
Comment un outil météorologique pourrait être utilisé pour prédire l’infection par le virus du Nil occidental

 Téléchargez cet article en format PDF
Téléchargez cet article en format PDFPublié par : L'Agence de la santé publique du Canada
Numéro : Volume 48-5, mai 2022 : Infections acquises par transmission vectorielle–Partie 1 : tiques & moustiques
Date de publication : mai 2022
ISSN : 1481-8531
Soumettre un article
À propos du RMTC
Naviguer
Volume 48-5, mai 2022 : Infections acquises par transmission vectorielle–Partie 1 : tiques & moustiques
Science de la mise en œuvre
La prédiction des taux d'infection minimaux dans les moustiques Culex pipiens-restuans issus du système de surveillance intégrée du virus du Nil occidental du Québec, en fonction des données météorologiques
Julie Ducrocq1, Karl Forest-Bérard1, Najwa Ouhoummane1, Elhadji Laouan Sidi1, Antoinette Ludwig2, Alejandra Irace-Cima1
Affiliations
1 Institut national de santé publique du Québec, Montréal, QC
2 Opérations et interventions scientifiques, Laboratoire national de microbiologie, Agence de la santé publique du Canada, Saint-Hyacinthe, QC
Correspondance
Citation proposée
Ducrocq J, Forest-Bérard K, Ouhoummane N, Laouan Sidi E, Ludwig A, Irace-Cima A. La prédiction des taux d'infection minimaux dans les moustiques Culex pipiens-restuans issus du système de surveillance intégrée du virus du Nil occidental du Québec, en fonction des données météorologiques. Relevé des maladies transmissibles au Canada 2022;48(5):220–32. https://doi.org/10.14745/ccdr.v48i05a03f
Mots-clés : épidémiologie, éclosions, outil de prévision, Québec, virus du Nil occidental
Résumé
Contexte : Le ministère de la Santé et des Services sociaux (MSSS) du Québec s'est montré intéressé par le développement d'un outil d'alerte précoce pour identifier les éclosions humaines saisonnières d'infection par le virus du Nil occidental, afin de moduler ses interventions en santé publique. L'objectif de cette étude était de déterminer si un outil de prédiction des taux d'infection minimaux dans les moustiques du complexe Culex pipiens-restuans, un indicateur du risque humain, avant la saison des moustiques, pouvait être développé et facilement utilisable, en employant les données météorologiques.
Méthodes : Le taux d'infection minimal annuel (nombre de lots positifs/nombre de moustiques) a été calculé pour 856 pièges entomologiques déployés de 2003 à 2006, et de 2013 à 2018 dans le sud du Québec. Les coefficients de détermination (R2) ont été estimés à l'aide du jeu de données de validation (un tiers des données sélectionnées aléatoirement) avec des équations d'estimation généralisées. Celles-ci ont été préalablement ajustées avec une méthode descendante et des termes polynomiaux, en utilisant le restant des données (représentant le jeu d'entrainement contenant les deux tiers des données), afin de minimiser les critères d'information bayésienne. Les températures moyennes et les précipitations ont été regroupées selon cinq échelles temporelles (par mois, par saison et par groupes de 4, 6 et 10 mois).
Résultats : Les températures moyennes et les précipitations cumulatives des mois précédents de mars (R2 = 0,37), mai (R2 = 0,36), décembre (R2 = 0,35) et la saison automnale (R2 = 0,38) ont expliqué environ 40 % des variations des taux d'infection minimaux annuels dans les Culex pipiens-restuans. L'inclusion de la variable « année d'échantillonnage » dans tous les modèles de régression a augmenté la capacité prédictive (R2 entre 0,42 et 0,57).
Conclusion : Tous les modèles de régression explorés ont des capacités prédictives trop faibles pour être utiles comme outil de santé publique. L'ajout d'autres facteurs associés à l'épidémiologie du virus du Nil occidental à ce type de modèle devrait être exploré.
Introduction
Le virus du Nil occidental (VNO) est la plus importante infection transmise par les moustiques au Québec depuis les deux dernières décenniesNote de bas de page 1. Depuis 2000, un système provincial de surveillance spécifique à cet arbovirus a été mis en placeNote de bas de page 2, un an après sa première détection en Amérique du Nord, à la suite d'une éclosion de maladies neuroinvasives dans l'État de New York, aux États-UnisNote de bas de page 3. Ce système est composé d'une surveillance passive rehaussée des humains, des oiseaux sauvages malades ou morts (qui agissent comme réservoirs ou hôtes accidentels) et d'une surveillance active des moustiquesNote de bas de page 4. L'introduction du VNO au Québec a été confirmé en 2002 par le système de surveillance avec des premiers cas d'infection chez des oiseaux et des cas humains acquis localementNote de bas de page 5Note de bas de page 6. Les cas neurologiques d'infection par le VNO représentent généralement une très faible proportion (environ 1 %) des personnes infectées puisque 70 % à 80 % des infections demeurent asymptomatiques, tandis que les autres présentent des symptômes non spécifiques (i.e. : syndrome grippal)Note de bas de page 3. Entre 2003 et 2018, un total de 541 cas (dont 24 décès) ont été déclarés au Québec avec les présentations cliniques suivantes : syndrome neurologique (70 %), non neurologique (23 %), asymptomatique (6 %) et inconnue (1 %)Note de bas de page 1. Deux éclosions ont été observés en 2012 (n = 134; 30 % des cas de partout au Canada) et 2018 (n = 200; 46 % des cas de partout au Canada)Note de bas de page 1Note de bas de page 7. Des fluctuations annuelles du nombre de cas humains ont été observées dans d'autres régionsNote de bas de page 8Note de bas de page 9 et semblent être associées à des phénomènes se produisant à différentes échelles géographiques et temporellesNote de bas de page 10 (i.e. selon les interactions locales entre les moustiques, les oiseaux et les humains, les facteurs environnementaux et les fluctuations climatiques à grande échelle comme le phénomène d'oscillation australe El Niño)Note de bas de page 11Note de bas de page 12Note de bas de page 13Note de bas de page 14.
Les scientifiques travaillent encore à comprendre et à identifier les facteurs qui influencent les éclosions d'infections causées par le VNO, et à élaborer des modèles qui seraient en mesure de prédire quand et où ces éclosions pourraient se produire. Le ministère de la Santé et des Services sociaux (MSSS) s'est montré intéressé par le développement d'un outil d'alerte précoce pour identifier les éclosions à l'avance et utiliser ces résultats pour atténuer les risques et réduire l'exposition humaine en modulant leurs interventions. Ces outils sont généralement basés sur les données météorologiques afin de prédire une éclosion potentielle avec un délai allant de quelques semaines à quelques mois avant que les indicateurs du système de surveillance confirment la circulation du VNONote de bas de page 15. La température et les précipitations sont les prédicteurs des infections associés au VNO les plus couramment utilisésNote de bas de page 16. L'objectif principal de ce projet est de déterminer si un outil d'alerte précoce basé sur les données météorologiques pourrait prédire les taux d'infection par le VNO dans les moustiques.
