamna ditropis) et la maraîche (L. nasus).
D’après la séparation géographique, les différences génétiques avec les autres populations mondiales et le manque de preuves d’une structuration dans l’Atlantique Nord, les requins-taupes bleus de l’Atlantique Nord sont considérés comme appartenant à la grande population de l’Atlantique.
Même si l’espèce n’est pas directement ciblée au Canada, un petit nombre d’individus sont capturés et débarqués en tant que prises accessoires par certaines pêches canadiennes. Le requin-taupe bleu est aussi très prisé par les pêcheurs sportifs états-uniens, et parfois canadiens, parce qu’il se débat énergiquement et qu’il est comestible.
Le requin-taupe bleu est répandu dans les eaux tempérées et tropicales de tous les océans, entre le 50e degré de latitude nord (60e degré de latitude nord dans l’Atlantique Nord-Est) et le 50e degré de latitude sud approximativement. On rencontre l’espèce dans l’ensemble de l’Atlantique Nord, depuis le 60e degré de latitude nord jusqu’à l’équateur. Les individus présents dans les eaux canadiennes sont de grands migrateurs qui visitent généralement les eaux chaudes du Gulf Stream l’été et l’automne et qui se trouvent à la limite septentrionale de l’aire de répartition de la grande population de l’Atlantique Nord. L’espèce a été observée dans les bancs Georges et Brown, le long du plateau continental de la Nouvelle-Écosse, dans les bancs de Terre-Neuve et même dans le golfe du Saint-Laurent.
La température semble être le principal facteur de la répartition du requin-taupe bleu. La plage de températures de l’eau privilégiée étant de 17-22 °C, il est peu probable que le requin-taupe bleu fasse des séjours prolongés en eaux canadiennes. Le manque de données a empêché la détermination des habitats nécessaires à la réalisation des fonctions essentielles (p. ex. accouplement, mise bas) de l’espèce au Canada, en plus de nuire aux études visant à établir si l’habitat avait changé au fil du temps.
Le requin-taupe bleu est aplacentaire et vivipare, et les embryons en développement se nourrissent des œufs non fécondés pendant la période de gestation de 15 à 18 mois. Les femelles donnent naissance à 11 petits en moyenne, et ce, tous les 3 ans. L’âge auquel la moitié des individus sont matures a été estimé à 8 ans chez les mâles et à 18 ans chez les femelles. L’espèce a une faible productivité, comparativement à d’autres espèces de requins, et la durée d’une génération est d’environ 25 ans. Il semble que les femelles migrent vers les 20 à 30º de latitude nord pour mettre bas puisqu’aucune femelle gravide n’a été capturée en dehors de ces latitudes.
L’espèce est vraisemblablement capable de s’adapter aux changements naturels de son environnement, les adultes pouvant rapidement parcourir de longues distances et se nourrir d’un vaste éventail d’espèces. D’après des études sur l’alimentation des adultes, les requins-taupes bleus ont une grande variété de proies, principalement des poissons, dont le tassergal, le stromaté, le thon, le maquereau, la bonite et l’espadon.
Pour les eaux canadiennes, une série de taux de capture provenant des pêches à la palangre pélagique pour la période de 1996 à 2014 est le seul indice d’abondance disponible. Les données les plus récentes montrent un déclin des taux de capture par rapport aux données du début de la série chronologique, déclin toutefois non significatif. Les eaux canadiennes représentent la limite septentrionale de l’aire de répartition de l’espèce, et, par conséquent, les variations de l’indice canadien peuvent refléter des changements de la répartition.
À l’échelle internationale, six indices de taux de capture sont considérés par la Commission internationale pour la conservation des thonidés de l’Atlantique (CICTA) comme étant les plus représentatifs de l’abondance. En les interprétant ensemble, on conclut que la plupart de ces indices montrent un déclin pendant les années 1990, suivi d’une hausse après 2000.
Une évaluation des stocks menée par la CICTA en 2012 a permis de conclure que les indices d’abondance actualisés montraient des tendances à la hausse ou stables. En dépit des incertitudes, la Commission a conclu que la probabilité de surpêche était faible et que le statut de la population est au-dessus du rendement équilibré maximal.
Les prises accessoires dans les pêches commerciales à la palangre ciblant les thons et les espadons pélagiques sont la principale cause de mortalité en eaux canadiennes et dans l’ensemble de l’aire de répartition du requin-taupe bleu. Le taux de survie après remise à l’eau des requins-taupes capturés à la palangre au Canada a récemment été estimé à 49 %, ce qui donne un taux de mortalité total moyen annuel historique d’environ 69 t/an. La remise à l’eau volontaire en 2015 de requins-taupes bleus vivants dans la Région des Maritimes, appuyée par l’industrie de la pêche à la palangre, devrait réduire la mortalité dans les eaux canadiennes. Les femelles matures forment moins de 1 % des individus capturés par les pêches à la palangre pélagique dans la région canadienne des Maritimes; la couverture par les observateurs en mer représente en moyenne quelque 5 % de l’effort de pêche annuel.
On ne connaît pas bien la mortalité totale due aux pêches dans tout l’Atlantique Nord à cause de la mauvaise comptabilisation des prises, en particulier pendant les années précédant 1996. De 1996 à 2010, les débarquements moyens déclarés s’élevaient à environ 2 400 t, mais ce nombre a été considéré comme une sous-estimation dans une récente étude de la CICTA, qui a estimé que les captures (débarquements et rejets) pendant cette période étaient en moyenne de 3 500 t.
En raison des caractéristiques de leur cycle vital, par exemple la croissance relativement lente, l’âge tardif de la maturité et les faibles taux de reproduction, les populations de requins-taupes bleus présentent une productivité relativement faible; c’est pourquoi leur capacité de se rétablir est limitée une fois qu’elles sont appauvries.
Aucune pêche ne cible le requin-taupe bleu dans l’Atlantique canadien, et le Canada interdit l’ablation des nageoires de requins (où les nageoires sont retirées et conservées alors que le corps du requin est rejeté en mer). Les individus accidentellement capturés peuvent être conservés dans le cadre de certaines pêches. Des règlements de pêche et des mesures de protection de la Région des Maritimes prévoient l’adoption d’une limite annuelle non restrictive de débarquements de requins-taupes bleus de 100 t, l’utilisation d’hameçons circulaires corrodables pour réduire les prises accessoires et la mortalité après remise à l’eau dans les pêches à la palangre pélagique ainsi que la remise à l’eau volontaire des requins-taupes bleus vivants. Les règlements actuels ne limitent pas la mortalité totale par pêche ou les rejets en mer.
Lors de sa dernière évaluation du requin-taupe bleu (sous-population de l’Atlantique Nord) en 2004, l’Union internationale pour la conservation de la nature (UICN) a désigné l’espèce « vulnérable ». La National Oceanographic and Atmospheric Administration (NOAA) des États-Unis décrit le stock comme étant sain puisqu’il n’est pas surexploité et que la probabilité de surpêche est faible. Le COSEPAC a précédemment désigné la population de l’Atlantique du requin-taupe bleu espèce menacée en 2006, mais la décision d’inscrire l’espèce à l’annexe 1 de la Loi sur les espèces en péril doit encore être prise.
| Sujet | Information |
|---|---|
| Durée d’une génération d’après G = âge à maturité/mortalité naturelle | 25 ans |
| Y a-t-il un déclin continu observé du nombre d’individus matures? | Non |
| Pourcentage estimé de déclin continu du nombre total d’individus matures sur [cinq ans ou deux générations]. | Déclin continu non observé |
Pourcentage [observé, estimé, inféré ou présumé] de [réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix dernières années ou trois dernières générations].
|
Aucune tendance pour l’UD entière selon les indices |
| Pourcentage [prévu ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours des [dix prochaines années ou trois prochaines générations]. | Inconnu |
| Pourcentage [observé, estimé, inféré ou présumé] [de réduction ou d’augmentation] du nombre total d’individus matures au cours de toute période de [dix ans ou trois générations] commençant dans le passé et se terminant dans le futur. | Inconnu |
| Est-ce que les causes du déclin sont clairement a) réversibles et b) comprises et c) ont effectivement cessé? | Aucun déclin évident a) s.o. b) s.o. c) s.o. |
| Y a-t-il des fluctuations extrêmes du nombre d’individus matures? | Non, il s’agit d’une espèce à faible productivité |
| Sujet | Information |
|---|---|
| Superficie estimée de la zone d’occurrence | 1 060 000 km2 |
| Indice de zone d’occupation (IZO) (Fournissez toujours une valeur établie à partir d’une grille à carrés de 2 km de côté). |
>> 2 000 km2 |
| La population est-elle gravement fragmentée, c’est-à-dire que plus de 50 % de sa zone d’occupation totale se trouvent dans des parcelles d’habitat qui sont a) plus petites que la superficie nécessaire au maintien d’une population viable et b) séparées d’autres parcelles d’habitat par une distance supérieure à la distance de dispersion maximale présumée pour l’espèce? | a) Non b) Non |
| nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme. (utilisez une fourchette plausible pour refléter l’incertitude, le cas échéant) |
> 10. Vaste aire de répartition, capture dans le cadre de plusieurs pêches internationales dans l’ensemble de l’Atlantique Nord |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de la zone d’occurrence? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de l’indice de zone d’occupation? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de sous-populations? | Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
Non |
| Y a-t-il un déclin continu [observé, inféré ou prévu] de [la superficie, l’étendue et/ou la qualité] de l’habitat? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de sous-populations? | Non |
| Y a-t-il des fluctuations extrêmes du nombre de localités? Voir « Définitions et abréviations » sur le Site Web du site web du COSEPAC et IUCN (février 2014; en anglais seulement) pour obtenir des précisions sur ce terme.? |
Non |
| Y a-t-il des fluctuations extrêmes de la zone d’occurrence? | Non |
| Y a-t-il des fluctuations extrêmes de l’indice de zone d’occupation? | Non |
| Sous-populations (utilisez une fourchette plausible) | Nombre d’individus matures |
|---|---|
| - | Inconnu |
| Total | - |
| Sujet | Information |
|---|---|
| La probabilité de disparition de l’espèce à l’état sauvage est d’au moins [20 % sur 20 ans ou 5 générations, ou 10 % sur 100 ans] | Aucune analyse disponible |
| Sujet | Information |
|---|---|
| Un calculateur des menaces a-t-il été rempli pour l’espèce? | Non Quels autres facteurs limitatifs sont pertinents? |
| Sujet | Information |
|---|---|
| Situation des populations de l’extérieur les plus susceptibles de fournir des individus immigrants au Canada | Une seule population, dont font partie les individus du Canada; la situation de la population est considérée comme étant au-dessus du point de référence de gestion. |
| Une immigration a-t-elle été constatée ou est-elle possible? | Possible, preuves génétiques chez des mâles |
| Des individus immigrants seraient-ils adaptés pour survivre au Canada? | Oui |
| Y a-t-il suffisamment d’habitat disponible au Canada pour les individus immigrants? | Oui |
| Les conditions se détériorent-elles au Canada Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe)? |
Inconnu |
| Les conditions de la population source se détériorent-elles Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe)? |
Inconnu |
| La population canadienne est-elle considérée comme un puits Voir le tableau 3 (Lignes directrices pour la modification de l’évaluation de la situation d’après une immigration de source externe)? |
Non |
| La possibilité d’une immigration depuis des populations externes existe-t-elle? | Possible |
| Sujet | Information |
|---|---|
| L’information concernant l’espèce est-elle de nature délicate? | Non |
| Sujet | Information |
|---|---|
| Espèce désignée « menacée » en avril 2006. | Réexamen du statut : l’espèce a été désignée « préoccupante » en avril 2017. |
| Sujet | Information |
|---|---|
| Statut | Espèce préoccupante |
| Code alphanumérique | Sans objet |
| Justification de la désignation | Cette espèce est plus vulnérable que de nombreuses autres espèces de requins de l’Atlantique en raison de sa longue durée de vie et de sa faible productivité. L’espèce compte une seule population fortement migratrice dans l’Atlantique Nord, dont une partie de cette population est présente dans les eaux canadiennes de façon saisonnière. Les taux de prise totaux dans l’Atlantique Nord ainsi que l’abondance sont stables. La plupart des analyses réalisées dans le cadre d’une récente évaluation des stocks ont montré une tendance stable ou à la hausse depuis 1971, résultant en ce qui a amené une amélioration du statut comparativement au précédent rapport de situation du COSEPAC. Toutefois, le statut de l’espèce n’est pas sécurisé étant donné les incertitudes entourant l’évaluation des stocks, la courte période d’augmentation des taux de prise relativement à la durée de génération de 25 ans et la faible productivité de l’espèce. |
| Sujet | Information |
|---|---|
| Critère A (déclin du nombre total d’individus matures) | Ne correspond pas aux critères. La cause du déclin antérieur a cessé, est comprise et est réversible. La réduction du nombre total d’individus matures est inférieure à 50 %. |
| Critère B (petite aire de répartition, et déclin ou fluctuation) | Ne correspond pas aux critères, car la zone d’occurrence et l’IZO dépassent les seuils. |
| Critère C (nombre d’individus matures peu élevé et en déclin) | Ne correspond pas aux critères, car la taille de la population dépasse les seuils. |
| Critère D (très petite population ou répartition restreinte) | Ne correspond pas aux critères. |
| Critère E (analyse quantitative) | Analyse non réalisée. |
| cellule vide | cellule vide |
| Sujet | Information |
|---|---|
| b. « l’espèce sauvage peut devenir ‘‘menacée’’ si les facteurs dont on craint l’influence négative sur sa longévité ne sont ni renversés, ni gérés de façon efficace ». | - |
Plusieurs éléments d’information se sont ajoutés depuis la rédaction du rapport de situation du COSEPAC sur le requin-taupe de 2006, notamment :
- des études confirmant la structure génétique de la population;
- toute l’information provenant des activités de marquage dans l’Atlantique Nord;
- les données issues des étiquettes de collecte de données détachables (ECDD) posées sur les requins-taupes canadiens capturés;
- l’information sur la survie après remise à l’eau provenant des pêches canadiennes à la palangre pélagique;
- l’évaluation de 2012 de la CICTA, qui comprenait une analyse des meilleurs indices des tendances, estimations des captures non déclarées et estimations de la productivité disponibles, ainsi qu’une évaluation des stocks;
- l’âge des femelles à maturité, qui a été substantiellement révisé d’après de nouvelles études de validation de l’âge et qui est passé de 7-8 ans à 18 ans (selon de nouvelles données), ce qui laisse entrevoir une résilience plus faible qu’on ne le croyait;
- de meilleures estimations de la mortalité par pêche au Canada.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) a été créé en 1977, à la suite d’une recommandation faite en 1976 lors de la Conférence fédérale-provinciale sur la faune. Le Comité a été créé pour satisfaire au besoin d’une classification nationale des espèces sauvages en péril qui soit unique et officielle et qui repose sur un fondement scientifique solide. En 1978, le COSEPAC (alors appelé Comité sur le statut des espèces menacées de disparition au Canada) désignait ses premières espèces et produisait sa première liste des espèces en péril au Canada. En vertu de la Loi sur les espèces en péril (LEP) promulguée le 5 juin 2003, le COSEPAC est un comité consultatif qui doit faire en sorte que les espèces continuent d’être évaluées selon un processus scientifique rigoureux et indépendant.
Le Comité sur la situation des espèces en péril au Canada (COSEPAC) évalue la situation, au niveau national, des espèces, des sous-espèces, des variétés ou d’autres unités désignables qui sont considérées comme étant en péril au Canada. Les désignations peuvent être attribuées aux espèces indigènes comprises dans les groupes taxinomiques suivants : mammifères, oiseaux, reptiles, amphibiens, poissons, arthropodes, mollusques, plantes vasculaires, mousses et lichens.
Le COSEPAC est composé de membres de chacun des organismes responsables des espèces sauvages des gouvernements provinciaux et territoriaux, de quatre organismes fédéraux (le Service canadien de la faune, l’Agence Parcs Canada, le ministère des Pêches et des Océans et le Partenariat fédéral d’information sur la biodiversité, lequel est présidé par le Musée canadien de la nature), de trois membres scientifiques non gouvernementaux et des coprésidents des sous-comités de spécialistes des espèces et du sous-comité des connaissances traditionnelles autochtones. Le Comité se réunit au moins une fois par année pour étudier les rapports de situation des espèces candidates.
Remarque : Catégorie « DSIDD » (données insuffisantes pour donner une désignation) jusqu’en 1994, puis « indéterminé » de 1994 à 1999. Définition de la catégorie (DI) révisée en 2006.
Le Service canadien de la faune d’Environnement et Changement climatique Canada assure un appui administratif et financier complet au Secrétariat du COSEPAC.
Le requin-taupe (Isurus oxyrinchus) est l’une des deux espèces du genre Isurus (l’autre étant le petit requin-taupe [I. paucus]) et l’une des cinq espèces de la famille des Lamnidés (requins-taupes). Les autres lamnidés qui vivent au Canada sont le grand requin blanc (Carcharodon carcharias), la taupe du Pacifique (Lamna ditropis; aussi appelé requin-taupe saumon) et la maraîche (Lamna nasus; aussi appelé requin-taupe commun). Aucune sous-population de requins-taupes n’est reconnue.
Le requin-taupe bleu se distingue par son museau pointu, ses yeux relativement petits, ses longues dents lisses ressemblant à des dagues et dépourvues de cuspide latérale (sur les deux mâchoires) et sa bouche en forme de U (figure 1). Les dents antérieures du maxillaire inférieur font saillie sur un plan horizontal, même lorsque la bouche est fermée. Les nageoires pectorales sont légèrement incurvées, et le bout se termine en pointe; le bord antérieur, qui fait de 16 à 22 % de la longueur totale de l’animal, est plus court que la longueur de la tête. La première nageoire dorsale prend naissance vis-à-vis de la pointe arrière libre des nageoires pectorales ou juste derrière. Le bout est largement arrondi chez les jeunes, mais il est plus anguleux et étroitement arrondi chez les juvéniles de grande taille et les adultes. La première nageoire dorsale est plus longue que la largeur de sa base chez les gros individus, mais elle est de longueur égale ou inférieure chez les jeunes de moins de 185 cm. La nageoire caudale en forme de croissant est pourvue d’une carène principale horizontale, mais pas de carène secondaire. La coloration dorsolatérale est bleu vif ou violacée, et le dessous du museau est blanc chez les jeunes et les adultes. La tête, de couleur foncée, couvre en partie les septa branchiaux. La couleur foncée des flancs ne s’étend pas jusqu’à l’abdomen. Les nageoires pelviennes sont de couleur foncée sur la moitié antérieure et blanches sur la moitié postérieure, et le dessous est blanc également.
Des erreurs d’identification se sont produites en eaux chaudes, où les aires de répartition des deux espèces de requins-taupes se chevauchent. Cependant, dans les eaux canadiennes, où le petit requin-taupe est extrêmement rare, il est peu probable que les deux espèces soient régulièrement confondues. Dans l’Atlantique canadien, le requin-taupe bleu a déjà été confondu avec la maraîche, ce qui a peut-être contribué à la sous-estimation des débarquements de requins-taupes bleus avant 1996 (Campana et al., 2004).

Description longue de la figure 1
Illustration du requin-taupe bleu (Isurus oxyrinchus). Le requin-taupe bleu se distingue par son museau pointu et ses yeux relativement petits. Ses nageoires pectorales sont légèrement incurvées, et le bout se termine en pointe; le bord antérieur fait de 16 à 22 pour cent de la longueur totale de l’animal. La première nageoire dorsale prend naissance à la hauteur de l’extrémité arrière libre des nageoires pectorales ou juste derrière. La nageoire caudale en forme de croissant est pourvue d’une carène principale horizontale, mais pas de carène secondaire. La coloration dorsolatérale du requin taupe bleu est bleu vif ou violacée, et le dessous du museau est blanc. La tête, de couleur foncée, couvre en partie les septa branchiaux.
Les connaissances actuelles sur la structure spatiale de la population de requins-taupes bleus à l’échelle mondiale et, plus précisément, dans l’Atlantique Nord, ont été acquises grâce à plus de 50 années d’activités de marquage traditionnelles, à la pose récente d’étiquettes satellites et aux études génétiques menées ces 2 dernières décennies (ICCAT, 2012; Campana et al., 2015).
En 2012, la Commission internationale pour la conservation des thonidés de l’Atlantique (CICTA) a colligé toutes les données obtenues à partir des activités de marquage traditionnelles (micromarques) dans l’Atlantique Nord. Depuis 1962, un total de 9 218 micromarques ont été posées, et 1 203 requins marqués ont été recapturés (figure 2). La plupart de ces micromarques ont été déployées au large de la côte nord-est des États-Unis. Bien que l’on ait observé des requins-taupes bleus parcourir de longues distances (jusqu’à 3 400 km) dans l’Atlantique, la plupart des déplacements se faisaient entre le sud et l’est de l’Atlantique Nord-Ouest, et très peu de captures ont été réalisées au sud du 20e parallèle et aucune au sud du 5e parallèle.
Le marquage de requins au moyen d’étiquettes classiques a été pratiqué en eaux canadiennes de 1961 à 1986 (n = 110) et plus récemment de 2006 à 2015 (n = 32), et 5 et 2 captures ont été réalisées lors de ces deux périodes, respectivement (figure 3; Showell et al., 2017). De 2010 à 2014, le Canadian Shark Laboratory a déployé 43 étiquettes satellites sur des requins-taupes bleus sains et blessés, principalement sur le plateau néo-écossais, parmi lesquelles 34 ont été récupérées ou ont transmis des données (Campana et al., 2015; Showell et al., 2017; figure 4). Les données tirées des micromarques traditionnelles et des étiquettes satellites posées au Canada l’été laissent croire à des déplacements généraux vers le sud, vers l’est ou vers le sud-est ainsi que des déplacements sur de longues distances vers l’est en direction du centre de l’Atlantique ou vers le sud en direction des Caraïbes.
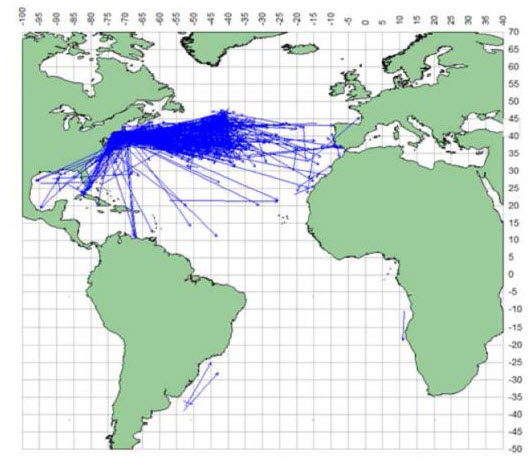
Description longue de la figure 2
Carte illustrant la répartition des sites de marquage et de remise à l’eau de requins taupes bleus dans l’océan Atlantique. Les lignes droites relient les sites de remise à l’eau aux sites de recapture.
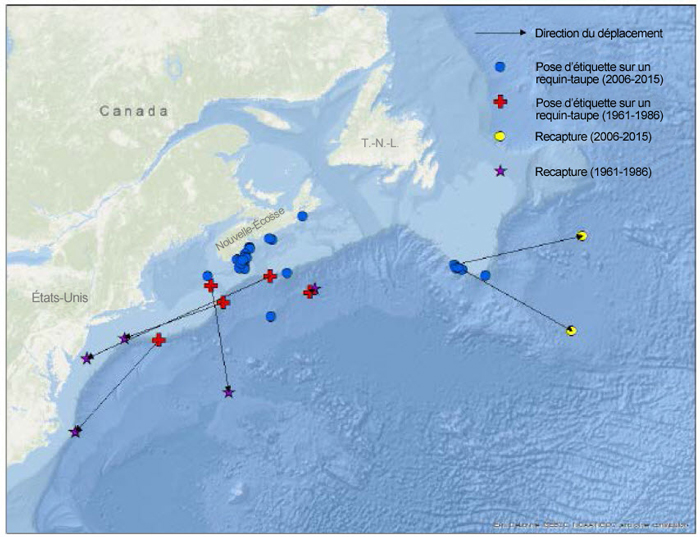
Description longue de la figure 3
Carte illustrant les sites de remise à l’eau et de recapture de requins-taupes bleus marqués au moyen de méthodes traditionnelles sur deux périodes (de 2006 à 2015, et de 1961 à 1986).