Méthodes
L'historique du système de surveillance intégrée du VNO au Québec (avec les composantes humaines, animales et entomologiques) est décrite ailleursNote de bas de page 17. Une revue de la littérature scientifique a été faite pour élaborer un graphe acyclique dirigé (figure A1) afin de conceptualiser les relations entre les facteurs potentiellement associés au VNO et identifier les variables importantes à inclure dans les modèles prédictifs, à l'aide de l'outil DAGittyNote de bas de page 18.
Les données de 13 830 lots de moustiques du complexe Culex pipiens-restuans, car ces deux espèces (Cx. pipiens et Cx. restuans) ne peuvent être différenciées morphologiquementNote de bas de page 19, ont été extraites du Système intégré des données de vigie sanitaire du virus du Nil occidentalNote de bas de page 20 pour calculer le taux d'infection minimal (TIM) annuel (nombre de lots positifs/nombre de moustiques dans chaque lot) (figure 1). Les TIM ont été calculés pour chacun des 856 lots de moustiques, regroupées pour chaque piège déployé chaque année (i.e. le piège-année) et distancés par un rayon supérieur ou égal à 1 km2 puisque cela représente la distance moyenne de déplacement de Cx. pipiens-restuans)Note de bas de page 21 et la résolution spatiale des données météorologiques (1 km2). La surveillance des moustiques a été effectuée de façon non homogène dans 12 des 18 régions sociosanitaires (RSS), avec un plus grand nombre de lots de moustiques testés pour les RSS de Montréal et de la Montérégie (tableau 1), sauf depuis 2017, où 49 pièges entomologiques ont été déployés chaque année, dans le sept mêmes RSS (Montréal = 5, Laval = 4, Montérégie = 5, Outaouais = 7, Lanaudière = 7, Mauricie-Centre-du-Québec = 7 et Capitale-Nationale = 14).
Figure 1 : Organigramme des données entomologiques extraites du Système intégré des données de vigie sanitaire du virus du Nil occidental du Québec
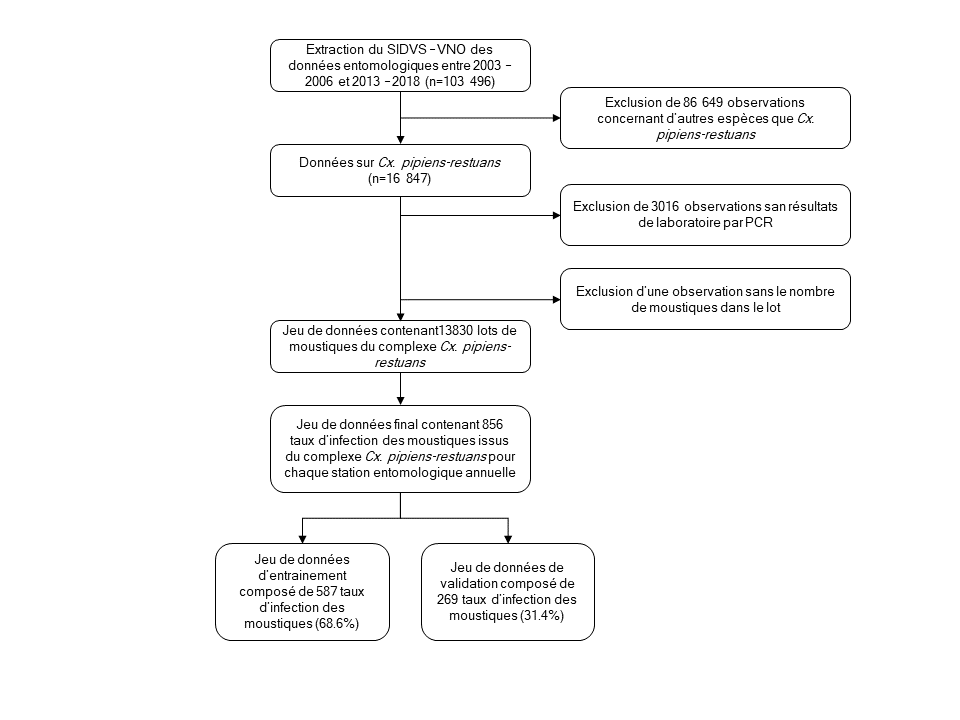
Description textuelle : Figure 1
Cette figure est un organigramme expliquant pourquoi certaines données entomologiques ont été exclues de l'analyse finale. Sur les 103 496 lots de moustiques recensés entre 2003 et 2018, 86 649 observations ont été exclues parce que les lots étaient composés d'autres espèces de moustiques que le principal vecteur du virus du Nil occidental (Culex pipiens-restuans), 3 016 observations ont été exclues parce que les lots de moustiques n'ont pas été envoyés au laboratoire et une observation a été exclue en raison de l'absence du nombre de moustiques présents dans ce lot. Ainsi, le jeu final de données est composé de 13 830 lots de moustiques du complexe Culex pipiens-restuans qui ont été regroupés en 856 taux d'infection minimaux par le VNO pour représenter chacune des stations entomologiques annuellement déployées depuis 2003. Ce jeu de données a ensuite été séparé en un jeu de données d'entrainement composé de 587 taux d'infection (2/3 des données) et en un jeu de données de validation composé de 269 taux d'infection (1/3 des données) afin d'évaluer la capacité prédictive des modèles.