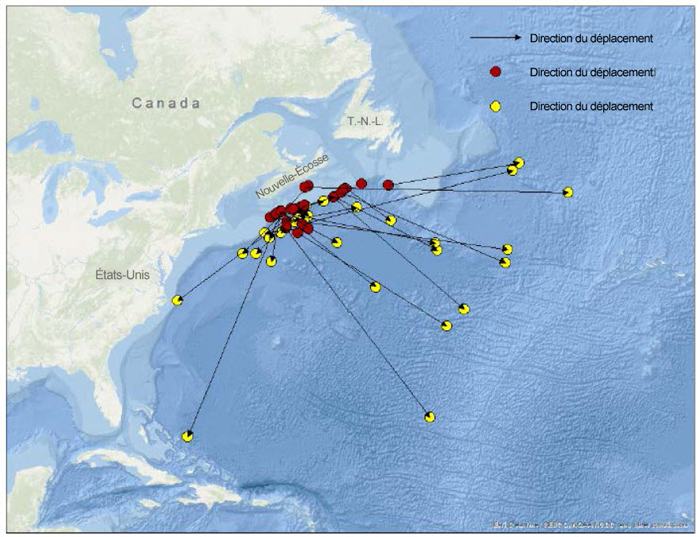
Description longue de la figure 4
Carte illustrant les sites de pose d’étiquettes de collecte de données détachables (ECDD) sur des requins-taupes bleus et les sites de remise à l’eau au Canada.
Ensemble, les études de marquage au Canada et aux États-Unis indiquent que les individus marqués sont de grands migrateurs qui fréquentent principalement les eaux à l’ouest du 40e degré de longitude et au nord du 30e degré de latitude nord.
Des données génétiques, principalement tirées d’analyses de l’ADN mitochondrial (ADNmt), appuient l’hypothèse d’une population de l’Atlantique Nord distincte et d’une grande séparation spatiale de celle-ci par rapport aux autres populations. Heist et al. (1996) ont utilisé l’ADNmt pour analyser la structure des populations de requins-taupes bleus de l’Atlantique et du Pacifique. Ils ont constaté que la population de l’Atlantique Nord différait considérablement des populations de l’Atlantique Sud et du Pacifique Nord et Sud (Fst global = 0,15), et ont ainsi conclu que la population de l’Atlantique Nord recevait un flux génique très limité à partir des populations d’autres zones et, par conséquent, qu’il était justifié qu’elle fasse l’objet de considérations de gestion distinctes. Schrey et Heist (2003) ont étudié l’ADN microsatellite (nucléaire) du requin-taupe bleu à 4 locus en utilisant 433 échantillons provenant de l’Atlantique Nord, de l’Atlantique Sud, du Pacifique Nord, du Pacifique Sud et des côtes sud-africaines de l’Atlantique et du Pacifique. Cette étude a révélé de très faibles degrés de différenciation, et ce, même entre les principaux bassins océaniques (FST global < 0,003), et les résultats permettent difficilement d’infirmer l’hypothèse selon laquelle les requins-taupes bleus forment une seule et même population à l’échelle mondiale. Selon un modèle de mutation, la valeur de P était légèrement inférieure à 0,05, alors que, selon un autre modèle de mutation, elle était légèrement supérieure à 0,05. L’analyse de puissance indiquait une très grande capacité de détecter la structure de la population au niveau indiqué par l’étude de l’ADNmt. Selon Schrey et Heist (2003), les deux jeux de données pourraient s’expliquer par le fait que les femelles affichent un haut degré de philopatrie (d’où les grandes différences observées dans l’ADNmt, hérité de la mère) et que les mâles ont davantage tendance à se disperser (d’où la faible différenciation observée dans les marqueurs de l’ADN nucléaire). Grâce à des techniques d’ADNmt semblables (n = 106), Tagchi et al. (2011) ont confirmé de nouveau la séparation génétique des populations de requins-taupes bleus de l’Atlantique Nord et du Pacifique.
Les eaux canadiennes se trouvent à la limite de l’aire de répartition de l’espèce dans l’Atlantique Nord (voir la section Répartition ci-dessous). D’après la séparation biogéographique, les différences génétiques par rapport aux autres populations du monde et l’absence de preuves d’une structuration au sein de l’Atlantique Nord, les requins-taupes bleus de l’Atlantique Nord sont considérés comme formant une population, et la seule UD présente au Canada fait partie d’une population de l’Atlantique Nord plus étendue. Aucune information sur le pourcentage de la population de l’Atlantique Nord qui vit au Canada n’est disponible.
Même s’il n’est pas directement ciblé au Canada, le requin-taupe bleu est une espèce capturée et vendue en raison de la qualité de sa chair. Ce poisson est aussi très prisé par les pêcheurs sportifs, surtout aux États-Unis, parce qu’il se débat énergiquement et qu’il est comestible. À l’échelle de l’aire de répartition, y compris en eaux canadiennes, la chair est utilisée à l’état frais, congelé, fumé et séché salé aux fins de consommation humaine; l’huile est extraite pour les vitamines; les nageoires sont servies dans des soupes; la peau est transformée en cuir; les mâchoires et les dents servent d’ornements. Le requin-taupe bleu est un prédateur opportuniste situé au sommet de la chaîne trophique, et il a de nombreuses proies. Par conséquent, il joue probablement un rôle important dans la structure des écosystèmes marins.
Le requin-taupe bleu se rencontre dans toutes les mers tempérées et tropicales de la planète. La population de requins-taupes de l’Atlantique Nord fréquente toutes les eaux comprises entre le 60e degré de latitude nord et l’équateur (figure 5a) (Showell et al., 2017).
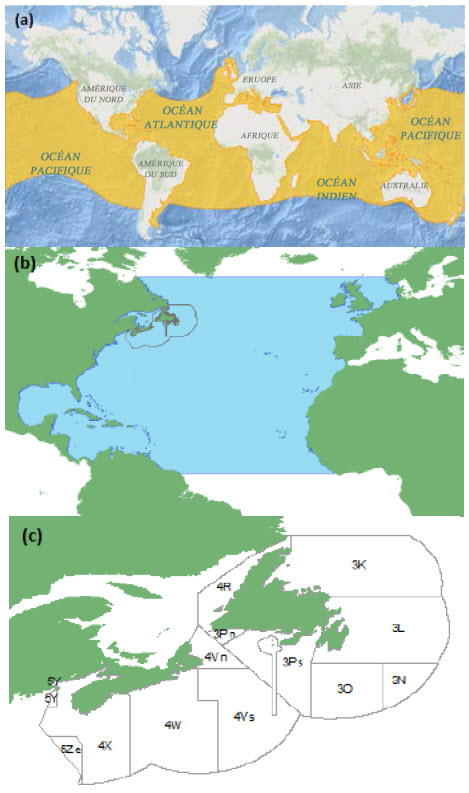
Description longue de la figure 5
Cartes indiquant (a) l’aire de répartition mondiale approximative du requin taupe bleu (toutes les eaux au sud du 60e parallèle et au nord de l’équateur); (b) l’aire de répartition de l’unité désignable dans l’Atlantique Nord; (c) la zone d’occupation canadienne de l’unité désignable du requin-taupe bleu.
L’aire de répartition canadienne a été estimée à partir de la répartition de l’ensemble des observations connues recueillies par les pêches commerciales (ZIFF [fichier informatisé sur les échanges entre les zones] et SIPMAR [Système d’information sur les pêches des Maritimes]), des données canadiennes d’observateurs en mer (programmes des observateurs en mer des Régions des Maritimes et de Terre-Neuve-et-Labrador), des relevés scientifiques et des données de marquage qui chevauchent une division de pêche de l’OPANO (figure 6; Showell et al., 2017). Dans les eaux canadiennes, le requin-taupe bleu est généralement associé à des eaux chaudes telles que celles au sein ou à proximité du Gulf Stream. L’espèce a été repérée dans les bancs Georges et Browns, le long du plateau continental de la Nouvelle-Écosse, dans les bancs de Terre-Neuve et même dans le golfe du Saint-Laurent (Showell et al., 2017). Les données fournies par les observateurs à bord de bateaux canadiens, féroïens et japonais révèlent que les requins-taupes bleus sont capturés dans les eaux tant côtières qu’extracôtières entre la partie sud de la zone économique exclusive (ZEE) du Canada et le 50e degré de latitude nord. Cette espèce est un grand migrateur qui visite les côtes canadiennes de l’Atlantique de façon saisonnière (à la fin de l’été et à l’automne). Les requins-taupes des eaux canadiennes représentent une extension nord-ouest de la population de l’Atlantique Nord et, bien que l’on ne dispose d’aucune information sur le pourcentage d’individus résidant au Canada, ils ne représentent probablement qu’une petite partie cette grande population. Il n’existe aucune donnée témoignant d’une expansion ou d’une contraction de l’aire de répartition du requin-taupe bleu dans les eaux canadiennes de l’Atlantique.
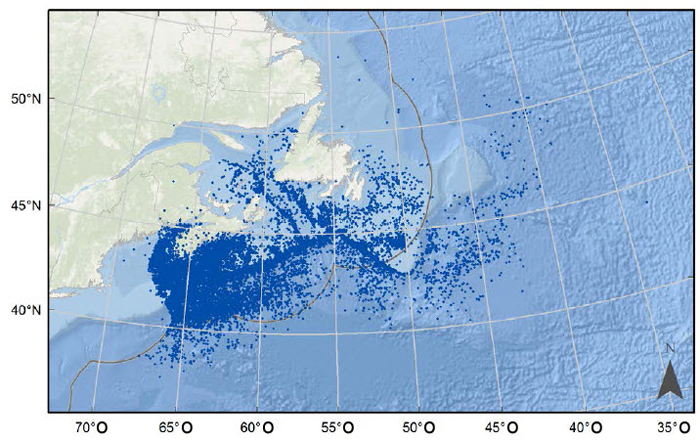
Description longue de la figure 6
Carte illustrant les observations combinées de requins-taupes bleus en eaux canadiennes provenant des bases de données des observateurs ZIFF (Zonal Interchange File Format) et SIPMAR (Système d’information sur les pêches des maritimes) pour la période allant de 1998 à 2014.
La zone d’occurrence dans la portion canadienne de cette UD a été calculée en tant que la somme de la partie des divisions ou sous-divisions 3KL+3NOP+4R+ 4VWX+5Y+5Ze de l’OPANO se trouvant dans la ZEE du Canada (1,06 million de km2) (figure 5b; Showell et al., 2017). La zone d’occupation actuelle, représentée par des observations fréquentes ou des captures, mesure environ 800 000 km2 (Campana et al., 2004a).
Bien que plusieurs étiquettes satellites aient été déployées sur des requins-taupes bleus dans les eaux canadiennes depuis le rapport de situation du COSEPAC de 2006, les nouvelles données n’ont pas modifié notre compréhension des besoins en matière d’habitat dans l’Atlantique Nord (Campana et al., 2015). La température semble être le principal facteur de la répartition du requin-taupe bleu. Le requin-taupe bleu préfère les eaux de 17 à 22 °C, de sorte que, dans l’océan Atlantique, il est souvent associé au Gulf Stream (Compagno, 2001), lequel se trouve surtout en dehors des eaux canadiennes. Des enregistreurs de température et de profondeur sur des émetteurs satellites indiquent que le requin-taupe bleu fréquente des eaux de 10,4 à 28,6 °C et la couche d’eau entre la surface et 556 m (Loefer et al., 2005).
En général, le requin-taupe bleu se rencontre en zone extracôtière, sur le rebord du plateau continental, sur le plateau continental et parfois près des côtes. Dans la partie ouest de l’Atlantique Nord, le requin-taupe bleu se déplace vers le plateau continental lorsque les températures dépassent les 17 °C à la surface de l’eau, habituellement de juin à décembre.
Le manque de données empêche la détermination des habitats nécessaires à la réalisation des fonctions essentielles du cycle vital (p. ex. accouplement, mise bas) de l’espèce en eaux canadiennes. Godin et al. (données inédites, 2015) ont repéré des zones à hauts taux de capture (débarquements + rejets) (figure 7), mais aucun élément de l’habitat n’a été associé à ces zones.