| Variables spatio-temporelles | Nombre d'entrées de données | Taux d'infection minimal pour 1 000 moustiques | Température moyenne aux pièges entomologiques | Précipitations moyennes aux pièges entomologiques | |||
|---|---|---|---|---|---|---|---|
| n | IC à 95 % | n | min.; max. | n | min.; max. | ||
| Province du Québec | 856 | 3.1Tableau 1 Note de bas de page a | 2,2; 4.0Tableau 1 Note de bas de page a | 6,3 | 2,1; 8,2 | 3,0 | 2,2; 4,3 |
| Régions sociosanitaires (RSS) | |||||||
| 01 – Bas-Saint-Laurent | 0 | – | – | – | – | – | – |
| 02 – Saguenay – Lac-Saint-Jean | 6 | 0 | 0; 45,9 | 2,5 | 2,1; 2,9 | 2,6 | 2,4; 2,8 |
| 03 – Capitale Nationale | 40 | 4,4 | 1,5; 7,3 | 5,0 | 3,5; 7,8 | 3,5 | 2,9; 4,3 |
| 04 – Mauricie et Centre-du-Québec | 28 | 1,6 | 0; 4,0 | 5,4 | 4,4; 7,2 | 3,1 | 2,3; 3,5 |
| 05 – Estrie | 8 | 0 | 0; 36,9 | 5,8 | 5,0; 7,0 | 3,5 | 3,0; 3,8 |
| 06 – Montréal | 265 | 3,1 | 2,2; 3,9 | 6,4 | 5,7; 8,2 | 3,0 | 2,4; 3,9 |
| 07 – Outaouais | 47 | 3,1 | 0; 7,3 | 6,0 | 3,5; 7,8 | 2,9 | 2,2; 3,9 |
| 08 – Abitibi-Témiscamingue | 3 | 0 | 0; 70,8 | 3,2 | 3,1; 3,3 | 2,7 | 2,7; 2,8 |
| 09 – Côte-Nord | 0 | – | – | – | – | – | – |
| 10 – Nord-du-Québec | 0 | – | – | – | – | – | – |
| 11 – Gaspésie-îles-de-la-Madeleine | 0 | – | – | – | – | – | – |
| 12 – Chaudière-Appalaches | 8 | 0 | 0; 36,9 | 4,6 | 3,7; 6,2 | 3,1 | 2,7; 3,6 |
| 13 – Laval | 82 | 4,9 | 3,1; 6,7 | 6,5 | 5,7; 8,0 | 3,0 | 2,5; 4,0 |
| 14 – Lanaudière | 36 | 3,5 | 0,6; 6,5 | 6,7 | 5,4; 7,8 | 3,1 | 2,3; 4,0 |
| 15 – Laurentides | 70 | 4,8 | 2,0; 7,7 | 6,4 | 5,3; 8,0 | 3,2 | 2,6; 4,2 |
| 16 – Montérégie | 263 | 3,9 | 2,9; 5,0 | 6,5 | 5,3; 8,1 | 2,9 | 2,4; 3,9 |
| 17 – Nunavik | 0 | – | – | – | – | – | – |
| 18 – Terres-Cries-de-la-Baie-James | 0 | – | – | – | – | – | – |
| Analyse globale de type 3 : Khi-carré (valeur-p)Tableau 1 Note de bas de page b | – | 9,9 | 0,5 | 23,3 | 0,02 | 17,2 | 0,1 |
| Année | |||||||
| 2003 | 118 | 3,0 | 1,7; 4,4 | 5,5 | 3,1; 6,3 | 2,6 | 2,3; 3,4 |
| 2004 | 86 | 0,9 | 0; 2,1 | 5,7 | 2,1; 6,5 | 3,1 | 2,4; 3,8 |
| 2005 | 146 | 3,4 | 2,0; 4,8 | 6,5 | 3,7; 7,0 | 3,1 | 2,4; 4,3 |
| 2006 | 65 | 0,2 | 0; 0,4 | 7,5 | 6,8; 7,9 | 3,8 | 3,1; 4,2 |
| 2007 | 0 | – | – | – | – | – | – |
| 2008 | 0 | – | – | – | – | – | – |
| 2009 | 0 | – | – | – | – | – | – |
| 2010 | 0 | – | – | – | – | – | – |
| 2011 | 0 | – | – | – | – | – | – |
| 2012 | 0 | – | – | – | – | – | – |
| 2013 | 63 | 5,5 | 3,2; 7,8 | 7,1 | 6,4; 7,5 | 2,8 | 2,4; 3,1 |
| 2014 | 199 | 4,2 | 2,7; 5,6 | 6,0 | 5,7; 6,2 | 3,0 | 2,8; 3,2 |
| 2015 | 45 | 5,6 | 2,6; 8,7 | 6,2 | 4,5; 6,5 | 2,7 | 2,2; 3,4 |
| 2016 | 47Tableau 1 Note de bas de page c | 5,6 | 2,3; 9,0 | 7,0 | 3,1; 8,2 | 3,0 | 2,5; 3,8 |
| 2017 | 46Tableau 1 Note de bas de page c | 5,4 | 3,2; 7,7 | 6,5 | 5,1; 7,7 | 3,5 | 3,2; 3,9 |
| 2018 | 41Tableau 1 Note de bas de page c | 11,8 | 0; 91,1 | 6,2 | 4,7; 7,5 | 3,1 | 2,6; 3,5 |
| Analyse globale de type 3 : Khi-carré (valeur-p)Tableau 1 Note de bas de page b | – | 28,5 | 0,008 | 61,1 | < 0,000 1 | 53,9 | < 0,000 1 |
| MoisTableau 1 Note de bas de page d | |||||||
| Janvier | 0 | – | – | -10,8 | −19,7; −4,2 | 2,0 | 0,6; 5,3 |
| Février | 0 | – | – | -8,8 | −16,9; −4,1 | 2,1 | 0,9; 5,4 |
| Mars | 0 | – | – | -3,8 | −7,3; 0,5 | 2,0 | 0,8; 5,0 |
| Avril | 0 | – | – | 5,4 | −3,0; 7,8 | 3,6 | 0,2; 7,4 |
| Mai | 96 | 0 | 0; 37,7 | 13,4 | 7,5; 16,6 | 3,2 | 0,7; 6,8 |
| Juin | 1 518 | 0 | 0; 0,1 | 18,7 | 13,1; 21,4 | 4,2 | 1,5; 7,6 |
| Juillet | 3 611 | 0,8 | 0,4; 1,2 | 21,1 | 18,0; 23,5 | 3,2 | 1,6; 7,2 |
| Août | 4 688 | 5,5 | 4,3; 6,7 | 20,1 | 16,4; 22,3 | 3,5 | 1,7; 6,2 |
| Septembre | 3 675 | 6,0 | 4,4; 7,5 | 16,1 | 12,4; 18,6 | 2,9 | 0,8; 7,7 |
| Octobre | 242 | 1,9 | 0; 4,4 | 9,0 | 3,4; 12,9 | 3,4 | 1,2; 6,8 |
| Novembre | 0 | – | – | 1,4 | −2,3; 4,4 | 3,0 | 0,3; 5,6 |
| Décembre | 0 | – | – | −6,5 | −11,8; 1,8 | 3,2 | 0,6; 7,1 |
| Analyse globale de type 3 : Khi-carré (valeur-p)Tableau 1 Note de bas de page b | – | 15,6 | < 0,000 1 | s.o.Tableau 1 Note de bas de page e | s.o.Tableau 1 Note de bas de page e | s.o.Tableau 1 Note de bas de page e | s.o.Tableau 1 Note de bas de page e |
Les variables explicatives comprennent les températures journalières moyennes (Tmoyenne = Tmaximale+Tminimal/2) (°C) et les précipitations (mm) journalières extraites du Oak Ridge National Laboratory Distributed Active Archive Center (http://daymet.ornl.org) selon les coordonnées du système de géopositionnement par satellite (GPS) de chaque piège-année (latitude et longitude). Ainsi, les températures journalières moyennes et la somme des précipitations ont été regroupées selon cinq échelles temporelles différentes : 1) les mois précédents; 2) saisons précédentes : été (juin et juillet), printemps (mars à mai), hiver (décembre à février) et automne (septembre à novembre); ainsi que des regroupement de 3) quatre mois précédents (novembre de l'année précédente à février de l'année en cours); 4) six mois précédents (septembre de l'année précédente à février de l'année en cours) et 5) dix mois précédents (septembre de l'année précédente à juillet de l'année en cours), d'après la littérature scientifique suggérant que les conditions météorologiques ont une incidence sur la transmission du VNO dans les mois à venirNote de bas de page 11Note de bas de page 22. Pour les huit pièges avec données météorologiques manquantes, les données des pièges voisins les plus proches leurs ont été attribuées, avec la moitié de ces pièges séparés par plus d'un kilomètre de leur piège apparié. Pour tenir compte des variations dans la force d'échantillonnage géographique et temporel des moustiques, deux nouvelles variables de « forces d'échantillonnage » ont été créées pour chaque RSS (figure 2) en fonction du nombre de lots de moustiques testés : 1) annuellement; et 2) au cours de la période de 2003 à 2018 (tableau 1).
Figure 2 : Carte des 18 régions sociosanitaires (RSS) de la province de Québec selon le nombre de lots de moustiques testés entre les périodes de 2003 à 2006 et de 2013 à 2018
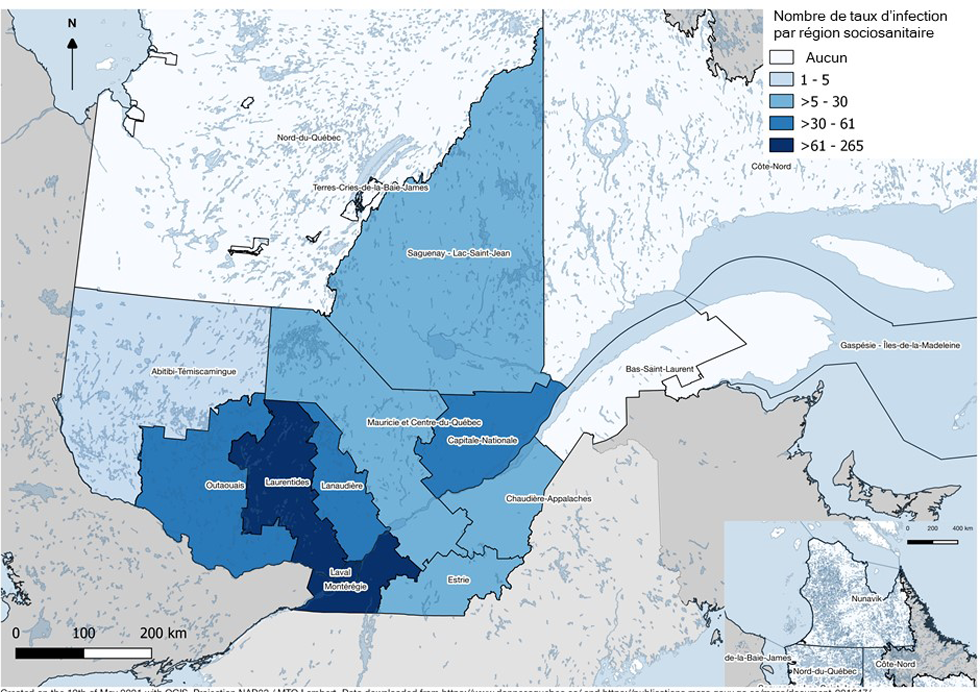
Description textuelle : Figure 2
Il s'agit d'une carte des 18 régions sociosanitaires du Québec qui sont colorées en fonction du nombre de lots de moustiques qui ont été analysés, afin de montrer les biais d'échantillonnage géographique. Du jeu de données final, composés des 856 taux d'infection minimaux chez les moustiques, quatre régions sociosanitaires (Montréal, Laval, Laurentides et Montérégie) représentent entre 62 et 265 observations chacune. Les régions sociosanitaires de l'Outaouais, Lanaudière et Capitale-Nationale représentent entre 31 et 61 observations chacune, celles de l'Estrie, la Mauricie-Centre-du-Québec, Chaudière-Appalaches et Saguenay-Lac-Saint-Jean représentent entre 6 et 30 observations tandis que l'Abitibi-Témiscamingue représente entre 1 et 5 observations. Aucune surveillance entomologique n'a eu lieu dans les régions du Bas-Saint-Laurent, Gaspésie-Iles-de-la-Madeleine, Côte-Nord, Nord-du-Québec, Nunavik et des Terres-Cries-de-la-Baie-James.
Des analyse bivariées entre les TIM annuels de Cx. pipiens-restuans (issue) et les variables explicatives (RSS, année et mois) ont été effectuées à l'aide d'un modèle de régression linéaire, employant une matrice de covariance autorégressive et où les pièges entomologiques déployés dans la même ville ont été imbriqués dans chaque RSS, pour tenir compte de leur proximité. Ensuite, des modèles de régression multivariée ont évalué la capacité prédictive de chaque regroupement temporel de température et de précipitation, à estimer les TIM, grâce aux observations réparties aléatoirement entre les jeux de données d'entraînement (n = 2/3) et de validation (n = 1/3). Une sélection descendante de variables, avec un maximum de quatre termes polynomiaux, pour obtenir les critères d'information bayésienne les plus bas, a été effectuée avec les données d'entrainement. Ensuite, les variables sélectionnées ont été employées dans un modèle de régression de Poisson, à l'aide des données de validationNote de bas de page 23. Les TIM ont été estimés en utilisant le nombre de lots positifs de Cx. pipiens-restuans sur le logarithme du nombre de moustiques testés, tandis que des pièges entomologiques dans la même ville ont été imbriqués dans chaque RSS pour tenir compte des données corrélées, à l'aide d'une matrice de covariance autorégressive. La méthodologie associée aux analyses de sensibilité et post hoc sont décrites en annexe. La macro SAS (%RsquareV) a été utilisée pour calculer le coefficient de détermination (R2), reflétant la proportion de la variance du TIM expliquée par les prédicteursNote de bas de page 24 et seuls les R2 ≥ 0,30 sont décrits dans le texte. Les intervalles de confiance (IC) à 95 % et les valeurs-p sont présentés, et tous les postulats associés aux modèles de régression ont été inspectés visuellementNote de bas de page 25. L'analyse statistique a été effectuée à l'aide de la version 9.4 du logiciel SAS/STATMD (SAS Institute, Cary, Caroline du Nord, États-Unis).
Résultats
Depuis 2003, 172 094 moustiques Cx. pipiens-restuans ont été analysés par réaction en chaîne par polymérase (PCR). Pour les 856 pièges-année à l'étude, le TIM moyen était de de 3,1 pour 1 000 moustiques (IC à 95 %; 2.2–4.0) et a varié en variaient en fonction de l'année (χ2 = 28,5; p = 0,0008) et du mois (χ2 = 15,6; p < 0,0001) d'échantillonnage, mais pas selon les RSS (χ2 = 9,9; p = 0,5) (tableau 1). Pour la période de l'étude, les températures mensuelles moyennes à proximité des pièges entomologiques variaient de 23,5°C (juillet) à −19,7°C (janvier), tandis que les précipitations mensuelles cumulatives variaient de 0,3 mm (novembre) à 7,7 mm (septembre).