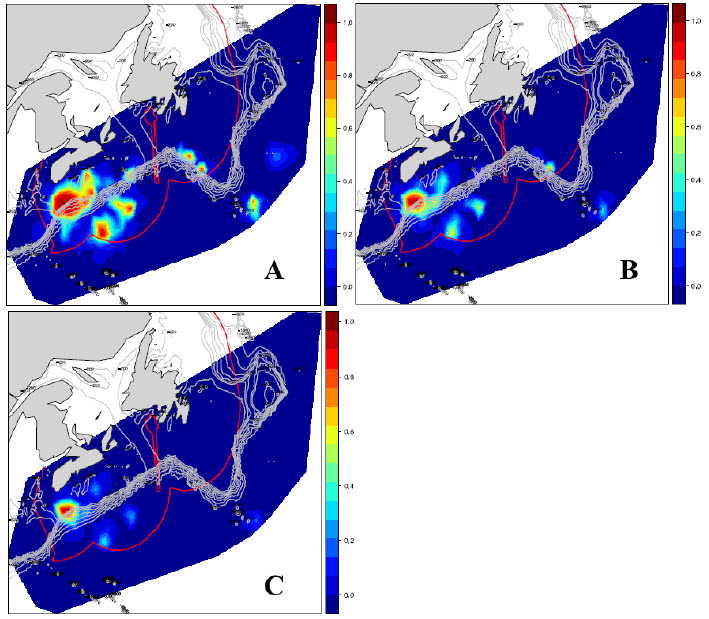
Description longue de la figure 7
Carte à trois volets illustrant les points chauds principaux pour la capture de requins taupes bleus (débarquements + rejets) par les pêches canadiennes à la palangre pélagique de 2003 à 2013. Les volets montrent les captures multipliées par deux fois (volet a), par cinq fois (volet b) et par dix fois (volet c) le nombre moyen de requins par mouillage.
Le manque de données a nui aux études visant à déterminer si l’habitat du requin-taupe bleu a changé au fil du temps dans l’UD de l’Atlantique Nord ou dans les eaux canadiennes. Bien que l’Atlantique Nord ait subi des anomalies de température positives ces dernières décennies, aucune recherche sur le moment des migrations vers les zones extracôtières et côtières et les tendances en matière de répartition n’a été réalisée.
Le requin-taupe bleu est aplacentaire et vivipare, ses embryons en développement se nourrissant des œufs non fécondés pendant la période de gestation, laquelle varie à l’échelle planétaire. Dans l’Atlantique Nord, elle varie de 15 à 18 mois environ, et la taille des portées est de 11 petits en moyenne, et ce, tous les 3 ans (Campana et al., 2004a). La période de mise bas s’étend de la fin de l’hiver au milieu de l’été, et les petits mesurent environ 70 cm de longueur (Mollet al., 2000). Après la parturition, les femelles se reposeraient pendant une période de 18 mois, ce qui donne un cycle de reproduction présumé de 3 ans (Mollet et al., 2000; ICCAT, 2012).
La validation de l’âge et de la croissance du requin-taupe a donné des résultats variés, selon la technique utilisée et le bassin océanique étudié. Dans l’Atlantique Nord, d’après la validation par la méthode de datation au radiocarbone, l’âge auquel la moitié des individus sont matures est estimé à 8 ans chez les mâles (185 cm de longueur à la fourche [LF]) et à 18 ans chez les femelles (275 cm LF) (Natanson et al., 2006). Selon Campana et al. (2004a), qui ont obtenu des estimations semblables de l’âge à la maturité, les femelles restent immatures jusqu’à 18 ans (272 cm LF).
D’après une couverture annuelle d’environ 5 % par les observateurs en mer, les données sur la fréquence des longueurs rapportées par les observateurs des pêches à la palangre pélagique de la Région des Maritimes donnent à penser que les individus matures sont rares dans les eaux canadiennes de l’Atlantique ou qu’ils ne sont pas capturés par les engins de pêche commerciale, ou encore qu’ils sont vraisemblablement en mesure de briser la ligne grâce à leur taille quand ils sont capturés (DFO, 2016). Le pourcentage de mâles matures (longueur totale [LT] > 185 cm) et de femelles matures (LT > 275 cm) rapporté par les observateurs de la flottille de pêche commerciale entre 2006 et 2015 est estimé respectivement à 7 (n = 1 114) et à moins de 1 % (n = 1 025). La composition selon la taille observée dans les captures des pêches japonaises à la palangre pélagique pratiquées en eaux canadiennes de 1986 à 1996 a augmenté au cours de cette période, tandis que celle des pêches canadiennes à la palangre pélagique pratiquées de 1999 à 2014 a varié, sans qu’une tendance ressorte (figure 8).
Selon Bishop et al. (2006), la mortalité naturelle (M) du requin-taupe bleu de l’Atlantique Nord serait de 0,10 à 0,15. Smith et al. (1998) a, quant à lui, établi la mortalité naturelle à 0,16.
La durée d’une génération a été calculée au moyen de la formule suivante : 1/mortalité des adultes + âge où 50 % des femelles sont en âge de se reproduire. La durée d’une génération est donc d’environ 25 ans [18+(1/0,15)].
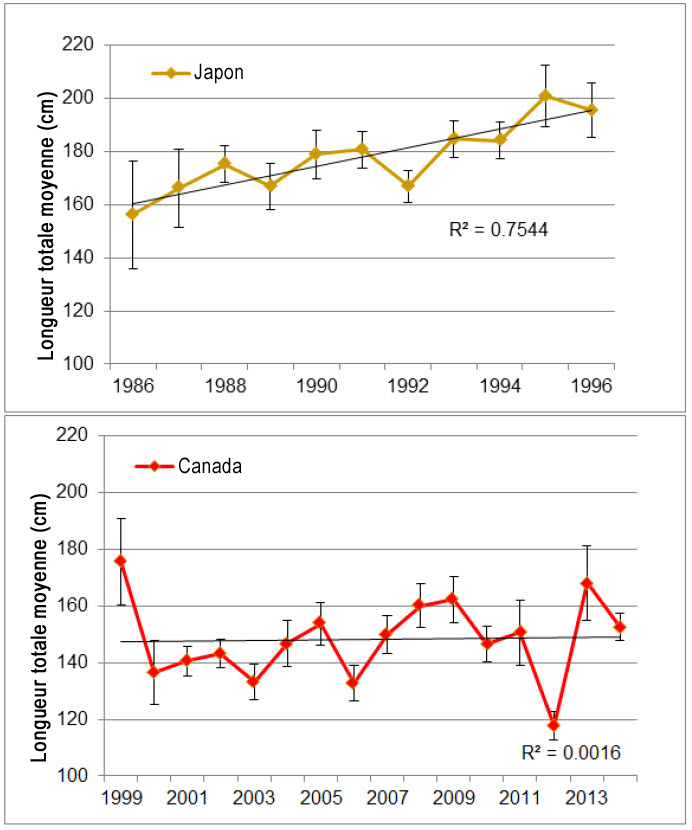
Description longue de la figure 8
Graphiques illustrant la distribution de la fréquence des longueurs des requins-taupes bleus capturés par des pêches japonaises à la palangre en eaux canadiennes de 1986 à 1996 (graphique du haut), ainsi que par des pêches canadiennes à la palangre de 1999 à 2014 (graphique du bas), répertoriées par les programmes canadiens d’observateurs en mer.
Les requins-taupes bleus adultes ont probablement des adaptations leur permettant de supporter l’étendue actuelle des changements climatiques, les changements du type de proie et les hausses de la température de l’eau puisqu’ils peuvent se déplacer sur de longues distances et se nourrir d’une grande variété d’espèces. De plus, ils occupent un vaste territoire, ce qui les rend moins vulnérables aux phénomènes stochastiques localisés. La physiologie de l’espèce lui permet de demeurer très active en eaux froides (Carey et al., 1981).
Le requin-taupe bleu est un grand migrateur; des déplacements sur une distance allant jusqu’à 3 400 km ont été observés (Casey et Kohler, 1992). Les études de marquage classiques montrent que la plupart des recaptures se font à moins de 500 km du site de marquage (Kohler et al., 1998; ICCAT, 2012). Compte tenu de sa préférence pour les eaux chaudes et de sa grande mobilité, il est peu probable que l’espèce passe de longues périodes dans les eaux canadiennes, sauf en été et au début de l’automne. Les requins-taupes bleus marqués et recapturés (Nmarqués =9 218; Nrecapturés =1 203) de 1962 à 2012 dans l’Atlantique Nord-Ouest ont révélé une vaste gamme de profils de déplacement, mais surtout des déplacements vers le sud, le sud-est et l’est à partir du site de marquage (figure 2).
Une étude récente de suivi des déplacements des requins-taupes bleus marqués au large de la péninsule du Yucatan, au Mexique (n = 12), et au large du Maryland, aux États-Unis (n = 14), a montré des déplacements propres à la région et peu de chevauchement des aires de répartition qui fournirait la preuve d’une structuration spatiale à des échelles plus petites que celle actuellement considérée (Vaudo et al., 2016).
Il n’existe aucun modèle accepté de la migration du requin-taupe bleu dans l’Atlantique Nord. Maia et al. (2007) ont résumé les connaissances sur la dispersion et la migration dans l’Atlantique Nord. Il semble que les femelles migrent vers les 20 à 30°de latitude nord pour mettre bas, d’après les données selon lesquelles aucune femelle n’a été capturée en dehors de ces latitudes (bien que de telles inférences soient limitées par la disponibilité des données des observateurs scientifiques). Les mâles ont tendance à fréquenter davantage les latitudes élevées que les femelles, d’après la fréquence des données sur les captures observées, mais cette tendance pourrait être en partie due à la séparation dans la colonne d’eau, où les femelles passeraient plus de temps en profondeur, ou à un effet de la sélectivité des engins de pêche. Schrey et Heist (2003) ont émis l’hypothèse voulant que les femelles soient philopatriques à des lieux de mise bas (qu’il reste à repérer) et que les mâles parcourent de plus grandes distances, en se fondant sur la structure de la population déterminée par les microsatellites et l’ADNmt (voir la section Structure spatiale et variabilité de la population). Il est difficile de mettre au point un modèle de migration accepté, car l’on manque d’information sur les lieux de mise bas et que l’on ne comprend pas entièrement les profils de déplacement des mâles/femelles et des individus immatures/matures sur un cycle annuel.
D’après des études sur l’alimentation des adultes, le requin-taupe bleu se nourrit d’une grande variété d’espèces, principalement le calmar et des poissons osseux (ostéichthyens), dont le thon, le maquereau, la bonite et l’espadon (Bowman et al., 2000). Certains sont d’avis que les requins-taupes bleus de grande taille tendent à chasser des proies plus grosses, par exemple d’autres requins, de petits cétacés et des tortues. D’après deux méthodes d’échantillonnage utilisées dans l’ouest de l’Atlantique Nord, le tassergal (Pomatomus saltatrix) et le stromaté (Peprilus triacanthus) étaient les plus importantes proies, composant respectivement 78 et 31 % de l’alimentation de l’espèce (Bowman et al., 2000).
L’information sur l’abondance et les tendances provient principalement d’indices qui dépendent des pêches tant de l’intérieur que de l’extérieur du Canada. Le requin-taupe bleu est le plus abondant en eaux canadiennes de juin à décembre, période où la température de l’eau est la plus élevée. Les eaux canadiennes ne représentent qu’environ 2,5 % de l’aire de répartition géographique mondiale de la population de l’Atlantique Nord. Par conséquent, les indices provenant de l’extérieur du Canada sont pertinents dans l’évaluation de la situation de l’espèce au Canada. Le MPO (Showell et al., 2017) et la CICTA (ICCAT, 2012) ont tous deux fourni des documents résumant les données dépendantes des pêches existantes dans l’aire de répartition de l’Atlantique Nord du requin-taupe bleu, principales sources sur lesquelles est basé le présent rapport de situation.
En eaux canadiennes, les prises accessoires de requins-taupes bleus déterminées à partir de sorties de pêche à la palangre pélagique contrôlées par des observateurs dans la Région des Maritimes de 1996 à 2014 sont les seules données disponibles pour définir un indice d’abondance et comprendre la démographie de la population (Showell et al., 2017). La couverture par les observateurs en mer des pêches à la palangre pélagique est d’environ 5 % depuis 2004. Les autres pêches qui interagissent avec le requin-taupe bleu (p. ex. au filet maillant et au chalut) ont une couverture de 1 à 18 %, selon la région et le type d’engin. Il convient aussi de noter que les captures canadiennes se font à la périphérie de l’aire de répartition de l’UD de l’Atlantique Nord et que, par conséquent, les indices ne doivent pas être considérés séparément des indices qui couvrent la totalité de l’aire de répartition du requin-taupe bleu dans l’Atlantique Nord.