La capacité prédictive des températures moyennes et des précipitations antérieures à prédire les TIM saisonniers était la plus élevée en utilisant les mois précédents de mars (R2 = 0,37), mai (R2 = 0,36) et décembre (R2 = 0,35) précédents, ainsi que pour la saison automnale précédente (R2 = 0,38). Les autres coefficients de détermination étaient tous inférieurs à 0,30 (tableau 2). Les résultats des analyses de sensibilité et post hoc sont décrits dans l'annexe. En résumé, l'inclusion de la variable « année d'échantillonnage » dans les principaux modèles de régression a augmenté tous les R2 (intervalle de 0,42 à 0,57) alors que cette augmentation était moins importante lorsque la variable géographique « RSS » a été incluse (intervalle : 0,18 à 0,46) (tableau A1).
| Modèles de régression et équations de l'ensemble de données de formation | Coefficient de détermination de l'ensemble de données de validation (Penalised R2) |
|
|---|---|---|
| A) Par mois précédent | ||
| Juillet | TIM = -915,67 + 130.81Temp - 6.45Temp2 + 0.11Temp3 + 35.05Prec – 14.65Prec2 + 2.58Prec3 – 0.17Prec4 + ε | 0,28 |
| JuinTableau 2 Note de bas de page a | TIM = -40,79 + 3.54Temp – 0.10Temp2 + 0.95Prec – 0.09Prec2 + ε | 0,15 |
| MaiTableau 2 Note de bas de page a | TIM = 13,60 – 2.13Temp + 0.09Temp2 – 7.98Prec + 2.48Prec2 – 0.24Prec3 + ε | 0,36 |
| Avril | TIM = -7,64 – 0.27Temp + 2.47Prec – 0.69Prec2 + 0.08Prec3 – 0.003Prec4 + ε | 0,19 |
| Mars | TIM = -22,03 – 4.28Temp – 2.31Temp2 - 0.44Temp3 – 0.03Temp4 + 30.83Prec – 22.71Prec2 + 6.79Prec3 – 0.70Prec4 + ε | 0,37 |
| Février | TIM = -2,85 + 2.38Temp + 0.20Temp2 – 0.01Temp3 + ε | 0,10 |
| Janvier | TIM = -47,75 — 17.42Temp – 2.65Temp2 - 0.18Temp3 – 0.004Temp4 + 0.34Prec – 0.04Prec2 + ε | 0,24 |
| Décembre | TIM = -6,93 + 0.21Temp – 0.05Temp2 – 0.01Temp3 + 2.67Prec – 0.79Prec2 – 0.06Prec3 + ε | 0,35 |
| Novembre | TIM = -4,74 – 0.07Temp – 0.46Temp2 + 0.12Temp3 + 2.61Prec + 2.74Prec2 – 0.65Prec3 + 0.09Prec4 + ε | 0,18 |
| Octobre | TIM = -7,48 + 0.20Temp – 0.83Prec + 0.47Prec2 + 0.06Prec3 + ε | 0,28 |
| Septembre | TIM = -6,02 + 0.16Temp – 1.75Prec + 0.26Prec2 + ε | 0,28 |
| Août | TIM = -13,09 + 0.40Temp – 0.24Prec + ε | 0,08 |
| B) Par saisonTableau 2 Note de bas de page b | ||
| ÉtéTableau 2 Note de bas de page a | TIM = -45,53 + 4.37Temp – 0.11Temp2 – 3.25Prec + 0.46Prec2 + ε | 0,15 |
| PrintempsTableau 2 Note de bas de page c | TIM = -206,81 + 84.33Temp – 25.56Temp2 + 3.36Temp3 – 0,16Temp4 + 132.67Prec – 64.01Prec2 + 13.26Prec3 – 1.00Prec4 + ε | 0,09 |
| Hiver | TIM = 2,31 + 0.89Temp + 0.05Temp2 – 6.40Prec + 3.04Prec2 – 0.46Prec3 + ε | 0,24 |
| Automne | TIM = -438,86 + 153.19Temp – 17.32Temp2 + 0.65Temp3 – 15.35Prec + 4.79Prec2 – 0.50Prec3 + ε | 0,38 |
| C) De novembre de l'année précédente à février de la même année | ||
| Quatre mois regroupés | TIM = -48,07 – 2.87Temp – 1.24Temp2 – 0.18Temp3 – 0.01Temp4 + 54.97Prec – 26.15Prec2 + 15.35Prec3 – 0.41Prec4 + ε | 0,24 |
| D) De septembre de l'année précédente à février de la même année | ||
| Six mois regroupés | TIM = 9,37 + 0.48Temp – 0.23Prec + 13.46Prec2 – 3.51Prec3 + 0.33Prec4 + ε | 0,27 |
| E) De septembre de l'année précédente à juillet de la même annéeTableau 2 Note de bas de page c | ||
| Dix mois regroupés | TIM = -119,02 — 113.69Temp – 41.82Temp2 + 6.68Temp3 – 0.39Temp4 + ε | 0,12 |
Discussion
C'est la première fois qu'un outil d'alerte précoce basé sur les données météorologiques pour prédire les TIM annuels chez Cx. pipiens-restuans au Québec est exploré, utilisant les données de surveillance provinciale du VNO. La capacité des températures moyennes et de l'accumulation des précipitations à prédire les TIM était la plus élevée lorsque ces données ont été regroupées pour les mois précédents de mars, de décembre et de mai, ainsi que pour la saison précédente d'automne (septembre à novembre de l'année précédente). Cependant, la capacité prédictive du modèle est probablement trop faible pour être utile et employé comme système d'alerte précoce. Le fait que seulement ~40 % de la variance des TIM annuels de Cx. pipiens-restuans peuvent être expliquées par la température moyenne et les précipitations, suggère que d'autres facteurs liés à l'épidémiologie du VNO doivent être intégrés à ce type d'outil prédictifNote de bas de page 15.
De nombreux systèmes d'alerte ou de détection précoce, basé principalement ou uniquement sur des données météorologiques, ont été mis au point pour prédire d'autres maladies transmises par les moustiques (e.g. Chikungunya, dengue, malaria, fièvre jaune et Zika) afin d'anticiper les activités de contrôle antivectorielleNote de bas de page 24. Nos résultats appuient que les capacités de prédiction insuffisantes de ces modèles ne peuvent être utiles en temps réel pour le MSSS. L'épidémiologie du VNO est complexe et l'ajout de variables supplémentaires, comme les facteurs environnementaux (e.g. l'habitat potentiel ou la distribution des vecteurs et des réservoirs, l'indice de végétation, l'utilisation du territoire) et des données concernant les hôtes aviaires (abondance, migrations), améliore habituellement les prédictionsNote de bas de page 15. Cependant, l'intégration des données du système de surveillance en cours de saison et le raffinement de l'échelle géographique mène généralement au développement d'outils de prédiction moins facile à utiliser pour les autorités de santé publiqueNote de bas de page 26.