Il existe plusieurs indices de taux de capture provenant des débarquements des pêches commerciales à la palangre dans d’autres parties de l’aire de répartition. L’analyse la plus récente de tous les taux de capture a été revue et publiée par la CICTA (ICCAT, 2012). L’analyse de la CICTA a relevé les six séries de taux de capture les plus représentatives : une des pêches à la palangre du Japon (1994-2010); deux des pêches à la palangre des États-Unis (données d’observateurs de 1992 à 2010, données de journaux de bord de 1986 à 2010); une des pêches récréatives des États-Unis (1981-2010); une des pêches à la palangre du Portugal (2000-2010); une des pêches à la palangre de l’Espagne (1990-2010) (figure 9 a, b).
La CICTA (ICCAT, 2012) a mis au point deux modèles de population, soit un modèle bayésien de production excédentaire et un modèle « sans capture », au moyen des quatre indices de taux de capture des pêches à la palangre (États-Unis, Japon, Portugal, Espagne). De plus, la CICTA a appliqué un facteur d’estimation des captures pour mieux tenir compte de la mauvaise comptabilisation des captures dans les jeux de données historiques.
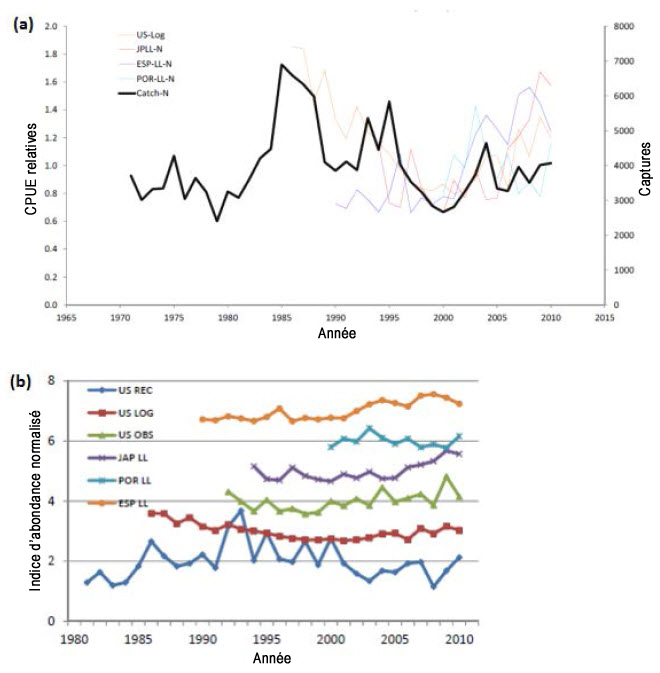
Description longue de la figure 9
Graphiques indiquant les indices d’abondance du requin-taupe bleu dans l’Atlantique Nord et les captures totales (graphique a) et les indices de taux de capture normalisés de requins-taupes bleus produits par Showell et al. (2017) (graphique b).
Un indice d’abondance fondé sur les indices de taux de capture canadiens normalisés provenant de la pêche à la palangre pélagique dans la région des Maritimes a été élaboré par Campana et al. (2004a) et a ensuite été utilisé pour mettre à jour la série de taux de capture (figure 10; Fowler et Campana, 2009; Showell et al., 2017). Après application des taux de capture à un modèle linéaire généralisé, la mise à jour la plus récente (figure 10) montre une baisse des taux de capture comparativement aux données du début de la série, mais cette baisse n’est pas significative. Les variables qui expliquent l’utilité limitée de l’indice canadien sont notamment le petit nombre de sorties annuelles en mer avec observateurs au cours desquelles des prises de requins-taupes bleus ont été réalisées (fourchette : 11-95) et la faible couverture spatiale non seulement dans l’aire de répartition canadienne, mais aussi dans l’aire de répartition à l’échelle de l’Atlantique Nord.
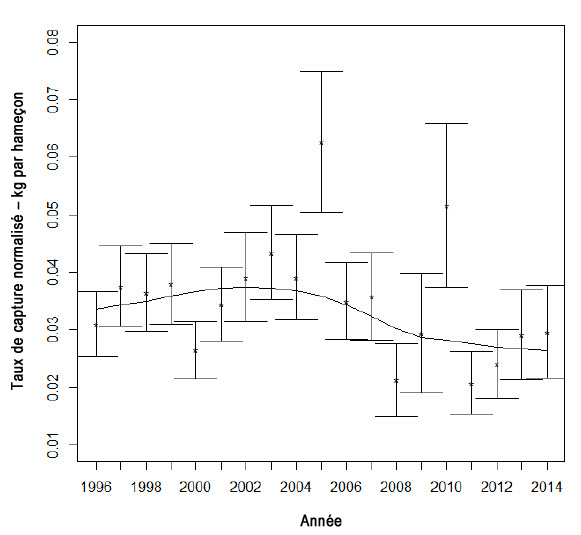
Description longue de la figure 10
Graphique indiquant les taux de capture normalisés de requins-taupes bleus des pêches à la palangre pélagique dans la Région des Martitimes, de 1996 à 2014, sur le plateau néo-écossais.
Les six indices du taux de capture les plus fiables relevés par la CICTA (ICCAT, 2012) ont été sélectionnés dans le cadre d’une évaluation systématique basée sur 12 questions visant à examiner la suffisance des séries de captures par unité d’effort (CPUE) (couverture géographique, durée des séries chronologiques, etc.; voir le tableau 14, ICCAT, 2012). Bien que les six séries résultantes soient considérées comme les meilleures disponibles, elles ont de grandes limites parce qu’elles dépendent des pêches et que, dans certains cas, elles sont très variables. Showell et al. (2017) ont normalisé les six indices au moyen des données disponibles dans l’évaluation de 2012 de la CICTA. Les indices ne montrent aucune tendance uniforme pour la population de l’Atlantique Nord (figure 9a, b). Selon l’interprétation générale de la CICTA, les dernières années de chaque série de CPUE affichent une tendance à la hausse ou stable, alors que les captures demeurent relativement constantes (figures 9 et 14).
| Année | Région | Palangre | Ligne à main | Filet maillant | Chalut à panneaux | Autre | Tournoi | Total régional | Total annuel |
|---|---|---|---|---|---|---|---|---|---|
| 1993 | Maritimes | cellule vide | cellule vide | 0,3 | cellule vide | cellule vide | cellule vide | 0,3 | 3,71 |
| cellule vide | T.-N. | 1,1 | cellule vide | 2,3 | cellule vide | 0,0 | cellule vide | 3,41 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| 1994 | Maritimes | 117,6 | 2,3 | 9,5 | 1,7 | 0,1 | cellule vide | 131,2 | 142,4 |
| cellule vide | T.-N. | 6,5 | cellule vide | 4,5 | cellule vide | cellule vide | cellule vide | 11 | cellule vide |
| cellule vide | Québec | cellule vide | 0,2 | cellule vide | cellule vide | cellule vide | cellule vide | 0,2 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| 1995 | Maritimes | 88,0 | 0,2 | 13,4 | 0,7 | 0,5 | cellule vide | 102,8 | 111,2 |
| cellule vide | T.-N. | 5,9 | cellule vide | 2,4 | cellule vide | cellule vide | cellule vide | 8,3 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| 1996 | Maritimes | 50,5 | 0,3 | 7,8 | 1,0 | cellule vide | 0,1 | 59,6 | 67,51 |
| cellule vide | T.-N. | 5,6 | cellule vide | 2,3 | cellule vide | 0,0 | cellule vide | 7,91 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,0 | 0 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| 1997 | Maritimes | 90,2 | 0,2 | 9,3 | 1,5 | cellule vide | cellule vide | 101,2 | 109,5 |
| cellule vide | T.-N. | 4,0 | cellule vide | 4,0 | 0,1 | cellule vide | cellule vide | 8,1 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,2 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,2 | cellule vide |
| 1998 | Maritimes | 46,2 | 0,2 | 8,0 | 2,2 | 0,6 | cellule vide | 57,2 | 70,9 |
| cellule vide | T.-N. | 9,5 | cellule vide | 4,0 | cellule vide | cellule vide | cellule vide | 13,5 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,2 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,2 | cellule vide |
| 1999 | Maritimes | 45,8 | cellule vide | 4,8 | 1,8 | 0,7 | cellule vide | 53,1 | 70,4 |
| cellule vide | T.-N. | 7,8 | 0,1 | 9,2 | 0,1 | cellule vide | cellule vide | 17,2 | cellule vide |
| cellule vide | Québec | 0,0 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| 2000 | Maritimes | 48,2 | 0,1 | 5,3 | 0,4 | 0,8 | 0,49 | 54,8 | 79,5 |
| cellule vide | T.-N. | 10,7 | cellule vide | 12,9 | 0,1 | 0,5 | cellule vide | 24,2 | cellule vide |
| cellule vide | Québec | 0,0 | cellule vide | cellule vide | cellule vide | cellule vide | 0,3 | 0,3 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,2 | 0,2 | cellule vide |
| 2001 | Maritimes | 51,2 | 0,2 | 5,2 | 0,2 | 0,4 | cellule vide | 57,2 | 69,7 |
| cellule vide | T.-N. | 8,6 | cellule vide | 3,6 | 0,1 | cellule vide | cellule vide | 12,3 | cellule vide |
| cellule vide | Québec | 0,0 | 0,1 | cellule vide | cellule vide | 0,0 | cellule vide | 0,1 | cellule vide |
| cellule vide | Golfe | 0,0 | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide | 0,1 | cellule vide |
| 2002 | Maritimes | 54,3 | 0,3 | 9,8 | 0,8 | 1,3 | 0,67 | 66,5 | 79,3 |
| cellule vide | T.-N. | 6,4 | 0,1 | 4,5 | cellule vide | cellule vide | cellule vide | 11 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | 0,1 | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| cellule vide | Golfe | 0,8 | cellule vide | 0,2 | cellule vide | cellule vide | 0,7 | 1,7 | cellule vide |
| 2003 | Maritimes | 57,6 | 0,2 | 6,8 | 0,5 | 1,4 | 0,40 | 66,5 | 74 |
| cellule vide | T.-N. | 6,0 | cellule vide | 1,4 | cellule vide | 0,1 | cellule vide | 7,5 | cellule vide |
| cellule vide | Québec | 0,0 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| 2004 | Maritimes | 62,1 | 6,8 | 0,1 | 0,1 | 1,0 | 1,00 | 70,2 | 81,4 |
| cellule vide | T.-N. | 8,0 | 3,0 | cellule vide | cellule vide | cellule vide | cellule vide | 11 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,2 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,2 | cellule vide |
| 2005 | Maritimes | 71,3 | 11,9 | 0,9 | 0,9 | 0,9 | 0,39 | 85,5 | 95,7 |
| cellule vide | T.-N. | 5,3 | 4,4 | 0,1 | 0,1 | cellule vide | cellule vide | 9,8 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,4 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,4 | cellule vide |
| 2006 | Maritimes | 61,5 | 4,9 | 0,3 | 0,3 | cellule vide | 0,39 | 66,7 | 70,4 |
| cellule vide | T.-N. | 2,4 | 1,2 | cellule vide | cellule vide | cellule vide | cellule vide | 3,6 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide | 0,1 | cellule vide |
| 2007 | Maritimes | 61,3 | 6,0 | 0,8 | 0,8 | cellule vide | 0,20 | 68,1 | 71,3 |
| cellule vide | T.-N. | 1,9 | 1,0 | cellule vide | cellule vide | 0,0 | cellule vide | 2,9 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,2 | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide | 0,3 | cellule vide |
| 2008 | Maritimes | 39,3 | 2,3 | 0,7 | 0,7 | 1,3 | cellule vide | 43,6 | 45,8 |
| cellule vide | T.-N. | 2,0 | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | 2,1 | cellule vide |
| cellule vide | Québec | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| 2009 | Maritimes | 46,6 | 1,7 | 0,2 | 0,2 | cellule vide | 0,49 | 48,5 | 53 |
| cellule vide | T.-N. | 3,5 | 0,9 | cellule vide | cellule vide | cellule vide | cellule vide | 4,4 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| 2010 | Maritimes | 37,0 | 0,5 | 0,1 | 0,1 | 0,3 | 0,25 | 37,9 | 41,3 |
| cellule vide | T.-N. | 1,5 | 1,5 | cellule vide | cellule vide | cellule vide | cellule vide | 3 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Golfe | 0,2 | 0,2 | cellule vide | cellule vide | cellule vide | cellule vide | 0,4 | cellule vide |
| 2011 | Maritimes | 35,6 | 0,1 | cellule vide | cellule vide | 0,1 | 0,15 | 35,8 | 37,6 |
| cellule vide | T.-N. | 1,3 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 1,3 | cellule vide |
| cellule vide | Québec | 0,2 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,2 | cellule vide |
| cellule vide | Golfe | 0,2 | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide | 0,3 | cellule vide |
| 2012 | Maritimes | 28,4 | 0,2 | 0,5 | 0,5 | cellule vide | 0,42 | 29,1 | 29,7 |
| cellule vide | T.-N. | cellule vide | 0,4 | cellule vide | cellule vide | cellule vide | cellule vide | 0,4 | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide | 0,1 | cellule vide |
| cellule vide | Golfe | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| 2013* | Maritimes | 34,4 | 0,4 | cellule vide | cellule vide | cellule vide | 0,32 | 35,1 | 35,3 |
| cellule vide | T.-N. | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0 | cellule vide |
| cellule vide | Québec | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| cellule vide | Golfe | 0,1 | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | 0,1 | cellule vide |
| 2014* | Maritimes | 53,2 | 1,5 | cellule vide | cellule vide | cellule vide | 0,32 | 35,1 | 35,3 |
| cellule vide | T.-N. | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| cellule vide | Québec | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
| cellule vide | Golfe | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide | cellule vide |
*Les données pour les Régions de Terre-Neuve, du Québec et du Golfe étaient incomplètes au moment de la publication.