Forces et faiblesses
L'une des forces de ce projet est que la proximité géographique des pièges entomologiques a été prise en compte dans la plupart des modèles de régression, car cela peut biaiser les résultats. De plus, les analyses de sensibilité multiples et post hoc ont amené de la robustesse aux résultats. Les équations polynomiales reflètent les associations non linéaires entre les variables météorologiques et les TIM, permettant une meilleure représentation de leur relation et une meilleure adéquation du modèle. De plus, la sélection descendante des variables explicatives s'est avérée aussi efficace que l'inspection visuelle des modèles additifs généralisés. Étant donné que les TIM représentent de faibles valeurs numériques, la puissance statistique a été augmentée en calculant les TIM annuels, seulement à l'aide de moustiques collectés en août et septembre. Bien que les TIM annuels moyens aient augmenté pour la plupart des regroupements de lots de moustiques positifs, le coefficient de détermination pour la plupart des modèles de régression a diminué de façon inattendue. Il est possible que la taille d'échantillonnage se limitant aux mois d'août et de septembre ait réduit la puissance statistique, puisque 13 pièges entomologiques n'avaient pas de moustiques capturés.
D'après notre cadre conceptuel, un biais de surajustement a pu s'introduire lors de l'ajustement pour « l'année d'échantillonnage » dans les modèles de régressionNote de bas de page 27. Malgré de multiples analyses de sensibilité avec des variables de confusion potentielles, de la confusion résiduelle ne peut être écartéeNote de bas de page 28. Comme les lots de moustiques étaient regroupés pour chaque année de piégeage, il n'était pas possible de tenir compte des différents types de pièges employés. Cette limitation a probablement un effet négligeable, puisque 85,2 % des moustiques ont été capturés par les pièges « CDC-light ». Aussi, il n'a pas été possible de tenir compte des applications de larvicides pendant les saisons 2003–2005 et 2013–2014, ce qui aurait pu influencer les estimés. Étant donné que les tests par PCR sont effectués indépendamment du piégeage des moustiques, si un biais de mesure devait être présent, une diminution de la force de l'association serait attendue.
Un biais de sélection aurait pu s'introduire dans les résultats en raison de la répartition géographique des pièges qui est liée à 1) l'incidence antérieure du VNO et 2) l'influence de la météo sur l'activité physique humaine et l'exposition aux moustiquesNote de bas de page 29Note de bas de page 30. De plus, l'emplacement fixe des pièges entomologiques depuis 2017 se fait au détriment de la détection de l'émergence possible du VNO dans les RSS moins densément peuplés ou situées à des latitudes plus septentrionales. Le nombre limité de pièges entomologiques sur un territoire aussi vaste et l'absence de surveillance des moustiques pendant les mois les plus froids diminuent la puissance des analyses statistique. Ainsi, un plus grand nombre de pièges répartis sur une zone géographique plus large au Québec, permettrait d'obtenir des données météorologiques plus hétérogènes et d'augmenter la puissance des analyses. Puisque les vecteurs de VNO varient selon les régions et que sa transmission semble être influencées par des variables environnementales et géographiques à plus fines échelles que les RSS, les résultats de ce projet ne peuvent être généralisés à d'autres situations. Malgré ces limites, les données de surveillance du VNO au Québec continueront d'être explorées. Les prochaines étapes consisteront à modéliser directement l'incidence du VNO chez l'humain employant les données météorologiques afin de contourner les données entomologiques manquantes entre 2007 et 2012 et d'explorer l'élaboration d'un outil de système de détection précoce.
Conclusion
Tous les modèles de régression étudiés ont des capacités prédictives trop faibles pour être utiles comme outil de santé publique. D'autres facteurs associés à l'épidémiologie du virus du Nil occidental doivent être ajoutés au modèle d'alerte précoce basé sur les données météorologiques pour qu'il soit utile aux autorités sanitaires provinciales.
Déclaration des auteurs
K. F. B., N. O., A. L. et A. I. C. — Conception du projet
E. S. L. — Gestion de la base de données et des analyses statistiques
J. D. — Conception du projet, gestion de la base de données et des analyses statistiques, interprétation des résultats, écriture de l'ébauche originale
Tous les coauteurs ont révisé et commenté le texte.
Intérêts concurrents
Aucun.
Remerciements
Nous tenons à remercier le ministère de la Santé et des Services qui finance le système de surveillance du virus du Nil occidental depuis 2003, qui comprend l'échantillonnage annuel de moustiques par une entreprise privée, l'analyse par PCR des lots de moustiques effectuée par le Laboratoire de santé publique du Québec (LSPQ), ainsi que l'hébergement, la gestion et l'analyse des données par l'Institut national de santé publique du Québec (INSPQ).
Financement
Ce projet a été financé par l'Agence de la santé publique du Canada par le « Protocole d'accord relatif à la recherche sur les risques d'émergence de maladies à transmission vectorielle en raison des changements climatiques 2020–2022 » conclu avec l'Institut national de santé publique du Québec (INSPQ).
Annexe
Méthodes
Étant donné que les cas humains d'infection causées par le virus du Nil occidental (VNO) sont habituellement déclarés au début du mois d'aoûtNote de bas de page 1 et que l'objectif est de mettre au point un outil d'alerte précoce capable de prédire ces cas humains quelques mois avant qu'ils ne se produisent, les températures et les précipitations du mois d'août n'ont pas été incluses dans les analyses.
Une revue de la littérature scientifique a été effectuée pour élaborer un graphe acyclique dirigé (figure A1) afin de conceptualiser les relations entre les facteurs potentiellement associés au VNO et d'identifier les variables qui doivent être ajustées dans le modèle à l'aide de l'outil DAGitty.
Figure A1 : Cadre conceptuel utilisant un graphe acyclique dirigé entre les variables météorologiques et les principaux indicateurs du système de surveillance intégrée du virus du Nil occidental du Québec

Description textuelle : Figure A1
Cette figure représente un graphe acyclique dirigé montrant les différentes relations entre les températures journalières moyennes et les précipitations cumulatives, qui sont les variables explicatives et l'issue, qui est le taux d'infection minimal des moustiques du complexe Culex pipiens-restuans. Basé sur la littérature scientifique, ce graphe représente le cadre conceptuel qui aide à guider l'analyse statistique, car il permet d'identifier toutes les autres variables à inclure dans les modèles. Les autres variables qui sont incluses dans le graphique sont l'année d'échantillonnage, le nombre d'oiseaux infectés (agissant comme réservoir du virus), le nombre de cas humains et animaux (agissant comme hôtes accidentels), le plan d'échantillonnage entomologique (géographique et temporel) et les variables environnementales non mesurées.
Analyses de sensibilité et post hoc
Sept analyses de sensibilité ont été effectuées comme suit : 1) des modèles de régression ont été ajustés à l'aide d'une inspection visuelle des graphiques générés par un modèle additif généralisé avec un maximum de quatre degrés de lissage effectué par une validation croisée généralisée; 2) les principaux modèles ont été réanalysés sans les données des quatre pièges entomologiques (VBL 034, VBL 904, SHA 002 et WEN 001) qui ont hérité des données météorologiques de leur voisin le plus proche avec un rayon supérieur à 1 km2; 3) en incluant la variable de force d'échantillonnage 2003–2018 pour chaque RSS; 4) en incluant la force d'échantillonnage annuelle pour chaque RSS; 5) en attribuant le TIM des moustiques piégés seulement pendant le mois d'août et de septembre au lieu de pendant tout l'été; 6) en ajoutant l'année d'échantillonnage dans le modèle; et 7) en ajoutant le RSS dans le modèle plutôt que comme mesure répétée. À la suite des résultats de la 6e analyse de sensibilité, l'année d'échantillonnage a été utilisée comme substitut pour les facteurs de confusion non mesurés. Une analyse post hoc avec un modèle de Poisson a été effectuée en modélisant directement le TIM en utilisant l'année d'échantillonnage comme variable explicative catégorique. Pour mesurer si les variables météorologiques étaient des facteurs de confusion de cette association, l'année d'échantillonnage a été incluse comme variable continue afin de faciliter le calcul et l'interprétation de la différence et du rapport entre les deux estimations (βannée d'échantillonnage). La confusion était présente en fonction d'une différence ou d'un rapport des estimations supérieures à 0,1 décimal ou 10 %Note de bas de page 31. Quatre analyses bivariées additionnelles ont été effectuées avec l'année de l'échantillonnage comme variable explicative continue et, respectivement, la température annuelle moyenne, la précipitation annuelle moyenne, la force d'échantillonnage annuelle et la force d'échantillonnage du RSS comme issues principales. Une distribution normale, un lien « identité » et d'une matrice de covariance autorégressive avec une imbrication des pièges entomologiques déployés dans la même ville à l'intérieur de chaque RSS pour tenir compte des corrélations, a permis d'obtenir des estimations représentant des différences dans les taux d'infection minimaux (TIM).
Résultats
Analyses de sensibilité
Les modèles de régression ajustés en fonction d'une inspection visuelle des graphiques provenant des modèles additifs généralisés ont conduit à des résultats semblables à ceux de l'analyse principale, sauf pour le mois de février (R2 est passé à 0,22) et août (R2 est passé à 0,24), ainsi que pour les dix mois antécédents regroupés (R2 est passé à 0,18), alors que toutes les autres comparaisons présentaient une différence de cinq décimales ou moins (tableau A1). Si les quatre pièges entomologiques avec des données météorologiques empruntées au piège voisin le plus proche au-delà de 1km2 sont exclu, les résultats ont varié pour les mois de mai (R2 a diminué à 0,19) et de septembre (R2 a augmenté à 0,38), l'été (R2 a augmenté à 0,29) et l'hiver (R2 a diminué à 0,16) (tableau A1). Lorsque la force d'échantillonnage pour chaque RSS a été incluse, tous les R2 se situaient à l'intérieur de cinq décimales des principaux résultats (tableau A1). Inclure la variable reflétant la force annuelle de l'échantillonnage a augmenté les capacités prédictives d'au moins cinq décimales pour neuf regroupements (juin, avril, février, novembre, août; été et printemps; de six et dix mois). L'utilisation des taux d'infection minimaux de Cx. pipiens-restuans piégés en août et septembre au lieu de tout l'été, ont influencé les R2 pour 14 regroupements temporels (diminuée pour mai, avril, décembre, novembre, septembre, été, hiver, automne, quatre et six mois et augmenté pour juin, octobre, août et printemps). Lorsque l'année d'échantillonnage a été incluse dans les modèles de régression de l'analyse principale, tous les coefficients de détermination ont augmenté entre 0,42 et 0,57. L'inclusion de la variable des RSS dans les principaux modèles de régression a augmenté la capacité prédictive d'au moins cinq décimales pour tous les regroupements temporels sauf juillet, mai et l'automne.
| Modèles de régression | Analyses de sensibilitéTableau A1 Note de bas de page a | ||||||
|---|---|---|---|---|---|---|---|
| Utilisant des modèles d'additifs généralisés pour les spécifications du modèle | Sans les quatre pièges entomologiques | Incluant la force d'échantillonnage pour chaque RSS | Incluant la force d'échantillonnage annuelle | Utilisant les TIM d'août et de septembre | Ajoutant l'année d'échantillonnage dans le modèle principal | Ajoutant le RSS dans le modèle principal | |
| A) Par mois précédent | |||||||
| Juillet | 0,28 | 0,28 | 0,27 | 0,30 | 0,31 | 0,56↑ | 0,29 |
| JuinTableau A1 Note de bas de page b | 0,18 | 0,15 | 0,17 | 0,21↑ | 0,24↑ | 0,53↑ | 0,24↑ |
| MaiTableau A1 Note de bas de page b | 0,36 | 0,19↓ | 0,35 | 0,39 | 0,30↓ | 0,53↑ | 0,35 |
| Avril | 0,23 | 0,19 | 0,19 | 0,31↑ | 0,13↓ | 0,55↑ | 0,31↑ |
| Mars | 0,37 | 0,37 | 0,38 | 0,34 | 0,41 | 0,53↑ | 0,46↑ |
| Février | 0,22↑ | 0,22↑ | 0,10 | 0,20↑ | 0,09 | 0,53↑ | 0,22↑ |
| Janvier | 0,26 | 0,24 | 0,24 | 0,27 | 0,21 | 0,57↑ | 0,30↑ |
| Décembre | 0,33 | 0,34 | 0,37 | 0,36 | 0,29↓ | 0,53↑ | 0,41↑ |
| Novembre | 0,18 | 0,18 | 0,19 | 0,33↑ | 0,04↓ | 0,54↑ | 0,36↑ |
| Octobre | 0,28 | 0,28 | 0,27 | 0,30 | 0,34↑ | 0,51↑ | 0,41↑ |
| SeptembreTableau A1 Note de bas de page b | 0,29 | 0,38↑ | 0,30 | 0,35↑ | 0,21↓ | 0,57↑ | 0,38↑ |
| Août | 0,24↑ | 0,11 | 0,09 | 0,23↑ | 0,24↑ | 0,48↑ | 0,20↑ |
| B) Par saisonTableau A1 Note de bas de page c | |||||||