L’évaluation la plus récente de la population de requins-taupes bleus de l’Atlantique Nord a été entreprise par la CICTA (ICCAT, 2012) et utilise les données allant jusqu’à 2010. Étant donné le manque d’uniformité de la comptabilisation, les captures rapportées n’ont pas été considérées comme étant fiables pour être utilisées dans le modèle. On a donc dérivé une nouvelle série de captures, dans laquelle les débarquements et les rejets sont rapportés au prorata afin de combler les lacunes dans les séries chronologiques (tableau 8 dans ICCAT, 2012). Ainsi, le nombre de captures estimé (débarquements + rejets) dans les séries chronologiques utilisées aux fins de modélisation était considérablement plus élevé que les estimations tirées des débarquements déclarés.
On a utilisé un modèle bayésien de production excédentaire pour estimer la situation de la population en incorporant les captures estimées ainsi que quatre des indices susmentionnés : série tirée des journaux de bord des pêches à la palangre des États-Unis, série tirée des pêches à la palangre du Japon, série tirée des pêches à la palangre du Portugal et série tirée des pêches à la palangre de l’Espagne (figure 9a). Seize formules de modèle ont été exécutées au moyen de diverses combinaisons d’indices et d’options de pondération. Le rendement équilibré maximal (REM), ou rendement maximal durable (RMD), calculé à partir des diverses exécutions variait de 5 300 à 24 000 tonnes, la moyenne étant d’environ 19 000 t. Dans tous les cas, les estimations de la biomasse dépassaient la BREM (biomasse qui produirait le REM), la plupart étant d’environ 1,75. Quatre exécutions du modèle montrant la tendance à long terme du rapport B/BREM révèlent une tendance à la hausse ou stable depuis 1971, mais l’évaluation a révélé une mauvaise correspondance entre les indices et les prévisions du modèle (figure 11). Les récentes estimations des captures, soit environ 4 000 t/an, sont moins élevées que la limite inférieure de la fourchette de valeurs du REM (figure 14; voir la section Menaces).
Un modèle « sans capture » intégrant seulement les données sur les CPUE de 1956 à 2010 a également été étudié. Dix formules ont été exécutées pour l’Atlantique Nord, et elles ont produit des résultats semblables à ceux obtenus avec le modèle bayésien de production excédentaire; toutefois, aucun rendement n’a pu être estimé, et aucun intervalle de confiance n’a été fourni. Deux des dix exécutions représentant la fourchette la plus élevée ont été incluses dans le rapport de la CICTA (ICAAT, 2012) (figure 12). Le pire scénario indiquait un rapport BSR/BSR0 de 0,55, tandis que le meilleur scénario indiquait un rapport de 0,71. Pour toutes les exécutions, la CICTA a conclu que la biomasse estimée correspondait mal à la série de CPUE, ce qui donne à penser qu’une amélioration des connaissances sur les paramètres et les facteurs biologiques influant sur la série de CPUE s’impose.
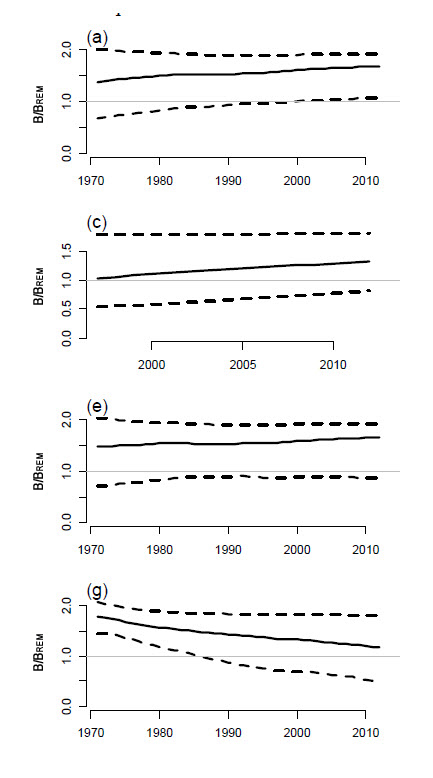
Description longue de la figure 11
Graphique à quatre volets illustrant la médiane du rapport actuel entre la biomasse fondée sur le rendement équilibré maximal (BREM) du requin-taupe bleu, avec les intervalles de confiance à 80 % de quatre exécutions du modèle bayésien de production excédentaire. L’exécution 1 correspond à une pondération égale à partir de 1971, l’exécution 3, à une pondération égale à partir de 1997, l’exécution 6, à une pondération fondée sur la superficie à partir de 1971, et l’exécution 13, au nombre de captures estimé d’après l’effort jusqu’en 1996.
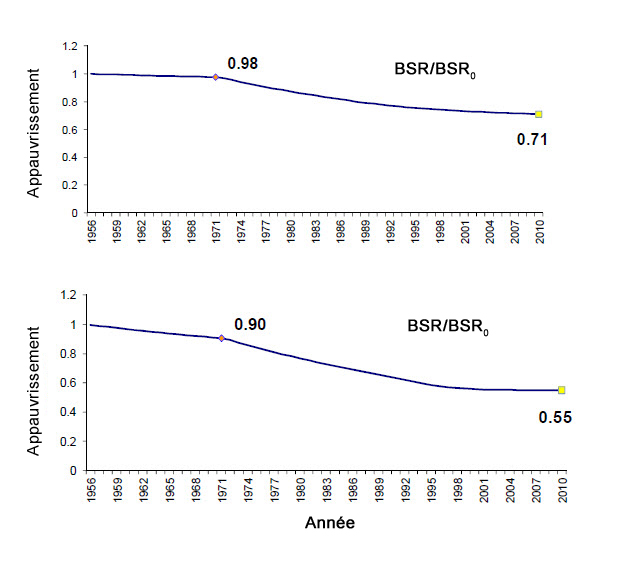
Description longue de la figure 12
Graphiques illustrant les indices d’appauvrissement de la biomasse relative des stocks reproducteurs de requin-taupe bleu dans l’Atlantique Nord selon deux exécutions du modèle « sans capture », de 1956 à 2010.
Dans l’ensemble, la CICTA (ICCAT, 2012) a conclu que les indices d’abondance et les données sur les débarquements affichaient des tendances à la hausse ou stables depuis l’évaluation précédente (2008). Les signes de surpêche potentielle qui ressortaient de l’évaluation des stocks de 2008 ont diminué, et les taux de capture actuels pourraient être considérés comme durables. Les résultats indiquaient que la probabilité de surpêche était faible; toutefois, vu l’incertitude entourant les données sur les captures, les intervalles de confiance des tendances et des autres paramètres produits par les analyses sont étendus.
Les individus font partie d’une seule population répartie sur un vaste territoire dans la moitié nord de l’océan Atlantique. Les eaux canadiennes, en périphérie de l’aire de répartition géographique totale, représentent seulement 2,5 % de celle-ci. Les menaces au Canada représentent seulement une petite portion de toutes les menaces (Showell et al., 2017). Par conséquent, la disparition de l’espèce du pays est très improbable. Par contre, si elle devait survenir, une immigration de source externe serait possible à partir de la grande population, à moins que toute la population de l’UD subisse elle aussi un grave déclin.
La mortalité directe due aux diverses pêches dirigées et aux prises accessoires est la seule menace qui pèse sur le requin-taupe bleu dans l’ensemble de son aire de répartition. Bien que l’espèce ne soit pas ciblée au Canada, elle fait l’objet de captures, et les débarquements en tant que prises accessoires sont permis dans le cadre de certaines pêches. Dans les eaux canadiennes, les prises accessoires de requins-taupes bleus dans les pêches à la palangre pélagique ciblant le thon et l’espadon sont la principale cause de mortalité; les pêches au filet maillant et au chalut visant les poissons de fond rapportent un moins grand nombre d’interactions avec l’espèce (tableau 1; figure 13a). Les prises par les pêches récréatives en eaux canadiennes sont considérées comme négligeables (Campana et al., 2004b). Environ 89 % des débarquements déclarés au Canada entre 1994 et 2014 proviennent de la Région des Maritimes (figure 13b). Les données sur les débarquements au Canada sont considérées comme une forte sous-estimation du total des prises accessoires puisque les rejets ne sont pas déclarés. Le nombre de débarquements a baissé depuis 2008. Cette baisse pourrait être notamment expliquée par le fait que la flottille canadienne de palangriers est passée des hameçons en J aux hameçons circulaires (DFO, 2016). À l’extérieur du Canada, les flottilles de palangriers commerciaux des pays étrangers sont les principales sources de mortalité. Les autres types d’engins et de pêches, dont la pêche récréative aux États-Unis, sont responsables d’un nombre moins élevé de cas de mortalité (figure 14).
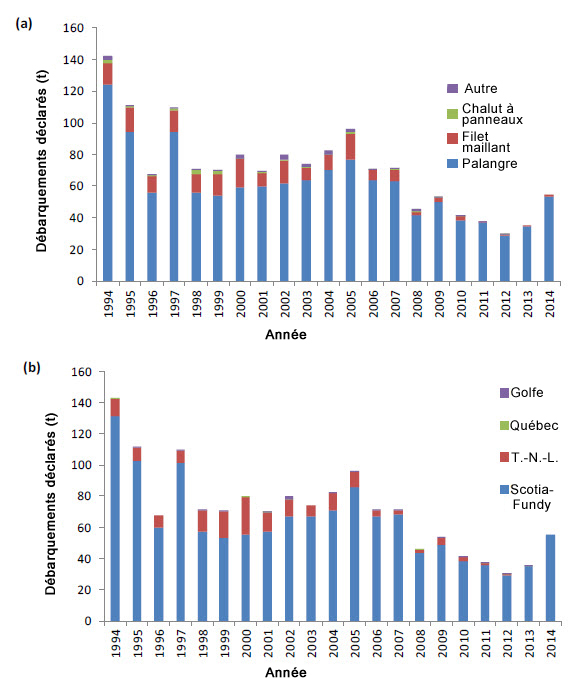
Description longue de la figure 13
Graphiques illustrant les débarquements canadiens de requins-taupes bleus (tonnes) par type d’engin de pêche (graphique a) et par région de gestion (graphique b), de 1994 à 2014.
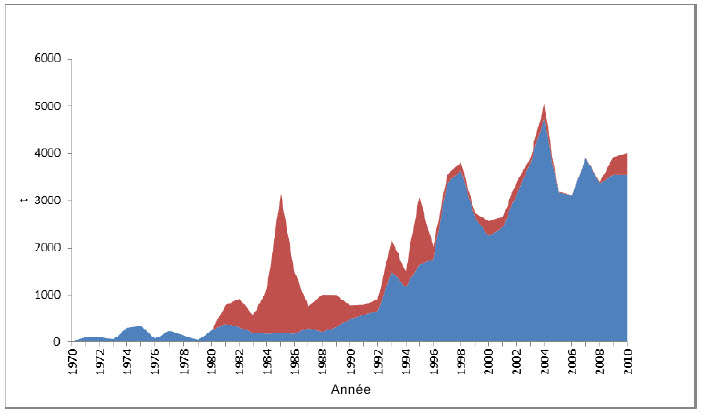
Description longue de la figure 14
Graphique illustrant les captures estimées de requins-taupes bleus (débarquements et rejets) dans l’Atlantique Nord par des palangriers et d’autres engins, de 1970 à 2010.
À partir de 2015, la flottille canadienne de palangriers dans la Région des Maritimes a volontairement appuyé les mesures de gestion visant à relâcher les requins-taupes encore vivants après le retrait de l’engin de pêche. La survie après remise à l’eau des requins-taupes bleus capturés par une palangre a été estimée seulement récemment (Campana et al., 2015). La survie des individus remis à l’eau dépend des conditions au moment de leur capture et de leur remise à l’eau, qui peuvent varier en fonction de la méthode de capture, des techniques de mouillage des engins, du temps avant le retrait de l’engin, de la taille des requins, du traitement des requins à bord des bateaux et des conditions environnementales (Campana et al., 2015). Campana et al. (2015) ont estimé le taux de mortalité des requins-taupes bleus remis à l’eau dans le cadre des pêches canadiennes à la palangre pélagique à 49 % en se basant sur les estimations de la condition des individus au moment de leur remise à l’eau (morts, blessés ou sains). La survie après remise à l’eau des individus blessés et sains a été estimée au moyen des étiquettes satellites (n = 33) (figure 15).
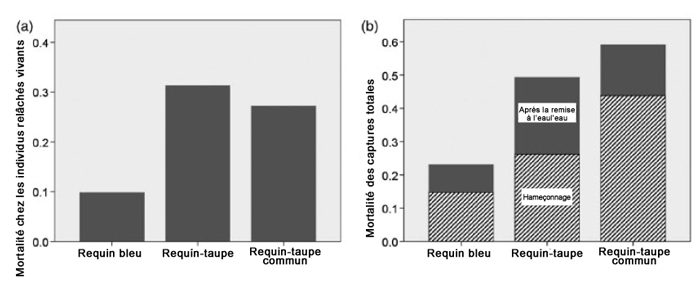
Description longue de la figure 15
Graphiques illustrant la mortalité des requins due à la capture ou à l’hameçonnage dans les pêches commerciales canadiennes à la palangre pélagique, répartie par espèce (requin bleu, requin-taupe et requin-taupe commun). Le graphique a montre la proportion de requins qui meurent après la remise à l’eau, d’après les ECDD. Le graphique b montre la proportion des captures totales qui meurent pendant l’hameçonnage et après la remise à l’eau.
La mortalité annuelle totale dans la Région des Maritimes (1996-2014) a été calculée en appliquant un taux de mortalité de 49 % aux estimations des rejets totaux (à partir des rapports des observateurs) des pêches à la palangre pélagique, des pêches au chalut à panneaux visant les poissons de fond, des pêches à la palangre visant les poissons de fond et des pêches au filet maillant visant les poissons de fond (Showell et al., 2017). Les rejets estimés de requins morts ont été ajoutés aux débarquements déclarés, ce qui a permis d’obtenir une mortalité moyenne totale de 69 t/an (fourchette : 42-115 t/an; 1996-2014) (Showell et al., 2017).
Dans la Région de Terre-Neuve-et-Labrador, les estimations des prises accessoires de requins-taupes bleus ont été calculées en extrapolant le rapport des sorties de pêche contrôlées par des observateurs sur les sorties de pêche non contrôlées par des observateurs de 1998 à 2010 de plusieurs pêches (figure 16; Showell et al., 2017). Les estimations annuelles des prises étaient très variables à cause de la faible couverture par les observateurs (de 0 à 3 %). Les estimations vont de quasi nulles à 174 t, avec une moyenne annuelle de 80 t (figure 16). La plupart des prises estimées sont tirées de la pêche au filet maillant ciblant la morue, qui aurait un taux de mortalité de 100 %. Par conséquent, la mortalité annuelle totale au Canada est estimée à 150 t/an.
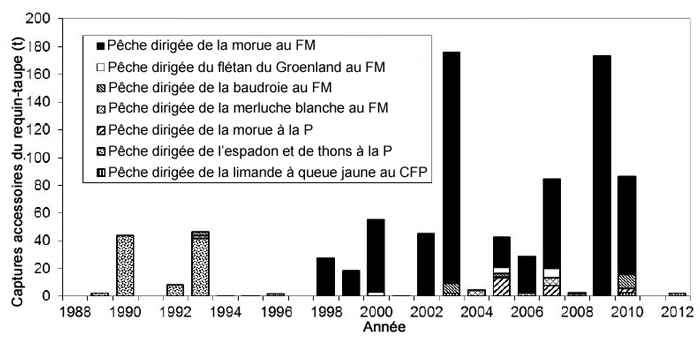
Description longue de la figure 16
Graphique illustrant les prises accessoires annuelles estimées (tonnes) de requins-taupes bleus par espèce ciblée et par engin (filet maillant, palangre et chalut de fond à panneaux) dans la zone économique exclusive du Canada se trouvant dans les divisions 3LNOP, de 1998 à 2010.
La forte incertitude entourant la mortalité par pêche à l’échelle de l’Atlantique Nord découle de la mauvaise comptabilisation des prises, les estimations des rejets en mer étant pratiquement inexistantes et les débarquements ne faisant pas l’objet de déclarations complètes. Par exemple, l’Organisation des pêches de l’Atlantique Nord-Ouest (OPANO) n’exige pas de ses pays membres qu’ils consignent les rejets de requins dans le cadre des pêches menées dans la zone qu’elle réglemente (soit à l’extérieur des ZEE des pays), ce qui explique pourquoi on ignore le nombre total de rejets de requins-taupes bleus en haute mer. L’exactitude des débarquements déclarés s’est améliorée après 1996, année où la CICTA a exigé la déclaration des débarquements de requins.
De 1996 à 2010, les débarquements moyens déclarés s’élevaient à quelque 2 395 t, mais ce nombre est considéré comme une sous-estimation. Étant donné la sous-déclaration des débarquements et des rejets, la CICTA (ICCAT, 2012) a fourni de meilleures estimations en calculant le rapport entre les débarquements de requins-taupes bleus et les débarquements totaux de thons et d’espadons de chaque flottille ces dernières années, puis en le multipliant par la somme des débarquements de thons et d’espadons chaque année historique. De 1996 à 2010, les estimations fondées sur ce rapport étaient en moyenne de 3 527 t (fourchette : 2 668-4 648 t). Toutefois, ces estimations fondées sur les débarquements doivent être interprétées avec prudence, car elles ne tiennent pas compte des rejets de requins en mer et ne représentent ainsi que des taux de mortalité minimale. Si l’on utilise la mortalité moyenne annuelle totale estimée pour le Canada (soit 150 t), les pêches canadiennes seraient responsables d’environ 4 % de la mortalité globale en termes de poids. La remise à l’eau volontaire des individus vivants depuis 2015 dans le cadre des pêches à la palangre pélagique dans la Région des Maritimes devrait faire diminuer encore plus la mortalité en eaux canadiennes, mais pas le nombre d’interactions avec des engins de pêche.
Comme le décrit la section Biologie ci-dessus, les femelles matures sont rarement capturées dans les pêches canadiennes. Les données des observateurs recueillies de 2006 à 2015 indiquent que seulement 1 % des femelles observées capturées ont atteint la maturité sexuelle. Toutefois, ce constat doit être interprété avec prudence, car la majorité des prises accessoires de requins-taupes bleus demeurent non déclarées à cause de la faible couverture annuelle par les observateurs en mer des pêches de l’Atlantique canadien qui ont des interactions avec des requins.
Les requins-taupes bleus renferment des teneurs élevées en contaminants, dont des PCB, des DDT, des pesticides et du mercure (Lyons et al. 2013). Toutefois, l’on ne sait pas si ces contaminants représentent une source de mortalité pour la population.
Les populations de requins-taupes bleus, comparativement à celles d’autres espèces de requins, ont une faible productivité, qui limite leur capacité de se rétablir une fois qu’elles sont appauvries. La CICTA (ICCAT, 2012) a réalisé une analyse de productivité et de sensibilité de 20 populations de requins et déterminé que la faible productivité par rapport à celle d’autres espèces de requins était principalement due au temps que les femelles prennent pour atteindre la maturité et à leur cycle de reproduction de trois ans (tableau 2; ICCAT, 2012).
| Stock | Productivité (r) | LCI | LCS | Durée d’une génération (ans) |
|---|---|---|---|---|
| Requin bleu (Atlantique Nord) | 0,314 | 0,279 | 0,345 | 8,2 |
| Requin bleu (Atlantique Sud) | 0,299 | 0,264 | 0,327 | 9,8 |
| Pastenague violette (Atlantique Nord) | 0,230 | 0,181 | 0,279 | 6,2 |
| Requin-marteau commun | 0,225 | 0,213 | 0,237 | 13,4 |
| Requin-tigre | 0,190 | 0,180 | 0,200 | 15,6 |
| Requin longimane | 0,121 | 0,104 | 0,137 | 10,4 |
| Requin-marteau halicorne (Atlantique Sud) | 0,121 | 0,110 | 0,132 | 21,6 |
| Requin-renard | 0,121 | 0,099 | 0,143 | 11,0 |
| Requin-marteau halicorne (Atlantique Nord) | 0,096 | 0,093 | 0,107 | 21,6 |
| Requin soyeux (Atlantique Nord) | 0,078 | 0,065 | 0,090 | 14,4 |
| Grand requin-marteau | 0,070 | 0,069 | 0,071 | 27,1 |
| Requin-taupe bleu | 0,058 | 0,049 | 0,068 | 25,0 |
| Maraîche | 0,052 | 0,044 | 0,059 | 20,3 |
| Pastenague violette (Atlantique Sud) | 0,051 | 0,004 | 0,096 | 6,6 |
| Requin obscur | 0,043 | 0,035 | 0,050 | 29,6 |
| Requin soyeux (Atlantique Sud) | 0,042 | 0,029 | 0,054 | 16,5 |
| Requin de nuit | 0,041 | 0,028 | 0,053 | 14,9 |
| Petit requin-taupe | 0,029 | 0,020 | 0,038 | 25,2 |
| Requin gris | 0,010 | 0,005 | 0,024 | 21,8 |
| Requin-renard à gros yeux | 0,009 | 0,001 | 0,018 | 17,8 |
Le requin-taupe bleu a une vaste aire de répartition dans tout l’Atlantique Nord. Il est susceptible d’être capturé dans plusieurs (probablement plus de 10) pêches internationales. Au sens accordé par le COSEPAC, chaque pêcherie pourrait être considérée comme une « localité », où une menace particulière pesant sur la population serait associée à chaque localité.
Le requin-taupe bleu ne fait l’objet d’aucune pêche dirigée dans l’Atlantique canadien, et il n’y a plus de plan de gestion intégrée des pêches (PGIP) qui supervise la gestion des requins. L’espèce est plutôt gérée dans le cadre des PGIP des pêches qui capturent des requins en tant que prises accessoires (p. ex. la pêche à la palangre pélagique visant l’espadon dans les Maritimes). De plus, l’Atlantic Canada Conservation Action Plan for Selected Shark Species (CAP) a récemment été approuvé. Ce plan d’action a été élaboré afin de mettre à jour le Plan d’action national pour la conservation et la gestion des requins du Canada (DFO, 2007) en réponse au Plan d’action international pour la conservation et la gestion des requins de la FAO (FAO, 1999). Le CAP a pour objectif de gestion principal de s’assurer que les activités humaines n’exercent pas d’effets néfastes inacceptables sur l’écosystème, sans toutefois fournir de plans d’action pour aider à conserver les espèces de requins pélagiques.