| Été | 0,14 | 0,29↑ | 0,15 | 0,24Tableau A1 Note de bas de page d↑ | 0,02↓ | 0,55↑ | 0,27Tableau A1 Note de bas de page e↑ |
| Printemps | 0,09 | 0,09 | 0,09 | 0,18↑ | 0,15Tableau A1 Note de bas de page e↑ | 0,53↑ | 0,23Tableau A1 Note de bas de page e↑ |
| Hiver | 0,20 | 0,16↓ | 0,23 | 0,26 | 0,08↓ | 0,51↑ | 0,32↑ |
| Automne | 0,38 | 0,38 | 0,38 | 0,40 | 0,27↓ | 0,53↑ | 0,41 |
| C) Groupe de quatre mois | |||||||
| De novembre de l'année précédente à février de l'année en cours | 0,26 | 0,24 | 0,24 | 0,23 | 0,14↓ | 0,57↑ | 0,35↑ |
| D) Groupe de six mois | |||||||
| De septembre de l'année précédente à février de l'année en cours | 0,27 | 0,26 | 0,29 | 0,34↑ | 0,19↓ | 0,56↑ | 0,36↑ |
| E) Dix mois regroupésTableau A1 Note de bas de page dTableau A1 Note de bas de page bTableau A1 Note de bas de page d | |||||||
| De septembre de l'année précédente à juillet de l'année en cours | 0,18↑ | 0,12Tableau A1 Note de bas de page e | 0,13Tableau A1 Note de bas de page e | 0,20↑ | 0,17 | 0,42↑ | 0,18↑ |
Analyse post hoc
Lorsqu'on modélise directement le TIM en utilisant seulement la variable « année de d'échantillonnage », le R2 était de 0,53. L'estimé associée à l'année d'échantillonnage, sous forme de variable linéaire continue, et comme seule variable explicative des TIM de Cx. pipiens-restuans étaient de 1,09 (IC à 95 %; 1,06–1 117; p < 0,0001) tandis que des fluctuations annuelles sont observées lorsqu'on les traite sous forme catégorique (résultats non présentés). Les résultats des estimés obtenues dans tous les principaux modèles de régression à l'aide des variables météorologiques et de l'année d'échantillonnage variaient entre 1,06 et 1,15, avec des effets de confusion très faibles sur les échelles de différence et de rapport (tableau A2). Lorsqu'on compare les TIM entre les années et en utilisant 2018 comme référence, toutes les années d'échantillonnage antérieures avaient des valeurs inférieures sauf en 2016, puisque l'IC supérieur de 95 % de l'estimation chevauche la valeur nulle (tableau A3). Lorsque l'année d'échantillonnage est traitée comme variable continue, le nombre de pièges entomologiques a diminué de façon linéaire de 2003 à 2018 (β = −1.13 [−1,45–0,81; p < 0,0001]). Par conséquent, le nombre de pièges entomologiques et le TIM sont inversement corrélés entre 2003 et 2018, avec une augmentation de 2 % dans les TIM pour chaque piège entomologique déployé de moins, dans le temps (β = 0.98 [0,98–0,99; p < 0,0001]). Au niveau de chaque piège entomologique déployé au Québec, les températures annuelles moyennes ont augmenté (β = 0,04 [0,02–0,06; p = 0,0003]), tandis que les précipitations annuelles moyennes ont diminué (β = −0,008 [−0,02–0,006; p = 0,07]) de 2003 à 2018.
| Modèles de régression | Estimés pour l'année d'échantillonnage en tant que variable continue avec les données météorologiques | Valeur-p | Différence entre les estimésTableau A2 Note de bas de page a | Rapport de risque dans les estimés pour l'année d'échantillonnageTableau A2 Note de bas de page b | |
|---|---|---|---|---|---|
| n | IC à 95 % | ||||
| A) Par mois précédent | |||||
| Juillet | 1,08 | 1,04–1,12 | < 0,000 1 | 0,01 | 1,01 |
| JuinTableau A2 Note de bas de page c | 1,11 | 1,07–1,15 | < 0,000 1 | −0,02 | 0,98 |
| MaiTableau A2 Note de bas de page c | 1,09 | 1,06–1,12 | < 0,000 1 | 0,00 | 1,00 |
| Avril | 1,07 | 1,04–1,11 | 0,000 1 | 0,02 | 1,02 |
| Mars | 1,15 | 1,12–1,18 | < 0,000 1 | −0,06 | 0,95 |
| Février | 1,08 | 1,05–1,11 | < 0,000 1 | 0,01 | 1,01 |
| Janvier | 1,14 | 1,10–1,18 | < 0,000 1 | −0,05 | 0,96 |
| Décembre | 1,11 | 1,06–1,14 | 0,000 7 | −0,02 | 0,98 |
| Novembre | 1,06 | 1,03–1,09 | 0,000 3 | 0,03 | 1,03 |
| Octobre | 1,08 | 1,04–1,13 | < 0,000 1 | 0,01 | 1,01 |
| Septembre | 1,09 | 1,06–1,13 | < 0,000 1 | 0,00 | 1,00 |
| Août | 1,09 | 1,07–1,12 | < 0,000 1 | 0,00 | 1,00 |
| B) Par saisonTableau A2 Note de bas de page d | |||||
| ÉtéTableau A2 Note de bas de page c | 1,12 | 1,10–1,15 | < 0,000 1 | −0,03 | 0,97 |
| PrintempsTableau A2 Note de bas de page e | 1,09 | 1,06–1,1 | < 0,000 1 | 0,00 | 1,00 |
| Hiver | 1,12 | 1,07–1,16 | < 0,000 1 | −0,03 | 0,97 |
| Automne | 1,08 | 1,05–1,11 | < 0,000 1 | 0,01 | 1,01 |
| C) Groupe de quatre mois | |||||
| De novembre de l'année précédente à février de l'année en cours | 1,09 | 1,05–1,13 | < 0,000 1 | 0,00 | 1,00 |
| D) Groupe de six mois | |||||
| De septembre de l'année précédente à février de l'année en cours | 1,09 | 1,06–1,11 | < 0,000 1 | 0,00 | 1,00 |
| E) Groupe de dix moisTableau A2 Note de bas de page f | |||||
| De septembre de l'année précédente à juillet de l'année en cours | 1,08 | 1,06–1,10 | < 0,000 1 | 0,01 | 1,01 |
| Année d'échantillonnage | Taux d'infection minimal de Cx. pipiens-restuans pour 1 000 moustiques | Rapport du taux d'infection minimal | Valeur-p | ||
|---|---|---|---|---|---|
| n | Taux | n | Taux | ||
| 2003 | 3,0 | 1,7–4,4 | 0,24 | 0,16–0,34 | < 0,0001Tableau A3 Note de bas de page a |
| 2004 | 0,9 | 0–2,1 | 0,06 | 0,04–0,10 | < 0,0001Tableau A3 Note de bas de page a |
| 2005 | 3,4 | 2,0–4,8 | 0,21 | 0,12–0,36 | < 0,0001Tableau A3 Note de bas de page a |
| 2006 | 0,2 | 0–0,4 | 0,05 | 0,03–0,10 | < 0,0001Tableau A3 Note de bas de page a |
| 2013 | 5,5 | 3,2–7,8 | 0,41 | 0,26–0,66 | 0,0002Tableau A3 Note de bas de page a |
| 2014 | 4,2 | 2,7–5,6 | 0,31 | 0,20–0,46 | < 0,0001Tableau A3 Note de bas de page a |
| 2015 | 5,6 | 2,6–8,7 | 0,36 | 0,21–0,65 | 0,0005Tableau A3 Note de bas de page a |
| 2016 | 5,6 | 2,3–9,0 | 0,50 | 0,22–1,13 | 0,1 |
| 2017 | 5,4 | 3,2–7,7 | 0,53 | 0,35–0,80 | 0,002Tableau A3 Note de bas de page a |
| 2018 (Référence) | 11,8 | 0–91,1 | 1,00 | 1,00 | – |
| Analyse globale de type 3 (avec 9 degrés de liberté) | – | – | Χ2 = 17,98 | Χ2 = 17,98 | 0,04 |

Cette œuvre est mise à disposition selon les termes de la Licence Creative Commons Attribution 4.0 International