Une limite non restrictive de 100 t de requins-taupes bleus vise les pêches à la palangre pélagique dans la Région des Maritimes. Les pêches canadiennes à la palangre pélagique dans les Maritimes a récemment soutenu la remise à l’eau volontaire des requins-taupes bleus vivants, ce qui devrait réduire les débarquements et la mortalité dans les années à venir, mais aucune mesure ne limite les interactions de requins avec les engins de pêche. Pour soutenir cette mesure volontaire, le Fonds mondial pour la nature-Canada (WWF-Canada) a élaboré le guide Shark Fishing – Best Catch, Handle and Release Practices, qui a été remis aux responsables de tournois de pêche aux requins et aux groupes de pêche de la Région des Maritimes pour aider à réduire la mortalité des requins après leur remise à l’eau. Pour obtenir leur permis de pêche, les palangriers pélagiques canadiens dans la Région des Maritimes doivent consigner dans des journaux de bord les prises de requins-taupes bleus, et ils doivent utiliser des hameçons circulaires corrodables pour réduire les prises accessoires et la mortalité après remise à l’eau. Dans les pêches aux poissons de fond dans la Région de Terre-Neuve-et-Labrador, les débarquements de requins sont réglementés de sorte que les requins ne représentent pas plus que 10 % des pêches ciblant les espèces de poissons de fond (en poids); aucune autre mesure de protection n’existe.
Les prises accessoires de requins-taupes bleus sont débarquées en tant que produits alimentaires, et le requin-taupe bleu a plus de valeur que le requin bleu (Prionace glauca), espèce souvent capturée par les mêmes pêches. Dans l’Atlantique canadien, les pêches aux poissons de fond utilisant des filets maillants et des palangres doivent observer une politique sur le débarquement de requins entiers adoptée par le Canada, qui vise à s’assurer que les requins pélagiques ont encore les ailerons naturellement fixés à leur carcasse, et ce, avant la pesée et la comptabilisation par un vérificateur à quai au point de débarquement. Une autre mesure de gestion des requins du Canada est la réglementation basée sur le pourcentage de 5 %, qui autorise un poids maximal de nageoires équivalent à 5 % du poids des carcasses débarquées par sortie de pêche. Les pêches dans l’Atlantique canadien qui utilisent des chaluts de fond ou des sennes coulissantes ne sont pas autorisées à conserver les prises accessoires de requins-taupes bleus. Cette interdiction ne réduit cependant pas les interactions entre les requins avec les engins de pêche. L’ablation des nageoires de requin (où les nageoires sont retirées et conservées alors que le corps du requin, parfois vivant, est rejeté en mer) est interdite depuis 1994 au Canada. Il convient toutefois de souligner que, en raison de la valeur de la chair du requin-taupe bleu, celui-ci est moins touché par le problème d’ablation des nageoires. Néanmoins, il est à noter que le Canada autorise la vente de nageoires de requin à l’intérieur de ses frontières, tout comme il permet leur exportation vers des marchés étrangers.
Le COSEPAC a évalué le requin-taupe bleu pour la première fois en 2006 et l’avait alors désigné espèce menacée, mais celui-ci n’a pas reçu de protection juridique aux termes de la Loi sur les espèces en péril du Canada. La dernière évaluation du requin-taupe bleu (sous-population de l’Atlantique) a été réalisée en 2004 par l’Union internationale pour la conservation de la nature (UICN), qui l’a coté « vulnérable » (Cailliet et al., 2009) et ajouté une annotation indiquant qu’une mise à jour était requise sur le site Web. La National Oceanographic and Atmospheric Administration (NOAA) des États-Unis a décrit le statut de la population comme suit : le stock est sain, car il ne fait pas l’objet de surpêche et la probabilité de surpêche est faible (NOAA, 2015).
L’habitat du requin-taupe bleu se trouve dans la zone économique exclusive (ZEE) du Canada, au large de la côte atlantique et est gérée par Pêches et Océans Canada. Aucune mesure de protection spécifique n’est actuellement appliquée à l’habitat occupé par le requin-taupe bleu. En eaux internationales à l’extérieur du Canada, aucune zone n’est actuellement désignée aux fins de protection de l’habitat du requin-taupe bleu.
Le rédacteur remercie Mark Showell, Mark Fowler, Warren Joyce, Mike McMahon, Mark Simpson et Carolyn Miri, qui ont mené les travaux pour le document de recherche du Secrétariat canadien de consultation scientifique (SCCS). Merci à Christie Whelan de son aide et à Aurélie Cosandey-Godin, de la WWF, d’avoir fourni ses analyses.
Bishop, S.D.H., M.P. Francis, C. Duffy, et J.C. Montgomery. 2006. Age, growth, longevity and natural mortality of the shortfin mako shark (Isurus oxyrinchus) in New Zealand waters. Marine and Freshwater Research. 57 : 143-154.
Bowman, R.E., C. E. Stillwell, W.L. Michaels, et M.D. Grosslein. 2000. Food of northwest Atlantic fishes and two common species of squid. NOAA Technical Memorandum. NMFS-NE 155, 138 p.
Cailliet, G.M., R.D. Cavanagh, D.W. Kulka, J.D. Stevens, A. Soldo, S. Clo, D. Macias, J. Baum, S. Kohin, A. Duarte, J.A. Holtzhausen, E. Acuña, A. Amorim, et A. Domingo. 2009. Isurus oxyrinchus (Atlantic subpopulation). The IUCN Red List of Threatened Species 2009 : e.T161749A5494807. [consulté le 5 janvier 2016].
Campana, S.E., L. Marks, et W. Joyce. 2004a. Biology, fishery and stock status of shortfin mako sharks (Isurus oxyrinchus) in Atlantic Canadian Waters. Canadian Science Advisory Secretariat Research Document 2004/094.
Campana, S.L. Marks, W. Joyce, et N. Kohler. 2 004 b. Influence of recreational and commercial fishing on blue shark (Prionace glauca) population in Atlantic Canadian waters. Canadian Science Advisory Secretariat Research Document 2004/069. 67 p.
Campana, S.E., W. Joyce, M. Fowler, et M. Showell. 2015. Discards, hooking, and post-release mortality of porbeagle (Lamna nasus), shortfin mako (Isurus oxyrinchus), and blue shark (Prionace glauca) in the Canadian pelagic longline fishery. ICES Journal of Marine Science, doi:10.1093/icesjms/fsv234
Carey, F.G., J.M. Teal, et J.W. Kanwisher. 1981. The visceral temperature of mackerel sharks (Lamnidae). Physiological Zoology 54: 334-344.
Casey, J.G., et N.E. Kohler. 1992. Tagging studies on the shortfin mako shark (Isurus oxyrinchus) in the western North Atlantic. Australian Journal of Marine and Freshwater Research 43: 45-60.
Compagno, L.J.V. 2001. Sharks of the world. An annotated and illustrated catalogue of shark species known to date. Volume 2. Bullhead, mackerel and carpet sharks (Heterodontiformes, Lamniformes and Orectolobiformes). United Nations Food and Agricultural Organization Species Catalogue for Fishery Purposes 1(2) Rome, FAO. 269 pp.
DFO. 2007. National plan of action for the conservation and management of Sharks. Ottawa, Canada. 31 p. (Également disponible en français : MPO. 2007. Plan d’action national pour la conservation et la gestion des requins. Ottawa, Canada. 34 p.)
DFO. 2016. Proceedings of the Zonal Peer Review Pre-COSEWIC Assessment for Shortfin Mako (Isurus oxyrinchus) in Atlantic Canada ; September 16-17, 2015. DFO Canadian Science Advisory Secretariat Proceedings Series. 2015/062.
Fowler, G.M., et S.E. Campana 2009. Commercial by-catch rates of Shortfin Mako (Isurus oxyrinchus) from longline fisheries in the Canadian Atlantic. Collective Volume of Scientific Papers. International Commission for the Conservation of Atlantic Tunas 64(5) 1668-1676.
Heist, E.J., J. A. Musick, et J. E. Graves. 1996. Genetic population structure of the shortfin mako (Isurus oxyrinchus) inferred from restriction fragment length polymorphism analysis of mitochondrial DNA. Canadian Journal of Fisheries and Aquatic Sciences 53:583–588.
Godin, A., T. Wimmer, et B. Worm. 2015. Pre-COSEWIC Assessment for Shortfin Mako: Bycatch and discard in Canadian fisheries. Unpublished paper presented to Pre-COSEWIC meeting, September 16-17, 2015, Bedford Institute of Oceanography, Bedford, Nova Scotia, Canada Correia.
FAO. 1999. International Plan of Action for reducing incidental catch of seabirds in longline fisheries. International Plan of Action for the conservation and management of sharks. International Plan of Action for the management of fishing capacity. Rome, FAO. 26p. (Également disponible en français : FAO. 1999. Plan d’action international visant à réduire les captures accidentelles d’oiseaux de mer par les palangriers. Plan d’action international pour la conservation et la gestion des requins. Plan d’action international pour la gestion de la capacité de pêche. Rome, FAO. 27 p.)
ICCAT. 2012. Shortfin mako stock assessment and ecological risk assessment meeting (Version PDF; 5,8 Mo) . [consulté en juin 2016]. (Également disponible en français : ICCAT. 2012. Réunion de 2012 d’évaluation du stock de requin-taupe bleu et d’évaluation des risques écologiques (Version PDF; 122 Ko)).
Kohler, N. E., J.G. Casey, et P.A. Turner. 1998. NMFS cooperative shark tagging program, 1962-93: an atlas of shark tag and recapture data. Marine Fisheries Review 60:1-87.
Loefer, J.K., G. R. Sedberry, et J.C. McGovern. 2005. Vertical movements of a shortfin mako in the western North Atlantic as determined by pop-up satellite tagging. Southeastern Naturalist 4:237–246.
Lyons, K., A. Carlisle, A. Preti, C. Mull, M. Blasius, J. O’Sullivan, C. Winkler, et C.G. Lowe. 2013. Effects of trophic ecology and habitat use on maternal transfer of contaminants in four species of young of the year lamniform sharks. Marine Environmental Research 90 : 27-38.
Maia, A., N. Queiroz, H.N. Cabral, A. M. Santos, et J.P. Correia. 2007. Reproductive biology and population dynamics of the shortfin mako, Isurus oxyrinchus Rafinesque, 1810, off the southwest Portuguese coast, eastern North Atlantic. Journal of Applied Ichthyology 23: 246–251.
Mollet, H.F., G. Cliff, H.L. Pratt Jr., et J.D. Stevens. 2000. Reproductive biology of the female shortfin mako, Isurus oxyrinchus, with comments on the embryonic development of lamnoids. Fisheries Bulletin 98:299-318.
Natanson, L.J., N.E. Kohler, D. Ardizzone, G.M. Cailliet, S.P. Wintner, et H.F. Mollet 2006. Validated age and growth estimates for the shortfin mako, Isurus oxyrinchus, in the North Atlantic Ocean. Environmental Biology of Fishes. 77 : 376-383.
NOAA. 2015. North Atlantic shortfin Mako shark stock status [consulté en janvier 2016].
Schrey, A.W., et E.J. Heist. 2003. Microsatellite analysis of population structure in the shortfin mako (Isurus oxyrinchus). Canadian Journal of Fisheries and Aquatic Sciences 60:670-675.
Showell, M.A., G.M. Fowler, W. Joyce, M. McMahon, C.M. Miri, et M.R. Simpson. 2017. Current Status and Threats to the North Atlantic Shortfin Mako Shark (Isurus oxyrinchus) Population in Atlantic Canada. DFO Can. Sci. Advis. Sec. Res. Doc. 2017/039.
Smith, S.E., D.W. Au, et C. Show. 1998. Intrinsic rebound potentials of 26 species of Pacific sharks. Marine and Freshwater Research 49:663-678.
Taguchi, M., T. Kitamura, et K. Yokawa. 2011. Genetic population structure of shortfin mako (Isurus oxyrinchus) inferred from mitochondrial DNA on inter-oceanic scale. International Scientific Committee for Tuna and Tuna-Like Species in the North Pacific Ocean Shark Working Group Workshop, 19-21 July 2011.
Vaudo, J. J., M.E. Byrne, B.M. Wetherbee, G.M. Harvey, et M.S. Shivji. 2016. Long-term satellite tracking reveals region-specific movements of a large pelagic predator, the shortfin mako shark, in the western North Atlantic Ocean. Journal of Applied Ecology. 10.1111/1365-2664.12852
Scott Wallace travaille dans les domaines de la conservation et de la gestion des océans depuis 1995 et se spécialise dans les espèces en péril et les pêches durables. Il a travaillé pour le gouvernement, des ONGE et des universités. Il a rédigé le rapport de situation du COSEPAC sur le requin-taupe bleu de 2006. Il a été membre du Sous-comité de spécialistes des poissons marins du COSEPAC de 2007 à 2015. Il est titulaire d’un doctorat de l’Université de la Colombie-Britannique.